
Личинки моллюсков (глохидии) — паразиты рыб
Личинки моллюсков (глохидии) в течение определенного отрезка времени паразитируют на рыбах.
На рыбах СССР паразитируют глохидии двустворчатых моллюсков класса Bivalvia. Наиболее широко распространены представители родов Anodonta (беззубка) и Unio (перловица).
Возбудитель. У глохидиев беззубок и перловиц тонкая раковина овально-треугольной Формы с двумя створками, которые соединены мышечной связкой (лигаментом) и могут закрепляться на рыбе при помощи мускула замыкателя. Вершина створок переходит в острый «клюв», снабженный мелкими зубчиками или шипами. Размеры и строение створок глохидиев у разных видов варьируют. Размеры раковин беззубки 0,34X0,28 мм, перловицы обыкновенной — 0,22X0,19.
Развитие моллюсков проходит при участии промежуточных хозяев — рыб. Из оплодотворенных яиц, находящихся в жаберной полости моллюска, формируются личинки — глохидии. В момент приближения рыбы моллюск выталкивает глохидиев через выводной сифон в воду и они прикрепляются к жабрам, плавникам и кожным покровам рыбы, защемляя участки ткани створками.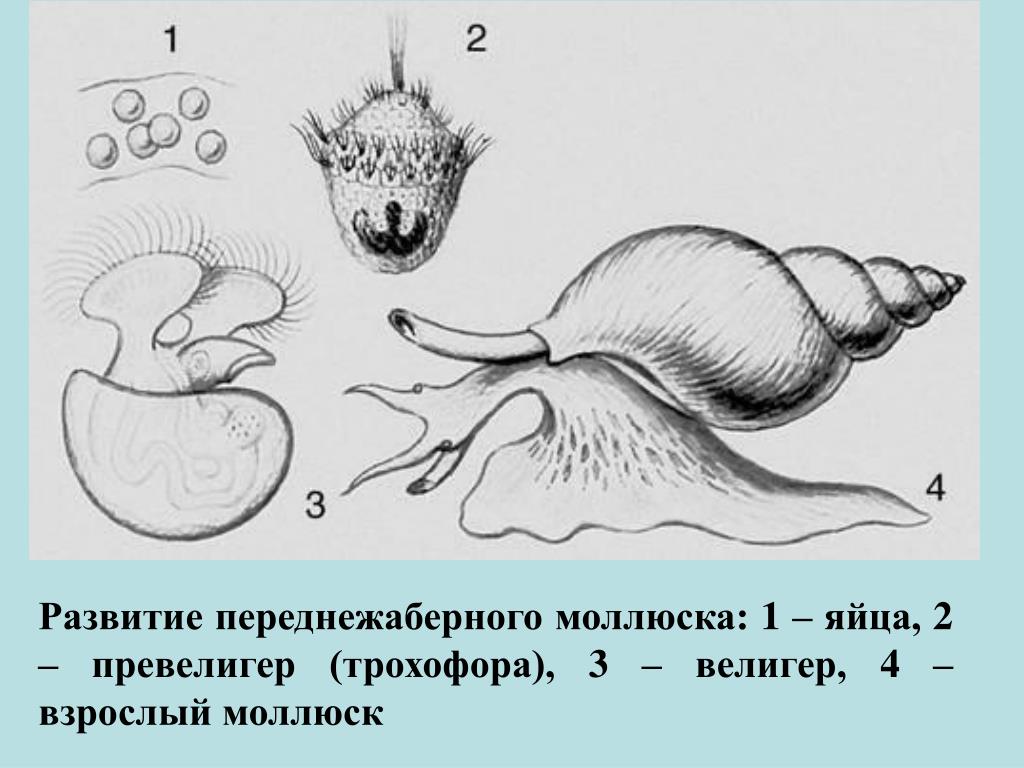
Развитие глохидиев на рыбе в зависимости от температуры продолжается от 8—10 дней до 1—2 мес. Личинка увеличивается в размерах, превращается в молодого моллюска и, разорвав стенку капсулы, выходит в воду и падает на дно.
Эпизоотология. Заражению глохидиями подвержены различные виды пресноводных рыб, особенно щука, окунь, реже — хариус, горчак и др. У перловиц образование глохидиев и массовое заражение рыбы происходит с конца мая по август, у беззубок — весной.
Глохидии перловиц выбрасываются из моллюска сотнями. На одной рыбе может находиться огромное число глохидиев. У окуня длиной около 8 см насчитывалось до 400 личинок, у щуки — до 3—5 тыс. личинок.
Заражение рыбы личинками перловиц происходит обычно в реках, так как перловицы чувствительны к недостатку кислорода в воде. Заражение личинками беззубок наблюдается в прудах, озерах, речных заводях и затонах.
Клинические признаки. В месте прикрепления глохидия эпителий разрастается и образует вокруг личинки капсулу. Находящийся внутри капсулы глохидий питается осмотически за счет хозяина. После выхода молодого моллюска из капсулы пораженное место быстро заживает. Гибели рыб не отмечено. После первого массового заражения у рыб вырабатывается иммунитет. У подуста имеется врожденный иммунитет. В экспериментах отмечено, что хотя на подусте могут поселяться до 600 глохидиев, однако через 2—3 дня рыба освобождается от них полностью.
Диагноз. Диагноз ставят на основании паразитологического анализа и обнаружения большого количества капсул с глохидиями.
Меры борьбы. Ввиду редкости заболевания меры борьбы не предлагаются.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Чужие «хищники»: гигантские китайские моллюски захватили Волгу | Статьи
Двустворчатые моллюски родом из Китая захватили экосистему Нижней Волги.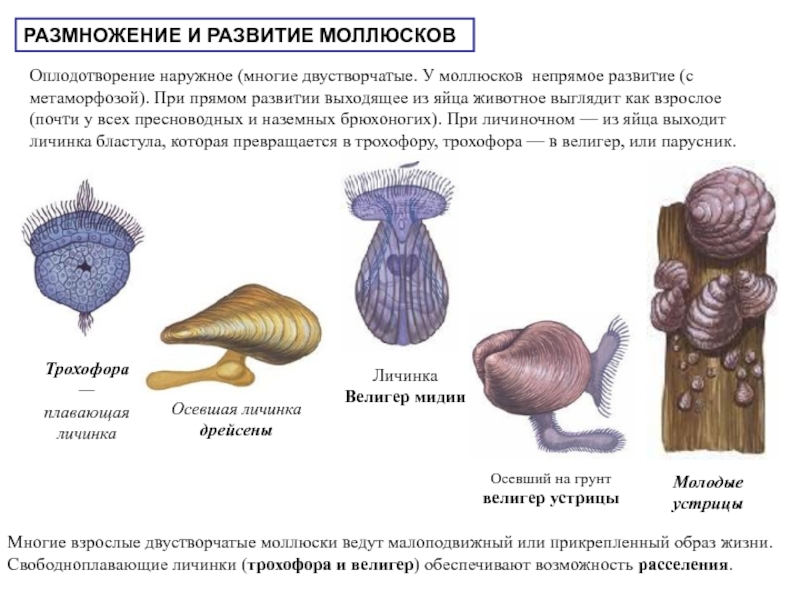 Беззубки диаметром до 25 см используют рыб для воспроизводства и распространения своих личинок (глохидий), прикрепляя их к жабрам и телу обитателей реки. Это приводит к асфиксии у молоди и ухудшению общего состояния взрослых рыб. Кроме того, «чужие» вытесняют местных беспозвоночных животных, приводя к изменению биологического разнообразия и сокращению кормовой базы обитающих в Волге рыб. Ученые выявили также локальные очаги этих видов в реках Обь и Енисей. Неизвестно, сколько рыбоводных хозяйств России сейчас заражено чужеродными моллюсками, но счет может идти на десятки.
Беззубки диаметром до 25 см используют рыб для воспроизводства и распространения своих личинок (глохидий), прикрепляя их к жабрам и телу обитателей реки. Это приводит к асфиксии у молоди и ухудшению общего состояния взрослых рыб. Кроме того, «чужие» вытесняют местных беспозвоночных животных, приводя к изменению биологического разнообразия и сокращению кормовой базы обитающих в Волге рыб. Ученые выявили также локальные очаги этих видов в реках Обь и Енисей. Неизвестно, сколько рыбоводных хозяйств России сейчас заражено чужеродными моллюсками, но счет может идти на десятки.
Транзитом через Казахстан
Биологи Федерального исследовательского центра комплексного изучения Арктики имени академика Н.П. Лавёрова Уральского отделения РАН (Архангельск) обнаружили присутствие в реках и озерах несвойственных российским водоемам китайских беззубок из рода синанодонта (научные названия — Sinanodonta lauta и Sinanodonta woodiana), сообщила пресс-служба Минобрнауки России.
Это гигантские моллюски — их раковины могут достигать размера суповой тарелки (20–25 см в диаметре). Факты инвазий были подтверждены в ходе экспедиций 2017–2019 годов и анализа собранных материалов. Новые для отечественных пресноводных экосистем виды моллюсков, по всей видимости, были завезены из Казахстана вместе с рыбопосадочным материалом предприятиями, занимающимися разведением рыбы.
Факты инвазий были подтверждены в ходе экспедиций 2017–2019 годов и анализа собранных материалов. Новые для отечественных пресноводных экосистем виды моллюсков, по всей видимости, были завезены из Казахстана вместе с рыбопосадочным материалом предприятиями, занимающимися разведением рыбы.
Фото: commons.wikimedia.org
Туда они попали также вместе с рыбой. Известно, что в период 1960–1970-х годов в водоемы Казахской ССР заселялся толстолобик, карп и амурский бычок. Китайские беззубки впервые были обнаружены в озере Балхаш в 1973 году.
Архангельские ученые собрали генетические данные, подтверждающие, что беззубки попали в Казахстан из Китая: Sinanodonta woodiana — из бассейна реки Янцзы, Sinanodonta lauta — из Сунгари, китайского притока Амура.
Неизвестно, сколько рыбоводных хозяйств России сейчас заражено чужеродными моллюсками, но счет может идти на десятки. В 2019 году эти крупные дальневосточные беззубки были обнаружены и в Волге. Анализ возраста собранных моллюсков по годовым кольцам прироста раковины показывает, что инвазия в реке могла начаться в первой половине 2000-х годов.
Анализ возраста собранных моллюсков по годовым кольцам прироста раковины показывает, что инвазия в реке могла начаться в первой половине 2000-х годов.
Чужие в Волге
Сравнительно теплый климат Нижней Волги благоприятствует вселенцам. В этом заключается серьезная экологическая проблема. По сути, сейчас Нижняя Волга наводнена гигантскими чужеродными моллюсками, которых, по сведениям ученых, практически невозможно убрать из экосистемы.
— Уже сегодня можно говорить о сильном преобразовании водных биоценозов Нижней Волги из-за появления этих чужеродных видов, — заявил директор Федерального исследовательского центра комплексного изучения Арктики имени академика Н.П. Лавёрова УрО, член-корреспондент РАН Иван Болотов. — Это может привести к катастрофическим последствиям для экосистем, поскольку массовые поселения таких крупных беззубок могут способствовать вымиранию местных видов моллюсков и других беспозвоночных, сокращению кормовой базы бентосоядных рыб и снижению численности фитопланктона.
Бентосоядными считаются те рыбы, которые питаются тем, что находится на дне и в грунте. Их пища — любые представители макрозообентоса: ракообразные, черви, личинки насекомых и моллюски.
— Бентосоядные рыбы — это основные промысловые животные Волжского бассейна, — пояснил ихтиолог, директор Института комплексных исследований Арктики ФИЦКИА УрО РАН Александр Носовелов. — В основном это осетровые и карповые, составляющие основу промысла в Нижней Волге, где они, согласно особенностям биологии, питаются бентосом.
Фото: ИЗВЕСТИЯ/Константин Кокошкин
Пресноводные двустворчатые моллюски на ранней стадии развития паразитируют на рыбах. Глохидии (их личинки. — «Известия») крепятся к жабрам или к телу рыб и таким образом могут массово мигрировать в естественной среде обитания.
— Большое скопление глохидий на жабрах может привести к банальной асфиксии рыб, — глохидии нарушают процесс естественного поглощения кислорода из воды, и молодь гибнет от этого в первую очередь, а взрослые рыбы испытывают определенный дискомфорт, — подчеркнул Александр Новоселов. — Глохидии, в большом количестве оседая на теле рыб, также являются фактором беспокойства, проявляя себя как внешний паразит.
— Глохидии, в большом количестве оседая на теле рыб, также являются фактором беспокойства, проявляя себя как внешний паразит.
По мнению ученых, снижение численности осетровых в Волге в какой-то мере может быть связано с прогрессирующим глохидиозом — массовым паразитированием личинок моллюсков на рыбе, не адаптированной к такого рода заболеваниям.
Нужна проверка
Уже сегодня на Нижней Волге в районе Астрахани доля особей этих беззубок в благоприятных для них местах составляет от 20 до 40% от общего количества отобранных со дна двустворчатых животных. Многочисленные сообщения о находках гигантских моллюсков в СМИ и социальных сетях Астраханской области в последние годы также свидетельствуют о тенденции. Местные виды никогда не достигают таких величин, как инвазивные.
Фото: inaturalist.org
По мнению ученых, высока вероятность дальнейшего распространения инвазивных моллюсков вместе с молодью рыб из зараженных хозяйств. Более того, поскольку на Нижней Волге «китайские поселенцы» натурализовались и встречаются повсеместно, вероятно массовое заражение волжской рыбы глохидиями.
Более того, поскольку на Нижней Волге «китайские поселенцы» натурализовались и встречаются повсеместно, вероятно массовое заражение волжской рыбы глохидиями.
— Эту рыбу нельзя использовать для переселения в незараженные реки, в которых чужеродные беззубки отсутствуют, — уверен Иван Болотов. — Одним из основных путей распространения инвазии по стране могут служить массовые выпуски молоди с целью компенсации экологических ущербов. Нужны обязательные проверки рыбы на заражение личинками беззубок.
Также необходимо срочно провести проверку максимального количества рыбоводных хозяйств и водоемов России для выяснения истинных масштабов инвазии китайских беззубок. Это позволит разработать программу мер по снижению негативных последствий для экономики и экосистем нашей страны, считают ученые.
— Данный вид моллюсков никогда не рассматривался в России в качестве промыслового объекта, поэтому наше подведомственное научное учреждение ФГБНУ ВНИРО не занималось его изучением, — говорится в ответе Росрыболовства на запрос «Известий». — Упомянутые виды относятся к нежелательным инвазивным видам, и контроль за их распространением находится в ведении Министерства природных ресурсов и экологии Российской Федерации.
— Упомянутые виды относятся к нежелательным инвазивным видам, и контроль за их распространением находится в ведении Министерства природных ресурсов и экологии Российской Федерации.
В Минприроды «Известиям», в свою очередь, сказали, что «данный вопрос находится в компетенции Росрыболовства».
Ученые нашли в реках России опасных моллюсков из Китая — Российская газета
Биологи Федерального исследовательского центра комплексного изучения Арктики обнаружили в реках и озерах России гигантских моллюсков родом из Китая, которые паразитируют на рыбах, угрожая их существованию.
Как сообщает Минобрнауки РФ, двустворчатые моллюски размером с суповую тарелку — китайские беззубки — были обнаружены в ходе экспедиций на нижней Волге и в сибирских реках Обь и Енисей.
Ученые проследили возможный путь чужеродных моллюсков. Предполагается, что в 1960-е годы они попали из Китая в Казахстан вместе с рыбой, которой заселяли водоемы этой республики. Оттуда беззубок завезли на территорию России с посадочным материалом для рыбоводных хозяйств. Первым местом обитания моллюсков могло стать Беловское водохранилище на Оби в Кемеровской области, где занимаются промышленным разведением рыбы.
Первым местом обитания моллюсков могло стать Беловское водохранилище на Оби в Кемеровской области, где занимаются промышленным разведением рыбы.
По словам директора ФИЦКИА УрО РАН Ивана Болотова, зараженная аквакультура перевозилась в другие хозяйства в разные регионы страны. Это привело к неконтролируемому распространению моллюсков, не свойственных для российских водоемов. Ученые полагают, что беззубками могут быть заражены десятки рыбоводных хозяйств.
В Волге они появились предположительно в начале 2000-х годов и с тех пор размножились в огромных количествах. По мнению исследователей, это создает серьезную экологическую проблему. Чужеродных моллюсков невозможно убрать, они уже изменили донные сообщества реки. Беззубки могут вытеснить ее привычных обитателей и привести к сокращению рыбных ресурсов.
На ранней стадии развития эти моллюски паразитируют на разных видах рыб. Личинки — глохидии — крепятся к жабрам или телу рыб и массово мигрируют. Как отмечает Иван Болотов, снижение численности осетровых в Волге может быть связано именно с прогрессирующим глохидиозом — массовым паразитированием личинок моллюсков на рыбах.
При этом в районе Астрахани доля беззубок доходит уже до 40 процентов от общего количества двустворчатых моллюсков. Местные виды не способны достичь таких показателей. Распространение «чужаков» может привести к катастрофическим последствиям для экосистем, сокращению кормовой базы рыб, считают биологи.
По мнению исследователей, вся волжская рыба уже заражена глохидиями, и ее нельзя использовать для переселения в реки, где этих моллюсков пока нет. Для того чтобы избежать дальнейшего распространения китайской беззубки, необходимо проверять молодь рыб на наличие личинок перед ее массовым выпуском в водоемы, утверждает ученые.
Они также говорят о необходимости проверки максимального количества рыбоводных хозяйств и водоемов страны, которая позволит установить истинные масштабы бедствия. Затем надо будет разработать меры по снижению негативных последствий для экономики и экосистем. Ученые представят свои предложения на этот счет.
Многим российским водным экосистемам угрожает реальная опасность от гигантских моллюсков
— Для нас, конечно, это был шок. В том плане, что они не только натурализовались, они достигли высокой численности.
В том плане, что они не только натурализовались, они достигли высокой численности.
Если у учёных шок, то дело действительно серьёзное. Речь идёт о гигантских моллюсках, раковины которых размером с тарелку — до 25 сантиметров в диаметре и больше. Они уже представляют опасность для ряда экосистем российских рек и озёр. И, в первую очередь, угрожают нижней Волге. К такому выводу пришли архангельские биологи по результатам экспедиций последних двух лет. По словам доктора биологических наук Ивана Болотова, чужеродные двустворчатые беззубки образуют очень плотные поселения в грунте, — их раковины стоят вертикально, зарывшись наполовину или на две трети — одна к одной, что внешне очень напоминает паркет или черепицу.
Иван Болотов, директор Федерального исследовательского центра комплексного изучения Арктики Уральского отделения РАН, доктор биологических наук:
— Такие плотные поселения они мешают водообмену в верхних частях грунта, происходят застойные явления, ну и происходит вытеснение местных видов моллюсков, насекомых и других водных животных, которые служат пищей для рыб.
На раковинах можно смело ставить штамп «маde in China», — уже доказано родина инвазивных пришельцев — Китай, реки Янцзы и Сунгари. Правда, завезены гигантские беззубки в Россию из водоёмов Казахстана в 60–70-е годы прошлого века. Сравнительно тёплый климат Волги благоприятствовал новоселам-азиатам. Новые виды моллюсков из одной экосистемы попадали в другую вместе с рыбопосадочным материалом от предприятий, разводящих рыбу. Как правило на ранней стадии развития моллюсков их глохидии — личинки паразитируют на рыбах, прикрепляясь к жабрам или к телу, потом отделяются. В условиях родной экосистемы, к примеру у северной жемчужницы, весь этот процесс для той же сёмги протекает безболезненно. Но в отношениях чужеродных гигантов и волжских рыб есть проблемы.
Иван Болотов, директор Федерального исследовательского центра комплексного изучения Арктики Уральского отделения РАН, доктор биологических наук:
— Я уверен, что этот глохидиоз не будет так легко проходить, потому что глохидии эти очень крупные личинки у китайских беззубок, и рыба совсем к ним не адаптирована. То есть, это может вызывать или асфиксию какую-то, или общее ухудшение состояния рыб.
То есть, это может вызывать или асфиксию какую-то, или общее ухудшение состояния рыб.
По мнению биолога, снижение численности осетровых в Волге может быть связано с прогрессирующим глохидиозом — массовым паразитированием личинок моллюсков на рыбе, не адаптированной к такого рода заболеваниям. Поэтому большие поселения крупных беззубок будут способствовать вымиранию местных видов. А, как известно, моллюски являются индикаторами чистоты среды в целом.
Александр Кондаков, ведущий научный сотрудник Института биогеографии и генетических ресурсов, кандидат технических наук:
— Они по-разному ассимилируют тяжёлые металлы. Наши в большей степени очищают воду, и соответственно в большей степени забирают как раз тяжёлые металлы, какие-то компоненты на себя. И когда вода сильно грязная, соответственно, они начинают активно погибать. А эти виды будут чувствовать себя более комфортно, они быстрее становятся половозрелыми, быстрее развиваются. У них ряд преимуществ, которыми они обладают по-сравнению с нашими видами.
