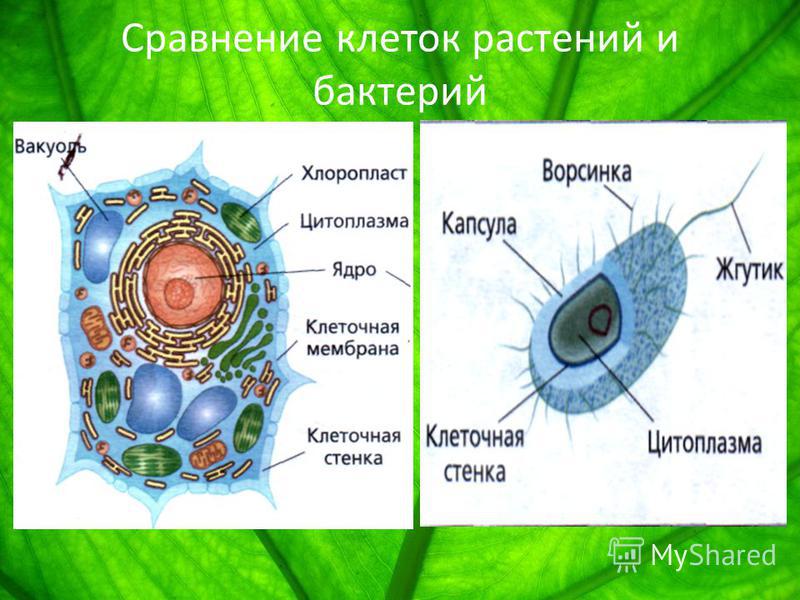
Сходство и различия в строении клеток растений, животных и грибов | Биология
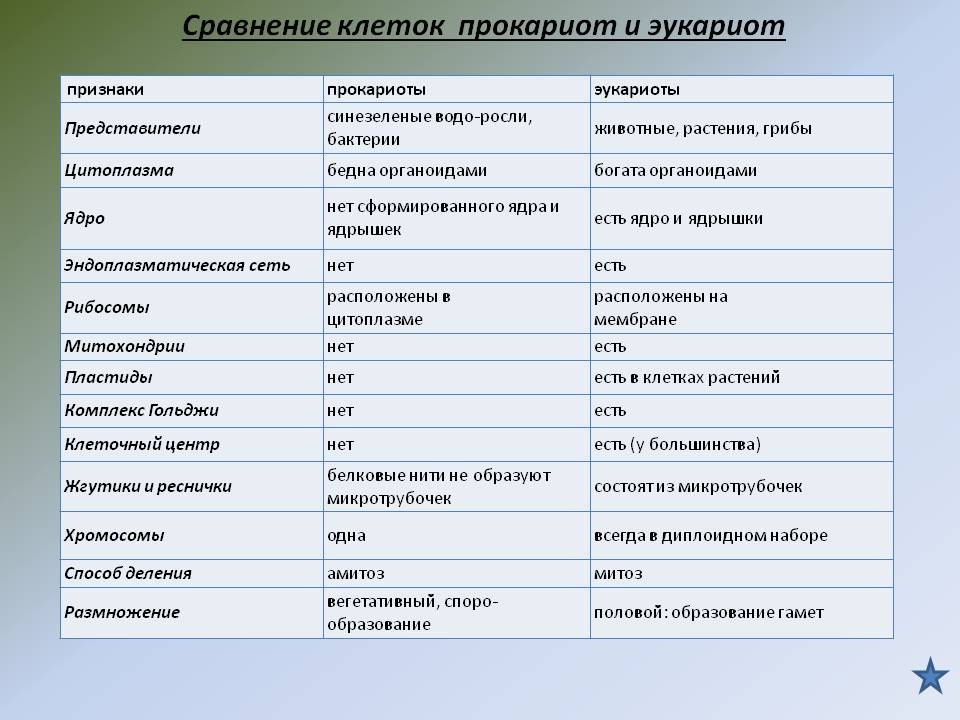
Сходство в строении клеток эукариот. Сейчас нельзя с полной уверенностью сказать, когда и как возникла на Земле жизнь. Мы также точно не знаем, как питались первые живые существа на Земле: ав-тотрофно или гетеротрофно. Но в настоящее время на нашей планете мирно сосуществуют представители нескольких царств живых существ. Несмотря на большое различие в строении и образе жизни, очевидно, что между ними сходств больше, чем различий, и все они, вероятно, имеют общих предков, живших в далекой архейской эре. О наличии общих «дедушек» и «бабушек» свидетельствует целый ряд общих признаков у клеток эукариот: простейших, растений, грибов и животных. К этим признакам можно отнести:
— общий план строения клетки: наличие топлазмы, ядра, органоидов;
— принципиальное сходство процессов в клетке;
— кодирование наследственной информ иновых кислот;
— единство химического состава клеток;
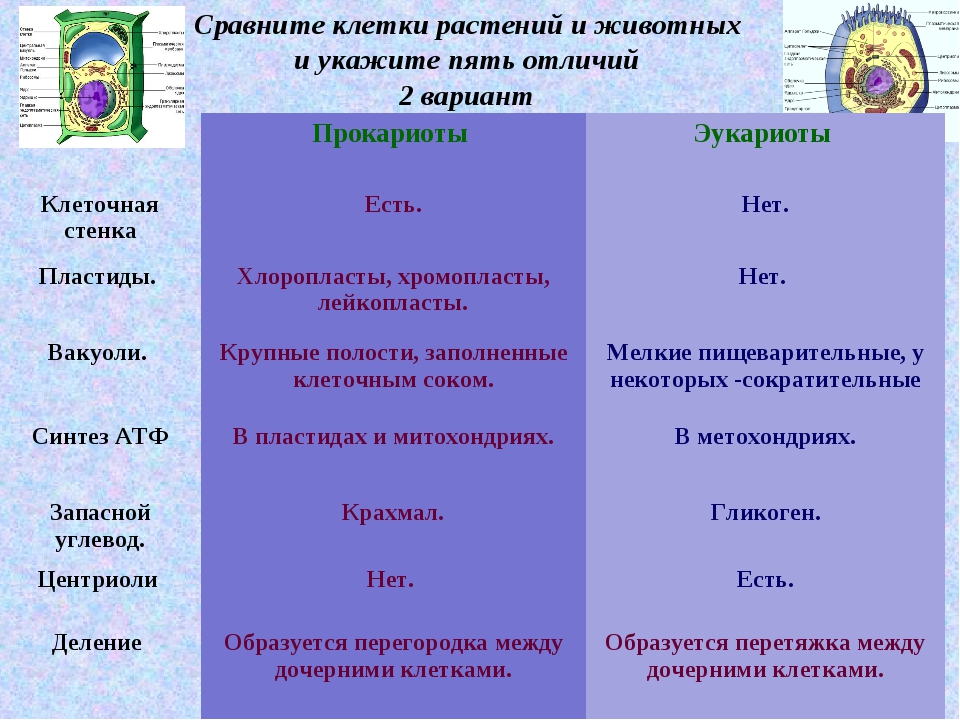
— сходные процессы деления клеток. Различия в строении клеток растений
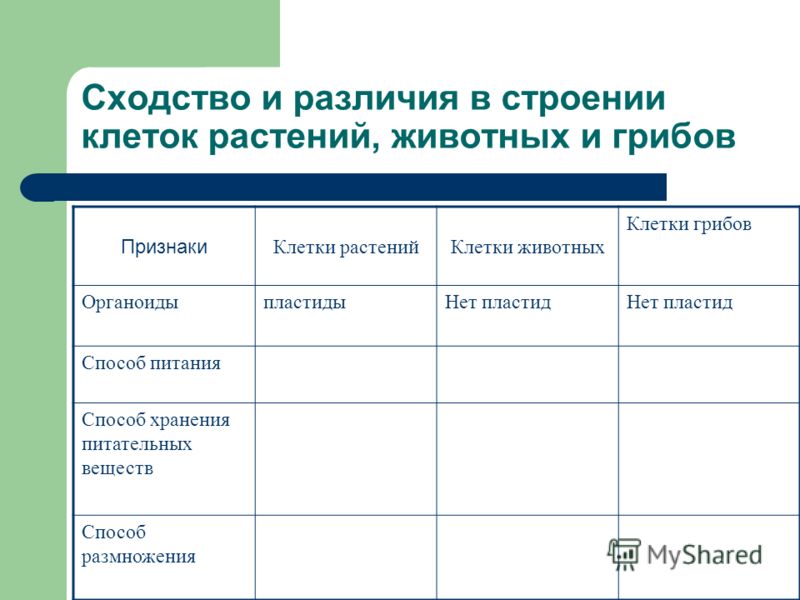
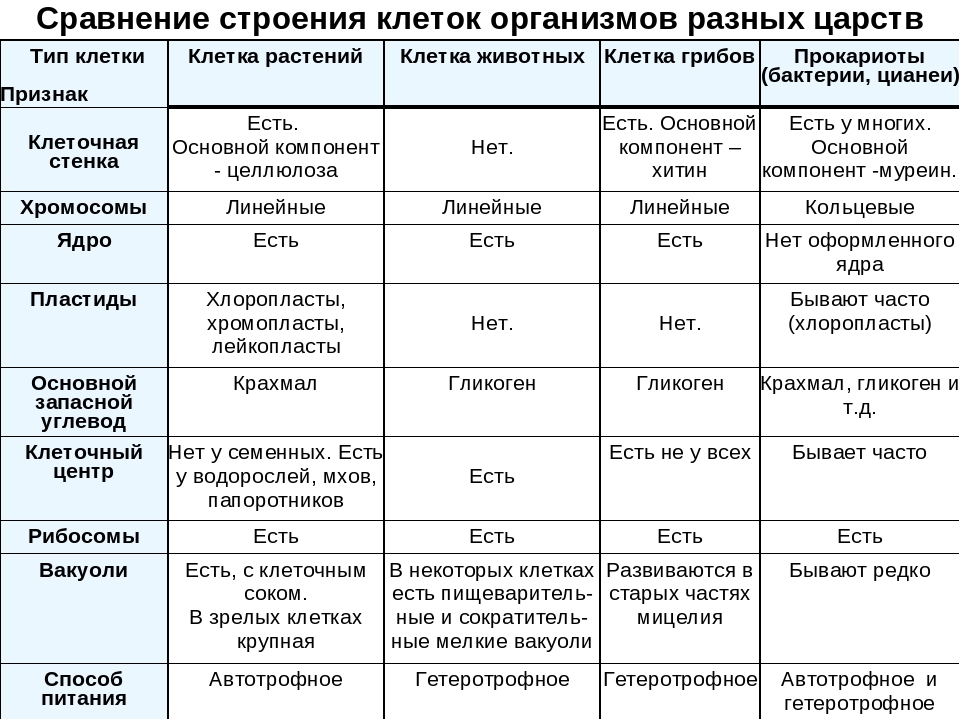
Различия в строении клеток растений
эволюции, в связи с неодинаковыми услоЕ ток представителей различных царств жив] жество отличий. Сравним строение и жизв тений и животных (табл. 4).
Главное отличие между клетками этих двух царств заключается в способе их питания. Клетки растений, содержащие хлоропласты, являются автотрофами, т. е. сами синтезируют необходимые для жизнедеятельности органические вещества за счет энергии света в процессе фотосинтеза. Клетки животных — гетеротрофы, т. е. источником углерода для синтеза собственных органических веществ для них являются органические вещества, поступающие с пищей. Эти же пищевые вещества, например углеводы, служат для животных источником энергии. Есть и исключения, такие как зеленые жгутиконосцы, которые на свету способны к фотосинтезу, а в темноте питаются готовыми органическими веществами. Для обеспечения фотосинтеза в клет-ках растений содержатся пластиды, несущие хлорофилл и другие пигменты.
Так как растительная клетка имеет клеточную стенку, защищающую ее содержимое и обеспечивающую постоянную ее форму, то при делении между дочерними клетками образуется перегородка, а животная клетка, не имеющая такой стенки, делится с образованием перетяжки.
Особенности клеток грибов. Еще совсем недавно грибы относили к растениям, однако сейчас эта весьма своеобразная и большая по числу видов группа живых существ выделена в отдельное царство. Грибы, так же как и животные, — гетеротрофы, питаются готовыми органическими соединениями. Они могут быть сапротрофами, т. е. питаться органикой мертвых существ, паразитами, т. е. питаться живой органикой, или симбионтами высших растений, находясь с ними во взаимовыгодной связи. Пластид и хлорофилла клетки грибов не содержат. Среди грибов существуют и «хищники», образующие в почве клейкие петли, в которых запутываются мелкие круглые черви. После этого клетки грибницы проникают в пойманного червя, разрастаются в нем и высасывают его содержимое. У клеток грибов, как и у растений, есть клеточная стенка поверх плазматической мембраны. Часто в состав клеточной стенки у грибов входит хитин — вещество, образующее наружные покровы у членистоногих. Запасным питательным веществом в клетках грибов является углевод гликоген, как у животных, а не крахмал, как у растений. Тело гриба образовано нитевидными структурами в один ряд клеток — гифами. У некоторых грибов перегородки между клетками утрачиваются, и возникает грибница, состоящая из одной гигантской многоядерной клетки. Грибы не способны к активному движению, зато они могут расти неограниченно — это признаки, которые объединяют грибы с растениями.
Тело гриба образовано нитевидными структурами в один ряд клеток — гифами. У некоторых грибов перегородки между клетками утрачиваются, и возникает грибница, состоящая из одной гигантской многоядерной клетки. Грибы не способны к активному движению, зато они могут расти неограниченно — это признаки, которые объединяют грибы с растениями.
Различие и сходство растений и животных
Различие между растениями и животными является не качественным, а количественным. То есть оно выражается в том, что преобладают определенные особенности строения тех или иных организмов. Нельзя вести речь об исключительной их свойственности растениям или животным.
Строение тела
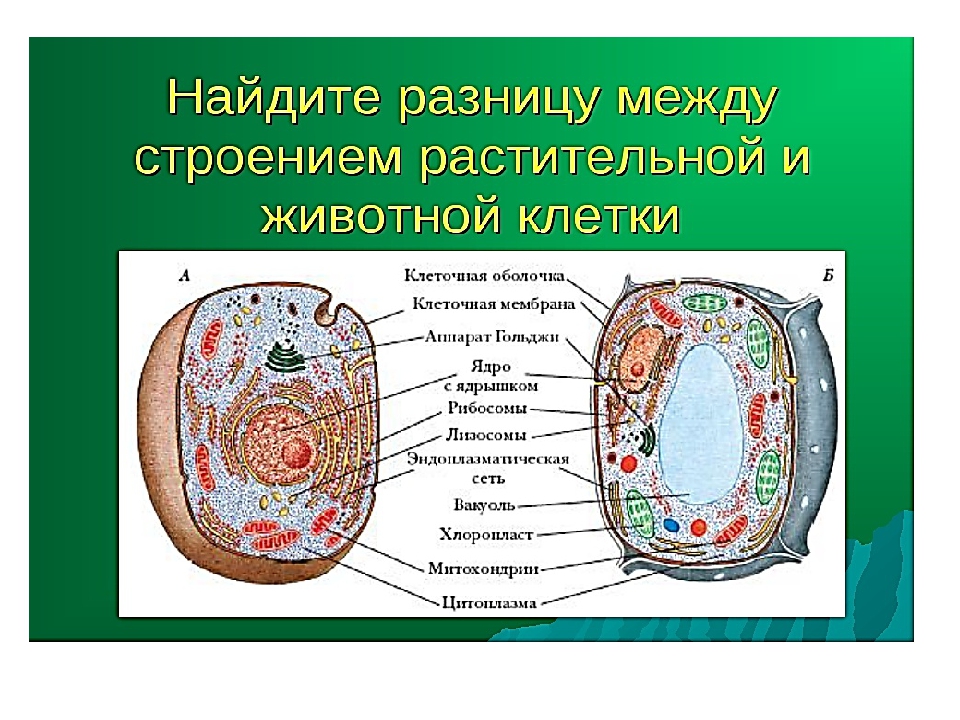
В строении тела наблюдается сходство и различие животных и растений. В чем же они состоят? Имеются сходства клеток растений и животных. Низшие растения и животные состоят из простых клеток. При этом они часто являются подвижными. Сходства и различия клеток растений и животных требуют детального рассмотрения. Предлагаем углубиться в этот вопрос.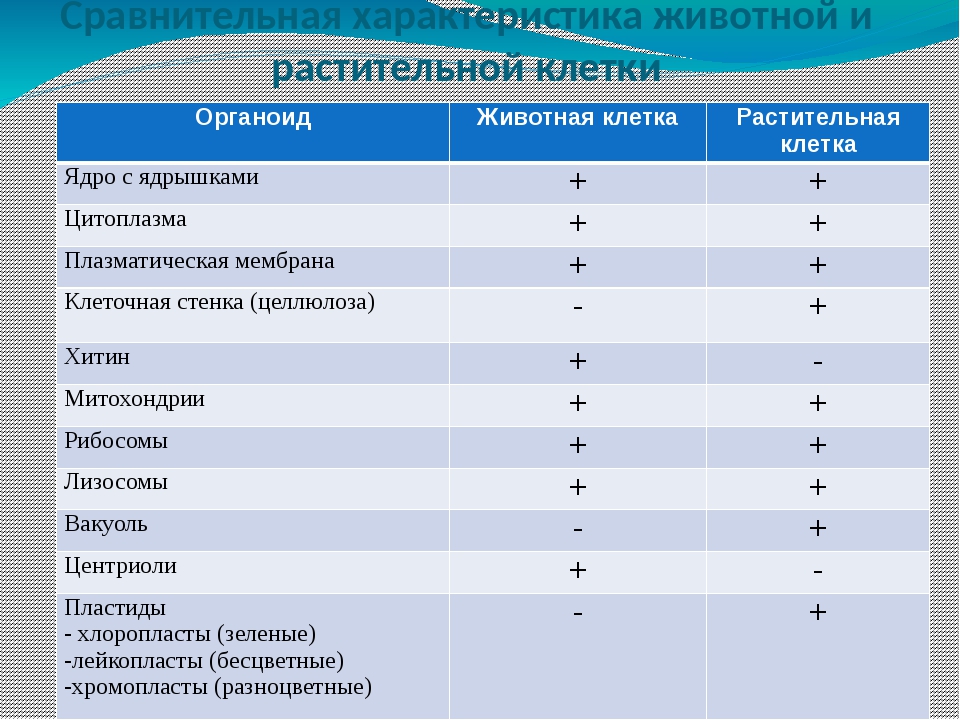
Клеточное строение
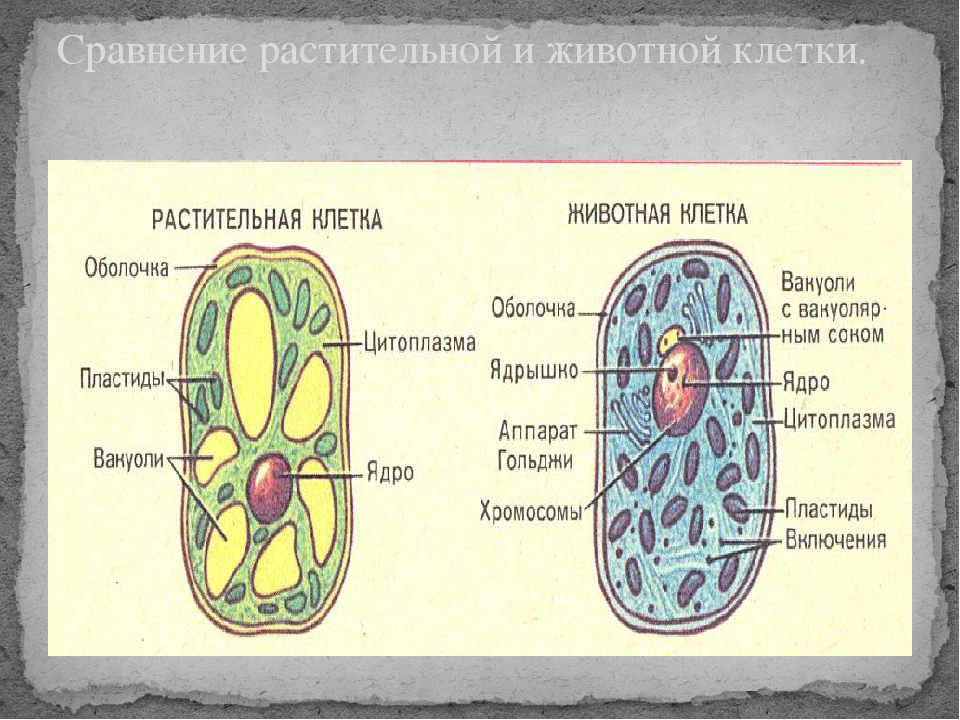
То, что между ними наблюдается сходство, является результатом общности происхождения жизни. Как животные, так и растительные клетки обладают следующими свойствами: они живые, делятся, растут, в них происходит обмен веществ. В клетках и тех и других организмов есть цитоплазма, ядро, митохондрии, эндоплазматическая сеть, аппарат Гольджи, рибосомы.
