
Растения, царство
Царство Растения. В это царство входит подавляющее большинство современных фотосинтезирующих автотрофных живых организмов. Это определяет их главенствующую роль в биосфере как продуцирующих первичное органическое вещество и высвобождающих молекулярный кислород. Если в одной среде водоросли делят эту функцию с цианобактериями (фотосинтезирующие бактерии кислород не выделяют), то в наземных экосистемах продукция кислорода осуществляется исключительно растениями.[ …]
РАСТЕНИЯ [Р1аШае, или VegetaЫ-Иа] — группа (царство) автотроф-ных (кроме паразитов) организмов, размножающихся спорами, семенами и вегетативными частями, обладающих целлюлозными стенками и потому не способных к активному передвижению. Как правило, растут всю жизнь , представлены на Земле около 350 тыс. видов. См. Типы растительности. РАСТИТЕЛЬНОСТЬ АЗОНАЛЬНАЯ — растительность, встречающаяся везде, не принадлежащая исключительно к.-л. конкретной ландшафтной зоне.[ …]
Царство Грибы — Mycota (Fungi).
Это царство представлено организмами, клетки которых обладают плотными клеточными стенками и которые способны к фотосинтезу. Растения этого царства классифицируют на три подцарства, а именно: багрянки (РЬусоЬю а), настоящие водоросли (РЬусоЫо а) и высшие растения (Embryophyta).
Царство растений включает более 300 тыс. видов, из них покрытосеменных — 200 тыс. видов, голосеменных — 600—770 видов, папоротникообразных—10—11 тыс. видов, мхов — 25—27 тыс. видов, лишайников — 30 тыс. видов, водорослей — 25 тыс. видов.[ …]
Растения — эукариотические автотрофные фотосинтезирующие организмы. Царство растений насчитывает около 500 тыс. видов. Растения являются продуцентами органических веществ и основным источником энергии для других живых организмов. Любые пищевые цепи начинаются с зеленых растений. Они же определяют характер биоценоза, защищают почву от эрозии. Растения служат источником кислорода воздуха и оказывают значительное влияние на климат Земли. Человек использует около 1,5 тыс. видов культурных растений как пищевые, технические и лекарственные ресурсы.[ …]
Растения являются автотрофными (потребляющими органические вещества, полученными путем преобразования из неорганических) живыми организмами, для которых характерны способность к фотосинтезу и наличие плотных клеточных оболочек, состоящих обычно из целлюлозы. Они, как правило, не способны к активному передвижению. Растения являются основными поставщиками кислорода в атмосферу и потребителями углекислого газа. Они также составляют значительную часть пищевого рациона многих видов животных и людей. Царство растений включает более 350 тыс. научно описанных видов.[ …]
Они, как правило, не способны к активному передвижению. Растения являются основными поставщиками кислорода в атмосферу и потребителями углекислого газа. Они также составляют значительную часть пищевого рациона многих видов животных и людей. Царство растений включает более 350 тыс. научно описанных видов.[ …]
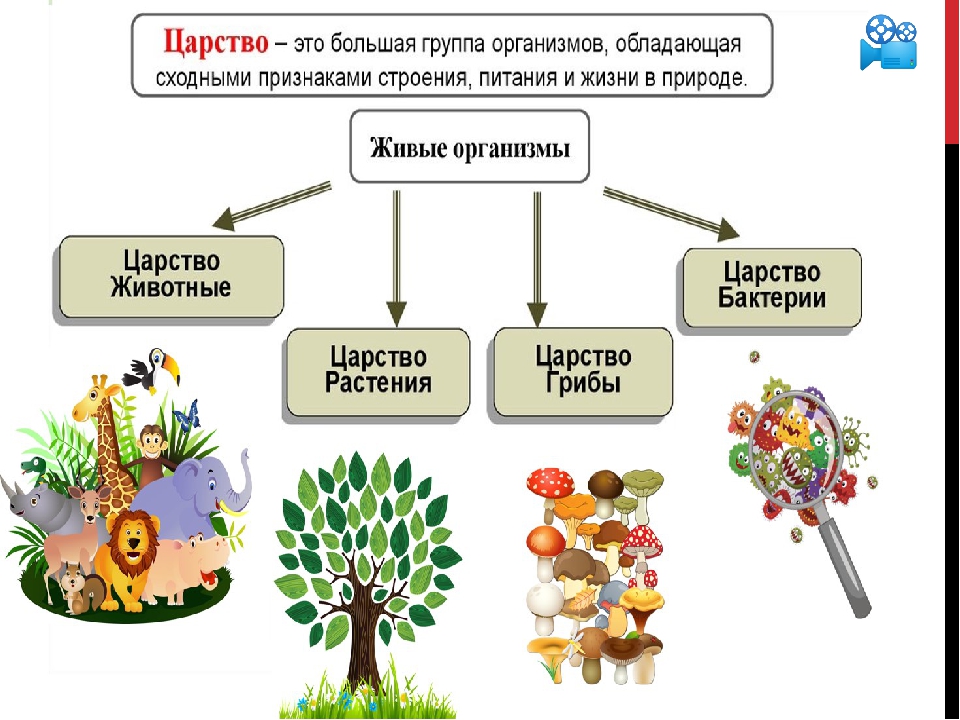
ЦАРСТВО — в биологии высшая таксономическая категория в системе организмов (Ц. животных, Ц. растений).[ …]
РАСТЕНИЯ (Р.) — организмы, образующие отдельное царство живых организмов. Характеризуются разной сложностью организации. Размножаются спорами (мхи, плауны, хвощи, папоротники) или семенами (голосеменные, цветковые). К Р. также относятся разнообразные водоросли, входящие в состав водных экосистем и обитающие в почве. Условно к Р. относятся лишайники.[ …]
Царство растений разделяется примерно на дюжину главных отделов. Наиболее развитым, т. е. эволюционировавшим в наиболее позднее время, является отдел, с которым непосредственно связан садовод. Он включает так называемые высшие растения, т. е. растения, имеющие корни, стебли, листья и сосудистую систему, или трахеи-ды (этим и объясняется их название ТгасЬеорЬу1а, т е. сосудистые). Остальные отделы почти не содержат садовых растений как таковых (за исключением съедобных грибов) и поэтому представляют для нас преимущественно косвенный интерес. Зато они ответственны за болезни многих наших культур.[ …]
е. растения, имеющие корни, стебли, листья и сосудистую систему, или трахеи-ды (этим и объясняется их название ТгасЬеорЬу1а, т е. сосудистые). Остальные отделы почти не содержат садовых растений как таковых (за исключением съедобных грибов) и поэтому представляют для нас преимущественно косвенный интерес. Зато они ответственны за болезни многих наших культур.[ …]
В царствах дробянок, грибов и животных, как правило, существует иная закономерность: потенциальный запас жизненных ресурсов огромен — обычно используется не более 30—60% наземной луговой растительности (крупные млекопитающие) и 10% лесной растительности1. Более интенсивно и полно утилизируется растительноядными животными чистая продукция водных растений. Как хорошо известно, пирамида биомасс в водной среде как бы перевернута — продуценты размножаются столь интенсивно, что консументы могут превышать их по биомассе. Однако это касается лишь мелких обитателей водной среды. Для рыб, птиц и млекопитающих общая их биомасса всегда намного ниже, чем потребляемых ими растительных кормов.
Капское царство — самое маленькое среди флористических царств Земли, но благодаря исключительному своеобразию флоры и самостоятельности ее развития все географы растений единодушно отделяют его от остальной флоры Африки. Оно состоит лишь из одной области — Капской.[ …]
Экологию (царство) растений, в соответствии с их общепринятой (ботанической) классификацией, очевидно, следует разделить на экологии (подцарства) багрянок, настоящих водорослей, высших растений.[ …]
В животном царстве мейоз ведет к образованию половых клеток — гамет и обычно только эти клетки содержат гаилоидный набор хромосом. У растений мейоз может происходить на разных этапах жизненного цикла, причем в качестве гаплоидных продуктов у них образуются как половые клетки — гаметы, так и бесполовые споры. Продолжительность жизни гамет ограничена и составляет от нескольких минут до нескольких суток, после чего неоплодотворенные гаметы пропадают.[ …]
Итак, из двух царств эукариот растения являются создателями органических веществ, а грибы замыкают круговорот, подготавливая питание для автотрофов. [ …]
[ …]
Существуют три царства: царство бактерий, царство животных, царство растений; место первого из них — самая нижняя граница двух других. Вирусы — большие молекулы нуклеиновой кислоты в комбинации с белком — стоят на грани между живой и неживой природой. Они образуют свой собственный мир, совершенно отличный от «клеточной» жизни, представленной тремя уже упомянутыми царствами.[ …]
Функциональные царства сообществ не могут быть идентичными царствам в систематике, но они в значительной степени составляют их эволюционную основу. В традиционной системе Из двух царств царство животных включало одновременно и одноклеточные, и многоклеточные организмы, характеризующиеся способностью к заглатыванию пищи и подвижностью, а обширное царство растений объединяло главным образом фотосинтезирующие группы и (несколько произвольно скооперированные с ними) бактерии и грибы. В современном альтернативном делении форм жизни бактерии (вместе с синезелеными водорослями) образуют царство Мопега, являясь прокариотами, тогда как высшие растения и животные также составляют царства, характеризующиеся направлением эволюции, которое рассмотрено выше.
Семена многих высших растений распространяются птицами. Птиц привлекает наличие мясистой сахаристой ткани, которую они поедают и переваривают. В ткань эту погружены семена, защищенные толстыми оболочками; они проходят через пищеварительный тракт неповрежденными, а иногда пребывание в нем даже стимулирует их прорастание. У различных представителей растительного царства мякоть плодов ведет свое происхождение от совершенно разных структур. Ряд примеров приведен на рис. 1.10. Многие из изображенных растений принадлежат к далеко отстоящим друг от друга филогенетическим линиям, например тисс — голосеменное растение, а все остальные — покрытосеменные. Впрочем, даже в пределах одного лишь семейства розоцветных для образования привлекательной мясистой оболочки семени «задействованы» весьма разнообразные структуры. Земляника, яблоня, персик — все они образуют привлекательную для птиц мякоть и (или) защищают свои семена совершенно различными способами.
Животные питаются либо растениями, либо другими животными, так что в конечном счете вещества, из которых строится их организм, берутся из воздуха или из минерального царства.[ …]
В современной классификации растений и животных имеется много спорных вопросов, т. к. одни биологи склонны укрупнять систематические единицы, тогда как другие стремятся их детализировать. Поэтому существует несколько классификаций как растений, так и животных. В приводимом ниже описании разнообразия организг ов используется классификация, исходным моментом которой является разделение живого мира на царства растений и животных.[ …]
В пределах этого надцарства у растений выделяют царство грибов и царство растений.[ …]
Эволюция многоклеточных животных. Царство животных н менее разнообразно, чем царство растений, а по числу видов животные превосходят растения. В настоящее время описано более 1,5 миллиона видов животных.[ …]
Эукариотов чаще всего делят на два царства — животных и растений. С царством животных дело обстоит сравнительно просто, если не считать вопроса о положении отдельных групп жгутиконосцев, в том числе эвглено-вых, которых некоторые зоологи продолжают по традиции относить к простейшим животным. Гораздо сложнее обстоит дело с царством растений. Мы уже видели, что из царства растений должны быть исключены все прокариоты, в том числе цианеи (сине-зеленые водоросли), что теперь редко у кого вызывает возражения. Более спорно положение грибов в системе организмов. Хотя в учебниках ботаники грибы все еще относятся по традиции к растениям, еще в первой половине прошлого столетия известный миколог Е. Фриз (микология — наука о грибах) предложил выделить их в отдельное самостоятельное царство грибов, что впоследствии было принято целым рядом крупных ботаников и специалистов-микологов. В настоящее время большинство микологов считает, что происхождение грибов от какой бы то ни было группы растений чрезвычайно мало вероятно. Гораздо более вероятным признается происхождение грибов от исконно гетеротрофных, амебоидных двужгутиковых простейших, о чем писал еще в 1884 г.
С царством животных дело обстоит сравнительно просто, если не считать вопроса о положении отдельных групп жгутиконосцев, в том числе эвглено-вых, которых некоторые зоологи продолжают по традиции относить к простейшим животным. Гораздо сложнее обстоит дело с царством растений. Мы уже видели, что из царства растений должны быть исключены все прокариоты, в том числе цианеи (сине-зеленые водоросли), что теперь редко у кого вызывает возражения. Более спорно положение грибов в системе организмов. Хотя в учебниках ботаники грибы все еще относятся по традиции к растениям, еще в первой половине прошлого столетия известный миколог Е. Фриз (микология — наука о грибах) предложил выделить их в отдельное самостоятельное царство грибов, что впоследствии было принято целым рядом крупных ботаников и специалистов-микологов. В настоящее время большинство микологов считает, что происхождение грибов от какой бы то ни было группы растений чрезвычайно мало вероятно. Гораздо более вероятным признается происхождение грибов от исконно гетеротрофных, амебоидных двужгутиковых простейших, о чем писал еще в 1884 г.
