
прямая ссылка 30 июля 2018 | 11:13
прямая ссылка 01 января 2017 | 03:15
В оцифрованном доисторическом мире
прямая ссылка 20 января 2016 | 13:58
И до динозавров была жизнь
прямая ссылка 15 марта 2014 | 21:11
Я злой и страшный стрекоза…
прямая ссылка 13 сентября 2013 | 21:18
прямая ссылка 11 августа 2013 | 00:34
Время, когда ещё ни один динозавр не ступал ногой на землю…
прямая ссылка 15 марта 2013 | 18:16
прямая ссылка 05 февраля 2009 | 15:04
прямая ссылка 25 августа 2008 | 10:58 |


 Во-первых, ясно, что простой переводчик никогда бы до такой подмены не додумался. Следовательно, переводчика консультировал палеонтолог, причем он, как видим, не согласился сохранить оригинальные названия. Это может ничего и не значить. Но вообще говоря, замечание в тему: среди палеонтологов нет желаемого единства взглядов. Имеется в виду следующее: в целом ряде случаев даже такие, казалось бы, чисто условные обозначения довольно жестко привязаны к определенным представлениям, теориям. И смена теорий нередко приводит к смене названий вымерших существ…
Во-первых, ясно, что простой переводчик никогда бы до такой подмены не додумался. Следовательно, переводчика консультировал палеонтолог, причем он, как видим, не согласился сохранить оригинальные названия. Это может ничего и не значить. Но вообще говоря, замечание в тему: среди палеонтологов нет желаемого единства взглядов. Имеется в виду следующее: в целом ряде случаев даже такие, казалось бы, чисто условные обозначения довольно жестко привязаны к определенным представлениям, теориям. И смена теорий нередко приводит к смене названий вымерших существ… .
.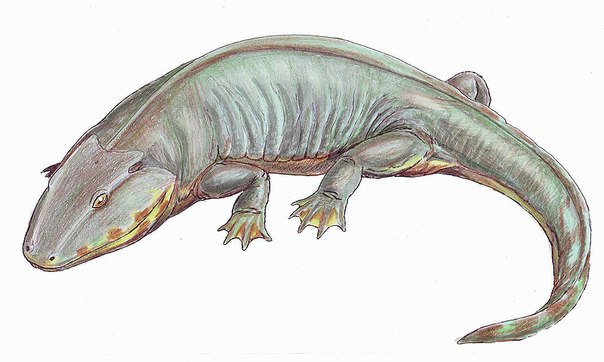

 Тут нет какопса, зато есть ринезух.
Тут нет какопса, зато есть ринезух. Пользуясь неусмиренным желанием публики заглянуть не только на расстояние, но во время, Британская широковещательная корпорация предоставила развлекательный аттракцион, имитирующий жизнь животного мира до динозавров. Так далеко обычно не заглядывали, ограничившись пробегом по учебнику биологии по инфузориям-туфелькам, выползающим на сушу водорослям-папоротникам и сразу переходя к милым сердцу гигантским ящерам.
Пользуясь неусмиренным желанием публики заглянуть не только на расстояние, но во время, Британская широковещательная корпорация предоставила развлекательный аттракцион, имитирующий жизнь животного мира до динозавров. Так далеко обычно не заглядывали, ограничившись пробегом по учебнику биологии по инфузориям-туфелькам, выползающим на сушу водорослям-папоротникам и сразу переходя к милым сердцу гигантским ящерам. 
 О’кей, Google – и вот тебе развлекательная картина, где специалисты так наложили в отснятые локации придуманных рептилий, что и не подкопаешься. Хотя, справедливости ради, передвижение тварей уж больно плавное и замедленное. Зато эффект присутствия камеры как конфетка, то брызги содержимого разорванного кишечника очередного диметродона на неё брызнут, то любопытная ящерка в объектив вытаращится «А это чаво здесь?». Кто-то из фанатов «Прогулок с …» даже за деревьями человека разглядел. А вдруг на машине времени прилетели и сняли, реалистически же всё получилось.
О’кей, Google – и вот тебе развлекательная картина, где специалисты так наложили в отснятые локации придуманных рептилий, что и не подкопаешься. Хотя, справедливости ради, передвижение тварей уж больно плавное и замедленное. Зато эффект присутствия камеры как конфетка, то брызги содержимого разорванного кишечника очередного диметродона на неё брызнут, то любопытная ящерка в объектив вытаращится «А это чаво здесь?». Кто-то из фанатов «Прогулок с …» даже за деревьями человека разглядел. А вдруг на машине времени прилетели и сняли, реалистически же всё получилось. И основное сожаление, что занимаются таким производством все забугорные капиталисты, у которых нет великого духовного мира и сколковских нанотехнологий.
И основное сожаление, что занимаются таким производством все забугорные капиталисты, у которых нет великого духовного мира и сколковских нанотехнологий. Казалось бы, о чем можно было еще снять, когда уже вышли фильмы о динозаврах, доисторических монстрах, первобытных людях. В 2005 г. вышел последний (пока что) фильм Хлои Лелэнд и Тиа Хайнса «Прогулки с монстрами. Жизнь до динозавров», который фактически являлся приквелом ко всем фильмам данного цикла.
Казалось бы, о чем можно было еще снять, когда уже вышли фильмы о динозаврах, доисторических монстрах, первобытных людях. В 2005 г. вышел последний (пока что) фильм Хлои Лелэнд и Тиа Хайнса «Прогулки с монстрами. Жизнь до динозавров», который фактически являлся приквелом ко всем фильмам данного цикла.

 В общем, расстарались.
В общем, расстарались. 
 Так, со следами уже совсем не заморачиваются, а пыль/песок/грязь летят все меньше и меньше. Кроме того, сэкономили на крупных планах, на которых компьютерную графику в ‘динозаврах’ заменяли куклы аниматроникс, добавлявшие правдивости и какого-то особого очарования показываемым животным. Здесь такого не наблюдается, зато зачем-то показывают внутренние процессы — изгибание позвоночника, сокращения сердца, трение камней в желудке, работу легких. На фоне остальных весьма условных объяснений это кажется каплей влаги в пустыне, к тому же — соленой.
Так, со следами уже совсем не заморачиваются, а пыль/песок/грязь летят все меньше и меньше. Кроме того, сэкономили на крупных планах, на которых компьютерную графику в ‘динозаврах’ заменяли куклы аниматроникс, добавлявшие правдивости и какого-то особого очарования показываемым животным. Здесь такого не наблюдается, зато зачем-то показывают внутренние процессы — изгибание позвоночника, сокращения сердца, трение камней в желудке, работу легких. На фоне остальных весьма условных объяснений это кажется каплей влаги в пустыне, к тому же — соленой.  .. Хотя, конечно, теоретически и те, и другие могли жить раньше и в Сибири, но для этого есть Спилберг и его ‘Юрский парк’, где как раз все ‘теоретически’, фантастически и вообще сказочно здорово. А от документальных фильмов ВВС ждешь немного другого.
.. Хотя, конечно, теоретически и те, и другие могли жить раньше и в Сибири, но для этого есть Спилберг и его ‘Юрский парк’, где как раз все ‘теоретически’, фантастически и вообще сказочно здорово. А от документальных фильмов ВВС ждешь немного другого. 

 И так далее.
И так далее. На этот раз — о жизни до динозавров. Лично мне фильм понравился, но лишь из-за общей тематики — жизнь до динозавров, то, что я люблю. Но лапов в нём — море.
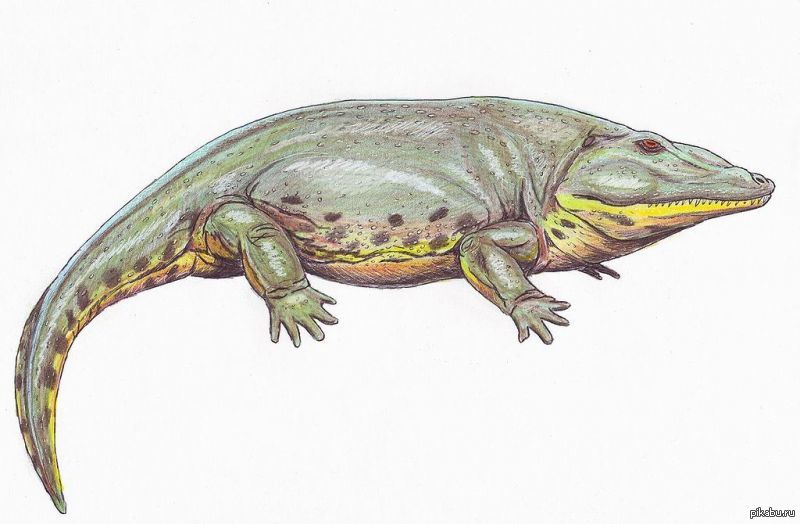
На этот раз — о жизни до динозавров. Лично мне фильм понравился, но лишь из-за общей тематики — жизнь до динозавров, то, что я люблю. Но лапов в нём — море. Раньше считали, что это был паук. Протерогиринус, древняя амфибия, показан вполне реалистично и достоверно, но каким образом он оказался в позднем карбоне, да ещё сразился с артроплеврой, столкнув её на удобно пристроенный колышек — непонятно. Петролакозавр тоже показан нормально, но в фильме от него происходит синапсид эдафозавр. В действительности он не был предком звероящеров. В качестве местообитания диметродона указывается Германия. Покащанный вид — Dimetrodon teutonis, в фильме он показан слишком крупным, так как в реальности этот вид был всего 1,4 метра в длину. Для тамошнего горгонопсида указывается род Gorgonops, который в реальности не был таким огромным, как та пятиметровая самка. К тому же горгонопсы не были собаконогими. Про скутозавра вообще молчу. Тероцефал из фильма ядовит, соотвественно он может быть эучамберсией, которая в действительности жила в пермском периода.
Раньше считали, что это был паук. Протерогиринус, древняя амфибия, показан вполне реалистично и достоверно, но каким образом он оказался в позднем карбоне, да ещё сразился с артроплеврой, столкнув её на удобно пристроенный колышек — непонятно. Петролакозавр тоже показан нормально, но в фильме от него происходит синапсид эдафозавр. В действительности он не был предком звероящеров. В качестве местообитания диметродона указывается Германия. Покащанный вид — Dimetrodon teutonis, в фильме он показан слишком крупным, так как в реальности этот вид был всего 1,4 метра в длину. Для тамошнего горгонопсида указывается род Gorgonops, который в реальности не был таким огромным, как та пятиметровая самка. К тому же горгонопсы не были собаконогими. Про скутозавра вообще молчу. Тероцефал из фильма ядовит, соотвественно он может быть эучамберсией, которая в действительности жила в пермском периода. Птеригот назван эвриптеридом, но это уже ближе к истине. Гинерпетон назван стегоцефалом. ‘Стегоцефалы’ — устаревший термин. Хайкоуихтис вообще своим именем не назван.
Птеригот назван эвриптеридом, но это уже ближе к истине. Гинерпетон назван стегоцефалом. ‘Стегоцефалы’ — устаревший термин. Хайкоуихтис вообще своим именем не назван.  Оно вышло после «Прогулок с динозаврами» и «Прогулок с чудовищами» и как бы завершило рассмотрение эволюции животных на нашей планете. Однако, несмотря на великолепную графику, слаженную работу аниматоров и исследователей и непревзойденную реалистичность животных, фильм имеет множество упущений и недостатков. Это заключается, прежде всего, в некоторых неверно изображенных деталях, составляющих достаточно весомые ошибки.
Оно вышло после «Прогулок с динозаврами» и «Прогулок с чудовищами» и как бы завершило рассмотрение эволюции животных на нашей планете. Однако, несмотря на великолепную графику, слаженную работу аниматоров и исследователей и непревзойденную реалистичность животных, фильм имеет множество упущений и недостатков. Это заключается, прежде всего, в некоторых неверно изображенных деталях, составляющих достаточно весомые ошибки.
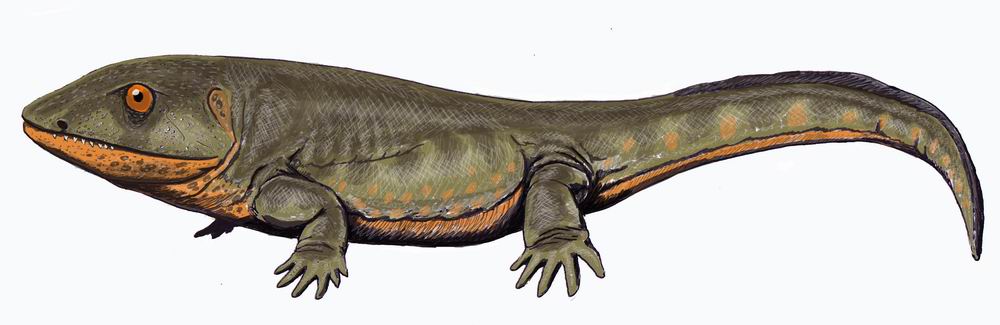 Эти пробелы современные исследователи привыкли заполнять посредством сравнения вымерших форм животных с рецентными, более близкими в своем происхождении или развивающимися конвергентно.
Эти пробелы современные исследователи привыкли заполнять посредством сравнения вымерших форм животных с рецентными, более близкими в своем происхождении или развивающимися конвергентно..jpg/1280px-Loxomma1DB_(flipped).jpg) Именно поэтому считают, что сибирский род Scutosaurus, следовательно, характерный для территории Лавразии, продолжающей в то время (255 млн. лет назад) отделяться от Гондваны, (слитых около 360 млн. лет тому назад в единый суперконтинент Пангея, и окончательно разделившихся около 150 млн. лет назад), никак не мог встретиться с представителями родов Diictodon, Gorgonops и Rhinesuchus, обитавших на территории современной Южной Африки, в прошлом Гондваны.
Именно поэтому считают, что сибирский род Scutosaurus, следовательно, характерный для территории Лавразии, продолжающей в то время (255 млн. лет назад) отделяться от Гондваны, (слитых около 360 млн. лет тому назад в единый суперконтинент Пангея, и окончательно разделившихся около 150 млн. лет назад), никак не мог встретиться с представителями родов Diictodon, Gorgonops и Rhinesuchus, обитавших на территории современной Южной Африки, в прошлом Гондваны. 
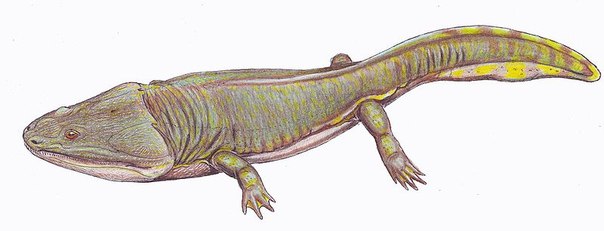 Или тем, что приелось. Или тем, что все монстры — очень неизвестны по сравнению с динозаврами и мамонтами с саблезубыми тиграми. Всё-таки малоизвестная жизнь до динозавров! Посмотрите фильмы, почитайте книги — везде либо динозавры, либо древние млекопитающие (мамонты, энтелодонт из одной экранизации ‘Затерянного мира ‘). То есть неизвестные незнакомцы с трудными именами.
Или тем, что приелось. Или тем, что все монстры — очень неизвестны по сравнению с динозаврами и мамонтами с саблезубыми тиграми. Всё-таки малоизвестная жизнь до динозавров! Посмотрите фильмы, почитайте книги — везде либо динозавры, либо древние млекопитающие (мамонты, энтелодонт из одной экранизации ‘Затерянного мира ‘). То есть неизвестные незнакомцы с трудными именами.
Стегоцефалы
Стегоцефа́лы[2][3], или панцирноголовые[2][3] (лат. Stegocephali, или Stegocephalia, от др.-греч. στέγος — «кровля» и κεφᾰλή — «голова»), — группа челюстноротых из клады тетраподоморф. Традиционно в таксон включали различных ранних «земноводных»; в рамках филогенетической номенклатуры стегоцефалами считаются все позвоночные с тетраподными конечностями (а также вторично их утратившие), в том числе и современные четвероногие (Tetrapoda)[1][4]. Характерной особенностью ранних стегоцефалов является строение черепа, получившее название стегального. Сплошной панцирь из кожных костей покрывал черепную коробку сверху и с боков, оставляя отверстия лишь для ноздрей, глаз и теменного органа[5].
Характерной особенностью ранних стегоцефалов является строение черепа, получившее название стегального. Сплошной панцирь из кожных костей покрывал черепную коробку сверху и с боков, оставляя отверстия лишь для ноздрей, глаз и теменного органа[5].
