
ВПР по химии на тему «10 класс-проф»
Спецификация
контрольных измерительных материалов для проведения
в 2016 году итоговой диагностической работы
по ОРГАНИЧЕСКОЙ ХИМИИ
10 класс, профильный уровень
1. Назначение КИМ – оценить уровень общеобразовательной подготовки по курсу органической химии 10 класса на углублённом уровне. Результаты диагностической работы могут быть использованы для аттестации обучающихся за курс химии 10 класса.
2. Документы, определяющие содержание КИМ
Содержание диагностической работы определяется Федеральным компонентом государственного стандарта основного общего образования (приказ Минобразования России от 05.03.2004 № 1089 «Об утверждении Федерального компонента государственных стандартов начального общего, основного общего и среднего (полного) общего образования»).
3. Подходы к отбору содержания, разработке структуры КИМ диагностической работы
Разработка КИМ диагностической работы по химии осуществлялась с учётом следующих общих положений:
рассматривается в качестве инвариантного ядра содержания действующих программ по химии для средней школы. В Федеральном компоненте государственного образовательного стандарта по химии эта система знаний представлена в виде требований к подготовке выпускников.
Учебный материал, на базе которого строятся задания, отбирается по признаку его значимости для общеобразовательной подготовки учащихся X класса.
Отбор содержания, а также разработка структуры КИМ осуществляются в соответствии с указанными положениями, а также с учётом нормативных документов, определяющих структуру и содержание КИМ для проведения единого государственного экзамена (ЕГЭ) по химии ( www.fipi.ru).
Тексты заданий в КИМ в целом соответствуют формулировкам, принятым в учебниках, включённых в Федеральный перечень учебников, рекомендуемых Министерством образования и науки РФ к использованию при реализации имеющих государственную аккредитацию образовательных программ среднего (полного) общего образования.
4. Характеристика структуры и содержания КИМ
Каждый вариант экзаменационной работы содержит 27 заданий. В числе этих заданий:
задания с порядковыми номерами 1 –20 базового уровня сложности, ответ на которые записывается в виде одной цифры;
задания с порядковыми номерами 21 – 25 повышенного уровня сложности на установление соответствия между элементами двух множеств, ответ на которые записывается в виде последовательности трёх или четырёх цифр;
задания с порядковыми номерами 26 – 27 высокого уровня сложности, которые предполагают запись развёрнутого ответа.
5. Распределение заданий КИМ по содержанию, проверяемым умениям
и видам деятельности
В работе представлены задания, которые проверяют усвоение знаний по следующим элементам содержания курса органической химии:
Классификация органических веществ.
 Общие формулы классов органических веществ. Номенклатура органических веществ (тривиальная и международная).
Общие формулы классов органических веществ. Номенклатура органических веществ (тривиальная и международная).Теория строения органических соединений: гомология и изомерия (структурная и пространственная).
Характерные химические свойства углеводородов.
Характерные химические свойства кислородсодержащих органических веществ.
Характерные химические свойства азотсодержащих органических соединений: аминов и аминокислот.
Природные источники углеводородов, их переработка.
Высокомолекулярные соединения. Реакции полимеризации и поликонденсации. Полимеры.
Качественные реакции органических соединений.
Взаимосвязь органических соединений.
Расчёты массы вещества или объёма газов по известному количеству вещества, массе или объёму одного из участвующих в реакции веществ.
Нахождение молекулярной формулы вещества.
В процессе выполнения заданий диагностической работы учащиеся продемонстрируют сформированность следующих метапредметных и предметных умений:
называть изученные вещества по тривиальной или международной номенклатуре;
определять принадлежность веществ к различным классам органических соединений; гомологи и изомеры;
характеризовать строение и химические свойства изученных органических соединений;
объяснять зависимость свойств неорганических и органических веществ от их состава и строения;
планировать эксперимент по получению и распознаванию важнейших неорганических и органических соединений;
проводить вычисления по химическим формулам и уравнениям.

6. Распределение заданий КИМ по уровням сложности
В КИМ включены задания трёх уровней сложности (см. таблицу 1).
Таблица 1
Распределение заданий итоговой работы по уровням сложности
Уровень сложностизаданий
Количество
заданий
Максимальный
первичный балл
Процент от максимального пер-вичного балла за всю работу
Базовый (Б)
20
20
55,5
Повышенный (П)
5
10
27,8
Высокий (В)
2
6
16,7
Итого
27
36
100
7. Система оценивания выполнения отдельных заданий и итоговой диагностической работы в целом
Верное выполнение каждого из заданий 1 – 20 оценивается 1 баллом.
Верное выполнение каждого из заданий 21 – 25 максимально оценивается 2 баллами. Если в ответе на эти задания допущена только одна ошибка, то задание считается выполненным частично и оценивается 1 баллом.
Задания с развёрнутым ответом могут быть выполнены обучающимися разными способами. Поэтому приведённые в критериях оценивания образцы ответов следует рассматривать лишь как один из возможных вариантов ответа. Каждый верно выполненный элемент ответа оценивается 1 баллом, причём максимальный балл за каждое из заданий с порядковыми номерами 26 и 27 составляет 3 балла.
Таблица перевода баллов в отметки по пятибалльной шкале
Отметка по пятибалльной шкале«2»
«3»
«4»
«5»
Первичные баллы
1 – 10
11 – 20
21 – 31
32 – 36
8. Продолжительность итоговой диагностической работы
Продолжительность итоговой диагностической работы
На выполнение итоговой диагностической работы отводится 90 минут.
9. Дополнительные материалы и оборудование
В процессе выполнения итоговой диагностической работы по химии разрешается использовать следующие материалы и оборудование:
– Периодическая система химических элементов Д.И. Менделеева;
– таблица растворимости солей, кислот и оснований в воде;
– электрохимический ряд напряжений металлов;
– непрограммируемый калькулятор.
Приложение 1
Обобщенный план варианта контрольных измерительных материалов
для проведения итоговой диагностической работы
по ХИМИИ
10 класс, профильный уровень
Уровни сложности заданий: Б – базовый; П – повышенный, В – высокий.
| | Проверяемые элементы содержания | Коды прове-ряемых элементов содержа-ния по кодифи-катору | Коды требо-ваний | Уро-вень слож-ности зада- ния | Макс. | При-мерное время выпол-нения задания (мин.) |
| 1 | Классификация органических веществ. Общие формулы классов органических веществ. | 3.3 | 2.2.6 | Б | 1 | 2 |
| 2 | Функциональная группа | 3.2 | 1.1.1 | Б | 1 | 2 |
| 3 | Типы связей в молекулах органических веществ. | 3.2 | 2.2.2 | Б | 1 | 2 |
| 4 | Гибридизация атомных орбиталей углерода. | 3.2 | 2.2.3 | Б | 1 | 2 |
| | Номенклатура органических веществ (тривиальная и международная) | 3.3 | 2.2.7 | Б | 1 | 2 |
| 6 | Теория строения органических соединений: гомология и изомерия (структурная и пространственная). | 3.1 | 2.2.7 | Б | 1 | 2 |
| 7 | Характерные химические свойства и способы получения углеводородов: алканов | 3. | 2.3.4 | Б | 1 | 2 |
| 8 | Характерные химические свойства углеводородов: алкенов. Основные способы получения углеводородов | 3.4 4.1.7 | 2.3.4 1.3.4 | Б | 1 | 2 |
| 9 | Характерные химические свойства углеводородов: алкинов. Основные способы получения углеводородов | 3.4 4.1.7 | 2.3.4 1.3.4 | Б | 1 | 2 |
| 10 | Характерные химические свойства углеводородов: диенов. Основные способы получения углеводородов | 3.4 4.1.7 | 2.3.4 1.3.4 | Б | 1 | 2 |
| 11 | Характерные химические свойства углеводородов: аренов. Основные способы получения углеводородов | 3.4 4.1.7 | 2.3.4 1.3.4 | Б | 1 | 2 |
| 12 | Характерные химические свойства предельных одноатомных и многоатомных спиртов, фенола. Основные способы получения кислородсодержащих соединений. | 3.5 4. | 2.3.4 2.5.1 | Б | 1 | 2 |
| 13 | Характерные химические свойства альдегидов. Основные способы получения кислородсодержащих соединений. | 3.6 4.1.8 | 2.3.4 | Б | 1 | 2 |
| 14 | Характерные химические свойства предельных карбоновых кислот. Основные способы получения кислородсодержащих соединений. | 3.6 4.1.8 | 2.3.4 | Б | 1 | 2 |
| 15 | Характерные химические свойства сложных эфиров. Основные способы получения кислородсодержащих соединений. | 3.6 4.1.8 | 2.3.4 | Б | 1 | 2 |
| 16 | Природные источники углеводородов, их переработка Высокомолекулярные соединения. Реакции полимеризации и поликонденсации. Полимеры. | 4.2.3 4.2.4 | 1.3.3 | Б | 1 | 2 |
| 17 | Биологически важные вещества: жиры, белки, углеводы (моносахариды, дисахариды, полисахариды) | 3.8 | 2.3.4 | Б | 1 | 2 |
| 18 | Характерные химические свойства азотсодержащих органических соединений: аминов | 3. | 2.3.4 | Б | 1 | 2 |
| 19 | Характерные химические свойства азотсодержащих органических соединений: аминокислот | 3.7 | 2.3.4 | Б | 1 | 2 |
| 20 | Расчёты массы вещества или объёма газов по известному количеству вещества, массе или объёму одного из участвующих в реакции веществ | 4.3.3 | 2.5.2 | Б | 1 | 2 |
| 21 | Ионный (правило В.В. Марковникова) и радикальный механизмы реакций в органической химии | 1. | 2.2.3 | П | 2 | 5 |
| 22 | Характерные химические свойства углеводородов | 3.4 | 2.3.4 | П | 2 | 5 |
| 23 | Характерные химические свойства кислородсодержащих органических соединений | 3.5 | 2.3.4 | П | 2 | 5 |
| 24 | Классификация органических веществ. Номенклатура органических веществ (тривиальная и международная) | 3. | 2.1.1 | П | 2 | 5 |
| 25 | Качественные реакции органических соединений | 4.1.5 | 2.5.1 | П | 2 | 5 |
| 26 | Взаимосвязь органических соединений. | 3.9 | 2.3.4 2.4.3 | В | 3 | 10 |
| 27 | Нахождение молекулярной формулы вещества | 4.3.7 | 2.5.2 | В | 3 | 10 |
 балл за вы-полнение зада-ния
балл за вы-полнение зада-ния
 4
4
 1.8
1.8
 7
7 4.10
4.10 3
3ВПР
В МОУ Ликино-Дулёвская гимназия в марте — мае 2022 учебного года пройдут всероссийские проверочные работы в следующих классах:
4 класс – русский язык, математика, окружающий мир
5 класс – русский язык, математика, история, биология
6 класс – русский язык, математика, два предмета по выбору МО (география, обществознание, история, биология)
7 класс – русский язык, математика, иностранный язык (английский язык), обществознание, биология, география, физика, история
8 класс – русский язык, математика, два предмета по выбору МО (география, обществознание, история, биология, химия, физика)
4 класс
Математика | Русский | Окружающий мир
5 класс
Математика | История | Биология | Русский язык
6 класс
Математика | Обществознание | География | История | Биология | Русский язык
7 класс
Математика | Обществознание | География | История | Иностранные языки | Русский язык | Биология | Физика.
8 класс
Математика | Обществознание | География | История | Химия | Русский язык | Биология | Физика.
Перспективы ВПР на 2021-2022 учебный год
→ Формирование работ из банка заданий ВПР для 4-9, 11 классов, проведение в любой день в заданный промежуток.
→ Использование банка заданий ВПР при проведении процедур государственного контроля качества образования на региональном уровне.
ВПР пройдут в сентябре, октябре. ВПР не стоит бояться, к ним не нужно специально готовиться, нанимать репетиторов. По результатам ВПР не принимается никаких решений, влияющих на дальнейшую судьбу ребенка, получение аттестата, перевод в следующий класс, поступление в вуз. Нет никаких оснований бояться и переживать из-за ВПР больше, чем из-за самой обычной контрольной работы в школе.
ВПР не являются государственной итоговой аттестацией. ВПР учащиеся пишут в своих школах. Рекомендуемое время их проведения – второй-третий урок в школьном расписании; продолжительность – от одного до двух уроков.
РЕКОМЕНДАЦИИ РОДИТЕЛЯМ!
Обеспечьте дома удобное место для занятий, проследите, чтобы никто из домашних не мешал. Помогите детям распределить время для подготовки по дням. Во время интенсивного умственного напряжения необходима питательная и разнообразная пища. Найдите различные варианты заданий по предмету и потренируйте ребёнка.
Постарайтесь справиться со своими эмоциями, чтобы Ваше волнение не передалось ребенку и не увеличило его тревожность. Подбадривайте детей, хвалите их за то, что они делают хорошо. Повышайте их уверенность в себе, так как чем больше ребенок боится неудачи, тем больше вероятности допуска ошибок.
Говорите чаще детям:
- Ты у меня все сможешь.
- Я уверена, ты справишься с экзаменами.
- Я тобой горжусь.
- Что бы не случилось, ты для меня самый лучший
В беседах с ребенком старайтесь повысить его уверенность в хорошем результате ВПР.
И помните: самое главное — это снизить напряжение и тревожность ребенка и обеспечить подходящие условия для занятий.
Телефон «горячей линии» по вопросам проведения ВПР
8 (496) 422-46-08 – Жигунова Екатерина Павловна, муниципальный координатор проведения ВПР
8 (496) 414-17-06 – Ершова Ольга Владимировна, школьный координатор проведения ВПР
ВПР и НИКО в 2017-2018 учебном году
Муниципальное бюджетное общеобразовательное учреждение
«Средняя школа № 53»
Директор: Пузрякова Юлия Борисовна
Юридический адрес: 153034, город Иваново, улица Смирнова, дом 103
Фактический адрес: 153034, город Иваново, улица Смирнова, дом 103
Телефон доверия: 8 (4932) 32-90-63
Телефон: 8 (4932) 32-90-63
Режим работы: Понедельник-пятница с 8. 00 до 18.45
00 до 18.45
Адрес электронной почты: Этот адрес электронной почты защищен от спам-ботов. У вас должен быть включен JavaScript для просмотра.
Наименование банка: ОТДЕЛЕНИЕ ИВАНОВО БАНКА РОССИИ // УФК ПО ИВАНОВСКОЙ ОБЛАСТИ г. Иваново
Единый казначейский счет (ЕКС): 40102810645370000025
Номер казначейского счета: 03234643247010003300
БИК: 012406500
Учредитель: Управление образования администрации города Иваново
Начальник управления: Арешина Елена Васильевна
Адрес: 153000, г. Иваново, пл. Революции, д. 6
- Подробности
-
Создано 11.
 09.2017 10:00
09.2017 10:00
В интервью «РИА Новости» Сергей Кравцов, глава Рособрнадзора, сообщил о том, какие нововведения ожидаются в средних общеобразовательных учреждениях РФ в 2017-2018 учебном году в части процедур оценивания знаний школьников.
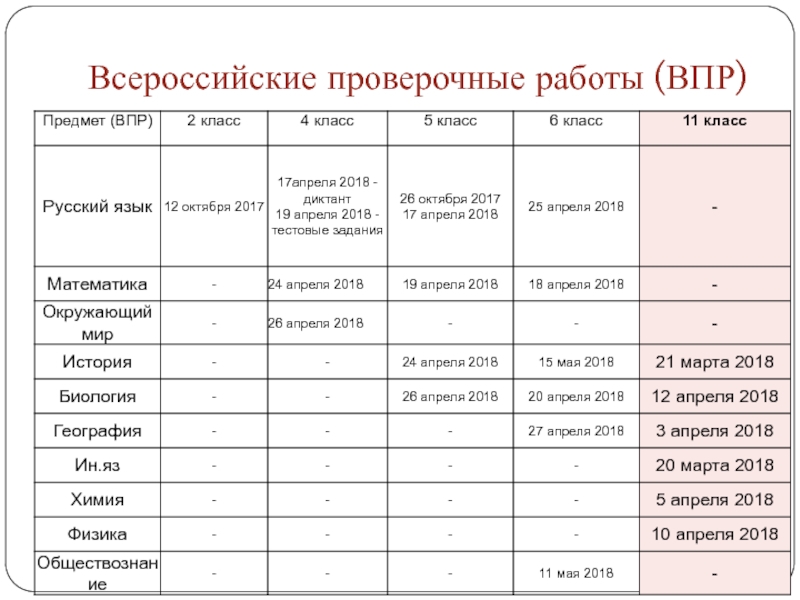
Как отметил руководитель надзорной службы, произойдёт увеличение количества участников Всероссийских проверочных работ (ВПР) и расширение перечня предметов. Так, помимо учащихся 2, 4, 5 и 11 классов в ВПР примут участие и шестиклассники, а к предыдущему списку предметов добавят обществознание и географию в 6-ом классе и иностранный язык – в 11 классе.
В соответствии с проектом график проведения ВПР выглядит следующим образом:
|
Предмет (ВПР) |
2 класс |
4 класс |
5 класс |
6 класс |
|
Русский язык |
3 октября 2017 |
17апреля 2018 – диктант 19 апреля 2018 — тестовые задания |
5октября 2017 |
25 апреля 2018 |
|
Математика |
|
24 апреля 2018 |
19 апреля 2018 |
18 апреля 2018 |
|
Окружающий мир |
|
26 апреля 2018 |
|
|
|
История |
|
|
24 апреля 2018 |
15 мая 2018 |
|
Биология |
|
|
26 апреля 2018 |
20 апреля 2018 |
|
География |
|
|
|
27 апреля 2018 |
|
Обществознание |
|
|
|
11 мая 2018 |
Даты проведения ВПР в 11 классах пока уточняются.
Кроме того, в текущем учебном году проведут национальные исследования качества образования (НИКО) по биологии и химии (10 класс), по литературе и мировой художественной культуре (6 и 8 классы).
Что же касается ЕГЭ, то, по словам Сергея Кравцова, изменений не ожидается.
В Рособрнадзоре планируют повсеместное распространение технологий печати КИМов (контрольно-измерительных материалов) в экзаменационных аудиториях и сканирования готовых работ участников ЕГЭ в ППЭ (пунктах проведения экзаменов).
Сайт учителя биологии, химии, географии.
Полезная информация ВПР по химии 2018 год.
Назначение ВПР по химии – оценить уровень общеобразовательной подготовки обучающихся 11 класса в соответствии с требованиями ФГОС. ВПР позволяют осуществить диагностику достижения предметных и метапредметных результатов, в том числе уровня сформированности универсальных учебных действий (УУД) и овладения межпредметными понятиями. Результаты ВПР в совокупности с имеющейся в образовательной организации информацией, отражающей индивидуальные образовательные траектории обучающихся, могут быть использованы для оценки личностных результатов обучения.
Официальная дата проведения экзамена: 5 апреля 2018 года
Тематика заданий ВПР по химии:
Варианты ВПР для самостоятельного решения с ответами:
Обязательные варианты ВПР по химии для решения на каникулах:
Вариант ВПР — 16 (Смотреть/скачать)
Вариант ВПР — 17 (Смотреть/скачать)
Вариант ВПР — 18 (Смотреть/скачать)
Сайт информационной поддержки проведения ВПР (образцы проверочных работ, ответы на часто задаваемые вопросы, др.) https://vpr.statgrad.org/
Скачать бесплатно описание ВПР 2018 по химии с ответами. 11 класс
Скачать бесплатно демоверсию ВПР 2018 по химии с ответами. 11 класс
ИНСТРУКЦИЯ ПО ВЫПОЛНЕНИЮ ВПР ПО ХИМИИ:
Всероссийская проверочная работа включает в себя 15 заданий. На выполнение работы по химии отводится 1 час 30 минут (90 минут). Ответы на задания записывайте в отведённом для них поле. В случае записи неверного ответа зачеркните его и запишите рядом новый.
На выполнение работы по химии отводится 1 час 30 минут (90 минут). Ответы на задания записывайте в отведённом для них поле. В случае записи неверного ответа зачеркните его и запишите рядом новый.
При выполнении работы разрешается использовать:
- Периодическую систему химических элементов Д.И. Менделеева;
- Таблицу растворимости солей, кислот и оснований в воде;
- Электрохимический ряд напряжений металлов;
- Непрограммируемый калькулятор.
При выполнении заданий Вы можете использовать черновик. Записи в черновике проверяться и оцениваться не будут. Советуем выполнять задания в том порядке, в котором они даны. Для экономии времени пропускайте задание, которое не удаётся выполнить сразу, и переходите к следующему. Если после выполнения всей работы у Вас останется время, Вы сможете вернуться к пропущенным заданиям. Баллы, полученные Вами за выполненные задания, суммируются. Постарайтесь выполнить как можно больше заданий и набрать наибольшее количество баллов.
Постарайтесь выполнить как можно больше заданий и набрать наибольшее количество баллов.
Всероссийские проверочные работы (ВПР) — Блог — Подгоренская СОШ №2
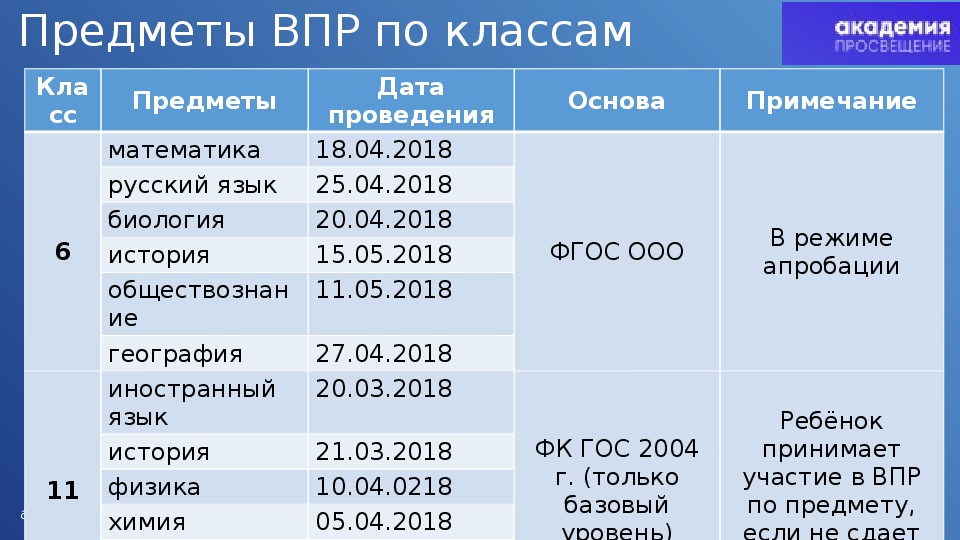
Всероссийские проверочные работы (ВПР) весной 2018 года будут проведены для учащихся 4 и 5 классов во всех школах России, в 6 и 11 классах – по решению школ.
ВПР – это итоговые контрольные работы, проводимые по отдельным учебным предметам для школьников всей страны. ВПР не являются аналогом государственной итоговой аттестации. Они проводятся на региональном или школьном уровне.
Первыми предстоит написать ВПР по шести учебным предметам выпускникам. В 11 классах ВПР проводятся по решению школы для учащихся, которые не выбрали данные предметы для сдачи в форме ЕГЭ. 20 марта для одиннадцатиклассников пройдет ВПР по иностранным языкам (английскому, немецкому и французскому), 21 марта – по истории, 3 апреля – по географии (ВПР по географии может проводиться как в 11, так и в 10 классах в зависимости от учебного плана школы), 5 апреля – по химии, 10 апреля – по физике, 12 апреля – по биологии.
ВПР для 4 классов откроет русский язык: первую часть работы учащиеся напишут 17 апреля, вторую – 19 апреля. ВПР по математике пройдет в 4 классах 24 апреля, по окружающему миру – 26 апреля. Пятиклассникам 17 апреля предстоит написать ВПР по русскому языку, 19 апреля – по математике, 24 апреля – по истории, 26 апреля – по биологии. Весной 2018 года ВПР в режиме апробации пройдут также для учащихся 6 классов. 18 апреля они напишут проверочную работу по математике, 20 апреля – по биологии, 25 апреля – по русскому языку, 27 апреля – по географии, 11 мая – по обществознанию, 15 мая – по истории.
ВПР являются самой массовой оценочной процедурой в системе образования: с момента их введения в 2015 году российские школьники написали около 17 миллионов проверочных работ.
ВПР учащиеся пишут в своих школах. Рекомендуемое время их проведения – второй-третий урок в школьном расписании; продолжительность – от одного до двух уроков. Работы выполняются по заданиям, разработанным на федеральном уровне, и проверяются по единым критериям. Проверка работ участников ВПР осуществляется в день проведения работы коллегиально учителями школы. После проверки результаты вносятся школами в единую информационную систему, с данными которой могут работать эксперты.
Проверка работ участников ВПР осуществляется в день проведения работы коллегиально учителями школы. После проверки результаты вносятся школами в единую информационную систему, с данными которой могут работать эксперты.
Результаты ВПР могут использоваться для формирования программ развития образования на уровне муниципалитетов, регионов и в целом по стране, для совершенствования методики преподавания предметов в конкретных школах, а также для индивидуальной работы с учащимися по устранению имеющихся пробелов в знаниях. По результатам ВПР не принимаются никакие обязательные решения, важные для определения дальнейшей судьбы и образовательной траектории школьника. Оценки за ВПР не влияют на получение аттестата и на перевод в следующий класс. С образцами и описаниями ВПР 2018 года для 11 классов можно ознакомиться на сайте ФИПИ, для остальных классов – на портале ВПР.
Химия | Олимпиадные задания
Задания, ответы, критерии оценивания Общие указания: если в задаче требуются расчёты, они обязательно должны быть приведены в решении. Ответ, приведённый без расчётов или иного обоснования, не засчитывается. Решения и система оценивания В итоговую оценку из 6 задач засчитываются 5 решений, за которые участник набрал наибольшие баллы, то есть одна из задач с наименьшим баллом не […]
Ответ, приведённый без расчётов или иного обоснования, не засчитывается. Решения и система оценивания В итоговую оценку из 6 задач засчитываются 5 решений, за которые участник набрал наибольшие баллы, то есть одна из задач с наименьшим баллом не […]
6629 07.02.2018
Задания, ответы, критерии оценивания Общие указания: если в задаче требуются расчёты, они обязательно должны быть приведены в решении. Ответ, приведённый без расчётов или иного обоснования, не засчитывается. Решения и система оценивания В итоговую оценку из 6 задач засчитываются 5 решений, за которые участник набрал наибольшие баллы, то есть одна из задач с наименьшим баллом не […]
5926 06.02.2018
Задания, ответы, критерии оценивания Общие указания: если в задаче требуются расчёты, они обязательно должны быть приведены в решении. Ответ, приведённый без расчётов или иного обоснования, не засчитывается. Решения и система оценивания В итоговую оценку из 6 задач засчитываются 5 решений, за которые частник набрал наибольшие баллы, то есть одна из задач с наименьшим баллом не […]
Решения и система оценивания В итоговую оценку из 6 задач засчитываются 5 решений, за которые частник набрал наибольшие баллы, то есть одна из задач с наименьшим баллом не […]
6293 06.02.2018
Задания, ответы, критерии оценивания Общие указания: если в задаче требуются расчёты, они обязательно должныбыть приведены в решении. Ответ, приведённый без расчётов или иногообоснования, не засчитывается. Решения и система оценивания. В итоговую оценку из 6 задач засчитываются 5 решений, за которые участник набрал наибольшие баллы, то есть одна из задач с наименьшим баллом не учитывается. Содержание Задание […]
5952 06.02.2018
Содержание Задача 1. Элемент-хамелеон Решение Задача 2. Свойства гомологов Решение Задача 3. Синтез ванадата Решение Задача 4. Гидратация углеводородов Решение Задача 5. Идентификация кислородсодержащего соединения Решение Задача 6. Получение и свойства неизвестной жидкости Решение Задача 1. Элемент-хамелеон Содержание ↑ На приведённой ниже схеме представлены превращения соединений одного химического элемента: Вещества Б, Д и Е нерастворимы в […]
Получение и свойства неизвестной жидкости Решение Задача 1. Элемент-хамелеон Содержание ↑ На приведённой ниже схеме представлены превращения соединений одного химического элемента: Вещества Б, Д и Е нерастворимы в […]
8407 07.11.2017
Содержание Задача 1. Левые части Решение Задача 2. От азотной кислоты к алюминату Решение Задача 3. Получение и окисление алкена Решение Задача 4. Запах газа Решение Задача 5. «Медный всадник» Решение Задача 6. Осторожное получение газа Решение В итоговую оценку из 6 задач засчитываются 5 решений, за которые участник набрал наибольшие баллы, то есть одна […]
12911 07.11.2017
Содержание Задача 1. Два газа Решение Задача 2. Соединения с водородом Решение Задача 3. Свойства неизвестного газа Решение Задача 4. Хорошо растворимый газ Решение Задача 5. Ионное соединение без металлов Решение Задача 6. Расширение и сжатие газа Решение В итоговую оценку из 6 задач засчитываются 5 решений, за которые участник набрал наибольшие баллы, то есть […]
Расширение и сжатие газа Решение В итоговую оценку из 6 задач засчитываются 5 решений, за которые участник набрал наибольшие баллы, то есть […]
8633 06.11.2017
Содержание Задача 1. Реакции соединения Решение Задача 2. Вещество из водорода и кислорода Решение Задача 3. Фториды в природе и в быту Решение Задача 4. Новое ракетное топливо Решение Задача 5. Реакция горения Решение Задача 6. Эксперименты с газами Решение В итоговую оценку из 6 задач засчитываются 5 решений, за которые участник набрал наибольшие […]
9778 06.11.2017
Задание 1. (10 баллов) Правые части По правой части с коэффициентами восстановите формулы веществ и коэффициенты в левой части уравнений реакций: … + … + … = 2Na2CrO4 + 3NaNO2 + 2CO2 … + … = Na2Cr2O7+ 2NaCl + h3O … + … + … = 2Na3[Cr(OH)6] + 3S + 4NaOH … + … = […]
6717
21. 01.2017
01.2017
Задание 1. (10 баллов) «Правые части» По правой части с коэффициентами восстановите формулы веществ и коэффициенты в левой части уравнений реакций: … + … = Sih5↑ + 2MgBr2 + 4Nh4↑ … + … = Ca3N2 + 3Cah3 … + … = 2KCl + 3Ca(OH)2 … + … + … = SiCl4 […]
13569 20.01.2017
Задание 1. (10 баллов) «Ионные реакции» Завершите приведённые ниже сокращённые ионные уравнения реакций с коэффициентами. Все неизвестные частицы обозначены многоточиями. … + 2… → Cu(OH)2↓ … + 2OH– + … → BaSO3↓ + … Pb2+ + … → … + 2H+ H+ + … → CO2↑ + … 3H+ + … → Al3+ + 3… […]
11508 19.01.2017
Задание 1 (10 баллов) «Реакции кислорода» Приведите примеры реакций с участием кислорода, в результате которых образуются: а) вода и кислотный оксид; б) вода и простое вещество; в) кислотный и основный оксиды; г) два кислотных оксида; д) основание. Напишите уравнения реакций. Решение а) Ch5 + 2O2 = CO2 + 2h3O; б) 4Nh4 + 3O2 = 2N2 […]
Напишите уравнения реакций. Решение а) Ch5 + 2O2 = CO2 + 2h3O; б) 4Nh4 + 3O2 = 2N2 […]
7835 19.01.2017
Задача 1 «Гаснущие свечки» В прибор, изображённый на рисунке, поместили небольшие кусочки кристаллического вещества Х белого цвета и налили жидкость Y. После того как открыли кран, жидкость Y опустилась из воронки в нижнюю часть прибора и пришла в соприкосновение с веществом X, началась реакция, сопровождающаяся выделением бесцветного газа Z. Газ Z по газоотводной трубке поступал […]
6981 18.01.2017
Задание 1 «Десять порошков» В десяти пронумерованных стаканах выданы порошки следующих веществ: медь, оксид меди(II), древесный уголь, красный фосфор, сера, железо, хлорид натрия, сахар, мел, малахит (оснóвный карбонат меди(II)). Ученики исследовали свойства выданных порошкообразных веществ, результаты своих наблюдений представили в таблице. Номер стакана Цвет исследуемого вещества «Поведение» порошка при помещении его в стакан с водой […]
Номер стакана Цвет исследуемого вещества «Поведение» порошка при помещении его в стакан с водой […]
12064 18.01.2017
Задача 1. «Полезная соль» Нерастворимая в воде соль X входит в состав множества полезных веществ – белых красок, огнеупорных материалов, жидкостей для бурения скважин, контрастных веществ для рентгенографии. Она состоит из трёх элементов, один из которых – сера. При прокаливании с избытком угля X превращается в растворимую соль Y, которая состоит всего из двух элементов […]
8666 17.01.2017
продуктов CBD Выдающаяся часть растущей конопляной промышленности Вермонта
vpr-news-cbd-vermont-20181010.mp3
Нажмите на кнопку воспроизведения вверху, чтобы прослушать вторую часть этой истории.
Поскольку сбор урожая подходит к концу, 2018 год оказывается ключевым для промышленной конопли в Вермонте. Его выращивает больше людей, чем когда-либо прежде, и миллионы долларов инвестируются в конопляные фермы и в промышленные лаборатории, необходимые для извлечения КБД из конопли.
CBD, что означает каннабидиол, является полностью легальным соединением, содержащимся в конопле, и не является психоактивным, а это означает, что он не поднимет вам настроение. Он появляется в розничных магазинах по всей стране.
Нигде бурно развивающаяся конопляная сцена Вермонта не проявляется так ярко, как в Хардвике, где базируется компания Green Mountain CBD. В этом году компания вырастила 40 акров конопли в Хардвике и Гайд-парке. Но в отличие от других производителей конопли в Вермонте, Green Mountain CBD также перерабатывает конопляное масло и производит капсулы и настойки CBD на ферме.
«Я думаю, что на самом деле мы просто работаем с тем, что существует уже много веков», — сказал генеральный директор Алехандро Бергад, имеющий опыт выращивания конопли в Колорадо.
Бергад и его партнер Джейкоб Гольдштейн сделали конопляную ферму Hardwick лидером среди производителей конопли в Вермонте. Они построили два производственных здания на ферме Хардвик и сушильный цех площадью 20 000 квадратных футов, который сушит растения конопли за 24 часа — коноплю нужно высушить перед переработкой.
Бергад использует сенсорный экран, чтобы запустить сборочную линию стоимостью 150 000 долларов, которая укупоривает и маркирует бутылки с капсулами CBD.«Эта линия прямо сейчас выплевывает около тысячи бутылок в час», — объяснил Бергад, когда мимо катились белые пластиковые бутылки.
Кредит Джон Калиш / Для VPR
/
Для VPR
Алехандро Бергад (слева) и Джейкоб Гольдштейн из центрального делового района Грин-Маунтин перед конопляным полем в Хардвике в июле. Green Mountain CBD в настоящее время производит 60 000 капсул в день, но планирует увеличить производство до миллиона капсул в день в следующем году. Компания, которая изменит свое название на Sun Soil, открыла офис в Берлингтоне и планирует удвоить свою площадь в 2019 году — возможно, за счет долины Шамплейн — благодаря инвестициям в размере 5 миллионов долларов от иностранной венчурной компании.
Компания, которая изменит свое название на Sun Soil, открыла офис в Берлингтоне и планирует удвоить свою площадь в 2019 году — возможно, за счет долины Шамплейн — благодаря инвестициям в размере 5 миллионов долларов от иностранной венчурной компании.
Расширение выращивания и переработки конопли происходит по всему штату.
«Наш регистр конопли взорвался», — сообщает Кэри Жигер, руководитель программы конопли Сельскохозяйственного агентства Вермонта. «В этом году у нас около 400 человек выращивают коноплю, и это почти 3000 акров.”
Жигер также занимается выращиванием конопли в округе Вашингтон, где он сотрудничает с защитником конопли из Нью-Мексико Дугом Файном.
Законодательное собрание Вермонта разрешило выращивание конопли в 2013 году, а в 2014 году было выращено несколько участков конопли. По всему штату площади, отведенные под коноплю в этом году, увеличились в четыре раза по сравнению с прошлым годом. Посевные площади, отведенные под коноплю в США, в этом году увеличились более чем вдвое, а в паре западных штатов, как сообщается, утроились.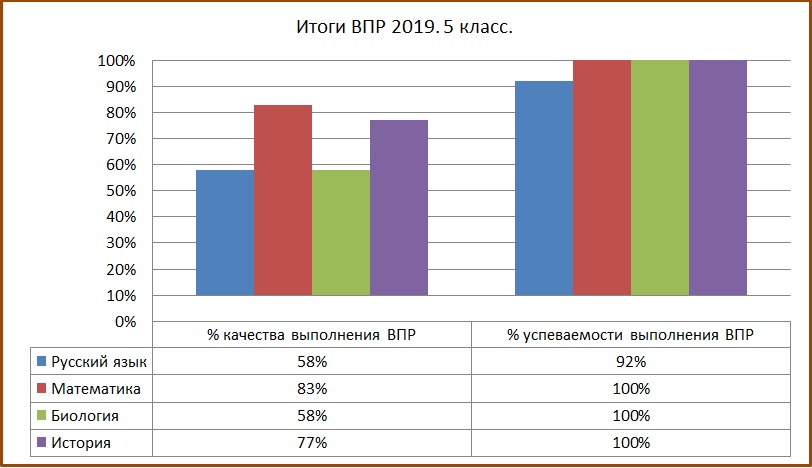
Кредит Джон Калиш / Для VPR
/
Для VPR
Кэри Жигер — директор программы конопли Агентства сельского хозяйства Вермонта, а также сам выращивает коноплю.В Вермонте фермеры, выращивающие коноплю, платят до 500 долларов за фунт, чтобы получить КБД, извлеченный из высушенного цветка конопли с использованием метода, в котором используется углекислый газ.
До этого года экстракторы обслуживали как медицинскую марихуану, так и коноплю в Уотербери и Милтоне. Но ожидается, что новые экстракторы, открывающиеся в Южном Берлингтоне и Браттлборо как раз к осеннему сбору конопли, снизят стоимость добычи КБД.
