
Ресничные черви — класс, строение, питание
Ресничные черви, или тубеллярии – класс типа плоские черви, включающий более 10 отрядов, которые объединяют более 3, 5 тысяч видов. Подавляющее большинство этих червей свободноживущие, лишь некоторые виды паразитируют в организмах животных – моллюсков, ракообразных, иглокожих, некоторых других животных. Свободноживущие формы встречаются в пресных и соленых водоемах, иногда в тропическом поясе с повышенной влажностью на суше в почве. Самые известные представители данного класса – виды отряда планарии (черная, белая, бурая, т.д.).
Ресничные черви, не являющиеся паразитами, питаются как хищники мелкими беспозвоночными. Они передвигаются, плавая в воде или ползая по дну водоемов или на суше по поверхности почвы.
Размеры тела ресничных червей составляют от миллиметра до 30 см. Это самые примитивные двусторонне-симметричные животные. Тело может быть овальной, вытянутой или уплощенной формы, покрыто ресничным эпителием.
Внутреннее строение ресничных червей имеет характерные особенности. Так, полости тела нет, все промежутки между внутренними органами заполнены паренхимой. Ротовое отверстие находится на брюшной стороне, чаще в средней части тела, но может быть на переднем или заднем его конце. Проглоченная пища попадает в мускулистую глотку. У примитивных видов ресничных червей пищеварительный процесс осуществляется в специальных пищеварительных клетках либо в отдельных частях паренхимы. У более развитых ресничных червей имеется слепо оканчивающийся разветвленный или мешковидный кишечник.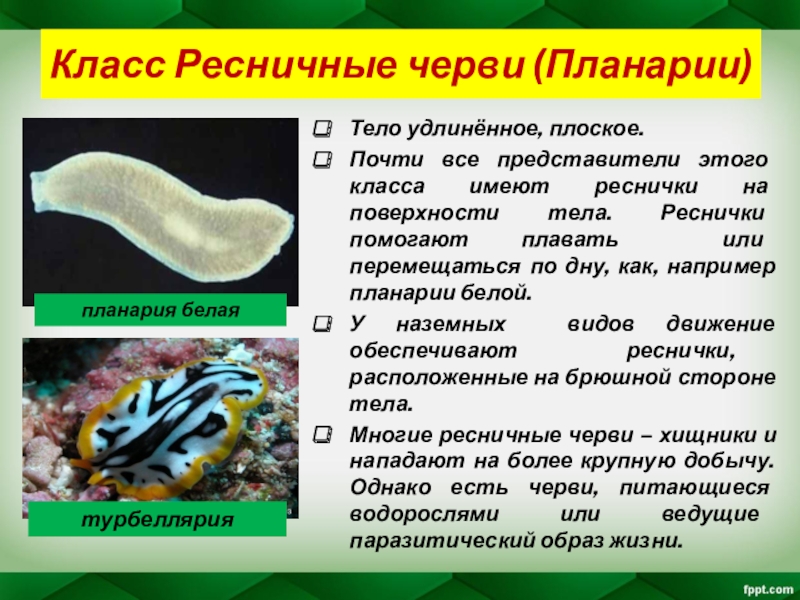
Органы кровообращения у ресничных червей отсутствуют. Дыхание осуществляется всей поверхностью тела. Выделительная система представлена протонефридиями, примитивные виды лишены органов выделения.
Нервная система у ресничных червей может быть двух типов. У низших червей она диффузная, нервные клетки расположены в толще кожного эпителия. У высших представителей данного класса нервная ткань представлена головными нервными узлами и парными нервными стволами, связанными перемычками. Органы чувств ресничных червей – это обонятельные ямки, глаза, щетинки и жгутики как органы осязания, у некоторых видов имеются статоцисты как органы равновесия.
Все ресничные черви являются гермафродитами, то есть у каждой особи имеются и женские, и мужские половые железы. Мужские гаметы формируются в множественных семенниках, они через семявыносящие протоки выбрасываются наружу. Женский половой аппарат включает яичники, желточники, образующие желточные клетки для питания зародыша, и женские половые протоки.
Развитие ресничных червей прямое. Из яйца появляется личинка, внешне сходная со взрослой особью. У редких видов развитие протекает с метаморфозом. Некоторые черви способны размножаться бесполым путем – делением пополам.
Значение в природе ресничных червей обусловлено их участием в сложных пищевых цепочках. Эти животные – сами хищники и одновременно являются пищей для других животных.
Статьи по теме:
1. Черви
2. Плоские черви
3. Планарии
Особенности строения ресничных червей таблица. Класс ресничные черви или турбеллярии
Класс ресничные черви или турбеллярии — относятся к типу плоские черви. Тип плоские черви включает такие классы, как ресничные, сосальщики, ленточные и насчитывает около 26000 видов.
Представитель ресничных червей
Происхождение . Ближайшими предками ресничных червей считаются кишечнополостные животные. В ходе эволюции черви перешли к активному способу жизни, как следствие у них усовершенствовалась половая система, выделительная, нервная, сформировалась билатеральная симметрия (зеркальное отражение противоположных сторон тела).
Особенности строения ресничных червей
Форма тела ресничных червей продолговатая, иногда в виде листа. Окрашено оно в белый цвет или иногда вовсе остается бесцветным. Часто внешние покровы имеют разнообразный окрас, в зависимости от содержащегося пигмента.
Снаружи черви укрыты однослойным ресничным эпителием. Во время взросления реснички теряются, поэтому взрослые особи вовсе их лишены. В эпителиальном слое находятся железистые структуры, хаотично раскиданы по телу или сосредоточены в группах.
Черви обладают специфическими железами, которые состоят из рабдитных клеток, в них есть особые палочки, что преломляют лучи света. Если к животному прикоснутся, рабдиты выходят наружу и набухают. В следствии, на теле червя формируется слизистый слой. Считают, что он необходим для защиты от врагов. На боковой части тела находятся железы, выделяющие токсические вещества, парализующие жертв.
Если к животному прикоснутся, рабдиты выходят наружу и набухают. В следствии, на теле червя формируется слизистый слой. Считают, что он необходим для защиты от врагов. На боковой части тела находятся железы, выделяющие токсические вещества, парализующие жертв.
Под наружным покровом расположена базальная мембрана, которая придает телу ту или иную форму, служит местом крепления мышц. Вместе, мышцы и эпителиальный шар, формируют кожно-мускульный мешок. Гладкомышечные волокна расположены слоями, снаружи находятся кольцевые мышцы, в среднем слое — продольные, во внутреннем — диагональные. Также ресничные черви обладают спинно-брюшными или дорзовентральными мышцами.
Активное перемещение возможно у небольших червей благодаря движению ресничек, другие же представители передвигаются с помощью кожно-мускульного мешка.
Внутренняя полость наполнена соединительной тканью. Мелкие промежутки между ее клетками содержат жидкость, при помощи которой передаются питательные вещества с кишечной полости к тканям, и происходит транспорт метаболических продуктов к органам выделения. Также соединительная ткань выполняет опорную функцию.
Также соединительная ткань выполняет опорную функцию.
Система пищеварения . Пищеварительный тракт слепозамкнутый. Ротовое отверстие находится на задней части тела, предназначено для захвата пищи и для выведения непереваренных частиц. Рот переходит в глотку, которая имеет складчатое строение. От нее отходит несколько ветвей кишечника. У крупных видов ветви располагаются следующим образом: одна – впереди, где пищеварение идет в первую очередь, и две — сзади, расположенные ближе к боковым частям тела.
В глоточном отделе находятся железы, которые играют особую роль в добыче пищи. Так, ресничные черви нападают на моллюсков и вводят в их тело пищеварительные ферменты, и только после частичного переваривания, они поглощают еду.
Большинство представителей хищники и употребляют мелких беспозвоночных. Они накрывают жертву телом и проглатывают ее. У некоторых видов для этого есть выдвижная глотка.
Нервная система
ресничных червей. Она представлена совокупностью нервных клеток, сосредоточенных в передней части тела. Они образуют двойной узел — мозговой ганглий (служит своеобразным координирующим звеном). От узла идут параллельные нервные стволы, которые связываются между собой с помощью перемычек.
Они образуют двойной узел — мозговой ганглий (служит своеобразным координирующим звеном). От узла идут параллельные нервные стволы, которые связываются между собой с помощью перемычек.
Дыхание ресничных червей происходит всей поверхностью тела, так как они лишены специфических дыхательных органов. Системы кровообращения у ресничных червей нет.
Органы чувств у ресничных червей достаточно развиты. Есть тактильная чувствительность благодаря рецепторам кожи. Статоцисты обеспечивают поддержание равновесия, также содержат внутри примитивный слуховой аппарат. В передней части тела размещены обонятельные ямки, которые воспринимают запахи и необходимы червям во время охоты. Рядом расположены глаза, от одной пары до нескольких.
Система выделения
. Выделительные органы состоят из двух или больше каналов, которые через наружное отверстие сообщаются с внешней средой. В теле они разветвляются, формируя сеть. Концевые канальцы содержат звездчатые клетки с пучками ресничек.
Размножение ресничных червей. Ресничные черви имеют половые признаки обоих полов. Они содержат как мужские, так и женские гаметы. Большинство видов размножаются половым путем. Формирование взрослой особи возможно прямым способом, но несколько обитателей морей развиваются с метаморфозом, когда на свет появляется личинка не похожая на зрелый организм. Некоторым представителям ресничных червей свойственно бесполое размножение. При этом если разделить червя на две части, из каждой разовьется полноценная особь.
Значение ресничных червей в природе и жизни человека
Вызывают тяжелые заболевания у человека и животных, названные гельминтозами. Вред наносят аквариумным рыбкам, поселяясь в недостаточно очищенной воде, они поедают икру, а взрослых особей парализуют токсическими веществами.
Также ресничные черви являются неотъемлемой частью пищевых цепочек.
Среда обитания ресничных червей . Представители класса распространены повсеместно. В соленых и пресных водах, меньше их в почве, тропических широтах, с влажным климатом. Попадая в живой организм, селятся в полости кишечника.
Представителей мира простейших микроорганизмов большое количество. К ним относятся ресничные черви, которых в природе насчитывается около 3500 типов.
Некоторые из полосатых червей совершенно безопасны, а другие могут нести опасность для жизни и здоровья человека. Их размеры зависят от места среды, где они проживают.
Чтобы рассмотреть некоторых из них, необходим микроскоп, а есть особи, которые достигают 40 см в длину. Они могут свободно обитать в окружающей среде, но при этом являются хищными.
Среда обитания — это преимущественно в близи водоемов, или на его берегу, земле, под камнями.
Также им подходит для проживания увлажненная почва. Отличительным классом считаются планарии, так как имеют самую разнообразную расцветку.
Отличительным классом считаются планарии, так как имеют самую разнообразную расцветку.
Эти особи названы не просто так ресничными, все их тело покрыто именно ими. Реснички необходимы для того, чтобы передвигаться. Ресничные особи перемещаются в пространстве словно змеи, могут ползти или плыть.
Строение и форма ресничных червей овального типа, немного приплющенная, но встречаются особи удлиненного вида. Все представители этих червей имеют двустороннее симметричное строение, спереди у них есть ротовая полость, которая расположена на брюхе.
Также в передней части тела они имеют органы чувств.
Плоские черви имеют свои характерные особенности строения ресничного покрова, и обладают мерцательным эпителием. Выделяют два типа – это слипшиеся и раздельные реснички. Однако не все представители этого класса обладают ими.
Червь передвигается при помощи секретной жидкости, которая вырабатывается железами секреции. Ее выделение происходит в передней части тела особи, и она помогает удерживать равновесие и свободно перемещаться по объекту, на котором находиться. По краям ресничные имеют железы одноклеточного происхождения.
По краям ресничные имеют железы одноклеточного происхождения.
Слизь, которая ими выделяется, обладает некоторой токсичностью. В природе она им необходима для защиты от представителей более крупных микроорганизмов. С возрастом черви начинают терять свои реснички, и внешне это напоминает линьку, как у кошки или собаки.
Внутреннее строение ресничных особей полностью совпадает с плоским типом микроорганизмов. Их кожно-мускульный мешок формируется с помощью мышечного органа.
Он состоит из 3 основных волокон.
Первый – это кольцевой слой, он располагается с внешней стороны особи.
Второй – диагональный, он находиться под небольшим углом, а последний – это продольный слой, который образуется в нижней части организма.
При сокращении мышечных тканей ускоряется процесс перемещения, а крупные ресничные особи увеличивают скорость скольжения.
Пищеварительная и выделительная система
Пищеварительная система ресничных червей имеет сложное внутреннее строение, состоящее из многочисленных органов. По ним внутрь поступают требуемые для жизнедеятельности полезные вещества.
По ним внутрь поступают требуемые для жизнедеятельности полезные вещества.
Существуют ресничные особи, не имеющие полноценной пищеварительной системы, их называют без кишечными. Особи, обладающие четко сформированным кишечным трактом подразделяются на два вида, это ветвистокишечные и прямокишечные.
В зависимости от этого имеются отклонения в строении, у ветвистых червей ротовая полость находиться неподалеку от задней части туловища.
А у особей с прямокишечным типом строения, рот расположен в передней части, он примыкает к глотке, имеющей слепые ветки кишечного тракта. На теле червей имеются железы, которые обеспечивают наружное переваривание продуктов питания, и называются они глоточными.
Микроорганизмы имеют наружное пищеварение, поэтому получать полезные для себя питательные вещества они могут путем прикрепления рта к жертве.
В процессе он выделяет секрет, который способен переварить пищу снаружи, а потом уже всасывает в свой организм. Питаются они в основном мелкими микроорганизмами, которые не имеют позвонков, например рачками.
Питаются они в основном мелкими микроорганизмами, которые не имеют позвонков, например рачками.
У них не совсем простая выделительная система, она состоит из значительного количества пор, через которые из организма выходят переработанные вещества. Многочисленные ветви, из которых состоит выделительная система, соединяются с одним или двумя основными каналами, присоединенные к кишечному тракту.
Когда он отсутствует накопление ненужных веществ происходит на верхней поверхности туловища в некоторых клетках. После того как накопиться достаточное количество экскретов, они сами со временем отсоединяются от червя.
Нервная система и органы чувств
Ресничные черви отличаются особенностью строения нервной системы.
У некоторых представителей этого класса имеется несколько нервных окончаний, которые располагаются только в передней части туловища. А есть плоские черви, которые обладают значительным количеством нейронов, соединяющиеся в основные стволы, их может быть до 8 штук.
Реснитчатым существам присуще иметь систему органов чувств. На их теле присутствуют реснички, которые не имеют подвижность. Существуют представители этого класса, которые умеют чувствовать равновесие, так как у них есть статоцист. Этим органом являются пузырьки или ямки, отличие составляет тип особи.
На окружающую среду они реагируют с помощью сенсилл, это такие неподвижные реснички, которые распределены по всему телу. При наличии органа статициста возможно наличие ортогона.
То есть плоские черви имеют мозговую систему, состоящую из множества каналов, которая относиться к виду решетчатых.
Характеристика строения органов обоняния весьма интересна. Благодаря тому, что существует развитое обоняние, турбеллярия может находить для себя еду. На теле у хищника есть специальные ямки, в которые поступает сигнал от окружающей среды, и передается он в мозговую систему.
Несмотря на то, что на теле около мозговых ганглий имеется несколько глаз, они не имеют зрения. Некоторые ученые утверждают, что крупные плоские черви все-таки могут различать окружающие предметы. По проведенным исследованиям выяснилось, что у них существует хрусталик, который помогает видеть.
Некоторые ученые утверждают, что крупные плоские черви все-таки могут различать окружающие предметы. По проведенным исследованиям выяснилось, что у них существует хрусталик, который помогает видеть.
Реснитчатые особи отличаются от плоских тем, что могут дышать. Плоские черви живут в бескислородной среде, где и размножаются. Реснитчатым необходим кислород, так как они обитают в другой среде. Дышит хищник с помощью своего тела, то есть на его поверхности есть поры, которые и поглощают кислород.
Половая система и способ размножения
Ресничные черви одновременно имеют женские и мужские половые органы, то есть являются гемофрадитами. В семенниках у них располагаются мужские, а в яичниках женские органы.
Семенная жидкость для оплодотворения проходит по специальным каналам и попадает в яйцеклетку. Затем уже поступает в яйцевод, и образовывает клоаку. Размножение класса осуществляется перекрестным методом.
Особи поочередно оплодотворяют друг другу, жидкость спермы они вспрыскивают в отверстие клоаки. Семена образуют яйца, которые покрыты скорлупой. После они откладывают яйца, и впоследствии из них вылупливается новая особь. По внешнему виду они, как взрослые глисты.
Семена образуют яйца, которые покрыты скорлупой. После они откладывают яйца, и впоследствии из них вылупливается новая особь. По внешнему виду они, как взрослые глисты.
Размножение турбулярии несколько отличается от других представителей класса. У них из яйца вылупливается малая личинка, которая уже растет в месте обитания.
Ресничные черви могут размножаться бесполовым способом.
На теле появляется перетяжка, которая через некоторое время разделяет туловище на две отдельные части. И постепенно они приобретают все жизненно необходимые органы.
Ресничные черви необходимы для поддержания баланса в популяции беспозвоночных животных. Хищники поедают их, и тем самым не дают им разрастаться в большом количестве.
Строение кожно-мускульного мешка ресничных червей.
Профилактика.
Диагностика
1.Личная. Соблюдение правил личной гигиены.
2.Общественная. Санитарное обустройство мест общественного пользования, наблюдение за источниками общественного водоснабжения, санитарно-просветительская работа с населением, борьба с грызунами, гигиеническое содержание свиней.
Кожно-мускульный мешок . Под эпителием находится базальная мембрана , служащая для придания телу определенной формы и для прикрепления мышц. Совокупность мышц и эпителия образует единый комплекс —кожно-мускульный мешок . Мышечная система состоит из нескольких слоев гладких мышечных волокон . Наиболее поверхностно располагаются кольцевые мышцы , несколько глубже — продольные и самые глубокие — диагональные мышечные волокна . Кроме перечисленных видов мышечных волокон для ресничных червей характерны спинно-брюшные , или дорзовентральные , мышцы . Это пучки волокон, идущие от спинной стороны тела к брюшной.
Движение осуществляется за счет биения ресничек (у мелких форм) или сокращения кожно-мускульного мешка (у крупных представителей).
Четко выраженной полости тела
ресничные черви не имеют. Все промежутки между органами заполненыпаренхимой
— рыхлой соединительной тканью. Небольшие пространства между клетками паренхимы заполнены водянистой жидкостью, благодаря чему может осуществляться передача продуктов от кишечника к внутренним органам и перенос продуктов обмена к выделительной системе. Кроме этого, паренхиму можно рассматривать как опорную ткань.
Небольшие пространства между клетками паренхимы заполнены водянистой жидкостью, благодаря чему может осуществляться передача продуктов от кишечника к внутренним органам и перенос продуктов обмена к выделительной системе. Кроме этого, паренхиму можно рассматривать как опорную ткань.
Мышечная часть кожно-мускульного мешка образована несколькими слоями мышечных волокон. Снаружи располагается кольцевой слой, под ним продольный и самый нижний — диагональный, волокна которого идут под углом друг к другу. Именно совокупность покровов и лежащих ниже слоев мышечных волокон и образует кожно-мускульный мешок.
Пищеварительная система
ресничных червей слепозамкнутая
. Рот
служит и для проглатывания пищи
, и для выбрасывания непереваренных остатков пищи
. Рот располагается обычно на брюшной стороне тела и ведет вглотку
. У некоторых крупных ресничных червей, например у пресноводной планарии, ротовое отверстие открывается в глоточный карман
, в котором находится мускулистая глотка
, способная вытягиваться и высовываться через рот наружу. Средняя кишка
у мелких форм ресничных червей представляет собой ветвящиеся во все стороны каналы
, а у крупных форм кишечник представлен тремя ветвями
: одной передней
, идущей к переднему концу тела, и двумя задними
, идущими по бокам к заднему концу тела.
Средняя кишка
у мелких форм ресничных червей представляет собой ветвящиеся во все стороны каналы
, а у крупных форм кишечник представлен тремя ветвями
: одной передней
, идущей к переднему концу тела, и двумя задними
, идущими по бокам к заднему концу тела.
Большинство турбеллярии — хищники, питающиеся различными мелкими животными. Пищеварительная система состоит из передней и средней кишки, замыкающейся слепо. Рот служит не только для проглатывания пищи, но и для выбрасывания непереваренных твердых остатков. Рот помещается обычно на брюшной стороне тела: на небольшом расстоянии от переднего конца тела, посредине брюшной стороны, или ближе к заднему полюсу. Рот ведет в эктодермальную глотку, которая, в свою очередь, переходит в среднюю кишку. У некоторых крупных турбеллярии (у пресноводных планарий отр. Tricladida и у морских турбеллярии отр. Polycladida) ротовое отверстие открывается не в глотку, а в особое глубокое впячивание наружных покровов, называемое глоточным карманом (рис. 124 , рис. 135). Со дна кармана в его полость вдается мускулистая глотка. Она имеет вид трубки, которая может сильно вытягиваться и высовываться через рот наружу, служа для захвата добычи.
124 , рис. 135). Со дна кармана в его полость вдается мускулистая глотка. Она имеет вид трубки, которая может сильно вытягиваться и высовываться через рот наружу, служа для захвата добычи.
Энтодермальная средняя кишка устроена у многих форм Turbellaria различно. У мелких турбеллярий (отр. Rhabdocoela, Macrostomida и др.) она имеет вид простого мешка или слепо замкнутой трубки (рис. 134). У крупных же форм кишечник, как правило, разветвлен. Так, у многоветвистых (Polycladida) глотка ведет в желудок, от которого во все стороны к краям тела отходят ветвящиеся каналы, слепо замкнутые на концах (рис. 135). У трехветвистых (Tricladida) от глотки, находящейся вблизи средины тела, отходят сразу три главные ветви кишечника (рис. 123 , рис. 124): одна идет прямо вперед, другие две загибаются и по бокам глотки направляются назад; каждая из ветвей дает побочные слепые веточки.
Такое строение кишечника не случайно. У мелких турбеллярий, размер которых не превышает нескольких миллиметров, продукты пищеварения легко распределяются по всему телу через рыхлую паренхиматозную ткань. У крупных же трикладид и поликладид, иногда достигающих 1-3 и даже 30 см, этот процесс чрезвычайно затруднен. У таких турбеллярий функцию распределения продуктов пищеварения по телу животного выполняют ветви кишечника, пронизывающие паренхиму во всех направлениях и непосредственно соприкасающиеся со всеми тканями и органами.
У крупных же трикладид и поликладид, иногда достигающих 1-3 и даже 30 см, этот процесс чрезвычайно затруднен. У таких турбеллярий функцию распределения продуктов пищеварения по телу животного выполняют ветви кишечника, пронизывающие паренхиму во всех направлениях и непосредственно соприкасающиеся со всеми тканями и органами.
В процессе переваривания пищи у турбеллярий, так же как у кишечнополостных, большое место занимает внутриклеточное пищеварение. Частицы пищи, предварительно подвергающиеся обработке секретом глоточных желез, поступают в кишечник и захватываются клетками кишечного эпителия, в которых формируются многочисленные пищеварительные вакуоли. У отряда бескишечных турбеллярий выраженная средняя кишка вообще отсутствует (рис. 126). Глотка у них вдается в несколько обособленный участок паренхимы (пищеварительная паренхима), в которой происходит внутриклеточное переваривание пищи
Пищеварительная система ресничных червей довольно разнообразна в пределах класса и варьирует от примитивной — без оформленного кишечника до сравнительно сложной — с ветвистым кишечником. Основные отряды ресничных червей отличаются прежде всего формой кишечника.
Основные отряды ресничных червей отличаются прежде всего формой кишечника.
Довольно сложный кишечник имеется у многоветвистых турбеллярий — поликлад (отряд Polycladida). Ротовое отверстие у них, как правило, ближе к заднему концу тела и ведет в складчатую глотку, от которой радиально расходятся множество слепых ветвей энтодермального кишечника.
В подотряде трехветвистых, или планарий — триклад (Tricladida ) от глотки, расположенной посередине тела, отходят три ветви кишечника. Одна ветвь направлена к переднему концу тела, а две — назад. При этом пища сначала поступает в передний вырост, а оттуда перетекает в задние. Таким образом, питательными веществами первоначально обеспечиваются органы, расположенные в передней части тела.
У отряда прямокишечных (Rhabdocoela) глотка расположена на переднем конце тела и кишечник прямой, неветвистый.
У ресничных червей с оформленным кишечником большую роль в переваривании пищи выполняют глоточные железы. У многих видов наблюдается внекишечное пищеварение.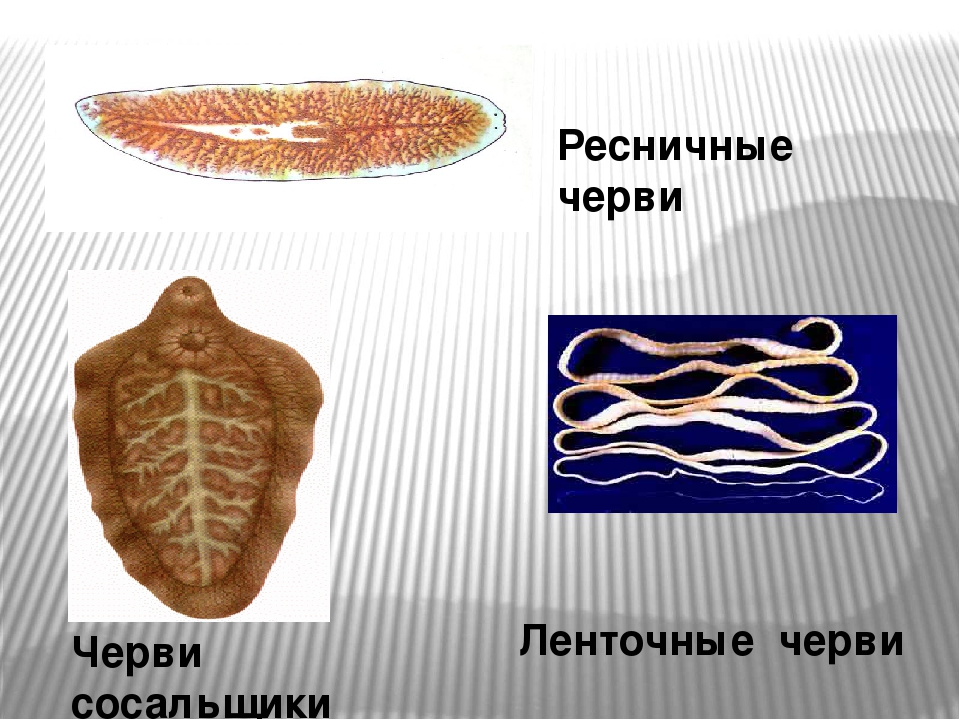 Нередко планарии нападают на довольно крупные жертвы (моллюсков, рачков). В тело жертвы они выделяют пищеварительные ферменты, а затем всасывают полупереваренную пищу.
Нередко планарии нападают на довольно крупные жертвы (моллюсков, рачков). В тело жертвы они выделяют пищеварительные ферменты, а затем всасывают полупереваренную пищу.
Несмотря на определенную сложность пищеварительной системы, усвоение пищи у турбеллярий в значительной мере осуществляется за счет внутриклеточного пищеварения, то естьфагоцитоза эпителиальных клеток средней кишки.
Большинство турбеллярий являются хищниками и питаются различными мелкими беспозвоночными. Обнаружив жертву, червь накрывает ее своим телом, после чего заглатывает. У планарий для этого служит выдвижная глотка, которая выбрасывается из глубокого впячивания наружного покрова — глоточного кармана. Если размер добычи слишком велик и ее невозможно проглотить целиком, червь отрывает куски сильными сосательными движениями глотки, после чего заглатывает их. Однако мелкие членистоногие (например, рачки) покрыты жестким панцирем и разорвать их планария не может. В таком случае она выделяет из глотки наружу пищеварительные ферменты, которые расщепляют ткани жертвы вне организма червя, после чего размягченная пища всасывается глоткой. Таким образом, у турбеллярий возможно и наружное пищеварение. Некоторые турбеллярии (планарии) обладают своеобразной способностью использовать «трофейное оружие». Ученые установили, что при поедании гидр планарией стрекательные клетки не разрушаются, а мигрируют в покровы червя и защищают его.
Таким образом, у турбеллярий возможно и наружное пищеварение. Некоторые турбеллярии (планарии) обладают своеобразной способностью использовать «трофейное оружие». Ученые установили, что при поедании гидр планарией стрекательные клетки не разрушаются, а мигрируют в покровы червя и защищают его.
Признаками класс ресничные черви имеют свои специфические особенности, которые хорошо выражены у планарий и достаточно подробно описаны в школьном учебнике зоологии, где приведены схематические рисунки внешнего и внутреннего строения белой планарии.
Планарии представляют собой доступный объект для постановки, ряда наблюдений и опытов в уголках живой природы, позволяя учителю существенно дополнить, уточнить и расширить сведения учащихся о классе ресничных червей, полученные ими на уроках.
В наших пресных водоемах можно добыть планарий, которые принадлежат к разным видам и отличаются друг от друга формой головы, количеством и расположением глазков, величиной тела. Обычно их находят вблизи берега под камнями, под опавшими в воду листьями, на стеблях подводных растений.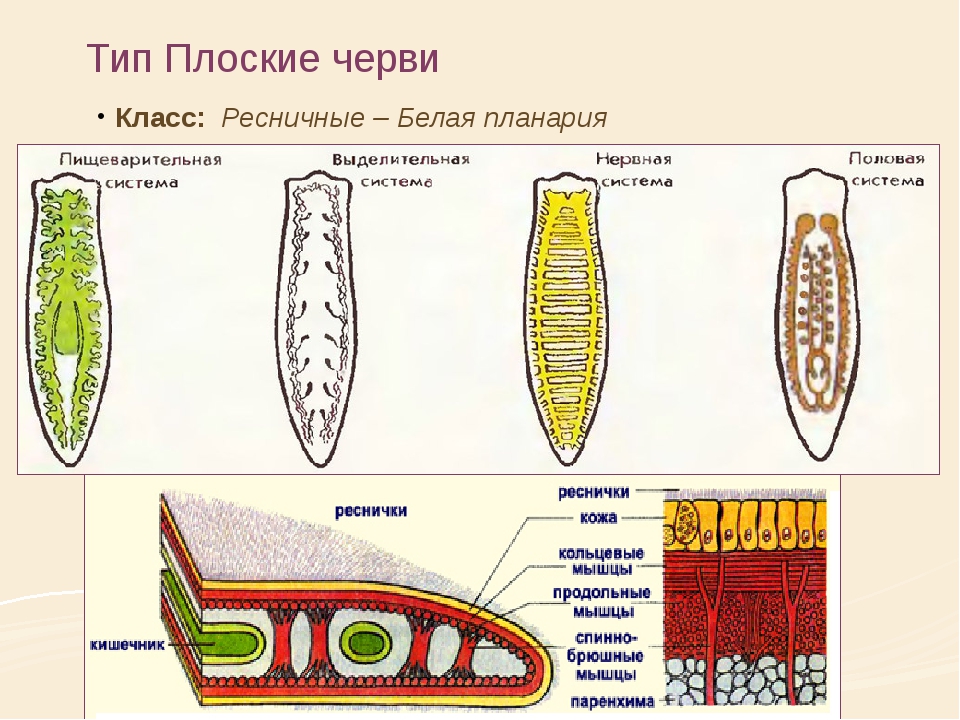
Белая, или молочная, планария. Это — наиболее крупная форма с просвечивающимися покровами. За мутно-белый цвет тела ее называют молочной планарией. Ее легко содержать в аквариуме (в плоском сосуде), но надо часто менять воду. Кормить можно водяными осликами, предварительно отрезая у них конечности. Молочная планария живет в неволе долго и даже откладывает яйца.
На часовом или предметном стекле в капле при малом увеличении микроскопа или в лупу хорошо видны следующие детали: два глазка на переднем конце тела в виде черных пятен, ротовое отверстие и глотка на брюшной стороне (позади середины тела), трехветвистый кишечник, просвечивающий сквозь стенки тела (два канала, соединяющихся в один ствол). Реснички червя увидеть трудно. В лучшем случае при рассматривании под сильным увеличением микроскопа у какой-либо мелкой планарии, накрытой покровным стеклом, можно обнаружить по краю тела ресничный эпителий в виде нежной пленки.
Мышечная система ресничных червей
Наблюдая за движениями планарий в аквариуме, учащиеся должны обратить внимание на ее плавное скольжение по стеклу. Причина заключается в следующем. Кожные железы ресничного червя выделяют слизь, которая создает благоприятные условия для работы ресничек. Их согласованные движения перемещают тело планарии по слизистой дорожке со скоростью, которую можно измерить секундомером. Кроме того, поступательному движению способствует волнообразное сокращение продольных мышц брюшной стороны тела. Временами планария может переходить к скольжению в воде по нижней стороне пленки поверхностного натяжения подобно тому, как это делают другие водные организмы ( , актинии, ). Следует сообщить учащимся, что в коже ресничного червя наряду с железами, выделяющими слизистые вещества, есть еще особые органы — рабдиты. Это палочковидные образования, которые в воде растворяются и превращаются в клейкую слизь, покрывающую все тело червя. Рабдиты, с одной стороны, можно рассматривать как органы нападения, вернее, как ловчий аппарат, который способствует прилипанию добычи к телу планарии, а с другой стороны, как органы защиты, сковывающие движения нападающего врага.
Причина заключается в следующем. Кожные железы ресничного червя выделяют слизь, которая создает благоприятные условия для работы ресничек. Их согласованные движения перемещают тело планарии по слизистой дорожке со скоростью, которую можно измерить секундомером. Кроме того, поступательному движению способствует волнообразное сокращение продольных мышц брюшной стороны тела. Временами планария может переходить к скольжению в воде по нижней стороне пленки поверхностного натяжения подобно тому, как это делают другие водные организмы ( , актинии, ). Следует сообщить учащимся, что в коже ресничного червя наряду с железами, выделяющими слизистые вещества, есть еще особые органы — рабдиты. Это палочковидные образования, которые в воде растворяются и превращаются в клейкую слизь, покрывающую все тело червя. Рабдиты, с одной стороны, можно рассматривать как органы нападения, вернее, как ловчий аппарат, который способствует прилипанию добычи к телу планарии, а с другой стороны, как органы защиты, сковывающие движения нападающего врага.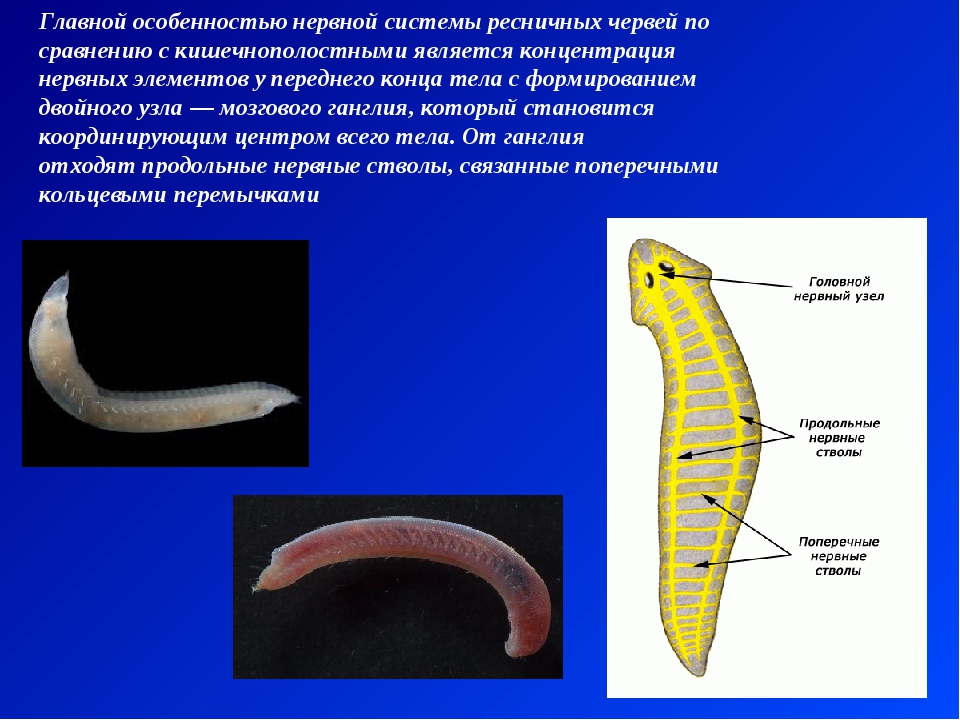 Интересно отметить защитную роль нематоцистов кишечнополостных в случае попадания их в организм планарий. Если какая-либо планария съест несколько гидр, то в ее коже можно обнаружить стрекательные клетки, занявшие место в эктодерме среди других клеток червя и продолжающие здесь свойственную им деятельность.
Интересно отметить защитную роль нематоцистов кишечнополостных в случае попадания их в организм планарий. Если какая-либо планария съест несколько гидр, то в ее коже можно обнаружить стрекательные клетки, занявшие место в эктодерме среди других клеток червя и продолжающие здесь свойственную им деятельность.
Эктодерма ресничных червей содержит нервные окончания, которые воспринимают химические, термические и механические раздражения, передавая их в виде возбуждения в центральную нервную систему (в стволы и головной ганглий). Передняя часть планарии особенно чувствительна к прикосновениям, так как здесь имеются разветвления осязательных нервных окончаний. К глазам подходят зрительные нервы, при посредстве которых планария воспринимает свет. Все это указывает на более высокую организацию планарий по сравнению с кишечнополостными и определяет их более сложное поведение.
В естественных условиях ресничные черви живут под камнями или прячутся под другими предметами, находясь в темноте, поэтому на свет у них существует определенная реакция. Если в аквариуме создать две зоны — освещенную и затемненную, то легко наблюдать переползание планарий в темную часть, т. е. демонстрировать отрицательный фототаксис. Отсюда можно сделать вывод, что ресничные черви не только отличают свет от тени, но и направление света, от которого они уклоняются. Однако примитивное строение глаз не позволяет планарии видеть предмет.
Если в аквариуме создать две зоны — освещенную и затемненную, то легко наблюдать переползание планарий в темную часть, т. е. демонстрировать отрицательный фототаксис. Отсюда можно сделать вывод, что ресничные черви не только отличают свет от тени, но и направление света, от которого они уклоняются. Однако примитивное строение глаз не позволяет планарии видеть предмет.
Питание ресничных червей
Интересно пищевое поведение ресничных червей, основанное на восприятии химических раздражений. Так, например, если поместить в определенное место аквариума пищу, то прежде всего планария покинет свое убежище и начнет перемещаться типичным для нее скольжением, делая при этом поисковые движения покачиванием головы из стороны в сторону, как бы ловя запах, исходящий от добычи. Двустороннее расположение обонятельных нервов помогает ей определить нужное направление и правильно выбрать путь, ведущий к пище. Если, например, добыча находится слева, то нервные окончания левой стороны головы получат более сильное раздражение, чем правая, и планария поползет влево, т. е. туда, где находится пища.
е. туда, где находится пища.
Нервная система ресничных червей
Велика роль нервной системы в двигательных функциях ресничного червя. Если его обезглавить, червь становится неподвижным. Это указывает на роль головного ганглия, от которого организм получает информацию. Кроме того, способность к движениям находится в зависимости от целости нервных стволов. Если перерезать тело планарии, но оставить неповрежденным нервный тяж, планария будет двигаться; в противном случае наступит паралич задней части тела, лишенной связи с передней. Отсюда следует, что концентрация нервных клеток на переднем конце тела планарии в виде ганглия образует нечто подобное мозгу, который возбуждает активность мускулатуры только в том случае, если сигналы от воспринимающих участков поступают по нервным стволам к рабочим органам. Здесь важно подчеркнуть появление у плоских червей по сравнению с кишечнополостными прогрессивного усложнения нервной системы. Концентрация нервных на переднем конце тела повысила его значение в осуществлении контроля над другими подчиненными ему участками.
Воздействие различных факторов внешней среды на организм планарий вызывает у них стереотипную реакцию двух типов: пассивную и активную. В качестве пассивной реакции можно назвать переход к неподвижности в темноте как ответ на прекращение освещения или сокращение тела при раздражении как пассивно-оборонительный рефлекс. Активные реакции выражаются в передвижении планарий к источнику раздражения (например, к пище) или от него (например, от лучей света), а также обильное выделение слизи как активно-оборонительный рефлекс в ответ на прикосновение.
