
Транспорт питательных веществ | справочник Пестициды.ru
Выделяют несколько типов транспортных систем, которые позволяют различным веществам преодолевать преграду цитоплазматической мембраны (ЦПМ) и попадать внутрь клетки микроорганизма. Это пассивная диффузия, облегченная диффузия, активный транспорт. Отмечается, что только активный транспорт способствует аккумуляции (накоплению) веществ внутри клетки[1].
Схема процесса пассивной диффузииСхема процесса пассивной диффузии
Пассивная диффузия
Пассивная или простая диффузия – неспецифический процесс. Он происходит за счет разницы концентраций. Передвижение молекул осуществляется из более концентрированного раствора в менее концентрированный (по градиенту их концентрации).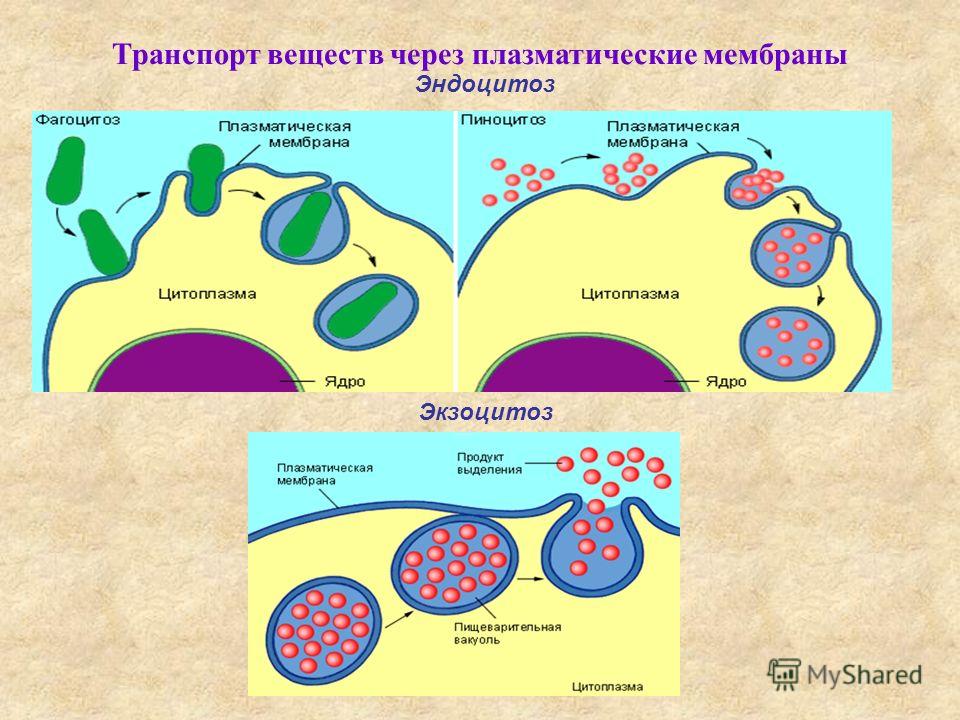
Схема процесса облегченной диффузии
Облегченная диффузия
Облегченная диффузия – перенос веществ через цитоплазматическую мембрану по градиенту их концентрации с участием пермеаз (транслоказ) – специфических мембранных белков, способствующих прохождению веществ через цитоплазматическую мембрану[3]
[1].
Пермеаза фиксирует на себе молекулу переносимого вещества, вместе с ней преодолевает цитоплазматичекую мембрану. После этого комплекс «вещество-пермеаза» диссоциирует. Освободившаяся пермеаза, диффундирует к наружной поверхности, присоединяет новую молекулу вещества и транспортирует ее внутрь клетки[1].
Облегченная диффузия не требует расхода энергии, если наружная концентрация вещества выше внутренней, поскольку в таком случае вещество перемещается «вниз» по химическому градиенту. Скорость процесса зависит от концентрации вещества в наружном растворе. Предполагается, что выход продуктов обмена веществ из микробной клетки может также происходит по методу облегченной диффузии с помощью переносчиков[1].
Параллельно отмечается, что облегченная диффузия более характерна для эукариотических организмов[4].
Схема активного транспорта
Схема активного транспорта
Активный транспорт
Активный транспорт является основным механизмом избирательного переноса вещества через цитоплазматическую мембрану в клетку против градиента концентрации. Этот процесс протекает при участии локализованных в цитоплазматической мембране переносчиков – пермеаз. Это вещества белковой природы, высокочувствительные к субстрату[3].
Этот процесс протекает при участии локализованных в цитоплазматической мембране переносчиков – пермеаз. Это вещества белковой природы, высокочувствительные к субстрату[3].
Активным транспортом в цитоплазму бактериальной клетку поступает подавляющее большинство разнообразных веществ (ионы, углеводы, аминокислоты, липиды)[4].
Для активного транспорта необходимы затраты энергии. Ее получают в виде АТФ, либо за счет протондвижущей силы энергизованной мембраны [3].
У многих микробов, чаще у грамотрицательных бактерий, в активном транспорте принимают участие связующие белки. Эти вещества не входят в структуру мембраны, не идентичны пермеазам. Они локализованы в периплазматическом пространстве. Связующие белки не имеют каталитической активности, но обладают высоким сродством к определенным питательным веществам, аминокислотам, углеводам, неорганическим ионам. Выделено и изучено более 100 различных связующих белков[3].
Активный транспорт осуществляется двумя путями:
- Без химической модификации переносимого вещества[4].

- С химической модификацией переносимого вещества[4].
В первом случае молекула питательного вещества образует комплекс с белком периплазматического пространства. Белок взаимодействует со специфической пермеазой цитоплазматической мембраны. После энергозависимого проникновения через цитоплазматическую мембрану комплекс «субстрат – белок периплазмы – пермеаза» диссоциирует и молекула субстрата освобождается
Во втором случае наблюдается следующие последовательные процессы:
- Фосфорилирование мембранного фермента-2 со стороны цитоплазмы фосфоенолпируватом[4].
- Молекула субстрата связывается на поверхности цитоплазматической мембраны фосфорилированным ферментом-2[4].
- Энергозависимый транспорт молекулы субстрата через мембрану в цитоплазму[4].
- Перенос фосфатной группы на молекулу сбстрата[4].
- Диссоциация в цитоплазме комплекса «субстрат-фермент»[4].

Отмечается, что молекулы субстрата аккумулируются в цитоплазме клеток и теряют способность выйти из них именно за счет фосфорилирования [4].
Отдельные авторы второй путь активного транспорта (с химической модификацией переносимого вещества) выделяют в отдельный (четвертый) способ транспорта питательных веществ – транслокацию (перенос) групп (радикалов)[1][3].
Движения внутри клеток — Справочник химика 21
Область применимости уравнения (8.18) ограничена плотностями,, существенно превышающими критическую плотность, поскольку при выводе формулы (8.17) было сделано предположение, что каждая молекула жидкости совершает колебательные движения внутри клетки , образованной ближайшими соседями этой молекулы. [c.239] У нас нет необходимости вдаваться во все детали. В общих чертах, прокариоты проще, у них отсутствуют специальные молекулы, которые позволяют более совершенным эукариотам осуществлять сложные процессы. Эти процессы позволяют эукариотам нести намного больше генетической информации (разрешая иметь набор хромосом вместо лишь одного отрезка ДНК), жить в других организмах и перемещать молекулы повсюду внутри самих себя с определенной целью. Если есть одно свойство, которое ставит эукариот выше прокариот, так это молекулярный аппарат для генерации и управления движением внутри клетки. Именно он привел к образованию мышц, весьма важных для животных, и допускает сложный танец хромосом, который мы наблюдаем в виде Митоза.
Эти процессы позволяют эукариотам нести намного больше генетической информации (разрешая иметь набор хромосом вместо лишь одного отрезка ДНК), жить в других организмах и перемещать молекулы повсюду внутри самих себя с определенной целью. Если есть одно свойство, которое ставит эукариот выше прокариот, так это молекулярный аппарат для генерации и управления движением внутри клетки. Именно он привел к образованию мышц, весьма важных для животных, и допускает сложный танец хромосом, который мы наблюдаем в виде Митоза.
Движение снаружи ко внутренним областям клетки может происходить тремя путями. Во время фагоцитоза клетка охватывает объект распластывающейся вокруг него мембраной [159]. Такое обволакивание объекта протекает обычно с участием микрофиламентов. Когда мембрана полностью охватит фагоцитируемую частицу, происходит слияние ее краев. Процесс фагоцитоза чувствителен к цитохалазину. При пиноцитозе образуются пузырьки диаметром 200—700 нм, в которые захватывается внеклеточная среда.
 Интенсивность пиноцитоза зависит от типа клетки. Пиноцитоз обеспечивает поступление питательных веществ и сигнальных молекул внутрь клетки. Как и фагоцитоз, он чувствителен к цитохалазинам [171]. Третий тип интернализации мы будет называть эндоцитозом. Это название использовалось нередко для всех трех типов движения внутрь клетки, но в на- [c.93]
Интенсивность пиноцитоза зависит от типа клетки. Пиноцитоз обеспечивает поступление питательных веществ и сигнальных молекул внутрь клетки. Как и фагоцитоз, он чувствителен к цитохалазинам [171]. Третий тип интернализации мы будет называть эндоцитозом. Это название использовалось нередко для всех трех типов движения внутрь клетки, но в на- [c.93] Анабиоз имеет место и при замораживании клеток, когда свободная вода внутри клетки превращается в лед, И в этом случае физиологические процессы максимально замедляются или даже прекращаются, так как биохимические реакции в твердой фазе льда идти не могут из-за отсутствия свободного движения молекул. При замораживании клеток, особенно медленном, образуются крупные кристаллы льда внутри клетки, которые могут вызвать повреждения клеточных структурных элементов. Следовательно, клетки надо обезвоживать или замораживать так, чтобы не допустить необратимые изменения в них, в противном случае наступает летальное состояние — смерть, а не анабиоз.
Структуру и динамику самых разнообразных жидкостей, начиная от жидкого водорода и кончая расплавленными силикатами, можно изучать посредством различных спектральных методов. Среди них наиболее важны дифракция рентгеновских лучей, нейтронография, ядерный магнитный резонанс, лазерная спектроскопия комбинационного рассеяния и рассеяние света. Одним из самых мощных новейших методов является импульсное лазерное возбуждение. В пикосекундном диапазоне (10 с) мы можем исследовать движение молекулы растворенного вещества внутри клетки молекул растворителя. Теперь можно непосредственно наблюдать за фундаментальными химическими событиями в реальном времени. Например, можно наблюдать, как два атома иода в жидкости соединяются в молекулу, как захватывается (сольватируется) жидкой водой свободный электрон, как энергия, поглощенная молекулой растворенного вещества (азот или бензол), передается от нее окружающим молекулам растворителя.
Сеансы в совершенно темной комнате вокруг стола, когда присутствующие держат друг друга за руки, а на столе поставлена клетка с запертым внутри ее колокольчиком. Ожидаемое спиритическое явление составляет движение всей клетки и отдельно звон колокольчика. [c.188]
Одна из проблем, связанных с процессом поступления элемента из внешнего раствора в растение, возникает в связи с тем, что это поступление может осуществляться против градиента концентрации, т. е. элемент может входить в клетку при условиях, когда концентрация внутри клетки превышает концентрацию во внешнем растворе. Множество исследований накопления ионов направлено на изучение сути процессов, включающих движение против градиента концентрации. Некоторые из них касаются природы носителей , которые участвуют в переходе ионов через мембрану. Накопление ионов в растениях против градиента концентрации обычно зависит от энергии, высвобождающейся при аэробном дыхании, но механизм перехода энергии в процессе накопления ионов понятен не полностью.
Ферменты и промежуточные продукты обмена веществ неравномерно распределены в отдельных элементах клеточной структуры. Упорядоченному движению молекул внутри клетки способствует высокая степень избирательности клеточной мембраны. Транспорт веществ через биологические мембраны, представляющие собой генетически детерминированные структуры, в которых заложена информация относительно тех процессов, выполнение которых они обеспечивают, является ферментативным процессом и обусловлен активностью мембранных ферментов. [c.439]
К оболочке вплотную прилегает цитоплазматическая мембрана. Она обладает избирательной проницаемостью, т. е. пропускает внутрь клетки и отводит из нее определенные вещества. Благодаря такой способности мембрана играет роль органеллы, концентрирующей питательные вещества внутри клетки и способствующей выведению наружу продуктов жизнедеятельности. Внутри клетки всегда наблюдается повышенное по сравнению о окружающей средой осмотическое давление. Цитоплазматическая мембрана обеспечивает его постоянство. Кроме того, она является местом локализации ряда ферментных систем, в частности окислительно-восстановительных ферментов, связанных с получением энергии (у эукариотов они находятся в митохондриях). В отличие от клеток эукариотов в прокариотической клетке отсутствует деление ее на отсеки. Клетки прокариотов не имеют ни комплекса Гольджи, ни митохондрий, не наблюдается у них и направленного движения цитоплазмы. Явления пиноцитоза и фагоцитоза прокариотам не свойственны. Из органелл только рибосомы аналогичны рибосомам эукариотов. [c.43]
Цитоплазматическая мембрана обеспечивает его постоянство. Кроме того, она является местом локализации ряда ферментных систем, в частности окислительно-восстановительных ферментов, связанных с получением энергии (у эукариотов они находятся в митохондриях). В отличие от клеток эукариотов в прокариотической клетке отсутствует деление ее на отсеки. Клетки прокариотов не имеют ни комплекса Гольджи, ни митохондрий, не наблюдается у них и направленного движения цитоплазмы. Явления пиноцитоза и фагоцитоза прокариотам не свойственны. Из органелл только рибосомы аналогичны рибосомам эукариотов. [c.43]
По вопросу о роли движения протоплазмы в жизни клетки нет единой точки зрения, однако важность этого процесса сомнений не вызывает. Большое значение могут иметь круговые движения протоплазмы как один из быстрых путей перемещения веществ внутри клетки и из клетки в клетку. [c.35]
Проникновение Са » внутрь клетки влияет на орган движения инфузории — реснички — точно так же, как у нас вхождение Са в мышечные клетки необходимо для их сокращения (мы рассказывали об этом, говоря про Са-насос). Кстати, и разрядка трихоцист у инфузорий связана с потоками Са в клетку. Большинство Са-каналов инфузорий расположено прямо на мембране ресничек. Если удар по передней части инфузории вызвал ПД, то откроется много Са-каналов, внутрь клетки войдет много Са , а от этого реснички инфузории меняют направление своего удара. Возникает реверс инфузория отплывает хвостом вперед от раздражителя, например от препятствия, на которое она натолкнулась. После того как Са-насос и митохондрии уберут излишки Са++ из цитоплазмы, нормальная работа ресничек восстанавливается. [c.262]
Кстати, и разрядка трихоцист у инфузорий связана с потоками Са в клетку. Большинство Са-каналов инфузорий расположено прямо на мембране ресничек. Если удар по передней части инфузории вызвал ПД, то откроется много Са-каналов, внутрь клетки войдет много Са , а от этого реснички инфузории меняют направление своего удара. Возникает реверс инфузория отплывает хвостом вперед от раздражителя, например от препятствия, на которое она натолкнулась. После того как Са-насос и митохондрии уберут излишки Са++ из цитоплазмы, нормальная работа ресничек восстанавливается. [c.262]
Для изучения структуры ацетилхолинового рецептора были использованы методы электронной микроскопии и малоугловой дифракции рентгеновских лучей, однако точный ответ на вопрос, как образуется трансмембранный гидрофильный канал, до сих пор не получен. Было предложено несколько моделей, основанных главным образом на аминокислотной последовательности субъединиц Одна из моделей представлена на рис. 6-64. То, что кластеры отрицательно заряженных аминокислотных остатков выстилают отверстие канала, объясняет, но-видимому, известный факт, что отрицательно заряженные ионы не способны проходить через канал, а положительно заряженные ионы с размером до 0,65 нм могут это делать. Через канал проходят преимущественно ионы Ка» и К», а также некоторое количество Са «. Строгих ограничений на вид катионов не существует, поэтому поток каждого из них через канал определяется главным образом их концентрациями и электрохимическими движущими силами. Так как градиент напряжения уравновешивает градиент концентрации К» через мембрану при наличии потенциала покоя, то и движущая сила для ионов К близка к нулю (см. схему 6-2). Напротив, для ионов Ка как градиент напряжения, так и градиент концентрации действуют в одном направлении, способствуя движению ионов внутрь клетки. Это же справедливо и для Са «, но его внеклеточная концентрация намного [c.404]
Через канал проходят преимущественно ионы Ка» и К», а также некоторое количество Са «. Строгих ограничений на вид катионов не существует, поэтому поток каждого из них через канал определяется главным образом их концентрациями и электрохимическими движущими силами. Так как градиент напряжения уравновешивает градиент концентрации К» через мембрану при наличии потенциала покоя, то и движущая сила для ионов К близка к нулю (см. схему 6-2). Напротив, для ионов Ка как градиент напряжения, так и градиент концентрации действуют в одном направлении, способствуя движению ионов внутрь клетки. Это же справедливо и для Са «, но его внеклеточная концентрация намного [c.404]
Трансмембранный потенциал частично обусловлен избирательной проницаемостью клеточной мембраны, что ограничивает скорость движения одного иона относительно другого. К+, например, может проникать через мембрану значительно быстрее,, чем С1 . Если оба этих иона нутри клетки имеют более высокую концентрацию, чем вокруг нее, то более быстрая нетто-диф-фузия ионов К+ наружу по градиенту концентрации в конечном итоге приведет к возникновению более высокого отрицательного заряда внутри клетки, так как там останется избыточное ко- личество С1 .
 [c.223]
[c.223] Движение ионов через мембраны происходит частично благодаря электрохимическим градиентам и частично с помощью локализованных в мембранах насосов. Когда транспорт осуществляется по электрохимическому градиенту, ионы сначала присоединяются к особым участкам на мембране (пермеазам). Затем они проникают в клетку в соответствии с уравнением Нернста, если общий эффект градиента их концентрации по обе стороны мембраны и электрический трансмембранный потенциал обеспечивают движущую силу, направленную внутрь. Транс-, мембранные потенциалы образуются двумя путями 1) в результате диффузии как анионов, так и катионов, которые, однако, движутся через мембрану с разными скоростями 2) благодаря электрогенному транспорту с прямым использованием энергии для прокачивания протонов, анионов или катионов через мембрану против их электрохимических градиентов. Оба этих процесса всегда действуют таким образом, что внутри клетки создается более отрицательный заряд по сравнению с зарядом юне ее. [c.238]
[c.238]
В отличие от рассмотренных ранее перемещений растений, которые, как правило, можно наблюдать только с помощью микроскопов, движения органов закрепленных в субстрате растений обнаруживаются обычно невооруженным глазом. Здесь мы встречаемся не только с различиями в размерах, но и с совершенно иными механизмами движений. Если движения внутри клеток, с помощью жгутиков и амебоидные, как и перемещения животных, основываются главным образом на способности белковых молекул сокращаться, органы прикрепленных растений изменяют свое положение в пространстве прежде всего благодаря процессам роста и колебаниям тур горного давления. Но несмотря на серьезные различия в механике движений, обнаруживается значительное сходство в проявлении основных физиологических закономерностей, связанных с раздражениями. Все, что мы обсуждали, рассматривая клетку и свободные перемещения растений, облегчит знакомство с изменением положения органов, которым вы, может быть, заинтересовались во время прогулок или экскурсий, посещения оранжерей или наблюдая растения у себя дома. [c.58]
[c.58]
Различают активный и пассивный перенос (транспорт) нейтральных молекул и ионов через биомембраны. Активный транспорт происходит при затрате химической энергии за счет гидролиза АТФ или переноса электрона по дыхательной цепи митохондрий. Пассивный транспорт не связан с затратой клеткой химической энергии он осуществляется в результате диффузии веществ в сторону меньшего электрохимического потенциала (рис. 48). Примером активного транспорта может служить перенос ионов калия и натрия через цитоплазматические мембраны (К» — внутрь клетки, а Ыа» — из нее), перенос кальция через мембраны саркоплазматического ретикулума скелетных и сердечной мышц внутрь пузырьков ретикулума, перенос ионов водорода через мембраны митохондрий из матрикса наружу. Все эти процессы происходят за счет энергии гидролиза АТФ и осуществляются особыми ферментами — транспортными АТФ-азами (рис. 49). Наиболее известный пример пассивного транспорта — это движение ионов натрия и калия через цитоплазматическую мембрану нервных волокон при распространении потенциала действия. Впрочем, и в покоящейся клетке существует утечка ионов через мембраны, обусловленная их проницаемостью проницаемость обычно возрастает при патологии. [c.122]
Впрочем, и в покоящейся клетке существует утечка ионов через мембраны, обусловленная их проницаемостью проницаемость обычно возрастает при патологии. [c.122]
Калий. Калий необходим не только как питательный элемент, но п как стимулятор размножения дрожжей. Стимулирующее дей-стБие объясняется его существенной ролью в окислительном фос-форилировании и в процессах гликолиза. Движение неорганического фосфора внутрь клетки специфично стимулируется калием. Калий активирует дрожжевую альдолазу, необходим для действия фермента иируваткарбоксилазы и влияет, так же как азот и сера, на липидный обмен дрожжевых клеток. [c.199]
Он образует цилиндрический канал, который с одной стороны выступает на 65 А в синаптическую щель, а с другой — пронизывает липидный бцслой мембраны, входя на 15 А внутрь клетки. Этот узкий канал (или пора) расширяется до 20 А при «посадке» на рецептор нейромедиатора (комплекс RAX) за счет резкого уменьшения вращательного (конформационного) движения субъединиц. Увеличение размера канала облегчает прохождение ионов К+ и Na+ через мембрану против электрохимического фадиента. При этом изменяется мембранный потенциал покоящегося нейрона 2, и в нем генерируется нервный импульс. После этого нейромедиатор гидролизуется ацетилхолинэстера-зой до неактивного холина, и ионофорныи канал закрывается. [c.31]
Увеличение размера канала облегчает прохождение ионов К+ и Na+ через мембрану против электрохимического фадиента. При этом изменяется мембранный потенциал покоящегося нейрона 2, и в нем генерируется нервный импульс. После этого нейромедиатор гидролизуется ацетилхолинэстера-зой до неактивного холина, и ионофорныи канал закрывается. [c.31]
Питание микроорганизмов осуществляется через поверхность их тела путем диффузии в результате разных концентраций веществ внутри и вне организма. Движение растворенных веществ лод действием осмотического давления происходит в сторону мень-щих концентраций, воды — в сторону больших. Так как поступающие в клетку вещества вовлекаются в биохимические процессы и усваиваются микроорганизмом, равновесия их внутри клетки и. вне ее практически не наступает. Однако проникновение вещества -В клетку не всегда объяснимо осмосом. Цитоплазматическая мембрана обладает избирательной способностью отличать нужные вещества от ненужных и извлекать их из растворов с малой концентрацией, не пропуская вредные для клетки вещества, содержащиеся в среде в значительных концентрациях (до определенных лределов). Так как поверхность клеток на единицу их массы лредставляет громадную величину, то процессы обмена и размножения микроорганизмов происходят с большими скоростями, и этим объясняются интенсивные биоповреждения некоторых материалов, на которых идут такие процессы. Давление в клетке создается поступившими в нее веществами, продуктами обмена и веществами клеточного синтеза. В связи с высоким осмотическим давлением внутри клетки создается постоянный приток в нее воды. Этим можно объяснить способность микроорганизмов развиваться на сравнительно сухих средах. Так, микрогрибы способны повреждать материалы, имеющие влажность 15…20 % и ниже. [c.15]
Так как поверхность клеток на единицу их массы лредставляет громадную величину, то процессы обмена и размножения микроорганизмов происходят с большими скоростями, и этим объясняются интенсивные биоповреждения некоторых материалов, на которых идут такие процессы. Давление в клетке создается поступившими в нее веществами, продуктами обмена и веществами клеточного синтеза. В связи с высоким осмотическим давлением внутри клетки создается постоянный приток в нее воды. Этим можно объяснить способность микроорганизмов развиваться на сравнительно сухих средах. Так, микрогрибы способны повреждать материалы, имеющие влажность 15…20 % и ниже. [c.15]
Характерным свойством живого материала является то, что он движется. Степень движения меняется от явного перемещения в потоке цитоплазмы до движения ионов, электролитов, молекул и макромолекул относительно друг друга внутри клетки. В результате обмена веществ биологический материал постоянно изменяется, разрушая и перестраивая функциональную архитектуру клетки. Эта выраженная нестабильность мешает проведению рентгеновского микроанализа, если не найдены пути мгновенного сдерживания активности клетки и удержания ее в этом состоянии до тех пор, пока выполняются исследования. Если бы это было сделано, то окружающая среда, в которой должен производиться рентгеновский микроанализ, полностью была бы лишена жизненных процессов. Типичный одноклеточный организм менее 2 мкм в поперечинке синтезирует много сотен соединений путем тонкого регулируемого процесса, способен воспроизводить сам себя и генетически эволюционировать и видоизменять эти процессы. Если захотелось бы найти быстрый способ разрушения этого уникального тончайшего механизма, то, вероятно, не нашлось бы ничего лучше потока быстрых электронов, который за одну секунду смог бы испарить количество воды, во много раз превышающее вес образца. [c.266]
Эта выраженная нестабильность мешает проведению рентгеновского микроанализа, если не найдены пути мгновенного сдерживания активности клетки и удержания ее в этом состоянии до тех пор, пока выполняются исследования. Если бы это было сделано, то окружающая среда, в которой должен производиться рентгеновский микроанализ, полностью была бы лишена жизненных процессов. Типичный одноклеточный организм менее 2 мкм в поперечинке синтезирует много сотен соединений путем тонкого регулируемого процесса, способен воспроизводить сам себя и генетически эволюционировать и видоизменять эти процессы. Если захотелось бы найти быстрый способ разрушения этого уникального тончайшего механизма, то, вероятно, не нашлось бы ничего лучше потока быстрых электронов, который за одну секунду смог бы испарить количество воды, во много раз превышающее вес образца. [c.266]
Изучение фотографических изображений клетки, полученных прн помощи микроскопа в разные моменты времени, позволили увидеть, что плазматическая мембрана, так же как и митохондрии и другие органеллы, постоянно находится в движении. Митохондрии скручиваются и поворачиваются, а поверхность мембраны постоянно совершает волнообразные движения. Пузырьки освобождают свое содержимое в окружающую среду, выводя его из клеток, а перенос веществ внутрь клетки осуществляется за счет процесса эндоцитоза (гл. 1, разд. Б.4). При помощи химических методов было показано также, что составляющие мембраны вещества транспортируются из эндоплаз1матического ретикулума в пузырьки аппарата Гольджи, в экскреторные гранулы и в плазматическую мембрану. Важным этапом биосинтетических процессов, протекающих в клетке, является присоединение углеводных (гликозильных) остатков к молекулам белка с образованием гликопротеидов и гликолипидов. Ферменты, катализирующие эти реакции, — гликозилтрансферазы (гл. 12)-—обнаружены в эндоплазматическом ретикулуме и в пузырьках а1ппарата Гольджи. Эти ферменты катализируют присоединение углеводных единиц (по одной в каждом акте реакции) к определенным местам молекул белков, липидов и других соединений, экскретируемых из клеток.