Систематика
Соотношение между определениями четвероногих. Звёздочкой выделено гипотетическое первое позвоночное с пятипалыми конечностями, которое также было одним из первых стегоцефалов. Синим цветом отмечены кроновые четвероногие.Отряд Stegocephali был введён американским палеонтологом Эдвардом Копом в 1868 году. Коп отнёс к стегоцефалам всех известных на тот момент ранних «земноводных», самые древние из которых происходят из отложений каменноугольного периода. Таксон широко использовался в XIX и начале XX века (часто под видоизменённым названием Stegocephalia), причём ключевым признаком стегоцефалов считалось наличие тетраподных конечностей. Когда были обнаружены девонские стволовые четвероногие ихтиостега (1932[6]) и акантостега (1952[7]), они сразу были отнесены к стегоцефалам[1][4].
К концу XX века — началу XXI века многие авторы отказались от выделения стегоцефалов как группы. В 1998 году палеонтолог Мишель Лорин[en] дал филогенетическое определение кладе Stegocephalia: все позвоночные, более родственные темноспондильным (Temnospondyli), чем пандерихту (Panderichthys)[8]. В 2020 году, в томе Phylonyms, с которым в силу вступил ФилоКод[en], Лорин отметил, что таксон Stegocephali имеет приоритет над Stegocephalia как введённый раньше. Он дал следующее определение: крупнейшая клада, включающая Eryops megacephalus Cope, 1877 (Temnospondyli), но не Tiktaalik roseae Daeschler et al., 2006, Panderichthys rhombolepis Gross, 1930 (Panderichthyidae) и Eusthenopteron foordi Whiteaves, 1881 (Osteolepiformes)[1][4]. Оба определения подразумевают включение современных четвероногих в состав стегоцефалов, что делает последних ныне живущей процветающей группой. «Переходная форма» тиктаалик попадает под первое определение[8], но заведомо исключается вторым[1][4].
В соответствии с определением 2020 года, стегоцефалы соответствуют примерно той же кладе, что и четвероногие в традиционном (апоморфном) понимании[1][4]. Лорин рассматривает четвероногих как кроновую группу, в которую не входят ранние позвоночные с тетраподной конечностью, такие как акантостега[9][10].
Филогенетическое положение стегоцефалов (Stegocephali sensu Laurin, 2020) может быть отображено следующей кладограммой[11][12][13][a]:
- ↑ Филогенетические определения:
- Eotetrapodiformes: ближайший общий предок Eusthenopteron и Ichthyostega и все его потомки[14];
- Стегоцефалы (Stegocephali): наибольшая клада, включающая Eryops megacephalus, но не Tiktaalik roseae, Panderichthys rhombolepis и Eusthenopteron foordi[1][1];
- Лабиринтодонты (Labyrinthodontia): наименьшая клада, включающая Acanthostega gunnari, Ichthyostega stensioei, Crassigyrinus scoticus, Mastodonsaurus giganteus, Baphetes kirkbyi и Anthracosaurus russelli[15][16];
- Четвероногие (Tetrapoda sensu lato): позвоночные с исходно пятипалыми конечностями[17][18][19];
- Четвероногие (Tetrapoda sensu stricto): наименьшая кроновая клада, включающая Homo sapiens, Caecilia tentaculata, Siren lacertina и Pipa pipa[9][10].
Элпистостега (Elpistostege) часто рассматривается как сестринский таксон тиктаалика. Однако в ревизии Клутье и соавторов 2020 года это род занимает более продвинутое положение и оказывается самым базальным стегоцефалом или четвероногим в широком смысле (если рассматривать неразветвлённые дистальные радиальные элементы как пальцы). Результаты анализа могут быть отображены следующей упрощённой кладограммой[20]:
Лабиринтодонты
За 18 лет до выделения стегоцефалов Герман Бурмейстер ввёл немного менее инклюзивную группу Labyrinthodontia (лабиринтодонты). Эти два названия использовались взаимозаменяемо, либо же лабиринтодонты считались подгруппой стегоцефалов. Буквально название Labyrinthodontia означает «лабиринтозубые». Дело в том, что на поперечном срезе зубов этих животных видно извилистые дентиновые складки, из-за чего зуб покрыт продольными бороздами. Такая же структура зубов характерна для более примитивных рыб-рипидистий. Высказывались предположения, что такая структура зубов изначально связана с наличием ядовитых желез в полости рта (в частности, нёбной железы). Но, возможно, лабиринтовая структура просто укрепляла зуб.
Но, возможно, лабиринтовая структура просто укрепляла зуб.
Таксон Labyrinthodontia долгое время не выделялся в рамках филогенетической номенклатуры. В 2020 году Лорин предложил определение, согласно которому лабиринтодонты представляют собой наименьшую кладу, включающую Acanthostega gunnari, Ichthyostega stensioei, Crassigyrinus scoticus, Mastodonsaurus giganteus, Baphetes kirkbyi и Anthracosaurus russelli. Таким образом, в данной классификации к лабиринтодонтам относится большинство стегоцефалов, в том числе четвероногие[15][16].
Примечания
- ↑ 1 2 3 4 5 6 7 8 9 de Queiroz, Cantino & Gauthier, 2020, Stegocephali E. D. Cope 1868 [M. Laurin], converted clade name, pp.
 741—745.
741—745.
- ↑ 1 2 Банников А. Г., Денисова М. Н., Даревский И. С. Класс Земноводные, или Амфибии (Amphibia) // Жизнь животных. В 7 т. / гл. ред. В. Е. Соколов. — 2‑е изд., перераб. — М. : Просвещение, 1985. — Т. 5 : Земноводные. Пресмыкающиеся / под ред. А. Г. Банникова. — С. 26. — 399 с. : ил.
- ↑ 1 2 Стегоцефалы / Буланов В. В. // Социальное партнёрство — Телевидение. — М. : Большая российская энциклопедия, 2016. — С. 211. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 31). — ISBN 978-5-85270-368-2.
- ↑ 1 2 3 4 5 6 Stegocephali (англ.). RegNum. Дата обращения: 29 июня 2021.
- ↑ Abel P.
 , Werneburg I. Morphology of the temporal skull region in tetrapods: research history, functional explanations, and a new comprehensive classification scheme (англ.) // Biological Reviews. — ISSN 1469-185X. — doi:10.1111/brv.12751.
, Werneburg I. Morphology of the temporal skull region in tetrapods: research history, functional explanations, and a new comprehensive classification scheme (англ.) // Biological Reviews. — ISSN 1469-185X. — doi:10.1111/brv.12751.
- ↑ Säve-Söderbergh G.ruen. Preliminary note on Devonian stegocephalians from East Greenland (англ.) // Meddelelser om Grønlandruen : journal. — 1932. — Vol. 94, no. 7. — P. 1—107. — doi:10.1017/S0016756800098769.
- ↑ Jarvik E.ruen. On the fish-like tail in the ichthyostegid stegocephalians with descriptions of a new stegocephalian and new crossopterygian from the Upper Devonian of East Greenland (англ.) // Meddelelser om Grønlandruen : journal.
 — 1952. — Vol. 114, no. 12. — P. 190.
— 1952. — Vol. 114, no. 12. — P. 190.
- ↑ 1 2 Laurin M.ruen. The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I. Systematics, middle ear evolution, and jaw suspension (англ.) // Ann. Sci. Nat., Zool., 13 Ser. — Paris, 1998. — Vol. 19. — P. 1—42.
- ↑ 1 2 de Queiroz, Cantino & Gauthier, 2020, Tetrapoda B. Hatschek and C. J. Cori 1896 [M. Laurin], converted clade name, pp. 759—764.
- ↑ 1 2 Tetrapoda (англ.). RegNum. Дата обращения: 21 июня 2021.
- ↑ Merck J. And Now For Something Completely Different: Sarcopterygii (англ.). GEOL 431 Vertebrate Paleobiology. Дата обращения: 1 июля 2021. Архивировано 24 апреля 2021 года.
- ↑ Merck J. Elpistostegalia becomes «digitized» (англ.). GEOL 431 Vertebrate Paleobiology. Дата обращения: 1 июля 2021. Архивировано 30 июня 2021 года.
- ↑ Swartz B. A Marine Stem-Tetrapod from the Devonian of Western North America (англ.) // PLOS One : journal. — 2012. — Vol. 7, iss. 3. — P. e33683. — ISSN 1932-6203. — doi:10.1371/journal.pone.0033683. — PMID 22448265. — S2CID 18496954.
- ↑ Coates M. I., Friedman M. Litoptychus bryanti and characteristics of stem tetrapod neurocrania (англ.) // Morphology, Phylogeny and Paleobiogeography of Fossil Fishes / In Elliot D. K., Maisey J. G., Yu X., Miao D. (eds.).. — 2010. — P. 389—416. — ISBN 978-3-89937-122-2.
- ↑ 1 2 3 de Queiroz, Cantino & Gauthier, 2020, Labyrinthodontia R. Owen 1859 [M. Laurin], converted clade name, pp. 747—750.
- ↑ 1 2 3 Labyrinthodontia (англ.
 ). RegNum. Дата обращения: 29 июня 2021.
). RegNum. Дата обращения: 29 июня 2021.
- ↑ Laurin M.ruen, Anderson J. S. Meaning of the Name Tetrapoda in the Scientific Literature: An Exchange (англ.) // Systematic Biology : journal. — 2004. — Vol. 53, iss. 1. — P. 68—80. — ISSN 1063-5157. — doi:10.1080/10635150490264716. — PMID 14965901. — S2CID 15922260.
- ↑ Lee M. S. Y. Stability of higher taxa in phylogenetic nomenclature — some comments on Moore (1998) (англ.) // Zoologica Scripta : journal. — 1999. — Vol. 28, iss. 3—4. — P. 361–366. — ISSN 1463-6409. — doi:10.1046/j.1463-6409.1999.00017.x. — S2CID 85990284.
- ↑ Anderson J. S. Use of Well-Known Names in Phylogenetic Nomenclature: A Reply to Laurin (англ.) // Systematic Biology : journal. — 2002. — Vol. 51, iss. 5. — P. 822—827. — ISSN 1063-5157. — doi:10.1080/10635150290102447. — PMID 12396594. — S2CID 45870887.
- ↑ Cloutier R.
 , Clement A. M., Lee M. S. Y., Noël R., Béchard I. Elpistostege and the origin of the vertebrate hand (англ.) // Nature : journal. — 2020. — Vol. 579, iss. 7800. — P. 549—554. — ISSN 1476-4687. — doi:10.1038/s41586-020-2100-8. — PMID 32214248. — S2CID 213171029.
, Clement A. M., Lee M. S. Y., Noël R., Béchard I. Elpistostege and the origin of the vertebrate hand (англ.) // Nature : journal. — 2020. — Vol. 579, iss. 7800. — P. 549—554. — ISSN 1476-4687. — doi:10.1038/s41586-020-2100-8. — PMID 32214248. — S2CID 213171029.
Литература
- Быстров А. П. и Ефремов И. А. Benthosuchus sushkini Efr. — лабиринтодонт из эотриаса реки Шарженги. Тр. Палеонтологического ин-та, Москва — Ленинград, изд. АН СССР, 1940, т. X, вып. 1, С. 1—152. Рис. 90. Тир. 850.
- Основы палеонтологии: Земноводные, пресмыкающиеся и птицы / под ред. А. К. Рождественского, Л. П. Татаринова. — М., 1964.
- Шмальгаузен И. И. Происхождение наземных позвоночных. — М., 1964.
- Carroll, R. L. 1995. Problems of the phylogenetic analysis of Paleozoic choanates. Bulletin du Museum national d’Histoire naturelle de Paris 4eme serie 17: 389—445.
- Laurin, M. & R. R. Reisz. 1997. A new perspective on tetrapod phylogeny. In S.
 Sumida and K. Martin (eds.) Amniotes Origins: Completing the Transition to Land: 9—59. London: Academic Press.
Sumida and K. Martin (eds.) Amniotes Origins: Completing the Transition to Land: 9—59. London: Academic Press. - Laurin M. & R. R. Reisz. 1999. A new study of Solenodonsaurus janenschi, and a reconsideration of amniote origins and stegocephalian evolution. Canadian Journal of Earth Sciences 36: 1239—1255.
- Vallin G. & M. Laurin. 2004. Cranial morphology and affinities of Microbrachis, and a reappraisal of the phylogeny and lifestyle of the first amphibians. Journal of Vertebrate Paleontology 24: 56—72.
- Phylonyms: A Companion to the PhyloCode (англ.) / de Queiroz K.ruen, Cantino P. D., Gauthier J. A., eds. — Boca Raton: Taylor & Francis Group, CRC Press, 2020. — 1352 p. — ISBN 978-1-138-33293-5.
Ссылки
Ароморфозы
Ароморфозы характеризуются. I. Общим повышением организации, II. развитием приспособлений широкого значения, III. расширением среды обитания.
1. Повышение организации. Под повышением организации понимаются морфо-физиологические изменения ее, которые: а) вызывают усиление жизнедеятельности организма, б) связаны с более глубокой функциональной диференцировкой частей, в) ведут к развитию более активных способов борьбы за существование.
У высших позвоночных, например, наблюдается следующая цепь ароморфозов:
- Разделение сердца на правую и левую половины и диференцировка двух кругов кровообращения. Одновременно: увеличение рабочей ёмкости легких.
- Следствия: более совершенное окисление крови и обильное снабжение органов кислородом, интенсификация функций органов.
- Диференцировка и специализация органов пищеварения. Следствия: более полное использование пищевых веществ, интенсификации обменных процессов, общее улучшение питания. Общее повышение активности жизнедеятельности (1, 2). Возникновение теплокровности. Усиление активности двигательных (локомоторных) органов. Усовершенствование конструкции локомоторных органов, соответствующее усилению их активности.

- Усложнение головного мозга и обострение органов чувств, развитие реакций поведения, в особенности — разумного типа, как ответов на быструю смену обстановки.
Все эти ароморфозы взаимно связаны. Конструктивно совершенные органы движения не имели бы значения без общего повышения активности; высокое развитие мозга и органов чувств было бы биологической нелепостью без высокой активности и совершенных органов движения и т. п. Теплокровность также связана с рядом особенностей — развитием пера или волоса, препятствующих отдаче тепла, с другой стороны, приспособлениями к саморегуляции тепла и т. д.
С точки зрения дарвиниста все эти гармонически сочетающиеся ароморфозы возникли не вдруг, а накапливались в результате выживания миксобиотипов с наиболее удачной комбинацией ароморфных признаков.
Перейдем теперь к оценке приспособительного значения ароморфозов.
2. Приспособления широкого значения. Описанные выше особенности высокой организации оказываются выгодными и полезными в весьма различных условиях среды и при относительно быстрых изменениях ее, почему при наличии указанных особенностей организм оказывается приспособленным не к каким-либо специальным (отдельным) условиям среды, а к многообразной жизненной обстановке.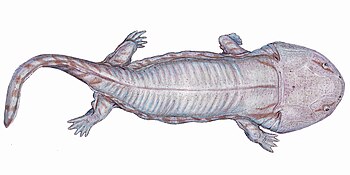 В этом смысле и говорится о приспособлениях «широкого значения». Например, обладание подвижными конечностями полезно (в особенности в виду их мультифункциональности) и в воде, и на горах, и в долине, и в лесу, и в пустыне и т. п. Высокое развитие головного мозга и органов чувств позволяет приспособляться к самой различной биотической обстановке и т. д.
В этом смысле и говорится о приспособлениях «широкого значения». Например, обладание подвижными конечностями полезно (в особенности в виду их мультифункциональности) и в воде, и на горах, и в долине, и в лесу, и в пустыне и т. п. Высокое развитие головного мозга и органов чувств позволяет приспособляться к самой различной биотической обстановке и т. д.
3. Расширение среды обитания. Приспособления «широкого значения» позволяют выйти за пределы первоначальной среды и занять новую среду с новыми свойствами. Выгода такого активного проникновения в новую среду самоочевидна, в особенности если последняя не включает равноценных конкурентов и освобождает организмы от конкурентов в прежней среде.
То, что было сказано по поводу приспособлений «широкого значения» и расширения среды обитания, показывает, что понятие ароморфоза включает не только морфо-физиологическую, но и экологическую характеристику.
Перейдем теперь к примерам ароморфозов.