У планарий можно выработать временную связь типа условного рефлекса, однако эта связь не отличается большой стабильностью. Так, например, в качестве условного раздражителя в одном из опытов был использован свет, а в качестве безусловного применен раствор хлорида натрия. Эта соль вызывает оборонительную реакцию планарии в виде сокращения мускулатуры. Если через 5 с после воздействия света раздражать планарию поваренной солью, то через 30-40 сочетаний у нее начнет формироваться условный рефлекс на свет, который через 120-170 сочетаний дает от 40 до 80% положительных ответов. Однако в дальнейшем полученная временная связь ослабевает, несмотря на продолжение эксперимента.
Однако в дальнейшем полученная временная связь ослабевает, несмотря на продолжение эксперимента.
Регенерация ресничных червей
В уголке живой природы вполне возможно поручить учащимся постановку опытов с регенерацией планарий. Для этого планарию разрезают на куски на любом уровне, и в каждом фрагменте голова регенерирует из переднего конца, а хвост — из заднего. Известно, что способность к регенерации у ресничных червей выражена в очень высокой степени. Даже из тела планарии можно восстановить целый организм.
С регенерацией у планарии тесно связано явление автотомии (самокалечения), которое имеет большое значение в жизни ресничных червей. При наступлении неблагоприятных условий (например, при резком снижении кислорода в воде или при сильном прогревании водоема) планарии распадаются на части, которые при этом не погибают. Когда среда нормализуется, каждая часть планарии регенерирует в целый организм. Некоторые виды планарий этим способом размножаются в обычных условиях, что можно рассматривать как одну из разновидностей бесполого размножения, дополняющего половое.
Можно начать их описание с плана организации, и прежде всего перечислить их морфологические признаки. Это, несомненно, важно. Однако не менее важно понимать причины, вызвавшие к жизни ту или иную организацию живых существ. Итак, первый и главнейший признак ресничных червей — это тот, что они свободноживущие организмы. Вот что их определяет! А если это так, то для них должны быть типичны признаки, присущие свободным организмам: это прежде всего органы свободного передвижения в пространстве, способность определять в нем положение собственного тела и, наконец, способность вовремя заметить других животных — и врагов, и добычу — и вовремя среагировать на тех и на других. Для ресничных червей прежде всего характерны следующие признаки. Тело их покрыто ресничками, согласованное движение которых обеспечивает плавное передвижение в пространстве. У ресничных червей имеется особый орган равновесия — статоцист
— в, виде пузырька с плотным свободным ядрышком внутри, подобный тем, кд- , кие имеются у кишечнополостных; этот орган позволяет червю ориентироваться в пространстве. Переверните ресничного червя, и он тотчас повернется брюшной стороной вниз. Информацию о своем положении в пространстве ресничный плоский, червь получил от статоциста. Ресничным червям присущи развитые opганы обоняния (химические органы) и органы восприятия света (фоторецепторы). Химические органы представлены обонятельными ямками по бокам головы, тогда как фоторецепторы представлены глазками, расположенными на переднем крае тела. Наконец, отметим еще один признак свободного существования ресничных червей, а именно покровы их тела окрашены в различные цвета — зеленый, желтый, розовый, светло- и темно-коричневый, почти черный, красный, фиолетовый, синевато-серый и др. Таковы главные признаки ресничных червей, определяемые их свободной жизнью. На таблице 17 показаны некоторые представители ресничных червей. Кто скажет, что они некрасивы, по крайней мере по окраске?
Переверните ресничного червя, и он тотчас повернется брюшной стороной вниз. Информацию о своем положении в пространстве ресничный плоский, червь получил от статоциста. Ресничным червям присущи развитые opганы обоняния (химические органы) и органы восприятия света (фоторецепторы). Химические органы представлены обонятельными ямками по бокам головы, тогда как фоторецепторы представлены глазками, расположенными на переднем крае тела. Наконец, отметим еще один признак свободного существования ресничных червей, а именно покровы их тела окрашены в различные цвета — зеленый, желтый, розовый, светло- и темно-коричневый, почти черный, красный, фиолетовый, синевато-серый и др. Таковы главные признаки ресничных червей, определяемые их свободной жизнью. На таблице 17 показаны некоторые представители ресничных червей. Кто скажет, что они некрасивы, по крайней мере по окраске?
В остальном ресничным червям свойственны признаки плоских червей: кожно-мускульный мешок, состоящий из кожных покровов и сложной системы продольных, кольцевых, косых и спинно-брюшных мышц; паренхима, заполняющая тело; продольные нервные стволы; брюшное ротовое отверстие; слепая кишка — прямая или разветвленная на два ствола; выделительные органы в виде ветвящихся трубочек с мерцательными колбочками на концах и экскреторным наружным отверстием на заднем конце тела; мощно развитая половая система, в которой комбинируются всегда мужские и женские половые органы. Все это как бы вправлено в общий «архитектонический план» двубоко-вой симметрии (рис. 192).
Все это как бы вправлено в общий «архитектонический план» двубоко-вой симметрии (рис. 192).
Ресничные черви — хищники. Они нападают на мелких животных, например мелких рачков, и высасывают их, или даже рвут нежное тельце рачков на части, или глотают целиком. Живут они в различных средах. Известны морские ресничные черви, пресноводные и, наконец, почвенные.
Отряд бескишечные ресничные черви (Acoela)
У берегов Бретани (Франция) начинается отлив. Отступает море. Обнажается морское дно. И на обнаженном дне появляются зеленые пятна. Они как живые. Постепенно пятна становятся темнее и изменяют свою форму. И действительно, это скопления живых существ, бесчисленных маленьких турбеллярий Convoluta roskoffensis, относящихся к роду конволют
(Convoluta). Зеленая окраска животных вызвана тем, что в их паренхиме живут зеленые водоросли — зоохлореллы
, принадлежащие к жгутиковым простейшим. Как только начнется прилив, конволюты зароются в песок, уходя от напора воды.
Этот ритм чередующегося зарывания в песок и появления на его поверхности конволют можно наблюдать и в аквариуме.
Как у всех бескишечных турбеллярий, у этого червя нет кишечника. Вместо него в паренхиме развита нежная плазматическая ткань с многочисленными ядрами в ней, но без клеточных границ. Ткань, в которой сливаются в общий комплекс клеточные элементы, лишённые клеточных границ, называют синцитием
* . Молодые конволюты способны питаться; они захватывают пищу при помощи ресничек, которыми усажено ротовое отверстие. Пищевые комочки попадают в синцитий, и здесь происходит процесс внутриклеточного пищеварения, столь характерный для всех турбеллярий, в том числе и имеющих кишечник. Однако взрослые конволюты питаются иначе. Они питаются веществами, ассимилируемыми водорослями, которые живут в их теле. Водоросли (зоохлореллы) проникают в организм конволют тогда, когда они еще только развиваются из яйца. Взрослые конволюты откладывают яйца в кокон. Эти коконы выделяют вещества, химически воздействующие на зоохлорелл и привлекающие их. Можно сказать, что яйца конволют буквально инфицированы (заражены) зоохлореллами. Но это заражение — источник жизни конволют. Из развивающихся яиц формируется молодая конволюта, и она оказывается носительницей зоохлорелл, которые размножаются в ее теле и как бы входят в его состав, обеспечивая процессы ассимиляции органических веществ и энергии солнечного света. Так зоохлореллы снабжают конволюту питанием. С другой стороны, зоохлореллы зависят от конволют и не могут жить без них. И конволюты ведут себя так, что существование зоохлорелл обеспечено. Зоохлореллы зеленого цвета. Они носители хлоропластов и нуждаются в солнечном свете. Конволюты в течение всего дня неподвижно сидят на месте, облитые яркими лучами солнца — источником жизни и энергии зоохлорелл, но вместе с тем и источником питания самих конволют. Конволюты полезны для зоохлорелл, а эти последние жизненно необходимы для конволют. Перед нами типичный пример симбиоза
— обоюдополезного сожительства свободных организмов.
Можно сказать, что яйца конволют буквально инфицированы (заражены) зоохлореллами. Но это заражение — источник жизни конволют. Из развивающихся яиц формируется молодая конволюта, и она оказывается носительницей зоохлорелл, которые размножаются в ее теле и как бы входят в его состав, обеспечивая процессы ассимиляции органических веществ и энергии солнечного света. Так зоохлореллы снабжают конволюту питанием. С другой стороны, зоохлореллы зависят от конволют и не могут жить без них. И конволюты ведут себя так, что существование зоохлорелл обеспечено. Зоохлореллы зеленого цвета. Они носители хлоропластов и нуждаются в солнечном свете. Конволюты в течение всего дня неподвижно сидят на месте, облитые яркими лучами солнца — источником жизни и энергии зоохлорелл, но вместе с тем и источником питания самих конволют. Конволюты полезны для зоохлорелл, а эти последние жизненно необходимы для конволют. Перед нами типичный пример симбиоза
— обоюдополезного сожительства свободных организмов.
Как уже говорилось, у бескишечных турбеллярий нет кишечника. На брюшной стороне тела конволют расположена воронка, ведущая в ротовое отверстие, а оно в свою очередь ведет в синцитий. Кпереди от ротового отверстия расположен маленький пузырек, просвечивающий сквозь покровы тела. Это статоцист
. Известный специалист по турбелляриям Бресла у так разъясняет функции статоциста: если конволют, содержащихся в сосуде, оставить в покое, все они соберутся на поверхности воды. Однако даже легкое сотрясение заставляет их опускаться на дно. Эти реакции обусловлены наличием статоциста. «Реакции,- пишет Бреслау,- исчезают, если животное обезглавлено или если соленая морская вода, в которой находится червь, быстро заменяется пресной. В последнем случае происходит повреждение тонкой структуры статоциста». Итак, статоцист, т. е. орган равновесия, определяет и упоминавшуюся выше реакцию конволют на приливы и отливы. На рисунке 194 показан другой вид конволюты — Convoluta convoluta, в теле которой видны зоохлореллы, а также округлые яйца. В отличие от ранее названного вида, эта конволюта питается не только за счет зоохлорелл, но и через ротовое отверстие.
На брюшной стороне тела конволют расположена воронка, ведущая в ротовое отверстие, а оно в свою очередь ведет в синцитий. Кпереди от ротового отверстия расположен маленький пузырек, просвечивающий сквозь покровы тела. Это статоцист
. Известный специалист по турбелляриям Бресла у так разъясняет функции статоциста: если конволют, содержащихся в сосуде, оставить в покое, все они соберутся на поверхности воды. Однако даже легкое сотрясение заставляет их опускаться на дно. Эти реакции обусловлены наличием статоциста. «Реакции,- пишет Бреслау,- исчезают, если животное обезглавлено или если соленая морская вода, в которой находится червь, быстро заменяется пресной. В последнем случае происходит повреждение тонкой структуры статоциста». Итак, статоцист, т. е. орган равновесия, определяет и упоминавшуюся выше реакцию конволют на приливы и отливы. На рисунке 194 показан другой вид конволюты — Convoluta convoluta, в теле которой видны зоохлореллы, а также округлые яйца. В отличие от ранее названного вида, эта конволюта питается не только за счет зоохлорелл, но и через ротовое отверстие.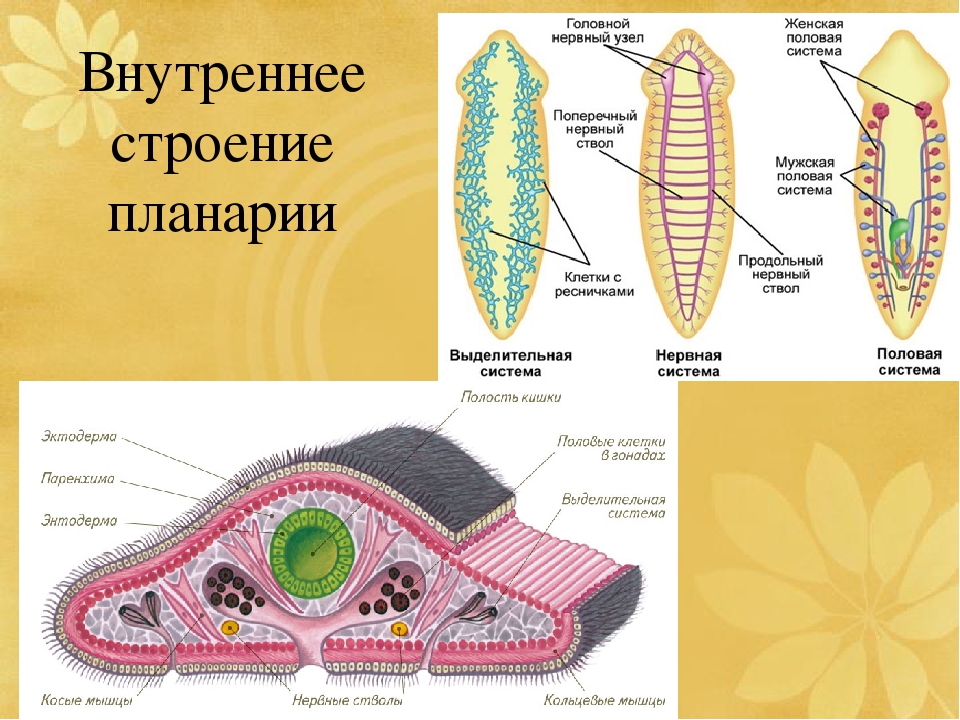
* (Синцитий — от греческих слов syn вместе и cytos — клетка. )
В заключение отметим, что бескишечные турбеллярий , или ацелы , различно трактуются специалистами. В системе их обычно ставят на первое место как наиболее примитивных турбеллярий. Однако некоторые специалисты считают, что ацелы скорее всего упрощенные формы. Известный специалист по турбелляриям Отто Штейнбок (1958) считает, что бескишечные турбеллярий образуют примитивную ветвь, от которой берут свое начало высшие турбеллярий. К ним мы и обратимся.
Отряд прямокишечные турбеллярий (Rhabdocoela)
И этот отряд представлен мелкими животными. Однако, в отличие от ацел , у прямокишечных турбеллярий имеется кишечник в виде прямой трубки, за что отряд и получил свое название. Ротовое отверстие, расположенное всегда на брюшной стороне, ведет в мускулистую глотку. Ротовое отверстие лежит то посередине брюшной стороны тела, то ближе к переднему или заднему концу его.
У прямокишечных турбеллярий имеются фоторецепторы
, т. е. органы, воспринимающие свет и обычно называемые глазами. И в самом деле, эти органы пигментированы и несут светопреломляющую линзу
— своего рода хрусталик. Глаза могут, однако, и отсутствовать. Около 400 видов этого отряда живет в пресной воде, в море и частью в почве.
е. органы, воспринимающие свет и обычно называемые глазами. И в самом деле, эти органы пигментированы и несут светопреломляющую линзу
— своего рода хрусталик. Глаза могут, однако, и отсутствовать. Около 400 видов этого отряда живет в пресной воде, в море и частью в почве.
Чтобы читатель составил себе некоторое представление о формах этого отряда, мы ограничимся характеристикой некоторых видов.
Род Mesostomum — один из наиболее богатых видами. У представителей этого рода обычно плоское тело. Рот помещается примерно в середине брюшной стороны. Ротовая полость снабжена сильным сосущим аппаратом, с помощью которого эти маленькие хищники хватают своих жертв л высасывают их.
Один из красивейших видов этого рода — Mesostoma ehrenbergii — достигает в длину 1 см. Назван так в честь знаменитого зоолога, опубликовавшего в первой, половине XIX в. (Эренберг, 1795-1876) ценные исследования, посвященные микроскопическим животным. Вид этот можно обнаружить весной на затопленных половодьем лугах, а также в прудах с илистым грунтом и в водоемах, заросших тростником и ситником. Животное прозрачно, как стекло, и кажется, что оно такое же хрупкое!
Животное прозрачно, как стекло, и кажется, что оно такое же хрупкое!
Наблюдая за животным, можно заметить, что в воде оно совершает медленные и, видимо, свободные колебания, оставаясь при этом подвешенным на тонкой и невидимой нити, образовавшейся из его собственных слизистых выделений (рис. 191).
Достаточно, однако, потревожить покой животного, как тотчас турбеллярия начинает вздрагивать и изгибаться с таким же проворством, как какая-нибудь пиявка.
Мезостома
— хищник. В высшей степени интересно наблюдать, как она атакует водяную блоху
— маленького пресноводного рачка. Мезостома хватает его примерно так же, как мы ловим рукой муху. При этом мезостома стремительно загибает задний конец тела до его соприкосновения с передним концом, в то время как боковые края тела прижимаются друг к другу. Рачок оказывается в живой ловушке. Он беспомощно бьется в ней некоторое время, затем утихает, а мезостома высасывает его содержимое. Маленькая трагедия заканчивается, и хищник вытягивается как бы в знак полного удовлетворения.
Некоторые мезостомы (других видов) способны действовать, как пауки, создавая слизистую паутину, в которую попадают их жертвы.
В средней части Европы водится другой интересный представитель того же рода — Mesostomum tetragonum. Эта мезостома достигает 7-10 мм в длину и обитает в маленьких пересыхающих или чистых, но заросших водной растительностью прудах. Она коричневого цвета, с небольшим красноватым отливом. По бокам тела развиты лопасти, волнообразно изгибающиеся от переднего к заднему концу тела. С помощью, этих лопастей и ресничек, покрывающих кожу, мезостома плавает в воде.
Одной из интересных особенностей биологии прямокишечных турбеллярий надо считать их способность жить в пересыхающих мелких водоемах. Видимо, они приспособлены к перенесению сухости. Некоторые виды были констатированы в мелких водоемах и даже маленьких лужах, на недели высыхавших жарким летом. Яйца одной из таких мезостом, извлеченные из засохшего ила, и положенные затем в воду, через несколько дней стали развиваться.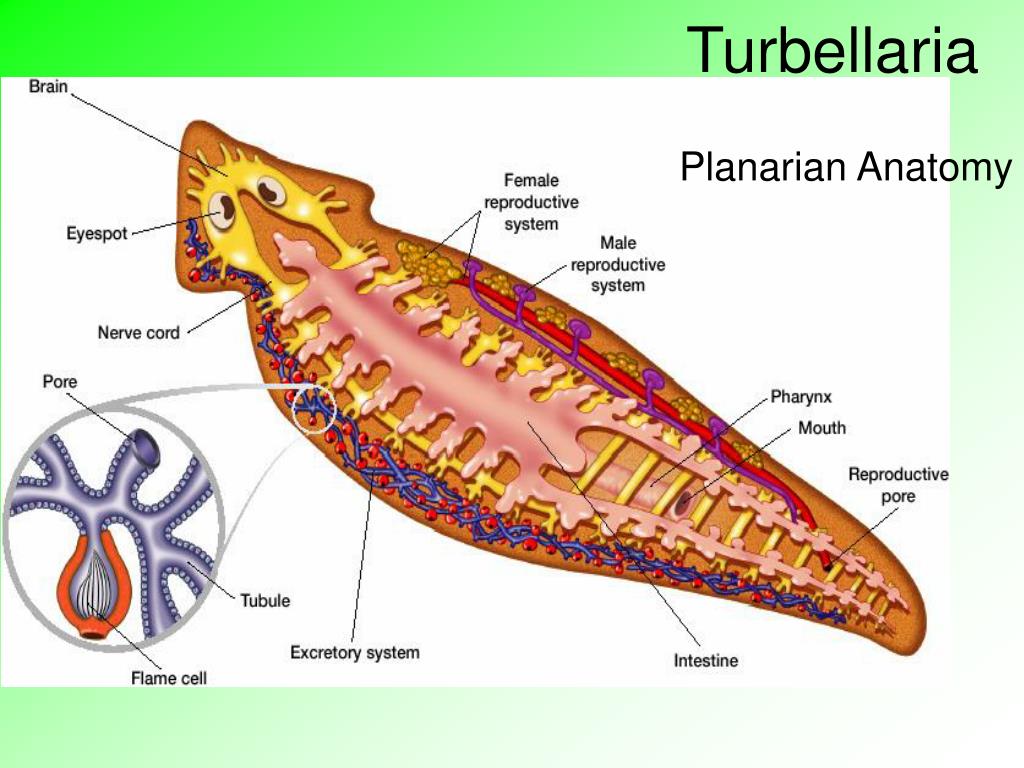
Отметим, что яйца мезостом бывают двух типов — с толстой и тонкой скорлупой. Последняя характерна для летних яиц. По-видимому, у мезостом намечается закономерная смена форм яиц: летние яйца развиваются в результате самооплодотворения мезостомы, а зимние — только в результате перекрестного оплодотворения двух особей. Яйца мезостом имеют обычно форму дисков с центральным углубление. Зимние яйца высвобождаются из тела матери только после ее смерти и могут переносить зимнюю стужу и летнюю сушь. Это приспособление имеет значение для выживания вида. Из зимних яиц рождаются зимние особи, встречающиеся ранней весной и летом. Особи зимней генерации летом откладывают яйца с тонкой скорлупкой, а в дальнейшем снова с плотной. Из летних яиц развиваются летние животные, дающие два сорта яиц — тонкоскорлуповые и толстоскорлуповые; таким образом, за зимними формами следуют летние, продуцирующие эти два сорта яиц. Когда наступают холода, из тонкоскорлуповых яиц рождаются осенние формы, развивающие зимние яйца. Из них и рождаются упомянутые выше зимние формы.
Из них и рождаются упомянутые выше зимние формы.
Все, что было сказано выше, станет нам понятнее, если мы хотя бы сжато познакомимся с морфологической организацией мезостом. Как у всех турбеллярий, тело их покрыто ресничками — органом движения. Как уже было сказано выше, кишечник их представлен прямой трубкой (рис. 195). Плотные отбросы питания выбрасываются через рот, жидкие — через специальную выделительную систему- протонефридии , представленные конечными клетками с «ресничным пламенем» и отходящими от них выделительными каналами (рис. 195).
Наиболее сложную организацию имеет половая система. Все мезостомы, да и прямокишечные турбеллярий вообще, как правило, гермафродиты , т. е. обоеполые формы.
На рисунке 195 показана половая система Mesostoma ehrenbergii. He вникая в подробности ее описания, укажем лишь на сложность организации половой системы и на совмещение в ней органов мужской и женской природы. На рисунке показан яичник, связанный с семеприемником. По бокам этой системы лежат семенники. От семенников идут два семяпровода, по которым сперматозоиды проникают в мужской совокупительный орган, имеющий у мезостомы форму опрокинутой реторты. Через «горлышко» этой «реторты» сперма попадает в так называемый общий проток
, из которого она поступает в упомянутый семеприемник, где семя сохраняется. Яйца также проникают в семеприемник и затем, уже оплодотворенные, перемещаются через общий проток в половую клоаку. В дальнейшем яйца, достигшие клоаки, покрываются здесь желтком из желточников и скорлупой. У мезостом происходит перекрестное оплодотворение, т. е. две особи взаимно оплодотворяют друг друга. Такое оплодотворение повышает жизнеспособность потомства и обеспечивает его большее, наследственно обусловленное многообразие. Все это показывает, что половые функции у прямокишечных турбеллярий достигают высокого уровня развития.
По бокам этой системы лежат семенники. От семенников идут два семяпровода, по которым сперматозоиды проникают в мужской совокупительный орган, имеющий у мезостомы форму опрокинутой реторты. Через «горлышко» этой «реторты» сперма попадает в так называемый общий проток
, из которого она поступает в упомянутый семеприемник, где семя сохраняется. Яйца также проникают в семеприемник и затем, уже оплодотворенные, перемещаются через общий проток в половую клоаку. В дальнейшем яйца, достигшие клоаки, покрываются здесь желтком из желточников и скорлупой. У мезостом происходит перекрестное оплодотворение, т. е. две особи взаимно оплодотворяют друг друга. Такое оплодотворение повышает жизнеспособность потомства и обеспечивает его большее, наследственно обусловленное многообразие. Все это показывает, что половые функции у прямокишечных турбеллярий достигают высокого уровня развития.
Следует отметить, что прямокишечным турбелляриям присуще не только половое размножение, но также и бесполое, достигаемое посредством деления особей. Вот как описывает Бреслау этот процесс у Microstomum lineare: «У микростомы деление начинается образованием поперечной перегородки, развивающейся между кожно-мускульным мешком и мышечным слоем кишечника, и притом примерно посередине тела. Последнее оказывается, таким образом, разделенным на две части, соответствующие двум будущим дочерним особям. В дальнейшем ходе процесса деления на месте возникшей перегородки тело перешнуровывается, перетяжка углубляется, так что в конце концов обе половины червя оказываются связанными только не разделившимся еще кишечником. Наконец и этот последний разделяется, и оба зооида (таково общее название образующихся подобным способом дочерних индивидуумов) оказываются свободными. Но прежде чем это произойдет, кроме описанного первого деления, подготовляются и дальнейшие. Обе дочерние особи, оставаясь еще связанными друг с другом, сами начинают делиться надвое, и из первоначально целостного животного получается цепочка из четырех зооидов, у которых, в свою очередь, развиваются поперечные перегородки, указывающие на наступление третьей и дальнейших стадий деления».
Вот как описывает Бреслау этот процесс у Microstomum lineare: «У микростомы деление начинается образованием поперечной перегородки, развивающейся между кожно-мускульным мешком и мышечным слоем кишечника, и притом примерно посередине тела. Последнее оказывается, таким образом, разделенным на две части, соответствующие двум будущим дочерним особям. В дальнейшем ходе процесса деления на месте возникшей перегородки тело перешнуровывается, перетяжка углубляется, так что в конце концов обе половины червя оказываются связанными только не разделившимся еще кишечником. Наконец и этот последний разделяется, и оба зооида (таково общее название образующихся подобным способом дочерних индивидуумов) оказываются свободными. Но прежде чем это произойдет, кроме описанного первого деления, подготовляются и дальнейшие. Обе дочерние особи, оставаясь еще связанными друг с другом, сами начинают делиться надвое, и из первоначально целостного животного получается цепочка из четырех зооидов, у которых, в свою очередь, развиваются поперечные перегородки, указывающие на наступление третьей и дальнейших стадий деления». Впрочем, обычно в естественных условиях наблюдаются цепочки из 6-8 зооидов, так как более длинные цепочки легко разрываются. «Развитие поперечных перегородок, превращающих животное в цепочку зооидов,- продолжает Бреслау,- само по себе не обеспечивает, конечно, жизнеспособности дочерних индивидуумов. Процесс деления необходимо дополняется новообразованиями, возникающими в результате регенерации и ведущими к тому, что отдельные участки делящегося червя достигают организации самостоятельных животных». Бреслау нашел, что обе формы размножения — половое и бесполое — чередуются: после неопределенного числа поколений, возникших в течение весны посредством деления, осенью происходит половая дифференцировка; с наступлением половозрелости бесполое размножение прекращается.
Впрочем, обычно в естественных условиях наблюдаются цепочки из 6-8 зооидов, так как более длинные цепочки легко разрываются. «Развитие поперечных перегородок, превращающих животное в цепочку зооидов,- продолжает Бреслау,- само по себе не обеспечивает, конечно, жизнеспособности дочерних индивидуумов. Процесс деления необходимо дополняется новообразованиями, возникающими в результате регенерации и ведущими к тому, что отдельные участки делящегося червя достигают организации самостоятельных животных». Бреслау нашел, что обе формы размножения — половое и бесполое — чередуются: после неопределенного числа поколений, возникших в течение весны посредством деления, осенью происходит половая дифференцировка; с наступлением половозрелости бесполое размножение прекращается.
Кроме упомянутых двух родов, к прямокишечным турбелляриям относится и ряд других. Среди них упомянем об интересных представителях рода Dalyellia, для которых характерны симбиотические отношения с зоохлореллами. Их присутствием в теле видов данного рода обусловлена зеленая окраска у Dalyellia viridis, особенно типичная для почвы луговых низин и для заболоченной воды.
Отряд трехветвистокишечные турбеллярий (Tricladida)
Роды и виды этого отряда обладают, несомненно, более высокой организацией. Прежде всего формы этого отряда крупнее. Так, широко распространенная молочная планария , названная так за цвет тела, достигает 15-26 мм в длину. Имеются и более крупные формы.
Расположенное на брюшной стороне тела ротовое отверстие ведет у этих форм в полость, в которой лежит выдвижная глотка, в состоянии покоя далеко оттянутая назад. Когда животное захватывает пищу, глотка наподобие хобота выдвигается наружу. Она снабжена не только собственной мускулатурой, но и собственной иннервацией. Если глотку оторвать, она продолжает двигаться и извиваться, как червь. Из глотки пища поступает в кишечник, состоящий из трех ветвей. Одна из них направлена вперед, а две — назад, справа и слева. Все они многократно делятся, и последние боковые разветвления заканчиваются слепо.
Форма тела трикладид
обычно удлиненная, даже листовидная, иногда тело расширенное в средней части или круглое, у некоторых форм лентовидное. Покровы и ткани прозрачны, но вместе с тем для трикладид достаточно характерны различные окраски, то более светлые, то более темные. Большинство форм имеет глазки, расположенные на спинной стороне переднего конца тела. Такие глазки имеют пресноводные формы и многие морские представители отряда. Однако у ряда форм имеются многочисленные мелкие глазки, распределенные по периферии переднего конца тела параллельно внешнему контуру его (рис. 191, 6). Иногда глазков нет. Глазки и органы химического чувства, также расположенные на головном конце в виде боковых ресничных ямок, иннервируются от нервных ядер «головного мозга», состоящего из скоплений нервных клеток. «Мозг» — парное образование. От него назад — справа и слева — идут нервные стволы, более мощные на брюшной стороне тела и несколько менее развитые на спинной. Из сказанного видно, что у турбеллярий развивается центральная нервная система, строго ориентированная и отличающаяся перемещением наиболее ответственных органов чувств на передний конец тела, ориентирующий животное в пространстве.
Покровы и ткани прозрачны, но вместе с тем для трикладид достаточно характерны различные окраски, то более светлые, то более темные. Большинство форм имеет глазки, расположенные на спинной стороне переднего конца тела. Такие глазки имеют пресноводные формы и многие морские представители отряда. Однако у ряда форм имеются многочисленные мелкие глазки, распределенные по периферии переднего конца тела параллельно внешнему контуру его (рис. 191, 6). Иногда глазков нет. Глазки и органы химического чувства, также расположенные на головном конце в виде боковых ресничных ямок, иннервируются от нервных ядер «головного мозга», состоящего из скоплений нервных клеток. «Мозг» — парное образование. От него назад — справа и слева — идут нервные стволы, более мощные на брюшной стороне тела и несколько менее развитые на спинной. Из сказанного видно, что у турбеллярий развивается центральная нервная система, строго ориентированная и отличающаяся перемещением наиболее ответственных органов чувств на передний конец тела, ориентирующий животное в пространстве. Большинство трикладид размножается половым путем и обладает сложной половой системой (рис. 195), включающей женские и мужские генитальные органы.
Большинство трикладид размножается половым путем и обладает сложной половой системой (рис. 195), включающей женские и мужские генитальные органы.
Наука знает свыше 500 видов этого отряда, заселяющих дно морей и пресных вод и известных также в почве. Около 100 видов трикладид живет в пресной воде. Остановимся лишь на немногих представителях этой группы турбеллярий.
Среди трикладид имеются настоящие гиганты. Байкальский представитель этого отряда Polycotylus достигает в длину 30 см.
Значительно уступает по размерам байкальскому поликотилусу другая трикладида — молочная планария (Dendrocoelum lacteum), которая достигает всего 15-26 мм в длину при поперечнике примерно в 6 мм. Головной конец тупо срезан, задний округлен. Тотчас позади переднего края тела расположены черные глаза. Под основанием переднего края развита присасывательная бороздка. Как и другие виды планарий, молочная планария прячется под камнями, среди листьев тростника или на нижней стороне листьев водяной лилии. Этот вид особенно хорош для изучения строения кишечника. В падающем свете он кажется почти черным, в проходящем — несколько светлее.
Этот вид особенно хорош для изучения строения кишечника. В падающем свете он кажется почти черным, в проходящем — несколько светлее.
Чаще темно-коричневая Euplanaria gonocephala имеет треугольный головной конец тела, по бокам которого расположены несколько выступающие углы, известные под названием ушей. Другая планария — Euplanaria polychroa — характеризуется индивидуальной изменчивостью окраски: наблюдаются особи коричневого цвета или темно-коричневые вплоть до черного, иногда зеленые темных оттенков, известны пестрые формы — черные со светлыми пятнами. В связи с этим точное определение вида достигается лишь изучением его анатомических признаков. Голова горной планарий (Crenobia alpina) украшена головными отростками, похожими на щупальца. Эти виды известны для Европы. От упомянутых видов рогатая многоглазка (Polycelis cornuta) и черная многоглазка (Polycelis nigra) отличаются тем, что у них по переднему краю головы видны серии глазков.
Планарий — хищники.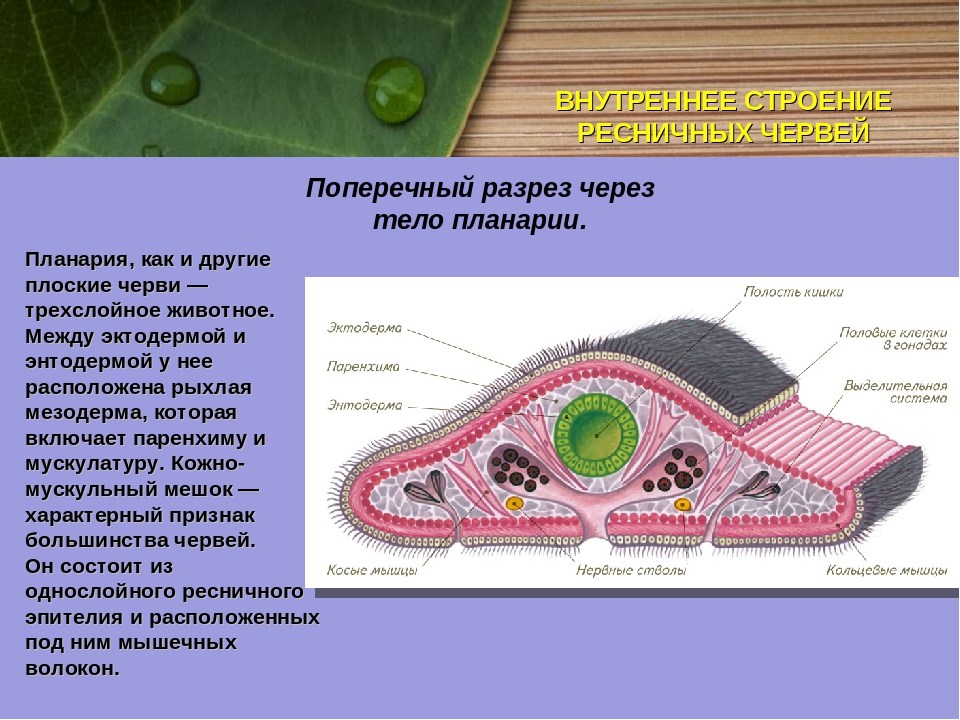 Они питаются мелкими рачками, нападают даже на улиток, на личинок некоторых насекомых, хотя не отказываются и от разлагающихся остатков других животных. У планарий сильно развито химическое чувство (обоняние). Почуяв добычу, планария направляется к ней, выдвигает свою глотку и сильными сосательными движениями рвет тело жертвы. Однако планарий выдерживают длительную голодовку и при этом «худеют», уменьшаясь в размерах, но не теряя типичных для данного вида пропорций тела.
Они питаются мелкими рачками, нападают даже на улиток, на личинок некоторых насекомых, хотя не отказываются и от разлагающихся остатков других животных. У планарий сильно развито химическое чувство (обоняние). Почуяв добычу, планария направляется к ней, выдвигает свою глотку и сильными сосательными движениями рвет тело жертвы. Однако планарий выдерживают длительную голодовку и при этом «худеют», уменьшаясь в размерах, но не теряя типичных для данного вида пропорций тела.
Яйца планарий заключены в плотную скорлупку: иногда они лежат в капсулах, сидящих на тонкой ножке, или в коконах, которые откладываются в защищенных местах. В каждой такой кладке содержится несколько десятков яйцевых и сотни желточных клеток, обеспечивающих питание развивающихся эмбрионов. Из яиц вылупляются белесые молодые формы, еще лишенные пигментов.
Замечательной чертой биологии планарий надо считать оригинальную реакцию на наступление неблагоприятных условий — повышение температуры воды, недостаток кислорода и т. п. В подобных случаях планарий способны распадаться на куски, регенерирующие при наступлении благоприятных условий в целых животных с полной и типичной организацией. Этот процесс известен под названием самокалечения
, или автотомии
. Многие формы даже в обычных условиях способны делиться на части, что можно рассматривать как особую форму размножения.
п. В подобных случаях планарий способны распадаться на куски, регенерирующие при наступлении благоприятных условий в целых животных с полной и типичной организацией. Этот процесс известен под названием самокалечения
, или автотомии
. Многие формы даже в обычных условиях способны делиться на части, что можно рассматривать как особую форму размножения.
В этой связи необходимо указать на поразительную способность планарий, да и турбеллярий вообще, к регенерации — реконструктивному восстановлению утраченных частей тела. Опыты показали, что у планарий даже 1 / 279 часть тела сохраняет способность к восстановлению полной организации со всеми присущими планариям органами. Едва ли можно сомневаться в том, что эта особенность имеет важное защитное значение, обеспечивающее сохранение жизни. Здесь уместно познакомить читателя и с другими защитными приспособлениями у турбеллярий. К ним принадлежат прежде всего кожные железы, позволяющие планарии плотно прикрепляться к поверхности, по которой она ползет. Какое же защитное значение имеют особые кожные образования турбеллярий, известные под названием рабдит
? Эти палочкообразные образования лежат в кожном эпителии и могут выбрасываться наружу, расплываясь здесь в слизь, покрывающую поверхность тела турбеллярий. Эта слизь также имеет защитное значение. Так, рабдиты выбрасываются животным из его кожи при поранениях; в этих случаях расплывающиеся в слизь рабдиты закрывают ранки, а способность к регенерации быстро обеспечивает заживление поврежденного участка тела. Интересно, что у некоторых турбеллярий (Mesostoma ehrenbergii) обнаружены стрекательные клетки в коже, совершенно сходные с такими же клетками кишечнополостных. Еще интереснее, что эти клетки, по-видимому, действительно принадлежат кишечнополостным, захваченным и съеденным турбелляриями. Известно, что планарии нападают на пресноводных гидр и охотно поедают их.
Какое же защитное значение имеют особые кожные образования турбеллярий, известные под названием рабдит
? Эти палочкообразные образования лежат в кожном эпителии и могут выбрасываться наружу, расплываясь здесь в слизь, покрывающую поверхность тела турбеллярий. Эта слизь также имеет защитное значение. Так, рабдиты выбрасываются животным из его кожи при поранениях; в этих случаях расплывающиеся в слизь рабдиты закрывают ранки, а способность к регенерации быстро обеспечивает заживление поврежденного участка тела. Интересно, что у некоторых турбеллярий (Mesostoma ehrenbergii) обнаружены стрекательные клетки в коже, совершенно сходные с такими же клетками кишечнополостных. Еще интереснее, что эти клетки, по-видимому, действительно принадлежат кишечнополостным, захваченным и съеденным турбелляриями. Известно, что планарии нападают на пресноводных гидр и охотно поедают их.
Турбеллярий живут не только в пресных водах. Известен целый ряд видов трёхветвистокишечных турбеллярий (трикладид), которые приспособлены к жизни на почве, обычно под камнями, во влажных местах. Таков, например, Rhynchodemus terrestris. Этот червь имеет цилиндрическое тело и достигает 16 мм в длину. Спинная сторона его окрашена в темно-серый цвет, тогда как брюшко окрашено в белый цвет. На переднем конце тела видны два черных глаза. Другой вид этого рода — Rh. bilineatus — попадается в цветочных горшках. Если земля в них недостаточно влажная на поверхности, то животное уползает в глубину. Как только земля увлажняется, ринходемус
вновь появляется на поверхности, ощупывая свой путь головным концом тела. Крупные экземпляры этого вида достигают 12 мм в длину. Спина у этого вида красно-коричневая с мраморным рисунком на этом фоне, причем вдоль спины видны продольные линии, окрашенные в красно-коричневый цвет. Примерно в середине тела, на спинной стороне, расположено темное пятно. Оно соответствует расположению глотки.