Митохондрии скручиваются и поворачиваются, а поверхность мембраны постоянно совершает волнообразные движения. Пузырьки освобождают свое содержимое в окружающую среду, выводя его из клеток, а перенос веществ внутрь клетки осуществляется за счет процесса эндоцитоза (гл. 1, разд. Б.4). При помощи химических методов было показано также, что составляющие мембраны вещества транспортируются из эндоплаз1матического ретикулума в пузырьки аппарата Гольджи, в экскреторные гранулы и в плазматическую мембрану. Важным этапом биосинтетических процессов, протекающих в клетке, является присоединение углеводных (гликозильных) остатков к молекулам белка с образованием гликопротеидов и гликолипидов. Ферменты, катализирующие эти реакции, — гликозилтрансферазы (гл. 12)-—обнаружены в эндоплазматическом ретикулуме и в пузырьках а1ппарата Гольджи. Эти ферменты катализируют присоединение углеводных единиц (по одной в каждом акте реакции) к определенным местам молекул белков, липидов и других соединений, экскретируемых из клеток. Другие ферменты катализируют присоединение сульфатных и ацетильных групп к углеводным фрагментам молекул глико Протеидов. [c.356]
Другие ферменты катализируют присоединение сульфатных и ацетильных групп к углеводным фрагментам молекул глико Протеидов. [c.356]
Мышечное сокращение — лучше всего изученное механохи-мическое явление. К таким же явлениям относится множество биологических процессов движения растений движения клеток с помощью жгутиков и ресничек-, вся совокупность движений в дроцессах митоза и мейоза движения внутри не делящейся клетки сократительные процессы в хвостах фаговых частиц механохимические процессы в мембранах движение рибосом относительно мРНК в полисомах акустическая и механорецепция. [c.411]
Концентрация и осмотическое давление различных жидкостей в организме поддерживаются на постоянном уровне действием специальных осморегуляторов. Осмотическое давлегше растворов является следствием теплового движения молекул растворенного вещества, стремящегося занять возможно больший объем. Плазма крови, лимфа, слезная и спинномозговая жидкость имеют постоянное осмотическое давление (гипертонический раствор) в результате разности осмотических давлений внутри эритроцитов и окружающей та плазмы осуществляется движение воды из эритроцитов, идущее до выравнивания осмотических давлений. Эритроциты при этом, лишаясь части водьт, сморщиваются (плазмолиз). Если вводится раствор с малым осмотическим давлением (гипотонический раствор), жидкость проникает внутрь клетки эритроцит разбухает, клеточная оболочка может нарушиться, а клетка погибнуть (гемолиз). Чтобы избежать указанных осмотических сдвигов необходимо изотонизировать раствор до уровня осмотического давления биологических жидкостей оргатшзма. Такие растворы называются изотоническими. [c.635]
Эритроциты при этом, лишаясь части водьт, сморщиваются (плазмолиз). Если вводится раствор с малым осмотическим давлением (гипотонический раствор), жидкость проникает внутрь клетки эритроцит разбухает, клеточная оболочка может нарушиться, а клетка погибнуть (гемолиз). Чтобы избежать указанных осмотических сдвигов необходимо изотонизировать раствор до уровня осмотического давления биологических жидкостей оргатшзма. Такие растворы называются изотоническими. [c.635]
Концентрация и осмотическое давление различных жидкостей в организме поддерживаются на постоянном уровне действием специальных осморегуляторов. Осмотическое давление растворов является следствием теплового движения молекул растворенного вещества, стремящегося занять возможно больший объем. Плазма крови, лимфа, слезная и спинномозговая жидкость имеют постоянное осмотическое давление (гипертонический раствор) в результате разности осмотических давлений внутри эритроцитов и окружающей их плазмы осуществляется движение воды из эритроцитов, идущее до выравнивания осмотических давлений. Эритроциты при этом, лишаясь воды, сморщиваются (плазмолиз). Если вводится раствор с малым осмотическим давлением (гипотонический раствор), жидкость проникает внутрь клетки эритроцит разбухает, клеточная оболочка может нарушиться, а [c.371]
Эритроциты при этом, лишаясь воды, сморщиваются (плазмолиз). Если вводится раствор с малым осмотическим давлением (гипотонический раствор), жидкость проникает внутрь клетки эритроцит разбухает, клеточная оболочка может нарушиться, а [c.371]
Молекулы воды, некоторых газов (например, О2, Н2, N2) и углеводородов, концентрации которых во внешней среде выше, чем в клетке, проходят через ЦПМ внутрь клетки посредством пассивной диффузии. Движущей силой этого процесса служит фадиент конценфации вещества по обе стороны мембраны. Основным соединением, поступающим в клетку и покидающим ее таким путем, является вода. Движение воды через мембрану, подчиняющееся законам пассивной диффузии, привело к выводу о существовании в мембране пор. Эти поры пока не удалось увидеть в элекфонный микроскоп, но некоторые данные о них были получены косвенными методами. Расчетным путем установлено, что поры должны быть очень мелкими и занимать небольшую часть поверхности ЦПМ. Высказывается предположение, что они не являются стабильными сфуктурными образованиями, а возникают в результате временных пересфоек молекулярной организации мембраны. [c.50]
[c.50]
Бактерии, имеющие палочковидную или цилиндрическую форму клетки, бывают разной длины от совсем коротких, почти кокков и коккобактерий (I—1,5 мкм), до длинных, иногда прямых, а иногда изогнутых или искривленных палочек и даже нитей (10—18 мкм и больше). Палочковидные формы бактерий широко распространены в природе. Их примерно в три раза больше, чем сферических. Это объясняется более выгодным соотношением массы и поверхности у цилиндра, чем у шара. Многие палочковидные формы подвижны. Органами движения служат так называемые жгутики. Среди палочковидных форм встречается довольно много видов, способных к образованию спор. Споры всегда образуются внутри клетки — эндоспоры. Спорообразующие палочковидные грамположительные аэробные формы называются бациллами и объединяются в род Ba illus. Споровые грамположительные анаэробные фор.мы, живущие без воздуха, [c.10]
Ван-дср Ваальс [297] недавно отметил, что невозможно строго записать значение потенциальной энергии клатратированных молекул в виде суммы значений координат положения и ориентации. Вклад вращения в свободную энергию будет зависеть от состояния вибрации. Это последнее состояние, — говорит он, — соответствует движению молекул внутри клетки . Ярко выраженным доказательством такой потери свободы ориентации многоатомными молекулами в клатрате является меньшая энтропия их клатратов по сравнению с энтропией многоатомшзьх газов. [c.93]
Вклад вращения в свободную энергию будет зависеть от состояния вибрации. Это последнее состояние, — говорит он, — соответствует движению молекул внутри клетки . Ярко выраженным доказательством такой потери свободы ориентации многоатомными молекулами в клатрате является меньшая энтропия их клатратов по сравнению с энтропией многоатомшзьх газов. [c.93]
Проницаемость в живых клетках представляет собой активный процесс и имеет мало общего с молекулярной диффузией или осмотическим потоком. Наоборот, активный транспорт осуществляется чаще всего против градиента концентрации, т. е. в направлении от мепьшей концентрации к большей. Ясно, что это — сложное явление, в котором обязательно должна потребляться энергия, так как движение веществ в направлении, обратном диффузии, связано с уменьшением энтропии. Активный перенос веществ как внутрь клетки из внешней среды, так и внутрь различных структурных элементов из заполяющей клетку гиалоплазмы осуществляется особыми нерастворимыми белками и белковыми комплексами, образующими наружную клеточную мембрану и различные структурные образования внутри клеток. Активный транспорт через мембраны и внутрь клеточных органелл связан с протеканием химических реакций, конечно, ферментативных. Поэтому проблема проницаемости и соответствующая функция белков тесно связана с их ферментативной функцией. С другой стороны, с помощью активного транспорта осуществляется один из механизмов автоматического регулирования. Как мы увидим дальше, регулирование проницаемости митохондрий осуществляется путем их сокращения пли расслабления. Причиной этого движения яляется сократительная реакция в особом белке, т. е. это явление вполне аналогично сокращению мышцы. [c.139]
Активный транспорт через мембраны и внутрь клеточных органелл связан с протеканием химических реакций, конечно, ферментативных. Поэтому проблема проницаемости и соответствующая функция белков тесно связана с их ферментативной функцией. С другой стороны, с помощью активного транспорта осуществляется один из механизмов автоматического регулирования. Как мы увидим дальше, регулирование проницаемости митохондрий осуществляется путем их сокращения пли расслабления. Причиной этого движения яляется сократительная реакция в особом белке, т. е. это явление вполне аналогично сокращению мышцы. [c.139]
Перейдем теперь к генетике бактериофагов, которые изучены гораздо лучше, чем все другие вирусы. Картина заражения клетки бактериофагом следующая. Бактериофаг адсорбируется своим хвостом на внешней поверхности клетки, проделывает в оболочке микроскопическое отверстие, для чего в его хвосте присутствует специальный фермент со свойствами лизоцима, затем инъецирует внутрь клетки свое содержимое, что у больших фагов сопровождается настоящим сократительным движением (рис. 124). В результате от фага остается нустая белковая оболочка, или тень . Отдельные эпизоды во всей этой последовательности удается хорошо заснять с помощью электронного микроскопа. Освободить бактериальную клетку от адсорбированных на ней пустых оболочек фагов легко с помощью быстрой мешалки. [c.364]
124). В результате от фага остается нустая белковая оболочка, или тень . Отдельные эпизоды во всей этой последовательности удается хорошо заснять с помощью электронного микроскопа. Освободить бактериальную клетку от адсорбированных на ней пустых оболочек фагов легко с помощью быстрой мешалки. [c.364]
Аминокислота глицин представляет собой тормозный нейромедиатор, открывающий хло-ридные каналы в постсинаптической мембране и приводящей к ее гаперполяризации (внутри клетки увеличивается отрицательный заряд). Глицин играет важную роль в спинном мозге, где способствует контролю движений скелетных мышц, поддерживая их расслабление (предотвращая стимуляцию). О том, каково значение глицина, можно судить по действию стрихнина, который блокирует глициновые рецепторы, подавляя тем самым эффект глицина. В этом случае даже очень слабая стимуляция вызывает мышечное сокращение. Пострадавший от стрихни-нового отравления задыхается, поскольку не способен расслабить мышцы, участвующие в дыхании. [c.294]
[c.294]
Пусть теперь внутри клетки имеется много свободных ионов какого-то элемента, например калия, а снаружи таких ионов нет или их гораздо меньше. Пусть клеточная мембрана пропускает только ионы и не пропускает никаких других ионов. Тогда ионы начнут выходить из клетки, где их много, наружу (двигаться по градиенту концентрации, диффундировать — все эти слова означают одно и то же). Вместе с ними будет выноситься наружу их положительный заряд. Внутрь через мембрану будет проходить мало ионов, так как снаружи мало калия. В реэультате на клеточной мембране будет возникать разность потенциалов снаружи клетки — плюс , а внутри — минус (рис. И). Эта разность потенциалов будет тормозить движение новых положительных заряженных ионов калия наружу и увеличивать поток этих ионов внутрь. Когда потоки ионов наружу и внутрь сравняются, установится динамическое равновесие и на мембране будет поддерживаться постоянная разность потенциалов. Это и есть потенциал покоя (ПП). Его величина описывается формулой Нернста (3.2). [c.61]
Его величина описывается формулой Нернста (3.2). [c.61]
Микротрубочки растут в одном направлении от специфических центров (центриолей) внутри клетки. На каждой хроматиде хромосомы (см. гл. 37) имеется кинетохор, откуда начинается рост микротрубочек. Многие нарущения в делении хромосом являются результатом аномалий в структуре или функции кинетохоров. Движение хромосом в анафазе митоза зависит от микротрубочек, но молекуляр- [c.345]
Механизм движения везикул в клетке, очевидно, не диффузионный. Как мы увидим в дальнейшем, внутри живой клетки, как и в отдельных ее органеллах, нет места для диффузионной диссипации энергии. Все движения в клетке управляются межмолекулярными взаимодействиями и локальными электрическими полями. Так организован и транс-цитоз — транспорт молекул через клетку. Этот процесс характерен для поляризованных клеток, таких как эпителиальные клетки кишечника, которые имеют базальную и апикальную поверхности (каждая со своим определенным фосфолипидным составом), создающие электрическое попе в клетке и определяющие направление транспорта везикул. Примером может служить адсорбция антител, содержащихся в молоке матери, клетками кишечника новорожденного. Эти антитела поглощаются апикальной поверхностью эндотелиальных клеток, переносятся внутри клетки к базальной поверхности и затем вьщеляются с базальной поверхности в кровь. Аналогично организован механизм секреции тирео-идного гормона. Сначала тиреоглобулин выделяется в просвет фолликула щитовидной железы, затем происходит эндоцитоз тиреоглобулина эпителиальными клетками, в составе везикул он транспортируется через клетку, одновременно подвергаясь частичному протеолизу, и образованный в везикулах низкомолекулярный гормон тироксин секретируется в ближайший кровеносный капилляр. [c.120]
Примером может служить адсорбция антител, содержащихся в молоке матери, клетками кишечника новорожденного. Эти антитела поглощаются апикальной поверхностью эндотелиальных клеток, переносятся внутри клетки к базальной поверхности и затем вьщеляются с базальной поверхности в кровь. Аналогично организован механизм секреции тирео-идного гормона. Сначала тиреоглобулин выделяется в просвет фолликула щитовидной железы, затем происходит эндоцитоз тиреоглобулина эпителиальными клетками, в составе везикул он транспортируется через клетку, одновременно подвергаясь частичному протеолизу, и образованный в везикулах низкомолекулярный гормон тироксин секретируется в ближайший кровеносный капилляр. [c.120]
Фаллоидин-высокотоксичный алкалоид гриба Amanita phalloides-ъ противоположность цитохалазинам стабилизирует актиновые филаменты и подавляет их деполимеризацию. Этот агент не может легко проходить через цитоплазматическую мембрану, поэтому его приходится инъецировать в клетку. Оказалось, что он блокирует миграцию не только амеб, но и различных клеток позвоночных в культуре по-видимому, процессы сборки и деполимеризации актиновых филаментов играют ключевую роль в амебоидном движении. Фаллоидин стабилизирует актиновые филаменты, высокоспецифичным образом связываясь с ними по всей их длине это позволяет использовать его флуоресцентные производные для окрашивания актиновых филаментов внутри клетки (см. рис. 10-78). [c.100]
Оказалось, что он блокирует миграцию не только амеб, но и различных клеток позвоночных в культуре по-видимому, процессы сборки и деполимеризации актиновых филаментов играют ключевую роль в амебоидном движении. Фаллоидин стабилизирует актиновые филаменты, высокоспецифичным образом связываясь с ними по всей их длине это позволяет использовать его флуоресцентные производные для окрашивания актиновых филаментов внутри клетки (см. рис. 10-78). [c.100]
В состоянии геля основная плазма представляет собой дисперсную систему, в которой частицы диспергированного вещества расположены в дисперсионной среде в виде сети или сот и связаны друг с другом в местах соприкосновения. Эта имеющая высокую вязкость основная плазма образует канальцы меняющейся ширины, в которые устремляется менее вязкая основная плазма, находящаяся в состоянии золя. Необходимая для сокращения энергия поставляется в форме АТФ (аденозинтри-фосфорной кислоты), т. е. освобождается при гидролитическом расщеплении богатой энергией АТФ. При этом после отщепления от АТФ концевой фосфатной группы возникает АДФ (аденозиндифосфор-ная кислота). Как вам, может быть, уже известно, АТФ/АДФ-система — это главная система передачи энергии внутри клетки. Движения цитоплазмы могут быть вызваны как внутренними (автономными), так и внешними раздражениями. В этих случаях мы говорим о динезах (например, о фото-, [c.33]
При этом после отщепления от АТФ концевой фосфатной группы возникает АДФ (аденозиндифосфор-ная кислота). Как вам, может быть, уже известно, АТФ/АДФ-система — это главная система передачи энергии внутри клетки. Движения цитоплазмы могут быть вызваны как внутренними (автономными), так и внешними раздражениями. В этих случаях мы говорим о динезах (например, о фото-, [c.33]
Что МЫ знаем о процессах, проходящих в сочленениях Как уже было упомянуто при обсуждении основных положений физиологии раздражений, возбуждение характеризуется проявлением потенциала действия. yMimosa pudi a он достигает примерно 140 мВ в это время потенциал покоя, как правило, равен —160 мВ, но после раздражения он увеличивается на —20 мВ. Согласно нащим современным представлениям электрофизиологические явления зависят прежде всего от ионов соединений хлора и калия, концентрация которых внутри клетки регулируется с помощью ионных насосов. Возбуждение моторных клеток приводит к временному нарушению структуры плазмалеммы. С этим связаны увеличение проницаемости мембраны, временная приостановка деятельности ионных насосов, выход ионов хлора и калия, а также резкое падение тургора. Кроме того, клеточный сок выходит из вакуоли в клеточную оболочку и в межклетники. Невооруженным глазом можно видеть, что нижняя сторона первичного сочленения становится явно более темной. Одновременно с потерей тургора моторными клетками, находящимися на нижней стороне, клетки верхней стороны частично теряют свою сопротивляемость. В уравнении сосущей силы это величина А. Следовательно, на верхней стороне увеличивается сосущая сила и соответственно поглощение воды, и лист опускается. Теперь можно также понять, почему при проявлении закона все или ничего подпороговые раздражения не суммируются, а ответная реакция наступает лишь после того, как будет превышен порог раздражения. Очевидно, это зависит от состояния мембраны. Возможно, что в выведении (выдавливании) воды участвуют и сократительные белки. На это указывает, в частности, высокое содержание АТФ в моторных клетках, сильно падающее во время движения.
С этим связаны увеличение проницаемости мембраны, временная приостановка деятельности ионных насосов, выход ионов хлора и калия, а также резкое падение тургора. Кроме того, клеточный сок выходит из вакуоли в клеточную оболочку и в межклетники. Невооруженным глазом можно видеть, что нижняя сторона первичного сочленения становится явно более темной. Одновременно с потерей тургора моторными клетками, находящимися на нижней стороне, клетки верхней стороны частично теряют свою сопротивляемость. В уравнении сосущей силы это величина А. Следовательно, на верхней стороне увеличивается сосущая сила и соответственно поглощение воды, и лист опускается. Теперь можно также понять, почему при проявлении закона все или ничего подпороговые раздражения не суммируются, а ответная реакция наступает лишь после того, как будет превышен порог раздражения. Очевидно, это зависит от состояния мембраны. Возможно, что в выведении (выдавливании) воды участвуют и сократительные белки. На это указывает, в частности, высокое содержание АТФ в моторных клетках, сильно падающее во время движения. [c.128]
[c.128]
Урок 8. передвижение веществ у животных — Биология — 6 класс
Биология, 6 класс
Урок 8. Передвижение веществ у животных
Перечень вопросов, рассматриваемых на уроке
- На сегодняшнем уроке мы продолжим изучение процессов жизнедеятельности живых организмов.
- Познакомимся с тем, как осуществляется транспорт веществ.
Тезаурус
Гемолимфа – жидкость, циркулирующая в сосудах и межклеточных полостях беспозвоночных животных с незамкнутой системой кровообращения (членистоногие, моллюски).
Кровь – жидкая и подвижная соединительная ткань внутренней среды организма, которая состоит из плазмы (жидкой среды) и взвешенных в ней клеток(форменных элементов: эритроцитов, лейкоцитов и тромбоцитов).
Сердце – полый мышечный орган, который обеспечивает с помощью повторных ритмичных сокращений движение крови по кровеносным сосудам.
*Артерии – кровеносные сосуды, несущие кровь от сердца к органам.
*Вены – кровеносный сосуд, по которому кровь движется к сердцу
*Капилляры – самым тонким сосудом в организме человека и других животных. он участвует в обмене веществ между кровью и тканями.
Основная и дополнительная литература по теме урока
- Биология. 5 – 6 класс. Линия жизни / В. В. Пасечник, С. В. Суматохин, Г. С. Калинова, Г. Г. Швецов, З. Г. Гапонюк. – М.: Просвещение, 2018.
- Биология в схемах и таблицах / А. Ю. Ионцева, А. В. Торгалов.
- Введение в биологию: Неживые тела. Организмы: учеб. Для уч — ся 5 – 6 кл. общеобразоват. учеб. заведений / А. И. Никишов. – М.: Гуманитар. изд. центр ВЛАДОС, 2012.