Грибы — это выделенные в отдельное царство живые организмы. Они объединяют около 100 тысяч видов и представляют собой гетеротрофные организмы, схожие по ряду признаков как с растениями, так и с животными. Они имеют клеточную оболочку, неподвижны и питаются за счет всасывания необходимых веществ, имеют неограниченный рост,— все это сближает их с растениями. В то же время грибы не способны синтезировать органические вещества из неорганических, не имеют хлоропластов и это делает их схожими с животными. Грибы делят на высшие и низшие. Из последних всем хорошо известны дрожжи и плесневелые, а высшие—это те грибы, которые в виде шляпочных произрастают повсеместно на Земле при наличии питательной среды и оптимальных условий. По способу питания выделяют сапрофиты, питающиеся различного рода органическими остатками, и паразиты, живущие на других организмах.[ …]
По способу питания выделяют сапрофиты, питающиеся различного рода органическими остатками, и паразиты, живущие на других организмах.[ …]
Суккулентами (лат. succus — сок) называют растения с сочными, мясистыми и утолщенными стеблями, листьями, цветками. Они встречаются в различных семействах растительного царства и обитают главным образом в засушливых областях Африки и Америки. Облик суккулентов, превративших свои надземные органы во вместилища запасной воды, обусловлен сходными путями морфологической эволюции под влиянием изменения условий внешней среды и становления засушливого климата. Обретенные особенности строения помогают суккулентам, экономно расходуя воду, выдерживать длительные периоды засухи и зноя.[ …]
Все разнообразие форм роста цветковых растений можно свести к двум основным типам — д р е в е с и о му и т р а вин и с т о м у. Древесные растения характерны для лесов и различного рода кустарниковых сообществ, по отсутствуют пли играют .лишь второстепенную роль в степях, лугах и родственных пм сообществах, где царствуют травм. Имеется огромное разнообразно древесных и особенно травянистых растений, которые, в свою очередь, подразделяются па отдельные типы форм роста, пли жизненных форм. Существуют также промежуточные формы между дроиосиыми и травяпистыми растениями, которые не всегда легко отнести к тому или иному типу. В таких случаях ответ часто может дать только специальное анатомическое исследование проводящей системы стебля, ее ксилемы. Дело в том, что для древесных растений, будь то огромные деревья или небольшие кустарнички, характерно Наличие активно действующего камбия, производящего в стеблях и корнях ясно выраженные слои вторичной ксилемы. У травянистых растений, как привило, активный камбий отсутствует или же имеется в зачаточной форме.[ …]
Имеется огромное разнообразно древесных и особенно травянистых растений, которые, в свою очередь, подразделяются па отдельные типы форм роста, пли жизненных форм. Существуют также промежуточные формы между дроиосиыми и травяпистыми растениями, которые не всегда легко отнести к тому или иному типу. В таких случаях ответ часто может дать только специальное анатомическое исследование проводящей системы стебля, ее ксилемы. Дело в том, что для древесных растений, будь то огромные деревья или небольшие кустарнички, характерно Наличие активно действующего камбия, производящего в стеблях и корнях ясно выраженные слои вторичной ксилемы. У травянистых растений, как привило, активный камбий отсутствует или же имеется в зачаточной форме.[ …]
Микробиологи несут ответственность за царство Protista, которое в массе превосходит массу животных и растений. Там, где зоологи и ботаники обязаны давать сигнал тревоги, микробиолог в состоянии помочь в осуществлении исправляющих и предупредительных мер, направленных на охрану биосферы.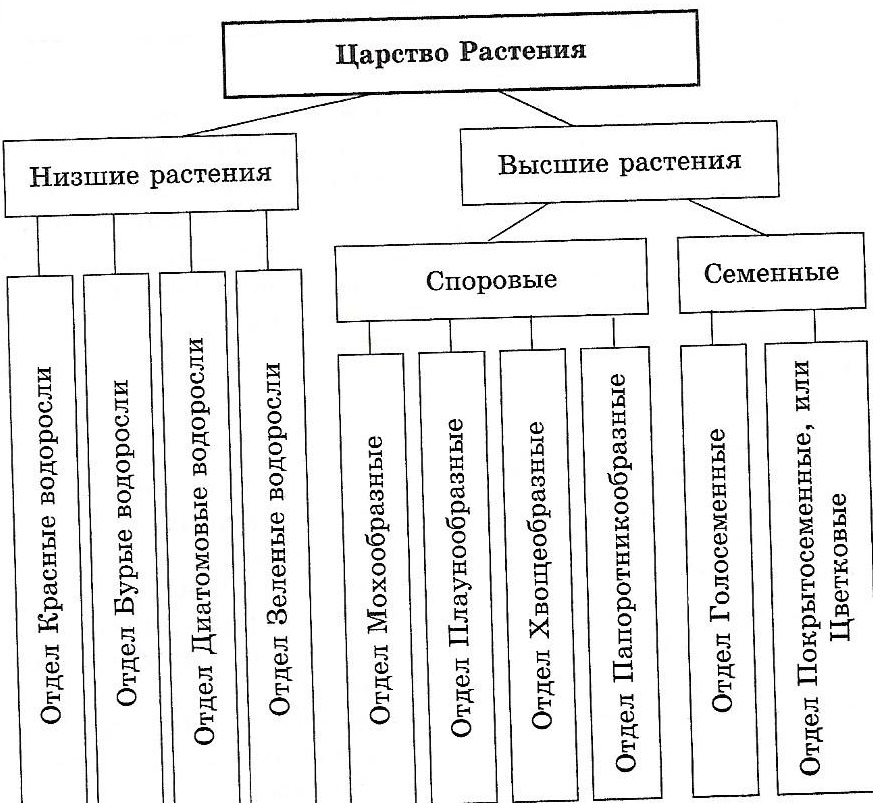 [ …]
[ …]
Онтогенез, или индивидуальное развитие, растений начинается с момента оплодотворения яйцеклетки или возникновения зачаточной ночки в репродуктивных органах и тканях материнского растения и закапчивается отмиранием растения. Таким образом, онтогенез представляет собой полный цикл жизнедеятельности растения, включает в себя все его жизненные процессы и проявления и продолжается в зависимости от вида растения от 5—6 под у миниатюрных эфемеров до 3—5 тыс. лет у гигантов растительного царства — мамонтовых деревьев, кедров и других видов.[ …]
Общее распределение вирусов в растительном царстве обсуждается в гл. Здесь мы рассмотрим факторы, связывающие жизнеспособность вируса и его распространение в нолевых условиях со спектром растений-хозяев.[ …]
В настоящее время начато изучение целых групп растений. Систематическое внимание уделяется в особенности вторичным метаболитам низкого молекулярного веса, которые являются второстепенными продуктами обмена веществ. Такие соединения, как алкалоиды, флавоноиды, небелковые аминокислоты, терпены, гликозиды, распространены в растительном царстве неравномерно. Во многих случаях их физиологическая роль до настоящего времени неясна, однако они оказывают вполне определенное специфическое действие на высшие организмы, и именно поэтому химики, фармакологи и врачи уделяют этим соединениям огромное внимание. В настоящее время химическое исследование растений стало неотъемлемой частью таксономического метода, позволяющего рассматривать целые роды и семейства в глобальном масштабе, а не только с региональной позиции. Для таксономии присутствие или отсутствие данного вещества в растении всегда имеет положительное значение. Химический анализ делает возможной объективную статистическую оценку и надежную интерпретацию результатов.[ …]
Во многих случаях их физиологическая роль до настоящего времени неясна, однако они оказывают вполне определенное специфическое действие на высшие организмы, и именно поэтому химики, фармакологи и врачи уделяют этим соединениям огромное внимание. В настоящее время химическое исследование растений стало неотъемлемой частью таксономического метода, позволяющего рассматривать целые роды и семейства в глобальном масштабе, а не только с региональной позиции. Для таксономии присутствие или отсутствие данного вещества в растении всегда имеет положительное значение. Химический анализ делает возможной объективную статистическую оценку и надежную интерпретацию результатов.[ …]
Однако такое деление живых организмов на четыре царства еще не легло в основу справочной и учебной литературы, поэтому при дальнейшем изложении материала мы придерживаемся традиционных классификаций, по которым бактерии, синезеленые водоросли и грибы являются отделами низших растений.[ …]
Фенольные соединения обнаруживаются уже в низших растениях -водорослях, грибах, лишайниках, мхах, плаунах, папоротниках. В высших голосеменных и покрытосеменных растениях эти вещества находятся уже в форме полифенолов, например дубильных веществ и лигнинов. У древесных растений они встречаются во всех частях организма — в коре, древесине, листьях, корнях, плодах и патологических образованиях. Распространенность фенолов и полифенолов в растительном царстве, их закономерное присутствие в определенных растительных тканях означает, что в растениях они играют определенную физиологическую роль.[ …]
В высших голосеменных и покрытосеменных растениях эти вещества находятся уже в форме полифенолов, например дубильных веществ и лигнинов. У древесных растений они встречаются во всех частях организма — в коре, древесине, листьях, корнях, плодах и патологических образованиях. Распространенность фенолов и полифенолов в растительном царстве, их закономерное присутствие в определенных растительных тканях означает, что в растениях они играют определенную физиологическую роль.[ …]
В главе IV было показано, как азот циркулирует между царствами минералов, растений и животных, переходя из одной формы в другую, и как важны эти превращения для сельского хозяйства.[ …]
Известно, что издавна органический мир делился на два царства — животных и растений. Однако в наше время его уже следует делить на две империи — доклеточных (вирусы и фаги) и клеточных (все остальные организмы). Империя доклеточных состоит из единственного царства — вирусов (фаги тоже вирусы-паразиты). Империя клеточных включает уже два над-царства, четыре царства, и еще семь подцарств (табл. 1.1).[ …]
1.1).[ …]
Все живые организмы биосферы подразделяются на четыре царства: прокариоты (доядерные), животные, грибы и растения.[ …]
В кембрий-ордовик суша постепенно осваивается примитивными растениями и животными; з морях царствуют трилобиты, грап-толиты, наутилоидеи, мшанки. В силуре появляются рыбы, в девоне— насекомые и земноводные. Сравнительно богатый наземный биостром девона говорит о возникновении к тому времени озонового экрана — этого удивительного «новообразования» географической оболочки. Располагаясь в стратосфере, на высоте 20— 25 км, озоновый экран поглощает коротковолновую часть ультрафиолетовой солнечной радиации, губительной для органической жизни. Он создан жизнью для того, чтобы защитить жизнь, дать ей новые, почти неограниченные возможности для развития на нашей Земле. И уже в следующем геологическом периоде — каменноугольном— суша одевается влажными густыми лесами высокой биологической продуктивности из древовидных папоротников, гигантских плаунов и хвощей. Это прямое доказательство высокого, близкого к современному содержания кислорода в атмосфере.[ …]
Это прямое доказательство высокого, близкого к современному содержания кислорода в атмосфере.[ …]
Отдел Покрытосеменные, или Цветковые (Angiospermae, или МаЁПо1юр11у1а). Растения этого отдела встречаются почти повсеместно. На их долю приходится 250 000—300 000 видов, т. е. почти две трети видов царства растений. В настоящее время они являются самой процветающей группой растений.[ …]
В нынешних представлениях к консументам относят огромное количество живых организмов из самых разных царств, типов, классов биологической системы. Например, среди растений есть формы, паразитирующие на других, как правило, это формы, не содержащие хлорофилла. Среди высших растений к консументам можно отнести насекомоядные и другие растения, способные к смешанному питанию. Царство животных представлено исключительно консументами. Позиции, которые занимают консументы в биологическом круговороте, весьма важны, хотя далеко неоднозначны. Биологический круговорот может обойтись и без консументов. Так, в некоторых условиях замкнутые модельные системы, созданные искусственно в лабораторных условиях из зеленых растений и микроорганизмов, при наличии минеральных веществ и воды могут существовать достаточно длительный период. -Это осуществимо из-за фотосинтеза и деструкции растительных остатков. Но, как пишет И.А. Шилов (2000), в природных условиях гибель таких систем становится весьма вероятной, и гарантами устойчивости биологического круговорота служат именно консументы.[ …]
-Это осуществимо из-за фотосинтеза и деструкции растительных остатков. Но, как пишет И.А. Шилов (2000), в природных условиях гибель таких систем становится весьма вероятной, и гарантами устойчивости биологического круговорота служат именно консументы.[ …]
Три основных типа питания, и соответственно способа использования- продукции, представляют три функциональных царства природных сообществ: продуценты, или зеленые растения, которые сами создают свою пищу и в результате дыхания расходуют часть ее для своих нужд; консументы, или животные, которые питаются за счет поглощения и внутреннего переваривания органического вещества, и редуценты — бактерии и грибы, живущие за счет поглощения пищи, осуществляя ее внешнее переваривание и разлагая органическое вещество до неорганических продуктов. Валовая первичная продукция за вычетом расходов на дыхание — это есть чистая первичная продукция, и ее могут потреблять консументы и редуценты. Ступени на пищевых цепях, по которым распределяются организмы в зависимости от их позиции в цепи, — это трофические уровни. [ …]
[ …]
Р. X. Уиттекер — биолог широкого профиля. Область его научных интересов чрезвычайно многогранна: проблемы классификации и ординации растительных сообществ, вопросы динамики и продуктивности растительности, циклы элементов минерального питания, экологические ниши, видовой состав фитоценозов и другие вопросы теории и практики экологии. Ему, в частности, принадлежит обоснование разделения организмов на пять царств: прокариоты, протисты, грибы, растения, животные. Эта система приводится в большинстве американских учебников по биологии.[ …]
Особенности царства растений
Зарождение и развитие растительного мира тесно связано с историей Земли. Ученые считают, что первоначально колыбелью жизни была вода. Она же была одновременно и средой ее дальнейшего развития.
Чтобы жить в воде, растениям не нужны специальные приспособления, так как сама вода поддерживает и питает организмы, живущие в ней. Сначала растения могли жить только в воде на определенной глубине от поверхности. На поверхность воды они смогли выйти лишь тогда, когда вокруг Земли образовался озоновый слой, который уменьшил губительное влияние на живые организмы ультрафиолетового излучения. Это и позволило растениям выйти на сушу.
На поверхность воды они смогли выйти лишь тогда, когда вокруг Земли образовался озоновый слой, который уменьшил губительное влияние на живые организмы ультрафиолетового излучения. Это и позволило растениям выйти на сушу.
Покинув гостеприимную водную среду, растения должны были приспособиться к жизни на суше. Для этого им пришлось образовать ряд новых структур, которые могли бы взять на себя выполнение многочисленных функций, осуществляемых прежде водой. Завоевание суши было, по-видимому, длительным и трудным процессом. Новые формы, пытавшиеся жить в почве, получали достаточное количество минеральных солей, воды и двуокиси углерода, но были лишены света, необходимою для фотосинтеза. Те же, которые пытались жить на поверхности, испытывали недостаток воды и минеральных солей. В результате длительного, жертвенного (многие растения погибали) процесса истинно наземными формами стали растения, которые смогли выжить в новых условиях благодаря развитию ряда специализированных органов: листьев, находящихся на воздухе, поглощающих свет и осуществляющих фотосинтез; корней, служащих для закрепления растений и поглощения воды и минеральных солей из почвы. Вследствие этою представители царства растений не способны к активному движению, они имеют неограниченный рост; их клеточная оболочка состоит из целлюлозы, что придает им прочность.
Вследствие этою представители царства растений не способны к активному движению, они имеют неограниченный рост; их клеточная оболочка состоит из целлюлозы, что придает им прочность.
Одной из основных особенностей царства растений является способ питания. Все они автотрофы, так как способны самостоятельно, используя энергию Солнца, создавать из неорганических веществ органические. Их способ питания называют еще фототрофным (питание на свету). Запасные питательные вещества откладываются в виде крахмала. Очень важным моментом в жизни представителей царства растений является то, что в их жизненном цикле развития происходит чередование половой и бесполой фаз (поколений): гаметофита и спорофита.
На спорофите (от греч. «спора» и «фитон» — растения) образуются органы бесполого размножения, а на гаметофите (от греч. «гамета» — половая клетка и «фитоп») — полового.
Царство растений условно подразделяют на низшие и высшие растения.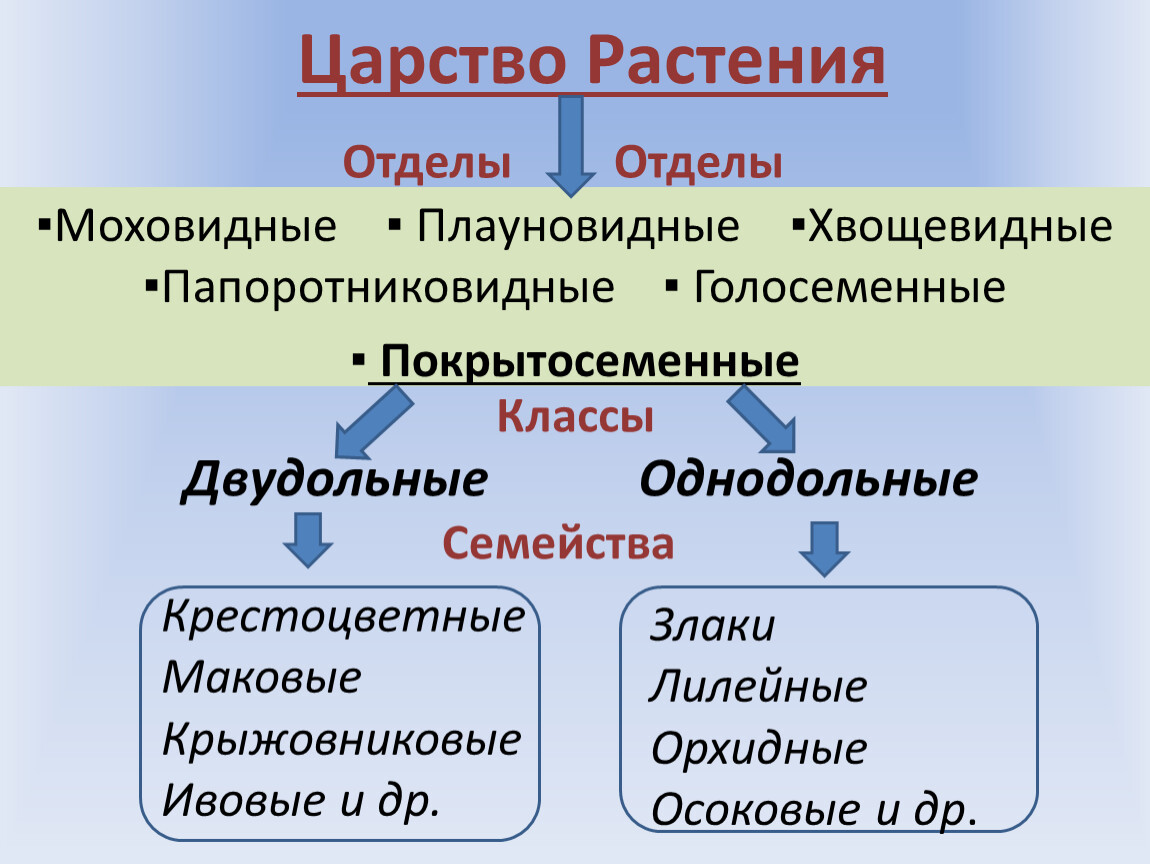 У низших растений органы полового и бесполого размножения обычно одноклеточные, у высших растений — многоклеточные. В царстве растений условно выделяют две большие группы: низшие и высшие растения.
У низших растений органы полового и бесполого размножения обычно одноклеточные, у высших растений — многоклеточные. В царстве растений условно выделяют две большие группы: низшие и высшие растения.