Один из новых экстракторов находится в бывшей пекарне, которая производила печенье Twinkies в Брэттлборо.Новое предприятие Northeast Processing затмевает мощности других экстракционных лабораторий Вермонта. Генеральный директор компании Карл Кристиансен имеет докторскую степень. по химии и контракты с 50 конопляными фермами Вермонта.
по химии и контракты с 50 конопляными фермами Вермонта.
«Мы рассматриваем коноплю как нечто, что может быть очень выгодно для штата, потому что государству нужны новые доходы, — сказал Кристиансен, — [Вермонту] нужна новая промышленность, но он не хочет отказываться от того, чем она является. И мы рассматриваем коноплю как действительно важный рынок для штата, потому что мы все еще можем оставаться верными тому, что представляет собой Вермонт.”
По словам Кристиансена, часть из 2 миллионов долларов, вложенных в его стартап, поступила от производителя конопли. Он ожидает, что среди клиентов Northeast Processing будут фермеры и производители продуктов CBD, которые не обязательно выращивают коноплю.
Кредит Джон Калиш / Для VPR
/
Для VPR
Карл Кристиансен на бывшей фабрике Twinkie в Брэттлборо, которая была преобразована в штаб-квартиру компании Northeast Processing.
«Есть много фермеров, которые просто хотят сосредоточиться на том, чтобы быть фермерами, и поэтому мы покупаем этот продукт и превращаем его в масла, а затем мы идем и затем продаем его производителям продукции, которые могут не захотеть заниматься сельским хозяйством, — сказал Кристиансен. «И мы считаем, что это также важная часть рынка».
Технология обработки конопли для извлечения CBD чрезвычайно высокотехнологична, но превращение конопляных семян в пищу — совсем не то.
На заводе Victory Hemp Foods площадью 7000 квадратных футов в Миддлбери семена конопли очищают от шелухи перед прессованием.Прессованные семена, известные как жмых, измельчают и просеивают, чтобы получить конопляную муку, которая на 50 процентов состоит из белка.
Оборудование на этом заводе в Миддлбери принадлежало Full Sun, местной компании по производству растительных масел, которая обанкротилась. Его приобрела компания Victory Hemp Foods из Кентукки, главным операционным директором которой является Крис Бейли, основатель Vermont Smoke & Cure в Хайнесбурге.
— У нас очень хороший спрос на конопляное масло, — сказал Бейли. «Все, от людей, которые используют его в качестве масла-носителя для CBD, до других производителей продуктов питания и людей, занимающихся здоровьем и красотой, используют его для мыла и ухода за кожей.
Победа Конопля нашла только одного фермера в Вермонте, желающего выращивать коноплю как зерновую культуру. Все остальные выращивают коноплю для КБР. 24-летний фермер по имени Сэм Беллаванс выращивал 10 акров на молочной ферме с тысячей коров в Олбурге. Victory Hemp покупает его урожай и планирует закупать семена на фермах в штате Мэн и северной части штата Нью-Йорк.
На данный момент, однако, почти все фермеры, выращивающие коноплю в Вермонте, надеются нажиться на «золотой лихорадке КБР». Исследовательская фирма New Frontier Data, специализирующаяся на индустрии каннабиса, оценивает, что в прошлом году американцы потратили 367 миллионов долларов на продукты CBD, и ожидает, что к 2020 году объем продаж превысит миллиард долларов.
Исследовательская компания New Frontier Data, специализирующаяся на производстве каннабиса, подсчитала, что в прошлом году американцы потратили 367 миллионов долларов на продукты CBD, и ожидает, что к 2020 году объем продаж превысит миллиард долларов.
На исследовательской ферме UVM в Олбурге в 2018 году было посажено 2 акра конопли. Не все из них предназначались для исследований CBD.
«У нас есть два разных сорта волокна и три разных сорта зерна», — объяснил Абха Гупта, координатор по сельскохозяйственным культурам и почве в UVM Extension . «Они были высажены в три разных срока посадки».
Агроном Хизер Дарби считает, что широкая доступность конопляного семени, наряду с официальной государственной программой исследований и разработок конопли, созданной законодателями штата, является причиной растущего интереса к этой культуре. Дарби сказала, что многие жители Вермонта звонили ей за советом.
— Большинство людей ищут рынки сбыта, — сказала Дарби. «Некоторые из вопросов: «Это реальная возможность или это просто пузырь, который мы видим, проплывая мимо?» Вы знаете, «Где мы можем на самом деле подключиться к этому?» И рынки действительно все еще развиваются, особенно если мы не говорим о производстве КБД.
«Некоторые из вопросов: «Это реальная возможность или это просто пузырь, который мы видим, проплывая мимо?» Вы знаете, «Где мы можем на самом деле подключиться к этому?» И рынки действительно все еще развиваются, особенно если мы не говорим о производстве КБД.
Но КБД, конечно же, это то, о чем все говорят. Хотя FDA запрещает заявления о предполагаемой пользе КБД для здоровья, спрос на КБД как средство от боли, беспокойства, бессонницы и других неприятностей растет.
Кредит Джон Калиш / Для VPR
/
Для VPR
Экстрактор сверхкритического диоксида углерода, используемый для извлечения КБД из цветков конопли в Лабораториях нутрицевтических исследований в Уотербери.У него есть лабораторное оборудование на полмиллиона долларов, и он планирует потратить еще сотни тысяч на новое оборудование. Калев Фриман, врач и ученый из Лаборатории нутрицевтических исследований в Уотербери, планирует провести исследование воздействия КБД на животных.
«КБР не соответствует самым строгим уровням доказательности, которые были бы рандомизированными контрольными испытаниями в нескольких центрах. КБД еще не соответствует этим критериям», — отметил Фриман. данные, которые настолько убедительно подтверждают преимущества КБД, что очень трудно сказать, что для этого нет никаких доказательств.
Неудивительно, что в Вермонте возникла кустарная промышленность, производящая широкий спектр продуктов CBD: капсулы, настойки, лакомства для собак, кремы для тела, напитки и конфеты.
В Nutty Steph’s в Миддлсексе вы можете купить CBD. -настоянный шоколад в различных дозировках и формах.
В июле Жаклин Фернандес-Рики, владеющая магазином, сказала, что продукты CBD составляли 30 процентов продаж Nutty Steph после того, как они были представлены шестью месяцами ранее.
На вопрос о ней 3- Фернандес-Рики сказал: «Шоколадный батончик CBD на унцию, который продается за 20 долларов: «Это гораздо больше, чем плитка шоколада.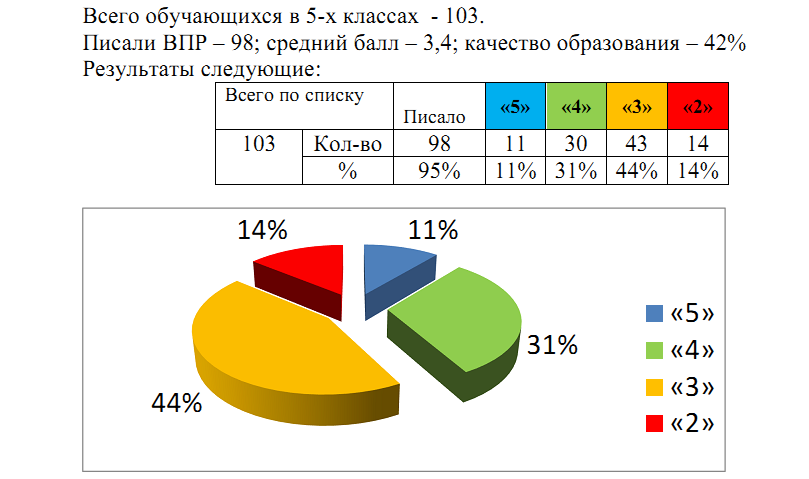
Хотя шоколад содержит кофеин, люди, страдающие бессонницей, обнаружили, что шоколад с КБД помогает им заснуть, сказал Фернандес-Рики.
CBD также присутствует в меде, кофе и чае.
Кредит Джон Калиш / Для VPR
/
Для VPR
Владелец Чокнутого Стефа Жаклин Фернандес-Рики держит тарелку с шоколадными сердечками CBD.Джо и Ребекка Пименталь выращивают коноплю на ферме Люс в Стокбридже.Они платят за извлечение КБД из конопли.
Pimentals открыли коммерческую кухню в Вефиле, чтобы производить бальзам, мед и настойку CBD. Они сотрудничали с пивоварней Long Trail Brewery над пивом CBD, получившим название «Medicator», но федеральные власти запретили его продавать.
Их CBD теперь используется в холодном напитке, производимом Abracadabra Coffee в Вудстоке, и является ингредиентом чая и смеси чая/сока, производимой Dobra Tea и Tomgirl в Берлингтоне.
«Обе эти компании используют наш мед, насыщенный КБД», — сказала Ребекка Пименталь. «Я думаю, вы увидите это повсюду».
«Я думаю, вы увидите это повсюду».
Владелец магазина Green State Gardener в Берлингтоне Дилан Раап говорит, что в этом году его компания продаст продукции CBD на миллион долларов. У него есть собственная линейка пищевых продуктов CBD, в которую входит бутылка жевательных конфет CBD за 50 долларов.
«Мы видим бумеров, приходящих из-за болей в суставах», — сказал Раап во время интервью в магазине на Пайн-стрит. «Но мы также видим много миллениалов, да и вообще всех, которые беспокоятся и просто стабилизируют настроение эффектом, который оказывает КБД.”
Raap скоро выпустит газированную воду и содовую, наполненные CBD.
«Я думаю, что для класса продуктов CBD нет предела возможностям», — сказал он.
В Берлингтоне есть еще один магазин, специализирующийся на продуктах CBD, которым управляет диспансер Champlain Valley. Другие магазины в Линдонвилле, Сент-Олбансе, Браттлборо и Миддлбери посвящены исключительно продуктам из конопли или КБД.
Шейн Линн, исполнительный директор диспансера Champlain Valley, у которого также есть магазин в Брэттлборо, где продаются продукты CBD, жалуется на низкую прибыль. Он подозревает, что цены на CBD будут падать по мере роста предложения конопли.
Он подозревает, что цены на CBD будут падать по мере роста предложения конопли.
«Вы должны думать, эй, Кентукки, тогда вы должны думать о Колорадо», — сказала Линн. «Это действительно мировой рынок. Итак, как фермеры Вермонта собираются конкурировать за пределами Вермонта?»
Еще один знаток каннабиса, который думает, что цены на коноплю упадут, — Джоэл Бедард, основатель The Vermont Hemp Company.
«Все так взволнованы каннабиноидами, и они хотят сначала попытаться заработать хорошие деньги, а затем, когда все стабилизируется, они поймут: «Хорошо, это на самом деле продовольственная культура».«Это гораздо больше, чем просто каннабиноиды», — Джоэл Бедард, The Vermont Hemp Company.
«Все так взволнованы каннабиноидами, и они хотят сначала попытаться заработать хорошие деньги, а затем, когда все стабилизируется, они поймут: «Хорошо, это на самом деле продовольственная культура», — сказал Бедард. «Это гораздо больше, чем просто каннабиноиды».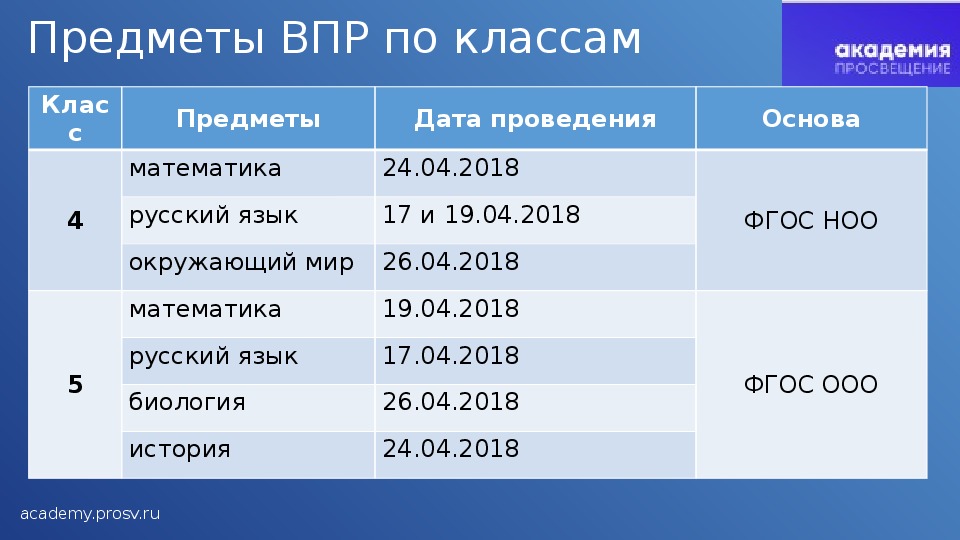
CBD, конечно, не единственный каннабиноид. Другие каннабиноиды с известными медицинскими преимуществами включают CBG и CBC (не путать с Канадской радиовещательной корпорацией).
На втором ежегодном собрании Hemp Fest, организованном Heady Vermont в прошлом месяце, с программной речью выступил Сет Кроуфорд, селекционер конопли из Орегона, который предсказывает, что CBG готово взлететь.
В интервью после своего выступления Кроуфорд отметил, что кристаллическая форма КБД продается по цене до 5000 долларов за фунт, а кристаллическая форма КБГ стоит втрое больше. Кроуфорд сказал, что кристаллический CBC продается примерно по 45 000 долларов за фунт.
«По мере появления новых каннабиноидов они становятся дорогими, — объяснил Кроуфорд.«Когда эти новые соединения появятся в сети, фермеры смогут заработать значительные суммы денег в этой отрасли».
Семена конопли Кроуфорд, выращенные в Орегоне, составляют около 50 акров урожая конопли в Вермонте в этом году.
Набор инструментов CRISPR–dCas для генной инженерии и синтетической биологии
https://doi.org/10.1016/j.jmb.2018.06.037Получить права и содержимое Платформа для регуляции генов и контроля эпигенома.. преимущества, проблемы и перспективы системы dCas как технологической платформы
Abstract
Программируемый контроль экспрессии генов необходим для понимания функций генов, конструирования клеточного поведения и разработки терапевтических средств.Помимо приложений для редактирования генов, обеспечиваемых нуклеазами CRISPR-Cas9 и CRISPR-Cas12a, изобретение молекул Cas без нуклеаз (dCas9 и dCas12a) предлагает платформу для точного контроля функции генома без редактирования генов. Были разработаны различные инструменты dCas, которые представляют собой всеобъемлющий набор инструментов, позволяющий исследовать функцию генов и модулировать клеточное поведение. В этом обзоре обобщаются текущие применения инструментов dCas для регуляции транскрипции, эпигенетической инженерии, визуализации генома, генетического скрининга и иммунопреципитации хроматина. Мы также подчеркиваем преимущества и существующие проблемы современных инструментов dCas в генной инженерии и синтетической биологии и представляем перспективы будущих направлений и приложений.
Мы также подчеркиваем преимущества и существующие проблемы современных инструментов dCas в генной инженерии и синтетической биологии и представляем перспективы будущих направлений и приложений.
Ключевые слова
DCAS9
CRISPRI / A
CRISPRI / A
Gene / A
Gene / A
Epigenome Engineering
Генетическое скрининг
Сокращения
Сокращения TALES
транскрипционные активаторы, подобные активаторы
CrisprКластерны для регулярных мерсрочных короткими палиндромическими повторениями
CRISPRICrispr Interference
SCFVодноцепочечный переменный фрагмент
SAMСинергетическая активация Mediator
HSF1Тепловой фактор человека 1
CIDSХимикогенецированные димеризующие домены
OIDSОптогенетически индуцирующие димеризующие домены
GPCRSг белковых рецепторов
РыбаФлуоресценция в гибридизации SITU
Рекомендуемые статьиСсылки на статьи (0)
© 2018 The Authors. Опубликовано Elsevier Ltd.
Опубликовано Elsevier Ltd.
Рекомендуемые статьи
Ссылки на статьи
Произошла ошибка при настройке файла cookie пользователя
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка браузера на прием файлов cookie
Существует множество причин, по которым файл cookie не может быть установлен правильно. Ниже приведены наиболее распространенные причины:
- В вашем браузере отключены файлы cookie.Вам необходимо сбросить настройки браузера, чтобы принять файлы cookie, или спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файл cookie.
- Ваш браузер не поддерживает файлы cookie.
 Попробуйте другой браузер, если вы подозреваете это.
Попробуйте другой браузер, если вы подозреваете это. - Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie.Чтобы это исправить, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Предоставить доступ без файлов cookie потребует от сайта создания нового сеанса для каждой посещаемой вами страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в файле cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только та информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, если вы не решите ввести его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступ к остальной части вашего компьютера, и только сайт, создавший файл cookie, может его прочитать.
Структурное понимание ремоделирования убиквитинлигазы Cullin4-RING с помощью Vpr из вирусов иммунодефицита обезьян
Abstract
В ходе эволюции вирусы выработали средства манипулирования убиквитин-протеасомной системой хозяина, чтобы подавлять противовирусные факторы хозяина. Семейство лентивирусных вспомогательных белков Vpx/Vpr узурпирует субстратный рецептор DCAF1 лигаз Cullin4-RING (CRL4) хозяина, семейства модульных убиквитинлигаз, участвующих в репликации ДНК, репарации ДНК и регуляции клеточного цикла. Модуляция специфичности CRL4 DCAF1 с помощью Vpx и Vpr некоторых вирусов иммунодефицита обезьян (SIV) приводит к рекрутированию, полиубиквитилированию и последующей протеасомной деградации фактора рестрикции хозяина SAMHD1, что приводит к усилению репликации вируса в дифференцированных клетках. Чтобы раскрыть механизм убиквитилирования SAMHD1, вызванного Vpr SIV, мы провели интегративный биохимический и структурный анализ белка Vpr из SIV, заражающих Cercopithecus cephus (SIV mus ).Рентгеновская кристаллография выявляет сходство между SIV mus Vpr и другими членами семейства Vpx/Vpr в отношении взаимодействия DCAF1, в то время как криоэлектронная микроскопия и масс-спектрометрия сшивки подчеркивают расходящийся молекулярный механизм рекрутирования SAMHD1. Кроме того, эти исследования демонстрируют, как SIV mus Vpr использует динамическую архитектуру мультисубъединичной сборки CRL4 DCAF1 для оптимизации убиквитилирования SAMHD1.
Модуляция специфичности CRL4 DCAF1 с помощью Vpx и Vpr некоторых вирусов иммунодефицита обезьян (SIV) приводит к рекрутированию, полиубиквитилированию и последующей протеасомной деградации фактора рестрикции хозяина SAMHD1, что приводит к усилению репликации вируса в дифференцированных клетках. Чтобы раскрыть механизм убиквитилирования SAMHD1, вызванного Vpr SIV, мы провели интегративный биохимический и структурный анализ белка Vpr из SIV, заражающих Cercopithecus cephus (SIV mus ).Рентгеновская кристаллография выявляет сходство между SIV mus Vpr и другими членами семейства Vpx/Vpr в отношении взаимодействия DCAF1, в то время как криоэлектронная микроскопия и масс-спектрометрия сшивки подчеркивают расходящийся молекулярный механизм рекрутирования SAMHD1. Кроме того, эти исследования демонстрируют, как SIV mus Vpr использует динамическую архитектуру мультисубъединичной сборки CRL4 DCAF1 для оптимизации убиквитилирования SAMHD1. В совокупности настоящая работа обеспечивает детальное молекулярное понимание изменчивости и видовой специфичности эволюционной гонки вооружений между рестрикцией SAMHD1 хозяина и противодействием лентивирусам посредством белков Vpx/Vpr.
В совокупности настоящая работа обеспечивает детальное молекулярное понимание изменчивости и видовой специфичности эволюционной гонки вооружений между рестрикцией SAMHD1 хозяина и противодействием лентивирусам посредством белков Vpx/Vpr.
Резюме автора Из-за ограниченного размера вирусных геномов репликация вируса в значительной степени зависит от компонентов клетки-хозяина. В дополнение к энергетическому метаболизму клетки-хозяина и ее аппарату репликации ДНК и синтеза белка механизм деградации белка является привлекательной мишенью для повторного присвоения вирусом. Некоторые вирусные факторы переключают специфичность убиквитинлигазы хозяина на противовирусные факторы хозяина, чтобы пометить их для разрушения протеасомой, чтобы снять внутриклеточные барьеры для репликации вируса.Здесь мы представляем молекулярные детали того, как вспомогательный белок вируса иммунодефицита обезьян Vpr взаимодействует с субстратным рецептором убиквитинлигазы хозяина Cullin4-RING и как это взаимодействие перенаправляет специфичность Cullin4-RING на противовирусный фактор хозяина SAMHD1. Исследования раскрывают механизм индуцированного Vpr рекрутирования SAMHD1 и последующего убиквитинирования. Более того, путем сравнения с родственными вспомогательными белками других видов вирусов иммунодефицита мы иллюстрируем удивительную изменчивость молекулярных стратегий противодействия SAMHD1, которые эти вирусы приняли во время эволюционной адаптации к своим хозяевам.Наконец, наша работа также обеспечивает более глубокое понимание внутренней работы механизма убиквитилирования Cullin4-RING хозяина.
Исследования раскрывают механизм индуцированного Vpr рекрутирования SAMHD1 и последующего убиквитинирования. Более того, путем сравнения с родственными вспомогательными белками других видов вирусов иммунодефицита мы иллюстрируем удивительную изменчивость молекулярных стратегий противодействия SAMHD1, которые эти вирусы приняли во время эволюционной адаптации к своим хозяевам.Наконец, наша работа также обеспечивает более глубокое понимание внутренней работы механизма убиквитилирования Cullin4-RING хозяина.
Введение
Большая часть вирусов разработала средства для кооптации механизма убиквитинирования своего хозяина с целью улучшения условий репликации либо путем введения вирусных убиквитинлигаз и деубиквитиназ, либо путем модификации белков хозяина, участвующих в убиквитилировании [1– 3]. В частности, убиквитинлигазы хозяина являются заметной мишенью для вирусной узурпации, чтобы перенаправить специфичность на противовирусные факторы рестрикции хозяина.Это приводит к рекрутированию факторов рестрикции в виде неэндогенных neo -субстратов, вызывая их полиубиквитилирование и последующую протеасомную деградацию [4-8]. Это противодействие противовирусному репертуару хозяина имеет важное значение для инфекционности и распространения вируса [9–12], и механистическое понимание этих изменений специфичности расширяет наше понимание вирусного патогенеза и может проложить путь к новым методам лечения.
Это противодействие противовирусному репертуару хозяина имеет важное значение для инфекционности и распространения вируса [9–12], и механистическое понимание этих изменений специфичности расширяет наше понимание вирусного патогенеза и может проложить путь к новым методам лечения.
Часто модифицирующие белки, кодируемые вирусом, связываются с убиквитинлигазами Cullin4-RING (CRL4) и адаптируются к ним [5].CRL4 состоит из каркаса Cullin4 (CUL4), который соединяет каталитическую субъединицу RING-домена ROC1 с адапторным белком DDB1, который, в свою очередь, связывается с рецепторами обменного субстрата (DCAF, DDB1- и CUL4-ассоциированные факторы) [13-17]. В некоторых случаях адаптер DDB1 служит якорем для вирусных белков, которые затем действуют как «вирусные DCAF» для рекрутирования антивирусного субстрата. Примерами являются обезьяний белок 5V вируса и мышиный цитомегаловирус M27, которые связываются с DDB1 и привлекают белки STAT1/2 для убиквитинирования, чтобы вмешиваться в интерфероновый ответ хозяина [18-20]. Точно так же CUL4-зависимое подавление передачи сигналов STAT важно для репликации вируса Западного Нила [21]. Кроме того, белок Х вируса гепатита В захватывает DDB1, вызывая протеасомную деструкцию комплекса структурного поддержания хромосомы (SMC), что способствует репликации вируса [22, 23].
Точно так же CUL4-зависимое подавление передачи сигналов STAT важно для репликации вируса Западного Нила [21]. Кроме того, белок Х вируса гепатита В захватывает DDB1, вызывая протеасомную деструкцию комплекса структурного поддержания хромосомы (SMC), что способствует репликации вируса [22, 23].
Вирусные факторы также связываются с рецепторами DCAF и модифицируют их, перенаправляя их на антивирусные субстраты. Яркими примерами являются вспомогательные белки лентивирусов Vpr и Vpx. Все современные вирусы иммунодефицита человека и обезьян (ВИЧ/ВИО) кодируют Vpr, в то время как только две линии, представленные ВИЧ-2 и ВИО-инфицирующими мандрилами, несут Vpx [24].Белки Vpr и Vpx упаковываются в дочерние вирионы и высвобождаются в клетку-хозяина при инфицировании, где они связываются с DCAF1 в ядре [25]. В данной работе соответствующие белки вируса иммунодефицита обезьян Vpx/Vpr будут указываться с видом их хозяина в качестве нижнего индекса с использованием следующих сокращений: mus – усатая обезьяна ( Cercopithecus cephus ), mnd – мандрил ( Mandrillus sphinx ), rcm – мангабей красношапочный ( Cercocebus torquatus ), см – мангабей сажистый ( Cercocebus atys ), деб – обезьяна де Бразза ( Cercopithecus ignoretus ), сык – обезьяна Сайка ( Cercopithecus ignores ), african – albogularis обезьяна ( Chlorocebus спец. ).Vpr HIV-1 важен для репликации вируса in vivo и в моделях инфекции макрофагов [26]. Недавний протеомный анализ показал, что модуляция специфичности DCAF1 белками Vpr HIV-1 приводит к подавлению сотен белков-хозяев DCAF1- и протеасом-зависимым образом [27], включая ранее описанную деградацию Vpr HIV-1 . нацелены на UNG2 [28], HLTF [29], MUS81 [30, 31], MCM10 [32] и TET2 [33]. Эта удивительная неразборчивость в мишенях деградации также частично сохраняется в более отдаленных кладах, примерами которых являются Vpr agm и Vpr mus [27].Однако плейотропность Vpr и отсутствие легкодоступных экспериментальных моделей не позволили определить, как именно эти события деградации способствуют репликации [26].
).Vpr HIV-1 важен для репликации вируса in vivo и в моделях инфекции макрофагов [26]. Недавний протеомный анализ показал, что модуляция специфичности DCAF1 белками Vpr HIV-1 приводит к подавлению сотен белков-хозяев DCAF1- и протеасом-зависимым образом [27], включая ранее описанную деградацию Vpr HIV-1 . нацелены на UNG2 [28], HLTF [29], MUS81 [30, 31], MCM10 [32] и TET2 [33]. Эта удивительная неразборчивость в мишенях деградации также частично сохраняется в более отдаленных кладах, примерами которых являются Vpr agm и Vpr mus [27].Однако плейотропность Vpr и отсутствие легкодоступных экспериментальных моделей не позволили определить, как именно эти события деградации способствуют репликации [26].
В отличие от этого, Vpx имеет гораздо более узкий диапазон субстратов. Недавно сообщалось, что он нацеливает стимулятор генов интерферона (STING) и компоненты комплекса концентратора молчания человека (HUSH) на деградацию, что приводит к ингибированию противовирусной cGAS-STING-опосредованной передачи сигналов и реактивации латентных провирусов соответственно [34–36]. ].Важно отметить, что Vpx также рекрутирует фактор рестрикции SAMHD1 в DCAF1, чтобы пометить его для протеасомной деструкции [37, 38]. SAMHD1 представляет собой дезоксинуклеотидтрифосфат (дНТФ) трифосфогидролазу, которая ограничивает репликацию ретровирусов в неделящихся клетках путем снижения пула дНТФ до уровней, которые не могут поддерживать вирусную обратную транскрипцию [39–46]. Ретровирусы, которые экспрессируют Vpx, способны ослаблять рестрикцию SAMHD1, обеспечивая репликацию в дифференцированных клетках миелоидного происхождения, покоящихся Т-клетках и Т-клетках памяти [38, 47, 48].В результате постоянной эволюционной гонки вооружений между рестрикцией SAMHD1 хозяина и его вирусным антагонистом Vpx механизм рекрутирования SAMHD1, опосредованного Vpx, в высокой степени зависит от вида и штамма вируса: клада Vpx, представленная Vpx ВИЧ-2 , распознает С-концевой домен (CtD) SAMHD1, тогда как Vpx mnd2/rcm связывает N-концевой домен (NtD) SAMHD1 принципиально другим образом [24, 49–52].
].Важно отметить, что Vpx также рекрутирует фактор рестрикции SAMHD1 в DCAF1, чтобы пометить его для протеасомной деструкции [37, 38]. SAMHD1 представляет собой дезоксинуклеотидтрифосфат (дНТФ) трифосфогидролазу, которая ограничивает репликацию ретровирусов в неделящихся клетках путем снижения пула дНТФ до уровней, которые не могут поддерживать вирусную обратную транскрипцию [39–46]. Ретровирусы, которые экспрессируют Vpx, способны ослаблять рестрикцию SAMHD1, обеспечивая репликацию в дифференцированных клетках миелоидного происхождения, покоящихся Т-клетках и Т-клетках памяти [38, 47, 48].В результате постоянной эволюционной гонки вооружений между рестрикцией SAMHD1 хозяина и его вирусным антагонистом Vpx механизм рекрутирования SAMHD1, опосредованного Vpx, в высокой степени зависит от вида и штамма вируса: клада Vpx, представленная Vpx ВИЧ-2 , распознает С-концевой домен (CtD) SAMHD1, тогда как Vpx mnd2/rcm связывает N-концевой домен (NtD) SAMHD1 принципиально другим образом [24, 49–52].
В ходе эволюционной адаптации к своим хозяевам-приматам и из-за селективного давления, направленного на уклонение от ограничения SAMHD1, две группы SIV, не имеющие Vpx, SIV agm и SIV deb/mus/syk , neo -функционализировал свой Vpr для связывания SAMHD1 и индукции его деградации [24, 49, 53].Следовательно, эти виды развили «гибридные» белки Vpr, которые сохраняют нацеливание на некоторые факторы хозяина, истощенные Vpr типа HIV-1 [27], и дополнительно индуцируют деградацию SAMHD1.
Чтобы раскрыть молекулярные механизмы DCAF1- и SAMHD1-взаимодействия такого «гибридного» Vpr, мы инициировали интегративный биохимический и структурный анализ белка Vpr из ВИО, заражающего Cercopithecus cephus , Vpr mus . Эти исследования выявляют сходство и различия с белками Vpx и Vpr из других видов лентивирусов и указывают на расходящийся молекулярный механизм Vpr mus -зависимого рекрутирования SAMHD1 в CUL4/ROC1/DDB1/DCAF1 (CRL4 DCAF1 ). Кроме того, криоэлектронная микроскопия (крио-ЭМ) реконструкция белкового комплекса CRL4 DCAF1 , модифицированного Vpr mus , позволяет получить представление о структурной пластичности всей сборки убиквитинлигазы CRL4 с последствиями для механизма переноса убиквитина.
Кроме того, криоэлектронная микроскопия (крио-ЭМ) реконструкция белкового комплекса CRL4 DCAF1 , модифицированного Vpr mus , позволяет получить представление о структурной пластичности всей сборки убиквитинлигазы CRL4 с последствиями для механизма переноса убиквитина.
Результаты
SAMHD1-CtD необходим и достаточен для связывания Vprmus и убиквитилирования in vitro -концевой домен (DCAF1-CtD), белковые комплексы восстанавливали
in vitro из очищенных компонентов и анализировали с помощью гель-фильтрационной (GF) хроматографии.Различные использованные белковые конструкции схематически показаны на рис. S1A. В отсутствие дополнительных партнеров по связыванию Vpr mus становится нерастворимым после удаления тега аффинной очистки GST (рис. S1B) и, соответственно, не может быть применен к колонке GF. . Взаимодействие SAMHD1 с DDB1/DCAF1-CtD не было обнаружено в отсутствие Vpr mus (рис. S1C). Анализ комбинаций бинарных белков (Vpr mus и DDB1/DCAF1-CtD; Vpr mus и SAMHD1) показывает, что Vpr mus элюируется вместе с DDB1/DCAF1-CtD (рис. S1D) или с SAMHD1 (рис. S1E).Инкубация Vpr mus с DDB1/DCAF1B и SAMHD1 с последующим GF привела к совместной элюции всех трех компонентов (рис. 1А). Вместе эти результаты показывают, что Vpr mus образует стабильные бинарные и тройные белковые комплексы с DDB1/DCAF1-CtD и/или SAMHD1 in vitro . Кроме того, инкубация с любым из этих партнеров по взаимодействию, по-видимому, стабилизирует Vpr mus , уменьшая его склонность к агрегации/нерастворимости. Рис. 1. Биохимический анализ Vpr mus , индуцированного перенаправлением специфичности CRL4 DCAF1 .
S1D) или с SAMHD1 (рис. S1E).Инкубация Vpr mus с DDB1/DCAF1B и SAMHD1 с последующим GF привела к совместной элюции всех трех компонентов (рис. 1А). Вместе эти результаты показывают, что Vpr mus образует стабильные бинарные и тройные белковые комплексы с DDB1/DCAF1-CtD и/или SAMHD1 in vitro . Кроме того, инкубация с любым из этих партнеров по взаимодействию, по-видимому, стабилизирует Vpr mus , уменьшая его склонность к агрегации/нерастворимости. Рис. 1. Биохимический анализ Vpr mus , индуцированного перенаправлением специфичности CRL4 DCAF1 . ( AC ) Анализ GF in vitro восстановление белковых комплексов, содержащих DDB1/DCAF1-CtD, Vpr mus и SAMHD1 ( A ), SAMHD1-ΔCtD ( B-D1C ) или T4L-SAMHD ( С ). Объемы элюирования стандартов молекулярной массы белка указаны над хроматограммой в A . Окрашенные кумасси синим анализы SDS-PAGE фракций, собранных во время анализов GF, показаны под хроматограммами с цветными прямоугольниками в соответствии с хроматограммами. SAM – стерильный α-мотивный домен, HD – гистидин-аспартатный домен, T4L – Т4 лизоцим. Звездочка и двойная звездочка указывают на незначительное загрязнение оставшейся протеазой GST-3C и тегом очистки GST соответственно. ( D-G ) Реакции убиквитилирования in vitro с очищенными белковыми компонентами в отсутствие ( D ) или в присутствии ( E-G ) Vpr mus с указанными конструкциями SAMHD1 в качестве субстрата. Реакции останавливали через указанное время, разделяли на SDS-PAGE и визуализировали окрашиванием кумасси синим.
SAM – стерильный α-мотивный домен, HD – гистидин-аспартатный домен, T4L – Т4 лизоцим. Звездочка и двойная звездочка указывают на незначительное загрязнение оставшейся протеазой GST-3C и тегом очистки GST соответственно. ( D-G ) Реакции убиквитилирования in vitro с очищенными белковыми компонентами в отсутствие ( D ) или в присутствии ( E-G ) Vpr mus с указанными конструкциями SAMHD1 в качестве субстрата. Реакции останавливали через указанное время, разделяли на SDS-PAGE и визуализировали окрашиванием кумасси синим.
Предыдущие клеточные анализы показали, что остатки 583-626 SAMHD1 макаки-резус (SAMHD1-CtD) необходимы для Vpr mus , индуцированной протеасомной деградации [49]. Чтобы проверить это открытие в нашей системе in vitro , конструкции, содержащие SAMHD1-CtD, слитые с лизоцимом T4 (T4L-SAMHD1-CtD), или содержащие только N-концевые домены SAMHD1 и не содержащие SAMHD1-CtD (SAMHD1-ΔCtD), , инкубировали с Vpr mus и DDB1/DCAF1-CtD, и комплексообразование оценивали с помощью GF-хроматографии. Анализ полученных хроматограмм с помощью SDS-PAGE показывает, что SAMHD1-ΔCtD не элюировался совместно с DDB1/DCAF1-CtD/Vpr mus (рис. 1B). Напротив, T4L-SAMHD1-CtD накапливался в том же пике элюирования, что и DDB1/DCAF1-CtD и Vpr mus (рис. 1C). Эти результаты подтверждают, что SAMHD1-CtD необходим для стабильной ассоциации с DDB1/DCAF1-CtD/Vpr mus in vitro , и демонстрируют, что SAMHD1-CtD достаточно для опосредованного Vpr mus рекрутирования T4L-SAMHD1- Слитая конструкция CtD с DDB1/DCAF1-CtD.
Анализ полученных хроматограмм с помощью SDS-PAGE показывает, что SAMHD1-ΔCtD не элюировался совместно с DDB1/DCAF1-CtD/Vpr mus (рис. 1B). Напротив, T4L-SAMHD1-CtD накапливался в том же пике элюирования, что и DDB1/DCAF1-CtD и Vpr mus (рис. 1C). Эти результаты подтверждают, что SAMHD1-CtD необходим для стабильной ассоциации с DDB1/DCAF1-CtD/Vpr mus in vitro , и демонстрируют, что SAMHD1-CtD достаточно для опосредованного Vpr mus рекрутирования T4L-SAMHD1- Слитая конструкция CtD с DDB1/DCAF1-CtD.
Чтобы соотнести эти данные с ферментативной активностью, было проведено анализов убиквитилирования in vitro путем инкубации SAMHD1, SAMHD1-ΔCtD или T4L-SAMHD1-CtD с очищенным CRL4 DCAF1-CtD , E1 (UBA1), E2 (UBCH5C), убиквитин и АТФ. Входные белки показаны на рис. S2A, а контрольные реакции на рис. S2B, C. В отсутствие Vpr mus убиквитилирование SAMHD1 не наблюдалось (рис. 1D и S2D), в то время как добавление Vpr mus приводило к надежному убиквитилированию SAMHD1. , о чем свидетельствует сдвиг SAMHD1 вверх в анализе SDS PAGE, вызванный ковалентной модификацией с увеличением количества молекул убиквитина, что приводит к почти полной потере полосы, соответствующей немодифицированному SAMHD1, после 15-минутной инкубации (рис. 1E и S2E).В соответствии с аналитическими данными GF, SAMHD1-ΔCtD не убиквитилировался в присутствии Vpr mus (рис. 1F и S2F). Напротив, T4L-SAMHD1-CtD был эффективно убиквитилирован, что привело к потере> 90% полосы, соответствующей немодифицированному T4L-SAMHD1-CtD, через 15 минут (рис. 1G и S2F). Опять же, эти данные подтверждают функциональную важность SAMHD1-CtD для опосредованного Vpr mus рекрутирования в убиквитинлигазу CRL4 DCAF1 .