Перемещение заражённой рыбы от хозяйства к хозяйству привело к бесконтрольному расселению китайских беззубок по России, а также и Европе. Казалось бы, для водоёмов северных широт проблема пока не актуальна. Но это может изменить глобальное потепление. Причём другой вид азиатского моллюска уже обнаружен в тёплой водной зоне Архангельской ТЭЦ. Учёные Поморья считают, что необходимо срочно провести проверку рыбоводных хозяйств и водоёмов страны, чтобы представить истинные масштабы расселения китайских беззубок. И разработать программу мер по снижению негативных последствий для экономики и экосистем России.
Чем опасны паразиты в морепродуктах
Юлия Макеева
Общеизвестно, что морепродукты богаты белком, йодом, витаминами и минералами. Но употреблять их нужно с осторожностью, предупреждает эксперт «ВиЖ» Ольга Иванова. Зараженность ракообразных и моллюсков глистами составляет примерно 28%. Большая часть паразитов погибает при термической обработке или уничтожается иммунной системой человека, однако некоторые могут вызвать у человека серьезные заболевания.
Анизакидоз
Паразиты анизакиды вызывают серьезное заболевание анизакидоз.
Это круглые черви, которые поражают крупных морских млекопитающих, а также кальмаров и каракатиц. Личинки анизакид могут жить в устрицах и прочих видах моллюсков, уточнила Ольга Иванова.
Этот паразит длиной до 5 см имеет мощный рот-присоску с тремя губами. Анизакиды –чрезвычайно жизнеспособные паразиты, которые могут существовать в суровых условиях. Их второе название – сельдяной червь.
Человек может заразиться анизакидозом при употреблении сырого или недостаточно термически обработанного мяса морских обитателей.
«Впервые анизакидоз был диагностирован в 1955 году в Голландии, где заболевание стало причиной смерти местного жителя. Заражение произошло в результате регулярного употребления малосольной рыбы», – рассказала Ольга Иванова.
В организме личинки анизакид не превращаются во взрослых червей и не способны к размножению. Однако даже в таком незрелом состоянии паразиты наносят непоправимый вред здоровью человека.
Глисты заселяют полностью весь пищеварительный тракт, вызывая серьезные последствия:
· воспалительные процессы, язвы, прободения;
· перфорация (пробуравливание) стенки желудка, кишечника;
· разрывы стенки кишечника, непроходимость;
· мигрируют в поджелудочную железу, печень;
· поселяются в желчных, почечных протоках.
В запущенных случаях большие колонии глистов проникают в брюшную полость, становятся причиной перитонита и летального исхода.
Ангиостронгилёз
Еще одно опасное для человека заболевание – ангиостронгилёз, возбудителем которого является нематода Angiostrongylus cantonensis. Человек заражается этими паразитами при употреблении в пищу различных видов моллюсков, крабов и креветок в сыром или полусыром виде.
«Паразит вызывает эозинофильные менингоэнцефалиты. Нередко личинок находят в спинном и головном мозге, передней камере глаза», – рассказала Ольга Иванова.
Заболевшего ангиостронгилёзом человека мучают сильные головные боли, наблюдается тошнота и рвота, лихорадка, поражение глазодвигательных и других черепных нервов, эпилептические припадки, параличи, спутанность сознания. В тяжелых случаях возможен смертельный исход.
В тяжелых случаях возможен смертельный исход.
Профилактика
Любителям морепродуктов эксперт советует придерживаться следующих правил:
· покупать морские деликатесы в крупных супермаркетах, где известные поставщики;
· не употреблять в пищу сырые, малосольные, копченые морепродукты;
· свежие тушки осматривать, подвергать тщательной чистке от внутренностей;
· дополнительно замораживать, термически обрабатывать.
В солевых растворах и уксусной кислоте личинки глистов сразу не умирают, уточняет эксперт. При температуре -40°С паразиты погибают за 40 минут, при температуре +60 °С – за 10 минут.
Общая характеристика Типа Моллюски — урок. Биология, Животные (7 класс).
Моллюски (или Мягкотелые) — один из самых крупных типов животных. Известно свыше \(130\) тысяч видов моллюсков. Моллюски живут в водной и наземно-воздушной среде. Есть среди них и паразиты.
Моллюски — животные с мягким телом, которое у большинства видов защищено раковиной. У них есть мантия, мантийная полость, незамкнутая кровеносная система. Двустворчатые и головоногие моллюски двусторонне-симметричные, а у брюхоногих тело асимметричное.
У зародышей моллюсков вторичная полость тела, но у взрослых особей она остаётся только в виде двух участков: полости околосердечной сумки и полости половой железы. Между органами в теле моллюсков находится паренхима (рыхлая соединительная ткань).
Тип Моллюски объединяет классы: Брюхоногие, Двустворчатые, Головоногие.
Рис. \(1\). Классы моллюсков
Внешнее строение
Тело моллюсков несегментировано и состоит из головы (её нет у Двустворчатых), туловища и ноги.
Рис. \(2\). Внешнее строение
На голове находится рот и органы чувств: щупальца и глаза.
Нога — это мускулистый орган движения, с помощью которого моллюск ползает, плавает, роет.
У большинства моллюсков имеется раковина.
Туловище Моллюсков покрыто кожной складкой — мантией (вещество, из которого строится раковина, выделяется клетками мантии). Пространство между стенками туловища и мантией называется мантийная полость. В ней расположены органы дыхания. В мантийную полость открываются анальное, половое и выделительное
отверстия.
Мантия моллюска — это кожная складка между телом и раковиной.
Мантийная полость — это пространство между стенками туловища и мантией.
Органы моллюсков объединены в системы: пищеварительную, дыхательную, кровеносную, нервную, выделительную, половую.
Пищеварительная системаСтроение пищеварительной системы зависит от типа питания моллюсков. В ней выделяют ротовую полость, глотку, пищевод, желудок и кишечник. В кишечник поступают соки, вырабатываемые пищеварительной железой (печенью). Заканчивается пищеварительный тракт анальным отверстием.
В кишечник поступают соки, вырабатываемые пищеварительной железой (печенью). Заканчивается пищеварительный тракт анальным отверстием.
Рис. \(3\). Пищеварительная система
Дыхание у моллюсков, живущих в воде, осуществляется жабрами, а у наземных — с помощью лёгкого. Некоторые водные моллюски (например, прудовики) также дышат лёгкими, периодически поднимаясь к поверхности воды, чтобы вдохнуть.
Рис. \(4\). Органы дыхания
Кровеносная система
В кровеносной системе моллюсков есть сердце — мышечный орган, перекачивающий кровь. Сердце состоит из камер: желудочка и одного или нескольких предсердий.
Моллюски имеют незамкнутую кровеносную систему (за исключением головоногих). У них кровь из кровеносных сосудов сначала поступает в промежутки между органами, а затем опять в сосуды, по которым переносится к органам дыхания (лёгким или жабрам) для обогащения кислородом.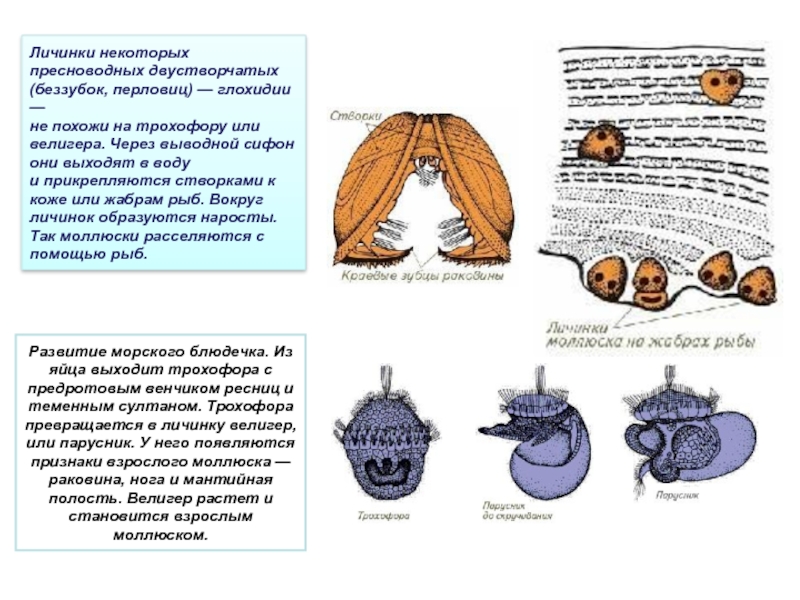
Рис. \(5\). Кровеносная система
Нервная система и органы чувств
У большинства моллюсков нервная система разбросанно-узлового типа. Она образована расположенными в разных частях тела парными ганглиями (нервными узлами) и отходящими от них нервами. Самая сложная нервная система у Головоногих моллюсков.Выделительная система
У моллюсков выделительная система представлена одной или несколькими почками. Продукты выделения (мочевая кислота) выводятся через выделительные отверстия в мантийную полость.
Рис. \(6\). Выделительная и половая системы
Моллюскам характерно только половое размножение. Среди них есть раздельнополые виды (двустворчатые, головоногие) и гермафродиты (брюхоногие). Размножаются моллюски, откладывая оплодотворённые яйца.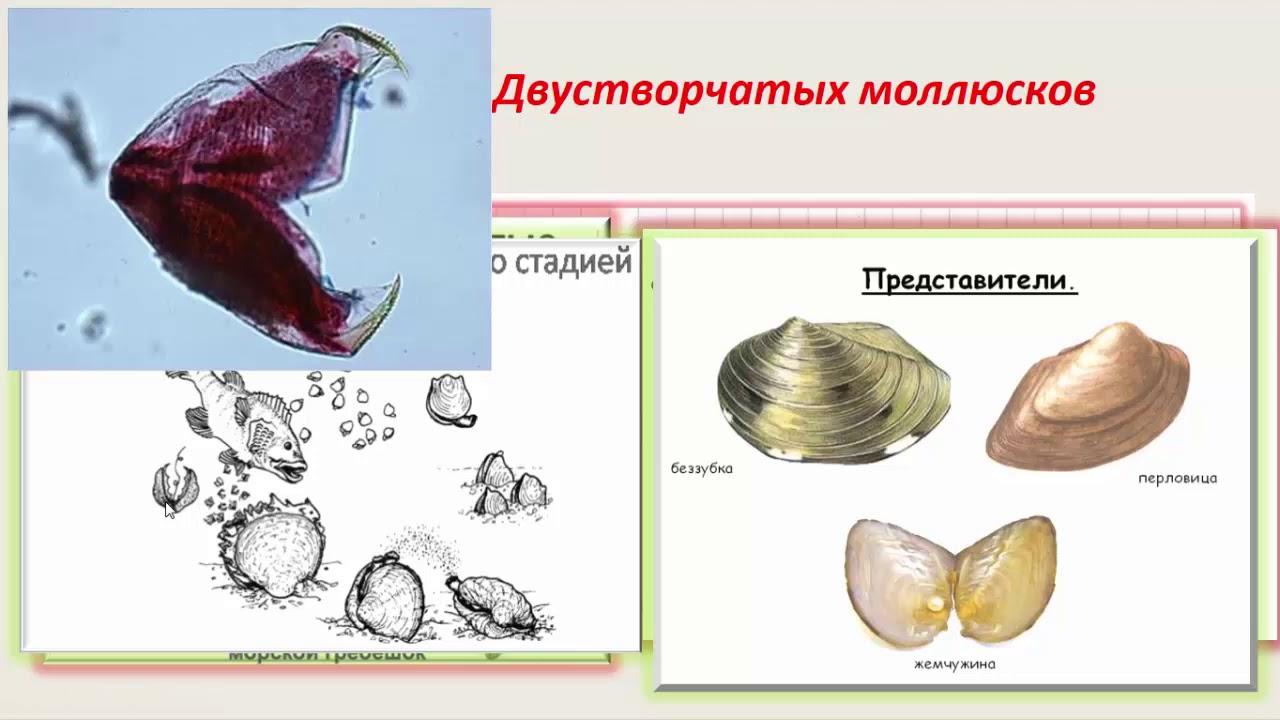
Оплодотворение у них наружное (у двустворчатых) или внутреннее (у брюхоногих и головоногих). Развитие непрямое или прямое в зависимости от образа жизни: из яйца выходит планктонная личинка или маленький моллюск.
Происхождение
Учёные предполагают, что Моллюски имеют общих предков с Кольчатыми червями, у которых ещё не было сегментации тела. Эволюционные связи между типами можно предположить, если сравнить их развитие.
Типичная личинка морских моллюсков (парусник) очень похожа на личинки многощетинковых червей.
Рис. \(7\). Парусник
Источники:
Рис. 1. Классы моллюсков. © ЯКласс.
Рис. 2. Внешнее строение. https://cdn.pixabay.com/photo/2017/07/30/21/15/snail-2555657_960_720.
Рис. 3. Пищеварительная система. © ЯКласс.
Рис. 4. Органы дыхания. © ЯКласс.
Рис. 5. Кровеносная система. © ЯКласс.
Рис. 6. Выделительная и половая системы. © ЯКласс.
Рис. 7. Парусник. © ЯКласс.
Аллерген p4 — Anisakis Larvae, IgE
Аллерген p4 (Anisakis Larvae, IgE) – это определение аллерген-специфических иммуноглобулинов класса E к белкам личинок паразитического червя Anisakis.
Синонимы русские
Аллерген-специфический IgE к анисакис ларве.
Синонимы английские
Anisakis, IgE(p4).
Метод исследования
Иммунохемилюминесцентный анализ.
Единицы измерения
МЕ/мл (международная единица на миллилитр).
Какой биоматериал можно использовать для исследования?
Венозную кровь.
Как правильно подготовиться к исследованию?
- Не курить в течение 30 минут до исследования.
Общая информация об исследовании
Anisakis simplex (анизакис, сельдяной червь) – это гельминт, паразитирующий у морских млекопитающих, попадающий в организм после употребления в пищу термически не обработанных инфицированных рыб (сельдь, скумбрия, анчоусы, сардины, треска) или моллюсков (кальмар).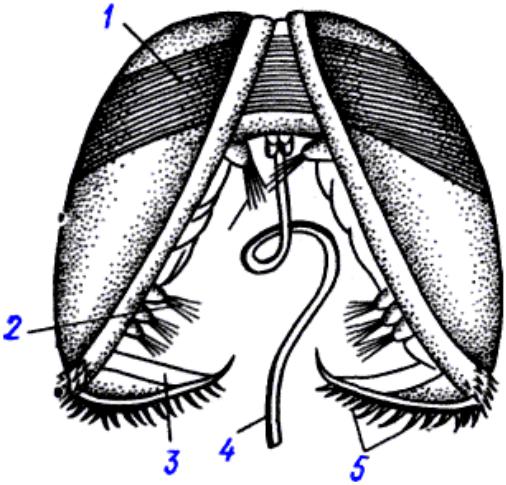
Живые личинки проникают в стенки желудка или кишечника, вызывая симптомы гастроинтестинального анизакидоза – тошноту, рвоту, острую боль в животе, диарею. В последнее время отмечается тенденция к росту числа случаев анизакидоза во всем мире в связи с популярностью блюд из свежей рыбы и моллюсков (суши).
Личинки Anisakis содержат множество аллергенов, запускающих IgE-опосредованные реакции у 35% пациентов с анизакидозом. Низкая перекрёстная реактивность отмечается к аллергенам аскарид, несмотря на родовую близость. Также аллергены имеют сходные эпитопы с некоторыми белками насекомых и ракообразных.
Симптомы аллергии включают в себя появление зуда и генерализованной сыпи на коже, отек слизистых, приступы бронхиальной астмы, отек Квинке, анафилактический шок.
В последующем аллергические реакции могут запускаться при употреблении в пищу продуктов, инфицированных гельминтами, в том числе и после заморозки, термической обработки, консервации, в связи с высокой устойчивостью аллергенов Anisakis.
Сенсибилизация к аллергенам личинок Anisakis подтверждается определением высокого титра специфических IgE в сыворотке крови пациентов. Это может быть полезно в случаях дифференциальной диагностики аллергических состояний, возникающих после употребления в пищу рыбы и морепродуктов. Пациенты с аллергией на белки личинок паразитического червя Anisakis имеют отрицательные результаты при определении сывороточных антител IgE к рыбе и положительные результаты уровня специфических антител IgE к Anisakis Larvae.
Для чего используется исследование?
- Для диагностики аллергических заболеваний, связанных с сенсибилизацией к белкам личинок паразитического червя Anisakis.
- Для дифференциальной диагностики аллергии на морскую рыбу и морепродукты.
Когда назначается исследование?
- При симптомах аллергии, возникающих сразу или через непродолжительное время после употребления морской рыбы и моллюсков, инфицированных личинками гельминта Anisakis.

Что означают результаты?
Референсные значения: 0 — 0,10 МЕ/мл.
Положительный результат
- Аллергия на белки личинок паразитического червя Anisakis.
- Сенсибилизация к аллергенам личинок Anisakis с высоким риском развития аллергической реакции после употребления инфицированной морской рыбы и моллюсков в пищу.
Отрицательный результат
- Отсутствие сенсибилизации к аллергенамличинок паразитического червя Anisakis.
- Исключение контакта с аллергеном.
Кто назначает исследование?
Аллерголог, педиатр, врач общей практики, терапевт, пульмонолог, гастроэнтеролог, хирург.
Литература
1) Pediatric Allergy: Principles and Practice, 18, 167-176.
2) Henry’s Clinical Diagnosis and Management by Laboratory Methods, Chapter 55, 1057-1070.
3) Detection of mosquito saliva–specific IgE and IgG4 antibodies by immunoblotting. Henrikki Brummer-Korvenkontio, Pekka Lappalainen, Timo Reunala, Timo Palosuo.
Henrikki Brummer-Korvenkontio, Pekka Lappalainen, Timo Reunala, Timo Palosuo.
4) Nelson Textbook of Pediatrics, Chapter 146, 1121-1124.
5) The Anisakis allergy debate: does an evolutionary approach help? Carmen Cuéllar and Marta Rodero. Trends in Parasitology, 2012-01-01, Volume 28, Issue 1, Pages 9-15.
Личинки устриц – обзор
Механизмы и моделирование распространения
Влияние гидрографии на распространение личинок является ключом к пониманию экологической связи между популяциями (Cowen, 2006; Cowen et al., 2007). Вернер, Коуэн и Пэрис (2007) перечисляют следующие физические процессы, среди прочего, как относящиеся к делу: течения, вызванные ветром и плавучестью, фронты и связанные с ними струи, приливы, а также поверхностные и придонные пограничные слои. Largier (2003) использовал модель адвекции с диффузией, чтобы показать, как взаимодействие между этими двумя компонентами потока может создавать широкий спектр вариантов рассредоточения.Если адвекция больше диффузии, то будет преобладать расселение вниз по течению от натальной популяции личинок; если диффузионный поток больше адвекции, перенос личинок будет разнонаправленным. Схема рассредоточения становится более сложной там, где противоположные течения сходятся в прибрежной зоне, вызывая отклонение воды от берега. Это создает барьер для расселения, который ограничивает личиночный обмен между сходящимися водными массами (Hare, Guenther, & Fagan, 2005; см. главу 1). Учитывая, что пространственное распределение адвективных потоков может меняться в зависимости от сезона, можно предположить, что организмы могут фактически влиять на свое собственное расселение, определяя время нереста по отношению к доминирующим схемам течения, а также изменяя продолжительность периода нереста (Morgan, 1995).
Схема рассредоточения становится более сложной там, где противоположные течения сходятся в прибрежной зоне, вызывая отклонение воды от берега. Это создает барьер для расселения, который ограничивает личиночный обмен между сходящимися водными массами (Hare, Guenther, & Fagan, 2005; см. главу 1). Учитывая, что пространственное распределение адвективных потоков может меняться в зависимости от сезона, можно предположить, что организмы могут фактически влиять на свое собственное расселение, определяя время нереста по отношению к доминирующим схемам течения, а также изменяя продолжительность периода нереста (Morgan, 1995).
В обзоре литературы Pelc et al. (2009) провели различие между видами с потенциалом расселения на большие расстояния и видами с более ограниченным (ограниченным) расселением. Первые виды демонстрируют ареалы, в которых филогеографические разрывы в целом совпадают с современными гидрографическими барьерами для расселения. Однако у видов с более ограниченным расселением связь между филогеографическими перерывами и океанографическими особенностями была менее очевидной, и для этих видов длительное воздействие исторических (включая эволюционных) факторов помогает объяснить разрозненное распространение существующих взрослых особей. Расселение некоторых эстуарных видов классифицируется как «ограниченное», несмотря на долгоживущую планктотрофную личиночную фазу. В пределах крупных эстуариев время пребывания водной массы может быть большим по сравнению с продолжительностью пелагической фазы . Моделирование распределения личинок устриц в Чесапикском заливе продемонстрировало взаимосвязь между плавательным поведением, соленостью и горизонтальными течениями, ограничивающими распространение (Dekshenieks, Hofmann, Klinck, & Powell, 1996; North et al., 2008).Процессы, ограничивающие распространение личинок устриц их натальными эстуариями, обсуждаются позже (глава 9), но следует отметить, что понимание распространения личинок, полученное в результате изучения 90 007 прибрежных 90 008 видов, не может безоговорочно трансформироваться в более четкое понимание распространения устриц в местах циркуляции 90 007 эстуариев 90 008 видов. применять. Однако в обоих сценариях необходимо учитывать влияние поведения личинок.