- Биология. Живой организм. 5 — 6 классы: учебник для общеобразовательных учреждений с приложением на электронном носителе / Л. Н. Сухорукова, В. С. Кучменко, И. Я. Колесникова. – М.: Просвещение, 2013.

- Биология. Обо всем живом. 5 класс: учебник / С. Н. Ловягин, А. А. Вахрушев, А. С. Раутиан. – М.: Баласс, 2014.
Теоретический материал для самостоятельного изучения
На сегодняшнем уроке мы продолжим изучение процессов жизнедеятельности живых организмов и познакомимся с тем, как осуществляется транспорт веществ у животных.
Вы уже знаете, что перенос веществ в организме – это жизненно-необходимый процесс. Если бы мы могли заглянуть внутрь живых организмов, то увидели бы следующее. В одноклеточных организмах животных (например, амеба, инфузория — туфелька) перемещение питательных веществ в клетке происходит за счет движения цитоплазмы. При этом у амебы происходит перекатывание цитоплазмы, а, следовательно, перемешивание питательных веществ. У инфузории — туфельки осуществляется круговое движение цитоплазмы, которое приводит к распределению веществ в клетке.
Многоклеточные животные для переноса веществ имеют особые системы органов.
У них перенос питательных веществ и газов выполняет кровь или гемолимфа, образуя особую систему – кровеносную. Она состоит из сердца и сосудов, по которым движется кровь. Например, дождевой червь имеет развитую кровеносную систему. Она состоит из сосудов, по которым движется кровь. Кровь – жидкость красного цвета, которая находится внутри кровеносных сосудов.
Она состоит из сердца и сосудов, по которым движется кровь. Например, дождевой червь имеет развитую кровеносную систему. Она состоит из сосудов, по которым движется кровь. Кровь – жидкость красного цвета, которая находится внутри кровеносных сосудов.
Кровь состоит из плазмы и клеток крови. Плазма – это бесцветная жидкость. Клетки крови делятся на красные – эритроциты, белые – лейкоциты и тромбоциты. Эритроциты придают крови красный цвет, так как в их состав входит особое вещество – пигмент гемоглобин. Соединяясь с кислородом, он разносит его по всему организму. Таким образом, осуществляя транспортную и дыхательную функции крови. Лейкоциты выполняют защитную функцию: они уничтожают попавшие в организм болезнетворные микроорганизмы. Тромбоциты участвуют в процессе свертывания крови, например, при ранении.
Примеры и разбор решения заданий тренировочного модуля
Задание 1. Установите последовательность движения крови по большому кругу кровообращения начиная с левого желудочка.
Варианты ответов:
правое предсердие
левый желудочек
аорта
артерия
капилляры
вена.
Правильный вариант ответа:
- левый желудочек
- аорта
- артерия
- капилляры
- правое предсердие
- вена.
Задание 2. Заполните пропуски в таблице «Строение пищеварительной системы».
Тип кровеносной системы | Особенности строения | Какая жидкость циркулирует | У каких животных имеется |
Незамкнутая кровеносная система | |||
Замкнутая кровеносная система |
Варианты ответов:
- Транспортная жидкость на определённых участках выходит из сосудов и попадает в полость тела. Потом она снова собирается в сосудах
- Транспортная жидкость циркулирует только по сосудам
- Гемолимфа
- Кровь
- Членистоногие
- Моллюски
- Кольчатые черви
- Позвоночные
Правильный вариант:
Тип кровеносной системы | Особенности строения | Какая жидкость циркулирует | У каких животных имеется |
Незамкнутая кровеносная система | Транспортная жидкость на определённых участках выходит из сосудов и попадает в полость тела. | Гемолимфа | Членистоногие, моллюски |
Замкнутая кровеносная система | Транспортная жидкость циркулирует только по сосудам | Кровь | Кольчатые черви Позвоночные |
 Потом она снова собирается в сосудах.
Потом она снова собирается в сосудах.Создан наноконвейер на основе белков-биомоторов
| Поделиться Ученым удалось установить, что молекула миозина-VI может работать, как самостоятельный наномотор-транспортер, и перевозить «на себе» грузы. На основе молекулы кинезина создан работающий конвейер, который передвигает макроскопические предметы. Практически во всех живых организмах находятся миниатюрные биомоторы — белки кинезин и миозин. Они отвечают за сокращение мышечных тканей, за транспортировку внутриклеточных грузов по микротрубкам, за деление клеток, а также за работу некоторых энзимов. Недавно ученые установили, что молекулы кинезина и миозина-VI могут быть полезны в исследованиях, проводимых в области наномашин. Более того, их можно «приручить» для приведения в движение таких сложных наносистем, как молекулярные конвейеры, транспортирующие молекулы, и даже живые клетки внутри биочипа-анализатора. Или же они могут служить основой двигательных систем нанороботов и микромашин, способных передвигаться в наноразмерном диапазоне.
Они отвечают за сокращение мышечных тканей, за транспортировку внутриклеточных грузов по микротрубкам, за деление клеток, а также за работу некоторых энзимов. Недавно ученые установили, что молекулы кинезина и миозина-VI могут быть полезны в исследованиях, проводимых в области наномашин. Более того, их можно «приручить» для приведения в движение таких сложных наносистем, как молекулярные конвейеры, транспортирующие молекулы, и даже живые клетки внутри биочипа-анализатора. Или же они могут служить основой двигательных систем нанороботов и микромашин, способных передвигаться в наноразмерном диапазоне.
Ученым из Иллинойского университета удалось доказать, что молекула миозина-VI может являться самостоятельным наномотором-транспортером, который сможет перевозить «на себе» грузы, сообщает NanotechBriefs.
Другая команда ученых из института им. Макса Планка создала на основе молекулы кинезина работающий конвейер, который передвигает макроскопические предметы на своей поверхности.
Молекула белка кинезина — один из известнейших биологических моторов. Это представитель класса линейных биомоторов. Среди механохимических преобразователей энергии, распространенных в живой природе, они играют важную роль. Эти белковые машины движутся вдоль полимерных нитей, используя в качестве «топлива» молекулы АТФ. К таким моторам относятся белки актомиозинового комплекса, входящего в состав сократительного аппарата мышц.
Движение микроворсинок (жгутиков и ресничек бактерий и простейших) определяется взаимодействием другой пары моторных белков — динеина и тубулина. Смещение головок динеина относительно тубулиновых микротрубочек белков обеспечивает волнообразные движения микроворсинок. Среди большого числа моторных белков миозин скелетных мышц и кинезин из клеток мозга являются наиболее изученными молекулярными моторами. Несмотря на то, что функции миозина и кинезина в клетке различаются, они удивительно похожи по своему строению и механизмам действия.
Молекула кинезина работает следующим образом. Совместно с микротрубками цитоскелета она выполняет транспорт веществ внутри клетки и перемещение везикул. Если провести аналогию с макромиром, то микротрубки играют роль рельсов, по которым перемещаются молекулы белков кинезина (вагоны), неся на себе полезный груз. Один конец этой молекулы прикрепляется к везикуле, которую необходимо транспортировать, а другой — к микротрубке, которая направляет движение.
Молекула кинезина представляет собой димер, образованный двумя одинаковыми полипептидными цепями. Подобно молекуле миозина, с одной стороны каждой полипептидной цепи кинезина формируется глобулярная головка, соединенная со сравнительно длинным хвостом. Линейные размеры головки сравнительно невелики, они составляют 7,5 x 4,5 x 4,5 нм. Длина молекулы — 50 нм. Хвосты двух мономерных цепей сплетены вместе, а наклоненные в разные стороны головки образуют своеобразную «рогатину», которая непосредственно взаимодействует с глобулярными мономерами микротрубочки, вдоль которой перемещается кинезин.
Молекула «шагает» вдоль микротрубки, делая 8-нанометровые шаги. На рисунке кинезиновый комплекс перемещает органеллу меланосому (ответственную за синтез меланина) вдоль микротрубки. Чтобы сделать такой «шаг», молекула использует в качестве топлива одну молекулу АТФ. При этом сила, развиваемая одной молекулой кинезина, составляет величину 6 пН. Если бы такой мощностью в расчете на единицу массы обладали автомобильные моторы, то они могли бы легко разгонять машины до скоростей, существенно превышающих скорость звука. Коэффициент полезного действия кинезинового мотора также велик — примерно 50%.
В процессе «ходьбы» молекула кинезина может расщепить за одну секунду до 100 молекул АТФ, переместившись на 800 нм. Работая в качестве индивидуального молекулярного извозчика, кинезин может совершать перемещения на очень большие расстояния (до 1 мм).
Почему администраторы не заметят миграцию данных в облака
Облака
Ученые из института им. Макса Планка попытались «приручить» кинезин вне клетки. Для этого они покрыли молекулами кинезина гладкую стеклянную поверхность, создав что-то вроде ковра, ворсинки которого представляли собой молекулы кинезина. Потом исследователи разместили на этой поверхности ряд микротрубок и микросфер. Исследователи добавили к среде раствор АТФ — и получился огромный «трубочный» конвейер. Если в клетке кинезин шагает вдоль микротрубки сам, то в искусственной системе молекула жестко закреплена, и свободные концы молекул «шагали» по микротрубкам, передвигая их.
Макса Планка попытались «приручить» кинезин вне клетки. Для этого они покрыли молекулами кинезина гладкую стеклянную поверхность, создав что-то вроде ковра, ворсинки которого представляли собой молекулы кинезина. Потом исследователи разместили на этой поверхности ряд микротрубок и микросфер. Исследователи добавили к среде раствор АТФ — и получился огромный «трубочный» конвейер. Если в клетке кинезин шагает вдоль микротрубки сам, то в искусственной системе молекула жестко закреплена, и свободные концы молекул «шагали» по микротрубкам, передвигая их.
Молекула миозина VI, напротив, — мономер, но ученые из Иллинойса считают, что полностью развернутая молекула этого энзима представляет собой димер. Это значит, что молекула может шагать по поверхности актина.
«Миозин VI — маленькая молекула, которая может превращать химическую энергию в механическое движение и таким образом перемещаться по поверхности филаментов другой макромолекулы — актина», — говорит доктор Пол Селвин (Paul Selvin), один из исследователей из Иллинойского университета. — Наши исследования показали, что молекула миозина-VI может быть не мономером, как это случается во всем миозиновых моторах, а димером. Это позволяет нам сконструировать транспортер грузов на его основе».
— Наши исследования показали, что молекула миозина-VI может быть не мономером, как это случается во всем миозиновых моторах, а димером. Это позволяет нам сконструировать транспортер грузов на его основе».
В 2004 году д-р Селвин и его коллеги создали на основе миозина-VI «ходока», который передвигался по актиновым филаментам с шагом в 60 нм. «В этом эксперименте две молекулы миозина VI были «сшиты» вместе, формируя димер, который перемещался по актиновой поверхности, — говорит д-р Селвин. — Однако теперь мы знаем, что для работоспособного нанодвигателя можно использовать всего одну молекулу миозина-VI в качестве димера».
В последних экспериментах ученые использовали флуоресцентную технику визуализации FIONA (Fluorescence Imaging with One Nanometer Accuracy), с помощью которой удалось отслеживать шаги миозинового «ходока» с точностью до 1,5 нм.
В дальнейшем обе команды планируют создать микрочипы со встроенными конвейерами, работающими в разных направлениях.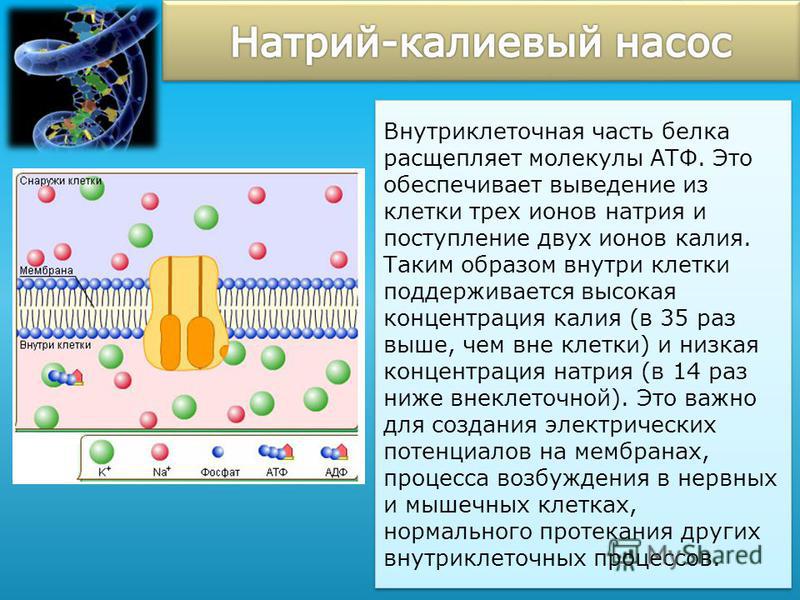 Это будет большим плюсом для будущих систем наносборки, так как НЭМС-конвейеры пока сделать достаточно трудно. Наверняка такие транспортные системы будут полезны и в «лабораториях на чипе», и в быстродействующих анализаторах.
Это будет большим плюсом для будущих систем наносборки, так как НЭМС-конвейеры пока сделать достаточно трудно. Наверняка такие транспортные системы будут полезны и в «лабораториях на чипе», и в быстродействующих анализаторах.
Технологии успешного SOC: тактика и стратегия сбора событий
БезопасностьPrimary Active Transport | Protocol (Translated to Russian)
5.11: Первичный активный транспорт
В отличие от пассивного транспорта, активный транспорт включает перемещение вещества через мембраны в направлении, противоположном его концентрации или электрохимическому градиенту. Существует два типа активного транспорта: первичный активный транспорт и вторичный активный транспорт. Первичный активный транспорт использует химическую энергию АТФ для работы белковых насосов, встроенных в клеточную мембрану. Используя энергию АТФ, насосы переносят ионы против их электрохимических градиентов — в направлении, в котором они обычно не перемещаются путем диффузии.
Используя энергию АТФ, насосы переносят ионы против их электрохимических градиентов — в направлении, в котором они обычно не перемещаются путем диффузии.
Взаимосвязь между концентрацией, электрическими и электрохимическими градиентами
Чтобы понять динамику активного переноса, важно сначала понять электрические градиенты и градиенты концентрации. Градиент концентрации — это разница в концентрации вещества на мембране или пространстве, которая движет движением из областей с высокой концентрацией в области с низкой концентрацией. Точно так же электрический градиент — это сила, возникающая в результате разницы между электрохимическими потенциалами на каждой стороне мембраны, которая приводит к перемещению ионов через мембрану до тех пор, пока заряды на обеих сторонах мембраны не станут одинаковыми. Электрохимический градиент создается, когда объединяются силы градиента химической концентрации и градиента электрического заряда.
Натриево-калиевый насос
Одним из важных переносчиков, отвечающих за поддержание электрохимического градиента в клетках, является натриево-калиевый насос. Первичная транспортная активность насоса происходит, когда он ориентирован так, что он охватывает мембрану с закрытой внеклеточной стороной, а его внутриклеточная область открыта и связана с молекулой АТФ. В этой конформации переносчик имеет высокое сродство к ионам натрия, обычно присутствующим в клетке в низких концентрациях, и три из этих иона входят в насос и присоединяются к нему. Такое связывание позволяет АТФ переносить одну из своих фосфатных групп на переносчик, обеспечивая энергию, необходимую для закрытия внутриклеточной стороны насоса и открытия внеклеточной области.
Первичная транспортная активность насоса происходит, когда он ориентирован так, что он охватывает мембрану с закрытой внеклеточной стороной, а его внутриклеточная область открыта и связана с молекулой АТФ. В этой конформации переносчик имеет высокое сродство к ионам натрия, обычно присутствующим в клетке в низких концентрациях, и три из этих иона входят в насос и присоединяются к нему. Такое связывание позволяет АТФ переносить одну из своих фосфатных групп на переносчик, обеспечивая энергию, необходимую для закрытия внутриклеточной стороны насоса и открытия внеклеточной области.
Изменение конформации снижает сродство насоса к ионам натрия, которые высвобождаются во внеклеточное пространство, но увеличивает его сродство к калию, позволяя ему связывать два иона калия, присутствующих в низкой концентрации во внеклеточной среде. Затем внеклеточная сторона насоса закрывается, и производная АТФ фосфатная группа на транспортере отсоединяется. Это позволяет новой молекуле АТФ связываться с внутриклеточной стороной насоса, которая открывается и позволяет ионам калия выходить в клетку, возвращая переносчик к его исходной форме, начиная цикл снова.
Из-за первичной транспортной активности насоса возникает дисбаланс в распределении ионов через мембрану. Внутри клетки больше ионов калия и больше ионов натрия вне клетки. Таким образом, внутренняя часть клеток оказывается более негативной, чем внешняя. Электрохимический градиент создается в результате ионного дисбаланса. Затем сила от электрохимического градиента запускает реакции вторичного активного транспорта. Вторичный активный транспорт, также известный как ко-транспорт, происходит, когда вещество транспортируется через мембрану в результате электрохимического градиента, установленного первичным активным транспортом, без необходимости дополнительного АТФ.
Литература для дополнительного чтения
Sahoo, Swagatika, Maike K. Aurich, Jon J. Jonsson, and Ines Thiele. “Membrane Transporters in a Human Genome-Scale Metabolic Knowledgebase and Their Implications for Disease.” Frontiers in Physiology 5 (March 11, 2014). [Source]
[Source]
Yang, Nicole J., and Marlon J. Hinner. “Getting Across the Cell Membrane: An Overview for Small Molecules, Peptides, and Proteins.” Methods in Molecular Biology (Clifton, N.J.) 1266 (2015): 29–53. [Source]
Клетка, транспорт воды и веществ |
Жизнедеятельность клетки связана с постоянным обменом ее содержимого с окружающей средой. Точно так же и внутри клетки происходит перемещение веществ между органеллами или компартментами. Все эти события связаны с преодолением основного барьера для веществ — мембраны, ограничивающей органеллу или саму клетку. При этом следует помнить, что главная функция биомембран — избирательность транспорта для различных — веществ и ионов.
Возможные способы транспорта через мембраны можно разделить на 4 основных типа: пассивная диффузия, облегченная диффузия, активный транспорт и цитозы.
Пассивная диффузия. Это процесс транспорта через мембраны веществ из области с большей их концентрацией в область с меньшей концентрацией (по химическому градиенту), в котором не принимают участия транспортные белки и не затрачивается энергия. С помощью такого способа через мембрану транспортируются малые незаряженные молекулы, например молекулы газов, некоторые анестезирующие вещества, а также вода. Чтобы пересечь бислой, молекула должна преодолеть поверхностное натяжение на границе мембраны, проникнуть в бислой, продиффундировать через него и выйти с противоположной стороны, вновь преодолев энергетический барьер на границе раздела фаз. Этим и объясняется избирательная проницаемость липидного бислоя для небольших молекул неэлектролитов. Удивительным является факт весьма легкого и быстрого проникновения воды через мембраны: показано, что молекуле воды требуется для пересечения бислоя всего 1 мкс. Для объяснения этого феномена в последнее время появляются основанные на некоторых экспериментальных данных предположения о том, что в мембранах все же существуют какие-то белковые проводящие пути для воды, либо молекулы воды пользуются локальными дефектами в структуре бислоев.
С помощью такого способа через мембрану транспортируются малые незаряженные молекулы, например молекулы газов, некоторые анестезирующие вещества, а также вода. Чтобы пересечь бислой, молекула должна преодолеть поверхностное натяжение на границе мембраны, проникнуть в бислой, продиффундировать через него и выйти с противоположной стороны, вновь преодолев энергетический барьер на границе раздела фаз. Этим и объясняется избирательная проницаемость липидного бислоя для небольших молекул неэлектролитов. Удивительным является факт весьма легкого и быстрого проникновения воды через мембраны: показано, что молекуле воды требуется для пересечения бислоя всего 1 мкс. Для объяснения этого феномена в последнее время появляются основанные на некоторых экспериментальных данных предположения о том, что в мембранах все же существуют какие-то белковые проводящие пути для воды, либо молекулы воды пользуются локальными дефектами в структуре бислоев.
Перемещение одних только молекул воды через полупроницаемую мембрану можно рассматривать как частный вид диффузии — осмос. Под осмосом понимают переход молекул воды из области с высоким водным потенциалом и низкой концентрацией растворенного вещества в область с низким водным потенциалом и высокой концентрацией растворенного вещества (рис. 4.9). В этом случае молекулы воды будут переходить из гипотонического раствора в гипертонический до тех пор, пока не наступит равновесие и оба раствора не станут изотоническими по отношению друг к другу.
Под осмосом понимают переход молекул воды из области с высоким водным потенциалом и низкой концентрацией растворенного вещества в область с низким водным потенциалом и высокой концентрацией растворенного вещества (рис. 4.9). В этом случае молекулы воды будут переходить из гипотонического раствора в гипертонический до тех пор, пока не наступит равновесие и оба раствора не станут изотоническими по отношению друг к другу.
Чтобы обозначить величину уменьшения водного потенциала, вызванного присутствием растворенных веществ, используют термин «осмотическое давление». Под осмотическим давлением понимают давление, которое следует приложить к раствору, чтобы остановить осмотическое поступление воды в него через полупроницаемую мембрану. Повышение концентрации растворенного вещества увеличивает осмотическое давление и уменьшает водный потенциал раствора.
Перемещение воды через плазматические мембраны клеток в соответствии с законами осмоса создает организмам немалые проблемы, особенно для водных обитателей. Поэтому осморегуляция (поддержание водного потенциала в клетке на постоянном уровне) является важной стороной функциональной деятельности большинства организмов, и на ее осуществление зачастую тратится значительная доля запасенной клеткой энергии.
Поэтому осморегуляция (поддержание водного потенциала в клетке на постоянном уровне) является важной стороной функциональной деятельности большинства организмов, и на ее осуществление зачастую тратится значительная доля запасенной клеткой энергии.
Скорость диффузии веществ через мембраны зависит от многих причин: растворимости вещества в мембране, коэффициента диффузии в мембране, а также разности концентрации вещества снаружи и внутри клетки (градиента концентрации) (рис. 4.10).
Облегченная диффузия. Этот вид транспорта осуществляется с помощью транспортных белков по электрохимическому градиенту (разность электрических потенциалов и концентраций веществ) без затрат энергии. Это селективный перенос веществ — вещество будет транспортировано через мембрану лишь в том случае, если для него в мембране имеется функционирующий транспортный белок.
Поскольку в облегченной диффузии задействованы белки, этот процесс, в отличие от пассивной диффузии, может достигать эффекта насыщения. Стадия насыщения (рис. 4.10) характеризует состояние, когда все транспортные белки для данного вещества насыщены субстратом и скорость транспорта этого вещества достигает максимума.
Стадия насыщения (рис. 4.10) характеризует состояние, когда все транспортные белки для данного вещества насыщены субстратом и скорость транспорта этого вещества достигает максимума.
С помощью облегченной диффузии через мембрану транспортируются многие вещества, в том числе гидрофильные молекулы: углеводы, аминокислоты, нуклеотиды, различные ионы и др. При этом скорость транспорта значительно превышает скорость пассивной диффузии (рис.4.10). Принципиально возможны два пути переноса веществ и ионов через мембрану: с помощью переносчиков и каналов. Поскольку трансмембранное перемещение белков в биомембранах не обнаружено, предложена модель, описывающая работу переносчиков — механизм «пинг-понг». Согласно этому механизму, транспорт веществ связан с конформационными изменениями в структуре белка-переносчика, которые индуцируются связыванием транспортируемого вещества (рис. 4.11).