Что касается различий, то они появились в результате разных путей развития, расхождения в питании, а также появления возможности у животных двигаться самостоятельно, в отличие от растений. У последних клеточная стенка есть, она состоит из целлюлозы. У животных же ее не наблюдается. Функция клеточной стенки состоит в том, что она придает дополнительную жесткость растениям, а также защищает эти организмы от потерь воды. У животных нет вакуоли, а у растений она есть. Хлоропласты имеются исключительно у представителей растительного царства. В них формируются из неорганических органические вещества, при этом происходит поглощение энергии. Животные же питаются готовыми органическими веществами. Они получают их с пищей.
Животные же питаются готовыми органическими веществами. Они получают их с пищей.
Развитие животных и растений
У многоклеточных животных имеется важная особенность. Она состоит в том, что тело этих организмов снабжено множеством полостей. Их можно рассматривать как результат того, что покровы оказались ввернуты внутрь тела животного. Эти полости в большинстве своем образуются именно так. Иногда они появляются в результате расщепления тканей, которые формируют тело животного. Развитие животного, таким образом, можно свести к появлению ряда складок, а также загибов внутрь организма. Что касается многоклеточных растений, то в этом смысле они лишены полостей. Если у них имеются сосуды, то они формируются с помощью прободения и слияния рядов клеток. Однако развитие растений сводится к тому, что у них образуются выступы наружу плотного зачатка. Это и приводит к тому, что появляются различные придатки организма, такие как корни, листья и т. д.
Подвижность
Сходство и различие животных и растений наблюдаются также в подвижности. Животные обладают большей подвижностью. Из-за этого клетки их в большинстве своем являются голыми.
Животные обладают большей подвижностью. Из-за этого клетки их в большинстве своем являются голыми.
У малоподвижных растений же, как мы уже говорили, они одеты плотной оболочкой. Она состоит из целлюлозы (клетчатки). Раздражительность и подвижность не являются исключительными свойствами животных. Однако эти особенности у них все-таки достигают высшего развития. Тем не менее подвижны не только одноклеточные, но и многоклеточные растения. Между одноклеточными растениями и животными или же зародышевыми стадиями многоклеточных наблюдается сходство даже в том, какие они используют способы движения. И тем и другим свойственны такие, которые осуществляются непостоянными отростками, иначе называемыми псевдоподиями. Это называется амебоидным движением. Сходство между растениями и животными состоит в том, что и те и другие могут перемещаться, используя жгуты.
Они также могут делать это при помощи выделений вещества из своего тела. Эти выделения позволяют организму двигаться в нужную сторону, противоположную направлению истечения вещества.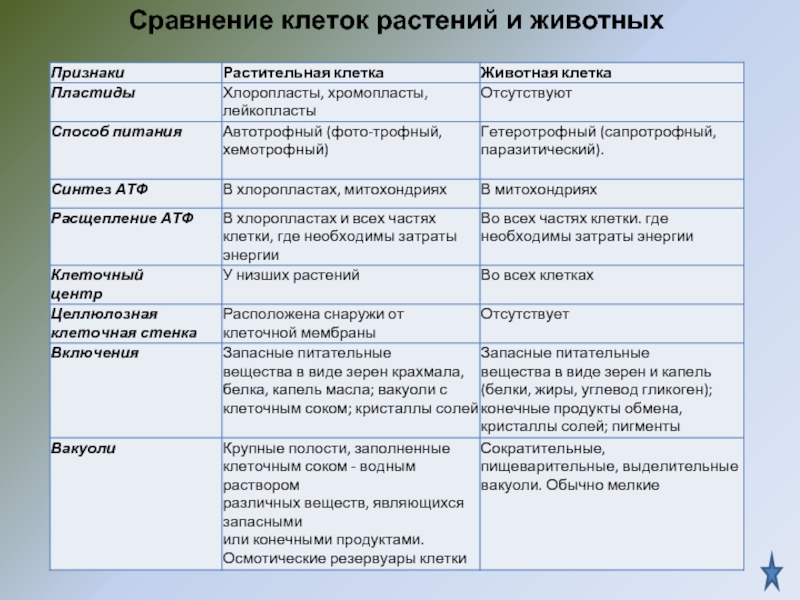 Данным свойством обладают, в частности, диатомовые водоросли и грегарины. Многоклеточные высшие растения поворачивают листья к свету определенным образом. Некоторые из них складывают их на ночь. В этом случае можно говорить о явлениях так называемого сна растений. Некоторые виды способны отвечать движениями на прикосновение, сотрясение и другие раздражения.
Данным свойством обладают, в частности, диатомовые водоросли и грегарины. Многоклеточные высшие растения поворачивают листья к свету определенным образом. Некоторые из них складывают их на ночь. В этом случае можно говорить о явлениях так называемого сна растений. Некоторые виды способны отвечать движениями на прикосновение, сотрясение и другие раздражения.
Весьма интересны эти черты сходства животных и растений. Однако многие другие не менее любопытны. Предлагаем вам узнать и о них.
Обособление мышечной и нервной ткани
Следующее сходство и отличие животных и растений связано с мышечной и нервной тканью. Чарльз Дарвин показал, что кончики корней и стеблей всех растений совершают вращательное движение. Однако лишь у многоклеточных животных существует обособление в качестве отдельной ткани сократимой мышечной, выполняющей функцию раздражимости, а также обособление специальных органов чувств, которые служат для восприятия различных раздражений. Но и среди многоклеточных животных имеются виды, не обладающие обособленными нервной и мышечной тканью, а также органами чувств. Это, к примеру, некоторые губки.
Это, к примеру, некоторые губки.
Способ питания растений
В питании также имеется сходство и отличие животных и растений. Однако здесь определенности все-таки больше. Считается, что основное отличие между растениями и животными сводится именно к типу их питания. Растения с помощью хлорофилла (зеленого пигмента) формируют органическое вещество из кислорода, углерода и водорода, которые они находят в воде и в воздухе. Так создается клетчатка, крахмал и другие вещества, не содержащие азота. А путем присоединения азота, находимого в почве в виде азотистых солей, растение строит и белковые вещества. Таким образом, эти организмы способны находить пищу везде. В жизни растений движение не может играть такой большой роли, как у животных.
Способ питания животных
Эти организмы могут существовать лишь за счет органических соединений, представленных в готовом виде. Они получают их или от растений, или от других животных, то есть в конечном счете от растений.
Животное должно суметь добыть себе пищу. Именно отсюда проистекает его большая подвижность. Растение формирует органические соединения, животное же их разрушает. Оно сжигает эти соединения в своем теле. В результате этого процесса выделяются продукты распада в виде мочи и углекислоты. Животное все время выделяет из атмосферы обратно в атмосферу угольную кислоту. При жизни своей оно освобождает азот через мочеиспускание, а после гибели – при разложении. Растение берет из атмосферы угольную кислоту. Нитрогенные бактерии осуществляют перевод азота в почву. Из нее он вновь потребляется растениями.
Именно отсюда проистекает его большая подвижность. Растение формирует органические соединения, животное же их разрушает. Оно сжигает эти соединения в своем теле. В результате этого процесса выделяются продукты распада в виде мочи и углекислоты. Животное все время выделяет из атмосферы обратно в атмосферу угольную кислоту. При жизни своей оно освобождает азот через мочеиспускание, а после гибели – при разложении. Растение берет из атмосферы угольную кислоту. Нитрогенные бактерии осуществляют перевод азота в почву. Из нее он вновь потребляется растениями.
Особенности дыхания
Сходства и различия между животными и растениями касаются также дыхания. По поводу того, которое сопровождается выделением углекислоты и поглощением кислорода, можно сказать, что оно свойственно в равной степени как растениям, так и животным. Однако у последних этот процесс совершается намного энергичнее.
У растений же такое дыхание заметно только тогда, когда процесс питания, противоположный этому процессу, не совершается. Питание – это поглощение углекислоты, при котором в атмосферу выбрасывается часть кислорода. Оно может не совершаться, к примеру, при прорастании семян или же в темноте.
Питание – это поглощение углекислоты, при котором в атмосферу выбрасывается часть кислорода. Оно может не совершаться, к примеру, при прорастании семян или же в темноте.
Поскольку процесс горения у животных происходит энергичнее, повышение температуры у них заметнее и сильнее, нежели у растений. Таким образом, дыхание у растений все-таки существует, однако основная роль этих организмов в круговороте веществ заключается в поглощении углекислоты, освобождении кислорода, а также потреблении азота, находящегося в атмосфере (с помощью бактерий). У животных же обратная роль. Они производят в атмосферу углекислоту и азот (тоже частично с помощью бактерий – при гниении), а кислород поглощают.
Питание: исключения из правил
Нередко наблюдается сходство растений и животных в том, как у них осуществляется питание. К примеру, не содержащие хлорофилла грибы используют в пищу уже готовые органические вещества. А некоторые жгутиковые и бактерии могут создавать органическое вещество, при этом они лишены хлорофилла. Ряд насекомоядных растений способен захватывать и перерабатывать животные ткани. Таким образом проявляется сходство растений и животных. Некоторые виды жгутиковых, которые содержат хлорофилл, вырабатывают на свету зерна, по своим свойствам сходные с крахмальными. Значит, они питаются тем же способом, что и растения. А в темноте их питание происходит сапрофитически, то есть оно осуществляется всей поверхностью тела за счет разлагающихся веществ.
Ряд насекомоядных растений способен захватывать и перерабатывать животные ткани. Таким образом проявляется сходство растений и животных. Некоторые виды жгутиковых, которые содержат хлорофилл, вырабатывают на свету зерна, по своим свойствам сходные с крахмальными. Значит, они питаются тем же способом, что и растения. А в темноте их питание происходит сапрофитически, то есть оно осуществляется всей поверхностью тела за счет разлагающихся веществ.
Нетипичный химический состав элементов
Сходство растений и животных наблюдается и в химическом составе элементов, из которых сложены их тела. Деятельный хлорофилл, правда, свойственен только растениям. В некоторых случаях его можно найти в организме высших животных. Однако при этом он принадлежит не им, а водорослям. Некоторые из них симбиотически живут в теле животных. Нам уже известно о том, что хлорофилла лишены многие растения. С другой стороны, Euglena, которая имеет деятельный хлорофилл, и другие формы, подобные ей, имеют чуть ли ни такое же право быть отнесенными к животному царству, как и к растительному. На сегодняшний день не доказано сходство с хлорофиллом зеленого пигмента, имеющегося в крыльях прямокрылых насекомых. Этот пигмент, во всяком случае, не функционирует в них как хлорофилл.
На сегодняшний день не доказано сходство с хлорофиллом зеленого пигмента, имеющегося в крыльях прямокрылых насекомых. Этот пигмент, во всяком случае, не функционирует в них как хлорофилл.
Похожие вещества
Сходство растений и животных проявляется и в похожих веществах, присутствующих в их телах. Для первых характерно наличие клетчатки. Однако оболочка, облекающая тела ряда морских животных, состоит из туницина. Данное вещество имеет сходство с клетчаткой. Для растений, как известно, свойственно такое вещество, как крахмал. Однако в жизни животных его изомер (гликоген) играет такую же важную роль. А у миксомицетов, или слизистых грибов, вместо крахмала имеется как раз гликоген.
Вывод
Все вышесказанное приводит нас к заключению о том, что различия между растениями и животными достаточно условны. Можно сделать вывод и о том, что и те и другие ведут свое происхождение из некоего общего источника, то есть от таких форм, которые могут быть с полным правом отнесены как к растениям, так и к животным.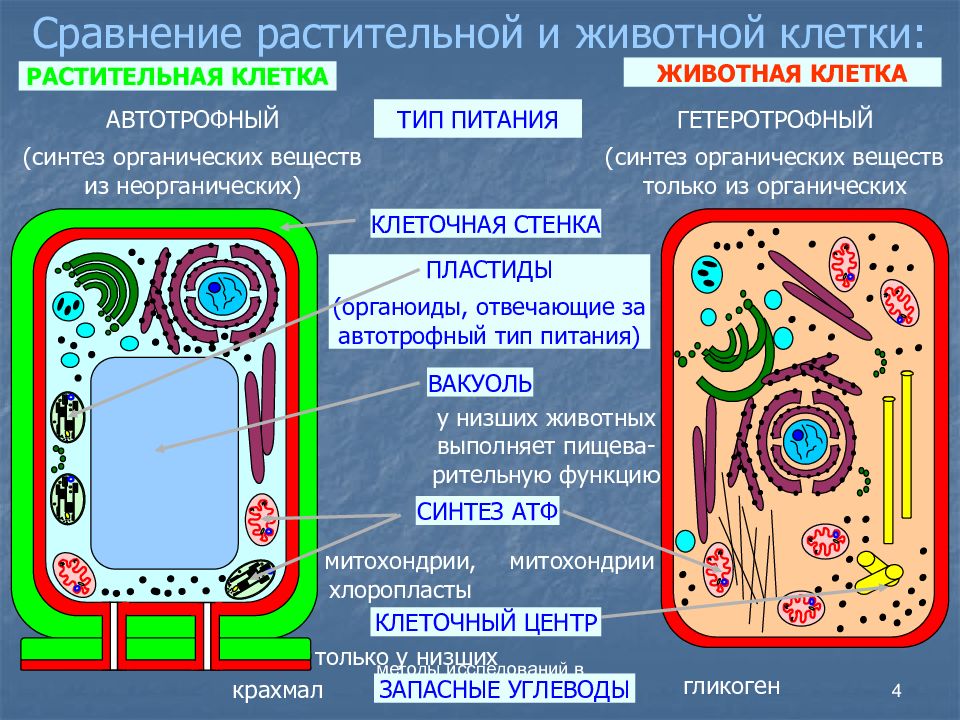 Эти формы отчасти сохранились на нашей планете.
Эти формы отчасти сохранились на нашей планете.
Сходство и различия клеток растений грибов и животных
10 класс (базовый уровень)Тема: Сходство и различия в
строении клеток
растений, животных и грибов
Максимова Л.А.
Учитель биологии
МБОУ СОШ № 166
Г. Самара
Клетка растений
Клетка животных
Клетка грибов
СВОЙСТВА
ПЛАСТИДЫ
ЛЕЙКОПЛАСТЫ
Из моно- и дисахари
дов синтезируется
крахмал, запасают
белки и жиры.
ХЛОРОПЛАСТЫ
ХРОМОПЛАСТЫ
Осуществляют
фотосинтез
Содержат
пигменты
каротиноиды,
обуславливают
яркую окраску
цветов и плодов.
ФУНКЦИИ
Типы питания
клеток
Автотрофы
Бактерии
Растения
Гетеротрофы
бактерии
грибы
животные
Доказательства родства царств живой природы
Растения
Грибы
Животные
Общий план строения клетки
Принципиальное сходство процессов обмена веществ и
энергии
Кодирование наследственной информации
Единство химического состава
Сходные процессы деления клеток
С растениями
С животными
Клеточная стенка
— хитин
Способ питания гетеротрофы
Не способны к
активному
движению
Рост
неограниченный
Гликоген –
запасное
вещество
Различия между царствами живой природы
Признаки
Способ
питания
Клеточная
стенка
Пластиды
вакуоли
Запасной
углевод
Центриоли
Деление
Растения
Автотрофы
+
целлюлоз
ная
+
крахмал
—
Перфорация
от центра к
оболочке
Грибы
Гетеротрофы
+
хитиновая
—
гликоген
+
Образуется
перетяжка
между
дочерними
клетками
Животные
Гетеротрофы
—
—
гликоген
+
Образуется
перетяжка
между
дочерними
клетками
Царства
принадлежат ::
которые принадлежат
клеток, которые
номера клеток,
Выберите номера
№1.
 Выберите
ВыберитеЗадание №1.
Задание
грибов
царству бактерий
– кк царству
В–
животных, В
царству животных,
–к царству
Б –к
растений, Б
царству растений,
А –– кк царству
А
№1
№2
№2
№3
№4
№4
№3
№1
№6
№6
№8
№9
№5
№5
№6 №7
№7
№8
№9
№10
Домашнее задание:
1.Изучить §19, ответить на вопросы
2.Подготовить сообщения:
1) История открытия вирусов
2) Д.И. Ивановский. Вклад в науку биологию
3)Синдром иммунодефицита (СПИД)
Профилактика заболевания.
19. Сходство и различия в строении клеток растений, животных и грибов
Параграф 19
Модуль 19
Сходство и различия в строении клеток растений, животных и грибов
Рассказать о сходстве в строении всех эукариотических клеток
• Царства эукариотических организмов.
• Общие предки эукариот.
•
Ядерные организмы.
• Другие общие части клеток.
• Сходство процессов обмена.
• Кодирование наследственной информации.
• Химический состав.
• Деление клеток.
Какими главными отличительными особенностями обладают растительные клетки?
• Способ питания.
• Строение клеточной стенки.
• Пластиды.
• Хранение питательных веществ.
• Клеточный центр.
• Деление клетки.
Какими главными отличительными особенностями обладают животные клетки?
• Способ питания.
• Отсутствие клеточной стенки.
• Особенности вакуолей.
• Хранение питательных веществ.
• Центриоли.
• Деление клетки.
Какими главными отличительными особенностями обладают клетки грибов?
• Способ питания.
• Типы питания.
•
Особенности клеточной стенки.
• Хранение питательных веществ.
• Наличие гифов.
• Многоядерные клетки.
• Неограниченный рост.
• Способы размножения.
Итоговые задания
№ 1
Дайте определения следующим понятиям: сапрофиты, паразиты, симбионты, гифы, автотрофы, фототрофы, гетеротрофы.