Расселение некоторых эстуарных видов классифицируется как «ограниченное», несмотря на долгоживущую планктотрофную личиночную фазу. В пределах крупных эстуариев время пребывания водной массы может быть большим по сравнению с продолжительностью пелагической фазы . Моделирование распределения личинок устриц в Чесапикском заливе продемонстрировало взаимосвязь между плавательным поведением, соленостью и горизонтальными течениями, ограничивающими распространение (Dekshenieks, Hofmann, Klinck, & Powell, 1996; North et al., 2008).Процессы, ограничивающие распространение личинок устриц их натальными эстуариями, обсуждаются позже (глава 9), но следует отметить, что понимание распространения личинок, полученное в результате изучения 90 007 прибрежных 90 008 видов, не может безоговорочно трансформироваться в более четкое понимание распространения устриц в местах циркуляции 90 007 эстуариев 90 008 видов. применять. Однако в обоих сценариях необходимо учитывать влияние поведения личинок.
Учитывая априори, что поведение личинок, вероятно, играет роль в распространении (Shanks, 2009), существуют ли обстоятельства, при которых модель прибрежной циркуляции, игнорирующая поведение личинок при плавании, может быть лучше, чем давать «грубый» прогноз? личиночного транспорта? Гилг и Хилбиш (2003) рассмотрели этот вопрос, применив двухмерную (интегрированную по глубине) модель прибрежных течений с высоким разрешением на юго-западе полуострова Англии для прогнозирования распределения личинок мидий, «выпущенных» (в рамках модели) в различные места вдоль побережья. Они сравнили результаты модели с оценками распространения, в которых использовались генетические маркеры для различения видов Mytilus по обе стороны хорошо охарактеризованной гибридной зоны. Океанографическая модель точно предсказала характер расселения личинок как по направлению, так и по расстоянию; смоделированные расстояния рассеивания составляли ≤ 60 км по сравнению с расчетами на основе генетических данных от 21 до 64 км. Эти результаты подтвердили предположение о том, что физические процессы преобладают над рассеиванием, по крайней мере, в некоторых прибрежных ситуациях.Обобщение этого и других подобных исследований способствовало выработке гипотезы апвеллинга/нисхождения, которая, проще говоря, предполагает, что личинки в прибрежных водах пассивно рассеиваются и, например, уносятся в море во время апвеллинга и возвращаются на берег во время нисхождения. Это было предложено не только как объяснение дисперсии перпендикулярно побережью, но и для объяснения распределения вдоль береговой линии, где происходят сезонные изменения баланса между событиями апвеллинга и нисхождения.
Они сравнили результаты модели с оценками распространения, в которых использовались генетические маркеры для различения видов Mytilus по обе стороны хорошо охарактеризованной гибридной зоны. Океанографическая модель точно предсказала характер расселения личинок как по направлению, так и по расстоянию; смоделированные расстояния рассеивания составляли ≤ 60 км по сравнению с расчетами на основе генетических данных от 21 до 64 км. Эти результаты подтвердили предположение о том, что физические процессы преобладают над рассеиванием, по крайней мере, в некоторых прибрежных ситуациях.Обобщение этого и других подобных исследований способствовало выработке гипотезы апвеллинга/нисхождения, которая, проще говоря, предполагает, что личинки в прибрежных водах пассивно рассеиваются и, например, уносятся в море во время апвеллинга и возвращаются на берег во время нисхождения. Это было предложено не только как объяснение дисперсии перпендикулярно побережью, но и для объяснения распределения вдоль береговой линии, где происходят сезонные изменения баланса между событиями апвеллинга и нисхождения.
Шанкс и Бринк (2005) оспорили эту гипотезу. Они взяли образцы личинок четырех видов двустворчатых моллюсков в прибрежных и прибрежных водах Северной Каролины, США. Данные показали, что вертикальное плавание способно поставить личинок под влияние различных водных масс в стратифицированной толще воды и отвечает за удержание личинок вблизи берега даже при наличии прибрежных течений (рис. 3.12). . Вертикальные скорости плавания того же порядка, что и вертикальные течения воды, и могут быть причиной ухода от морского транспорта в поверхностных течениях во время апвеллинга.Shanks and Shearman (2009; см. также Shanks, Largier, Brink, Brubaker, & Hoof, 2002; Shanks, Grantham, et al., 2003; Shanks, Largier, et al., 2003) взяли пробы личинок беспозвоночных (включая двустворчатых педивелигеров) на разрез между сушей и морем в штате Орегон, США. Как при апвеллинге, так и при опускании в толще воды личинки держались близко к берегу. Они были сосредоточены на средних глубинах, где, избегая переноса поверхностными течениями, была ограничена тенденция к расселению в открытом море. Шанкс и Ширман пришли к выводу, что гипотеза апвеллинга-даунвеллинга в ее самой основной форме (отсутствие личиночного поведения) оспаривается, и что плавательное поведение помогает личинкам поддерживаться на расстоянии от 1 до 4 км от берега, где значительные водные потоки подвержены влиянию прибрежная топография и динамика зоны прибоя. Авторы рекомендовали, чтобы для понимания переноса личинок вдоль побережья и к местам их возможного поселения требовалось больше знаний о гидрографии зоны прибоя и о том, как она взаимодействует с поведением личинок при плавании.
Шанкс и Ширман пришли к выводу, что гипотеза апвеллинга-даунвеллинга в ее самой основной форме (отсутствие личиночного поведения) оспаривается, и что плавательное поведение помогает личинкам поддерживаться на расстоянии от 1 до 4 км от берега, где значительные водные потоки подвержены влиянию прибрежная топография и динамика зоны прибоя. Авторы рекомендовали, чтобы для понимания переноса личинок вдоль побережья и к местам их возможного поселения требовалось больше знаний о гидрографии зоны прибоя и о том, как она взаимодействует с поведением личинок при плавании.
Рис. 3.12. Схематическая диаграмма поведения удерживаемых у берега личинок при подъеме и опускании водной массы. Чтобы личинки удерживались у берега, они должны реагировать на изменения давления, плавая вверх по мере падения давления и опускаясь по мере повышения давления, а скорость плавания должна быть такой же или большей, чем скорость вертикального течения.
(Из Shanks & Brink, 2005)., 2008). Бейкер и Манн (2003), например, заметили, что личинок C. virginica вели себя таким образом, что, вероятно, удерживали их в пределах эстуария — когда личинки приближались к метаморфозу, они оставались у дна водной толщи, в более легкой досягаемости субстраты, благоприятные для прикрепления и подверженные переносу вверх по устью, обусловленному плотностью. В лабораторных экспериментах личинки устриц увеличивали скорость вертикального плавания вниз при наличии химических раздражителей, вызывающих оседание (Turner, Zimmer-Faust, Palmer, Luckenbach, & Pentchef, 1994; обсуждается в главе 9).Этот тип поведения помогает ограничить рассеивание от родительских популяций. В моделях, протестированных North et al. (2008) различия в моделях поведения личинок между двумя видами устриц существенно повлияли как на расстояние расселения, так и на «успех» их транспортировки (доля выпущенных личинок, которые встретили подходящие места для поселения в конце пелагического периода). В гидрографической модели (отслеживание частиц), которая включает поведение личинок — личинки, способные к циклу метаморфоза между плаванием, «стремлением осесть», оседанием или не оседанием, — Broekhuizen, Lundquist, Hadfield, and Brown (2011) пришли к выводу, что большинство личинок выпущено к г.
virginica вели себя таким образом, что, вероятно, удерживали их в пределах эстуария — когда личинки приближались к метаморфозу, они оставались у дна водной толщи, в более легкой досягаемости субстраты, благоприятные для прикрепления и подверженные переносу вверх по устью, обусловленному плотностью. В лабораторных экспериментах личинки устриц увеличивали скорость вертикального плавания вниз при наличии химических раздражителей, вызывающих оседание (Turner, Zimmer-Faust, Palmer, Luckenbach, & Pentchef, 1994; обсуждается в главе 9).Этот тип поведения помогает ограничить рассеивание от родительских популяций. В моделях, протестированных North et al. (2008) различия в моделях поведения личинок между двумя видами устриц существенно повлияли как на расстояние расселения, так и на «успех» их транспортировки (доля выпущенных личинок, которые встретили подходящие места для поселения в конце пелагического периода). В гидрографической модели (отслеживание частиц), которая включает поведение личинок — личинки, способные к циклу метаморфоза между плаванием, «стремлением осесть», оседанием или не оседанием, — Broekhuizen, Lundquist, Hadfield, and Brown (2011) пришли к выводу, что большинство личинок выпущено к г. О.chilensis взрослых особей рассеялись менее чем на 1 км от мест их рождения (Тасман-Бей, Южный остров, Новая Зеландия) в течение 10 дней (предполагаемый максимальный период свободного плавания личинок). В ситуациях, когда водная толща хорошо перемешана, мелкомасштабные модели отслеживания частиц, которые игнорируют поведение при плавании, могут предоставить полезные модели дисперсии и связности (например, Haase, Eggleston, Luettich, Weaver, & Puckett, 2012), но решения не Включение личиночного поведения должно быть обосновано в большинстве случаев.
О.chilensis взрослых особей рассеялись менее чем на 1 км от мест их рождения (Тасман-Бей, Южный остров, Новая Зеландия) в течение 10 дней (предполагаемый максимальный период свободного плавания личинок). В ситуациях, когда водная толща хорошо перемешана, мелкомасштабные модели отслеживания частиц, которые игнорируют поведение при плавании, могут предоставить полезные модели дисперсии и связности (например, Haase, Eggleston, Luettich, Weaver, & Puckett, 2012), но решения не Включение личиночного поведения должно быть обосновано в большинстве случаев. Таинственные пути моллюсков к взрослой жизни – Океанографический институт Вудс-Хоул
С прохладным океанским бризом под безоблачным небом дети плетут по каналу маленькие парусники. Более крупные лодки снуют из залива Вакуит, чтобы порыбачить в проливе Нантакет или провести день на винограднике Марты. Те, кто не на воде, играют в теннис или отдыхают на пляже.
Типичный летний день в яхт-клубе Menauhant в Ист-Фалмуте, штат Массачусетс. Сцена, похожая на открытку, которую можно найти в любом яхт-клубе Кейп-Кода в июле, за исключением спутанного шланга, прикрепленного к насосу, и замусоренных фильтров и бутылки разложены на плавучем доке.Молодая женщина с косичками наклоняется над причалом, чтобы вытащить из воды два больших оранжевых мешка. По большей части люди слишком заняты, чтобы спрашивать, почему она здесь, но иногда некоторые задаются вопросом.
Сцена, похожая на открытку, которую можно найти в любом яхт-клубе Кейп-Кода в июле, за исключением спутанного шланга, прикрепленного к насосу, и замусоренных фильтров и бутылки разложены на плавучем доке.Молодая женщина с косичками наклоняется над причалом, чтобы вытащить из воды два больших оранжевых мешка. По большей части люди слишком заняты, чтобы спрашивать, почему она здесь, но иногда некоторые задаются вопросом.
Познакомьтесь с «девочкой-жуком»
В течение последних трех лет я проводил лето, собирая пробы в заливе Уокуит на наличие личинок моллюсков, морских гребешков и устриц. Пробую всегда и везде, где только можно. Я брал пробы с доков, лодок и каяков; на верфях, яхт-клубах и исследовательских станциях. Я пробовал еженедельно, ежедневно и даже в течение 48 часов подряд.Все эти пробы принесли мне прозвище «Девушка-жук» у некоторых начальников порта в бухте.
Личинки и прочий планктон подобны морским жукам. Подвешенные в воде, плавающие невидимыми стаями, личинки переносятся ветром и приливными течениями, пока не осядут и не вырастут во взрослых особей. Но этот критический этап жизненного цикла остается во многом загадкой. Личинки маленькие, и их пути трудно проследить. Личинки двустворчатых моллюсков практически невозможно идентифицировать даже специалисту.Но я надеюсь преодолеть эти препятствия с помощью своего докторского исследования в рамках совместной программы MIT/WHOI.
Но этот критический этап жизненного цикла остается во многом загадкой. Личинки маленькие, и их пути трудно проследить. Личинки двустворчатых моллюсков практически невозможно идентифицировать даже специалисту.Но я надеюсь преодолеть эти препятствия с помощью своего докторского исследования в рамках совместной программы MIT/WHOI.
Понимание личиночной стадии двустворчатых моллюсков может помочь обратить вспять сокращение популяции вкусных моллюсков. В городе Машпи на восточной стороне залива констебль моллюсков Ричард Йорк проявляет личный интерес к моему исследованию. Он выращивает миллионы взрослых моллюсков, чтобы ежегодно размещать их в заливе для коммерческого и любительского рыболовства, но он не может сказать, производят ли они личинок.
Чтобы найти личинок размером не больше мелкой песчинки, мне приходится фильтровать сотни литров воды и подсчитывать их под микроскопом. Когда люди застают меня за работой, они часто спрашивают меня о качестве воды. Некоторые хотят знать, есть ли поблизости рыба или почему этой весной падали синие крабы. Я не всегда знаю ответы на их вопросы; Я живу на Кейп-Коде всего три года. Многие из тех, кого я встречаю, проводят здесь всю свою жизнь и знают о заливе больше, чем я когда-либо узнаю.
Я не всегда знаю ответы на их вопросы; Я живу на Кейп-Коде всего три года. Многие из тех, кого я встречаю, проводят здесь всю свою жизнь и знают о заливе больше, чем я когда-либо узнаю.
В поисках гребешков
В этот конкретный день в клубе мужчина и его сын тянут свою лодку к причалу, пока я полоскаю оранжевые пакеты в ведре. Красно-бело-синяя тема их одежды напоминает мне о приближении народного праздника, что также отражается в увеличении движения лодок. Мужчина спрашивает меня, может ли он одолжить шланг, чтобы помыть свою лодку. — Могу я спросить, что вы делаете? он говорит.
«Ищу личинок моллюсков», — говорю ему; это мой стандартный ответ на этот распространенный вопрос.Я жду, чтобы оценить интерес человека, прежде чем добавлять больше деталей. На этот раз их интерес, похоже, пробудился, когда сын, которому на вид 10 или 12 лет, вылезает из лодки, чтобы заглянуть в мое ведро.
«Видите?» он спросил.
Я говорю ему, что они маленькие, и вам нужен микроскоп, чтобы их увидеть. Под микроскопом личинки выглядят как прозрачные полукольца с тканью, содержащей органы пищеварения и плавательные придатки. Как и у взрослых особей, у личинок есть панцирь из арагонита, минерала, содержащего кальций.При просмотре с использованием поляризационного фильтра кристаллы в личиночной оболочке заставляют ее отражать яркие цветовые узоры. Создаваемые узоры различны для каждого вида, и с помощью компьютера и программного обеспечения для распознавания изображений их можно использовать для различения отдельных видов.
Под микроскопом личинки выглядят как прозрачные полукольца с тканью, содержащей органы пищеварения и плавательные придатки. Как и у взрослых особей, у личинок есть панцирь из арагонита, минерала, содержащего кальций.При просмотре с использованием поляризационного фильтра кристаллы в личиночной оболочке заставляют ее отражать яркие цветовые узоры. Создаваемые узоры различны для каждого вида, и с помощью компьютера и программного обеспечения для распознавания изображений их можно использовать для различения отдельных видов.
Я говорю мальчику и его отцу, что меня особенно интересуют морские гребешки, и я использую оранжевые сетчатые мешочки для сбора личинок старшего возраста. Я объясняю эвтрофикацию: как возросшее количество септических систем в близлежащих районах загрузило залив азотом из отходов и удобрений, стимулируя безудержный рост крупных водорослей, которые блокируют попадание солнечного света на морскую траву на дне.Это привело к уменьшению морского морского гребешка, важной среды обитания, где личинки лаврового гребешка селятся, чтобы вырасти до взрослой жизни. Я надеюсь, что сетчатые мешки, свисающие с пристани, привлекут личинок морского гребешка, чтобы они поселились на них так же, как это сделали бы морские водоросли.
Я надеюсь, что сетчатые мешки, свисающие с пристани, привлекут личинок морского гребешка, чтобы они поселились на них так же, как это сделали бы морские водоросли.
– Гребешков здесь больше не увидишь, – соглашается сын. Даже 5-10 лет назад их было довольно сложно найти.
Отбор проб с приливами
Я жду встречи со своим кандидатом наук. советник, Скотт Галлагер, ученый из WHOI, и Крис Вайдман, директор по исследованиям в близлежащем Национальном эстуарном исследовательском заповеднике Уаквойт-Бэй.Десятилетия исследований в заповеднике сделали залив Уокуит известным на примере воздействия эвтрофикации на эстуарии. Потеря промысла морского гребешка — это лишь небольшая часть истории, но важная для любителей моллюсков.
Каждое лето взрослый морской гребешок выпускает в воду миллионы яйцеклеток и сперматозоидов. Через сутки после оплодотворения у личинки формируется панцирь, и она начинает свой путь. Личинка до двух недель дрейфует в толще воды, подвергаясь различным опасностям окружающей среды. Лишь очень небольшому проценту удается найти пищу, избежать того, чтобы быть съеденным (или быть засосанным в планктонный насос), и найти подходящее место для поселения и метаморфоза.
Лишь очень небольшому проценту удается найти пищу, избежать того, чтобы быть съеденным (или быть засосанным в планктонный насос), и найти подходящее место для поселения и метаморфоза.
Вода вымывается из залива Вакуит каждые три дня приливами. Если морской гребешок нерестится в бухте, он не может плыть быстрее приливных течений и вряд ли там останется. Он может оказаться вдали от берега, быть унесенным в соседний лиман или погибнуть, прежде чем найдет заросль взморья.
Существует вероятность того, что личинка может погрузиться на дно во время отлива, чтобы избежать движения с самыми сильными течениями, а личинки из прибрежных районов могут попасть в залив во время прилива.Чтобы исследовать эти гипотезы, мне нужно взять образцы за весь приливно-отливный цикл и на разных глубинах.
Скотт и Крис здесь, чтобы помочь получить образцы приливов. Мы устанавливаем автоматическую систему клапанов с фильтрами, подключенными к насосу, для непрерывного отбора проб воды при каждом приливе и отливе. У меня есть родственная система на стороне залива Машпи.
У меня есть родственная система на стороне залива Машпи.
Система запрограммирована на открытие или закрытие одного из четырех клапанов при каждом приливе. Это позволяет мне брать пробы в течение лунного приливного цикла (около 28 дней) и сравнивать поток личинок, которые входят и выходят из залива, но мне не нужно присутствовать, чтобы брать пробы при каждом приливе.
Работа с жуками и мопсами
После того, как мы обсудили планы установки, Скотт и Крис уходят, а я готовлюсь к еженедельному отбору проб. Ко мне подходит мужчина в рубашке поло яхт-клуба, за ним следует его собака — маленький мопс с соответствующим ошейником в морском стиле. Он видит, что я занимаюсь исследованием, и задает мне несколько вопросов, пока я пробую; сам ученый на пенсии, ему любопытно узнать об Океанографическом институте Вудс-Хоул и о проводимых там передовых исследованиях.Он удивлен и доволен тем, что я пользуюсь возможностью попробовать на собственном заднем дворе.
Чтобы применить новые технологии для решения научных вопросов, не обязательно отправляться в экзотические места. В заливе Вакуит теперь мы можем посмотреть на видовой состав моллюсков из каждой пробы планктона, которую я собираю. Это может определить, когда присутствуют виды, как они растут и выживают, и как их поведение при вертикальном плавании влияет на их перемещение с приливными течениями.
В заливе Вакуит теперь мы можем посмотреть на видовой состав моллюсков из каждой пробы планктона, которую я собираю. Это может определить, когда присутствуют виды, как они растут и выживают, и как их поведение при вертикальном плавании влияет на их перемещение с приливными течениями.
Такая фундаментальная недостающая информация необходима для усилий по поддержанию естественной популяции за счет восстановления взморника.С экономической точки зрения эти выборки позволяют менеджерам увидеть результаты своего труда по поддержанию любительского рыболовства. Пока мы заполняем пробелы между личиночной и взрослой стадиями развития моллюсков, мои новые друзья в яхт-клубе могут снова насладиться моллюсками с куахогами из залива.
Пока я разговариваю с этим человеком, его собака смотрит, как я качаю воду из шланга, и начинает тяжело дышать.
«Похоже, он хочет пить».