Работу каналов можно рассмотреть на примере ацетилхолинового рецептора. Этот интегральный белок находится в основном в мембранах нервномышечных соеди-нений скелетных мышц. Он состоит из пяти субъединиц четырех типов и открывается в ответ на связывание ацетилхолина (нейромедиатор). При взаимодействии с ацетилхолином канал открывается, что связано с изменением конформации субъединиц, и пропускает определенные ионы (Na+ , K+ , Ca2+ и некоторые другие), остается в таком положении 1 мс, а затем закрывается.
Он состоит из пяти субъединиц четырех типов и открывается в ответ на связывание ацетилхолина (нейромедиатор). При взаимодействии с ацетилхолином канал открывается, что связано с изменением конформации субъединиц, и пропускает определенные ионы (Na+ , K+ , Ca2+ и некоторые другие), остается в таком положении 1 мс, а затем закрывается.
Селективное перемещение катионов изменяет трансмембранный потенциал, в результате чего происходит электрическое возбуждение мышечной клетки, что приводит к сокращению мышцы. Изучение структуры ацетилхолинового рецептора показало, что пять белковых субъединиц встроены в бислой определенным образом: они организованы вокруг центральной поры диаметром 3 нм, через которую и транспортируются катионы. Непроницаемость канала для анионов и в три раза большую проницаемость для катионов, чем для незаряженных молекул, можно объяснить электростатическими взаимодействиями, возникающими благодаря присутствию в воротах канала биполярных или отрицательно заряженных групп.
Особым типом транспорта веществ в ходе облегченной диффузии является использование ионофоров, действие которых изучено на искусственных мембранах.
Под ионофорами понимают низкомолекулярные вещества пептидной природы, избирательно транспортирующие через мембраны ионы. Различают ионофорыканалообразователи (грамицидин А, амфотерицин B и др.) и ионофорыпереносчики (валиномицин, энниатины, боверицин).
Валиномицин представляет собой антибиотик депсипептидной природы, организованный по типу ионной «ловушки». В неполярных растворителях конформация валиномицина напоминает собой браслет, внутренняя полость которого точно подогнана под размеры ионов калия. Внешняя сфера валиномицина гидрофобна, в результате чего он способен перемещаться в липидном бислое и транспортировать через него ионы.
Хорошо изученным примером ионофоровканалообразователей служит грамицидин А. Это антибиотик пептидной природы, состоящий из 15 аминокислот. Две молекулы грамицидина могут пронизывать мембрану в виде двойной спирали или образуя димер «голова к голове». В таких конформациях молекулы грамицидина А формируют полый цилиндр, по которому могут перемещаться ионы металлов.
В таких конформациях молекулы грамицидина А формируют полый цилиндр, по которому могут перемещаться ионы металлов.
В биологических мембранах ионофорный тип транспорта до сих пор не обнаружен.
Активный транспорт. Это сопряженный с потреблением энергии перенос молекул или ионов через мембрану против электрохимического градиента, в котором задействованы транспортные белки. Благодаря активному транспорту в жизнеспособных клетках между двумя сторонами мембраны поддерживается разность потенциалов, т. е. электрический заряд, при этом у большинства изученных клеток внутреннее содержимое заряжено отрицательно по отношению к внешней среде.
Активный транспорт сопряжен со значительными затратами энергии: некоторые клетки тратят более трети всей запасенной энергии для создания ионного градиента на мембране. Это необходимо для таких жизненно важных процессов, как осморегуляция, генерация и передача нервных импульсов, перенос в клетки питательных веществ (сахаров, аминокислот и др. ).
).
Разнообразные системы активного транспорта отличаются друг от друга, в первую очередь тем, что служит для них источником энергии: АТР, ионный градиент, фосфоенолпируват, видимый свет. Наиболее хорошо изученной системой активного транспорта является натрийкалиевая (Na+/K+)АТРаза, функционирующая в плазматических мембранах животных клеток. Этот интегральный белок состоит из двух субъединиц: бульшая представлена полипептидом, имеющим участки связывания для ионов натрия и АТР на цито плазматической поверхности, а ионов калия — на наружной; меньшая субъединица является гликопротеином. Работа (Na+/K+)АТРазы заключается в следующем: при гидролизе одной молекулы АТР из клетки выкачивается 3 иона Na+ , а извне в клетку проводится 2 иона K+ , т. е. выводится больше положительных ионов, чем проводится внутрь клетки. Так на внутренней стороне мембраны создается избыточный отрицательный заряд, и клетка становится электрогенной. В мембранах обычно присутствуют проводящие пути для облегченной диффузии ионов натрия и калия по электрохимическому градиенту, и этот транспорт, хотя и с малой скоростью, со-вершается. Однако в жизнеспособной клетке не происходит уравнивания концентраций ионов, создающих электрохимический градиент на мембране, благодаря постоянной работе первичных активных переносчиков, таких, как (Na+/K+)АТРаза. Таким образом, ионные насосы, принимающие участие в первичном активном транспорте, осуществляют перемещение заряда на мембране и создают на ней электрохимический градиент, в котором заключена энергия.
Однако в жизнеспособной клетке не происходит уравнивания концентраций ионов, создающих электрохимический градиент на мембране, благодаря постоянной работе первичных активных переносчиков, таких, как (Na+/K+)АТРаза. Таким образом, ионные насосы, принимающие участие в первичном активном транспорте, осуществляют перемещение заряда на мембране и создают на ней электрохимический градиент, в котором заключена энергия.
Вторичные активные переносчики используют электрохимические градиенты в качестве движущей силы для транспорта растворимых веществ. Этот процесс можно проследить на примере клеток эпителия кишечника. Образуемые в кишечнике при переваривании пищи строительные блоки (аминокислоты, глюкоза и др.) поступают в кровь при диффузии через мембраны кровеносных сосудов, и эта диффузия осуществляется в ходе симпорта(однонаправленного транспорта) с ионами натрия.
Ионы натрия стремятся возвратиться в клетку согласно закономерностям облегченной диффузии и как бы тянут с собой молекулы питательных веществ. В мембранах обнаружены специфические переносчики сахаров и разных аминокислот, которые функционируют в системе активного транспорта, накапливая в клетке эти вещества, извлекая их даже из очень разбавленных растворов, т. е. против химического градиента. Эти же транспортные системы могут участвовать и в облегченной диффузии, если вещества транспортируются по химическому градиенту. Кроме описанного выше примера симпорта питательных веществ вместе с возвращающимися в клетку ионами натрия, существует и разнонаправленный транспорт — антипорт. Например, белок полосы 3 эритроцитов осуществляет сопряженный транспорт Cl и HCO3 в противоположных направлениях через эритроцитарную мембрану.
В мембранах обнаружены специфические переносчики сахаров и разных аминокислот, которые функционируют в системе активного транспорта, накапливая в клетке эти вещества, извлекая их даже из очень разбавленных растворов, т. е. против химического градиента. Эти же транспортные системы могут участвовать и в облегченной диффузии, если вещества транспортируются по химическому градиенту. Кроме описанного выше примера симпорта питательных веществ вместе с возвращающимися в клетку ионами натрия, существует и разнонаправленный транспорт — антипорт. Например, белок полосы 3 эритроцитов осуществляет сопряженный транспорт Cl и HCO3 в противоположных направлениях через эритроцитарную мембрану.
У аэробных бактерий транспорт питательных веществ в клетку осуществляется в ходе симпорта не с ионами Na+ , а с протонами. Наилучшим образом охарактеризованным примером подобного переносчика служит лактозопермеаза кишечной палочки. Этот интегральный белок использует протонный электрохимический градиент, созданный на мембране в результате окислительного фосфорилирования, для симпорта лактозы: с каждым возвращенным в клетку протоном переносится одна молекула лактозы.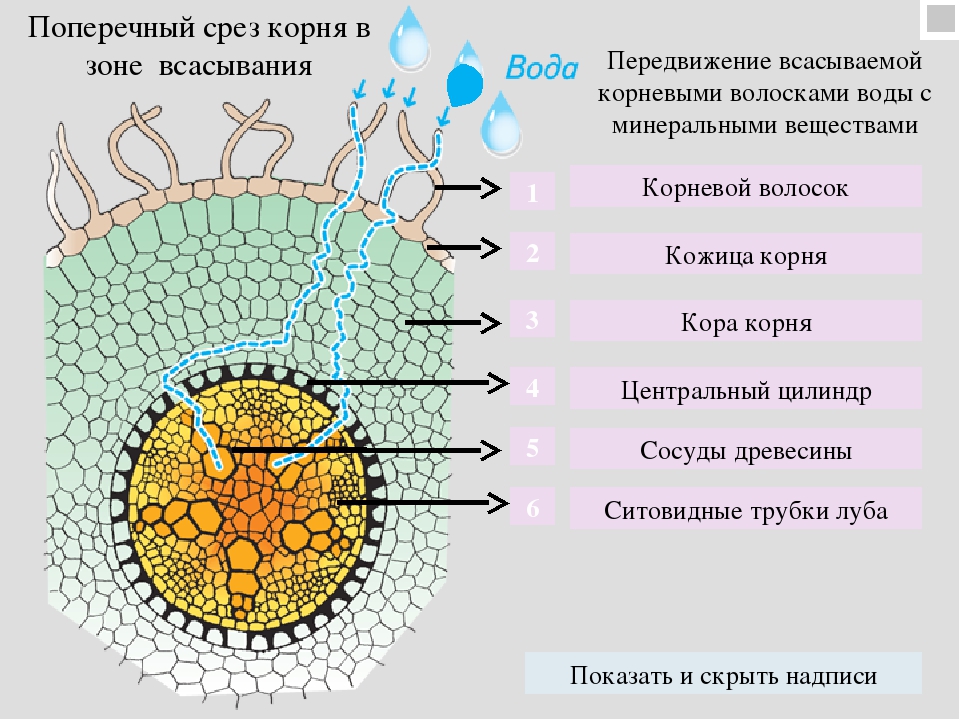
Следует отметить, что АТРазы представляют собой ферменты, катализирующие взаимообратимые реакции: при гидролизе АТР ионы транспортируются против электрохимического градиента, а перенос ионов по электрохимическому градиенту через каналы АТРазы может запускать синтез АТР.
Эндоцитоз и экзоцитоз. Эти способы переноса веществ через мембраны связаны с образованием впячиваний (инвагинаций) мембраны и формированием особых мембранных везикул, обеспечивающих прохождение через мембрану крупных макромолекул и частиц. При этом эндоцитоз обеспечивает поглощение клеткой веществ, а экзоцитоз — выделение из клетки. Принято делить цитозы еще на два типа: пиноцитоз и фагоцитоз. Пиноцитоз — это механизм, с помощью которого через мембрану проводятся белки и другие макромолекулы в жидкой фазе.
Фагоцитоз представляет собой поглощение клеткой крупных частиц, например бактерий, вирусов. Эти виды транспорта характерны в основном для эукариотических клеток, причем у животных фагоцитоз осуществляют только специализированные клетки, такие, например, как макрофаги. Для многих простейших, например амеб, фагоцитоз является основным способом питания.
Для многих простейших, например амеб, фагоцитоз является основным способом питания.
Важной особенностью цитозов является последовательное образование и слияние везикул, в которых заключено транспортируемое вещество, причем секретируемые и поглащаемые молекулы локализуются в везикулах и не смешиваются с другими макромолекулами или органеллами клетки. С помощью не установленного пока меха-низма каждый пузырек сливается только со специфическими мембранными структурами (рис. 4.12).
В основе цитозов лежит еще одно характерное свойство липидных слоев биомембран — способность к агрегации, в результате чего мелкие везикулы объединяются в более крупные или происходит объединение везикул с плазматической мембраной клетки. Такой механизм основан на универсальности структуры биомембран, участвующих в формировании клеточных органелл и протопластов. Аналогичное явление можно наблюдать в пенах, где мыльные пузыри, также состоящие из амфифильных молекул (мыла — соли жирных кислот), обладают тенденцией к объединению с образованием более крупных структур.
Способность мембран к агрегации лежит в основе такого широко распространенного способа переноса генетической информации, как слияние протопластов.
Скорость цитозов удивительно высокая. Показано, что клетки печени поглощают путем эндоцитоза за 1 ч количество жидкости, составляющее не менее 20% их объема, и количество мембранного материала, по площади превышающее в пять раз площадь их плазматической мембраны. Сходство цитозов с другими способами транспорта веществ через биомембраны состоит в том, что переносимое вещество должно быть «узнано» мембранными компонентами, иными словами, и в этом случае проявляется избирательная проницаемость мембран для различных соединений.
Тема урока: «Плазматическая мембрана. Транспорт веществ»
Планируемые результаты: Учащиеся систематизируют и углубляют знания о строении эукариотической клетки
Личностная значимость изучаемого для школьников: Животная клетка покрыта плазматической мембраной, поэтому необходимо представлять строение мембраны, чтобы изучить транспорт веществ через эту мембрану
Задачи урока:
- Образовательные: познакомить учащихся со строение и функциями плазматической мембраны
- Развивающие: развивать умения сравнивать строение клеточных структур, оценивать, объяснять причины сходства и различия
- Воспитывающие: воспитывать дух соревнования, коллективизма, точность и быстроту ответов; осуществлять эстетическое воспитание
Оборудование: компьютер, лабораторное оборудование, раздаточный материал, телевизор и видеомагнитофон, к\ф “Строение клетки”, СD-диск “Кирилл и Мефодий” 8 класс, наглядный материал исследовательской работы учеников
Тип урока: по дидактическим целям: изучения нового материала; по методам обучения: частично-поисковый, исследовательский
Формы работы: групповая, индивидуальная
Ход урока
Учитель читает:
Жизнь на Земле – это способ существования
материи в виде открытых систем, включая в себя
липиды, белки и нуклеиновые кислоты и способных к
самосохранению и саморегуляции и
самовоспроизведению; это сопряженная циркуляция
определенных химических элементов в водной
среде; направляемая энергией солнца в сторону
упорядочения и усложнения.
Учитель: Это современное представление о клетках появившиеся не так давно в науке и резко, отличающееся от теории Шванна подходит к теме нашего урока.
Наша цель: Рассмотреть особенности строения плазматической мембраны и определить особенности транспорта через нее веществ.
Учитель: Поработаем с терминами:
- гидрофильные вещества
- гидрофобные вещества
- макроэлементы
- биолэлементы
- микроэлементы
- денатурация
- ренатурация
- липиды
Учитель: Клеточная мембрана играет важную роль в клетке, т. к. она выполняет следующие функции:
- Отделяет клеточное содержимое от внешней среды
- Регулирует обмен между клеткой и внешней средой
- Делит клетки на отсеки
- На мембране некоторых органоидов происходят химические реакции
- На мембране располагаются и рецепторные
участки для распознавания внешних стимулов
(гормонов или других химических веществ),
поступающих из окружающей среды или из другой
части самого организма.

Учитель: В конце XVIII века стало известно, что клеточная мембрана ведет себя не как полупроницаемая мембрана, она обладает избирательной проницаемостью. Не пропускает, например аминокислоты и глюкозу (а они просто необходимы клетке). Сама мембрана активно регулирует этот процесс — одни вещества пропускает, а другие нет.
Ранние работы по проницаемости мембран показали, что органические растворители, например спирт, эфир проникают через мембрану быстрее, чем вода. Это свидетельствует о том, что в мембранах есть липиды, позже данные химического анализа подтвердили это, также как и то, что в мембранах есть белки.
Ученик: рассказывает о строении белков
Учитель: липиды подразделяются на фосфолипиды, гликолипиды и стиролы (ученики характеризуют особенности их строение)
Учитель: Группа учащихся занимается
проблемами биохимии клетки и в КГУ ведет
научно-исследовательскую работу, в частности они
проводили анализ фосфолипидного слоя.
Ученик: Если на поверхности воды распределить тонким слоем фосфолипиды, то их молекулы будут ориентированны таким образом, чтобы образовать монослой. Если количество липидов увеличить и встряхнуть, то образующиеся частицы будут называться мицеллами. В них гидрофобные хвосты внутри, а гидрофильные головы снаружи. Образуется билипидный слой. Такой билипидный слой входит в состав плазматической мембраны.
(Учащиеся показывают опыт, в мерный стакан наливают воды и на поверхность наливают оливковое масло, которое по своему составу очень походе на фосфолипиды, затем добавляют масло и встряхивают, показывая как образуются мицеллы. На доске ученик показывает схему образования мицеллы. В мицелле гидрофобные хвосты будут упорядочены внутри и тем самым защищены от контакта с водой.)
Учитель: Давайте представим на несколько
минут себя, не в обычном классе на уроке, а в
аудитории крупного вуза, где собрались ученые и
их ученики, занимающиеся проблемами строения
мембран и транспорта веществ.
Ученики: Давсон и Даниэли (1935 г.) высказали предположение, что билипидный слой окружен 2-я слоями белка
Робертсон (1959 г.) выдвигает гипотезу о строении элементарных мембран и вводит постулат, что мембрана трехслойна
Учитель: Эта гипотеза претерпела изменения, т. к. появились данные из разных источников, например при проведении метода скалывания. Клетка при мгновенном замораживании остается такой же как в живом виде и при скалывании было обнаружено, что в бислое липидов есть погруженные белки или пронизывающие этот бислой насквозь.
Ученик: Сингер и Николсон (1972) предложили жидкостно-мозаичную модель мембраны согласно которой, белковые молекулы, плавающие в жидком билипидном слое, образуют как бы своеобразную мозайку.
Учитель: Предполагается, что это
динамическая структура. Белки в билипидном слое
плавают подобно островам, а иногда их держат на
привязи нити цитоскелета, липиды также могут
менять свое положение. В мембране клетки есть
большое количество белков, но их все можно
разделить на группы из-за выполняемых функций.
В мембране клетки есть
большое количество белков, но их все можно
разделить на группы из-за выполняемых функций.
Например: есть структурные белки, есть белки переносчики транспортирующие через мембрану вещества, есть ферментативные белки, есть белки гликопротеины, их разветвленные антенны возвышаются над мембраной и за счет них идет распознавание других клеток, их правильная дифференцировка. Липиды находятся в жидком состоянии и похожи на оливковое масло.
Учитель: Перед организмами стоит серьезная проблема – транспорт веществ через клеточные мембраны, хотя толщина и небольшая, но они являются барьером для ионов и молекул (т. к. неполярные липиды мембраны отталкивают эти вещества)
Транспорт важен по ряду причин:
- Поддержание соответствующего Рн
- Поддержание определенной концентрации
- Поступление веществ является источником энергии
- Выведение токсических отходов, секреция полезных веществ
- Создание ионных градиентов необходимых для нервной и мышечной активности
Существует четыре основных механизма для поступления веществ в клетку или выхода из клетки наружу:
Разберем пассивный транспорт который идет по градиенту концентраций:
1. Диффузия – это движение молекул или ионов
из области с высокой концентрацией в область с
более низкой концентрацией по градиенту.
Диффузия – это движение молекул или ионов
из области с высокой концентрацией в область с
более низкой концентрацией по градиенту.
(Учитель может провести опыт, используя графопроектор, чашку Петри и КМпО4. В чашку Петри наливают воды и помещают несколько кристаллов КМпО4. Через некоторое время на экране мы видим, как кристаллы начинают растворяться, и происходит распространение вещества. Можно использовать холодную и горячую воду для опыта, тогда есть возможность сравнить скорость диффузии. )
Если к раствору медного купороса осторожно прилить сверху раствор воды, то через некоторое время бесцветный верхний слой воды окрасится в синий цвет, т. к. идет диффузия снизу молекул медного купороса. А если на пути поставить преграду?
2. Осмос – это движения растворителя из области высокой концентрации в область более низкой концентрации.
Учитель: В 1837 году французский ученый Дютроше
провел следующий опыт. Он взял животный пузырь и
наполнил его раствором сахара и опустил в воду.
Начало происходило засасывание воды в пузырь, и
он раздувался, раствор начинает подниматься
вверх по стеклянной трубке. Если отводной
стеклянной трубки не было, то мешок разрывался,
через некоторое время пузырь спадает, т. к.
сахароза начинает диффундировать. Такой прибор
Дютроше впоследствии получил названия
осмометра. (Учителю можно этот прибор
сконструировать и показать процесс осмоса. )
Он взял животный пузырь и
наполнил его раствором сахара и опустил в воду.
Начало происходило засасывание воды в пузырь, и
он раздувался, раствор начинает подниматься
вверх по стеклянной трубке. Если отводной
стеклянной трубки не было, то мешок разрывался,
через некоторое время пузырь спадает, т. к.
сахароза начинает диффундировать. Такой прибор
Дютроше впоследствии получил названия
осмометра. (Учителю можно этот прибор
сконструировать и показать процесс осмоса. )
Впоследствии опыт Дютроше модифицировали.
Пробочным сверлом делают отверстие в моркови и
наливают в это отверстие сахарный сироп
(подкрашенный свекольным соком) или вишневый, а в
углубление другого корнеплода – воду. Закрывают
оба отверстия пробкой со стеклянной трубкой.
Красители использовать нельзя, т. к. они убивают
живые клетки. Корни моркови снизу подрезают,
чтобы обнажить внутрении клетки. Первый
корнеплод помещают в стакан с водой и по трубке
поднимается сахарный сироп, а второй корнеплод
помещают в стакан с сиропом и из трубки вода
опускается.
Учитель: Использование этих видов транспорта в природе.
- подвядание листьев
- внесение избытка удобрений в цветы
- при борьбе с сорняками, когда междурядья поливают поваренной солью
Активный транспорт
3. Экзоцитоз (это движение веществ из клетки) и эндоцитоз (это движение веществ в клетку).
Активный транспорт идет с затратами энергии и против градиента концентрации, т. е. против естественной диффузии. Процесс экзо- и эндоцитоза распространен у простейших.
(Для изучения этого вида транспорта используется программа СD – диска “Кирилл и Мефодий. Биология 7 класс)
Учитель: Есть еще один вид активного транспорта это К +— Nа+ насос. Для изучения этого вида транспорта необходимо прочитать предложенный текст, написать конспект. (текст прилагается)
Закрепление:
Учащимся предлагается к/ф который
воспроизводиться без звука и который необходимо
озвучить, вся работа завершается тестом.
Тест:
1. Назовите химические соединения, которые мозаично расположены в наружной плазматической мембране и обеспечивают выполнение мембраной транспортной, ферментативной и рецепторной функций:
п. белки
б. полисахариды
к. липиды
г. ДНК
л. РНК
2. Липиды в клеточной мембране расположены послойно. Сколько таких липидных слоев содержится в мембране:
а. 1
р. 2
в. 3
б. 4
3. Назовите химические соединения, которые входят в состав наружной плазматической мембраны и, обладая гидрофобностью, служат основным барьером для проникновения в клетку воды и гидрофильных соединений:
с. белки
б. полисахариды
о. липиды
г. РНК
а. ДНК
4. Теплокровные и холоднокровные животные существенно различаются по составу жирных кислот в их клетках, в том числе в клеточных мембранах. Назовите вид жирных кислот, которых будет больше в клетках теплокровных животных, чем в клетках холоднокровных:
н. насыщенные
насыщенные
к. ненасыщенные
5. При возбуждении клетки ионы натрия и калия быстро перемещаются через наружную плазматическую мембрану клетки: ионы натрия – внутрь клетки, а ионы калия – наружу. Назовите вид транспорта этих ионов во время возбуждения.
к. осмос
и. диффузия
в. пиноцитоз
е. активный транспорт
д. фагоцитоз
6. Назовите процесс, посредством которого в желудке млекопитающего осуществляется секреция соляной кислоты из секреторных клеток слизистой оболочки в полость желудка:
а. диффузия
в. облегченная диффузия
к. активный транспорт
г. пиноцитоз
и. фагоцитоз
е. осмос
7. Концентрация раствора поваренной соли равна 0,5%. Клетки человека поместили в этот раствор. Что будет происходить с молекулами воды?