4. Примеры ароморфозов. Перечисленные выше признаки ароморфной организации возникли, как указывалось, не вдруг. Однако появление ароморфной организации способствовало освобождению обладателей ее от тесных и зависимых связей с частными условиями среды. Вследствие этого ароморфозы открыли возможность проникновения в новую среду. Фактор среды, в свою очередь, создал отношения, при которых дальнейшее повышение организации стало одной из форм жизненной (биологической) необходимости.
Перечисленные выше признаки ароморфной организации возникли, как указывалось, не вдруг. Однако появление ароморфной организации способствовало освобождению обладателей ее от тесных и зависимых связей с частными условиями среды. Вследствие этого ароморфозы открыли возможность проникновения в новую среду. Фактор среды, в свою очередь, создал отношения, при которых дальнейшее повышение организации стало одной из форм жизненной (биологической) необходимости.
Так, выход древних кистеперых рыб на сушу стал возможным благодаря приобретению плавниками новой функции — ползания по дну водоема. Парный плавательный пузырь приобрел функцию примитивного легкого. Эти особенности послужили предпосылками к переходу (на протяжении рядов поколений) к наземному существованию. Существенным моментом, стимулировавшим этот процесс, было использование прибрежной полосы суши в качестве кормовой базы (питание насекомыми).
Таким образом, расширение функций плавников и плавательных пузырей и наземное питание были важными факторами, содействовавшими эволюции первых наземных позвоночных примитивных амфибий — стегоцефалов. Стегоцефалы появляются в девоне. Они были весьма далеки от понятия «высокой организации». В отношении органов чувств и развития головного мозга они, несомненно, уступали рыбам, в особенности хищным. Однако, обладание примитивными конечностями наземного типа и не менее примитивными легочными мешками позволили удлинять пребывание на суше и медленно ползать в поисках пищи — насекомых. Береговая водная и наземная зона, на протяжении рядов поколений, стала нишей стегоцефалов, перешедших, таким образом, к жизни амфибионтов.
Стегоцефалы появляются в девоне. Они были весьма далеки от понятия «высокой организации». В отношении органов чувств и развития головного мозга они, несомненно, уступали рыбам, в особенности хищным. Однако, обладание примитивными конечностями наземного типа и не менее примитивными легочными мешками позволили удлинять пребывание на суше и медленно ползать в поисках пищи — насекомых. Береговая водная и наземная зона, на протяжении рядов поколений, стала нишей стегоцефалов, перешедших, таким образом, к жизни амфибионтов.
В девонских условиях жизнь позвоночного — амфибионта была своеобразной. Выход в новую среду снижал конкуренцию (наиболее сильные конкуренты и хищники остались в воде). На земле конкурентов — позвоночных еще не было. Поэтому первоначально выход на сушу не стимулировал появления более высоко организованных форм. Однако, эта благоприятная обстановка (а только в ее условиях могли выжить малоподвижные, примитивные стегоцефалы) содействовала быстром росту численности древнейших земноводных и возрастанию конкуренции между ними. Не трудно понять, что в популяции стегоцефалов возникали новые миксобиотипы. Какие же из их числа могли получить преимущества?
Не трудно понять, что в популяции стегоцефалов возникали новые миксобиотипы. Какие же из их числа могли получить преимущества?
Мы сейчас проследим только один путь достижения такой преимущественности — путь накопления дальнейших ароморфозов. В условиях наземного существования преимущества доставляет интенсификация жизненный функций. Это мы наблюдаем у потомков стегоцефалов — примитивных рептилий с их новыми ароморфозами. В чем же выражается ароморфная организация примитивных рептилий, весьма близких, по своей организации, к стегоцефалам?
Сеймурия (примитивная рептилия) во многом сходна с эриопсом (стегоцефал, амфибия). Однако, у первобытных рептилий, подобных сеймурии, имелись важные отличительные признаки. У них наблюдается явственное развитие грудных ребер, отличных от. брюшных ребер. Этот факт указывает на большее развитие легких, т. е.— интенсификацию дыхательного процесса. Это, в свою очередь, свидетельствует об усилении окислительных и обменных процессов, а следовательно, с повышении функциональной активности органов.
В полном соответствии со сказанным выше, у древних примитивных рептилий (судя по скелетам) была несколько сильнее развита мускулатура, связанная с органами передвижения по земле. Этот признак указывает на усиление способности к передвижениям (Ромер, 1939). Едва ли можно сомневаться в том, что эта черта улучшала возможности питания. Интенсификация функции передвижения косвенно указывает на повышение других сторон организации и в частности — сердца. Сердце рептилий имеет разобщенные предсердия, а в желудочке намечается неполная перегородка, признаки, несомненно, связанные с повышением окислительных процессов. Таким образом, у примитивных рептилий имелись следующие ароморфозы: 1) прогрессивные изменения в строение сердца, 2) увеличение легких (усиление окислительной функции), 3) интенсификаци, локомоторных органов. К этим основным ароморфозам у рептилий позднее присоединяются еще следующие: 4) развитие роговой чешуи (защита против высыхания). 5) развитие плотной яйцевой скорлупы (защита от высыхания в условиях наземной среды), 6) развитие зародышевых оболочек и 7) большие запасы желтка в яйце. Все эти особенности позволили рептилиям укрепиться в новой среде, освободиться от обязательных связей с водной средой, проникнуть в сухие местообитания. Мы видим, таким образом, что отмеченные приспособления «широкого значения» расширили ареал рептилий, сделали их менее зависимыми от частных условий и более конкурентноспособными. В результате в мезозое рептилии вытесняют амфибий. Работа отбора, протекая в описанном направлении, вела к переходу на более высокую ступень организации. Процесс этот вполне наметился примерно к перми. Следовательно, стегоцефалы, продолжая существовать, уже в этот период дают начало более высокой организации, развивающейся по пути описанных ароморфозов.
Все эти особенности позволили рептилиям укрепиться в новой среде, освободиться от обязательных связей с водной средой, проникнуть в сухие местообитания. Мы видим, таким образом, что отмеченные приспособления «широкого значения» расширили ареал рептилий, сделали их менее зависимыми от частных условий и более конкурентноспособными. В результате в мезозое рептилии вытесняют амфибий. Работа отбора, протекая в описанном направлении, вела к переходу на более высокую ступень организации. Процесс этот вполне наметился примерно к перми. Следовательно, стегоцефалы, продолжая существовать, уже в этот период дают начало более высокой организации, развивающейся по пути описанных ароморфозов.
В следующем периоде — триасе — наблюдается аналогичная картина. Пресмыкающиеся претерпевают внутреннюю диференцировку на различные жизненные формы. Однако, в пределах примитивной группы звероящеров наблюдается развитие новых ароморфных особенностей, вызревание которых, под творческим контролем отбора, приводит к накоплению признаков нового класса — млекопитающих. Подъем на новую ступень и в этом случае связан с ароморфными признаками, на первых шагах едва позволяющими отличить рептилию от млекопитающих. Примером может служить нижнетриасовый циногнатус (Cynognathus), обладавший целым рядом ароморфных черт.
Подъем на новую ступень и в этом случае связан с ароморфными признаками, на первых шагах едва позволяющими отличить рептилию от млекопитающих. Примером может служить нижнетриасовый циногнатус (Cynognathus), обладавший целым рядом ароморфных черт.
Можно утверждать, что дыхание у циногнатуса было интенсивнее, а легкие обладали большей рабочей поверхностью. На это указывает явственное обособление хорошо развитой грудной клетки. Окислительные и обменные процессы подвергались дальнейшему усилению, и мы имеем право утверждать, что у циногнатуса развивается теплокровность. Это доказывается следующими данными. Хоаны у типичных рептилий расположены в передней часта нёба, у млекопитающих — в задней его части. Эта особенность связана со способом дыхания. Пресмыкающееся, захватив добычу, может без вреда для себя задержать дыхание. Млекопитающее (наземное) нуждается в более или менее ритмичном, непрерывном дыхании во время жевания пищи. У рептилий возможность относительно длительной задержки дыхания связана с менее интенсивными окислительными и обменными процессами и холоднокровностью. У наземных млекопитающих неспособность к длительной задержке дыхания связана с более интенсивными окислительными и обменными процессами и теплокровностью. Таким образом, переднее положение хоан указывает на холоднокровность, а заднее — на теплокровность. У циногнатуса имелось заднее положение хоан, и можно думать, что подобные ему формы уже обладали некоторой собственной температурой тела. Эти отношения также свидетельствуют о повышении энергии жизнедеятельности.
У наземных млекопитающих неспособность к длительной задержке дыхания связана с более интенсивными окислительными и обменными процессами и теплокровностью. Таким образом, переднее положение хоан указывает на холоднокровность, а заднее — на теплокровность. У циногнатуса имелось заднее положение хоан, и можно думать, что подобные ему формы уже обладали некоторой собственной температурой тела. Эти отношения также свидетельствуют о повышении энергии жизнедеятельности.
Млекопитающие и происходят от звероящеров из группы териодонт (Theriodontia), к которым принадлежит и циногнатус.
Теплокровность — в высокой степени важный ароморфоз, делающий животное значительно менее зависимым от температурных колебаний. Она им ела огромное значение. Во-первых, расширялись рамки суточной деятельности животного: в холодную ночь активность пресмыкающегося снижается. Для млекопитающих ночной холод не играет подобной роли. Во-вторых, теплокровность, в областях с сезонными колебаниями температуры, расширяет рамки годовой деятельности: зимой рептилии, как правило, не активны, млекопитающие, как правило, активны (зимняя спячка ряда млекопитающих имеет иное происхождение, чем неактивность рептилий). В-третьих, теплокровность позволила млекопитающим проникнуть в области с любым климатом и фактически завоевать всю поверхность земли.
В-третьих, теплокровность позволила млекопитающим проникнуть в области с любым климатом и фактически завоевать всю поверхность земли.
Развитие теплокровности связано у млекопитающих с рядом других особенностей. Прежде всего — с важными ароморфными изменениями сердца. Полное разделение сердца (правое и левое), обособление артериальной крови от венозной — привели к дальнейшему усовершенствованию окислительных процессов. Уже у циногнатуса и других териодонтов происходит явственная диференцировка зубов на клыки и коренные. С этим признаком связана жевательная функция, отсутствующая у рептилий и улучшающая усвоение питательных веществ. Обменные процессы стали, следовательно, интенсивнее. Энергия жизнедеятельности резко поднялась. Органы чувств и головной мозг достигли высокой степени развития. Реакции на воздействия среды усовершенствовались, развивается поведение разумного типа, приобретающее огромное значение при быстрых изменениях среды. Возникло живорождение и забота о потомстве, поднявшая биотический потенциал класса.
В результате, если в мезозое амфибии уступили господство рептилиям, то к концу мезозоя последние уступили его млекопитающим. Мы видим, таким образом, типичные проявления конкуренции, в результате которой происходил очевидный отбор на повышение организации и накопление ароморфозов все более высокого жизненного значения.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Конспект урока по биологии для 5 класса «Как развивалась жизнь на Земле»
Тема урока: Как развивалась жизнь на Земле.
Цель урока: создать условия для активизации познавательной активности учащихся; для формирования умений самостоятельно приобретать знания, работать с источниками информации.
Задачи:
· Образовательные- сформировать представление о происхождении жизни, о живых организмах существовавших в разные периоды развития жизни на Земле.
· Коррекционно — развивающие–развивать навыки самостоятельной работы с текстом, умения анализировать, обобщать материал, делать выводы, сравнивать
·
Воспитательные– прививать навыки
коммуникативного общения, формировать уважительное отношение учащихся ко всему
живому на Земле, к природе.
Тип урока: урок изучения и первичного закрепления новых знаний
Оборудование: учебник, тетрадь, компьютер, мультимедийный проектор, раздаточный материал.
Ход урока:
1. Организационный момент.
2. Актуализация знаний.
Ребята, изучая тему «Земля», мы выяснили, что наша планета уникальная, единственная и неповторимая в Солнечной системе, я хочу узнать у вас, а в чем заключается уникальность нашей планеты? Правильно, на ней есть живые организмы, жизнь. Человек издавна задавался вопросом, а как на Земле развивалась жизнь. Попробуем и мы с вами выяснить этот вопрос.
Постановка цели урока совместно с учащимися. Где возникла жизнь, как она развивалась, животные прошлого.
3. Изучение нового материала.
Мне в руки попала записка палеонтолога. А вы знаете кто такой палеонтолог? Но в этой записке не хватает слов. Ваша задача прослушать и вставить недостающие слова..
550
млн. лет назад
лет назад
Развитие
жизни на Земле началось 3,5 млр. лет назад, в океане, где для этого были
благоприятные условия. Самыми древними ископаемыми живыми организмами считаются
строматолиты- скопления синезеленых водорослей. Строматолиты обогатили
атмосферу Земли кислородом. Жизнь в древних океанах Земли была сосредоточена у
самого дна. Здесь обитали разнообразные многоклеточные животные, питавшиеся
органическими остатками. Одни из них неподвижно прикреплялись к грунту, другие
неторопливо перемещались по дну или плавали в нескольких сантиметрах над ним.
Их общей чертой были мягкие покровы и отсутствие твердого скелета .Примерно 5оо
млн. лет назад в океане уже водились разнообразные водоросли и всевозможные
животные: медузы, кораллы, морские лилии. Особенно много было трилобитов.2-75
см. жили у дна, где медленно плавали и ползали. Древние рыбы-динихтисы.Первые
наземные растения- псилофиты появились на Земле 420 млн. лет назад. Они
заселили участки суши по берегам водоемах. Первые растения не имели листьев, а
их стебли были покрыты чешуйками.
Закрепление: Текст записки: «Жизнь на Земле зародилась ________________. Это произошло _______ млр. лет назад. 500 млн. лет назад в океане водились ________________________________________________________ .По дну ползали дальние родственники раков _______________________ . В воде плавали хищные рыбы ____________ .проверка – один читает, читаем по цепочке. остальные проверяют. Подсчет баллов.
А сейчас мы с вами перенесемся в следующий период развития жизни на 350 млн. лет назад и узнаем какие же изменения произошли на нашей Земле. А поможет нам в этом наш помощник – учебник. Чтение текста на стр. 96 4 абз.
Закрепление: беседа по вопросам
1. Какой климат был на Земле 350 млн. лет назад? (влажным и теплым, а значит было много болот)
2. Какие растения росли на Земле? (были пышные леса из хвощей, плаунов, папоротников. Слайда 8). А сейчас они есть на Земле? Они видоизменилисьПоказ соврем. Хвощей из гербария.
3. Как называется этот период в истории Земли? (Каменноугольный)
4. А почему он так
называется?
А почему он так
называется?
5. Какие животные жили в лесах?(пауки, скорпионы, стрекозы).
6. Какие животные жили в многочисленных болотах? (стегоцефалы, произошли от рыб).
Стегоцефалы- это древние пресмыкающиеся, родственники лягушек.
Шло
время… На смену земноводным пришли произошедшие от них пресмыкающиеся, которых
называют древними ящерами. Это период в развитии земли называют Юрским
периодом. Поначалу они были невелики и напоминали современных ящериц. Их сухая
кожа, покрытая плотными роговыми чешуйками, препятствовала испарению влаги. Именно
это позволило пресмыкающимся прочно обосноваться на суше. 130 миллионов лет
назад на Земле царствовали настоящие гиганты- динозавры, бродили стада
диплодоков. Динозавры это одна из групп древних наземных пресмыкающихся.
Существующие в то время летающие и морские ящеры к динозаврам не относятся. А
можем ли мы сейчас в зоопарке увидеть динозавра? Они вымерли 65 млн. лет назад.
Ученые выдвигают разные гипотезы (падение метеорита, изменение климата,
нехватка пищи, появление млекопитающих),но гипотезы – это всего
лишь предположения, а истинная причина вымирания динозавров – так и останется
тайной.
Развитие жизни на земле продолжилось, появились цветковые растения, птицы, млекопитающие, человек…Первые млекопитающие появились примерно 170 млн лет назад. Они были невелики и внешне напоминали современных грызунов. Примерно 150 млн лет назад на Земле появились первые птицы. Самой древней из них считается археоптерикс. Внешне он больше всего напоминал покрытого перьями маленького динозавра. Его челюсти были вооружены острыми зубами. А на каждом крыле было развито по три пальца с сильными когтями. Ученые предполагают, что он использовал их, залезая на стволы и ветви деревьев, с которых планировал на землю.Первые растения с цветами появились примерно 130—125 млн лет назад. Яркие цветки привлекали различных насекомых, которые обеспечили надежную транспортировку пыльцы с одного растения на другое
Зрительная гимнастика.