№ 2 (заполните пропуски)
Грибы не относятся ни к царству 1) …, так как у них нет
хлорофилла, ни к царству 2) …, так как ведут прикреплённый образ жизни, ни к
царству 3) …, так как многие из них являются 4) …. Они образуют особое царство,
которое и называется 5) … …. Главной особенностью их строения является то, что
их тело состоит из 6) …, нитевидных структур, образованных одним рядом 7) … Их
клеточные стенки состоят из 8) … — особого вещества, который образует также
наружные покровы у 9) … Все грибы делятся на несколько групп по типу питания.
Из них выделяются 10) …, питающиеся органикой мёртвых существ, 11) …,
питающиеся живой органикой, и 12) …, находящиеся во взаимовыгодной связи с 13)
… ….
№ 3 (исправьте предложения)
1. Главным отличием грибов от растений является их способность вести хищнический образ жизни.
2. Грибы не способны к автотрофному питанию, то есть все они являются сапрофитами, питающиеся готовыми органическими веществами.
3. Некоторые грибы могут быть симбионтами, то есть питаться за счёт животных, в организме которых они поселяются.
4. Тело грибов представляют собой мицелий, состоящий из большого количества плодовых тел, которые способны разрастаться на большие расстояния и отделяться от материнского организма, начиная самостоятельную жизнь.
5. Многие грибы необходимы для организма животных, так как они способны защищать их от многих опасных заболеваний.
6.
Особенностью, сближающей грибы с животными,
является отсутствие клеточной стенки, которая имеется только у растений и
состоит из хитина.
Сходство бактерий, грибов, животных — свидетельство единства живой природы — Учебник по Биологии. 7 класс. Запорожец
Учебник по Биологии. 7 класс. Запорожец — Новая программа
Мы с вами переворачиваем последние страницы учебника биологии для 7-го класса. Вы изучаете биологию в школе уже два года и продолжите изучение увлекательного мира живых организмов в течение всей жизни. Наверное, больше всего вас удивило многообразие жизненных форм: организмы отличаются и по форме, и по размерам, и по способам питания, и по приспособлению к определенным условиям. Подытоживая все, чему вы научились, все, о чем узнали во время интересных незабываемых уроков, давайте сделаем выводы.
Наряду с многочисленными отличиями мы видим определенное единство, общие черты, похожие приспособления у бактерий, растений, животных и грибов. Давайте подробнее остановимся именно на чертах сходства и их причинах.
Все они имеют общие признаки живых организмов. Для них характерны обмен веществ (питание, дыхание, выделение), рост, развитие, движение, размножение и раздражимость. Все они характеризуются сходством химического состава. В их телах по массе преобладает вода, которая выполняет важные функции в каждом организме. Липиды, белки, углеводы и нуклеиновые кислоты выполняют в организме важные функции, они являются определяющими в жизнедеятельности организма.
Все они характеризуются сходством химического состава. В их телах по массе преобладает вода, которая выполняет важные функции в каждом организме. Липиды, белки, углеводы и нуклеиновые кислоты выполняют в организме важные функции, они являются определяющими в жизнедеятельности организма.
Бактерии, растения, животные и грибы являются клеточными организмами — это одна из их основных общих черт. У многоклеточных растений и животных из клеток образуются ткани и органы. С помощью регуляторных систем все они действуют согласованно.
Рост и развитие многоклеточных организмов происходит путем деления клеток, которое проходит у растений и животных по одному принципу и схеме.
Всем этим организмам свойственно бесполое размножение, а растениям, грибам и животным — еще и половое. В основе полового размножения лежит образование половых клеток — яйцеклеток и сперматозоидов, которые сливаются при оплодотворении. Организм, образующийся в результате полового размножения, несет признаки материнского и отцовского организмов, что дает материал для дальнейшего исторического развития, т. е. эволюционного процесса.
е. эволюционного процесса.
Существуют определенные черты сходства в строении клеток бактерий, растений, животных и грибов: у них есть цитоплазма, мембрана. В клетках грибов, растений и животных есть ядро, которое управляет деятельностью клетки и сохраняет наследственный материал. Наследственный материал у этих организмов сохраняется и реализуется в соответствии с одинаковыми закономерностями, которые называют генетическим кодом. Генетический код един для всех организмов нашей планеты.
В клетках грибов, растений и животных есть органеллы, которые имеют сходное строение и выполняют одинаковые функции. Это, например, эндоплазматическая сеть, осуществляющая транспорт и синтез определенных веществ, рибосомы, синтезирующие белки, митохондрии, в которых расщепляются сложные органические вещества до простых, а энергия, которая при этом выделилась, аккумулируется в виде АТФ.
В чем же причина такого принципиального сходства? Конечно, мы не можем заглянуть в те времена, когда на нашей планете зарождались и развивались первые растения, животные, грибы. Но ученые, сравнивая разные научные факты, пришли к выводу, что, вероятно, у бактерий, растений, животных и грибов были общие предки, т. е. они имеют общее происхождение. Вероятнее всего, самыми древними организмами являются бактерии, от которых, очевидно, происходят предки современных растений, животных, грибов. На определенном этапе исторического развития они разделились и начали развиваться независимо. Так между ними накопились отличия. Дальнейшая их эволюция привела к увеличению численности, многообразия и степени приспособленности к окружающей среде.
Но ученые, сравнивая разные научные факты, пришли к выводу, что, вероятно, у бактерий, растений, животных и грибов были общие предки, т. е. они имеют общее происхождение. Вероятнее всего, самыми древними организмами являются бактерии, от которых, очевидно, происходят предки современных растений, животных, грибов. На определенном этапе исторического развития они разделились и начали развиваться независимо. Так между ними накопились отличия. Дальнейшая их эволюция привела к увеличению численности, многообразия и степени приспособленности к окружающей среде.
Светочувствительные белки архей и эукариот оказались родственниками
Живые организмы делятся на три основных группы (домена) по наличию ограниченных мембранами областей в составе клеток. Археи (первый домен) не содержат никаких мембранных частей в клетках. Более сложно устроены бактерии (второй домен) — в их клетках присутствуют органеллы, окруженные мембранами. Эукариоты (третий домен), в свою очередь, содержат и мембранные органеллы, и ядро в составе клеток.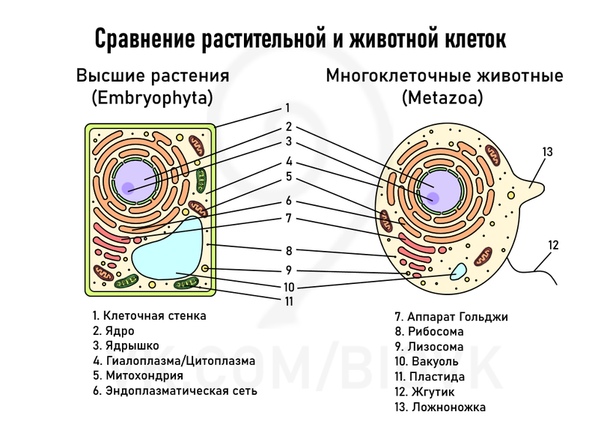
Ранее сравнение геномов многих животных показало, что светочувствительные белки в клетках выполняют чрезвычайно разные функции и присутствуют во всех царствах, а также во многих крупных вирусах. Наиболее распространенное семейство светочувствительных белков — родопсины. Родопсины первого типа состоят из семи пронизывающих мембрану клетки спиралей (Рисунок 1). Белки используют энергию света для активного и пассивного транспорта ионов через мембрану клетки, запуска сигнальных реакций и активации ферментов в клетках. Светочувствительные белки применяются в оптогенетике — методике исследования нервных импульсов, которые можно возбуждать при помощи света. Они считаются распространенными светособирающими белками на Земле и основными «световозахватителями» в океанах. Учитывая повсеместное распространение родопсинов и их важную экологическую роль, нет сомнений в том, что эти белки сыграли значительную роль в эволюции жизни на Земле. На ранних этапах эволюции многие организмы могли использовать родопсины в качестве одного из дополнительных источников энергии, что давало им эволюционное преимущество. Широкое разнообразие протонных помп, а также их древность позволяют исследовать глобальные эволюционные процессы по истории изменений этих белков. Однако родопсины эукариот исследованы менее обширно, так как их выделение и кристаллизация значительно сложнее.
Учитывая повсеместное распространение родопсинов и их важную экологическую роль, нет сомнений в том, что эти белки сыграли значительную роль в эволюции жизни на Земле. На ранних этапах эволюции многие организмы могли использовать родопсины в качестве одного из дополнительных источников энергии, что давало им эволюционное преимущество. Широкое разнообразие протонных помп, а также их древность позволяют исследовать глобальные эволюционные процессы по истории изменений этих белков. Однако родопсины эукариот исследованы менее обширно, так как их выделение и кристаллизация значительно сложнее.
Структура исследованного родопсина и схема соединения спиралей белка. Источник: Communications Biology
Авторы исследования использовали недавно разработанные ими же методы экспрессии (в системе LEXSY) для выделения родопсина 1 типа из одноклеточного гриба. Ученые кристаллизовали и получили структуру белка. Оказалось, что структура родопсина из гриба крайне похожа на структуру протонного насоса археи. Единственное значительное отличие было найдено во внутриклеточной части белка: петля ECL1 гораздо длиннее подобной у родопсина архей (см. рисунок 1). Авторы изучили функции этой петли и обнаружили, что она связывается с внутриклеточным концом, тем самым повышая стабильность белка. Для определения эволюционных отношений между родопсинами ученые сравнили известные структуры и последовательности белков. Оказалось, что последовательности и структуры родопсинов эукариот крайне схожи с таковыми у белков архей. Видимо, гены родопсинов подвергаются обширному горизонтальному переносу между организмами, что усложняет поиск общего предка. Однако высокое структурное сходство протонных помп архей и эукариот, наряду с функциональным сходством, представляет собой убедительное доказательство архейного происхождения эукариотических протонных родопсинов и, скорее всего, всех других эукариотических родопсинов.
Единственное значительное отличие было найдено во внутриклеточной части белка: петля ECL1 гораздо длиннее подобной у родопсина архей (см. рисунок 1). Авторы изучили функции этой петли и обнаружили, что она связывается с внутриклеточным концом, тем самым повышая стабильность белка. Для определения эволюционных отношений между родопсинами ученые сравнили известные структуры и последовательности белков. Оказалось, что последовательности и структуры родопсинов эукариот крайне схожи с таковыми у белков архей. Видимо, гены родопсинов подвергаются обширному горизонтальному переносу между организмами, что усложняет поиск общего предка. Однако высокое структурное сходство протонных помп архей и эукариот, наряду с функциональным сходством, представляет собой убедительное доказательство архейного происхождения эукариотических протонных родопсинов и, скорее всего, всех других эукариотических родопсинов.
«Мы получили первую кристаллическую структуру высокого разрешения светочувствительного протонного насоса из организма гриба и выяснили функциональную роль его N-концевой области.
Исследованный родопсин был экспрессирован в системе LEXSY, затем мы его кристаллизовали. Значит, система экспрессии LEXSY может быть сильным инструментом для получения мембранных белков эукариот для структурных исследований. Мы также сравнили последовательность и структуру полученного родопсина со светочувствительными протонными помпами из разных царств. Анализ показал, что эукариотические и архейные родопсины имеют глубокое структурное сходство, что подтверждает гипотезу об архейном происхождении родопсинов, найденных в геноме эукариот. Полученные результаты важны как для понимания эволюции животных, так и для дальнейших исследований родопсинов эукариот», — поясняет Дмитрий Забельский, сотрудник лаборатории химии и физики липидов Центра исследований молекулярных механизмов старения и возрастных заболеваний МФТИ, аспирант Физтех-школы физики и исследований им. Ландау МФТИ.
Работа выполнена при поддержке Министерства науки и высшего образования РФ, а также Российского научного фонда и Российского фонда фундаментальных исследований.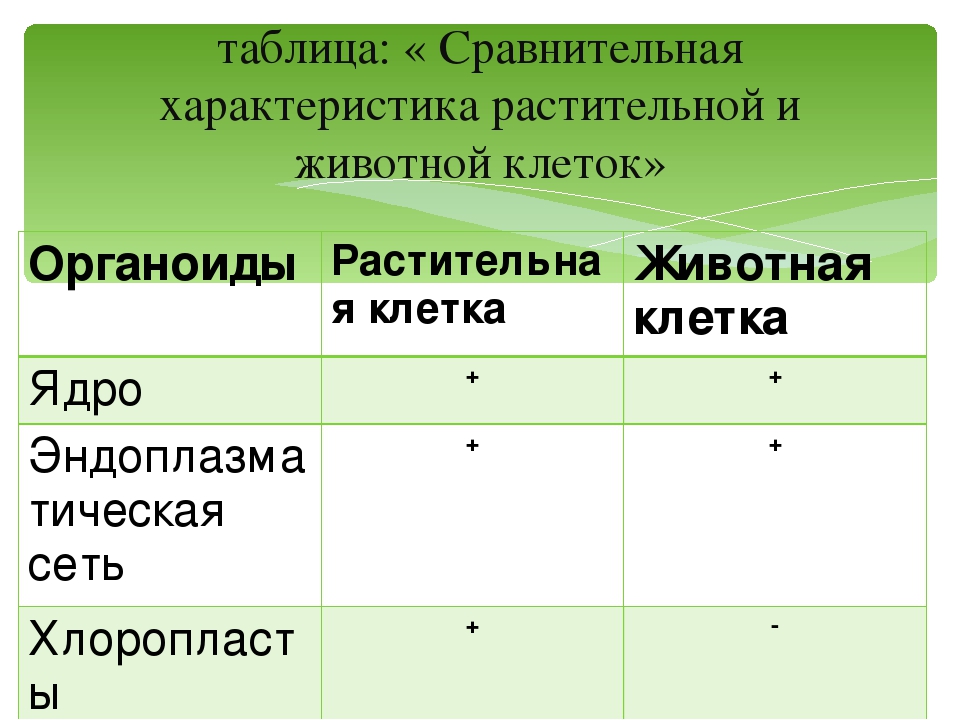
Клетки растений и клетки животных: объяснение 10 ключевых различий
Клетки растений и животных чрезвычайно важны для выживания миллионов видов, и без них жизнь не могла бы существовать. Несмотря на то, что у них много общего, у них также есть немало различий, и даже самая маленькая разница может иметь огромное значение для работы, которую они выполняют. Так чем же они похожи и что отличает клетки растений от клеток животных?
Ну, они разного размера и формы и хранят разные виды энергии.Оба имеют ядро, хотя в одном оно расположено в центре, а в другом нет. У них обоих есть клеточные мембраны, хотя у одного есть клеточная стенка, а у другого нет.
Сравнение клеток животных и клеток растений
Растительные и животные клетки имеют много различий, и каждая отдельная часть играет жизненно важную роль. Они настолько важны, что считаются строительными блоками всего живого на земле — как растений, так и животных.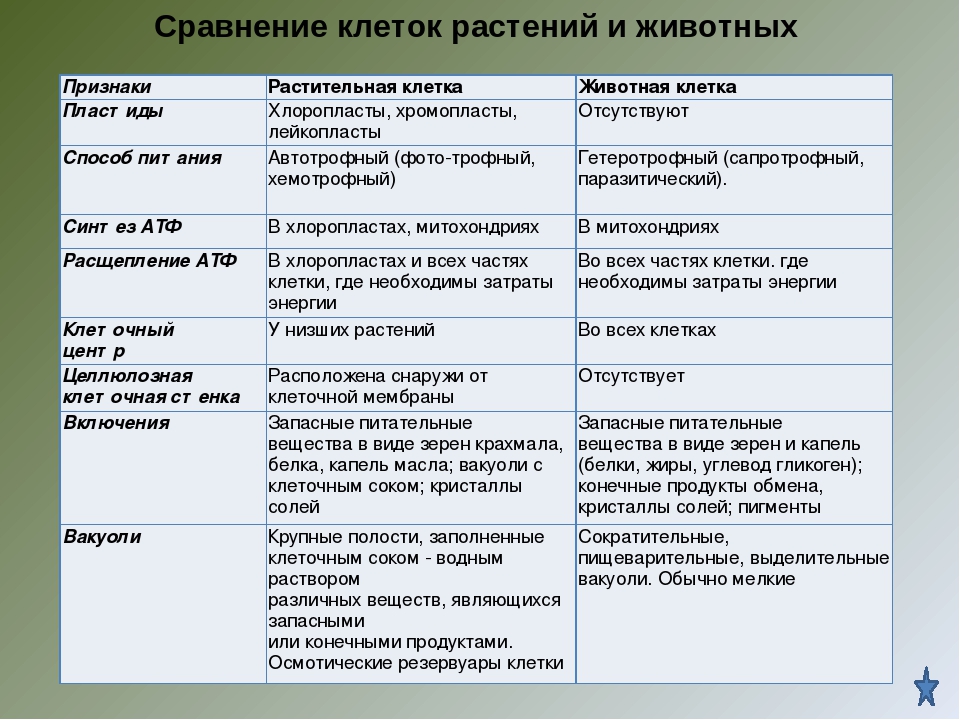 Хотя большинство клеток настолько крошечные, что их можно увидеть только в микроскоп, между ними есть некоторые важные различия.
Хотя большинство клеток настолько крошечные, что их можно увидеть только в микроскоп, между ними есть некоторые важные различия.
Ознакомьтесь с приведенной ниже таблицей, чтобы узнать о некоторых основных отличиях.
| 20015 клетки животных | клетки | ||
| Размер | |||
| 10 — 30 микрометров | 10 — 100 микрометров | 10 — 100 микрометров | |
| Форма | Нерегулярная форма | фиксированная, жесткая форма — часто Прямоугольный | |
| No | NO | Да | |
| Choroplasts | No | NO | Да |
| Nucleus Местоположение | Central | Central | |
| Vacuole | Multi вакуоли | Большая центральная вакуоля | |
| запасенная энергия | гликоген | Крахмал | |
| плазмодесмы | нет | Да | |
| лизосомы | Да | Редко | |
| Центриоль | Да | Редко |
 com
com 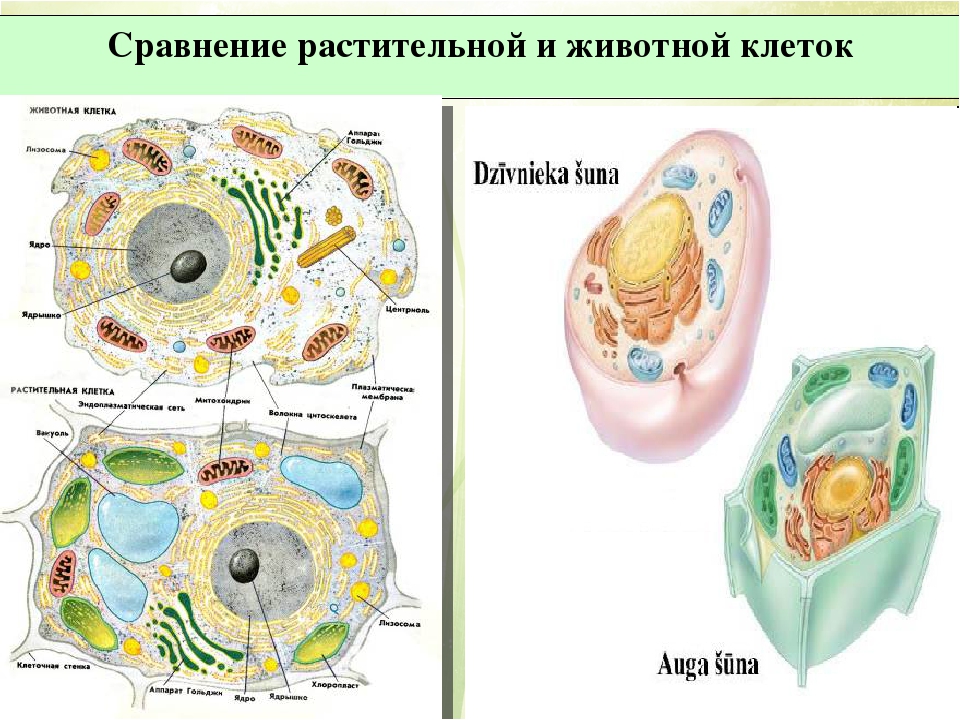 Вакуоли внутри растительной клетки иногда могут занимать до 90% ее объема.