Мужчина отводит его к ближайшему шлангу, чтобы напиться. Вскоре я заканчиваю свой образец и иду промывать фильтры. Собака все еще пьет, и мне интересно, сколько моих коллег должны делить оборудование с детьми и собаками. Я жду своей очереди.
Собака все еще пьет, и мне интересно, сколько моих коллег должны делить оборудование с детьми и собаками. Я жду своей очереди.
Исследование Кристин Минджионе было поддержано стипендией Национального эстуарного исследовательского заповедника через Национальное управление океанических и атмосферных исследований и грантом Института прибрежных океанов WHOI. Ее также поддержали Фонд Дж. Сьюарда Джонсона и Фонд Ричарда Ванстона. Эта статья была написана во время курса научного письма для аспирантов WHOI при финансовой поддержке Фонда Генри Л.и Грейс Доэрти, профессор океанографии.
Маленькие устрицы не могут строить здоровые раковины в подкисленных водах Вашингтона
Устрицы — один из знаковых продуктов Тихоокеанского Северо-Запада. Но их выживание находится под серьезной угрозой из-за закисления океана, которое иногда называют «злым близнецом» изменения климата.
Местные производители моллюсков наблюдают разрушительное воздействие на устриц и других моллюсков.
С 1989 года инкубаторий Taylor Shellfish вырастил миллиарды и миллиарды личинок устриц.Как и у любой другой фермы, у нее были свои неудачи — времена, когда большое количество устриц умирало по неизвестной причине. Но обычно все возвращалось.
Потом, в сезоне 2007-2008, такие провалы стали нормой.
«У нас не было личинок устриц, — вспоминал Билл Дьюи, директор компании по связям с общественностью. «Наше производство сократилось примерно на 75 процентов».
Вскоре они узнали, что у других производителей на Западном побережье такая же проблема. Дьюи сказал, что они знали, что происходит что-то серьезное.
«У нас был кризис семян устриц. В 2008-2009 годах не было семян для ферм вдоль и поперек Западного побережья», — сказал он.
Сначала они думали, что устриц убивают бактерии. Но даже после тщательной очистки резервуаров и разработки системы фильтрации для удаления бактерий устрицы продолжали умирать. Примерно в то же время Ричард Фили, ученый из Тихоокеанской морской экологической лаборатории NOAA в Сиэтле, связался с производителями и сообщил о недавней находке.
Примерно в то же время Ричард Фили, ученый из Тихоокеанской морской экологической лаборатории NOAA в Сиэтле, связался с производителями и сообщил о недавней находке.
«Он подошел к нам и сказал: «Вот что, я думаю, может происходить». Итак, мы начали изучать химический состав воды. О чудо… это была химия карбонатов. А последствия подкисления океана убивали наших личинок устриц».
Закисление океана происходит, когда морская вода со временем поглощает углекислый газ, снижая уровень pH воды. Маленьким устрицам труднее строить раковины.
Мы находимся в инкубаторе Тейлора в заливе Дабоб, где устрицы и другие моллюски начинают свою молодую жизнь.Дьюи останавливается у одного из многочисленных резервуаров в инкубатории. Он наполнен водой и миллионами личинок устриц, не превышающих в диаметре прядь волос. Это решающий этап молодой жизни устрицы.
«В первые 48 часов жизни устрица должна сделать две вещи: она должна построить раковину и этот орган, с помощью которого она питается и плавает, называется пергаментом», — сказал Дьюи.
Когда вода слишком кислая, яйцо деформируется и погибает. Он израсходовал свою ограниченную энергию на создание оболочки, и ее не осталось для создания пергамента.
Когда они поняли, что их молодые устрицы умирают, производители забили тревогу. Законодатели Олимпии обратили на это внимание. Перед тем, как губернатор Крис Грегуар покинул свой пост в 2011 году, она создала группу Blue Ribbon для решения этой проблемы. С тех пор работа группы была расширена.
Дженнифер Хеннесси, старший политический советник губернатора Джея Инсли, говорит, что закисление океана здесь происходит быстрее, чем в других частях страны.
«Потому что у нас есть апвеллинг на западном побережье Соединенных Штатов, который поднимает коррозионные воды, которые смешиваются с поверхностными водами.Так что мы наблюдаем эти изменения быстрее, чем, скажем, на восточном побережье США или в Мексиканском заливе».
Хеннесси сказал, что прогнозы показывают, что изменения будут происходить еще быстрее в следующие 30 лет, поэтому есть небольшое окно для решения этих проблем.
Тем временем производители моллюсков, такие как Taylor Shellfish, нашли обходной путь, позволяющий решить проблему в краткосрочной перспективе. Билл Дьюи демонстрирует большой пластиковый барабан с пластиковой трубкой. Это часть насосной системы.«Этот шум, который вы слышите, — это перистальтический насос, который впрыскивает карбонат натрия в воду», — сказал он.
Исследователь из Орегона создал специальное устройство, которое дает показания воды в режиме реального времени. Это позволило производителям регулировать воду в инкубаторе». То, что делает этот перистальтический насос, автоматически меняет химический состав воды, — сказал Дьюи, — добавляя в воду ионы карбоната, чтобы наши маленькие устрицы снова могли формировать свои раковины».
Дьюи сказал, что инструмент улучшил условия выращивания молодых устриц.Но как только они оказываются на фермах и в приливных зонах, все зависит от природы.
Дьюи сказал, что именно здесь приходит помощь общественности в сокращении выбросов углерода. Но это более сложное решение.
Но это более сложное решение.
Инкубаторий моллюсков Брант-Пойнт | Нантакет, Массачусетс,
Реконструкция инкубатора моллюсков Брант-Пойнт
В течение последних двух лет сотрудники отдела природных ресурсов работали над полной реставрацией и реконструкцией инкубатора моллюсков Брант-Пойнт. Объект, первоначально перемещенный береговой охраной США в начале 1900-х годов, нуждается в значительной реставрации и обновлении.Проект, финансируемый городом Нантакет, Фондом охраны природы и частными пожертвованиями, вступит в первую фазу строительства в конце лета 2015 года. Чтобы следить за проектом и получить более подробную информацию, перейдите по ссылке проекта в меню на левый.
Инкубаторные проекты
Контролируемое нерест и выпуск личинок морских гребешков залива Нантакет.
Проведен еженедельный нерест местного поголовья для нереста в условиях инкубатория. Морские гребешки Нантакетского залива кондиционируются и запускаются для нереста с собранной слюной. После выдержки полученных личинок в течение 10-14 дней их выпускают в места, считающиеся пригодными для обитания. Среда обитания оценивается подводными исследованиями, проводимыми сотрудниками Департамента природных ресурсов. На сегодняшний день в этом полевом сезоне инкубаторий произвел 50 миллионов личинок.
После выдержки полученных личинок в течение 10-14 дней их выпускают в места, считающиеся пригодными для обитания. Среда обитания оценивается подводными исследованиями, проводимыми сотрудниками Департамента природных ресурсов. На сегодняшний день в этом полевом сезоне инкубаторий произвел 50 миллионов личинок.
Нерестовые садки
Садки были размещены в непосредственной близости друг от друга в районах в начале гавани. Цель этого проекта — увеличить плотность гребешка в определенных районах, чтобы помочь максимизировать успех размножения местных популяций.
Исследование пополнения личинок
В районах вокруг гавани были размещены мешки для спака, чтобы помочь измерить количество личинок морского гребешка залива Нантакет по всей гавани. Личинки могут плавать, прикрепленные к материалу внутри, и не могут выплыть после того, как вырастут. Мешки со слюной собираются раз в две недели в течение всего сезона.
Картирование морских водорослей
Летом 2015 года Департамент природных ресурсов совместно с Чарльзом Костелло из MassDEP составил карту всех ресурсов морских водорослей в гаванях Нантакет и Мадакет. Окончательную наземную проверку планируется провести в последние две недели сентября. Мы надеемся получить окончательный вариант карты и отчета к концу года. Этот отчет может быть взят и повторен NRD в будущем, чтобы постоянно обновлять карту и видеть, как морская трава меняется из года в год.
Окончательную наземную проверку планируется провести в последние две недели сентября. Мы надеемся получить окончательный вариант карты и отчета к концу года. Этот отчет может быть взят и повторен NRD в будущем, чтобы постоянно обновлять карту и видеть, как морская трава меняется из года в год.
Подтверждение концепции восстановления устричных рифов
Совместно с Программой переработки раковин Департамент природных ресурсов начал проверку концепции размножения устриц и установки их на переработанные раковины, собранные в местных ресторанах.Личинки устриц и мешочки с раковинами хранятся в инкубаторе для измерения скорости роста и оседания. Эта информация необходима, поскольку мы рассчитываем разрешить большой проект по восстановлению устричных рифов.
Улучшение качества моллюсков
В сотрудничестве с местным рыбаком-промысловиком Карлом Сьолундом NRD в партнерстве с Адмиралтейской ассоциацией Мадакет, Ассоциацией моллюсков Нантакета, Земельным советом Нантакета и Ассоциацией охраны природы Мадакета разместили 180 000 семян куахога в гаванях Нантакета и Мадакета для рекреационного использования. когда они достигают размера урожая.Это первый год проекта, рассчитанного на три года.
когда они достигают размера урожая.Это первый год проекта, рассчитанного на три года.
Транспортировка личинок рыбы и моллюсков через заливы
Описание
М. П. Вайнштейн, редактор
165 страниц; Опубликовано Американским обществом рыболовства; Дата публикации: 1988
Резюме
Симпозиум 3
Прогресс в понимании того, как личинки преодолевают расстояния, течения, приливы и градиенты солености, чтобы добраться до своих мест обитания и оставаться там, и как на успех пополнения влияет строительство вокруг устьевых бухт.
Содержание
Предисловие, М. П. Вайнштейн, Д. Г. Кларк и Э. Дж. Пуллен
1. Водообмен между шельфом и эстуарием между Мексиканским заливом и заливом Мобил, Алабама У. Дж. Уайзман мл., В. В. Шредер и С. П. Диннел
2. Транспортная модель водообмена между прибрежной бухтой и открытым океаном Д.-П. Ван
3. Наблюдения за режимами течения на входе, полученные на основе исследований численного и физического моделирования W. C. Seabergh
C. Seabergh
4. Оптимизация отбора проб для изучения приливного переноса в эстуариях B.Кьерфве и Т. Г. Волавер
5. Физические океанографические процессы, влияющие на перемещение личинок вокруг заливов Северной Каролины и через них Л. Дж. Пьетрафеса и Г. С. Яновиц
6. Роль поведенческих и физических факторов в пополнении личинок и молоди рыб в эстуарных питомниках Г. В. Бёлерт и Б. К. Манди
7. Физические процессы и механизмы прибрежных миграций неполовозрелых морских рыб Дж. М. Миллер
8. Взаимодействие ихтиопланктона и нектона океан-эстуарий в северной части Мексиканского залива R.Ф. Шоу, Б. Д. Роджерс, Дж. Х. Коуэн-младший и У. Х. Херке
9. Экспорт и реинвазия личинок как регуляторов эстуарных популяций десятиногих моллюсков Дж. Р. МакКонауга
10. Перенос личинок беспозвоночных между эстуариями и континентальным шельфом C.E. Epifanio
11. Моделирование физических и поведенческих механизмов, влияющих на пополнение спотового и атлантического горбыля в эстуарии мыса Страх Дж. П. Лоулер, М. П. Вайнштейн, Х. Я. Чен и Т. Л. Энглерт
П. Лоулер, М. П. Вайнштейн, Х. Я. Чен и Т. Л. Энглерт
12.Распространение икры и личинок рыб и модели циркуляции воды в заливе Наррагансетт, 1972–1973 гг. Д. У. Борн и Дж. Дж. Говони
13. Нулевая гипотеза, модели и статистические планы в изучении транспорта личинок Д. Р. Колби
Эпилог, М. П. Вайнштейн
Дрейфующие по морю: ученые из гранта Sea Grant изучают личинок устриц
Автор: ANN GREEN
На проливе Памлико сегодня безветренный день. Стоя на носу лодки, Эми Хаазе фокусирует свой бинокль на плавающем ярко-оранжевом и белом объекте.
«Дрифтер находится впереди по правому борту», — говорит Хаазе, аспирант Университета штата Северная Каролина.
Два члена Вспомогательной службы береговой охраны США готовятся найти дрифтера с мячом и плавучим тормозом или поплавком с нейтральной плавучестью, подвешенным под поверхностью воды. Один человек бросает сеть в воду, а затем вычерпывает дрейфующий буй, известный как дрифтер. Затем оба мужчины поднимают 40-килограммовый бродягу на палубу.
Затем оба мужчины поднимают 40-килограммовый бродягу на палубу.
Хаазе подходит, отсоединяет тормоз и складывает его, как зонтик.
«Отличная работа», — кричит Хаазе. «Теперь нам нужно найти еще четырех бродяг».
Когда капитан увеличивает скорость, Хаазе записывает широту и долготу дрифтера, который двигался по течению в течение последних 24 часов.
Каждые 10 минут буй записывает свое положение и связывается с центральным компьютером через спутниковый сотовый телефон. Записанные местоположения измеряют течения, а прикрепленный прибор измеряет температуру воды.
Эти данные являются частью исследования морского гранта Северной Каролины по переносу личинок устриц из заповедников, находящихся под управлением N.C. Отдел морского рыболовства (DMF). Ученые штата Северная Каролина Синтия Кудабак, Дэйв Эгглстон и Лиан Се проводят исследование вместе со своими аспирантами.
Вспомогательная служба береговой охраны США и другие добровольцы помогают найти буи. Работа является хорошим учебным опытом.
Работа является хорошим учебным опытом.
«Мне нравится находиться на воде, — говорит Чак Форман, вспомогательный рулевой береговой охраны США. «Это хорошая тренировка с использованием GPS. Это помогает нам в поисково-спасательных работах».
НАТУРАЛЬНАЯ УСТРИЦА
Исследование Sea Grant сосредоточено на американской устрице ( Crassostrea virginica ), которая часто сталкивается с суровыми и постоянно меняющимися условиями в реках и проливах Северной Каролины.
В последние годы активизировались работы по восстановлению и сохранению устричных рифов в Северной Каролине и других штатах. Большая часть этого толчка исходит от лучшего понимания того, как устрицы фильтруют загрязненную воду и как рифы обеспечивают среду обитания для многих других видов.
«Одна устрица может фильтровать почти шесть литров воды в час», — объясняет Эгглстон, директор Центра морских наук и технологий штата Северная Каролина в Морхед-Сити.
Эта функция помогает удалять загрязняющие вещества в проливах, бухтах и других прибрежных водах.
Чтобы восстановить популяцию устриц в эстуариях штата, DMF строит рифы на заповедных участках, в которых уже давно выращивают устриц. Натуральная устричная раковина или известняковый бетон являются основой рифов. В заказниках запрещены ловля устриц и использование дномешающих снастей, чтобы обеспечить развитие маточного стада устриц.
DMF управляет девятью устричными заповедниками в устьевых водах округов Дэр и Картерет, еще два находятся на стадии планирования.Прибрежная федерация Северной Каролины и The Nature Conservancy также восстанавливают рифы по всему штату.
«Мы рассматриваем возможность строительства нового убежища на северном берегу реки Нойз, — объясняет Крейг Харди, начальник отдела увеличения ресурсов DMF. «Теперь у нас есть святилище на южном берегу реки».
Кроме того, DMF курирует Программу выращивания устриц под причалами, которая позволяет жителям выращивать устриц под своими прибрежными доками или пирсами. Квалифицированные владельцы разрешений могут прикрепить до 90 квадратных футов контейнеров для выращивания устриц к принадлежащему им причалу или пирсу.
ИССЛЕДОВАНИЕ ЛИЧИНОК
Каждая устрица ежегодно откладывает миллионы яиц, которые переносятся течениями и приливами в окружающие районы. За счет развития и защиты маточного стада увеличивается доступность местных устриц в прилегающих районах.
Устрицы нерестятся с мая по сентябрь, пик приходится на середину-конец июня. Затем личинки расселяются по рифам.
Прошлым летом ученые, финансируемые за счет гранта Sea Grant, начали отслеживать движение воды в ключевых заповедниках маточного стада в проливах Кроатан и Памлико.
По предварительным данным, ученые определили, что эстуарные течения примерно следуют направлению ветра, что, по словам Хаазе, похоже на то же самое для личинок устриц.
По словам Кудабака, ветры в эстуарной системе Альбемарль-Памлико обычно дуют на юго-запад летом и падают, когда личинки перемещаются в толще воды.
«Мы ожидаем увидеть связь между заповедником «Крабовая нора» к югу от острова Роанок и пятью другими заповедниками в основной части пролива Памлико», — добавляет она. «Однако святилище Вест-Бэй на реке Нойз может быть более изолированным от остального звука».
«Однако святилище Вест-Бэй на реке Нойз может быть более изолированным от остального звука».
Эстуарная система включает около 3000 квадратных миль солоноватой воды с дренажной системой площадью 30000 квадратных миль.
«Хотя устрицы имеют как коммерческое, так и экономическое значение в этом регионе, и несколько исследований устриц было проведено в других частях Северной Каролины, это первый случай изучения переноса личинок устриц в проливе Памлико», — говорит Кудабак.
Приливы в проливе Памлико относительно слабые, добавляет она.
Система Северной Каролины сильно отличается от Чесапикского залива, где было проведено больше исследований переноса личинок устриц. Исследователи обнаружили, что усилия по восстановлению Чесапикского залива увенчались успехом в районах, где циркуляция удерживает личинок вблизи рифов, а низкая соленость снижает заболеваемость некоторыми заболеваниями.
Cudaback сообщает, что исследователи штата Северная Каролина будут сравнивать данные с исследованиями Чесапикского залива, а также с результатами предыдущего проекта Sea Grant по переносу личинок синего краба в проливе Памлико.
Чтобы изучить, как глубина водной толщи и соленость влияют на перенос личинок устриц, ученые устанавливают акустические доплеровские профилировщики течений (ADCP) на твердом дне. Профилографы регистрируют течения на всех глубинах каждые пять минут и работают непрерывно до трех месяцев.
Чтобы получить полное представление о циркуляции, течения, измеренные ADCP, будут сравниваться с течениями, измеренными дрейфующим буем, добавляет Cudaback.
«Это исследование необычно тем, что оно объединяет биологов и геофизиков для решения проблемы с более чем одной точки зрения», — говорит заместитель директора North Carolina Sea Grant Стив Ребах.
Этим летом исследователи Sea Grant снова разместят дрейфующие буи в проливе Памлико и привлекут добровольцев из вспомогательной службы береговой охраны.
«Понимание схемы течения в эстуарии предоставит нашим менеджерам информацию для строительства новых убежищ и доставки личинок в районы, которые нуждаются в улучшении», — говорит Харди. «Если мы знаем, где будут находиться личинки из отдельных убежищ, когда они оседают, мы можем размещать материалы в этих областях, обеспечивая субстрат для увеличения их поселения.
«Если мы знаем, где будут находиться личинки из отдельных убежищ, когда они оседают, мы можем размещать материалы в этих областях, обеспечивая субстрат для увеличения их поселения.
Кроме того, данные о характере течения в эстуарии помогут управляющим DMF решить, где разместить личинок при строительстве новых убежищ.
«Исследование Sea Grant предоставляет жизненно важную информацию, которая связывает два важных проекта по восстановлению устриц, обеспечивая максимальный успех обоих», — добавляет Харди.
ФРГ ИССЛЕДОВАНИЕ
Получив грант Северной Каролины на рыболовные ресурсы (FRG), Эгглстон работает с коммерческим рыбаком из Внешних отмелей Юджином Баллансом, который обновил историческую карту устричных зарослей в проливе Памлико.