, о чем свидетельствует сдвиг SAMHD1 вверх в анализе SDS PAGE, вызванный ковалентной модификацией с увеличением количества молекул убиквитина, что приводит к почти полной потере полосы, соответствующей немодифицированному SAMHD1, после 15-минутной инкубации (рис. 1E и S2E).В соответствии с аналитическими данными GF, SAMHD1-ΔCtD не убиквитилировался в присутствии Vpr mus (рис. 1F и S2F). Напротив, T4L-SAMHD1-CtD был эффективно убиквитилирован, что привело к потере> 90% полосы, соответствующей немодифицированному T4L-SAMHD1-CtD, через 15 минут (рис. 1G и S2F). Опять же, эти данные подтверждают функциональную важность SAMHD1-CtD для опосредованного Vpr mus рекрутирования в убиквитинлигазу CRL4 DCAF1 .
Анализ кристаллической структуры белковых комплексов DDB1/DCAF1-CtD, связанных с апо- и Vprmus
Определяли комплекс DDB1/DCAF1-CtD и тройной комплекс слитого белка DDB1/DCAF1-CtD/T4L-Vpr mus (остатки 1-92).Структуры были решены с использованием молекулярной замены и уточнены до разрешений 3,1 Å и 2,5 Å соответственно (таблица S1). Vpr mus принимает трехспиральный пучок, стабилизированный координацией иона цинка остатками His и Cys на спирали-1 и на С-конце (рис. 2А). Наложение структур Vpr mus на ранее определенные структуры Vpx sm [50], Vpx mnd2 [51, 52] и Vpr HIV-1 [54] выявляет консервативную трехспиральную складку пучка и сходное положение. спиральных пучков на DCAF1-CtD (рис. S3A).Кроме того, большинство боковых цепей, участвующих во взаимодействии с DCAF1, являются консервативными по типу во всех белках Vpx и Vpr (рис. S3B-G и S6A), что убедительно свидетельствует об общем молекулярном механизме захвата CRL4-DCAF1 хозяина семейством Vpx/Vpr. вспомогательных белков. Однако существуют также значительные различия в длине и регистре спирали, а также конформационные вариации в области петли на N-конце спирали-1, в начале спирали-1 и в петле между спиралями-2 и -3 (рис. S3A). ).
Vpr mus принимает трехспиральный пучок, стабилизированный координацией иона цинка остатками His и Cys на спирали-1 и на С-конце (рис. 2А). Наложение структур Vpr mus на ранее определенные структуры Vpx sm [50], Vpx mnd2 [51, 52] и Vpr HIV-1 [54] выявляет консервативную трехспиральную складку пучка и сходное положение. спиральных пучков на DCAF1-CtD (рис. S3A).Кроме того, большинство боковых цепей, участвующих во взаимодействии с DCAF1, являются консервативными по типу во всех белках Vpx и Vpr (рис. S3B-G и S6A), что убедительно свидетельствует об общем молекулярном механизме захвата CRL4-DCAF1 хозяина семейством Vpx/Vpr. вспомогательных белков. Однако существуют также значительные различия в длине и регистре спирали, а также конформационные вариации в области петли на N-конце спирали-1, в начале спирали-1 и в петле между спиралями-2 и -3 (рис. S3A). ).

( A ) Общая структура комплекса DDB1/DCAF1-CtD/Vpr mus в двух проекциях. DCAF1-CtD показан серым рисунком и полупрозрачной поверхностью. Vpr mus показан в виде темно-зеленого изображения, а координированный ион цинка показан в виде серой сферы. T4L и DDB1 опущены для ясности. ( B ) Наложение апо-DCAF1-CtD (голубой рисунок) на связанный с Vpr mus DCAF1-CtD (серый/зеленый рисунок).Показаны только области DCAF1-CtD со значительными структурными различиями между формами, связанными с апо и Vpr mus . Неупорядоченные петли обозначены пунктирными линиями. ( C ) Сравнение бинарных комплексов Vpr mus /DCAF1-CtD и тройных комплексов Vpx sm /DCAF1-CtD/SAMHD1-CtD. Для DCAF1-CtD показана только N-концевая область «кислой петли». Vpr mus , DCAF1-CtD и связанный цинк окрашены, как в A ; Vpx sm представлен оранжевым рисунком, а SAMHD1-CtD — розовым. Выбранные боковые цепи Vpr/Vpx/DCAF1-CtD показаны в виде палочек, а электростатические взаимодействия между этими боковыми цепями обозначены пунктирными линиями. ( D ) Анализ GF in vitro воссоздания белковых комплексов, содержащих DDB1/DCAF1-CtD/Vpr mus или мутант Vpr mus R15E/R75E и SAMHD1. Анализы SDS-PAGE соответствующих фракций GF показаны под хроматограммой с цветными прямоугольниками в соответствии с хроматограммой. ( E-F ) In vitro восстановление белковых комплексов, содержащих SAMHD1 и Vpr mus R15E/R75E ( E ) или DDB1/DCAF1-CtD и Vpr mus R17 F 9027E (9027E).Анализы SDS-PAGE соответствующих фракций GF показаны под хроматограммой с цветными прямоугольниками в соответствии с хроматограммой. Звездочка и двойная звездочка указывают на незначительное загрязнение оставшейся протеазой GST-3C и тегом очистки GST соответственно.
Выбранные боковые цепи Vpr/Vpx/DCAF1-CtD показаны в виде палочек, а электростатические взаимодействия между этими боковыми цепями обозначены пунктирными линиями. ( D ) Анализ GF in vitro воссоздания белковых комплексов, содержащих DDB1/DCAF1-CtD/Vpr mus или мутант Vpr mus R15E/R75E и SAMHD1. Анализы SDS-PAGE соответствующих фракций GF показаны под хроматограммой с цветными прямоугольниками в соответствии с хроматограммой. ( E-F ) In vitro восстановление белковых комплексов, содержащих SAMHD1 и Vpr mus R15E/R75E ( E ) или DDB1/DCAF1-CtD и Vpr mus R17 F 9027E (9027E).Анализы SDS-PAGE соответствующих фракций GF показаны под хроматограммой с цветными прямоугольниками в соответствии с хроматограммой. Звездочка и двойная звездочка указывают на незначительное загрязнение оставшейся протеазой GST-3C и тегом очистки GST соответственно.
Vpr mus связывается сбоку и сверху дискообразного 7-лопастного β-пропеллера (BP) домена DCAF1-CtD с общей площадью контактной поверхности ~1600 Å 2 , включающей три основные области взаимодействия . Удлиненный N-конец Vpr mus присоединяется к щели между лопастями DCAF1 BP 1 и 2 посредством нескольких водородных связей, электростатических и гидрофобных взаимодействий (рис. S3B-D). Вторая, меньшая площадь контакта образуется за счет гидрофобного взаимодействия между остатками Vpr mus L31 и E34 из Helix-1 и DCAF1 W1156, расположенными в петле на вершине лопасти BP 2 (рис. S3E). Третья поверхность взаимодействия включает С-концевую половину Vpr mus Helix-3, которая вставляется в гребень на вершине DCAF1 (S3F, G Fig).
Удлиненный N-конец Vpr mus присоединяется к щели между лопастями DCAF1 BP 1 и 2 посредством нескольких водородных связей, электростатических и гидрофобных взаимодействий (рис. S3B-D). Вторая, меньшая площадь контакта образуется за счет гидрофобного взаимодействия между остатками Vpr mus L31 и E34 из Helix-1 и DCAF1 W1156, расположенными в петле на вершине лопасти BP 2 (рис. S3E). Третья поверхность взаимодействия включает С-концевую половину Vpr mus Helix-3, которая вставляется в гребень на вершине DCAF1 (S3F, G Fig).
Наложение кристаллических структур, связанных с apo-DDB1/DCAF1-CtD и Vpr mus , выявляет конформационные изменения в DCAF1 при ассоциации Vpr mus . Связывание N-концевого плеча Vpr с mus вызывает лишь незначительную перестройку петли в лезвии 2 BP (рис. S3C). Напротив, значительные структурные изменения происходят на верхней поверхности домена BP: полярные и гидрофобные взаимодействия остатков DCAF1 P1329, F1330, F1355, N1371, L1378, M1380 и T1382 с боковыми цепями Vpr mus T79, R83, R86 и E87 в Helix-3 приводит к стабилизации растяжения последовательности, соединяющей лопасти BP 6 и 7 («C-терминальная петля», рис. 2B и S3F).Более того, электростатические взаимодействия боковых цепей остатков R15, R75 и R76 Vpr mus с DCAF1 E1088, E1091 и E1093 фиксируют конформацию «кислотной петли» перед лезвием 1 BP, которая также неструктурирована и гибка в отсутствие Vpr. mus (рис. 2B, C и S3D, F).
2B и S3F).Более того, электростатические взаимодействия боковых цепей остатков R15, R75 и R76 Vpr mus с DCAF1 E1088, E1091 и E1093 фиксируют конформацию «кислотной петли» перед лезвием 1 BP, которая также неструктурирована и гибка в отсутствие Vpr. mus (рис. 2B, C и S3D, F).
Примечательно, что в ранее определенных структурах комплексов Vpx/DCAF1/SAMHD1 «кислотная петля» является центральной точкой тройного контакта, обеспечивая платформу для связывания положительно заряженных боковых цепей аминокислот либо на N-, либо на C-конце SAMHD1 [ 50–52].Например, Vpx sm позиционирует SAMHD1-CtD таким образом, что SAMHD1 K622 вступает в электростатическое взаимодействие с остатком «кислой петли» DCAF1 D1092 (рис. 2C, левая панель). Однако в кристаллической структуре Vpr mus связанный Vpr mus теперь блокирует доступ к соответствующему карману для связывания SAMHD1-CtD, в частности, за счет расположения расширенной N-концевой петли, которая предшествует Helix-1. Кроме того, боковые цепи Vpr mus R15, R75 и R76 нейтрализуют «кислотную петлю» DCAF1, предотвращая образование дополнительных солевых мостиков к основным остаткам в SAMHD1-CtD (рис. 2C, правая панель).
Кроме того, боковые цепи Vpr mus R15, R75 и R76 нейтрализуют «кислотную петлю» DCAF1, предотвращая образование дополнительных солевых мостиков к основным остаткам в SAMHD1-CtD (рис. 2C, правая панель).
Чтобы подтвердить важность остатков R15 и R75 Vpr mus для связывания DCAF1-CtD, путем сайт-направленного мутагенеза были созданы мутации с реверсированием заряда глутаматов. Спектр кругового дихроизма (CD) двойного мутантного GST-слитого белка Vpr mus R15E R75E был идентичен таковому дикого типа, что указывает на сходное содержание вторичной структуры и, следовательно, на отсутствие серьезных структурных нарушений, вызванных аминокислотными заменами (S3H Fig). Влияние двойного мутанта Vpr mus R15E R75E на сборку комплекса затем анализировали с помощью GF-хроматографии.SDS-PAGE анализ полученного хроматографического профиля показывает почти полную потерю пика комплекса DDB1/DCAF1-CtD/Vpr mus /SAMHD1 (рис. 2D, фракция 6) по сравнению с диким типом, одновременно с обогащением ( i) некоторая часть Vpr mus R15E R75E, связанная с DDB1/DCAF1-CtD (рис. 2D, фракция 7), (ii) свободный DDB1/DCAF1-CtD (фракция 7-8) и (iii) Vpr mus Бинарный комплекс R15E R75E/SAMHD1 (рис. 2D, фракции 8-9). Это предполагает, что изменение заряда боковых цепей Vpr mus R15 и R75 ослабляет сильную ассоциацию с DCAF1, наблюдаемую в Vpr mus дикого типа, из-за потери электростатического взаимодействия с «кислотной петлей», в соответствии с кристаллической структурой.Следовательно, некоторая часть SAMHD1, связанного с Vpr, диссоциирует. Это мнение дополнительно подтверждается анализом GF бинарных комбинаций двойного мутанта Vpr mus R15E R75E либо с SAMHD1, либо с DDB1/DCAF1-CtD. Инкубация Vpr mus R15E R75E с SAMHD1 с последующим GF приводит к совместной элюции обоих белков, что сопровождается сдвигом пика элюции в сторону более высокой кажущейся молекулярной массы по сравнению с одним SAMHD1 (рис. 2E, фракции 8–9). . Напротив, инкубация двойного мутанта Vpr mus с DDB1/DCAF1-CtD не изменяет объем элюции видов DDB/DCAF1-CtD, и при анализе SDS-PAGE в плазме SDS-PAGE невозможно обнаружить полосу, соответствующую Vpr.
2D, фракция 7), (ii) свободный DDB1/DCAF1-CtD (фракция 7-8) и (iii) Vpr mus Бинарный комплекс R15E R75E/SAMHD1 (рис. 2D, фракции 8-9). Это предполагает, что изменение заряда боковых цепей Vpr mus R15 и R75 ослабляет сильную ассоциацию с DCAF1, наблюдаемую в Vpr mus дикого типа, из-за потери электростатического взаимодействия с «кислотной петлей», в соответствии с кристаллической структурой.Следовательно, некоторая часть SAMHD1, связанного с Vpr, диссоциирует. Это мнение дополнительно подтверждается анализом GF бинарных комбинаций двойного мутанта Vpr mus R15E R75E либо с SAMHD1, либо с DDB1/DCAF1-CtD. Инкубация Vpr mus R15E R75E с SAMHD1 с последующим GF приводит к совместной элюции обоих белков, что сопровождается сдвигом пика элюции в сторону более высокой кажущейся молекулярной массы по сравнению с одним SAMHD1 (рис. 2E, фракции 8–9). . Напротив, инкубация двойного мутанта Vpr mus с DDB1/DCAF1-CtD не изменяет объем элюции видов DDB/DCAF1-CtD, и при анализе SDS-PAGE в плазме SDS-PAGE невозможно обнаружить полосу, соответствующую Vpr. соответствующие дроби (рис.2F, фракции 7-8). Эти данные ясно демонстрируют потерю взаимодействия с DDB1/DCAF1-CtD при изменении заряда остатков 15 и 75 Vpr mus , в то время как активность связывания SAMHD1 сохраняется.
соответствующие дроби (рис.2F, фракции 7-8). Эти данные ясно демонстрируют потерю взаимодействия с DDB1/DCAF1-CtD при изменении заряда остатков 15 и 75 Vpr mus , в то время как активность связывания SAMHD1 сохраняется.
Крио-ЭМ анализ CRL4-NEDD8
DCAF1-CtD /Vprmus/SAMHD1 конформационные состояния и динамика Чтобы получить представление о структуре Vpr mus в контексте полной сборки CRL4 и понять механизм рекрутирования SAMHD1 , мы начали крио-ЭМ анализ сборки CRL4 DCAF1-CtD /Vpr mus /SAMHD1.В этих исследованиях небольшой убиквитин-подобный белок NEDD8 был ферментативно присоединен к субъединице CUL4, чтобы получить ее активную форму (S4A Fig) [55]. Комплекс CRL4-NEDD8 DCAF1-CtD /Vpr mus /SAMHD1 был воссоздан in vitro и очищен с помощью GF-хроматографии (S4B Fig). 2D-классификация полученных изображений частиц выявила значительную композиционную и конформационную гетерогенность, особенно в отношении присутствия и положения субкомплекса CUL4-NEDD8/ROC1 (стебель) по отношению к DDB1/DCAF1/Vpr mus (ядро) (рис. S4C). .
S4C). .
Два последовательных раунда 3D-классификации дали три популяции частиц, что привело к 3D-реконструкциям с разрешением 8-10 Å, которые содержали как ядро CRL4, связанное с Vpr mus , так и стебель (конформационные состояния -1, -2 и -3 , рис. 3A и S4D-F). Качество трехмерных объемов было достаточным для соответствия кристаллографическим моделям ядра (рис. 2) и стебля (PDB 2hye) [15] как твердых тел (рис. 3B и S4G). Для субъединицы каталитического RING-домена ROC1 присутствовала только фрагментированная электронная плотность рядом с положением, которое она занимает в кристаллографической модели (рис. S4G).Во всех трех состояниях электронная плотность избирательно отсутствовала для С-концевого домена В (WHB) крылатой спирали CUL4 (остатки 674–759), содержащего сайт модификации NEDD8 (К705), и для предшествующей α-спирали, соединяющей N-концевой домен CUL4 к домену WHB (фиг. S4G). В соответствии с этим наблюдением положения NEDD8, присоединенного к CRL5, и домена CRL4 ROC1 RING стерически несовместимы при наложении их соответствующих кристаллических структур (S4H Fig) [56].
( A ) Два вида наложения CRL4-NEDD8 DCAF1-CtD /Vpr mus /SAMHD1 крио-ЭМ реконструкции (конформационное состояние-1 – светло-зеленый, состояние-2 – лосось, состояние-3 – фиолетовый). Части плотностей, соответствующие DDB1 BPA/BPC, DCAF1-CtD и Vpr mus , были наложены друг на друга. ( B ) Два вида суперпозиции молекулярных моделей DDB1/DCAF1-CtD/Vpr mus и CUL4/ROC1 (PDB 2hye) [15], которые были приспособлены как твердые тела к соответствующим крио-ЭМ плотностям; модели ориентированы как в A .DDB1/DCAF1-CtD/Vpr mus показан на рис. 2A, CUL4 показан в виде мультяшного изображения, окрашен как A , а ROC1 показан в виде голубого мультяшного изображения. ( C ) Сравнение самых внешних ориентаций стебля CUL4, наблюдаемых в представленном здесь крио-ЭМ анализе (состояния-1 и -3, окрашены как B , показывают вращение DDB1 BPB на 119,5 °) с двумя крайними положениями стебля. присутствует в предыдущих кристаллических структурах (PDB 4a0l [13], PDB 6dsz [115], окрашен в серый цвет, показывает вращение DDB1 BPB на 143,4°).
присутствует в предыдущих кристаллических структурах (PDB 4a0l [13], PDB 6dsz [115], окрашен в серый цвет, показывает вращение DDB1 BPB на 143,4°).
Выравнивание трехмерных объемов из состояний-1, -2 и -3 показывает, что плотности ядра, представляющие DDB1 BPA, BPC, DCAF1-CtD и Vpr mus , хорошо накладываются друг на друга, указывая на то, что эти компоненты не подвергаются значительным конформационным флуктуациям и, таким образом, образуют жесткая платформа для связывания субстрата и прикрепления стебля CRL4 (рис. 3). Однако вращение DDB1 BPB вокруг шарнира, соединяющего его с BPC, приводит к трем разным ориентациям стебельчатых областей состояния-1, -2 и -3 относительно ядра.Углы поворота BPB были измерены как 69° между состояниями-1 и -2 и 50° между состояниями-2 и -3.
Эти данные согласуются с предыдущим прогнозом, основанным на обширном сравнительном анализе кристаллической структуры, который постулировал прибл. Вращение ножки CRL4 вокруг ядра на 150° [13, 15, 16, 19, 57]. Однако крайние левая и правая ориентации CUL4, наблюдаемые здесь, состояния-1 и -3 из нашего крио-ЭМ-анализа, указывают на несколько более узкий диапазон вращения стебля (119 °) по сравнению с конформациями самого дальнего стебля, смоделированными на основе ранее определенных кристаллических структур. (143°) (рис. 3С).Возможное объяснение этого несоответствия возникает из проверки плотностей крио-ЭМ и подобранных моделей, показывающих, что наряду с основным интерфейсом взаимодействия на DDB1 BPB существуют дополнительные молекулярные контакты между CUL4 и DDB1. В частности, в состоянии-1 имеется контакт между петлей, соединяющей спирали D и E кулинового повтора CUL4 (CR)1 (остатки 161–169), и петлей, выступающей из лопасти 3 BP DDB1 BPC (остатки 795–801, S4I рис.). В состоянии 3 петля между спиралями D и E CUL4 CR2 (остатки 275–282) примыкает к области в С-концевом спиральном домене DDB1 (остатки 1110–1127, S4J Fig).Эти вспомогательные взаимодействия могут потребоваться для блокировки наблюдаемых здесь самых удаленных позиций стебля, чтобы ограничить диапазон вращения CUL4.
Однако крайние левая и правая ориентации CUL4, наблюдаемые здесь, состояния-1 и -3 из нашего крио-ЭМ-анализа, указывают на несколько более узкий диапазон вращения стебля (119 °) по сравнению с конформациями самого дальнего стебля, смоделированными на основе ранее определенных кристаллических структур. (143°) (рис. 3С).Возможное объяснение этого несоответствия возникает из проверки плотностей крио-ЭМ и подобранных моделей, показывающих, что наряду с основным интерфейсом взаимодействия на DDB1 BPB существуют дополнительные молекулярные контакты между CUL4 и DDB1. В частности, в состоянии-1 имеется контакт между петлей, соединяющей спирали D и E кулинового повтора CUL4 (CR)1 (остатки 161–169), и петлей, выступающей из лопасти 3 BP DDB1 BPC (остатки 795–801, S4I рис.). В состоянии 3 петля между спиралями D и E CUL4 CR2 (остатки 275–282) примыкает к области в С-концевом спиральном домене DDB1 (остатки 1110–1127, S4J Fig).Эти вспомогательные взаимодействия могут потребоваться для блокировки наблюдаемых здесь самых удаленных позиций стебля, чтобы ограничить диапазон вращения CUL4.
Молекулярный механизм нацеливания на SAMHD1
Повторный анализ данных крио-ЭМ, включающий отбор частиц на основе матрицы и обширную трехмерную классификацию, позволил выделить дополнительную однородную популяцию частиц (S5A, B Fig). Это подмножество изображений частиц дало трехмерную реконструкцию с номинальным разрешением 7,3 Å, которая содержала только электронную плотность, соответствующую ядру CRL4 (рис. S5A-D).Молекулярные модели доменов DDB1 BP A и C (BPA, BPC), DCAF1-CtD и Vpr mus , полученные из нашей кристаллической структуры (рис. 2), могут быть помещены в виде твердых тел в этот объем крио-ЭМ (рис. 4A). . Для большей части SAMHD1 не было видно явной электронной плотности. Однако при ближайшем рассмотрении был обнаружен дополнительный трубчатый, слегка дугообразный элемент плотности, ок. 35 Å в длину, расположен на верхней поверхности спирального пучка Vpr mus , примерно в 17 Å от и напротив интерфейса связывания Vpr mus / DCAF1-CtD (рис. 4A, красные стрелки).Один конец тубулярного объема контактирует с серединой Vpr mus Helix-1, а другой конец образует дополнительные контакты с С-концом Helix-2 и N-концом Helix-3 (рис. 4B). Локальное разрешение 7,5-8 Å (рис. S5C) исключало подгонку атомной модели. Принимая во внимание биохимические данные, показывающие, что SAMHD1-CtD достаточно для рекрутирования в DDB1/DCAF1/Vpr mus , мы предполагаем, что эта наблюдаемая особенность электронной плотности соответствует области SAMHD1-CtD, которая физически взаимодействует с Vpr mus .Учитывая его размеры, предполагаемая плотность SAMHD1-CtD может вместить ок. 10 аминокислотных остатков в полностью вытянутой конформации или до 23 остатков в изогнутой спиральной конфигурации. Все предыдущие анализы кристаллической структуры [46], а также предсказания вторичной структуры показывают, что остатки SAMHD1 на С-конце каталитического HD-домена и С-концевой доли (аминокислоты 599-626) неупорядочены в отсутствие дополнительных партнеров по связыванию.
4A, красные стрелки).Один конец тубулярного объема контактирует с серединой Vpr mus Helix-1, а другой конец образует дополнительные контакты с С-концом Helix-2 и N-концом Helix-3 (рис. 4B). Локальное разрешение 7,5-8 Å (рис. S5C) исключало подгонку атомной модели. Принимая во внимание биохимические данные, показывающие, что SAMHD1-CtD достаточно для рекрутирования в DDB1/DCAF1/Vpr mus , мы предполагаем, что эта наблюдаемая особенность электронной плотности соответствует области SAMHD1-CtD, которая физически взаимодействует с Vpr mus .Учитывая его размеры, предполагаемая плотность SAMHD1-CtD может вместить ок. 10 аминокислотных остатков в полностью вытянутой конформации или до 23 остатков в изогнутой спиральной конфигурации. Все предыдущие анализы кристаллической структуры [46], а также предсказания вторичной структуры показывают, что остатки SAMHD1 на С-конце каталитического HD-домена и С-концевой доли (аминокислоты 599-626) неупорядочены в отсутствие дополнительных партнеров по связыванию. Соответственно, N-концевые глобулярные домены молекулы SAMHD1 могут быть гибко связаны с C-концевой связью, идентифицированной здесь.В этом случае основная масса SAMHD1 выбирает множество позиций относительно ядра DDB1/DCAF1-CtD/Vpr mus и, следовательно, усредняется в процессе крио-ЭМ реконструкции.
Соответственно, N-концевые глобулярные домены молекулы SAMHD1 могут быть гибко связаны с C-концевой связью, идентифицированной здесь.В этом случае основная масса SAMHD1 выбирает множество позиций относительно ядра DDB1/DCAF1-CtD/Vpr mus и, следовательно, усредняется в процессе крио-ЭМ реконструкции.
( A ) Два вида крио-ЭМ реконструкции ядра CRL4-NEDD8 DCAF1-CtD /Vpr mus /SAMHD1. Кристаллическая структура комплекса DDB1/DCAF1-CtD/Vpr mus была вписана в крио-ЭМ-плотность как твердое тело и показана теми же цветами, что и на рис. 2А.Модель и плотность DDB1 BPB были удалены для ясности. Красными стрелками отмечена дополнительная плотность на верхней поверхности пучка спирали Vpr mus . ( B ) Подробное изображение электронной плотности SAMHD1-CtD. Модель имеет ту же ориентацию, что и в A , левая панель. Выбранные остатки Vpr mus W29 и A66, которые находятся в тесном контакте с дополнительной плотностью, показаны красным цветом. ( C ) Реконструкция in vitro белковых комплексов, содержащих DDB1/DCAF1-CtD, Vpr mus или мутант Vpr mus W29A/A66W, и SAMHD1, оцененная с помощью аналитического GF.Анализы SDS-PAGE соответствующих фракций GF показаны под хроматограммой с цветными прямоугольниками в соответствии с хроматограммой. ( D-E ) In vitro восстановление белковых комплексов, содержащих SAMHD1 и Vpr mus W29A/A66W ( D ) или DDB1/DCAF1-CtD и Vpr mus W29A/A666W ( 9026W). Анализы SDS-PAGE соответствующих фракций GF показаны под хроматограммой с цветными прямоугольниками в соответствии с хроматограммой.Звездочка и двойная звездочка указывают на незначительное загрязнение оставшейся протеазой GST-3C и тегом очистки GST соответственно.
Выбранные остатки Vpr mus W29 и A66, которые находятся в тесном контакте с дополнительной плотностью, показаны красным цветом. ( C ) Реконструкция in vitro белковых комплексов, содержащих DDB1/DCAF1-CtD, Vpr mus или мутант Vpr mus W29A/A66W, и SAMHD1, оцененная с помощью аналитического GF.Анализы SDS-PAGE соответствующих фракций GF показаны под хроматограммой с цветными прямоугольниками в соответствии с хроматограммой. ( D-E ) In vitro восстановление белковых комплексов, содержащих SAMHD1 и Vpr mus W29A/A66W ( D ) или DDB1/DCAF1-CtD и Vpr mus W29A/A666W ( 9026W). Анализы SDS-PAGE соответствующих фракций GF показаны под хроматограммой с цветными прямоугольниками в соответствии с хроматограммой.Звездочка и двойная звездочка указывают на незначительное загрязнение оставшейся протеазой GST-3C и тегом очистки GST соответственно.
Чтобы проверить эту гипотезу, аминокислотные остатки Vpr mus в непосредственной близости от предполагаемой плотности SAMHD1-CtD были заменены с помощью сайт-направленного мутагенеза.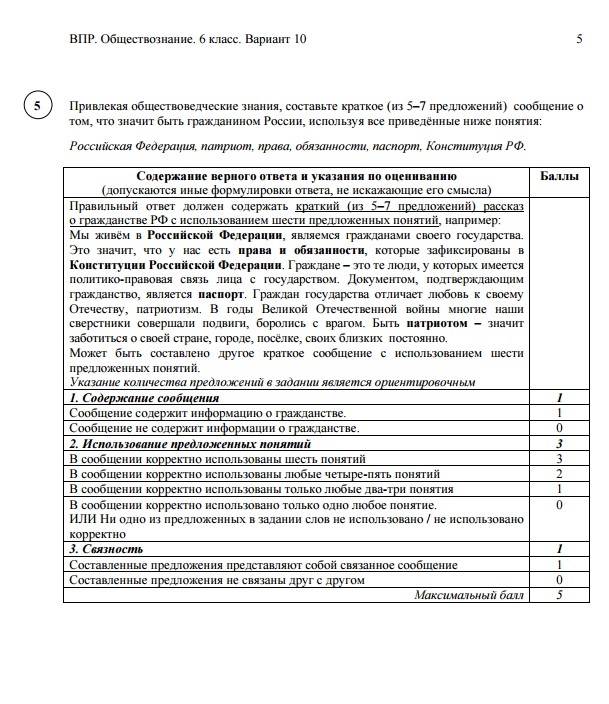 В частности, Vpr mus W29 был заменен на аланин, чтобы заблокировать гидрофобный контакт с SAMHD1-CtD с участием ароматической боковой цепи, а Vpr mus A66 был заменен на объемный триптофан, чтобы вызвать стерическое столкновение с SAMHD1-CtD. (рис. 4В).Структурная целостность двойного мутанта Vpr mus W29A A66W была подтверждена с помощью спектроскопии CD (S3H Fig), а затем было оценено образование комплекса с DDB1/DCAF1-CtD и SAMHD1 с помощью аналитического GF. По сравнению с Vpr mus дикого типа, мутант W29A A66W показал сниженную пиковую интенсивность комплекса DDB1/DCAF1-CtD/Vpr mus /SAMHD1 (рис. 4C, фракция 6) одновременно с (i) обогащением DDB1/DCAF1- Тройной комплекс CtD/Vpr mus , субстехиометрически связанный с SAMHD1 (рис. 4C, фракция 7), (ii) избыток комплекса DDB1/DCAF1-CtD (рис. 4C, фракция 8) и (iii) избыток SAMHD1 (рис. 4C) , дроби 9-10).Кроме того, с помощью GF также анализировали бинарные комбинации двойного мутанта Vpr mus W29A A66W с DDB1/DCAF1-CtD или SAMHD1.
В частности, Vpr mus W29 был заменен на аланин, чтобы заблокировать гидрофобный контакт с SAMHD1-CtD с участием ароматической боковой цепи, а Vpr mus A66 был заменен на объемный триптофан, чтобы вызвать стерическое столкновение с SAMHD1-CtD. (рис. 4В).Структурная целостность двойного мутанта Vpr mus W29A A66W была подтверждена с помощью спектроскопии CD (S3H Fig), а затем было оценено образование комплекса с DDB1/DCAF1-CtD и SAMHD1 с помощью аналитического GF. По сравнению с Vpr mus дикого типа, мутант W29A A66W показал сниженную пиковую интенсивность комплекса DDB1/DCAF1-CtD/Vpr mus /SAMHD1 (рис. 4C, фракция 6) одновременно с (i) обогащением DDB1/DCAF1- Тройной комплекс CtD/Vpr mus , субстехиометрически связанный с SAMHD1 (рис. 4C, фракция 7), (ii) избыток комплекса DDB1/DCAF1-CtD (рис. 4C, фракция 8) и (iii) избыток SAMHD1 (рис. 4C) , дроби 9-10).Кроме того, с помощью GF также анализировали бинарные комбинации двойного мутанта Vpr mus W29A A66W с DDB1/DCAF1-CtD или SAMHD1. Эти данные показывают потерю взаимодействия SAMHD1 (рис. 4D), в то время как способность связывать DDB1/DCAF1-CtD сохраняется (рис. 4E). Вместе эти биохимические анализы подтверждают расположение сайта связывания SAMHD1-CtD на верхней поверхности пучка спирали Vpr mus , как предполагается крио-ЭМ реконструкцией среднего разрешения.
Эти данные показывают потерю взаимодействия SAMHD1 (рис. 4D), в то время как способность связывать DDB1/DCAF1-CtD сохраняется (рис. 4E). Вместе эти биохимические анализы подтверждают расположение сайта связывания SAMHD1-CtD на верхней поверхности пучка спирали Vpr mus , как предполагается крио-ЭМ реконструкцией среднего разрешения.
Для получения дополнительных экспериментальных данных сборка CRL4 DCAF1-CtD /Vpr mus /SAMHD1 была дополнительно исследована с помощью масс-спектрометрии сшивки (CLMS) с использованием фотореактивного сшивающего агента сульфо-SDA.Это бифункциональное соединение содержит функциональную группу сложного эфира NHS на одном конце, которая реагирует с первичными аминами и гидроксильными группами, а другой конец ковалентно связывается с любой боковой цепью аминокислоты в пределах досягаемости при УФ-активации через промежуточный карбен [58]. Соответственно, инкубация белков или белковых комплексов с сульфо-SDA с последующим УФ-освещением позволяет сшивать с высокой плотностью боковые цепи лизина и, в меньшей степени, серина, треонина и тирозина с аминокислотами в пределах досягаемости SDA. спейсерная группа с более быстрой кинетикой, чем сшивающие агенты на основе чистых эфиров NHS, из-за короткого периода полураспада промежуточного соединения, активируемого УФ-излучением.Сшитые пептиды впоследствии идентифицируются с помощью масс-спектрометрии и дают представление о топологии и расстояниях между остатками белков и комплексов [59]. В случае сборки CRL4 DCAF1-CtD /Vpr mus /SAMHD1 большинство идентифицированных поперечных связей, которые могут быть отображены на структуру (468/519, 90,2%), находятся в пределах установленного порога нарушения 25 Å. по геометрии проставки SDA. Интересно, что 8 из 11 поперечных связей между DDB1 и CUL4 удовлетворяются моделью состояния 1, но все чаще нарушаются в состояниях 2 и -3 (S5E Fig), поддерживая в решении ротационную гибкость стебля CRL4 по отношению к DDB1/DCAF1-CtD, наблюдаемый в крио-ЭМ (рис. 3).
спейсерная группа с более быстрой кинетикой, чем сшивающие агенты на основе чистых эфиров NHS, из-за короткого периода полураспада промежуточного соединения, активируемого УФ-излучением.Сшитые пептиды впоследствии идентифицируются с помощью масс-спектрометрии и дают представление о топологии и расстояниях между остатками белков и комплексов [59]. В случае сборки CRL4 DCAF1-CtD /Vpr mus /SAMHD1 большинство идентифицированных поперечных связей, которые могут быть отображены на структуру (468/519, 90,2%), находятся в пределах установленного порога нарушения 25 Å. по геометрии проставки SDA. Интересно, что 8 из 11 поперечных связей между DDB1 и CUL4 удовлетворяются моделью состояния 1, но все чаще нарушаются в состояниях 2 и -3 (S5E Fig), поддерживая в решении ротационную гибкость стебля CRL4 по отношению к DDB1/DCAF1-CtD, наблюдаемый в крио-ЭМ (рис. 3).
Дополнительные 300 поперечных связей включали SAMHD1, простирающиеся до С-концевой половины CUL4, до участка последовательности DDB1, включающего аминокислотные остатки 900–1000, до частей DCAF1-CtD и до Vpr mus (рис. 5A, С5Е). Остатки CRL4 DCAF1-CtD /Vpr mus , демонстрирующие перекрестные связи с SAMHD1, были картированы на модели состояния 2 и показали наличие большой, но определенной поверхности взаимодействия (рис. 5B). Важно отметить, что были очевидны перекрестные связи между С-концом SAMHD1-CtD (остатки K622, K626) и областью в Vpr mus Helix-1 (остатки 27–36), которая образует часть предполагаемого SAMHD1-CtD. интерфейс связывания, наблюдаемый в крио-ЭМ, и который содержит Vpr mus W29, один из остатков, замененных в мутагенезе и биохимическом анализе, представленном выше (фиг. 5B, фиолетовые сферы).Кроме того, аминокислотные остатки из N-концевой части SAMHD1-CtD (остатки K595, K596, T602-S606) сшиты вблизи «кислой петли» DCAF1-CtD (остатки 1092-1096), которая иммобилизована Vpr рядом с предполагаемым сайтом связывания SAMHD1-CtD и с самым С-концом CUL4 (остатки Y744, A759), который также примыкает к предсказанному положению связывания SAMHD1-CtD (рис.