- Низшие растения
- Водоросли
- зеленые
- диатомовые
- бурые
- красные, или багрянки
- Водоросли
- Вычшие растения
- Споровые
- риниофиты
- мохообразные
- папоротникообразные
- псилофиты
- плаунообразные
- хвощеобразные
- Семенные
- голосеменные (пинофиты)
- покрытосеменные (цветковые)
- однодольные
- двудольные
- Споровые
|
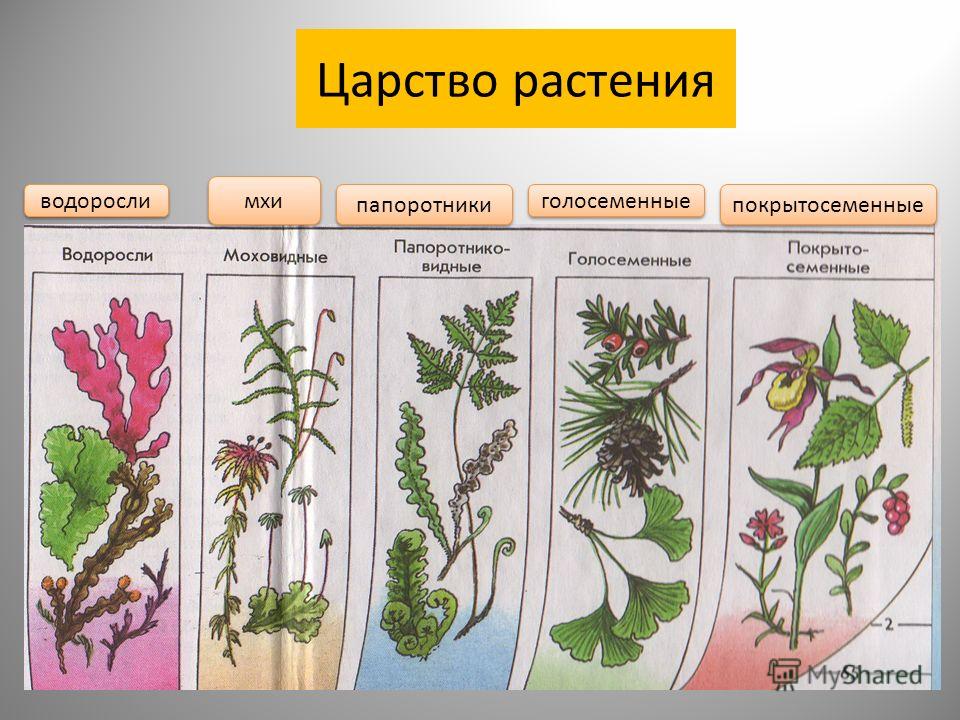
Царство растений объединяет живые организмы, обладающие характерными признаками. Все они являются автотрофами, то есть могут осуществлять фотосинтез. Схема. Классификация растений Чем выше уровень организации растений, тем более явно заметны различия между ними и другими живыми организмами. Классификация и систематические категории царства растений подверглись изменениям с середины 20-го века. До этого времени все растения были разделены на низшие и высшие. К низшим относились бактерии, грибы, водоросли, лишайники и слизевики, а к высшим – моховидные, риниевые, плауновидные, псилотовые, хвощевидные, голосеменные, папоротниковидные и покрытосеменные. На сегодняшний день в систематике растений существуют царство бактерий и царство грибов отдельно друг от друга. Отделы царства растенийВсе растения распределены по основным отделам царства растений. Это мхи, плауны, папоротники, голосеменные, хвощи и покрытосеменные (цветковые) растения. Представители отдела покрытосеменных (цветковых) растений, в свою очередь, разделены на два класса – двудольные и однодольные. Разные типы размножения растений обусловливают их деление на семенные растения и размножающиеся спорами. Значение растений на Земле огромно. Именно представители царства растений – первичные производители органических веществ. Доказано, что весь имеющийся в атмосфере кислород появился, благодаря жизнедеятельности растений, а точнее — фотосинтезу. Растительные сообщества являются естественной средой обитания животных и человека, при этом обеспечивая их едой, в том числе косвенно, участвуя в образовании почвы. Растения служат сырьем для получения разных технологических материалов, топлива, строительных материалов, лекарственных средств. Некоторые виды растений стали культурными, и из них получают ценные пищевые продукты. |
 Также все представители царства растений имеют плотные клеточные оболочки, основой которых является такое органическое вещество, как целлюлоза. Крахмал у растений – запасное вещество. Несмотря на то, что растениям-сапрофитам и паразитам свойственно гетеротрофное питание, они относятся к царству растений, так как такой тип питания вторичный. Имеются и другие характерные особенности у представителей царства растений. Это определенные циклы жизнедеятельности, пути закладки органов, неподвижный образ жизни, т.д. Хотя вышеуказанные признаки не являются общими для всех групп растений, но их сочетание позволяет выделить растения, в особенности высокоорганизованные, из всех живых организмов других царств. На низшем уровне развития растения легко можно перепутать с простейшими животными. Главная отличительная особенность растений на этом уровне – наличие хлоропластов и особенности структуры клеток.
Также все представители царства растений имеют плотные клеточные оболочки, основой которых является такое органическое вещество, как целлюлоза. Крахмал у растений – запасное вещество. Несмотря на то, что растениям-сапрофитам и паразитам свойственно гетеротрофное питание, они относятся к царству растений, так как такой тип питания вторичный. Имеются и другие характерные особенности у представителей царства растений. Это определенные циклы жизнедеятельности, пути закладки органов, неподвижный образ жизни, т.д. Хотя вышеуказанные признаки не являются общими для всех групп растений, но их сочетание позволяет выделить растения, в особенности высокоорганизованные, из всех живых организмов других царств. На низшем уровне развития растения легко можно перепутать с простейшими животными. Главная отличительная особенность растений на этом уровне – наличие хлоропластов и особенности структуры клеток.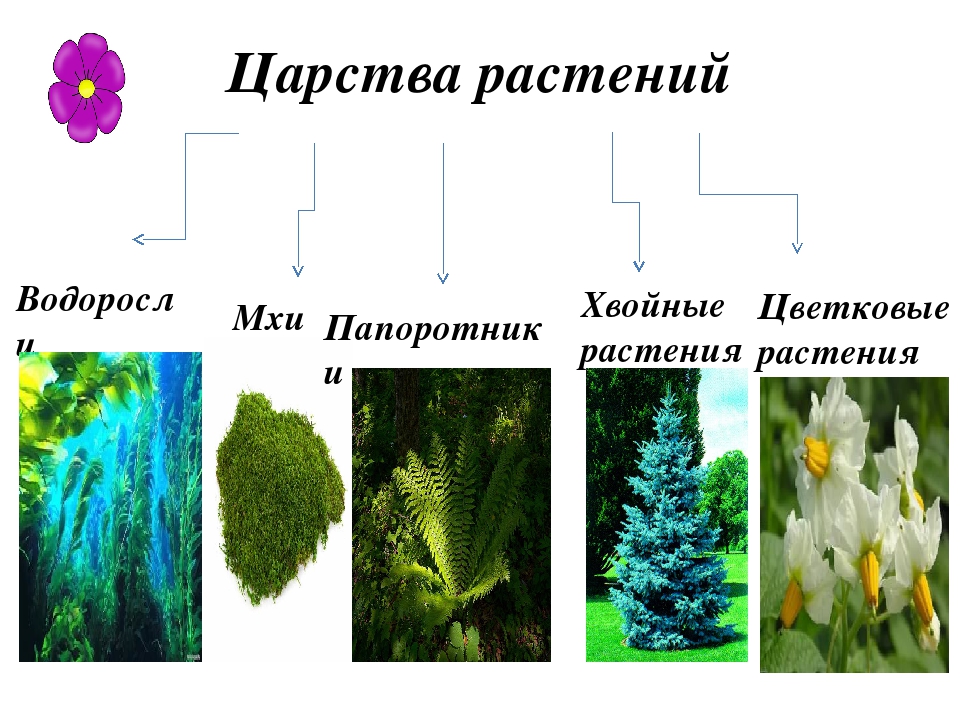 Основное большинство высокоорганизованных растений имеют слишком расчлененное тело, что обусловливает увеличение его поверхности для лучшего усвоения из окружающего пространства газов и жидкости с целью дальнейшего преобразования их в питательные вещества в ходе фотосинтеза. Наличие большого количества специализированных частей тела у высших растений стало возможным, именно благодаря расчленению и дифференцировке тела. Большинство значимых признаков строения растений обусловлены особенностями их размножения, развития и типом расселения.
Основное большинство высокоорганизованных растений имеют слишком расчлененное тело, что обусловливает увеличение его поверхности для лучшего усвоения из окружающего пространства газов и жидкости с целью дальнейшего преобразования их в питательные вещества в ходе фотосинтеза. Наличие большого количества специализированных частей тела у высших растений стало возможным, именно благодаря расчленению и дифференцировке тела. Большинство значимых признаков строения растений обусловлены особенностями их размножения, развития и типом расселения. Поэтому группа «низшие растения» канула в лету. В современной систематике царство растений делится на три подцарства: настоящие водоросли, багрянки (красные водоросли) и высшие растения (зародышевые). В эти три подцарства включены все произрастающие на Земле 350 тысяч видов растений. Они различаются размерами – от очень маленьких до огромных растений. Все представители царства растений отличаются друг от друга жизненными формами (травы, деревья, кустарники), длительностью периода жизнедеятельности (многолетние, однолетние, двулетние), требованиями к условиям окружающей среды, типами размножения.
Поэтому группа «низшие растения» канула в лету. В современной систематике царство растений делится на три подцарства: настоящие водоросли, багрянки (красные водоросли) и высшие растения (зародышевые). В эти три подцарства включены все произрастающие на Земле 350 тысяч видов растений. Они различаются размерами – от очень маленьких до огромных растений. Все представители царства растений отличаются друг от друга жизненными формами (травы, деревья, кустарники), длительностью периода жизнедеятельности (многолетние, однолетние, двулетние), требованиями к условиям окружающей среды, типами размножения. С учетом требований растений к условиям произрастания выделяют теплолюбивые растения и холодостойкие, теневыносливые и светолюбивые, засухоустойчивые и влаголюбивые. Те растения, средой обитания которых является вода, называются водными.
С учетом требований растений к условиям произрастания выделяют теплолюбивые растения и холодостойкие, теневыносливые и светолюбивые, засухоустойчивые и влаголюбивые. Те растения, средой обитания которых является вода, называются водными.примеры, основные систематические категории и специфические особенности их строения
Сравните систематические категории Царства растений, приведите примеры этих организмов, укажите особенности их строения. .. Если вы затрудняетесь с выполнением подобных заданий, наша статья поможет разобраться в данном вопросе.
.. Если вы затрудняетесь с выполнением подобных заданий, наша статья поможет разобраться в данном вопросе.
Признаки Царства растений
Все организмы, населяющие нашу планету, ученые объединили в Царства живой природы. Растения, примеры которых мы рассмотрим в нашей статье, имеют свои отличительные особенности. Прежде всего это автотрофный способ питания. Только растения способны производить органические вещества и кислород из воды и углекислого газа при наличии солнечного света. Местом, где происходит этот сложный фотохимический процесс, называемый фотосинтезом, являются зеленые органеллы растительных клеток — хлоропласты.
Характерными признаками представителей данного Царства является прикрепленный образ жизни, неограниченный рост, который продолжается в течение всей жизни, а также наличие углевода крахмала в качестве запасного питательного вещества.
Систематика растений
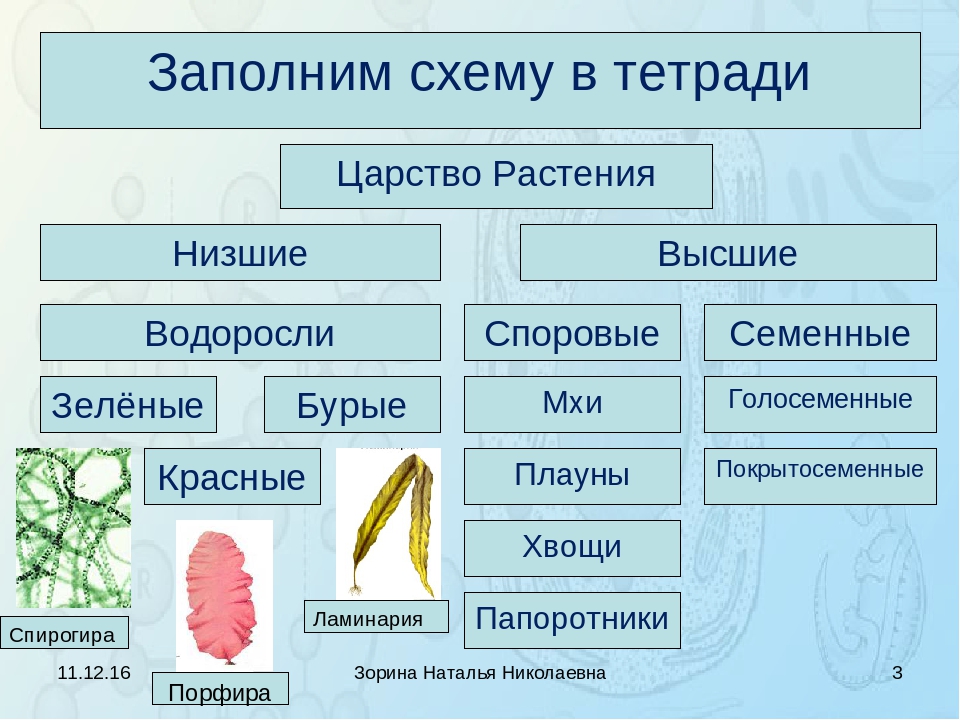
Систематические категории Царства растений, примеры которых будут рассмотрены нами далее, созданы на основе общих признаков строения.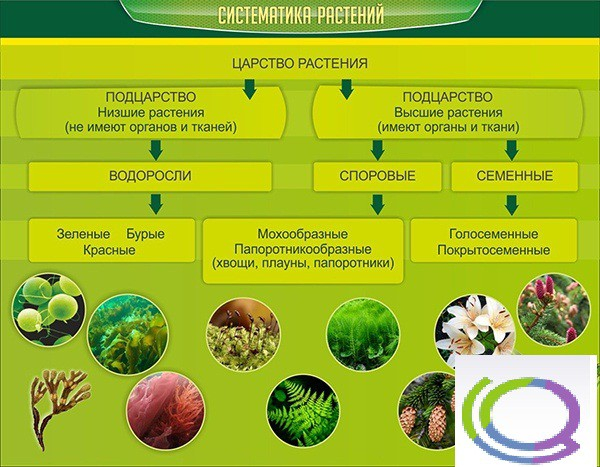 Первоначально их объединяют в две большие группы: низшие и высшие. Тело первых состоит из многочисленных неспециализированных клеток, которые не образуют ткани, и называется слоевище. А высшие имеют сложное листостебельное строение.
Первоначально их объединяют в две большие группы: низшие и высшие. Тело первых состоит из многочисленных неспециализированных клеток, которые не образуют ткани, и называется слоевище. А высшие имеют сложное листостебельное строение.
Низшие растения
Это самые простые организмы, которые представляют Царство растений. Примеры их известны всем. Это ламинария, или морская капуста, соргассум, хламидомонада, улотрикс, вольвокс, спирогира. Среди них есть одно-, многоклеточные и колониальные организмы. Все они обитают в воде. Кроме слоевища, которое представляет недифференцированное тело водорослей, у них есть ризоиды. Эти структуры частично выполняют функцию корней, но не образуют тканей, чем и отличаются от них. Поскольку вода сама поддерживает форму слоевища, у водорослей отсутствуют механические ткани.
Высшие растения
Растения, образующие настоящие ткани и органы, называются высшими. Первыми их представителями, которые вышли из воды на сушу, являются представители споровых. Это мхи, плауны, хвощи и папоротники. Вместо ризоидов у некоторых из них появляется мочковатая корневая система, которая растет пучком на подземном видоизмененном побеге. Они размножаются при помощи клеток бесполого размножения — спор. Еще одной характерной чертой является чередование поколений в их жизненном цикле. Более сложно устроены семенные растения, это дало им возможность занять господствующее положение на планете.
Это мхи, плауны, хвощи и папоротники. Вместо ризоидов у некоторых из них появляется мочковатая корневая система, которая растет пучком на подземном видоизмененном побеге. Они размножаются при помощи клеток бесполого размножения — спор. Еще одной характерной чертой является чередование поколений в их жизненном цикле. Более сложно устроены семенные растения, это дало им возможность занять господствующее положение на планете.