В настоящее время ученые составляют пространственную карту и топографию шести устричных заповедников в проливе Памлико. Программа FRG, финансируемая Генеральной Ассамблеей Северной Каролины, находится в ведении Морского гранта Северной Каролины.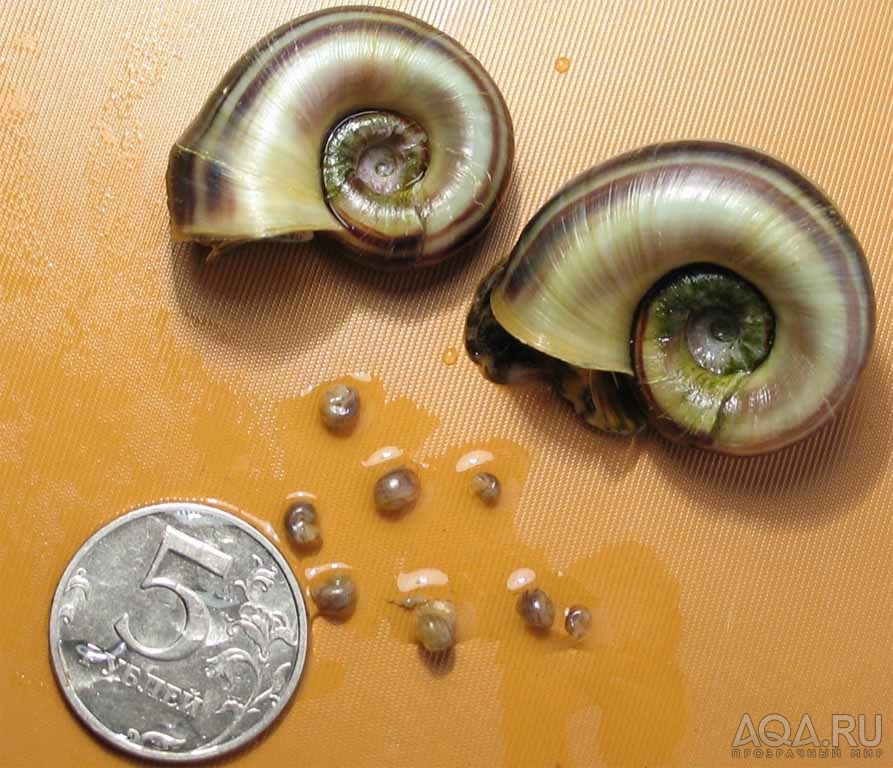
Чтобы измерить разнообразие поселений личинок устриц, исследователи используют веревки из ракушек, подвешенные к докам по всему проливу Памлико, которые еженедельно проверяют старшеклассники.
«Изменения в поселении личинок устриц в проливе Памлико в течение летнего и осеннего личиночного периода будут сравниваться с прогнозами математических моделей, чтобы увидеть, насколько надежны модели в отношении того, где оседают личинки», — говорит Эгглстон.— А дрифтеры дадут нам информацию, откуда берутся личинки.
В 2006 году 13 старшеклассников из Мантео, Ориентала, Энглхарта, Байборо, Хаттераса, Окракока и Сидар-Айленда участвовали в ряде исследований, в том числе в сборе раковин, развертывании новых личинок устриц и измерении температуры, солености и вода для анализа на хлорофилл.
«Это отличная образовательная возможность для старшеклассников познакомиться с наукой, — говорит аспирант штата Северная Каролина Брэндон Пакетт.
Предварительные результаты показывают, что заселение в юго-западной части залива Памлико низкое, говорит Эгглстон.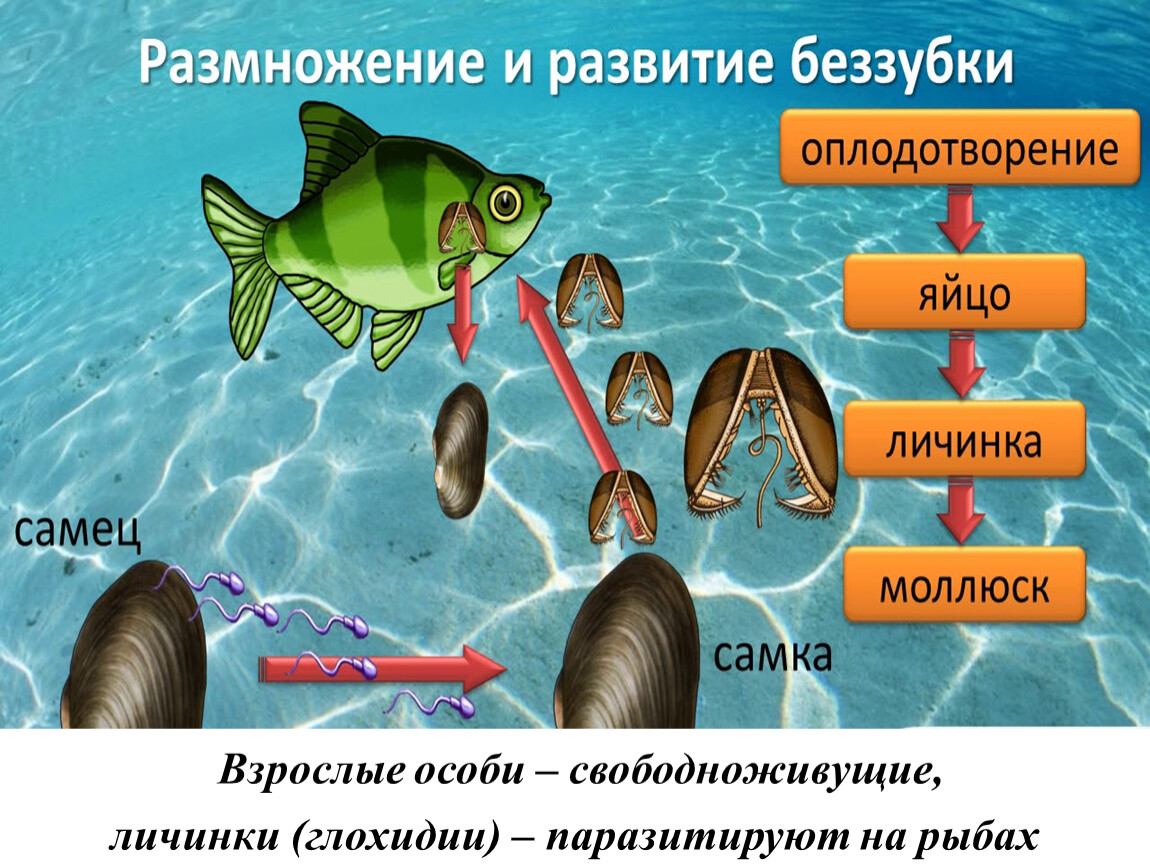 «Это та же картина, которая была обнаружена в проливе Памлико, когда мы измеряли поселение синих крабов за 10-летний период».
«Это та же картина, которая была обнаружена в проливе Памлико, когда мы измеряли поселение синих крабов за 10-летний период».
Команда также обнаружила, что пик расселения устриц приходится на начало лета вблизи Окракока и Хаттераса. Исследователи проанализируют относительную величину расселения устриц в зависимости от направления и скорости ветра, добавляет он.
Другая часть исследования посвящена судьбе личинок устриц после поселения.
Эгглстон и Балланс наблюдают за устрицами с течением времени, чтобы определить, в каких заповедниках устрицы растут быстрее всего и живут дольше всех.
DMF будет использовать данные для ранжирования лучших мест для приютов маточного стада.
«По мере того, как последующие поколения устриц оседают и растут, рифы образуют множество укромных уголков и
трещин, которые обеспечивают богатую микросреду обитания для многих различных видов животных», — говорит Сара Мирабилио, специалист по рыболовству из Северной Каролины Sea Grant.
Исследование, проведенное в 1961 году Гарри Уэллсом из Университета Дьюка, показало, что более 303 различных видов использовали устричные рифы в качестве среды обитания в Северной Каролине, добавляет она.
ЦЕННЫЕ ВИДЫ
С незапамятных времен устрицы были важным источником пищи на побережье Северной Каролины. Когда европейцы впервые прибыли, они были поражены количеством найденных устриц.
Пик индустрии устриц в Каролине пришелся на 1880-е годы, когда компании из Балтимора построили большие консервные заводы в прибрежных городах.В то же время коммерческие рыболовы начали использовать устричные земснаряды.
Несмотря на более поздние попытки ограничить дноуглубительные работы, большая часть ущерба устричным зарослям уже была нанесена к 1910 году, когда запасы устриц начали сокращаться.
«Сейчас устрицы высаживаются примерно на 1 процент столетней давности, — говорит Кудабак. «Этот коллапс имел разрушительные экономические и экологические последствия».
В 2005 году в штате было собрано более 377 000 фунтов мяса устриц стоимостью более 1 доллара США.6 миллионов по сравнению с 545 000 фунтов стерлингов в 1985 году на сумму более 1 миллиона долларов, по данным DMF.
В Северной Каролине устрицы водятся от крайней юго-восточной оконечности реки Альбемарл у северной оконечности острова Роанок на юг через проливы Кроатан, Роанок и Памлико до устьев южных районов штата недалеко от границы с Южной Каролиной.
Однако, по словам Харди, в последние три-четыре года популяция устриц имеет тенденцию к росту по сравнению с исторически низким уровнем добычи в середине 1990-х годов.
Hardy приписывает это увеличение упадку Dermo или Perkinus marinus , простейшего паразита, который легко передается от устрицы к устрице. «Произошло возвращение к более нормальному климату и уровню солености», — добавляет он. «В середине 90-х мягкие зимы и повышенная соленость вызвали рост заболеваемости дермо».
Устриц собирают с октября по март щипцами, граблями или вручную в приливных зонах с мелководьем вдоль побережья Северной Каролины. Коммерческие рыбаки также используют земснаряды в некоторых частях пролива Памлико.
Коммерческие рыбаки также используют земснаряды в некоторых частях пролива Памлико.
В Северной Каролине устрицы водятся от крайней юго-восточной оконечности пролива Альбемарл у северной оконечности острова Роанок на юг через проливы Кроатан, Роанок и Памлико до устьев рек южной части штата недалеко от границы с Южной Каролиной.
ИССЛЕДОВАНИЯ ПО ВОССТАНОВЛЕНИЮ УСТРИЦ
В течение нескольких лет Мартин Поузи и Трой Альфин, получившие морской грант Северной Каролины, проводили передовые исследования в области дизайна рифов, чтобы разработать передовые методы восстановления устриц как для рыбной промышленности, так и для функций экосистемы, включая качество воды и среду обитания рыб.
Ученые из Университета Северной Каролины в Уилмингтоне обнаружили, что рифы высокой сложности с многочисленными расщелинами больше использовались в качестве среды обитания для разнообразных рыб, креветок и крабов, включая синих крабов, чем рифы с меньшей структурой. Эти рифы также являются средой обитания для многих мелких беспозвоночных, которые являются источником пищи для важных рыбных видов.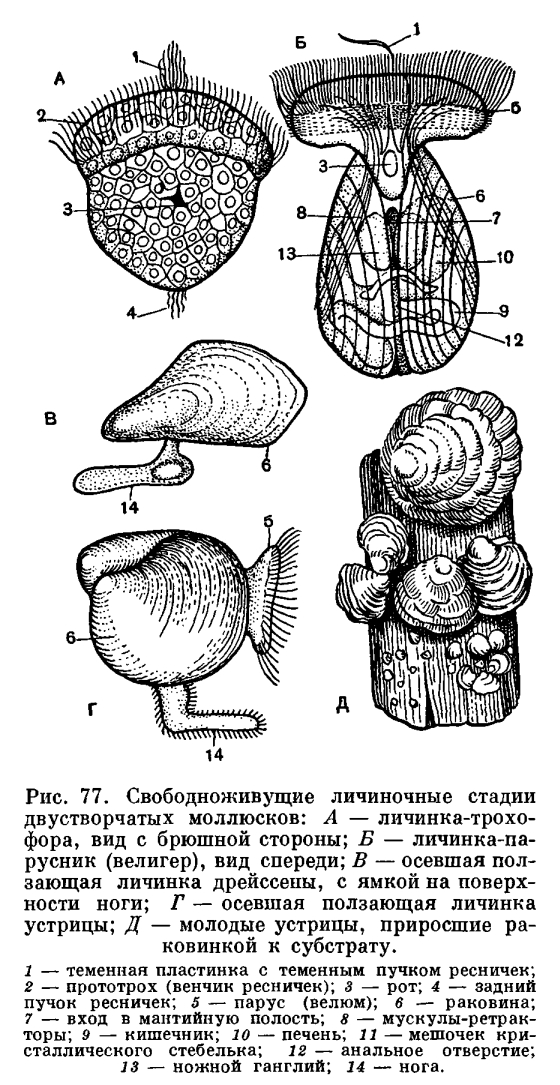
Кроме того, край рифа влияет на местные течения, органический материал и количество питательных веществ внутри рифа и на прилегающих отмелях.
Posey и Alphin использовали результаты исследования, чтобы помочь Прибрежной федерации Северной Каролины в выборе участка, проектировании рифов и определении размеров устричных рифов в ряде прибрежных проектов.
DMF также использовал данные для выращивания твердых материалов, таких как ракушки и известняк, на искусственных святилищах.
В новом исследовании Sea Grant Поузи и Альфин, а также аспирант UNCW Тед Уилгис анализируют успех пяти проектов по восстановлению устричных рифов на центральном и юго-восточном побережье.
«Наша цель — протестировать предлагаемые стандартизированные методы отбора проб, оценив использование проектов восстановления в качестве среды обитания и для улучшения качества воды», — говорит Поузи.
В работе также исследуется, как различия в толщине раковины влияют на создание рифов, влияние ландшафта на успех восстановления, а также влияние засева на недавно созданные раковинные рифы живыми устрицами, используемыми для ускорения развития.
Исследование включает оценку плотности и размера устриц, а также организмов, обитающих в пределах рифовой системы — рыб, креветок и крабов, которые используют риф как среду обитания и место для кормления.Проект, который вырос из усилий Национального морского гранта, включает в себя совместные усилия исследователей Северной Каролины и Южной Каролины.
«Предварительные результаты показывают, что не все усилия по восстановлению одинаково успешны, — говорит Поузи.
Некоторые искусственные рифы, по-видимому, сформировали сообщества, похожие на естественные районы в тех же окрестностях, всего за три года после создания, добавляет он.
«Однако на некоторых рифах все еще нет хорошо развитого рифового сообщества и низкая плотность устриц по сравнению с прилегающими районами», — говорит Поузи.«Ключом к восстановлению является понимание того, почему одни усилия более успешны, чем другие».
Эта информация будет полезна менеджерам по ресурсам Северной Каролины, а также всем, кто заинтересован в оценке эффективности проектов по восстановлению устриц, объясняет Поузи.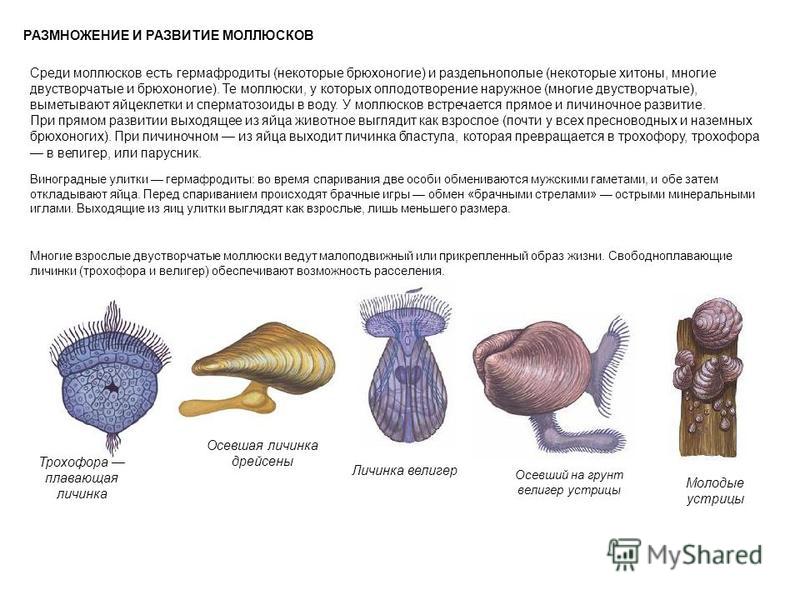
«Восстановление устриц является приоритетом для Северной Каролины, а также многих прибрежных штатов от Средней Атлантики до Мексиканского залива».
Чтобы узнать больше о приютах DMF, посетите Интернет: www.ncfisheries.net и нажмите «Моллюски и ракообразные», а затем «Заповедники моллюсков».
Эта статья была опубликована в весеннем выпуске журнала Coastwatch за 2007 год.
Для получения контактной информации и запросов на перепечатку посетите веб-сайт ncseagrant.ncsu.edu/coastwatch/contact/.
Влияние подкисления на обызвествление и рост личинок устриц в эстуариях
Аннотация
Фон
Деятельность человека увеличила концентрацию углекислого газа в атмосфере на 36% за последние 200 лет.Одна треть всего антропогенного CO 2 была поглощена океанами, снизив pH примерно на 0,1 единицы и значительно изменив химический состав их карбонатов. Широко распространено беспокойство по поводу того, что эти изменения серьезно изменяют морскую среду обитания, но мало или вообще не уделяется внимания биоте эстуариев и прибрежных районов, экосистемам, которые менее защищены от pH из-за естественного снижения щелочности.
Методология/основные выводы
Для устранения изменений, вызванных CO 2 , в эстуарной кальцификации, велигерные личинки двух видов устриц, восточной устрицы (Crassostrea virginica), и устрицы Сумино ( Crassostrea ariakensis) , были выращены в устьевой воде под четырьмя p CO 2 режимы, 280, 380, 560 и 800 мкатм, для имитации атмосферных условий в доиндустриальную эпоху, нынешние и прогнозируемые будущие концентрации через 50 и 100 лет соответственно.Манипуляции с CO 2 проводились с использованием автоматизированной системы управления с отрицательной обратной связью, которая позволяла непрерывно и точно контролировать p CO 2 в экспериментальных аквариумах. Рост личинок измеряли с помощью анализа изображений, а кальцификацию измеряли с помощью химического анализа кальция в их панцирях. C. virginica испытал уменьшение площади скорлупы на 16% и снижение содержания кальция на 42% при сравнении доиндустриальной обработки и обработки конца 21 st века p CO 2 . C. ariakensis не показал изменений ни в росте, ни в кальцификации. Оба вида продемонстрировали чистую кальцификацию и рост, даже когда арагонит был недонасыщен, что противоречит предыдущим ожиданиям для личинок беспозвоночных, которые производят арагонитовые раковины.
C. ariakensis не показал изменений ни в росте, ни в кальцификации. Оба вида продемонстрировали чистую кальцификацию и рост, даже когда арагонит был недонасыщен, что противоречит предыдущим ожиданиям для личинок беспозвоночных, которые производят арагонитовые раковины.
Выводы и значение
Наши результаты показывают, что эстуарные и прибрежные экосистемы умеренного пояса уязвимы к ожидаемым изменениям в химическом составе воды из-за повышенного содержания CO 2 в атмосфере и что биологические реакции на подкисление, особенно кальцификацию биоты, будут видоспецифичными и, следовательно, гораздо более изменчивыми и сложными. чем сообщалось ранее.
Образец цитирования: Миллер А.В., Рейнольдс А.С., Собрино С., Ридель Г.Ф. (2009) Моллюски сталкиваются с неопределенным будущим в условиях высокого содержания CO 2 Мир: Влияние подкисления на кальцификацию и рост личинок устриц в эстуариях. ПЛОС ОДИН 4(5):
е5661. https://doi.org/10.1371/journal.pone.0005661
https://doi.org/10.1371/journal.pone.0005661
Редактор: Зои Финкель, Университет Маунт Элисон, Канада
Поступила в редакцию: 4 февраля 2009 г.; Принято: 27 апреля 2009 г .; Опубликовано: 27 мая 2009 г.
Авторское право: © 2009 Miller et al.Это статья с открытым доступом, распространяемая в соответствии с лицензией Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания оригинального автора и источника.
Финансирование: Финансирование этого исследования было предоставлено Фондом Сьюарда Джонсона через Смитсоновскую сеть морских наук, а также федеральными ассигнованиями, выделенными Смитсоновскому институту.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
За последние 200 лет сжигание ископаемого топлива, вырубка лесов и освоение земель увеличили концентрацию углекислого газа в атмосфере на 36%, и ожидается, что уровень выбросов CO 2 увеличится в течение следующего столетия [1] – [3]. Примерно треть всего антропогенного CO 2 поглощена океанами Земли [4]–[5]. Хотя океан частично поглотил недавно высвободившийся антропогенный атмосферный CO 2 , это произошло за счет значительного снижения pH (закисления) и изменения химического состава карбонатов в поверхностных водах океана.С ок. 1760 г. рН поверхности океана (верхние 200 м водной толщи) уменьшился примерно на 0,1 единицы, и в ближайшие 100 лет ожидается дальнейшее снижение на 0,1–0,5 единицы [6]–[8]. Существует широко распространенное опасение, что эти изменения приведут к необратимым изменениям экологического режима в морской среде обитания, например, массовому сокращению среды обитания коралловых рифов и связанного с ними биоразнообразия, а также снижению доступности ионов карбоната для кальцификации биоты [9]–[11].
Примерно треть всего антропогенного CO 2 поглощена океанами Земли [4]–[5]. Хотя океан частично поглотил недавно высвободившийся антропогенный атмосферный CO 2 , это произошло за счет значительного снижения pH (закисления) и изменения химического состава карбонатов в поверхностных водах океана.С ок. 1760 г. рН поверхности океана (верхние 200 м водной толщи) уменьшился примерно на 0,1 единицы, и в ближайшие 100 лет ожидается дальнейшее снижение на 0,1–0,5 единицы [6]–[8]. Существует широко распространенное опасение, что эти изменения приведут к необратимым изменениям экологического режима в морской среде обитания, например, массовому сокращению среды обитания коралловых рифов и связанного с ними биоразнообразия, а также снижению доступности ионов карбоната для кальцификации биоты [9]–[11].