5A, С5Е). Остатки CRL4 DCAF1-CtD /Vpr mus , демонстрирующие перекрестные связи с SAMHD1, были картированы на модели состояния 2 и показали наличие большой, но определенной поверхности взаимодействия (рис. 5B). Важно отметить, что были очевидны перекрестные связи между С-концом SAMHD1-CtD (остатки K622, K626) и областью в Vpr mus Helix-1 (остатки 27–36), которая образует часть предполагаемого SAMHD1-CtD. интерфейс связывания, наблюдаемый в крио-ЭМ, и который содержит Vpr mus W29, один из остатков, замененных в мутагенезе и биохимическом анализе, представленном выше (фиг. 5B, фиолетовые сферы).Кроме того, аминокислотные остатки из N-концевой части SAMHD1-CtD (остатки K595, K596, T602-S606) сшиты вблизи «кислой петли» DCAF1-CtD (остатки 1092-1096), которая иммобилизована Vpr рядом с предполагаемым сайтом связывания SAMHD1-CtD и с самым С-концом CUL4 (остатки Y744, A759), который также примыкает к предсказанному положению связывания SAMHD1-CtD (рис.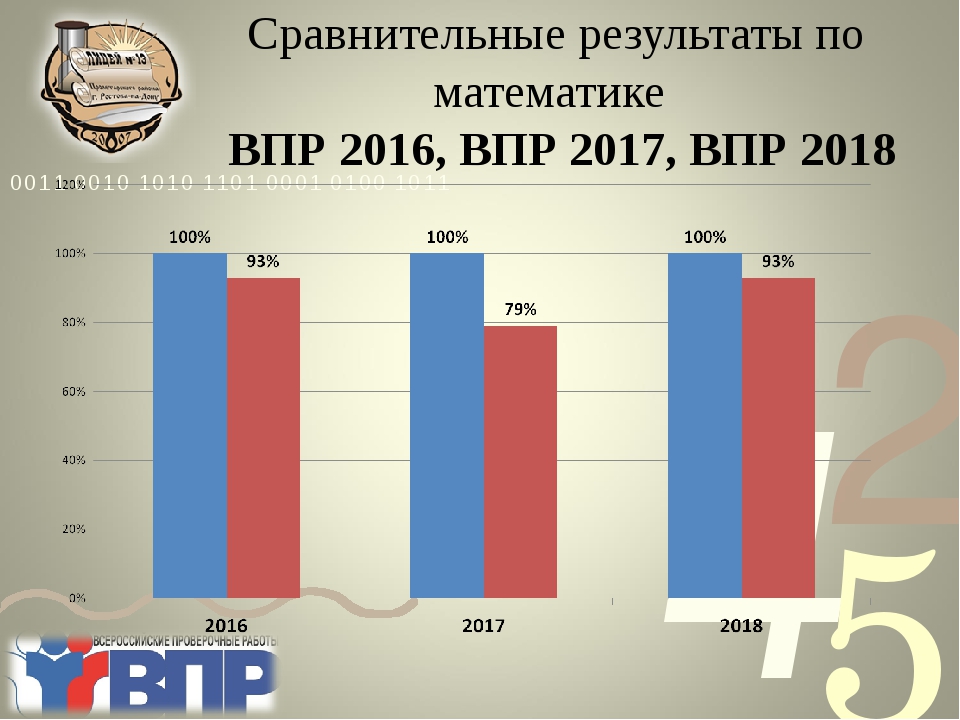 5B, розовые сферы). Наконец, перекрестные связи в N-концевом домене SAMHD1 и каталитическом домене HD почти исключительно включали участки поверхности CRL4 DCAF1-CtD /Vpr mus , окружающие предполагаемую точку прикрепления SAMHD1-CtD и обращенные к ней (рис.5В, светло-коричневые сферы). Эти наблюдения совместимы с рекрутированием SAMHD-CtD на верхней поверхности пучка спирали Vpr, на что указывает крио-ЭМ. Кроме того, пространственное распределение поперечных связей, включающих N-концевые домены SAMHD1, предполагает, что они гибко связаны с SAMHD1-CtD, что приводит к сильно изменчивому расположению относительно CRL4 DCAF1-CtD /Vpr mus и, таким образом, предлагает множество возможностей сшивки с соседними компонентами CRL4, опять же в соответствии с результатами крио-ЭМ-реконструкции, особенно с учетом позиционной неоднородности ножки CUL4 (рис.3).
5B, розовые сферы). Наконец, перекрестные связи в N-концевом домене SAMHD1 и каталитическом домене HD почти исключительно включали участки поверхности CRL4 DCAF1-CtD /Vpr mus , окружающие предполагаемую точку прикрепления SAMHD1-CtD и обращенные к ней (рис.5В, светло-коричневые сферы). Эти наблюдения совместимы с рекрутированием SAMHD-CtD на верхней поверхности пучка спирали Vpr, на что указывает крио-ЭМ. Кроме того, пространственное распределение поперечных связей, включающих N-концевые домены SAMHD1, предполагает, что они гибко связаны с SAMHD1-CtD, что приводит к сильно изменчивому расположению относительно CRL4 DCAF1-CtD /Vpr mus и, таким образом, предлагает множество возможностей сшивки с соседними компонентами CRL4, опять же в соответствии с результатами крио-ЭМ-реконструкции, особенно с учетом позиционной неоднородности ножки CUL4 (рис.3).
( A ) mus и SAMHD1, идентифицированные с помощью CLMS.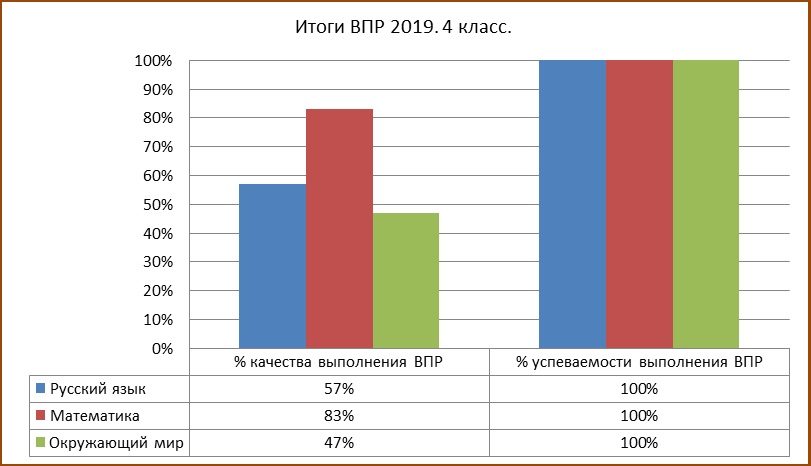 Белки имеют цветовую кодировку, как на рис. 3, 4, и черный/белый SAMHD1. SAMHD1-CtD выделен желтым цветом. Сшивки с N-концевыми глобулярными доменами SAM и HD SAMHD1 окрашены в светло-коричневый цвет, в то время как сшивки с N-концевой половиной SAMHD1-CtD выделены розовым цветом, а сшивки с С-концом SAMHD1-CtD отмечены окрашены в фиолетовый цвет.( B ) Поперечные связи Sulfo-SDA из A , в той же цветовой схеме, картированные на молекулярной модели CRL4-NEDD8 DCAF1-CtD /Vpr mus /SAMHD1 (состояние-2), получено из крио-ЭМ анализа (рис. 3). Плотность SAMHD1-CtD из крио-ЭМ-анализа CRL4-NEDD8 DCAF1-CtD /Vpr mus /SAMHD1 (основной) (рис. 4) показана желтой сеткой. ( C ) Доступное пространство взаимодействия SAMHD1-CtD, рассчитанное сервером DisVis [61], согласующееся как минимум с 14 из 26 наблюдаемых перекрестных связей, визуализируется серой сеткой.DCAF1-CtD и Vpr mus ориентированы и окрашены, как на рис.
Белки имеют цветовую кодировку, как на рис. 3, 4, и черный/белый SAMHD1. SAMHD1-CtD выделен желтым цветом. Сшивки с N-концевыми глобулярными доменами SAM и HD SAMHD1 окрашены в светло-коричневый цвет, в то время как сшивки с N-концевой половиной SAMHD1-CtD выделены розовым цветом, а сшивки с С-концом SAMHD1-CtD отмечены окрашены в фиолетовый цвет.( B ) Поперечные связи Sulfo-SDA из A , в той же цветовой схеме, картированные на молекулярной модели CRL4-NEDD8 DCAF1-CtD /Vpr mus /SAMHD1 (состояние-2), получено из крио-ЭМ анализа (рис. 3). Плотность SAMHD1-CtD из крио-ЭМ-анализа CRL4-NEDD8 DCAF1-CtD /Vpr mus /SAMHD1 (основной) (рис. 4) показана желтой сеткой. ( C ) Доступное пространство взаимодействия SAMHD1-CtD, рассчитанное сервером DisVis [61], согласующееся как минимум с 14 из 26 наблюдаемых перекрестных связей, визуализируется серой сеткой.DCAF1-CtD и Vpr mus ориентированы и окрашены, как на рис. 4А.
Для более количественной оценки информации о расстоянии, присущей перекрестным ссылкам SAMHD1-CtD, объем, доступный для SAMHD1-CtD для взаимодействия с CRL4 DCAF1-CtD /Vpr mus , в соответствии с расстоянием CLMS ограничений моделировали с помощью программного инструмента DisVis [60, 61]. Для этого анализа SAMHD1-CtD моделировали как пептид в расширенной конформации. Во время моделирования модель молекулярного состояния CRL4 DCAF1-CtD /Vpr mus сохранялась фиксированной, а шестимерный поиск всех возможных степеней свободы вращения и трансляции для модели SAMHD1-CtD в молекулярном контакте с CRL4 DCAF1-CtD /Vpr mus был рассчитан и ранжирован в соответствии с ограничениями расстояния CLMS.Чтобы визуализировать выходные данные, все возможные пространственные положения центра масс SAMHD1-CtD, которые удовлетворяют> 50% ограничений CLMS, были нанесены в виде карты плотности на структуру DCAF1-CtD/Vpr mus (рис. 5C). . В соответствии с крио-ЭМ-реконструкцией этот независимый вычислительный анализ также определяет местонахождение SAMHD1-CtD на вершине пучка спирали Vpr mus .
5C). . В соответствии с крио-ЭМ-реконструкцией этот независимый вычислительный анализ также определяет местонахождение SAMHD1-CtD на вершине пучка спирали Vpr mus .
Взятые вместе, структурные, биохимические данные и данные CLMS согласуются с моделью, в которой сам С-конец SAMHD1 рекрутируется Vpr mus , чтобы разместить оставшиеся домены SAMHD1 надлежащим образом для доступа к каталитическому механизму на дистальном конце стебля CRL4.
Эти данные позволяют провести структурное сравнение со способами связывания субстрата neo белков Vpx и Vpr из разных линий ретровирусов (рис. 6A-D). Vpx HIV-2 и Vpx sm позиционируют SAMHD1-CtD сбоку от домена DCAF1 BP посредством взаимодействия с N-концами Vpx Helices-1 и -3 (рис. 6B) [50]. Vpx mnd2 и Vpx rcm связывают SAMHD1-NtD, используя двусторонний интерфейс, включающий сторону DCAF1 BP и верхнюю поверхность пучка спирали Vpx (рис. 6C) [51, 52].Vpr HIV-1 взаимодействует со своим субстратом убиквитилирования UNG2, используя как верхнюю часть, так и верхний край пучка спирали Vpr HIV-1 (Fig.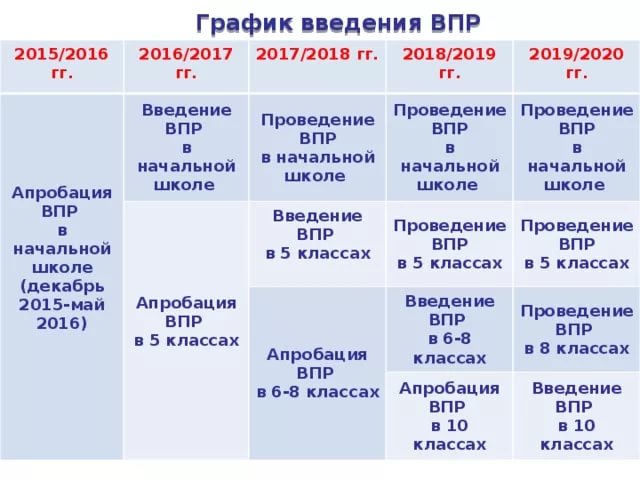 6D) [54]. Следует отметить, что эти интерфейсы взаимодействия верхней поверхности лишь частично перекрываются с интерфейсом связывания Vpr mus /SAMHD1-CtD, идентифицированным здесь, и используют принципиально разные наборы взаимодействующих аминокислотных остатков. Таким образом, оказывается, что интерфейсы молекулярного взаимодействия, управляющие Vpx/Vpr-опосредованным распознаванием и деградацией субстрата neo , не сохраняются между родственными вспомогательными белками SIV и Vpx/Vpr ВИЧ, даже в тех случаях, когда идентичные области SAMHD1-CtD нацелены на рекрутирование. .
6D) [54]. Следует отметить, что эти интерфейсы взаимодействия верхней поверхности лишь частично перекрываются с интерфейсом связывания Vpr mus /SAMHD1-CtD, идентифицированным здесь, и используют принципиально разные наборы взаимодействующих аминокислотных остатков. Таким образом, оказывается, что интерфейсы молекулярного взаимодействия, управляющие Vpx/Vpr-опосредованным распознаванием и деградацией субстрата neo , не сохраняются между родственными вспомогательными белками SIV и Vpx/Vpr ВИЧ, даже в тех случаях, когда идентичные области SAMHD1-CtD нацелены на рекрутирование. .
Сравнение Neo Моды распознавания Neo VPR MUS ( A ), VPX SM ( B ), VPX MND2 ( C ) и VPR ВИЧ-1 ( D ). DCAF1-CtD показан серым мультяшным цветом и полупрозрачной поверхностью, Vpr mus – зеленым, Vpx sm – оранжевым, Vpx mnd2 – синим и Vpr HIV-1 – светло-коричневым показаны мультяшными. Модели рекрутированных субстратов убиквитилирования показаны в виде сильно отфильтрованных, полупрозрачных расчетных карт электронной плотности со следующей схемой окраски: SAMHD1-CtD, связанный с Vpr mus – желтый, SAMHD1-CtD (связанный с Vpx sm , PDB 4cc9 ) [50] – мятно-зеленый, SAMHD1-NtD (Vpx mnd2 , PDB 5aja) [51] – пурпурный, UNG2 (Vpr HIV-1 , PDB 5jk7) [54] – светло-фиолетовый.
Модели рекрутированных субстратов убиквитилирования показаны в виде сильно отфильтрованных, полупрозрачных расчетных карт электронной плотности со следующей схемой окраски: SAMHD1-CtD, связанный с Vpr mus – желтый, SAMHD1-CtD (связанный с Vpx sm , PDB 4cc9 ) [50] – мятно-зеленый, SAMHD1-NtD (Vpx mnd2 , PDB 5aja) [51] – пурпурный, UNG2 (Vpr HIV-1 , PDB 5jk7) [54] – светло-фиолетовый.
Обсуждение
Наши рентгеноструктурные исследования сборки DDB1/DCAF1-CtD/Vpr mus обеспечивают первое понимание структуры класса «гибридных» белков Vpr SIV.Они присутствуют в линиях лентивирусов SIV agm и SIV mus/deb/syk и сочетают характеристики родственных вспомогательных белков Vpr HIV-1 и SIV Vpx.
Как и Vpx SIV, «гибридные» белки Vpr подавляют рестрикционный фактор хозяина SAMHD1, привлекая его к CRL4 DCAF1 для убиквитинирования и последующей протеасомной деградации. Однако, используя комбинацию рентгеновского, крио-ЭМ и CLMS анализов, мы показываем, что молекулярная стратегия, которую Vpr mus развила для нацеливания на SAMHD1, разительно отличается от Vpx-содержащих штаммов SIV.В двух кладах белков Vpx расходящаяся аминокислотная последовательность простирается непосредственно перед Helix-1 (вариабельная область (VR)1, S6A Fig) вместе с полиморфизмами в SAMHD1-N-конце соответствующего вида-хозяина определяют, является ли ВИЧ -2-тип или SIV mnd -тип Vpx распознают SAMHD1-CtD или SAMHD1-NtD соответственно. Эти механизмы распознавания приводят к позиционированию SAMHD1-CtD или -NtD на стороне домена DCAF1 BP таким образом, что обеспечиваются дополнительные контакты между SAMHD1 и DCAF1, образуя тройные сборки Vpx/SAMHD1/DCAF1 с очень низкой скоростью диссоциации [50]. –52, 62].В Vpr mus разные принципы определяют специфичность SAMHD1-CtD. Здесь VR1 вообще не участвует в связывании SAMHD1-CtD, но формирует дополнительные взаимодействия с DCAF1, которые не наблюдаются в белковых комплексах Vpx/DCAF1 (S6A Fig).
Однако, используя комбинацию рентгеновского, крио-ЭМ и CLMS анализов, мы показываем, что молекулярная стратегия, которую Vpr mus развила для нацеливания на SAMHD1, разительно отличается от Vpx-содержащих штаммов SIV.В двух кладах белков Vpx расходящаяся аминокислотная последовательность простирается непосредственно перед Helix-1 (вариабельная область (VR)1, S6A Fig) вместе с полиморфизмами в SAMHD1-N-конце соответствующего вида-хозяина определяют, является ли ВИЧ -2-тип или SIV mnd -тип Vpx распознают SAMHD1-CtD или SAMHD1-NtD соответственно. Эти механизмы распознавания приводят к позиционированию SAMHD1-CtD или -NtD на стороне домена DCAF1 BP таким образом, что обеспечиваются дополнительные контакты между SAMHD1 и DCAF1, образуя тройные сборки Vpx/SAMHD1/DCAF1 с очень низкой скоростью диссоциации [50]. –52, 62].В Vpr mus разные принципы определяют специфичность SAMHD1-CtD. Здесь VR1 вообще не участвует в связывании SAMHD1-CtD, но формирует дополнительные взаимодействия с DCAF1, которые не наблюдаются в белковых комплексах Vpx/DCAF1 (S6A Fig). Молекулярные контакты между Vpr mus и SAMHD1 рассредоточены по спиралям-1 и -3, обращены в сторону от места взаимодействия DCAF1 и иммобилизуют SAMHD1-CtD на верхней стороне пучка спирали Vpr mus (рис. S6A). Помещение SAMHD1-CtD в такое положение препятствует стабилизации тройного взаимодействия с DCAF1-CtD, но все же приводит к надежному убиквитилированию SAMHD1 in vitro и деградации SAMHD1 в клеточных анализах [24].
Молекулярные контакты между Vpr mus и SAMHD1 рассредоточены по спиралям-1 и -3, обращены в сторону от места взаимодействия DCAF1 и иммобилизуют SAMHD1-CtD на верхней стороне пучка спирали Vpr mus (рис. S6A). Помещение SAMHD1-CtD в такое положение препятствует стабилизации тройного взаимодействия с DCAF1-CtD, но все же приводит к надежному убиквитилированию SAMHD1 in vitro и деградации SAMHD1 в клеточных анализах [24].
Каталитическая дНТФазная активность SAMHD1 зависит от нуклеотид-зависимой олигомеризации, опосредованной двумя аллостерическими сайтами связывания нуклеотидов, где нуклеотиды на основе гуанина в первом сайте индуцируют образование димера, а связывание дНТФ со вторым сайтом приводит к сборке каталитически активный тетрамер. SAMHD1-CtD необходим для образования тетрамера, обеспечивая критические молекулярные контакты с соседними протомерами. Более того, дестабилизация тетрамера посредством CDK1/2-циклинА-зависимого фосфорилирования T592 в SAMHD1-CtD эндогенно ослабляет активность SAMHD1 в циклирующих клетках [46, 63-65]. Соответственно, возможно, что, изолируя SAMHD1-CtD, Vpr mus дестабилизирует тетрамер SAMHD1 и, таким образом, аннулирует активность SAMHD1, прежде чем вызвать его протеасомную деградацию, в соответствии с предыдущим наблюдением Vpx , опосредованного ВИЧ-2 . Разборка тетрамера SAMHD1 и ингибирование активности дНТФазы [62].
Соответственно, возможно, что, изолируя SAMHD1-CtD, Vpr mus дестабилизирует тетрамер SAMHD1 и, таким образом, аннулирует активность SAMHD1, прежде чем вызвать его протеасомную деградацию, в соответствии с предыдущим наблюдением Vpx , опосредованного ВИЧ-2 . Разборка тетрамера SAMHD1 и ингибирование активности дНТФазы [62].
Предсказания относительно молекулярного механизма связывания SAMHD1 другими «гибридными» ортологами Vpr затруднены из-за расхождения последовательностей.Даже в Vpr deb , наиболее близком по отношению к Vpr mus , сохраняется только приблизительно 50% боковых цепей аминокислот, выстилающих предполагаемый карман связывания SAMHD1-CtD (S6A Fig). Предыдущие эксперименты по убиквитинированию in vitro и клеточной деградации не показали явного предпочтения Vpr deb для рекрутирования либо SAMHD1-NtD, либо –CtD [24, 49]. Кроме того, спорным является вопрос о том, действительно ли Vpr deb связывает DCAF1 [66], что, возможно, можно объяснить аминокислотными вариациями на самом N-конце и/или в Helix-3 (S6A Fig).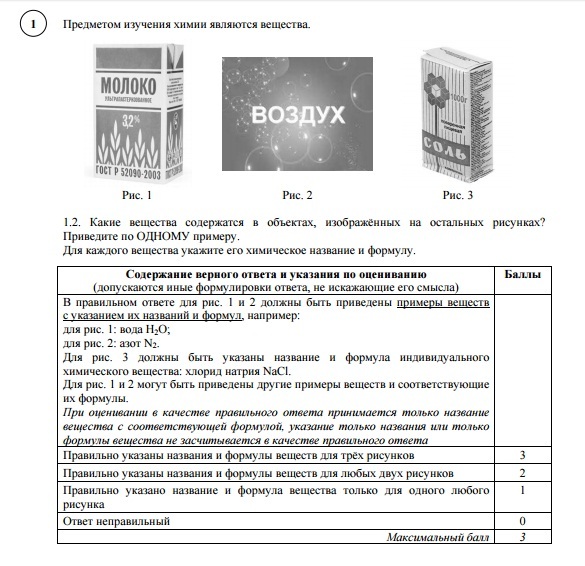 Vpr syk специфичен для SAMHD1-CtD [49], но большинство остатков, образующих платформу связывания SAMHD1-CtD, наблюдаемых в настоящем исследовании, не являются консервативными. Линия белков Vpr SIV agm еще более дивергентна, со значительными различиями не только в возможных остатках, контактирующих с SAMHD1, но и в участках последовательности, предшествующих Helix-1 и соединяющих спирали-2 и -3, а также в N-концевая половина Helix-3 (фиг. S6A). Кроме того, есть указания на то, что рекрутирование SAMHD1 с помощью Vpr agm.Подтип GRI включает молекулярное распознавание как SAMHD1-NtD, так и –CtD [49, 53]. В заключение, повторяющиеся раунды эволюционной адаптации лентивирусов к фактору рестрикции SAMHD1 хозяина с последующей повторной адаптацией хозяина привели к высоковидоспецифическим, разнообразным молекулярным способам взаимодействия Vpr-SAMHD1. Аналогичная гонка молекулярных вооружений между внутренними клеточными противовирусными факторами-хозяевами и вирусными антагонистами сформировала видоспецифический лентивирусный антагонизм, например.
Vpr syk специфичен для SAMHD1-CtD [49], но большинство остатков, образующих платформу связывания SAMHD1-CtD, наблюдаемых в настоящем исследовании, не являются консервативными. Линия белков Vpr SIV agm еще более дивергентна, со значительными различиями не только в возможных остатках, контактирующих с SAMHD1, но и в участках последовательности, предшествующих Helix-1 и соединяющих спирали-2 и -3, а также в N-концевая половина Helix-3 (фиг. S6A). Кроме того, есть указания на то, что рекрутирование SAMHD1 с помощью Vpr agm.Подтип GRI включает молекулярное распознавание как SAMHD1-NtD, так и –CtD [49, 53]. В заключение, повторяющиеся раунды эволюционной адаптации лентивирусов к фактору рестрикции SAMHD1 хозяина с последующей повторной адаптацией хозяина привели к высоковидоспецифическим, разнообразным молекулярным способам взаимодействия Vpr-SAMHD1. Аналогичная гонка молекулярных вооружений между внутренними клеточными противовирусными факторами-хозяевами и вирусными антагонистами сформировала видоспецифический лентивирусный антагонизм, например.
Предыдущее исследование структуры DDB1/DCAF1/Vpr HIV-1 в комплексе с neo -субстратом UNG2 показало, что Vpr HIV-1 взаимодействует с UNG2, имитируя фосфатный остов ДНК.Точнее, остатки UNG2, выступающие в большую бороздку его эндогенного ДНК-субстрата, встраиваются в гидрофобную щель, образованную Vpr HIV-1 Helices-1, -2 и N-концевой половиной Helix-3 [54]. . Этот механизм может объяснить необычайную неразборчивость связывания Vpr HIV-1 , поскольку список потенциальных субстратов деградации Vpr HIV-1 значительно обогащен ДНК- и РНК-связывающими белками [27]. Более того, было высказано предположение, что неразборчивая Vpr , индуцированная ВИЧ-1 , деградация факторов хозяина с ДНК- или РНК-связывающей активностью вызывает остановку клеточного цикла на границе фазы G2/M, что является наиболее подробно описанным фенотипом белков Vpr. далеко [26, 27, 70].В Vpr mus N-концевая половина Helix-1, а также объемный аминокислотный остаток W48, который также сохраняется в Vpr agm и Vpx, сужают гидрофобную щель (S6A, B Fig). Кроме того, удлиненный N-конец Vpr mus Helix-3 несовместим со связыванием UNG2 из-за стерического исключения (рис. S6C). В соответствии с этими наблюдениями, Vpr mus не подавляет UNG2 в линии Т-клеток человека [27]. Однако Vpr mus , Vpr syk и Vpr agm также вызывают остановку клеточного цикла G2/M в соответствующих клетках-хозяевах [66, 71, 72].
. Этот механизм может объяснить необычайную неразборчивость связывания Vpr HIV-1 , поскольку список потенциальных субстратов деградации Vpr HIV-1 значительно обогащен ДНК- и РНК-связывающими белками [27]. Более того, было высказано предположение, что неразборчивая Vpr , индуцированная ВИЧ-1 , деградация факторов хозяина с ДНК- или РНК-связывающей активностью вызывает остановку клеточного цикла на границе фазы G2/M, что является наиболее подробно описанным фенотипом белков Vpr. далеко [26, 27, 70].В Vpr mus N-концевая половина Helix-1, а также объемный аминокислотный остаток W48, который также сохраняется в Vpr agm и Vpx, сужают гидрофобную щель (S6A, B Fig). Кроме того, удлиненный N-конец Vpr mus Helix-3 несовместим со связыванием UNG2 из-за стерического исключения (рис. S6C). В соответствии с этими наблюдениями, Vpr mus не подавляет UNG2 в линии Т-клеток человека [27]. Однако Vpr mus , Vpr syk и Vpr agm также вызывают остановку клеточного цикла G2/M в соответствующих клетках-хозяевах [66, 71, 72]. Это убедительно указывает на существование дополнительных структурных детерминант у Vpr mus , Vpr syk , Vpr agm и потенциально Vpr HIV-1 , которые регулируют рекрутирование и убиквитилирование ДНК/РНК-связывающих факторов хозяина, кроме того к гидрофобной, имитирующей ДНК щели на вершине пучка из трех спиралей. Будущие усилия по структурной характеристике этих детерминант еще больше расширят наше понимание того, как спиральный каркас Vpx/Vpr связывается и, таким образом, адаптируется к множеству neo -субстратных эпитопов.
Это убедительно указывает на существование дополнительных структурных детерминант у Vpr mus , Vpr syk , Vpr agm и потенциально Vpr HIV-1 , которые регулируют рекрутирование и убиквитилирование ДНК/РНК-связывающих факторов хозяина, кроме того к гидрофобной, имитирующей ДНК щели на вершине пучка из трех спиралей. Будущие усилия по структурной характеристике этих детерминант еще больше расширят наше понимание того, как спиральный каркас Vpx/Vpr связывается и, таким образом, адаптируется к множеству neo -субстратных эпитопов.
Наши крио-ЭМ реконструкции CRL4 DCAF1-CtD /Vpr mus /SAMHD1, дополненные CLMS, также дают представление о структурной динамике сборок CRL4 до переноса убиквитина. Данные подтверждают ранее описанное вращательное движение ножки CRL4 в отсутствие ограничений, налагаемых кристаллической решеткой, создавая зону убиквитилирования вокруг субстратного рецептора, модифицированного Vpr mus (рис. 3 и 7А) [13, 15, 16, 19, 57]. Отсутствие плотности для недилированного домена CUL4 WHB и для каталитического домена ROC1 RING указывает на то, что эти дистальные элементы стебля очень подвижны и, вероятно, имеют множество ориентаций относительно каркаса CUL4 (рис. 7B).Эти наблюдения согласуются со структурным анализом CRL1 и CRL5, где недилирование CUL1/5 приводит к переориентации домена WHB кулина и высвобождению домена ROC1 RING из каркаса куллина одновременно со стимуляцией активности убиквитилирования [56]. ]. Более того, недавний крио-ЭМ анализ структуры CRL1 β-TRCP /IκBα продемонстрировал значительную подвижность прекаталитических доменов CUL1-NEDD8 WHB и ROC1 RING [73]. Такая гибкость, по-видимому, необходима для структурной организации множественных CRL1-зависимых процессов, в частности, зародышеобразования каталитической сборки, включающей сложные белок-белковые взаимодействия между NEDD8, CUL1, заряженным убиквитином E2 и субстратным рецептором.Эта синергетическая сборка затем направляет С-конец убиквитина к субстрату лизина для праймирования убиквитином [73].
3 и 7А) [13, 15, 16, 19, 57]. Отсутствие плотности для недилированного домена CUL4 WHB и для каталитического домена ROC1 RING указывает на то, что эти дистальные элементы стебля очень подвижны и, вероятно, имеют множество ориентаций относительно каркаса CUL4 (рис. 7B).Эти наблюдения согласуются со структурным анализом CRL1 и CRL5, где недилирование CUL1/5 приводит к переориентации домена WHB кулина и высвобождению домена ROC1 RING из каркаса куллина одновременно со стимуляцией активности убиквитилирования [56]. ]. Более того, недавний крио-ЭМ анализ структуры CRL1 β-TRCP /IκBα продемонстрировал значительную подвижность прекаталитических доменов CUL1-NEDD8 WHB и ROC1 RING [73]. Такая гибкость, по-видимому, необходима для структурной организации множественных CRL1-зависимых процессов, в частности, зародышеобразования каталитической сборки, включающей сложные белок-белковые взаимодействия между NEDD8, CUL1, заряженным убиквитином E2 и субстратным рецептором.Эта синергетическая сборка затем направляет С-конец убиквитина к субстрату лизина для праймирования убиквитином [73]. Соответственно, наши крио-ЭМ исследования могут указывать на то, что аналогичные принципы применимы к катализируемому CRL4 убиквитилированию. Однако, чтобы разгадать каталитическую архитектуру CRL4, должны быть проведены сложные процедуры перекрестного связывания, как в ссылке (73).
Соответственно, наши крио-ЭМ исследования могут указывать на то, что аналогичные принципы применимы к катализируемому CRL4 убиквитилированию. Однако, чтобы разгадать каталитическую архитектуру CRL4, должны быть проведены сложные процедуры перекрестного связывания, как в ссылке (73).
( A ) Вращение ножки CRL4 увеличивает пространство, доступное для каталитических элементов на дистальном конце ножки, образуя зону убиквитилирования вокруг ядра. ( B ) Модификация CUL4-WHB с помощью NEDD8 приводит к увеличению подвижности этих дистальных элементов стебля (CUL4-WHB, домен ROC1 RING) [56], дополнительно расширяя зону убиквитилирования и активируя формирование каталитической сборки для переноса убиквитина (см. также D ) [73]. ( C ) Гибкая привязка SAMHD1 к ядру с помощью Vpr mus помещает большую часть SAMHD1 в зону убиквитинирования и оптимизирует доступность поверхности. ( D ) Динамические процессы A — C вместе создают многочисленные возможности для сборки каталитического механизма (CUL4-NEDD8 WHB, ROC1, убиквитин-(уби-)заряженный E2) на открытых боковых цепях лизина SAMHD1. Здесь три из этих возможностей показаны схематически. Таким образом, покрытие убиквитином на SAMHD1 максимизируется.
( D ) Динамические процессы A — C вместе создают многочисленные возможности для сборки каталитического механизма (CUL4-NEDD8 WHB, ROC1, убиквитин-(уби-)заряженный E2) на открытых боковых цепях лизина SAMHD1. Здесь три из этих возможностей показаны схематически. Таким образом, покрытие убиквитином на SAMHD1 максимизируется.
Внутренняя подвижность элементов стебля CRL4 может способствовать размещению субстратов различных размеров и форм в зоне убиквитилирования CRL4 и может рационализировать широкий диапазон субстратов, доступных для убиквитилирования CRL4 через множественные рецепторы DCAF.Из-за селективного давления, направленного на противодействие ограничению SAMHD1 хозяина, ВИЧ-2 и некоторые SIV, среди других вирусов, воспользовались этой динамической архитектурой CRL4 путем модификации субстратного рецептора DCAF1 вспомогательными белками семейства Vpx/Vpr. Путем связывания либо SAMHD1-CtD, либо -NtD с DCAF1 и, таким образом, гибкого рекрутирования основной массы SAMHD1 доступность боковых цепей лизина как проксимальных, так и на глобулярных доменах SAMHD1 к каталитической сборке CRL4 может быть дополнительно улучшена (рис. 7С, Г).Это обеспечивает эффективное Vpx/Vpr-опосредованное праймирование SAMHD1, полиубиквитилирование и протеасомную деградацию для стимуляции репликации вируса. Кроме того, структурное понимание этого эволюционно оптимизированного, высокоспецифичного механизма деградации белка может дать информацию о позиционировании новых синтетических методов на основе CRL4 DCAF1 для направленной деградации белка, например. в виде химерных (PROTAC-) или соединений типа молекулярного клея, нацеленных на протеолиз [74, 75].
7С, Г).Это обеспечивает эффективное Vpx/Vpr-опосредованное праймирование SAMHD1, полиубиквитилирование и протеасомную деградацию для стимуляции репликации вируса. Кроме того, структурное понимание этого эволюционно оптимизированного, высокоспецифичного механизма деградации белка может дать информацию о позиционировании новых синтетических методов на основе CRL4 DCAF1 для направленной деградации белка, например. в виде химерных (PROTAC-) или соединений типа молекулярного клея, нацеленных на протеолиз [74, 75].
Наконец, хотя современные схемы высокоактивной антиретровирусной терапии (ВААРТ) способны контролировать репликацию ВИЧ-1 у инфицированных пациентов [76], они не могут уничтожить вирус из-за рецидива вируса после прекращения лечения и приводят к появлению резистентного вируса варианты [77].Соответственно, идентификация и ингибирование новых мишеней в дополнение к тем, которые уже охвачены ВААРТ, представляют большой интерес. В этом контексте вспомогательные белки ВИЧ долгое время рассматривались как перспективные мишени для лекарственных средств [78, 79]. Например, доклинические исследования, направленные на ингибирование Vif-опосредованной деградации APOBEC3, завершились выделением нескольких соединений, препятствующих репликации ВИЧ-1 [80–82]. Из-за беспорядочных взаимодействий с хозяином Vpr HIV-1 также считается привлекательной мишенью для антиретровирусных препаратов [83].Представленный здесь сравнительный структурный анализ взаимодействия Vpr/DCAF1-CtD может послужить основой для будущих усилий по разрушению этого интерфейса взаимодействия с малыми молекулами, чтобы сразу отменить все процессы деградации белка-хозяина, индуцированные Vpr, и максимизировать ингибирующий эффект на репликацию вируса.
Например, доклинические исследования, направленные на ингибирование Vif-опосредованной деградации APOBEC3, завершились выделением нескольких соединений, препятствующих репликации ВИЧ-1 [80–82]. Из-за беспорядочных взаимодействий с хозяином Vpr HIV-1 также считается привлекательной мишенью для антиретровирусных препаратов [83].Представленный здесь сравнительный структурный анализ взаимодействия Vpr/DCAF1-CtD может послужить основой для будущих усилий по разрушению этого интерфейса взаимодействия с малыми молекулами, чтобы сразу отменить все процессы деградации белка-хозяина, индуцированные Vpr, и максимизировать ингибирующий эффект на репликацию вируса.
Методы
Экспрессия и очистка белков
Конструкции амплифицировали с помощью ПЦР из матриц кДНК и встраивали в указанные экспрессионные плазмиды с использованием стандартных методов рестрикционных ферментов (таблица S2).pAcGHLT-B-DDB1 (плазмида № 48638) и pET28-UBA1 (плазмида № 32534) были получены от Addgene. Плазмида коэкспрессии pOPC-UBA3-GST-APPBP1 и плазмида pGex6P2-UBC12 были получены от MRC-PPU Reagents and Services (клоны 32498, 3879). Убиквитин бычьих эритроцитов и рекомбинантный hsNEDD8 были приобретены у Sigma-Aldrich (U6253) и BostonBiochem (UL-812) соответственно. Точечные мутации вводили путем сайт-направленного мутагенеза с использованием KOD-полимеразы (Novagen). Все конструкции и варианты суммированы в таблице S3.
Плазмида коэкспрессии pOPC-UBA3-GST-APPBP1 и плазмида pGex6P2-UBC12 были получены от MRC-PPU Reagents and Services (клоны 32498, 3879). Убиквитин бычьих эритроцитов и рекомбинантный hsNEDD8 были приобретены у Sigma-Aldrich (U6253) и BostonBiochem (UL-812) соответственно. Точечные мутации вводили путем сайт-направленного мутагенеза с использованием KOD-полимеразы (Novagen). Все конструкции и варианты суммированы в таблице S3.
Белки, экспрессированные из векторов pAcGHLT-B, pGex6P1/2, pOPC и pET49b, содержали N-концевую GST-His-метку; pHisSUMO – N-концевая His-SUMO-метка; pET28, pRSF-Duet-1 – N-концевая His-метка; pTri-Ex-6 – С-концевая His-метка. Конструкции в векторах pAcGHLT-B и pTri-Ex-6 экспрессировали в клетках Sf9, а конструкции в векторах pET28, pET49b, pGex6P1/2, pRSF-Duet-1 и pHisSUMO — в E. coli Rosetta 2(DE3).
Рекомбинантные бакуловирусы ( Autographa californica nucleopolyhedrovirus клон C6) были получены, как описано ранее [84].Клетки Sf9 культивировали в среде Insect-XPRESS (Lonza) при 28°С в шейкере-инкубаторе Innova 42R (New Brunswick) при скорости встряхивания 180 об/мин. В типичном препарате 1 л клеток Sf9 при 3×10 6 клеток/мл коинфицировали 4 мл вируса DDB1 с высоким титром и 4 мл вируса DCAF1-CtD с высоким титром в течение 72 часов.
В типичном препарате 1 л клеток Sf9 при 3×10 6 клеток/мл коинфицировали 4 мл вируса DDB1 с высоким титром и 4 мл вируса DCAF1-CtD с высоким титром в течение 72 часов.