н. будет перемещаться преимущественно в клетку
б. будет перемещаться преимущественно из клетки
к. перемещаться не будет
перемещаться не будет
г. будут в равном количестве перемещаться в обе стороны: в клетку и из клетки
8. В ряде случаев молекулы растворенного вещества попадают в клетку, находясь в составе капли жидкости внутри пузырьков, которые образовались после впячивания и последующего отшнуровывания плазматической мембраны. Назовите этот вид транспорта веществ через наружную плазматическую мембрану:
а. диффузия
и. облегченная диффузия
в. активный транспорт
о. пиноцитоз
д. фагоцитоз
ж. осмос
9. Предположим, что плазматическая мембрана проницаема для растворенного в воде химического соединения, причем оно находится в разных концентрациях по разные стороны мембраны. В какую сторону будут перемещаться молекулы этого вещества:
б. в сторону большей концентрации
в. в сторону меньшей концентрации
10. Назовите процесс, посредством которого происходит движение ионов натрия из межклеточной жидкости в нервную клетку в момент возбуждения:
е. диффузия
диффузия
к. облегченная диффузия
в. активный транспорт
ж. пиноцитоз
д. фагоцитоз
з. осмос
11. Какие по растворимости в воде участки молекул фосфолипидов расположены в наружной плазматической мембране на поверхности, обращенной в сторону цитоплазмы клеток:
в. гидрофильные
12. н. гидрофобные
В какую сторону перемещаются молекулы растворенного вещества в процессе диффузии:
о. в сторону большей концентрации растворенного вещества
и. в сторону меньшей концентрации растворенного вещества
13. Назовите процесс посредством которого дизентерийная амеба поглощает эритроциты человека:
о. диффузия
б. облегченная диффузия
к. активный транспорт
г. пиноцитоз
е. фагоцитоз
ключевое слово: проникновение
В конце урока, свои впечатления учащиеся выражают в виде синквейна.
Синквейн:
- Транспорт
- Пассивный, активный
- Отделять, регулировать, делить
- Зачем он нужен клетке?
- Жизнь.

Презентация
Пассивный и активный транспорт — The Biology Primer
Активный транспорт
Активный транспорт — это движение молекул через клеточную мембрану в направлении против градиента их концентрации, от низкой до высокой. Активный транспорт обычно связан с накоплением высоких концентраций молекул, в которых нуждается клетка, таких как ионы, глюкоза и аминокислоты. Если в процессе используется химическая энергия, например, из аденозинтрифосфата (АТФ), это называется первичным активным транспортом.Вторичный активный транспорт предполагает использование электрохимического градиента. Активный транспорт использует клеточную энергию, в отличие от пассивного транспорта, который не использует клеточную энергию. Активный транспорт — хороший пример процесса, для которого клеткам требуется энергия.
Почему клетки такие маленькие?
Клетки настолько малы, что для их исследования понадобится микроскоп. Почему? Чтобы ответить на этот вопрос, мы должны понять, что для выживания клетки должны постоянно взаимодействовать с окружающей их средой.Растворенные в воде газы и молекулы пищи должны абсорбироваться, а продукты жизнедеятельности должны удаляться. Для большинства клеток прохождение всех материалов внутрь и из клетки должно происходить через плазматическую мембрану. Каждая внутренняя область клетки должна обслуживаться частью клеточной поверхности. По мере роста клетки увеличивается ее внутренний объем и расширяется клеточная мембрана. К сожалению, объем увеличивается быстрее, чем площадь поверхности, и поэтому относительная площадь поверхности, доступная для пропускания материалов в единицу объема ячейки, неуклонно уменьшается.Наконец, в какой-то момент поверхности будет достаточно, чтобы обслужить весь интерьер; чтобы выжить, клетка должна перестать расти. Важным моментом является то, что отношение площади поверхности к объему становится меньше по мере увеличения ячейки. Таким образом, если клетка вырастет за определенный предел, недостаточное количество материала сможет пересечь мембрану достаточно быстро, чтобы приспособиться к увеличенному клеточному объему.
Почему? Чтобы ответить на этот вопрос, мы должны понять, что для выживания клетки должны постоянно взаимодействовать с окружающей их средой.Растворенные в воде газы и молекулы пищи должны абсорбироваться, а продукты жизнедеятельности должны удаляться. Для большинства клеток прохождение всех материалов внутрь и из клетки должно происходить через плазматическую мембрану. Каждая внутренняя область клетки должна обслуживаться частью клеточной поверхности. По мере роста клетки увеличивается ее внутренний объем и расширяется клеточная мембрана. К сожалению, объем увеличивается быстрее, чем площадь поверхности, и поэтому относительная площадь поверхности, доступная для пропускания материалов в единицу объема ячейки, неуклонно уменьшается.Наконец, в какой-то момент поверхности будет достаточно, чтобы обслужить весь интерьер; чтобы выжить, клетка должна перестать расти. Важным моментом является то, что отношение площади поверхности к объему становится меньше по мере увеличения ячейки. Таким образом, если клетка вырастет за определенный предел, недостаточное количество материала сможет пересечь мембрану достаточно быстро, чтобы приспособиться к увеличенному клеточному объему. Когда это происходит, клетка должна делиться на более мелкие клетки с подходящим соотношением площадь поверхности / объем или перестать функционировать.Вот почему клетки такие маленькие.
Когда это происходит, клетка должна делиться на более мелкие клетки с подходящим соотношением площадь поверхности / объем или перестать функционировать.Вот почему клетки такие маленькие.
3.5 Пассивный транспорт — Концепции биологии — 1-е канадское издание
К концу этого раздела вы сможете:
- Объясните, почему и как происходит пассивный перенос
- Понимать процессы осмоса и диффузии
- Определите тонус и опишите его отношение к пассивному транспорту
Плазменные мембраны должны позволять определенным веществам входить в клетку и выходить из нее, при этом предотвращая попадание вредных материалов и выход основных материалов.Другими словами, плазматические мембраны избирательно проницаемы — они пропускают одни вещества, но не пропускают другие. Если бы они потеряли эту избирательность, клетка больше не могла бы поддерживать себя и была бы разрушена. Некоторым клеткам требуется большее количество определенных веществ, чем другим клеткам; у них должен быть способ получения этих материалов из внеклеточных жидкостей. Это может происходить пассивно, поскольку определенные материалы движутся вперед и назад, или в ячейке могут быть специальные механизмы, обеспечивающие транспортировку.Большинство клеток тратят большую часть своей энергии в форме аденозинтрифосфата (АТФ) на создание и поддержание неравномерного распределения ионов на противоположных сторонах их мембран. Структура плазматической мембраны способствует этим функциям, но также создает некоторые проблемы.
Это может происходить пассивно, поскольку определенные материалы движутся вперед и назад, или в ячейке могут быть специальные механизмы, обеспечивающие транспортировку.Большинство клеток тратят большую часть своей энергии в форме аденозинтрифосфата (АТФ) на создание и поддержание неравномерного распределения ионов на противоположных сторонах их мембран. Структура плазматической мембраны способствует этим функциям, но также создает некоторые проблемы.
Наиболее прямые формы мембранного транспорта пассивны. Пассивный перенос — это естественное явление и не требует от клетки затрат энергии для выполнения движения. При пассивном переносе вещества перемещаются из области с более высокой концентрацией в область с более низкой концентрацией в процессе, называемом диффузия .Говорят, что физическое пространство, в котором имеется различная концентрация одного вещества, имеет градиент концентрации.
Плазменные мембраны асимметричны, что означает, что, несмотря на зеркальное отображение, образованное фосфолипидами, внутренняя часть мембраны не идентична внешней стороне мембраны. Интегральные белки, которые действуют как каналы или насосы, работают в одном направлении. Углеводы, присоединенные к липидам или белкам, также находятся на внешней поверхности плазматической мембраны. Эти углеводные комплексы помогают клетке связывать необходимые клетке вещества во внеклеточной жидкости.Это значительно усиливает избирательность плазматических мембран.
Интегральные белки, которые действуют как каналы или насосы, работают в одном направлении. Углеводы, присоединенные к липидам или белкам, также находятся на внешней поверхности плазматической мембраны. Эти углеводные комплексы помогают клетке связывать необходимые клетке вещества во внеклеточной жидкости.Это значительно усиливает избирательность плазматических мембран.
Напомним, что плазматические мембраны имеют гидрофильные и гидрофобные области. Эта характеристика помогает перемещению одних материалов через мембрану и препятствует перемещению других. Жирорастворимый материал может легко проскальзывать через гидрофобное липидное ядро мембраны . Такие вещества, как жирорастворимые витамины A, D, E и K, легко проходят через плазматические мембраны в пищеварительном тракте и других тканях.Жирорастворимые препараты также легко проникают в клетки и легко транспортируются в ткани и органы организма. Молекулы кислорода и углекислого газа не имеют заряда и проходят через простую диффузию.
Полярные вещества, за исключением воды, создают проблемы для мембраны. Хотя некоторые полярные молекулы легко соединяются с внешней стороной клетки, они не могут легко пройти через липидное ядро плазматической мембраны . Кроме того, в то время как небольшие ионы могут легко проскальзывать через промежутки в мозаике мембраны, их заряд мешает им это делать.Ионы, такие как натрий, калий, кальций и хлорид, должны иметь специальные средства проникновения через плазматические мембраны. Простые сахара и аминокислоты также нуждаются в помощи для транспорта через плазматические мембраны.
Распространение — это пассивный транспортный процесс. Отдельное вещество имеет тенденцию перемещаться из области с высокой концентрацией в область с низкой концентрацией , пока концентрация не станет одинаковой по всему пространству. Вы знакомы с диффузией веществ по воздуху. Например, представьте, как кто-то открывает флакон духов в комнате, заполненной людьми. Самая высокая концентрация духов находится во флаконе, а самая низкая — на краю комнаты. Пар духов будет рассеиваться или распространяться от флакона, и постепенно все больше и больше людей будут ощущать запах духов по мере их распространения. Материалы перемещаются в цитозоле клетки путем диффузии, а некоторые материалы перемещаются через плазматическую мембрану путем диффузии (рис. 3.24). Распространение не требует затрат энергии. Скорее, разные концентрации материалов в разных областях являются формой потенциальной энергии, а диффузия — это диссипация этой потенциальной энергии по мере того, как материалы движутся вниз по градиенту их концентрации, от высокого к низкому.
Самая высокая концентрация духов находится во флаконе, а самая низкая — на краю комнаты. Пар духов будет рассеиваться или распространяться от флакона, и постепенно все больше и больше людей будут ощущать запах духов по мере их распространения. Материалы перемещаются в цитозоле клетки путем диффузии, а некоторые материалы перемещаются через плазматическую мембрану путем диффузии (рис. 3.24). Распространение не требует затрат энергии. Скорее, разные концентрации материалов в разных областях являются формой потенциальной энергии, а диффузия — это диссипация этой потенциальной энергии по мере того, как материалы движутся вниз по градиенту их концентрации, от высокого к низкому.
Каждое отдельное вещество в среде, такой как внеклеточная жидкость, имеет свой собственный градиент концентрации, независимый от градиентов концентрации других материалов. Кроме того, каждое вещество будет распространяться в соответствии с этим градиентом.
Кроме того, каждое вещество будет распространяться в соответствии с этим градиентом.
На скорость диффузии влияют несколько факторов.
- Степень градиента концентрации: чем больше разница в концентрации, тем быстрее происходит диффузия.Чем ближе распределение материала к равновесию, тем медленнее становится скорость диффузии.
- Масса диффундирующих молекул: более массивные молекулы движутся медленнее, потому что им труднее перемещаться между молекулами вещества, через которое они движутся; поэтому они распространяются медленнее.
- Температура: более высокие температуры увеличивают энергию и, следовательно, движение молекул, увеличивая скорость диффузии.
- Плотность растворителя: по мере увеличения плотности растворителя скорость диффузии уменьшается.Молекулы замедляются, потому что им труднее проходить через более плотную среду.
Концепция в действии
Чтобы увидеть анимацию процесса диффузии в действии, просмотрите это короткое видео о транспорте через клеточную мембрану.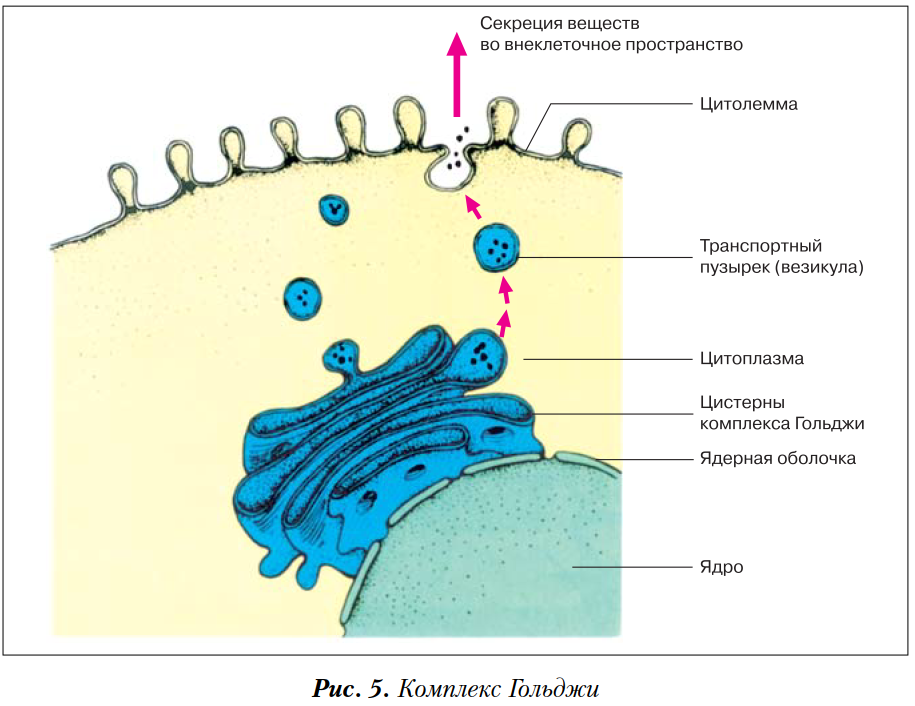
При облегченном транспорте, также называемом облегченной диффузией, материал перемещается через плазматическую мембрану с помощью трансмембранных белков вниз по градиенту концентрации (от высокой до низкой концентрации) без затрат клеточной энергии.Однако вещества, которые подвергаются облегченному транспорту, в противном случае не могли бы легко и быстро диффундировать через плазматическую мембрану. Решение проблемы перемещения полярных и других веществ через плазматическую мембрану лежит в белках, покрывающих ее поверхность. Транспортируемый материал сначала прикрепляется к рецепторам белка или гликопротеина на внешней поверхности плазматической мембраны. Это позволяет удалить материал, необходимый клетке, из внеклеточной жидкости. Затем вещества передаются определенным интегральным белкам, которые облегчают их прохождение, потому что они образуют каналы или поры, которые позволяют определенным веществам проходить через мембрану. Интегральные белки, участвующие в облегченном транспорте, вместе называются транспортными белками, и они функционируют либо как каналы для материала, либо как носители.
Интегральные белки, участвующие в облегченном транспорте, вместе называются транспортными белками, и они функционируют либо как каналы для материала, либо как носители.
— это диффузия воды через полупроницаемую мембрану в соответствии с градиентом концентрации воды через мембрану. В то время как диффузия переносит материал через мембраны и внутри клеток, осмос переносит только воду через мембрану, а мембрана ограничивает диффузию растворенных веществ в воде.Осмос — это особый случай диффузии. Вода, как и другие вещества, перемещается из области с более высокой концентрацией в область с более низкой концентрацией. Представьте себе стакан с полупроницаемой мембраной, разделяющей две стороны или половины (рис. 3.25). С обеих сторон мембраны уровень воды одинаков, но с каждой стороны существуют разные концентрации растворенного вещества или растворенного вещества, которое не может пересечь мембрану. Если объем воды такой же, но концентрации растворенных веществ различаются, то есть также разные концентрации воды, растворителя, по обе стороны от мембраны.
Принцип диффузии заключается в том, что молекулы перемещаются и при возможности равномерно распространяются по среде. Однако через нее будет диффундировать только материал, способный проникнуть через мембрану. В этом примере растворенное вещество не может диффундировать через мембрану, а вода может.Вода в этой системе имеет градиент концентрации. Следовательно, вода будет диффундировать вниз по градиенту концентрации, пересекая мембрану в сторону, где она менее концентрирована. Эта диффузия воды через мембрану — осмос — будет продолжаться до тех пор, пока градиент концентрации воды не станет равным нулю. Осмос в живых системах происходит постоянно.
Тоничность описывает количество растворенного вещества в растворе. Мера тоничности раствора или общего количества растворенных веществ, растворенных в определенном количестве раствора, называется его осмолярностью. Три термина — гипотонический, изотонический и гипертонический — используются для связи осмолярности клетки с осмолярностью внеклеточной жидкости, содержащей клетки. В гипотоническом растворе, таком как водопроводная вода, внеклеточная жидкость имеет более низкую концентрацию растворенных веществ, чем жидкость внутри клетки, и вода поступает в клетку. (В живых системах точкой отсчета всегда является цитоплазма, поэтому префикс hypo — означает, что внеклеточная жидкость имеет более низкую концентрацию растворенных веществ или более низкую осмолярность, чем цитоплазма клетки.Это также означает, что во внеклеточной жидкости концентрация воды выше, чем в клетке. В этой ситуации вода будет следовать градиенту своей концентрации и попадет в ячейку. Это может вызвать взрыв или лизис животной клетки.
Три термина — гипотонический, изотонический и гипертонический — используются для связи осмолярности клетки с осмолярностью внеклеточной жидкости, содержащей клетки. В гипотоническом растворе, таком как водопроводная вода, внеклеточная жидкость имеет более низкую концентрацию растворенных веществ, чем жидкость внутри клетки, и вода поступает в клетку. (В живых системах точкой отсчета всегда является цитоплазма, поэтому префикс hypo — означает, что внеклеточная жидкость имеет более низкую концентрацию растворенных веществ или более низкую осмолярность, чем цитоплазма клетки.Это также означает, что во внеклеточной жидкости концентрация воды выше, чем в клетке. В этой ситуации вода будет следовать градиенту своей концентрации и попадет в ячейку. Это может вызвать взрыв или лизис животной клетки.
В гипертоническом растворе (префикс hyper — относится к внеклеточной жидкости, имеющей более высокую концентрацию растворенных веществ, чем цитоплазма клетки), жидкость содержит меньше воды, чем клетка, например морская вода. Поскольку в ячейке более низкая концентрация растворенных веществ, вода будет покидать ячейку.Фактически растворенное вещество вытягивает воду из клетки. Это может вызвать сморщивание или образование зубцов в животной клетке.
Поскольку в ячейке более низкая концентрация растворенных веществ, вода будет покидать ячейку.Фактически растворенное вещество вытягивает воду из клетки. Это может вызвать сморщивание или образование зубцов в животной клетке.
В изотоническом растворе внеклеточная жидкость имеет ту же осмолярность, что и клетка. Если концентрация растворенных веществ в клетке совпадает с концентрацией внеклеточной жидкости, чистого движения воды внутрь или из клетки не будет. Клетки крови в гипертонических, изотонических и гипотонических растворах приобретают характерный вид (рис. 3.26).
Рис. 3.26. Осмотическое давление изменяет форму эритроцитов в гипертонических, изотонических и гипотонических растворах.Врач вводит пациенту изотонический физиологический раствор. Пациент умирает, и вскрытие показывает, что многие эритроциты были разрушены. Как вы думаете, раствор, который ввел врач, был действительно изотоническим?
 ->
->
У некоторых организмов, таких как растения, грибы, бактерии и некоторые простейшие, есть клеточные стенки, которые окружают плазматическую мембрану и предотвращают лизис клеток.Плазматическая мембрана может расширяться только до предела клеточной стенки, поэтому клетка не подвергается лизису. Фактически, цитоплазма растений всегда слегка гипертоническая по сравнению с клеточной средой, и вода всегда будет проникать в клетку, если она доступна. Этот приток воды создает тургорное давление, которое укрепляет клеточные стенки растения (рис. 3.27). У недревесных растений давление тургора поддерживает растение. Если клетки растения становятся гипертоническими, как это происходит во время засухи, или если растение не поливают надлежащим образом, вода будет уходить из клетки.В этом состоянии растения теряют тургорное давление и вянут.
Рис. 3.27. Тургорное давление внутри растительной клетки зависит от тоничности раствора, в котором она купается. Пассивные формы транспорта, диффузии и осмоса, перемещают материалы с малой молекулярной массой. Вещества диффундируют из областей с высокой концентрацией в области с низкой концентрацией, и этот процесс продолжается до тех пор, пока вещество не будет равномерно распределено в системе. В растворах, содержащих более одного вещества, каждый тип молекулы диффундирует в соответствии со своим градиентом концентрации.На скорость диффузии могут влиять многие факторы, включая градиент концентрации, размер диффундирующих частиц и температуру системы.
Вещества диффундируют из областей с высокой концентрацией в области с низкой концентрацией, и этот процесс продолжается до тех пор, пока вещество не будет равномерно распределено в системе. В растворах, содержащих более одного вещества, каждый тип молекулы диффундирует в соответствии со своим градиентом концентрации.На скорость диффузии могут влиять многие факторы, включая градиент концентрации, размер диффундирующих частиц и температуру системы.
В живых системах диффузия веществ внутрь и из клеток опосредуется плазматической мембраной. Некоторые материалы легко диффундируют через мембрану, но другие затрудняются, и их прохождение возможно только с помощью белковых каналов и носителей. Химия живых существ происходит в водных растворах, и балансировка концентраций этих растворов является постоянной проблемой.В живых системах диффузия некоторых веществ была бы медленной или затрудненной без мембранных белков.
градиент концентрации: зона высокой концентрации напротив зоны низкой концентрации
диффузия: пассивный процесс транспортировки низкомолекулярного материала вниз по градиенту его концентрации
облегченная транспортировка: процесс, при котором материал движется вниз по градиенту концентрации (от высокой до низкой концентрации) с использованием интегральных мембранных белков
гипертонический: описывает раствор, в котором внеклеточная жидкость имеет более высокую осмолярность, чем жидкость внутри клетки
гипотонический: описывает раствор, в котором внеклеточная жидкость имеет более низкую осмолярность, чем жидкость внутри клетки
изотонический: описывает раствор, в котором внеклеточная жидкость имеет ту же осмолярность, что и жидкость внутри клетки
осмолярность: общее количество веществ, растворенных в определенном количестве раствора
осмос: перенос воды через полупроницаемую мембрану из зоны с высокой концентрацией воды в зону с низкой концентрацией воды через мембрану
пассивная транспортировка: способ транспортировки материалов, не требующий энергии
избирательно проницаемая: характеристика мембраны, которая пропускает одни вещества, но не пропускает другие
растворенное вещество: вещество, растворенное в другом с образованием раствора
тоничность: количество растворенного вещества в растворе.
Атрибуция в СМИ
- Рисунок 3.24: модификация работы Марианы Руис Вильярреал
- Рисунок 3.26: модификация работы Марианы Руис Вильярреал
- Рисунок 3.27: модификация работы Марианы Руис Вильярреал
Транспорт малых молекул — Клетка
Внутренний состав клетки сохраняется, поскольку плазматическая мембрана избирательно проницаема для малых молекул. Большинство биологических молекул не могут диффундировать через бислой фосфолипидов, поэтому плазматическая мембрана образует барьер, который блокирует свободный обмен молекулами между цитоплазмой и внешней средой клетки.Затем специфические транспортные белки (белки-носители и канальные белки) опосредуют избирательный проход малых молекул через мембрану, позволяя клетке контролировать состав своей цитоплазмы.
Пассивная диффузия
Самым простым механизмом, с помощью которого молекулы могут пересекать плазматическую мембрану, является пассивная диффузия. Во время пассивной диффузии молекула просто растворяется в бислое фосфолипидов, диффундирует через него, а затем растворяется в водном растворе на другой стороне мембраны. Никакие мембранные белки не участвуют, и направление транспорта определяется просто относительной концентрацией молекулы внутри и вне клетки. Чистый поток молекул всегда ниже градиента их концентрации — от отсека с высокой концентрацией к отсеку с более низкой концентрацией молекулы.