Царство растений: примеры и классификация
Размножение при помощи семян дает растениям больше шансов выжить в неблагоприятных условиях среды обитания. Рассмотрим их особенности на примере хвойных деревьев. Они являются представителями отдела Голосеменные. Их характерными особенностями являются наличие тонких листьев хвоинок и смоляных ходов в стволе. А отличительной — расположение семян на чешуйках шишек голо или открыто. Отсюда и происходит их название. Конечно, оказавшись в условиях пониженных температур или недостатка влаги, не все они прорастут.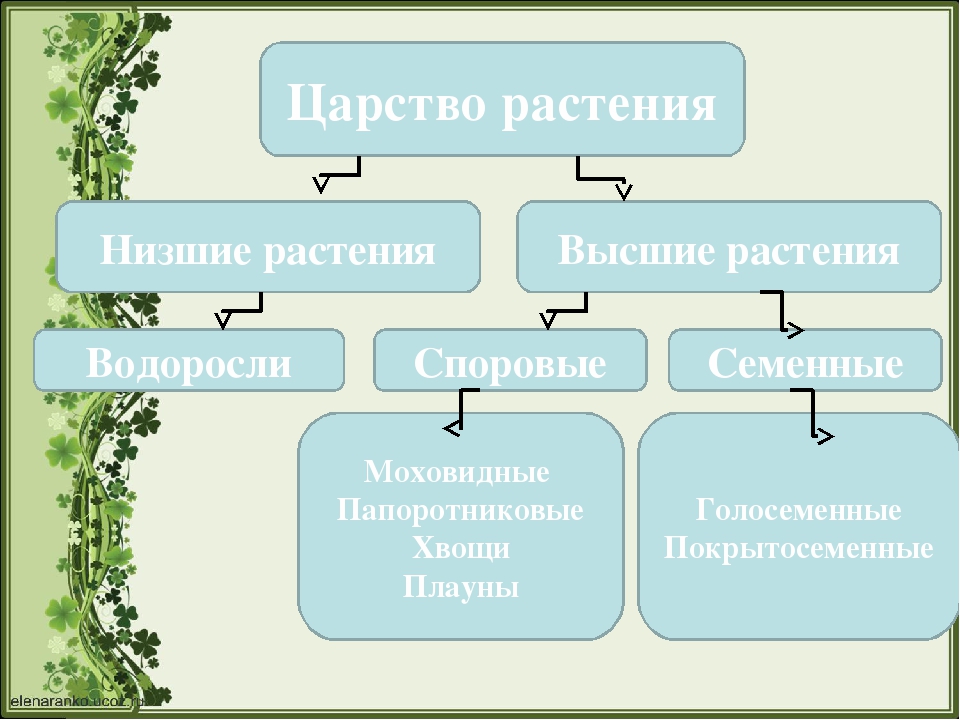 Но, в отличие от спор, семена представляют собой многоклеточные структуры и имеют запасные питательные вещества — эндосперм.
Но, в отличие от спор, семена представляют собой многоклеточные структуры и имеют запасные питательные вещества — эндосперм.
В более выгодном положении оказываются Покрытосеменные растения. Они образуют цветок и плод, в котором происходит развитие и созревание семян. Эта структура служит дополнительной защитой, источником тепла и питания. Отдел Покрытосеменные объединяет два класса: Одно- и Двудольные. Первые имеют одну семядолю в зародыше, мочковатую корневую систему, простые линейные листья. К ним относятся представители семейств Злаковые, Луковые и Лилейные. Все они являются травянистыми растениями из-за отсутствия камбия — боковой образовательной ткани. Более разнообразны и многочисленны Двудольные. Кроме количества семядолей их отличает корневая система стержневого типа, как сложные, так и простые листья, наличие камбия. Среди них встречаются и травы, и кустарники, и деревья. Они наиболее широко представляют Царство Растений. Примеры наиболее распространенных — это Розоцветные, Капустные, Пасленовые, Астровые, Бобовые.
Значение представителей растительного царства
Значение всех представителей растений сложно переоценить, ведь они обеспечивают все живое необходимым условием жизни — кислородом. Водоросли являются исходным материалом для производства биологически активных веществ, лекарств, пищевых добавок и красок. Голосеменные славятся наличием фитонцидов и ценной древесины. Человек употребляет в пищу плоды и части цветковых растений.
Таким образом, Царство растений, примеры и систематические особенности которого были рассмотрены в нашей статье, отличается точной иерархией развития, разнообразием и играет важнейшую роль в жизни планеты.
ЦАРСТВО РАСТЕНИЯ РАЗНООБРАЗИЕ, РАСПРОСТРАНЕНИЕ И ЗНАЧЕНИЕ РАСТЕНИЙ. НИЗШИЕ И ВЫСШИЕ РАСТЕНИЯ. ГОЛОСЕМЕННЫЕ
ЦАРСТВО РАСТЕНИЯ
РАЗНООБРАЗИЕ, РАСПРОСТРАНЕНИЕ И ЗНАЧЕНИЕ РАСТЕНИЙ. НИЗШИЕ И ВЫСШИЕ РАСТЕНИЯ. ГОЛОСЕМЕННЫЕ
1. К низшим растениям относят:
A. Мхи
Б. Водоросли
B. Мхи и водоросли
Г. Папоротникообразные
2. Для водорослей характерны следующие признаки:
Для водорослей характерны следующие признаки:
A. Имеют листья и стебли
Б. Обитают в водоемах и цветут
B. Размножаются семенами
Г. Имеют слоевище и ризоиды
3. Заполните таблицу «Признаки высших и низших растений».
4. Выберите один наиболее точный и полный ответ.
Водоросли – это:
A. Растения, обитающие в воде
Б. Одноклеточные растения, обитающие в воде
B. Самые древние растения на Земле
Г. Самые древние растения на Земле, тело которых одноклеточное или многоклеточные – слоевище
5. Выберите правильные ответы.
Хроматофор – это:
A. Название многоклеточной водоросли
Б. Название пигмента
B. Один крупный хлоропласт
Г. Чашеобразная пластида в клетке водоросли
Д. Органоиды – водоросли, содержащие пигмент
6. Выберите один правильный ответ.
Ризоиды – это:
A. Название растений
Б. Вид корня
B. Органоид клетки
Г. Ветвистые клетки, при помощи которых водоросли прикрепляются к субстрату
7. Хламидомонада получила такое название из-за того, что она:
Хламидомонада получила такое название из-за того, что она:
A. Небольших размеров
Б. Имеет грушевидную форму тела
B. Имеет орган передвижения – жгутик
Г. Простейший организм, покрытый оболочкой
8. Красные водоросли обитают:
A. На мелководьях морей
Б. На больших глубинах морей
B. Во всех водоемах
Г. Во всей тоще океанов
9. Морской капустой называют:
А. Ламинарию
Б. Порфиру
В. Ульву
Г. Филлофору
10. Подберите соответствующие названия для различных форм лишайников:
I. Кустистые
II. Листовые
III. Накипные
A. Ксантория настенная
Б. Бацидия
B. Олений мох
11. Вставьте пропущенные слова.
Слоевище лишайников состоит из… и…, живущих в…
12. Сходство с водорослями имеют мхи:
A. Риччия
Б. Маршанция
B. Сфагнум
Г. Кукушкин лен
13. У кукушкиного льна женские растения, в отличие от мужских имеют:
A. Сперматозоиды
Б.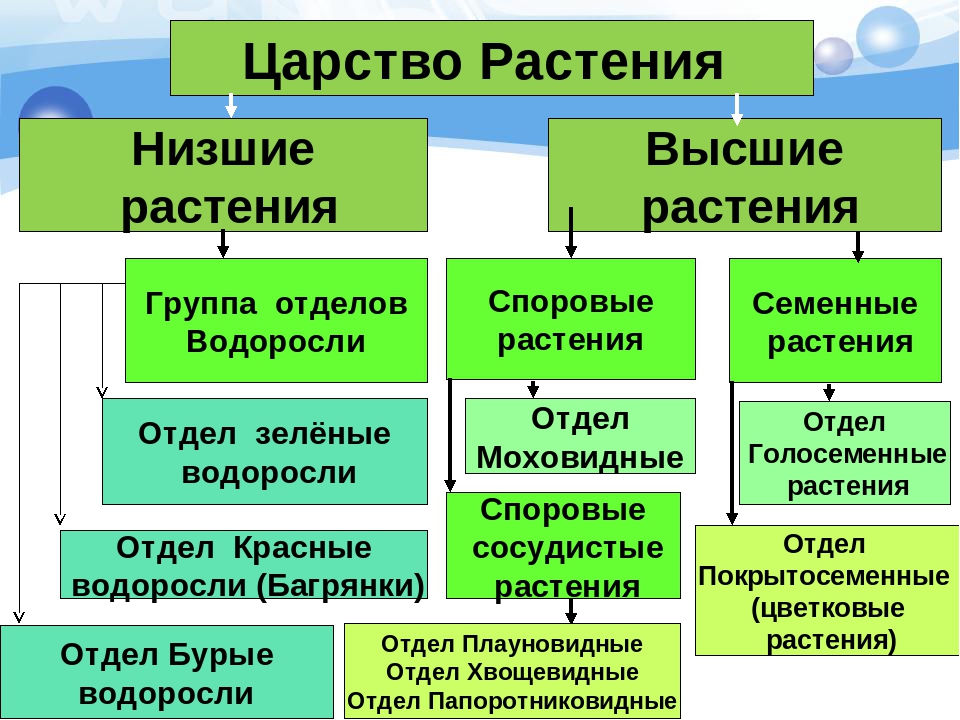 Яйцеклетки
Яйцеклетки
B. Коробочки на длинных ножках
Г. Листья
14. Сфагнум, в отличие от кукушкиного льна:
A. Быстро всасывает и проводит воду
Б. Не имеет ризоидов
B. Размножается спорами
Г. Не имеет стебля и листьев
15. Там, где растет сфагнум:
A. Деревья развиваются хорошо
Б. Деревья становятся угнетенными
B. Деревья расти не могут
Г. Лес заболачивается
16. Выберите правильное определение.
Торф – это:
A. Толща полуразложившихся растительных остатков
Б. Название мхов
B. Толща разложившихся папоротников
Г. Тип болота
17. Выберите наиболее полный ответ.
Плауны, хвощи и папоротники относят к высшим споровым растениям:
A. Они широко расселились по земле
Б. Размножаются спорами
B. Имеют корни, стебель, листья и размножаются спорами
Г. Размножаются семенами
18. Вайями называют:
A. Сильно рассеченные листья папоротника
Б. Вид папоротника
B. Корень папоротника
Корень папоротника
Г. Подземные побеги
19. Становится редким и нуждается в охране:
А. Папоротник
Б. Хвощ
В. Плаун
Г. Кукушкин лен
20. Показателем повышенной кислотности почв на пашне служит распространение на этой территории:
А. Папоротника
Б. Хвоща
В. Плауна
Г. Крапивы
21. Вставьте пропущенные слова.
Папоротник имеет подземные побеги – …, от них растут…, на нижней стороне которых находятся маленькие бугорки – …
22. Папоротникообразные относятся к высшим споровым растениям, так как они:
A. Широко расселились по земле
Б. Имеют корень
B. Имеют корень, стебель, листья и размножаются спорами
Г. Размножаются спорами
23. К голосеменным растениям относят:
A. Кукушкин лен и сосну
Б. Ель и хвощ
B. Пихту и лиственницу
Г. Можжевельник и плаун
24. Выберите более точный ответ.
Залежи каменного угля образовались:
A. Из отмерших древовидных папоротников
Б. Из отмерших частей мхов
Из отмерших частей мхов
B. Из большого скопления остатков растительности
Г. Из большого скопления отмерших водорослей
25. Хвойные растения хорошо приспособлены к неблагоприятным условиям:
A. Хвоя имеет плотную кожицу, покрытую восковым веществом, поэтому растения испаряют мало воды
Б. Имеют стебель, корень, хвою
B. Имеют шишки
Г. Образуют семена, с помощью которых размножаются
26. К названиям растений, обозначенным цифрами, допишите характерные для них признаки, обозначенные буквами.
I. Сосна
II. Можжевельник
III. Лиственница
IV. Кедр
V. Ель
A. На укороченном побеге имеются две хвоинки
Б. На укороченном побеге имеются 3–5 хвоинок
B. Имеются шишкоягоды
Г. На зиму сбрасывает хвою
Д. Хвоинки располагаются на побеге по одной, супротивно друг другу
27. Название голосеменные получили потому, что:
A. Семена лежат открыто на поверхности чешуи шишек
Б. Размножаются семенами
B. Имеют шишки
Имеют шишки
Г. Имеют хвою
28. Женские шишки в отличие от мужских:
A. Образуют семена
Б. Растут у оснований молодых побегов
B. Участвуют в размножении
Г. Зеленовато-желтые, собраны тесными группами
29. Рассеивание семян сосны и ели происходит:
А. Весной
Б. Летом
В. Зимой
Г. Осенью
30. Отличительным признаком покрытосеменных является:
A. Наличие цветка
Б. Размножение семенами
B. Наличие корня, стебля, листьев, плодов
Г. Занимают различные среды жизни
31. К деревьям относят:
A. Малину и рябину
Б. Ель и березу
B. Бамбук и орешник
Г. Липу и дуб
32. К однолетним растениям относят:
А. Крапиву
Б. Картофель
В. Капусту
Г. Одуванчик
33. Найдите соответствие.
Определите растения по продолжительности жизни.
I. Однолетние
II. Двулетние
III. Многолетние
A. Береза
Б. Рябина
B. Капуста
Г. Горох
Д. Морковь
Морковь
Е. Бегония
34. Выберите верное утверждение.
1. Водоросли обитают в воде и на суше в местах с повышенной влажностью.
2. Водоросли относятся к низшим растениям, т. к. они не имеют корней, стеблей и листьев.
3. Водоросли размножаются спорами.
4. Плеврококк – это одноклеточная водоросль.
5. Многоклеточные водоросли имеют тело – слоевище.
6. Водорослями питаются многие водные животные.
7. Ламинария – это морской салат.
8. Тело лишайника – слоевище состоит из водоросли и гриба.
9. Лишайники первыми заселяются на бесплодных землях.
10. Различают печеночные и листоватые мхи.
11. Мхи имеют стебель, листья и ризоиды, с помощью которых они прикрепляются к грунту и всасывают воду, например сфагнум.
12. Папоротников насчитывается более 100 000 видов.
13. Вайи – сильно расчлененные листья папоротников, растут прямо от корневищ.
14. Голосеменные – это высшие растения, они имеют стебель, корень, листья и образуют семена.
15. У голосеменных семена лежат открыто на поверхности чешуи шишек.
16. К голосеменным относятся ель, сосна, пихта, кипарис, щитовник, можжевельник, туя.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес4.4.1. Общая характеристика царства Растения
Ботаника изучает строение, жизнедеятельность, закономерности индивидуального и эволюционного развития, распространение и условия обитания растений.
Значение растений в биосфере огромно. Фотосинтезирующие растения являются источником органического вещества и кислорода, оказывают значительное влияние на климат. Жизнь современного человека зависит от использования культурных растений, которых в настоящее время насчитывается около 1500 видов. Человек широко использует природные и культивируемые растения для изготовления лекарственных препаратов или в декоративных целях.
Размеры и строение растений — от 2—3 мкм у одноклеточных водорослей (хлорелла) до сложно устроенных покрытосеменных с высокодифференцированными тканями и органами (эвкалипты до 150 м).
Общие черты растений:
Наличие в составе клеток твердой клеточной оболочки из целлюлозы.
Способ питания — адсорбционный (всасывательный).
Прикрепление к твердому субстрату и ограничение подвижности.
Расселение растений зачатками (спорами, семенами), находящимися в состоянии покоя.
По типу питания растения — автотрофы, запасное питательное вещество — крахмал.
Закономерное чередование двух поколений в жизненном цикле, проходящем от зиготы одного растения до зиготы другого: бесполого диплоидного поколения — спорофита, и полового гаплоидного — гаметофита.
При чередовании поколений гаметофит — растительный организм, на котором формируются гаметы, закономерно сменяется спорофитом: гаплоидные гаметы сливаются диплоидная зигота зародыш
взрослое растение — спорофит (имеет спорангий и производит споры, одинаковые или разные. При образовании спор происходит мейоз, поэтому споры гаплоидные) из спор вырастает гаметофит (гаплоидное поколение, есть половые органы, производящие сперматозоиды и яйцеклетки). Все наземные растения имеют половые органы одного типа: мужские называются антеридии, а женские — архегонии. В мужских органах образуется много сперматозоидов, а в женских — одна крупная неподвижная яйцеклетка.
При образовании спор происходит мейоз, поэтому споры гаплоидные) из спор вырастает гаметофит (гаплоидное поколение, есть половые органы, производящие сперматозоиды и яйцеклетки). Все наземные растения имеют половые органы одного типа: мужские называются антеридии, а женские — архегонии. В мужских органах образуется много сперматозоидов, а в женских — одна крупная неподвижная яйцеклетка.
Гаметофит и спорофит могут быть одинаковыми как по строению, так и по продолжительности жизни. Но у наземных растений оба поколения различны. При этом оба поколения могут быть самостоятельными или одно развивается на другом. Так, у мхов спорофит (коробочка со спорами) — часть одного растения, тело которого является гаметофитом. У семенных растений сильно редуцированный, лишенный хлорофилла гаметофит представлен клетками зародышевого мешка.