Для типичной экспрессии E. coli Rosetta 2 (DE3) 2 л среды LB инокулировали 20 мл ночной культуры и выращивали в шейкере-инкубаторе Multitron HT (Infors) при 37°C, 150 об/мин до OD 600 достиг 0.7. В этот момент температуру снижали до 18°C, экспрессию белка индуцировали добавлением 0,2 мМ IPTG и культуры выращивали еще 20 часов. Во время совместной экспрессии CUL4 и ROC1 из pRSF-Duet перед индукцией в среду для выращивания добавляли 50 мкМ сульфата цинка. Клетки
Sf9 осаждали центрифугированием при 1000 об/мин, 4°С в течение 30 мин с использованием центрифужного ротора JLA 9.1000 (Beckman). Клетки E. coli осаждали центрифугированием при 4000 об/мин, 4°С в течение 15 мин с использованием того же ротора.Осадок клеток ресуспендировали в буфере, содержащем 50 мМ трис, рН 7,8, 500 мМ NaCl, 4 мМ MgCl2, 0,5 мМ трис-(2-карбоксиэтил)фосфин (TCEP), мини-полные ингибиторы протеазы (1 таблетка на 50 мл) и 20 мМ имидазол (только для His-меченых белков). 100 мл лизирующего буфера использовали для ресуспендирования осадка из 1 л культуры Sf9 и 35 мл лизирующего буфера на осадок из 1 л культуры E. coli . Перед ресуспендированием осадков коэкспрессии CUL4/ROC1 рН буфера доводили до 8,5. Добавляли 5 мкл бензоназы (Merck) и клетки лизировали, пропуская суспензию не менее двух раз через Microfluidizer (Microfluidics).Лизаты осветляли центрифугированием при 48000 g в течение 45 мин при 4°С. Очистку белков проводили при 4°C на Äkta pure FPLC (GE) с использованием хроматографических колонок XK 16/20 (GE), содержащих 10 мл соответствующей аффинной смолы. GST-меченые белки были захвачены на глутатион-сефарозе (GSH-Sepharose FF, GE), промыты 250 мл промывочного буфера (50 мМ Tris-HCl, pH 7,8, 500 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ TCEP). и элюировали тем же буфером с добавлением 20 мМ восстановленного глутатиона.Белки, меченные His, иммобилизовали на Ni-Sepharose HP (GE), промывали 250 мл промывочного буфера с добавлением 20 мМ имидазола и элюировали промывочным буфером, содержащим 0,3 М имидазол.
100 мл лизирующего буфера использовали для ресуспендирования осадка из 1 л культуры Sf9 и 35 мл лизирующего буфера на осадок из 1 л культуры E. coli . Перед ресуспендированием осадков коэкспрессии CUL4/ROC1 рН буфера доводили до 8,5. Добавляли 5 мкл бензоназы (Merck) и клетки лизировали, пропуская суспензию не менее двух раз через Microfluidizer (Microfluidics).Лизаты осветляли центрифугированием при 48000 g в течение 45 мин при 4°С. Очистку белков проводили при 4°C на Äkta pure FPLC (GE) с использованием хроматографических колонок XK 16/20 (GE), содержащих 10 мл соответствующей аффинной смолы. GST-меченые белки были захвачены на глутатион-сефарозе (GSH-Sepharose FF, GE), промыты 250 мл промывочного буфера (50 мМ Tris-HCl, pH 7,8, 500 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ TCEP). и элюировали тем же буфером с добавлением 20 мМ восстановленного глутатиона.Белки, меченные His, иммобилизовали на Ni-Sepharose HP (GE), промывали 250 мл промывочного буфера с добавлением 20 мМ имидазола и элюировали промывочным буфером, содержащим 0,3 М имидазол. Фракции элюента анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и доводили до 5 мл с помощью центрифужных фильтрующих устройств (Vivaspin). Если применимо, добавляли 100 мкг протеазы GST-3C или 50 мкг тромбина на мг общего белка, и образец инкубировали в течение 12 часов на льду для отщепления аффинных меток. В качестве второго этапа очистки выполняли гель-фильтрационную хроматографию (ГФ) на Äkta prime plus FPLC (GE) с колонками Superdex 200 16/600 (GE), уравновешенными в 10 мМ Tris-HCl, pH 7.8, 150 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ буфер TCEP, скорость потока 1 мл/мин. Для очистки комплекса CUL4/ROC1 рН всех буферов для очистки доводили до 8,5. Пиковые фракции анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и концентрировали до прибл. 20 мг/мл, мгновенно замороженные в жидком азоте небольшими аликвотами и хранящиеся при температуре -80°C. Концентрации белков определяли с помощью спектрофотометра NanoDrop (ND 1000, Peqlab) с использованием теоретических коэффициентов поглощения, рассчитанных на основе аминокислотной последовательности с помощью ProtParam на веб-сервере ExPASy [85].
Фракции элюента анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и доводили до 5 мл с помощью центрифужных фильтрующих устройств (Vivaspin). Если применимо, добавляли 100 мкг протеазы GST-3C или 50 мкг тромбина на мг общего белка, и образец инкубировали в течение 12 часов на льду для отщепления аффинных меток. В качестве второго этапа очистки выполняли гель-фильтрационную хроматографию (ГФ) на Äkta prime plus FPLC (GE) с колонками Superdex 200 16/600 (GE), уравновешенными в 10 мМ Tris-HCl, pH 7.8, 150 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ буфер TCEP, скорость потока 1 мл/мин. Для очистки комплекса CUL4/ROC1 рН всех буферов для очистки доводили до 8,5. Пиковые фракции анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и концентрировали до прибл. 20 мг/мл, мгновенно замороженные в жидком азоте небольшими аликвотами и хранящиеся при температуре -80°C. Концентрации белков определяли с помощью спектрофотометра NanoDrop (ND 1000, Peqlab) с использованием теоретических коэффициентов поглощения, рассчитанных на основе аминокислотной последовательности с помощью ProtParam на веб-сервере ExPASy [85].
Аналитический гель-фильтрационный анализ
Перед гель-фильтрационным анализом аффинные метки удаляли путем инкубации 30 мкг протеазы GST-3C с 6 мкМ каждого белкового компонента в объеме 150 мкл промывочного буфера с последующей инкубацией на льду в течение 12 ч. . Чтобы удалить расщепленную GST-метку и протеазу GST-3C, добавляли 20 мкл гранул GSH-Sepharose FF (GE) и образец вращали при 4 °C в течение одного часа. Гранулы GSH-сефарозы удаляли центрифугированием при 4°C, 3500 об/мин в течение 5 мин и 120 мкл супернатанта загружали на аналитическую колонку GF (Superdex 200 10/300 GL, GE), уравновешенную в 10 мМ Tris-HCl. рН 7.8, 150 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ TCEP, скорость потока 0,5 мл/мин. Фракции объемом 1 мл собирали и анализировали с помощью SDS-PAGE. Для следующих контрольных образцов 120 мкл очищенного белка наносили непосредственно на колонку GF, потому что не нужно было удалять метку очистки путем расщепления: SAMHD1, T4L-SAMHD1-CtD, SAMHD1-ΔCtD. Для этих образцов концентрация была доведена до 18 мкМ, 37 мкМ и 30 мкМ, соответственно, для учета более низкого коэффициента экстинкции этих выделенных белковых компонентов, чтобы обеспечить лучшую визуализацию пика элюции. Колонку GF калибровали с использованием калибровочного набора для гель-фильтрации (GE) с высоким молекулярным весом (HMW) в соответствии с инструкциями производителя.
Колонку GF калибровали с использованием калибровочного набора для гель-фильтрации (GE) с высоким молекулярным весом (HMW) в соответствии с инструкциями производителя.
Анализы убиквитилирования in vitro
160 мкл реакционной смеси, содержащей 0,5 мкМ субстрата (указанные конструкции SAMHD1, S2 рис.), 0,125 мкМ DDB1/DCAF1-CtD, 0,125 мкМ pr-TUL4/SUV-SUV-ROC1, 0,12MOC1, 0,12MOC1 мус (остатки 1-92), 0,25 мкМ UBCH5C, 15 мкМ убиквитина в 20 мМ трис-HCl, рН 7,8, 150 мМ NaCl, 2.5 мМ MgCl 2 , 2,5 мМ АТФ. В контрольных реакциях некоторые компоненты были исключены, как показано на рис. S2. Был взят образец объемом 30 мкл для анализа SDS-PAGE (t=0). Реакции инициировали добавлением 0,05 мкМ UBA1, инкубировали при 37°C и через 1 мин, 2 мин, 5 мин и 15 мин отбирали 30 мкл образцов SDS-PAGE, сразу смешивали с 10 мкл буфера для образцов 4x SDS и кипятили при 95°С в течение 5 мин. Образцы анализировали с помощью SDS-PAGE.
In vitro Неддилирование CUL4/ROC1
Для начальных испытаний неддилирования была подготовлена 200 мкл реакционная смесь, содержащая 8 мкМ CUL4/ROC1, 1. 8 мкМ UBC12, 30 мкМ NEDD8 в 50 мМ Tris-HCl pH 7,8, 150 мМ NaCl, 2,5 мМ MgCl 2 , 2,5 мМ АТФ. Для SDS-PAGE отбирали 2 образца по 30 мкл, один немедленно смешивали с 10 мкл 4x буфера для образцов SDS, другой инкубировали в течение 60 мин при 25°C. Реакцию инициировали добавлением 0,7 мкМ APPBP1/UBA3, инкубировали при 25°C, и через 1 мин, 5 мин, 10 мин, 30 мин и 60 мин отбирали 30 мкл образцов SDS-PAGE, сразу же смешивали с 10 мкл 4x буфер для образца SDS и кипятят при 95°C в течение 5 мин. Образцы анализировали с помощью SDS-PAGE.На основании этого теста объем реакции увеличили до 1 мл и инкубировали в течение 5 мин при 25°C. Реакцию гасили добавлением 5 мМ TCEP и немедленно загружали в колонку Superdex 200 16/600 GF (GE), уравновешенную 10 мМ Tris-HCl, pH 7,8, 150 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ TCEP при скорость потока 1 мл/мин. Пиковые фракции анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и концентрировали до ~20 мг/мл, мгновенно замораживали в жидком азоте небольшими аликвотами и хранили при -80°C.
8 мкМ UBC12, 30 мкМ NEDD8 в 50 мМ Tris-HCl pH 7,8, 150 мМ NaCl, 2,5 мМ MgCl 2 , 2,5 мМ АТФ. Для SDS-PAGE отбирали 2 образца по 30 мкл, один немедленно смешивали с 10 мкл 4x буфера для образцов SDS, другой инкубировали в течение 60 мин при 25°C. Реакцию инициировали добавлением 0,7 мкМ APPBP1/UBA3, инкубировали при 25°C, и через 1 мин, 5 мин, 10 мин, 30 мин и 60 мин отбирали 30 мкл образцов SDS-PAGE, сразу же смешивали с 10 мкл 4x буфер для образца SDS и кипятят при 95°C в течение 5 мин. Образцы анализировали с помощью SDS-PAGE.На основании этого теста объем реакции увеличили до 1 мл и инкубировали в течение 5 мин при 25°C. Реакцию гасили добавлением 5 мМ TCEP и немедленно загружали в колонку Superdex 200 16/600 GF (GE), уравновешенную 10 мМ Tris-HCl, pH 7,8, 150 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ TCEP при скорость потока 1 мл/мин. Пиковые фракции анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и концентрировали до ~20 мг/мл, мгновенно замораживали в жидком азоте небольшими аликвотами и хранили при -80°C.
Подготовка образцов для рентгеновской кристаллографии, кристаллизация, сбор данных и структурное решение
Комплекс DDB1/DCAF1-CtD
Кристаллы DDB1/DCAF1-CtD выращивали методом диффузии паров висячей капли путем смешивания равных объемов (1 мкл) раствора DDB1/DCAF1-CtD в концентрации 10 мг/мл с резервуарным раствором, содержащим 100 мМ цитрата Tri-Na, рН 5.5, 18% ПЭГ 1000 и суспендируют над резервуаром объемом 500 мкл. Кристаллы росли в течение ночи при 18°С. Кристаллы подвергали криозащите в резервуарном растворе с добавлением 20% глицерина и криоохлаждению в жидком азоте. Набор данных с монокристалла был собран в Diamond Light Source (Didcot, Великобритания) при длине волны 0,
Å. Данные обрабатывали с помощью XDS [86] (S1 Table), а структуру расшифровывали методом молекулярной замены с помощью программы MOLREP [87] и доступных структур DDB1 (PDB 3e0c) и DCAF1-CtD (PDB 4cc9) [50] в качестве поиска. модели.Итерационные циклы корректировки модели с помощью программы Coot [88] с последующим уточнением с помощью программы PHENIX [89] дали окончательное соотношение R/R без факторов 22,0%/27,9% (таблица S1). В модели 94,5 % остатков имеют двугранные углы остова в предпочтительной области графика Рамачандрана, остальные попадают в разрешенные области, и ни один из них не является выбросом. Детали сбора данных и уточнения статистики представлены в таблице S1. Координаты и структурные факторы депонированы в PDB под инвентарным номером 6zue.
В модели 94,5 % остатков имеют двугранные углы остова в предпочтительной области графика Рамачандрана, остальные попадают в разрешенные области, и ни один из них не является выбросом. Детали сбора данных и уточнения статистики представлены в таблице S1. Координаты и структурные факторы депонированы в PDB под инвентарным номером 6zue. Комплекс DDB1/DCAF1-CtD/T4L-Vprmus (1-92)
Комплекс DDB1/DCAF1-CtD/Vpr мус был собран путем инкубации очищенных DDB1/DCAF1-CtD и HisSUMO-T4L-Vpr мус (остатки 1-92), в молярном соотношении 1:1, в буфере, содержащем 50 мМ бис-трис-пропана, рН 8,5, 0,5 М NaCl, 4 мМ MgCl 2 , 0,5 мМ TCEP, содержащий 1 мг HRV- Протеаза 3C для удаления HisSUMO-tag. После инкубации на льду в течение 12 часов образец загружали в колонку Superdex 200 16/600 GF (GE) с колонкой 1 мл GSH-Sepharose FF (GE), подключенной в линию.Колонку уравновешивали 10 мМ бис-трис-пропана, рН 8,5, 150 мМ NaCl, 4 мМ MgCl2 и 0,5 мМ TCEP. Скорость потока через колонку составляла 1 мл/мин.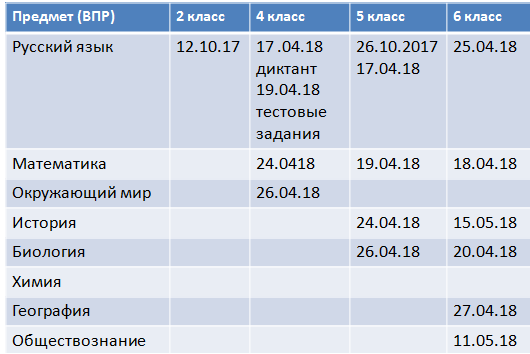 Фракции GF анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и концентрировали до 4,5 мг/мл.
Фракции GF анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и концентрировали до 4,5 мг/мл.
Кристаллы были приготовлены методом диффузии пара сидячей каплей путем смешивания равных объемов (200 нл) белкового комплекса с концентрацией 4,5 мг/мл и резервуарного раствора, содержащего 8-10% ПЭГ 4000 (масса/объем), 200 мМ MgCl 2 , 100 мМ HEPES-NaOH, pH 7.0-8,2. Объем резервуара составлял 75 мкл. Кристаллы вырастали после по меньшей мере 4 недель инкубации при 4°С. Кристаллы подвергали криозащите в резервуарном растворе с добавлением 20% глицерина и криоохлаждению в жидком азоте. Были собраны наборы данных с двух монокристаллов, сначала в BESSY II (Helmholtz-Zentrum Berlin, HZB) при длине волны 0,
Å, а затем в ESRF (Гренобль) при длине волны 1 Å. Наборы данных обрабатывались отдельно с использованием XDS [86] и XDSAPP [90]. Структура расшифрована путем молекулярной замены с использованием исходного набора данных BESSY с помощью программы PHASER [91] и следующих структур в качестве поисковых моделей: DDB1/DCAF1-CtD (настоящая работа) и вариант T4L E11H (PDB 1qt6) [92]. ].После оптимизации исходной модели и уточнения по сравнению с набором данных ESRF с более высоким разрешением, Vpr mus был помещен вручную в плотность, используя ЯМР-модель Vpr HIV-1 (PDB 1m8l) [93] в качестве ориентира. Итерационные циклы настройки модели с помощью программы Coot [88] с последующим уточнением с помощью программы PHENIX [89] дали окончательное соотношение R/R без факторов 21,61%/26,05%. В модели 95,1 % остатков имеют двугранные углы остова в предпочтительной области графика Рамачандрана, остальные попадают в разрешенные области, и ни один из них не является выбросом.Детали сбора данных и уточнения статистики представлены в таблице S1. Координаты и структурные факторы депонированы в PDB под инвентарным номером 6zx9.
].После оптимизации исходной модели и уточнения по сравнению с набором данных ESRF с более высоким разрешением, Vpr mus был помещен вручную в плотность, используя ЯМР-модель Vpr HIV-1 (PDB 1m8l) [93] в качестве ориентира. Итерационные циклы настройки модели с помощью программы Coot [88] с последующим уточнением с помощью программы PHENIX [89] дали окончательное соотношение R/R без факторов 21,61%/26,05%. В модели 95,1 % остатков имеют двугранные углы остова в предпочтительной области графика Рамачандрана, остальные попадают в разрешенные области, и ни один из них не является выбросом.Детали сбора данных и уточнения статистики представлены в таблице S1. Координаты и структурные факторы депонированы в PDB под инвентарным номером 6zx9.
Крио-ЭМ подготовка проб и сбор данных
Сборка комплекса
Очищенные CUL4-NEDD8/ROC1, DDB1/DCAF1-CtD, GST-Vpr mus и SAMHD1 макака-резуса, 1 мкМ каждый, инкубировали в конечном объеме 1 мл 10 мМ Tris-HCl pH 7,8, 150 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ TCEP, дополненный 1 мг протеазы GST-3C. После инкубации на льду в течение 12 часов образец загружали на колонку Superdex 200 16/600 GF (GE), уравновешенную тем же буфером со скоростью 1 мл/мин, с колонкой GSH-Sepharose FF (GE) объемом 1 мл, соединенной в линия. Фракции GF анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и концентрировали до 2,8 мг/мл.
Подготовка сетки
3,5 мкл белкового раствора, содержащего 0,05 мкМ CUL4-NEDD8/ROC1/DDB1/DCAF1-CtD/Vpr мус /SAMHD1 и 0,25 мкМ конъюгата UBCH5C-убиквитин (S4 A, B рис.), наносили на Перфорированная углеродная сетка Quantifoil R2/4 Cu/Rh 300 меш (Quantifoil Micro Tools GmbH), покрытая дополнительной тонкой углеродной пленкой в качестве подложки для образца и окрашенная 2% уранилацетатом для начальной характеристики.Для крио-ЭМ свежую углеродную сетку Quantifoil R1.2/1.3 Cu с отверстиями 400 меш (Quantifoil Micro Tools GmbH) подвергали тлеющему разряду в течение 30 с с использованием плазменной очистки Harrick с техническим воздухом при 0,3 мбар и 7 Вт. Белок 3,5 мкл. раствор, содержащий 0,4 мкМ комплекса CUL4-NEDD8/ROC1/DDB1/DCAF1-CtD/Vpr mus /SAMHD1 и 2 мкМ конъюгата UBCH5C-убиквитин, наносили на сетку, инкубировали в течение 45 с, блотировали с помощью устройства Vitrobot Mark II (FEI , Thermo Fisher Scientific) в течение 1-2 с при температуре 8°C и влажности 80% и погружали в жидкий этан.Сетки хранились в жидком азоте до визуализации.
Белок 3,5 мкл. раствор, содержащий 0,4 мкМ комплекса CUL4-NEDD8/ROC1/DDB1/DCAF1-CtD/Vpr mus /SAMHD1 и 2 мкМ конъюгата UBCH5C-убиквитин, наносили на сетку, инкубировали в течение 45 с, блотировали с помощью устройства Vitrobot Mark II (FEI , Thermo Fisher Scientific) в течение 1-2 с при температуре 8°C и влажности 80% и погружали в жидкий этан.Сетки хранились в жидком азоте до визуализации.
Сбор данных крио-ЭМ
Исходные наборы данных отрицательного окрашивания и крио-ЭМ были собраны автоматически для контроля качества образцов и реконструкции с низким разрешением на крио-ЭМ Tecnai Spirit 120 кВ (FEI, Thermo Fisher Scientific), оснащенном F416 CMOS камеры (TVIPS) с использованием Leginon [94, 95]. Затем изображения частиц были проанализированы с помощью 2D-классификации и первоначальной реконструкции модели с использованием SPHIRE [96], cisTEM [97] и Relion 3.07 [98]. Эти данные выявили наличие комплексов, содержащих как DDB1/DCAF1-CtD/Vpr mus (ядро), так и CUL4/ROC1 (стебель). Данные высокого разрешения были собраны на крио-ЭМ Tecnai Polara 300 кВ (FEI, Thermo Fisher Scientific), оснащенном детектором прямых электронов K2summit (Gatan) при номинальном увеличении 31000x с размером пикселя 0,625 Å/пиксель на масштаб объекта. Всего было собрано 3644 видеостека в режиме суперразрешения с использованием Leginon [94, 95] со следующими параметрами: диапазон расфокусировки 0.5-3,0 мкм, 40 кадров на фильм, время экспозиции 10 с, доза электронов 1,25 е/Å 2 /с и кумулятивная доза 50 е/Å 2 на фильм.
Данные высокого разрешения были собраны на крио-ЭМ Tecnai Polara 300 кВ (FEI, Thermo Fisher Scientific), оснащенном детектором прямых электронов K2summit (Gatan) при номинальном увеличении 31000x с размером пикселя 0,625 Å/пиксель на масштаб объекта. Всего было собрано 3644 видеостека в режиме суперразрешения с использованием Leginon [94, 95] со следующими параметрами: диапазон расфокусировки 0.5-3,0 мкм, 40 кадров на фильм, время экспозиции 10 с, доза электронов 1,25 е/Å 2 /с и кумулятивная доза 50 е/Å 2 на фильм.
Крио-ЭМ вычислительный анализ
Видеоролики были совмещены и взвешены по дозе с использованием MotionCor2 [99], а начальная оценка функции передачи контраста (CTF) была выполнена с помощью пакета CTTFind4 [100]. Полученные микрофотографии были проверены вручную, чтобы исключить изображения со значительными загрязнениями (как правило, большими агрегатами белка или загрязнениями льда) или артефактами сетки.Спектры мощности проверялись вручную, чтобы исключить изображения с астигматическими, слабыми или плохо определенными спектрами. После этих этапов контроля качества набор данных включал 2322 микрофотографии (63% от общего числа). На этом этапе набор данных был выбран дважды и обработан отдельно, чтобы получить реконструкции состояний-1, -2 и -3 (анализ 1) и ядра (анализ 2).
После этих этапов контроля качества набор данных включал 2322 микрофотографии (63% от общего числа). На этом этапе набор данных был выбран дважды и обработан отдельно, чтобы получить реконструкции состояний-1, -2 и -3 (анализ 1) и ядра (анализ 2).
Для анализа 1 положения частиц были определены с использованием программы гауссовского пикинга cisTEMs, в результате чего было получено в общей сложности 959 155 изображений частиц.После двух раундов 2D-классификации для дальнейшей обработки было отобрано 227 529 изображений частиц (S4C, D Fig). Используя эти данные, была создана исходная модель с использованием Relion 3.07. Полученная карта дала сильный сигнал для ядра, но только для фрагментированной плотности стебля, что указывает на большую неоднородность области стебля в наборе данных. Эта большая степень композиционной (+/- ножка) и конформационной неоднородности (движение ножки относительно ядра) усложняла классификацию.Соответственно, выравнивание и классификация проводились одновременно. Первая цель состояла в том, чтобы разделить набор данных на три категории: «мусор», «ядро» и «ядро+стебель».
Первая цель состояла в том, чтобы разделить набор данных на три категории: «мусор», «ядро» и «ядро+стебель».
Поэтому ножка была удалена из исходной модели с помощью инструмента «Ластик» в Chimera [101]. Эта основная карта использовалась в качестве исходной модели для 3D-классификации уровня 1 с Relion 3.07 с уменьшенным размером пикселя 2,5 Å/пиксель. Использовались следующие параметры: количество классов K=6, T=10, глобальный шаг поиска=7.5°, количество итераций = 25. Классификация дала два класса, содержащих стебель (классы 3 и 5, содержащие 23% и 22% изображений частиц соответственно) (рис. S4D). Эти частицы были объединены и направлены на 3D-классификацию уровня 2 с использованием следующих параметров: количество классов K=6, T=10, глобальный шаг поиска=7,5°, количество итераций=25. Три из этих классов дали карты среднего разрешения с интерпретируемыми функциями (состояния-1, -2 и -3, рис. S4D). Эти три класса были уточнены по отдельности с помощью 3D-автоматического уточнения в Relion 3. 07, в результате чего были получены карты с разрешением от 7,8 Å до 8,9 Å (рис. S4E-G).
07, в результате чего были получены карты с разрешением от 7,8 Å до 8,9 Å (рис. S4E-G).
Для анализа 2 положения частиц определяли с помощью сопоставления шаблона с отфильтрованной картой, включающей ядро и основу, с использованием программного обеспечения Gautomatch (https://www2.mrc-lmb.cam.ac.uk/research/locally-developed- программное обеспечение/чжан-программное обеспечение/). Было найдено 712 485 изображений частиц, извлеченных с помощью Relion 3.07 и впоследствии 2D-классифицированных с помощью cryoSPARC [102], в результате чего после отбора было получено 505 342 изображения частиц (S5A, B Fig).Эти изображения частиц были разделены на два подмножества одинакового размера, и для уменьшения вычислительной нагрузки была выполнена трехмерная классификация уровня 1 с использованием Relion 3.07 для обоих из них (рис. S5B). Были использованы следующие параметры: начальная модель = «ядро», количество классов K = 4, T = 10, глобальный шаг поиска = 7,5 °, количество итераций = 25, размер пикселя 3,75 Å/px. Из них были отобраны те, у которых есть и сердцевина, и стебель. Классы, изображающие одинаковую ориентацию стебля относительно ядра, были объединены и направлены на уровень 2 в виде трех разных субпопуляций, содержащих 143 172, 193 059 и 167 666 изображений частиц соответственно (рис. S5B).
Из них были отобраны те, у которых есть и сердцевина, и стебель. Классы, изображающие одинаковую ориентацию стебля относительно ядра, были объединены и направлены на уровень 2 в виде трех разных субпопуляций, содержащих 143 172, 193 059 и 167 666 изображений частиц соответственно (рис. S5B).
Для уровня 2 каждая субпопуляция была классифицирована отдельно по 4 классам. Из этих 12 классов все изображения частиц, демонстрирующие четко определенные плотности ядра и стебля, были объединены и помечены как «ядро + стебель», в результате чего в общей сложности было получено 310 801 изображение частиц. 193 096 изображений частиц, представляющих классы, содержащие только ядро, были объединены и помечены как «ядро» (рис. S5B).Подмножество «ядро + стебель» было разделено на 6 классов, 5 из которых содержали как стебель, так и сердцевину (рис. S5B), а один класс состоял только из ядра со связанным Vpr mus . 5 классов со стеблем показали ориентацию стебля, аналогичную той, что была получена в результате анализа 1 (см. выше, S4 рис.), но были уточнены по отдельности до более низкого разрешения, как в анализе 1, и были отброшены. Однако индивидуальное уточнение класса уровня 3 только для ядра дало реконструкцию 7,3 Å (S5C, рис. D).
выше, S4 рис.), но были уточнены по отдельности до более низкого разрешения, как в анализе 1, и были отброшены. Однако индивидуальное уточнение класса уровня 3 только для ядра дало реконструкцию 7,3 Å (S5C, рис. D).
Молекулярная визуализация, подгонка твердого тела, трехмерное структурное выравнивание, анализ вращения и интерфейса
Карты плотности и атомные модели были визуализированы с помощью Coot [88], PyMOL (Schrödinger) и UCSF Chimera [101].Подгонка жесткого тела и структурное выравнивание выполнялись с использованием программы UCSF Chimera [101]. Углы поворота между крайними положениями домена DDB1 BPB измеряли с помощью сервера DynDom [103] (http://dyndom.cmp.uea.ac.uk/dyndom/runDyndom.jsp). Молекулярные интерфейсы анализировали с помощью сервера EBI PDBePISA [104] (https://www.ebi.ac.uk/msd-srv/prot_int/cgi-bin/piserver).
Масс-спектрометрия сшивки (CLMS)
Сборка комплекса
Очищенные CUL4/ROC1, DDB1/DCAF1-CtD, GST-Vpr mus и SAMHD1 макака-резуса, по 1 мкМ каждого, инкубировали в объеме 3 мл буфер, содержащий 10 мМ HEPES pH 7. 8, 150 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ TCEP, дополненный 1 мг протеазы GST-3C. После инкубации на льду в течение 12 ч образец загружали на колонку Superdex 200 16/600 GF (GE), уравновешенную тем же буфером, со скоростью потока 1 мл/мин с колонкой 1 мл GSH-Sepharose FF ( GE) подключены в линию. Фракции GF анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и концентрировали до 6 мг/мл.
8, 150 мМ NaCl, 4 мМ MgCl 2 , 0,5 мМ TCEP, дополненный 1 мг протеазы GST-3C. После инкубации на льду в течение 12 ч образец загружали на колонку Superdex 200 16/600 GF (GE), уравновешенную тем же буфером, со скоростью потока 1 мл/мин с колонкой 1 мл GSH-Sepharose FF ( GE) подключены в линию. Фракции GF анализировали с помощью SDS-PAGE, соответствующие фракции объединяли и концентрировали до 6 мг/мл.
Photo-Crosslinking
Сшивающий агент сульфо-SDA (сульфосукцинимидил-4,4′-азипентаноат) (Thermo Scientific) растворяли в сшивающем буфере (10 мМ HEPES pH 7.8, 150 мМ NaCl, 4 мМ MgCl2, 0,5 мМ TCEP) до 100 мМ перед использованием. Стадию мечения проводили путем инкубации аликвот по 18 мкг комплекса в концентрации 1 мг/мл с добавлением 2, 1, 0,5, 0,25, 0,125 мМ сульфо-SDA, соответственно, в течение часа. Затем образцы облучали УФ-светом с длиной волны 365 нм для образования поперечных связей в течение 20 мин и гасили 50 мМ Nh5HCO3 в течение 20 мин. Все шаги выполнялись на льду. Продукты реакции разделяли на геле Novex Bis-Tris 4–12% SDS-PAGE (Life Technologies). Полосу геля, соответствующую сшитому комплексу, вырезали и расщепляли трипсином (Thermo Scientific Pierce) [107], а полученные триптические пептиды экстрагировали и обессоливали с помощью C18 StageTips [108].Элюированные пептиды фракционировали на колонке Superdex Peptide 3,2/300 (GE Healthcare) при скорости потока 10 мкл/мин с использованием 30% (об./об.) ацетонитрила и 0,1% (об./об.) трифторуксусной кислоты в качестве подвижной фазы. Фракции по 50 мкл собирали и сушили в вакууме.
Продукты реакции разделяли на геле Novex Bis-Tris 4–12% SDS-PAGE (Life Technologies). Полосу геля, соответствующую сшитому комплексу, вырезали и расщепляли трипсином (Thermo Scientific Pierce) [107], а полученные триптические пептиды экстрагировали и обессоливали с помощью C18 StageTips [108].Элюированные пептиды фракционировали на колонке Superdex Peptide 3,2/300 (GE Healthcare) при скорости потока 10 мкл/мин с использованием 30% (об./об.) ацетонитрила и 0,1% (об./об.) трифторуксусной кислоты в качестве подвижной фазы. Фракции по 50 мкл собирали и сушили в вакууме.
Получение методом CLMS
Образцы для анализа ресуспендировали в 0,1% (об./об.) муравьиной кислоте, 3,2% (об./об.) ацетонитриле. Анализ ЖХ-МС/МС выполняли на масс-спектрометре Orbitrap Fusion Lumos Tribrid (Thermo Fisher), соединенном в режиме реального времени с системой ВЭЖХ Ultimate 3000 RSLCnano (Dionex, Thermo Fisher).Образцы разделяли на колонке EASY-Spray длиной 50 см (Thermo Fisher). Подвижная фаза А состояла из 0,1% (об. /об.) муравьиной кислоты, а подвижная фаза В состояла из 80% (об./об.) ацетонитрила с 0,1% (об./об.) муравьиной кислоты. Скорость потока составляла 0,3 мкл/мин с использованием градиентов, оптимизированных для каждой хроматографической фракции автономного фракционирования, в диапазоне от 2% подвижной фазы B до 55% подвижной фазы B в течение 90 минут. Данные МС были получены в режиме зависимости от данных с использованием настройки максимальной скорости с временем цикла 3 с. Для каждого цикла с помощью Orbitrap записывали полный сканирующий масс-спектр с разрешением 120 000 в диапазоне от 400 до 1 500 м/з.Ионы с исходным зарядовым состоянием от 3+ до 7+ были выделены и фрагментированы. Фрагментация аналита была достигнута с помощью высокоэнергетической столкновительной диссоциации (HCD) [109], после чего спектры фрагментации были записаны в Orbitrap с разрешением 50 000. Динамическое исключение было включено с подсчетом одиночных повторов и длительностью исключения 60 с.
/об.) муравьиной кислоты, а подвижная фаза В состояла из 80% (об./об.) ацетонитрила с 0,1% (об./об.) муравьиной кислоты. Скорость потока составляла 0,3 мкл/мин с использованием градиентов, оптимизированных для каждой хроматографической фракции автономного фракционирования, в диапазоне от 2% подвижной фазы B до 55% подвижной фазы B в течение 90 минут. Данные МС были получены в режиме зависимости от данных с использованием настройки максимальной скорости с временем цикла 3 с. Для каждого цикла с помощью Orbitrap записывали полный сканирующий масс-спектр с разрешением 120 000 в диапазоне от 400 до 1 500 м/з.Ионы с исходным зарядовым состоянием от 3+ до 7+ были выделены и фрагментированы. Фрагментация аналита была достигнута с помощью высокоэнергетической столкновительной диссоциации (HCD) [109], после чего спектры фрагментации были записаны в Orbitrap с разрешением 50 000. Динамическое исключение было включено с подсчетом одиночных повторов и длительностью исключения 60 с.
Обработка CLMS
Повторная калибровка предшественника m/z была проведена на основе высокодостоверных (<1% доля ложных открытий (FDR)) линейных идентификаций пептидов. Повторно откалиброванные списки пиков сравнивали с последовательностями и обратными последовательностями (в качестве ловушек) перекрестно-сшитых пептидов с использованием пакета программного обеспечения Xi (v.1.7.5.1) для идентификации [110]. Окончательные списки перекрестных связей были составлены с использованием идентифицированных кандидатов, отфильтрованных до <1% FDR на уровне связи с xiFDR v.2.0 [111], обеспечивающим минимальное покрытие последовательностей 20% и 4 наблюдаемых фрагмента на пептид.
Повторно откалиброванные списки пиков сравнивали с последовательностями и обратными последовательностями (в качестве ловушек) перекрестно-сшитых пептидов с использованием пакета программного обеспечения Xi (v.1.7.5.1) для идентификации [110]. Окончательные списки перекрестных связей были составлены с использованием идентифицированных кандидатов, отфильтрованных до <1% FDR на уровне связи с xiFDR v.2.0 [111], обеспечивающим минимальное покрытие последовательностей 20% и 4 наблюдаемых фрагмента на пептид.
Анализ CLMS
Для того чтобы выбрать доступный объем взаимодействия SAMHD1-CtD в соответствии с данными CLMS, модель SAMHD1 была создана с использованием I-TASSER [112].Из модели был извлечен SAMHD1-CtD со случайной конфигурацией катушки. Чтобы отобразить все перекрестные связи, недостающие петли в сложной структуре были сгенерированы с помощью MODELLER [113]. Затем поиск объема взаимодействия был отправлен на веб-сервер DisVis [61] с допустимым расстоянием от 1,5 Å до 22 Å для каждого ограничения с использованием опции «полное сканирование». Интервал выборки вращения был установлен на 9,72 °, а шаг вокселя сетки — на 1 Å. Доступный объем взаимодействия визуализировали с помощью UCSF Chimera [101].
Интервал выборки вращения был установлен на 9,72 °, а шаг вокселя сетки — на 1 Å. Доступный объем взаимодействия визуализировали с помощью UCSF Chimera [101].
Спектроскопия кругового дихроизма (CD)
Для оценки содержания вторичной структуры Vpr mus , меченного GST, и двойных мутантов R15E/R75E и W29A/A66W была проведена спектроскопия CD. Образцы белка подвергали диализу в течение ночи при 4°C против буфера CD (смесь 100 мМ NaF, 10 мМ K 2 HPO 4 /KH 2 PO 4 , pH 8,5) и концентрацию белка доводили до 0,2 мг. /мл. Для каждого образца регистрировали три повторных спектра КД в 1.Шаг 0 нм в диапазоне 190-260 нм. Измерения проводились в кварцевых кюветах с длиной оптического пути 0,05 см (Hellma, Мюльхайм, Германия) при 20°C на спектрополяриметре Chirascan (Applied Photophysics, Лондон, Великобритания). Эталонный спектр буфера CD вычитали из усредненного спектра образца белка. Измеренная эллиптичность в миллиградусах (m°) была преобразована в молярную эллиптичность [Θ] в соответствии с уравнением Θ=m°*M/(10xLxC), где M — средняя молекулярная масса белка, L — длина пути измерительной ячейки, а C — молярная концентрация белка. Молярную эллиптичность строили в зависимости от длины волны с использованием SigmaPlot версии 14.0 (Systat Software Inc., Сан-Хосе, Калифорния, США).
Молярную эллиптичность строили в зависимости от длины волны с использованием SigmaPlot версии 14.0 (Systat Software Inc., Сан-Хосе, Калифорния, США).
Доступность данных
Координаты и структурные факторы для кристаллических структур были депонированы в Банке белковых данных (PDB) с кодами доступа 6ZUE (DDB1/DCAF1-CtD) и 6ZX9 (DDB1/DCAF1-CtD/T4L-Vpr муз 1-92). Крио-ЭМ-реконструкции были депонированы в банке данных электронной микроскопии (EMDB) под кодами доступа EMD-10611 (основной), EMD-10612 (конформационное состояние-1), EMD-10613 (состояние-2) и EMD-10614 ( состояние-3).Данные CLMS депонированы в базе данных PRIDE [114] под кодом доступа PXD020453, пароль рецензента fCrQG2u8.