Таков, например, Rhynchodemus terrestris. Этот червь имеет цилиндрическое тело и достигает 16 мм в длину. Спинная сторона его окрашена в темно-серый цвет, тогда как брюшко окрашено в белый цвет. На переднем конце тела видны два черных глаза. Другой вид этого рода — Rh. bilineatus — попадается в цветочных горшках. Если земля в них недостаточно влажная на поверхности, то животное уползает в глубину. Как только земля увлажняется, ринходемус
вновь появляется на поверхности, ощупывая свой путь головным концом тела. Крупные экземпляры этого вида достигают 12 мм в длину. Спина у этого вида красно-коричневая с мраморным рисунком на этом фоне, причем вдоль спины видны продольные линии, окрашенные в красно-коричневый цвет. Примерно в середине тела, на спинной стороне, расположено темное пятно. Оно соответствует расположению глотки.
Описанные турбеллярий принадлежат к группе почвенных трёхветвистокишечных турбеллярий
(Tricladida terricola). Большая часть видов этой экологической группы приурочена к тропическим и субтропическим странам, где они обитают во влажной почве. Путешествия замечательного биолога XIX в. Чарлза Дарвина познакомили нас с богатейшей фауной почвенных планарии влажных лесов Южной Америки. Среди экзотических планарии встречаются настоящие гиганты мира ресничных червей, достигающие 60 мм в длину. Следует отметить, что у таких крупных форм, да и более мелких, но все же превосходящих скромных турбеллярий европейских стран, реснички теряют значение органов движения. Чем крупнее турбеллярия, тем большее значение в ее движениях приобретают мышцы кожно-мускульного мешка. Их волнообразные сокращения позволяют крупным турбелляриям скользить по поверхности субстрата примерно так же, как это делают улитки.
Путешествия замечательного биолога XIX в. Чарлза Дарвина познакомили нас с богатейшей фауной почвенных планарии влажных лесов Южной Америки. Среди экзотических планарии встречаются настоящие гиганты мира ресничных червей, достигающие 60 мм в длину. Следует отметить, что у таких крупных форм, да и более мелких, но все же превосходящих скромных турбеллярий европейских стран, реснички теряют значение органов движения. Чем крупнее турбеллярия, тем большее значение в ее движениях приобретают мышцы кожно-мускульного мешка. Их волнообразные сокращения позволяют крупным турбелляриям скользить по поверхности субстрата примерно так же, как это делают улитки.
Большинство упомянутых выше почвенных форм размножается половым путем. Известно около 400 видов почвенных трикладид. На острове Цейлон живет интересный представитель этой группы из рода бипалий
(Bipalium). Этот ресничный червь выделяет слизь из кожи, причем солидная капля ее вытягивается под тяжестью турбеллярий в нить, и ресничный червь оказывается подвешенным во влажном воздухе цейлонских лесов.
Один из видов бипалий — Bipalium nеwence — хороший объект для изучения механизма движений с помощью ресничек. Названная бипалия завезена чуть ли не во все страны света и встречается в оранжереях с экзотической флорой. Здесь можно наблюдать, как бипалий легко ползают по наклонным плоскостям и даже по вертикальной поверхности.
Движения обеспечиваются, с одной стороны, змеевидными изгибаниями тела, а с другой- волнообразными сокращениями мышц брюшной стороны тела и работой ребничек, покрывающих его. При этом выясняется, что животное не может использовать для движения реснички, если брюшная сторона тела («подошва») не выделяет слизи. Таким образом, работа ресничек обеспечивается наличием выделяемой из кожи слизи. Поэтому позади червя всегда виден слизистый след. Интересно, что если животное намеревается спуститься вниз, то слизистые выделения кожного покрова собираются, как уже указывалось, в комок: червь опускается отвесно вниз, а комок превращается в нить, удлиняющуюся по мере этого движения. Естественно, что для такого «путешествия» слизи нужно много. И если турбеллярий заставляли несколько раз проделать такой путь, движение задерживалось из-за недостатка слизи. Расход ее был слишком велик! Таково значение слизистых выделений кожных покровов турбеллярий в движениях этих животных.
Естественно, что для такого «путешествия» слизи нужно много. И если турбеллярий заставляли несколько раз проделать такой путь, движение задерживалось из-за недостатка слизи. Расход ее был слишком велик! Таково значение слизистых выделений кожных покровов турбеллярий в движениях этих животных.
Бипалии, как и другие турбеллярий,- хищники, способные хватать и поедать маленьких дождевых червей.
Отряд многоветвисто-кишечные турбеллярий (Polycladida)
В этот отряд входит примерно 300 видов. Все они живут в море. Это довольно крупные ресничные черви, достигающие 16 см в длину. Они имеют широкое листовидное тело, обычно окрашенное в великолепные яркие цвета. Наиболее типичный признак организации этих животных — оригинальное строение кишечника. Пищеварительная система начинается мощной глоткой, которая переходит в кишечник, имеющий форму трубки с отходящими от нее по радиусам разветвленными стволами. Разумеется, и у этих форм ануса нет, однако у некоторых форм он присутствует. Личинка поликлад покрыта ресничками и способна плавать в воде. Это так называемая мюллеровская личинка
, снабженная мускулистой глоткой и восемью лопастями. В дальнейшем она превращается во взрослую поликладу. Таким образом, для поликлад характерно развитие с превращением. К этой группе принадлежит ряд семейств, представленных многими родами и видами. У одних поликлад развита брюшная присоска, у других ее нет. К формам с брюшной присоской принадлежит, например, живущая в Северном и Средиземном морях Planocera folium, несущая два затылочных щупальца. К этой же группе поликлад относится в европейских морях и известная для фауны СССР Leptoplana tremellaris, способная плавать. Во время отлива она прячется в песке или под камнями, вновь появляясь с, наступлением прилива. Одним из наиболее интересных представителей поликлад с брюшной присоской можно считать Thysanozoon brochii, или мохнатую планарию
. Русское название обусловлено тем, что на спинной стороне этого животного (рис.
Личинка поликлад покрыта ресничками и способна плавать в воде. Это так называемая мюллеровская личинка
, снабженная мускулистой глоткой и восемью лопастями. В дальнейшем она превращается во взрослую поликладу. Таким образом, для поликлад характерно развитие с превращением. К этой группе принадлежит ряд семейств, представленных многими родами и видами. У одних поликлад развита брюшная присоска, у других ее нет. К формам с брюшной присоской принадлежит, например, живущая в Северном и Средиземном морях Planocera folium, несущая два затылочных щупальца. К этой же группе поликлад относится в европейских морях и известная для фауны СССР Leptoplana tremellaris, способная плавать. Во время отлива она прячется в песке или под камнями, вновь появляясь с, наступлением прилива. Одним из наиболее интересных представителей поликлад с брюшной присоской можно считать Thysanozoon brochii, или мохнатую планарию
. Русское название обусловлено тем, что на спинной стороне этого животного (рис. 197) вся поверхность кожи покрыта своеобразными кожными сосочками. На головном конце тела развиты две похожие на уши складки, имеющие, по-видимому, значение органов осязания. Брюшная сторона тела белая. Интересно, что в спинные сосочки проникают разветвления кишечника.
197) вся поверхность кожи покрыта своеобразными кожными сосочками. На головном конце тела развиты две похожие на уши складки, имеющие, по-видимому, значение органов осязания. Брюшная сторона тела белая. Интересно, что в спинные сосочки проникают разветвления кишечника.
Некоторое представление о многообразии и красоте окрасок поликлад читатель получит, рассмотрев цветную таблицу 17.
Фауна нашей страны богата представителями класса турбеллярий
, и притом всех четырех охарактеризованных выше отрядов. Особенно поражает фауна турбеллярий озера Байкал, которая включает 13 родов и 90 видов, в огромном большинстве относящихся к отряду трехветвистокишечных
(Tricladida). В этой группе очень резко выражен эндемизм * фауны Байкала. Все 13 родов и 90 видов — эндемики Байкала, т. е. нигде, кроме Байкала, не встречаются. Столь же резкий эндемизм свойствен байкальским губкам, олигохетам, брюхоногим моллюскам, некоторым группам рыб и ряду других обитателей Байкала. Эндемизм байкальской фауны — свидетельство большой ее древности и своеобразия этого самого глубокого озера в мире, его температурного режима и химических свойств воды.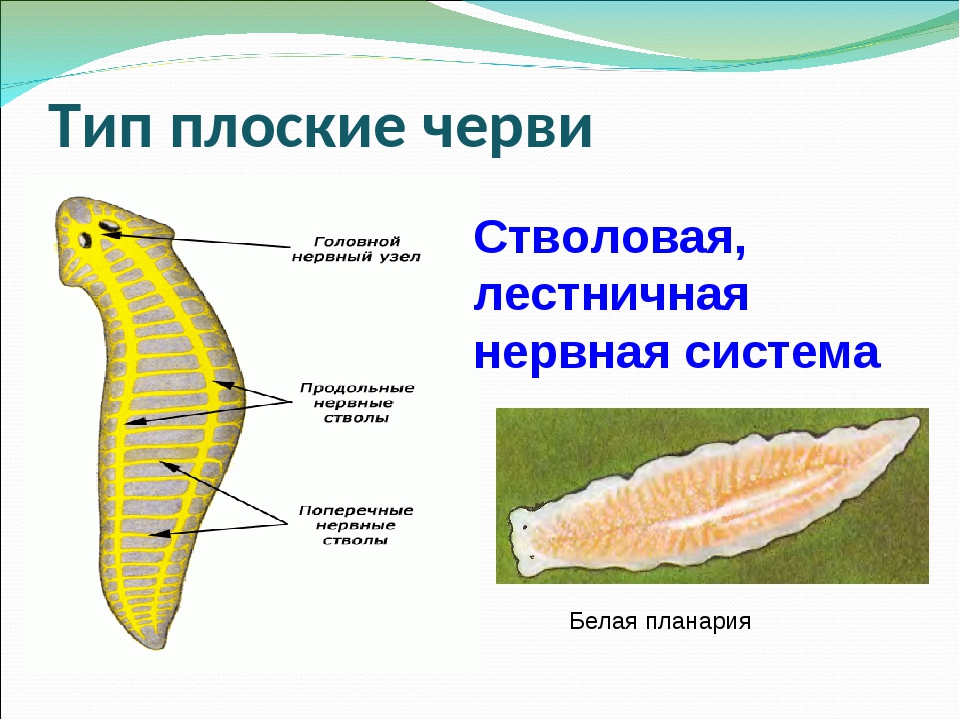
* (Эндемизм — от греческого endemos, что значит местный, постоянно живущий. )
Происхождение турбеллярий
У турбеллярий сохраняются некоторые элементы сходства с гребневиками. В частности, у бескишечных турбеллярий на одной из стадий формообразования молодого животного обнаруживаются четыре зачатка «мозга». Четыре корешка брюшных и спинных нервов наблюдаются у взрослых форм. Таким образом, у бескишечных турбеллярий обнаруживаются элементы лучевой симметрии в организации нервной системы. Известный советский специалист по турбелляриям и крупный зоолог В. Н. Беклемишев, анализируя организацию бескишечных турбеллярий, приходит к выводу, что «симметрия турбеллярий легко выводится из симметрии гребневиков». В. Н. Беклемишев, следуя в этом отношении за Графом, приходит к выводу, что и гребневики и турбеллярий, и в частности бескишечные ацелы, произошли от планулоподобных предков, т. е. форм, сходных с личинкой (планулой) кишечнополостных животных. Планула кишечнополостных — двуслойный организм, тело которого состоит из наружной эктодермы и внутренней энтодермы, образующей стенку примитивного кишечника. Тело планулы покрыто ресничками. Примерно так рисуется Графу и предок турбеллярий, отличающийся от планулоподобной формы развитием половых органов.
Тело планулы покрыто ресничками. Примерно так рисуется Графу и предок турбеллярий, отличающийся от планулоподобной формы развитием половых органов.
Класс ресничные черви (Turbellaria) | Биология животных
Общая характеристика. В большинстве случаев свободноживущие в водной среде или почве плоские черви с нерасчлененным телом, покрытым мерцательным эпителием. На спинной стороне переднего конца тела имеется обычно несколько примитивных глазков. Рот расположен у большинства видов посредине брюшной поверхности тела.
Известно около 3 тыс. видов ресничных червей, из которых несколько десятков видов ведут паразитический образ жизни. Обитают в морях, озерах и реках, но иногда также на суше. Практического значения почти не имеют, но интересны при изучении процессов регенерации, сильно развитой у них, и при решении вопроса о филогенетических взаимоотношениях классов тина плоских червей.
Строение и жизненные отправления. Форма тела ресничных червей различна (рис.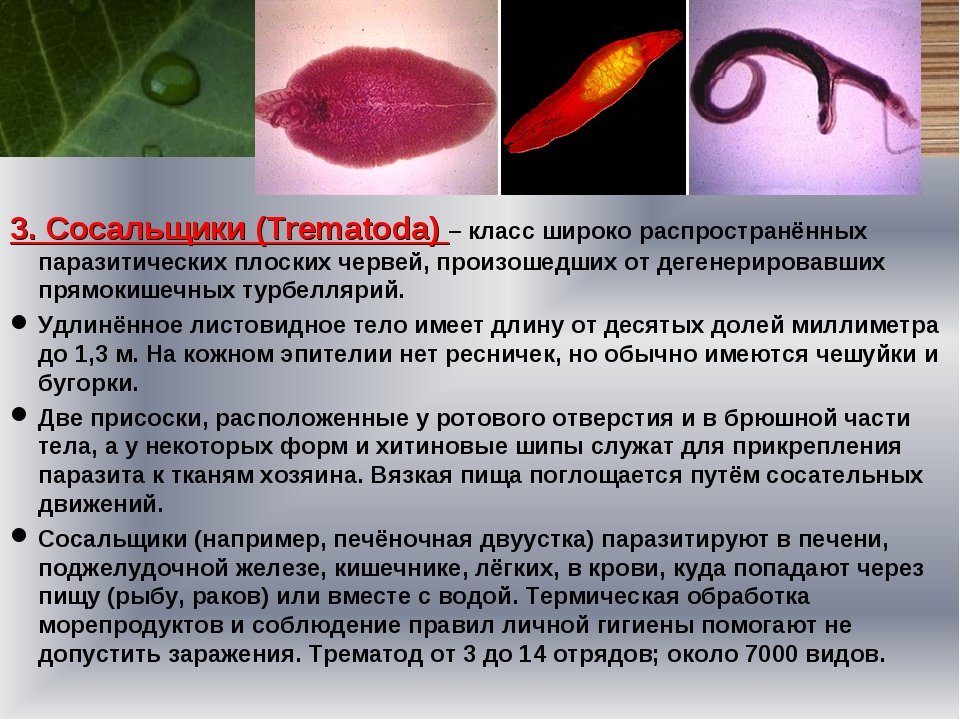 70). У некоторых по бокам тела тянутся складки в виде оторочки. Длина тела колеблется от 0,2 мм до 35 см и более (у наземных).
70). У некоторых по бокам тела тянутся складки в виде оторочки. Длина тела колеблется от 0,2 мм до 35 см и более (у наземных).
Покровы образованы ресничным (мерцательным) однослойным эпителием. Клетки его несут на своей поверхности мелкие реснички, движение которых способствует плаванию червя. Внутри клеток эпителия включены мельчайшие блестящие тельца — рабдиты, выполняющие защитную функцию, иногда служат для добывания пищи. Они периодически выбрасываются наружу и, растворяясь в воде, одевают животное рыхлой клейкой оболочкой. В покровах ресничных червей заложено много железистых клеток. Некоторые из них выделяют слизь, другие — особые ядовитые белковые вещества. Так, у молочных планарий ядовитых железок много на брюшной стороне тела. Найдя какое-либо мелкое животное, червь накрывает его своим плоским телом и убивает ядом.
Нервная система ресничных червей различна по степени сложности. У некоторых примитивных форм она диффузная. У других вдоль тела идет несколько нервных тяжей, дающих ветви к различным органам. У третьих имеются (обычно в головной части тела) скопления нервных клеток—ганглии, от которых отходят продольные нервные тяжи (рис. 71). Но, как показали исследования, интегрирующая роль этих головных ганглиев у ресничных червей невелика.
У третьих имеются (обычно в головной части тела) скопления нервных клеток—ганглии, от которых отходят продольные нервные тяжи (рис. 71). Но, как показали исследования, интегрирующая роль этих головных ганглиев у ресничных червей невелика.
Органы чувств представлены примитивными глазками, органами равновесия — статоцитами — осязательными клетками, разбросанными в покровах. Число глазков и их строение варьируют.
Органы пищеварения имеют весьма разнообразное строение. У большинства видов рот расположен в средней части брюшной стороны тела. Он ведет в объемистую глотку, способную у многих видов выпячиваться наружу, присасываться к добыче и высасывать ее. От глотки отходит кишечник, иногда — прямой, но чаще ветвящийся на две, три или много ветвей. Анальное отверстие отсутствует, и остатки пищи выбрасываются через рот. Некоторые ресничные черви лишены кишечника, и пища, поступающая через рот, попадает в рыхлую массу клеток паренхимы, которые ее поглощают и переваривают. У форм, имеющих кишечник, пища переваривается как в его просвете, так и клетками стенок, захватывающими кусочки корма. Следовательно, ресничным червям свойственно как внеклеточное, так и внутриклеточное пищеварение.
Следовательно, ресничным червям свойственно как внеклеточное, так и внутриклеточное пищеварение.
Рис. 70. Виды вей:
/-молочная планарпи; 2 — — многоглав к а
Рис. 71. Пищеварительная и пернпая системы молочной нлаиарии:
/ — рот; 2 — глотка; 3 — передняя ветвь кишечника; 4, 5 ……задние ветки кишечники; б — головной нервный узел; 7 —боковой нервный тяж; 8 – глазок
Внутриклеточное пищеварение у ресничных червей впервые было описано выдающимся русским ученым И. И. Мечниковым в 1865 г. Позднее он установил, что внутриклеточное переваривание пищи широко распространено среди многоклеточных животных. Он обнаружил, что у самых различных животных имеются особые клетки — фагоциты, способные захватывать и переваривать проникшие в их тело различные микроорганизмы (в том числе и болезнетворные). На основе этих данных И. И. Мечниковым была создана фагоцитарная теория борьбы организмов с возбудителями болезней, имеющая большое значение для медицины и ветеринарии.
Органы дыхания отсутствуют. Газообмен происходит через покровы всей поверхности тела.
Органы выделения у большинства видов ресничных червей представлены протонефридиями. У многих морских видов особых органов выделения нет или они слабо развиты и остаточные продукты обмена веществ удаляются из тела через покровы и стенки кишечника.
Органы размножения ресничных червей имеют разное строение. Большая часть этих животных — гермафродиты, но самооплодотворение устраняется разновременным созреванием яиц и сперматозоидов. Оплодотворение внутреннее.
Развитие ресничных червей происходит у морских форм обычно с превращением, а у пресноводных — без него.
Ресничным червям присуща исключительная способность регенерации. Недаром их назвали бессмертными под ножом хирурга. У некоторых видов этих животных при их разделении на 1500 частей удавалось получить из каждой части новую особь.
После открытия А. О. Ковалевским в Красном море своеобразных ползающих гребневиков (Coeloplana), имеющих уплощенное тело, ресничные покровы и ряд других признаков, свойственных ресничным червям, филогенетическая близость последних к гребневикам, а через них к кишечнополостным представляется вероятной.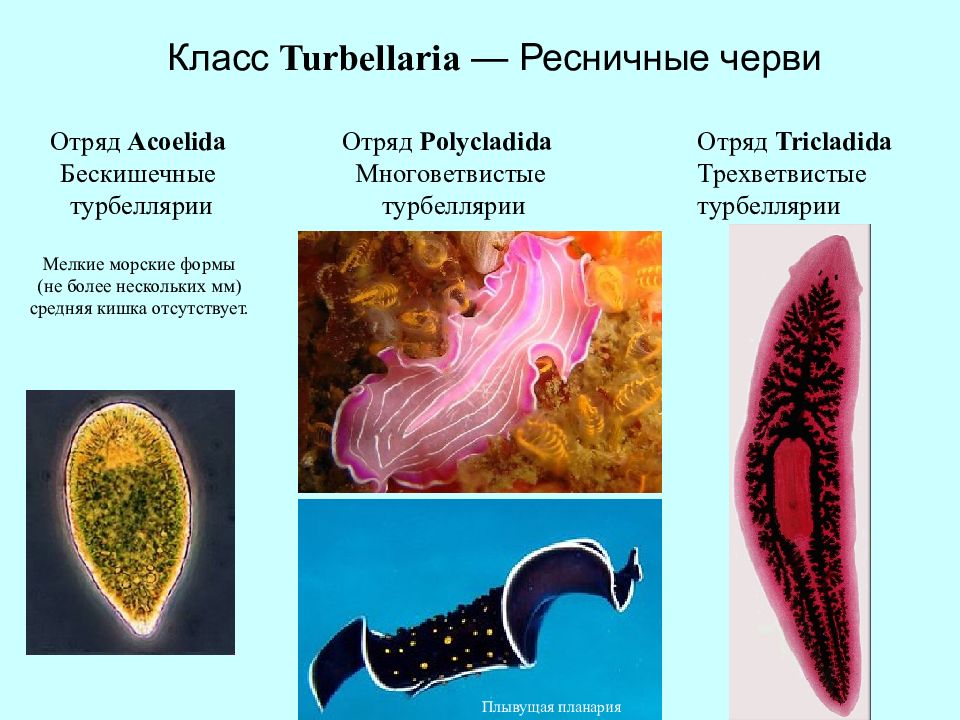
особенности строения, виды и общая характеристика
Внутреннее строение ресничных червей таблица. Плоские черви: особенности строения, виды и общая характеристика
Теория для подготовки к блоку №4 ЕГЭ по биологии: с
истема и многообразие органического мира.Тип Плоские черви
Плоские черви — тип наиболее примитивных трехслойных животных. В отличие от кишечнополостных, у них формируется третий (средний) зародышевый листок – мезодерма .
Форма тела плоских червей, как следует из названия типа, уплощённая. Они билатерально симметричны, то есть через тело можно провести только одну плоскость симметрии. Этот тип симметрии впервые появляется в ходе эволюции именно у плоских червей.
Тело не сегментировано, на переднем конце находится ротовое отверстие, которое ведёт в кишечную полость. В этом плоские черви схожи с кишечнополостными. Однако, в отличии от них, в теле плоских червей можно выделить не просто диффузно разбросанные клетки разных типов, а уже чётко сформированные ткани. Ткани образуют органы, органы составляют системы: пищеварительную, выделительную, нервную
и половую
.
Ткани образуют органы, органы составляют системы: пищеварительную, выделительную, нервную
и половую
.
Органы дыхания и кровеносная система отсутствуют. Обмен газов осуществляется непосредственно через покровы тела, поэтому плоская форма тела выгодно увеличивает площадь поверхности газообмена.
Пространство между внутренними органами и стенкой тела заполнено паренхимой – неспециализированной тканью из среднего зародышевого листка, мезодермы. Паренхима служит для запасания и транспорта веществ, поддерживает форму тела червя и служит опорой для внутренних органов.
Покровы и мускулатура
Эпителиальная и мышечная ткани обособлены, их разделяет прослойка соединительной ткани. Вместе эти три ткани образуют стенку тела червей, которая называется кожно-мускульным мешком
. Обычно наружные слои мышечных клеток являются кольцевидными, то есть при их сокращении тело червя сужается и вытягивается. Внутренние слои мышц имеют продольное расположение, с их помощью червь может укорачиваться и изгибаться в разные стороны.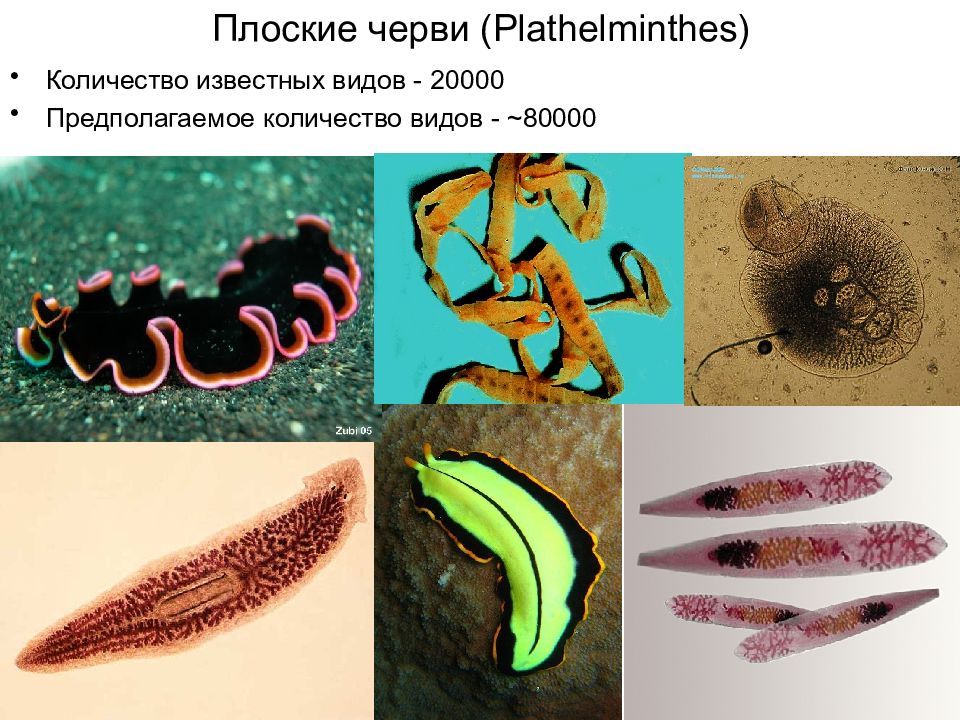 Кроме того, имеются дорзовентральные (спинно-брюшные)
пучки
мышц – они соединяют брюшную и спинную части животного. При их сокращении тело уплощается.
Кроме того, имеются дорзовентральные (спинно-брюшные)
пучки
мышц – они соединяют брюшную и спинную части животного. При их сокращении тело уплощается.
Пищеварительная система
Пищеварительная система состоит из передней кишки (глотки ), образованной эктодермой, и средней энтодермальной кишки, в которой собственно и происходит пищеварение. Задней кишки и анального отверстия нет, поэтому остатки непереваренной пищи возвращаются в окружающую среду через ротовое отверстие.
Нервная система плоских червей устроена гораздо сложнее, чем у кишечнополостных. Вот её характерные отличия:
- нервные клетки собраны в ганглии, которые в свою очередь связаны в нервные стволы;
- нервные клетки расположены глубже в теле, что позволяет их обезопасить;
- происходит цефализация , то есть ганглии, расположенные ближе к голове, играют более важную роль в управлении телом;
- олигомеризация
нервных центров, то есть уменьшение их количества по мере усложнения организма.

В переднем отделе тела располагается крупный мозговой ганглий, от которого кзади отходят два нервных ствола. Стволы соединяются поперечными перемычками, отчего такая система получила название ортогона (имеется в виду ортогональное, то есть перпендикулярное расположение нервных стволов).
Выделительная система
Продукты жизнедеятельности, часто токсичные для клеток, накапливаются в тканевой жидкости. В отличии от кишечнополостных, плоские черви не имеют возможности выделять продукты обмена непосредственно во внешнюю среду, для этого необходима отдельная система.
Выделительная система состоит из ветвящихся канальцев эктодермального происхождения – протонефридиев
. Каждый каналец заканчивается клеткой звёздчатой формы – циртоцитом
. На циртоцитах находятся пучки ресничек. При биении ресничек, напоминающих мерцательное пламя
, происходит движении тканевой жидкости в канальцы протонефридиев. Все канальцы впадают в более крупные протоки, которые открываются на поверхности тела выделительными отверстиями. Таким образом жидкость с продуктами обмена попадает наружу.
Таким образом жидкость с продуктами обмена попадает наружу.
У некоторых видов в задней части тела выделительные каналы расширяются, образуя мочевой пузырь. В нём накапливаются и концентрируются продукты обмена. С помощью выделительной системы из тела червя может также выводиться лишняя жидкость, что особенно важно для пресноводных форм. Без этого механизма пресноводные черви просто не смогли бы поддерживать водно-солевой баланс.
Половая система
Большая часть плоских червей – гермафродиты. Половые железы у них расположены в глубине тела, наружу половые клетки выводятся по протокам. Организация половой системы может значительно отличаться у представителей разных видов.
Мужские половые железы – семенники
. От них к совокупительному органу (циррусу
) идут семяпроводы. В женской половой системе есть яичники, желточники
, яйцеводы и влагалище, открывающееся в половую клоаку. Желточники схожи по строению с яичниками, но содержат желточные клетки
– стерильные яйцеклетки с большим запасом питательных веществ для будущей яйцеклетки.
Классификация
К плоским червям относят пять классов, из которых в школьном курсе рассматривают только три.
Класс Ресничные черви (Turbellaria)
В классе насчитывается более 3500 видов. В отличии от других плоских червей, большинство турбеллярий свободноживущие. Характерные представители класса – планарии (молочная, бурая, траурная, чёрная и т.д.). Они живут в пресной воде, в большом количестве водятся в стоячих и медленнотекущих водоёмах, скрываются под камнями или листьями растений. Размеры ресничных червей – от 2-3 мм до 30 см.
Тело плоское, утолщенное в середине. На переднем конце могут находиться выросты. С помощью ресничек и кожно-мускульного мешка черви могут ползать по различным поверхностям или плыть. Ротовое отверстие обычно находится в средней части тела.
В эпителии турбеллярий рассеяны одноклеточные железы, которые выделяют слизистый или белковый секрет. Слизь, вероятно, помогает перемещению и прикреплению к субстрату, служит для защиты. Белковый секрет может быть токсичен, что отпугивает других хищных животных.
Белковый секрет может быть токсичен, что отпугивает других хищных животных.
Большая часть ресничных червей – хищники. Они обладают выдвигающейся глоткой, с помощью которой можно заглатывать добычу или отрывать от неё куски. Если тело жертвы покрыто хитиновым панцирем, червь выбрасывает пищеварительные ферменты наружу и размягчает жёсткие покровы. Интересно, что планарии могут использовать «оружие» кишечнополостных: когда червь поедает гидру, её стрекательные клетки не расщепляются, а мигрируют через стенку тела, оказываясь в эпителии червя, защищая его от врагов.
Так как турбеллярии ведут активный образ жизни, у них достаточно хорошо развиты органы чувств. Всё тело покрыто специальными длинными чувствительными ресничками, сенсиллами . Они воспринимают механические или химические раздражения. Также почти у всех ресничных есть органы равновесия и два или более светочувствительных глазка , которые расположены в области головы или равномерно по краю тела.
Ресничные черви гермафродиты, оплодотворение внутреннее, чаще всего перекрёстное, то есть партнёры по очереди оплодотворяют друг друга. Сперма обычно вводится в половую клоаку, но иногда и непосредственно в тело червя (в этом случае совокупительный орган протыкает покровы партнёра). После этого сперматозоиды движутся к яйцеклеткам и оплодотворяют их.
Сперма обычно вводится в половую клоаку, но иногда и непосредственно в тело червя (в этом случае совокупительный орган протыкает покровы партнёра). После этого сперматозоиды движутся к яйцеклеткам и оплодотворяют их.
Развитие может быть прямым (из яйца выходит особь, похожая на взрослую) или с превращением (из яйца выходит личинка с ресничками).
Турбеллярии хорошо регенерируют: из небольшого куска тела может развиться полноценный взрослый организм. При наступлении неблагоприятных условий планарии имеют обыкновение распадаться на части и в таком виде пережидать длительное время. После улучшения условий из кусков регенерируют новые организмы. Это пример бесполого размножения ресничных червей.
Класс Сосальщики (Trematoda)
Нервная система образована парой головных ганглиев. Две перемычки, соединяющие ганглии, образуют окологлоточное нервное кольцо. От кольца вперёд и назад отходят нервные стволы.
Трематоды гермафродиты. У всех сосальщиков женская половая система представлена одним ветвящимся яичником, желточниками и скорлуповыми железами
. Их протоки впадают в мешкообразную полость, которая переходит в матку. Матка открывается в половую клоаку. Рядом расположен совокупительный орган, куда поступают сперматозоиды из двух семенников (редко из одного).
Их протоки впадают в мешкообразную полость, которая переходит в матку. Матка открывается в половую клоаку. Рядом расположен совокупительный орган, куда поступают сперматозоиды из двух семенников (редко из одного).
При оплодотворении семя попадает в половую клоаку, откуда сперматозоиды движутся к яйцеклеткам. Оплодотворённые яйцеклетки окружаются желточными клетками, покрываются скорлупой и начинают движение из матки наружу.
Жизненный цикл сосальщиков сложный: червь проходит несколько стадий развития со сменой хозяев. Взрослое животное (марита ), способное к половому размножению, живёт в основном хозяине – позвоночном. После оплодотворения яйца выходят во внешнюю среду и попадают в воду (чаще всего с фекалиями хозяина). В воде из яйца выходит мирацидий , личинка с ресничками.
Мирацидий активно плавает и ищет себе промежуточного хозяина, моллюска определённого вида. Например, для печёночного сосальщика промежуточный хозяин – малый прудовик
. Проникнув внутрь моллюска с помощью специального хоботка, личинка теряет реснички и становится неподвижной спороцистой
. Спороциста делится бесполым путём, в результате образуется множество личинок нового поколения. Они питаются тканями моллюска и продолжают размножаться. В итоге из моллюска выходят церкарии
– личинки с хвостиками, похожие на взрослых марит. Церкарии прикрепляются к листьям прибрежных растений и инцистируются. Циста
может длительное время ожидать, пока его не съест животное-хозяин. Человек может заразиться, если выпьет сырую воду с оторвавшимися цистами.
Спороциста делится бесполым путём, в результате образуется множество личинок нового поколения. Они питаются тканями моллюска и продолжают размножаться. В итоге из моллюска выходят церкарии
– личинки с хвостиками, похожие на взрослых марит. Церкарии прикрепляются к листьям прибрежных растений и инцистируются. Циста
может длительное время ожидать, пока его не съест животное-хозяин. Человек может заразиться, если выпьет сырую воду с оторвавшимися цистами.
Тело напоминает тонкую ленту, состоит из головки, шейки и множества члеников. Из-за членистого строения ленточных червей также называют цепнями . Длина червей может достигать 20-30 м. Такие крупные особи называются солитёрами , потому что встречаются обычно только по одиночке.
На головке расположены присоски и крючья, с помощью которых червь крепко цепляется к стенке кишечника. За шейкой следует множество члеников, каждый из которых живёт и развивается самостоятельно.
Пищеварительная система у ленточных червей полностью редуцирована: животные обитают в кишечнике и всасывают поверхностью тела пищу, обработанную ферментами хозяина.
Дыхание анаэробного типа, поэтому при окислении питательных веществ глюкоза расщепляется не полностью. Продукты неполного расщепления выводятся наружу и отравляют организм хозяина.
В каждом членике червя находятся органы выделительной и половой систем. Нервная система развита крайне слабо: по бокам проходят два нервных ствола, а в эпителии разбросаны осязательные клетки.
Ленточные черви гермафродиты. Половые органы развиваются постепенно: самые молодые членики, расположенные рядом с головкой, могут вообще не иметь их. В паренхиме формируется большое число семенников с протоками, которые сливаются в общий семяпровод. Яичник один, крупный, состоит из нескольких долек.
Возможно как перекрёстное оплодотворение, так и самооплодотворение, при котором сперматозоиды вводятся во влагалище соседнего или даже своего членика. По мере созревания яиц членик созревает и в итоге может оторваться от тела червя. Яйца попадают наружу с фекалиями хозяина и могут оседать на листьях растений.
Когда яйцо глотает промежуточный хозяин, из него выходит онкосфера , личинка с шестью крючьями. Для бычьего цепня (Taeniarhynchus saginatus) промежуточными хозяевами являются парнокопытные, для свиного цепня (Taenia solium) – свиньи, собаки, зайцы и кролики. Попадая в кишку животного, онкосфера пробуравливает её стенку и выходит в кровоток, оседая в каком-либо органе. Там личинка преобразуется в финну и дожидается, когда она попадёт в тело следующего хозяина. Обычно заражение происходит, когда основной хозяин поедает промежуточного. Человек может заразиться, употребляя в пищу плохо прожаренное мясо.
В кишечнике из финны выворачивается головка червя и закрепляется на стенке кишки. Молодые членики обособляются от шейки, тело цепня растёт.
Все черви можно поделить на три типа (плоские, кольчатые, круглые), каждый из которых отличается своими характерными особенностями. Данный тип относится к беспозвоночным животным, лишенным полости тела и обладающим двусторонней симметрией.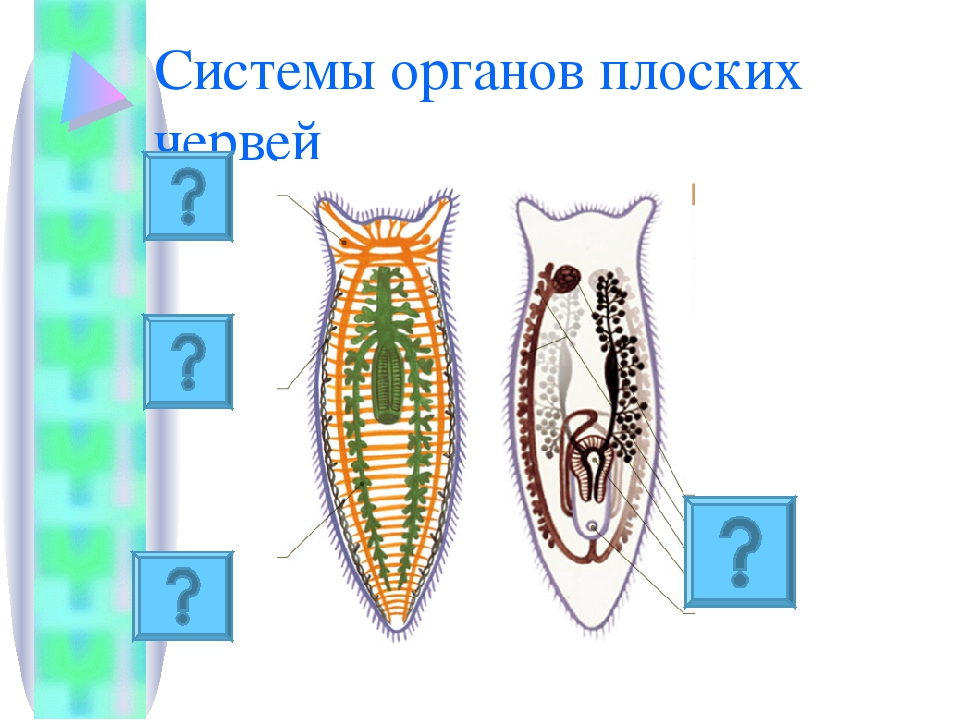
Пациенты, которые хотят избавиться от гельминтов, часто спрашивают натуральные препараты с наименьшими побочными эффектами. В таких случаях рекомендую это средство.
Основные признаки типа плоские черви
- пищеварительная;
- нервная;
- половая;
- выделительная.
Данный тип обладает наличием нескольких систем и даже зачатками органов
Кровеносная система
Не имеется, но функцию крови выполняет паренхима, состоящая из соединительных клеток. Именно она и транспортирует питательные вещества в организме.
Пищеварительная система
Довольно упрощенная, состоит из глотки и кишки.
Глотка мощная, может:
- засасывать;
- выворачиваться и обволакивать свою жертву.
Кишка из двух отделов — передний и средний, чаще всего разветвлена. Имеет замкнутое строение, так что все непереваренные отходы выходят через рот. Ротовое отверстие находится ближе к середине тела червя.
Свободные черви в своем большинстве хищники и у них даже бывает своеобразное приспособление захвата жертвы. Эта система наблюдается не у всех классов, у более примитивных червей ее нет. Например, ленточные черви питаются всей поверхностью.
Эта система наблюдается не у всех классов, у более примитивных червей ее нет. Например, ленточные черви питаются всей поверхностью.