Быстро или медленно развивалась жизнь на Земле? (очень медленно, миллиарды лет). Ребята, а как вы думаете, развитие жизни на земле закончилось?
Жизнь
продолжает развиваться, ученые выводят новые сорта растений, новые виды
животных. А некоторые оказались под угрозой исчезновения, в том числе по
вине человека, и наша задача – не допустить исчезновения и вымирания
современных растений и животных, а оберегать и охранять их.
А некоторые оказались под угрозой исчезновения, в том числе по
вине человека, и наша задача – не допустить исчезновения и вымирания
современных растений и животных, а оберегать и охранять их.
1. Диплодок. Он жил 140 млн. лет назад в Северной
Америке. Название переводят как «двуотростковый» (на позвонках хвоста внизу
были двойные отростки). Длина ящера до 27 м., причём хвост – 14 м., а шея – 7
м. Голова очень небольшая, диаметром 60 см. Масса около 60 тонн. Питался
растениями.
2. Брахиозавр. Жил 140 млн. лет назад в
Африке. Название переводится как «плечистый ящер». Длина почти 23 м., высота до
12 м., масса до 75 тонн. Благодаря своей длинной шее он мог доставить листья с
деревьев.
3. Компсогнат. Жил в Северной Америке.
Название переводят как «изящная челюсть». Это один из самых маленьких
динозавров. Животное с длинным хвостом и полыми костями, длина до 74 см., вес 3
кг. Питался ящерицами и насекомыми.
4. Трицератопс. Жил в Северной Америке.
Название переводят как «трёхрогий». Длина до 9 м., масса до 12 тонн. Рога
достигают 1 м. Питался растениями.5. Стегозавр. Жил в Северной Америке.
Название переводят как «ящер под крышей». Длина более 7 м., масса 4 тонны.
Голова диаметром 40 см. Питался растениями.
Длина до 9 м., масса до 12 тонн. Рога
достигают 1 м. Питался растениями.5. Стегозавр. Жил в Северной Америке.
Название переводят как «ящер под крышей». Длина более 7 м., масса 4 тонны.
Голова диаметром 40 см. Питался растениями.
6. Эвоплоцефал. Название означает «хорошо
защищённая голова». Под кожей имел прочный панцирь из костных пластин. На конце
хвоста нарост в форме гантели, что служило видимо, защитой от хищников. Длина
более 5 м., масса 3 тонны. Растительноядный.
7. Плезиозавр. Морской ящер. Имел широкую
голову и короткий хвост. 4 плавника помогали ему передвигаться в воде. Длина
12-15 м., вес до 10 тонн. Плезиозавра сравнивают со змеёй, продетой в туловище
черепаха. Питался моллюсками и рыбой.
8. Тираннозавр. Название переводят как
«ящер – тиран», «ящер – разбойник». Длина около 12 м., масса 6 тонн. Был
хищником, но не брезговал падалью. Особый вид тираннозавра – тираннозавр – рэкс
(король).
9. Птерозавр. Крылатый ящер. Внешне
напоминает птицу, но тело его, вероятно, было голым, без перьев. Крылья похожи
на крылья летучих мышей. Разные виды птерозавров имели разные размеры. Длина от
50 см., до 8 м.. Вес от 4 кг., до 3 тонн.
Крылья похожи
на крылья летучих мышей. Разные виды птерозавров имели разные размеры. Длина от
50 см., до 8 м.. Вес от 4 кг., до 3 тонн.
4. Подведение итогов урока. Рефлексия.
Итог урока: ребята, похвалите себя, что вы узнали на уроке.
5. Домашнее задание изучить п.9 стр. 52-56. Ответить на вопросы.
Класс пресмыкающиеся, подготовка к ЕГЭ по биологии
Пресмыкающиеся, или рептилии — класс наземных позвоночных животных типа хордовые, размножение и развитие которых более не зависят (как у земноводных) от водоема. Относятся к амниотам. Рептилии достигли наибольшего расцвета в мезозое.
На настоящий момент класс насчитывает около 10 тысяч видов.
Рептилии произошли от древних панцирноголовых земноводных — стегоцефалов, которые приспособились к защите тела от высыхания и
смогли освоить отдаленные от водоемов пространства.
Независимость от водоема досталась пресмыкающимся с трудом, для этого их организм приобрел ряд прогрессивных черт — ароморфозов:
- Зародышевые оболочки
- Внутреннее оплодотворение
- Сухая кожа
- Ячеистые легкие и грудная клетка
- Неполная перегородка в сердце
- Кора больших полушарий
- Вторичная почка
Как сказано выше, пресмыкающиеся относятся к амниотам. Это значит, что развитие их зародышей происходит в особом зародышевом пузыре с амниотической жидкостью — амнионе. Поразительно! Им более не нужен водоем, этот водоем всегда с собой, прямо в организме самки вокруг зародыша.
К зародышевым оболочкам относится наружная, серозная оболочка (сероза), внутренняя амниотическая оболочка и аллантоис. Аллантоис — зародышевый орган, с помощью которого осуществляется дыхание. Эти оболочки позволили пресмыкающимся расселиться от водоемов вглубь материков, занять огромные территории.
У рыб и земноводных оплодотворение было наружным, малоэффективным, с небольшой вероятностью встречи половых клеток. При
внутреннем оплодотворении, которое появляется у рептилий, семенная жидкость самца со сперматозоидами вводится непосредственно в
половые пути самки, что значительно увеличивает вероятность встречи мужских и женских гамет.
При
внутреннем оплодотворении, которое появляется у рептилий, семенная жидкость самца со сперматозоидами вводится непосредственно в
половые пути самки, что значительно увеличивает вероятность встречи мужских и женских гамет.
Кожа пресмыкающихся сухая, практически лишена желез (которых так много у земноводных). Верхние слои эпителия ороговевают, на поверхности кожи формируются роговые чешуйки. Эта защита необходима от пересыхания, помогает эффективнее удерживать воду в организме.
У пресмыкающихся происходит усложнение строения дыхательной системы: появляются ячеистые легкие, обеспечивающие более эффективный газообмен. Кожное дыхание (доминировавшее у амфибий) у пресмыкающихся сведено к минимуму или отсутствует вовсе.
Дыхательные пути у рептилий дифференцируются на гортань, трахею и бронхи. Дыхание реберного типа — в нем участвуют появившиеся вместе с ребрами межреберные мышцы. Ребра образуют
новую структуру скелета — грудную клетку.
Дыхание реберного типа — в нем участвуют появившиеся вместе с ребрами межреберные мышцы. Ребра образуют
новую структуру скелета — грудную клетку.
В сердце у пресмыкающихся развивается неполная межжелудочковая перегородка, способствующая более эффективному разделению артериальной и венозной крови. Это, в свою очередь, повышает эффективность обмена веществ (син. — метаболизм).
Тем не менее, над перегородкой кровь смешанная, так что пресмыкающиеся, как и земноводные, относятся к пойкилотермным (холоднокровным) животным.
У пресмыкающихся впервые возникают зачатки новой коры головного мозга, совершенствуется воспринимающая и интегрирующая
функции головного мозга. Становится возможным более сложное поведение.
Становится возможным более сложное поведение.
Возникает тазовая (вторичная) почка — метанефрос, выделительный каналец которой гораздо длиннее: становится возможным обратное всасывание веществ. Засушливый климат, в котором живут рептилии, располагает к экономии воды, их моча становится более концентрированной.
Отряды пресмыкающихся
В составе класса можно выделить 4 отряда, каждый из которых мы вкратце обсудим. Рептилии традиционно изучаются на примере типичного представителя — прыткой ящерицы, входящей в состав отряда чешуйчатые. С него мы и начнем знакомство с рептилиями.
Отряд чешуйчатые — прыткая ящерица
- Покровы, опорно-двигательная система
- Пищеварительная система
- Дыхательная система
- Кровеносная система
- Выделительная система
- Нервная система
- Половая система
Тело ящерицы покрыто сухой кожей, практически лишенной желез, с роговыми чешуями и щитками. Такое строение
покровов тела предотвращает высыхание организма, защищает от потери воды. Испарение через кожу
сохраняется, но в минимальном объеме.
Такое строение
покровов тела предотвращает высыхание организма, защищает от потери воды. Испарение через кожу
сохраняется, но в минимальном объеме.
Тело подразделяется на голову, туловище, хвост и две пары конечностей. Конечности расположены по бокам туловища, поэтому поднять голову высоко над землей пресмыкающиеся не могут. Конечности пятипалого типа, перепонки между пальцами отсутствуют.
Предполагаю, вам с детства известно о том, что ящерицы могут отбрасывать свои хвосты. Это действительно так, для ящериц характерна автотомия (от греч. autos — сам и греч. tome — отсечение) — способность животного самопроизвольно отбрасывать части своего тела.
В случае нападения
хищника автотомия может спасти жизнь ящерицы, так как отброшенный хвост приковывает к себе внимание и хищник
перестает преследовать ящерицу.
Скелет почти полностью окостеневший, более прочный, чем у земноводных. Позвоночник состоит из 5 отделов: шейный (8 позвонков), грудной, поясничный, крестцовый и хвостовой. Ребра, сочлененные одним концом с грудными позвонками, а другим — с грудиной, образуют замкнутую костную структуру — грудную клетку.
К ребрам крепятся впервые возникшие у пресмыкающихся межреберные мышцы, участвующие в дыхании.
За счет наличия 8 шейных позвонков значительно увеличивается подвижность головы, что имеет крайне важное значения для добывания пищи и ориентировке в окружающем мире. Полость тела рептилий — целом.
Скелеты поясов конечностей служат опорой для самих конечностей. Передний (плечевой) пояс состоит из парных ключиц, лопаток, вороньих
костей (коракоидов) и надгрудинника (надключицы), а также хрящей. Задний (тазовый) пояс конечностей образован тремя костями: седалищной,
лобковой и подвздошной, также включает в себя хрящевые прослойки между ними.
Задний (тазовый) пояс конечностей образован тремя костями: седалищной,
лобковой и подвздошной, также включает в себя хрящевые прослойки между ними.
Скелеты передних и задних свободных конечностей аналогичны по строению конечностям амфибий. Однако стоит заметить, что голень и предплечье не состоят из одной единой кости: они подразделяются на лучевую и локтевую (составляют предплечье), малоберцовую и большеберцовую (составляют голень).
Строение пищеварительной системы рептилий более дифференцировано по сравнению с таковым у земноводных.
Конические зубы прочно срастаются с челюстями.
Хорошо развиты слюнные железы, которые участвуют не только в смачивании пищи (как у амфибий), но и ферментативно обрабатывают пищевой комок за счет фермента — амилазы, расщепляющей углеводы.
Ротовая полость переходит в короткую глотку, которая продолжается в пищевод. Пищевод расширяется и переходит в желудок, от которого начинается тонкий, а затем толстый отдел кишечника. В тонкую кишку открываются протоки печени, поджелудочной железы и желчного пузыря.
Граница между тонкой и толстой кишкой особенно хорошо выражена: на ее месте располагается зачаток слепой кишки. Оканчивается пищеварительная система клоакой.
Кожа не участвует в дыхании, эту функцию берут на себя возникшие у рептилий ячеистые легкие. По сравнению с легочными мешками амфибий, ячеистые легкие имеют бо́льшую дыхательную поверхность.
Воздух присасывается в легкие благодаря движениям грудной клетки, которые происходят за счет сокращения межреберных
мышц. Через воздухоносные пути: хоаны, глотку, гортань, трахею и бронхи — воздух достигает пузырьков, альвеол,
стенка которых густо оплетена капиллярами, в которых и происходит газообмен.
Через воздухоносные пути: хоаны, глотку, гортань, трахею и бронхи — воздух достигает пузырьков, альвеол,
стенка которых густо оплетена капиллярами, в которых и происходит газообмен.
Поскольку рептилии холоднокровные животные, то интенсивность их обмена веществ напрямую зависит от температуры окружающей среды. Частота дыхательных движений также коррелирует с метаболизмом, поэтому чем выше температура окружающей среды, тем чаще дышит животное.
Кровеносная система замкнутая. Сердце трехкамерное: два предсердия и один желудочек с неполной перегородкой. Разделение крови в желудочке эффективнее, чем у земноводных, но над перегородкой кровь смешанная, поэтому рептилии — холоднокровные.
От сердца отходят 3 сосуда: легочная артерия, по которой венозная кровь поступает в легкие, где насыщается кислородом,
и две дуги аорты — правая и левая. От правой дуги аорты отходят сонные артерии, направляющиеся к головному мозгу.
От правой дуги аорты отходят сонные артерии, направляющиеся к головному мозгу.
Правая дуга аорты начинается от левой части желудка — вследствие чего у рептилий анатомически обусловлено поступление более насыщенной кислородом — артериальной крови — к головному мозгу, остальные органы получают смешанную кровь из левой дуги аорты.
По сравнению с земноводными выделительная система у рептилий более прогрессивного строения, представлена тазовыми (вторичными) почками, называемыми также — метанефрос, позволяющими гораздо эффективнее удерживать воду в организме, препятствуя ее избыточному удалению.
Выделительные канальцы становятся длиннее, за счет чего реабсорбция происходит лучше: из их просвета большее количество воды успевает обратно всосаться
в кровеносное русло. Это нужно для того, чтобы удержать воду в организме, ведь рептилии обитают в засушливых местах. Конечный продукт
обмена веществ у пресмыкающихся — мочевая кислота.
Конечный продукт
обмена веществ у пресмыкающихся — мочевая кислота.
Сравнивая головной мозг земноводных и пресмыкающихся, становятся заметны несколько существенных отличий.
Мозжечок развит лучше, чем у земноводных. У рептилий появляются зачатки новой коры переднего мозга, он значительно увеличивается в объеме и перестает быть исключительно обонятельным центром, вбирая в себя новые функции.
Для рептилий характерно более сложное (чем для амфибий) поведение, помимо заложенных генетически безусловных рефлексов, они способны вырабатывать адаптивные, условные рефлексы.
Органы чувств также претерпевают изменения. Аккомодация, настройка глаза на наилучшее видение объекта, теперь достигается не только
перемещением хрусталика вперед-назад (так было у амфибий), но и изменением кривизны хрусталика. Это называется — двойная аккомодация,
настройка глаза на резкость, так что видят рептилии лучше амфибий.
Это называется — двойная аккомодация,
настройка глаза на резкость, так что видят рептилии лучше амфибий.
Веки и мигательная перепонка служат для увлажнения поверхности глаза и предохраняют от высыхания.
Орган слуха также совершенствуется: появляется круглое окно во внутреннем ухе, улучшается передача звуковых колебаний, благодаря чему слух рептилий становится острее. Среднее ухо по-прежнему представлено стремечком и барабанной перепонкой.
Имеются слаборазвитые органы вкуса, локализующиеся в глотке, и органы тепловой чувствительности, особенно хорошо развитые у змей, расположенные между глазом и носом на лице. Органы обоняния находятся в хоанах. Хоаны — внутренние носовые отверстия, с помощью которых полость носа сообщается с глоткой.
Мужская половая система представлена семенниками и отходящими от них семяпроводами, впадающими в мочеточники..jpg/1280px-Seymouria2_(flipped).jpg) Мочеточники открываются в клоаку. У самцов имеются особые совокупительные органы, предназначенные для введения семенной жидкости
в половые пути самки. Оплодотворение у рептилий внутреннее.
Мочеточники открываются в клоаку. У самцов имеются особые совокупительные органы, предназначенные для введения семенной жидкости
в половые пути самки. Оплодотворение у рептилий внутреннее.
У самок половая система состоит из яичников, от которых отходят яйцеводы, впадающие в клоаку. Сперматозоиды оплодотворяют яйцеклетки в верхней части яйцевода.
По мере продвижения яйца вниз по яйцеводу оно покрывается сначала белковой оболочкой, а затем скорлуповой. Эти оболочки — приспособление к жизни на суше, обеспечивающие всестороннюю защиту зародыша.
Напомню, что пресмыкающиеся (также, как птицы и млекопитающие) относятся к группе амниот, зародыш у которых окружен зародышевыми оболочками: наружной — серозой, внутренней — амниотической. Имеется особый зародышевый орган дыхания — аллантоис.
На этом наша беседа о прыткой ящерице подходит к концу. Впереди ждут другие представители рептилий. Из отряда чешуйчатых, помимо
прыткой ящерицы, хочется отметить гекконов — мелких примитивных ящериц, ведущих ночной образ жизни, отлично лазающих по деревьям,
скалам и стенам домов. Самые крупные представители чешуйчатых — вараны, обитающие в Южной Азии, Африке, — достигают в длину до 3,5 метров.