В процессе эволюции при приспособлении к дефициту воды происходили постепенная редукция (сокращение продолжительности жизненных циклов) полового поколения — гаметофита, упрощение половых органов и увеличение размеров бесполого поколения — спорофита.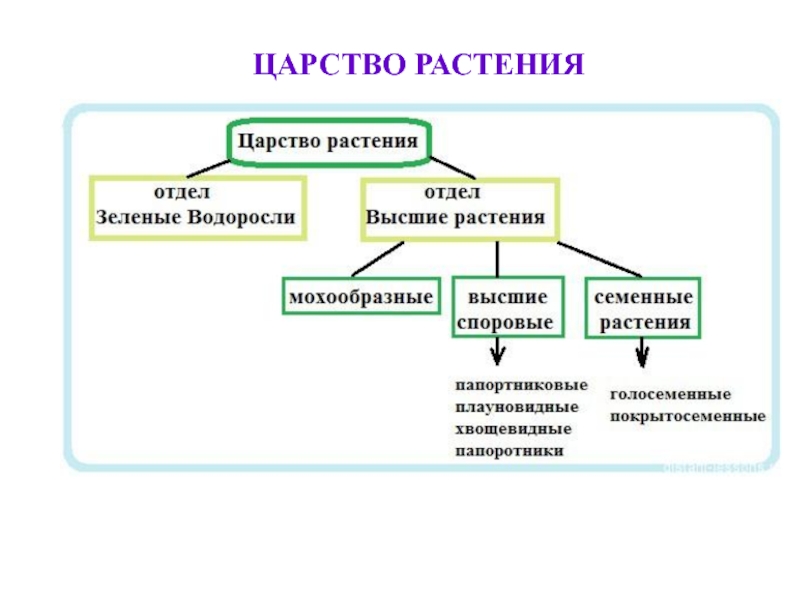
Около 600 тыс. видов растений условно делят на:
низшие —тело не расчленено на органы и ткани и называется слоевищем, или талломом. К этой группе относятся водоросли и лишайники.
высшие — имеются органы (корень, стебель и лист), образованные сложно дифференцированными тканями. Зигота высших растений развивается в многоклеточный зародыш. К высшим растениям относятся отделы моховидные, плауновидные, хвощевидные, папоротниковидные, голосеменные и покрытосеменные (цветковые).
Растения появились на Земле около 2 млрд лет назад. Первоначально развитие растительных организмов происходило в водной среде, что привело к появлению – водорослей. Затем растения стали осваивать сушу. Этому способствовало возникновение следующих ароморфозов:
– возникновение фотосинтеза;
– возникновение эукариотического строения клеток;
– возникновение мейоза и оплодотворения;
– возникновение многоклеточности и дифференциации клеток с образованием тканей и органов;
– возникновение чередования гаплоидного и диплоидного поколений;
– возникновение семени;
– возникновение цветка.
Эволюция растений шла в направлении от споровых к семенным, от низших к высшим.
Контрольная работа «Царство Растения
Контрольная работа «Царство Растения» Вариант №1.
1.Низшие растения — это растения:
1. Имеющие ткани
2. Не имеющие тканей и органов
3. Имеющие ткани и органы
2.Водоросли – это растения имеющие:
1. Корень
2. Листья
3. Слоевище
3.Плауны имеют:
1. Ползучий стебель с множеством побегов
2. Короткий стебель, неветвящийся
3. Древесный стебель, с множеством листьев
4. Мхи размножаются:
а) спорами б) семенами в)корнями г) стеблями
5. Какое значение играют папоротники в жизни человека?
а) участвуют в образовании торфа
б) из них получают агар-агар
в) используют как строительный материал
г) из них образовался каменный уголь
6.Залежи каменного угля образовались:
1. Из древних мхов
Из древних мхов
2. Из древних папоротников
3. Из древних голосеменных
7.В медицине применяются споры:
1. Плауны
2. Хвоща
3. Папоротника
8.Голосеменные – растения:
1. Травянистые и деревья
2. Кустарники и деревья
3. Травы
9.Цветок и плод развиваются:
1. У голосеменных
2. У папоротника
3. У покрытосеменных
10. Весенние побеги хвоща:
а) бурые со спороносными колосками
б) зеленые, называются вайи
в) имеют длинный ползучий стебель
г) имеют ветвящийся зеленый побег
11. Цветок — орган размножения растений, так как:
а) в нем образуются споры
б) его опыляют насекомые
в) в нем образуется нектар
г) в нем образуются гаметы и происходит оплодотворение
12. К одноклеточным водорослям относится:
а) спирогира в) улотрикс
б) хлорелла г) вайи
13. Агар-агар получают из водорослей, относящихся
к
Агар-агар получают из водорослей, относящихся
к
отделу:
а) зеленых в) бурых
б) красных г) колониальных
14. Для семян растений класса однодольных в отличие от семян класса двудольных характерно наличие:
а) семенной кожуры б) зародыша с одной семядолей
в) зародыша с двумя семядолями г) эндосперма
15. К двудольным растениям относится:
а) пшеница б) лук в) кукуруза г) яблоня
16. Охарактеризуйте жизненный цикл Папоротниковидных.-1б
Контрольная работа «Царство Растения» Вариант №2.
1.Высшие растения имеют:
1. Слоевище
2. Вегетативные органы
3. Органы и ткани
2.Кукушкин лен – это:
1. Листостебельное растение, имеет ризоиды
2. Растение, не имеющее стебля и листьев
3. Растение, имеющее корень
3.Вайи папоротников растут:
1. От корневища
2. От стебля
3. От корней
4.Торф образуется из:
1. Папоротников
Папоротников
2. Мхов
3. Плаунов
5.Споры, в отличии от семян:
1. Имеют запас питательных веществ
2. Не имеют запаса питательных веществ
3. Являются защитной оболочкой растений
6.К низшим растениям относятся:
1. Водоросли и некоторые мхи
2. Мхи и плауны
3. Плауны и папоротники
7.Папоротники в основном:
1. Травянистые растения
2. Травянистые, но имеются древовидные формы
3. Древовидные формы
8. Весенние побеги хвоща:
а) бурые со спороносными колосками б) зеленые, называются вайи в) имеют длинный ползучий стебель г) имеют ветвящийся зеленый побег
9. Для семян растений класса однодольных в отличие от семян класса двудольных характерно наличие:
а) семенной кожуры б) зародыша с одной семядолей в) зародыша с двумя семядолями г) эндосперма
10. К двудольным растениям относится:
а) пшеница б) лук в) кукуруза г) яблоня
11. Где находится запас питательных веществ в
семени фасоли?
Где находится запас питательных веществ в
семени фасоли?
а) в эндосперме б) в семядолях в) в корешке г) в семенной кожуре
12. Назовите главные признаки растений семейства крестоцветные:
а. корневая система стержневая, жилкование листьев сетчатое, зародыш с двумя семядолями.
б. цветок пятичленного типа, плод боб, на корнях — клубеньковые бактерии.
в. цветок четырехчленного типа, околоцветник двойной, соцветие кисть, плод стручок или стручочек.
г. цветок пятичленного типа, околоцветник двойной, плод ягода или коробочка, содержат ядовитые веществ
13. К одноклеточным водорослям относится:
а) спирогира в) улотрикс
б) хлорелла г) вайи
14. Агар-агар получают из водорослей, относящихся
к
отделу:
а) зеленых в) бурых
б) красных г) колониальных
15. Назовите главные признаки отдела покрытосеменных:
а. корневая система стержневая, жилкование листьев
сетчатое, зародыш с двумя семядолями. б. наличие цветка, плода с семенами,
хорошо развитая проводящая система.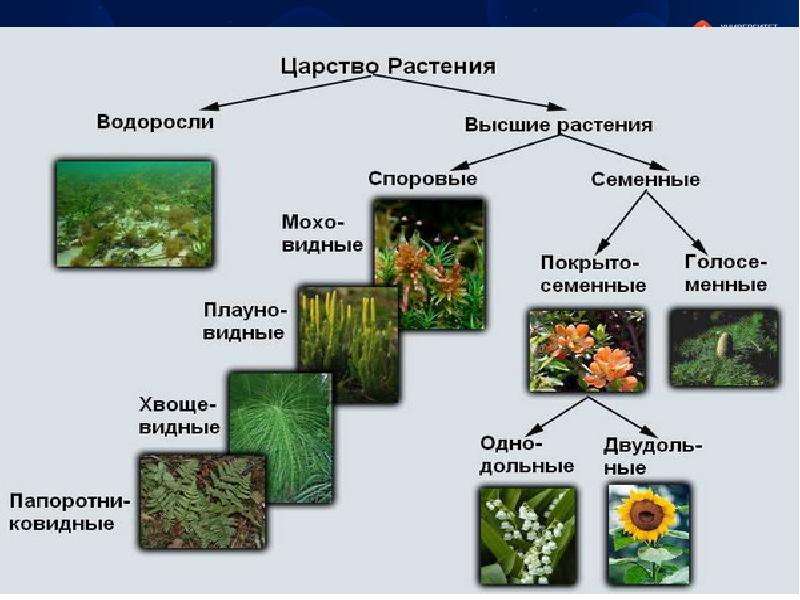
в. корневая система мочковатая, жилкование листьев дуговое или параллельное, зародыш с одной семядолей. г. хорошо развитые вегетативные органы, отсутствие цветка и плода.
16. Охарактеризуйте жизненный цикл Моховидных.-1б.
Kingdom Plantae – обзор
Половой диморфизм у растений
Двудомность у растений встречается гораздо реже, чем у животных. Приблизительно 96 % видов царства Plantae являются цветковыми семенными растениями (покрытосеменными; Roskov et al. , 2014), и только около 6 % видов покрытосеменных двудомными (Renner and Ricklefs, 1995; Barrett, 2002; Vamosi et al. , 2003). Среди остальных семенных растений (голосеменных) 50 % видов двудомные, в том числе 37 % хвойных (Givnish, 1980; Bateman et al., 2011). Среди спорообразующих растений двудомность очень редка у папоротников (Jesson, Garnock-Jones, 2012), но относительно обычна у мохообразных (мхов, печеночников и роголистников), где встречается у 50–60% видов (Hedenas, Bisang, 2011). ; Jesson and Garnock-Jones, 2012; McDaniel и др. , 2013).
; Jesson and Garnock-Jones, 2012; McDaniel и др. , 2013).
У большинства раздельнополых растений пол различают только репродуктивные структуры. Они состоят из первичных репродуктивных органов (например, тычинок и пестиков цветков покрытосеменных) плюс окружающих соматических тканей, часто образованных из видоизмененных стеблей или листьев (например,г., чашечка и венчик покрытосеменных). Половой диморфизм в этих соматических тканях обычно интерпретируется как адаптация самцов к рассеиванию пыльцы или сперматозоидов, а самок к захвату пыльцы или сперматозоидов, а также к защите и обеспечению эмбрионов (Givnish, 1980; Eckhart, 1999; Geber, 1999; Barrett and Hough, 2013). ; McDaniel и др. , 2013). Это преобладающие вторичные половые диморфизмы у растений, и они наиболее выражены у видов, которые зависят от ветра или воды для распространения пыльцы, но производят крупные семена или плоды, которые разносятся животными (Givnish, 1980; Renner and Ricklefs, 1995; Vamosi и др. ). др., 2003; Бернаски, 2010; Бейтман и др. , 2011).
). др., 2003; Бернаски, 2010; Бейтман и др. , 2011).
Вторичный половой признак, наиболее вероятно отличающийся между мужскими и женскими репродуктивными структурами, — это размер. Диморфизм размеров цветков встречается более чем у 80% раздельнополых видов покрытосеменных растений, при этом мужские цветки несколько чаще бывают крупнее, чем наоборот (Eckhart, 1999). Напротив, у раздельнополых голосеменных женские репродуктивные органы (стробилы) крупнее, и только женские стробилы развиваются в большие долгоживущие шишки, типичные для хвойных и саговниковых.У покрытосеменных количество цветков и размеры соцветий (соцветий) также могут различаться между полами: у самцов чаще бывает больше цветков или более крупные соцветия, чем у самок. В нескольких семействах также были описаны различия в форме цветков и в положении или ориентации цветков на растении (Eckhart, 1999; Barrett and Hough, 2013).
Помимо репродуктивных структур, наиболее распространенным SDM у растений является общий размер. Как у покрытосеменных, так и у голосеменных самцы, как правило, крупнее самок у крупных, долгоживущих древесных видов (например,g., деревья и кустарники), тогда как самки, как правило, крупнее у мелких недолговечных травянистых видов (Obeso, 2002; Barrett and Hough, 2013). Размерный диморфизм наиболее выражен у плеврокарпных мхов (класс Bryophyta), где примерно у трети видов карликовые самцы развиваются из спор, которые приземляются на стебли или листья самок и остаются прикрепленными к самкам на протяжении всей их жизни (Hedenas and Bisang, 2011). ).
Как у покрытосеменных, так и у голосеменных самцы, как правило, крупнее самок у крупных, долгоживущих древесных видов (например,g., деревья и кустарники), тогда как самки, как правило, крупнее у мелких недолговечных травянистых видов (Obeso, 2002; Barrett and Hough, 2013). Размерный диморфизм наиболее выражен у плеврокарпных мхов (класс Bryophyta), где примерно у трети видов карликовые самцы развиваются из спор, которые приземляются на стебли или листья самок и остаются прикрепленными к самкам на протяжении всей их жизни (Hedenas and Bisang, 2011). ).
Половой диморфизм в других аспектах вегетативной морфологии растений встречается гораздо реже, но был отмечен по крайней мере у нескольких видов.Примеры включают размер листа (самки чаще крупнее), форму листа, размер стебля (самки чаще толще) и структуру ветвления (самцы обычно более разветвлены) (Dawson and Geber, 1999; Kavanagh et al. , 2011; Barrett and Хью, 2013). Как и у животных, мужские и женские растения также часто различаются по экологическим характеристикам и признакам жизненного цикла (Dudley, 2006; Geber et al.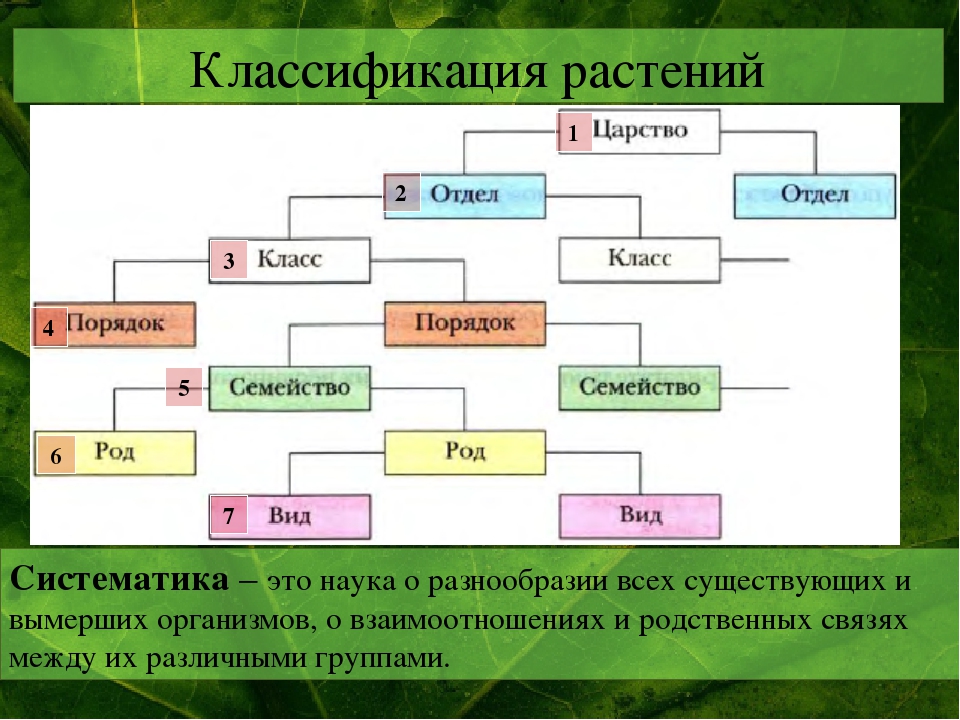 , 1999; Barrett and Hough, 2013). Полы иногда частично разделены средой обитания: самки более ограничены участками с большим количеством воды или питательных веществ, а самцы часто превосходят самок в своей способности к клональному воспроизведению, размножаются в более раннем возрасте и стареют раньше, чем самки.Эти различия отражают специфические для пола компромиссы между ростом и размножением и, как и у животных, интерпретируются как приспособления для максимизации приспособленности за счет мужских или женских репродуктивных функций.
, 1999; Barrett and Hough, 2013). Полы иногда частично разделены средой обитания: самки более ограничены участками с большим количеством воды или питательных веществ, а самцы часто превосходят самок в своей способности к клональному воспроизведению, размножаются в более раннем возрасте и стареют раньше, чем самки.Эти различия отражают специфические для пола компромиссы между ростом и размножением и, как и у животных, интерпретируются как приспособления для максимизации приспособленности за счет мужских или женских репродуктивных функций.