Выделительная система
Выделительная система довольно масштабна и состоит из множества канальцев, которые объединяются и ведут к выделительным порам.
В паренхиме содержатся специальные клетки, которые загоняют вредные вещества в канальцы. Для человека эти продукты выделения очень опасны и токсичны наравне с ядом.
Ресничные червиВнешнее строение ресничных червей
Размеры ресничных червей
колеблются чаще всего в пределах нескольких миллиметров, реже сантиметров, среди них очень много мелких форм, размеры которых не превышают 1-2 мм. Однако среди турбеллярий имеются и более крупные черви. Так, байкальский червь Polycotylus достигает 30 см, а некоторые наземные тропические формы имеют длину 50-60 см.
Тело турбеллярий в большинстве случаев сплющено в дорзовентральном направлении, листовидной формы, одна-ко среди мелких видов некоторые имеют более пли менее веретенообразную форму.
У большинства турбеллярий на теле нет никаких придатков. Только у некоторых на головном конце имеются два выроста в виде небольших щупалец. Движения турбеллярий разнообразны
. Они обусловлены, с одной стороны, движением ресничек, покрывающих тело турбеллярий, а с другой — сокращением мускулатуры.
Кожно-мускульный мешок
Ресничные червиПоверхность тела
ресничных червей покрыта однослойным ресничным мерцательным эпителием. Под ним расположены многочисленные одноклеточные (реже многоклеточные) слизистые, клейкие и белковые железы, протоки которых открываются наружу среди эпителиальных клеток. Слизистые железы
выделяют слизь, облегчающую скольжение турбеллярий. Выделения клейких желез застывают в виде нитей, на которых животные могут временно подвешиваться на поверхностной пленке воды или подводных предметах. Белковые железы образуют ядовитый секрет, имеющий защитное значение.
Многие эпителиальные клетки содержат так называемые рабдиты. Это сильно преломляющие свет палочки, расположенные внутри клеток. Они представляют собой «оформленный секрет» клеток. Рабдиты
формируются непосредственно в эпителиальных клетках или в клетках, расположенных глубже,- в паренхиме. Последние связаны с эпителиальными клетками цитоплазматическими мостиками, по которым рабдиты продвигаются к поверхности.
Они представляют собой «оформленный секрет» клеток. Рабдиты
формируются непосредственно в эпителиальных клетках или в клетках, расположенных глубже,- в паренхиме. Последние связаны с эпителиальными клетками цитоплазматическими мостиками, по которым рабдиты продвигаются к поверхности.
При малейшем раздражении рабдиты выбрасываются из клеток наружу и расплываются в слизистую массу. Они состоят из ядовитых веществ и представляют собой средство защиты и нападения. Во всяком случае, известно, что многие ресничные черви несъедобны для других животных.
Под кожным эпителием, отделенные от него тонкой базальной перепонкой, расположены слоями мускульные волокна. Непосредственно под эпителием расположен сплошной кольцевой, или поперечный, слой мышечных волокон. Этот слой называется так потому, что оси мускульных клеток расположены поперек оси тела червя. Сокращение этих мышц обусловливает сжатие тела. Под кольцевым слоем обычно расположен слой так называемых косых, или диагональных, мышц. Оси мышечных волокон, составляющих этот слой, расположены перпендикулярно друг к другу и под углом к кольцевому слою. Наконец, третий слой состоит из мышечных волокон, вытянутых вдоль тела животного. Это слой продольных мышечных волокон. Все мышечные слои состоят из гладких мышечных волокон. Мышцы вместе с кожным эпителием образуют кожно-мускульный мешок, очень характерный не только для плоских, но и для других типов червей, хотя число мышечных слоев и их последовательность могут быть различны.
Оси мышечных волокон, составляющих этот слой, расположены перпендикулярно друг к другу и под углом к кольцевому слою. Наконец, третий слой состоит из мышечных волокон, вытянутых вдоль тела животного. Это слой продольных мышечных волокон. Все мышечные слои состоят из гладких мышечных волокон. Мышцы вместе с кожным эпителием образуют кожно-мускульный мешок, очень характерный не только для плоских, но и для других типов червей, хотя число мышечных слоев и их последовательность могут быть различны.
Кроме мышц, входящих в состав кожно-мускульного мешка, у них имеются еще пучки мышц, тянущихся от спинной части кожно-мускульного мешка к брюшной. Это — дорзовентральные мышечные пучки. Совокупность всей описанной мускулатуры обусловливает все довольно сложные движения тела турбеллярий.
Паренхима
Как уже было отмечено, внутри кожно-мускульного мешка все пространство между различными органами заполнено паренхимой, состоящей в основном из рыхло расположенных клеток неопределенной формы; часто эти клетки снабжены отростками, с межклеточным веществом между ними.
Паренхима представляет собой рыхлую соединительную ткань мезодермического происхождения. Среди основных клеток паренхимы расположены многочисленные мускульные волокна, железистые, рабдитные клетки и т. п.
Пищеварительная система
У турбеллярий, как у кишечнополостных и гребневиков, Пищеварительная система замкнутая, т. е. рот является единственным отверстием, через которое поглощается нища и выбрасываются её «непереваренные остатки — экскременты. Для большинства турбеллярий характерно разделение кишечника на два отдела: переднюю эктодермическую глотку и среднюю слепозамкнутую энтодермическую кишку. Рот всегда помещается на брюшной стороне, но может быть ближе к переднему или заднему концу, а иногда может располагаться в центре брюшной поверхности.
У некоторых турбеллярий глотка может отсутствовать или иметь форму короткой простой трубки, средняя кишка у них совсем отсутствует, а пищеварительные клетки расположены в паренхиме, не образуя пищеварительной полости. Такое весьма простое строение органов пищеварения свойственно низшим турбелляриям, преимущественно обитающим в морях и объединяемым в отряд бескишечных (Acoela).
У всех прочих ресничных червей (турбеллярий) глотка хорошо развита, причем чаще всего она представляет собой трубку с очень мускулистыми стенками, помещающуюся в особом влагалище, из которого глотка может высовываться наружу. Такая глотка представляет собой ловчий или сосательный аппарат.
Средняя кишка может иметь различное строение. У одних турбеллярий глотка ведет в среднюю кишку мешковидной формы, не имеющую никаких разветвлений. Это имеет место у мелких турбеллярий.
У крупных турбеллярий кишечник более или менее сильно разветвлен, от мешковидной части кишки отходят ветви: одна вперед к голове и много парных ветвей, идущих во все стороны. Эти выросты кишечника, в свою очередь, разветвляются. Такое строение кишечника наблюдается у морских турбеллярий, относящихся к отряду многоветви-стокишечных (Polycladida). Разветвленность средней кишки и радиальное расположение ее ветвей у многоветвистокишечных дало повод сравнивать среднюю кишку этих турбеллярий с гастроваскулярной системой кишечно-полостных.

Наконец, у турбеллярий из подотряда трехветвистокишечных (Tricladida) главной кишки нет и прямо от глотки отходят три ветви средней кишки. Одна ветвь идет вперед к головному отделу, а две направлены к заднему концу тела. Все эти ветви кишечника, в свою очередь, разветвляются. К этому подотряду относятся многие пресноводные турбеллярий.
Степень разветвленности кишечника различных турбеллярий, несомненно, связана с размерами животных. Не считая бескишечных, наиболее мелкими среди турбеллярий будут формы с не разветвленным кишечником.
Наибольшей степени разветвленности кишечник достигает у более крупных — многоветвистокишечных и трехветвистокишечных турбеллярий. Это объясняется отсутствием у турбеллярий кровеносной системы. Средняя кишка не только является органом пищеварения, но и выполняет функцию распределения пищи по всему телу, подобно гастроваскулярной системе медуз и гребневиков. Стенки средней кишки выстланы однослойным эпителием, состоящим из клеток с закругленными расширенными концами, среди которых расположены особые железистые клетки.
 Эти клетки выделяют в полость кишки пищеварительные ферменты. Однако переваривание пищи в полости кишки происходит только частично. Мелкие частицы пищи захватываются клетками эпителия кишечника и перевариваются внутри этих клеток.
Эти клетки выделяют в полость кишки пищеварительные ферменты. Однако переваривание пищи в полости кишки происходит только частично. Мелкие частицы пищи захватываются клетками эпителия кишечника и перевариваются внутри этих клеток.Таким образом, в отношении процесса пищеварения турбеллярий мало отличаются от кишечнополостных. Клетки средней кишки обладают фагоцитарной функцией, и пищеварение у турбеллярий также, в значительной степени, внутриклеточное.
Анального отверстия и задней кишки у турбеллярий, как и у всех плоских червей, нет. Однако у некоторых турбеллярий имеются особые поры, при помощи которых полость кишечника сообщается с внешней средой. Значение этих пор не выяснено.
Выделительная система
Органы выделения впервые возникают у ресничных червей. Они представлены системой сильно разветвленных каналов, часто образующих перемычки, или анастомозы. Наиболее тонкие канальцы замыкаются слепо концевыми, или терминальными, клетками, а главные каналы открываются выделительными отверстиями. Терминальные клетки грушевидной формы, часто со звездчатыми отростками, расположены они непосредственно в паренхиме. Внутри клеток имеется полость, в которой помещается пучок длинных ресничек. Пучок ресничек находится в непрерывном колебательном движении, напоминающем колебания пламени свечи, за что эти клетки называют пламенными клетками. Полость терминальной клетки продолжается в ее отростке. Это и есть начало выделительного канала. Далее к отростку клетки примыкает ряд удлиненных клеток, сквозь которые проходит канал. Канальцы, отходящие от близко расположенных пламенных клеток, соединяются в более крупные протоки, затем эти протоки впадают в еще более крупные, открывающиеся одним или несколькими отверстиями наружу.
Терминальные клетки грушевидной формы, часто со звездчатыми отростками, расположены они непосредственно в паренхиме. Внутри клеток имеется полость, в которой помещается пучок длинных ресничек. Пучок ресничек находится в непрерывном колебательном движении, напоминающем колебания пламени свечи, за что эти клетки называют пламенными клетками. Полость терминальной клетки продолжается в ее отростке. Это и есть начало выделительного канала. Далее к отростку клетки примыкает ряд удлиненных клеток, сквозь которые проходит канал. Канальцы, отходящие от близко расположенных пламенных клеток, соединяются в более крупные протоки, затем эти протоки впадают в еще более крупные, открывающиеся одним или несколькими отверстиями наружу.
Описанные органы выделяют из тела избыток воды, а также жидкие продукты диссимиляции. Продукты распада органических веществ диффузно проникают из паренхимы в полость выделительной клетки и движением мерцательного пламени гонятся по каналам, которые также выстланы ресничками, и, наконец, выделяются наружу.
Важнейшей чертой выделительных органов ресничных червей (и всех плоских червей) является наличие особых терминальных клеток, замыкающих выделительные каналы. Такого типа выделительные органы беспозвоночных называют протонефридиями.
У различных турбеллярий выделительные органы развиты неодинаково. Они менее развиты у морских форм (многоветвистокишечные и бескишечные турбеллярии), вероятно, потому, что в условиях жизни в соленых водах организм не перегружается водой.
Нервная система
У некоторых наиболее примитивных ресничных червей из отряда бескишечных нервная система представляет собой диффузное нервное сплетение, более густое на переднем конце тела помещается скопление нервных клеток, образующих зачаточный головной ганглий, от которого отходят нервные стволы, идущие почти радиально.
У многоветвистокишечных ресничных червей мозговой ганглий расположен близко от центра тела (у округлых форм) или сдвинут к переднему концу (у вытянутых в длину форм). От него радиально расходится до 11 пар нервных стволов, связанных поперечными перемычками, или комиссурами. Обычно наиболее развита задняя пара нервных стволов. В результате образуется довольно правильная нервная сеть, особенно ясно выраженная у форм с центрально-расположенным нервным ганглием.
Обычно наиболее развита задняя пара нервных стволов. В результате образуется довольно правильная нервная сеть, особенно ясно выраженная у форм с центрально-расположенным нервным ганглием.
Органы чувств ресничных червей, глаза
Органы чувств представлены прежде всего осязательными клетками, особенно многочисленными на переднем конце и по бокам тела. Имеющиеся у некоторых ресничных червей или турбеллярий головные щупальца служат органами химического чувства.
У многих турбеллярий (бескишечных, некоторых Catenulida, Seriata и др.) в тесной связи с головным ганглием расположен статоцист, в виде замкнутого пузырька со статолитом внутри. Статоцист орган ориентации животного в пространстве. При изменении положения тела червя сигнал от статоциста передается через нервную систему мускулатуре турбеллярий до тех пор, пока последняя не примет нормального положения.
У большинства турбеллярий имеется одна или несколько пар глаз (у некоторых наземных планарий более 1000) иного строения, чем уже известные нам глаза медуз. Глаза помещаются непосредственно под кожным эпителием и состоят из пигментного бокала и зрительных клеток. Пигментный бокал, чаще состоящий из одной гигантской клетки, имеет форму чаши, обращенной вогнутой частью к периферии. Клетка (или клетки, если бокал многоклеточный) наполнена пигментом, и ядро помещается в ее выпуклой части. В пигментный бокал погружена одна или несколько зрительпых клеток своеобразной, булавовидной формы. Расширенные концы этих клеток заканчиваются светочувствительными палочками или колбочками. Изогнутые части зрительных клеток обращены к поверхности тела, и к ним подходят нервы головного ганглия. Благодаря такому расположению клеток лучи света сначала проходят через плазму зрительной клетки, а затем уже попадают на светочувствительную часть клетки. (У других животных светочувствительная часть клетки обращена непосредственно к свету). Поэтому глаза такого строения, как у турбеллярий, называются обращенными или инвертированными.
Глаза помещаются непосредственно под кожным эпителием и состоят из пигментного бокала и зрительных клеток. Пигментный бокал, чаще состоящий из одной гигантской клетки, имеет форму чаши, обращенной вогнутой частью к периферии. Клетка (или клетки, если бокал многоклеточный) наполнена пигментом, и ядро помещается в ее выпуклой части. В пигментный бокал погружена одна или несколько зрительпых клеток своеобразной, булавовидной формы. Расширенные концы этих клеток заканчиваются светочувствительными палочками или колбочками. Изогнутые части зрительных клеток обращены к поверхности тела, и к ним подходят нервы головного ганглия. Благодаря такому расположению клеток лучи света сначала проходят через плазму зрительной клетки, а затем уже попадают на светочувствительную часть клетки. (У других животных светочувствительная часть клетки обращена непосредственно к свету). Поэтому глаза такого строения, как у турбеллярий, называются обращенными или инвертированными.
Размножение
Подавляющее большинство ресничных червей — гермафродиты. Половые органы у ресничных червей устроены крайне сложно и разнообразно в разных группах. Они отличаются по числу гонад, их структуре, присутствию многих дополнительных образований половой системы. Так, мужские половые железы — семенники — могут быть крупными одиночными или парными либо мелкими многочисленными образованиями. Женские половые железы — яичники — обычно парные, но могут быть и одиночными, и многочисленными. У более примитивных турбеллярий яичники простые. В них формируются яйца, которые содержат некоторое количество желтка, а также скорлупкового вещества. Такие яйца называют энтолецитальными. У более высокоорганизованных турбеллярий яичники дифференцируются на отделы: один из них, большой, продуцирует только питательные желточные клетки, а другой, маленький,- яйцеклетки. Эти отделы могут превращаться в самостоятельные парные органы: собственно яичники и желточники. Образующиеся яйцеклетки совсем лишены желтка. После оплодотворения их окружают желточные клетки, а затем вокруг них формируется общая оболочка.
Половые органы у ресничных червей устроены крайне сложно и разнообразно в разных группах. Они отличаются по числу гонад, их структуре, присутствию многих дополнительных образований половой системы. Так, мужские половые железы — семенники — могут быть крупными одиночными или парными либо мелкими многочисленными образованиями. Женские половые железы — яичники — обычно парные, но могут быть и одиночными, и многочисленными. У более примитивных турбеллярий яичники простые. В них формируются яйца, которые содержат некоторое количество желтка, а также скорлупкового вещества. Такие яйца называют энтолецитальными. У более высокоорганизованных турбеллярий яичники дифференцируются на отделы: один из них, большой, продуцирует только питательные желточные клетки, а другой, маленький,- яйцеклетки. Эти отделы могут превращаться в самостоятельные парные органы: собственно яичники и желточники. Образующиеся яйцеклетки совсем лишены желтка. После оплодотворения их окружают желточные клетки, а затем вокруг них формируется общая оболочка. Такие яйца называют эктолецитальными.
Такие яйца называют эктолецитальными.
Протоки половых желез — семяпроводы и яйцеводы — обычно парные, в нижнем отделе сливаются в непарные образования. Они могут открываться самостоятельно мужским и женским половыми отверстиями на брюшной стороне тела или в общую половую клоаку.
Низшие турбеллярий лишены женских выводных протоков. Так, у некоторых бескишечных ресничных червей яйцеводы отсутствуют. Сперма вносится партнером, который прорывает покровы червя совокупительным органом. Сперма попадает в паренхиму и оплодотворяет находящиеся там яйца. Откладка яиц возможна через разрыв стенок тела или через рот, как у кишечнополостных.
Сложное строение гермафродитной половой системы ресничных червей разберем на примере обычной в пресных водах молочной планарии (Dendrocoelum lacteum).
Мужские половые органы состоят из многочисленных мелких семенников, расположенных в паренхиме по бокам всего тела. От семенников отходят тончайшие семявыносящие канальцы, которые впадают в два семяпровода, направляющихся назад. Позади глотки семяпроводы впадают в семенной мешок. В задней части семенной мешок переходит в совокупительный орган, пронизанный семяизвергательным каналом. Совокупительный орган при совокуплении выдвигается через половую клоаку и вводится в половое отверстие другой особи.
Позади глотки семяпроводы впадают в семенной мешок. В задней части семенной мешок переходит в совокупительный орган, пронизанный семяизвергательным каналом. Совокупительный орган при совокуплении выдвигается через половую клоаку и вводится в половое отверстие другой особи.
Женский половой аппарат состоит чаще всего из одной пары яичников, расположенных в передней части тела. От яичников отходят два длинных яйцевода, направляющихся по бокам тела назад и сливающихся в непарный яйцевод, который открывается в половую клоаку рядом с карманом совокупительного органа.
На всем протяжении парных яйцеводов в них открываются протоки многочисленных желточников, в которых образуются особые желточные клетки, богатые питательными материалами.
В половую клоаку открываются еще два органа: копулятивная сумка — складчатый мешок с довольно тонким стебельчатым каналом и мускулистый железистый орган. Его значение не выяснено.
При спаривании молочных планарий совокупительный орган вводится в половое отверстие и через половую клоаку в копулятивную сумку другой особи. Таким образом, сперма прежде всего попадает в копулятивную сумку, а из нее — в яйцеводы, в ту их часть, которая расположена вблизи яичников. Оплодотворение происходит при выходе яиц из яичника в яйцевод. Затем яйца, продвигаясь по яйцеводам мимо отверстий желточных протоков, окружаются желточными клетками и попадают в половую клоаку. Здесь вокруг яиц вместе с желточными клетками образуется кокон из выделений желточных клеток и особых скорлуповых желез. Отложенный кокон подвешивается к подводным предметам.
Таким образом, сперма прежде всего попадает в копулятивную сумку, а из нее — в яйцеводы, в ту их часть, которая расположена вблизи яичников. Оплодотворение происходит при выходе яиц из яичника в яйцевод. Затем яйца, продвигаясь по яйцеводам мимо отверстий желточных протоков, окружаются желточными клетками и попадают в половую клоаку. Здесь вокруг яиц вместе с желточными клетками образуется кокон из выделений желточных клеток и особых скорлуповых желез. Отложенный кокон подвешивается к подводным предметам.
Развитие
У ресничных червей с энтолецитальными яйцами происходит полное неравномерное дробление по спиральному типу, напоминающее дробление яиц кольчатых червей, немертин и моллюсков.
Развитие турбеллярий обычно прямое, лишь у некоторых групп наблюдается метаморфоз. У морских многоветвистокишечных ресничных червей из яйца выходит своеобразная мюллеровская личинка яйцевидной формы. Вначале она обнаруживает черты радиальной симметрии, а затем все более приобретает билатеральную симметрию. Впереди рта, расположенного на брюшной стороне, имеется 8 лопастных выростов, покрытых ресничками. Такая личинка ведет планктонный образ жизни, и этим обеспечивается расселение морских турбеллярий. Личинки морских турбеллярий переносятся морскими течениями на большие расстояния и постепенно превращаются во взрослых животных. При этом их рот сдвигается вперед, околоротовые лопасти уменьшаются, а все тело сплющивается. Личинка опускается на дно и окончательно приобретает билатеральную симметрию.
Впереди рта, расположенного на брюшной стороне, имеется 8 лопастных выростов, покрытых ресничками. Такая личинка ведет планктонный образ жизни, и этим обеспечивается расселение морских турбеллярий. Личинки морских турбеллярий переносятся морскими течениями на большие расстояния и постепенно превращаются во взрослых животных. При этом их рот сдвигается вперед, околоротовые лопасти уменьшаются, а все тело сплющивается. Личинка опускается на дно и окончательно приобретает билатеральную симметрию.
Развитие эктолецитальных яиц проходит по-иному. У описанной выше молочной планарии в коконе содержится от 20 до 40 яиц и около 80- 90 тысяч желточных клеток. Последние окружают каждую яйцеклетку, а позднее сливаются и образуют синцитий. Бластомеры оказываются разобщенными и погруженными в общую массу желтка. Они образуют три группы клеток, две из которых обеспечивают поглощение желтка зародышем, а из третьей формируется сам зародыш. Развитие прямое: из кокона отрождаются маленькие планарии.
Бесполое размножение наблюдается у некоторых турбеллярий из отрядов Macrostomida, Catenulida и Seriata (подотряд Tricladida). Оно заключается в поперечном делении червей. У некоторых форм, например у Microstomum lineare, бесполое размножение происходит все лето и лишь осенью сменяется половым. При бесполом размножении посередине тела появляется перетяжка, и у задней половины начинается образование рта и глотки. Задолго до разделения червя надвое дочерние особи также начинают делиться и появляются перетяжки II, III и т. п. порядков. Так образуется цепочка из делящихся зооидов.
Галлерея
Ресничные черви (от лат. turbellaria или турбеллярии) – это такой класс червей, который зоологи относят к типу плоские черви. Общее количество всех представителей данного класса колеблется в пределах 3,5 тысяч особей , из которых подавляющее большинство — это свободноживущие существ а.
Особенности
Тело турбеллярий покрыто мерцательным ресничным эпителием
. Благодаря работе ресничек и сокращению мускульной части тела турбеллярия передвигается в пространстве ползком или вплавь.
Благодаря работе ресничек и сокращению мускульной части тела турбеллярия передвигается в пространстве ползком или вплавь.
Многие виды турбеллярий показали удивительные способности регенерации.
Особенно выделилась в этом умении планария – обитатель водоемов и один из крупных типичных представителей класса ресничные черви. Эта турбеллярия из отряда трехветвистых умеет полностью восстановиться и сформироваться в полноценного червя даже если от ее тела останется 0,01 часть.
При неблагоприятных для жизни условиях планария самопроизвольно распадается на мельчайшие кусочки, чтобы в последующем ожить в более приятной для существования среде.
Размеры особей колеблются от крошечных и невидимых до вполне внушительных размеров – 30-40 см .
Классификация ресничных червей
Новая, основательно переработанная и усложненная классификация турбеллярий на данный момент включает в себя 12 отрядов
. Если раньше турбеллярий разделяли 4 отряда, основываясь лишь на различии строения кишечной системы, то теперь отличительными признаками являются: типы яиц и способы дробления, строение половой и выделительных систем и т. д.
д.
Специалисты выделяют несколько главных отрядов, которые считаются основными:
- Отряд Бескишечные (Acoela) – крошечные и преимущестенно морские обитатели с примитивной формой строения тела: нет кишечника (его заменяет пищеварительная паренхема), отсутствуют органы выделения, типичные для других представителей низших беспозвоночных, примитивное устройство половых органов и т.д. Есть статоцист – орган чувства равновесия.
- Отряд Макростомиды (Macrostomida) – примитивные мелкие черви. Обитают в морской и пресной воде. Кишечник мешковидный. Половая система примитивная – отсутствуют обособленные желточники. Отдельные представители размножаются путем поперечного деления.
- Гнатостомулиды (Gnathostomulida) – мелкие турбеллярии, обитающие в песках морских побережий. Кишечник мешковидный. В глотке присутствует пара хитиновых челюстей. Характерная особенность – вместо ресничек имеют жгутики, происходящие из каждой клетки эпителия.
- Многоветвистые (Polyclaida)
– крупные (до 15 см) морские обитатели с листовидной формой тела.
 Кишечник разветвленный, множественное количество половых желез. Примитивные черты – нет обособленных желточников, а у некоторых представителей и половых притоков и т.п.
Кишечник разветвленный, множественное количество половых желез. Примитивные черты – нет обособленных желточников, а у некоторых представителей и половых притоков и т.п. - Трехветвистые, или планарии (Tricladida) – пресноводные особи с листовидным и лентовидным строением тел. Половая система развита, присутствуют многочисленные семенники, пара яичников и желточники. Трехветвистое строение средней кишки.
- Прямокишечные (Rhabdocoela, или Neorhabdocoela) – животные маленького размера: от 0,5 до 5 мм. Тело почти не приплюснуто, практически цилиндрическое. Плавают в воде с помощью ресничек. Характерная черта – прямой и слепо замкнутый кишечник. Рот находится на передней части тела. Присутствуют органы чувств – пара глазков. Выделительная система состоит из пары каналов с двумя отверстиями.
- Темноцефалы (Themnocephalida) – черви несколько миллиметров в длину с вытянутым туловищем. Имеют щупальцы и присоски с железами. Эпителий лишен ресничек.
Строение и физиология
Для представителей турбеллярий характерны общие черты строения, как, например, вытянутое в длину тело листовидной или лентовидной формы, часто не имеющее дополнительных придатков. Лишь редкие особи могут иметь щупальцеобразные выросты. Ресничные черви отличаются разнообразной пятнистой окраской и редко встречаются особи бесцветные или одного сплошного цвета.
Лишь редкие особи могут иметь щупальцеобразные выросты. Ресничные черви отличаются разнообразной пятнистой окраской и редко встречаются особи бесцветные или одного сплошного цвета.
Характерная черта ресничных червей, которая и дала название этому классу – это особое строение мерцательного эпителия в виде высокорасположенных клеток цилиндрической формы, на краях которых располагаются реснички. Биение ресничек обеспечивает передвижение турбеллярии в пространстве, как и сокращение кожно-мускульного мешка.
Все внутренние органы, которые существуют у того или иного представителя ресничных червей, соединены рыхлой тканью – паренхимой . Она играет роль посредника в пищеварительной системе и от одного к другому органу передает продукты пищеварения в кишечнике и продукты обмена веществ к выделительной системе. Паренхиму специалисты рассматривают и как соединительную ткань
Кожные покровы часто умеют множество желез, выделяющие различные по составу и свойствам жидкости. Зачастую это отравляющие вещества, играющие защитную роль.
Зачастую это отравляющие вещества, играющие защитную роль.
Пищеварительная система и питание
Между отдельными представителями отрядов ресничных червей существуют основополагающие различия в строении их пищеварительной системы. Существуют виды червей с очень примитивным устройством переваривания пищи – бескишечные .
- Темноцефалы.
- Удонеллиды.
Примитивные турбеллярии переваривают пищу в специальных пищеварительных клетках или паренхиме.
Особи с более эволюционно развитой пищеварительной системой подразделяются на виды:
- Ветвистокишечные.
- Прямокишечные.
Нервная система, органы чувств и дыхание
Типичное строение нервной системы для представителей класса турбуллярий – мозговой ганглий и ответвленные от него нервные стволы. Нервные окончания концентрируются в основном у передней части тела, формируя своеобразный двойной узел – ганглий
. Это примитивный мозг беспозвоночных ресничных червей работает как нервный центр.
Отдельные примитивные представители турбуллярий имеют просто устроенную нервную систему, которая состоит лишь из сплетения небольшого количества нервных окончаний и расположенных близко к поверхности тела. А другие могут обладать большим количеством нейронов, более утолщенным и чувствительным к внешним раздражителям.
Органы чувств хорошо развиты . Для осязания турбеллярии используют всю поверхность своей кожи. Осязают с помощью ресничек. Некоторые черви имеют орган статосцист – орган чувства равновесия. Почти у всех червей есть глаза.
Отдельно сформированных органов дыхания у турбеллярий нет. Вся поверхность их кожи имеет способность поглощать кислород, содержащийся в воде. Именно способность к дыханию путем поглощения кислорода всей поверхностью тела отличает свободноживущих ресничных червей от других плоских червей, большая часть которых анаэробы. Что означает их возможность существовать в среде без кислорода. Дыхание для ресничных червей – важная часть существования.
Размножение и половые признаки
Турбеллярии в большинстве своем гермафродиты, т.е. имеют как мужские половые органы, так и женские. Спаривание происходит перекрестно и оба червя оплодотворяют друг друга в роле самцов, и потом становятся оплодотворенными как самки.
После удачного спаривания формируется яйцо, из которого выходит личинка, напоминающая внешним видом взрослую особь. Отдельные виды способны размножаться делением пополам, после чего из каждой части формируется отдельная полноценная особь.
Строение кожно-мускульного мешка ресничных червей.
Профилактика.
Диагностика
1.Личная. Соблюдение правил личной гигиены.
2.Общественная. Санитарное обустройство мест общественного пользования, наблюдение за источниками общественного водоснабжения, санитарно-просветительская работа с населением, борьба с грызунами, гигиеническое содержание свиней.
Кожно-мускульный мешок
.
Под эпителием находится базальная мембрана
, служащая для придания телу определенной формы и для прикрепления мышц. Совокупность мышц и эпителия образует единый комплекс —кожно-мускульный мешок
. Мышечная система состоит из нескольких слоев гладких мышечных волокон
. Наиболее поверхностно располагаются кольцевые мышцы
, несколько глубже — продольные
и самые глубокие — диагональные мышечные волокна
. Кроме перечисленных видов мышечных волокон для ресничных червей характерны спинно-брюшные
, или дорзовентральные
, мышцы
. Это пучки волокон, идущие от спинной стороны тела к брюшной.
Совокупность мышц и эпителия образует единый комплекс —кожно-мускульный мешок
. Мышечная система состоит из нескольких слоев гладких мышечных волокон
. Наиболее поверхностно располагаются кольцевые мышцы
, несколько глубже — продольные
и самые глубокие — диагональные мышечные волокна
. Кроме перечисленных видов мышечных волокон для ресничных червей характерны спинно-брюшные
, или дорзовентральные
, мышцы
. Это пучки волокон, идущие от спинной стороны тела к брюшной.
Движение осуществляется за счет биения ресничек (у мелких форм) или сокращения кожно-мускульного мешка (у крупных представителей).
Четко выраженной полости тела
ресничные черви не имеют. Все промежутки между органами заполненыпаренхимой
— рыхлой соединительной тканью. Небольшие пространства между клетками паренхимы заполнены водянистой жидкостью, благодаря чему может осуществляться передача продуктов от кишечника к внутренним органам и перенос продуктов обмена к выделительной системе. Кроме этого, паренхиму можно рассматривать как опорную ткань.
Кроме этого, паренхиму можно рассматривать как опорную ткань.
Мышечная часть кожно-мускульного мешка образована несколькими слоями мышечных волокон. Снаружи располагается кольцевой слой, под ним продольный и самый нижний — диагональный, волокна которого идут под углом друг к другу. Именно совокупность покровов и лежащих ниже слоев мышечных волокон и образует кожно-мускульный мешок.
Пищеварительная система
ресничных червей слепозамкнутая
. Рот
служит и для проглатывания пищи
, и для выбрасывания непереваренных остатков пищи
. Рот располагается обычно на брюшной стороне тела и ведет вглотку
. У некоторых крупных ресничных червей, например у пресноводной планарии, ротовое отверстие открывается в глоточный карман
, в котором находится мускулистая глотка
, способная вытягиваться и высовываться через рот наружу. Средняя кишка
у мелких форм ресничных червей представляет собой ветвящиеся во все стороны каналы
, а у крупных форм кишечник представлен тремя ветвями
: одной передней
, идущей к переднему концу тела, и двумя задними
, идущими по бокам к заднему концу тела.
Большинство турбеллярии — хищники, питающиеся различными мелкими животными. Пищеварительная система состоит из передней и средней кишки, замыкающейся слепо. Рот служит не только для проглатывания пищи, но и для выбрасывания непереваренных твердых остатков. Рот помещается обычно на брюшной стороне тела: на небольшом расстоянии от переднего конца тела, посредине брюшной стороны, или ближе к заднему полюсу. Рот ведет в эктодермальную глотку, которая, в свою очередь, переходит в среднюю кишку. У некоторых крупных турбеллярии (у пресноводных планарий отр. Tricladida и у морских турбеллярии отр. Polycladida) ротовое отверстие открывается не в глотку, а в особое глубокое впячивание наружных покровов, называемое глоточным карманом (рис. 124 , рис. 135). Со дна кармана в его полость вдается мускулистая глотка. Она имеет вид трубки, которая может сильно вытягиваться и высовываться через рот наружу, служа для захвата добычи.
Энтодермальная средняя кишка устроена у многих форм Turbellaria различно. У мелких турбеллярий (отр. Rhabdocoela, Macrostomida и др.) она имеет вид простого мешка или слепо замкнутой трубки (рис. 134). У крупных же форм кишечник, как правило, разветвлен. Так, у многоветвистых (Polycladida) глотка ведет в желудок, от которого во все стороны к краям тела отходят ветвящиеся каналы, слепо замкнутые на концах (рис. 135). У трехветвистых (Tricladida) от глотки, находящейся вблизи средины тела, отходят сразу три главные ветви кишечника (рис. 123 , рис. 124): одна идет прямо вперед, другие две загибаются и по бокам глотки направляются назад; каждая из ветвей дает побочные слепые веточки.
У мелких турбеллярий (отр. Rhabdocoela, Macrostomida и др.) она имеет вид простого мешка или слепо замкнутой трубки (рис. 134). У крупных же форм кишечник, как правило, разветвлен. Так, у многоветвистых (Polycladida) глотка ведет в желудок, от которого во все стороны к краям тела отходят ветвящиеся каналы, слепо замкнутые на концах (рис. 135). У трехветвистых (Tricladida) от глотки, находящейся вблизи средины тела, отходят сразу три главные ветви кишечника (рис. 123 , рис. 124): одна идет прямо вперед, другие две загибаются и по бокам глотки направляются назад; каждая из ветвей дает побочные слепые веточки.
Такое строение кишечника не случайно. У мелких турбеллярий, размер которых не превышает нескольких миллиметров, продукты пищеварения легко распределяются по всему телу через рыхлую паренхиматозную ткань. У крупных же трикладид и поликладид, иногда достигающих 1-3 и даже 30 см, этот процесс чрезвычайно затруднен. У таких турбеллярий функцию распределения продуктов пищеварения по телу животного выполняют ветви кишечника, пронизывающие паренхиму во всех направлениях и непосредственно соприкасающиеся со всеми тканями и органами.
В процессе переваривания пищи у турбеллярий, так же как у кишечнополостных, большое место занимает внутриклеточное пищеварение. Частицы пищи, предварительно подвергающиеся обработке секретом глоточных желез, поступают в кишечник и захватываются клетками кишечного эпителия, в которых формируются многочисленные пищеварительные вакуоли. У отряда бескишечных турбеллярий выраженная средняя кишка вообще отсутствует (рис. 126). Глотка у них вдается в несколько обособленный участок паренхимы (пищеварительная паренхима), в которой происходит внутриклеточное переваривание пищи
Пищеварительная система ресничных червей довольно разнообразна в пределах класса и варьирует от примитивной — без оформленного кишечника до сравнительно сложной — с ветвистым кишечником. Основные отряды ресничных червей отличаются прежде всего формой кишечника.
Довольно сложный кишечник имеется у многоветвистых турбеллярий — поликлад (отряд Polycladida). Ротовое отверстие у них, как правило, ближе к заднему концу тела и ведет в складчатую глотку, от которой радиально расходятся множество слепых ветвей энтодермального кишечника.
В подотряде трехветвистых, или планарий — триклад (Tricladida ) от глотки, расположенной посередине тела, отходят три ветви кишечника. Одна ветвь направлена к переднему концу тела, а две — назад. При этом пища сначала поступает в передний вырост, а оттуда перетекает в задние. Таким образом, питательными веществами первоначально обеспечиваются органы, расположенные в передней части тела.
У отряда прямокишечных (Rhabdocoela) глотка расположена на переднем конце тела и кишечник прямой, неветвистый.
У ресничных червей с оформленным кишечником большую роль в переваривании пищи выполняют глоточные железы. У многих видов наблюдается внекишечное пищеварение. Нередко планарии нападают на довольно крупные жертвы (моллюсков, рачков). В тело жертвы они выделяют пищеварительные ферменты, а затем всасывают полупереваренную пищу.
Несмотря на определенную сложность пищеварительной системы, усвоение пищи у турбеллярий в значительной мере осуществляется за счет внутриклеточного пищеварения, то естьфагоцитоза эпителиальных клеток средней кишки.
Большинство турбеллярий являются хищниками и питаются различными мелкими беспозвоночными. Обнаружив жертву, червь накрывает ее своим телом, после чего заглатывает. У планарий для этого служит выдвижная глотка, которая выбрасывается из глубокого впячивания наружного покрова — глоточного кармана. Если размер добычи слишком велик и ее невозможно проглотить целиком, червь отрывает куски сильными сосательными движениями глотки, после чего заглатывает их. Однако мелкие членистоногие (например, рачки) покрыты жестким панцирем и разорвать их планария не может. В таком случае она выделяет из глотки наружу пищеварительные ферменты, которые расщепляют ткани жертвы вне организма червя, после чего размягченная пища всасывается глоткой. Таким образом, у турбеллярий возможно и наружное пищеварение. Некоторые турбеллярии (планарии) обладают своеобразной способностью использовать «трофейное оружие». Ученые установили, что при поедании гидр планарией стрекательные клетки не разрушаются, а мигрируют в покровы червя и защищают его.
Класс Ресничные черви, или Турбеллярии (Turbellaria)
Турбеллярии — основной класс плоских червей; в основном они свободноживущие формы, населяющие моря, пресноводные бассейны и влажную почву. У турбеллярий значительного развития достигает нервная система, хорошо развиты органы чувств, они быстро реагируют на внешние факторы и в частности на свет и присутствие пищи.
Все они — хищники, и этот факт несомненно способствовал более интенсивному развитию органов чувств, ловчих и защитных приспособлений, интенсификации двигательных реакций. Как правило, все они — мелкие формы. Кожа несет ресничный покров. Рот всегда на брюшной стороне. Ануса нет. Кожно-мускульный мешок состоит из эпителия и сложной системы мышц. Центральная нервная система представлена продольными стволами и головными ганглиями. Кишечник либо отсутствует, либо имеет вид прямой или ветвящейся трубки. Пищеварение, как правило, внутриклеточное. Дыхание кожное. Кровеносная система отсутствует. Функция выделения принадлежит кишечнику и специальным органам выделения — протонефридиям. Как правило, гермафродиты. Половое отверстие на брюшной стороне. Развитие прямое или с превращением.
Дыхание кожное. Кровеносная система отсутствует. Функция выделения принадлежит кишечнику и специальным органам выделения — протонефридиям. Как правило, гермафродиты. Половое отверстие на брюшной стороне. Развитие прямое или с превращением.