Вакуоли внутри растительной клетки иногда могут занимать до 90% ее объема. 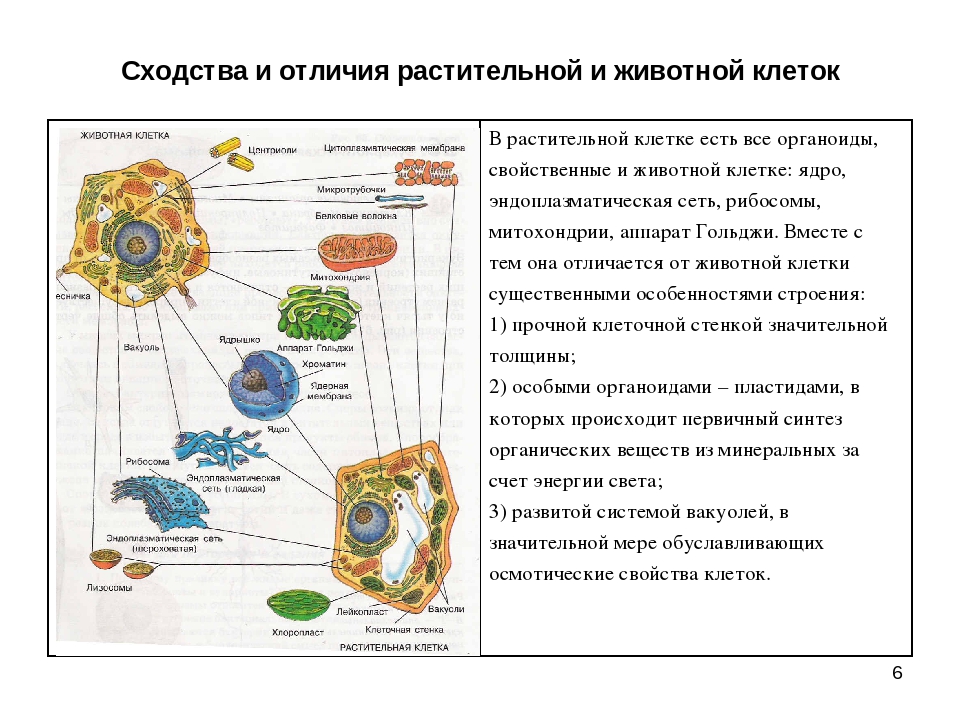 Клетки растений нуждаются в хлоропластах, чтобы они могли преобразовывать энергию солнечного света в полезную энергию посредством фотосинтеза. Животные этого не делают, поэтому хлоропласты им не нужны.
Клетки растений нуждаются в хлоропластах, чтобы они могли преобразовывать энергию солнечного света в полезную энергию посредством фотосинтеза. Животные этого не делают, поэтому хлоропласты им не нужны. 
 Эукариотические клетки представляют собой клетки, содержащие ядро и другие мембраносвязанные органеллы.
Эукариотические клетки представляют собой клетки, содержащие ядро и другие мембраносвязанные органеллы. 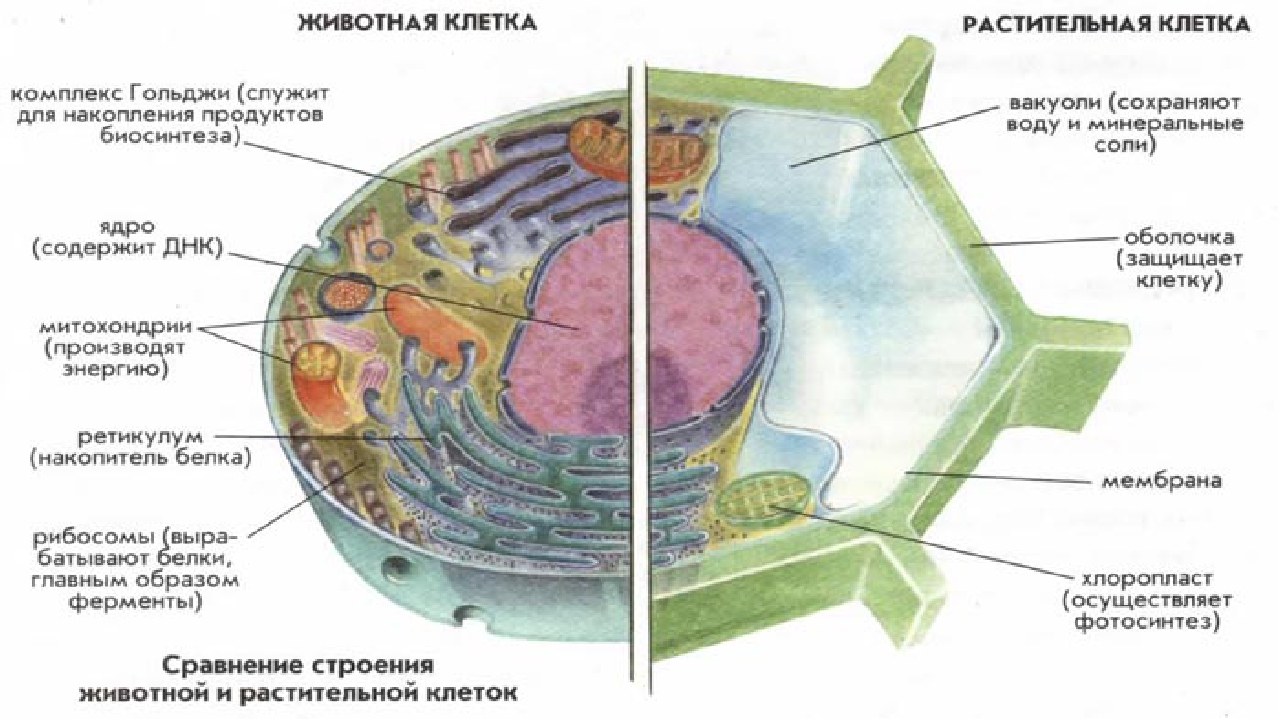 Вот почему растения известны как производители, а животные — как потребители.
Вот почему растения известны как производители, а животные — как потребители. , 2014). За исключением животных и простейших, большинство типов клеток покрыты клеточной стенкой (CW), сложной сетью белков и углеводов, в которой также могут откладываться фенольные соединения во время определенных физиологических процессов (Keegstra, 2010; Bellincampi et al.). др., 2014). Название CW описывает характеристики жесткости, поддержки и реальной формы, придаваемые этим конкретным ECM растительным клеткам (Guerriero et al., 2014). Помимо структурных функций, растительные КС играют важнейшую физиологическую роль, среди которых создание тургорного давления, контроль межклеточной коммуникации и защитная реакция от вредителей и патогенов (Lionetti and Metraux, 2014; Lionetti et al., 2014a, 2015; Whitehill et al., 2016). ). Совсем недавно убедительные доказательства изображают CW растений как динамическую структуру, в значительной степени реконструированную для решения новых физиологических функций (Ebine and Ueda, 2015; Lionetti et al., 2017).
, 2014). За исключением животных и простейших, большинство типов клеток покрыты клеточной стенкой (CW), сложной сетью белков и углеводов, в которой также могут откладываться фенольные соединения во время определенных физиологических процессов (Keegstra, 2010; Bellincampi et al.). др., 2014). Название CW описывает характеристики жесткости, поддержки и реальной формы, придаваемые этим конкретным ECM растительным клеткам (Guerriero et al., 2014). Помимо структурных функций, растительные КС играют важнейшую физиологическую роль, среди которых создание тургорного давления, контроль межклеточной коммуникации и защитная реакция от вредителей и патогенов (Lionetti and Metraux, 2014; Lionetti et al., 2014a, 2015; Whitehill et al., 2016). ). Совсем недавно убедительные доказательства изображают CW растений как динамическую структуру, в значительной степени реконструированную для решения новых физиологических функций (Ebine and Ueda, 2015; Lionetti et al., 2017).  Им необходимо проникнуть в живые клетки хозяина и вступить в контакт с цитоплазмой (Димитров, 2004). Самая ранняя и наиболее важная стадия вирусной инфекции — проникновение в клетку и последующая передача вирусного генетического материала (Smith and Helenius, 2004; Alsteens et al., 2017). После репликации вирусы могут перемещаться непосредственно между соседними клетками и проникать в хозяина с помощью механизмов распространения, которые могут сильно зависеть от конкретного способа проникновения вируса в клетку-хозяина и выхода из нее.CW растений представляет собой физический барьер для проникновения вирусов и повышает уровень сложности межклеточного перемещения вирусов (Lionetti et al., 2012; Knox and Benitez-Alfonso, 2014). Противоположное положение имеет место в случае ВКМ животных клеток, компоненты которого могут действовать как вирусные рецепторы, способствуя распознаванию, прикреплению и проникновению вируса в клетку.
Им необходимо проникнуть в живые клетки хозяина и вступить в контакт с цитоплазмой (Димитров, 2004). Самая ранняя и наиболее важная стадия вирусной инфекции — проникновение в клетку и последующая передача вирусного генетического материала (Smith and Helenius, 2004; Alsteens et al., 2017). После репликации вирусы могут перемещаться непосредственно между соседними клетками и проникать в хозяина с помощью механизмов распространения, которые могут сильно зависеть от конкретного способа проникновения вируса в клетку-хозяина и выхода из нее.CW растений представляет собой физический барьер для проникновения вирусов и повышает уровень сложности межклеточного перемещения вирусов (Lionetti et al., 2012; Knox and Benitez-Alfonso, 2014). Противоположное положение имеет место в случае ВКМ животных клеток, компоненты которого могут действовать как вирусные рецепторы, способствуя распознаванию, прикреплению и проникновению вируса в клетку.  , 2010; Кулар и др., 2014; Теохарис и др., 2016). ECM также обогащены белками, такими как коллагены (основной структурный белок соединительной ткани), эластин, фибронектин, ламинины и гликопротеины. ГАГ представляют собой длинные и отрицательно заряженные гетерополисахариды, характеризующиеся дисахаридными повторами N -ацетилированных гексозаминов и D -/ L -гексуроновой кислоты, которые замещены сульфатными группами в различных положениях. Основными ГАГ являются галактозаминогликаны хондроитинсульфат (CS) и дерматансульфат (DS), а также гликозаминогликаны кератансульфат (KS), гепарин (Hep) и гепарансульфат (HS).Гиалуронан (ГК) также является важным компонентом ЭМС. Уникальная среди ГАГ, ГК биосинтезируется на клеточной мембране, а не в аппарате Гольджи, не сульфатирована и не связана с белками. Клетки, встроенные в ECM, взаимодействуют с этой макромолекулярной сетью через свои поверхностные рецепторы, такие как интегрины, рецепторы домена дискоидина (DDR), PG клеточной поверхности и рецептор HA CD44 (Bosman and Stamenkovic, 2003).
, 2010; Кулар и др., 2014; Теохарис и др., 2016). ECM также обогащены белками, такими как коллагены (основной структурный белок соединительной ткани), эластин, фибронектин, ламинины и гликопротеины. ГАГ представляют собой длинные и отрицательно заряженные гетерополисахариды, характеризующиеся дисахаридными повторами N -ацетилированных гексозаминов и D -/ L -гексуроновой кислоты, которые замещены сульфатными группами в различных положениях. Основными ГАГ являются галактозаминогликаны хондроитинсульфат (CS) и дерматансульфат (DS), а также гликозаминогликаны кератансульфат (KS), гепарин (Hep) и гепарансульфат (HS).Гиалуронан (ГК) также является важным компонентом ЭМС. Уникальная среди ГАГ, ГК биосинтезируется на клеточной мембране, а не в аппарате Гольджи, не сульфатирована и не связана с белками. Клетки, встроенные в ECM, взаимодействуют с этой макромолекулярной сетью через свои поверхностные рецепторы, такие как интегрины, рецепторы домена дискоидина (DDR), PG клеточной поверхности и рецептор HA CD44 (Bosman and Stamenkovic, 2003).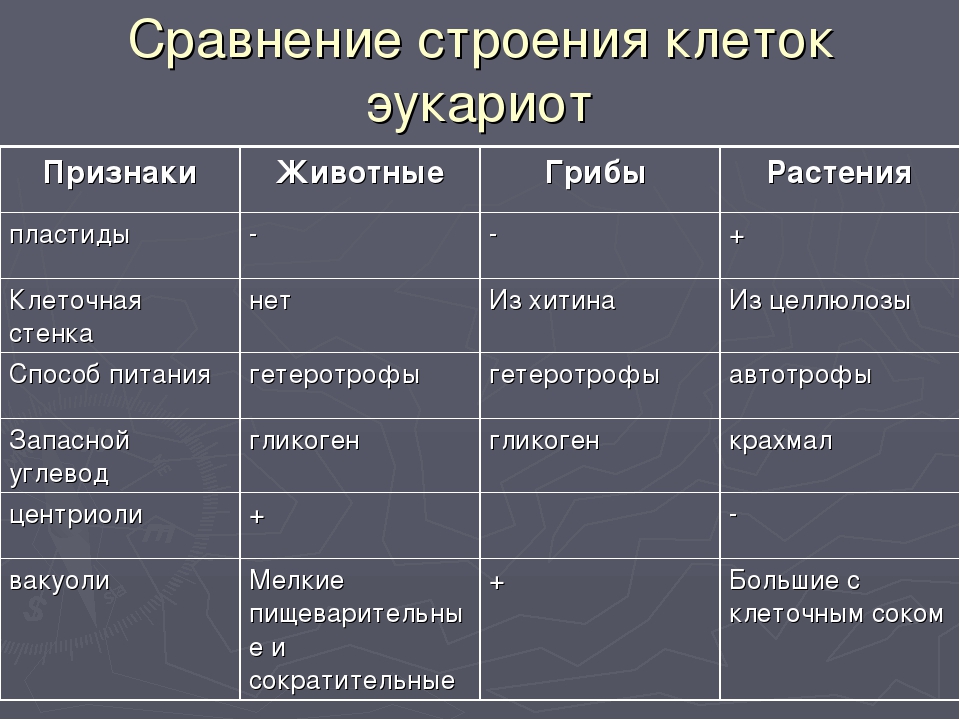 Различные типы клеток синтезируют и секретируют макромолекулы матрикса под контролем множества сигналов.Изменения в составе и структуре ВКМ, которые могут быть эндогенно опосредованы протеиназами, такими как матриксные металлопротеиназы (ММП), влияют как на общую структуру, так и на биомеханические свойства сформированной сети, а также на сигналы, передаваемые клеткам, тем самым модулируя их ответы (Bonnans et al., 2014).
Различные типы клеток синтезируют и секретируют макромолекулы матрикса под контролем множества сигналов.Изменения в составе и структуре ВКМ, которые могут быть эндогенно опосредованы протеиназами, такими как матриксные металлопротеиназы (ММП), влияют как на общую структуру, так и на биомеханические свойства сформированной сети, а также на сигналы, передаваемые клеткам, тем самым модулируя их ответы (Bonnans et al., 2014). 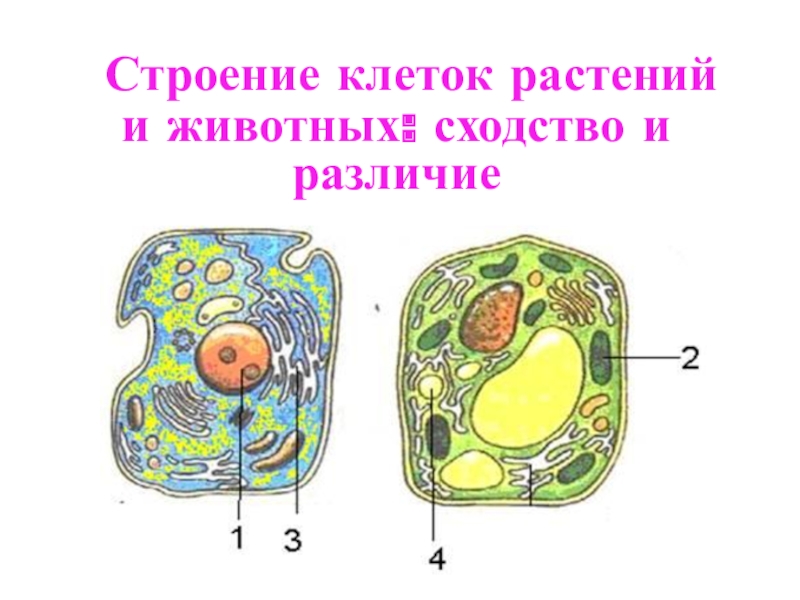 Пектины представляют собой сложную группу полисахаридов, состоящую из гомогалактуронана (HG), рамногалактуронана I (RGI), рамногалактуронана II (RGII) и ксилогалактуронана.HG, линейный полимер остатков (1,4)-α-связанной галактуроновой кислоты (GalA), является преобладающим компонентом пектинов CW листьев и имеет решающее значение для целостности тканей, пластичности стенок и адгезии клеток (Lionetti et al., 2010, 2014а; Погорелко и др., 2013). В то время как целлюлоза синтезируется на плазматической мембране (ПМ) (McNamara et al., 2015; Maleki et al., 2016), секреторный путь растений играет функциональную роль в биосинтезе КС нецеллюлозных полисахаридов, гликопротеинов и ПГ, которые синтезируется в аппарате Гольджи гликозилтрансферазами (Kim and Brandizzi, 2016; Temple et al., 2016).