Никакие мембранные белки не участвуют, и направление транспорта определяется просто относительной концентрацией молекулы внутри и вне клетки. Чистый поток молекул всегда ниже градиента их концентрации — от отсека с высокой концентрацией к отсеку с более низкой концентрацией молекулы.
Таким образом, пассивная диффузия представляет собой неселективный процесс, с помощью которого любая молекула, способная растворяться в фосфолипидном бислое, способна пересечь плазматическую мембрану и уравновеситься между внутренней и внешней частью клетки.Важно отметить, что только небольшие, относительно гидрофобные молекулы способны диффундировать через бислой фосфолипидов со значительной скоростью (). Таким образом, газы (такие как O 2 и CO 2 ), гидрофобные молекулы (такие как бензол) и небольшие полярные, но незаряженные молекулы (такие как H 2 O и этанол) могут диффундировать через плазматическую мембрану. . Однако другие биологические молекулы не могут растворяться в гидрофобной внутренней части фосфолипидного бислоя. Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересекать плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl — ).Вместо этого прохождение этих молекул через мембрану требует активности специфических транспортных и канальных белков, которые, следовательно, контролируют движение большинства биологических молекул в клетку и из клетки.
Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересекать плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl — ).Вместо этого прохождение этих молекул через мембрану требует активности специфических транспортных и канальных белков, которые, следовательно, контролируют движение большинства биологических молекул в клетку и из клетки.
Рисунок 12.15
Проницаемость фосфолипидных бислоев. Газы, гидрофобные молекулы и небольшие полярные незаряженные молекулы могут диффундировать через бислои фосфолипидов. Более крупные полярные и заряженные молекулы не могут.
Облегченная диффузия и белки-носители
Облегченная диффузия, как и пассивная диффузия, включает движение молекул в направлении, определяемом их относительной концентрацией внутри и снаружи клетки.Внешний источник энергии не предусмотрен, поэтому молекулы перемещаются через мембрану в направлении, определяемом их градиентами концентрации, а в случае заряженных молекул — электрическим потенциалом на мембране. Однако облегченная диффузия отличается от пассивной диффузии тем, что транспортируемые молекулы не растворяются в фосфолипидном бислое. Вместо этого их прохождение опосредуется белками, которые позволяют транспортируемым молекулам пересекать мембрану, не взаимодействуя напрямую с ее гидрофобным внутренним пространством.Таким образом, облегченная диффузия позволяет полярным и заряженным молекулам, таким как углеводы, аминокислоты, нуклеозиды и ионы, проходить через плазматическую мембрану.
Однако облегченная диффузия отличается от пассивной диффузии тем, что транспортируемые молекулы не растворяются в фосфолипидном бислое. Вместо этого их прохождение опосредуется белками, которые позволяют транспортируемым молекулам пересекать мембрану, не взаимодействуя напрямую с ее гидрофобным внутренним пространством.Таким образом, облегченная диффузия позволяет полярным и заряженным молекулам, таким как углеводы, аминокислоты, нуклеозиды и ионы, проходить через плазматическую мембрану.
Обычно различают два класса белков, которые обеспечивают облегченную диффузию: белки-носители и канальные белки. Белки-носители связывают определенные молекулы, которые транспортируются на одной стороне мембраны. Затем они претерпевают конформационные изменения, которые позволяют молекуле проходить через мембрану и высвобождаться с другой стороны.Напротив, канальные белки (см. Следующий раздел) образуют открытые поры через мембрану, обеспечивая свободную диффузию любой молекулы соответствующего размера и заряда.
Белки-носители ответственны за облегченную диффузию сахаров, аминокислот и нуклеозидов через плазматические мембраны большинства клеток. Поглощение глюкозы, которая служит основным источником метаболической энергии, является одной из наиболее важных транспортных функций плазматической мембраны, а переносчик глюкозы представляет собой хорошо изученный пример белка-носителя.Переносчик глюкозы первоначально был идентифицирован как белок массой 55 кДа в эритроцитах человека, в которых он составляет примерно 5% от общего мембранного белка. Последующее выделение и анализ последовательности клона кДНК выявили, что переносчик глюкозы имеет 12 α-спиральных трансмембранных сегментов — структуру, типичную для многих белков-носителей (2). Эти трансмембранные α-спирали содержат преимущественно гидрофобные аминокислоты, но некоторые из них также содержат полярные аминокислотные остатки, которые, как считается, образуют сайт связывания глюкозы внутри белка.
Рисунок 12.16
Структура переносчика глюкозы. Переносчик глюкозы имеет 12 трансмембранных α-спиралей. Полярные аминокислотные остатки, расположенные внутри фосфолипидного бислоя, показаны темно-фиолетовыми кружками. (По материалам GI Bell, CF Burant, J. Takeda (подробнее …)
Переносчик глюкозы имеет 12 трансмембранных α-спиралей. Полярные аминокислотные остатки, расположенные внутри фосфолипидного бислоя, показаны темно-фиолетовыми кружками. (По материалам GI Bell, CF Burant, J. Takeda (подробнее …)
Как и для многих мембранных белков, трехмерная структура переносчика глюкозы неизвестна, поэтому молекулярный механизм транспорта глюкозы остается открытым вопросом Однако кинетические исследования показывают, что переносчик глюкозы функционирует путем чередования двух конформационных состояний ().В первой конформации сайт связывания глюкозы обращен к внешней стороне клетки. Связывание глюкозы с этим внешним сайтом вызывает конформационные изменения в транспортере, так что сайт связывания глюкозы теперь обращен внутрь клетки. Затем глюкоза может быть выпущена в цитозоль с последующим возвращением переносчика к его исходной конформации.
Рисунок 12.17
Модель для облегченной диффузии глюкозы. Переносчик глюкозы чередуется между двумя конформациями, в которых сайт связывания глюкозы поочередно открыт снаружи и внутри клетки.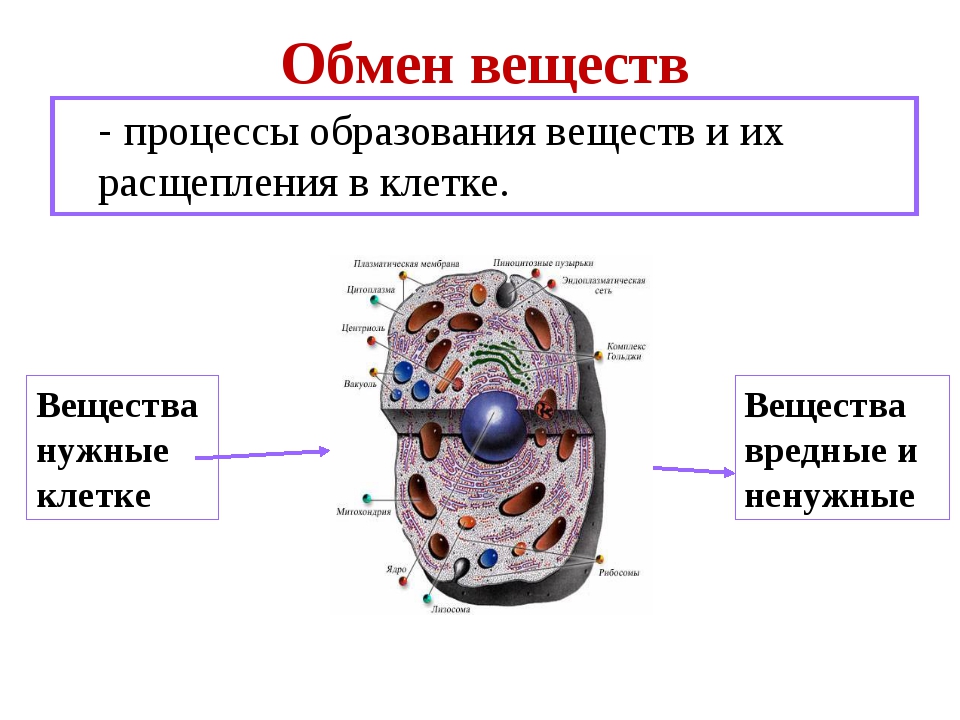 В первой показанной конформации (A) глюкоза (подробнее …)
В первой показанной конформации (A) глюкоза (подробнее …)
Большинство клеток, включая эритроциты, подвергаются воздействию внеклеточной глюкозы с более высокими концентрациями глюкозы, чем внутри клетки, поэтому облегченная диффузия приводит к общему внутреннему транспорту глюкозы. . Как только глюкоза поглощается этими клетками, она быстро метаболизируется, поэтому внутриклеточные концентрации глюкозы остаются низкими, а глюкоза продолжает транспортироваться в клетку из внеклеточных жидкостей. Однако, поскольку конформационные изменения переносчика глюкозы обратимы, глюкоза может транспортироваться в противоположном направлении, просто меняя шаги в обратном направлении.Такой обратный поток происходит, например, в клетках печени, в которых глюкоза синтезируется и попадает в кровоток.
Ионные каналы
В отличие от белков-носителей, канальные белки просто образуют открытые поры в мембране, позволяя небольшим молекулам подходящего размера и заряда свободно проходить через липидный бислой. Одна группа канальных белков, обсуждавшаяся ранее, — это порины, которые обеспечивают свободный проход ионов и небольших полярных молекул через внешние мембраны бактерий (см.).Канальные белки также позволяют молекулам перемещаться между клетками, соединенными щелевыми соединениями, которые обсуждаются позже в этой главе. Плазматические мембраны многих клеток также содержат белки водных каналов (аквапорины), через которые молекулы воды могут пересекать мембрану намного быстрее, чем они могут диффундировать через бислой фосфолипидов. Однако наиболее охарактеризованными белками каналов являются ионные каналы, которые обеспечивают прохождение ионов через плазматические мембраны. Хотя ионные каналы присутствуют в мембранах всех клеток, они особенно хорошо изучены в нервах и мышцах, где их регулируемое открытие и закрытие отвечает за передачу электрических сигналов.
Одна группа канальных белков, обсуждавшаяся ранее, — это порины, которые обеспечивают свободный проход ионов и небольших полярных молекул через внешние мембраны бактерий (см.).Канальные белки также позволяют молекулам перемещаться между клетками, соединенными щелевыми соединениями, которые обсуждаются позже в этой главе. Плазматические мембраны многих клеток также содержат белки водных каналов (аквапорины), через которые молекулы воды могут пересекать мембрану намного быстрее, чем они могут диффундировать через бислой фосфолипидов. Однако наиболее охарактеризованными белками каналов являются ионные каналы, которые обеспечивают прохождение ионов через плазматические мембраны. Хотя ионные каналы присутствуют в мембранах всех клеток, они особенно хорошо изучены в нервах и мышцах, где их регулируемое открытие и закрытие отвечает за передачу электрических сигналов.
Три свойства ионных каналов являются центральными для их функции (). Во-первых, транспортировка по каналам происходит очень быстро. Более миллиона ионов в секунду проходят через открытые каналы — скорость потока примерно в тысячу раз превышает скорость переноса белков-носителей. Во-вторых, ионные каналы обладают высокой избирательностью, поскольку узкие поры в канале ограничивают прохождение ионами соответствующего размера и заряда. Таким образом, специфические канальные белки позволяют проходить через мембрану Na + , K + , Ca 2+ и Cl —.В-третьих, большинство ионных каналов не открыты постоянно. Вместо этого открытие ионных каналов регулируется «воротами», которые временно открываются в ответ на определенные стимулы. Некоторые каналы (называемые лиганд-зависимыми каналами ) открываются в ответ на связывание нейротрансмиттеров или других сигнальных молекул; другие ( потенциалозависимых каналов ) открываются в ответ на изменения электрического потенциала через плазматическую мембрану.
Рисунок 12.18
Модель ионного канала. В закрытой конформации поток ионов блокируется затвором. Открытие затвора позволяет ионам быстро проходить через канал. Канал содержит узкую пору, которая ограничивает прохождение ионов подходящего размера (подробнее …)
Открытие затвора позволяет ионам быстро проходить через канал. Канал содержит узкую пору, которая ограничивает прохождение ионов подходящего размера (подробнее …)
Фундаментальная роль ионных каналов в передаче электрических импульсов была выяснена с помощью серии элегантных экспериментов, представленных Аланом Ходжкином и Эндрю Хаксли. в 1952 году. Эти исследователи использовали гигантские нервные клетки кальмаров в качестве модели. Аксоны этих гигантских нейронов имеют диаметр около 1 мм, что позволяет вставлять электроды и измерять изменения мембранного потенциала, которые происходят во время передачи нервных импульсов.Используя этот подход, Hodgkin и Huxley продемонстрировали, что эти изменения мембранного потенциала являются результатом регулируемого открытия и закрытия каналов Na + и K + в плазматической мембране. Впоследствии стало возможным изучать активность отдельных ионных каналов, используя методику патч-клампа , разработанную Эрвином Нехером и Бертом Сакманном в 1976 году (). В этом методе микропипетка с диаметром кончика около 1 мкм используется для изоляции небольшого участка мембраны, позволяя анализировать поток ионов через единственный канал и значительно повышая точность, с которой может быть измерена активность ионных каналов. изучал.
В этом методе микропипетка с диаметром кончика около 1 мкм используется для изоляции небольшого участка мембраны, позволяя анализировать поток ионов через единственный канал и значительно повышая точность, с которой может быть измерена активность ионных каналов. изучал.
Рисунок 12.19
Метод патч-зажима. Небольшой участок мембраны изолирован на кончике микропипетки. Затем из пипетки можно подавать стимулы, позволяя измерить поведение захваченного канала. (По материалам E. Neher и B. Sakmann, 1992. (подробнее …)
Поток ионов через мембранные каналы зависит от установления ионных градиентов через плазматическую мембрану. Все клетки, включая нервы и мышцы, содержат ионные насосы (обсуждаемые в следующем разделе), которые используют энергию, полученную в результате гидролиза АТФ, для активного переноса ионов через плазматическую мембрану.В результате ионный состав цитоплазмы существенно отличается от такового внеклеточных жидкостей (). Например, Na + активно откачивается из клеток, в то время как K + закачивается. Следовательно, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточных жидкостях, чем внутри клетки, тогда как концентрация K + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Следовательно, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточных жидкостях, чем внутри клетки, тогда как концентрация K + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Таблица 12.1
Концентрации внеклеточных и внутриклеточных ионов.
Поскольку ионы электрически заряжены, их транспорт приводит к созданию электрического градиента через плазматическую мембрану. При покоящихся аксонах кальмаров на плазматической мембране существует электрический потенциал около 60 мВ, при этом внутренняя часть клетки отрицательна по отношению к внешней (). Этот электрический потенциал возникает как от ионных насосов, так и от потока ионов через каналы, открытые в плазматической мембране покоящейся клетки.Плазматическая мембрана покоящихся аксонов кальмаров содержит открытые каналы K + , поэтому она более проницаема для K + , чем для Na + или других ионов. Следовательно, поток K + вносит наибольший вклад в мембранный потенциал покоя.
Рисунок 12.20
Ионный градиент и мембранный потенциал покоя аксона гигантского кальмара. Показаны только концентрации Na + и K + , потому что это ионы, которые участвуют в передаче нервных импульсов.Na + откачивается из ячейки, в то время как K + закачивается, (подробнее …)
Как обсуждалось в главе 10, поток ионов через мембрану управляется как составляющими концентрации, так и напряжением электрохимический градиент. Например, в 20 раз более высокая концентрация K + внутри аксона кальмара по сравнению с внеклеточной жидкостью вытесняет поток K + из клетки. Однако, поскольку K + заряжен положительно, этот отток K + из клетки генерирует электрический потенциал через мембрану, при этом внутренняя часть клетки становится отрицательно заряженной.Этот мембранный потенциал противодействует продолжающемуся потоку K + из клетки, и система приближается к состоянию равновесия, в котором мембранный потенциал уравновешивает градиент концентрации K + .
Количественно связь между концентрацией ионов и мембранным потенциалом определяется уравнением Нернста:
, где В, — равновесный потенциал в вольтах, R — газовая постоянная, T — абсолютная температура, z — заряд иона, F — постоянная Фарадея, а C o и C i — концентрации иона снаружи и внутри ячейки соответственно.Равновесный потенциал существует отдельно для каждого иона, а мембранный потенциал определяется потоком всех ионов, пересекающих плазматическую мембрану. Однако, поскольку аксоны кальмаров в состоянии покоя более проницаемы для K + , чем для Na + или других ионов (включая Cl —), мембранный потенциал покоя (-60 мВ) близок к равновесному потенциалу, определяемому внутриклеточным и внеклеточные концентрации K + (-75 мВ).
Когда нервные импульсы ( потенциалов действия ) проходят вдоль аксонов, мембрана деполяризуется (). Мембранный потенциал изменяется от -60 мВ до примерно +30 мВ менее чем за миллисекунду, после чего он снова становится отрицательным и возвращается к своему исходному значению. Эти изменения являются результатом быстрого последовательного открытия и закрытия управляемых напряжением каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию каналов Na + . Это позволяет Na + течь в клетку под действием как градиента его концентрации, так и мембранного потенциала.Внезапное попадание Na + приводит к большому изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивированы, и управляемые по напряжению каналы K + открываются, существенно увеличивая проницаемость мембраны до K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ).
Мембранный потенциал изменяется от -60 мВ до примерно +30 мВ менее чем за миллисекунду, после чего он снова становится отрицательным и возвращается к своему исходному значению. Эти изменения являются результатом быстрого последовательного открытия и закрытия управляемых напряжением каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию каналов Na + . Это позволяет Na + течь в клетку под действием как градиента его концентрации, так и мембранного потенциала.Внезапное попадание Na + приводит к большому изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивированы, и управляемые по напряжению каналы K + открываются, существенно увеличивая проницаемость мембраны до K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ). Управляемые по напряжению каналы K + затем инактивируются, и мембранный потенциал возвращается к своему уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Управляемые по напряжению каналы K + затем инактивируются, и мембранный потенциал возвращается к своему уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Рисунок 12.21
Мембранный потенциал и ионные каналы во время потенциала действия. (A) Изменения мембранного потенциала в одной точке гигантского аксона кальмара после стимула. E Na и E K — потенциалы равновесия для Na + и K + соответственно.(B) Мембранный потенциал (подробнее …)
Деполяризация соседних областей плазматической мембраны позволяет потенциалам действия перемещаться по аксонам нервных клеток в виде электрических сигналов, что приводит к быстрой передаче нервных импульсов на большие расстояния. Например, аксоны мотонейронов человека могут быть более метра в длину. Прибытие потенциалов действия к концам большинства нейронов затем сигнализирует о высвобождении нейротрансмиттеров, таких как ацетилхолин, которые переносят сигналы между клетками в синапсе ().Нейротрансмиттеры, высвобождаемые пресинаптическими клетками, связываются с рецепторами на мембранах постсинаптических клеток, где они действуют, открывая лиганд-зависимые ионные каналы. Одним из наиболее хорошо охарактеризованных каналов является ацетилхолиновый рецептор мышечных клеток. Связывание ацетилхолина открывает канал, проницаемый как для Na + , так и для K + . Это обеспечивает быстрый приток Na + , который деполяризует мембрану мышечной клетки и запускает потенциал действия. Затем потенциал действия приводит к открытию потенциалозависимых каналов Ca 2+ , что приводит к увеличению внутриклеточного Ca 2+ , что сигнализирует о сокращении (см.).
Рисунок 12.22
Передача сигналов посредством высвобождения нейромедиатора в синапсе. Прибытие нервного импульса к концу нейрона сигнализирует о слиянии синаптических пузырьков с плазматической мембраной, что приводит к высвобождению нейромедиатора из пресинаптической клетки в (подробнее …)
рецептор ацетилхолина, первоначально выделенный из Электрический орган Торпеда лучей в 1970-е годы, является прототипом лиганд-закрытых каналов. Рецептор состоит из пяти субъединиц, расположенных в мембране в виде цилиндра ().Считается, что в закрытом состоянии поры канала заблокированы боковыми цепями гидрофобных аминокислот. Связывание ацетилхолина вызывает конформационное изменение рецептора, так что эти гидрофобные боковые цепи смещаются из канала, открывая поры, которые позволяют проходить положительно заряженным ионам, включая Na + и K + . Однако канал остается непроницаемым для отрицательно заряженных ионов, таких как Cl —, потому что он выстлан отрицательно заряженными аминокислотами.
Рисунок 12.23
Модель рецептора ацетилхолина. Рецептор состоит из пяти субъединиц, расположенных вокруг центральной поры. Связывание ацетилхолина с сайтом во внеклеточной области рецептора вызывает аллостерические изменения, которые открывают ворота канала. Канал (подробнее …)
Большую степень ионной селективности демонстрируют потенциалозависимые каналы Na + и K + . Каналы Na + более чем в десять раз проницаемы для Na + , чем для K + , тогда как каналы K + более чем в тысячу раз проницаемы для K + , чем для Na + .Селективность канала Na + можно объяснить, по крайней мере частично, на основе узкой поры, которая действует как размерный фильтр. Ионный радиус Na + (0,95 Å) меньше, чем у K + (1,33 Å), и считается, что поры канала Na + достаточно узкие, чтобы препятствовать прохождению K +. или более крупные ионы ().
Рисунок 12.24
Ионная селективность каналов Na + . Узкая пора позволяет прохождению Na + , связанного с одной молекулой воды, но препятствует прохождению ионов K + или более крупных.Каналы
K + также имеют узкие поры, которые препятствуют прохождению более крупных ионов. Однако, поскольку Na + имеет меньший ионный радиус, это не учитывает избирательную проницаемость этих каналов для K + . Селективность канала K + основана на другом механизме, который был выяснен при определении трехмерной структуры канала K + методом рентгеновской кристаллографии в 1998 г. (). Пора канала содержит фильтр с узкой селективностью, который выстлан карбонильными атомами кислорода (C = O) из основной цепи полипептида.Когда ион K + попадает в фильтр селективности, взаимодействия с этими карбонильными атомами кислорода вытесняют молекулы воды, с которыми связан K + , позволяя дегидратированному K + проходить через поры. Напротив, дегидратированный Na + слишком мал, чтобы взаимодействовать с этими карбонильными атомами кислорода в фильтре селективности, который остается жестко открытым. Следовательно, Na + остается связанным с молекулами воды в гидратированном комплексе, который слишком велик для прохождения через канал.
Рисунок 12.25
Избирательность каналов K + . Канал K + содержит узкий селективный фильтр, облицованный карбонильными атомами кислорода (C = O). Пора достаточно широка, чтобы пропустить дегидратированный K + , из которого были вытеснены все связанные молекулы воды, как (подробнее …)
Управляемый напряжением Na + , K + и Ca 2+ каналов принадлежат к большому семейству родственных белков (). Например, последовательность генома с.elegans выявил около 200 генов, кодирующих ионные каналы, которые предположительно необходимы для выполнения различных ролей в передаче сигналов в клетках. Каналы K + состоят из четырех идентичных субъединиц, каждая из которых содержит две или шесть трансмембранных α-спиралей. Каналы Na + и Ca 2+ состоят из одной полипептидной цепи, но каждый полипептид содержит четыре повторяющихся домена, которые соответствуют субъединицам канала K + . Стробирование напряжения опосредуется одной из трансмембранных α-спиралей, которая содержит несколько положительно заряженных аминокислот.Деполяризация мембраны вызывает движение этих положительных зарядов к внешней стороне клетки, смещая положение этого трансмембранного сегмента и открывая канал. Быстрая инактивация каналов Na + и K + во время распространения потенциалов действия затем опосредуется цитоплазматическими частями полипептидной цепи, которые связываются с цитоплазматическим устьем поры канала и предотвращают дальнейший поток ионов ().