Представитель класса ресничных червей — молочная планария (Dendrocoelum lacteum)
Форма тела и внешняя организация. Молочная планария хорошо отличается от других видов молочно-белым цветом тела. У наевшихся особей на белом фоне тела ясно просвечивает темный ветвистый кишечник. Эта относительно крупная форма достигает 15—26 мм в длину при поперечнике тела примерно 6 мм. Головной конец тела поперечно срезан, задний — заостренно округлен. Позади переднего края головного конца расположены явственные черные глаза, а под основанием его развита присасывательная бороздка. Все тело молочной планарии покрыто равномерным ресничным покровом. Работа ресничек обеспечивает плавное скольжение по субстрату, которое столь характерно для этого червя, как и для других мелких видов.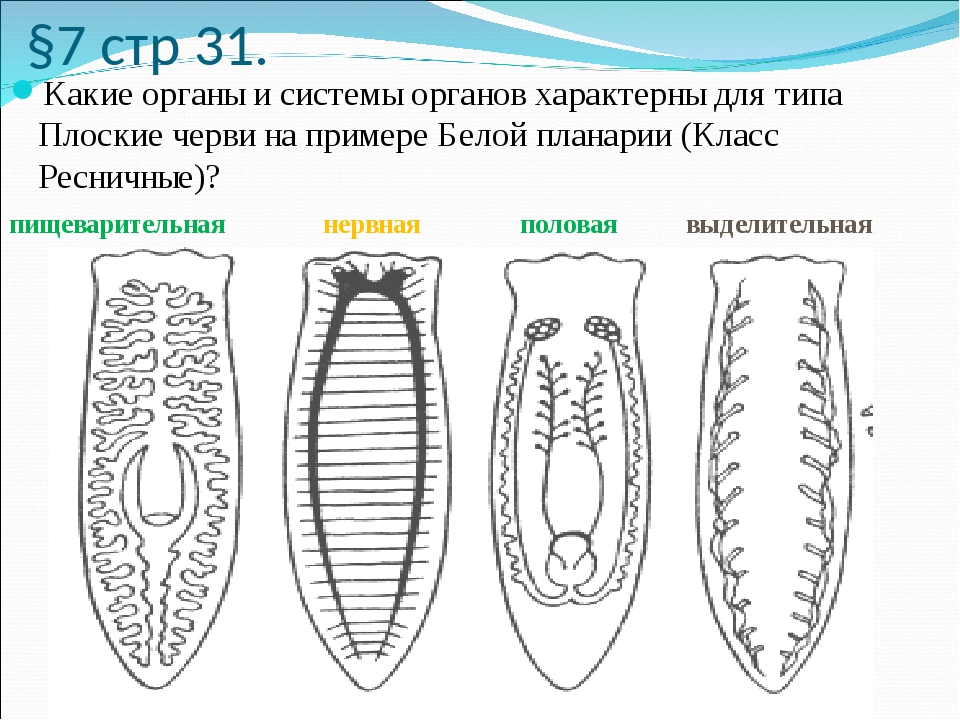
На брюшной стороне, несколько кзади от середины тела, расположено ротовое отверстие, а примерно посередине между ним и задним концом тела — половое. Мельчайшие боковые выделительные отверстия в количестве 8 пар расположены дорзально. Обитающая в наших пресных водах молочная планария обычно держится под различными подводными предметами, где или неподвижно сидит на месте, или плавно ползет по субстрату в поисках пищи. Тело молочной планарии отличается присущей всем турбелляриям гибкостью и эластичностью, способно удлиняться и сокращаться. Животное относительно быстро и плавно скользит по субстрату, по подводным листьям, по затененной поверхности погруженных камней, по нижней стороне пленки поверхностного натяжения воды.
Еще интересные статьи по теме:
Ответы | § 5. Строение и образ жизни ресничных червей — Биология, 8 класс
1. Где живут молочные планарии?
Молочная планария обитает в пресных водоёмах под камнями, листьями растений и других укрытий.
2. Какие группы мышц есть у планарии?
Тело червя покрыто эпителием, с которым срастаются три слоя мышц.
Группы мышц планарии
- Поперечные (кольцевые) мышцы. Они расположены вокруг тела червя.
- Продольные мышцы. Находятся под поперечными и тянутся от головной части червя до хвостовой.
- Косые (диагональные) мышцы. Соединяют спинные и брюшные мышцы, иногда проходят сквозь тело червя.
Все группы мышц срастаются с эпителием и образуют кожно-мускульный мешок.
3. Из каких отделов состоит пищеварительная система ресничного червя?
Отделы пищеварительной системы:
- рот (находится посередине тела),
- глотка (соединена со ртом),
- кишечник (связана с глоткой).
4. Как происходит выделение продуктов обмена у планарии?
Кишечник червя осуществляет доставку пищи ко всем участкам тела. Организм не имеет анального отверстия, поэтому продукты обмена выходят через рот.
5. Чем отличается строение и образ жизни гидры и планарии?
- Гидра является двухслойным животным, планария — трёхслойным.
- Гидра относится к типу кишечнополостных, планария — к типу плоских червей.
- Гидра не имеет сформировавшихся тканей, планария имеет покровную, мышечную ткани и паренхиму.
- Гидра размножается половым способом и бесполым, планария имеет мужские и женские органы, является гермафродитом.
- Гидра имеет лучевую симметрию, планария двухстороннюю симметрию тела.
- Гидра не имеет систем органов, планария имеет пищеварительную, нервную, половую и выделительную системы.
- У гидры два вида пищеварения, у планарий — одно.
- Гидра имеет цилиндрическую форму тела, планария — листовидную.
6. Как происходит размножение и развитие молочной планарии?
Планария, являясь гермафродитом, имеет мужские и женские половые клетки: два яичника и семенники, содержащие сперматозоиды. Поэтому оплодотворение происходит внутри организма. Планария откладывается яйца в защитной оболочке — коконе — который прикрепляет к различным предметам в воде. Затем через 2-3 недели маленькие планарии разрывают кокон.
Поэтому оплодотворение происходит внутри организма. Планария откладывается яйца в защитной оболочке — коконе — который прикрепляет к различным предметам в воде. Затем через 2-3 недели маленькие планарии разрывают кокон.
7. Почему плоские черви, несмотря на примитивное строение, широко распространены и многочисленны?
Местом обитания червей являются пресные и морские водоёмы, тропические леса. Также они ведут паразитический образ жизни. Способ размножения — двуполый. За счёт этого они получили широкое распространение.
8. Если положить планарию на спинную сторону, то она перевернётся на брюшную сторону. При ярком свете планария стремится спрятаться в затенённых местах. Объясните такое поведение планарии.
Планария имеет реакцию на определённый раздражитель (таксис). Если её перевернуть на брюшную сторону, сработает реакция — планария примет прежнее положение. Также она имеет отрицательный фототаксис, который срабатывает при определённом свете.
Класс ресничные черви, подготовка к ЕГЭ по биологии
Класс ресничные черви представлен исключительно свободноживущими (хищными) формами. Такой образ жизни предрасполагает к более прогрессивному строению мышечной, нервной системы, хорошо развитым органам чувств (по сравнению с паразитами). Всего класс ресничные черви насчитывает около 3500 видов. Представители класса: молочная планария, черная планария, турбеллярии.
Изучать класс ресничные черви мы будем на примере типичного представителя — молочной планарии.
Предпочитает молочная планария водоемы с чистой, богатой кислородом водой, медленно текучие или стоячие. Могут встречаться и в загрязненных местах — лужах, при этом их излюбленное место — затененные места у дна.
Средние размеры животного весьма скромные — 2,6 см в длину и 6 мм в толщину. Тело уплощено (тип все же называется «плоские» черви), задний конец заострен.
Поверхность тела планарии покрыта эпителием, на котором находится множество ресничек, что облегчает движение планарии.
Тело уплощено (тип все же называется «плоские» черви), задний конец заострен.
Поверхность тела планарии покрыта эпителием, на котором находится множество ресничек, что облегчает движение планарии.
В толще эпителиального слоя находится множество выводных протоков желез, из которых выделяется клейкий секрет, покрывающий тело планарии. За счет этого секрета к планарии приклеиваются мелкие животные: мальки рыб, беспозвоночные, которые становятся ее добычей. Часть желез, расположенных по краям тела (белковые железы) выделяют токсический секрет.
Особые железы в эпителии — рабдитные клетки, выполняют важную защитную функцию, благодаря им планария почти не поедается другими животными. При возникновении опасности рабдиты раздражаются и выстреливаются на поверхность тела, где образуют защитную слизь.
Передвигаются планарии не только за счет биения ресничек, но и за счет сокращение мышц, которых
у них три слоя: кольцевые, продольные и косые. Спинно-брюшные мышцы, тянущиеся от верхней стороны тела к нижней, не относятся к кожно-мускульному мешку.
При сокращении спинно-брюшных мышц тело сплющивается. За счет развитой мышечной системы тело планарии отличается гибкостью и эластичностью.
Спинно-брюшные мышцы, тянущиеся от верхней стороны тела к нижней, не относятся к кожно-мускульному мешку.
При сокращении спинно-брюшных мышц тело сплющивается. За счет развитой мышечной системы тело планарии отличается гибкостью и эластичностью.
Полость тела у молочной планарии, как и у всех плоских червей — отсутствует. Пространство между органами занято клетками неправильной формы, отростки которых переплетаются друг с другом — такая структура носит название паренхима (мезенхима). Высокая способность к регенерации обусловлена именно клетками мезенхимы.
Ротовое отверстие располагается с нижней стороны тела, продолжается в мускулистую глотку. Когда планария пытается поймать добычу, глоточный аппарат способен резко выдвигаться, чтобы увеличить шансы на успешный обед.
Если обед пойман и его размеры меньше, чем глоточное отверстие, далее он поступает в разветвленный кишечник (обычно имеет 2-3 ответвления).
В случае больших размеров добычи, планария начинает выделять ферменты во внешнюю среду и жертва подвергается внеклеточному пищеварению.
Анальное отверстие отсутствует, поэтому удаление непереваренных остатков пищи происходит через ротовое отверстие.
Кровеносная система у плоских червей отсутствует, поэтому кишечник имеет разветвленную конфигурацию, помогающую доставить необходимые питательные вещества ко всем тканям и органам.
Специализированные органы дыхания отсутствуют, осуществляется дыхание всей поверхность тела. Напомню, что дыхание — процесс поглощение кислорода и выделение углекислого газа.
Выделение из организма жидких продуктов обмена веществ осуществляется с помощью специальных органов — протонефридий, представляющих собой
систему каналов.
Нервная система стволового (лестничного, или ортогонального) типа. Как и у плоских червей вообще, представлена двумя головными нервными узлами (ганглиями), от которых отходят нервные стволы, имеющие множество ответвлений, иннервирующих все внутренние органы и кожу.
Органы чувств хорошо развиты у свободноживущих форм, в частности у молочной планарии. Имеются головные щупальца с хеморецепторами, помогающими находить пищу. Щупальца (головные лопасти) расположены на переднем конце тела. Рядом с щупальцами располагаются два глаза, с помощью которых планария оценивает уровень освещенности.
Между глазами расположен орган равновесия — статоцисты.
К органам чувств, располагающимся на переднем конце тела, от головных узлов направляются короткие нервы.
Молочная планария относится к гермафродитам. На одной особи имеются как мужские, так и женские половые органы. Размножаются преимущественно половым способом.
Мужская половая система состоит из пузырьковидных семенников, в которых созревают двужгутиковые сперматозоиды. От семенников отходят множество канальцев, впадающих в семяпровод, а семяпроводы, в свою очередь, соединяются в семенной пузырь. Из семенного пузыря начинается семяизвергательный канал, который продолжается в особый совокупительный орган — циррус.
Женская половая система представлена парными яичниками, отходящими от них яйцеводами, желточников, в которых образуются желточные клетки, и
скорлуповых желез, где происходит оплодотворение, окружение яйца желточными клетками с запасом питательных веществ и формирование у яйца твердой оболочки —
скорлупы.
Оплодотворение у планарий перекрестное: в ходе этого процесса две совокупляющиеся особи вводят циррусы (совокупительные органы) в копулятивные сумки друг друга. После оплодотворения яйца покрываются плотными оболочками и образуются «коконы», которые планария прикрепляет к различным подводным предметам.
Развитие планарии прямое, без метаморфоза. Из коконов появляются молодые планарии, похожие на взрослую особь, но значительно меньше ее в размерах.
передвижение | поведение | Британника
Принципы
Для передвижения всем животным необходимы как движущие, так и управляющие механизмы. Разнообразные движущие механизмы животных включают сократительную структуру — в большинстве случаев мышцу — для создания движущей силы. Количество, качество и положение сокращений инициируются и координируются нервной системой: благодаря этой координации ритмические движения придатков или тела вызывают локомоцию.
Животные успешно занимают большую часть огромного количества различных физических сред (экологических ниш) на Земле; однако при обсуждении передвижения эти среды можно разделить на четыре типа: воздушные (включая древесные), водные, роющие (подземные) и наземные.Физические ограничения движения — гравитация и сопротивление — одинаковы в каждой среде: они различаются только степенью. Сила тяжести здесь рассматривается как вес и инерция (сопротивление движению) тела, сопротивление как любая сила, уменьшающая движение. Хотя это и не определения физика, они достаточны для общего понимания сил, препятствующих передвижению животных.
Чтобы противодействовать силе гравитации, которая особенно важна при передвижении по воздуху, земле и земле, все животные, живущие в этих трех средах, развили скелетные системы, поддерживающие их тело и предотвращающие его разрушение.Скелетная система может быть внутренней или внешней и действовать как жесткий каркас или как гибкая гидравлическая (жидкостная) опора.
Чтобы начать движение, воздушные, роющие и наземные животные должны выполнить достаточное количество мышечной работы для преодоления инерции. Водные животные также должны преодолевать инерцию; однако плавучесть воды уменьшает влияние гравитации на движение. В самом деле, поскольку многие водные животные невесомы, т. е. обладают нейтральной плавучестью, вытесняя объем воды, вес которого равен их весу в сухом состоянии, для преодоления инерции требуется небольшая мышечная работа.Но не все водные животные невесомы. Те, у кого отрицательная плавучесть, тонут из-за своего веса; следовательно, чем больше их вес, тем больше мышечной энергии они должны затратить, чтобы оставаться на данном уровне. И наоборот, животное с положительной плавучестью всплывает на поверхность и отдыхает на ней и должно расходовать мышечную энергию, чтобы оставаться под водой.
В воде основной силой, которая замедляет или препятствует движению вперед, является сопротивление, величина которого зависит от формы животного и от того, как эта форма рассекает воду. Сопротивление возникает в основном из-за трения воды, когда она течет по поверхности животного, и прилипания воды к поверхности животного (т. Е. Из-за вязкости воды). Из-за вязкости воды ее течение имеет пластинчатый характер; т. е. разные слои воды текут с разной скоростью, при этом самым медленным является слой течения, примыкающий к поверхности тела. По мере увеличения скорости потока ламеллярный рисунок теряется и развивается турбулентность, что увеличивает сопротивление.
Сопротивление возникает в основном из-за трения воды, когда она течет по поверхности животного, и прилипания воды к поверхности животного (т. Е. Из-за вязкости воды). Из-за вязкости воды ее течение имеет пластинчатый характер; т. е. разные слои воды текут с разной скоростью, при этом самым медленным является слой течения, примыкающий к поверхности тела. По мере увеличения скорости потока ламеллярный рисунок теряется и развивается турбулентность, что увеличивает сопротивление.
Еще одним компонентом сопротивления является замедление движения вперед из-за обратного притяжения водоворотов за хвостом животного.Когда они стекают с животного, слои воды с каждой стороны встречаются и смешиваются. Если животное обтекаемой формы (например, имеет веретенообразную форму), турбулентность низкая; если, однако, слои воды с боков встречаются резко и с разной скоростью, турбулентность высока, вызывая сильное обратное притяжение или сопротивление животного.
Передвижение по воздуху также сталкивается с сопротивлением сопротивления, но поскольку вязкость и плотность воздуха намного меньше, чем у воды, сопротивление также меньше. Однако ламеллярный поток воздуха через поверхности крыла чрезвычайно важен.Восходящая сила полета или подъемная сила возникает из-за того, что воздух течет быстрее по верхней поверхности, чем по нижней поверхности крыла. Поскольку эта разница в потоке создает более низкое давление воздуха на верхней поверхности, животное поднимается. Подъемная сила также создается потоком воды по поверхности, но водные животные используют подъемную силу как средство управления, а не как источник движения.
Однако ламеллярный поток воздуха через поверхности крыла чрезвычайно важен.Восходящая сила полета или подъемная сила возникает из-за того, что воздух течет быстрее по верхней поверхности, чем по нижней поверхности крыла. Поскольку эта разница в потоке создает более низкое давление воздуха на верхней поверхности, животное поднимается. Подъемная сила также создается потоком воды по поверхности, но водные животные используют подъемную силу как средство управления, а не как источник движения.
Сопротивление обычно считается незначительным влиянием на наземное передвижение; а в роющем передвижении трение и плотность (рыхлость) почвы являются двумя основными ограничениями.Если почва чрезвычайно рыхлая, как и песок, некоторые животные могут «плавать» по ней. Однако такое роющее передвижение встречается довольно редко; большинству ископаемых животных приходится с большим трудом прокладывать туннели в почве, и после этого они зависят от туннелей для активного передвижения.
Движение у животных осуществляется двумя типами локомоции: осевой и аппендикулярной. При осевой локомоции, которая включает в себя гидравлический прямоточный метод выброса воды (например, кальмар), создание объемной волны (угорь) или метод сокращения-якоря-вытягивания (пиявка), форма тела изменяется, и взаимодействие все тело с окружающей средой обеспечивает движущую силу.При аппендикулярной локомоции специальные придатки тела взаимодействуют с окружающей средой, создавая движущую силу.
При осевой локомоции, которая включает в себя гидравлический прямоточный метод выброса воды (например, кальмар), создание объемной волны (угорь) или метод сокращения-якоря-вытягивания (пиявка), форма тела изменяется, и взаимодействие все тело с окружающей средой обеспечивает движущую силу.При аппендикулярной локомоции специальные придатки тела взаимодействуют с окружающей средой, создавая движущую силу.
Есть также много видов животных, которые зависят от окружающей среды для передвижения, типа мобильности, называемого пассивным передвижением. У некоторых медуз, например, есть структуры, называемые поплавками, которые возвышаются над поверхностью воды и действуют как паруса. Несколько пауков разработали сложные способы кайтинга; когда нить их паутинного шелка после вытягивания в воздух достигает определенной длины, сопротивления нити ветру достаточно, чтобы унести ее вместе с прикрепленным пауком.У одной рыбы, реморы, спинной плавник переместился на макушку головы и превратился в присоску; прикрепившись к более крупной рыбе, ремора может добраться до следующего приема пищи.
Морфологическая и ультраструктурная карта передних сенсорных ресничек и глии Caenorhabditis elegans с высоким разрешением
Животные должны чувствовать и реагировать на многочисленные сигналы окружающей среды в широком диапазоне интенсивности сигнала. Сложность внешних сигналов частично отражается в удивительном разнообразии морфологий и функций сенсорных нейронов.Многие основные типы сенсорных нейронов содержат первичные реснички на основе микротрубочек (МТ), которые содержат молекулы передачи сигнала и необходимы для сенсорных свойств нейронов (Perkins et al., 1986; Inglis et al., 2007; McEwen et al., 2008; Ramamurthy and Cayouette, 2009; Pifferi et al., 2010). Хотя основные механизмы сборки, основные структуры и общие функции ресничек в высокой степени консервативны, реснички сенсорных нейронов структурно разнообразны и специализированы для выполнения своих уникальных ролей (Berbari et al., 2009; Silverman and Leroux, 2009).Напр., обонятельные нейроны позвоночных содержат пучок из 6-17 ресничек, исходящих из дендритных бугорков и содержащих обонятельные сигнальные белки (Menco, 1980; McEwen et al. , 2008; Pifferi et al., 2010). Сходным образом фоторецепторы палочек и колбочек содержат высокосложные наружные сегменты ресничек, различающиеся по морфологии и локализующие молекулы, необходимые для фототрансдукции (Berbari et al., 2009; Yildiz and Khanna, 2012). Морфология и функция сенсорных ресничек дополнительно регулируются посредством взаимодействия с ненейрональными клетками, такими как глия (Young and Bok, 1969; Strauss, 2005; Bacaj et al., 2008; Проко и др., 2011). Дефекты структуры и функции ресничек приводят к изменению клеточной передачи сигналов и способствуют системным нарушениям, которые в совокупности называются цилиопатиями (Green and Mykytyn, 2010; Louvi and Grove, 2011; Waters and Beales, 2011). Таким образом, анализ архитектуры сенсорных ресничек, а также их организации и взаимодействия с поддерживающими клетками необходим для полного описания и понимания механизмов, лежащих в основе функциональной специализации сенсорной нервной системы.
, 2008; Pifferi et al., 2010). Сходным образом фоторецепторы палочек и колбочек содержат высокосложные наружные сегменты ресничек, различающиеся по морфологии и локализующие молекулы, необходимые для фототрансдукции (Berbari et al., 2009; Yildiz and Khanna, 2012). Морфология и функция сенсорных ресничек дополнительно регулируются посредством взаимодействия с ненейрональными клетками, такими как глия (Young and Bok, 1969; Strauss, 2005; Bacaj et al., 2008; Проко и др., 2011). Дефекты структуры и функции ресничек приводят к изменению клеточной передачи сигналов и способствуют системным нарушениям, которые в совокупности называются цилиопатиями (Green and Mykytyn, 2010; Louvi and Grove, 2011; Waters and Beales, 2011). Таким образом, анализ архитектуры сенсорных ресничек, а также их организации и взаимодействия с поддерживающими клетками необходим для полного описания и понимания механизмов, лежащих в основе функциональной специализации сенсорной нервной системы.
Нематода Caenorhabditis elegans представляет собой превосходную модельную систему для изучения молекулярных и нейронных механизмов, с помощью которых животные обнаруживают сигналы окружающей среды и реагируют на них (например, Bargmann, 2006; Goodman, 2006; Zhang, 2008; Garrity et al. , 2010). Гермафродит C. elegans имеет компактную сенсорную нервную систему, состоящую по крайней мере из 78 нейронов 34 типов, которые различаются по своей морфологии, связям и сенсорным свойствам (Ward et al., 1975; Ware et al., 1975; Перкинс и др., 1986; White et al., 1986) (www.wormatlas.org). Многие из основных хемо-, механо- и термосенсорных нейронов присутствуют в органах чувств, расположенных в голове и хвосте червя; 60 из этих сенсорных нейронов покрыты ресничками (Ward et al., 1975; Ware et al., 1975; Perkins et al., 1986; White et al., 1986; Bargmann, 2006; Inglis et al., 2007) (www. www.wormatlas.org). Как и у других организмов, развитие и поддержание правильной структуры ресничек необходимы для уникальных сенсорных свойств каждого типа нейронов у C.elegans (Dusenbery et al., 1975; Lewis, Hodgkin, 1977; Culotti, Russell, 1978; Perkins et al., 1986). В голове реснитчатые сенсорные нейроны организованы в несколько сенсилл, каждая из которых содержит различные типы сенсорных нейронов и связанные с ними глиальные вспомогательные клетки (Ward et al.
, 2010). Гермафродит C. elegans имеет компактную сенсорную нервную систему, состоящую по крайней мере из 78 нейронов 34 типов, которые различаются по своей морфологии, связям и сенсорным свойствам (Ward et al., 1975; Ware et al., 1975; Перкинс и др., 1986; White et al., 1986) (www.wormatlas.org). Многие из основных хемо-, механо- и термосенсорных нейронов присутствуют в органах чувств, расположенных в голове и хвосте червя; 60 из этих сенсорных нейронов покрыты ресничками (Ward et al., 1975; Ware et al., 1975; Perkins et al., 1986; White et al., 1986; Bargmann, 2006; Inglis et al., 2007) (www. www.wormatlas.org). Как и у других организмов, развитие и поддержание правильной структуры ресничек необходимы для уникальных сенсорных свойств каждого типа нейронов у C.elegans (Dusenbery et al., 1975; Lewis, Hodgkin, 1977; Culotti, Russell, 1978; Perkins et al., 1986). В голове реснитчатые сенсорные нейроны организованы в несколько сенсилл, каждая из которых содержит различные типы сенсорных нейронов и связанные с ними глиальные вспомогательные клетки (Ward et al. , 1975; Ware et al., 1975; Perkins et al., 1986). Многим сенсорным нейронам были приписаны специфические функции (Bargmann, 2006) (www.wormatlas.org), что обеспечивает экспериментальную систему, в которой можно не только сопоставлять структурную и функциональную специализацию, но и подробно описывать степень разнообразия сенсорных нейронов. Морфология ресничек и ультраструктуры, присутствующие у многоклеточных животных.
, 1975; Ware et al., 1975; Perkins et al., 1986). Многим сенсорным нейронам были приписаны специфические функции (Bargmann, 2006) (www.wormatlas.org), что обеспечивает экспериментальную систему, в которой можно не только сопоставлять структурную и функциональную специализацию, но и подробно описывать степень разнообразия сенсорных нейронов. Морфология ресничек и ультраструктуры, присутствующие у многоклеточных животных.
Пионерская работа 1970-х и 1980-х годов предоставила начальное морфологическое и ультраструктурное описание передней сенсорной анатомии C. elegans (Ward et al., 1975; Ware et al., 1975; Perkins et al., 1986). В этих исследованиях для описания структуры и организации органов чувств головы использовалась просвечивающая электронная микроскопия серийных срезов (ssTEM) химически фиксированных животных. В частности, в этих исследованиях описаны морфологически разнообразные реснички, присутствующие на дендритных окончаниях сенсорных типов нейронов в этих сенсиллах (Ward et al. , 1975; Уэр и др., 1975; Перкинс и др., 1986). Например, было показано, что восемь сенсорных нейронов органов амфид содержат структурно простые реснички, которые напрямую контактируют с окружающей средой через «канал», образованный глиальными клетками, тогда как реснички других сенсорных нейронов амфид оказались более сложными и встроены в глиальные клетки. клеточные процессы (Ward et al., 1975; Ware et al., 1975; Perkins et al., 1986).
, 1975; Уэр и др., 1975; Перкинс и др., 1986). Например, было показано, что восемь сенсорных нейронов органов амфид содержат структурно простые реснички, которые напрямую контактируют с окружающей средой через «канал», образованный глиальными клетками, тогда как реснички других сенсорных нейронов амфид оказались более сложными и встроены в глиальные клетки. клеточные процессы (Ward et al., 1975; Ware et al., 1975; Perkins et al., 1986).
Хотя эти исследования послужили основой для обширных дальнейших исследований сенсорной биологии C.elegans , две проблемы побудили нас пересмотреть описание передней сенсорной анатомии дикого типа. Во-первых, достижения в области методов визуализации и сохранения тканей привели к разработке методов и инструментов для ультраструктурной визуализации, которые обеспечивают гораздо более подробные изображения с более высоким разрешением, чем это было возможно ранее (Rostaing et al., 2004; Weimer, 2006; Muller-Reichert et al.). и др., 2008; Холл и др. , 2012). Например, электронная томография с последовательными срезами (ssET) теперь позволяет выполнять трехмерную (3D) реконструкцию структур и выявляет новые особенности благодаря почти изотропному разрешению в рамках реконструкции, а не ограничению в направлении z с помощью толщина пластиковых секций (McEwen and Marko, 2001; Muller-Reichert et al., 2010). Во-вторых, ультраструктурные особенности некоторых сенсорных ресничек остаются плохо описанными, частично из-за ограничений в методах визуализации, но также и из-за проблем, возникающих при обычной химической фиксации биологических структур. Напр., морфология и ультраструктура ресничек в газочувствительных BAG и ноцицептивных нейронах FLP плохо описаны, и паттерн распределения аксонемных МТ в сильно разветвленных ресничках, таких как таковые в хемосенсорных нейронах AWA, не был полностью охарактеризован.
, 2012). Например, электронная томография с последовательными срезами (ssET) теперь позволяет выполнять трехмерную (3D) реконструкцию структур и выявляет новые особенности благодаря почти изотропному разрешению в рамках реконструкции, а не ограничению в направлении z с помощью толщина пластиковых секций (McEwen and Marko, 2001; Muller-Reichert et al., 2010). Во-вторых, ультраструктурные особенности некоторых сенсорных ресничек остаются плохо описанными, частично из-за ограничений в методах визуализации, но также и из-за проблем, возникающих при обычной химической фиксации биологических структур. Напр., морфология и ультраструктура ресничек в газочувствительных BAG и ноцицептивных нейронах FLP плохо описаны, и паттерн распределения аксонемных МТ в сильно разветвленных ресничках, таких как таковые в хемосенсорных нейронах AWA, не был полностью охарактеризован.
Здесь мы предоставляем морфологический и ультраструктурный анализ с высоким разрешением передних окончаний сенсорных нейронов и глии у взрослого гермафродита C. elegans с использованием ssTEM и ssET замороженных и замещенных под высоким давлением (HPF-FS) взрослых животные. Этот анализ теперь позволил нам надежно смоделировать морфологию всех передних сенсорных ресничек, включая 50 из 60 реснитчатых нейронов у взрослых гермафродитов C. elegans . Мы описываем ультраструктуры различных типов ресничек с беспрецедентными подробностями, включая таковые ранее не охарактеризованных ресничек, и описываем новые особенности глиально-ресничных взаимодействий.Мы также характеризуем их межклеточные отношения и идентифицируем важные ультраструктурные особенности, включая МТ, волокна, везикулы и соединения. Более того, эта карта с высоким разрешением обнаруживает три различных и новых процесса распределения MT в разветвленных ресничках, указывая на различные способы транспорта и доставки цилиарных белков. Вместе с обширным анализом путей спецификации клеток, возможных у C. elegans , это полное ультраструктурное описание передней сенсорной анатомии позволит детально понять молекулярные и клеточные механизмы, с помощью которых устанавливается морфологическое разнообразие, а также вклад этого разнообразия к клеточной функциональной специализации.
elegans с использованием ssTEM и ssET замороженных и замещенных под высоким давлением (HPF-FS) взрослых животные. Этот анализ теперь позволил нам надежно смоделировать морфологию всех передних сенсорных ресничек, включая 50 из 60 реснитчатых нейронов у взрослых гермафродитов C. elegans . Мы описываем ультраструктуры различных типов ресничек с беспрецедентными подробностями, включая таковые ранее не охарактеризованных ресничек, и описываем новые особенности глиально-ресничных взаимодействий.Мы также характеризуем их межклеточные отношения и идентифицируем важные ультраструктурные особенности, включая МТ, волокна, везикулы и соединения. Более того, эта карта с высоким разрешением обнаруживает три различных и новых процесса распределения MT в разветвленных ресничках, указывая на различные способы транспорта и доставки цилиарных белков. Вместе с обширным анализом путей спецификации клеток, возможных у C. elegans , это полное ультраструктурное описание передней сенсорной анатомии позволит детально понять молекулярные и клеточные механизмы, с помощью которых устанавливается морфологическое разнообразие, а также вклад этого разнообразия к клеточной функциональной специализации.
Ciliary Rab28 и BBSome негативно регулируют выделение внеклеточных везикул
Реснички представляют собой консервативные органеллы на основе микротрубочек (МТ), которые отходят от поверхности большинства типов эукариотических клеток. Реснички выполняют множество функций, включая подвижность и передачу сигналов, а также способность выделять внеклеточные везикулы (EV) (Wang and Barr, 2018; Carter and Blacque, 2019; Nachury and Mick, 2019). Дефекты первичных ресничек приводят к моносимптомным и синдромальным нарушениям у человека, которые в совокупности называются цилиопатиями.К ним относятся такие заболевания, как аутосомно-доминантная поликистозная болезнь почек (АДПБП), пигментный ретинит, колбочковая дистрофия и синдром Барде-Бидля (СББ) (Waters and Beales, 2011).
Реснички зависят от нескольких способов внутриклеточного транспорта для установления и контроля их молекулярного состава (Jensen and Leroux, 2017). Внутрижгутиковый транспорт (IFT) управляется кинезин-II антероградно (от основания реснички к кончику) и цитоплазматическим динеином 2 ретроградно (от кончика реснички к основанию) моторами (Rosenbaum and Witman, 2002). Грузовые адаптеры, такие как комплексы IFT-A/B и BBSome, позволяют IFT транспортировать структурные и сигнальные белки в реснички и из них (Liem et al., 2012; Bhogaraju et al., 2013; Lechtreck, 2015; Nachury, 2018). . BBSome регулирует сигнальные каскады ресничек, удаляя различные мембранные белки из ресничек с помощью ретроградной IFT (Lechtreck et al., 2009; Liu and Lechtreck, 2018; Ye et al., 2018) и состоит из октамерного комплекса (BBS1/2/4/ 5/7/8/9 и BBS18), который рекрутируется на мембраны небольшим G-белком Arl6 (Liew et al., 2014). Липидированные мембранные белки переносятся в реснички с помощью белков, связывающих пренил- (например, PDE6D) и миристоил- (например, UNC119), и затем высвобождаются из этих челноков с помощью факторов смещения груза, таких как Arl3 (Ismail et al., 2012; Fansa et al. ., 2016; Дженсен и Леру, 2017).
Грузовые адаптеры, такие как комплексы IFT-A/B и BBSome, позволяют IFT транспортировать структурные и сигнальные белки в реснички и из них (Liem et al., 2012; Bhogaraju et al., 2013; Lechtreck, 2015; Nachury, 2018). . BBSome регулирует сигнальные каскады ресничек, удаляя различные мембранные белки из ресничек с помощью ретроградной IFT (Lechtreck et al., 2009; Liu and Lechtreck, 2018; Ye et al., 2018) и состоит из октамерного комплекса (BBS1/2/4/ 5/7/8/9 и BBS18), который рекрутируется на мембраны небольшим G-белком Arl6 (Liew et al., 2014). Липидированные мембранные белки переносятся в реснички с помощью белков, связывающих пренил- (например, PDE6D) и миристоил- (например, UNC119), и затем высвобождаются из этих челноков с помощью факторов смещения груза, таких как Arl3 (Ismail et al., 2012; Fansa et al. ., 2016; Дженсен и Леру, 2017).
Помимо своей роли сигнального узла, ресничка также является эволюционно консервативным местом биогенеза и отщепления внеклеточных везикул (EV) (Wood et al. , 2013; Wang et al., 2014a; Salinas et al., 2017). В большинстве исследований ЭВ использовались культивируемые клетки или биожидкости, и было приложено много усилий для улучшения методологии выделения и характеристики различных подтипов ЭВ с использованием белковых и липидных маркеров. Экзосомы (диаметром 30–150 нм) образуются в результате слияния мультивезикулярных телец. Микровезикулы или эктосомы (100–1000 нм) отпочковываются от плазматической мембраны (Maas et al., 2017; Raposo, Stoorvogel, 2013; Meldolesi, 2018). ЭВ гетерогенны и обычно выделяются в зависимости от размера и плотности и имеют неизвестное биогенное происхождение: доступные маркеры не являются строго специфичными для экзосом или микровезикул/эктосом.Таким образом, обычно используются термины малые ВВ (<200 нм) и большие ВВ (>200 нм), а термин «маленький ВВ» охватывает как экзосомы, так и малые эктосомы/микровезикулы (Théry et al., 2018). Чтобы преодолеть эти проблемы в области EV, мы разработали генетически кодированную систему отслеживания груза EV, меченную флуоресцентным белком, которая позволяет изучать биогенез, выделение и высвобождение цилиарного EV у живых животных (Wang et al.
, 2013; Wang et al., 2014a; Salinas et al., 2017). В большинстве исследований ЭВ использовались культивируемые клетки или биожидкости, и было приложено много усилий для улучшения методологии выделения и характеристики различных подтипов ЭВ с использованием белковых и липидных маркеров. Экзосомы (диаметром 30–150 нм) образуются в результате слияния мультивезикулярных телец. Микровезикулы или эктосомы (100–1000 нм) отпочковываются от плазматической мембраны (Maas et al., 2017; Raposo, Stoorvogel, 2013; Meldolesi, 2018). ЭВ гетерогенны и обычно выделяются в зависимости от размера и плотности и имеют неизвестное биогенное происхождение: доступные маркеры не являются строго специфичными для экзосом или микровезикул/эктосом.Таким образом, обычно используются термины малые ВВ (<200 нм) и большие ВВ (>200 нм), а термин «маленький ВВ» охватывает как экзосомы, так и малые эктосомы/микровезикулы (Théry et al., 2018). Чтобы преодолеть эти проблемы в области EV, мы разработали генетически кодированную систему отслеживания груза EV, меченную флуоресцентным белком, которая позволяет изучать биогенез, выделение и высвобождение цилиарного EV у живых животных (Wang et al. , 2014a; Wang et al. , 2015; Maguire et al., 2015; Silva et al., 2017; O’Hagan et al., 2017).
, 2014a; Wang et al. , 2015; Maguire et al., 2015; Silva et al., 2017; O’Hagan et al., 2017).
EVs, высвобождаемые из кончиков ресничек, явно имеют эктосомное происхождение (Wood et al., 2013; Phua et al., 2017). Было предложено несколько функций цилиарных ВВ, включая передачу сигналов между клетками и/или организмами (Wang et al., 2014a; Cao et al., 2015), удаление отходов (Nager et al., 2017), резорбцию и разборку ресничек (Long et al., 2016; Phua et al., 2017), и морфогенез сложных цилиарных мембран (Salinas et al., 2017). Дефекты биогенеза и высвобождения ЭВ связаны с цилиарной дисфункцией (Wang et al., 2015; Магуайр и др., 2015 г.; Ван и др., 2014а; О’Хаган и др., 2017 г.; Сильва и др., 2017; Дилан и др., 2018). Аномальные фенотипы EV также наблюдались при цилиопатиях, таких как поликистозная болезнь почек (PKD), поликистозная болезнь печени и дегенерация сетчатки (Hogan et al., 2009; Salinas et al., 2017; Dilan et al., 2018; Masyuk et al. ., 2010). Однако взаимосвязь между цилиарными EVs и функцией ресничек остается неизвестной.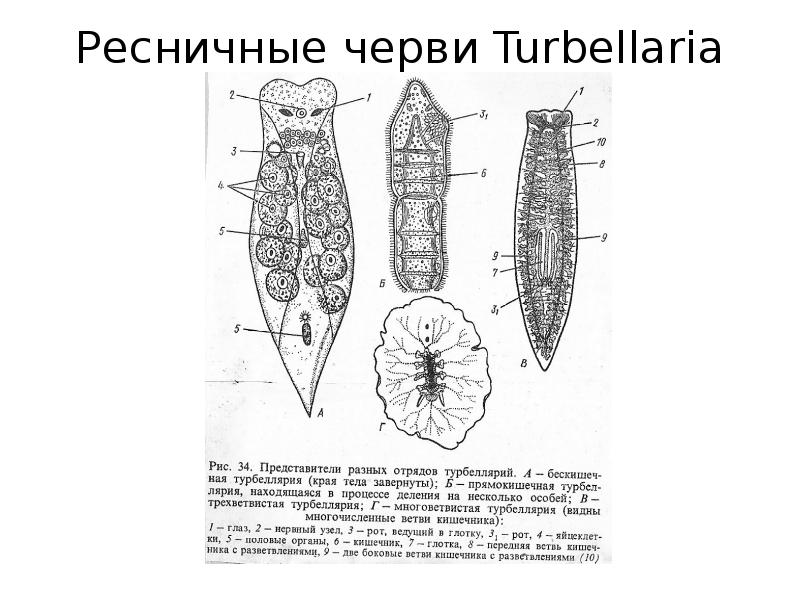
Нематода C. elegans представляет собой превосходную систему in vivo для изучения формирования, специализации и функции ресничек, включая выделение EV (Bargmann, 2006).У C. elegans сенсорных ресничек присутствуют на кончиках дендритов сенсорных нейронов, причем большинство из них находится в сенсорных органах (сенсиллах). Большинство этих органов подвергается воздействию окружающей среды через кутикулярные поры, происходящие из окружающих отростков глиальных клеток (Singhvi and Shaham, 2019). Из 302 нейронов, составляющих нервную систему гермафродита, 60 являются реснитчатыми (Ward et al., 1975; Perkins et al., 1986; Doroquez et al., 2014). C. elegans самцов обладают дополнительными 54 реснитчатыми нейронами (Sulston et al., 1980; Кук и др., 2019 г.; Молина-Гарсия и др., 2019). Реснички C. elegans обладают схожим базовым структурным планом, простирающимся от дегенеративного базального тельца до переходной зоны (TZ), и аксонемной структурой, которая демонстрирует огромное разнообразие с точки зрения их расположения МТ между различными типами клеток (Perkins et al. ., 1986; Doroquez et al., 2014; Silva et al., 2017; Akella et al., 2019). На самом дистальном конце дендрита компартмент перицилиарной мембраны (PCMC) действует как транспортный узел для сортировки белков в ресничку и из нее (Kaplan et al., 2012; Нечипуренко и др., 2017; Сервас и др., 2017).