Благодарности
Мы благодарим MPI-MG за предоставление доступа к инструментам TEM группы обслуживания микроскопии и крио-ЭМ. Мы благодарим доктора Манфреда Вайса и научный персонал BESSY-MX (Макромолекулярная рентгеновская кристаллография)/Helmholtz Zentrum Berlin für Materialien und Energie на каналах BL14. 1, BL14.2 и BL14.3, эксплуатируемых Joint Berlin MX. — Лаборатория на кольце хранения электронов BESSY II (Берлин-Адлерсхоф, Германия), а также научный персонал ESRF (Гренобль, Франция) на каналах ID30A-3, ID30B, ID23-1, ID23-2 и ID29 для непрерывного служба поддержки.Мы благодарим компанию Diamond Light Source (Didcot, Великобритания) за доступ и поддержку синхротронного луча I04 и крио-ЭМ-оборудования в Национальном центре электронной биовизуализации Великобритании (eBIC). Кроме того, авторы признательны Северогерманскому альянсу суперкомпьютеров (HLRN) и кластеру высокопроизводительных вычислений для исследований Берлинского института здравоохранения за предоставление ресурсов для высокопроизводительных вычислений. Мы благодарны профессору Удо Хайнеманну и Цзяньхуэй Вану (Max-Delbrück-Centrum, Берлин, Германия) за доступ и помощь во время спектроскопии КД.Плазмида pHisSUMO была щедрым подарком доктора Евангелоса Христодулу (Институт Фрэнсиса Крика, Лондон, Великобритания). Матрица кДНК SAMHD1 макаки-резуса была щедрым подарком профессора Майкла Эмермана (Центр исследования рака им.
1, BL14.2 и BL14.3, эксплуатируемых Joint Berlin MX. — Лаборатория на кольце хранения электронов BESSY II (Берлин-Адлерсхоф, Германия), а также научный персонал ESRF (Гренобль, Франция) на каналах ID30A-3, ID30B, ID23-1, ID23-2 и ID29 для непрерывного служба поддержки.Мы благодарим компанию Diamond Light Source (Didcot, Великобритания) за доступ и поддержку синхротронного луча I04 и крио-ЭМ-оборудования в Национальном центре электронной биовизуализации Великобритании (eBIC). Кроме того, авторы признательны Северогерманскому альянсу суперкомпьютеров (HLRN) и кластеру высокопроизводительных вычислений для исследований Берлинского института здравоохранения за предоставление ресурсов для высокопроизводительных вычислений. Мы благодарны профессору Удо Хайнеманну и Цзяньхуэй Вану (Max-Delbrück-Centrum, Берлин, Германия) за доступ и помощь во время спектроскопии КД.Плазмида pHisSUMO была щедрым подарком доктора Евангелоса Христодулу (Институт Фрэнсиса Крика, Лондон, Великобритания). Матрица кДНК SAMHD1 макаки-резуса была щедрым подарком профессора Майкла Эмермана (Центр исследования рака им. Фреда Хатчинсона, Сиэтл, США). Рекомбинантная бакмида BAC10:1629KO была щедрым подарком профессора Яна Джонса (Университет Рединга, Великобритания). pAcGHLT-B-DDB1 был подарен профессором Нин Чжэн (плазмида Addgene 48638). pET28-mE1 был подарен профессором Хорхе Эдуардо Азеведо (плазмида Addgene 32534).
Фреда Хатчинсона, Сиэтл, США). Рекомбинантная бакмида BAC10:1629KO была щедрым подарком профессора Яна Джонса (Университет Рединга, Великобритания). pAcGHLT-B-DDB1 был подарен профессором Нин Чжэн (плазмида Addgene 48638). pET28-mE1 был подарен профессором Хорхе Эдуардо Азеведо (плазмида Addgene 32534).
Принципы дизайна регуляторов транскрипции на основе CRISPR с дефицитом нуклеаз | Исследование дрожжей FEMS
Аннотация
Разработка кластеризованных регулярно расположенных коротких палиндромных повторов (CRISPR) — CRISPR-ассоциированных белков продолжает расширять набор инструментов, доступных для редактирования генома, перепрограммирования регуляции генов, визуализации генома и эпигенетических исследований живых организмов. В этом обзоре будут представлены новые принципы дизайна с использованием перепрограммирования экспрессии генов на основе CRISPR с дефицитом нуклеаз.В обзоре основное внимание будет уделено конструкциям, реализованным в дрожжах как на уровне белков CRISPR, так и на направляющей РНК (гРНК), но, где это уместно, будут отданы должное исходным исследованиям, проведенным на других видах. В дополнение к принципам дизайна в этом обзоре также освещаются приложения, извлекающие выгоду из использования регуляции транскрипции, опосредованной CRISPR, и обсуждаются будущие направления дальнейшего расширения набора инструментов для перепрограммирования геномов с дефицитом нуклеаз. Таким образом, этот обзор должен представлять общий интерес для экспериментаторов, чтобы ознакомиться с параметрами, лежащими в основе возможностей перепрограммирования геномных функций с использованием технологий CRISPR с дефицитом нуклеаз.
В дополнение к принципам дизайна в этом обзоре также освещаются приложения, извлекающие выгоду из использования регуляции транскрипции, опосредованной CRISPR, и обсуждаются будущие направления дальнейшего расширения набора инструментов для перепрограммирования геномов с дефицитом нуклеаз. Таким образом, этот обзор должен представлять общий интерес для экспериментаторов, чтобы ознакомиться с параметрами, лежащими в основе возможностей перепрограммирования геномных функций с использованием технологий CRISPR с дефицитом нуклеаз.
ВВЕДЕНИЕ
Живые клетки регулируют экспрессию генов за счет координированных действий ДНК-связывающих регуляторов транскрипции, РНК-полимераз и арсенала вспомогательных коактиваторов (Hahn 2004). Сложная сеть механизмов транскрипции контролирует основные функции, такие как клеточная дифференцировка, клеточное деление, реакции на условия окружающей среды и метаболизм. Наше механистическое понимание генов и путей, подтверждающих своевременное и адекватное выполнение этих важных функций, в значительной степени опиралось на исследования функциональной геномики, часто сопровождаемые эффективными методологиями для точного контроля нарушений экспрессии генов (Khalil et al. 2012; Si и др. 2015).
2012; Si и др. 2015).
РНК-интерференция, посттранскрипционный механизм молчания генов, запускаемый малыми интерферирующими РНК или короткими шпилечными РНК, образованными в результате эндонуклеазно-опосредованной РНКазы III деградации двухцепочечных РНК, является одной из таких методологий (Drinnenberg et al. 2009). С помощью итеративной РНКи у микробов был оптимизирован нокдаун нескольких генов, связанных с химической устойчивостью и производством гетерологичных метаболитов (Crook, Schmitz and Alper, 2013; Si et al. 2015). Другой метод, используемый для изменения экспрессии сотен генов, называемый глобальная инженерия транскрипционных механизмов (gTME), основан на введении мутантных библиотек общих факторов транскрипции, регулирующих специфичность промотора, а затем на скрининге определенных фенотипов с последующей характеристикой и подтверждением мутантного контекста гена. фактор транскрипции и анализ транскриптома (Alper et al. 2006). Кроме того, для целенаправленной регуляции генов восходящая инженерия синтетических факторов транскрипции на основе белков гибридных цинковых пальцев (ZF) и промоторов для ортогонального контроля экспрессии генов выявила важные параметры для скоординированной, настроенной и пространственной регуляции экспрессии генов (Khalil и др. 2012). В совокупности разработка методов исследования условной потери функции путем нарушений экспрессии нескольких генов оказалась важной для нашего понимания функции генов, особенно при изучении функции основных генов и полигенных признаков (например, химической устойчивости). . Однако, хотя вышеупомянутые методы поддерживают простое нацеливание на множественные гены для нокдауна и сверхэкспрессии, недостатки очевидны. Это включает в себя отсутствие специфичности и ограниченный регуляторный потенциал (RNAi), необходимость введения синтетического геномного материала (ZF) или потребность в системе скрининга для отбора глобальных транскрипционных изменений, которые не предполагались a priori (gTME).
Кроме того, для целенаправленной регуляции генов восходящая инженерия синтетических факторов транскрипции на основе белков гибридных цинковых пальцев (ZF) и промоторов для ортогонального контроля экспрессии генов выявила важные параметры для скоординированной, настроенной и пространственной регуляции экспрессии генов (Khalil и др. 2012). В совокупности разработка методов исследования условной потери функции путем нарушений экспрессии нескольких генов оказалась важной для нашего понимания функции генов, особенно при изучении функции основных генов и полигенных признаков (например, химической устойчивости). . Однако, хотя вышеупомянутые методы поддерживают простое нацеливание на множественные гены для нокдауна и сверхэкспрессии, недостатки очевидны. Это включает в себя отсутствие специфичности и ограниченный регуляторный потенциал (RNAi), необходимость введения синтетического геномного материала (ZF) или потребность в системе скрининга для отбора глобальных транскрипционных изменений, которые не предполагались a priori (gTME).
С 2013 г. система бактериальных коротких палиндромных повторов с регулярными интервалами (CRISPR)–Cas вдохновила на рациональную разработку ортогональных синтетических стратегий перепрограммирования транскрипции, основанных на РНК-опосредованном нацеливании белков Cas с дефицитом нуклеаз на заранее определенные геномные локусы (Larson et и др. , 2013 г.; Qi и др., , 2013 г.). Вкратце, системы CRISPR-Cas основаны на древней бактериальной адаптивной иммунной системе, в которой CRISPR-ассоциированный белок (Cas) направляется в геномные локусы с помощью направляющей РНК (гРНК) с последовательностью 20 нуклеотидов, комплементарной геномному сайту-мишени (Jinek и др. 2012; Конг и др. 2013). На этой платформе появились две основные системы для (i) редактирования генома с использованием гРНК-опосредованных эндонуклеазно-опосредованных разрывов ДНК (DSB) для облегчения как нокаута, так и нокаута гена (Jinek et al. 2012; Cong et al. 2013), и (ii) dCas-опосредованная транскрипционная и посттранскрипционная регуляция с дефицитом нуклеаз, выяснение эпигенетических ландшафтов и редактирование оснований с дефицитом DSB, и это лишь некоторые из них (Qi et al. 2013; Lenstra и другие. 2015; Фу и др. 2016; Нисида и др. 2016; Кокс и др. 2017). С точки зрения регуляции транскрипции, нуклеазо-дефицитные формы CRISPR-ассоциированного белка Cas9 типа II, называемого dCas9, из Streptococcus pyogenes , были признаны мощной платформой для перепрограммирования экспрессии генов и геномной функции. По сути, dCas9 представляет собой мутант Cas9, нуклеазная активность которого снижена за счет мутаций в нуклеазных доменах RuvC и HNH, но при этом сохраняется способность связывания ДНК, запрограммированная гРНК (Qi et al. 2013). Первоначально было продемонстрировано, что dCas9 и гРНК могут опосредовать эффективную репрессию генов в бактериях, когда dCas9 направляется в проксимальные положения промотора ниже сайта начала транскрипции (TSS), механизм, созданный интерференцией CRISPR (CRISPRi) (Larson et al.
2013), и (ii) dCas-опосредованная транскрипционная и посттранскрипционная регуляция с дефицитом нуклеаз, выяснение эпигенетических ландшафтов и редактирование оснований с дефицитом DSB, и это лишь некоторые из них (Qi et al. 2013; Lenstra и другие. 2015; Фу и др. 2016; Нисида и др. 2016; Кокс и др. 2017). С точки зрения регуляции транскрипции, нуклеазо-дефицитные формы CRISPR-ассоциированного белка Cas9 типа II, называемого dCas9, из Streptococcus pyogenes , были признаны мощной платформой для перепрограммирования экспрессии генов и геномной функции. По сути, dCas9 представляет собой мутант Cas9, нуклеазная активность которого снижена за счет мутаций в нуклеазных доменах RuvC и HNH, но при этом сохраняется способность связывания ДНК, запрограммированная гРНК (Qi et al. 2013). Первоначально было продемонстрировано, что dCas9 и гРНК могут опосредовать эффективную репрессию генов в бактериях, когда dCas9 направляется в проксимальные положения промотора ниже сайта начала транскрипции (TSS), механизм, созданный интерференцией CRISPR (CRISPRi) (Larson et al. 2013 г. ; Qi и др. , 2013 г.).
2013 г. ; Qi и др. , 2013 г.).
В последние годы стало очевидным, что по сравнению с упомянутыми выше традиционными подходами к перепрограммированию функции генома с помощью ненативных регуляторов транскрипции нуклеазо-дефицитные варианты Cas9 и бактерии Lachnospiraceae ND2006 Cpf1 являются мощными РНК-управляемыми технологии регуляции генома дрожжей.В частности, широкое распространение получили удобство, специфичность, надежность и масштабируемость активации и репрессии эндогенных генов (Фарзадфард, Перли и Лу, 2013; Гилберт, и др., , 2013; Залатан, и др., , 2015; Лиан, и др., ). 2017). Кроме того, CRISPR-опосредованная регуляция транскрипции представляет собой мощный подход к целевому, комбинаторному и настраиваемому интерфейсу перепрограммирования транскрипции, особенно с учетом простоты синтеза и экспрессии гРНК без длительной генетической модификации геномов хозяина видов, сопротивляющихся трансформации и целенаправленному редактированию генома.
В этом обзоре будет рассмотрен огромный прогресс CRISPR-опосредованных систем, применяемых для репрограммирования регуляции транскрипции у дрожжей, включая обширный список факторов, влияющих на эффективность гРНК, и принципы дизайна для оптимальной реконфигурации dCas9 и dCpf1. В конце обзора будут освещены будущие перспективы использования белков Cas с дефицитом нуклеаз в сочетании с другими дополнительными появляющимися технологиями для перепрограммирования функций генома без необходимости экзогенной нуклеазной активности.Хотя этот обзор будет посвящен в основном опосредованному dCas9 перепрограммированию экспрессии генов в дрожжах, недавно был опубликован более независимый от хозяина обзор технологий CRISPR-dCas с дефицитом нуклеаз (Mitsunobu et al. 2017).
РЕГЛАМЕНТ ТРАНСКРИПЦИИ НА ОСНОВЕ CRISPR
Модуляция активности dCas9
Регуляция активности белка CRISPR слиянием белков
Регуляторы транскрипции имеют глобулярную форму. Чаще всего регуляторы включают два модульных домена, обеспечивающих (i) связывание ДНК и (ii) регуляторный домен, поддерживающий активацию или репрессию транскрипции (Jensen et al. 2010; Khalil et al. 2012). Благодаря этой модульности эксперименты по замене доменов оказались успешными для создания синтетических регуляторов транскрипции с определенной специфичностью связывания ДНК, слитых с различными регуляторными доменами, чтобы усилить транскрипционную активацию или репрессию как нативных, так и синтетических промоторов (Khalil et al. 2012; Фолчер и др. 2013). Модульность регуляторных доменов позволила разработать регуляторы транскрипции, которые могут регулировать экспрессию генов до гораздо более высоких уровней по сравнению с регуляторами, основанными только на нативной конструкции (Folcher et al. 2013).
Чаще всего регуляторы включают два модульных домена, обеспечивающих (i) связывание ДНК и (ii) регуляторный домен, поддерживающий активацию или репрессию транскрипции (Jensen et al. 2010; Khalil et al. 2012). Благодаря этой модульности эксперименты по замене доменов оказались успешными для создания синтетических регуляторов транскрипции с определенной специфичностью связывания ДНК, слитых с различными регуляторными доменами, чтобы усилить транскрипционную активацию или репрессию как нативных, так и синтетических промоторов (Khalil et al. 2012; Фолчер и др. 2013). Модульность регуляторных доменов позволила разработать регуляторы транскрипции, которые могут регулировать экспрессию генов до гораздо более высоких уровней по сравнению с регуляторами, основанными только на нативной конструкции (Folcher et al. 2013).
Когда дефицитный по нуклеазе dCas9 первоначально использовался в бактериях, сообщалось о репрессии генов до 99,9% (Qi et al. 2013). Однако при использовании только dCas9 и одной гРНК в дрожжах для регуляции экспрессии целевого гена сообщалось лишь об умеренных репрессиях, варьирующихся от отсутствия эффекта до 2-3-кратной репрессии (Farzadfard, Perli and Lu 2013; Deaner, Mejia and Alper 2017; Vanegas, Lehka and Mortensen 2017), хотя в одном исследовании сообщалось о 18-кратном снижении активности репортерного гена (Gilbert et al. 2013). Этот уровень регуляции сравним с исследованиями на других эукариотах и позволяет предположить, что одного комплекса гРНК с dCas9 недостаточно для стерического затруднения прогрессии РНК и/или блокировки инициации транскрипции (Gilbert et al. 2013; Lawhorn, Ferreira and Wang). 2014). Вдохновленные модульной конструкцией других синтетических регуляторов транскрипции и признавая, что белки CRISPR, связанные с гРНК, аналогичны простым молекулам, связывающимся с ДНК, исследования с использованием нарушений экспрессии, опосредованных dCas9 или dCpf1, в настоящее время включают дополнительные регуляторные домены, слитые с dCas9 и/или dCpf1 для улучшения потенциалов репрессии и активации (рис.
2013). Однако при использовании только dCas9 и одной гРНК в дрожжах для регуляции экспрессии целевого гена сообщалось лишь об умеренных репрессиях, варьирующихся от отсутствия эффекта до 2-3-кратной репрессии (Farzadfard, Perli and Lu 2013; Deaner, Mejia and Alper 2017; Vanegas, Lehka and Mortensen 2017), хотя в одном исследовании сообщалось о 18-кратном снижении активности репортерного гена (Gilbert et al. 2013). Этот уровень регуляции сравним с исследованиями на других эукариотах и позволяет предположить, что одного комплекса гРНК с dCas9 недостаточно для стерического затруднения прогрессии РНК и/или блокировки инициации транскрипции (Gilbert et al. 2013; Lawhorn, Ferreira and Wang). 2014). Вдохновленные модульной конструкцией других синтетических регуляторов транскрипции и признавая, что белки CRISPR, связанные с гРНК, аналогичны простым молекулам, связывающимся с ДНК, исследования с использованием нарушений экспрессии, опосредованных dCas9 или dCpf1, в настоящее время включают дополнительные регуляторные домены, слитые с dCas9 и/или dCpf1 для улучшения потенциалов репрессии и активации (рис. 1а и б).
1а и б).
Рисунок 1.
Модуляция активности Cas9 и Cpf1 с дефицитом нуклеаз в дрожжах путем слияния доменов, регулирующих транскрипцию. ( a) Схематическая иллюстрация доменов репрессии транскрипции, которые были успешно слиты с активацией dCas9 CRISPR с дефицитом нуклеаз в дрожжах. ( b ) Схематическая иллюстрация доменов активации транскрипции, которые были успешно слиты с дефицитными по нуклеазе dCas9 и dCpf1 для активации CRISPR в дрожжах.
Рисунок 1.
Модуляция активности Cas9 и Cpf1 с дефицитом нуклеаз в дрожжах путем слияния доменов, регулирующих транскрипцию. ( a) Схематическая иллюстрация доменов репрессии транскрипции, которые были успешно слиты с активацией dCas9 CRISPR с дефицитом нуклеаз в дрожжах. ( b ) Схематическая иллюстрация доменов активации транскрипции, которые были успешно слиты с дефицитными по нуклеазе dCas9 и dCpf1 для активации CRISPR в дрожжах.
В своем основополагающем исследовании dCas9-опосредованной регуляции транскрипции у эукариот Gilbert et al. сравнили эффект слияния домена репрессора транскрипции млекопитающих, Mxi1, который, как сообщается, взаимодействует с модифицирующим хроматин гомологом гистоновой деацетилазы Sin3 в дрожжах, с dCas9 (Schreiber-Agus et al. 1995; Gilbert et al. 2013) ( Рис. 1а). Нацелившись на промотор TEF1 , dCas9-Mxi1 репрессировал активность репортерного гена в 53 раза по сравнению с упомянутым выше 18-кратным использованием только dCas9.Этот результат сравним с эффектом, о котором недавно сообщалось в исследовании Yarrowia lipolytica (Schwartz et al. 2017) . Здесь Schwartz et al. сообщили о 10-кратной репрессии уровней транскриптов MIh2 при использовании dCas9, но при прямом сравнении эффектов использования dCas9 по сравнению с dCas9-Mxi1 на гены Ku70 и Ku80 , связанные с негомологичным соединением концов, Шварц и др. наблюдали самый высокий уровень репрессии (87%) для Ku80 , когда гибрид dCas9-Mxi1 сравнивали с dCas9 (38%) (Schwartz et al.
сравнили эффект слияния домена репрессора транскрипции млекопитающих, Mxi1, который, как сообщается, взаимодействует с модифицирующим хроматин гомологом гистоновой деацетилазы Sin3 в дрожжах, с dCas9 (Schreiber-Agus et al. 1995; Gilbert et al. 2013) ( Рис. 1а). Нацелившись на промотор TEF1 , dCas9-Mxi1 репрессировал активность репортерного гена в 53 раза по сравнению с упомянутым выше 18-кратным использованием только dCas9.Этот результат сравним с эффектом, о котором недавно сообщалось в исследовании Yarrowia lipolytica (Schwartz et al. 2017) . Здесь Schwartz et al. сообщили о 10-кратной репрессии уровней транскриптов MIh2 при использовании dCas9, но при прямом сравнении эффектов использования dCas9 по сравнению с dCas9-Mxi1 на гены Ku70 и Ku80 , связанные с негомологичным соединением концов, Шварц и др. наблюдали самый высокий уровень репрессии (87%) для Ku80 , когда гибрид dCas9-Mxi1 сравнивали с dCas9 (38%) (Schwartz et al. 2017). Для дальнейшего изучения дизайна слияния dCas9 для оптимальной репрессии Schwartz et al. и Гандер и др. (Gander et al. 2017) также тестировали слияния между dCas9 и Krüppel-ассоциированным блоком (или доменом KRAB) из геномов четвероногих позвоночных (Witzgall et al. 1994). Здесь Schwartz et al. обнаружили сопоставимые уровни обилия транскриптов в порядке 2–3-кратной репрессии для dCas9-KRAB, что также наблюдается для dCas9, в то время как Gander et al. наблюдали примерно 2,5-кратную репрессию для dCas9-KRAB по сравнению с примерно 12-кратной репрессией при использовании dCas9-Mxi1 для контроля экспрессии синтетического промотора на основе CYC1 (Gander et al. 2017). Эти результаты также подтверждаются математическими моделями, предсказывающими, что репрессия только через dCas9 пропускает больше, чем репрессия через dCas9-Mxi1 (Gander et al. 2017). Помимо Mxi1 и KRAB, Gander et al.
2017). Для дальнейшего изучения дизайна слияния dCas9 для оптимальной репрессии Schwartz et al. и Гандер и др. (Gander et al. 2017) также тестировали слияния между dCas9 и Krüppel-ассоциированным блоком (или доменом KRAB) из геномов четвероногих позвоночных (Witzgall et al. 1994). Здесь Schwartz et al. обнаружили сопоставимые уровни обилия транскриптов в порядке 2–3-кратной репрессии для dCas9-KRAB, что также наблюдается для dCas9, в то время как Gander et al. наблюдали примерно 2,5-кратную репрессию для dCas9-KRAB по сравнению с примерно 12-кратной репрессией при использовании dCas9-Mxi1 для контроля экспрессии синтетического промотора на основе CYC1 (Gander et al. 2017). Эти результаты также подтверждаются математическими моделями, предсказывающими, что репрессия только через dCas9 пропускает больше, чем репрессия через dCas9-Mxi1 (Gander et al. 2017). Помимо Mxi1 и KRAB, Gander et al. также тестировали домены репрессии GAL80, LUG, TPLRD1, TUP1 и XTC1 (Flick and Johnston 1990; Edmondson, Smith and Roth 1996; Wu et al. 2001; Травен и др. 2002; Пьер-Жером и др. 2014), при этом LUG и TPLRD1 демонстрируют такой же потенциал репрессии, как и KRAB, в то время как ни слияния GAL80, TUP1 и XTC1 не показали никакой репрессии (Fig. 1a). Точно так же Lian et al. (2017) тестировали варианты репрессорных доменов TUP1, MIG1, CRT1, XTC1 и UME6 (Edmondson, Smith and Roth 1996; Ostling, Carlberg and Ronne 1996; Kadosh and Struhl 1997; Zhang and Reese 2005; Traven et al. 2002). ), и сообщили, что трехкомпонентный домен репрессии, сконструированный из UME6, MIG1 и TUP1, является наиболее успешной конструкцией для dCas9-опосредованной репрессии (до 5 раз более сильной репрессии по сравнению с dCas9-Mxi1), тогда как слияние с dCpf1 не было эффективным. для CRISPRi (Lian et al. 2017).
также тестировали домены репрессии GAL80, LUG, TPLRD1, TUP1 и XTC1 (Flick and Johnston 1990; Edmondson, Smith and Roth 1996; Wu et al. 2001; Травен и др. 2002; Пьер-Жером и др. 2014), при этом LUG и TPLRD1 демонстрируют такой же потенциал репрессии, как и KRAB, в то время как ни слияния GAL80, TUP1 и XTC1 не показали никакой репрессии (Fig. 1a). Точно так же Lian et al. (2017) тестировали варианты репрессорных доменов TUP1, MIG1, CRT1, XTC1 и UME6 (Edmondson, Smith and Roth 1996; Ostling, Carlberg and Ronne 1996; Kadosh and Struhl 1997; Zhang and Reese 2005; Traven et al. 2002). ), и сообщили, что трехкомпонентный домен репрессии, сконструированный из UME6, MIG1 и TUP1, является наиболее успешной конструкцией для dCas9-опосредованной репрессии (до 5 раз более сильной репрессии по сравнению с dCas9-Mxi1), тогда как слияние с dCpf1 не было эффективным. для CRISPRi (Lian et al. 2017).
Помимо слияния репрессорных доменов, в нескольких исследованиях проводилось слияние белков CRISPR с одиночными и множественными доменами активации транскрипции, чтобы обеспечить CRISPR-опосредованную активацию экспрессии генов, называемую CRISPRa (Gilbert et al. 2014). У дрожжей Farzadfard et al. были первыми, кто показал, что dCas9 можно использовать в качестве активатора транскрипции при слиянии с активационным доменом (Farzadfard, Perli and Lu 2013). Здесь они первоначально протестировали dCas9-VP64, направленный либо на смысловую, либо на антисмысловую цепь минимального промотора CYC1 , и обнаружили несколько положений гРНК, обеспечивающих статистически значимое повышение репортерной флуоресценции порядка 1.в 5–3,0 раза (Фарзадфард, Перли и Лу, 2013). Аналогичные кратные изменения наблюдались для dCas9-VP64, нацеленного на промоторы GAL1 и ADE2 (Farzadfard, Perli and Lu 2013; Vanegas, Lehka and Mortensen 2017), в то время как Naranjo et al. сообщили о >100- и >250-кратном увеличении уровней транскриптов при использовании dCas9-VP64 и GAL4-dCas9-VP64, соответственно, для мишени FRM2 (Naranjo et al. 2015). активность под управлением любой из протестированных гРНК.
2014). У дрожжей Farzadfard et al. были первыми, кто показал, что dCas9 можно использовать в качестве активатора транскрипции при слиянии с активационным доменом (Farzadfard, Perli and Lu 2013). Здесь они первоначально протестировали dCas9-VP64, направленный либо на смысловую, либо на антисмысловую цепь минимального промотора CYC1 , и обнаружили несколько положений гРНК, обеспечивающих статистически значимое повышение репортерной флуоресценции порядка 1.в 5–3,0 раза (Фарзадфард, Перли и Лу, 2013). Аналогичные кратные изменения наблюдались для dCas9-VP64, нацеленного на промоторы GAL1 и ADE2 (Farzadfard, Perli and Lu 2013; Vanegas, Lehka and Mortensen 2017), в то время как Naranjo et al. сообщили о >100- и >250-кратном увеличении уровней транскриптов при использовании dCas9-VP64 и GAL4-dCas9-VP64, соответственно, для мишени FRM2 (Naranjo et al. 2015). активность под управлением любой из протестированных гРНК. Помимо тестирования dCas9-VP64 на CRISPRa, Farzadfard et al. также протестировали возможность управления несколькими копиями dCas9-VP64 и, таким образом, настройки активности репортерного промотора. Исходя из этого, авторы наблюдали, что активность репортерного гена увеличивалась до 70 раз при нацеливании dCas9-VP64 максимум на 12 идентичных позиций оператора с использованием одной гРНК (Farzadfard, Perli and Lu 2013). Одно интересное наблюдение, признанное уже на этой ранней стадии CRISPR-опосредованного репрограммирования транскрипции, заключалось в сильном влиянии, оказываемом положением гРНК по отношению к влиянию регуляции, основанной на dCas9.В частности, Farzadfard et al. обнаружили, что, хотя dCas9-VP64 мог служить активатором транскрипции, когда гРНК располагались выше ТАТА-бокса, наблюдалась значительная репрессия активности репортерного гена порядка 2–3 раз, когда слитый белок направлялся в положения, перекрывающиеся или ниже по течению от коробки ТАТА (Фарзадфард, Перли и Лу, 2013).
Помимо тестирования dCas9-VP64 на CRISPRa, Farzadfard et al. также протестировали возможность управления несколькими копиями dCas9-VP64 и, таким образом, настройки активности репортерного промотора. Исходя из этого, авторы наблюдали, что активность репортерного гена увеличивалась до 70 раз при нацеливании dCas9-VP64 максимум на 12 идентичных позиций оператора с использованием одной гРНК (Farzadfard, Perli and Lu 2013). Одно интересное наблюдение, признанное уже на этой ранней стадии CRISPR-опосредованного репрограммирования транскрипции, заключалось в сильном влиянии, оказываемом положением гРНК по отношению к влиянию регуляции, основанной на dCas9.В частности, Farzadfard et al. обнаружили, что, хотя dCas9-VP64 мог служить активатором транскрипции, когда гРНК располагались выше ТАТА-бокса, наблюдалась значительная репрессия активности репортерного гена порядка 2–3 раз, когда слитый белок направлялся в положения, перекрывающиеся или ниже по течению от коробки ТАТА (Фарзадфард, Перли и Лу, 2013). Эффекты, специфичные для позиций гРНК, будут более подробно описаны в разделе «Модуляция активности гРНК».
Эффекты, специфичные для позиций гРНК, будут более подробно описаны в разделе «Модуляция активности гРНК».
В дополнение к однодоменному VP64 белки CRISPR также недавно были успешно слиты с комбинациями активаторов транскрипции, включая VPR, который сконструирован из четырехкратных копий вирусного белка простого герпеса (VP16), трансактивационного домена NF. субъединица p65 -kB (p65AD) и трансактиватор R вируса Эпштейна-Барр (Rta) (Chavez et al. 2016; Динер и Альпер, 2017 г.; Дженсен и др. 2017). Как свидетельствует Chavez et al. , сравнивая уровни экспрессии репортерного гена с использованием гРНК, нацеленных на промоторы дрожжей GAL7 и HED1 , dCas9-VPR опосредовал примерно 100- и 40-кратную активацию соответственно по сравнению со скромным 14- и 8-кратным увеличением, наблюдаемым при использовании dCas9. -VP64 (рис. 1b) (Чавес и др. , 2015 г.). Помимо использования дефицитного по нуклеазе Cas9 из S. pyogenes , Lian et al. систематически тестировали новые CRISPR-опосредованные активаторы транскрипции путем слияния нескольких белков CRISPR с дефицитом нуклеаз с активационными доменами (Lian et al. 2017). Здесь авторы обнаружили, что оптимальный домен активации зависел от белка Cas, протестированного с наиболее эффективным вариантом S. pyogenes dCas9, демонстрирующим до 12-кратной активации активности репортерного гена при слиянии с VPR, в то время как наиболее эффективный Вариант dCpf1 индуцировал до 8-кратной активации активности гена при слиянии с VP64-p65AD (Lian et al. 2017).
систематически тестировали новые CRISPR-опосредованные активаторы транскрипции путем слияния нескольких белков CRISPR с дефицитом нуклеаз с активационными доменами (Lian et al. 2017). Здесь авторы обнаружили, что оптимальный домен активации зависел от белка Cas, протестированного с наиболее эффективным вариантом S. pyogenes dCas9, демонстрирующим до 12-кратной активации активности репортерного гена при слиянии с VPR, в то время как наиболее эффективный Вариант dCpf1 индуцировал до 8-кратной активации активности гена при слиянии с VP64-p65AD (Lian et al. 2017).
Признавая результаты исследований слияния dCas9, несколько групп с тех пор успешно применили dCas9-VPR для CRISPRa у дрожжей (Deaner and Alper 2017; Deaner, Mejia and Alper 2017; Jensen et al. 2017). Несмотря на то, что большинство наблюдаемых активаций находятся в диапазоне от 2 до 10 раз, Deaner et al. наблюдали более чем 160-кратные изменения экспрессии гена NDE2 при сравнении наилучшего положения гРНК для опосредования CRISPRi с использованием dCas9-Mxi1 и наиболее сильного положения гРНК для опосредования CRISPRa с использованием dCas9-VPR (Deaner and Alper 2017). В соответствии с выводами Farzadfard et al. , Deaner и Alper также сообщили о позиционно-специфическом потенциале dCas9-VPR для умеренной репрессии экспрессии генов (Farzadfard, Perli and Lu 2013; Deaner and Alper 2017).
В соответствии с выводами Farzadfard et al. , Deaner и Alper также сообщили о позиционно-специфическом потенциале dCas9-VPR для умеренной репрессии экспрессии генов (Farzadfard, Perli and Lu 2013; Deaner and Alper 2017).
В совокупности несколько исследований показали, что CRISPRi/a у дрожжей приводит к изменениям экспрессии и активности генов в порядке >50-кратного подавления и >100-кратного повышения, при этом dCas9-Mxi1 и dCas9-VPR в настоящее время являются наиболее частыми. принятые регуляторы.В общем, dCas9 является универсальным партнером по слиянию как для доменов активации, так и для доменов репрессии, однако оптимальный выбор регуляторных доменов, которые будут использоваться для репрограммирования транскрипции, может зависеть от белка CRISPR. Это открывает возможности для многофункционального перепрограммирования, опосредованного CRISPR, с использованием ортогональных последовательностей PAM различных белков CRISPR в качестве аналогов для вышестоящих активирующих (UAS) или репрессирующих последовательностей (Lian et al. 2017).
2017).
Регуляция экспрессии генов, кодирующих белки CRISPR
При использовании дефицитных по нуклеазе синтетических регуляторов на основе CRISPR важно понимать, что регуляторный потенциал регуляторов транскрипции неотъемлемо зависит от уровня экспрессии самого регулятора транскрипции, при этом более высокая экспрессия чаще всего обеспечивает наивысший уровень репрессии и/или активации рассматриваемого гена(ов)-мишени (Skjoedt et al. 2016). В соответствии с этим в большинстве исследований на дрожжах используются сильные конститутивные или гликолитические промоторы для управления экспрессией гена, кодирующего dCas9 и его варианты (Gilbert et al. 2013; Lian et al. 2017). Из S. Cerevisiae , это включает в себя TDH4 (или GDP1), TEF1 и PDC1 промоутами для использования в S. Cerevisiae и Kluyveromyces Marxianus (Gilbert et al. 2013; Chavez и другие. 2015; Смит и др. 2016; Намбу-Нишида и др. 2017), тогда как в Y. lipolytica Schwartz et al. использовали ранее сконструированный сильный конститутивный промотор, основанный на укороченном промоторе TEF1 , слитом с восемью копиями 105-п. Blanchin-Roland, Cordero Otero and Gaillardin 1994; Blazeck и др. 2011; Schwartz et al. 2016; Шварц и др. 2017).
2013; Chavez и другие. 2015; Смит и др. 2016; Намбу-Нишида и др. 2017), тогда как в Y. lipolytica Schwartz et al. использовали ранее сконструированный сильный конститутивный промотор, основанный на укороченном промоторе TEF1 , слитом с восемью копиями 105-п. Blanchin-Roland, Cordero Otero and Gaillardin 1994; Blazeck и др. 2011; Schwartz et al. 2016; Шварц и др. 2017).
В дополнение к конститутивной экспрессии dCas9 способность программировать начало регуляции гена-мишени побудила использовать индуцибельную экспрессию dCas9 для условного перепрограммирования транскрипции у дрожжей. При использовании синтетического промотора, первоначально разработанного Ellis et al. , была достигнута индуцируемая галактозой и ангидротетрациклином (aTc) экспрессия dCas9-VP64 (Ellis, Wang and Collins 2009; Farzadfard, Perli and Lu 2013), что позволило добиться 70-кратной индуцируемой экспрессии минимального промотора CYC1 с выходами сравним с активационным потенциалом других обычно используемых эндогенных промоторов GAL1 и CUP1 (Farzadfard, Perli and Lu 2013). Вместе с дополнительным набором свето- и аллостерически регулируемых систем CRISPR/Cas9, описанных в клетках млекопитающих (Gao et al. 2016; Oakes et al. 2016), такая галактозо- и aTc-индуцируемая экспрессия dCas9 и его вариантов позволяет контролировать начало экспрессии целевого гена.
Вместе с дополнительным набором свето- и аллостерически регулируемых систем CRISPR/Cas9, описанных в клетках млекопитающих (Gao et al. 2016; Oakes et al. 2016), такая галактозо- и aTc-индуцируемая экспрессия dCas9 и его вариантов позволяет контролировать начало экспрессии целевого гена.
Модуляция активности гРНК
Рекрутирование эффекторов слитыми с аптамерами гРНК
Неотъемлемая взаимосвязь между dCas9 и гРНК ограничивает dCas9-опосредованное программирование мультигенных генных цепей, основанных на транскрипции, только одним направлением регуляции (т.е. репрессия или активация) на уровне одной клетки. Это не выравнивает сложность и изощренность, лежащие в основе нативных транскрипционных сетей. Однако, по аналогии со слиянием регуляторных доменов с dCas9, инженерия самой гРНК оказалась модульной и настраиваемой платформой для диверсификации не только геномных сайтов-мишеней (исходной последовательности), но и функции CRISPR-опосредованной регуляции транскрипции.