Рисунок 12.26
Структуры потенциалозависимых катионных каналов.Каналы K + , Na + и Ca 2+ принадлежат к семейству родственных белков. Канал K + формируется из ассоциации четырех идентичных субъединиц, одна из которых показана. Канал Na + состоит из одного полипептида (подробнее …)
Рисунок 12.27
Инактивация каналов K + и Na + . После открытия, управляемого по напряжению, каналы K + и Na + быстро инактивируются за счет связывания цитоплазматических частей полипептидных цепей с порами.Для канала K + инактивация опосредуется цепочкой (подробнее …)
Широкий спектр ионных каналов (включая каналы Ca 2+ и Cl —) отвечает на различные нейротрансмиттеры. или открываются и закрываются с различной кинетикой после деполяризации мембраны. Согласованные действия этих множественных каналов ответственны за сложность передачи сигналов в нервной системе. Более того, как обсуждается в следующей главе, роль ионных каналов не ограничивается электрически возбудимыми клетками нервов и мышц; они также играют важную роль в передаче сигналов в других типах клеток.Таким образом, регулируемое открытие и закрытие ионных каналов обеспечивает клетки чувствительным и универсальным механизмом реагирования на различные раздражители окружающей среды.
Активный транспорт, управляемый гидролизом АТФ
Чистый поток молекул за счет облегченной диффузии через белки-носители или канальные белки всегда энергетически нисходящий в направлении, определяемом электрохимическими градиентами через мембрану. Однако во многих случаях клетка должна транспортировать молекулы против их градиентов концентрации.При активном переносе энергия, обеспечиваемая другой сопряженной реакцией (такой как гидролиз АТФ), используется для управления восходящим транспортом молекул в энергетически невыгодном направлении.
Ионные насосы, отвечающие за поддержание градиентов ионов через плазматическую мембрану, представляют собой важные примеры активного транспорта, управляемого непосредственно гидролизом АТФ. Как обсуждалось ранее (см.), Концентрация Na + примерно в десять раз выше снаружи, чем внутри клеток, тогда как концентрация K + выше внутри, чем снаружи.Эти ионные градиенты поддерживаются насосом Na + -K + (также называемым Na + -K + ATPase ), который использует энергию, полученную из Гидролиз АТФ для переноса Na + и K + против их электрохимических градиентов. Этот процесс является результатом управляемых АТФ конформационных изменений в помпе (). Во-первых, ионы Na + связываются с сайтами с высоким сродством внутри клетки.Это связывание стимулирует гидролиз АТФ и фосфорилирование помпы, вызывая конформационные изменения, которые открывают сайты связывания Na + за пределы клетки и снижают их сродство к Na + . Следовательно, связанный Na + высвобождается во внеклеточные жидкости. В то же время на поверхности клетки экспонируются высокоаффинные сайты связывания K + . Связывание внеклеточного K + с этими сайтами затем стимулирует гидролиз фосфатной группы, связанной с насосом, что вызывает второе конформационное изменение, подвергая сайты связывания K + цитозолю и снижая их сродство к связыванию, так что K + выпущен внутри ячейки.Насос имеет три сайта связывания для Na + и два для K + , поэтому каждый цикл транспортирует три Na + и два K + через плазматическую мембрану за счет одной молекулы АТФ.
На важность насоса Na + -K + указывает тот факт, что он, по оценкам, потребляет почти 25% АТФ, используемого многими клетками животных. Одной из важнейших ролей градиентов Na + и K + , создаваемых помпой, является распространение электрических сигналов по нервам и мышцам.Как будет вскоре обсуждено, градиент Na + , устанавливаемый насосом, также используется для управления активным транспортом множества других молекул. Еще одна важная роль насоса Na + -K + в большинстве клеток животных заключается в поддержании осмотического баланса и объема клеток. Цитоплазма содержит высокую концентрацию органических молекул, включая макромолекулы, аминокислоты, сахара и нуклеотиды. В отсутствие противовеса это привело бы внутрь потока воды за счет осмоса, что, если его не остановить, привело бы к набуханию и, в конечном итоге, к взрыву ячейки.Необходимый противовес обеспечивается ионными градиентами, устанавливаемыми насосом Na + -K + (). В частности, насос устанавливает более высокую концентрацию Na + снаружи, чем внутри ячейки. Как уже обсуждалось, поток K + через открытые каналы дополнительно устанавливает электрический потенциал на плазматической мембране. Этот мембранный потенциал, в свою очередь, вытесняет Cl — из клетки, поэтому концентрация Cl — (как и Na + ) примерно в десять раз выше во внеклеточных жидкостях, чем в цитоплазме.Эти различия в концентрациях ионов уравновешивают высокие концентрации органических молекул внутри клеток, выравнивая осмотическое давление и предотвращая чистый приток воды.
Рис. 12.29
Градиенты ионов через плазматическую мембрану типичной клетки млекопитающего. Концентрации Na + и Cl — выше снаружи, чем внутри клетки, тогда как концентрация K + выше внутри, чем снаружи. Низкие концентрации Na + и Cl — уравновешивают (более…)
Активный транспорт Ca 2+ через плазматическую мембрану управляется насосом Ca 2+ , который структурно связан с насосом Na + -K + и аналогичным образом питается от АТФ. гидролиз. Насос Ca 2+ транспортирует Ca 2+ из клетки, поэтому внутриклеточные концентрации Ca 2+ чрезвычайно низкие: приблизительно 0,1 мкМ, по сравнению с внеклеточными концентрациями около 1 м M .Эта низкая внутриклеточная концентрация Ca 2+ делает клетку чувствительной к небольшому увеличению внутриклеточных уровней Ca 2+ . Такое временное повышение внутриклеточного Ca 2+ играет важную роль в передаче сигналов в клетке, как уже отмечалось в отношении сокращения мышц (см.) И обсуждается далее в следующей главе.
Подобные ионные насосы в плазматических мембранах бактерий, дрожжей и растительных клеток ответственны за активный транспорт H + из клетки.Кроме того, H + активно выкачивается из клеток, выстилающих желудок, что приводит к повышению кислотности желудочного сока. Структурно различные насосы ответственны за активный транспорт H + в лизосомы и эндосомы (см.). И все же третий тип насоса H + представлен АТФ-синтазами митохондрий и хлоропластов: в этих случаях насосы можно рассматривать как работающие в обратном направлении, с движением ионов вниз по электрохимическому градиенту, используемому для управления синтезом АТФ.
Самое большое семейство мембранных транспортеров состоит из транспортеров ABC, названных так потому, что они характеризуются высококонсервативными АТФ-связывающими доменами или A TP- b , включая c assettes (). Как в прокариотических, так и в эукариотических клетках идентифицировано более сотни членов семей. У бактерий переносчики ABC используют энергию, полученную в результате гидролиза АТФ, для транспортировки широкого спектра молекул, включая ионы, сахара и аминокислоты. В эукариотических клетках первый переносчик ABC был обнаружен как продукт гена (называемого геном множественной лекарственной устойчивости или mdr , ген), который делает раковые клетки устойчивыми к различным лекарствам, используемым в химиотерапии.В настоящее время идентифицированы два переносчика МЛУ. Обычно они экспрессируются в различных клетках, где их функция заключается в удалении потенциально токсичных чужеродных соединений. Например, экспрессия переносчика МЛУ в капиллярных эндотелиальных клетках головного мозга, по-видимому, играет важную роль в защите мозга от токсичных химических веществ. К сожалению, переносчики МЛУ часто экспрессируются на высоком уровне в раковых клетках, где они распознают множество лекарств и выкачивают их из клеток, тем самым делая раковые клетки устойчивыми к широкому спектру химиотерапевтических агентов и создавая серьезное препятствие для успешного лечения рака. .
Рисунок 12.30
Конструкция транспортера ABC. Основная структурная единица транспортеров ABC состоит из шести трансмембранных доменов, за которыми следует АТФ-связывающая кассета. В транспортерах плазматической мембраны (таких как проиллюстрированный транспортер MDR) две из этих единиц (подробнее …)
Другим важным с медицинской точки зрения членом семейства транспортеров ABC является ген, ответственный за муковисцидоз. Хотя он является членом семейства ABC, продукт этого гена (называемый регулятором трансмембранной проводимости кистозного фиброза или CFTR) функционирует как канал Cl — в эпителиальных клетках, а дефектный транспорт Cl — характерен для болезнь.Канал CFTR Cl — также необычен тем, что для его открытия, по-видимому, требуется как гидролиз АТФ, так и цАМФ-зависимое фосфорилирование. Структурная основа функции CFTR как регулируемого ионного канала еще предстоит выяснить в будущих исследованиях.
Активный перенос, управляемый ионными градиентами
Ионные насосы и переносчики ABC, описанные в предыдущем разделе, используют энергию, полученную непосредственно в результате гидролиза АТФ, для переноса молекул против их электрохимических градиентов.Другие молекулы транспортируются против их градиентов концентрации с использованием энергии, полученной не за счет гидролиза АТФ, а за счет сопряженного транспорта второй молекулы в энергетически выгодном направлении. Градиент Na + , установленный насосом Na + -K + , обеспечивает источник энергии, который часто используется для обеспечения активного транспорта сахаров, аминокислот и ионов в клетках млекопитающих. Сходные роли играют градиенты H + , установленные насосами H + бактерий, дрожжей и растительных клеток.
Эпителиальные клетки, выстилающие кишечник, представляют собой хороший пример активного транспорта, управляемого градиентом Na + . Эти клетки используют системы активного транспорта в апикальных доменах своих плазматических мембран для поглощения пищевых сахаров и аминокислот из просвета кишечника. Поглощение глюкозы, например, осуществляется транспортером, который согласованно транспортирует два Na + и одну глюкозу в клетку (). Поток Na + вниз по его электрохимическому градиенту обеспечивает энергию, необходимую для поглощения глюкозы с пищей и для накопления высоких внутриклеточных концентраций глюкозы.Затем глюкоза высвобождается в нижележащую соединительную ткань (которая содержит кровеносные капилляры) на базолатеральной поверхности кишечного эпителия, где она транспортируется вниз по градиенту концентрации за счет облегченной диффузии (). Поглощение глюкозы из просвета кишечника и ее выпуск в кровоток, таким образом, является хорошим примером поляризованной функции эпителиальных клеток, которая является результатом специфической локализации активного транспорта и облегченных переносчиков диффузии в апикальные и базолатеральные домены плазматической мембраны. , соответственно.
Рисунок 12.31
Активный транспорт глюкозы. Активный транспорт, управляемый градиентом Na + , отвечает за захват глюкозы из просвета кишечника. Транспортер координированно связывает и транспортирует одну глюкозу и два Na + в клетку. Транспорт Na + (подробнее …)
Рисунок 12.32
Транспорт глюкозы эпителиальными клетками кишечника. Транспортер в апикальном домене плазматической мембраны отвечает за активное поглощение глюкозы (путем котранспорта с Na + ) из просвета кишечника.В результате, глюкоза с пищей абсорбируется и (подробнее …)
Координатное поглощение глюкозы и Na + является примером симпорта, транспорта двух молекул в одном направлении. Напротив, облегченная диффузия глюкозы является примером унипорта, транспорта только одной молекулы. Активный транспорт также может происходить с помощью антипорта, в котором две молекулы транспортируются в противоположных направлениях (). Например, Ca 2+ экспортируется из клеток не только насосом Ca 2+ , но и антипортером Na + -Ca 2+ , который транспортирует Na + в клетку и Ca 2. + из.Другой пример — обменный белок Na + -H + , который функционирует в регуляции внутриклеточного pH. Антипортер Na + -H + связывает транспорт Na + в клетку с экспортом H + , тем самым удаляя избыток H + , образующийся в результате метаболических реакций, и предотвращая подкисление цитоплазмы.
Рисунок 12.33
Примеры антипорт. Ca 2+ и H + экспортируются из клеток антипортерами, которые связывают свой экспорт с энергетически выгодным импортом Na + .
Box
Молекулярная медицина: муковисцидоз.
Вопросы
Случайное перемещение веществ из регионов с низкой концентрацией в регионы с более высокой концентрацией называется диффузией . ( T rue / F alse)
Небольшие локальные концентрации молекул перемещаются и рассеиваются, пока не заполнят доступный объем. ( T rue / F alse)
Обычно вне клетки больше воды (более высокая концентрация), чем внутри.( T rue / F alse)
Обычно вода пытается диффундировать снаружи клетки внутрь. ( T rue / F alse)
Клеточная мембрана разработана таким образом, чтобы позволить практически любому веществу проходить через нее довольно свободно. ( T rue / F alse)
Термин « полупроницаемый » относится к клеточной стенке, которая окружает клетки животных и растений и предотвращает их утечку.( T rue / F alse)
Осмос — это прохождение воды через полупроницаемый барьер. ( T rue / F alse)
Белки-носители расположены на внешних мембранах клеток. ( T rue / F alse)
Белки-носители предотвращают перемещение больших молекул снаружи внутрь клетки. ( T rue / F alse)
Белки-носители работают против эффектов осмоса.( T rue / F alse)
Когда желаемые вещества активно перекачиваются извне внутрь клеток, необходима энергия в виде АТФ. ( T rue / F alse)
Очень большие куски твердых материалов могут быть обернуты частью клеточной мембраны, чтобы стать внутренними пузырьками. ( T rue / F alse)
Эндоцитоз происходит только в растительных клетках.( T rue / F alse)
Фагоцитоз запускается, когда твердый материал вступает в контакт с внешней клеточной мембраной. ( T rue / F alse)
Лизосомы содержат пищеварительные ферменты. ( T rue / F alse)
Пассивный транспорт Определение и примеры
Пассивный транспорт — это тип клеточного транспорта, при котором вещества, такие как ионы и молекулы, перемещаются вниз по их соответствующим градиентам концентрации.Это означает, что вещество имеет тенденцию перемещаться из области с более высокой концентрацией в область с более низкой концентрацией . Вещества имеют тенденцию перемещаться в тот регион, где их мало. Поскольку их движение составляет вниз по или вдоль их градиента концентрации, процесс не требует метаболической энергии (например, АТФ) в отличие от активного транспорта, другого типа клеточного транспорта, который по существу требует, чтобы АТФ перемещал вещества против их градиента концентрации.Четыре основных типа пассивного транспорта: (1) простая диффузия, (2) облегченная диффузия, (3) фильтрация и (4) осмос.
Определение пассивного транспорта
Иллюстрация, показывающая, как происходит пассивный транспорт. Водорастворимые молекулы движутся вниз по градиенту концентрации через мембрану через белок канала (пример облегченной диффузии). Жирорастворимые молекулы легко перемещаются через двухслойную липидную мембрану (пример беспрепятственной диффузии).
Пассивный транспорт определяется как вид транспорта, который перемещает вещества из зоны с большей концентрацией в зону с меньшей концентрацией.Таким образом, движение веществ происходит по градиенту концентрации.
Пассивный транспорт против активного
И пассивный транспорт, и активный транспорт — это механизмы клеточного транспорта, используемые клеткой для перемещения веществ через биологическую мембрану. Пассивный транспорт, однако, отличается от активного транспорта тем, что вещества перемещаются на вдоль в направлении их соответствующего градиента концентрации, в отличие от движения веществ, которое противоречит их градиенту при активном переносе.Движение веществ при пассивном транспорте происходит в направлении, противоположном активному транспорту.
При пассивном переносе вещества (например, ионы и растворенные вещества) перемещаются через мембрану из области их более высокой концентрации в область их более низкой концентрации. Движение описывается как , спуск . По этой причине клеточная энергия (например, АТФ) не требуется, поскольку она находится в активном транспортном состоянии. Скорее, пассивный транспорт использует кинетическую и естественную энергию для движения.И наоборот, в активном транспорте движение составляет в гору , что означает, что они перемещаются из области, где их меньше, в область, где их уже больше. Из-за этого процесс обычно использует АТФ в качестве движущей силы.
Типы пассивного транспорта
При перемещении веществ через биологическую мембрану пассивный транспорт может нуждаться или не нуждаться в помощи мембранного белка. Существует четыре основных типа пассивного транспорта: (1) простая диффузия, (2) облегченная диффузия, (3) фильтрация и (4) осмос.Простая и облегченная диффузия относится к чистому перемещению молекул от более высоких концентраций к более низким. Осмос относится к диффузии растворителя (обычно молекул воды) через полупроницаемую мембрану от более низких до более высоких концентраций растворенных веществ. Фильтрация — это движение молекул воды и растворенных веществ через клеточную мембрану под действием гидростатического давления, создаваемого сердечно-сосудистой системой.
Биологическое значение
Пассивный транспорт важен для правильного функционирования растений и животных.У растений, например, такие газы, как углекислый газ и кислород, диффундируют в растительную клетку и из нее через устьичные отверстия в соответствии с их соответствующими градиентами концентрации. Двуокись углерода важна для растений, поскольку она является одним из основных реагентов фотосинтеза. Одним из источников углекислого газа являются животные, выделяющие газ на выдохе. Этот выброс углекислого газа в окружающую среду опосредуется пассивным транспортом. В частности, диффузия углекислого газа происходит в капиллярных слоях между кровью и тканевой жидкостью.По мере того, как он диффундирует из тканей в кровь, он затем попадает в легкие, где снова диффундирует из крови в альвеолы для выдоха. По мере того, как углекислый газ выходит наружу, кислород, в свою очередь, перемещается в легкие и переносится в ткани тела также путем простой диффузии.
См. Также
Дополнительная литература
© Biology Online. Контент предоставлен и модерируется Biology Online Editors
Клеточная мембрана — анатомия и физиология
Одним из величайших чудес клеточной мембраны является ее способность регулировать концентрацию веществ внутри клетки.Эти вещества включают ионы, такие как Ca ++ , Na + , K + и Cl — ; питательные вещества, включая сахара, жирные кислоты и аминокислоты; и продукты жизнедеятельности, особенно диоксид углерода (CO 2 ), которые должны покинуть ячейку.
Двухслойная липидная структура мембраны обеспечивает первый уровень контроля. Фосфолипиды плотно упакованы вместе, и мембрана имеет гидрофобную внутреннюю часть. Эта структура делает мембрану избирательно проницаемой.Мембрана с избирательной проницаемостью позволяет только веществам, отвечающим определенным критериям, проходить через нее без посторонней помощи. В случае клеточной мембраны только относительно небольшие неполярные материалы могут перемещаться через липидный бислой (помните, липидные хвосты мембраны неполярны). Некоторыми примерами этого являются другие липиды, кислород и углекислый газ, а также спирт. Однако водорастворимые материалы, такие как глюкоза, аминокислоты и электролиты, нуждаются в некоторой помощи для прохождения через мембрану, потому что они отталкиваются гидрофобными хвостами фосфолипидного бислоя.Все вещества, которые проходят через мембрану, делают это одним из двух общих методов, которые классифицируются в зависимости от того, требуется ли энергия или нет. Пассивный транспорт — это движение веществ через мембрану без затрат клеточной энергии. Напротив, активный транспорт — это движение веществ через мембрану с использованием энергии аденозинтрифосфата (АТФ).
Пассивный транспорт
Чтобы понять , как веществ пассивно перемещаются через клеточную мембрану, необходимо понимать градиенты концентрации и диффузию.Градиент концентрации — это разница в концентрации вещества в пространстве. Молекулы (или ионы) будут распространяться / диффундировать от места, где они более сконцентрированы, к месту, где они менее концентрированы, до тех пор, пока они не будут равномерно распределены в этом пространстве. (Когда молекулы движутся таким образом, они, как говорят, перемещаются на вниз на их градиента концентрации.) Диффузия — это перемещение частиц из области с более высокой концентрацией в область с более низкой концентрацией. Несколько общих примеров помогут проиллюстрировать эту концепцию.Представьте, что вы находитесь в закрытой ванной. Если распылить флакон духов, молекулы аромата естественным образом распространятся из места, где они оставили флакон, во все углы ванной комнаты, и это распространение будет продолжаться до тех пор, пока градиент концентрации не исчезнет. Другой пример — ложка сахара, помещенная в чашку чая. В конце концов сахар будет распространяться по всему чаю, пока не исчезнет градиент концентрации. В обоих случаях, если в комнате теплее или чай горячее, диффузия происходит еще быстрее, поскольку молекулы сталкиваются друг с другом и распространяются быстрее, чем при более низких температурах.Таким образом, внутренняя температура тела около 98,6 ° F также способствует диффузии частиц внутри тела.
Посетите эту ссылку, чтобы увидеть диффузию и то, как она приводится в движение кинетической энергией молекул в растворе. Как температура влияет на скорость диффузии и почему?
Когда какое-либо вещество существует в большей концентрации на одной стороне полупроницаемой мембраны, такой как клеточные мембраны, любое вещество, которое может двигаться вниз по градиенту своей концентрации через мембрану, будет делать это.Рассмотрим вещества, которые могут легко диффундировать через липидный бислой клеточной мембраны, такие как газы кислород (O 2 ) и CO 2 . O 2 обычно диффундирует в клетки, потому что он более сконцентрирован вне них, а CO 2 обычно диффундирует из клеток, потому что он более сконцентрирован внутри них. Ни один из этих примеров не требует энергии со стороны клетки, и поэтому они используют пассивный транспорт для перемещения через мембрану.
Прежде чем двигаться дальше, необходимо рассмотреть газы, которые могут диффундировать через клеточную мембрану.Поскольку клетки быстро потребляют кислород во время метаболизма, обычно внутри клетки концентрация O 2 ниже, чем снаружи. В результате кислород будет диффундировать из межклеточной жидкости непосредственно через липидный бислой мембраны в цитоплазму внутри клетки. С другой стороны, поскольку клетки продуцируют CO 2 в качестве побочного продукта метаболизма, концентрации CO 2 повышаются в цитоплазме; следовательно, CO 2 будет перемещаться из клетки через липидный бислой в интерстициальную жидкость, где его концентрация ниже.Этот механизм движения молекул через клеточную мембрану со стороны, где они более сконцентрированы, в сторону, где они менее сконцентрированы, представляет собой форму пассивного транспорта, называемого простой диффузией ([ссылка]).
Простая диффузия через клеточную (плазменную) мембрану
Структура липидного бислоя позволяет небольшим незаряженным веществам, таким как кислород и углекислый газ, и гидрофобным молекулам, таким как липиды, проходить через клеточную мембрану вниз по градиенту их концентрации путем простой диффузии.
Большие полярные или ионные молекулы, которые являются гидрофильными, не могут легко пересечь фосфолипидный бислой. Очень маленькие полярные молекулы, такие как вода, могут пересекаться посредством простой диффузии из-за своего небольшого размера. Заряженные атомы или молекулы любого размера не могут пересечь клеточную мембрану посредством простой диффузии, поскольку заряды отталкиваются гидрофобными хвостами внутри бислоя фосфолипидов. Растворенные вещества, растворенные в воде по обе стороны от клеточной мембраны, будут иметь тенденцию диффундировать вниз по градиенту их концентрации, но поскольку большинство веществ не могут свободно проходить через липидный бислой клеточной мембраны, их движение ограничивается белковыми каналами и специализированными транспортными механизмами в мембране. .Облегченная диффузия — это процесс диффузии, используемый для тех веществ, которые не могут пересекать липидный бислой из-за своего размера, заряда и / или полярности ([ссылка]). Типичным примером облегченной диффузии является перемещение глюкозы в клетку, где она используется для производства АТФ. Хотя глюкоза может быть более концентрированной вне клетки, она не может пересекать липидный бислой посредством простой диффузии, потому что он является одновременно большим и полярным. Чтобы решить эту проблему, специальный белок-носитель, называемый переносчиком глюкозы, будет переносить молекулы глюкозы в клетку, чтобы облегчить ее внутреннюю диффузию.