., 1986; Doroquez et al., 2014; Silva et al., 2017; Akella et al., 2019). На самом дистальном конце дендрита компартмент перицилиарной мембраны (PCMC) действует как транспортный узел для сортировки белков в ресничку и из нее (Kaplan et al., 2012; Нечипуренко и др., 2017; Сервас и др., 2017).
Цефалические мужские (CEM) и общие внутренние губные (IL2) нейроны являются единственными известными реснитчатыми EV-высвобождающими нейронами (EVN), как определено с помощью визуализации грузов EV с флуоресцентной меткой, высвобождаемых вне животного (Wang et al., 2014a). Эти выпущенные из окружающей среды электромобили способствуют общению между животными (Wang et al., 2014a; Silva et al., 2017). У C. elegans EVs «сбрасываются» у основания ресничек и остаются в просветах головных и внутренних губных органов чувств, что визуализируется с помощью просвечивающей электронной микроскопии (TEM) и томографических реконструкций.ЭВ также «высвобождаются» из ресничек непосредственно в окружающую среду за пределами сенсорных органов, что наблюдается с помощью флуоресцентно-меченых грузов ЭВ, таких как полицистин TRP PKD-2::GFP и миристоилированный белок периферической мембраны CIL-7 (Wang et al.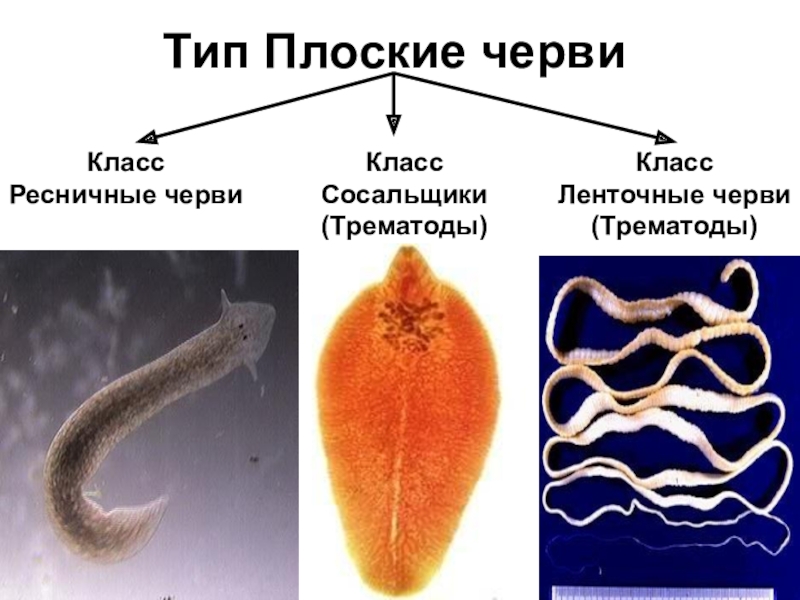 , 2014a; Магуайр и др., 2015). Высвобождение цилиарного EV контролируется авторами, ластиками и ридерами тубулинового кода (Wang et al., 2014a; Wang et al., 2015; Maguire et al., 2015; Silva et al., 2017; O’Hagan et al., 2017). У этих мутантов EVs сбрасываются в основании ресничек и накапливаются в просветах органов чувств, но не выделяются в окружающую среду.В отличие от цилиарного механизма высвобождения EV, механизм шеддинга основания ресничного EV остается плохо охарактеризованным.
, 2014a; Магуайр и др., 2015). Высвобождение цилиарного EV контролируется авторами, ластиками и ридерами тубулинового кода (Wang et al., 2014a; Wang et al., 2015; Maguire et al., 2015; Silva et al., 2017; O’Hagan et al., 2017). У этих мутантов EVs сбрасываются в основании ресничек и накапливаются в просветах органов чувств, но не выделяются в окружающую среду.В отличие от цилиарного механизма высвобождения EV, механизм шеддинга основания ресничного EV остается плохо охарактеризованным.
Малые G-белки, в том числе белки семейства Rab, регулируют формирование и функцию ресничек, а также биогенез ЭВ (Li and Hu, 2011; Blacque et al., 2018; Knödler et al., 2010; Blanc and Vidal, 2018; Meldolesi). , 2018). Здесь мы сосредоточимся на Rab28, который связан с аутосомно-рецессивной колбочково-стержневой дистрофией (arCRD) (Roosing et al., 2013; Riveiro-Alvarez et al., 2015; Lee et al., 2017). Дегенерация фоторецепторов также обнаруживается при многих цилиопатиях, включая синдром Барде-Бидля (Waters and Beales, 2011). В ресничках C. elegans амфид и фазмид RAB-28 является BBS-8-зависимым IFT-грузом, который связывается с перицилиарной мембраной (PCM) в конформации, связанной с GTP (Jensen et al., 2016). Функционально нарушение C. elegans RAB-28 приводит к дефектам в размере сенсорного компартмента амфид посредством предполагаемого неавтономного клеточного механизма, который, как ранее предполагалось, включает ЭВ (Jensen et al., 2016). Связанная с EV функция RAB-28 также подтверждается его значительной гиперпредставленностью в транскриптоме C. elegans EVN (Wang et al., 2015). Играет ли RAB-28 роль в биологии EV, и как RAB-28 функционирует в ресничках, неизвестно.
В ресничках C. elegans амфид и фазмид RAB-28 является BBS-8-зависимым IFT-грузом, который связывается с перицилиарной мембраной (PCM) в конформации, связанной с GTP (Jensen et al., 2016). Функционально нарушение C. elegans RAB-28 приводит к дефектам в размере сенсорного компартмента амфид посредством предполагаемого неавтономного клеточного механизма, который, как ранее предполагалось, включает ЭВ (Jensen et al., 2016). Связанная с EV функция RAB-28 также подтверждается его значительной гиперпредставленностью в транскриптоме C. elegans EVN (Wang et al., 2015). Играет ли RAB-28 роль в биологии EV, и как RAB-28 функционирует в ресничках, неизвестно.
Здесь мы анализируем транспортный путь, который регулирует локализацию C. elegans RAB-28 в не-EVN (амфидные/фазмидные нейроны) и ресничках EVN. Мы обнаружили, что ассоциация PCM RAB-28 зависит от BBSome и, в меньшей степени, от его регулятора ARL-6, в то время как для его транспорта IFT требуются BBSome, PDL-1 (ортолог PDE6D) и ARL-3. Мы также выявляем клеточно-специфические различия в том, как развернут транспортный механизм RAB-28, при этом PDL-1 незаменим для поддержания уровней PCM RAB-28 в амфидных/фазмидных нейронах, но не в EVN. Функциональный анализ показал, что мутанты BBSome фенокопируют дефекты морфогенеза сенсорного компартмента амфид, наблюдаемые у червей с нарушением RAB-28. Важно отметить, что потеря генов RAB-28 или BBSome, но не PDL-1, вызывает эктопическое накопление EV в головном сенсорном органе EVN, указывая на то, что RAB-28 и BBSome отрицательно регулируют биогенез EV и/или выделение EV в PCM.Это исследование выявило сеть BBSome-ARL-6-PDL-1 для нацеливания RAB-28, критического регулятора EV, на специфические мембраны ресничек, что позволяет регулировать EV в разных местах. Члены этой транспортной сети, а также RAB-28 и EV-груз PKD-2 представляют собой хорошо зарекомендовавшие себя цилиарные белки, связанные с несколькими генетическими заболеваниями человека (Waters and Beales, 2011; Reiter and Leroux, 2017).
Мы также выявляем клеточно-специфические различия в том, как развернут транспортный механизм RAB-28, при этом PDL-1 незаменим для поддержания уровней PCM RAB-28 в амфидных/фазмидных нейронах, но не в EVN. Функциональный анализ показал, что мутанты BBSome фенокопируют дефекты морфогенеза сенсорного компартмента амфид, наблюдаемые у червей с нарушением RAB-28. Важно отметить, что потеря генов RAB-28 или BBSome, но не PDL-1, вызывает эктопическое накопление EV в головном сенсорном органе EVN, указывая на то, что RAB-28 и BBSome отрицательно регулируют биогенез EV и/или выделение EV в PCM.Это исследование выявило сеть BBSome-ARL-6-PDL-1 для нацеливания RAB-28, критического регулятора EV, на специфические мембраны ресничек, что позволяет регулировать EV в разных местах. Члены этой транспортной сети, а также RAB-28 и EV-груз PKD-2 представляют собой хорошо зарекомендовавшие себя цилиарные белки, связанные с несколькими генетическими заболеваниями человека (Waters and Beales, 2011; Reiter and Leroux, 2017). Мы предполагаем, что нарушение регуляции цилиарной продукции EV может быть фактором патологии некоторых цилиопатий (Wang and Barr, 2016).
Мы предполагаем, что нарушение регуляции цилиарной продукции EV может быть фактором патологии некоторых цилиопатий (Wang and Barr, 2016).
Radiolar Глаза серпулидных червей (Annelida, Serpulidae): структуры, функция и фототранспределение
ABILDGARD, PC 1789. BESCHREIBUNG EINER TORSEN SEEBLASE ( HOLOTHURIA PRAPUS LINN.) ZWEN ARTEN DES STEINBORERS ( TEREBELLA LINN. ), einer Grossen Sandröhre ( Sabella Linn.). шр. Гэс. Натурфр. Фрейнде Берл. 9: 133–146.
Эндрюс, Э. А. 1891. Сложные глаза кольчатых червей. Дж. Морфол . 5: 271–299.
Арендт, Д., К. Тессмар-Райбл, Х. Снайман, А. В. Доррестейн и Дж. Виттбродт. 2004. Цилиарные фоторецепторы с опсином позвоночного типа в мозге беспозвоночных. Наука 306: 869–871.
Одуэн, Дж. В., и Х. Милн Эдвардс. 1834. Аннелидес , pt. 1. Recherches pour servir à l’Historie naturelle du litoral de la France, ou Recueil de memoires sur l’anatomie, la physiologie, la classification et les moeurs des animaux des nos cotes; ouvrage accompagné de planches faites d’après nature , vol.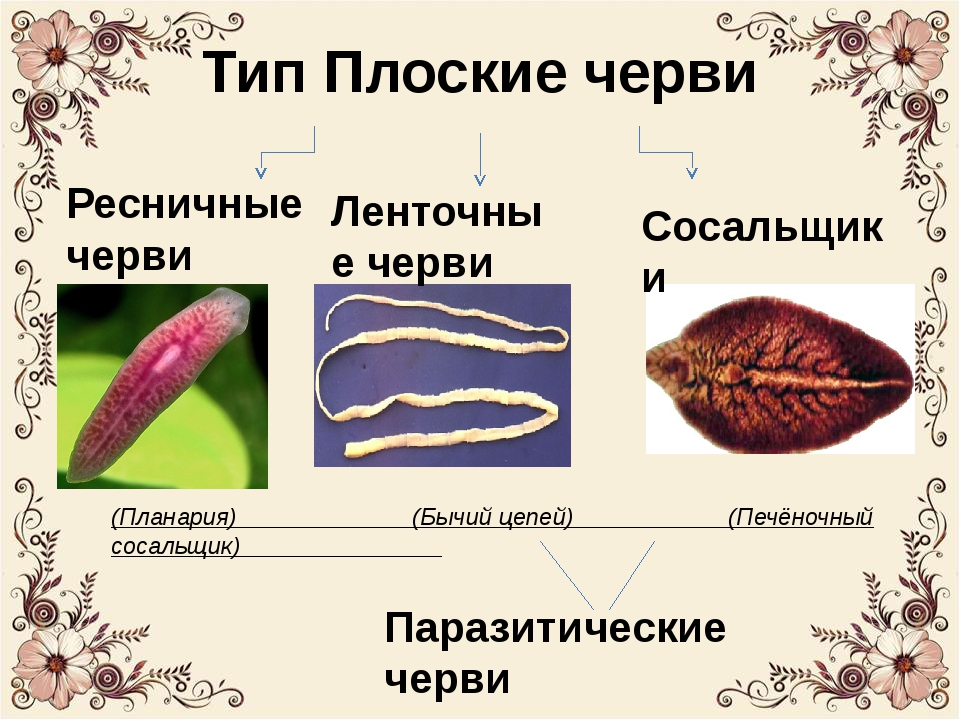 2. Крошар, Париж.
2. Крошар, Париж.
Baird, W. 1865. О новых трубчатых кольчатых червях, в коллекции Британского музея, pt. 2. Зоол. Дж. Линн. соц. 8: 157–160.
Бен-Элиаху, М. Н. и Х. А. тен Хоув. 2011. Serpulidae (Annelida: Polychaeta) из Суэцкого канала — с точки зрения лессепской миграции (монография). Zootaxa 2848: 1–147.
Бок, М. Дж.и Д.-Э. Нильссон. 2016. Глаза веерного червя. Курс. биол. 26: Р907–Р908.
Бок, М.Дж., М.Л. Портер, А.Р. Плейс и Т.В. Кронин. 2014. Биологические солнцезащитные средства настраивают полихроматическое ультрафиолетовое зрение креветок-богомолов. Курс. биол. 24: 1636–1642.
Бок, М. Дж., М. Капа и Д.-Э. Нильссон. 2016. Здесь, там и везде: радиолярные глаза веерных червей (Annelida, Sabellidae). Интегр. Комп. биол. 56: 784–795.
Бок, М. Дж., М.Л. Портер и Д.-Э. Нильссон. 2017. Фототрансдукция в радиолярных глазах веерных червей. Курс. биол. 27: R698–R699.
Дж., М.Л. Портер и Д.-Э. Нильссон. 2017. Фототрансдукция в радиолярных глазах веерных червей. Курс. биол. 27: R698–R699.
Бринкманн Н. и А. Ваннингер. 2009. Нейрогенез предполагает независимую эволюцию покрышек у серпулидных полихет. BMC Evol. биол. 9: 270.
Капа, М.и А. Мюррей. 2009. Обзор рода Megalomma (Polychaeta: Sabellidae) в Австралии с описанием трех новых видов, новыми записями и примечаниями о некоторых особенностях, имеющих филогенетические последствия. Рек. Ауст. Мус. 61: 201–224.
Капа М., П. Хатчингс, М. Тереза Агуадо и Н. Дж. Ботт. 2011. Филогения Sabellidae (Annelida) и отношения с другими таксонами, выведенные из морфологии и множества генов. Кладистика 27: 449–469.
Cavanaugh, GM 1956. Формулы и методы IV Морской биологической лаборатории Химический кабинет . Морская биологическая лаборатория, Вудс-Хоул, Массачусетс.
Морская биологическая лаборатория, Вудс-Хоул, Массачусетс.
Claparède, E. 1868. Les Annélides Chétopodes du Golfe de Naples, Première et Séconde party . Рамбо и Шухардт, Женева и Базель.
Cronin, T.W. 1985. Зрительный пигмент ротоногих ракообразных, Squilla empusa . Дж. Комп. Физиол. А 156: 679–687.
Кронин Т.В. и М.Дж. Бок. 2016. Фоторецепция и зрение в ультрафиолете. Дж. Экспл. биол. 219: 2790–2801.
Кронин Т. В. и Н. Дж. Маршалл. 1989. Множественные спектральные классы фоторецепторов в сетчатке гонодактилоидных ротоногих ракообразных. Дж. Комп. Физиол. А 166: 261–275.
Дэй, Дж.H. 1955. Polychaeta Южной Африки, pt. 3. Оседлые виды с берегов мыса и эстуариев. Зоол. Дж. Линн. соц. 42: 407–452.
ДеВантье, Л. М., Р. Э. Райхельт и Р. Х. Брэдбери. 1986. Защищает ли Spirobranchus giganteus хозяина Porites от хищничества со стороны Acanthaster planci : давление хищников как механизм совместной эволюции? Мар. Экол. прог. сер. 32: 307–310.
Брэдбери. 1986. Защищает ли Spirobranchus giganteus хозяина Porites от хищничества со стороны Acanthaster planci : давление хищников как механизм совместной эволюции? Мар. Экол. прог. сер. 32: 307–310.
Dragesco-Kernéis, A. 1980. Taches oculaires segmentaires chez Dasychone (Annélides Polychètes) etude ultrastructurale. Ках. биол. 21 марта: 287–302.
Ермак, Т. Х. и Р. М. Икин. 1976. Тонкое строение мозговых и пигидиальных глазков у Chone ecaudata (Polychaeta: Sabellidae). Дж. Ультраструктур. Рез. 54: 243–260.
Фейн Г.Л., Р. Харди и С. Б. Лафлин. 2010. Фототрансдукция и эволюция фоторецепторов. Курс. биол. 20: Р114–Р124.
Фовель, П. 1927. Polychètes se’dentaires. Addenda aux errantes, Archiannelides, Myzostomaires. Фавн о. 16: 1–494.
Fauvel, P. 1953. Фауна Индии, Пакистана, Цейлона, Бирмы и Малайи: Annelida polychaeta. Индийская пресса, Аллахабад.
1953. Фауна Индии, Пакистана, Цейлона, Бирмы и Малайи: Annelida polychaeta. Индийская пресса, Аллахабад.
Фишер, А. 1963. Über den Bau und die Hell-Dunkel-Adaptation der Augen des Polychäten Platynereis dumerilii . Сотовый. Ткань Res. 61: 338–353.
Гарм А. и Д.-Э. Нильссон. 2014. Визуальная навигация у морских звезд: первые доказательства использования зрения и глаз у морских звезд. Проц. Р. Соц. биол. науч. B 281: 20133011.
Грубе А.E. 1862. Mittheilungen ueber die Serpulen, mit besonderer Beruecksichtigungihrer Deckel. часов. Гэс. Бреслау Яресбер. 39: 53–69.
Grube, AE 1878. Annulata Semperiana: Beiträge zur Kenntnis der Annelidenfauna der Philippinen, nach den von Herrn Prof. Semper mitgebrachten Sammlungen. Мем. акад. Имп. науч. Санкт-Петербургб. сер. 7 25: 1–300.
Hartline, H. K. 1938. Выделение импульсов в зрительном нерве Pecten в ответ на освещение глаза. Дж. Сотовый. Комп. Физиол. 11: 465–478.
K. 1938. Выделение импульсов в зрительном нерве Pecten в ответ на освещение глаза. Дж. Сотовый. Комп. Физиол. 11: 465–478.
Hesse, R. 1899. Untersuchungen über die Organe der Lichtempfindung bei niederen Tieren. V. Die Augen der polychäten Anneliden. З. Висс. Зоол. 65: 446–516.
Имадзима М. и О. Хартман. 1964. Многощетинковые кольчатые черви Японии. Оккас. Пап. Аллан Хэнкок найден. 26: 1–452.
Имадзима, М.и Х. А. тен Хоув. 1984. Serpulinae (Annelida, Polychaeta) с островов Трук, Понапе и атолла Маджуро, с некоторыми другими новыми находками в Индо-Тихоокеанском регионе. Проц. Япония. соц. Сис. Зоол. 27: 35–66.
Iroso, I. 1921. Revisione dei Serpulidi e Sabellidi del Golfo di Napoli. Опубл. стн. Зоол. Неаполь 3: 47–91.
Като К. и Д. М. Стэндли. 2014. MAFFT: итеративное уточнение и дополнительные методы. Методы Мол. биол. 1079: 131–246.
MAFFT: итеративное уточнение и дополнительные методы. Методы Мол. биол. 1079: 131–246.
Кирс, М., Р. Мойр, А. Уилсон, С. Стоунз-Хавас, М. Ченг, С. Старрок, С. Бакстон, А. Купер, С. Марковиц, К. Дюран и др. . 2012 . Geneious Basic: интегрированная и расширяемая настольная программная платформа для организации и анализа данных о последовательностях. Биоинформатика 28: 1647–1649.
Куприянова Е.К. и Т.А. Макдональд.2006. Филогенетические отношения внутри Serpulidae (Sabellida, Annelida), полученные на основе молекулярных и морфологических данных. Зоол. Скр. 35: 421–439.
Куприянова Е.К., Ниши Э. 2010. Serpulidae (Annelida, Polychaeta) с подводных гор Паттон-Мюррей, залив Аляска, северная часть Тихого океана. Zootaxa 2665: 51–68.
Куприянова Е.К. и Г.В. Роуз. 2008. Еще один пример парафилии у Annelida: молекулярные доказательства того, что Sabellidae содержит Serpulidae. Мол. Филогенет. Эвол. 46: 1174–1181.
Мол. Филогенет. Эвол. 46: 1174–1181.
Куприянова, Э. К., Ю. Сун, Х. А. тен Хоув, Э. Вонг и Г. В. Роуз. 2015. Serpulidae (Annelida) острова Лизард, Большой Барьерный риф, Австралия. Zootaxa 4019: 275–353.
Лангмид Б. и С. Л. Зальцберг. 2012. Быстрое выравнивание с промежутками чтения с помощью Bowtie 2. Nat. Методы 9: 357–359.
Лоуренс П.А., Красне Ф.Б. 1965. Цилиарные фоторецепторы кольчатых червей. Наука 148: 965–966.
Лерке Дж., Х. А. тен Хоув, Т. А. Макдональд, Т. Бартоломеус и К. Блейдорн. 2007. Филогенетические взаимоотношения Serpulidae (Annelida: Polychaeta), основанные на данных о последовательности 18S рДНК, и значение для эволюции глазных яблок. Орг. Дайверы. Эвол. 7: 195–206.
Leutscher-Hazelhoff, J. T.1984. Цилиарные клетки развились для гиперполяризации зрения — почему? Naturwissenschaften 71: 213–214.
Li, L., M.J. Connors, M. Kolle, G.T. England, D.I. Speiser, X. Xiao, J. Aizenberg и C. Ortiz. 2015. Многофункциональность хитонового биоминерализованного доспеха со встроенной зрительной системой. Наука 350: 952–956.
Mak, PMS 1982. Полихеты, связанные с кораллами Гонконга, с особой ссылкой на серпулидов.Стр. 596–617 в Труды первого Международного семинара по морской биологии: Морская флора и фауна Гонконга и Южного Китая , Б. С. Мортон и С. К. Ценг, ред. Издательство Гонконгского университета, Гонконг.
Миллер, М. А., В. Пфайффер и Т. Шварц. 2010. Создание научного портала CIPRES для построения больших филогенетических деревьев. Семинар по шлюзовым вычислительным средам (GCE), 2010 г., Новый Орлеан, Луизиана. [Онлайн]. Цифровая библиотека IEEE Xplore.Доступно: http://ieeexplore.ieee.org/document/5676129/ [2017, 26 сентября].
Monro, C.C.A. 1933. Polychaeta Sedentaria, собранная доктором К. Кросслендом в Колоне в районе Панамы и на Галапагосских островах во время экспедиции S.Y. Святой Георгий. Проц. Зоол. соц. Лонд. 2: 1039–1092.
Кросслендом в Колоне в районе Панамы и на Галапагосских островах во время экспедиции S.Y. Святой Георгий. Проц. Зоол. соц. Лонд. 2: 1039–1092.
Монтегю, Г. 1803. Testacea Britannica или Естественная история британских раковин, морских, наземных и пресноводных, включая самые мелкие: систематически расположенные и украшенные рисунками .Дж. Уайт, Лондон.
Montagu, G. 1813. Описания нескольких новых или редких животных, преимущественно морских, обнаруженных на южном побережье Девоншира. Пер. Линн. соц. Лонд. 11: 1–26.
Mörch, OAL 1863. Revisio critica Serpulidarum. Et Bidrag til Rørormenes Naturhistorie. Naturhistorisk Tidsskrift København 1: 347–470.
Никол, Дж.A. C. 1948. Гигантские аксоны кольчатых червей. Q. Rev. Biol. 23: 291–323.
Nicol, J. A. C. 1950. Ответы Branchiomma vesiculosum (Montagu) на световую стимуляцию. J. Mar. Biol. доц. Великобритания 29: 303–320.
J. Mar. Biol. доц. Великобритания 29: 303–320.
Нильссон Д.-Э. 1994. Глаза как оптические сигнальные системы у веерных червей и анадаров. Филос. Транс. Р. Соц. биол. науч. В 346: 195–212.
Нильссон Д.-Э. 2009. Эволюция глаз и зрительного поведения. Филос. Транс. Р. Соц. биол. науч. В 364: 2833–2847.
Нильссон Д.-Э. 2013. Эволюция глаза и его функциональные основы. Виз. Неврологи. 30: 5–20.
Orrrhage, L. 1980. О строении и гомологах переднего конца полихет семейств Sabellidae и Serpulidae. Зооморфология 96: 113–168.
Оррейдж Л. и М.К.М. Мюллер. 2005. Морфология нервной системы Polychaeta (Annelida). Hydrobiologia 535: 79–111.
Pallas, PS 1766. Miscellanea Zoologica: quibus novae imprimis atque obscurae animalium, виды, описанные и наблюдательные, иконибусные, иллюстрированные . Петрум ван Клиф, Hagae Comitum.
Петрум ван Клиф, Hagae Comitum.
Пезнер А.К., А. Р. Лим, Дж. Дж. Канг, Т. К. Армента и Д. Т. Блюмштейн. 2017. Скрывающееся поведение елочных червей в разных временных масштабах. Поведение. Экол. 28: 154–163.
Pillai, T.G. 1971. Исследования коллекции морских и солоноватоводных многощетинковых кольчатых червей семейства Serpulidae с Цейлона. Цейлон J. Sci. биол. науч. 9: 88–130.
Pixell, H.L.M. 1913. Polychaeta семейств Serpulidae и Sabellidae, собранные Шотландской национальной антарктической экспедицией. Пер. Р. Соц. Эдинб. 49: 347–358.
Портер, М. Л., Дж. Р. Бласик, М. Дж. Бок, Э. Г. Кэмерон, Т. Прингл, Т. В. Кронин и П. Р. Робинсон. 2012. Проливая новый свет на эволюцию опсинов. Проц. Р. Соц. Лонд. биол. науч. В 279: 3–14.
Прентисс, Н. К., К. Василеяду, С. Фолветтер, К. Арванитидис и Х.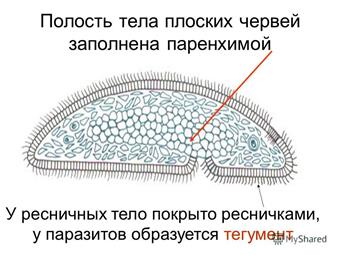 А. тен Хоув. 2014. Новый род и вид Serpulidae (Annelida, Polychaeta, Sabellida) из Карибского моря. Zootaxa 3900: 204–222.
А. тен Хоув. 2014. Новый род и вид Serpulidae (Annelida, Polychaeta, Sabellida) из Карибского моря. Zootaxa 3900: 204–222.
Purschke, G. 2005. Органы чувств полихет (Annelida). Стр. 53–78 в Морфология, молекулы, эволюция и филогения полихет и родственных таксонов , Т. Бартоломеус и Г. Пуршке, ред. Спрингер, Берлин.
Пуршке Г., Д. Арендт, Х. Хаузен и М.К.М. Мюллер. 2006. Фоторецепторные клетки и глаза кольчатых червей. Структура членистоногих. Дев. 35: 211–230.
Радл, Э. 1912. Neue Lehre vom Zentralen Nervensystem . Энгельманн, Лейпциг.
Рамирес, М. Д., А. Н. Пайретт, М. С. Панки, Дж. М. Серб, Д. И. Спайзер, А. Дж. Сваффорд и Т. Х. Окли. 2016. Последний общий предок большинства билатеральных животных обладал как минимум девятью опсинами. Геном Биол. Эвол. 8 : 3640–3652.
Reynolds, E. S. 1963. Использование цитрата свинца при высоком pH в качестве электронно-непрозрачного красителя в электронной микроскопии. Дж. Сотовый. биол. 17 : 208–212.
S. 1963. Использование цитрата свинца при высоком pH в качестве электронно-непрозрачного красителя в электронной микроскопии. Дж. Сотовый. биол. 17 : 208–212.
Rioja, E. 1931. Estudio de los poliquetos de la Península Ibérica. Мем. Р. акад. Cиенц. 2: 1–472.
Rullier, F. 1948. La vision et l’habitude chez Mercierella enigmatica Fauvel. Бык. лаборатория Марит. Динар 30: 21–27.
Schmarda, L.K. 1861. Neue Wirbellose Thiere: Beobachted und Gesammelt auf einer Reise um die Erdr 1853 bis 1857 , I, 03den 0 TurbellariundenВ. Энгельманн, Лейпциг.
Смит, Р. К. и К. С. Бейкер. 1981. Оптические свойства чистейших природных вод (200–800 нм). Заяв. Опц. 20: 177–184.
Смит, Р. С. 1984a. Развитие и расселение Spirobranchus giganteus (Polychaeta: Serpulidae). Стр. 461–483 в Трудах Первой международной конференции по полихетам , П.А. Хатчингс, изд. Линнеевское общество Нового Южного Уэльса, Сидней.
Стр. 461–483 в Трудах Первой международной конференции по полихетам , П.А. Хатчингс, изд. Линнеевское общество Нового Южного Уэльса, Сидней.
Смит, Р. С. 1984b. Новые ассоциации органелл в фоторецепторах серпулидного многощетинкового червя. Tissue Cell 16: 951–956.
Smith, R.S. 1985. Фоторецепторы серпулидных полихет. Кандидат наук. диссертация, Университет Джеймса Кука, Таунсвилл, Австралия.
Спайзер, Д. И. и С. Джонсен. 2008а. Сравнительная морфология вогнутых зеркальных глаз гребешков (Pectinoidea). утра. Малакол. Бык. 26: 27–33.
Спайзер, Д. И. и С. Джонсен. 2008б. Гребешки визуально реагируют на размер и скорость виртуальных частиц. Дж. Экспл. биол. 211: 2066–2070.
Спайзер, Д. И., Д. Дж. Эрнисс и С. Джонсен. 2011. Хитон использует арагонитовые линзы для формирования изображений.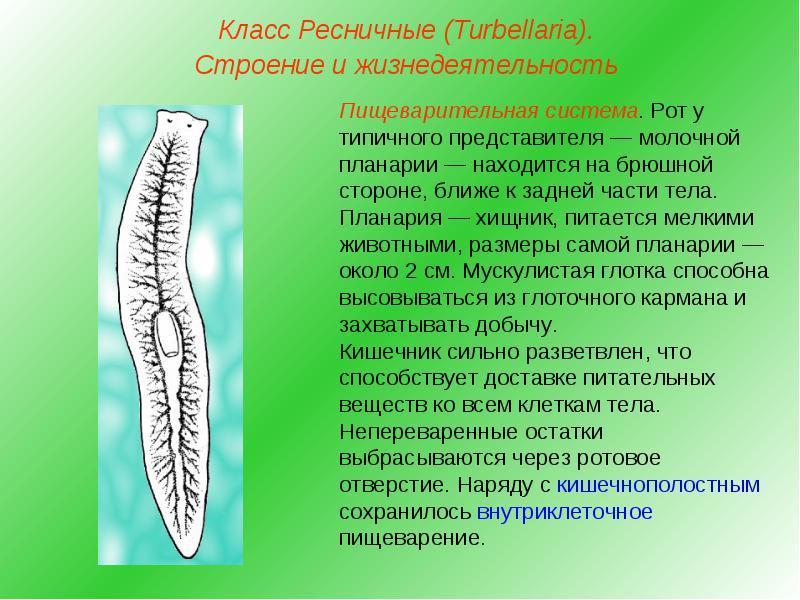 Курс. биол. 21: 665–670.
Курс. биол. 21: 665–670.
Спайзер, Д.I., M.S. Pankey, A.K. Zaharoff, B.A. Battelle, H.D. Bracken-Grissom, J.W. Breinholt, S.M. Bybee, T.W. Cronin, A.Garm, A.R. Lindgren et al. . 2014. Использование филогенетически обоснованной аннотации (PIA) для поиска взаимодействующих со светом генов в транскриптомах немодельных организмов. BMC Биоинформатика 15: R114–R126.
Stamatakis, A. 2006. RAxML-VI-HPC: филогенетический анализ на основе максимального правдоподобия с тысячами таксонов и смешанных моделей. Биоинформатика 22: 2688–2690.
Ставенга Д.Г., Р.П. Смитс и Б.Дж. Хендерс. 1993. Простые экспоненциальные функции, описывающие полосы поглощения спектров зрительного пигмента. Виз. Рез. 33: 1011–1017.
Straughan, D. 1967. Marine Serpulidae (Annelida: Polychaeta) из восточного Квинсленда и Нового Южного Уэльса. австр. Дж. Зул. 15: 201–261.
австр. Дж. Зул. 15: 201–261.
Вс, С.-Ю., Д.-Г. Луо, А. Теракита, Ю. Шичида, Х.-В. Ляо, М.А. Казми, Т.П. Сакмар и К.-В. Яу. 2006. Компоненты фототрансдукции теменного глаза и их потенциальные эволюционные последствия. Наука 311: 1617–1621.
тен Хоув Х.А. и Куприянова Е.К. 2009. Таксономия Serpulidae (Annelida, Polychaeta): состояние дел. Zootaxa 2036: 1–126.
тен Хоув, Х.А. и Ф. Дж. А. Пантус. 1985. Различение родов Apomatus Philippi, 1844 и Protula Risso, 1826 (Polychaeta: Serpulidae): еще один призыв к методическому подходу к таксономии серпулид. Зоол. Мед. 59: 419–437.
Цукамото Х., И. С. Чен, Ю. Кубо и Ю. Фурутани. 2017. Ресничный опсин в мозгу морского кольчатого зоопланктона чувствителен к УФ-излучению, и чувствительность регулируется одним аминокислотным остатком. Дж. Биол. хим. 292: 12971–12980.
хим. 292: 12971–12980.
Uchida, H. 1978. Серпулидные трубчатые черви (Polychaeta, Sedentaria) из Японии с систематическим обзором группы. Бык. Мар Парк Рез. стн. 2: 1–98.
Ульрих-Лютер, Э. М., С. Дюпон, Э. Арболеда, Х. Хаузен и М. И. Арноне. 2011. Уникальная система фоторецепторов в трубчатых ножках морского ежа. Проц. Натл. акад. науч. США 108: 8367–8372.
Вайн, П. Дж. и Дж. Х. Бейли-Брок. 1984. Таксономия и экология трубчатых червей коралловых рифов (Serpulidae, Spirorbidae) в Суданском Красном море. Зоол. Дж. Линн. соц. 80: 135–156.
Vöcking, O. 2016. Ларвальные фоторецепторы моллюска Leptochiton asellus . Кандидат наук. диссертация, Бергенский университет, Норвегия.
Уолд Г. и С. Рэйпорт. 1977. Зрение кольчатых червей. Наука 196: 1434–1439.
ЧЕРВЯКИ. 2017. Всемирный регистр морских видов. [Онлайн]. Доступно: http://www.marinespecies.org [2017, 24 октября].
2017. Всемирный регистр морских видов. [Онлайн]. Доступно: http://www.marinespecies.org [2017, 24 октября].
Йеррамилли Д. и С. Джонсен. 2010. Пространственное зрение пурпурного морского ежа Strongylocentrotus purpuratus (Echinoidea). Дж. Экспл. биол. 213: 249–255.
Zibrowius, H. 1969. Quelques nouvelles recoltes de Serpulidae (Polychaeta Sedentaria) dans le Golfe de Gabes et en Tripolitaine.Описание Vermiliopsis pomatostegoides n. сп. Бык. Инст. Натл. науч. Технол. Океаногр. Пеше 1: 123–137.
Сигнальные и связанные с ресничками нарушения
Wheatley DN . Первичные реснички в нормальных и патологических тканях. Патобиология 1995; 63 :222–238.
КАС Google Scholar
Уитли Д.Н., Ван А.М., Страгнелл Г.Э.Экспрессия первичных ресничек в клетках млекопитающих. Cell Biol Int 1996; 20 :73–81.
Cell Biol Int 1996; 20 :73–81.
КАС Google Scholar
Преториус HA, Spring KR . Физиологический вид первичной реснички. Annu Rev Physiol 2004.
Corey DP, Garcia-Anoveros J, Holt JR, et al. TRPA1 является кандидатом на механочувствительный трансдукционный канал волосковых клеток позвоночных. Природа 2004; 432 :723–730.
КАС Google Scholar
Пазур Г.Дж., Розенбаум Д.Л. Внутрижгутиковый транспорт и реснично-зависимые заболевания. Trends Cell Biol 2002; 12 :551–555.
КАС Google Scholar
Пазур Г.Дж. Внутрижгутиковый транспорт и заболевание почек, зависящее от ресничек: цилиарная гипотеза поликистоза почек. J Am Soc Нефрол 2004; 15 :2528–2536.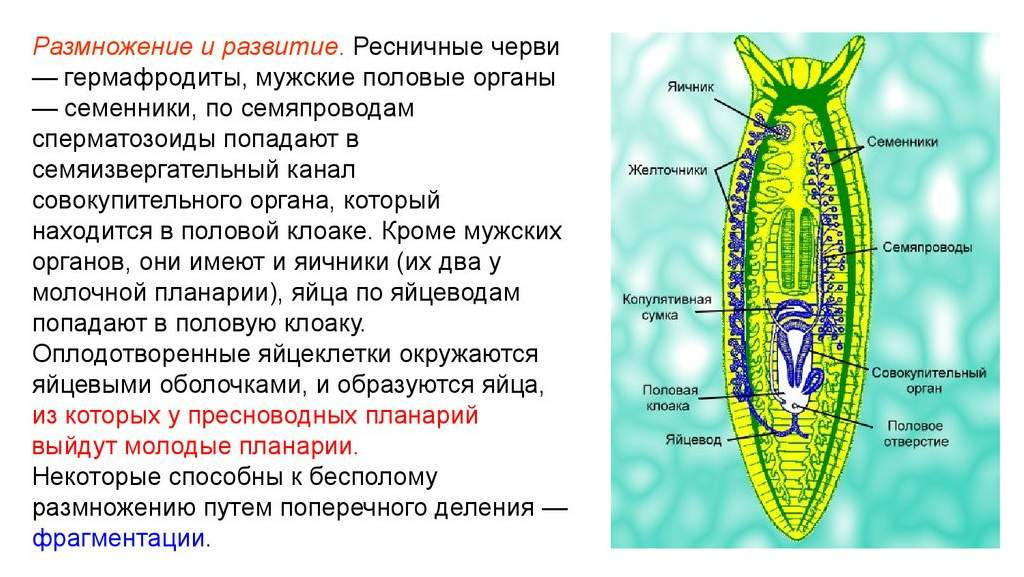
Google Scholar
Silflow CD, Lefebvre PA . Сборка и подвижность ресничек и жгутиков эукариот. Уроки Chlamydomonas reinhardtii . Завод Физиол 2001; 127 :1500–1507.
КАС пабмед ПабМед Центральный Google Scholar
Ридер К.Л., Фаруки С., Ходжаков А. . Центросома у позвоночных: больше, чем центр организации микротрубочек. Trends Cell Biol 2001; 11 :413–419.
КАС Google Scholar
Бейссон Дж., Райт М. . Сборка базального тельца/центриоли и непрерывность. Curr Opin Cell Biol 2003; 15 :96–104.
КАС Google Scholar
Датчер SK . Выяснение функций базальных телец и центриолей у Chlamydomonas reinhardtii . Трафик 2003; 4 :443–451.
Трафик 2003; 4 :443–451.
КАС Google Scholar
Афзелиус Б.А. Синдром человека, вызванный неподвижными ресничками. Наука 1976; 193 :317–319.
КАС Google Scholar
Бердон В.Е., Вилли У. Situs inversus, бронхоэктазы и синусит и их связь с неподвижными ресничками: история болезней и их первооткрыватели — Манес Картагенер и Бьорн Афцелиус. Педиатр Радиол 2004; 34 : 38–42.
Google Scholar
Афзелиус Б.А. Заболевания, связанные с ресничками. Дж Патол 2004; 204 :470–477.
КАС Google Scholar
Геремек М., Витт М. . Первичная цилиарная дискинезия: гены, гены-кандидаты и хромосомные регионы. J Appl Genet 2004; 45 :347–361.
Google Scholar
МакГрат Дж., Брюкнер М. . Реснички лежат в основе лево-правой асимметрии позвоночных. Curr Opin Genet Dev 2003; 13 :385–392.
КАС Google Scholar
McGrath J, Somlo S, Makova S, et al. Две популяции узловых моноцилий инициируют лево-правую асимметрию у мышей. Сотовый 2003; 114 : 61–73.