Пектины представляют собой сложную группу полисахаридов, состоящую из гомогалактуронана (HG), рамногалактуронана I (RGI), рамногалактуронана II (RGII) и ксилогалактуронана.HG, линейный полимер остатков (1,4)-α-связанной галактуроновой кислоты (GalA), является преобладающим компонентом пектинов CW листьев и имеет решающее значение для целостности тканей, пластичности стенок и адгезии клеток (Lionetti et al., 2010, 2014а; Погорелко и др., 2013). В то время как целлюлоза синтезируется на плазматической мембране (ПМ) (McNamara et al., 2015; Maleki et al., 2016), секреторный путь растений играет функциональную роль в биосинтезе КС нецеллюлозных полисахаридов, гликопротеинов и ПГ, которые синтезируется в аппарате Гольджи гликозилтрансферазами (Kim and Brandizzi, 2016; Temple et al., 2016). 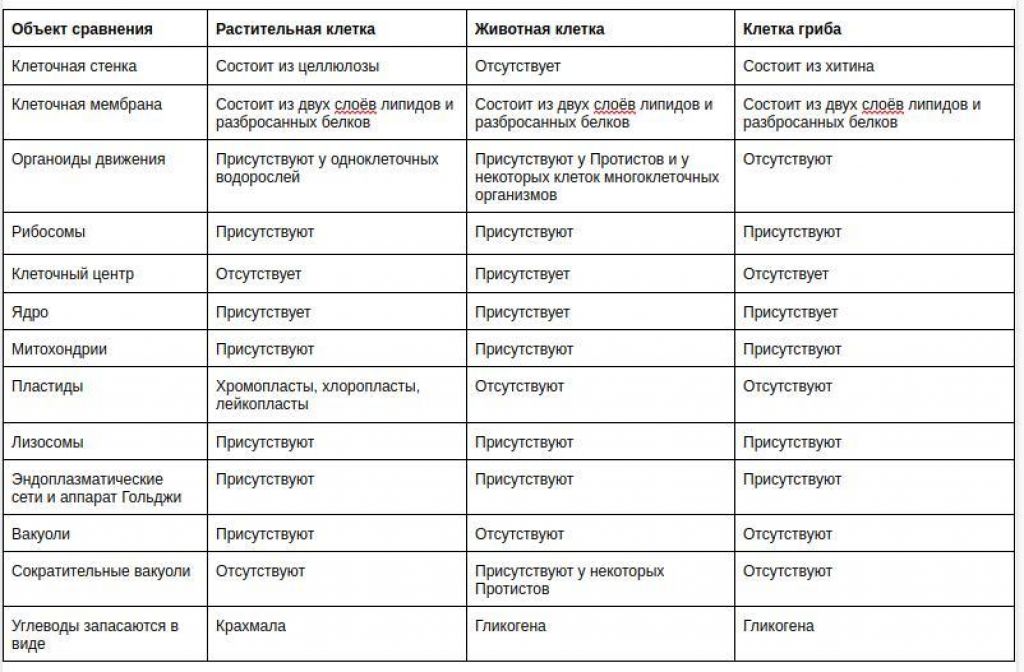 ЕСМ представляет собой огромный барьер, но разные вирусы выработали определенные стратегии для его преодоления и даже использования для проникновения в клетку. Проникновение вируса начинается с прикрепления к рецепторам клеточной поверхности и заканчивается переносом вирусного генома в цитоплазму (Димитров, 2004). После распознавания и связывания с рецепторами клеточной поверхности, которыми могут быть белки, углеводы или липиды, вирусы могут проникать в клетки посредством эндоцитоза.ECM, по-видимому, участвует в прикреплении, первых шагах проникновения вируса (рис. 1А). Большинство папилломавирусов (PV) используют HS в качестве первичных рецепторов прикрепления (Sapp and Bienkowska-Haba, 2009; DiGiuseppe et al., 2017). Количество и тип сульфатации могут влиять на прикрепление вируса и инфекцию (Knappe et al., 2007). Ламинин 5, высокомолекулярный белок внеклеточного матрикса, проявляет высокую аффинность к вирионам вируса папилломы человека типа 11 (HPV11) и, помимо HS, может опосредовать связывание с ECM (Richards et al.
ЕСМ представляет собой огромный барьер, но разные вирусы выработали определенные стратегии для его преодоления и даже использования для проникновения в клетку. Проникновение вируса начинается с прикрепления к рецепторам клеточной поверхности и заканчивается переносом вирусного генома в цитоплазму (Димитров, 2004). После распознавания и связывания с рецепторами клеточной поверхности, которыми могут быть белки, углеводы или липиды, вирусы могут проникать в клетки посредством эндоцитоза.ECM, по-видимому, участвует в прикреплении, первых шагах проникновения вируса (рис. 1А). Большинство папилломавирусов (PV) используют HS в качестве первичных рецепторов прикрепления (Sapp and Bienkowska-Haba, 2009; DiGiuseppe et al., 2017). Количество и тип сульфатации могут влиять на прикрепление вируса и инфекцию (Knappe et al., 2007). Ламинин 5, высокомолекулярный белок внеклеточного матрикса, проявляет высокую аффинность к вирионам вируса папилломы человека типа 11 (HPV11) и, помимо HS, может опосредовать связывание с ECM (Richards et al.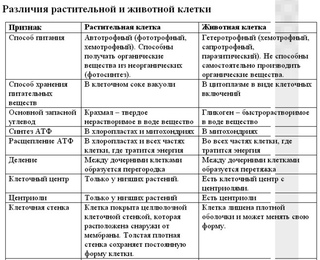 , 2014). Считается, что HS и гликосфинголипиды, а также белки, связывающие углеводы, такие как лектины, действуют как корецепторные молекулы, которые повышают эффективность проникновения вируса денге, вызывая лихорадку и геморрагические расстройства у людей и приматов (Hidari and Suzuki, 2011). Предполагается, что некоторые гликопротеины вируса гепатита С (ВГС) прикрепляются к лектинам на поверхности клетки-хозяина (клетки печени) для инфицирования (Bartenschlager and Sparacio, 2007). Начальное взаимодействие вируса простого герпеса (ВПГ) опосредовано взаимодействием с HS (Akhtar and Shukla, 2009).Также интегрины были задействованы в качестве предполагаемых рецепторов ВПГ и вируса иммунодефицита человека (ВИЧ) (Parry et al., 2005; Ding et al., 2015). Содержащие сиаловую кислоту гликаны используются многими вирусами, такими как вирусы гриппа, парагриппа, паротита, короны, норовируса, ротавируса и ДНК-опухолевых вирусов, в качестве рецепторов для проникновения в клетку (Stencel-Baerenwald et al.
, 2014). Считается, что HS и гликосфинголипиды, а также белки, связывающие углеводы, такие как лектины, действуют как корецепторные молекулы, которые повышают эффективность проникновения вируса денге, вызывая лихорадку и геморрагические расстройства у людей и приматов (Hidari and Suzuki, 2011). Предполагается, что некоторые гликопротеины вируса гепатита С (ВГС) прикрепляются к лектинам на поверхности клетки-хозяина (клетки печени) для инфицирования (Bartenschlager and Sparacio, 2007). Начальное взаимодействие вируса простого герпеса (ВПГ) опосредовано взаимодействием с HS (Akhtar and Shukla, 2009).Также интегрины были задействованы в качестве предполагаемых рецепторов ВПГ и вируса иммунодефицита человека (ВИЧ) (Parry et al., 2005; Ding et al., 2015). Содержащие сиаловую кислоту гликаны используются многими вирусами, такими как вирусы гриппа, парагриппа, паротита, короны, норовируса, ротавируса и ДНК-опухолевых вирусов, в качестве рецепторов для проникновения в клетку (Stencel-Baerenwald et al.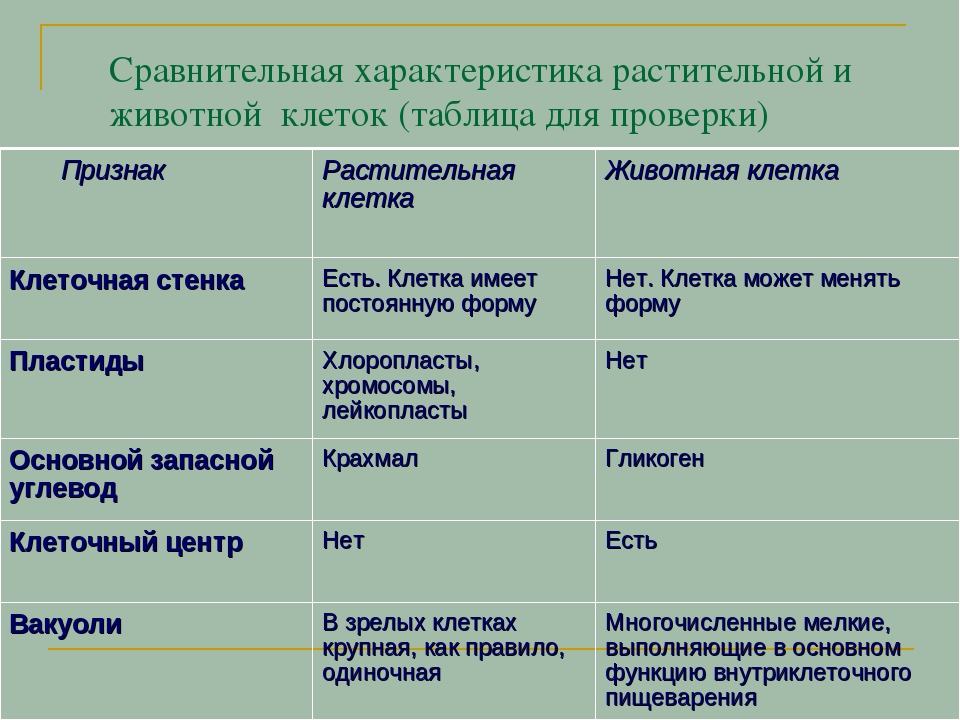 , 2014; Matrosovich). и др., 2015).
, 2014; Matrosovich). и др., 2015). 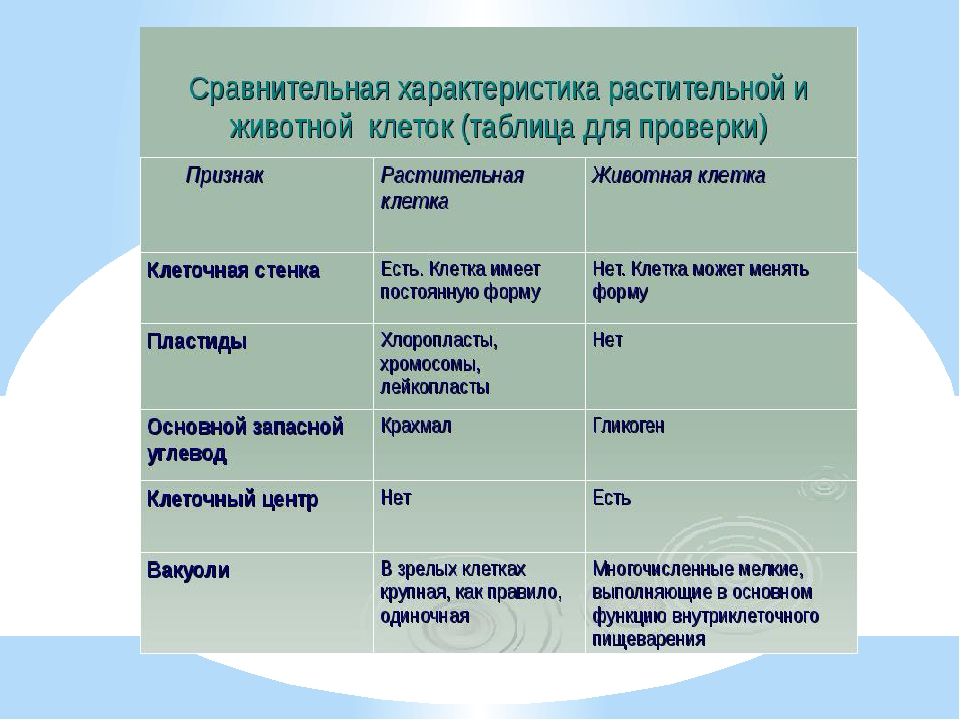 Пересечение CW представляет собой серьезную проблему для вирусов, и такой сложный процесс еще полностью не изучен. Вирусы могут проникать в клетки-хозяева и вступать в контакт с цитоплазмой только при механическом повреждении, включающем частичное разрушение КС и перфорацию ПМ, или при питании беспозвоночными переносчиками, такими как грибы, нематоды или насекомые (Hull, 2013) (рис. 1Б). .Кроме того, вирусы могут передаваться вертикально через семена или путем вегетативного размножения (Blanc, 2007). Оказавшись внутри цитоплазмы растительной клетки, вирусы освобождаются от оболочки и реплицируются по признакам, сходным с описанными для вирусов животных. Во второй половине 20 века был проведен ряд исследований для раскрытия механизма(ов) проникновения вируса в растительные клетки (Shaw, 1985). Попытки выяснить, проникают ли вирусы в клетки растений через пиноцитоз или прикрепление к специфическим рецепторам клеточной поверхности после инокуляции, остались безрезультатными.Наблюдения за палочковидными частицами вируса табачной мозаики (ВТМ) и вируса табачной погремушки (ТРВ), концы которых прикреплены к внешней поверхности КС или к протопластам после ручной инокуляции, позволили предположить, что внеклеточный сайт прикрепления будет способствовать проникновению в клетку вириона или генома РНК вируса.
Пересечение CW представляет собой серьезную проблему для вирусов, и такой сложный процесс еще полностью не изучен. Вирусы могут проникать в клетки-хозяева и вступать в контакт с цитоплазмой только при механическом повреждении, включающем частичное разрушение КС и перфорацию ПМ, или при питании беспозвоночными переносчиками, такими как грибы, нематоды или насекомые (Hull, 2013) (рис. 1Б). .Кроме того, вирусы могут передаваться вертикально через семена или путем вегетативного размножения (Blanc, 2007). Оказавшись внутри цитоплазмы растительной клетки, вирусы освобождаются от оболочки и реплицируются по признакам, сходным с описанными для вирусов животных. Во второй половине 20 века был проведен ряд исследований для раскрытия механизма(ов) проникновения вируса в растительные клетки (Shaw, 1985). Попытки выяснить, проникают ли вирусы в клетки растений через пиноцитоз или прикрепление к специфическим рецепторам клеточной поверхности после инокуляции, остались безрезультатными.Наблюдения за палочковидными частицами вируса табачной мозаики (ВТМ) и вируса табачной погремушки (ТРВ), концы которых прикреплены к внешней поверхности КС или к протопластам после ручной инокуляции, позволили предположить, что внеклеточный сайт прикрепления будет способствовать проникновению в клетку вириона или генома РНК вируса. Гаард и де Зутен, 1979). Однако не было доказано, что прикрепление вируса специфично для восприимчивых хозяев, и до сих пор не было получено окончательных доказательств проникновения вируса при прикреплении. Насколько нам известно, растительные вирусы не могут активно разрушать CW, и хотя у растений наблюдались пути, подобные эндоцитозу (Kitakura et al., 2011), вирусы не могут ни использовать эндоцитозный путь для проникновения в клетки, окруженные КС, ни покидать их путем почкования. Отсутствие липопротеиновой оболочки у большинства растительных вирусов, вероятно, представляет собой адаптацию к эволюции КС, в отличие от оболочечных вирусов, проникающих в клетки животных без КС. У нескольких родов оболочечных вирусов растений, то есть у Tospovirus, Cytorhabdovirus, Nucleorhabdovirus и Emaravirus , оболочка способствует передаче вирионов, опосредованной вектором, но не требуется для проникновения в клетку и межклеточного движения (Adkins, 2000; Jackson et al., 2005; Альборноз и др.
Гаард и де Зутен, 1979). Однако не было доказано, что прикрепление вируса специфично для восприимчивых хозяев, и до сих пор не было получено окончательных доказательств проникновения вируса при прикреплении. Насколько нам известно, растительные вирусы не могут активно разрушать CW, и хотя у растений наблюдались пути, подобные эндоцитозу (Kitakura et al., 2011), вирусы не могут ни использовать эндоцитозный путь для проникновения в клетки, окруженные КС, ни покидать их путем почкования. Отсутствие липопротеиновой оболочки у большинства растительных вирусов, вероятно, представляет собой адаптацию к эволюции КС, в отличие от оболочечных вирусов, проникающих в клетки животных без КС. У нескольких родов оболочечных вирусов растений, то есть у Tospovirus, Cytorhabdovirus, Nucleorhabdovirus и Emaravirus , оболочка способствует передаче вирионов, опосредованной вектором, но не требуется для проникновения в клетку и межклеточного движения (Adkins, 2000; Jackson et al., 2005; Альборноз и др.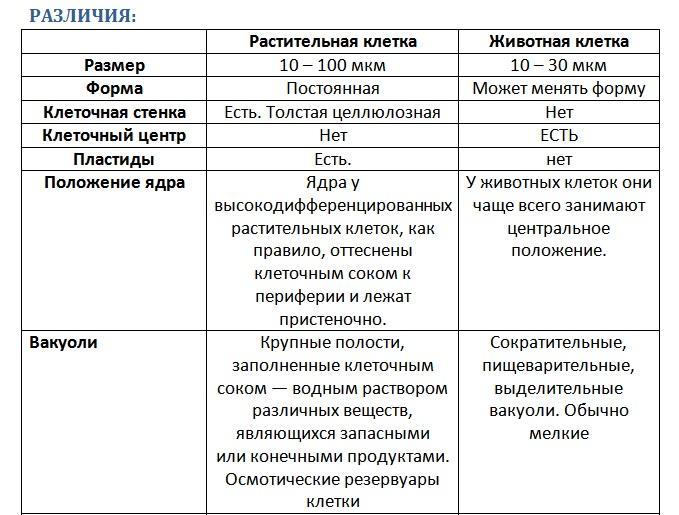 , 2016). Интересно, что вирус хлореллы paramecium bursaria (PBCV-1) может активно проникать в КС Chlorella spp., одноклеточных зеленых водорослей, имеющих схожую клеточную структуру с высшими растениями. После ферментативного расщепления CW PBCV-1 сливается с клеточной мембраной через липидную двухслойную мембрану под наружным гликопротеиновым капсидом и транслоцирует свой геном в водоросль-хозяина (Van Etten, 2003).
, 2016). Интересно, что вирус хлореллы paramecium bursaria (PBCV-1) может активно проникать в КС Chlorella spp., одноклеточных зеленых водорослей, имеющих схожую клеточную структуру с высшими растениями. После ферментативного расщепления CW PBCV-1 сливается с клеточной мембраной через липидную двухслойную мембрану под наружным гликопротеиновым капсидом и транслоцирует свой геном в водоросль-хозяина (Van Etten, 2003).  2А). Способность использовать и манипулировать межклеточным контактом способствует успеху вирусных инфекций. Многие вирусы, в том числе ВПГ, ВИЧ и Т-лимфотропный вирус человека (HTLV), могут образовывать так называемые вирусологические синапсы: вызываемые вирусом специализированные области контакта между клетками, которые способствуют межклеточной передаче (Vasiliver-Shamis et al., 2008; Абаитуа и др., 2013). Вирусные инфекции могут активировать эндогенные молекулы клеточной адгезии (CAM), такие как белок ICAM-1, а также другие компоненты внеклеточного матрикса (Nakachi et al., 2011; Gross and Thoma-Kress, 2016). Некоторые вирусы также могут продуцировать свои собственные белки адгезии. Различные вирусы экспрессируют гликопротеин Env, который может действовать как молекула вирусной адгезии (VAM), имитируя поведение CAM (Mothes et al., 2010).