Впереди ждут другие представители рептилий. Из отряда чешуйчатых, помимо
прыткой ящерицы, хочется отметить гекконов — мелких примитивных ящериц, ведущих ночной образ жизни, отлично лазающих по деревьям,
скалам и стенам домов. Самые крупные представители чешуйчатых — вараны, обитающие в Южной Азии, Африке, — достигают в длину до 3,5 метров.
Змеи также принадлежат к отряду чешуйчатые. Это безногие животные, адаптировавшиеся к перемещениям в густом лесном покрове, кронах деревьев. У змей взгляд немигающий: отсутствуют подвижные веки. Однако их отсутствие не означает, что поверхность глаз не нуждается в увлажнении: у них имеется мигательная перепонка, смачивающая поверхность глаза.
Змеи подразделяются на ядовитых и неядовитых. Ядовитыми являются гадюка обыкновенная, песчаная эфа, черная мамба, кобра,
грюза, гремучие и морские змеи. К неядовитым относятся полозы, ужи, удавы.
К неядовитым относятся полозы, ужи, удавы.
Черепахи
Имеют уплощенное тело, покрытое панцирем из двух щитков: брюшного и спинного. Примечательно, что на задних конечностях имеются межпальцевые перепонки для плавания. Морские черепахи продвинулись еще дальше: у них задние конечности видоизменены в ласты, которые служат приспособлением к водному образу жизни.
Есть растительноядные и хищные формы. Настоящих зубов у черепах нет, их челюсти снабжены режущим роговым краем — клювом.
Крокодилы
Крокодилы — еще один отряд класса рептилий. Крокодилы населяют реки и озера тропических стран. Это крупные животные,
тело которых покрывают роговые чешуи. На задних конечностях крокодилы имеют межпальцевые перепонки. Зубы остроконечные, крупные,
располагаются в альвеолах челюсти, как и у млекопитающих.
Легкие имеют более прогрессивное строение — губчатое. Кровеносная система включает в себя четырех (!) камерное сердце, кровь в сердце не смешивается, так как желудочек полностью разделяется межжелудочковой перегородкой на две камеры.
Однако, крокодилы все равно остаются холоднокровными животными: кровь смешивается в спинной аорте, образованной в результате слияния левой и правой дуги аорты.
Вероятно, с детства многим известна история про птицу — крокодилова сторожа, которая питается остатками пищи из пасти крокодила. Спешу сообщить, что это скорее миф, чем правда. Крокодилы вовсе не нуждаются в подобной «чистке», а птицы предпочитают питаться насекомыми, в том числе мухами, которые слетаются на остатки мяса в пасти крокодила.
Клювоголовые
Мы добрались до самого древнего отряда рептилий. На сегодняшний день остался единственный его представитель — гаттерия,
обитающая на нескольких островах Новой Зеландии. По внешнему виду напоминает игуану, ведет ночной образ жизни, питается насекомыми,
моллюсками, червями. Гаттерия является вымирающим видом.
На сегодняшний день остался единственный его представитель — гаттерия,
обитающая на нескольких островах Новой Зеландии. По внешнему виду напоминает игуану, ведет ночной образ жизни, питается насекомыми,
моллюсками, червями. Гаттерия является вымирающим видом.
Значение пресмыкающихся
Как и все живые организмы, пресмыкающиеся являются звеном в цепи питания (консументами). Регулируют численность насекомых, питаясь ими. Среди рептилий встречаются как хищники, так и растительноядные формы.
Рептилии имеют пищевое значение. В Африке и Азии употребляют в пищу около сотни различных блюд из мяса змей; яйца, жир и мясо морской черепахи также употребляются в пищу. Из кожи крокодилов изготавливают различные изделия.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
| Исходные данные | |
|---|---|
| | |
| Ранг | |
| Taxonyyy (GBIF) | Жизнь: Animalia: Arthropoda: Malacostraaca: Amphipoda: STEGOCEPHALIDAE: Stegocephalus |
| таксономический Статус (GBIF) | принято |
| классификация (GBIF) | |
| Научное название | |
| STEGOCHALUS KROYER, 1842 | |
| Имя Опубликовано в | Naturhist. Tidsskrift, 4 (2) Tidsskrift, 4 (2) |
| Общее имя(я) | フクレソコエビ |
| Барнард, Дж. Лоренс и Гордон С. Караман, 1991: The Marine Amphipodan Gammariden Generas of Families and Gammarodan Gammaroide. Записи Австралийского музея, приложение 13, pt. 2. 419-866. — через Интегрированную таксономическую информационную систему (ITIS) | |
| De Broyer, C.; Лоури, Дж. К.; Язджевски, К. и Роберт, Х. (2007). Каталог Gammaridean и Corophiidean Amphipoda (Crustacea) Южного океана с данными о распространении и экологии.В: Де Бройер К. (ред.). Перепись антарктической морской жизни: обзор амфипод Южного океана. Том. I. Бюллетень Королевского института естественных наук Бельгии, Биология. 77, доп. 1: 1-325. — через Всемирный регистр морских видов | |
| Интегрированная таксономическая информационная система (ITIS). — через Всемирный реестр морских видов | |
Нив, Шеффилд Эйри. (1939-1996). Номенклатор Zoologicus vol. 1-10 онлайн. [разработано uBio, размещено онлайн в библиотеке MBLWHOI].- через Всемирный реестр морских видов [разработано uBio, размещено онлайн в библиотеке MBLWHOI].- через Всемирный реестр морских видов | |
| CoL2006 — через Временный реестр морских и неморских родов | |
| Nomenclator Zoologicus. Список названий родов и подродов в зоологии от десятого издания Линнея, 1758 г. до конца 2004 г. Оцифровано uBio из тт. 1–9 Neave (ред.), 1939–1996, плюс дополнительный цифровой том. http://ubio.org/NomenclatorZoologicus (по состоянию на 2006 г.). — через Временный реестр морских и неморских родов | |
| SN2000 непроверенный — через Временный реестр морских и неморских родов | |
| WoRMS (март 2013 г.) , Ганс Г.(2008) MESS — Marina Evertebrater från Södra Skandinavien (Морские беспозвоночные из Южной Скандинавии) — учебное пособие для студентов. .tmbl.gu.se/staff/HansGHanssonP.html» — через Dyntaxa. Шведские таксономические базы данных | |
Кройер, Х. (1842) Nye nordiske Arter og Slaegter af Amphipodernes Orden, henhørende til Familien Gammarina (Foreløbigt Uddrag af ett større Arbeide) — через Dyntaxa. Свенские таксономические базы данных Свенские таксономические базы данных | |
| Неопубликованный список Лоури 2006 г. — через Временный регистр морских и неморских родов | |
| Натурист. Тидскрифт, 4 (2) — через Временный реестр морских и неморских родов |
Stegocephalus inflatus
Stegocephalus inflatusК сожалению, GBIF не работает должным образом без включенного JavaScript.
Наш веб-сайт обнаружил, что вы используете устаревший небезопасный браузер, который не позволяет вам пользоваться сайтом.Мы предлагаем вам перейти на современный браузер.
- Набор данных
- Таксономия Европейского нуклеотидного архива (ENA)
- Классифицировать
- РАЗНОВИДНОСТЬ
Классификация
- царство
- Метазоа
- тип
- Членистоногие
- класс
- Малакострака
- приказ
- Амфипода
- семья
- Стегоцефалы
- род
- Стегоцефал
- разновидность
- Стегоцефал инфлатус
Имя
- Омонимы
- Стегоцефал инфлатус
Функциональная интерпретация JSTOR
Абстрактный Богатые железом октаэдрические кристаллы были описаны старшим автором в клетках слепой кишки кишечника амфиподы Stegocephaloides christianiensis. Настоящее исследование выявило их присутствие у других видов семейства Stegocephalidae (Bathystegocephalus inflatus, Euandania ingens, Parandania boecki, Stegocephaloides auratus, S. vanhoffeni, Stegocephalus infaltus, Phippsiella spp. и Parandaniexis sp. (ср. mirabilis). Кристаллы не обнаружены. у Andaniopsis nordlandica, Tetradeion crassum или Andaniexis abyssi, хотя последний давал тканевую реакцию на железо Fe-клетки содержат только по одному кристаллу у всех видов, причем кристаллы постоянно увеличиваются в размерах проксимально в каждой слепой кишке.Самая дистальная часть слепой кишки была лишена кристаллов. Подробная работа была ограничена Stegocephaloides christianiensis, Stegocephalus inflatus и Parandaniexis sp. (ср. мирабилис). Описана ультраструктура слепой кишки S. christianiensis: различимы две фации клеток (R/F и В-клетки). Клетки R/F (= клетка Fe) столбчатые, с терминальной щеточной каймой из длинных микроворсинок. В этих клетках обнаруживаются липидные глобулы, гликоген, кристаллы железа и гранулы кальция.
Настоящее исследование выявило их присутствие у других видов семейства Stegocephalidae (Bathystegocephalus inflatus, Euandania ingens, Parandania boecki, Stegocephaloides auratus, S. vanhoffeni, Stegocephalus infaltus, Phippsiella spp. и Parandaniexis sp. (ср. mirabilis). Кристаллы не обнаружены. у Andaniopsis nordlandica, Tetradeion crassum или Andaniexis abyssi, хотя последний давал тканевую реакцию на железо Fe-клетки содержат только по одному кристаллу у всех видов, причем кристаллы постоянно увеличиваются в размерах проксимально в каждой слепой кишке.Самая дистальная часть слепой кишки была лишена кристаллов. Подробная работа была ограничена Stegocephaloides christianiensis, Stegocephalus inflatus и Parandaniexis sp. (ср. мирабилис). Описана ультраструктура слепой кишки S. christianiensis: различимы две фации клеток (R/F и В-клетки). Клетки R/F (= клетка Fe) столбчатые, с терминальной щеточной каймой из длинных микроворсинок. В этих клетках обнаруживаются липидные глобулы, гликоген, кристаллы железа и гранулы кальция. В-клетки имеют люминальную границу коротких коротких микроворсинок с апикальным комплексом мембраносвязанных везикул различной степени слияния.Состав кристаллов Fe описан с помощью рентгенозондового анализа. Выявлены сильные пики Fe вместе с небольшими пиками для Si, P, S, Cl, K, Ca, Cu и Zn. Эти элементы были идентифицированы и в окружающей цитоплазме. Кристаллический состав однородный, без отдельного ядра. Кристалл состоит из гексагонально расположенных электронно-плотных ядер диаметром 5,8 ± 0,3 нм с межъядерными расстояниями 10,5 ± 0,5 нм, 7,5 ± 0,5 нм и 9,5 ± 0,5 нм. Анализ ядер с помощью широкоугольной электронной дифракции дал четыре кольца с расстоянием d, равным 0.250, 0,223, 0,191 и 0,145 нм (все ± 0,003 нм). По этим основаниям вещество кристаллов идентифицируется как ферритин. Кристаллы ферритина выделяются с фекалиями Stegocephaloides christianiensis, что указывает на роль в экскреции железа, возможно, как часть процесса регуляции содержания в организме. Содержание железа в S.
В-клетки имеют люминальную границу коротких коротких микроворсинок с апикальным комплексом мембраносвязанных везикул различной степени слияния.Состав кристаллов Fe описан с помощью рентгенозондового анализа. Выявлены сильные пики Fe вместе с небольшими пиками для Si, P, S, Cl, K, Ca, Cu и Zn. Эти элементы были идентифицированы и в окружающей цитоплазме. Кристаллический состав однородный, без отдельного ядра. Кристалл состоит из гексагонально расположенных электронно-плотных ядер диаметром 5,8 ± 0,3 нм с межъядерными расстояниями 10,5 ± 0,5 нм, 7,5 ± 0,5 нм и 9,5 ± 0,5 нм. Анализ ядер с помощью широкоугольной электронной дифракции дал четыре кольца с расстоянием d, равным 0.250, 0,223, 0,191 и 0,145 нм (все ± 0,003 нм). По этим основаниям вещество кристаллов идентифицируется как ферритин. Кристаллы ферритина выделяются с фекалиями Stegocephaloides christianiensis, что указывает на роль в экскреции железа, возможно, как часть процесса регуляции содержания в организме. Содержание железа в S. christianiensis и ряде других прибрежных амфипод было исследовано с помощью атомно-абсорбционной спектрофотометрии. Содержание железа нелинейно связано с сухой массой тела у S. christianiensis и не может быть просто объяснено как следствие поверхностной абсорбции.Уровни железа у S. christianiensis были выше, чем у многих других исследованных видов. Морфология ротового аппарата S. christianiensis была исследована с помощью сканирующей электронной микроскопии. Анализ свежего содержимого желудка выявил книдариевые нематоцисты, которые по размеру и форме соответствовали таковым из Adamsia carciniopados, Pennatula Phosphorea и Hydractinia echinata. Наблюдения за поведением живых S. christianiensis показали, что Pennatula была вероятной добычей. Исследования ареала книдарий и нескольких известных хищников книдарий (Pycnogonum, Hyperia) подтвердили, что отделяемый аконтий Adamsia и мягкие ткани Pennatula содержат необычно высокие концентрации железа.Предполагается, что производство и выделение кристаллов ферритина S.
christianiensis и ряде других прибрежных амфипод было исследовано с помощью атомно-абсорбционной спектрофотометрии. Содержание железа нелинейно связано с сухой массой тела у S. christianiensis и не может быть просто объяснено как следствие поверхностной абсорбции.Уровни железа у S. christianiensis были выше, чем у многих других исследованных видов. Морфология ротового аппарата S. christianiensis была исследована с помощью сканирующей электронной микроскопии. Анализ свежего содержимого желудка выявил книдариевые нематоцисты, которые по размеру и форме соответствовали таковым из Adamsia carciniopados, Pennatula Phosphorea и Hydractinia echinata. Наблюдения за поведением живых S. christianiensis показали, что Pennatula была вероятной добычей. Исследования ареала книдарий и нескольких известных хищников книдарий (Pycnogonum, Hyperia) подтвердили, что отделяемый аконтий Adamsia и мягкие ткани Pennatula содержат необычно высокие концентрации железа.Предполагается, что производство и выделение кристаллов ферритина S. christianiensis и другими видами семейства Stegocephalidae, потребляющими книдарии, представляет собой систему регуляции железа у животных, испытывающих диетическую нехватку железа.
christianiensis и другими видами семейства Stegocephalidae, потребляющими книдарии, представляет собой систему регуляции железа у животных, испытывающих диетическую нехватку железа.
Королевское общество является самоуправляемым объединением многих самых выдающихся ученых мира, представляющих все области науки, техники и медицины, и является старейшей непрерывно существующей научной академией.Фундаментальная цель Общества, отраженная в его учредительных Уставах 1660-х годов, состоит в том, чтобы признавать, продвигать и поддерживать превосходство в науке, а также поощрять развитие и использование науки на благо человечества. Общество сыграло роль в некоторых из самых фундаментальных, значительных и судьбоносных открытий в истории науки, и ученые Королевского общества продолжают вносить выдающийся вклад в науку во многих областях исследований.
Кристаллы ферритина в слепой кишке кишечника stegocephaloides christianiensis boeck и других стегоцефаловых (amphipoda gammaridea): функциональная интерпретация
Богатые железом октаэдрические кристаллы были описаны старшим автором в клетках слепой кишки кишечника амфиподы Stegocephaloides christianiensis . Настоящее исследование показало их присутствие в других видах семейства Stegocephalidae ( Bathystegocephalus inflatus , Euandania ingens , Parandania boecki , Stegocephaloides Auratus , С. vanhojfeni , Stegocephalus inflatus , Phippsiella SPP. и Parandaniexis sp.(ср. mirabilis ).У Andaniopsis nordlandica , T etradeion crassum или Andaniexis abyssi кристаллы не обнаружены, хотя последний дал тканевую реакцию на железо.Клетки Fe содержат только один кристалл у всех видов, и кристаллы постоянно увеличиваются в размерах проксимально в каждой слепой кишке. Самая дистальная часть слепой кишки была лишена кристаллов. Подробная работа была ограничена Stegocephaloides christianiensis , Stegocephalus inflatus и Parandaniexis sp. (ср. мирабилис ). Описана ультраструктура слепой кишки S. christianiensis : различимы две фации клеток (R/F и В-клетки).Клетки R/F (=клетки Fe) столбчатые, с концевой щеточной каймой из длинных микроворсинок.