На сегодняшний день большинство исследований сосредоточено на подкислении, вызванном CO 2 , в полностью морских водах, в то время как мало внимания уделялось эстуариям с более низкой соленостью и прибрежным экосистемам умеренного пояса. Но прибрежные и эстуарные биомы являются одними из наиболее продуктивных с биологической точки зрения и поддерживают некоторые из наиболее обширных и измеримых экосистемных услуг (например, коммерческое и любительское рыболовство, места нагула рыбы и беспозвоночных, очистка воды, защита от наводнений и штормовых нагонов, отдых человека).Поскольку они более мелкие, менее соленые и имеют более низкую щелочность [12], эстуарии и прибрежные морские среды обитания более восприимчивы к изменениям рН, чем открытый океан. Эстуарии также подвержены значительному обогащению CO 2 , образующемуся в результате дыхания как природного, так и антропогенного углерода. В то время как многие эстуарии уже имеют высокое и изменчивое pCO 2 [13], обогащение атмосферы CO 2 сдвинет базовую линию в сторону еще более высоких значений. По этим причинам эти экосистемы, вероятно, испытают более острое воздействие повышенного содержания CO 2 в ближайшие десятилетия.
Но прибрежные и эстуарные биомы являются одними из наиболее продуктивных с биологической точки зрения и поддерживают некоторые из наиболее обширных и измеримых экосистемных услуг (например, коммерческое и любительское рыболовство, места нагула рыбы и беспозвоночных, очистка воды, защита от наводнений и штормовых нагонов, отдых человека).Поскольку они более мелкие, менее соленые и имеют более низкую щелочность [12], эстуарии и прибрежные морские среды обитания более восприимчивы к изменениям рН, чем открытый океан. Эстуарии также подвержены значительному обогащению CO 2 , образующемуся в результате дыхания как природного, так и антропогенного углерода. В то время как многие эстуарии уже имеют высокое и изменчивое pCO 2 [13], обогащение атмосферы CO 2 сдвинет базовую линию в сторону еще более высоких значений. По этим причинам эти экосистемы, вероятно, испытают более острое воздействие повышенного содержания CO 2 в ближайшие десятилетия.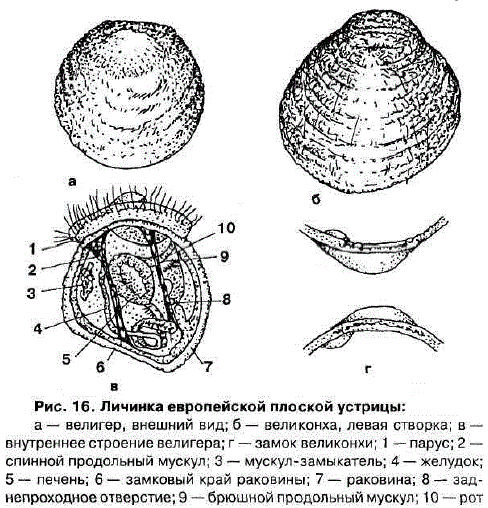
Прямым следствием подкисления, вызванного CO 2 , является снижение концентрации ионов карбоната [] в толще воды. Особую озабоченность в морских и эстуарных системах вызывает влияние повышения CO 2 на состояние насыщения воды по отношению к карбонату кальция (Ω), где K′ sp представляет собой произведение кажущейся растворимости [14]–[15] Таким образом, :Ω является косвенным показателем легкости, с которой организмы могут производить карбонат кальция (CaCO 3 ). Состояния насыщения воды по отношению к арагониту и кальциту, двум наиболее часто биоминерализуемым формам CaCO 3 , уменьшаются при повышении парциального давления CO 2 ( p CO 2 ) [15].
В эстуариях Ω естественным образом уменьшается с уменьшением солености из-за градиентов pH, Ca 2+ и вызванного разбавлением морской воды речной водой (см. Cai and Wang [13]). Недавнее исследование Salisbury et al. [16] указывает, что сезонные сбросы кислых речных вод еще больше усугубят закисление в эстуариях, предполагая возможность негативного воздействия на промысел моллюсков.
В Чесапикском заливе Вонг [12] показал, что вода реки Джеймс и океана консервативно смешиваются и что соленость и общая щелочность (ТА) линейно связаны между собой примерно от 5 psu до 32 psu.При ∼18 psu ТА в заливе составляет ∼1250 мкмоль/кг-SW. Если предположить консервативное перемешивание воды и равновесие с текущим атмосферным CO 2 (380 мкатм), то точка компенсации арагонита (т. е. Ω arag = 1,0) при летних температурах должна лежать вблизи изоплеты 18 psu. По мере увеличения атмосферного CO 2 , при условии, что Ca 2+ и щелочность останутся на нынешнем уровне, точка компенсации арагонита в эстуариях сместится в сторону моря в сторону более высокой солености. Мы утверждаем, что точка компенсации арагонита уже значительно сместилась в сторону устья залива с начала индустриальной эры (рис.1). Точка компенсации для кальцита, который примерно в 1,5 раза менее растворим, чем арагонит [14], возникает при меньшей солености и аналогичным образом будет смещаться в сторону моря.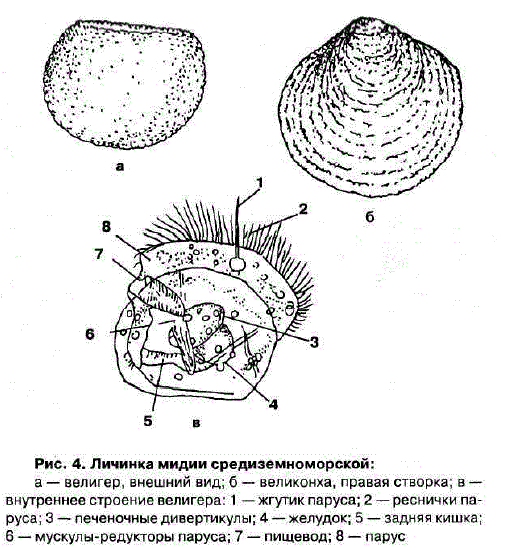 В действительности эстуарии часто не находятся в равновесии с атмосферой, однако изменение атмосферного p CO 2 сместит исходное равновесие, к которому стремятся эстуарии, и приведет к более широкому распространению p CO 2 .
В действительности эстуарии часто не находятся в равновесии с атмосферой, однако изменение атмосферного p CO 2 сместит исходное равновесие, к которому стремятся эстуарии, и приведет к более широкому распространению p CO 2 .
Рис. 1. Прогноз среднего летнего положения компенсационных точек арагонита (Ω arag = 1.0) для Чесапикского залива.
Теоретические положения компенсации арагонита были нанесены при прошлом, настоящем и прогнозируемом атмосферном CO 2 . Насыщенность арагонита рассчитывали по программе CO 2 SYS.XLS по щелочности и атмосферному p CO 2 при 25°C и давлении 0 мбар, предполагая консервативное смешение щелочностей Атлантики и реки Саскуэханна и средней летней солености ( 1985–2006 гг. – данные программы Чесапикского залива [47]). Арагонит перенасыщен мористее компенсационных линий.
https://doi.org/10.1371/journal.pone.0005661. g001
g001
Мы ожидаем, что личинки двустворчатых моллюсков будут более уязвимы, чем взрослые особи, к повышенному содержанию CO 2 , поскольку личинки биоминерализуют арагонит, более растворимую форму CaCO 3 , а не кальцит, преобладающий материал, используемый в раковинах взрослых особей. Вайс и др. [17] также указывают, что во время биоминерализации некоторые двустворчатые личинки (например, Crassostrea gigas и Mercenaria mercenaria ) генерируют еще более растворимый аморфный CaCO 3 в качестве эфемерного предшественника кристаллического арагонита, и поскольку личинки, как правило, менее устойчивы. чем взрослые к различным стрессам.Если личинки действительно более восприимчивы к подкислению, это может привести к снижению их производительности или даже выходу из строя, что в конечном итоге приведет к негативным последствиям для устриц и других популяций моллюсков. Курихара и др. при очень высоких значениях p CO 2 (2268 мкатм) эмбриональное развитие и формирование раковины у Crassostrea gigas подавлялось в течение первых 48 ч после оплодотворения [18]. Каким образом меняющийся химический состав карбонатов изменит экологическую структуру и функции бентических сообществ, остается критическим пробелом в наших знаниях [1]–[2].
Каким образом меняющийся химический состав карбонатов изменит экологическую структуру и функции бентических сообществ, остается критическим пробелом в наших знаниях [1]–[2].
Мы провели эксперименты на личинках велигеров двух близкородственных эвригалинных видов устриц, восточной устрицы ( Crassostrea virginica [Gmelin 1791]) и устрицы суминоэ ( Crassostrea ariakensis [Fujita 1913]), в условиях постоянного контроля рН. /pCO 2 на срок до 28 дней. C. virginica произрастает в Западной Атлантике, но недавние высадки составляют всего 1% от их исторического максимума в Чесапикском заливе, в значительной степени из-за перелова и болезней [19]–[20].Интродукция C. ariakensis в Чесапикский залив рассматривалась в надежде на восстановление промысла устриц и важных экосистемных функций, таких как фильтрация воды и снижение содержания питательных веществ и отложений в толще воды [21]. Мы предполагаем, что (а) личинки устриц будут демонстрировать снижение роста и кальцификацию при повышенном уровне p CO 2 и (б) развитие личинок будет неблагоприятно и ощутимо затронуто, когда Ω arag <1,0. В более широком смысле ожидается, что подкисление негативно повлияет на промысел устриц, других моллюсков и кальцифицирующую придонную фауну.
В более широком смысле ожидается, что подкисление негативно повлияет на промысел устриц, других моллюсков и кальцифицирующую придонную фауну.
Материалы и методы
Все протестированные личинки устриц были получены из Лаборатории Восточного побережья Института морских наук Вирджинии, Вачапрег, Вирджиния, где C. virginica и C. ariakensis были выведены в карантинных помещениях. Эксперимент Личинки Ca 1 были получены из запасов C. ariakensis , происходящих из Японского моря Ариаке, но случайно перенесенных на западное побережье США с партиями C.gigas в 1970-х годах [22]. Cv 1 были получены из запасов Wachapreague seaside C. virginica . В таблице 1 обобщены все экспериментальные параметры для экспериментов Cv 1 и Ca 1. Через 72 часа после оплодотворения личинки стадии D были перенесены из Вашапрега в карантинную лабораторию Смитсоновского центра экологических исследований в Эджуотере, штат Мэриленд, и помещены в культура. Чтобы проверить влияние повышенного p CO 2 , пониженного pH и состояния насыщения арагонита на рост личинок и кальцификацию, мы культивировали личинок устриц в контролируемых условиях, которые имитировали типичную летнюю эстуарную среду в верхних мезогалинных пределах Чесапикского залива: соленость = 18 psu, температура = 25°C, цикл день/ночь = 14:10 часов.Экспериментальные обработки были заблокированы в двух инкубаторах (камеры с контролируемой средой Percival I-36 с флуоресцентными лампами Philips серии 700 мощностью 32 Вт) и отдельных аквариумах, произвольно расположенных в каждом инкубаторе. Инкубатор A содержал 280 мкатм и 380 мкатм p CO 2 обработки, а инкубатор B содержал 560 мкатм и 800 мкатм для всех экспериментов. Перед экспериментами излучение ФАР было измерено внутри каждого инкубатора, и было показано, что оно одинаково (A = 167,7±5,3 мкЭ м −2 с −1 по сравнению с B = 169.2 ± 7,3 мкЭ м 90 416 -2 90 417 с 90 416 -1 90 417 (среднее значение ± стандартная ошибка среднего).
Чтобы проверить влияние повышенного p CO 2 , пониженного pH и состояния насыщения арагонита на рост личинок и кальцификацию, мы культивировали личинок устриц в контролируемых условиях, которые имитировали типичную летнюю эстуарную среду в верхних мезогалинных пределах Чесапикского залива: соленость = 18 psu, температура = 25°C, цикл день/ночь = 14:10 часов.Экспериментальные обработки были заблокированы в двух инкубаторах (камеры с контролируемой средой Percival I-36 с флуоресцентными лампами Philips серии 700 мощностью 32 Вт) и отдельных аквариумах, произвольно расположенных в каждом инкубаторе. Инкубатор A содержал 280 мкатм и 380 мкатм p CO 2 обработки, а инкубатор B содержал 560 мкатм и 800 мкатм для всех экспериментов. Перед экспериментами излучение ФАР было измерено внутри каждого инкубатора, и было показано, что оно одинаково (A = 167,7±5,3 мкЭ м −2 с −1 по сравнению с B = 169.2 ± 7,3 мкЭ м 90 416 -2 90 417 с 90 416 -1 90 417 (среднее значение ± стандартная ошибка среднего).
Природная морская вода была собрана из Sinepuxent Bay, MD (~28 psu) и Delaware Bay, DE (~24 psu), профильтрована через фильтр 0,2 мкм и разбавлена деионизированной водой до 18 psu. Двухточечное титрование с открытой ячейкой [23] определяло ТА воды непосредственно перед началом каждого эксперимента. Общая щелочность и P CO 2 Цели для допромышленного ( P CO 2 = 280 мкВТМ), современный день ( P CO 2 = 380 мкВТМ), 2050 год ( P CO 2 = 560 мкатм) и 2100 года ( p CO 2 = 800 мкатм) настройки, рассчитанные по сценарию IS92a «обычный бизнес» [24], затем были введены в компьютерную программу CO 2 .XLS [25]. Программа была специально параметризована двумя константами диссоциации K 1 и K 2 для угольной кислоты в устьевых водах Cai и Wang [13] для определения соответствующих экспериментальных уровней pH.
Личинки выращивали в поликарбонатных аквариумах объемом 4 л при четырех обработках p CO 2 , и каждая обработка включала три повторных аквариума с независимым контролем p CO 2 /pH (n = 12). Пробы фильтрованной воды (0.2 мкм) собирали из каждого аквариума каждые несколько дней в ходе экспериментов по анализу общего растворенного неорганического углерода (TDIC). (Для поддержания качества воды воду меняли во всех аквариумах каждые два дня. Пробы TDIC брали через 24–48 часов после замены воды.) Пробы воды закрывали крышками без свободного пространства и немедленно анализировали или хранили при 48 часов. TDIC измеряли с использованием анализатора общего органического углерода, оснащенного реакционной камерой для реагентов на основе фосфорной кислоты и неорганического углерода и недисперсионным инфракрасным детектором (Schimadzu TOC-V с комплектом реактора IC).Затем TDIC разделяли на pCO 2 , бикарбонат и карбонат, а TA рассчитывали с помощью CO2SYS.XLS [25] (таблица 1). пятна.
Пробы фильтрованной воды (0.2 мкм) собирали из каждого аквариума каждые несколько дней в ходе экспериментов по анализу общего растворенного неорганического углерода (TDIC). (Для поддержания качества воды воду меняли во всех аквариумах каждые два дня. Пробы TDIC брали через 24–48 часов после замены воды.) Пробы воды закрывали крышками без свободного пространства и немедленно анализировали или хранили при 48 часов. TDIC измеряли с использованием анализатора общего органического углерода, оснащенного реакционной камерой для реагентов на основе фосфорной кислоты и неорганического углерода и недисперсионным инфракрасным детектором (Schimadzu TOC-V с комплектом реактора IC).Затем TDIC разделяли на pCO 2 , бикарбонат и карбонат, а TA рассчитывали с помощью CO2SYS.XLS [25] (таблица 1). пятна.
Эксперименты Cv 1 ( Crassostrea virginica ) и Ca 1 ( C. ariakensis ) инокулировали 15 000 четырехдневных личинок D-стадии соответственно и выращивали до 28 дней. Аквариумы получали контролируемое количество микроводорослей Isochrysis galbana (Prymnesiophyceae) в качестве корма. Микроводоросли выращивали в полунепрерывных культурах на среде f/2-Guillard. Ежедневно пипеткой вносили равные количества микроводорослей из культур известной плотности в каждый аквариум, при этом суточные дозы увеличивались по мере роста личинок. Целевая плотность посева в каждом аквариуме увеличивалась с 1,5·10 4 кл·мл -1 в начале эксперимента до 1,0·10 5 кл·мл -1 в заключительные дни, когда личинки были крупнее. и прием большего количества пищи.
Аквариумы получали контролируемое количество микроводорослей Isochrysis galbana (Prymnesiophyceae) в качестве корма. Микроводоросли выращивали в полунепрерывных культурах на среде f/2-Guillard. Ежедневно пипеткой вносили равные количества микроводорослей из культур известной плотности в каждый аквариум, при этом суточные дозы увеличивались по мере роста личинок. Целевая плотность посева в каждом аквариуме увеличивалась с 1,5·10 4 кл·мл -1 в начале эксперимента до 1,0·10 5 кл·мл -1 в заключительные дни, когда личинки были крупнее. и прием большего количества пищи.
p CO 2 в каждом аквариуме периодически барботировали воздухом, обогащенным CO 2 (коммерчески доступный, сертифицированный 1,0% CO 2 ) и непрерывной аэрацией воздухом, очищенным от CO 2 . Для очистки CO 2 использовались натронно-известковые фильтры. Непрерывная аэрация очищенным от CO 2 воздухом насыщала воду кислородом и перемешивала воду, одновременно удаляя CO 2 из воды и повышая pH. Каждый аквариум постоянно контролировали с помощью pH-зонда (шкала NBS) и системы управления.Как только целевое значение pH было превышено, автоматический контроллер открывал соленоид и подавал воздушную смесь 1% CO 2 в аквариум, тем самым снижая pH. Поток CO 2 автоматически прекращался при достижении заданного значения pH. Основываясь на ежечасных измерениях pH, было показано, что система контроля с отрицательной обратной связью поддерживает pH (и p CO 2 ) на почти постоянном уровне до 4 недель. Используемые четыре заданных значения pH соответствовали желаемой цели обработки воды с известной общей щелочностью.Зонды были откалиброваны перед экспериментом с использованием прослеживаемых буферов NIST (pH = 4,00 и 7,00), повторно откалиброваны по крайней мере один раз во время эксперимента и еще раз по завершении эксперимента для обеспечения правильной работы.
Каждый аквариум постоянно контролировали с помощью pH-зонда (шкала NBS) и системы управления.Как только целевое значение pH было превышено, автоматический контроллер открывал соленоид и подавал воздушную смесь 1% CO 2 в аквариум, тем самым снижая pH. Поток CO 2 автоматически прекращался при достижении заданного значения pH. Основываясь на ежечасных измерениях pH, было показано, что система контроля с отрицательной обратной связью поддерживает pH (и p CO 2 ) на почти постоянном уровне до 4 недель. Используемые четыре заданных значения pH соответствовали желаемой цели обработки воды с известной общей щелочностью.Зонды были откалиброваны перед экспериментом с использованием прослеживаемых буферов NIST (pH = 4,00 и 7,00), повторно откалиброваны по крайней мере один раз во время эксперимента и еще раз по завершении эксперимента для обеспечения правильной работы.
В конце каждого эксперимента личинки из каждого аквариума собирали и фиксировали в 95% этаноле. Из каждой повторности отбирали случайные образцы и фотографировали при 20-кратном увеличении; (n = 205±6,4 оболочки на повторность (среднее значение ± стандартная ошибка среднего)). Панцири личинок клали на бок и фотографировали в силуэте.Площади внешней оболочки измеряли с помощью анализа цифровых изображений [26] (Scion Image, версия 4.0.3.2). Все площади представлены как общие площади внешней поверхности (т. е. 2-кратная сфотографированная площадь = общая площадь поверхности обоих клапанов). Те же образцы были промыты деионизированной водой для удаления морских солей и этанола, а раковины растворены в HCl с содержанием следов металлов и разбавлены до известного объема. Индуктивно-связанная плазменная/оптическая эмиссионная спектроскопия (ICP/OES) использовалась для измерения среднего содержания кальция на скорлупу в каждом повторном образце после обработки.
Из каждой повторности отбирали случайные образцы и фотографировали при 20-кратном увеличении; (n = 205±6,4 оболочки на повторность (среднее значение ± стандартная ошибка среднего)). Панцири личинок клали на бок и фотографировали в силуэте.Площади внешней оболочки измеряли с помощью анализа цифровых изображений [26] (Scion Image, версия 4.0.3.2). Все площади представлены как общие площади внешней поверхности (т. е. 2-кратная сфотографированная площадь = общая площадь поверхности обоих клапанов). Те же образцы были промыты деионизированной водой для удаления морских солей и этанола, а раковины растворены в HCl с содержанием следов металлов и разбавлены до известного объема. Индуктивно-связанная плазменная/оптическая эмиссионная спектроскопия (ICP/OES) использовалась для измерения среднего содержания кальция на скорлупу в каждом повторном образце после обработки.