2А). Способность использовать и манипулировать межклеточным контактом способствует успеху вирусных инфекций. Многие вирусы, в том числе ВПГ, ВИЧ и Т-лимфотропный вирус человека (HTLV), могут образовывать так называемые вирусологические синапсы: вызываемые вирусом специализированные области контакта между клетками, которые способствуют межклеточной передаче (Vasiliver-Shamis et al., 2008; Абаитуа и др., 2013). Вирусные инфекции могут активировать эндогенные молекулы клеточной адгезии (CAM), такие как белок ICAM-1, а также другие компоненты внеклеточного матрикса (Nakachi et al., 2011; Gross and Thoma-Kress, 2016). Некоторые вирусы также могут продуцировать свои собственные белки адгезии. Различные вирусы экспрессируют гликопротеин Env, который может действовать как молекула вирусной адгезии (VAM), имитируя поведение CAM (Mothes et al., 2010). 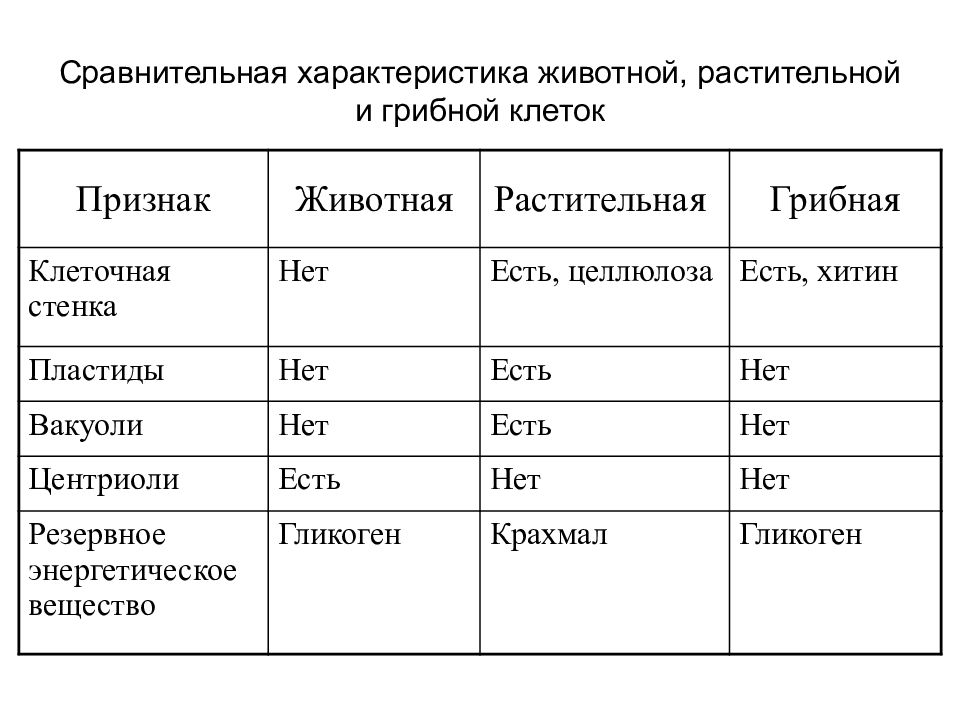 Оболочка ВИЧ-1 на поверхности инфицированной клетки связывается с α4β7 на клетке-мишени. Гликопротеины оболочки ВИЧ-1 (Env) экспрессируются на инфицированных клетках PM и взаимодействуют с рецепторами CD4 и CCR5 или CXCR4 на клетке-мишени. Молекулы адгезии, молекула межклеточной адгезии 1 (ICAM1) и антиген 1, ассоциированный с функцией лимфоцитов (LFA1), взаимодействуют с интегрином для стабилизации клеточного конъюгата. (B) Схематическое изображение согласованного действия некоторых эндогенных и экзогенных факторов, способствующих перемещению вируса при БП. После проникновения вируса растения снижают предел исключения размера PD за счет локального отложения каллозы в области шейки. Вирусная инфекция запускает активность β 1-3 глюканаз, которые способствуют закрытию поры PD в сотрудничестве с PME и MP. Сверхэкспрессия PMEI или замалчивание PME задерживает распространение вируса от клетки к клетке, противодействуя этим процессам и ограничивая PME/MP-опосредованное расширение пор PD.
Оболочка ВИЧ-1 на поверхности инфицированной клетки связывается с α4β7 на клетке-мишени. Гликопротеины оболочки ВИЧ-1 (Env) экспрессируются на инфицированных клетках PM и взаимодействуют с рецепторами CD4 и CCR5 или CXCR4 на клетке-мишени. Молекулы адгезии, молекула межклеточной адгезии 1 (ICAM1) и антиген 1, ассоциированный с функцией лимфоцитов (LFA1), взаимодействуют с интегрином для стабилизации клеточного конъюгата. (B) Схематическое изображение согласованного действия некоторых эндогенных и экзогенных факторов, способствующих перемещению вируса при БП. После проникновения вируса растения снижают предел исключения размера PD за счет локального отложения каллозы в области шейки. Вирусная инфекция запускает активность β 1-3 глюканаз, которые способствуют закрытию поры PD в сотрудничестве с PME и MP. Сверхэкспрессия PMEI или замалчивание PME задерживает распространение вируса от клетки к клетке, противодействуя этим процессам и ограничивая PME/MP-опосредованное расширение пор PD.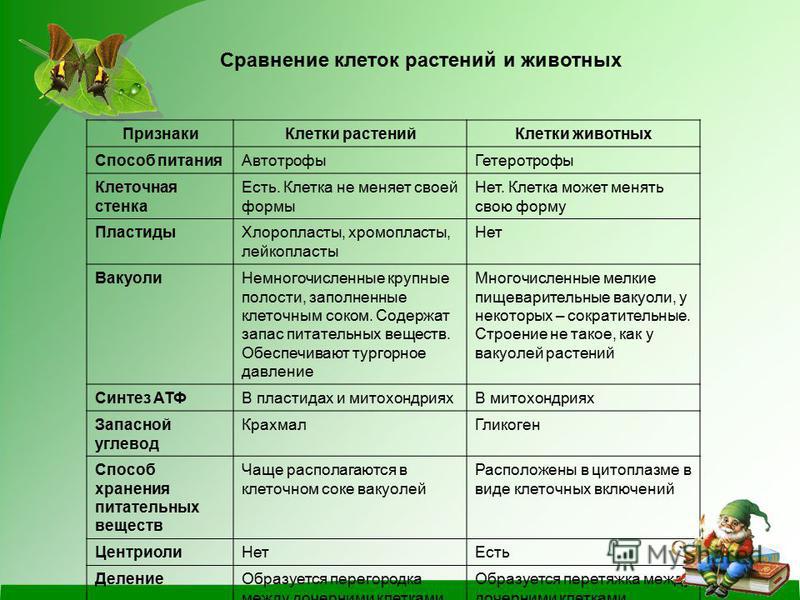 ПМ, плазматическая мембрана; ER, эндоплазматический ретикулум; CW, клеточная стенка; ПД, плазмодесмы; CP, белок оболочки; МП, двигательный белок; ВТМ, вирус табачной мозаики.
ПМ, плазматическая мембрана; ER, эндоплазматический ретикулум; CW, клеточная стенка; ПД, плазмодесмы; CP, белок оболочки; МП, двигательный белок; ВТМ, вирус табачной мозаики.  В любом случае они кодируют один или несколько транспортных белков (MPs), обеспечивающих PD-затвор через молекулярные механизмы, которые еще полностью не изучены, и с помощью цитоскелета и/или эндомембран хозяина, которые облегчают перемещение вируса по симпластным соединениям (Lucas, 2006; Харрис и др., 2010; Гаррис и Дин, 2011).
В любом случае они кодируют один или несколько транспортных белков (MPs), обеспечивающих PD-затвор через молекулярные механизмы, которые еще полностью не изучены, и с помощью цитоскелета и/или эндомембран хозяина, которые облегчают перемещение вируса по симпластным соединениям (Lucas, 2006; Харрис и др., 2010; Гаррис и Дин, 2011). 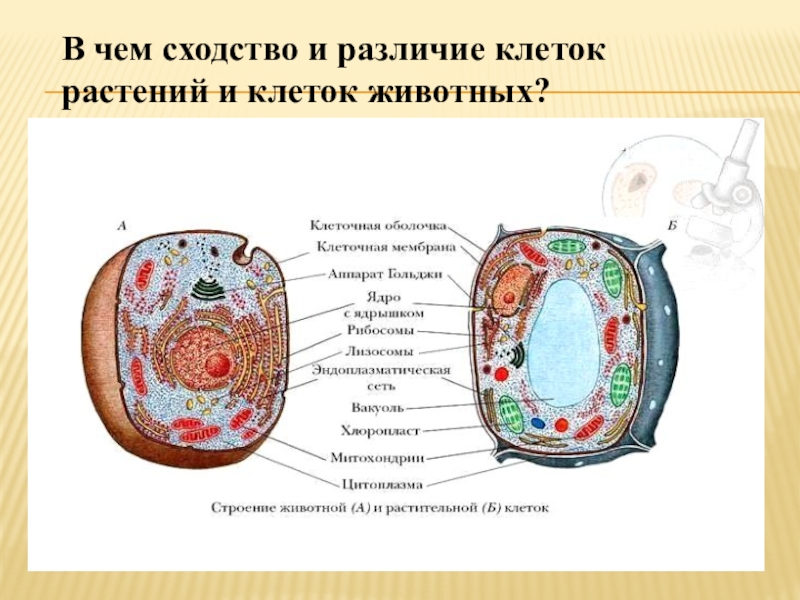 ММП, также называемые матриксинами, являются кальций-зависимыми цинксодержащими эндопептидазами, способными расщеплять белки ВКМ и обрабатывать биологически активные молекулы при патологических состояниях, таких как воспаление и повреждение тканей после воспалительных сигналов.Они могут опосредовать изменения в ECM и влиять на поведение иммунных и провоспалительных клеток (Liu et al., 2006; Wells et al., 2015; Peeters et al., 2017). ММП также влияют на тяжесть заболевания у младенцев с инфекцией респираторно-синцитиального вируса (РСВ). MMP и их ингибиторы вносят вклад в баланс между деградацией и отложением ECM, координируя заживление тканей (Schuurhof et al., 2012). Иммунные ответы возникают в контексте опосредованных интегрином адгезивных взаимодействий с ЕСМ. Например, при гриппозной инфекции интегрин α1β1, который связывает коллагены I и IV, опосредует удержание Т-клеток памяти в легких после элиминации вируса, что важно для вторичного иммунитета (Ray et al., 2004).
ММП, также называемые матриксинами, являются кальций-зависимыми цинксодержащими эндопептидазами, способными расщеплять белки ВКМ и обрабатывать биологически активные молекулы при патологических состояниях, таких как воспаление и повреждение тканей после воспалительных сигналов.Они могут опосредовать изменения в ECM и влиять на поведение иммунных и провоспалительных клеток (Liu et al., 2006; Wells et al., 2015; Peeters et al., 2017). ММП также влияют на тяжесть заболевания у младенцев с инфекцией респираторно-синцитиального вируса (РСВ). MMP и их ингибиторы вносят вклад в баланс между деградацией и отложением ECM, координируя заживление тканей (Schuurhof et al., 2012). Иммунные ответы возникают в контексте опосредованных интегрином адгезивных взаимодействий с ЕСМ. Например, при гриппозной инфекции интегрин α1β1, который связывает коллагены I и IV, опосредует удержание Т-клеток памяти в легких после элиминации вируса, что важно для вторичного иммунитета (Ray et al., 2004). 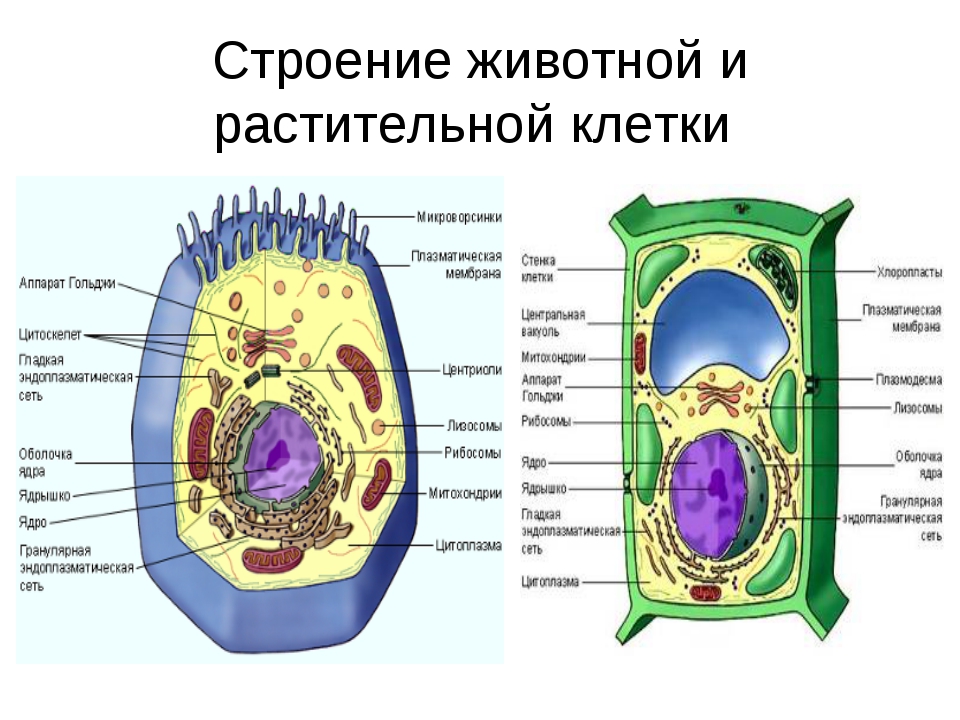 Каллоза (β-1,3-глюкан) – полисахарид, синтезируемый в КС каллозсинтазами и расщепляемый β-1,3-глюканазами (Завалиев и др., 2011), накапливается вокруг шейки БП в виде воротника и ее оборота. контролирует транспортную способность ФД (Fitzgibbon et al., 2010; Guseman et al., 2010; Vaten et al., 2011; Han et al., 2014). ECM растений может противодействовать PD-гейтирующей функции вирусных MP за счет локализованного апопластического накопления каллозы вокруг шейки PD и быстрого и эффективного снижения предела исключения размера PD (De Storme and Geelen, 2014).Доказательства, такие как взаимодействие МЧ ВТМ с белком ANK, содержащим анкириновый повтор, и вируса картофеля X TGB2 с белками, ассоциированными с β-1,3-глюканазой, позволяют предположить, что некоторые вирусные МЧ развили контрреакцию. стратегия уменьшения накопления каллозы и PD ворот для межклеточного движения (Fridborg et al., 2003; Ueki et al., 2010; Zavaliev et al., 2011). В более общем плане демонстрация того, что успех вирусной инфекции и накопление каллозы вокруг БП обратно пропорциональны (Iglesias and Meins, 2000; Zavaliev et al.
Каллоза (β-1,3-глюкан) – полисахарид, синтезируемый в КС каллозсинтазами и расщепляемый β-1,3-глюканазами (Завалиев и др., 2011), накапливается вокруг шейки БП в виде воротника и ее оборота. контролирует транспортную способность ФД (Fitzgibbon et al., 2010; Guseman et al., 2010; Vaten et al., 2011; Han et al., 2014). ECM растений может противодействовать PD-гейтирующей функции вирусных MP за счет локализованного апопластического накопления каллозы вокруг шейки PD и быстрого и эффективного снижения предела исключения размера PD (De Storme and Geelen, 2014).Доказательства, такие как взаимодействие МЧ ВТМ с белком ANK, содержащим анкириновый повтор, и вируса картофеля X TGB2 с белками, ассоциированными с β-1,3-глюканазой, позволяют предположить, что некоторые вирусные МЧ развили контрреакцию. стратегия уменьшения накопления каллозы и PD ворот для межклеточного движения (Fridborg et al., 2003; Ueki et al., 2010; Zavaliev et al., 2011). В более общем плане демонстрация того, что успех вирусной инфекции и накопление каллозы вокруг БП обратно пропорциональны (Iglesias and Meins, 2000; Zavaliev et al. , 2013) указывает на то, что накопление каллозы при БП является ранним барьером, который ECM активирует, чтобы блокировать проникновение вируса в соседние клетки и распространение в хозяине (Epel, 2009).
, 2013) указывает на то, что накопление каллозы при БП является ранним барьером, который ECM активирует, чтобы блокировать проникновение вируса в соседние клетки и распространение в хозяине (Epel, 2009).  , 2007; Peaucelle et al., 2011; Chebli and Geitmann, 2017).
, 2007; Peaucelle et al., 2011; Chebli and Geitmann, 2017).  Ключевым ограничением является нацеливание вируса на опухоль. ECM предотвращает не только распространение вируса, но и попадание вируса в опухоль. Таким образом, в настоящее время активно исследуется вооружение вирусов ферментами, разлагающими ВКМ, и распространение вирусной пермиссивности на неопухолевые стромальные клетки для улучшения распространения вируса и постоянного нацеливания вируса.Вирус болезни Ньюкасла (NDV) представляет собой парамиксовирус птиц, обладающий избирательным онколитическим действием на опухолевые клетки в культуре и на животных моделях (Матвеева и др., 2015). ECM ограничивает распространение NDV и других вирусов, но удаление тканевого коллагена и гепарансульфата с помощью обработки коллагеназой и гепариназой до инфицирования увеличивает распространение вируса (Yaacov et al., 2012). Коллаген, HA и HS также влияют на онколитическую активность аденовируса и HSV-1 (McKee et al., 2006; Guedan et al., 2010; Ватанабе, 2010).
Ключевым ограничением является нацеливание вируса на опухоль. ECM предотвращает не только распространение вируса, но и попадание вируса в опухоль. Таким образом, в настоящее время активно исследуется вооружение вирусов ферментами, разлагающими ВКМ, и распространение вирусной пермиссивности на неопухолевые стромальные клетки для улучшения распространения вируса и постоянного нацеливания вируса.Вирус болезни Ньюкасла (NDV) представляет собой парамиксовирус птиц, обладающий избирательным онколитическим действием на опухолевые клетки в культуре и на животных моделях (Матвеева и др., 2015). ECM ограничивает распространение NDV и других вирусов, но удаление тканевого коллагена и гепарансульфата с помощью обработки коллагеназой и гепариназой до инфицирования увеличивает распространение вируса (Yaacov et al., 2012). Коллаген, HA и HS также влияют на онколитическую активность аденовируса и HSV-1 (McKee et al., 2006; Guedan et al., 2010; Ватанабе, 2010).  Одной из возможностей может быть разработка растения CW, более устойчивого к насекомым-переносчикам вируса. Фактически слюнные выделения тлей содержат некоторые ферменты, разлагающие КС, такие как полигалактуроназы и пектинметилэстеразы, которые насекомые используют для проникновения стилета (Dreyer and Campbell, 1984).ХО, более устойчивый к ферментативной деградации, может уменьшить передачу вируса. Доказательства того, что PMEI ограничивают распространение вируса, позволяют предположить, что этот класс ингибиторов также можно использовать в программах селекции, направленных на получение сортов растений, менее восприимчивых к вирусным заболеваниям. Интересно, что пектиновые олигогалактурониды и фрагменты ГК могут восприниматься как молекулярные паттерны, ассоциированные с повреждением (DAMP), при повреждении тканей или патогенной инфекции, активирующие врожденную иммунную систему растений и животных соответственно (Ferrari et al., 2013; Каваси и др., 2017). Следует изучить потенциальную роль этих фрагментов в ответ на вирусную инфекцию и соответствующее биотехнологическое применение.