Настоящее исследование показало их присутствие в других видах семейства Stegocephalidae ( Bathystegocephalus inflatus , Euandania ingens , Parandania boecki , Stegocephaloides Auratus , С. vanhojfeni , Stegocephalus inflatus , Phippsiella SPP. и Parandaniexis sp.(ср. mirabilis ).У Andaniopsis nordlandica , T etradeion crassum или Andaniexis abyssi кристаллы не обнаружены, хотя последний дал тканевую реакцию на железо.Клетки Fe содержат только один кристалл у всех видов, и кристаллы постоянно увеличиваются в размерах проксимально в каждой слепой кишке. Самая дистальная часть слепой кишки была лишена кристаллов. Подробная работа была ограничена Stegocephaloides christianiensis , Stegocephalus inflatus и Parandaniexis sp. (ср. мирабилис ). Описана ультраструктура слепой кишки S. christianiensis : различимы две фации клеток (R/F и В-клетки).Клетки R/F (=клетки Fe) столбчатые, с концевой щеточной каймой из длинных микроворсинок. В этих клетках обнаруживаются липидные глобулы, гликоген, кристаллы железа и гранулы кальция. В-клетки имеют люминальную границу коротких коротких микроворсинок с апикальным комплексом мембраносвязанных везикул различной степени слияния. Состав кристаллов Fe описан с помощью рентгенозондового анализа. Выявлены сильные пики Fe вместе с небольшими пиками для Si, P, S, Cl, K, Ca, Cu и Zn. Эти элементы были идентифицированы и в окружающей цитоплазме.Кристаллический состав однородный, без отдельного ядра. Кристалл состоит из гексагонально расположенных электронно-плотных ядер диаметром 5,8 + 0,3 нм с межъядерными расстояниями 10,5 ± 0,5 нм, 7,5 + 0,5 нм и 9,5 + 0,5 нм. Широкоугольный электронографический анализ ядер дал четыре кольца с интервалами d 0,250, 0,223, 0,191 и 0,145 нм (все + 0,003 нм). По этим основаниям вещество кристаллов идентифицируется как ферритин. Кристаллы ферритина выделяются с фекалиями Stegocephaloides christianiensis , что указывает на роль в экскреции железа, возможно, как часть процесса регуляции содержания в организме.
В этих клетках обнаруживаются липидные глобулы, гликоген, кристаллы железа и гранулы кальция. В-клетки имеют люминальную границу коротких коротких микроворсинок с апикальным комплексом мембраносвязанных везикул различной степени слияния. Состав кристаллов Fe описан с помощью рентгенозондового анализа. Выявлены сильные пики Fe вместе с небольшими пиками для Si, P, S, Cl, K, Ca, Cu и Zn. Эти элементы были идентифицированы и в окружающей цитоплазме.Кристаллический состав однородный, без отдельного ядра. Кристалл состоит из гексагонально расположенных электронно-плотных ядер диаметром 5,8 + 0,3 нм с межъядерными расстояниями 10,5 ± 0,5 нм, 7,5 + 0,5 нм и 9,5 + 0,5 нм. Широкоугольный электронографический анализ ядер дал четыре кольца с интервалами d 0,250, 0,223, 0,191 и 0,145 нм (все + 0,003 нм). По этим основаниям вещество кристаллов идентифицируется как ферритин. Кристаллы ферритина выделяются с фекалиями Stegocephaloides christianiensis , что указывает на роль в экскреции железа, возможно, как часть процесса регуляции содержания в организме. Содержание железа в S. christianiensis и ряде других прибрежных амфипод было исследовано с помощью атомно-абсорбционной спектрофотометрии. Содержание железа было нелинейно связано с сухой массой тела у S. christianiensis и не может быть просто объяснено как следствие поверхностной адсорбции. Уровни железа у S. christianiensis были выше, чем у многих других исследованных видов. Морфология ротового аппарата S. christianiensis была исследована с помощью сканирующей электронной микроскопии.Анализ свежего содержимого желудка выявил книдариевые нематоцисты, которые по размеру и форме соответствовали таковым из Adamsia carciniopados , Pennatula фосфореа и Hydractinia echinata . Поведенческие наблюдения за живым S. christianiensis показали, что Pennatula был вероятной добычей. Исследования ряда книдарий и нескольких известных хищников книдарий ( Pycnogonum , Hyperia ) подтвердили, что отделяемый аконций Adamsia и мягкие ткани Pennatula содержали необычно высокие концентрации железа.
Содержание железа в S. christianiensis и ряде других прибрежных амфипод было исследовано с помощью атомно-абсорбционной спектрофотометрии. Содержание железа было нелинейно связано с сухой массой тела у S. christianiensis и не может быть просто объяснено как следствие поверхностной адсорбции. Уровни железа у S. christianiensis были выше, чем у многих других исследованных видов. Морфология ротового аппарата S. christianiensis была исследована с помощью сканирующей электронной микроскопии.Анализ свежего содержимого желудка выявил книдариевые нематоцисты, которые по размеру и форме соответствовали таковым из Adamsia carciniopados , Pennatula фосфореа и Hydractinia echinata . Поведенческие наблюдения за живым S. christianiensis показали, что Pennatula был вероятной добычей. Исследования ряда книдарий и нескольких известных хищников книдарий ( Pycnogonum , Hyperia ) подтвердили, что отделяемый аконций Adamsia и мягкие ткани Pennatula содержали необычно высокие концентрации железа. Предполагается, что производство и выделение кристаллов ферритина S. christianiensis и другими видами семейства Stegocephalidae, потребляющими книдарии, представляет собой систему регуляции железа у животных, испытывающих диетическую нехватку железа.
Предполагается, что производство и выделение кристаллов ферритина S. christianiensis и другими видами семейства Stegocephalidae, потребляющими книдарии, представляет собой систему регуляции железа у животных, испытывающих диетическую нехватку железа.
Страница не найдена | Отдел научных публикаций
— Любой -MFR 83(1-2), 2021MFR 82(3-4), 2020MFR 82(1-2), 2020MFR 81(3-4), 2019MFR 81(2), 2019MFR 81(1), 2019MFR 80( 4), 2018MFR 80(3), 2018MFR 80(2), 2018MFR 80(1), 2018MFR 79(3-4), 2017MFR 79(2), 2017MFR 79(1), 2017MFR 78(3-4), 2016MFR 78(1-2), 2016MFR 77(4), 2015MFR 77(3), 2015MFR 77(2), 2015MFR 77(1), 2015MFR 76(4), 2014MFR 76(3), 2014MFR 76(1-2) , 2014МФР 75(4), 2013МФР 75(3), 2013МФР 75(1-2), 2013МФР 74(4), 2012МФР 74(3), 2012МФР 74(2), 2012МФР 74(1), 2012МФР 73(4) , 2011 MFR 73 (3), 2011 MFR 73 (2), 2011 MFR 73 (1), 2011 MFR 72 (4), 2010 MFR 72 (3), 2010 MFR 72 (2), 2010 MFR 72 (1), 2010 MFR 71 (4), 2009 MFR 71(3), 2009MFR 71(2), 2009MFR 71(1), 2009MFR 70(3-4), 2008MFR 70(2), 2008MFR 70(1), 2008MFR 69(1-4), 2007MFR 68(1- 4), 2006 MFR 67 (4), 2005 MFR 67 (3), 2005 MFR 67 (2), 2005 MFR 67 (1), 2005 MFR 66 (4), 2004 MFR 66 (3), 2004 MFR 66 (2), 2004 MFR 66 (1) , 2004 MFR 65 (4), 2003 MFR 65 (3), 2003 MFR 65 (2), 2003 MFR 65 (1), 2003 MFR 64 (4), 2002 MFR 64 (3), 2002 MFR 64 (2), 2002 MFR 64 (1), 2002 MFR 63 (4), 2001 ППР 63 (3), 2001 ППР 63 (2) , 2001 MFR 63 (1), 2001 MFR 62 (4), 2000 MFR 62 (3), 2000 MFR 62 (2), 2000 MFR 62 (1), 2000 MFR 61 (4), 1999 MFR 61 (3), 1999 MFR 61 (2), 1999 MFR 61 (1), 1999 ПТР 60 (4), 1998 ППР 60 (3), 1998 ППР 60 (2), 1998 ППР 60 (1), 1998 ППР 59 (4), 1997 ППР 59 (3), 1997 ППР 59 (2), 1997 ППР 59 ( 1), ПТР 1997 58(4), ПТР 1996 58(3), ПТР 1996 58(1-2), ПТР 1996 57(3-4), ПТР 1995 57(2), ПТР 1995 57(1), ПТР 1995 56(4), ПТР 1994 56(3), 1994ПЗ 56(2), 1994ПЗ 56(1), 1994ПЗ 55(4), 1993ПЗ 55(3), 1993ПЗ 55(2), 1993ПЗ 55(1), 1993ПЗ 54(4), 1992ПЗ 54( 3), 1992MFR 54(2), 1992MFR 54(1), 1992MFR 53(4), 1991MFR 53(3), 1991MFR 53(2), 1991MFR 53(1), 1991MFR 52(4), 1990MFR 52(3) , 1990МФР 52(2), 1990МФР 52(1), 1990МФР 51(4), 1989МФР 51(3), 1989МФР 51(2), 1989МФР 51(1), 1989МФР 50(4), 1988МФР 50(3), 1988МФР 50(2), 1988МФР 50(1), 1988МФР 49(4), 1987МФР 49(3), 1987МФР 49(2), 1987МФР 49(1), 1987МФР 48(4), 1986МФР 48(3), 1986МФР 48( 2), 1986MFR 48(1), 1986MFR 47(4), 1985MFR 47(3), 1985MFR 47(2), 1985MFR 47(1), 1985MFR 46(4), 1984MFR 46(3), 1984MFR 46(2) , 1984 г. ПТР 46(1), 1984ПТР 45(10-12), 1983ПТР 45(7-9), 1983ПТР 45(4-6), 1983ПТР 45(3), 1983ПТР 45(2), 1983ПТР 45(1), 1983ПТР 44 (12), 1982МФР 44(11), 1982МФР 44(8), 1982МФР 44(6-7), 1982МФР 44(5), 1982МФР 44(4), 1982МФР 44(3), 1982МФР 44(2), 1982МФР 44 (1), 1982 ПТР 44 (9-10), 1982 ППР 43 (12), 1981 ППР 43 (11), 1981 ППР 43 (10), 1981 ППР 43 (9), 1981 ППР 43 (8), 1981 ППР 43 (7), 1981 ППР 43 (6), 1981MFR 43(5), 1981MFR 43(4), 1981MFR 43(3), 1981MFR 43(2), 1981MFR 43(1), 1981MFR 42(12), 1980MFR 42(11), 1980MFR 42(9) -10), 1980МФР 42(7-8), 1980МФР 42(6), 1980МФР 42(5), 1980МФР 42(3-4), 1980МФР 42(2), 1980МФР 42(1), 1980МФР 41(12), 1979МФР 41(11), 1979МФР 41(10), 1979МФР 41(9), 1979МФР 41(8), 1979МФР 41(7), 1979МФР 41(5-6), 1979МФР 41(4), 1979МФР 41(3), 1979МФР 41(1-2), 1979МФР 40(12), 1978МФР 40(11), 1978МФР 40(10), 1978МФР 40(9), 1978МФР 40(8), 1978МФР 40(7), 1978МФР 40(5-6) ), 1978 ПТР 40 (4), 1978 ПТР 40 (3), 1978 ПТР 40 (2), 1978 ПТР 40 (1), 1978 ПТР 39 (12) ПТР 39 (11) ПТР 39 (10) ПТР 39 (9) ПТР 39 (8 ) ПТР 39 (7) ПТР 39 (6) ПТР 39 (5)ПТ 39 (4)ПТ 39 (3)ПТ 39 (2)ПТ 39 (1)ПТ 38 (12)ПТ 38 (11)ПТ 38 (10)ПТ 38 (9)ПТ 38 (8)ПТ 38 (7)ПТ 38 (6)ПТ 38 (5)ПТ 38 (4)ПТ 38 (3)ПТ 38 (2)ПТ 38 (1)ПТ 37 (12)ПТ 37 (11)ПТ 37 (10)ПТ 37 (9)ПТ 37 (8)ПТ 37 (7)ПТ 37 (5-6)ПТ 