Для каждой экспериментальной обработки определяли среднюю массу CaCO 3 на единицу площади (мкг/мкм 2 ) путем деления среднего содержания CaCO 3 в скорлупе на среднюю площадь скорлупы. Затем эти значения делили на плотность кристаллического арагонита (2,93 г/см 3 ) [27] для оценки средней кажущейся толщины арагонита личиночных панцирей. Тем не менее, это равномерно завышает толщину оболочки, потому что анализ изображения вычисляет площадь оболочки в силуэте, тем самым занижая истинную трехмерную поверхность оболочки.Эта погрешность не должна влиять на сравнение с лечением одного и того же организма.
Затем эти значения делили на плотность кристаллического арагонита (2,93 г/см 3 ) [27] для оценки средней кажущейся толщины арагонита личиночных панцирей. Тем не менее, это равномерно завышает толщину оболочки, потому что анализ изображения вычисляет площадь оболочки в силуэте, тем самым занижая истинную трехмерную поверхность оболочки.Эта погрешность не должна влиять на сравнение с лечением одного и того же организма.
Результаты
Развитие личинок
Развитие личинок продолжалось с 96 ч после оплодотворения до стадий велигера с глазами и педивелигера (т. е. до достижения компетентности) среди самых крупных размерных классов во всех вариантах лечения и во всех экспериментах. Эксперименты Cv 1 и Ca 1 проводились до возраста личинок 30 и 32 дня соответственно.
Зона оболочки
Crassostrea virginica росла медленнее при p CO 2 , чем при 280 или 380 мкатм, как показывает анализ дисперсии605, n = 12, P = 0,015, рис. 2а). Никаких существенных различий в росте раковины не наблюдалось на уровне P CO 2 процедуры для C. ariakensis ( CA 1: F 3,8 = 0,024, N = 12, p = 12, p = 0,995, рис. 2б). Совокупная частота размеров раковин для каждого вида и обработки показывает: (1) что девяносто процентов раковин C. virginica превышают средний размер класса C. ariakensis и (2) явные эффекты обработки у C.virginica , но не C. ariakensis (рис. 3).
2а). Никаких существенных различий в росте раковины не наблюдалось на уровне P CO 2 процедуры для C. ariakensis ( CA 1: F 3,8 = 0,024, N = 12, p = 12, p = 0,995, рис. 2б). Совокупная частота размеров раковин для каждого вида и обработки показывает: (1) что девяносто процентов раковин C. virginica превышают средний размер класса C. ariakensis и (2) явные эффекты обработки у C.virginica , но не C. ariakensis (рис. 3).
Рисунок 2. Влияние обработки p CO 2 на рост и кальцификацию раковин личинок.
Средние площади скорлупы ± стандартная ошибка среднего (мкм 2 ) (панели A и B) и среднее содержание скорлупы CaCO 3 ± стандартная стандартная ошибка среднего (мкг/скорлупа) (панели C и D), полученные при обработке p CO 2 ±SEM (мкатм) для двух видов устриц. Для каждой обработки указаны соответствующие состояния насыщения арагонита (Ω arag ).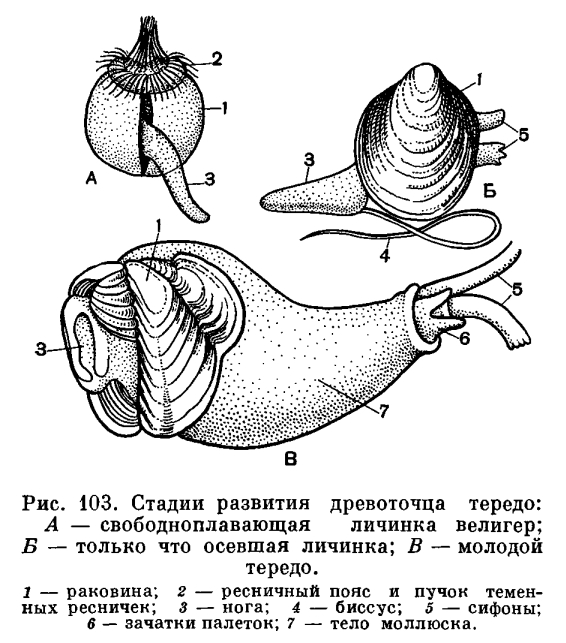
https://doi.org/10.1371/journal.pone.0005661.g002
Рисунок 3. Влияние обработки p CO 2 на совокупную частоту размеров раковин личинок (мкм 2 /раковина) для двух устриц разновидность.
Черные символы и кривые справа показывают результаты для Crassostrea virginica , эксперимент Cv 1 , возраст = 30 d. Незакрашенные символы и левые кривые отображают результаты C. ariakensis (эксперимент Ca 1 , возраст = 32 дня). PCO 2 обработки = 280, 380, 560, 800 мкатм. Количество снарядов на обработку = 615±26,7 (среднее ± стандартная ошибка среднего). C. virginica вырос до заметно больших размеров, чем C. ariakensis . Девяносто процентов раковин C. virginica были такого же размера или больше, чем средний размерный класс C. ariakensis (пунктирная линия). На рост C. virginica повлияли обработка p CO 2 и C.ariakensis не пострадал.
Незакрашенные символы и левые кривые отображают результаты C. ariakensis (эксперимент Ca 1 , возраст = 32 дня). PCO 2 обработки = 280, 380, 560, 800 мкатм. Количество снарядов на обработку = 615±26,7 (среднее ± стандартная ошибка среднего). C. virginica вырос до заметно больших размеров, чем C. ariakensis . Девяносто процентов раковин C. virginica были такого же размера или больше, чем средний размерный класс C. ariakensis (пунктирная линия). На рост C. virginica повлияли обработка p CO 2 и C.ariakensis не пострадал.
https://doi.org/10.1371/journal.pone.0005661.g003
Обызвествление
Количество CaCO 3 , биоминерализуемого личинками Crassostrea virginica , уменьшалось по мере увеличения p CO 2 (рис. 2в). Анализ дисперсии подтвержден значительные различия между P CO 2 лечение ( CV 1: F 3,8 = 11. 8026, N = 12, p = 0.0026). Напротив, четкие тенденции или различия в биоминерализации были очевидны для C. ariakensis ( Ca 1: F 3,7451, N = 11, p = 0,6103, рис. 2d ).
8026, N = 12, p = 0.0026). Напротив, четкие тенденции или различия в биоминерализации были очевидны для C. ariakensis ( Ca 1: F 3,7451, N = 11, p = 0,6103, рис. 2d ).
Насыщенность арагонита
В каждом эксперименте условия для арагонита варьировались от перенасыщения (Ω arag = 1,2−1,3) до недонасыщения (Ω arag = 0,6). На C. ariakensis не влияли различия в насыщенности арагонита, но рост и кальцификация у C.virginica были значительно сокращены, когда p CO 2 было высоким, а Ω arag <1,0 (рис. 2). Важно отметить, что оба вида достигли чистого роста, даже когда состояние насыщения арагонита было значительно ниже 1,0.
Толщина оболочки
Хотя измерения поперечного сечения панцирей личинок напрямую не проводились, мы получили оценку средней толщины панциря, которая теоретически связана с объемом арагонита. Для каждой экспериментальной обработки среднее содержание CaCO 3 на скорлупу делили на соответствующую среднюю площадь скорлупы (мкг/мкм 2 ).Затем это значение делили на плотность кристаллического арагонита (2,93×10 -6 мкг/мкм 3 ), получая среднюю кажущуюся толщину арагонита в мкм. Средняя кажущаяся толщина арагонита была поразительно одинаковой для разных вариантов лечения и экспериментов (от 9 до 11 мкм) независимо от вида или возраста личинок (рис. 4).
Для каждой экспериментальной обработки среднее содержание CaCO 3 на скорлупу делили на соответствующую среднюю площадь скорлупы (мкг/мкм 2 ).Затем это значение делили на плотность кристаллического арагонита (2,93×10 -6 мкг/мкм 3 ), получая среднюю кажущуюся толщину арагонита в мкм. Средняя кажущаяся толщина арагонита была поразительно одинаковой для разных вариантов лечения и экспериментов (от 9 до 11 мкм) независимо от вида или возраста личинок (рис. 4).
Рисунок 4. Влияние обработки p CO 2 на кажущуюся толщину арагонитовой раковины двух видов устриц.
Видимая толщина арагонита (мкм) панцирей личинок = среднее содержание CaCO 3 (мкг)/средняя площадь раковины (мкм 2 )/плотность арагонита (2.93×10 −6 мкг/мкм 3 ). Толщина арагонита была одинакова для разных видов и мало менялась при обработках p CO 2 .
https://doi. org/10.1371/journal.pone.0005661.g004
org/10.1371/journal.pone.0005661.g004
Вторая пара аналогичных экспериментов была проведена с использованием модифицированного дизайна (т. е. питание кормом из двух видов водорослей, использование другого штамма C .ariakensis и большую начальную плотность личинок). Из-за контаминации инфузориями одного из штаммов C.virginica , эксперименты пришлось прекратить досрочно. Тем не менее, для личинок в возрасте 14 дней результаты были аналогичны приведенным выше, указывая на значительное снижение площади раковины и содержания кальция у C. virginica при повышенной p CO 2 , но без значительного p CO 2 эффекты для C. ariakensis .
Обсуждение
Большинство исследований закисления океана были сосредоточены либо на тепловодных кораллах, либо на пелагической биоте [1]–[2], [28].В целом, морская фауна демонстрирует уменьшение кальцификации на возвышенностях p CO 2 (например, склератиниевые кораллы, Gattuso et al. [29], Langdon [9]; птероподы, Feely et al. [15], Orr et al. , [8]; фораминиферы, Bijima и др. [30]; см. Fabry и др. [28] для обзора реакций морской фауны на повышенное содержание p CO 2 ). Как группа, кокколитофориды реагируют более разнообразно, по-видимому, проявляя видоспецифические реакции на повышенное содержание CO 2 [31], при этом Iglesias-Rodriguez et al.[32] сообщают о повышенной кальцификации в Emiliania huxleyi при p CO 2 при 750 мкатм. На сегодняшний день эксперименты в значительной степени имитировали повышенный уровень CO 2 в атмосфере в полностью морских условиях (т. е. при высокой солености), что означает, что вода была перенасыщена CaCO 3 при всех обработках по вызову из-за относительно высокой общей щелочности морской воды.
[29], Langdon [9]; птероподы, Feely et al. [15], Orr et al. , [8]; фораминиферы, Bijima и др. [30]; см. Fabry и др. [28] для обзора реакций морской фауны на повышенное содержание p CO 2 ). Как группа, кокколитофориды реагируют более разнообразно, по-видимому, проявляя видоспецифические реакции на повышенное содержание CO 2 [31], при этом Iglesias-Rodriguez et al.[32] сообщают о повышенной кальцификации в Emiliania huxleyi при p CO 2 при 750 мкатм. На сегодняшний день эксперименты в значительной степени имитировали повышенный уровень CO 2 в атмосфере в полностью морских условиях (т. е. при высокой солености), что означает, что вода была перенасыщена CaCO 3 при всех обработках по вызову из-за относительно высокой общей щелочности морской воды.
В тех случаях, когда морские моллюски подвергались недостаточному насыщению арагонитом, результаты, как правило, были неблагоприятными.Например, Курихара и др. [18] наблюдали, что кальцификация и образование раковин ингибировались на раннем этапе развития личинок Crassostrea gigas , когда морская вода была недонасыщена по отношению к арагониту (Ω arag = 0,68). Когда взрослые птероподы ( Clio pyryata) подвергались воздействию морской воды, недостаточно насыщенной арагонитом, их раковины начинали растворяться в течение 48 часов [8], [15], [28]. Грин и др. [33] показали, что недавно отложенная молодь моллюсков с твердым панцирем (Mercenaria mercenaria , <2.0 мм ) , выращенных в бентических отложениях с недонасыщением поровой воды по CaCO 3 , показали признаки коррозии и значительно увеличили смертность. Холл-Спенсер и др. [34] наблюдали, что раковины взрослых брюхоногих моллюсков, обитающих в сверхвысоких p CO 2 местообитаниях, связанных с естественными подводными вулканическими жерлами CO 2 в Средиземном море, также растворяются, когда Ω≪1,0.
[18] наблюдали, что кальцификация и образование раковин ингибировались на раннем этапе развития личинок Crassostrea gigas , когда морская вода была недонасыщена по отношению к арагониту (Ω arag = 0,68). Когда взрослые птероподы ( Clio pyryata) подвергались воздействию морской воды, недостаточно насыщенной арагонитом, их раковины начинали растворяться в течение 48 часов [8], [15], [28]. Грин и др. [33] показали, что недавно отложенная молодь моллюсков с твердым панцирем (Mercenaria mercenaria , <2.0 мм ) , выращенных в бентических отложениях с недонасыщением поровой воды по CaCO 3 , показали признаки коррозии и значительно увеличили смертность. Холл-Спенсер и др. [34] наблюдали, что раковины взрослых брюхоногих моллюсков, обитающих в сверхвысоких p CO 2 местообитаниях, связанных с естественными подводными вулканическими жерлами CO 2 в Средиземном море, также растворяются, когда Ω≪1,0.
Напротив, когда личинок C. virginica и C. ariakensis непрерывно культивировали с 96 ч после оплодотворения (D-стадия) до ∼30 дней в типичных эстуарных условиях (соленость = 18 psu, TA ≈1225 мкмоль/кг ) и при воздействии повышенных уровней p CO 2 оба вида росли, обызвествлялись и развивались нормально без явных морфологических деформаций, несмотря на условия значительного недонасыщения арагонитом (Ω араг = 0.6−0,7). Эти результаты демонстрируют физиологическую способность личинок устриц выдерживать длительное воздействие (до 28 дней) высоких температур p CO 2 и недостаточного насыщения арагонитом, и противоречат ожиданиям, что личинки в панцире арагонита должны быть особенно склонны к растворению при высоких температурах . р СО 2 [28]. Неудивительно, что некоторые моллюски могут расти в ненасыщенных условиях; большинство пресноводных моллюсков явно хорошо приспособлены к таким условиям.Тем не менее, поскольку содержание CO 2 в атмосфере продолжает расти, все кальцифицирующие организмы будут сталкиваться с условиями, которых они не испытывали в недавней геологической истории.
virginica и C. ariakensis непрерывно культивировали с 96 ч после оплодотворения (D-стадия) до ∼30 дней в типичных эстуарных условиях (соленость = 18 psu, TA ≈1225 мкмоль/кг ) и при воздействии повышенных уровней p CO 2 оба вида росли, обызвествлялись и развивались нормально без явных морфологических деформаций, несмотря на условия значительного недонасыщения арагонитом (Ω араг = 0.6−0,7). Эти результаты демонстрируют физиологическую способность личинок устриц выдерживать длительное воздействие (до 28 дней) высоких температур p CO 2 и недостаточного насыщения арагонитом, и противоречат ожиданиям, что личинки в панцире арагонита должны быть особенно склонны к растворению при высоких температурах . р СО 2 [28]. Неудивительно, что некоторые моллюски могут расти в ненасыщенных условиях; большинство пресноводных моллюсков явно хорошо приспособлены к таким условиям.Тем не менее, поскольку содержание CO 2 в атмосфере продолжает расти, все кальцифицирующие организмы будут сталкиваться с условиями, которых они не испытывали в недавней геологической истории.
Когда Каррикер и Палмер [35] изучали нормальный рост и развитие раковин личинок C. virginica с помощью сканирующей электронной микроскопии, они обнаружили однородные, чрезвычайно тонкие раковины, 4–6 мкм, у личинок длиной 100–350 мкм. . Толщина раковин для их площади, ок. 2–4% от общей длины, вероятно, является приспособлением к планктонному образу жизни.В нашем исследовании оба вида устриц образовали личиночные раковины одинаковой средней толщины, независимо от p CO 2 , Ω arag или площади раковины. Мы интерпретируем образец одинаковой толщины раковины как еще одно свидетельство нормального развития раковины личинок.
Несмотря на их общую физиологическую толерантность к недонасыщенному арагониту, C. virginica и C. ariakensis по-разному реагируют на повышенный уровень p CO 2 . C. virginica демонстрирует стабильно сниженный рост и кальцификацию при высоком уровне p CO 2 . Наоборот, личинок C. ariakensis не по-разному воздействуют на p CO 2 , но растут и кальцифицируются медленнее, чем C. virginica при всех обработках p CO 2 . Эти различия позволяют предположить, что подкисление, вызванное CO 2 , зависит от вида и будет иметь непредсказуемые последствия в эстуариях и прибрежных экосистемах даже для близкородственных видов.
Наоборот, личинок C. ariakensis не по-разному воздействуют на p CO 2 , но растут и кальцифицируются медленнее, чем C. virginica при всех обработках p CO 2 . Эти различия позволяют предположить, что подкисление, вызванное CO 2 , зависит от вида и будет иметь непредсказуемые последствия в эстуариях и прибрежных экосистемах даже для близкородственных видов.
Из строго термодинамических соображений процесс кальцификации в недонасыщенных средах (Ω arag <1,0) должен требовать поступления экзогенной энергии, чтобы идти вперед. Weiner и Dove [36] описывают два преобладающих процесса биоминерализации, широко используемые морскими и эстуарными беспозвоночными. Внеклеточная минерализация используется моллюсками, склератиниевыми кораллами, мшанками и некоторыми фораминиферами и требует активной перекачки ионов или секреции и активного транспорта везикул, содержащих водные растворы с высокой концентрацией ионов, через клеточные мембраны для создания условий, благоприятных для кальцификации. Кокколитофориды, иглокожие и другие формаминиферы используют внутриклеточную биоминерализацию, при которой зародышеобразование минеральных структур происходит непосредственно внутри везикул с использованием аналогичных систем активного транспорта. Полученные карбонатные структуры затем секретируются через клеточную мембрану (Weiner and Dove [36] и ссылки в ней). По мере того, как арагонит становится менее насыщенным, энергетические затраты на биоминерализацию должны постепенно увеличиваться.
Кокколитофориды, иглокожие и другие формаминиферы используют внутриклеточную биоминерализацию, при которой зародышеобразование минеральных структур происходит непосредственно внутри везикул с использованием аналогичных систем активного транспорта. Полученные карбонатные структуры затем секретируются через клеточную мембрану (Weiner and Dove [36] и ссылки в ней). По мере того, как арагонит становится менее насыщенным, энергетические затраты на биоминерализацию должны постепенно увеличиваться.
Мы не воспринимаем недосыщенность арагонита как «формальный» барьер для обызвествления морскими или эстуарными организмами в целом.Состояние насыщения арагонита является скорее удобным показателем легкости, с которой может быть осуществлена биоминерализация. Чем выше состояние насыщения, тем больше относительная доступность как Ca 2+ , так и , и тем ниже потребность в энергии для биохимической накачки для создания локального пересыщения, используемого при производстве материала оболочки. Подобно тому, как виды различаются в зависимости от многих требований к окружающей среде, они, вероятно, различаются по своей способности или эффективности поставлять энергию для кальцификации.Морские таксоны, такие как кораллы и птероподы, которые хорошо приспособлены к морским условиям, где как Ca 2+ , так и были обильны и относительно постоянны в течение миллионов лет, могут иметь проблемы с кальцификацией по мере сокращения из-за антропогенного обогащения CO 2 , даже хотя их вода остается выше насыщения как для арагонита, так и для кальцита. Напротив, личинки эвригалинных устриц C. virginica и C. ariakensis , культивируемых в мезогалинных условиях с низким содержанием Ca 2+ и CO 3 2+ , способны генерировать и поддерживать арагонитовые раковины, когда несколько ниже насыщения.Хотя кальцификация подавляется в состояниях с более низким насыщением для C. virginica , C. ariakensis менее подвержен этим условиям.
Подобно тому, как виды различаются в зависимости от многих требований к окружающей среде, они, вероятно, различаются по своей способности или эффективности поставлять энергию для кальцификации.Морские таксоны, такие как кораллы и птероподы, которые хорошо приспособлены к морским условиям, где как Ca 2+ , так и были обильны и относительно постоянны в течение миллионов лет, могут иметь проблемы с кальцификацией по мере сокращения из-за антропогенного обогащения CO 2 , даже хотя их вода остается выше насыщения как для арагонита, так и для кальцита. Напротив, личинки эвригалинных устриц C. virginica и C. ariakensis , культивируемых в мезогалинных условиях с низким содержанием Ca 2+ и CO 3 2+ , способны генерировать и поддерживать арагонитовые раковины, когда несколько ниже насыщения.Хотя кальцификация подавляется в состояниях с более низким насыщением для C. virginica , C. ariakensis менее подвержен этим условиям. Эстуарии и открытые океаны существенно различаются по своим физическим и химическим характеристикам и динамике, как и история эволюции их биоты. По этим причинам ожидания в отношении биологической реакции на повышение содержания CO 2 в атмосфере и подкисление также должны быть разными.
Эстуарии и открытые океаны существенно различаются по своим физическим и химическим характеристикам и динамике, как и история эволюции их биоты. По этим причинам ожидания в отношении биологической реакции на повышение содержания CO 2 в атмосфере и подкисление также должны быть разными.