Одной из возможностей может быть разработка растения CW, более устойчивого к насекомым-переносчикам вируса. Фактически слюнные выделения тлей содержат некоторые ферменты, разлагающие КС, такие как полигалактуроназы и пектинметилэстеразы, которые насекомые используют для проникновения стилета (Dreyer and Campbell, 1984).ХО, более устойчивый к ферментативной деградации, может уменьшить передачу вируса. Доказательства того, что PMEI ограничивают распространение вируса, позволяют предположить, что этот класс ингибиторов также можно использовать в программах селекции, направленных на получение сортов растений, менее восприимчивых к вирусным заболеваниям. Интересно, что пектиновые олигогалактурониды и фрагменты ГК могут восприниматься как молекулярные паттерны, ассоциированные с повреждением (DAMP), при повреждении тканей или патогенной инфекции, активирующие врожденную иммунную систему растений и животных соответственно (Ferrari et al., 2013; Каваси и др., 2017). Следует изучить потенциальную роль этих фрагментов в ответ на вирусную инфекцию и соответствующее биотехнологическое применение.
 Поразительно, что в обеих матрицах вирусы могут использовать эти соединения для проникновения в клетку-хозяина, хотя и с разными стратегиями. Необходимы дальнейшие усилия, чтобы понять роль специфических растительных полисахаридов CW вокруг PD в перемещении вирусной клетки к клетке, а также выяснить возможную роль CW во проникновении вируса. Эти знания могут предоставить новые цели для генетического улучшения устойчивости растений к вирусам.
Поразительно, что в обеих матрицах вирусы могут использовать эти соединения для проникновения в клетку-хозяина, хотя и с разными стратегиями. Необходимы дальнейшие усилия, чтобы понять роль специфических растительных полисахаридов CW вокруг PD в перемещении вирусной клетки к клетке, а также выяснить возможную роль CW во проникновении вируса. Эти знания могут предоставить новые цели для генетического улучшения устойчивости растений к вирусам. 
 FEBS J. 276, 7228–7236. doi: 10.1111/j.1742-4658.2009.07402.x
FEBS J. 276, 7228–7236. doi: 10.1111/j.1742-4658.2009.07402.x 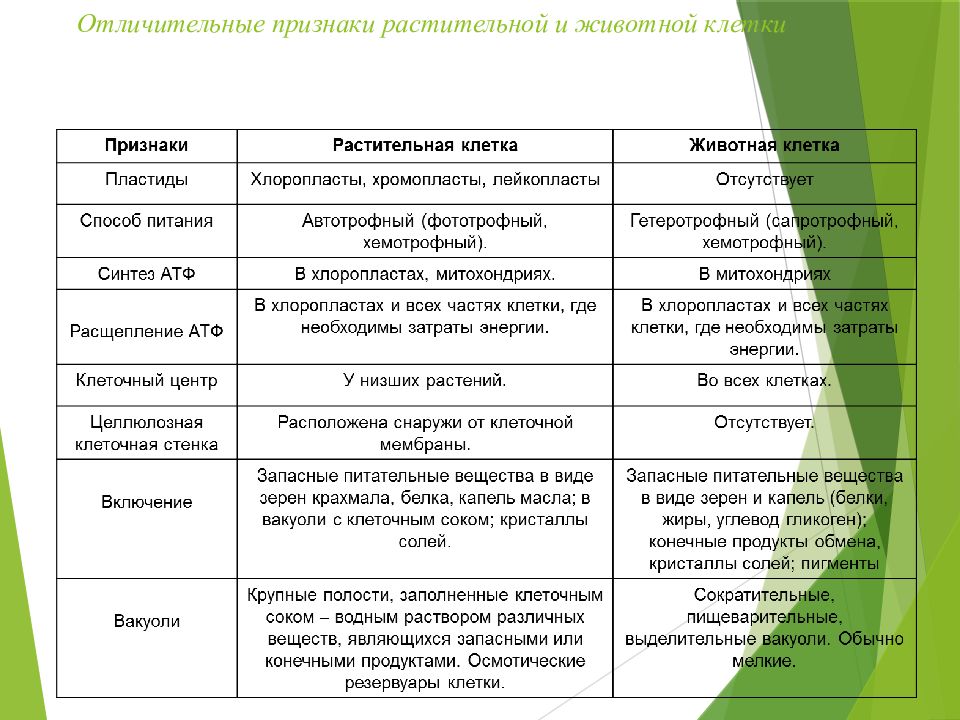 (1990). Гликопротеиновый ингибитор пектинметилэстеразы в плодах киви ( Actinidia chinensis ). евро. Дж. Биохим. 193, 183–187. doi: 10.1111/j.1432-1033.1990.tb19321.x
(1990). Гликопротеиновый ингибитор пектинметилэстеразы в плодах киви ( Actinidia chinensis ). евро. Дж. Биохим. 193, 183–187. doi: 10.1111/j.1432-1033.1990.tb19321.x  Хайнлайн (Берлин: Springer), 1–28. дои: 10.1007/7089_2006_099
Хайнлайн (Берлин: Springer), 1–28. дои: 10.1007/7089_2006_099  евро. Дж. Плант Патол. 141, 339–347. doi: 10.1007/s10658-014-0546-y
евро. Дж. Плант Патол. 141, 339–347. doi: 10.1007/s10658-014-0546-y  5.913
5.913  Дж. Эквайр. Иммунный дефицит. Синдр. 69, 509–518. doi: 10.1097/QAI.0000000000000676
Дж. Эквайр. Иммунный дефицит. Синдр. 69, 509–518. doi: 10.1097/QAI.0000000000000676  Фронт. Растениевод. 4:49. doi: 10.3389/fpls.2013.00049
Фронт. Растениевод. 4:49. doi: 10.3389/fpls.2013.00049  Мол. тер. 18, 1275–1283. doi: 10.1038/mt.2010.79
Мол. тер. 18, 1275–1283. doi: 10.1038/mt.2010.79  Дев. Ячейка 28, 132–146. doi: 10.1016/j.devcel.2013.12.008
Дев. Ячейка 28, 132–146. doi: 10.1016/j.devcel.2013.12.008 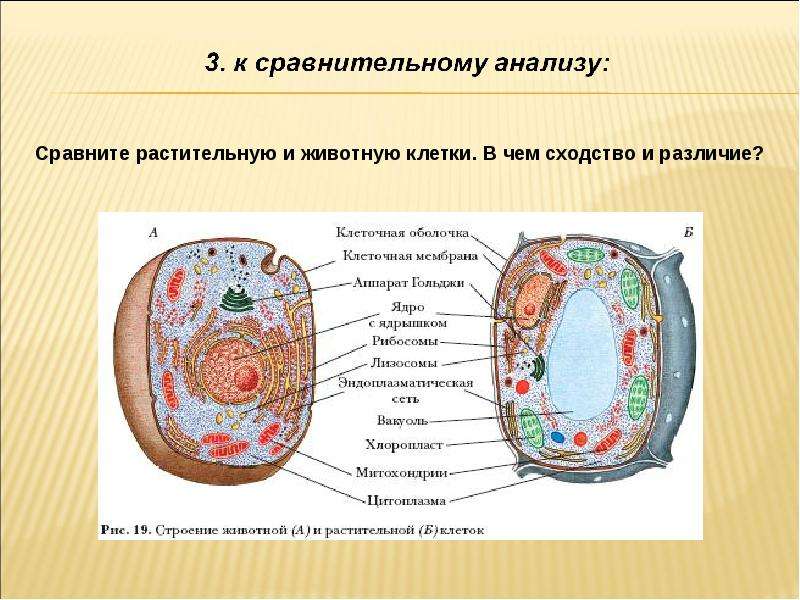 Нац. Преподобный Мол. Клеточная биол. 15, 802–812. doi: 10.1038/nrm3896
Нац. Преподобный Мол. Клеточная биол. 15, 802–812. doi: 10.1038/nrm3896  101, 128–138. doi: 10.1016/j.fct.2017.01.012
101, 128–138. doi: 10.1016/j.fct.2017.01.012 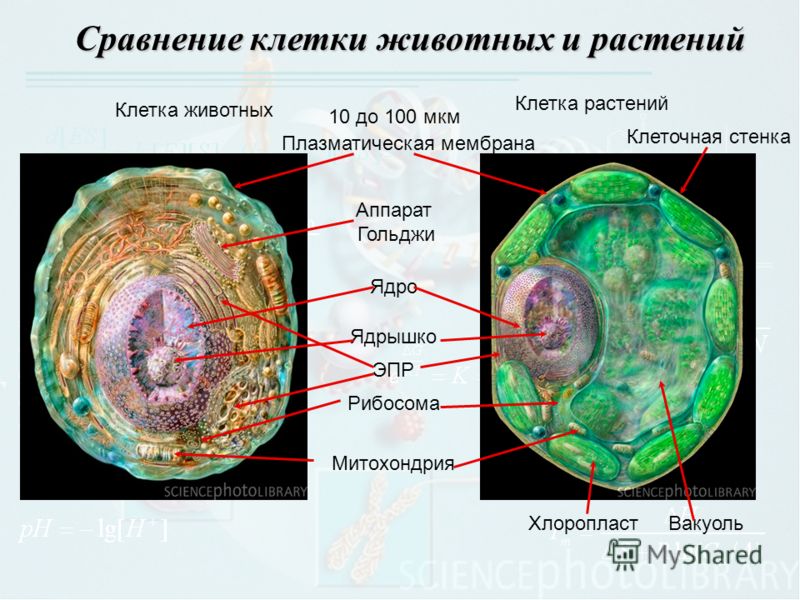 Биол. хим. 282, 27913–27922. doi: 10.1074/jbc.M705127200
Биол. хим. 282, 27913–27922. doi: 10.1074/jbc.M705127200  jplph.2012.05.006
jplph.2012.05.006 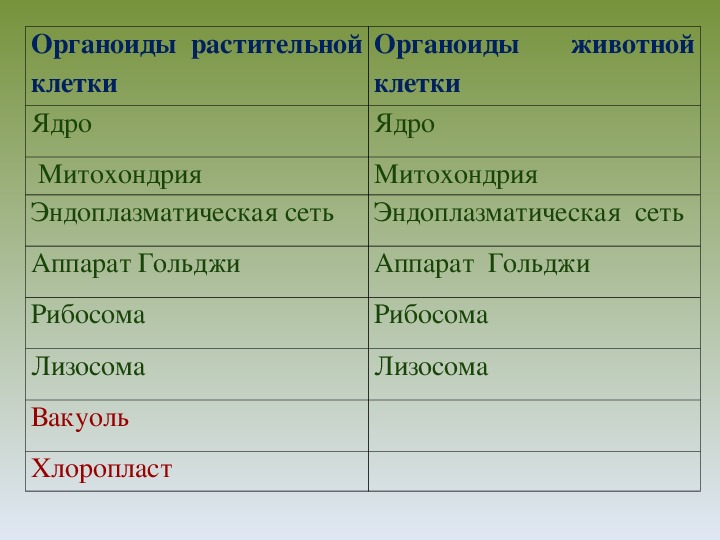 Завод физиол. 173, 1844–1863 гг. doi: 10.1104/pp.16.01185
Завод физиол. 173, 1844–1863 гг. doi: 10.1104/pp.16.01185  Кардиол. 22(Прил. Б), 25Б–30Б. doi: 10.1016/S0828-282X(06)70983-7
Кардиол. 22(Прил. Б), 25Б–30Б. doi: 10.1016/S0828-282X(06)70983-7 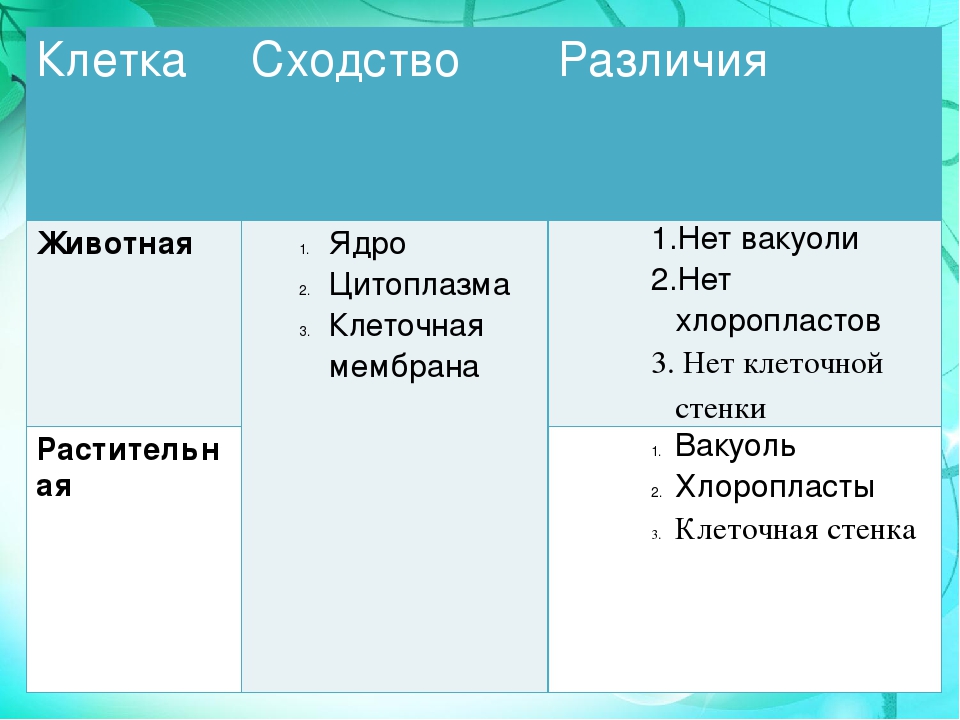 1146/annurev-biochem-060614-033930
1146/annurev-biochem-060614-033930 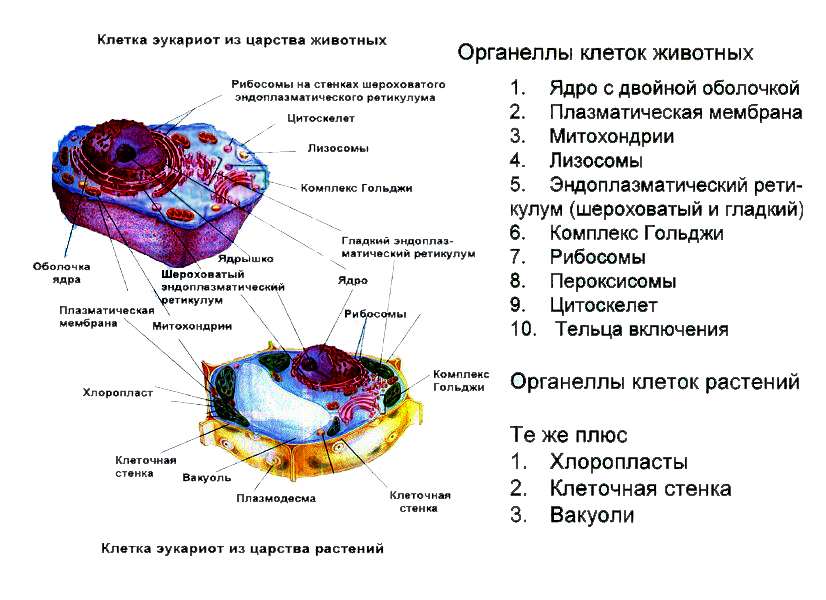 , и Нокс, Дж. П. (2000). Пространственная регуляция пектиновых полисахаридов по отношению к полям ямок в клеточных стенках околоплодника плодов томата. Завод физиол. 122, 775–781. doi: 10.1104/pp.122.3.775
, и Нокс, Дж. П. (2000). Пространственная регуляция пектиновых полисахаридов по отношению к полям ямок в клеточных стенках околоплодника плодов томата. Завод физиол. 122, 775–781. doi: 10.1104/pp.122.3.775  , Jorsal, A., Parving, H.H., Tarnow, L., et al. (2017). Металлопротеиназы матрикса плазмы связаны с сердечно-сосудистыми заболеваниями и общей смертностью у пациентов с диабетом 1 типа: 12-летнее последующее исследование. Кардиоваскл. Диабетол. 16, 55. doi: 10.1186/s12933-017-0539-1
, Jorsal, A., Parving, H.H., Tarnow, L., et al. (2017). Металлопротеиназы матрикса плазмы связаны с сердечно-сосудистыми заболеваниями и общей смертностью у пациентов с диабетом 1 типа: 12-летнее последующее исследование. Кардиоваскл. Диабетол. 16, 55. doi: 10.1186/s12933-017-0539-1 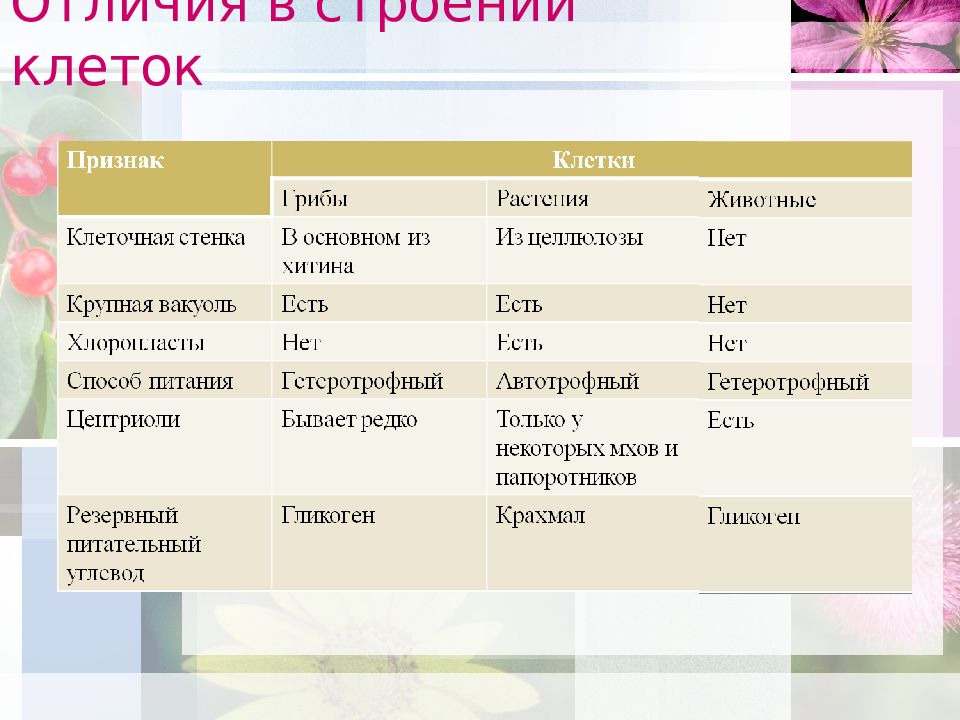 J., Franki, S.N., Pierce, R.H., Dimitrova, S., Koteliansky, V., Sprague, A.G., et al. (2004). Связывающий коллаген альфа1бета1 интегрин VLA-1 регулирует опосредованную CD8 Т-клетками иммунную защиту от гетерологичной инфекции гриппа. Иммунитет 20, 167–179. doi: 10.1016/S1074-7613(04)00021-4
J., Franki, S.N., Pierce, R.H., Dimitrova, S., Koteliansky, V., Sprague, A.G., et al. (2004). Связывающий коллаген альфа1бета1 интегрин VLA-1 регулирует опосредованную CD8 Т-клетками иммунную защиту от гетерологичной инфекции гриппа. Иммунитет 20, 167–179. doi: 10.1016/S1074-7613(04)00021-4 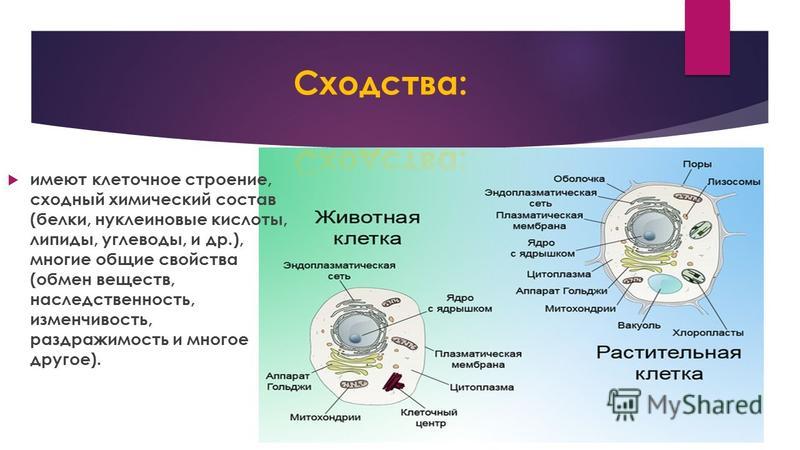 , Bont, L., Hodemaekers, H.M., de Klerk, A., de Groot, H., Hofland, R.W., et al. (2012). Белки, участвующие в динамике внеклеточного матрикса, связаны с тяжестью респираторно-синцитиального вирусного заболевания. евро. Дыхание J. 39, 1475–1481. дои: 10.1183/0
, Bont, L., Hodemaekers, H.M., de Klerk, A., de Groot, H., Hofland, R.W., et al. (2012). Белки, участвующие в динамике внеклеточного матрикса, связаны с тяжестью респираторно-синцитиального вирусного заболевания. евро. Дыхание J. 39, 1475–1481. дои: 10.1183/0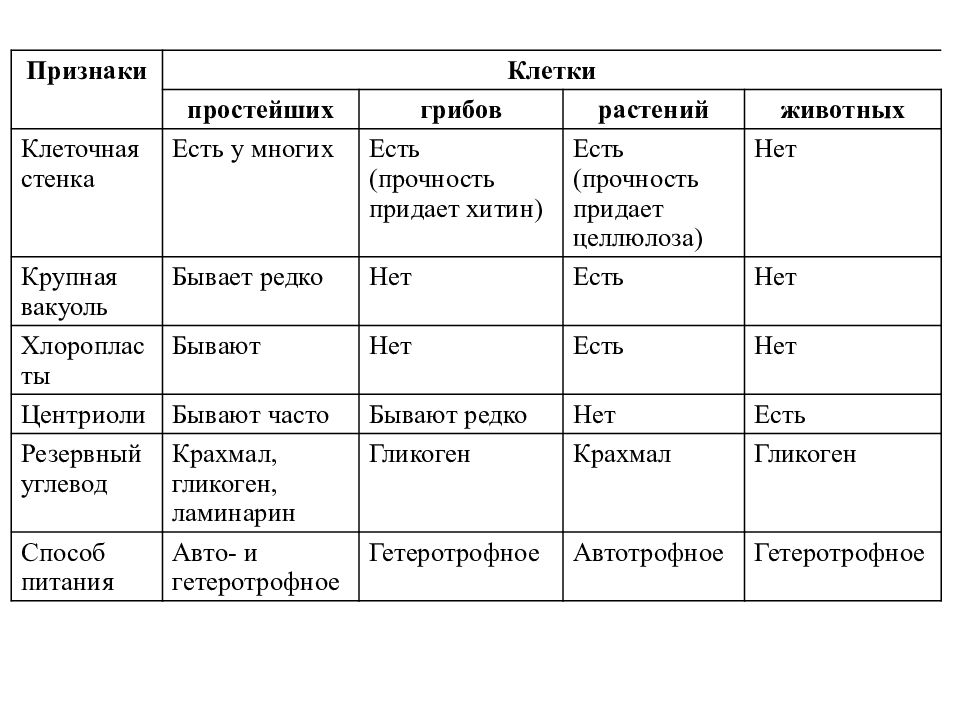 Гликобиология 26, 913–925. doi: 10.1093/гликоб/cww054
Гликобиология 26, 913–925. doi: 10.1093/гликоб/cww054  , Yadav, S.R., et al. (2011). Биосинтез каллозы регулирует симпластический транспорт во время развития корня. Дев. Мобильный 21, 1144–1155. doi: 10.1016/j.devcel.2011.10.006
, Yadav, S.R., et al. (2011). Биосинтез каллозы регулирует симпластический транспорт во время развития корня. Дев. Мобильный 21, 1144–1155. doi: 10.1016/j.devcel.2011.10.006  Дж. Генерал Вирол. 93 (часть 8), 1664–1672. doi: 10.1099/vir.0.043281-0