37 (4)ПТ 37 (3)ПТ 37 (2)ПТ 37 (1)ПТ 36 (12)ПТ 36 (11) )ПТ 36 (10)ПТ 36 (9)ПТ 36 (8)ПТ 36 (7)ПТ 36 (6)ПТ 36 (5)ПТ 36 (4)ПТ 36 (3)ПТ 36 (2)ПТ 36 (1) )ПТ 35 (12)ПТ 35 (11)ПТ 35 (10)ПТ 35 (9)ПТ 35 (8)ПТ 35 (7)ПТ 35 (5-6)ПТ 35 (3-4)ПТ 35 (1- 2)ПТР 34 (11-12)ПТР 34 (9-10)ПТР 34 (7-8)ПТР 34 (5-6)ПТР 34 (3-4)ПТР 34 (1-2)ПТР 33 (11-12) )ПТ 33 (10)ПТ 33 (9)ПТ 33 (7-8)ПТ 33 (6)ПТ 33 (5)ПТ 33 (4)ПТ 33 (3)ПТ 33 (2)ПТ 33 (1)ПТ 32 (12)ПТР 32 (11)ПТР 32 (10)ПТР 32 (8-9)ПТР 32 (7)ПТР 32 (6)ПТР 32 (5)ПТР 32 (4)ПТР 32 (3)ПТР 32 (2)ПТР 32(1)ПТ 32 (1)ПТ 31 (12)ПТ 31 (11)ПТ 31 (10)ПТ 31 (8-9)ПТ 31 (7)ПТ 31 (6)ПТ 31 (5)ПТ 31 (4) )ПТ 31 (3)ПТ 31 (2)ПТ 31 (1)ПТ 30 (12)ПТ 30 (11)ПТ 30 (10)ПТ 30 (8-9)ПТ 30 (7)ПТ 30 (6)ПТ 30 ( 5) ПТР 30 (4) ПТР 30 (3) ПТР 30 (2) ПТР 30 (1) ПТР 29 (12) ПТР 29 (11) ПТР 29 (10) ПТР 29 (8-9) ПТР 29 (7) ПТР 29 (6)ПТР 29 (5)ПТР 29 (4)ПТ 29 (3)ПТ 29 (2)ПТ 29 (1)ПТ 28 (12)ПТ 28 (11)ПТ 28 (10)ПТ 28 (9)ПТ 28 (8)ПТ 28 (7)ПТ 28 (6)ПТ 28 (5)ПТ 28 (4)ПТ 28 (3)ПТ 28 (2)ПТ 28 (1)ПТ 27 (12)ПТ 27 (11)ПТ 27 (10)ПТ 27 (9)ПТ 27 (8)ПТ 27 (7)ПТ 27 (6)ПТ 27 (5)ПТ 27 (4)ПТ 27 (3)ПТ 27 (2)ПТ 27 (1)ПТ 26 (12)ПТ 26 (11a)ПТ 26 (11)ПТ 26 (10)ПТ 26 (9)ПТ 26 (8)ПТ 26 (7)ПТ 26 (6)ПТ 26 (5)ПТ 26 (4)ПТ 26 (3)ПТР 26 (2)ПТР 26 (1)ПТР 25 (12)ПТР 25 (11)ПТР 25 (10)ПТР 25 (9)ПТР 25 (8)ПТР 25 (7)ПТР 25 (6)ПТ 25 (5)ПТ 25 (4)ПТ 25 (3)ПТ 25 (2)ПТ 25 (1)ПТ 24 (12)ПТ 24 (11)ПТ 24 (10)ПТ 24 (9)ПТ 24 (8)ПТ 24 (7)ПТ 24 (6)ПТ 24 (5)ПТ 24 (4)ПТ 24 (3)ПТ 24 (2)ПТ 24 (1)ПТ 23 (12)ПТ 23 (11)ПТ 23 (10)ПТ 23 (9)ПТ 23 (8)ПТ 23 (7)ПТ 23 (6)ПТ 23 (5)ПТ 23 (4)ПТ 23 (3)ПТ 23 (2)ПТ 23 (1)ПТ 22 (12)ПТ 22 (11)ПТР 22 (10)ПТР 22 (9)ПТР 22 (8)ПТР 22 (7)ПТР 22 (6)ПТР 22 (5)ПТР 22 (4) ПТР 22 (3) ПТР 22 (2) ПТР 22 (1) ПТР 21 (12) ПТР 21 (11) ПТР 21 (10) ПТР 21 (9) ПТР 21 (8) ПТР 21 (7) ПТР 21 (6) ПТР 21 (5) ПТР 21 (4) ПТР 21 (3) ПТР 21 (2a) ПТР 21 (2) ПТР 21 (1) ПТР 20 (12) ПТР 20 (11a) ПТР 20 (11) ПТР 20 (10) ПТР 20 (9) ПТР 20 (8) ПТР 20 (7) ПТР 20 (6) ПТР 20 (5) ПТР 20 (4) ПТР 20 (3) ПТР 20 (2) ПТР 20 (1) ПТР 19 (12) ПТР 19 (11) ПТР 19 (10) ПТР 19 (9) ПТР 19 (8) ПТР 19 (7) ПТР 19 (6) ПТР 19 (5a) ПТР 19 (5) ПТР 19 (4a) ПТР 19 (4) ПТР 19 (3) ПТР 19 (2) ПТР 19 (1) ПТР 18 (12) ПТР 18 (11) ПТР 18 (10) ПТР 18 (9) ПТР 18 (8) ПТР 18 (7) ПТР 18 (6) ПТР 18 (5) ПТР 18 (4) ПТР 18 (3) ПТР 18 (2) ПТР 18 (1) ПТР 17 (12) ПТР 17 (11) ПТР 17 (10) ПТР 17 (9) ПТР 17 (8) ПТР 17 (7) ПТР 17 (6) ПТР 17 (5) ПТР 17 (4) ПТР 17 (3) ПТР 17 (2) ПТР 17 (1) ПТР 16 (12) ПТР 16 (11) ПТР 16 (10) ПТР 16 (9) ПТР 16 (8) ПТР 16 (7) ПТР 16 (6) ПТР 16 (5) ПТР 16 (4) ПТР 16 (3) ПТР 16 (2) ПТР 16 (1) ПТР 15 (12) ПТР 15 (11) ПТР 15 (10) ПТР 15 (9) ПТР 15 (8) ПТР 15 (7) ПТР 15 (6) ПТР 15 (5) ПТР 15 (4) ПТР 15 (3) ПТР 15 (2) ПТР 15 (1) ПТР 14 (12) ПТР 14 (12a) ПТР 14 (11) ПТР 14 (10) ПТР 14 (9) ПТР 14 ( 8)ПТР 14 (7)ПТР 14 (6)ПТР 14 (5)ПТР 14 (4)ПТР 14 (3)ПТР 14 (2)ПТР 14 (1)ПТР 13 (12)ПТР 13 (11a)ПТР 13 ( 11)ПТР 13 (10)ПТР 13 (9)ПТР 13 (8)ПТР 13 (7)ПТР 13 (6)ПТР 13 (5)ПТР 13 (4)ПТР 13 (3)ПТР 13 (2)ПТР 13 ( 1)ПТР 12 (12)ПТР 12 (11a)ПТР 12 (11)ПТР 12 (10)ПТР 12 (9)ПТР 12 (8)ПТР 12 (7)ПТР 12 (6)ПТР 12 (5)ПТР 12 ( 4)ПТР 12 (3)ПТР 12 (2)ПТР 12 (1)ПТР 11 (12)ПТР 11 (11)ПТР 11 (10)ПТР 11 (9)ПТР 11 (8)ПТР 11 (7)ПТР 11 ( 6)ПТР 11 (5)ПТР 11 (4)ПТР 11 (3)ПТР 11 (2)ПТР 11 (1)ПТР 10 (12)ПТР 10 (11)ПТР 10 (10)ПТР 10 (9)ПТР 10 ( 8)ПТР 10 (7)ПТР 10 (6)ПТР 10 (5)ПТР 10 (4)ПТР 10 (3)ПТР 10 (2)ПТР 10 (1)ПТР 9 (12)ПТР 9 (11)ПТР 9 ( 10)ПТ 9 (9)ПТ 9 (8)ПТ 9 (7)ПТ 9 (6)ПТ 9 (5)ПТ 9 (4)ПТ 9 (3)ПТ 9 (2)ПТ 9 (1)ПТ 8 ( 12)ПТ 8 (11a)ПТ 8 (11)ПТ 8 (10)ПТ 8 (9)ПТ 8 (8)ПТ 8 (7)ПТ 8 (6)ПТ 8 (5)ПТ 8 (4)ПТ 8 ( 3) ПТР 8 (2) ПТР 8 (1)
ПТР 46(1), 1984ПТР 45(10-12), 1983ПТР 45(7-9), 1983ПТР 45(4-6), 1983ПТР 45(3), 1983ПТР 45(2), 1983ПТР 45(1), 1983ПТР 44 (12), 1982МФР 44(11), 1982МФР 44(8), 1982МФР 44(6-7), 1982МФР 44(5), 1982МФР 44(4), 1982МФР 44(3), 1982МФР 44(2), 1982МФР 44 (1), 1982 ПТР 44 (9-10), 1982 ППР 43 (12), 1981 ППР 43 (11), 1981 ППР 43 (10), 1981 ППР 43 (9), 1981 ППР 43 (8), 1981 ППР 43 (7), 1981 ППР 43 (6), 1981MFR 43(5), 1981MFR 43(4), 1981MFR 43(3), 1981MFR 43(2), 1981MFR 43(1), 1981MFR 42(12), 1980MFR 42(11), 1980MFR 42(9) -10), 1980МФР 42(7-8), 1980МФР 42(6), 1980МФР 42(5), 1980МФР 42(3-4), 1980МФР 42(2), 1980МФР 42(1), 1980МФР 41(12), 1979МФР 41(11), 1979МФР 41(10), 1979МФР 41(9), 1979МФР 41(8), 1979МФР 41(7), 1979МФР 41(5-6), 1979МФР 41(4), 1979МФР 41(3), 1979МФР 41(1-2), 1979МФР 40(12), 1978МФР 40(11), 1978МФР 40(10), 1978МФР 40(9), 1978МФР 40(8), 1978МФР 40(7), 1978МФР 40(5-6) ), 1978 ПТР 40 (4), 1978 ПТР 40 (3), 1978 ПТР 40 (2), 1978 ПТР 40 (1), 1978 ПТР 39 (12) ПТР 39 (11) ПТР 39 (10) ПТР 39 (9) ПТР 39 (8 ) ПТР 39 (7) ПТР 39 (6) ПТР 39 (5)ПТ 39 (4)ПТ 39 (3)ПТ 39 (2)ПТ 39 (1)ПТ 38 (12)ПТ 38 (11)ПТ 38 (10)ПТ 38 (9)ПТ 38 (8)ПТ 38 (7)ПТ 38 (6)ПТ 38 (5)ПТ 38 (4)ПТ 38 (3)ПТ 38 (2)ПТ 38 (1)ПТ 37 (12)ПТ 37 (11)ПТ 37 (10)ПТ 37 (9)ПТ 37 (8)ПТ 37 (7)ПТ 37 (5-6)ПТ 37 (4)ПТ 37 (3)ПТ 37 (2)ПТ 37 (1)ПТ 36 (12)ПТ 36 (11) )ПТ 36 (10)ПТ 36 (9)ПТ 36 (8)ПТ 36 (7)ПТ 36 (6)ПТ 36 (5)ПТ 36 (4)ПТ 36 (3)ПТ 36 (2)ПТ 36 (1) )ПТ 35 (12)ПТ 35 (11)ПТ 35 (10)ПТ 35 (9)ПТ 35 (8)ПТ 35 (7)ПТ 35 (5-6)ПТ 35 (3-4)ПТ 35 (1- 2)ПТР 34 (11-12)ПТР 34 (9-10)ПТР 34 (7-8)ПТР 34 (5-6)ПТР 34 (3-4)ПТР 34 (1-2)ПТР 33 (11-12) )ПТ 33 (10)ПТ 33 (9)ПТ 33 (7-8)ПТ 33 (6)ПТ 33 (5)ПТ 33 (4)ПТ 33 (3)ПТ 33 (2)ПТ 33 (1)ПТ 32 (12)ПТР 32 (11)ПТР 32 (10)ПТР 32 (8-9)ПТР 32 (7)ПТР 32 (6)ПТР 32 (5)ПТР 32 (4)ПТР 32 (3)ПТР 32 (2)ПТР 32(1)ПТ 32 (1)ПТ 31 (12)ПТ 31 (11)ПТ 31 (10)ПТ 31 (8-9)ПТ 31 (7)ПТ 31 (6)ПТ 31 (5)ПТ 31 (4) )ПТ 31 (3)ПТ 31 (2)ПТ 31 (1)ПТ 30 (12)ПТ 30 (11)ПТ 30 (10)ПТ 30 (8-9)ПТ 30 (7)ПТ 30 (6)ПТ 30 ( 5) ПТР 30 (4) ПТР 30 (3) ПТР 30 (2) ПТР 30 (1) ПТР 29 (12) ПТР 29 (11) ПТР 29 (10) ПТР 29 (8-9) ПТР 29 (7) ПТР 29 (6)ПТР 29 (5)ПТР 29 (4)ПТ 29 (3)ПТ 29 (2)ПТ 29 (1)ПТ 28 (12)ПТ 28 (11)ПТ 28 (10)ПТ 28 (9)ПТ 28 (8)ПТ 28 (7)ПТ 28 (6)ПТ 28 (5)ПТ 28 (4)ПТ 28 (3)ПТ 28 (2)ПТ 28 (1)ПТ 27 (12)ПТ 27 (11)ПТ 27 (10)ПТ 27 (9)ПТ 27 (8)ПТ 27 (7)ПТ 27 (6)ПТ 27 (5)ПТ 27 (4)ПТ 27 (3)ПТ 27 (2)ПТ 27 (1)ПТ 26 (12)ПТ 26 (11a)ПТ 26 (11)ПТ 26 (10)ПТ 26 (9)ПТ 26 (8)ПТ 26 (7)ПТ 26 (6)ПТ 26 (5)ПТ 26 (4)ПТ 26 (3)ПТР 26 (2)ПТР 26 (1)ПТР 25 (12)ПТР 25 (11)ПТР 25 (10)ПТР 25 (9)ПТР 25 (8)ПТР 25 (7)ПТР 25 (6)ПТ 25 (5)ПТ 25 (4)ПТ 25 (3)ПТ 25 (2)ПТ 25 (1)ПТ 24 (12)ПТ 24 (11)ПТ 24 (10)ПТ 24 (9)ПТ 24 (8)ПТ 24 (7)ПТ 24 (6)ПТ 24 (5)ПТ 24 (4)ПТ 24 (3)ПТ 24 (2)ПТ 24 (1)ПТ 23 (12)ПТ 23 (11)ПТ 23 (10)ПТ 23 (9)ПТ 23 (8)ПТ 23 (7)ПТ 23 (6)ПТ 23 (5)ПТ 23 (4)ПТ 23 (3)ПТ 23 (2)ПТ 23 (1)ПТ 22 (12)ПТ 22 (11)ПТР 22 (10)ПТР 22 (9)ПТР 22 (8)ПТР 22 (7)ПТР 22 (6)ПТР 22 (5)ПТР 22 (4) ПТР 22 (3) ПТР 22 (2) ПТР 22 (1) ПТР 21 (12) ПТР 21 (11) ПТР 21 (10) ПТР 21 (9) ПТР 21 (8) ПТР 21 (7) ПТР 21 (6) ПТР 21 (5) ПТР 21 (4) ПТР 21 (3) ПТР 21 (2a) ПТР 21 (2) ПТР 21 (1) ПТР 20 (12) ПТР 20 (11a) ПТР 20 (11) ПТР 20 (10) ПТР 20 (9) ПТР 20 (8) ПТР 20 (7) ПТР 20 (6) ПТР 20 (5) ПТР 20 (4) ПТР 20 (3) ПТР 20 (2) ПТР 20 (1) ПТР 19 (12) ПТР 19 (11) ПТР 19 (10) ПТР 19 (9) ПТР 19 (8) ПТР 19 (7) ПТР 19 (6) ПТР 19 (5a) ПТР 19 (5) ПТР 19 (4a) ПТР 19 (4) ПТР 19 (3) ПТР 19 (2) ПТР 19 (1) ПТР 18 (12) ПТР 18 (11) ПТР 18 (10) ПТР 18 (9) ПТР 18 (8) ПТР 18 (7) ПТР 18 (6) ПТР 18 (5) ПТР 18 (4) ПТР 18 (3) ПТР 18 (2) ПТР 18 (1) ПТР 17 (12) ПТР 17 (11) ПТР 17 (10) ПТР 17 (9) ПТР 17 (8) ПТР 17 (7) ПТР 17 (6) ПТР 17 (5) ПТР 17 (4) ПТР 17 (3) ПТР 17 (2) ПТР 17 (1) ПТР 16 (12) ПТР 16 (11) ПТР 16 (10) ПТР 16 (9) ПТР 16 (8) ПТР 16 (7) ПТР 16 (6) ПТР 16 (5) ПТР 16 (4) ПТР 16 (3) ПТР 16 (2) ПТР 16 (1) ПТР 15 (12) ПТР 15 (11) ПТР 15 (10) ПТР 15 (9) ПТР 15 (8) ПТР 15 (7) ПТР 15 (6) ПТР 15 (5) ПТР 15 (4) ПТР 15 (3) ПТР 15 (2) ПТР 15 (1) ПТР 14 (12) ПТР 14 (12a) ПТР 14 (11) ПТР 14 (10) ПТР 14 (9) ПТР 14 ( 8)ПТР 14 (7)ПТР 14 (6)ПТР 14 (5)ПТР 14 (4)ПТР 14 (3)ПТР 14 (2)ПТР 14 (1)ПТР 13 (12)ПТР 13 (11a)ПТР 13 ( 11)ПТР 13 (10)ПТР 13 (9)ПТР 13 (8)ПТР 13 (7)ПТР 13 (6)ПТР 13 (5)ПТР 13 (4)ПТР 13 (3)ПТР 13 (2)ПТР 13 ( 1)ПТР 12 (12)ПТР 12 (11a)ПТР 12 (11)ПТР 12 (10)ПТР 12 (9)ПТР 12 (8)ПТР 12 (7)ПТР 12 (6)ПТР 12 (5)ПТР 12 ( 4)ПТР 12 (3)ПТР 12 (2)ПТР 12 (1)ПТР 11 (12)ПТР 11 (11)ПТР 11 (10)ПТР 11 (9)ПТР 11 (8)ПТР 11 (7)ПТР 11 ( 6)ПТР 11 (5)ПТР 11 (4)ПТР 11 (3)ПТР 11 (2)ПТР 11 (1)ПТР 10 (12)ПТР 10 (11)ПТР 10 (10)ПТР 10 (9)ПТР 10 ( 8)ПТР 10 (7)ПТР 10 (6)ПТР 10 (5)ПТР 10 (4)ПТР 10 (3)ПТР 10 (2)ПТР 10 (1)ПТР 9 (12)ПТР 9 (11)ПТР 9 ( 10)ПТ 9 (9)ПТ 9 (8)ПТ 9 (7)ПТ 9 (6)ПТ 9 (5)ПТ 9 (4)ПТ 9 (3)ПТ 9 (2)ПТ 9 (1)ПТ 8 ( 12)ПТ 8 (11a)ПТ 8 (11)ПТ 8 (10)ПТ 8 (9)ПТ 8 (8)ПТ 8 (7)ПТ 8 (6)ПТ 8 (5)ПТ 8 (4)ПТ 8 ( 3) ПТР 8 (2) ПТР 8 (1)
Доисторические ящерицы, превратившиеся в птиц – Исторические статьи и иллюстрацииИсторические статьи и иллюстрации
Эта отредактированная статья о предыстории первоначально появилась в выпуске Look and Learn № 164, опубликованном 6 марта 1965 года.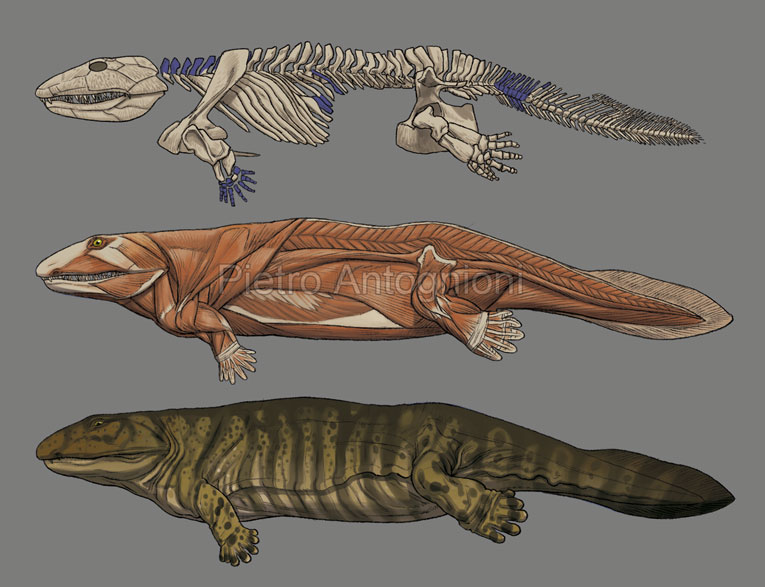
Эпоха ужасных ящериц
В каменноугольный период, начавшийся около 280 миллионов лет назад, землю начала покрывать богатая растительность в виде папоротников и пальмоподобных деревьев. Климат во всем мире был жарким и влажным, а по берегам континентов простирались обширные болота.