Используя преимущество 3 ΄ -конца гРНК, Zalatan et al. и Киани и др. были первыми, кто сконструировал гРНК с помощью РНК-аптамеров, взаимодействующих с белком (Kiani et al. 2015; Zalatan et al. 2015). У дрожжей это включало gRNAs, которые действительно могли контролировать не только локализацию dCas9 (и Cas9), но также и функцию. В своих основополагающих исследованиях они показали, что слияние РНК-аптамеров с tracr-частью гРНК позволяет связывать РНК-связывающие белки и тем самым контролировать регуляторный потенциал в зависимости от партнера по взаимодействию с белком, прикрепленного к РНК-связывающему белку (рис.2а) (Киани и др. 2015 г.; Залатан и др. 2015 г.). В частности, чтобы преобразовать как специфичность целевой последовательности, так и функцию в эти каркасные РНК (scRNAs), Zalatan et al. тестировали (i) разные аптамеры, (ii) 5 ΄ — по сравнению с 3 ΄ -концевыми слияниями, (iii) различное количество аптамеров, (iv) длину линкера между гРНК 5 ΄ -концом и аптамером, и (v) ортогональность между аптамером и родственными ему партнерами по связыванию РНК. Систематическая характеристика выявила три мощных РНК-связывающих модуля, каждый из которых состоит из аптамера и его партнера по РНК-связывающему белку, слитых либо с доменом активации VP64, либо с доменом репрессии Mxi1 (Fig. 2a). Более того, авторы показали, что несколько аптамеров могут быть введены в одиночные scRNAs, и не наблюдалось перекрестных помех между компонентами РНК-связывающих модулей, что в конечном итоге делает возможной как dCas9-опосредованную активацию, так и регуляцию в отдельных клетках только в зависимости от последовательности семян и кодируемого аптамера. в scRNA(s) (Zalatan et al. 2015). Что наиболее важно, при использовании стратегии scRNA вместе с РНК-связывающими модулями на основе VP64 у дрожжей наблюдалась более чем 50-кратная активация синтетического репортерного промотора по сравнению со скромной 2-3-кратной активацией, наблюдаемой для dCas9-VP64. Используя две разные scRNA для целевой активации генов вместе с репрессией, опосредованной dCas9, Zalatan et al.
Систематическая характеристика выявила три мощных РНК-связывающих модуля, каждый из которых состоит из аптамера и его партнера по РНК-связывающему белку, слитых либо с доменом активации VP64, либо с доменом репрессии Mxi1 (Fig. 2a). Более того, авторы показали, что несколько аптамеров могут быть введены в одиночные scRNAs, и не наблюдалось перекрестных помех между компонентами РНК-связывающих модулей, что в конечном итоге делает возможной как dCas9-опосредованную активацию, так и регуляцию в отдельных клетках только в зависимости от последовательности семян и кодируемого аптамера. в scRNA(s) (Zalatan et al. 2015). Что наиболее важно, при использовании стратегии scRNA вместе с РНК-связывающими модулями на основе VP64 у дрожжей наблюдалась более чем 50-кратная активация синтетического репортерного промотора по сравнению со скромной 2-3-кратной активацией, наблюдаемой для dCas9-VP64. Используя две разные scRNA для целевой активации генов вместе с репрессией, опосредованной dCas9, Zalatan et al. обеспечил синтетический контроль над потоками в точках ветвления пути биосинтеза виолацеина (рис. 2b), в то время как Jensen et al. продемонстрировали комбинаторное перепрограммирование генов мевалонатного и каротиноидного путей с использованием модулей активации MCP:VPR и репрессии PCP:Mxi1, что в конечном итоге привело к значительным изменениям уровней каротиноидов (рис. 2c) (Zalatan et al. 2015; Jensen et al. ). 2017).
обеспечил синтетический контроль над потоками в точках ветвления пути биосинтеза виолацеина (рис. 2b), в то время как Jensen et al. продемонстрировали комбинаторное перепрограммирование генов мевалонатного и каротиноидного путей с использованием модулей активации MCP:VPR и репрессии PCP:Mxi1, что в конечном итоге привело к значительным изменениям уровней каротиноидов (рис. 2c) (Zalatan et al. 2015; Jensen et al. ). 2017).
Рисунок 2.
Дизайн и применение каркасных РНК, контролирующих как геномную целевую последовательность, так и регуляторную функцию. ( a ) Пять примеров каркасных РНК (scRNAs), используемых в дрожжах.ScRNAs представляют собой сконструированные gRNA с РНК-аптамерами, взаимодействующими с белками. Белок и аптамер вместе относятся к РНК-связывающим модулям. Связывающие аптамеры белки MCP, PCP и COM ортогонально взаимодействуют с аптамерами MS2, PP7 и com соответственно. MCP, PCP и COM могут быть слиты с доменами активации или репрессии, что позволяет scRNAs определять целевой локус генома и регуляторную функцию. ( b ) Пример, иллюстрирующий репрограммирование одной клетки экспрессии трех генов, кодирующих часть пути биосинтеза виолацеина.( c ) Пример, иллюстрирующий перепрограммирование одной клетки экспрессии трех генов, кодирующих белки, регулирующие метаболический поток через мевалонатный и каротиноидный пути.
( b ) Пример, иллюстрирующий репрограммирование одной клетки экспрессии трех генов, кодирующих часть пути биосинтеза виолацеина.( c ) Пример, иллюстрирующий перепрограммирование одной клетки экспрессии трех генов, кодирующих белки, регулирующие метаболический поток через мевалонатный и каротиноидный пути.
Рисунок 2.
Дизайн и применение каркасных РНК, контролирующих как геномную целевую последовательность, так и регуляторную функцию. ( a ) Пять примеров каркасных РНК (scRNAs), используемых в дрожжах. ScRNAs представляют собой сконструированные gRNA с РНК-аптамерами, взаимодействующими с белками. Белок и аптамер вместе относятся к РНК-связывающим модулям.Связывающие аптамеры белки MCP, PCP и COM ортогонально взаимодействуют с аптамерами MS2, PP7 и com соответственно. MCP, PCP и COM могут быть слиты с доменами активации или репрессии, что позволяет scRNAs определять целевой локус генома и регуляторную функцию. ( b ) Пример, иллюстрирующий репрограммирование одной клетки экспрессии трех генов, кодирующих часть пути биосинтеза виолацеина. ( c ) Пример, иллюстрирующий перепрограммирование одной клетки экспрессии трех генов, кодирующих белки, регулирующие метаболический поток через мевалонатный и каротиноидный пути.
( c ) Пример, иллюстрирующий перепрограммирование одной клетки экспрессии трех генов, кодирующих белки, регулирующие метаболический поток через мевалонатный и каротиноидный пути.
Таким образом, инженерия гРНК в scРНК предлагает многонаправленное перепрограммирование экспрессии генов на основе CRISPR и имеет особое значение для изучения и улучшения нашего понимания полигенных признаков и комбинированных эффектов ключевых точек ветвления метаболического пути.
Регуляция экспрессии гРНК
Было показано, что уровни экспрессии гРНК коррелируют с эффективностью CRISPR/Cas9-опосредованной инженерии генома в клетках млекопитающих (Hsu et al. 2013). Чтобы соответствовать стехиометрии dCas9 или dCpf1, экспрессируемых из сильных конститутивных промоторов полимеразы II (см. раздел «Регулирование экспрессии генов, кодирующих белки CRISPR»), были тщательно исследованы способы оптимизации экспрессии гРНК и scРНК. Как правило, промоторы полимеразы III используются для управления экспрессией гРНК, поскольку промоторы РНК-полимеразы II добавляют дополнительные нуклеотиды к 5΄- и 3΄-концам гРНК и, таким образом, считается, что они прерывают функцию гРНК (Yoshioka et al. 2015). Первоначально промоторы полимеразы III SNR52 и RPR1 были адаптированы для конститутивной доставки гРНК в дрожжи (рис. 3) (DiCarlo et al. 2013; Farzadfard, Perli and Lu 2013; Gilbert et al. 2013). ). В частности, использование промотора SNR52 широко используется из-за его нативных сайтов расщепления транскриптов, что приводит к вырезанию гРНК из первичных транскриптов (DiCarlo et al. 2013). Затем, чтобы обеспечить большую гибкость в дизайне и силе экспрессии гРНК, в двух исследованиях сразу после вышеупомянутых исследований по конститутивной доставке гРНК было протестировано слияние саморасщепляющегося дельта-вируса гепатита (HDV) и рибозимов типа «голова-молот» с гРНК, таким образом возможность редактирования генома, полученного из промоторов полимеразы II (Gao and Zhao 2014; Ryan et al.
Как правило, промоторы полимеразы III используются для управления экспрессией гРНК, поскольку промоторы РНК-полимеразы II добавляют дополнительные нуклеотиды к 5΄- и 3΄-концам гРНК и, таким образом, считается, что они прерывают функцию гРНК (Yoshioka et al. 2015). Первоначально промоторы полимеразы III SNR52 и RPR1 были адаптированы для конститутивной доставки гРНК в дрожжи (рис. 3) (DiCarlo et al. 2013; Farzadfard, Perli and Lu 2013; Gilbert et al. 2013). ). В частности, использование промотора SNR52 широко используется из-за его нативных сайтов расщепления транскриптов, что приводит к вырезанию гРНК из первичных транскриптов (DiCarlo et al. 2013). Затем, чтобы обеспечить большую гибкость в дизайне и силе экспрессии гРНК, в двух исследованиях сразу после вышеупомянутых исследований по конститутивной доставке гРНК было протестировано слияние саморасщепляющегося дельта-вируса гепатита (HDV) и рибозимов типа «голова-молот» с гРНК, таким образом возможность редактирования генома, полученного из промоторов полимеразы II (Gao and Zhao 2014; Ryan et al. 2014). Гао и Чжао были первыми, кто обратил внимание на использование фланкированных рибозимами гРНК, позволяющих использовать промоторы pol II для управления экспрессией пре-гРНК, предназначенных для самокатализируемого процессинга (рис. 3) (Gao and Zhao 2014; Zhang et al. 2017). В дополнение к этому исследованию с использованием промотора ADh2 pol II для управления экспрессией гРНК, фланкированных минимальной головкой молотка 5 ΄ и рибозимами 3 ΄ HDV на концах 5 ΄ и 3 ΄ соответственно. , Райан и др. тестировали в общей сложности одиннадцать промоторов pol III для доставки функциональных гРНК (Ryan et al. 2014). Исследование пришло к выводу, что, хотя промоторы тРНК были совместимы со слиянием рибозима HDV, обеспечивая почти 100% инженерную эффективность, промотор snoRNA SNR52 был единственным промотором, не относящимся к тРНК, который выравнивал такую эффективность при слиянии с рибозимом HDV (Ryan et al.
 ). 2014). Эти результаты добавляются к более позднему эталону синтетических промоторов слияния pol III, промоторов pol II (управляющих экспрессией фланкированных рибозимом гРНК (RGR)) и промоторов pol III, не являющихся тРНК, управляющих экспрессией гРНК в S.cerevisiae и Y. lipolytica (рис. 3) (Schwartz et al. 2016; Deaner, Mejia and Alper 2017). Здесь было обнаружено, что уровни экспрессии в значительной степени коррелируют с инженерной эффективностью различных конструкций, при этом синтетические промоторы слияния между укороченными промоторами pol III и промоторами тРНК давали самые высокие оценки (> 90%) в Y. lipolytica , в то время как сильные Подход pol II TEF1-RGR продуцировал почти в 4 раза больше гРНК по сравнению с SNR52, что также коррелирует с более сильным регуляторным потенциалом (Schwartz et al. 2016; Динер, Мехиа и Альпер, 2017 г.). Также Гандер и др. использовали минимальный промотор CYC1 для создания набора промоторов гРНК-чувствительной полимеразы II (pGRR), управляющих экспрессией RGR (Gander et al.
). 2014). Эти результаты добавляются к более позднему эталону синтетических промоторов слияния pol III, промоторов pol II (управляющих экспрессией фланкированных рибозимом гРНК (RGR)) и промоторов pol III, не являющихся тРНК, управляющих экспрессией гРНК в S.cerevisiae и Y. lipolytica (рис. 3) (Schwartz et al. 2016; Deaner, Mejia and Alper 2017). Здесь было обнаружено, что уровни экспрессии в значительной степени коррелируют с инженерной эффективностью различных конструкций, при этом синтетические промоторы слияния между укороченными промоторами pol III и промоторами тРНК давали самые высокие оценки (> 90%) в Y. lipolytica , в то время как сильные Подход pol II TEF1-RGR продуцировал почти в 4 раза больше гРНК по сравнению с SNR52, что также коррелирует с более сильным регуляторным потенциалом (Schwartz et al. 2016; Динер, Мехиа и Альпер, 2017 г.). Также Гандер и др. использовали минимальный промотор CYC1 для создания набора промоторов гРНК-чувствительной полимеразы II (pGRR), управляющих экспрессией RGR (Gander et al. 2017). В их исследовании была сконструирована библиотека из 400 pGRR с двумя целевыми сайтами вместе с 20 RGR, в общей сложности 8000 NOR (либо один, либо оба) логических вентилей, включая как конститутивные, так и индуцируемые эстрадиолом промоторы pol II, чтобы управлять экспрессией RGR, в конечном итоге уступая до 12-кратной регуляции с помощью одиночных репортерных промоторов, контролируемых гРНК (McIsaac et al. 2013; Гандер и др. 2017).
2017). В их исследовании была сконструирована библиотека из 400 pGRR с двумя целевыми сайтами вместе с 20 RGR, в общей сложности 8000 NOR (либо один, либо оба) логических вентилей, включая как конститутивные, так и индуцируемые эстрадиолом промоторы pol II, чтобы управлять экспрессией RGR, в конечном итоге уступая до 12-кратной регуляции с помощью одиночных репортерных промоторов, контролируемых гРНК (McIsaac et al. 2013; Гандер и др. 2017).
Рисунок 3.
Экспрессия направляющих РНК (гРНК), каркасных РНК (scRNA) и фланкированных рибозимами гРНК. Сообщается о примерах нативных, гибридных и сконструированных промоторов, которые управляют экспрессией гРНК, scРНК и фланкированных рибозимами гРНК у дрожжей.
Рисунок 3.
Экспрессия направляющих РНК (гРНК), каркасных РНК (скРНК) и фланкированных рибозимом гРНК. Сообщается о примерах нативных, гибридных и сконструированных промоторов, которые управляют экспрессией гРНК, scРНК и фланкированных рибозимами гРНК у дрожжей.
Помимо нативных промоторов pol III и индуцируемых pol II, контролирующих экспрессию гРНК и RGR, другие группы использовали сконструированный нативный промотор RPR1 pol III, чтобы включить сайт связывания TetO для aTc-индуцируемой депрессии экспрессии гРНК, когда коэкспрессируя конститутивно экспрессируемый репрессор TetR, тем самым обеспечивая нарушения экспрессии в 2–20 раз (рис. 3) (Farzadfard et al. 2013; Smith et al. 2016; Jensen et al. 2017; Феррейра и др. 2018). Интересно, что в исследовании Ferreira et al. , три кассеты гРНК были экспрессированы из одного сконструированного промотора RPR1 pol III, а затем эндорибонуклеаза Csy4 была использована для расщепления транскрипта на субэлементы и усиления опосредованной dCas9-VPR экспрессии HMG1 , OLE1 и ACS1. промоторов примерно в 2 раза (Ferreira et al. 2018). Этот элегантный подход позволяет легко обойти необходимость повторного использования одного и того же промотора или необходимости использования нескольких разных промоторов при попытке перепрограммировать транскрипцию нескольких генов (рис. 3).
3).
Таким образом, несмотря на то, что нативные промоторы pol III изначально были предпочтительной конструкцией, простая разработка промоторов pol II, управляющих экспрессией саморасщепляющихся RGR, позволяет контролировать перепрограммирование генома, основанное практически на любом промоторе pol II (Zhang и др. , 2017 г.). Кроме того, стерические препятствия, предлагаемые индуцируемыми репрессорами, могут быть использованы для создания промоторов pol III для функциональной, своевременной и эффективной доставки гРНК.
Множественные гРНК для перепрограммирования геномных функций
Регулирование нативных и синтетических промоторов с помощью эндогенных или сконструированных факторов транскрипции зависит от их способности связывать родственные ТФ-связывающие сайты в таких промоторах (Khalil et al. 2012). По аналогии с этим и как упоминалось ранее (раздел «Регулирование экспрессии генов, кодирующих белки CRISPR»), Farzadfard et al. показали, что синергетические эффекты на регуляцию транскрипции могут наблюдаться при использовании нескольких гРНК, направляющих dCas9-опосредованный контроль промоторов-мишеней. Например, каждая из двух отдельных гРНК вызывала 2-кратную репрессию, тогда как их комбинация показала 7-кратную репрессию. Более того, Farzadfard et al. также тестировали управление несколькими копиями dCas9-VP64 и тем самым регулировали активность репортерного промотора, и таким образом наблюдали, что активность репортерного гена увеличивалась до 70 раз при нацеливании dCas9-VP64 максимум на 12 позиций (Farzadfard, Perli and Lu 2013).Точно так же Gilbert et al. протестировали 7× гРНК на промоторе TetO , продемонстрировав самую высокую из когда-либо зарегистрированных репрессий активности репортерного гена при использовании dCas9-Mxi1 (153×, рис. 1), в то время как Deaner et al. использовали двойные гРНК, экспрессированные из промоторов SNR52- и TEF1 , чтобы усилить регуляторный потенциал dCas9-VPR (Gilbert et al.
показали, что синергетические эффекты на регуляцию транскрипции могут наблюдаться при использовании нескольких гРНК, направляющих dCas9-опосредованный контроль промоторов-мишеней. Например, каждая из двух отдельных гРНК вызывала 2-кратную репрессию, тогда как их комбинация показала 7-кратную репрессию. Более того, Farzadfard et al. также тестировали управление несколькими копиями dCas9-VP64 и тем самым регулировали активность репортерного промотора, и таким образом наблюдали, что активность репортерного гена увеличивалась до 70 раз при нацеливании dCas9-VP64 максимум на 12 позиций (Farzadfard, Perli and Lu 2013).Точно так же Gilbert et al. протестировали 7× гРНК на промоторе TetO , продемонстрировав самую высокую из когда-либо зарегистрированных репрессий активности репортерного гена при использовании dCas9-Mxi1 (153×, рис. 1), в то время как Deaner et al. использовали двойные гРНК, экспрессированные из промоторов SNR52- и TEF1 , чтобы усилить регуляторный потенциал dCas9-VPR (Gilbert et al. 2013; Deaner, Mejia and Alper 2017). Напротив, Schwartz et al. также использовали две гРНК в области TSS –120 п.н., чтобы проверить, усиливает ли это репрессию Ku70 и Ku80 , однако они обнаружили лишь незначительные эффекты от использования двух гРНК по сравнению с возмущениями, наблюдаемыми при использовании только одной гРНК (Schwartz ). и другие. 2017).
2013; Deaner, Mejia and Alper 2017). Напротив, Schwartz et al. также использовали две гРНК в области TSS –120 п.н., чтобы проверить, усиливает ли это репрессию Ku70 и Ku80 , однако они обнаружили лишь незначительные эффекты от использования двух гРНК по сравнению с возмущениями, наблюдаемыми при использовании только одной гРНК (Schwartz ). и другие. 2017).
В совокупности, как и в нативных и других синтетических сетях регуляции транскрипции, количество регуляторов, привязанных к целевому регулону, представляет собой модульный клапан для настройки воздействия перепрограммирования, опосредованного CRISPR-dCas9. Тем не менее, использование нескольких гРНК должно быть тщательно разработано с особым вниманием к положению существующих регуляторных элементов и нуклеосом, чтобы настроить регуляторный потенциал путем простого увеличения количества гРНК, нацеленных на такие области (см. позиционирование и доступность хроматина»).
Смещение цепи в сравнении с нормативным потенциалом
Большое внимание привлекло механистическое понимание CRISPRi в отношении позиционирования гРНК. Первоначально основной механизм dCas9-опосредованной репрессии транскрипции был выяснен с использованием NET-seq в Escherichia coli (Churchman and Weissman 2011; Qi et al. 2013). В E. coli , Qi et al. определили, что гРНК вызывают сильную паузу транскрипции выше локуса-мишени гРНК на нематричной цепи, что привело к гипотезе о том, что физическое столкновение между удлиняющейся РНК-полимеразой и комплексом dCas9:гРНК приводит к блокированию транскрипции (Qi et al. 2013). Однако у дрожжей Farzadfard et al. были первыми, кто показал, что размещение гРНК в сходных положениях ниже TSS, но на разных цепях промотора, оказывает сходное негативное влияние на экспрессию генов. Более того, размещение gRNAs на любой из цепей выше TATA-бокса и TSS приводит к аналогичной dCas9-VP64-опосредованной активации генов (Farzadfard, Perli and Lu 2013). Точно так же Gilbert et al. позже сообщили, что целевая цепь ДНК и содержание гуанин-цитозина в гРНК не были определяющими факторами для успешного CRISPRi в их исследовании (Gilbert et al.
Первоначально основной механизм dCas9-опосредованной репрессии транскрипции был выяснен с использованием NET-seq в Escherichia coli (Churchman and Weissman 2011; Qi et al. 2013). В E. coli , Qi et al. определили, что гРНК вызывают сильную паузу транскрипции выше локуса-мишени гРНК на нематричной цепи, что привело к гипотезе о том, что физическое столкновение между удлиняющейся РНК-полимеразой и комплексом dCas9:гРНК приводит к блокированию транскрипции (Qi et al. 2013). Однако у дрожжей Farzadfard et al. были первыми, кто показал, что размещение гРНК в сходных положениях ниже TSS, но на разных цепях промотора, оказывает сходное негативное влияние на экспрессию генов. Более того, размещение gRNAs на любой из цепей выше TATA-бокса и TSS приводит к аналогичной dCas9-VP64-опосредованной активации генов (Farzadfard, Perli and Lu 2013). Точно так же Gilbert et al. позже сообщили, что целевая цепь ДНК и содержание гуанин-цитозина в гРНК не были определяющими факторами для успешного CRISPRi в их исследовании (Gilbert et al. 2014). Наконец, в более позднем исследовании, приняв подход с гораздо большей библиотекой гРНК для определения химико-генетического взаимодействия, Smith et al. сконструировал 383 гРНК в диапазоне от +500 п.н. до –500 п.н. области TSS пяти генов (Smith et al. 2016). Здесь авторы не обнаружили смещения цепи в отношении эффективности гРНК в тестируемом окне размером 1 т.п.н.
2014). Наконец, в более позднем исследовании, приняв подход с гораздо большей библиотекой гРНК для определения химико-генетического взаимодействия, Smith et al. сконструировал 383 гРНК в диапазоне от +500 п.н. до –500 п.н. области TSS пяти генов (Smith et al. 2016). Здесь авторы не обнаружили смещения цепи в отношении эффективности гРНК в тестируемом окне размером 1 т.п.н.
В соответствии с этими выводами недавно было дополнительно выяснено, что, в отличие от результатов CRISPRi в E.coli (Qi et al. 2013), dCas9 в дрожжах может действовать не как простой механизм блокирования транскрипции для РНК-полимеразы специфичным для цепи образом, а, скорее, комплекс гРНК:dCas9 поддерживает образование пермиссивные образования транскриптов, включая преждевременную терминацию и образование нового транскрипта как в смысловой, так и в антисмысловой ориентации (Howe et al. 2017). В совокупности это подчеркивает, что дрожжи не только сопротивляются потенциальному смещению цепей CRISPRi, но и что выводы, сделанные в исследованиях CRISPRi, должны учитывать целостность целевых транскриптов.
Позиционные эффекты гРНК
В отличие от исследований потенциальных эффектов, специфичных для цепей, существуют гораздо более убедительные доказательства из больших наборов данных о позиционно-специфических эффектах гРНК в промоторах.
В целом, гРНК, нацеленные на область выше ТАТА-бокса и TSS, в значительной степени коррелируют с активацией генов, опосредованной как dCas9-VP64, так и dCas9-VPR, в то время как расположение вариантов dCas9 ниже или в непосредственной близости от ТАТА-боксов отрицательно влияет на экспрессию генов (Farzadfard, Perli and Lu 2013; Deaner and Alper 2017).Например, нацеливание dCas9-VP64 на положение выше TATA-бокса обеспечивало почти 5-кратную активацию минимального промотора GAL1 , в то время как нацеливание гРНК на TATA-бокс или элемент kozak ниже него приводило к CRISPRi, вероятно, из-за интерференции. с комплексом инициации транскрипции, как также наблюдал Deaner et al. при использовании dCas9-VPR для CRISPRi (рис. 1) (Farzadfard, Perli and Lu, 2013; Deaner, Mejia and Alper, 2017). Кроме того, Deaner и Alper представили подробное исследование систематического тестирования чувствительности к возмущению ферментов (STEPS) путем размещения гРНК в окне ~0–750 п.н. выше TATA-бокса различных промоторов нативных дрожжей.Наблюдая за изменениями в экспрессии генов, поскольку dCas9-Mxi1 расположен дальше от блока TATA, а dCas9-VPR расположен ближе к блоку TATA, авторы смогли вывести карты чувствительности к потоку, нанеся изменения в образовании глицерина в зависимости от Градуированная экспрессия 5 генов (Динер и Альпер, 2017). С точки зрения применения авторы использовали STEPS, чтобы показать, что уровни экспрессии генов GPD1 и TPI1 положительно и отрицательно коррелируют с титрами глицерина соответственно.В конечном итоге эти опросы привели к простой стратегии сверхэкспрессии GPD1 / GPP1 , дающей более чем 5-кратное увеличение титров глицерина (4,89–28,0 г/л). Аналогичным образом, использование STEPS на пяти ключевых генах пентозофосфатного пути для увеличения потока через путь ароматических аминокислот привело к примерно 8-кратному увеличению титров 3-DHS (до 126,4 г/л) на фоне делеции zwf1 (Deaner and Alper 2017). ).
Вышеупомянутые исследования эффектов положения гРНК в значительной степени подтверждаются другим недавним исследованием.Здесь Smith et al. использовали CRISPRi на основе dCas9-Mxi1 для тестирования примерно 1000 гРНК, направленных против 20 генов, уровни экспрессии которых, по прогнозам, влияют на чувствительность к специфическим ингибиторам роста (Smith et al. 2016). Здесь авторы обнаружили, что медианный направляющий эффект для dCas9-Mxi1 был максимальным в окне от –200 п.н. до TSS, в то время как гРНК, расположенные вне окна –200 п.н. до TSS, только в некоторых случаях могли эффективно репрессировать транскрипцию, но менее эффективно ( Смит и др. 2016). Эти результаты отличаются от исследований, проведенных на клетках млекопитающих, в которых область от –50 до +300 по отношению к TSS оказалась наиболее эффективной для CRISPRi (Gilbert et al. 2014). Тем не менее, для дрожжей Smith et al. разработал инструмент для дизайна гРНК (http://lp2.github.io/yeast-crispri/), принимая во внимание как положение в геноме, доступность хроматина (раздел «Положение нуклеосом и доступность хроматина»), нуклеосомы (раздел «Положение нуклеосом и доступность хроматина»), длина и последовательность гРНК (раздел «специфичность и длина гРНК»), а также занятость транскрипционным фактором сайта-мишени (раздел «Другие релевантные признаки — активность базального промотора, интерференция связывания ТФ и вторичная структура РНК» .) (Смит и др. 2016 г.). Основываясь на этих и других выводах, Schwartz et al. идентифицировали гРНК для эффективной репрессии экспрессии генов у Y. lipolytica (Schwartz et al. 2017). В крупнейшем на сегодняшний день исследовании Smith et al. нацелен на опосредованную dCas9-Mxi1 репрессию более 1500 генов, необходимых для роста (Smith et al. 2017). Проанализировав > 9000 штаммов, содержащих уникальную гРНК с проверенной последовательностью, авторы усовершенствовали свои более ранние результаты (Smith et al. 2016), теперь выделяя положения гРНК в области между TSS и примерно 125 п.н. выше TSS, которые особенно эффективны для опосредованной CRISPR репрессии (Smith et al. 2017).
При этом, несмотря на то, что Jensen et al. нацелены на 88 гРНК в окне от –200 п.н. до TSS 12 нативных промоторов дрожжей, авторы обнаружили, что несколько гРНК не функционируют при использовании dCas9-VPR и dCas9-Mxi1 для перепрограммирования транскрипции (Jensen et al. 2017).
Обобщение позиционирования гРНК относительно TATA и TSS предлагает простую настраиваемую и портативную стратегию для нарушения активности экспрессии генов как для CRISPRi, так и для CRISPRa, хотя конкретное позиционирование должно также учитывать другие локальные стерические и регуляторные особенности эукариотических промоторов (см. разделы «Положение нуклеосом и доступность хроматина» и «специфичность и длина гРНК»).
Позиционирование нуклеосом и доступность хроматина
Было показано, чтонуклеосомы эффективно мешают действию ДНК-связывающих регуляторов транскрипции (Griesenbeck et al. 2003; Mao et al. 2011). Системы CRISPR, основанные на связывании ДНК, широко использовались в царстве эукариот, но, в отличие от бактерий, ДНК у эукариот в значительной степени закручена вокруг гистонов с образованием нуклеосом, что делает эукариотическую ДНК более плотно упакованной и менее доступной для других ДНК-связывающих белков. Рандо и Чанг, 2009 г.; Рандо и Уинстон, 2012 г.).Как было рассмотрено выше, гРНК, нацеленные на один и тот же промотор, могут по-разному влиять на транскрипцию (Smith et al. 2016), даже близко расположенные гРНК могут иметь разную эффективность, не строго коррелирующую с их расстоянием от TSS (Farzadfard, Perli and Lu 2013; Jensen et al. 2017; Vanegas, Lehka and Mortensen 2017). Это привело Smith et al. , чтобы выяснить, может ли доступность хроматина и расположение нуклеосом также влиять на эффективность направляющей для dCas9-опосредованной регуляции транскрипции.По аналогии с факторами транскрипции, канонически связывающими свободную от нуклеосом ДНК в промоторах, имеющих решающее значение для регуляции экспрессии генов, Smith et al. воспользовались результатами исследования Schep et al. , в котором они идентифицировали высокоструктурированный образец длин фрагментов ДНК и положения вокруг нуклеосом у дрожжей с использованием анализа доступного для транспозазы хроматина (ATAC-seq). Используя данные ATAC-seq вместе с другими наборами данных о положении нуклеосом по всему геному (Lee et al. 2007), Smith и др. обнаружили положительную корреляцию между эффективностью направляющей и показателями доступности хроматина в окне TSS от -400 п.н. до TSS +400 п.н. Несмотря на то, что исследования показали, что расположение гРНК ниже TSS может быть эффективным для перепрограммирования транскрипции (Farzadfard, Perli and Lu 2013; Deaner and Alper 2017), Smith et al. при тестировании сотен гРНК наблюдали, что взаимосвязь между эффективностью направляющей и плотностью чтения ATAC-seq распространяется на область, обычно занятую нуклеосомами, ниже по течению от TSS (Yuan et al. 2005 г.; Ли и др. 2007; Zaugg and Luscombe 2012), подтверждая мнение о том, что эффективность гРНК не определяется в строгом смысле ее близостью к TSS. Эти наблюдения согласуются с биохимическими исследованиями, показывающими, что Cas9 и dCas9 не могут стабильно взаимодействовать с PAM, когда они расположены в ядре нуклеосомы, что указывает на то, что доступность PAM является критическим фактором, определяющим активность белка CRISPR с дефицитом нуклеазы (Hinz, Laughery and Wyrick 2015; Исаак и др. 2016), что еще раз подтверждает наблюдение, что определение того, какие области-мишени с низкой занятостью нуклеосом и высокой доступностью хроматина, вероятно, будут более эффективными (Smith et al. 2016, 2017). Кроме того, в нескольких сообщениях о клетках человека подчеркивается, что расположение эффективных гРНК для dCas9-опосредованной репрессии транскрипции коррелирует с хроматиновыми метками, связанными с активной транскрипцией и открытым хроматином (h4K27ac, h4K9ac, h4K4me3, h4K4me2 и h4K79me2) (Horlbeck et al. 2016; Радзишевская и др. 2016).
Взятые вместе, биохимические данные и данные in vivo позволяют предположить, что стратегии дизайна гРНК должны избегать нацеливания на гРНК вблизи ядра нуклеосомы. Более того, поскольку существует несколько наборов данных о крупномасштабных картах позиционирования нуклеосом и доступности ДНК (Jiang and Pugh 2009; Schep et al. 2015), разработка будущих инструментов компьютерного проектирования для создания специфических и высокоэффективных гРНК должна оцениваться. включение таких наборов данных при выводе выбора гРНК.
Специфичность и длина гРНК
Длина гРНК является решающим фактором для специфичности мишени для нуклеазного Cas9, при этом минимальная длина нуклеазной активности составляет 17 нуклеотидов (Fu et al. 2014b). Для CRISPRi и CRISPRa в нескольких исследованиях оценивалось влияние укороченных гРНК по сравнению с полноразмерными 20-нуклеотидными спейсерными областями гРНК. Первоначально Ци и соавт. обнаружили, что для CRISPRi самая сильная репрессия наблюдалась при использовании полноразмерных гРНК, что подтверждается Kiani et al., которые обнаружили, что активация, опосредованная dCas-VPR, увеличивается с 2- до 100-кратной активации, когда длина семени сдвигается с 8 до 20 нуклеотидов (Kiani et al. 2015). Аналогично, у дрожжей Smith et al. обнаружили, что несовпадения, расположенные в семенной области, расположенной 1–10 относительно PAM, плохо переносятся как полноразмерными, так и укороченными гРНК (Smith et al. 2016), что также согласуется с результатами, полученными при нацеливании на Cas9. in vitro и в клетках млекопитающих (Hsu et al. 2013; Ву и др. 2014; Фу и др. 2014a), а также наблюдение, что всего лишь одного несовпадения пар оснований достаточно для перенаправления нацеливания dCas9 в дрожжах (Farzadfard, Perli and Lu 2013).
В целом, выводы, сделанные на основе этих исследований, показывают, что усечение гРНК снижает эффективность опосредованной CRISPR-dCas9 регуляции транскрипции как в отношении идеально совпадающих, так и не полностью совпадающих последовательностей-мишеней по сравнению с полноразмерными гРНК из 20 нуклеотидов (Kiani et al. 2015; Смит и др. 2016), хотя существует некоторая степень гибкости в конструировании дистальных положений гРНК, которые можно учитывать при конструировании гРНК, нацеленных на промоторные области, плотные в нуклеосомах и вышестоящих активирующих последовательностях.
Другие важные признаки — активность базального промотора, интерференция связывания TF и вторичная структура РНК
В предыдущих разделах некоторые принципы проектирования выделяются как имеющие особое значение для эффективного перепрограммирования транскрипции, опосредованного CRISPR.Для гРНК это включает (i) положительную корреляцию между уровнем экспрессии гРНК и инженерной эффективностью (Schwartz et al. 2016; Deaner et al. 2017), (ii) нацеливание гРНК на окно между –125 п.н. выше по течению TSS и TSS для CRISPRi и (iii) расположение гРНК в бедных нуклеосомами областях промоторов-мишеней (Smith et al. 2016, 2017). В дополнение к этим критериям проектирования следует упомянуть несколько дополнительных исследований для разработки оптимального CRISPR-опосредованного зондирования функции генома.
Во-первых, при выборе интересующих генов стоит учитывать наблюдаемую обратную зависимость между базовыми уровнями экспрессии интересующих генов и относительными нарушениями экспрессии, которые могут быть достигнуты с помощью dCas9-опосредованного перепрограммирования (т.е. высокая базальная экспрессия часто может быть лишь незначительной). активируется и наоборот) (Чавес и др. 2015 г.; Дженсен и др. 2017 г.). В соответствии с этим, другой интересующий фактор связан с регулирующей организацией целевого промотора(ов).Об использовании dCas9 для блокирования связывания ДНК синтетического регулятора транскрипции rTA на синтетическом репортерном промоторе TetON-Venus Gilbert et al. обнаружили, что 115-кратное подавление rtTA-индуцированной активации может быть достигнуто при совместной экспрессии dCas9 и гРНК, что позволяет предположить, что dCas9 может стерически конкурировать с факторами транскрипции, в противном случае контролирующими регуляцию целевого промотора, что указывает на возможность использования CRISPRi и CRISPRa. для определения регуляторных функций вышестоящих активирующих и вышестоящих репрессивных последовательностей (Gilbert et al. 2013). Однако, исходя из своего крупномасштабного библиотечного подхода, Smith et al. (2016) обнаружили лишь небольшое количество случаев, когда перекрытие с сайтом связывания активатора транскрипции коррелировало с повышенной эффективностью CRISPRi, что указывает на то, что этот параметр конструкции может зависеть от нативного регуляторного контекста целевых промоторов. В связи с этим Jensen et al. продемонстрировал опосредованное CRISPR повышение и понижение активности гена OLE1 в течение 48 часов, что коррелирует с количественным анализом с временным разрешением, демонстрирующим, что OLE1 сильно экспрессируется в течение ранней фазы до средней экспоненциальной фазы и подавляется от поздняя экспоненциальная фаза (Jensen et al. 2017). Наконец, еще один важный принцип проектирования гРНК, который следует упомянуть, исходит из ранее упомянутого крупномасштабного исследования CRISPRi, проведенного Smith et al. (2017). Здесь авторы выявили значительную корреляцию между энергией фолдинга в ккал/моль для предсказанной структуры РНК (лидер, целевая последовательность гРНК из 20 нуклеотидов и структурная часть) гРНК и эффективностью гРНК (т. е. больше фолдинга, меньшая эффективность) (Smith и др. , 2017 г.).
В совокупности были выяснены многочисленные параметры дизайна для оптимальной CRISPR-опосредованной регуляции транскрипции.Некоторые из параметров определены в ходе крупномасштабных исследований и считаются не зависящими от генов. Аналогичным образом, как видно из нескольких исследований, CRISPR-опосредованный регуляторный потенциал промоторов-мишеней может поддерживаться в течение длительного времени (Deaner and Alper, 2017; Jensen et al. , 2017), что подчеркивает надежность и ортогональность технологии.