14.1 Царство растений — концепции биологии
Цели обучения
К концу этого раздела вы сможете:- Описать основные характеристики царства растений
- Обсудите проблемы растительной жизни на суше
- Опишите приспособления, позволившие растениям заселить сушу
Растения представляют собой большую и разнообразную группу организмов.Существует около 300 000 видов каталогизированных растений. Из них около 260 000 растений, дающих семена. Мхи, папоротники, хвойные и цветковые растения — все это представители царства растений. Царство растений состоит в основном из фотосинтезирующих организмов; некоторые паразитические формы утратили способность к фотосинтезу. В процессе фотосинтеза используется хлорофилл, который находится в органеллах, называемых хлоропластами. У растений есть клеточные стенки, содержащие целлюлозу. Большинство растений размножаются половым путем, но у них также есть различные способы бесполого размножения.Растения демонстрируют неопределенный рост, что означает, что они не имеют окончательной формы тела, но продолжают наращивать массу тела, пока не умрут.
Из них около 260 000 растений, дающих семена. Мхи, папоротники, хвойные и цветковые растения — все это представители царства растений. Царство растений состоит в основном из фотосинтезирующих организмов; некоторые паразитические формы утратили способность к фотосинтезу. В процессе фотосинтеза используется хлорофилл, который находится в органеллах, называемых хлоропластами. У растений есть клеточные стенки, содержащие целлюлозу. Большинство растений размножаются половым путем, но у них также есть различные способы бесполого размножения.Растения демонстрируют неопределенный рост, что означает, что они не имеют окончательной формы тела, но продолжают наращивать массу тела, пока не умрут.
Адаптация растений к жизни на суше
Поскольку организмы приспосабливаются к жизни на суше, им приходится сталкиваться с рядом проблем в земной среде. Вода была описана как «вещество жизни». Внутренняя часть клетки — среда, в которой растворяется и диффундирует большинство малых молекул и в которой протекает большинство химических реакций обмена веществ, — представляет собой водянистый суп. Иссушение, или высыхание, представляет постоянную опасность для организма, подвергающегося воздействию воздуха. Даже когда части растения находятся близко к источнику воды, их надземные части, скорее всего, высохнут. Вода обеспечивает плавучесть организмам, обитающим в водной среде. На суше растениям необходимо развивать структурную поддержку в воздухе — среде, которая не дает такой же подъемной силы. Кроме того, мужские гаметы должны достичь женских гамет, используя новые стратегии, потому что плавание больше невозможно. Наконец, и гаметы, и зиготы должны быть защищены от высыхания.Успешные наземные растения разработали стратегии для решения всех этих проблем, хотя не все приспособления появились сразу. Одни виды не ушли далеко от водной среды, тогда как другие покинули воду и продолжили завоевывать самые засушливые места на Земле.
Иссушение, или высыхание, представляет постоянную опасность для организма, подвергающегося воздействию воздуха. Даже когда части растения находятся близко к источнику воды, их надземные части, скорее всего, высохнут. Вода обеспечивает плавучесть организмам, обитающим в водной среде. На суше растениям необходимо развивать структурную поддержку в воздухе — среде, которая не дает такой же подъемной силы. Кроме того, мужские гаметы должны достичь женских гамет, используя новые стратегии, потому что плавание больше невозможно. Наконец, и гаметы, и зиготы должны быть защищены от высыхания.Успешные наземные растения разработали стратегии для решения всех этих проблем, хотя не все приспособления появились сразу. Одни виды не ушли далеко от водной среды, тогда как другие покинули воду и продолжили завоевывать самые засушливые места на Земле.
Чтобы сбалансировать эти проблемы выживания, жизнь на суше предлагает несколько преимуществ. Во-первых, много солнечного света. На суше спектральное качество света, поглощаемого фотосинтетическим пигментом, хлорофиллом, не фильтруется водой или конкурирующими фотосинтезирующими видами в толще воды выше. Во-вторых, углекислый газ более доступен, поскольку его концентрация в воздухе выше, чем в воде. Кроме того, наземные растения эволюционировали раньше наземных животных; поэтому, пока суша не была заселена животными, никакие хищники не угрожали благополучию растений. Эта ситуация изменилась, когда животные вышли из воды и нашли обильные источники питательных веществ в установившейся флоре. В свою очередь, растения выработали стратегии отпугивания хищников: от шипов и шипов до ядовитых химикатов.
Во-вторых, углекислый газ более доступен, поскольку его концентрация в воздухе выше, чем в воде. Кроме того, наземные растения эволюционировали раньше наземных животных; поэтому, пока суша не была заселена животными, никакие хищники не угрожали благополучию растений. Эта ситуация изменилась, когда животные вышли из воды и нашли обильные источники питательных веществ в установившейся флоре. В свою очередь, растения выработали стратегии отпугивания хищников: от шипов и шипов до ядовитых химикатов.
Ранние наземные растения, как и ранние наземные животные, не жили далеко от обильного источника воды и разработали стратегии выживания для борьбы с засухой.Одной из таких стратегий является засухоустойчивость. Мхи, например, могут высохнуть и превратиться в коричневый и ломкий коврик, но как только дождь сделает воду доступной, мхи впитают ее и восстановят свой здоровый зеленый вид. Другая стратегия заключается в колонизации среды с высокой влажностью, где засухи случаются редко. Папоротники, ранняя линия растений, процветают во влажных и прохладных местах, например, в подлеске умеренных лесов. Позже растения отошли от водной среды, используя устойчивость к высыханию, а не толерантность.Эти растения, как и кактус, сводят к минимуму потерю воды до такой степени, что могут выжить в самых сухих условиях на Земле.
Позже растения отошли от водной среды, используя устойчивость к высыханию, а не толерантность.Эти растения, как и кактус, сводят к минимуму потерю воды до такой степени, что могут выжить в самых сухих условиях на Земле.
В дополнение к приспособлениям, характерным для жизни на суше, наземные растения демонстрируют приспособления, ответственные за их разнообразие и преобладание в наземных экосистемах. У многих наземных растений обнаруживаются четыре основных приспособления: чередование поколений, спорангий, в котором образуются споры, гаметангий, образующий гаплоидные клетки, а у сосудистых растений — апикальная меристема в корнях и побегах.
Смена поколений
Чередование поколений описывает жизненный цикл, в котором организм имеет как гаплоидные, так и диплоидные многоклеточные стадии (рис. 14.2).
Фигура
14,2
Показано чередование поколений между гаплоидным (1 n ) гаметофитом и диплоидным (2 n ) спорофитом. (кредит: модификация работы Питера Коксхеда)
(кредит: модификация работы Питера Коксхеда)
Гаплонтический относится к жизненному циклу, в котором присутствует доминирующая гаплоидная стадия.Диплоидный относится к жизненному циклу, в котором диплоидная стадия является доминирующей стадией, а гаплоидное число хромосом наблюдается только в течение короткого времени в жизненном цикле во время полового размножения. Например, люди диплонтны. У большинства растений наблюдается чередование поколений, которое описывается как гаплодиплонтическое: за гаплоидной многоклеточной формой, известной как гаметофит, следует в последовательности развития многоклеточный диплоидный организм, спорофит. Гаметофит дает начало гаметам или репродуктивным клеткам путем митоза.Это может быть наиболее очевидная фаза жизненного цикла растения, как у мхов, или она может проявляться в микроскопической структуре, такой как пыльцевое зерно у высших растений (собирательный термин для сосудистых растений). У низших растений стадия спорофита едва заметна (собирательный термин для растительных групп мхов, печеночников и роголистников). Высокие деревья представляют собой диплотическую фазу жизненного цикла таких растений, как секвойи и сосны.
Высокие деревья представляют собой диплотическую фазу жизненного цикла таких растений, как секвойи и сосны.
Спорангии бессемянных растений
Спорофит бессемянных растений является диплоидным и возникает в результате сингамии или слияния двух гамет (рис. 14.2). Спорофит несет спорангии (единственные, спорангии), органы, впервые появившиеся у наземных растений. Термин «спорангий» буквально означает «спора в сосуде», поскольку это репродуктивный мешок, содержащий споры. Внутри многоклеточных спорангиев диплоидные спороциты, или материнские клетки, производят гаплоидные споры в результате мейоза, который уменьшает число хромосом с 2 n до 1 n . Позже споры высвобождаются из спорангиев и рассеиваются в окружающей среде. У наземных растений образуются споры двух разных типов, что приводит к разделению полов на разных этапах жизненного цикла.Бессемянные несосудистые растения (более уместно называемые «бессемянные бессосудистые растения с доминирующей фазой гаметофита») производят только один вид спор и называются гомоспоровыми. После прорастания из споры гаметофит производит как мужские, так и женские гаметангии, обычно у одной и той же особи. Напротив, гетероспоровые растения производят два морфологически разных типа спор. Мужские споры называются микроспорами из-за их меньшего размера; сравнительно более крупные мегаспоры разовьются в женский гаметофит.Гетероспория наблюдается у некоторых бессемянных сосудистых растений и у всех семенных растений.
После прорастания из споры гаметофит производит как мужские, так и женские гаметангии, обычно у одной и той же особи. Напротив, гетероспоровые растения производят два морфологически разных типа спор. Мужские споры называются микроспорами из-за их меньшего размера; сравнительно более крупные мегаспоры разовьются в женский гаметофит.Гетероспория наблюдается у некоторых бессемянных сосудистых растений и у всех семенных растений.
Когда гаплоидная спора прорастает, она образует многоклеточный гаметофит путем митоза. Гаметофит поддерживает зиготу, образовавшуюся в результате слияния гамет, и образовавшуюся молодую спорофитную или вегетативную форму, и цикл начинается заново (рис. 14.3 и рис. 14.4).
Фигура 14.3 Этот жизненный цикл папоротника показывает чередование поколений с доминирующей стадией спорофита.(кредит «папоротник»: модификация работы Кори Занкера; кредит «гаметофит»: модификация работы «Влмастра»/Wikimedia Commons)
Фигура
14,4
Этот жизненный цикл мха показывает чередование поколений с доминирующей стадией гаметофита. (кредит: модификация работы Марианы Руис Вильярреал)
(кредит: модификация работы Марианы Руис Вильярреал)
Споры бессемянных растений и пыльца семенных растений окружены толстыми клеточными стенками, содержащими прочный полимер, известный как спорополленин. Это вещество характеризуется длинными цепочками органических молекул, связанных с жирными кислотами и каротиноидами, и придает большей части пыльцы желтый цвет.Спорополленин необычайно устойчив к химическому и биологическому разложению. Его прочность объясняет существование хорошо сохранившихся окаменелостей пыльцы. Спорополленин когда-то считался нововведением наземных растений; однако теперь известно, что зеленые водоросли Coleochaetes образуют споры, содержащие спорополленин.
Защита зародыша является основным требованием для наземных растений. Уязвимый эмбрион должен быть защищен от высыхания и других опасностей окружающей среды. Как у бессемянных, так и у семенных растений женский гаметофит обеспечивает питание, а у семенных растений зародыш также защищен, поскольку он развивается в новое поколение спорофита.
Гаметангии бессемянных растений
Гаметангии (единственное число, гаметангии) представляют собой структуры на гаметофитах бессемянных растений, в которых гаметы образуются путем митоза. Мужской гаметангий, антеридий, выделяет сперму. Многие бессемянные растения производят сперму, оснащенную жгутиками, которые позволяют им плавать во влажной среде к архегониям, женским гаметангиям. Зародыш развивается внутри архегония как спорофит.
Апикальные меристемы
Побеги и корни растений увеличиваются в длину за счет быстрого деления клеток в ткани, называемой апикальной меристемой (рис. 14.5). Верхушечная меристема представляет собой клеточный колпачок на кончике побега или корня, состоящий из недифференцированных клеток, которые продолжают размножаться на протяжении всей жизни растения. Меристематические клетки дают начало всем специализированным тканям растения. Удлинение побегов и корней позволяет растению получить доступ к дополнительному пространству и ресурсам: свету в случае побега и воде и минеральным веществам в случае корней. Отдельная меристема, называемая латеральной меристемой, производит клетки, увеличивающие диаметр стеблей и стволов деревьев.Апикальные меристемы представляют собой адаптацию, позволяющую сосудистым растениям расти в направлениях, необходимых для их выживания: вверх, к большей доступности солнечного света, и вниз, в почву, для получения воды и необходимых минералов.
Отдельная меристема, называемая латеральной меристемой, производит клетки, увеличивающие диаметр стеблей и стволов деревьев.Апикальные меристемы представляют собой адаптацию, позволяющую сосудистым растениям расти в направлениях, необходимых для их выживания: вверх, к большей доступности солнечного света, и вниз, в почву, для получения воды и необходимых минералов.
Фигура 14,5 Этот саженец яблони является примером растения, у которого верхушечная меристема дает начало новым побегам и росту корней.
Дополнительные адаптации наземных растений
По мере того, как растения приспосабливались к суше и становились независимыми от постоянного присутствия воды во влажных местообитаниях, у них появлялись новые органы и структуры.Ранние наземные растения не вырастали выше нескольких дюймов от земли, и на этих низких циновках они конкурировали за свет. Развивая побег и становясь выше, отдельные растения захватывают больше света. Поскольку воздух обеспечивает значительно меньшую поддержку, чем вода, наземные растения содержат более жесткие молекулы в своих стеблях (а позже и в стволах деревьев). Эволюция сосудистой ткани для распределения воды и растворенных веществ была необходимой предпосылкой для появления у растений более крупных тел. Сосудистая система состоит из тканей ксилемы и флоэмы.Ксилема проводит воду и минеральные вещества, взятые из почвы, к побегу; флоэма переносит пищу, полученную в результате фотосинтеза, по всему растению. Корневая система, которая эволюционировала, чтобы поглощать воду и минералы, также закрепляла все более высокие побеги в почве.
Эволюция сосудистой ткани для распределения воды и растворенных веществ была необходимой предпосылкой для появления у растений более крупных тел. Сосудистая система состоит из тканей ксилемы и флоэмы.Ксилема проводит воду и минеральные вещества, взятые из почвы, к побегу; флоэма переносит пищу, полученную в результате фотосинтеза, по всему растению. Корневая система, которая эволюционировала, чтобы поглощать воду и минералы, также закрепляла все более высокие побеги в почве.
У наземных растений восковой водостойкий покров, называемый кутикулой, покрывает надземные части растения: листья и стебли. Кутикула также предотвращает поглощение углекислого газа, необходимого для синтеза углеводов посредством фотосинтеза.Устьица, или поры, которые открываются и закрываются, чтобы регулировать движение газов и водяного пара, поэтому появились у растений, когда они переместились в более сухие места обитания.
Растения не могут избежать нападения хищных животных. Вместо этого они синтезируют большое количество ядовитых вторичных метаболитов: сложные органические молекулы, такие как алкалоиды, чей ядовитый запах и неприятный вкус отпугивают животных.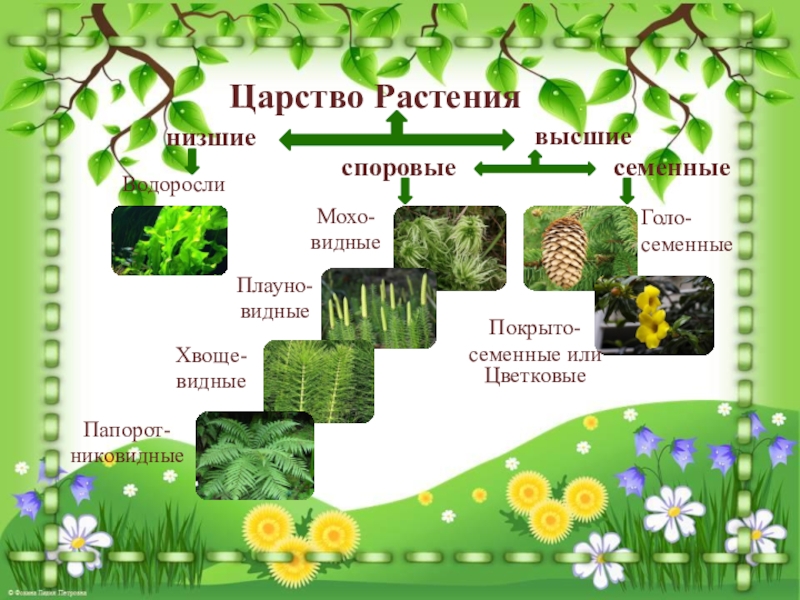 Эти токсичные соединения могут вызывать тяжелые заболевания и даже смерть.
Эти токсичные соединения могут вызывать тяжелые заболевания и даже смерть.
Кроме того, поскольку растения эволюционировали вместе с животными, были разработаны сладкие и питательные метаболиты, чтобы заманить животных и оказать ценную помощь в распространении пыльцевых зерен, фруктов или семян.Растения эволюционировали вместе с животными на протяжении сотен миллионов лет (рис. 14.6).