КАС пабмед Google Scholar
Нонака С., Ширатори Х., Сайджох Ю. и др. Определение лево-правого паттерна эмбриона мыши с помощью искусственного узлового потока. Природа 2002; 418 :96–99.
КАС пабмед Google Scholar
Ибанез-Таллон И., Хайнц Н., Омран Х. Бить или не бить: роль ресничек в развитии и заболевании. Хум Мол Жене 2003; 12 :R27–R35.
Хум Мол Жене 2003; 12 :R27–R35.
КАС Google Scholar
Саэки Х., Кондо С., Морита Т. и др. Синдром неподвижных ресничек, связанный с поликистозом почек. Дж Урол 1984; 132 :1165–1166.
КАС Google Scholar
Розенбаум Д.Л., Витман Г.Б. Внутрижгутиковый транспорт. Nat Rev Mol Cell Biol 2002; 3 :813–825.
КАС Google Scholar
Снелл В.Дж., Пан Дж., Ван К. Выявлены реснички и жгутики: от сборки жгутиков у Chlamydomonas до нарушений ожирения у человека. Сотовый 2004; 117 :693–697.
КАС Google Scholar
Козмински К.Г., Джонсон К.А., Форшер П. и др. Подвижность жгутика эукариот, не связанная с биением жгутиков. Proc Natl Acad Sci USA 1993; 90 :5519–5523.
Proc Natl Acad Sci USA 1993; 90 :5519–5523.
КАС Google Scholar
Шоли Дж.М. Внутрижгутиковый транспорт. Annu Rev Cell Dev Biol 2003; 19 :423–443.
КАС Google Scholar
Цинь Х., Динер Д.Р., Геймер С. и др. Внутрижгутиковый транспорт (IFT) груз: IFT транспортирует жгутиковые предшественники к кончику и продукты оборота к телу клетки. J Cell Biol 2004; 164 :255–266.
КАС пабмед ПабМед Центральный Google Scholar
Маршалл В.Ф., Розенбаум Дж.Л. Внутрижгутиковый транспорт уравновешивает непрерывный оборот наружных дублетных микротрубочек: последствия для контроля длины жгутиков. J Cell Biol 2001; 155 :405–414.
КАС пабмед ПабМед Центральный Google Scholar
Коул Д. Г.Внутрижгутиковый транспортный аппарат Chlamydomonas reinhardtii . Трафик 2003; 4 :435–442.
Г.Внутрижгутиковый транспортный аппарат Chlamydomonas reinhardtii . Трафик 2003; 4 :435–442.
КАС Google Scholar
Пиперно Г., Мид К., Хендерсон С. . Внутренние плечи динеина, но не внешние плечи динеина, нуждаются в активности белка-гомолога кинезина KHP1(FLA10) для достижения дистальной части жгутиков у Chlamydomonas . J Cell Biol 1996; 133 :371–379.
КАС Google Scholar
Цинь Х., Розенбаум Д.Л., Барр М.М. Гомолог аутосомно-рецессивного гена поликистозной болезни почек участвует в внутрижгутиковом транспорте в C. elegans реснитчатых сенсорных нейронах. Карр Биол 2001; 11 :457–461.
КАС Google Scholar
Berman SA, Wilson NF, Haas NA, et al. Новая киназа MAP регулирует длину жгутиков у Chlamydomonas . Карр Биол 2003; 13 :1145–1149.
Карр Биол 2003; 13 :1145–1149.
КАС Google Scholar
Тэм Л.В., Дентлер В.Л., Лефевр П.А. Дефектная сборка жгутиков и регуляция длины у нулевых мутантов LF3 у Chlamydomonas . J Cell Biol 2003; 163 :597–607.
КАС пабмед ПабМед Центральный Google Scholar
Пан Дж., Ван К., Снелл В.Дж.Киназа aurora необходима для разборки жгутиков у Chlamydomonas . Dev Cell 2004; 6 :445–451.
КАС Google Scholar
Уилсон Н.Ф., Лефевр П.А. Регуляция сборки жгутиков киназой 3 гликогенсинтазы у Chlamydomonas reinhardtii . Эукариотическая ячейка 2004 г.; 3 :1307–1319.
КАС пабмед ПабМед Центральный Google Scholar
Педерсен Л. Б., Геймер С., Слобода Р.Д., и др.Микротрубочки плюс отслеживающий концы белок EB1 локализованы на кончике жгутика и базальных тельцах Chlamydomonas reinhardtii . Карр Биол 2003; 13 : 1969–1974.
Б., Геймер С., Слобода Р.Д., и др.Микротрубочки плюс отслеживающий концы белок EB1 локализованы на кончике жгутика и базальных тельцах Chlamydomonas reinhardtii . Карр Биол 2003; 13 : 1969–1974.
КАС Google Scholar
Тирнауэр Дж.С., Бирер Б.Е. Белки EB1 регулируют динамику микротрубочек, полярность клеток и стабильность хромосом. J Cell Biol 2000; 149 :761–766.
КАС пабмед ПабМед Центральный Google Scholar
Нгуен Р.Л., Там Л.В., Лефевр П.А.Ген LF1 Chlamydomonas reinhardtii кодирует новый белок, необходимый для контроля длины жгутиков. Генетика 2004.
Дукат Д, Чжэн Ю . Киназы Aurora в сборке веретена и сегрегации хромосом. Exp Cell Res 2004; 301 :60–67.
КАС Google Scholar
Сарпал Р. , Тоди С.В., Сиван-Лукьянова Е. и др. Drosophila KAP взаимодействует с моторной субъединицей кинезина II KLP64D для сборки хордотональных сенсорных ресничек, но не хвостов сперматозоидов. Карр Биол 2003; 13 :1687–1696.
, Тоди С.В., Сиван-Лукьянова Е. и др. Drosophila KAP взаимодействует с моторной субъединицей кинезина II KLP64D для сборки хордотональных сенсорных ресничек, но не хвостов сперматозоидов. Карр Биол 2003; 13 :1687–1696.
КАС Google Scholar
Хан Ю.Г., Квок Б.Х., Кернан М.Дж. Внутрижгутиковый транспорт необходим у Drosophila для дифференцировки сенсорных ресничек, но не сперматозоидов. Карр Биол 2003; 13 :1679–1686.
КАС Google Scholar
Витман ГБ . Подвижность клеток: глухая Дрозофила держать ритм. Карр Биол 2003; 13 :R796–R798.
КАС Google Scholar
Браун Дж.М., Файн Н.А., Пандиян Г. и др. Гипоксия регулирует сборку ресничек у супрессоров Tetrahymena , у которых отсутствует ген внутрижгутиковой транспортной субъединицы. Мол Биол Селл 2003; 14 :3192–3207.
Мол Биол Селл 2003; 14 :3192–3207.
КАС пабмед ПабМед Центральный Google Scholar
Sun Z, Amsterdam A, Pazour GJ и др.Генетический скрининг у рыбок данио идентифицирует гены ресничек как основную причину кистозной почки. Разработка 2004; 131 :4085–4093.
КАС Google Scholar
Цудзикава М., Малики Дж. Внутрижгутиковые транспортные гены необходимы для дифференцировки и выживания сенсорных нейронов позвоночных. Нейрон 2004; 42 :703–716.
КАС Google Scholar
Гуай-Вудфорд Л.М.Мышиные модели поликистоза почек: молекулярные и терапевтические идеи. Am J Physiol Renal Physiol 2003; 285 :F1034–F1049.
КАС Google Scholar
Cole DG, Diener DR, Himelblau AL, et al. Chlamydomonas кинезин-II-зависимый внутрижгутиковый транспорт (IFT): частицы IFT содержат белки, необходимые для сборки ресничек в сенсорных нейронах Caenorhabditis elegans . J Cell Biol 1998; 141 :993–1008.
Chlamydomonas кинезин-II-зависимый внутрижгутиковый транспорт (IFT): частицы IFT содержат белки, необходимые для сборки ресничек в сенсорных нейронах Caenorhabditis elegans . J Cell Biol 1998; 141 :993–1008.
КАС пабмед ПабМед Центральный Google Scholar
Барр М.М., Штернберг П.В. Гомолог гена поликистозной болезни почек необходим для брачного поведения самцов C. elegans . Природа 1999; 401 :386–389.
КАС Google Scholar
Пазур Г.Дж., Дикерт Б.Л., Вучика Ю. и др. Chlamydomonas IFT88 и ее мышиный гомолог, ген поликистозной болезни почек tg737, необходимы для сборки ресничек и жгутиков. J Cell Biol 2000; 151 :709–718.
КАС пабмед ПабМед Центральный Google Scholar
Лин Ф. , Хисбергер Т., Кордес К. и др. Специфическая для почек инактивация субъединицы KIF3A кинезина-II ингибирует почечный цилиогенез и вызывает поликистозную болезнь почек. Proc Natl Acad Sci USA 2003; 100 :5286–5291.
, Хисбергер Т., Кордес К. и др. Специфическая для почек инактивация субъединицы KIF3A кинезина-II ингибирует почечный цилиогенез и вызывает поликистозную болезнь почек. Proc Natl Acad Sci USA 2003; 100 :5286–5291.
КАС Google Scholar
Полиция Уилсона .Поликистоз почек. N Engl J Med 2004; 350 :151–164.
КАС пабмед Google Scholar
Hughes J, Ward CJ, Peral B, et al. Ген поликистозной болезни почек 1 (PKD1) кодирует новый белок с множественными доменами распознавания клеток. Нат Жене 1995; 10 :151–160.
КАС Google Scholar
Мочизуки Т., Ву Г., Хаяши Т. и др.PKD2, ген поликистоза почек, кодирующий интегральный мембранный белок. Наука 1996; 272 :1339–1342.
КАС Google Scholar
Болетта А. , Жермино Г.Г. Роль полицистинов в почечном тубулогенезе. Trends Cell Biol 2003; 13 :484–492.
, Жермино Г.Г. Роль полицистинов в почечном тубулогенезе. Trends Cell Biol 2003; 13 :484–492.
КАС Google Scholar
Игараши П., Сомло С. . Генетика и патогенез поликистоза почек. J Am Soc Нефрол 2002; 13 :2384–2398.
КАС Google Scholar
Онучич Л.Ф., Фуру Л., Нагасава Ю. и др. PKHD1, ген поликистоза почек и заболевания печени 1, кодирует новый большой белок, содержащий множественные иммуноглобулиноподобные домены фактора транскрипции плексина и параллельные повторы бета-спирали 1. Am J Hum Genet 2002; 70 :1305–1317.
КАС пабмед ПабМед Центральный Google Scholar
Ward CJ, Hogan MC, Rossetti S, et al.Ген, мутировавший при аутосомно-рецессивном поликистозе почек, кодирует большой рецептороподобный белок. Нат Жене 2002; 30 :259–269.
Нат Жене 2002; 30 :259–269.
Google Scholar
Хисбергер Т., Бай Ю., Шао Х. и др. Мутация ядерного фактора-1бета гепатоцитов ингибирует экспрессию гена Pkhd1 и приводит к образованию почечных кист у мышей. J Clin Invest 2004; 113 :814–825.
КАС пабмед ПабМед Центральный Google Scholar
Кано Д.А., Мурсия Н.С., Пазур Г.Дж. и др.Модель поликистоза почек у мышей Orpk выявляет существенную роль первичных ресничек в организации ткани поджелудочной железы. Разработка 2004; 131 :3457–3467.
КАС Google Scholar
Хильдебрандт Ф . Нефронофтиз-медуллярная кистозная болезнь почек. В: Avner ED, Harmon WE, Niaudet P (eds.) Pediatric Nephrology , 5th edn. Липинкотт, Уильямс и Уильямс: Филадельфия, 2004 г. , стр. 665–673.
, стр. 665–673.
Google Scholar
Сонье С., Каладо Дж., Хейлиг Р. и др. Новый ген, который кодирует белок с доменом предполагаемой гомологии src 3, является геном-кандидатом для семейного ювенильного нефронофтиза. Hum Mol Genet 1997; 6 :2317–2323.
КАС Google Scholar
Hildebrandt F, Otto E, Rensing C, et al. Новый ген, кодирующий белок домена Sh4, мутирует при нефронофтизе 1 типа. Нат Жене 1997; 17 :149–153.
КАС Google Scholar
Отто Э.А., Шермер Б., Обара Т. и др. Мутации в INVS, кодирующем инверсин, вызывают нефронофтиз 2 типа, связывая почечную кистозную болезнь с функцией первичных ресничек и детерминацией левой-правой оси. Нат Жене 2003; 34 :413–420.
КАС пабмед ПабМед Центральный Google Scholar
Ольбрих Х. , Флигауф М., Хофеле Дж. и др.Мутации в новом гене NPHP3 вызывают у подростков нефронофтиз, тапеторетинальную дегенерацию и фиброз печени. Нат Жене 2003; 34 :455–459.
, Флигауф М., Хофеле Дж. и др.Мутации в новом гене NPHP3 вызывают у подростков нефронофтиз, тапеторетинальную дегенерацию и фиброз печени. Нат Жене 2003; 34 :455–459.
КАС Google Scholar
Отто Э., Хофеле Дж., Руф Р. и др. Ген, мутировавший при нефронофтизе и пигментном ретините, кодирует новый белок нефроретинин, законсервированный в эволюции. Am J Hum Genet 2002; 71 :1161–1167.
КАС пабмед ПабМед Центральный Google Scholar
Молле Г., Саломон Р., Грибуваль О. и др.Ген, мутировавший при ювенильном нефронофтизе 4 типа, кодирует новый белок, взаимодействующий с нефроцистином. Нат Жене 2002; 32 :300–305.
КАС Google Scholar
Наули С.М., Аленгхат Ф.Дж., Луо Ю. и др. Полицистины 1 и 2 опосредуют механоощущение в первичных ресничках почечных клеток. Нат Жене 2003; 33 :129–137.
Нат Жене 2003; 33 :129–137.
КАС Google Scholar
Йодер Б.К., Хоу С., Гуай-Вудфорд Л.М.Белки поликистоза почек, полицистин-1, полицистин-2, полярис и цистин локализованы в почечных ресничках. J Am Soc Нефрол 2002; 13 :2508–2516.
КАС Google Scholar
Уорд С.Дж., Юань Д., Масюк Т.В. и др. Клеточная и субклеточная локализация белка ARPKD; фиброцистин экспрессируется на первичных ресничках. Хум Мол Жене 2003; 12 :2703–2710.
КАС Google Scholar
Wang S, Luo Y, Wilson PD, et al.Аутосомно-рецессивный белок поликистозной болезни почек локализуется в первичных ресничках с концентрацией в базальной области тела. J Am Soc Нефрол 2004; 15 :592–602.
Google Scholar
Zhang MZ, Mai W, Li C, et al. Белок PKHD1, кодируемый геном аутосомно-рецессивного поликистоза почек, ассоциирован с базальными тельцами и первичными ресничками в почечных эпителиальных клетках. Proc Natl Acad Sci USA 2004; 101 :2311–2316.
Белок PKHD1, кодируемый геном аутосомно-рецессивного поликистоза почек, ассоциирован с базальными тельцами и первичными ресничками в почечных эпителиальных клетках. Proc Natl Acad Sci USA 2004; 101 :2311–2316.
КАС Google Scholar
Морган Д., Элей Л., Сэйер Дж. и др. Анализы экспрессии и взаимодействия с анафазным активирующим комплексным белком Apc2 указывают на роль inversin в первичных ресничках и участие в клеточном цикле. Хум Мол Жене 2002; 11 :3345–3350.
КАС Google Scholar
Ватанабе Д., Сайджо Ю., Нонака С. и др.Лево-правый детерминант Inversin является компонентом моноцилий узла и других ресничек 9+0. Разработка 2003; 130 :1725–1734.
КАС Google Scholar
Хоу С., Мруг М., Йодер Б. К. и др. Cystin, новый белок, ассоциированный с ресничками, разрушается в cpk мышиной модели поликистоза почек. J Clin Invest 2002; 109 :533–540.
К. и др. Cystin, новый белок, ассоциированный с ресничками, разрушается в cpk мышиной модели поликистоза почек. J Clin Invest 2002; 109 :533–540.
КАС пабмед ПабМед Центральный Google Scholar
Йодер Б.К., Туссон А., Милликан Л. и др.Polaris, белок, разрушенный у orpk мутантных мышей, необходим для сборки почечной реснички. Am J Physiol Renal Physiol 2002; 282 :F541–F552.
КАС Google Scholar
Браун NE, Мурсия NS . Задержка кистогенеза и повышенный цилиогенез связаны с повторной экспрессией polaris у мышей с мутацией Tg737. Kidney Int 2003; 63 :1220–1229.
Google Scholar
Taulman PD, Haycraft CJ, Balkovetz DF, et al.Polaris, белок, участвующий в формировании паттерна лево-правой оси, локализуется в базальных тельцах и ресничках. Мол Биол Селл 2001; 12 :589–599.
Мол Биол Селл 2001; 12 :589–599.
КАС пабмед ПабМед Центральный Google Scholar
Ханаока К., Цянь Ф., Болетта А. и др. Совместная сборка полицистина-1 и -2 создает уникальные токи, проницаемые для катионов. Природа 2000; 408 :990–994.
КАС Google Scholar
Roitbak T, Ward CJ, Harris PC, et al.Мультибелковый комплекс полицистина-1 разрушается в клетках поликистозной болезни почек. Мол Биол Селл 2004; 15 :1334–1346.
КАС пабмед ПабМед Центральный Google Scholar
Преториус HA, Spring KR . Первичная ресничка почечной клетки функционирует как датчик потока. Curr Opin Nephrol Hypertens 2003; 12 :517–520.
Google Scholar
Преториус HA, Spring KR . Изгиб первичной реснички клеток MDCK увеличивает внутриклеточный кальций. J Membr Biol 2001; 184 : 71–79.
Изгиб первичной реснички клеток MDCK увеличивает внутриклеточный кальций. J Membr Biol 2001; 184 : 71–79.
КАС Google Scholar
Преториус Х.А., Фрокиар Дж., Нильсен С. и др. Изгиб первичной реснички открывает Ca 2+ -чувствительные K+ каналы промежуточной проводимости в клетках MDCK. J Membr Biol 2003; 191 :193–200.
КАС Google Scholar
Преториус HA, Spring KR .Удаление первичной реснички клеток MDCK отменяет определение потока. J Membr Biol 2003; 191 :69–76.
КАС Google Scholar
Болетта А., Цянь Ф., Онучич Л.Ф. и др. Полицистин-1, продукт гена PKD1 , индуцирует устойчивость к апоптозу и спонтанный тубулогенез в клетках MDCK. Мол Ячейка 2000; 6 :1267–1273.
КАС пабмед Google Scholar
Бензинг Т., Герке П., Хопкер К. и др.Нефроцистин взаимодействует с Pyk2, p130(Cas) и тензином и запускает фосфорилирование Pyk2. Proc Natl Acad Sci USA 2001; 98 :9784–9789.
КАС Google Scholar
Дональдсон Дж. К., Демпси П. Дж., Редди С. и др. Crk-ассоциированный субстрат p130(Cas) взаимодействует с нефроцистином, и оба белка локализуются в межклеточных контактах поляризованных эпителиальных клеток. Exp Cell Res 2000; 256 : 168–178.
КАС Google Scholar
Элей Л., Тернпенни Л., Йейтс Л.М. и др. Взгляд на инверсин. Cell Biol Int 2004; 28 :119–124.
КАС Google Scholar
Pennekamp P, Karcher C, Fischer A, et al. Ионный канал полицистин-2 необходим для определения оси лево-право у мышей. Карр Биол 2002; 12 :938–943.
Ионный канал полицистин-2 необходим для определения оси лево-право у мышей. Карр Биол 2002; 12 :938–943.
КАС Google Scholar
Уотник Т.Дж., Джин И., Матунис Э. и др. Жгутиковый гомолог полицистина-2 необходим для мужской фертильности Drosophila . Карр Биол 2003; 13 :2179–2184.
КАС Google Scholar
Греш Л., Фишер Э., Рейманн А. и др. Транскрипционная сеть при поликистозе почек. EMBO J 2004; 23 :1657–1668.
КАС пабмед ПабМед Центральный Google Scholar
Menezes LF, Cai Y, Nagasawa Y, et al. Полидуктин, продукт гена PKHD1, включает изоформы, экспрессируемые в плазматической мембране, первичных ресничках и цитоплазме. Kidney Int 2004; 66 :1345–1355.
КАС Google Scholar
Хагивара Х., Овада Н., Таката К. . Клеточная биология нормального и аномального цилиогенеза в мерцательном эпителии. Int Rev Cytol 2004; 234 :101–141.
Google Scholar
Уотник Т., Жермино Г. . От ресничек к кисте. Нат Жене 2003; 34 :355–356.
КАС Google Scholar
Никель С., Бензинг Т., Селлин Л. и др. С-концевой фрагмент полицистина-1 запускает морфогенез ветвления и миграцию эпителиальных клеток канальцев почки. J Clin Invest 2002; 109 :481–489.
КАС пабмед ПабМед Центральный Google Scholar
Chauvet V, Xin T, Husson H, et al. Механические стимулы вызывают расщепление и ядерную транслокацию С-конца полицистина-1. J Clin Invest 2004; 114 :1433–1443.
J Clin Invest 2004; 114 :1433–1443.
КАС пабмед ПабМед Центральный Google Scholar
Кацанис Н .Олигогенные свойства синдрома Барде-Бидля. Хум Мол Жене 2004; 13 :R65–R71.
КАС Google Scholar
Ansley SJ, Badano JL, Blacque OE и др. Дисфункция базальных телец является вероятной причиной плейотропного синдрома Барде-Бидля. Природа 2003; 425 :628–633.
КАС Google Scholar
Kim JC, Badano JL, Sibold S, et al.Белок Bardet-Biedl BBS4 направляет груз в перицентриолярную область и необходим для закрепления микротрубочек и прогрессирования клеточного цикла. Нат Жене 2004; 36 :462–470.
КАС Google Scholar
Микитин К. , Маллинс Р.Ф., Эндрюс М. и др. Мыши с синдромом Барде-Бидля типа 4 (BBS4)-нулевые мыши участвуют Bbs4 в формировании жгутиков, но не в глобальной сборке ресничек. Proc Natl Acad Sci USA 2004; 101 :8664–8669.
, Маллинс Р.Ф., Эндрюс М. и др. Мыши с синдромом Барде-Бидля типа 4 (BBS4)-нулевые мыши участвуют Bbs4 в формировании жгутиков, но не в глобальной сборке ресничек. Proc Natl Acad Sci USA 2004; 101 :8664–8669.
КАС Google Scholar
Kulaga HM, Leitch CC, Eichers ER и др. Потеря белков BBS вызывает аносмию у людей и дефекты структуры и функции обонятельных ресничек у мышей. Нат Жене 2004; 36 :994–998.
КАС Google Scholar
Микитин К., Шеффилд В.К. Установление связи между ресничками и синдромом Барде-Бидля. Trends Mol Med 2004; 10 :106–109.
КАС Google Scholar
Li JB, Gerdes JM, Haycraft CJ и др. Сравнительная геномика идентифицирует протеом жгутика и базального тела, который включает ген болезни человека BBS5.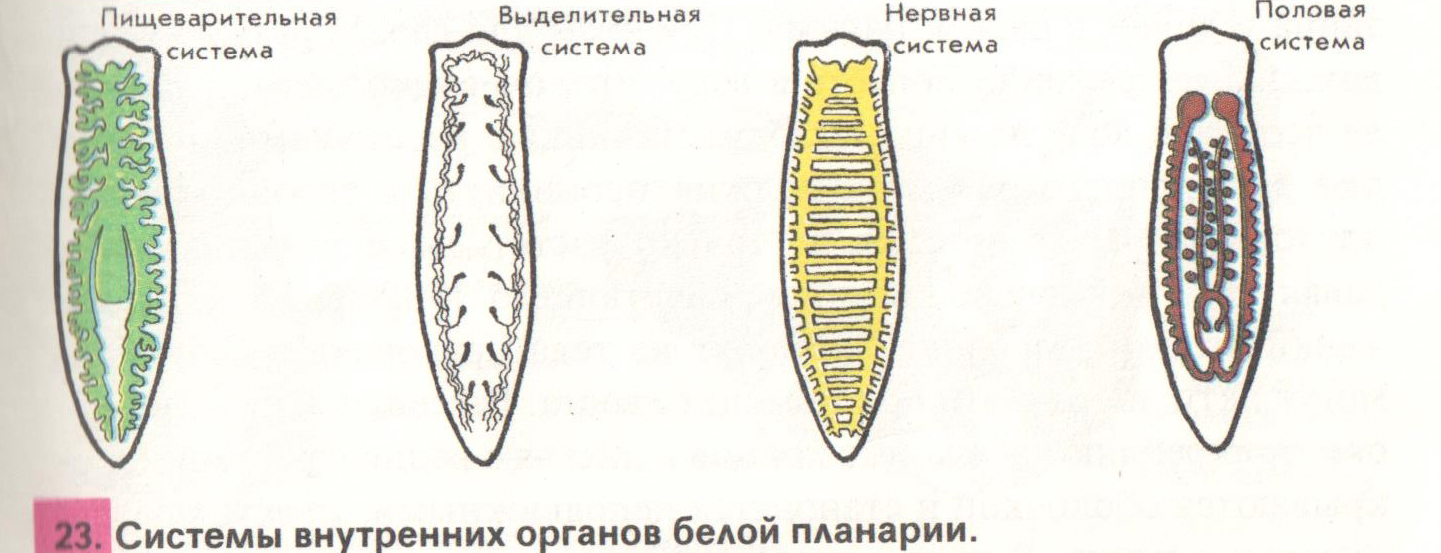 Сотовый 2004; 117 :541–552.
Сотовый 2004; 117 :541–552.
КАС Google Scholar
Блак О.Е., Рирдон М.Дж., Ли С. и др.Потеря функции белка C. elegans BBS-7 и BBS-8 приводит к дефектам ресничек и нарушению внутрижгутикового транспорта. Гены Дев 2004; 18 :1630–1642.
КАС пабмед ПабМед Центральный Google Scholar
Преториус Х.А., Преториус Дж., Нильсен С. и др. Бета1-интегрины в первичной ресничке клеток MDCK потенцируют индуцированную фибронектином передачу сигналов Ca 2+ . Am J Physiol Renal Physiol 2004; 287 :F969–F978.
КАС Google Scholar
Гендель М., Шульц С., Станариус А. и др. Селективное нацеливание рецептора соматостатина 3 на нейрональные реснички. Неврология 1999; 89 :909–926.
КАС Google Scholar
Браилов И., Банкила М., Бризоргей М.Дж. и др. Локализация рецепторов 5-НТ(6) на плазматической мембране ресничек нейронов головного мозга крысы. Мозг Рес 2000; 872 : 271–275.
КАС Google Scholar
Степанян З., Кочарян А., Пырский М. и др. Нейроны-мишени для лептина гипоталамуса крысы экспрессируют рецепторы соматостатина. J Нейроэндокринол 2003; 15 :822–830.
КАС Google Scholar
Хуанфу Д., Лю А., Ракеман А.С. и др. Для передачи сигналов Hedgehog у мышей требуются внутрижгутиковые транспортные белки. Природа 2003; 426 : 83–87.
КАС Google Scholar
Пул, Калифорния, Флинт, М. Х., Бомонт, Б. В. . Анализ морфологии и функции первичных ресничек соединительной ткани: клеточный кибернетический зонд? Селл Мотил 1985; 5 :175–193.
КАС Google Scholar
Пазур Г.Дж., Уитман Г.Б. Первичная ресничка позвоночных представляет собой сенсорную органеллу. Curr Opin Cell Biol 2003; 15 :105–110.
КАС Google Scholar
Солтер К.М., Гибор А. . Доказательства роли жгутиков в качестве сенсорных преобразователей при спаривании Chlamydomonas reinhardii . Природа 1977; 265 :444–445.
КАС Google Scholar
Пан Дж., Снелл В.Дж. Кинезин-II необходим для жгутиковой сенсорной передачи во время оплодотворения у Chlamydomonas . Мол Биол Селл 2002; 13 :1417–1426.
КАС пабмед ПабМед Центральный Google Scholar
Pan J, Misamore MJ, Wang Q и др. Транспорт белка и передача сигнала во время оплодотворения у Chlamydomonas . Трафик 2003; 4 :452–459.
КАС Google Scholar
Ван К., Снелл В.Дж. Жгутиковая адгезия между гаметами типа спаривания плюс и гаметами типа спаривания минус активирует жгутиковую протеин-тирозинкиназу во время оплодотворения у Chlamydomonas . J Biol Chem 2003; 278 :32936–32942.
КАС Google Scholar
Jensen CG, Poole CA, McGlashan SR, et al. Ультраструктурное, томографическое и конфокальное изображение первичной реснички хондроцитов in situ . Cell Biol Int 2004; 28 :101–110.
КАС Google Scholar
Кессель Р.Г., Кардон Р.Х. Ткани и органы: текст-атлас сканирующей электронной микроскопии .WH Freeman and Company: Сан-Франциско, Калифорния, 1979 г.
Google Scholar
Балантидиаз
Возбудители
Balantidium (= Neobalantidium ) (= Balantioides ) coli, крупное реснитчатое простейшее, является единственной известной инфузорией, способной заражать людей. Это часто связано со свиньями, основным хозяином-резервуаром. Недавний молекулярный анализ показал необходимость таксономического пересмотра, и теперь его иногда называют Neobalantidium coli или Balantioides coli, , хотя эта номенклатура не была ни утверждена, ни широко принята в медицинском сообществе.
Жизненный цикл
Кисты являются стадией передачи балантидиаза. Хозяин чаще всего приобретает кисту при приеме зараженной пищи или воды. После приема внутрь в тонком кишечнике происходит эксцистация, а трофозоиты колонизируют толстый кишечник. Трофозоиты обитают в просвете толстой кишки и червеобразном отростке человека и животных, где они размножаются путем бинарного деления, во время которого может происходить конъюгация. Трофозоиты подвергаются инцистации с образованием инфекционных цист.Некоторые трофозоиты внедряются в стенку толстой кишки и размножаются, вызывая язвенную патологию в стенке толстой кишки. Некоторые возвращаются в просвет и распадаются. Зрелые кисты выходят с калом.
Хосты
Свиньи являются основными резервуарными хозяевами. Люди также могут быть резервуарами, а другие потенциальные животные-хозяева включают грызунов и нечеловеческих приматов.
Географическое распространение
Balantidium coli встречается во всем мире. Поскольку свиньи являются основным резервуаром, инфицирование людей чаще происходит в районах, где выращиваются свиньи, а санитария неадекватна.
В большинстве случаев заболевание протекает бессимптомно. Клинические проявления, если они есть, могут быть острыми или хроническими с абдоминальными симптомами. Осложнения сопутствующей диареи или дизентерии могут возникать при затяжных инфекциях. Симптомы могут быть тяжелыми или смертельными у ослабленных людей/лиц с ослабленным иммунитетом.
Внекишечная инфекция встречается редко, но потенциально серьезна и обычно развивается вторично по отношению к кишечной инфекции. Перитонит и абсцессы печени отмечаются после перфорации кишечника или разрыва молниеносной язвы толстой кишки.Инвазия урогенитального тракта может быть вызвана контаминацией из анальной области или через свищи, вызванные тяжелой инфекцией.
Acoela: об их родстве и родстве, особенно с немертодерматидами и ксенотурбеллидами (Bilateria incertae sedis)
Abildgaard, P. C. (1806). Zoologica danica seu animalium Danicae et Norwegiae rariorum ac mines notorum descriptorum et historia. В OF Müller (Ed.), Zoologica Danica 4 (стр. 26). Копенгаген (Хавния): Н. Мёллери.
Google Scholar
Ахатц, Дж. Г., Хуге, М., Уоллберг, А., Джонделиус, У., и Тайлер, С. (2010). Систематический пересмотр акоэлей с ультраструктурой сперматозоидов 9 + 0 (Convolutida) и влияние полового конфликта на морфологию. Журнал зоологической систематики и эволюционных исследований, 48 (1), 9–32.
Артикул Google Scholar
Окессон, Б., Гшвентнер Р., Хендельберг Дж., Ладурнер П., Мюллер Дж. и Ригер Р. (2001). Деление в Convolutriloba longifissura : повторное рассмотрение бесполого размножения бескрылых турбеллярий. Acta Zoologica (Копенгаген), 82 (3), 231–239.
Google Scholar
Апельт, Г. (1969). Fortpflanzungsbiologie, Entwicklungszyklen und vergleichende Frühentwicklung acoeler Turbellarien. Морская биология (Берлин), 4 , 267–325.
Google Scholar
Акс, П., и Дорджес, Дж. (1966). Oligocherus limnophilus нояб. spec., ein Kaspisches Faunenelement als erster Süsswasservertreter der Turbellaria Acoela in Flussen Mitteleuropas. Internationale Revue der Gesamten Hydrobiologie, 51 , 15–44.
Артикул Google Scholar
Багунья, Дж., и Риуторт, М. (2004).Рассвет билатеральных животных: случай ацеломорфных плоских червей. Bioessays, 26 (10), 1046–1057.
ПабМед Статья КАС Google Scholar
Барнеа О., Брикнер И., Хуге М., Вейс В. М. и Бенаяху Ю. (2007). Первые свидетельства материнской передачи эндосимбионтов водорослей на стадии ооцитов в триплобластном хозяине с наблюдениями за размножением у Waminoa brickneri (Acoelomorpha). Биология беспозвоночных, 126 (2), 113–119.
Артикул Google Scholar
Бартоломеус Т. и Бальцер И. (1997). Convolutriloba longifissura , nov. спец. (Acoela) — первый случай продольного деления у Plathelminthes. Микрофауна Марина, 11 , 7–18.
Google Scholar
Бери А., Кардона А., Мартинес П. и Хартенштейн В.(2010). Строение центральной нервной системы ювенильного акоэля, Symsagittifera roscoffensis . Гены развития и эволюция, 220 (3–4), 61–76.
ПабМед Статья Google Scholar
Бери, А., и Мартинес, П. (2011). Активность ацетилхолинэстеразы в развивающейся и регенерирующей нервной системе acoel Symsagittifera roscoffensis . Acta Zoologica (Копенгаген), 92 (4), 383–392.
Артикул Google Scholar
Бун М., Виллемс М., Клэйс М. и Артуа Т. (2011). Сперматогенез и строение семенников Isodiametra pulchra (Isodiametridae, Acoela). Acta Zoologica (Копенгаген), 92 (2), 101–108.
Артикул Google Scholar
Бурла, С. Дж., Джулиусдоттир, Т., Лоу, С. Дж., Фримен, Р., Aronowicz, J., Kirschner, M., et al. (2006). Филогения вторичноротых выявляет монофилетических хордовых и новый тип Xenoturbellida. Nature (Лондон), 444 (7115), 85–88.
КАС Статья Google Scholar
Бурла, С.Дж., Нильсен, К., Локьер, А.Е., Литтлвуд, Д.Т.Дж., и Телфорд, М.Дж. (2003). Xenoturbella — вторичноротый, питающийся моллюсками. Nature (Лондон), 424 (6951), 925–928.
КАС Статья Google Scholar
Бурла, С. Дж., Рота-Стабелли, О., Ланфир, Р., и Телфорд, М. Дж. (2009). Структура митохондриального генома Xenoturbella bocki (филум Xenoturbellida) является предковой внутри вторичноротых. BMC Эволюционная биология, 9 (107), 1–14.
Google Scholar
Бойер, Британская Колумбия, Генри, Дж. К.и Мартиндейл, М. К. (1996). Модифицированное спиральное дробление: паттерн дуэтного дробления и судьбы ранних бластомеров у acoel turbellarian Neochildia fusca . Биологический бюллетень Wood’s Hole, 191 (2), 285–286.
Google Scholar
Бресслау, Э. (1909). Die Entwicklung der Acoelen. Verhandlungen der Deutschen Zoologischen Gesellschaft Leipzig, 19 , 314–324.
Google Scholar
Брюггеманн, Й.(1985а). Ультраструктура и формирование загубника бурсы Philocelis cellata (Plathelminthes[Platyhelminthes], Acoela). Гидробиология, 128 (1), 23–30.
Артикул Google Scholar
Брюггеманн, Дж. (1985b). Ultrastruktur und Bildungsweise penialer Hartstrukturen bei freilebenden Plathelminthen. Зооморфология (Берлин), 105 (3), 143–189.
Артикул Google Scholar
Буэно, Д., Багунья, Дж., и Ромеро, Р. (1997). Клеточно-, ткане- и позиционно-специфические моноклональные антитела против планарии Dugesia ( Girardia ) tigrina . Гистохимия и клеточная биология, 107 (2), 139–149.
ПабМед КАС Статья Google Scholar
Буш, Л. (1975). Биология Neochilda fusca n. ген. н. сп. с северо-восточного побережья США (Platyhelminthes — Turbellaria). Биологический бюллетень Wood’s Hole, 148 (1), 35–48.
КАС Статья Google Scholar
Карранса, С., Багунья, Дж., и Риуторт, М. (1997). Являются ли платигельминты монофилетической примитивной группой? Оценка с использованием последовательностей 18S рДНК. Молекулярная биология и эволюция, 14 (5), 485–497.
ПабМед КАС Статья Google Scholar
Чиодин М., Ахатц, Дж. Г., Ваннингер, А., и Мартинес, П. (2011). Молекулярная архитектура мышц Acoel и ее эволюционные последствия. Journal of Experimental Zoology Part B Molecular and Developmental Evolution, 316B (6), 427–439.
Артикул КАС Google Scholar
Костелло, Х.М., и Костелло, Д.П. (1938). Копуляция у Acoelous Turbellarian Polychoerus carmelensis . Биологический бюллетень Wood’s Hole, 75 , 85–98.
Артикул Google Scholar
Костелло, Х.М., и Костелло, Д.П. (1939). Откладка яиц у бескрылых турбеллярий Polychoerus carmelensis . Биологический бюллетень Wood’s Hole, 76 , 80–89.
Артикул Google Scholar
Крезе, М. (1975). Монография Solenofilomorphidae (Turbellaria: Acoela). Internationale Revue der Gesamten Hydrobiologie, 60 (6), 769–845.
Артикул Google Scholar
Дэвидсон, Э. Х. (2011). Эволюционная биология как биология регуляторных систем. Биология развития, 357 (1), 35–40. doi: 10.1016/j.ydbio.2011.02.004.
ПабМед КАС Статья Google Scholar
Де Мендоса, А., и Руис-Трильо, И. (2011). Таинственное эволюционное происхождение гена GNE и корня Bilateria. Молекулярная биология и эволюция, 28 (11), 2987–2991. doi:10.1093/Molbev/Msr142.
ПабМед Статья КАС Google Scholar
Де Малдер, К. (2009). Молекулярная характеристика исключительной системы стволовых клеток плоских червей . Диссертация, Гентский университет, Бельгия.
Де Малдер, К., Куалес, Г., Пфистер, Д., Виллемс, М., Эггер, Б., Сальвенмозер, В., и др. (2009).Характеристика системы стволовых клеток acoel Isodiametra pulchra . BMC Биология развития, 9 . дои: 10.1186/1471-213x-9-69.
Де Робертис, Э. М. (2008). Эво-Дево: Вариации на темы предков. Сотовый, 132 (2), 185–195. doi: 10.1016/J.Cell.2008.01.003.
ПабМед Статья КАС Google Scholar
Доу, Д. А. (1981). Сравнительная ультраструктура простой глотки у Turbellaria. Зооморфология (Берлин), 97 (1–2), 133–192.
Артикул Google Scholar
Дёрес, Дж. (1966). Paratomella unichaeta нояб. ген. ноябрь spec., Vertreter einer neuen Familie der Turbellaria Acoela mit asexueller Fortpflanzung durch Paratomie. Veröffentlichungen des Institutes für Meeresforschung Bremerhafen Sonderberichte, 2 , 187–200.
Google Scholar
Дёрес, Дж.(1968). Die Acoela (Turbellaria) der Deutschen Nordseekuste und ein neues System der Ordnung. Zeitschrift fuer Zoologische Systematik und Evolutionsforschung, 6 , 56–452.
Артикул Google Scholar
дю Буа-Реймон Маркус, Э. (1955). На Turbellaria и Polygordius с побережья Бразилии. Boletim da Faculdade de Filosofia Ciencias e Letras Universidade de Sao Paulo Zoologica, 20 , 19–65.