ПРОГНОЗ
Как видно из предыдущих разделов, при использовании CRISPR для исследования функций генома с помощью CRISPRi и CRISPRa необходимо учитывать множество соображений.Тем не менее, для нарушений транскрипции, по сравнению с другими методами, такими как РНКи, gTEM и направленная сверхэкспрессия, CRISPRi/a предлагает простой дизайн, программируемое РНК-опосредованное нацеливание и регуляторное направление как отдельных, так и нескольких генов на уровне одной клетки. Это мощно и использует природу многофакторной нативной регуляции транскрипции для нарушений транскрипции. Действительно, для перепрограммирования транскрипции подходы на основе dCas9 использовались для быстрого анализа динамики метаболических путей и выяснения ограничивающих скорость ферментативных стадий без необходимости редактирования генома (Zalatan et al. 2015; Динер и Альпер, 2017 г.; Дженсен и др. 2017). Кроме того, отдельные наборы экспериментов по трансформации (мультиплексы) могут быть легко реализованы, а однократный синтез наборов гРНК позволяет быстро проходить через повторяющиеся инженерные циклы, а именно путем быстрой оценки комбинаторных эффектов возмущений экспрессии для определения первичных и вторичных мишеней. которые не могли быть известны априори из нарушений экспрессии одного гена.
Однако, несмотря на то, что несколько белков CRISPR и версий гРНК были протестированы в крупномасштабных исследованиях на дрожжах, относительные изменения экспрессии, наблюдаемые при использовании dCas9-опосредованной регуляции транскрипции, по-прежнему часто наблюдаются как минимум на порядок меньше наблюдаемых для усилий по перепрограммированию бактерий и млекопитающих, часто в 100–20 000 раз (Qi et al. 2013; Чавес и др. 2015), тогда как самые высокие изменения транскриптов, зарегистрированные у дрожжей, примерно в 100–250 раз (Gilbert et al. 2013; Chavez et al. 2015; Naranjo et al. 2015). Для улучшения регуляторного потенциала CRISPR-dCas9 у дрожжей и дальнейшего усиления набора инструментов, доступных для исследования функций генома, по-прежнему существует потребность в дальнейшей разработке технологий перепрограммирования.
Недавно сообщалось об одной новой в своем классе технологии CRISPR, имеющей отношение к функциональным геномным исследованиям, с использованием ортологов РНКазы Cas13 с дефицитом нуклеаз из систем CRISPR-Cas типа VI, которые могут управляться одноэффекторными гРНК для нацеливания на большее количество более 70% посттранскрипционного нокдауна экспрессии генов в клетках млекопитающих и растений с высокой специфичностью к мишени (Abudayyeh et al. 2017; Кокс и др. 2017). Кроме того, лаборатория Чжана показала, что dCas13 может быть слит с ферментами семейства аденозиндезаминаз, действующих на РНК (ADAR), и тем самым позволяет редактировать РНК (Cox et al. 2017). Таким образом, считается, что dCas13, нацеленный на РНК, продвигает функциональную геномику на посттранскрипционном уровне, поддерживая функциональные исследования, например. Варианты сплайсинга мРНК, редактирование оснований на уровне РНК и выяснение процессинга мРНК с помощью вариантов dCas13, слитых с регуляторными доменами, аналогично принципам конструирования вариантов dCas9.
Кроме того, хотя это и отличается от CRISPR, следует отметить, что Barbieri et al. недавно сообщили, что подавление механизмов репарации ДНК дрожжей и замедление репликации усиливает множественное редактирование генома с помощью 90-нуклеотидных одноцепочечных олигодезоксинуклеотидов (ssODN) в дрожжах, что позволяет одновременно интегрировать более 10 ssODN с 60 мутациями на трансформацию (Barbieri). и др. , 2017 г.). Что наиболее важно, эта стратегия не зависит как от DSB ДНК, так и от гомологичной рекомбинации, и в ближайшем будущем станет возможным объединить мультилокусное разрешение и разрешение одной пары оснований этого подхода с опосредованным CRISPR-dCas9 перепрограммированием транскрипции для быстрой идентификации. отпечатков генома и экспрессии, связанных с желаемыми признаками.
Наконец, нативная регуляция транскрипции зависит от интегрирования многогенных пространственно-временных нарушений экспрессии. Для дальнейшего обеспечения синтетического контроля над транскрипцией полигенных признаков, особенно тех, которые зависят от основных генов, исследования в рамках контролируемых систем CRISPR должны использовать преимущества и дальнейшее развитие стратегий перепрограммирования, совместимых с оптогенетикой, тем самым обходя ограниченную обратимость химических реакций. индуцированные (например, aTc) системы (Xiaofeng et al. 2017). Точно так же следует учитывать аллостерическую регуляцию активности белка CRISPR для условного переключения клеточного принятия решений, например. состояния роста и метаболизма (Oakes et al. 2016). В конечном счете, предполагается, что такие методы значительно поддержат наше понимание и ортогональный контроль транскрипционных и посттранскрипционных регуляций для желаемых клеточных и метаболических результатов.
Благодарности
Авторы благодарят Эмиля Дамгаарда Йенсена за плодотворные обсуждения и комментарии к рукописи во время подготовки.
ФИНАНСИРОВАНИЕ
Работа выполнена при поддержке фонда Ново Нордиск.
Конфликт интересов . Не объявлено.
ССЫЛКИ
Абудайе
ОО
,Gootenberg
JS
,Essletzbichler
P
и др.Нацеливание на РНК с помощью CRISPR-Cas13
.Природа
2017
;550
:280
–4
.
Альпер
Х
,Moxley
J
,Nevoigt
E
и др.Инженерное оборудование для транскрипции дрожжей для повышения толерантности к этанолу и производства
.Наука
2006
;314
:1565
–8
.
Барбьери
ЭМ
,Муир
П
,Ахуэти-Они
БО
и др.Точное редактирование репликационных вилок ДНК позволяет проводить мультиплексную инженерию генома у эукариот
.Сотовый
2017
;171
:1453
–1467.e13
.
Бланшен-Роланд
С
,Кордеро Отеро
RR
,Гайярдин
C
.Две расположенные выше последовательности активации контролируют экспрессию гена XPR2 в дрожжах Yarrowia lipolytica
.Мол Селл Биол
1994
;14
:327
–38
.
Блазек
Дж
,Лю
L
,Редден
H
и др.Настройка экспрессии генов в Yarrowia lipolytica с помощью гибридного промотора
.Appl Environ Microbiol
2011
;77
:7905
–14
.
Чавес
А
,Scheiman
J
,Вора
S
и др.Высокоэффективное Cas9-опосредованное программирование транскрипции
.Nat Methods
2015
;12
:326
–8
.
Чавес
А
,Tuttle
M
,Pruitt
BW
и др.Сравнение активаторов Cas9 у нескольких видов
.Nat Methods
2016
;13
:563
–7
.
Черчмен
ЛС
,Вайсман
JS
.Секвенирование зарождающихся транскриптов позволяет визуализировать транскрипцию с разрешением нуклеотидов
.Природа
2011
;469
:368
–73
.
Конг
л
,Ran
FA
,Cox
D
и др.Мультиплексная инженерия генома с использованием систем CRISPR/Cas
.Наука
2013
;339
:819
–23
.
Кокс
ДБТ
,Gootenberg
JS
,Abudayyeh
OO
et al.Редактирование РНК с помощью CRISPR-Cas13
.Наука
2017
;358
1019
–27
.
Крук
НЗ
,Schmitz
AC
,Alper
HS
.Оптимизация системы интерференции дрожжевой РНК для контроля экспрессии генов и обеспечения быстрой метаболической инженерии
.ACS Synth Biol
2013
;3
:307
–13
.
Дай Х
,Chen X
,Fang Q
и др.Индуцируемый инструмент редактирования генома CRISPR: классификации и будущие тенденции
.Crit Rev Biotechnol
2017
;38
:573
–86
.
Динер
М
,Альпер
ГС
.Систематическое тестирование чувствительности ферментов к возмущению посредством дифференцированной модуляции dCas9 в Saccharomyces cerevisiae
.Metab Eng
2017
;40
:14
–22
.
Динер
М
,Мехия
J
,Альпер
HS
.Включение дифференцированного и крупномасштабного мультиплексирования желаемых генов с использованием двухрежимного активатора dCas9 в Saccharomyces cerevisiae
.ACS Synth Biol
2017
;6
:1931
–43
.
ДиКарло
JЕ
,Norville
JE
,Мали
P
и др.Геномная инженерия в Saccharomyces cerevisiae с использованием систем CRISPR-Cas
.Рез. нуклеиновых кислот
2013
;41
:4336
–43
.
Дринненберг
ИА
,Weinberg
DE
,Xie
KT
и др.РНКи в почкующихся дрожжах
.Наука
2009
;326
:544
–50
.
Эдмондсон
ДГ
,Смит
ММ
,Рот
SY
.Репрессионный домен дрожжевого глобального репрессора Tup1 напрямую взаимодействует с гистонами h4 и h5
.Гены Дев
1996
;10
:1247
–59
.
Эллис
Т
,Ван
X
,Коллинз
Дж.Дж.
.Конструирование синтетических генных сетей с предсказанными функциями на основе разнообразия и моделей
.Нат Биотехнолог
2009
;27
:465
–71
.
Фарзадфард
Ф
,Перли
SD
,Лу
ТК
.Настраиваемые и многофункциональные эукариотические факторы транскрипции на основе CRISPR/Cas
.ACS Synth Biol
2013
;2
:604
–13
.
Феррейра
Р
,Skrekas
C
,Nielsen
J
и др.Мультиплексное редактирование генома CRISPR/Cas9 и регуляция генов с использованием Csy4 в Saccharomyces cerevisiae
.ACS Synth Biol
2018
;77
:10
–15
.
Флик
JS
,Джонстон
М
.Две системы глюкозной репрессии промотора GAL1 в Saccharomyces cerevisiae
.Мол Селл Биол
1990
;10
:4757
–69
.
Фолчер
М
,Xie
M
,Spinnler
A
и др.Синтетические управляемые триггером двудольные факторы транскрипции млекопитающих
.Рез. нуклеиновых кислот
2013
;41
:e134
.
Фу
БХХ
,Hansen
LL
,Artiles
KL
и др.Ландшафт эффектов гомологии target:guide на Cas9-опосредованное расщепление
.Рез. нуклеиновых кислот
2014a
;42
:13778
–87
.
Фу
Д
,Rocha
PP
,Luo
VM
и др.Каркасы CRISPR-dCas9 и sgRNA обеспечивают двухцветную визуализацию сателлитных последовательностей и обогащенных повторами отдельных локусов в режиме реального времени
.Нац Коммуна
2016
;7
:11707
.
Фу
Д
,Sander
JD
,Reyon
D
и др.Повышение специфичности нуклеазы CRISPR-Cas с помощью укороченных направляющих РНК
.Нат Биотехнолог
2014b
;32
:279
–84
.
Гусак
МВт
,Врана
JD
,Вое
ЗЕ
и др.Цифровые логические схемы в дрожжах с вентилями CRISPR-dCas9 NOR
.Национальная связь
2017
;8
:15459
.
Гао
Д
,Xiong
X
,Wong
S
и др.Комплексная модуляция транскрипции с помощью ортогональных и индуцируемых регуляторов dCas9
.Nat Methods
2016
;13
:1043
–9
.
Гао
Д
,Чжао
Y
.Самопроцессинг фланкированных рибозимом РНК в направляющие РНК in vitro и in vivo для редактирования генома с помощью CRISPR
.J Integr Plant Biol
2014
;56
:343
–9
.
Гилберт
ЛА
,Horlbeck
MA
,Adamson
B
и др.CRISPR-опосредованный контроль репрессии и активации генов в масштабе генома
.Сотовый
2014
;159
:647
–61
.
Гилберт
ЛА
,Ларсон
MH
,Морсут
L
и др.Опосредованная CRISPR модульная РНК-управляемая регуляция транскрипции у эукариот
.Сотовый
2013
;154
:442
–51
.
Гризенбек
Дж
,Boeger
H
,Strattan
JS
и др.Аффинная очистка специфических сегментов хроматина из хромосомных локусов дрожжей
.Мол Селл Биол
2003
;23
:9275
–82
.
Хан
С
.Структура и механизм механизма транскрипции РНК-полимеразы II
.Nat Struct Mol Biol
2004
;11
:394
–403
.
Хинц
ДжМ
,Логери
МФ
,Вайрик
Дж.Дж.
.Нуклеосомы ингибируют активность эндонуклеазы Cas9 in vitro
.Биохимия
2015
;54
:7063
–6
.
Хорлбек
МА
,Gilbert
LA
,Villalta
JE
и др.Компактные и высокоактивные библиотеки следующего поколения для CRISPR-опосредованной репрессии и активации генов
.Элиф
2016
;5
. ДОИ: .
Хоу
ФС
,Russell
A
,Lamstaes
AR
и др.CRISPRi не является нитей-специфическим во всех локусах и переопределяет ландшафт транскрипции
.Элиф
2017
;6
:e29878
.
Хсу
ПД
,Scott
DA
,Weinstein
JA
et al.Специфичность нацеливания ДНК на РНК-управляемые нуклеазы Cas9
.Нат Биотехнолог
2013
;31
:827
–32
.
Исаак
РС
,Jiang
F
,Doudna
JA
и др.Дыхание и ремоделирование нуклеосом ограничивают функцию CRISPR-Cas9
.Элиф
2016
;5
:e13450
.
Дженсен
ЭД
,Ferreira
R
,Jakočiūnas
T
et al.Транскрипционное перепрограммирование у дрожжей с использованием dCas9 и комбинаторных стратегий гРНК
.Факт о микробных клетках
2017
;16
:46
.
Дженсен
МК
,Kjaersgaard
T
,Nielsen
MM
и др.Семейство факторов транскрипции NAC Arabidopsis thaliana: структурно-функциональные отношения и детерминанты передачи сигналов стресса ANAC019
.Biochem J
2010
;426
:183
–96
.
Цзян
С
,Пью
БФ
.Составленная и систематизированная справочная карта позиций нуклеосом в геноме Saccharomyces cerevisiae
.Геном Биол
2009
;10
:R109
.
Джинек
М
,Чилинский
К
,Фонфара
I
и др.Программируемая ДНК-эндонуклеаза, управляемая двойной РНК, в адаптивном бактериальном иммунитете
.Наука
2012
;337
:816
–21
.
Кадош
Д
,Штруль
К.
Репрессия с помощью Ume6 включает рекрутирование комплекса, содержащего корепрессор Sin3 и гистондеацетилазу Rpd3, на промоторы-мишени
.Сотовый
1997
;89
:365
–71
.
Халил
КАК
,Лу
ТК
,Башор
CJ
и др.Основа синтетической биологии для программирования эукариотических функций транскрипции
.Сотовый
2012
;150
:647
–58
.
Киани
С
,Чавес
А
,Таттл
М
и др.Разработка гРНК Cas9 для редактирования, активации и подавления генома
.Nat Methods
2015
;12
:1051
–4
.
Ларсон
МЗ
,Gilbert
LA
,Wang
X
и др.Интерференция CRISPR (CRISPRi) для специфичного для последовательности контроля экспрессии генов
.Nat Protoc
2013
;8
:2180
–96
.
Лоухорн
ИЭБ
,Феррейра
JP
,Ван
CL
.Оценка сайтов-мишеней sgRNA для CRISPR-опосредованной репрессии TP53
.PLoS One
2014
;9
:e113232
.
Ли
Вт
,Tillo
D
,Bray
N
и др.Атлас заполнения нуклеосом у дрожжей с высоким разрешением
.Нат Жене
2007
;39
:1235
–44
.
Ленстра
турецких лир
,Кулон
А
,Чоу
CC
и др.Визуализация одиночных молекул показывает переключение между ложной и функциональной транскрипцией нкРНК
.Мол Ячейка
2015
;60
:597
–610
.
Лиан
Дж
,HamediRad
M
,Hu
S
и др.Комбинаторная метаболическая инженерия с использованием ортогональной таифункциональной системы CRISPR
.Nat Comm
2017
;8
:1688
.
Мао
С
,Браун
CR
,Гризенбек
J
и др.Окклюзия регуляторных последовательностей промоторными нуклеосомами in vivo
.PLoS One
2011
;6
:e17521
.
МакИсаак
РС
,Оукс
BL
,Ван
X
и др.Системы возмущения экспрессии синтетических генов с быстрой, настраиваемой специфичностью к одному гену у дрожжей
.Рез. нуклеиновых кислот
2013
;41
:e57-
.
Мицунобу
Х
,Teramoto
J
,Nishida
K
и др.Помимо нативного Cas9: управление геномной информацией и функциями
.Trends Biotechnol
2017
;35
:983
–96
.
Намбу-Нисида
Д
,Нисида
К
,Хасунума
Т
и др.Разработка комплексного набора инструментов для геномной инженерии холодо- и термоустойчивых дрожжей Kluyveromyces marxianus , штамм
.Научный представитель
2017
;7
:8993
.
Наранхо
С
,Smith
JD
,Artieri
CG
и др.Анализ генетической основы сложной цис-регуляторной адаптации
.PLoS Genet
2015
;11
:e1005751
.
Нисида
К
,Arazoe
T
,Yachie
N
и др.Направленное редактирование нуклеотидов с использованием гибридных прокариотических и позвоночных адаптивных иммунных систем
.Наука
2016
;353
:aaf8729
.
Оукс
БЛ
,Nadler
DC
,Flamholz
A
и др.Профилирование инженерных горячих точек идентифицирует аллостерический переключатель CRISPR-Cas9
.Нат Биотехнолог
2016
;34
:646
–51
.
Остлинг
Дж
,Карлберг
М
,Ронне
Н
.Функциональные домены репрессора Mig1
.Мол Селл Биол
1996
;16
:753
–61
.
Пьер-Жером
Е
,Jang
SS
,Havens
KA
и др.Повторение пути прямого ответа ядерного ауксина у дрожжей
.P Natl Acad Sci USA
2014
;111
:9407
–12
.
Ци
ЛС
,Ларсон
MH
,Гилберт
ЛА
и др.Перепрофилирование CRISPR в качестве управляемой РНК платформы для последовательно-специфического контроля экспрессии генов
.Сотовый
2013
;152
:1173
–83
.
Радзишевская
А
,Шлюева
Д
,Мюллер
И
и др.Оптимизация положения sgRNA заметно повышает эффективность репрессии транскрипции, опосредованной CRISPR/dCas9
.Рез. нуклеиновых кислот
2016
;44
:e141
.
Рандо
ОЖ
,Чанг
HY
.Полногеномные представления структуры хроматина
.Annu Rev Biochem
2009
;78
:245
–71
.
Рандо
ОЖ
,Уинстон
Ф
.Хроматин и транскрипция у дрожжей
.Генетика
2012
;190
:351
–87
.
Райан
ОВ
,Skerker
JM
,Maurer
MJ
и др.Отбор библиотек хромосомной ДНК с использованием мультиплексной системы CRISPR
.Элиф
2014
;3
:e03703
.
Схема
АН
,Буэнростро
JD
,Денни
SK
и др.Структурированные нуклеосомные отпечатки позволяют картировать хроматиновую архитектуру в регуляторных областях с высоким разрешением
.Genome Res
2015
;25
:1757
–70
.
Шрайбер-Агус
Н
,Чин
Л
,Чен
К
и др.Аминоконцевой домен Mxi1 опосредует онкогенную активность против Myc и взаимодействует с гомологом дрожжевого репрессора транскрипции SIN3
.Сотовый
1995
;80
:777
–86
.
Шварц
С
,Frogue
K
,Рамеш
A
и др.Репрессия CRISPRi негомологичного соединения концов для улучшенной инженерии генома посредством гомологичной рекомбинации в Yarrowia lipolytica
.Биотехнолог Биоэнг
2017
;114
:2896
–906
.
Шварц
см
,Hussain
MS
,Blenner
M
и др.Промоторы синтетической РНК-полимеразы III облегчают высокоэффективное редактирование генома с помощью CRISPR-Cas9 в Yarrowia lipolytica
.ACS Synth Biol
2016
;5
:356
–9
.
Си
Т
,Луо
Y
,Бао
Z
и др.Эволюция генома с помощью РНК-интерференции у Saccharomyces cerevisiae для сложной инженерии фенотипа
.ACS Synth Biol
2015
;4
:283
–91
.
Скьёдт
мл
,Snoek
T
,Kildegaard
KR
и др.Разработка прокариотических активаторов транскрипции в качестве биосенсоров метаболитов в дрожжах
.Nat Chem Biol
2016
;12
:951
–8
.
Смит
ДД
,Суреш
S
,Schlecht
U
и др.Количественные интерференционные скрининги CRISPR в дрожжах выявляют химико-генетические взаимодействия и новые правила дизайна направляющей РНК
.Геном Биол
2016
;17
:45
.
Смит
ДД
,Schkecht
U
,Xu
W
и др.Метод высокопроизводительного производства библиотек ДНК с подтвержденной последовательностью и коллекций штаммов
.Мол Сист Биол
2017
;13
:913
.
Травен
А
,Старесинчич
Л
,Арнерич
М
и др.Белок дрожжей Xtc1 действует как прямой репрессор транскрипции
.Рез. нуклеиновых кислот
2002
;30
:2358
–64
.
Ванегас
кг
,Лехка
БЖ
,Мортенсен
УХ
.SWITCH: динамический инструмент CRISPR для геномной инженерии и управления метаболическими путями для создания фабрики клеток в Saccharomyces cerevisiae
.Факт о микробных клетках
2017
;16
:25
.
Витцгалл
Р
,O’Leary
E
,Leaf
A
и др.Kruppel-ассоциированный box-A (KRAB-A) домен белков цинковых пальцев опосредует репрессию транскрипции
.P Natl Acad Sci USA
1994
;91
:4514
–8
.
Ву
Дж
,Suka
N
,Carlson
M
и др.TUP1 использует гистон h4/h3B-специфическую деацетилазу HDA1 для подавления активности генов у дрожжей
.Мол Ячейка
2001
;7
:117
–26
.
Ву
х
,Scott
DA
,Kriz
AJ
и др.Полногеномное связывание эндонуклеазы CRISPR Cas9 в клетках млекопитающих
.Нат Биотехнолог
2014
;32
:670
–6
.
Йошиока
С
,Fujii
W
,Ogawa
T
и др.Разработка управляемой монопромотором системы CRISPR/Cas9 в клетках млекопитающих
.Научный представитель
2015
;5
:18341
.
юаня
Г-К
,Liu
Y-J
,Dion
MF
и др.Идентификация положения нуклеосом в масштабе генома у S. cerevisiae
.Наука
2005
;309
:626
–30
.
Залатан
ДжГ
,Lee
ME
,Almeida
R
и др.Разработка комплексных программ синтетической транскрипции с использованием каркасов РНК CRISPR
.Сотовый
2015
;160
:339
–50
.
Заугг
ДЖБ
,Ласкомб
Нью-Мексико
.Геномная модель зависимого от состояния поведения нуклеосом объясняет активность транскрипции у дрожжей
.Genome Res
2012
;22
:84
–94
.
Чжан
З
,Риз
JC
.Молекулярно-генетический анализ дрожжевого репрессора Rfx1/Crt1 выявил новый двухстадийный регуляторный механизм
.Мол Селл Биол
2005
;25
:7399
–411
.
Чжан
Т
,Гао
Y
,Ван
R
и др.Производство направляющих РНК in vitro и in vivo для CRISPR с использованием рибозимов и промоторов РНК-полимеразы II
.БиоПроток
2017
;7
:e2148
.© ФЭМС 2018.
Новости и Объявления — Портфолио Бурджу Гуркана
Новости
Январь 2022
Добро пожаловать в новых участников группы: Саудагар (постдоктор) из Индии и Огуз (аспирант) из Турции!
Наша совместная статья, опубликованная в Nature Communications: «Эволюция микроскопической неоднородности и динамики в глубоких эвтектических растворителях на основе хлорида холина».Работа от команды EFRC BEES!
декабрь 2021 г.
Наша статья с соавторами Сяо Су и Шаамой Малликарджун Шарада на тему «Перспективы и проблемы электрохимических подходов к реактивному разделению CO2» опубликована в iScience . Спасибо профессору Марте Хатцелл и Мишель Муццио за приглашение внести свой вклад в специальный выпуск журнала Negative Emission Science.
ноябрь 2021
Айдам, Дрейс, Уилл и Ли представили свою работу на конференции AIChE в Бостоне.Они также привезли домой несколько наград с конкурса Beer Brewing.
Доктор Гуркан и его сотрудники Гетман, Киддер, Пентцер и Парк получили награду DAC Министерства энергетики за разработку материалов и способов нетермической регенерации для улавливания углекислого газа из воздуха.
Сентябрь 2021
Мы приветствуем двух новых аспирантов: Рут и Сандуни! Рут будет изучать улавливание СО2 из воздуха, а Сандуни будет изучать электровосстановление СО2.
Первая статья Дрейса от первого автора недавно была принята и будет опубликована в специальном выпуске Frontiers in Energy Research: Rising Stars .Очень горжусь тем, что он провел это вычислительное исследование во время covid для своей в основном экспериментальной темы докторской диссертации!
Июль 2021
Лаборатория получает престижную награду NSF CAREER.
Д-р Гуркан повышен до заслуженного доцента Севера!
Май 2021
Поздравляем Джеффа с получением выдающейся докторской степени! Мы желаем ему всего наилучшего в его новой роли постдокторского исследователя в Лос-Аламосской национальной лаборатории.
Добро пожаловать в Харрисон! Харрисон, студент CWRU, этим летом будет изучать ионные жидкие электролиты для хранения энергии.
март 2021
Джефф был в ORNL и проводил эксперименты на линии луча! Очень взволнован нашей следующей рукописью, описывающей границы раздела электрод-электролит под приложенными потенциалами, как это видно по отражательной способности нейтронов.
Доктор Гуркан виртуально присутствовал на мероприятии NASA Space Tech Day on the Hill.
Д-р Гуркан модерировала серию бесед «Женщины чудес» с участием д-ра Дебры Уилфонг, выпускницы CWRU, в рамках празднования Месяца истории женщин; организован Ассоциацией выпускников Case и Женским центром Flora Stone Mather.
январь 2021
Мы начинаем новый год с множеством интересных новостей!
Присоединились два новых исследователя с докторской степенью! Добро пожаловать, Кевин и Мелина (Разия)!
Ли опубликовал свое исследование в ACS Sus Chem& Eng. Проверьте это; это открытый доступ! По-настоящему совместная работа группы Gurkan!
Уилл и Джефф опубликовали свою совместную работу в J ElectrochemicalSociety. Финансируется DOE EFRC BEES.
Д-р Гуркан получил премию Scialog Award 2020 от RSCA в области науки об отрицательном излучении!
Декабрь 2020 г.
Мы приняли участие во всестороннем обзоре Deep Eutectic Solvents, опубликованном в Chemical Reviews.Финансируется DOE EFRC BEES.
ноябрь 2020 г.
Д-р Гуркан выступил с приглашенным докладом на онлайн-сессии виртуального собрания Общества исследования материалов 2020: открытые электрохимические системы на основе потока.
октябрь 2020 г.
Уилл и Джефф представляют свое исследование межфазного взаимодействия электрод-электролит в растворителях глубокой эвтектики на ECS PRiME! Проверьте это здесь!
Сентябрь 2020
Новые участники присоединяются к команде: Добро пожаловать, Ася, Эмили и Ханна! Ася учится на инженерном факультете CWRU.Эмили и Ханна учатся искусству в Кливлендском институте искусств.
августа 2020153
августа
Минжи Эйдан и Ли Обезвременные транспортные мембраны с ионные жидкости публикуются в Границы в химии: https://doi.org/10.3389/fchem.2020.00637
Июль 2020 г.
Доктор Гуркан выбран в качестве стипендиата Scialog для участия в инициативе Scialog: Negative Emissions Science (NES) 2020 года, совместно спонсируемой Исследовательской корпорацией по развитию науки и Альфредом П.Фонд Слоуна.
июнь 2020 г.
Статья Джеффа и Генри о сольватации в глубоких эвтектических растворителях теперь доступна по адресу Journal of Physical Chemistry B: https://doi.org/10.1021/acs.jpcb.0c03296
MayПоздравляем Генри с дипломом бакалавра и магистра с дипломной работой! Он будет получать докторскую степень в Калифорнийском университете в Беркли!
Поздравляем Брайана с получением степени магистра!
Апрель 2020 г.
Статья Ли об улавливании диоксида углерода с помощью капсул с ионными жидкостями опубликована по адресу ACS Applied Materials & Interfaces .Сотрудничество с группой Pentzer в TAMU. Ознакомьтесь с ним на странице https://doi.org/10.1021/acsami.0c01622
, декабрь 2019 г.
Статья Джеффа об интерфейсе электрод-ионная жидкость от SERS только что была принята. Проверьте это на обложке Journal of Physical Chemistry C : https://doi.org/10.1021/acs.jpcc.9b08016
Наша приглашенная перспективная статья теперь доступна в Journal of Physical Chemistry Letters (doi.org/10.1021/acs.jpclett.9б01980). Обложка и видео будут!
Доктор Гуркан представил наши исследования в области космических технологий на мероприятии НАСА по космическим технологиям на Столичном холме.
Добро пожаловать в Сундар! Он присоединяется к группе в качестве постдока из Университета Южной Калифорнии.
ноябрь 2019 г.
Поздравляем Джеффа с получением награды DOE SCGSR !! Он будет заниматься нейтронными исследованиями в Окриджской национальной лаборатории в течение 6 месяцев, начиная с января 2020 года.
Д-р Гуркан выступит с докладом на собрании AIChE Orlando : «Проточные окислительно-восстановительные батареи с эталиновым глубоким эвтектическим растворителем» на сессиях: 226 — Проточные окислительно-восстановительные батареи для хранения энергии
Д-р Гуркан посещает Университет Тафтса (11/18) для участия в семинаре на факультете химической и биологической инженерии.
октябрь 2019 г.
Поздравляем I&EC Research’s Класс влиятельных исследователей 2019 года. Д-р Гуркан входит в число 32 мировых исследователей, получивших признание в виртуальном специальном выпуске.
Статья Вивиан и Ли доступна в Industrial & Engineering Chemistry Research (doi.org/10.1021/acs.iecr.9b03202), в специальном выпуске Chuck Eckert Festschrift
Джефф представил свою работу над Surface Усовершенствованная рамановская спектроскопия для исследования границ раздела электрод-электролит на конференции ECS в Атланте.
Генри представил плакат о глубоких эвтектических растворителях на конференции ECS в Атланте.
Д-р Гуркан посетил Университет Теннесси в Ноксвилле на семинаре кафедры химической и биомолекулярной инженерии.
Доктор Гуркан выступил с докладом в Исследовательском центре Эймса НАСА (25 октября), на семинарах XO.
Сентябрь 2019
Дрейс Пэнли из Университета штата Огайо (MS 2019) присоединяется к группе в качестве аспиранта.
Уильям Дин из Миннесотского университета (бакалавр наук, 2018 г.) присоединяется к группе в качестве аспиранта.
Добро пожаловать в Дрейс и Уилл!
Д-р Гуркан посетил Технологический институт Стивенса на семинаре: Серия «Нанотехнологии»
Август 2019 г.
Д-р Гуркан выступил с приглашенным докладом на исследовательской конференции Гордона по химии и физике жидкостей в Школе Холдернесса.
июнь 2019 г.
В этом году в CWRU стартовала программа REU. К нам присоединяется Виктория Зубисаррета из Университета Питтсбурга. Виктория будет с нами в течение 10 недель, работая над динамикой ионов в пористых средах.
май 2019 г.
1-й аспирант Energy Lab: Вивиан защитила докторскую диссертацию и официально закончила ее весной 2019 г.
апрель 2019 г. Лаборатория Гуркан. Профессор Бреннеке также читает лекцию Томаса Белла в Кливлендском государственном университете.
Март 2019 г.
DOE EFRC Прорывные электролиты для хранения энергии (BEES) провел свое первое всестороннее собрание в CWRU.Д-р Гуркан провел презентацию Thrust 1: Deep Eutectic Solvents.
Февраль 2019 г.
Хунчао Мао присоединяется к группе в качестве постдока Политехнического института Ренсселера. Хода Амани Хамедани присоединяется к группе в качестве старшего научного сотрудника Nano Precision Medical Inc. Добро пожаловать в Хунчао и Хода!
Малышка Гуркан #2: В феврале у доктора Гуркана родилась девочка! Добро пожаловать, Малышка Талия!
Декабрь 2018
Статья Вивиан опубликована в Journal of Physical Chemistry B .
Статья Джеффа по PCCP выделена на задней обложке журнала.
ноябрь 2018 г.
Вивиан представила два доклада на встрече AIChE в Питтсбурге в 2018 г.: одна посвящена батареям, а другая — суперконденсаторам.
Октябрь 2018
Статья Джеффа только что была опубликована по адресу Physical Chemistry Chemical Physics !
Джефф представил свою работу на встрече ECS AiMES в Канкуне. Он также представил нашу работу по поверхностно-активным ионным жидкостям.
Статья Карен только что опубликована в Журнале прикладной электрохимии !
Доктор Гуркан получает награду ACS-PRF для нового исследователя!
Сентябрь 2018
Брайан Чен присоединился к группе из Городского колледжа Нью-Йорка!
Юн-Ян Ли присоединился к группе из Национального университета Цин-Хуа, Тайвань!
Добро пожаловать в Брайан и Ли!
август 2018 г.
Доктор Гуркан получает награду НАСА за раннюю карьеру.
Доктор Гуркан выступил с докладом на исследовательской конференции Гордона по ионным жидкостям.
июль 2018 г.
Мы открываем новый Исследовательский центр EnergyFrontier Министерства энергетики США BEES: революционные электролиты для хранения энергии. Д-р Савинелл является директором Центра, а д-р Гуркан является руководителем одного из направлений и ответственным за интеграцию исследований в двух направлениях.
май 2018 г.
Поздравляем Карен и Сучитру с получением степени магистра!
Доктор Гуркан получает награду CWRU VPR Catalyst Award за субсидию на публикацию.
Апрель 2018 г.
Нашу статью только что приняли по адресу ACS Materials & Interfaces . https://pubs.acs.org/doi/10.1021/acsami.8b01285
Qianwen и Ningjin представляют свою работу на ShowCASE CWRU 20 апреля 2018 г.
Ссылка: http://case.edu/research/showcase /
Март 2018 г.
Д-р Гуркан избран директором отдела транспортных и энергетических процессов Айше.
Доктор Гуркан представляет на встрече ACS в Новом Орлеане 2018 года: Физическая химия ионных жидкостей .
Февраль 2018 г.
Д-р Гуркан выступает с докладом на семинаре на факультете химической и биомолекулярной инженерии Университета Клемсона.
Наше предложение по времени луча принято. Джефф отправится в NIST для проведения нейтронных измерений с целью изучения структуры ионных жидкостей.
ноябрь 2017 г.
Доктор Гуркан выступает с докладом на семинаре в Кливлендском государственном университете (16 ноября).
Лаборатория энергетики приняла участие во Всемирном дне науки для мира и развития в CWRU, демонстрируя наши исследования (10 ноября).
Октябрь 2017 г.
Д-р Гуркан выступит с основным докладом на сессии по технологиям перезаряжаемых/вторичных батарей для хранения энергии в AIChE, Миннеаполис.
Д-р Гуркан является исполняющим обязанности заместителя председателя по программированию отдела транспортных и энергетических процессов на ежегодном осеннем собрании AIChE 2017 года. Она также является модератором группы аспирантов Комитета женской инициативы.
Джозеф Уильямс из Кливлендского института искусств присоединяется к группе в качестве стажера-аниматора.
Сентябрь 2017 г.
Джеффри Кляйн из Школы горного дела Колорадо (бакалавр наук, 2017 г.) присоединяется к группе в качестве аспиранта.
Члены ENERGY LAB будут присутствовать на 232-м национальном собрании ECS. Тезисы Qianwen (Vivian) были приняты для устной презентации на Симпозиуме L03: Физическая и аналитическая электрохимия ионных жидкостей 6. Она представит свое исследование 4 октября 2017 г.: 8:40–9:00
Август 2017 г.
Д-р Гуркан присутствовал Летняя школа химического машиностроения ASEE 2017 года в Северной Каролине.
Д-р Гуркан получил грант Nord от UCITE.
июнь 2017 г.
VeAnn Pham из Аризонского университета присоединяется к группе в качестве участника NSF REU.
Нейл получает финансирование из ИСТОЧНИКА. Нейл будет работать в лаборатории летом 2017 года над процессами электровосстановления в ионных жидкостях. Его исследовательская поддержка исходит от Централизованного офиса студенческих исследований в CWRU.
ноябрь 2016 г.
Д-р Гуркан председательствовал на следующих сессиях ежегодного собрания AIChE в Сан-Франциско в 2016 г.: Технологии перезаряжаемых и вторичных батарей для аккумуляторов и технологий хранения энергии
, август 2016 г. учредил.
Домашний
Добро пожаловать в группу Ян!
Исследования в лаборатории Янга направлены на разработку передовых материалов для производства чистой энергии и хранения возобновляемой энергии устройств.
О докторе Ян Ян
Доктор. Ян Ян изучал материаловедение и инженерию и получил докторскую степень.D. из Университета Цинхуа в 2010 году. С 2010 по 2012 год он получил поддержку стипендии Александра фон Гумбольдта и работал с профессором доктором Патриком Шмуки в Университете Эрлангена-Нюрнберга, Германия. С 2012 по 2015 год он получил поддержку стипендии Питера М. и Рут Л. Николас и работал с профессором доктором Джеймсом М. Туром в Институте наноразмерных наук Ричарда Э. Смолли. & Tech., Университет Райса. С 2015 года он был доцентом в Технологическом центре нанонауки и на факультете материаловедения и инженерии Университета Центральной Флориды.Его текущие исследовательские интересы охватывают программируемое нанопроизводство, инженерию поверхностей и интерфейсов энергетических материалов, производство и хранение чистой энергии, сбор солнечной энергии и интеллектуальную/гибкую электронику, уделяя особое внимание передовым материалам и их электрохимическим применениям.
Избранные статьи и новости на обложках
Вакансии
Мы ищем аспирантов, студентов бакалавриата и докторантов с опытом работы и интересом к передовым материалам и возобновляемым источникам энергии.