Дж. Генерал Вирол. 93 (часть 8), 1664–1672. doi: 10.1099/vir.0.043281-0  1007/s00709-010-0247-0
1007/s00709-010-0247-0  Однако одно из существенных различий между растительной и животной клеткой заключается в том, что растения являются автотрофными, а животные гетеротрофными в зависимости от производства пищи. Автотрофный означает независимый (auto = self), что указывает на то, что этим живым организмам не нужно зависеть от какого-либо другого организма в отношении пищевых веществ, которые они могут производить самостоятельно. С другой стороны, гетеротрофы означают зависимые организмы, которым необходимо получать пищу от других организмов, таких как растительные клетки. Ниже приводится более подробное объяснение, описывающее основные различия между растительными и животными клетками: —
Однако одно из существенных различий между растительной и животной клеткой заключается в том, что растения являются автотрофными, а животные гетеротрофными в зависимости от производства пищи. Автотрофный означает независимый (auto = self), что указывает на то, что этим живым организмам не нужно зависеть от какого-либо другого организма в отношении пищевых веществ, которые они могут производить самостоятельно. С другой стороны, гетеротрофы означают зависимые организмы, которым необходимо получать пищу от других организмов, таких как растительные клетки. Ниже приводится более подробное объяснение, описывающее основные различия между растительными и животными клетками: — 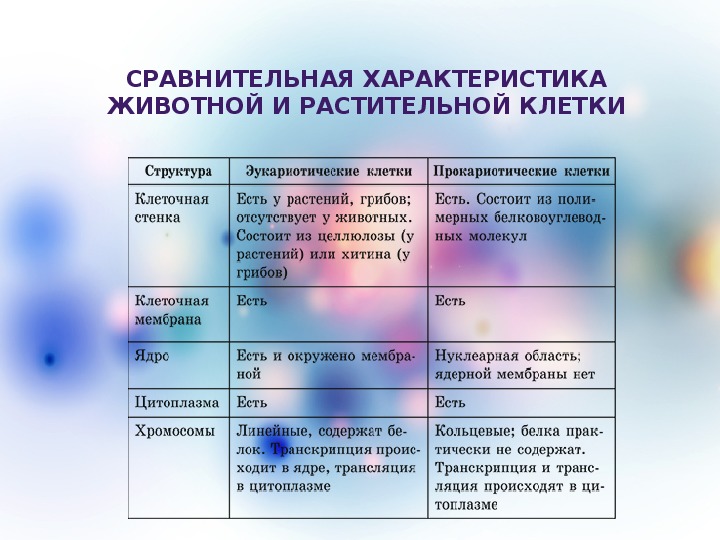
 Их нет в клетках животных.
Их нет в клетках животных.  Дополнительные мембраны внутри хлоропластов содержат структуры, непосредственно осуществляющие фотосинтез.
Дополнительные мембраны внутри хлоропластов содержат структуры, непосредственно осуществляющие фотосинтез.  Вакуоли могут занимать до 90% объема клетки и иметь одну мембрану. Их основная функция заключается в заполнении пространства в клетке, но они также могут выполнять пищеварительные функции, подобные лизосомам (которые также присутствуют в растительных клетках). Вакуоли содержат ряд ферментов, выполняющих различные функции, а их внутреннее пространство может использоваться в качестве хранилища питательных веществ или, как уже упоминалось, служить местом для деградации нежелательных веществ.
Вакуоли могут занимать до 90% объема клетки и иметь одну мембрану. Их основная функция заключается в заполнении пространства в клетке, но они также могут выполнять пищеварительные функции, подобные лизосомам (которые также присутствуют в растительных клетках). Вакуоли содержат ряд ферментов, выполняющих различные функции, а их внутреннее пространство может использоваться в качестве хранилища питательных веществ или, как уже упоминалось, служить местом для деградации нежелательных веществ.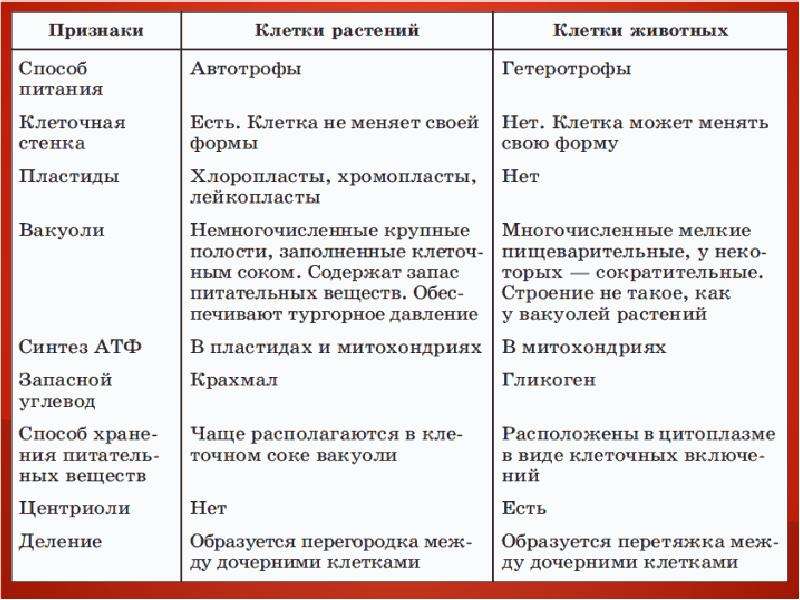 Хлорофилл также делает хлоропласты зелеными.
Хлорофилл также делает хлоропласты зелеными. 

 Большие клетки служили домами для маленьких клеток, а маленькие клетки снабжали большие клетки пищей или энергией. Считается, что эти маленькие клетки были предками того, что мы сейчас знаем как хлоропласты и митохондрии.Мы верим в это, потому что хлоропласты и митохондрии содержат свою собственную ДНК, уникальную от ДНК крупных клеток.
Большие клетки служили домами для маленьких клеток, а маленькие клетки снабжали большие клетки пищей или энергией. Считается, что эти маленькие клетки были предками того, что мы сейчас знаем как хлоропласты и митохондрии.Мы верим в это, потому что хлоропласты и митохондрии содержат свою собственную ДНК, уникальную от ДНК крупных клеток.  Это позволило бактериям избежать внешней защиты эмбриональной клетки. Для двух других методов ученым нужно было модернизировать бактериальные клетки, чтобы иметь возможность избежать защиты животных клеток.Клетки, которые они использовали, были клетками яичника хомяка (женский репродуктивный орган) и иммунными клетками мыши, особенно макрофагами. Ученые дали цианобактериям специальные гены, которые помогли им избежать обнаружения этими клетками млекопитающих.
Это позволило бактериям избежать внешней защиты эмбриональной клетки. Для двух других методов ученым нужно было модернизировать бактериальные клетки, чтобы иметь возможность избежать защиты животных клеток.Клетки, которые они использовали, были клетками яичника хомяка (женский репродуктивный орган) и иммунными клетками мыши, особенно макрофагами. Ученые дали цианобактериям специальные гены, которые помогли им избежать обнаружения этими клетками млекопитающих. 

 Кроме того, процесс цитокинеза различается между растительными и животными клетками. В этой статье делается попытка подчеркнуть разницу между цитокинезом растений и животных.
Кроме того, процесс цитокинеза различается между растительными и животными клетками. В этой статье делается попытка подчеркнуть разницу между цитокинезом растений и животных.  и другой материал из клеточной пластинки и, наконец, сливая его со стенкой родительской клетки.
и другой материал из клеточной пластинки и, наконец, сливая его со стенкой родительской клетки. 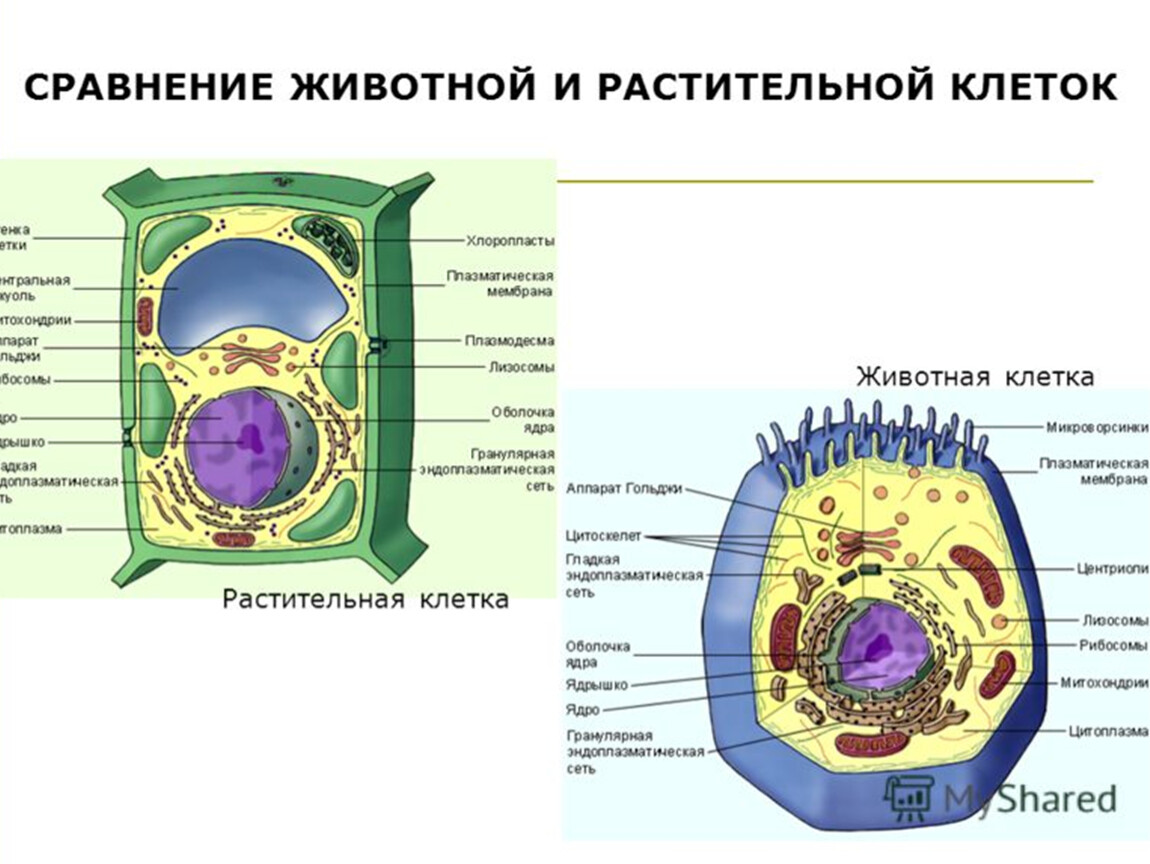
 Кроме того, аппарат Гольджи высвобождает везикулы, содержащие материалы клеточной стенки, которые сливаются в экваториальной плоскости, образуя клеточную пластинку во время цитокинеза растений. С другой стороны, немышечные миозиновые II и актиновые филаменты собираются экваториально, в середине клетки в клеточной коре, образуя сократительное кольцо во время цитокинеза животных.Таким образом, это также существенное различие между растительным и животным цитокинезом.
Кроме того, аппарат Гольджи высвобождает везикулы, содержащие материалы клеточной стенки, которые сливаются в экваториальной плоскости, образуя клеточную пластинку во время цитокинеза растений. С другой стороны, немышечные миозиновые II и актиновые филаменты собираются экваториально, в середине клетки в клеточной коре, образуя сократительное кольцо во время цитокинеза животных.Таким образом, это также существенное различие между растительным и животным цитокинезом. 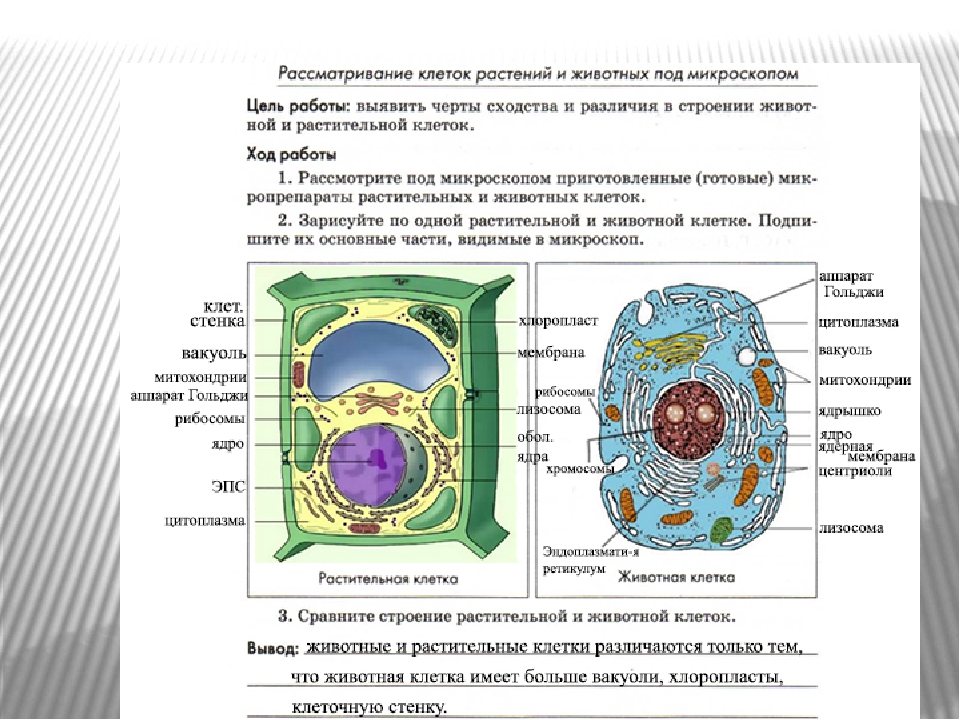 В клетках животных клеточные мембраны на противоположных сторонах клетки защемлены внутрь. Он позволяет клетке делиться. Борозда при дроблении продолжается до тех пор, пока обе стороны не соприкоснутся. При прикосновении в конце появляются две новые клетки. Таким образом, это суммирует разницу между цитокинезом растений и животных.
В клетках животных клеточные мембраны на противоположных сторонах клетки защемлены внутрь. Он позволяет клетке делиться. Борозда при дроблении продолжается до тех пор, пока обе стороны не соприкоснутся. При прикосновении в конце появляются две новые клетки. Таким образом, это суммирует разницу между цитокинезом растений и животных.