Облегченная диффузия
(a) Облегченная диффузия веществ через клеточную (плазматическую) мембрану происходит с помощью белков, таких как канальные белки и белки-носители. Канальные белки менее избирательны, чем белки-носители, и обычно легко различают свой груз в зависимости от размера и заряда. (б) Белки-носители более селективны, часто позволяя пересекать только один конкретный тип молекулы.
В качестве примера, хотя ионы натрия (Na + ) сильно сконцентрированы вне клеток, эти электролиты заряжены и не могут проходить через неполярный липидный бислой мембраны.Их диффузии способствуют мембранные белки, которые образуют натриевые каналы (или «поры»), так что ионы Na + могут перемещаться вниз по градиенту их концентрации из-за пределов клеток внутрь клеток. Есть много других растворенных веществ, которые должны пройти через облегченную диффузию, чтобы попасть в клетку, например, аминокислоты, или выйти из клетки, например, отходы. Поскольку облегченная диффузия — это пассивный процесс, он не требует затрат энергии клеткой.
Вода также может свободно перемещаться через клеточную мембрану всех клеток либо через белковые каналы, либо скользя между липидными хвостами самой мембраны.Осмос — это диффузия воды через полупроницаемую мембрану ([ссылка]).
Осмос
Осмос — это диффузия воды через полупроницаемую мембрану вниз по градиенту ее концентрации. Если мембрана проницаема для воды, но не для растворенного вещества, вода будет выравнивать свою концентрацию, диффундируя в сторону более низкой концентрации воды (и, следовательно, в сторону более высокой концентрации растворенного вещества). В стакане слева раствор с правой стороны мембраны гипертонический.
Движение молекул воды само по себе не регулируется клетками, поэтому важно, чтобы клетки подвергались воздействию среды, в которой концентрация растворенных веществ вне клеток (во внеклеточной жидкости) равна концентрации растворенных веществ внутри клеток. (в цитоплазме). Два раствора с одинаковой концентрацией растворенных веществ называются изотоническими (равное натяжение). Когда клетки и их внеклеточная среда изотоничны, концентрация молекул воды одинакова снаружи и внутри клеток, и клетки сохраняют свою нормальную форму (и функцию).
Осмос возникает, когда существует дисбаланс растворенных веществ вне клетки по сравнению с внутри клетки. Раствор, который имеет более высокую концентрацию растворенных веществ, чем другой раствор, называется гипертоническим, а молекулы воды имеют тенденцию диффундировать в гипертонический раствор ([ссылка]). Клетки в гипертоническом растворе будут сморщиваться, когда вода покидает клетку посредством осмоса. Напротив, раствор, который имеет более низкую концентрацию растворенных веществ, чем другой раствор, считается гипотоническим, а молекулы воды имеют тенденцию диффундировать из гипотонического раствора.Клетки в гипотоническом растворе будут поглощать слишком много воды и набухать, что в конечном итоге может привести к разрыву. Важнейшим аспектом гомеостаза живых существ является создание внутренней среды, в которой все клетки тела находятся в изотоническом растворе. Различные системы органов, особенно почки, работают над поддержанием этого гомеостаза.
Концентрация растворов
Гипертонический раствор имеет более высокую концентрацию растворенного вещества, чем другой раствор. Изотонический раствор имеет концентрацию растворенного вещества, равную другому раствору.Гипотонический раствор имеет меньшую концентрацию растворенного вещества, чем другой раствор.
Другой механизм помимо диффузии для пассивной транспортировки материалов между отсеками — фильтрация. В отличие от диффузии вещества, от более концентрированного до менее концентрированного, фильтрация использует градиент гидростатического давления, который выталкивает жидкость — и растворенные в ней вещества — из области с более высоким давлением в область с более низким давлением. Фильтрация — чрезвычайно важный процесс в организме.Например, кровеносная система использует фильтрацию для перемещения плазмы и веществ через эндотелиальную выстилку капилляров в окружающие ткани, снабжая клетки питательными веществами. Давление фильтрации в почках обеспечивает механизм удаления отходов из кровотока.
Активный транспорт
Для всех способов транспортировки, описанных выше, ячейка не расходует энергию. Мембранные белки, которые помогают в пассивном переносе веществ, делают это без использования АТФ.Во время активного транспорта АТФ требуется для перемещения вещества через мембрану, часто с помощью белков-переносчиков, и обычно против его градиента концентрации.
Один из наиболее распространенных типов активного транспорта включает белки, которые служат насосами. Слово «насос», вероятно, вызывает в воображении мысли об использовании энергии для накачки шины велосипеда или баскетбольного мяча. Точно так же энергия АТФ требуется этим мембранным белкам для переноса веществ — молекул или ионов — через мембрану, как правило, против градиентов их концентрации (из области с низкой концентрацией в область с высокой концентрацией).
Натрий-калиевый насос, который также называется Na + / K + АТФаза, транспортирует натрий из клетки, одновременно перемещая калий в клетку. Насос Na + / K + — это важный ионный насос, обнаруженный в мембранах многих типов клеток. Эти насосы особенно распространены в нервных клетках, которые постоянно выкачивают ионы натрия и притягивают ионы калия для поддержания электрического градиента через клеточные мембраны. Электрический градиент — это разница в электрическом заряде в пространстве.В случае нервных клеток, например, существует электрический градиент между внутренней и внешней частью клетки, при этом внутренняя часть заряжена отрицательно (около -70 мВ) относительно внешней стороны. Отрицательный электрический градиент сохраняется, потому что каждый насос Na + / K + перемещает три иона Na + из клетки и два иона K + в клетку для каждой используемой молекулы АТФ ([ссылка] ). Этот процесс настолько важен для нервных клеток, что на него приходится большая часть использования ими АТФ.
Натрий-калиевый насос
Натрий-калиевый насос находится во многих клеточных (плазматических) мембранах. Насос, работающий от АТФ, перемещает ионы натрия и калия в противоположных направлениях, каждый против своего градиента концентрации. За один цикл насоса три иона натрия вытесняются из ячейки, а два иона калия импортируются в ячейку.
Активные транспортные насосы могут также работать вместе с другими активными или пассивными транспортными системами для перемещения веществ через мембрану.Например, натрий-калиевый насос поддерживает высокую концентрацию ионов натрия вне клетки. Следовательно, если клетке нужны ионы натрия, все, что ей нужно сделать, это открыть пассивный натриевый канал, поскольку градиент концентрации ионов натрия заставит их диффундировать в клетку. Таким образом, действие активного транспортного насоса (натрий-калиевый насос) обеспечивает пассивный транспорт ионов натрия, создавая градиент концентрации. Когда активный транспорт обеспечивает перенос другого вещества таким образом, это называется вторичным активным транспортом.
Симпортеры — это вторичные активные переносчики, которые перемещают два вещества в одном направлении. Например, симпортер натрий-глюкоза использует ионы натрия, чтобы «втягивать» молекулы глюкозы в клетку. Поскольку клетки накапливают глюкозу для получения энергии, глюкоза обычно находится в более высокой концентрации внутри клетки, чем снаружи. Однако из-за действия натрий-калиевого насоса ионы натрия легко диффундируют в клетку при открытии симпортера. Поток ионов натрия через симпортер обеспечивает энергию, которая позволяет глюкозе перемещаться через симпортер в клетку против градиента ее концентрации.
И наоборот, антипортеры — это вторичные активные транспортные системы, которые транспортируют вещества в противоположных направлениях. Например, антипортер ионов натрия и водорода использует энергию поступающего внутрь потока ионов натрия для перемещения ионов водорода (H +) из клетки. Натрий-водородный антипортер используется для поддержания pH внутри клетки.
Другие формы активного транспорта не связаны с мембранными переносчиками. Эндоцитоз (попадание «в клетку») — это процесс поглощения клеткой материала путем охвата его частью своей клеточной мембраны, а затем отщипывания этой части мембраны ([ссылка]).После защемления часть мембраны и ее содержимое становятся независимыми внутриклеточными пузырьками. Везикула — это перепончатый мешок — сферическая полая органелла, ограниченная двухслойной липидной мембраной. Эндоцитоз часто приносит в клетку материалы, которые необходимо расщепить или переварить. Фагоцитоз («поедание клеток») — это эндоцитоз крупных частиц. Многие иммунные клетки участвуют в фагоцитозе вторгающихся патогенов. Как и маленькие пак-человечки, их работа — патрулировать ткани тела на предмет нежелательных веществ, таких как вторжение в бактериальные клетки, фагоцитировать и переваривать их.В отличие от фагоцитоза, пиноцитоз («питье клетки») переносит жидкость, содержащую растворенные вещества, в клетку через мембранные везикулы.
Три формы эндоцитоза
Эндоцитоз — это форма активного транспорта, при котором клетка окружает внеклеточные материалы, используя свою клеточную мембрану. (а) При фагоцитозе, который является относительно неселективным, клетка поглощает крупную частицу. (б) При пиноцитозе клетка поглощает мелкие частицы жидкости. (c) Напротив, рецептор-опосредованный эндоцитоз довольно селективен.Когда внешние рецепторы связывают определенный лиганд, клетка отвечает эндоцитозом лиганда.
Фагоцитоз и пиноцитоз захватывают большие части внеклеточного материала, и они, как правило, не обладают высокой селективностью в отношении веществ, которые они вносят. Клетки регулируют эндоцитоз определенных веществ через рецептор-опосредованный эндоцитоз. Рецептор-опосредованный эндоцитоз — это эндоцитоз части клеточной мембраны, содержащей множество рецепторов, специфичных для определенного вещества.Как только поверхностные рецепторы свяжут достаточное количество специфического вещества (лиганда рецептора), клетка будет эндоцитозировать часть клеточной мембраны, содержащую комплексы рецептор-лиганд. Таким образом эритроциты эндоцитируют железо, необходимый компонент гемоглобина. Железо связано с белком, который называется трансферрином в крови. Специфические рецепторы трансферрина на поверхности эритроцитов связывают молекулы железо-трансферрин, и клетка эндоцитирует комплексы рецептор-лиганд.
В отличие от эндоцитоза, экзоцитоз («извлечение из клетки») — это процесс экспорта материала клеткой с использованием везикулярного транспорта ([ссылка]). Многие клетки производят вещества, которые необходимо секретировать, как фабрика, производящая продукт на экспорт. Эти вещества обычно упакованы в мембраносвязанные везикулы внутри клетки. Когда мембрана везикулы сливается с клеточной мембраной, везикула выпускает свое содержимое в интерстициальную жидкость. Затем везикулярная мембрана становится частью клеточной мембраны.Клетки желудка и поджелудочной железы производят и секретируют пищеварительные ферменты посредством экзоцитоза ([ссылка]). Эндокринные клетки производят и секретируют гормоны, которые разносятся по всему телу, а определенные иммунные клетки производят и секретируют большое количество гистамина, химического вещества, важного для иммунных реакций.
Экзоцитоз
Экзоцитоз во многом похож на эндоцитоз в обратном направлении. Материал, предназначенный для экспорта, упаковывается в пузырьки внутри клетки. Мембрана везикулы сливается с клеточной мембраной, и содержимое выходит во внеклеточное пространство.
Ферментные продукты клеток поджелудочной железы
Ацинарные клетки поджелудочной железы производят и секретируют множество ферментов, переваривающих пищу. Крошечные черные гранулы на этой электронной микрофотографии представляют собой секреторные везикулы, заполненные ферментами, которые будут выводиться из клеток посредством экзоцитоза. LM × 2900. (Микрофотография предоставлена Медицинской школой Риджентс Мичиганского университета © 2012)
Болезни…
Cell: Cystic Fibrosis
Муковисцидоз (CF) поражает примерно 30 000 человек в Соединенных Штатах, при этом ежегодно регистрируется около 1000 новых случаев.Это генетическое заболевание наиболее известно своим поражением легких, вызывающим затрудненное дыхание и хронические легочные инфекции, но оно также поражает печень, поджелудочную железу и кишечник. Всего около 50 лет назад прогноз для детей, рожденных с МВ, был очень мрачным — ожидаемая продолжительность жизни редко превышала 10 лет. Сегодня, с развитием медицины, многие пациенты с МВ доживают до 30 лет.
Симптомы CF являются результатом неисправности мембранного ионного канала, называемого регулятором трансмембранной проводимости при муковисцидозе, или CFTR.У здоровых людей белок CFTR является интегральным мембранным белком, который переносит ионы Cl — из клетки. У человека с CF ген CFTR мутирован, таким образом, клетка производит дефектный белок канала, который обычно не включается в мембрану, а вместо этого разрушается клеткой.
CFTR требует для работы АТФ, что делает его Cl — транспортной формой активного транспорта. Эта характеристика долгое время озадачивала исследователей, потому что ионы Cl — на самом деле текут на вниз по градиенту их концентрации на , когда они транспортируются из клеток.Активный транспорт обычно перекачивает ионы против их градиента концентрации, но CFTR представляет собой исключение из этого правила.
В нормальной легочной ткани движение Cl — из клетки поддерживает обогащенную Cl — отрицательно заряженную среду непосредственно за пределами клетки. Это особенно важно в эпителиальной выстилке дыхательной системы. Клетки респираторного эпителия выделяют слизь, которая улавливает пыль, бактерии и другой мусор.Реснички (множественное число = реснички) — это один из волосовидных придатков, обнаруженных на определенных клетках. Реснички на эпителиальных клетках перемещают слизь и захваченные ею частицы по дыхательным путям от легких к внешней стороне. Для эффективного продвижения вверх слизь не может быть слишком вязкой; скорее, он должен иметь жидкую водянистую консистенцию. Транспорт Cl — и поддержание электроотрицательной среды вне клетки привлекают положительные ионы, такие как Na + , во внеклеточное пространство.Накопление ионов Cl — и Na + во внеклеточном пространстве создает богатую растворенными веществами слизь с низкой концентрацией молекул воды. В результате через осмос вода перемещается из клеток и внеклеточного матрикса в слизь, «разжижая» ее. Вот так в нормальной дыхательной системе слизь остается достаточно разбавленной, чтобы ее можно было вытолкнуть из дыхательной системы.
Если канал CFTR отсутствует, ионы Cl — не выводятся из клетки в достаточном количестве, что препятствует захвату ими положительных ионов.Отсутствие ионов в секретируемой слизи приводит к отсутствию нормального градиента концентрации воды. Таким образом, отсутствует осмотическое давление, втягивающее воду в слизь. Образующаяся слизь густая и липкая, и мерцательный эпителий не может эффективно удалить ее из дыхательной системы. Проходы в легких блокируются слизью вместе с мусором, который она переносит. Бактериальные инфекции возникают легче, потому что бактериальные клетки не выводятся из легких.
4.7 Пассивный транспорт — Биология человека
Создал: CK-12 / Адаптировал Кристин Миллер
Рис. 4.7.1. Подобно тому, как окна в доме пропускают свет, клеточная мембрана пропускает определенные вещества в клетку и из нее.Посмотрите на большие окна в этом доме (рис. 4.7.1). Представьте себе весь свет, который они должны пропускать в солнечный день. А теперь представьте, что вы живете в доме, в стенах которого нет окон и дверей. Ничто не могло войти или выйти. Или представьте, что вы живете в доме с дырами в стенах вместо окон и дверей.Вещи могли входить или выходить, но вы не могли контролировать, что входило или выходило. Только когда в доме есть стены с окнами и дверьми, которые можно открывать или закрывать, вы можете контролировать, что входит или выходит. Окна и двери позволяют, например, пропускать свет и домашних собак, а также защищать от дождя и насекомых.
Если бы клетка была домом, плазматическая мембрана была бы стенами с окнами и дверями. Перемещение предметов внутрь и из клетки — важная функция плазматической мембраны. Он контролирует все, что входит и выходит из клетки.Есть два основных пути, которыми вещества могут пересекать плазматическую мембрану: пассивный транспорт, который не требует затрат энергии клеткой, и активный транспорт, который требует энергии от клетки.
Пассивный транспорт происходит, когда вещества пересекают плазматическую мембрану без какого-либо ввода энергии со стороны клетки. Энергия не требуется, потому что вещества перемещаются из области с более высокой концентрацией в область с более низкой концентрацией. Концентрация означает количество частиц вещества на единицу объема.Чем больше частиц вещества в заданном объеме, тем выше концентрация. Вещество всегда перемещается из области с большей концентрацией в область с меньшей концентрацией.
Существует несколько различных типов пассивного транспорта, включая простую диффузию, осмос и облегченную диффузию. Каждый тип описан ниже.
Простая диффузия
Распространение — это движение вещества из-за разницы в концентрации.Это происходит без помощи других молекул. Вещество просто перемещается из области с большей концентрацией в область с меньшей концентрацией. Представьте, как кто-то распыляет духи в углу комнаты. Молекулы духов остаются в углу? Нет, они распространяются или распространяются по комнате, пока не распространятся равномерно. На рис. 4.7.2 показано, как происходит диффузия через клеточную мембрану. Вещества, которые могут протискиваться между молекулами липидов в плазматической мембране путем простой диффузии, обычно представляют собой очень маленькие гидрофобные молекулы, такие как молекулы кислорода и углекислого газа.
Рис. 4.7.2. Молекулы диффундируют через мембрану из области с более высокой концентрацией в область с более низкой концентрацией, пока концентрация не станет одинаковой с обеих сторон мембраны. Рисунок 4.7.3 Осмос — это тип диффузии, при котором только вода может пересекать плазматическую мембрану.Осмос
Осмос — это особый тип диффузии — диффузия молекул воды через мембрану. Как и другие молекулы, вода перемещается из области с более высокой концентрацией в область с более низкой концентрацией.Вода перемещается в клетку или выходит из нее до тех пор, пока ее концентрация не станет одинаковой с обеих сторон плазматической мембраны. На рисунке 4.7.3 пунктирной красной линией показана полупроницаемая мембрана. В первом стакане концентрация растворенных веществ неодинакова по обе стороны от мембраны, но растворенное вещество не может пересекаться — не может происходить диффузия растворенного вещества. В этом случае вода будет двигаться, чтобы выровнять концентрацию, как это произошло в стакане с правой стороны. Уровни воды неравномерны, но процесс осмоса выровнял градиент концентрации.
Облегченная диффузия
Вода и многие другие вещества не могут просто диффундировать через мембрану. Гидрофильные молекулы, заряженные ионы и относительно большие молекулы (например, глюкоза) нуждаются в помощи для диффузии. Эта помощь исходит от специальных белков в мембране, известных как транспортных белков . Диффузия с помощью транспортных белков называется облегченной диффузией . Существует несколько типов транспортных белков, включая канальные белки и белки-переносчики.Оба показаны на рисунке 4.7.4.
- Канальные белки образуют поры (или крошечные отверстия) в мембране. Это позволяет молекулам воды и небольшим ионам проходить через мембрану, не контактируя с гидрофобными хвостами липидных молекул внутри мембраны.
- Белки-носители связываются с определенными ионами или молекулами. При этом они меняют форму. Когда белки-носители меняют форму, они переносят ионы или молекулы через мембрану.
Для нормальной работы клетки ее внутренняя часть должна поддерживать стабильное состояние. Концентрации солей, питательных веществ и других веществ должны поддерживаться в определенных пределах. Состояние, в котором стабильные условия поддерживаются внутри клетки (или всего организма), называется гомеостазом .Гомеостаз требует постоянной корректировки, потому что условия всегда меняются как внутри, так и вне клетки. Транспорт веществ в клетки и из клеток, как описано в этом разделе, играет важную роль в гомеостазе. Позволяя веществам перемещаться в клетки и из них, транспорт поддерживает условия в пределах нормы внутри клеток и во всем организме в целом.
Посмотрите это видео «Транспортировка клеток» от сестер Амеба:
Транспорт клеток с сестрами Амеба, 2016.
- Управление движением предметов внутрь и из клетки — важная функция плазматической мембраны. Есть два основных пути, которыми вещества могут пересекать плазматическую мембрану: пассивный транспорт, который не требует затрат энергии клеткой, и активный транспорт, который требует энергии.
- Клетка не требует энергии для пассивного переноса, потому что это происходит, когда вещества естественным образом перемещаются из области с более высокой концентрацией в область с более низкой концентрацией.
- Простая диффузия — это движение вещества из-за разницы в концентрации. Это происходит без помощи других молекул. Вот как очень маленькие гидрофобные молекулы (например, кислород и углекислый газ) входят и покидают клетку.
- Осмос — это диффузия молекул воды через мембрану. Вода перемещается в клетку или выходит из нее путем осмоса до тех пор, пока ее концентрация не станет одинаковой с обеих сторон плазматической мембраны.
- Облегченная диффузия — это движение вещества через мембрану из-за различий в концентрации, но это происходит только с помощью транспортных белков (таких как канальные белки или белки-переносчики) в мембране.Так большие или гидрофильные молекулы и заряженные ионы входят в клетку и покидают ее.
- Процессы пассивного транспорта играют важную роль в гомеостазе. Позволяя веществам перемещаться в клетку и из клетки, они поддерживают условия в пределах нормы внутри клетки и организма в целом.
- В чем основное отличие пассивного транспорта от активного?
- Обобщите три различных способа пассивной передачи. Приведите пример вещества, которое транспортируется в разные стороны.
- Объясните, как транспорт через плазматическую мембрану связан с гомеостазом клетки.
- В общем, почему только очень маленькие гидрофобные молекулы могут пересекать клеточную мембрану путем простой диффузии?
- Объясните, как облегченная диффузия способствует осмосу в клетках. В своем ответе определите осмос и облегченную диффузию.
- Представьте себе гипотетическую клетку с более высокой концентрацией глюкозы внутри клетки, чем снаружи. Ответьте на следующие вопросы об этой клетке, предполагая, что весь транспорт через мембрану пассивен, а не активен.
- Может ли глюкоза просто диффундировать через клеточную мембрану? Почему или почему нет?
- Если предположить, что в клеточной мембране есть белки, транспортирующие глюкозу, каким путем глюкоза будет течь — в клетку или из нее? Поясните свой ответ.
- Если бы концентрация глюкозы внутри и снаружи клетки была одинаковой, как вы думаете, возник бы чистый поток глюкозы через клеточную мембрану в том или ином направлении? Поясните свой ответ.
- Каковы сходства и различия между канальными белками и белками-носителями?
Осмос и водный потенциал, Amoeba Sisters, 2018.
Структура клеточной мембраны — активный и пассивный транспорт, объясняет профессор Дэйв, 2016.
Атрибуты
Рисунок 4.7.1
Windows / The Oyster Suite в Эврика, Калифорния, автор — Дрю Коффман на Unsplash, используется в соответствии с лицензией Unsplash (https://unsplash.com/license).
Рисунок 4.7.2
Диффузия / Схема простой диффузии в клеточной мембране Мариана Руис Вильярреал [LadyofHats] передана в общественное достояние (https: // en.wikipedia.org/wiki/Public_domain).
Рисунок 4.7.3
Osmosis от OpenStax на Викискладе используется по лицензии CC BY 3.0 (https://creativecommons.org/licenses/by/3.0).
Рисунок 4.7.4
Схема облегченной диффузии в клеточной мембране Марианы Руис Вильярреал [LadyofHats] опубликована в открытом доступе (https://en.wikipedia.org/wiki/Public_domain).
Список литературы
Сестры Амеба. (2016, 24 июня).Клеточный транспорт. YouTube. https://www.youtube.com/watch?v=Ptmlvtei8hw&feature=youtu.be
Сестры Амеба. (2018, 27 июня). Осмос и водный потенциал. YouTube. https://www.youtube.com/watch?v=L-osEc07vMs&feature=youtu.be
Беттс, Дж. Г., Янг, К. А., Уайз, Дж. А., Джонсон, Э., По, Б., Круз, Д. Х., Корол, О., Джонсон, Д. Э., Уомбл, М., Де Сикс, П. (2013, 25 апреля ). Рисунок 3.7 Осмос [цифровое изображение]. В Анатомия и физиология . OpenStax. https: // openstax.организация / книги / анатомия и физиология / страницы / 3-1-the-cell-мембрана
Объясняет профессор Дэйв.