Из-за крайней пространственной и временной неоднородности эстуариев и прибрежных систем единообразное равновесие с атмосферным CO 2 не ожидается и не достигается [37]–[38].Эстуарные местообитания, где селятся и живут моллюски, часто испытывают локальные высокие концентрации p CO 2 , вплоть до 6000 мкатм, из-за бентического дыхания [39]. Наши результаты показывают, что для одного ключевого вида, C. virginica , рост и кальцификация уменьшаются при относительно низких повышениях p CO 2 , условиях, которые попадают в пределы изменчивости, которые в настоящее время существуют в его естественной среде обитания. В сочетании с существующим естественным воздействием будущее увеличение до p CO 2 может значительно повлиять на успех личинок и, в конечном итоге, на успех и распространение этих устриц в Чесапикском заливе и подобных системах.
Мы прогнозируем, что по мере увеличения концентрации CO 2 в атмосфере условия в эстуариях будут менее благоприятными для кальцификации во всех зонах солености и создадут новые экологические стрессы для их обитателей. Для некоторых кальцинирующих видов дополнительная энергетическая нагрузка, связанная с образованием скорлупы в менее благоприятных условиях, может ограничивать рост. Наши результаты предполагают, что это будет иметь место для C. virginica , но не обязательно для C. ariakensis. По мере роста затрат на энергию кальцифицирующая биота может столкнуться с компромиссом: либо выделять больше энергии на кальцификацию за счет роста и сохранения запасов энергии, либо соглашаться на меньшую степень кальцификации в обмен на больший рост и большее накопление энергия.
Когда личинки беспозвоночных испытывают физиологический стресс из-за химических и физических факторов, встречающихся в окружающей их среде, они иногда замедляют оседание и метаморфоз.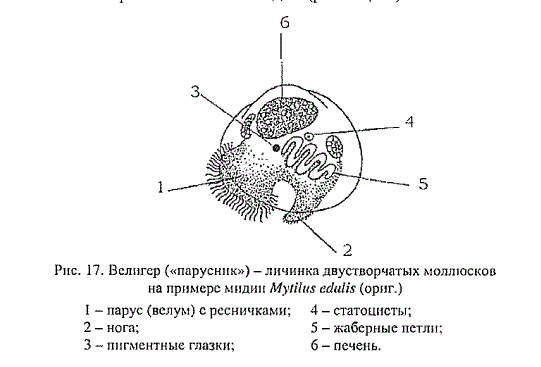 Печеник [40] перечисляет температуру, соленость, питание, низкий уровень растворенного кислорода, загрязнение и ультрафиолетовое излучение в качестве возможных стрессоров, которые могут вызвать замедленный метаморфоз, и далее указывает, что личинки более чувствительны к таким стрессорам, чем взрослые особи (Печеник [41] и ссылки в этой работе). ). Высокое насыщение p CO 2 и пониженное насыщение CaCO 3 могут давать аналогичные эффекты.Для планктотрофных личинок даже незначительное увеличение расхода энергии на производство раковины может уменьшить запасы энергии, которые имеют решающее значение для роста и выживания личинок. Метаморфоз требует больших энергетических затрат и полностью зависит от запасов энергии, накопленных на личиночной стадии [41]–[42]. Если подкисление, вызванное CO 2 , неблагоприятно влияет на рост и кальцификацию личинок, личинки вполне могут оказаться в невыгодном энергетическом отношении, поскольку они пытаются метаморфозом, что приводит к снижению выживаемости и приспособленности.
Печеник [40] перечисляет температуру, соленость, питание, низкий уровень растворенного кислорода, загрязнение и ультрафиолетовое излучение в качестве возможных стрессоров, которые могут вызвать замедленный метаморфоз, и далее указывает, что личинки более чувствительны к таким стрессорам, чем взрослые особи (Печеник [41] и ссылки в этой работе). ). Высокое насыщение p CO 2 и пониженное насыщение CaCO 3 могут давать аналогичные эффекты.Для планктотрофных личинок даже незначительное увеличение расхода энергии на производство раковины может уменьшить запасы энергии, которые имеют решающее значение для роста и выживания личинок. Метаморфоз требует больших энергетических затрат и полностью зависит от запасов энергии, накопленных на личиночной стадии [41]–[42]. Если подкисление, вызванное CO 2 , неблагоприятно влияет на рост и кальцификацию личинок, личинки вполне могут оказаться в невыгодном энергетическом отношении, поскольку они пытаются метаморфозом, что приводит к снижению выживаемости и приспособленности. Кроме того, Печеник [41] указывает, что латентные личиночные эффекты (т. е. влияние на постметаморфический рост и смертность из-за опыта личинок) могут быть общими и видоспецифичными среди морских организмов.
Кроме того, Печеник [41] указывает, что латентные личиночные эффекты (т. е. влияние на постметаморфический рост и смертность из-за опыта личинок) могут быть общими и видоспецифичными среди морских организмов.
Наши результаты показывают, что по мере роста содержания CO 2 в атмосфере в ближайшие десятилетия личинки C. virginica и, возможно, других эстуарных видов будут расти и кальцифицироваться медленнее, чем сегодня. Если личинки не смогут более эффективно оседать/метаморфизировать в будущем, замедление роста и кальцификация приведут к более длительному пребыванию в толще воды, что может привести к более высокой смертности личинок перед поселением.Смертность от хищничества и болезней увеличивается, чем дольше личинки остаются в толще воды [43]–[44]. Чтобы проиллюстрировать влияние относительно небольших изменений в толще воды и смертности на пополнение, Кеннеди [45] смоделировал теоретическую судьбу потомков личинок C. virginica от одной самки, используя данные о естественной суточной смертности от различных исследователей. Было постулировано снижение успешного пополнения на 89%, когда метаморфоз был отложен на пять дней (с 20 до 25 дней), а ежедневная смертность увеличивалась с 20% до 25%.Если более высокий уровень смертности в толще воды сочетается с более низким качеством личинок при поселении (т. е. личинки оседают с меньшими запасами энергии), то можно ожидать, что темпы пополнения в будущем снизятся.
Было постулировано снижение успешного пополнения на 89%, когда метаморфоз был отложен на пять дней (с 20 до 25 дней), а ежедневная смертность увеличивалась с 20% до 25%.Если более высокий уровень смертности в толще воды сочетается с более низким качеством личинок при поселении (т. е. личинки оседают с меньшими запасами энергии), то можно ожидать, что темпы пополнения в будущем снизятся.
Неизвестно, влияют ли продолжительное пребывание в планктоне и повышенные затраты энергии во время роста и развития личинок из-за повышенного содержания CO 2 на более поздних стадиях жизни и, в конечном счете, на динамику популяции и экологию C. virginica или других видов.Ясно, что личинки обоих изученных нами видов Crassostrea обладают физиологической способностью расти и кальцифицироваться в условиях недостаточного насыщения, но если и каким образом увеличение p CO 2 во всех точках их обитания отрицательно повлияет на выживаемость в планктоне. а также метаморфический успех и динамику пополнения еще предстоит проверить.
а также метаморфический успех и динамику пополнения еще предстоит проверить.
На тихоокеанском побережье США коммерческие предприятия по разведению устриц испытывают тревожные и широко распространенные трудности с содержанием тихоокеанской устрицы (C.gigas) живых личинок в культуре, при этом два крупнейших инкубатория сообщают о снижении производительности на целых 80%. Более того, «естественное» пополнение в течение нескольких лет было незначительным или отсутствовало в районах, где C. gigas ранее создали натурализованные популяции. (Р. Дауни, Ассоциация производителей моллюсков Тихоокеанского побережья, личное сообщение). В районах апвеллинга вдоль континентального шельфа западной части Северной Америки Feely et al. [46] определили, что pH и Ω arag поверхностных вод более кислые и имеют меньшую насыщенность арагонитом, чем ожидалось.Сообщалось, что на глубине воды 40–120 м во многих местах вдоль побережья, но на глубине 0 м в районе около границы Калифорнии и Орегона pH составлял ~7,75, а Ω arag <1,0. Неизвестно, связаны ли недавние неудачи пополнения и аквакультуры с изменениями в химическом составе карбонатов, но это должно быть исследовано.
Неизвестно, связаны ли недавние неудачи пополнения и аквакультуры с изменениями в химическом составе карбонатов, но это должно быть исследовано.
Естественная пространственная и временная неоднородность солености, щелочности, рН и p CO 2 в эстуариях налагает более сложные экологические нагрузки на кальцифицирующую биоту, чем в открытом океане, где окружающая среда гораздо менее изменчива.По мере увеличения концентрации CO 2 в атмосфере будет увеличиваться доля ненасыщенных для арагонита эстуарных местообитаний. Мы предсказываем, что в мире с высоким содержанием CO 2 точка компенсации арагонита в эстуариях (изоплет солености, где Ω arag = 1,0) сместится в сторону моря в сторону более высокой солености. Поскольку эстуарные виды эволюционировали в таких изменчивых средах, они могут обладать большей физиологической способностью реагировать на подкисление, вызванное CO 2 , чем полностью морские таксоны.Тем не менее адаптивной физиологической толерантности личинок может быть недостаточно для поддержания популяций кальцифицирующих бентосных видов, включая широко распространенные важные с экономической точки зрения промыслы, перед лицом изменения атмосферного CO 2 .
Благодарности
Мы благодарим J.P. Megonigal, E. Jewett, P. Neale и R. Osman за консультации и технические советы. Мы благодарим S. Bonniwell и M. Luckenbach за предоставление личинок устриц и советы по культивированию, а также K.Эразмус, Р. Ловенталь и К. Стулл, Г. Ридель, Д. Бутера, С. Сантагата, К. Клуг, М. Хаган и Г.М. Ruiz за их вклад в эти эксперименты и рукопись.
Вклад авторов
Задумал и спроектировал эксперименты: AWM CS GFR. Проведены эксперименты: AWM ACR GFR. Проанализированы данные: AWM ACR CS GFR. Предоставленные реагенты/материалы/инструменты для анализа: AWM ACR CS GFR. Написал статью: AWM ACR CS GFR.
Каталожные номера
- 1. Королевское общество (2005 г.) Закисление океана из-за увеличения содержания углекислого газа.Программный документ 12/05. Королевское общество.
- 2.
Клейпас Дж.А., Фили Р.А., Фабри В.Дж., Лэнгдон С., Сабина С.Л. и др. (2006) 88. Воздействие закисления океана на коралловые рифы и другие морские известняки: руководство для будущих исследований, отчет о семинаре, проведенном 18–20 апреля 2005 г.
 , Санкт-Петербург, Флорида, при поддержке Национального научного фонда, NOAA и Геологической службы США. Опрос.
, Санкт-Петербург, Флорида, при поддержке Национального научного фонда, NOAA и Геологической службы США. Опрос. - 3. Терли С., Блэкфорд Дж., Виддикомб С., Лоу Д., Найтингейл П.Д. и др. (2006) Обзор воздействия повышенного содержания CO2 в атмосфере на рН океана и морскую экосистему.Шеллнхубер Х.Дж., Крамер В., Накиценович Н., Вигли Т., Йохе Г., редакторы. Предотвращение опасного изменения климата, издательство Кембриджского университета, 8. стр. 65–70.
- 4. Сабин С.Л., Фили Р.А., Грубер Н., Ки Р.М., Ли К. и др. (2004) Океанический поглотитель антропогенного CO2. Наука 305: 367–371.
- 5. Канаделл Дж. Г., Ле Кве ре К., Раупах М. Р., Филд С. Б., Буйтенхуис Э. Т. и др. (2007) Вклад в ускорение роста атмосферного CO2 в результате экономической деятельности, углеродоемкости и эффективности естественных поглотителей.Proc Nat Acad Sci USA 104 (47): 18866–18870.
- 6.
Caldeira K, Wickett ME (2003) Антропогенный углерод и рН океана.
Природа 425: 365.
- 7. Caldeira K, Wickett ME (2005) Предсказания модели океана изменений химического состава в результате выбросов углекислого газа в атмосферу и океан, J Geophys Res Oceans 110(C9), C09504:
- 8. Orr JC, Fabry VJ, Aumont O, Bopp L, Doney SC, et al. (2005) Антропогенное закисление океана в двадцать первом веке и его влияние на кальцифицирующие организмы.Природа 437 (7059): 681–686.
- 9. Langdon C (2002) Обзор экспериментальных данных о влиянии CO2 на кальцификацию рифостроителей. Материалы 9-го -го -го Международного симпозиума по коралловым рифам 2: 1091–1098.
- 10. Gazeau F, Quiblier C, Jansen JM, Gattuso J-P, Middelburg JJ, et al. (2007) Влияние повышенного содержания CO2 на кальцификацию моллюсков. Geophys Res Lett 34: L07603.
- 11.
Карпентер К.Е., Абрар М., Эби Г., Аронсон Р.Б., Бэнкс С. и др.(2008) Одна треть рифообразующих кораллов сталкивается с повышенным риском исчезновения из-за изменения климата и местных воздействий.
 Наука 321: 560–563.
Наука 321: 560–563. - 12. Wong GT (1979) Щелочность и pH в южной части Чесапикского залива и устье реки Джеймс. Лимнол Океаногр 24: 970–977.
- 13. Cai WJ, Wang Y (1998) Химический состав, потоки и источники углекислого газа в устьевых водах рек Сатилла и Альтамаха, Джорджия. Лимнол Океаногр 43: 657–668.
- 14.Дони С.К., Фабри В.Дж., Фили Р.А., Клейпас Дж.А. (2009) Подкисление океана: другая проблема CO 2 . Annu Rev Mar Sci 1: 169–192.
- 15. Фили Р.А., Сабина К.Л., Ли К., Берелсон В., Клейпас Дж. и др. (2004) Воздействие антропогенного CO 2 на систему CaCO 3 в океанах. Наука 305: 362–366.
- 16. Солсбери Дж., Грин М., Хант С., Кэмпбелл Дж. (2008) Подкисление побережья реками: новая угроза моллюскам? ЭОС Транс АГУ 89: 513.
- 17.
Вайс И.М., Туросс Н., Аддади Л., Вайнер С. (2002)Формирование раковины личинок моллюсков: аморфный карбонат кальция является предшественником арагонита.
 J Exp Biol 293: 478–491.
J Exp Biol 293: 478–491. - 18. Курихара Х., Като С., Ишимацу А. (2007) Влияние повышенного pCO 2 в морской воде на раннее развитие устрицы Crassostrea gigas . Акват Биол 1: 91–98.
- 19. Kirby MX (2004) Рыбалка у побережья: Историческое расширение и крах промысла устриц вдоль континентальных окраин.Proc Acad Nat Sci USA 101: 13096–13099.
- 20. Брейтбург Д.Л., Ридель Г.Ф. (2005) Множественные стрессоры в морских системах. Биология сохранения морской среды: наука о поддержании биоразнообразия моря (MCB). стр. 167–182. Island Press, Вашингтон, округ Колумбия
- 21. OSB (2004) Неместные устрицы в Чесапикском заливе. Совет по изучению океана, Отдел исследований Земли и жизни, Национальный исследовательский совет национальных академий, The National Academy Press, Вашингтон, Д.С., 325 с.
- 22.
Бриз В.П., Малуф Р.Е. (1977) Методы выращивания в инкубаторе Crassostrea rivularis.
 Аквакультура 12: 123–126.
Аквакультура 12: 123–126. - 23. Эдмонд Дж.М. (1970) Высокоточное определение титрационной щелочности и общего содержания углекислого газа в морской воде методом потенциометрического титрования. Deep Sea Res 17: 737–750.
- 24. Леггетт Дж., Пеппер В.Дж., Сварт Р.Дж. (1992) Сценарии выбросов для МГЭИК: обновление. Изменение климата, 1992 г.: Дополнительный отчет к научной оценке МГЭИК.стр. 69–95. Издательство Кембриджского университета, Кембридж.
- 25. Пеллетье Г., Льюис Э., Уоллес Д. (2007) CO2sys.xls: калькулятор для системы CO 2 в морской воде для Microsoft Excel/VBA, Департамент экологии штата Вашингтон/Брукхейвенская национальная лаборатория, Олимпия, Вашингтон/Аптон, Нью-Йорк, США
- 26. Образ Scion для Windows. Альфа-версия 4.0.3.2, © 2000–2001, Scion Corporation, www.scioncorp.com.
- 27.
Lide DR (2007) Справочник CRC по химии и физике, 88-е издание (Справочник CRC по химии и физике).
 CRC.
CRC. - 28. Фабри В.Дж., Сейбель Б.А., Фили Р.А., Орр Дж.К. (2008) Воздействие закисления океана на морскую фауну и экосистемные процессы. ICES J Mar Sci 65: 414–432.
- 29. Gattuso JP, Frankignoulle M, Bourge I, Romaine S, Buddemeir RW (1998) Влияние насыщения морской воды карбонатом кальция на кальцификацию кораллов. Глобальное изменение планеты 18: 37–46.
- 30. Bijima J, Honisch B, Zeebe RE (2002) Влияние химического состава карбонатов океана на вес раковин живых фораминифер: комментарий к «Концентрации ионов карбоната в глубинных водах Карибского моря ледникового периода» В. С. Брокера и Э. Кларка.Геохим. Геофиз. Геосист 3: 1064.
- 31. Fabry VJ (2008) Морские кальцификаторы в океане с высоким содержанием CO2. Наука 320: 120–122.
- 32.
Iglesias-Rodriguez MD, Halloran PR, Rickaby REM, Hall IR, Colmenero-Hidalgo E, et al. (2008) Кальцификация фитопланктона в мире с высоким содержанием CO2.
 Наука 320: 336–340.
Наука 320: 336–340. - 33. Грин М.А., Джонс М.Е., Будро К.Л., Мур Р.Л., Вестман Б.А. (2004) Смертность от растворения молодых двустворчатых моллюсков в прибрежных морских отложениях.Лимнол Океаногр 49(3): 727–734.
- 34. Холл-Спенсер Дж. М., Родольфо-Металпа М., Мартин С., Рэнсом Э., Файн М. и др. (2008) Вулканические жерла углекислого газа демонстрируют воздействие окисления океана на экосистему. Природа-онлайн.
- 35. Каррикер М.Р., Палмер Р.Е. (1979) Ультраструктурный морфогенез створок продиссоконхов и ранних диссоконхов устрицы Crassostrea virginica . Proc Natl Моллюски. Доц. 69: 102–128.
- 36. Weiner S, Dove PM (2003) Обзор процессов биоминерализации и проблемы жизненного эффекта.Биоминерализация 1–29. Обзоры по минералогии и геохимии 54.
- 37.
Смит С.В., Холлибо Дж.Т. (1997) Годовой цикл и межгодовая изменчивость метаболизма экосистемы в заливе с умеренным климатом. Экол Моногр 67: 509–533.

- 38. Frankignoulle M, Abril G, Borges A, Bourge I, Canon C и др. (1998) Выбросы двуокиси углерода из устьев рек Европы. Наука 282: 434–436.
- 39. Cai WJ, Pomeroy LR, Moran MA, Wang Y (1999) Баланс массы кислорода и углекислого газа для эстуарно-приливного болотного комплекса пяти рек на юго-востоке США.С. Лимно. Океаногр 44: 639–649.
- 40. Печеник Ю.А. (1999) О преимуществах и недостатках личиночных стадий в жизненных циклах донных морских беспозвоночных. Mar Ecol Prog Ser 177: 269–297.
- 41. Печеник Ю.А. (2006) Личиночный опыт и латентные эффекты — метаморфозы — это не новое начало. Интегр Комп Биол 46: 323–333.
- 42. Burke K, Batller E, Miron G, Ouellette M, Tremblay R (2008) Качество личинок неместных видов двустворчатых моллюсков (европейская устрица, Ostrea edulis ) у восточного побережья Канады.J Shellfish Res 27 (4): 701–710.
- 43.
Морган С.Г. (1996) Жизнь и смерть в планктоне: лавальная смертность и адаптация.
 Экология личинок морских беспозвоночных. стр. 279–321. CRC Press, Нью-Йорк, штат Нью-Йорк.
Экология личинок морских беспозвоночных. стр. 279–321. CRC Press, Нью-Йорк, штат Нью-Йорк. - 44. Андервуд А.Дж., Фэйрвезер П.Г. (1989) Экология снабжения и бентосные морские сообщества. Тенденции Ecol Evol 4: 16–20.
- 45. Кеннеди В.С. (1996) Биология личинок и спата. Устрица восточная, Crassostrea virginica .стр. 371–441. Мэриленд Си Грант, Колледж-Парк, Мэриленд.
- 46. Фили Р.А., Сабин К.Л., Эрнандес-Айон Дж.М., Янсон Д., Хейлз Б. (2008) Доказательства подъема коррозионно-кислой воды на континентальный шельф. Наука 320 (5882): 1490–1492.
- 47. Программа Чесапикского залива. База данных качества воды программы Чесапикского залива с 1984 г. по настоящее время. По состоянию на 20 октября 2008 г., с http://www.chesapeakebay.net/wquality.htm.