Эти условия сохранялись и в пермском периоде, который иногда называют верхнекаменноугольным периодом. Теплый климат и обилие растительности — вот все, что было необходимо для развития наземных животных (рисунок вверху).
Этими первыми наземными животными были амфибии, которые делили свое время между сушей и водой.
Один из них, стегоцефал, был бы нелишним в современном мире. Это было похожее на тритона существо около трех футов длиной, которое ползало по мягкой земле или плескалось в неглубоких заводях.
Его тело было покрыто доспехами из прочной чешуи, а плоская голова была вооружена острыми зубами. Ибо стегоцефал был свирепым каннибалом, который сражался и поедал себе подобных.
Другие животные того далекого периода были совсем не похожи ни на одно из известных нам существ. Одним из самых крупных и странных был эдафозавр.
Эдафозавр был гигантом своего времени, иногда достигая в длину десяти футов и более. Его наиболее характерной чертой был огромный гребень или оборка из шипов на теле, идущих вдоль его спины.
Несмотря на устрашающий вид, эдафозавры были робкими и безобидными существами. Из-за своих крошечных зубов они не ели ничего, кроме растений.
И стегоцефалы, и эдафозавры вылупились из яиц, отложенных самками в болотистых прудах, и провели раннюю часть своей жизни в воде. Как и все земноводные, они произошли от рыб девонского периода, которые за миллионы лет приспособились к жизни на суше.
Мир стегоцефалов и эдафозавров тоже увидел первое насекомое. Это была меганевра, гигантская стрекоза с размахом крыльев в два фута.
Как только животные появились на земле, они развивались с поразительной скоростью не только в количестве, но и в размерах. Ко времени триасового периода (нижняя иллюстрация), начавшегося 190 миллионов лет назад, стали появляться самые фантастические животные.
Ко времени триасового периода (нижняя иллюстрация), начавшегося 190 миллионов лет назад, стали появляться самые фантастические животные.
Это были динозавры или большие рептилии. Название «динозавр» происходит от двух греческих слов: deinos, что означает «ужасный», и sauros, что означает «ящерица». Именно такими были динозавры, огромные ящерицы.
Но динозавры были не так страшны, как можно предположить по их названию и внешнему виду. Они были очень глупы, и большинство из них были робкими и безобидными.Все, чего они хотели, это чтобы их оставили в покое и съели тонны зелени, которые составляли их ежедневную еду.
Еще одним фантастическим существом эпохи динозавров был струтиомим. Это животное было намного меньше других монстров. У него были длинные задние ноги, очень короткие передние и длинный хвост. Он бегал и прыгал почти так же, как современные кенгуру.
Именно в эпоху динозавров, которая длилась многие миллионы лет и включала в себя несколько геологических периодов, появились первые птицы. Одним из них был рамфоринх, но если бы вы увидели его сегодня, вы бы вряд ли узнали в нем предка всех изящных и красочных птиц, летающих сейчас в мире.
Одним из них был рамфоринх, но если бы вы увидели его сегодня, вы бы вряд ли узнали в нем предка всех изящных и красочных птиц, летающих сейчас в мире.
Рамфоринх был просто динозавром-ящерицей, который за миллионы лет научился летать. Скорее всего, он произошел от небольшого вида динозавров, похожих на кенгуру.
Постоянно убегая от плотоядных рептилий и бья воздух своими короткими передними конечностями, они развили на своих передних конечностях бахрому из чешуи.Это помогло снять вес их тел с земли.
В конце концов они стали жить на деревьях в целях безопасности, а затем научились планировать с ветки на ветку. В течение миллионов лет они перешли от скольжения по воздуху к полету в нем.
Возможно, вам трудно представить, что современные птицы произошли от неуклюжих летающих рептилий 180 миллионов лет назад. Тем не менее, в наших птицах до сих пор сохранились следы их предков-рептилий.
Перья птиц лежат плоско на их телах, как перекрывающиеся чешуйки, а их клювы мало чем отличаются от беззубого рта ящерицы, хотя у рамфоринха в клюве был ряд огромных зубов. Но одним из самых поразительных доказательств того, что сегодняшние птицы произошли от рептилий, является чешуя на их ногах и пальцах ног.
Но одним из самых поразительных доказательств того, что сегодняшние птицы произошли от рептилий, является чешуя на их ногах и пальцах ног.
Любовь во времена хасмозавров: винтажное искусство динозавров: путешествия с динозаврами
После долгого перерыва в сериале я вернулся со своим первым постом о винтажном искусстве динозавров почти за два года. Вдохновленный мультяшным стилем поста Марка о динозаврах! Книга-головоломка «Найди отличия» , я отсканировал недавнее собственное приобретение, Путешествия с динозаврами .Сама книга содержит скудную информацию об издании, но поиск в Интернете приводит меня к дате публикации 1997 года, что делает ее не совсем винтажной, но, безусловно, по духу, так что поехали. Книга была написана детским писателем Вецио Мельгари и проиллюстрирована Джованни Джаннини и Виолайн Хюльне. К сожалению, нет указаний на то, как Джаннини и Халне работали вместе над иллюстрациями, которые сильно напоминают работы Ричарда Скарри.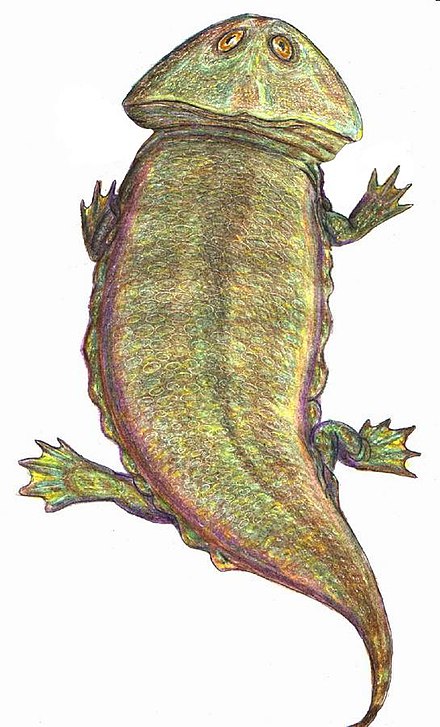 История Мельгари рассказывает о группе молодых животных — в основном собачьего происхождения, с одной или двумя кошками в смеси — которых профессор Альфред С.Вольфсбейн, «специалист в нескольких науках и волшебник компьютерного мира». Это были девяностые! Конечно, для наших целей нас больше заботит то, как представлены динозавры, чем сама история.
История Мельгари рассказывает о группе молодых животных — в основном собачьего происхождения, с одной или двумя кошками в смеси — которых профессор Альфред С.Вольфсбейн, «специалист в нескольких науках и волшебник компьютерного мира». Это были девяностые! Конечно, для наших целей нас больше заботит то, как представлены динозавры, чем сама история.
Первое путешествие — посетить «плавающих гигантов», также известных как зауроподы. Хотя Апатозавр изображен в своей классической привычке середины века стоять наполовину погруженным в воду, он упоминается по правильному имени, и в тексте даже упоминается устаревание «бронтозавра».Разворот является хорошим введением в основную цветовую эстетику книги и странное сочетание устаревших и современных идей, а также в его замечательную приверженность включению в смесь более мелкой фауны, как правило, хорошо помеченной, как, например, рыба, плавающая вокруг ног зауроподов здесь. .
Зауроподы из «Путешествий с динозаврами»: апатозавр, диплодок и брахиозавр.
Далее следуют различные гадрозавры, известные своими «странными головами», так как различные головные уборы гадрозавров всегда были популярным предметом в книжках с картинками.Самым интересным здесь является «Анатозавр», который к концу девяностых был официально свернут в Эдмонтозавра примерно на десятилетие. Он стоит в знакомой рыцарской позе на двух ногах (и неприятным образом ползет по древовидному папоротнику). Пейзаж снова сочетает в себе старое и новое, повторяя старый образ бесплодных, населенных вулканами ландшафтов, но также намекая на расцвет покрытосеменных в меловой период.
Странные головы: эдмонтозавр FKA, анатозавр, коритозавр и паразауролоф
Тиреофораны затем получают свое внимание. Стегозавр изображен спереди и в центре, как и ожидалось, стоящим на двух ногах и жующим листву по старой традиции. Появляется Велоцираптор , которому угрожает Анкилозавр , и он уводит его обратно в Азию (и, предположительно, в свое время, пару миллионов лет назад). По крайней мере, его можно узнать как животное, похожее на велоцираптора, а не как обычного хищника после JP. Kentrosaurus лишен зубов, чего никогда не было; хотя его первоначальное описание включало только один зуб и только фрагменты зубов или появляющиеся зубы после этого, я не уверен, что беззубый Kentrosaurus когда-либо был большой теорией — с радостью поправлю, если это так.
По крайней мере, его можно узнать как животное, похожее на велоцираптора, а не как обычного хищника после JP. Kentrosaurus лишен зубов, чего никогда не было; хотя его первоначальное описание включало только один зуб и только фрагменты зубов или появляющиеся зубы после этого, я не уверен, что беззубый Kentrosaurus когда-либо был большой теорией — с радостью поправлю, если это так.
Ankylosaurus, Kentrosaurus и Stegosaurus представляют Thyreophora
Соответственно, Triceratops занимает центральное место среди цератопсов, а его товарищи-старые резервные роговые лица Monoclonius и Styracosaurus подыгрывают. Посмотрите на довольно хорошо прорисованную морфологию головы детеныша трицератопса , спрятавшегося в правом нижнем углу.
Styracosaurus, Triceratops и Monoclonius, естественно
Плотоядные животные, что вполне уместно, представлены этими титанами-близнецами ужаса, Tyrannosaurus и… Игуанодон . Ага, Игуанодон , рвущийся на Мозазавр . Профессор Вольфсбейн называет его «одним из крупнейших хищников… среди наземных динозавров», уступив только тираннозавру . Я не знаю, откуда это взялось, кроме знаменитой анахроничной битвы мегалозавра против игуанодона Луи Фигье, в которой можно с уверенностью предположить, что игуанодон просто борется за свою жизнь, а не пытается выжить. его противник.Если кто-нибудь знает о каких-либо других изображениях хищного игуанодона , пожалуйста, дайте мне знать, что я пропустил. Это почти как если бы Мельгари смешал мегалозавра и игуанодона в одного ваззлезавра со стереотипной крадущейся походкой и хищным характером первого и шипом большого пальца второго. Мне нравится ликование на лице Рекси, как будто он просто в восторге от того, что Игуанодон сменил команду и хочет участвовать в действии.
Ага, Игуанодон , рвущийся на Мозазавр . Профессор Вольфсбейн называет его «одним из крупнейших хищников… среди наземных динозавров», уступив только тираннозавру . Я не знаю, откуда это взялось, кроме знаменитой анахроничной битвы мегалозавра против игуанодона Луи Фигье, в которой можно с уверенностью предположить, что игуанодон просто борется за свою жизнь, а не пытается выжить. его противник.Если кто-нибудь знает о каких-либо других изображениях хищного игуанодона , пожалуйста, дайте мне знать, что я пропустил. Это почти как если бы Мельгари смешал мегалозавра и игуанодона в одного ваззлезавра со стереотипной крадущейся походкой и хищным характером первого и шипом большого пальца второго. Мне нравится ликование на лице Рекси, как будто он просто в восторге от того, что Игуанодон сменил команду и хочет участвовать в действии.
Ужасные хищники, тираннозавры и игуанодоны.
Птерозавры в книге были отлиты по старому образцу кожистых демонов, при этом мало внимания уделялось масштабу или временной точности. И эй, так как мы находимся на берегу моря, Ichthyornis готовится к посадке на воду, и есть типичный Hesperornis . Смотри, ныряй.
И эй, так как мы находимся на берегу моря, Ichthyornis готовится к посадке на воду, и есть типичный Hesperornis . Смотри, ныряй.
«Эй, большие летучие мыши!»
Археоптерикс в классическом костюме блестящего раптора относится к развороту, посвященному различным птицам (и паре тапиров). Предположительно сейчас кайнозой, поэтому мы скорее зеленые, свежие и привлекательные, чем вулканические и бесплодные. Palaelodus стоит на заднем плане, розовый и с ошибками. Сын профессора Вольфсбейна, Уолт, пропал без вести, и некоторые из его предполагаемых друзей решили вообразить, что его преследует хищный Tyrannosaurus , которому угрожает Mastodonosaurus , или, что лучше всего, кормит Brachiosaurus до Mosasaurus . Бедный профессор Волчий яд, могу только представить его реакцию.
Птицы, тапиры и темные фантазии
Последний разворот, которым я поделюсь, — это «рекордсмен», в котором сомнительные факты и заезженные старые слухи передаются желающим и впечатлительным юным читателям.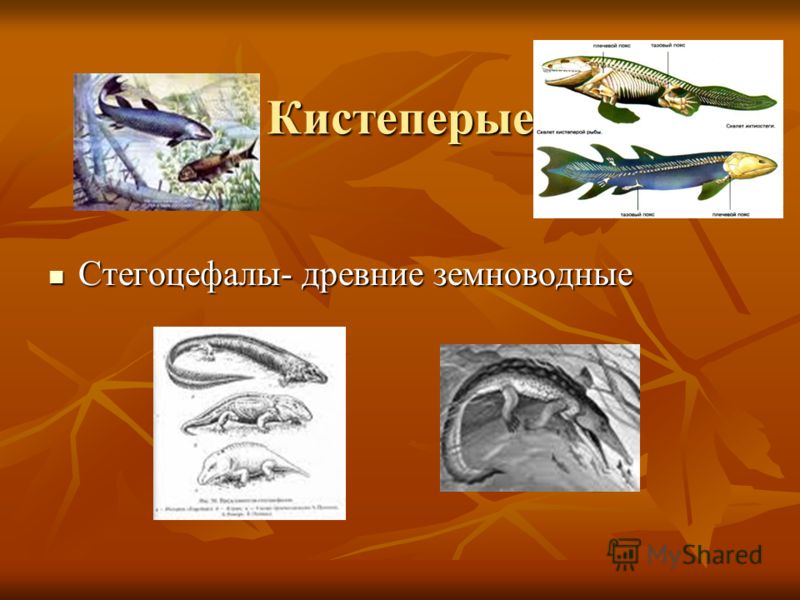 Здесь мы узнаем, что Аллозавр был «самым свирепым и прожорливым» динозавром, потому что наука, и это, безусловно, кажется взволнованным этими ловкачами Джемми. Ouranosaurus имеет самый длинный гребень, Tanystropheus причислен к динозаврам, потому что почему бы и нет, и ей-богу: Composognathus размером с курицу.
Здесь мы узнаем, что Аллозавр был «самым свирепым и прожорливым» динозавром, потому что наука, и это, безусловно, кажется взволнованным этими ловкачами Джемми. Ouranosaurus имеет самый длинный гребень, Tanystropheus причислен к динозаврам, потому что почему бы и нет, и ей-богу: Composognathus размером с курицу.
Наука!
Итак, резюмируем: причудливые иллюстрации, сочетающие старые взгляды, несколько современные знания и головокружительные неточности.Если бы Мельгари пошел с более легким приключенческим повествованием, которое не претендует на звание энциклопедии, поэтическая вольность была бы понятна, но вместо этого у нас есть книга, которая в основном запоминается своими причудливыми не фактами.
.