Фигура 14,6 Растения развили различные приспособления к жизни на суше. (а) Ранние растения росли близко к земле, как этот мох, чтобы избежать высыхания. (b) Более поздние растения развили восковую кутикулу, чтобы предотвратить высыхание. (c) Чтобы вырасти выше, как эти клены, растения должны были выработать новые структурные химические вещества, чтобы укрепить их стебли и сосудистые системы для транспортировки воды и минералов из почвы и питательных веществ из листьев.(d) Растения выработали физическую и химическую защиту, чтобы избежать поедания животными. (кредит a, b: модификация работы Кори Занкера; кредит c: модификация работы Кристин Симала; кредит d: модификация работы Джо Нейлор)
Связь эволюции
Эволюция в действии
Палеоботаника Каким образом организмы приобрели признаки, позволяющие им колонизировать новые среды, и как формируется современная экосистема, — это фундаментальные вопросы эволюции.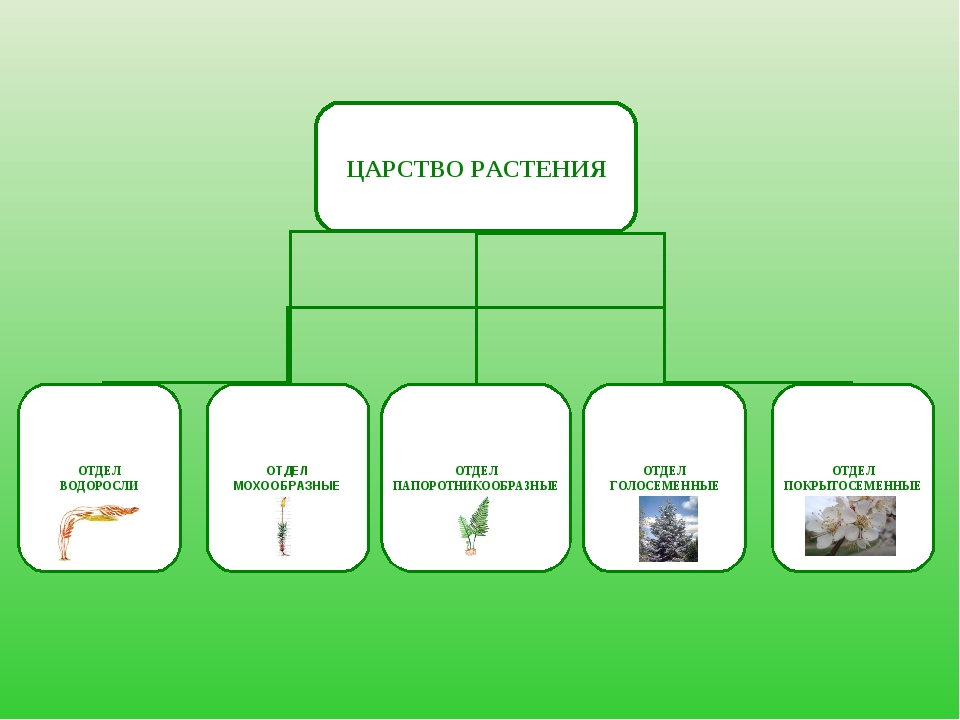 Палеоботаника решает эти вопросы, специализируясь на изучении вымерших растений.Палеоботаники анализируют образцы, извлеченные из полевых исследований, воссоздавая морфологию давно исчезнувших организмов. Они прослеживают эволюцию растений, следя за изменениями в морфологии растений, и проливают свет на связь между существующими растениями, идентифицируя общих предков, обладающих одинаковыми чертами. Эта область направлена на поиск переходных видов, которые преодолевают пробелы на пути к развитию современных организмов. Окаменелости образуются, когда организмы попадают в отложения или среду, где сохраняется их форма (рис. 14.7). Палеоботаники определяют геологический возраст образцов и характер среды их обитания, используя геологические отложения и окружающие их ископаемые организмы. Работа требует большой осторожности, чтобы сохранить целостность хрупких окаменелостей и слоев, в которых они найдены.
Палеоботаника решает эти вопросы, специализируясь на изучении вымерших растений.Палеоботаники анализируют образцы, извлеченные из полевых исследований, воссоздавая морфологию давно исчезнувших организмов. Они прослеживают эволюцию растений, следя за изменениями в морфологии растений, и проливают свет на связь между существующими растениями, идентифицируя общих предков, обладающих одинаковыми чертами. Эта область направлена на поиск переходных видов, которые преодолевают пробелы на пути к развитию современных организмов. Окаменелости образуются, когда организмы попадают в отложения или среду, где сохраняется их форма (рис. 14.7). Палеоботаники определяют геологический возраст образцов и характер среды их обитания, используя геологические отложения и окружающие их ископаемые организмы. Работа требует большой осторожности, чтобы сохранить целостность хрупких окаменелостей и слоев, в которых они найдены.
Одним из самых захватывающих последних достижений в палеоботанике является использование аналитической химии и молекулярной биологии для изучения окаменелостей.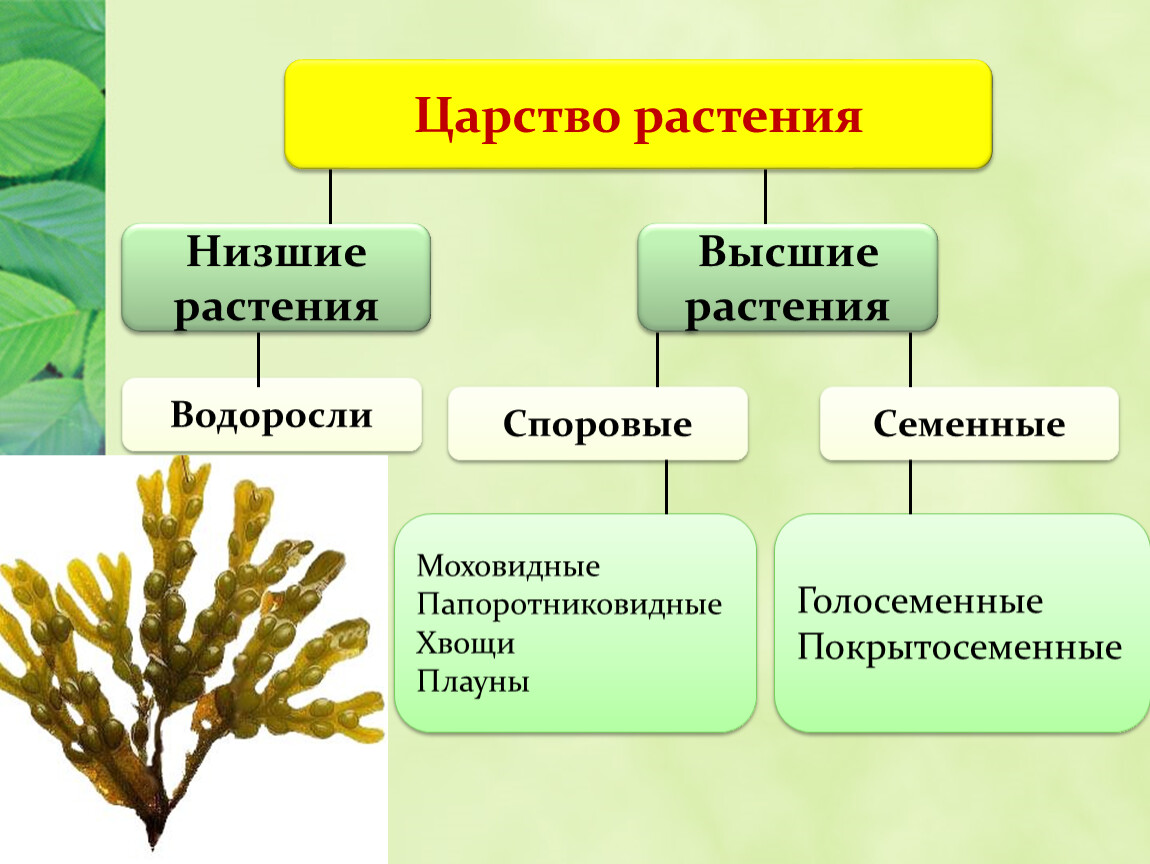 Для сохранения молекулярных структур требуется среда, свободная от кислорода, поскольку окисление и разложение материала в результате деятельности микроорганизмов зависят от присутствия кислорода.Одним из примеров использования аналитической химии и молекулярной биологии является идентификация олеанана, соединения, которое отпугивает вредителей и которое до сих пор казалось уникальным для цветковых растений. Олеанан был извлечен из отложений, датируемых пермским периодом, намного раньше современных дат появления первых цветковых растений. Ископаемые нуклеиновые кислоты — ДНК и РНК — содержат больше всего информации. Их последовательности анализируются и сравниваются с последовательностями живых и родственных организмов.Благодаря этому анализу можно построить эволюционные отношения для линий растений.
Для сохранения молекулярных структур требуется среда, свободная от кислорода, поскольку окисление и разложение материала в результате деятельности микроорганизмов зависят от присутствия кислорода.Одним из примеров использования аналитической химии и молекулярной биологии является идентификация олеанана, соединения, которое отпугивает вредителей и которое до сих пор казалось уникальным для цветковых растений. Олеанан был извлечен из отложений, датируемых пермским периодом, намного раньше современных дат появления первых цветковых растений. Ископаемые нуклеиновые кислоты — ДНК и РНК — содержат больше всего информации. Их последовательности анализируются и сравниваются с последовательностями живых и родственных организмов.Благодаря этому анализу можно построить эволюционные отношения для линий растений.
Некоторые палеоботаники скептически относятся к выводам, сделанным на основе анализа молекулярных окаменелостей. Во-первых, представляющие интерес химические материалы быстро разлагаются во время первоначальной изоляции при воздействии воздуха, а также при дальнейших манипуляциях. Всегда существует высокий риск загрязнения образцов посторонними материалами, в основном микроорганизмами. Тем не менее, по мере совершенствования технологий анализ ДНК окаменелых растений предоставит бесценную информацию об эволюции растений и их адаптации к постоянно меняющейся среде.
Всегда существует высокий риск загрязнения образцов посторонними материалами, в основном микроорганизмами. Тем не менее, по мере совершенствования технологий анализ ДНК окаменелых растений предоставит бесценную информацию об эволюции растений и их адаптации к постоянно меняющейся среде.
Фигура 14,7 Эта окаменелость пальмового листа ( Palmacites sp.), обнаруженная в Вайоминге, датируется примерно 40 миллионами лет назад.
Основные подразделения наземных растений
Наземные растения подразделяются на две основные группы в зависимости от наличия или отсутствия сосудистой ткани, как подробно показано на рис. 14.8. Растения, у которых отсутствует сосудистая ткань, образованная специализированными клетками для транспорта воды и питательных веществ, называются несосудистыми растениями.Мохообразные, печеночники, мхи и роголистники бессемянные и несосудистые и, вероятно, появились на ранних этапах эволюции наземных растений.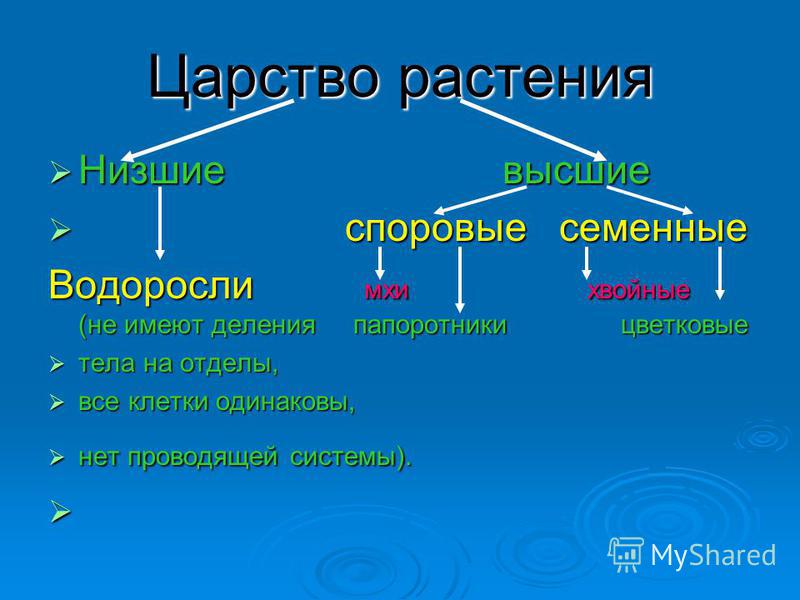 Сосудистые растения развили сеть клеток, которые проводят воду и растворенные вещества через тело растения. Первые сосудистые растения появились в позднем ордовике (461–444 млн лет назад) и, вероятно, были сходны с ликофитами, к которым относятся плауны (не путать со мхами) и птерофиты (папоротники, хвощи, метелки). Ликофиты и птерофиты относятся к бессемянным сосудистым растениям.Они не производят семян, которые представляют собой зародыши с запасами пищи, защищенными твердой оболочкой. Семенные растения составляют самую большую группу всех существующих растений и, следовательно, доминируют в ландшафте. К семенным растениям относятся голосеменные, в первую очередь хвойные, дающие «голые семена», и самые успешные растения, цветковые или покрытосеменные, которые защищают свои семена внутри камер в центре цветка. Стенки этих камер позже развиваются в плоды.
Сосудистые растения развили сеть клеток, которые проводят воду и растворенные вещества через тело растения. Первые сосудистые растения появились в позднем ордовике (461–444 млн лет назад) и, вероятно, были сходны с ликофитами, к которым относятся плауны (не путать со мхами) и птерофиты (папоротники, хвощи, метелки). Ликофиты и птерофиты относятся к бессемянным сосудистым растениям.Они не производят семян, которые представляют собой зародыши с запасами пищи, защищенными твердой оболочкой. Семенные растения составляют самую большую группу всех существующих растений и, следовательно, доминируют в ландшафте. К семенным растениям относятся голосеменные, в первую очередь хвойные, дающие «голые семена», и самые успешные растения, цветковые или покрытосеменные, которые защищают свои семена внутри камер в центре цветка. Стенки этих камер позже развиваются в плоды.
Фигура 14.8 В этой таблице показаны основные подразделения растений.
Plantae Archives — Неукротимая наука
Растения — это живые организмы, принадлежащие к царству Plantae. Они включают знакомые организмы, такие как деревья, травы, кусты, злаки, виноградные лозы, папоротники, мхи и зеленые водоросли. Научное исследование растений, известное как ботаника, выявило около 350 000 существующих видов растений, определяемых как семенные растения, мохообразные, папоротники и их союзники. По состоянию на 2004 г. было идентифицировано около 287 655 видов, из которых 258 650 цветковых и 18 000 мохообразных (см. таблицу ниже).Зеленые растения, иногда называемые Viridiplantae, получают большую часть своей энергии от солнечного света посредством процесса, называемого фотосинтезом.
Они включают знакомые организмы, такие как деревья, травы, кусты, злаки, виноградные лозы, папоротники, мхи и зеленые водоросли. Научное исследование растений, известное как ботаника, выявило около 350 000 существующих видов растений, определяемых как семенные растения, мохообразные, папоротники и их союзники. По состоянию на 2004 г. было идентифицировано около 287 655 видов, из которых 258 650 цветковых и 18 000 мохообразных (см. таблицу ниже).Зеленые растения, иногда называемые Viridiplantae, получают большую часть своей энергии от солнечного света посредством процесса, называемого фотосинтезом.
РАЗНООБРАЗИЕ РАСТЕНИЙ – ПОТРЯСАЮЩИЙ АССОРТИМЕНТ
Наземные растения удивительно разнообразны и легко приспосабливаются, но своим выпрыгиванием из пруда они обязаны наследственной помощи 400 млн лет назад от своих ближайших ныне живущих родственников, зеленых водорослей. Подумайте на минуту о переходе, который претерпели наземные растения, от предкового состояния роста, частично или полностью погруженного под воду, к росту на суше, окруженном не водой, а… воздухом! Это была авантюра, потому что там может быть суровый, сухой, иссушающий мир, но он окупился, и растения процветают в невероятно разнообразных средах по всей Земле.
Прежде чем мы погрузимся в разнообразие растений, давайте сначала совершим небольшое путешествие по закоулкам памяти эволюционного прошлого Земли. Чтобы понять эволюционные отношения группы организмов, биологи строят филогению. Это похоже на генеалогическое древо, за исключением того, что оно фокусируется не только на нескольких поколениях; он уменьшает масштаб и смотрит на отношения в течение эволюционного времени. Полезно знать, какие организмы появились раньше (предковые типы), а какие позже (потомки).Как только биологи выясняют отношения предков и потомков, они строят филогенетическое дерево, используя различные типы информации, такие как структура и форма растения (морфологические данные) и последовательности ДНК определенных генов (молекулярные данные). Растения организованы в филогенетическое дерево на основе сходств и различий в этих данных.
Наземные растения должны были претерпеть захватывающие структурные и репродуктивные изменения, чтобы адаптироваться от водного к наземному образу жизни. Учитывая их огромное разнообразие, все наземные растения имеют по крайней мере одну очень важную общую черту — они проходят уникальный жизненный цикл, называемый чередованием поколений.Это означает, что жизненный цикл любого наземного растения имеет две отдельные фазы. Количество времени, затрачиваемое на каждую фазу, варьируется в зависимости от группы, к которой относится растение.