Google Scholar
Эггер, Б., Гшвентнер, Р., и Ригер, Р. (2007). Свободноживущие плоские черви под ножом: прошлое и настоящее. Гены развития и эволюция, 217 (2), 89–104.
ПабМед Статья Google Scholar
Эггер Б., Штайнке Д., Таруи Х., Де Малдер К., Арендт Д., Боргони Г. и др. (2009). Быть или не быть плоским червем: полемика Acoel. Plos One, 4 (5), doi:10.1371/journal.pone.0005502.
Артикул КАС Google Scholar
Элерс, У. (1985). Das Phylogenetische System der Plathelminthes . Штутгарт: Густав Фишер.
Google Scholar
Элерс, У. (1992a). О тонкой структуре Paratomella rubra Rieger & Ott (Acoela) и месте таксона Paratomella Dörjes в филогенетической системе Acoelomorpha (Plathelminthes). Микрофауна Марина, 7 , 265–293.
Google Scholar
Элерс, У. (1992b). Пульсирующие тельца у Anaperus tvaerminnensis (Luther, 1912) (Acoela, Plathelminthes) представляют собой дегенерирующие эпидермальные клетки. Микрофауна Марина, 7 , 295–310.
Google Scholar
Элерс, У. (1992c). Эпидермальные клетки, модифицированные дермонефридиями, с вероятной выделительной функцией в Paratomella rubra (Acoela, Plathhelminthes). Микрофауна Марина, 7 , 253–264.
Google Scholar
Элерс, У. (1997). Ультраструктура статоцист трепанга Leptosynapta inhaerens (Holothuroidea, Echinodermata). Acta Zoologica (Копенгаген), 78 (1), 61–68.
Артикул Google Scholar
Фобель А. и Коласа Дж. (1978). К анатомии и морфологии пресноводного вида Acoela (Turbellaria): Limnoposthia polonica (Kolasa et Faubel, 1974). Bulletin de l’Academie Polonaise des Sciences Serie des Sciences Biologiques, 26 (6), 393–397.
Google Scholar
Fransen, ME (1980a). Ультраструктура целомической организации кольчатых червей. 1. Архианнелиды и другие мелкие полихеты. Зооморфология (Берлин), 95 (3), 235–249.
Артикул Google Scholar
Франсен, М. Э.(1980б). Вариации выстилки полости тела полихет. Американский зоолог, 20 (4), 751–751.
Google Scholar
Франсен, А., и Афзелиус, Б.А. (1987). Реснитчатый эпидермис Xenoturbella bocki (Platyhelminthes, Xenoturbellida) с некоторыми филогенетическими соображениями. Zoologica Scripta, 16 (1), 9–17.
Артикул Google Scholar
Фройденштайн, Дж.В. (2005). Характер, состояния и гомологии. Систематическая биология, 54 (6), 965–973. дои: 10.1080/10635150500354654.
ПабМед Статья Google Scholar
Гербер, К.В., Сальвенмозер, В., Ригер, Р.М., и Гшвентнер, Р. (2007). Нервная система Convolutriloba (Acoela) и ее формирование при регенерации после бесполого размножения. Зооморфология (Берлин), 126 (2), 73–87.doi:10.1007/S00435-007-0039-Z.
Артикул Google Scholar
Графф, Л. В. (1882 г.). Monography der Turbellarien. I. Рабдоцелида . Лейпциг: Вильгельм Энгельманн.
Google Scholar
Графф, Л. В. (1891 г.). Die Organization der Turbellaria Acoela . Лейпциг: Вильгельм Энгельманн.
Google Scholar
Графф Л.против (1904–08). Акоэла и рабдоцелида. В HG Bronn (Ed.), Klassen und Ordnungen des Thier-Reichs (том 4, стр. 1732–2597). Лейпциг: CF Winter’sche Verlagshandlung.
Графф, Л. В. (1905). Турбеллария I. Акоэла. В FE Schulze (Ed.), Das Tierreich 23 (стр. 23–34). Берлин: Königliche Preussische Akademie der Wissenschaften.
Google Scholar
Гшвентнер Р., Ладурнер П., Нимет, К., и Ригер, Р. (2001). Стволовые клетки у базальных билатерий: S-фаза и митотические клетки у Convolutriloba longifissura (Acoela, Platyhelminthes). Cell & Tissue Research, 304 (3), 401–408.
КАС Статья Google Scholar
Хэнсон, Э. Д. (1960). Бесполое размножение у бесклеточных турбеллярий. Йельский журнал биологии и медицины, 33 (2), 107–111.
ПабМед КАС Google Scholar
Харш, С.и Ваннингер, А. (2010). Эволюция нервной системы беспозвоночных: Chaetognatha на примере. Acta Zoologica (Копенгаген), 91 (1), 35–43. doi:10.1111/J.1463-6395.2009.00423.X.
Артикул Google Scholar
Хаспрунар, Г. (1996). Plathelminthes и Plathelminthomorpha — парафилетические таксоны. Журнал зоологической систематики и эволюционных исследований, 34 (1), 41–47.
Google Scholar
Хейнол, А.и Мартиндейл, М. К. (2008a). Разработка Acoel поддерживает простой урбилатерий, похожий на планулу. Philosophical Transactions of the Royal Society B-Biological Sciences, 363 (1496), 1493–1501. doi:10.1098/Rstb.2007.2239.
Артикул Google Scholar
Хейнол, А., и Мартиндейл, М. К. (2008b). Развитие Acoel указывает на независимую эволюцию рта и ануса билатерий. Nature (Лондон), 456 (7220), 382–U345.дои: 10.1038/Nature07309.
КАС Статья Google Scholar
Хейнол, А., и Мартиндейл, М. К. (2009). Координированная пространственная и временная экспрессия генов Hox во время эмбриогенеза у acoel Convolutriloba longifissura . БМС Биология, 7 . дои: 10.1186/1741-7007-7-65.
Hejnol, A., Obst, M., Stamatakis, A., Ott, M., Rouse, G.W., Edgecombe, G.D., et al. (2009). Оценка корня билатеральных животных с помощью масштабируемых филогеномных методов. Труды Королевского общества биологических наук, 276 (1677), 4261–4270. doi: 10.1098 / Rspb.2009.0896.
Артикул Google Scholar
Хендельберг, Дж. (1969). О развитии разных типов сперматозоидов из сперматид с двумя жгутиками у Turbellaria с замечаниями об ультраструктуре жгутиков. Zoologiska Bidrag fran Uppsala, 38 , 2–50.
Google Scholar
Хендельберг, Дж.(1977). Сравнительная морфология сперматозоидов турбеллярий, изученная с помощью электронной микроскопии. Acta Zoologica Fennica , 149–162.
Хендельберг, Дж., и Окессон, Б. (1988). Convolutriloba retrogemma род. и др. n., турбеллярий (Acoela, Platyhelminthes) с обратной полярностью репродуктивных почек. Прогресс в зоологии, 36 , 321–327.
Google Scholar
Хендельберг, Дж.и Хедлунд, К.О. (1974). К морфологии эпидермальной цилиарной корешковой системы бескрылой турбеллярии Childia groenlandica . Зун (Уппсала), 2 (1), 13–24.
Google Scholar
Хенниг, В. (1950). Основы теории филогенетической систематики . Берлин: Deutscher Zentral Verlag.
Google Scholar
Хенниг, В.(1965). Филогенетическая систематика. Ежегодный обзор энтомологии, 10 , 97–116.
Артикул Google Scholar
Генри, Дж. К., Мартиндейл, М. К., и Бойер, Британская Колумбия (2000). Уникальная программа развития плоского червя acoel, Neochildia fusca . Биология развития, 220 (2), 285–295.
ПабМед КАС Статья Google Scholar
Голландия, Н.Д. (2003). Ранняя эволюция центральной нервной системы: эпоха кожных мозгов? Nature Reviews Neuroscience, 4 (8), 617–627. дои: 10.1038/Nrn1175.
ПабМед КАС Статья Google Scholar
Hooge, MD (2001). Эволюция мускулатуры стенок тела Platyhelminthes (Acoelomorpha, Catenulida, Rhabditophora). Журнал морфологии, 249 (3), 171–194.
ПабМед КАС Статья Google Scholar
Хуге, М.Д., Хэй П.А., Тайлер С., Литвайтис М.К. и Корнфилд И. (2002). Молекулярная систематика Acoela (Acoelomorpha, Platyhelminthes) и ее соответствие морфологии. Молекулярная филогенетика и эволюция, 24 (2), 333–342.
ПабМед КАС Статья Google Scholar
Hooge, MD, & Rocha, CEF (2006). Acoela (Acoelomorpha) с северных пляжей штата Сан-Паулу, Бразилия, и систематический пересмотр семейства Otocelididae. Zootaxa, 1335 , 1–50.
Google Scholar
Hooge, MD, & Tyler, S. (2005). Новые инструменты для решения филогений: систематический пересмотр Convolutidae (Acoelomorpha, Acoela). Журнал зоологической систематики и эволюционных исследований, 43 (2), 100–113.
Артикул Google Scholar
Гроуда, М. (2007). Молекулярный анализ эволюции осей тела Bilaterian: передача сигналов Wnt и Bmp у Isodiamtera pulchra и Macrostomum lignano (Acoelomorpha, Macrostomorpha; Platyhelminthes) .Диссертация, Университет Инсбрука, Австрия.
Хайман, Л. Х. (1937). Репродуктивная система и совокупление у Amphiscolops langerhansi (Turbellaria acoela). Биологический бюллетень Wood’s Hole, 72 , 319–326.
Артикул Google Scholar
Хайман, Л. Х. (1951). Беспозвоночные: Platyhelminthes и Rhynchocoela. Акоеломат Bilateria (Том 2). Нью-Йорк: McGraw Hill Book Co.
Google Scholar
Хайман, Л. Х. (1959). Беспозвоночные: малые группы кишечнополостных (Том 5). Нью-Йорк: McGraw Hill Book Co.
Google Scholar
Исраэльссон, О. (1999). Новый взгляд на загадочный Xenoturbella (тип неизвестен): онтогенез и филогенез. Труды Лондонского Королевского общества, серия B, биологические науки, 266 (1421), 835–841.
Артикул Google Scholar
Ягерстен, Г. (1972). Эволюция жизненного цикла многоклеточных животных. Комплексная теория . Лондон и Нью-Йорк: Academic Press.
Google Scholar
Дженнингс, Дж. Б. (1971). Паразитизм и комменсализм у турбеллярий. Adv Parasitology, 9 , 1–32.
Артикул Google Scholar
Йонделиус, У., Ларссон, К., и Райкова, О. (2004). Дробление Nemertoderma westbladi (Nemertodermatida) и его филогенетическое значение. Зооморфология (Берлин), 123 (4), 221–225. doi: 10.1007/S00435-004-0105-8.
Артикул Google Scholar
Jondelius, U., Ruiz-Trillo, I., Baguñà, J., & Riutort, M. (2002). Nemertodermatida являются базальными билатериями, а не представителями Platyhelminthes. Zoologica Scripta, 31 (2), 201–215.
Артикул Google Scholar
Jondelius, U., Wallberg, A., Hooge, M., & Raikova, O.I. (2011). Как червь получил глотку: филогения, классификация и байесовская оценка эволюции признаков у Acoela. Систематическая биология, 60 (6), 845–871. doi: 10.1093/Sysbio/Syr073.
ПабМед Статья Google Scholar
Карлинг Т.Г.(1974). Об анатомии и родстве турбеллярских отрядов. В NW Riser & MP Morse (Eds.), Biology of the Turbellaria (стр. 1–16). Нью-Йорк: Макгроу-Хилл.
Google Scholar
Клаузер, доктор медицины, Смит, Дж. П. С., III, и Тайлер, С. (1986). Ультраструктура лобного органа у Convoluta и Macrostomum spp.: значение для моделей турбеллярного архетипа. Гидробиология, 132 , 47–52.
Артикул Google Scholar
Кнаусс, Э. Б. (1979). Указание на наличие анальной поры у Gnathostomulida. Zoologica Scripta, 8 (3), 181–186.
Артикул Google Scholar
Кристенсен, Р. М., и Фанч, П. (2000). Micrognathozoa: новый класс со сложными челюстями, такими как у Rotifera и Gnathostomulida. Журнал морфологии, 246 (1), 1–49.
ПабМед КАС Статья Google Scholar
Ладурнер, П., Пфистер, Д., Зайфарт, К., Шарер, Л., Малкнехт, М., Сальвенмозер, В., и др. (2005). Получение и характеристика клеточно- и тканеспецифичных моноклональных антител к плоскому червю Macrostomum sp. Гистохимия и клеточная биология, 123 (1), 89–104.
ПабМед КАС Статья Google Scholar
Ладурнер, П.и Ригер, Р. (2000). Эмбриональное развитие мышц Convoluta pulchra (Turbellaria — Acoelomorpha, Platyhelminthes). Биология развития, 222 (2), 359–375.
ПабМед КАС Статья Google Scholar
Ланфранки, А. (1990). Ультраструктура эпидермальных пятен платигельминта acoel. Tissue & Cell, 22 (4), 541–546.
КАС Статья Google Scholar
Лундин, К.(1998). Повторное посещение эпидермальных цилиарных корешков Xenoturbella bocki (Xenoturbellida): новая поддержка возможного родства с Acoelomorpha (Platyhelminthes). Zoologica Scripta, 27 (3), 263–270.
Артикул Google Scholar
Лундин, К. (2001). Дегенерирующие клетки эпидермиса у Xenoturbella bocki (тип неизвестен), Nemertodermatida и Acoela (Platyhelminthes). Бельгийский журнал зоологии, 131 , 153–157.
Google Scholar
Лундин, К., и Хендельберг, Дж. (1996). Дегенерирующие эпидермальные тельца («пульсирующие тельца») у Meara stichopi (Plathelminthes, Nemertodermatida). Зооморфология (Берлин), 116 (1), 1–5.
Артикул Google Scholar
Лютер, А. (1912). Studien über acöle Turbellarien aus dem Finnischen Meerbusen. Acta Societatis pro Fauna et Flora Fennica, 36 (5), 1–60.
Google Scholar
Майниц, М. (1983). Гнатостомулида. В Репродуктивная биология беспозвоночных. Том 1. Оогенез, яйцекладка и оосорбция (стр. 169–180). Чичестер, Нью-Йорк и др.: John Wiley & Sons
Маллатт, Дж., Крейг, К.В., и Йодер, М.Дж. (2010). Почти полные гены рРНК, собранные у многоклеточных животных: влияние большего количества таксонов, структурного выравнивания и эволюционных моделей с парными сайтами на реконструкцию филогении. Молекулярная филогенетика и эволюция, 55 (1), 1–17. doi:10.1016/J.Ympev.2009.09.028.
ПабМед Статья Google Scholar
Мамкаев Ю.В. (1967). Очерки морфологии бескрылых Turbellaria. Труди зоол. Инст. Ленингр., 44 , 26–108.
Google Scholar
Маркус, Э. (1947). Turbellarios marinhos do Brasil. Boletim Faculdade de Philosophia e Letras Universidade de Sao Paulo Zoologica, 12 , 99–215.
Google Scholar
Маркус, Э. (1948). Турбеллярия из Бразилии. Boletim da Faculdade de Filosofia Ciencias e Letras Universidade de Sao Paulo Zoologica, 13 , 111–243.
Google Scholar
Маркус, Э. (1949). Турбеллярия бразильская (7). Boletim da Faculdade de Filosofia Ciencias e Letras Universidade de Sao Paulo Zoologica, 14 , 7–155.
Google Scholar
Маркус, Э. (1950). Турбеллария Бразилейрос (8). Boletim da Faculdade de Filosofia Ciencias e Letras Universidade de Sao Paulo Zoologica, 15 , 5–191.
Google Scholar
Маркус, Э. (1951). Турбеллария Бразилейрос. Boletim da Faculdade de Filosofia Ciencias e Letras Universidade de Sao Paulo Zoologica, 16 , 1–215.
Google Scholar
Маркус, Э. (1952). Турбеллария Бразилейрос (10). Boletim da Faculdade de Filosofia Ciencias e Letras Universidade de Sao Paulo Zoologica, 17 , 5–187.
Google Scholar
Маркус, Э. (1954). Турбеллария Бразилейрос. Papéis Avulsos Departamento de Zoologia Secretaria de Agricultura São Paolo, 11 , 419–489.
Google Scholar
Маркус, Э.и Макнэ, В. (1954). Архитомия у вида Convoluta . Nature (Лондон), 173 , 130.
CAS Статья Google Scholar
Марк, Э. Л. (1892 г.). Polychoerus caudatus, nov. ген. и нояб. сп. (Festschrift zum 70. Geburtstage Rudolf Leuckarts) . Лейпциг: Вильгельм Энгельман.
Google Scholar
Марлета Ф., Жиль А., Caubit, X., Perez, Y., Dossat, C., Samain, S., et al. (2008). Транскриптом Chaetognath выявляет наследственные и уникальные особенности билатеральных. Биология генома, 9 (6), doi: 10.1186/Gb-2008-9-6-R94.
Артикул КАС Google Scholar
Марлоу, Х.К., Сривастава, М., Матус, Д.К., Рохсар, Д., и Мартиндейл, М.К. (2009). Анатомия и развитие нервной системы Nematostella vectensis , Anthozoan Cnidarian. Нейробиология развития, 69 (4), 235–254. doi:10.1002/Dneu.20698.
ПабМед КАС Статья Google Scholar
Морено, Э., Де Малдер, К., Сальвенмозер, В., Ладурнер, П., и Мартинес, П. (2010). Вывод о наследственной функции заднего гена Hox в билатериях: контроль за поддержанием репродуктивных структур, мускулатуры и нервной системы у плоского червя acoel Isodiametra pulchra . Эволюция и развитие, 12 (3), 258–266. doi:10.1111/J.1525-142x.2010.00411.X.
КАС Статья Google Scholar
Морено, Э., Надаль, М., Багунья, Дж., и Мартинес, П. (2009). Отслеживание происхождения билатеральной системы формирования Hox-паттернов: идеи плоского червя acoel Symsagittifera roscoffensis . Эволюция и развитие, 11 (5), 574–581. doi: 10.1111/J.1525-142x.2009.00363.Х.
КАС Статья Google Scholar
Морено, Э., Перманьер, Дж., и Мартинес, П. (2011). Происхождение систем формирования паттернов у билатерий — понимание генов Hox и ParaHox у Acoelomorpha. Геномика Протеомика Биоинформатика, 9 (3), 65–76. doi: 10.1016/S1672-0229(11)60010-7.
ПабМед КАС Статья Google Scholar
Мвиньи, А., Bailly, X., Bourlat, SJ, Jondelius, U., Littlewood, DTJ, & Podsiadlowski, L. (2010). Филогенетическое положение Acoela, выявленное с помощью полного митохондриального генома Symsagittifera roscoffensis . BMC Эволюционная биология, 10 . дои: 10.1186/1471-2148-10-309.
Pardos, F. (1988) Тонкая структура и функция ресничек глотки у Glossobalanus minutus Kowalewsky (Enteropneustea). Acta Zoologica (Копенгаген), 69 (1), 1–12.
Артикул Google Scholar
Педерсен, К.Дж., и Педерсен, Л.Р. (1986). Тонкие ультраструктурные наблюдения внеклеточного матрикса (ECM) Xenoturbella bocki Westblad, 1949. Acta Zoologica (Copenhagen), 67 (2), 103–113.
Артикул Google Scholar
Педерсен, К.Дж., и Педерсен, Л.Р. (1988). Ультраструктурные наблюдения эпидермиса Xenoturbella bocki Westblad, 1949; с обсуждением систем эпидермальных цитоплазматических филаментов беспозвоночных. Acta Zoologica (Копенгаген), 69 (4), 231–246.
Артикул Google Scholar
Петерсон, К. Дж., Кэмерон, Р. А., Тагава, К., Сато, Н., и Дэвидсон, Э. Х. (1999). Сравнительный молекулярный подход к формированию мезодермального паттерна у базальных вторичноротых: паттерн экспрессии Brachyury в полухордовых enteropneust, Ptychodera flava . Девелопмент (Кембридж), 126 (1), 85–95.
КАС Google Scholar
Петров А., Хуге, М., и Тайлер, С. (2004). Ультраструктура сперматозоидов Acoela (Acoelomorpha) и ее соответствие молекулярной систематике. Биология беспозвоночных, 123 (3), 183–197.
Артикул Google Scholar
Петров А., Хуге М. и Тайлер С. (2006). Сравнительная морфология бурсальных сопел у акоэлей (Acoela, Acoelomorpha). Журнал морфологии, 267 (5), 634–648.
ПабМед Статья Google Scholar
Филипп, Х., Brinkmann, H., Copley, R.R., Moroz, L.L., Nakano, H., Poustka, A.J., et al. (2011). Ацеломорфные плоские черви — вторичноротые, родственные Xenoturbella . Nature (Лондон), 470 (7333), 255–258.
КАС Статья Google Scholar
Плейель, Ф. (1995). О кодировании символов для реконструкции филогении. Кладистика, 11 (3), 309–315.
Артикул Google Scholar
Попова Н.В. и Мамкаев Ю.В. (1985). Ультраструктура и примитивные признаки глаз Convoluta convoluta (Turbellaria Acoela). ДАН СССР, 283 (3), 756–759.
Google Scholar
Райкова О.И., Фаллени А. и Жюстин Ж.-Л. (1997). Спермиогенез у Paratomella rubra (Platyhelminthes, Acoela): ультраструктурные, иммуноцитохимические, цитохимические исследования и филогенетические последствия. Acta Zoologica (Копенгаген), 78 (4), 295–307.
Артикул Google Scholar
Райкова О.И., Рейтер М., Густафссон М.К.С., Мауле А.Г., Халтон Д.В. и Джонделиус У. (2004a). Эволюция нервной системы у Paraphanostoma (Acoela). Zoologica Scripta, 33 (1), 71–88.
Артикул Google Scholar
Райкова О.И., Ройтер, М., Густафссон, М.К.С., Мауле, А.Г., Халтон, Д.В., и Джонделиус, У. (2004b). Базиэпидермальная нервная система у Nemertoderma westbladi (Nemertodermatida): GYIRFамидная иммунореактивность. Зоология, 107 (1), 75–86. doi: 10.1016/J.Zool.2003.12.002.
ПабМед Статья Google Scholar
Райкова О.И., Рейтер М., Йонделиус У. и Густафссон М.К.С. (2000a). Мозг Nemertodermatida (Platyhelminthes), выявленный иммуноокрашиванием анти-5HT и анти-FMRFамидом. Ткани и клетки, 32 (5), 358–365.
КАС Статья Google Scholar
Райкова О.И., Рейтер М., Йонделиус У. и Густафссон М.К.С. (2000b). Иммуноцитохимическое и ультраструктурное исследование нервной и мышечной систем Xenoturbella westbladi (Bilateria inc. sed.). Зооморфология (Берлин), 120 (2), 107–118.
Артикул Google Scholar
Райкова О.И., Рейтер М. и Жюстин Ж.-Л. (2001). Вклад в филогению и систематику Acoelomorpha. В DTJ Littlewood & RA Bray (Eds.), Interrelationships of the Platyhelminthes (том 60, стр. 13–23). Лондон и Нью-Йорк: Серия специальных томов Ассоциации систематики.
Google Scholar
Райкова О.И., Рейтер М., Котикова Е.А. и Густафссон М.К.С. (1998). Комиссуральный мозг! Характер иммунореактивности 5-НТ у Acoela (Plathelminthes) [Plathelminthes]. Зооморфология (Берлин), 118 (2), 69–77.
Артикул Google Scholar
Рамачандра, Н. Б., Гейтс, Р. Д., Ладурнер, П., Джейкобс, Д. К., и Хартенштейн, В. (2002). Эмбриональное развитие у примитивных билатеральных Neochildia fusca : нормальный морфогенез и изоляция генов POU Brn-1 и Brn-3. Гены развития и эволюция, 212 (2), 55–69.
ПабМед КАС Статья Google Scholar
Райзингер, Э.(1960). Был ли ист Ксенотурбелла? Zeitschrift fuer Wissenschaftliche Zoologie, 164 , 188–198.
Google Scholar
Райзингер, Э. (1961). Allgemeine Morphologie der Metazoen. Morphologie der Coelenteratcn, acoelomaten und pseudocoelomaten Würmer. Fortschritte der Zoologie, 13 , 1–82.
Google Scholar
Ремане А.(1963). Эволюционное происхождение целома. В EC Dougherty (Ed.), Нижние Metazoa . Лос-Анджелес: Издательство Калифорнийского университета.
Google Scholar
Рейтер, М., Райкова, О.И., и Густафссон, М.К.С. (1998). Эндокринный мозг? Характер иммунореактивности FMRF-амида у Acoela (Plathelminthes). Ткани и клетки, 30 (1), 57–63.
КАС Статья Google Scholar
Рейтер, М., Райкова, О.И., и Густафссон, М.К.С. (2001a). Закономерности нервной и мышечной систем у низших плоских червей. Бельгийский журнал зоологии, 131 , 47–53.
Google Scholar
Рейтер, М., Райкова, О.И., Йонделиус, У., Густафссон, М.К.С., Мауле, А.Г., и Халтон, Д.В. (2001b). Организация нервной системы у Acoela: иммуноцитохимическое исследование. Ткани и клетки, 33 (2), 119–128.
КАС Статья Google Scholar
Рихтер С., Loesel, R., Purschke, G., Schmidt-Rhaesa, A., Scholtz, G., Stach, T., et al. (2010). Нейрофилогения беспозвоночных: предлагаемые термины и определения для нейроанатомического глоссария. Frontiers in Zoology, 7 , 29.
PubMed Статья Google Scholar
Ригер, Р. М. (1980). Новая группа интерстициальных червей Lobatocerebridae nov. фам. (Annelida) и его значение для филогении многоклеточных животных. Зооморфология (Берлин), 95 (1), 41–84.
Артикул Google Scholar
Ригер, Р. М. (1985). Филогенетический статус организации acoelomat внутри Bilateria: гистологическая перспектива. В S. Conway Morris, JD George, R. Gibson, & HM Platt (Eds.), Происхождение и взаимоотношения низших беспозвоночных (Том 28, стр. 101–122). Оксфорд: Кларендон Пресс.
Google Scholar
Ригер Р.М. (1986). Über den Ursprung der Bilateria: die Bedeutung der Ultrastrukturforschung für ein neues Verstehen der Metazoenevolution. Verhandlungen der Deutschen Zoologischen Gesellschaft, 79 , 31–50.
Google Scholar
Ригер, Р. М., и Ладурнер, П. (2003). Значение мышечных клеток для происхождения мезодермы Bilateria. Интегративная и сравнительная биология, 43 (1), 47–54.
ПабМед Статья Google Scholar
Ригер Р.М., Тайлер С., Смит, Дж. П. С., III, и Ригер, Г. Э. (1991). Платигельминты: турбеллярии. В FW Harrison & BJ Bogitsh (Eds.), Микроскопическая анатомия беспозвоночных. Том 3. Platyhelminthes и Nemertinea (Том 3, стр. 7–140). Нью-Йорк: Wiley-Liss Inc.
Google Scholar
Райзер, СЗ (1987). Nemertinoides elongatus род. п., сп. н. (Turbellaria: Nemertodermatida) с крупных песчаных пляжей в западной части Северной Атлантики. Труды Вашингтонского гельминтологического общества, 54 (1), 60–67.
Google Scholar (1999). Плоские черви Acoel: самые ранние из существующих билатеральных многоклеточных животных, а не представители Platyhelminthes. Science (Вашингтон, округ Колумбия), 283 (5409), 1919–1923.
КАС Статья Google Scholar
Руис-Трильо, М., Риуторт, М., Фуркад, Х.М., Багунья, Дж., и Бур, Дж.Л. (2004). Данные митохондриального генома подтверждают базальное положение Acoelomorpha и полифилию Platyhelminthes. Молекулярная филогенетика и эволюция, 33 (2), 321–332.
ПабМед КАС Статья Google Scholar
Рупперт, Э. Э., Фокс, Р. С., и Барнс, Р. Д. (2004). Зоология беспозвоночных, функциональный эволюционный подход (7.ред.). Белмонт, Калифорния ; Лондон: Томсон Брукс/Коул.
Салвенмозер В., Эггер Б., Ахац Дж. Г., Ладурнер П. и Хесс М. В. (2010). Электронная микроскопия плоских червей: стандартный и криопрепаративный методы. В Т. Мюллер-Райхерт (ред.), Methods in Cell Biology (том 96, стр. 307–330): Academic Press.
Сальвини-Плавен, Л. (1978). О происхождении и эволюции низших Metazoa. Zeitschrift fuer Zoologische Systematik und Evolutionsforschung, 16 (1), 40–88.
Артикул Google Scholar
Шерер, Л., Литтлвуд, Д.Т.Дж., Вашенбах, А., Йошида, В., и Визосо, Д.Б. (2011). Брачное поведение и эволюция дизайна сперматозоидов. Труды Национальной академии наук Соединенных Штатов Америки, 108 (4), 1490–1495. doi:10.1073/Pnas.10138.
ПабМед Статья Google Scholar
Шимкевич, В.(1891 г.). Versuch Einer Klassiffikation des Tierreichs. Biologisches Zentralblatt, 11 , 291–295.
Google Scholar
Шухерт П. и Ригер Р. М. (1990). Ультраструктурные наблюдения карликового самца Bonellia viridis (Echiura). Acta Zoologica (Копенгаген), 71 (1), 5–16.
Артикул Google Scholar
Земмлер, Х., Байи, X., и Ваннингер, А. (2008). Миогенез у базальных билатерий Symsagittifera roscoffensis (Acoela). Границы зоологии, 5 (14).
Земмлер, Х., Чиодин, М., Байи, X., Мартинес, П., и Ваннингер, А. (2010). Шаги к централизованной нервной системе у базальных билатеральных: выводы из нейрогенеза acoel Symsagittifera roscoffensis . Развитие, рост и дифференциация, 52 (8), 701–713.
КАС Статья Google Scholar
Семпере, Л.Ф., Коул, К.Н., МакПик, Массачусетс, и Петерсон, К.Дж. (2006). Филогенетическое распространение микроРНК многоклеточных животных: понимание эволюционной сложности и ограничений. Journal of Experimental Zoology Part B Molecular and Developmental Evolution, 306B (6), 575–588.
КАС Статья Google Scholar
Семпере, Л. Ф., Мартинес, П., Коул, К., Багуна, Дж., и Петерсон, К. Дж. (2007). Филогенетическое распределение микроРНК подтверждает базальное положение плоских червей acoel и полифилию Platyhelminthes. Эволюция и развитие, 9 (5), 409–415.
КАС Статья Google Scholar
Шеннон, Т., III, и Ахатц, Дж. Г. (2007). Convolutriloba macropyga sp. nov., необычайно плодовитый ацеломорф (Acoelomorpha), обнаруженный в тропических аквариумах. Zootaxa, 1525 , 1–17.
Google Scholar
Сайкс, Дж. М., и Белый, А.Э. (2008). Радикальная модификация оси AP и эволюция бесполого размножения у Convolutriloba acoels. Эволюция и развитие, 10 (5), 619–631.
КАС Статья Google Scholar
Сайкс, Дж. М., и Белый, А. Э. (2010). Изготовление головы из хвоста: развитие перевернутой передне-задней оси во время почкования у ацеля. Биология развития, 338 (1), 86–97.
ПабМед КАС Статья Google Scholar
Смит, Дж.PS (1981). Тонкоструктурная анатомия паренхимы Acoela и Nemertodermatida (Turbellaria) . Диссертация, Университет Северной Каролины, Чапел-Хилл.
Смит, Дж. П. С., и Тайлер, С. (1985a). Acoel turbellarians: главные стержни эволюции многоклеточных животных или специализированное ответвление. В S. Conway Morris, JD George, R. Gibson, & HM Platt (Eds.), Происхождение и взаимоотношения низших беспозвоночных (Том 27, стр. 123–142). Оксфорд: Кларендон Пресс.
Google Scholar
Смит, Дж. П. С., и Тайлер, С. (1985b). Тонкая структура и эволюционные последствия лобного органа Turbellaria Acoela. 1. Diopisthoporus gymnopharyngeus sp. н. Zoologica Scripta, 14 (2), 91–102.
КАС Статья Google Scholar
Смит, Дж. П. С., и Тайлер, С. (1986). Лобные органы Acoelomorpha (Turbellaria): ультраструктура и филогенетическое значение. Гидробиология, 132 , 71–78.
Артикул Google Scholar
Смит, Дж. П. С., Тайлер, С., и Ригер, Р. М. (1986). Является ли Turbellaria полифилетическим? Гидробиология, 132 , 13–21.
Артикул Google Scholar
Смит, Дж. П. С., Тайлер, С., Томас, М. Б., и Ригер, Р. М. (1982). Морфология турбеллярных рабдитов: филогенетические последствия. Труды Американского микроскопического общества, 101 (3), 209–228.
Артикул Google Scholar
Стах, Т., Дюпон, С., Израэльсон, О., Фовиль, Г., Накано, Х., Каннеби, Т., и др. (2005). Нервные клетки Xenoturbella bocki (филум неизвестен) и Harrimania kupfferi (Enteropneusta) обладают положительной иммунореактивностью к антителам, вырабатываемым против нейропептидов иглокожих. Журнал Морской биологической ассоциации Соединенного Королевства, 85 (6), 1519–1524.
КАС Статья Google Scholar
Стейнбёк, О. (1966). Die Hofsteniiden (Turbellaria Acoela). Основы эволюции турбеллярии. Zeitschrift für Zoologische Systematik und Evolutionsforschung, 4 , 58–195.
Артикул Google Scholar
Стеррер, В. (1998). Новые и известные Nemertodermatida (Platyhelminthes-Acoelomorpha) — ревизия. Бельгийский журнал зоологии, 128 (1), 55–92.
Google Scholar
Струк, Т. Х., Пол, К., Хилл, Н., Хартманн, С., Хозель, К., Кубе, М., и др. (2011). Филогеномный анализ раскрывает эволюцию кольчатых червей. Nature (Лондон), 471 (7336), 95–98.
КАС Статья Google Scholar
Тагучи С., Тагава К., Хамфрис Т. и Сато Н.(2002). Гены Sox группы B, которые вносят вклад в спецификацию мозга позвоночных, экспрессируются в апикальном органе и цилиарных тяжах личинок полухордовых. Zoological Science (Токио), 19 (1), 57–66.
КАС Статья Google Scholar
Текле Ю.И., Райкова О.И., Ахмадзаде А. и Джонделиус У. (2005). Пересмотр Childiidae (Acoela), комплексный подход к реконструкции филогении акоэлей с перевернутыми мышечными слоями. Журнал зоологической систематики и эволюционных исследований, 43 (1), 72–90.
Артикул Google Scholar
Текле Ю.И., Райкова О.И., Жюстин Дж.-Л. и Йонделиус У. (2007). Ультраструктура и иммуноцитохимия тубулина копулятивной стилетоподобной структуры у видов Childia (Acoela). Журнал морфологии, 268 (2), 166–180.
ПабМед Статья Google Scholar
Телфорд, М.Дж., Локьер, А.Е., Картрайт-Финч, К., и Литтлвуд, Д.Т.Дж. (2003). Комбинированные филогении рибосомных РНК больших и малых субъединиц подтверждают базальное положение ацеломорфных плоских червей. Труды Лондонского королевского общества, серия B-Biological Sciences, 270 (1519), 1077–1083. doi: 10.1098/Rspb.2003.2342.
КАС Статья Google Scholar
Тодт, К. (2009). Структура и эволюция глотки симплекса у плоских червей acoel (Acoela). Журнал морфологии, 270 (3), 271–290.
ПабМед Статья Google Scholar
Тодт, К., и Тайлер, С. (2007). Цилиарные рецепторы, связанные со ртом и глоткой Acoela (Acoelomorpha): сравнительное ультраструктурное исследование. Acta Zoologica (Копенгаген), 88 (1), 41–58.
Артикул Google Scholar
Тайлер, С. (1979).Отличительные черты ресничек метазоонов и их систематическое значение. Ткани и клетки, 11 (3), 385–400.
КАС Статья Google Scholar
Тайлер, С. (2001). Ранний червь: происхождение и взаимоотношения низших плоских червей. В DTJ Littlewood & RA Bray (Eds.), Interrelationships of the Platyhelminthes (том 60, стр. 3–12). Лондон и Нью-Йорк: Серия специальных томов Ассоциации систематики.
Google Scholar
Тайлер, С., Гримм, Дж. К., и Смит, Дж. П. С. (1989). Динамика заживления эпидермальных ран у Acoel Turbellarians – роль пульсирующих тел. Американский зоолог, 29 (4), A115–A115.
Google Scholar
Тайлер С. и Ригер Р. М. (1974). Одножгутиковые сперматозоиды Nemertoderma (Turbellaria) и их филогенетическое значение. Science (Вашингтон, округ Колумбия), 188 , 730–732.
Артикул Google Scholar
Тайлер С. и Ригер Р. М. (1977). Ультраструктурные доказательства систематического положения Nemertodermatida (Turbellaria). Acta Zoologica Fennica , 194–207.
Тайлер С. и Ригер Р. М. (1999). Функциональная морфология мускулатуры акоеломного червя Convoluta pulchra (Plathelminthes). Зооморфология (Берлин), 119 (3), 127–141.
Артикул Google Scholar
Тайлер С., Шиллинг С., Хуге М. и Буш Л. Ф. (2012). Таксономическая база турбеллярий. Версия 1.7. http://turbellaria.umaine.edu.
Ульянин В. Н. (1870). Die Turbellarien der Bucht von Sebastopol. Арбайтен дер 2 . Versammlung russischer Naturforscher zu Moskau, 1869 , 1–96.
Google Scholar
Ульрих В.(1951). Vorschlage zu einer Revision der Grosseinteilung des Tierreiches. Verhandlungen der Deutschen Zoologischen Gesellschaft, 1950 , 244–271.
Google Scholar
Валлберг, А. (2012). Стилет — разнообразие и систематика Acoela и Nemertodermatida. http://acoela.myspecies.info.
Валберг, А., Курини-Галетти, М., Ахмадзаде, А., и Джонделиус, У. (2007). Отказ от Acoelomorpha: Acoela и Nemertodermatida являются отдельными ранними кладами билатерий. Zoologica Scripta, 36 (5), 509–523.
Артикул Google Scholar
Webster, B.L., Copley, R.R., Jenner, R.A., Mackenzie-Dodds, J.A., Bourlat, S.J., Rota-Stabelli, O., et al. (2006). Митогеномика и филогеномика выявляют приапулидных червей как существующие модели предков экдизозоя. Эволюция и развитие, 8 (6), 502–510.
Артикул Google Scholar
Вестблад, Э.(1940). Studien über skandinavische Turbellaria Acoela. 1. Архив зоологии, 32А (20), 1–28.
Google Scholar
Вестблад, Э. (1942). Studien über skandinavische Turbellaria Acoela. 2. Архив зоологии, 33А (14), 1–48.
Google Scholar
Вестблад, Э. (1945). Studien über skandinavische Turbellaria Acolea. 3. Архив зоологии, 36А (5), 1–56.
Google Scholar
Вестблад, Э. (1946). Studien über skandinavische Turbellaria Acoela. 4. Архив зоологии, 38А (1), 1–56.
Google Scholar
Вестблад, Э. (1948). Studien über Skandinavische Turbellaria Acoela. 5. Архив зоологии, 41А (7), 1–82.
Google Scholar
Вестблад, Э.(1949). Xenoturbella bocki н. г., н. сп. своеобразный, примитивный турбеллярный тип. Архив зоологии, 1 , 11–29.
Google Scholar
Вестхайде, В., и Ригер, Р. М. (2007). Систематик-Плакат: Зоология . Гейдельберг: Spektrum Akademischer Verlag.
Google Scholar
Винс, Дж. Дж. (2001). Анализ признаков в морфологической филогенетике: проблемы и решения. Систематическая биология, 50 (5), 689–699.
ПабМед КАС Статья Google Scholar
Винзор, Л. (1988). Новый acoel (Convolutidae) с северного побережья Квинсленда, Австралия. Прогресс в зоологии, 36 , 391–394.
Google Scholar
Сян, К.Ю., и Томас, Д.Т. (2008).