Учитывая их огромное разнообразие, все наземные растения имеют по крайней мере одну очень важную общую черту — они проходят уникальный жизненный цикл, называемый чередованием поколений.Это означает, что жизненный цикл любого наземного растения имеет две отдельные фазы. Количество времени, затрачиваемое на каждую фазу, варьируется в зависимости от группы, к которой относится растение.
Первая фаза называется поколением гаметофитов (фит = растение, гамето = гамета). На этом этапе мы имеем в виду растение (многоклеточное, а не просто одноклеточное), имеющее один набор хромосом (гаплоидный), производящий гаметы (яйцеклетки и сперматозоиды), которые также являются гаплоидными. Вот действительно интересная часть, яйцеклетка и сперматозоиды производятся посредством митоза, клеточного деления, в результате которого образуются две клетки, идентичные исходной клетке.
Для тех из нас, кто больше знаком с нашим собственным жизненным циклом, чем с жизненным циклом растений, часть описания поколения гаметофита «яйцеклетки и сперматозоиды, образующиеся в результате митоза» должна звучать назойливо в вашем сознании как основное отличие от того, как человек производит собственные яйцеклетки и сперматозоиды. Наши гаметы образуются в результате мейоза или деления одной клетки на четыре клетки, которые содержат половину генетической информации исходной клетки.
Наши гаметы образуются в результате мейоза или деления одной клетки на четыре клетки, которые содержат половину генетической информации исходной клетки.
Вторая фаза называется генерацией спорофита (опять же фит=растение, споро=спора).В этой фазе мы имеем в виду растение (многоклеточное, а не просто одноклеточное), имеющее два набора хромосом (диплоидное), что является результатом слияния двух гамет (сперматозоида и яйцеклетки). В самом общем смысле мы говорим о растении, которое производит споры. Более конкретно, диплоидный спорофит производит гаплоидные споры. Как они это делают? Ну, есть только один тип клеточного деления, при котором общее число хромосом уменьшается во время деления, и это мейоз. Таким образом, спорофит подвергается мейозу с образованием гаплоидных спор.
Теперь, когда мы разобрались в основах чередования поколений, давайте начнем наше путешествие по разнообразию наземных растений и посмотрим, как этот жизненный цикл проявляется в каждой группе.
МОХОВЫЕ
Первой группой, возникшей после зеленых водорослей, были несосудистые растения или мохообразные.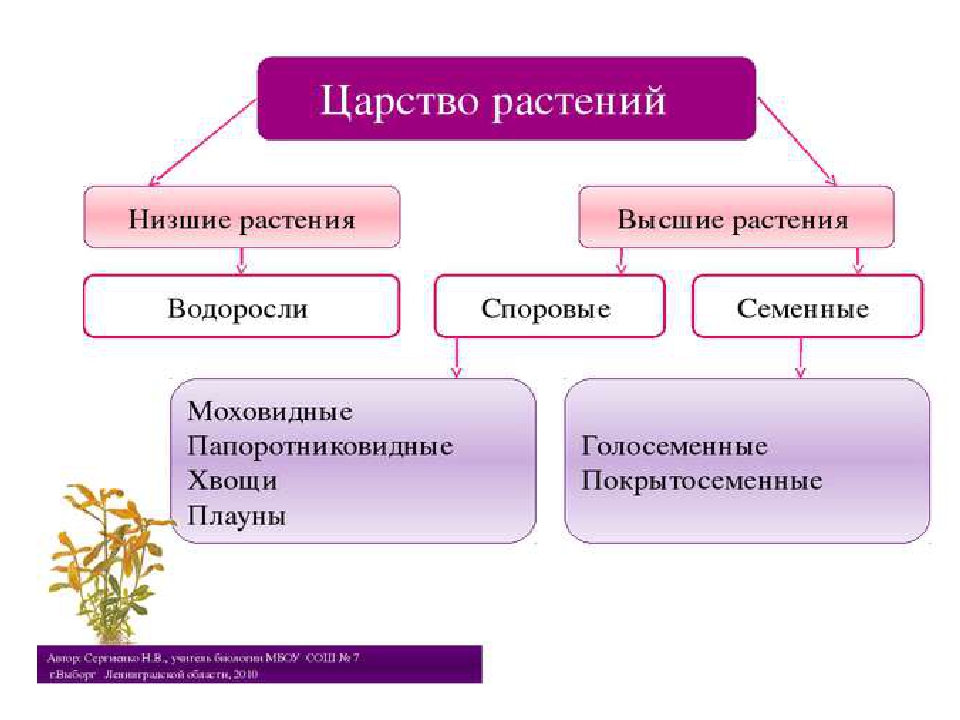 Помните, что сосудистая система — это «сантехника», по которой растения транспортируют воду, минералы и питательные вещества. Сосудистая ткань делится на два типа: ксилема переносит воду и минеральные вещества, а флоэма переносит питательные вещества.Поэтому несосудистые растения, такие как мхи, печеночники и роголистники, не имеют ксилемы или флоэмы, что также означает, что у них нет настоящих корней, стеблей или листьев. У некоторых есть ризоиды, которые могут выглядеть как маленькие корни, но они служат для закрепления растения. Ризоиды не всасывающие.
Помните, что сосудистая система — это «сантехника», по которой растения транспортируют воду, минералы и питательные вещества. Сосудистая ткань делится на два типа: ксилема переносит воду и минеральные вещества, а флоэма переносит питательные вещества.Поэтому несосудистые растения, такие как мхи, печеночники и роголистники, не имеют ксилемы или флоэмы, что также означает, что у них нет настоящих корней, стеблей или листьев. У некоторых есть ризоиды, которые могут выглядеть как маленькие корни, но они служат для закрепления растения. Ризоиды не всасывающие.
Без четко определенной системы транспортировки воды по всему телу, где бы вы догадались, что встречаются эти виды растений? А если бы вы никогда раньше не видели мохообразных, но знали, что у них нет сосудистой системы, вы бы заподозрили, что это растение высокое или низкорослое? Мохообразные и мхи встречаются во влажной среде и обычно низко над землей, поскольку у них нет стеблей или сосудистой системы для транспортировки воды.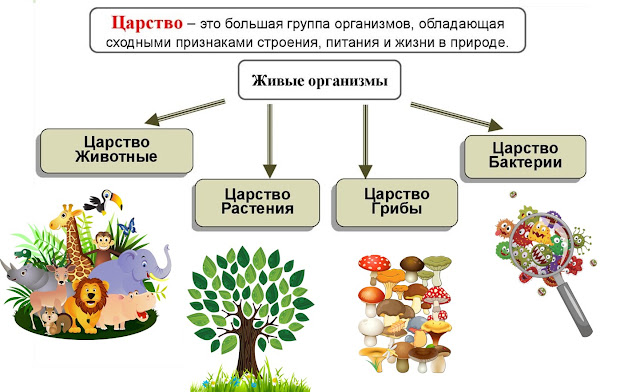
Есть несколько ключевых нововведений, через которые пришлось пройти нашим друзьям мохообразным, чтобы совершить эволюционный скачок из водной среды в наземную. Два структурных изменения позволили этим растениям выжить в сухой земной атмосфере. Во-первых, у мохообразных развилась кутикула, отличная от слоя кожи у основания наших ногтей на руках и ногах. Кутикула растения представляет собой восковой слой, покрывающий растение, который удерживает воду и предохраняет растение от высыхания. Во-вторых, у мохообразных развились устьица, которые представляют собой поры в кутикуле, обеспечивающие газообмен.Конечно, растения выделяют кислород, но им также необходимо поглощать углекислый газ для дыхания, поэтому поры имеют решающее значение для газообмена.
Последней ключевой инновацией мохообразных является репродуктивная адаптация. Напомним, что у мохообразных преобладает поколение гаметофитов. Поскольку это растение производит гаметы, они делают это стильно. Мохообразные развили гаметангии или специализированные структуры, образующие гаметы. Существует два типа: мужская структура, производящая сперму (антеридий), и женская структура, производящая яйцеклетку (архегоний).Мохообразные, несмотря на все успехи, которые они предприняли, чтобы приспособиться к жизни на суше, унаследовали от своих водных предков — им по-прежнему нужна вода для размножения. Сперматозоиды, выпущенные из антеридия, поплывут к архегонию, чтобы оплодотворить яйцеклетку. Развивающаяся зигота фактически вырастает из гаметофита в поколение, производящее споры (спорофит). Это похоже на помаду, вытекающую из тюбика. Выживание диплоидного спорофита полностью зависит от гаметофита. Помада не может выжить без футляра.
Существует два типа: мужская структура, производящая сперму (антеридий), и женская структура, производящая яйцеклетку (архегоний).Мохообразные, несмотря на все успехи, которые они предприняли, чтобы приспособиться к жизни на суше, унаследовали от своих водных предков — им по-прежнему нужна вода для размножения. Сперматозоиды, выпущенные из антеридия, поплывут к архегонию, чтобы оплодотворить яйцеклетку. Развивающаяся зигота фактически вырастает из гаметофита в поколение, производящее споры (спорофит). Это похоже на помаду, вытекающую из тюбика. Выживание диплоидного спорофита полностью зависит от гаметофита. Помада не может выжить без футляра.
БЕССЕМЕННЫЕ СОСУДИСТЫЕ
Ключевым нововведением для этой следующей группы и для всех растений, начиная с этого момента в нашем обзоре разнообразия растений, является сосудистая ткань: ксилема для транспортировки воды и минералов и флоэма для транспортировки сахара. Сосудистые растения имеют настоящие листья, стебли и корни. Сосудистые растения также содержат особое вещество, называемое лигнином, которое представляет собой соединение в клеточных стенках растений, придающее им дополнительную прочность и стабильность. Учитывая вертикальные и структурные преимущества сосудистых растений, неудивительно, что они в целом намного крупнее мохообразных.
Учитывая вертикальные и структурные преимущества сосудистых растений, неудивительно, что они в целом намного крупнее мохообразных.
Бессемянные сосудистые растения включают плауны, папоротники, хвощи и папоротники. Некоторым растениям этой группы по-прежнему нужна вода для оплодотворения. В отличие от мохообразных преобладает поколение спорофитов. Растение, производящее гаметы, или поколение гаметофитов, является свободноживущим, но очень маленьким.
СЕМЯННЫЕ РАСТЕНИЯ
Ключевым нововведением для остальных растений является развитие семян. Это может показаться простым, но развитие семян было главной адаптацией в эволюции растений.Семена крепкие, и самое главное, они могут переносить засушливые условия. Адаптация семени означала, что растения были свободны от зависимости от воды для размножения и, следовательно, могли колонизировать более сухую среду.
Немного о «семени». Это довольно замечательная адаптация, которая была бы невозможна без эволюции характеристики семенных растений, известной как гетероспория.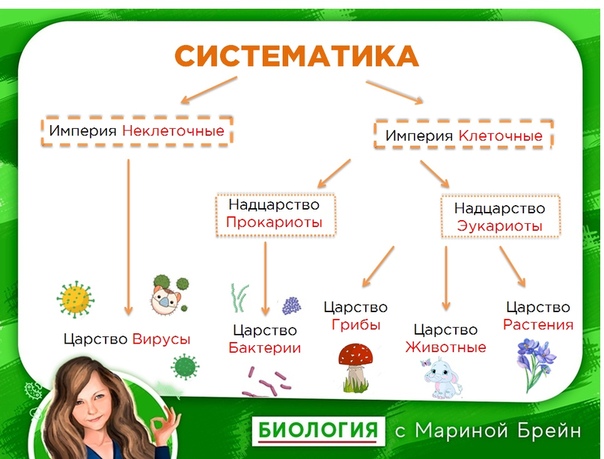 В основном это означает производство двух различных типов структур, производящих спор, и, следовательно, двух различных типов спор: микроспор и мегаспор.Микроспоры развиваются в пыльцу или мужской гаметофит. Мегаспора развивается в яйцо или женский гаметофит. Яйцеклетка и сперматозоид сливаются, образуя зиготу, которая развивается в зародыш, защищенный несколькими слоями и завернутый в защитную оболочку. Весь пакет является семенем!
В основном это означает производство двух различных типов структур, производящих спор, и, следовательно, двух различных типов спор: микроспор и мегаспор.Микроспоры развиваются в пыльцу или мужской гаметофит. Мегаспора развивается в яйцо или женский гаметофит. Яйцеклетка и сперматозоид сливаются, образуя зиготу, которая развивается в зародыш, защищенный несколькими слоями и завернутый в защитную оболочку. Весь пакет является семенем!
Мы разделяем семенные растения на две основные группы: голосеменные и покрытосеменные. Захватывающим аспектом этих двух групп является то, что спорофит является доминирующим, а гаметофит настолько редуцирован, что его выживание зависит от спорофита.Сперматозоид и яйцеклетка развиваются внутри спорофита, а женский гаметофит сохраняется в тканях спорофита. Есть три основных типа голосеменных растений: саговники, гинкго и хвойные. Все голосеменные для опыления полагаются на ветер. Ветер может быть привередливой вещью, и не всегда самой надежной, особенно когда на карту поставлено что-то столь же важное, как обеспечение жизнеспособного потомства (оплодотворение и производство семян)! Наша следующая группа действительно улучшила свою стратегию опыления парой замечательных нововведений.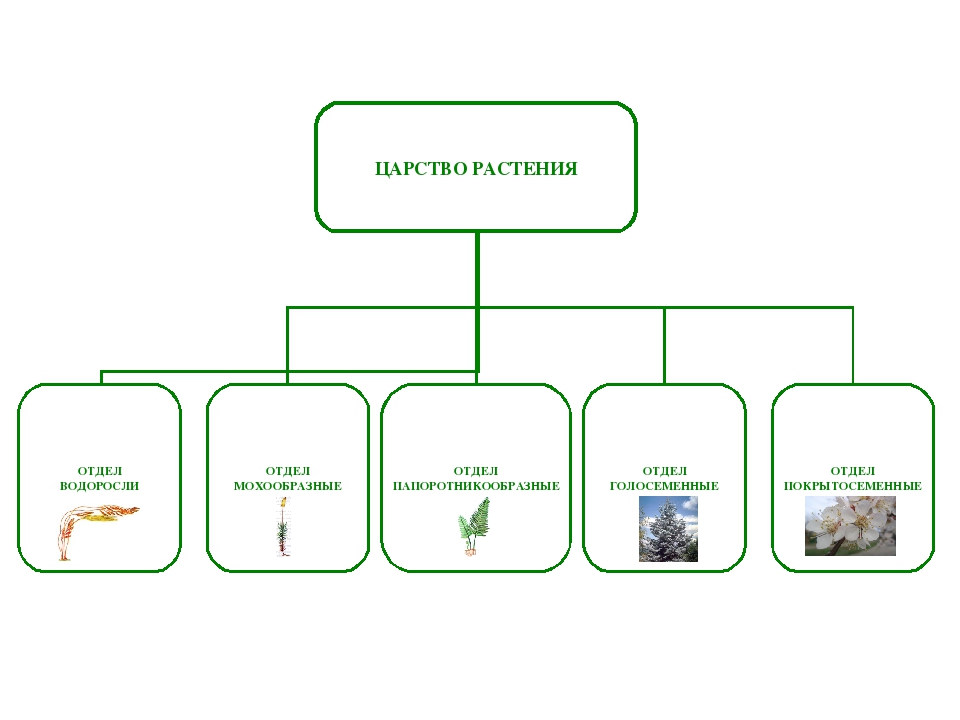
ПОКРЫТОСЕМЯННЫЕ
покрытосеменных – невероятно успешная группа, распространяющаяся по всему земному шару. Есть два секрета успеха покрытосеменных растений: цветы и плоды. Помимо привлечения людей (цветочная индустрия зарабатывает миллиарды долларов в год!) цветы также служат для привлечения опылителей. Это преимущество перед голосеменными растениями, которые полагаются на порывы ветра для переноса пыльцы!
Еще одно ключевое новшество покрытосеменных — плоды. Плоды сладкие, вкусные, а внутри них скрыт весь генетический материал для следующего поколения растений – семян! Это был очень умный маркетинговый ход — упаковать такой драгоценный груз во внешнее покрытие, которое было бы мягким, мясистым, сладким и почти неотразимым для животных.
Итак, как видите, растениям пришлось претерпеть множество структурных и репродуктивных изменений, чтобы адаптироваться от водного к наземному образу жизни. Но имейте в виду всеобъемлющую тенденцию в разнообразии растений – переход от растений с преобладанием гаметофитов к биофитам, к эволюционному промежуточному этапу, на котором доминирует поколение с преобладанием спорофитов, а поколение гамеофита является независимым, к семенным растениям, где поколение гаметофитов настолько снижено.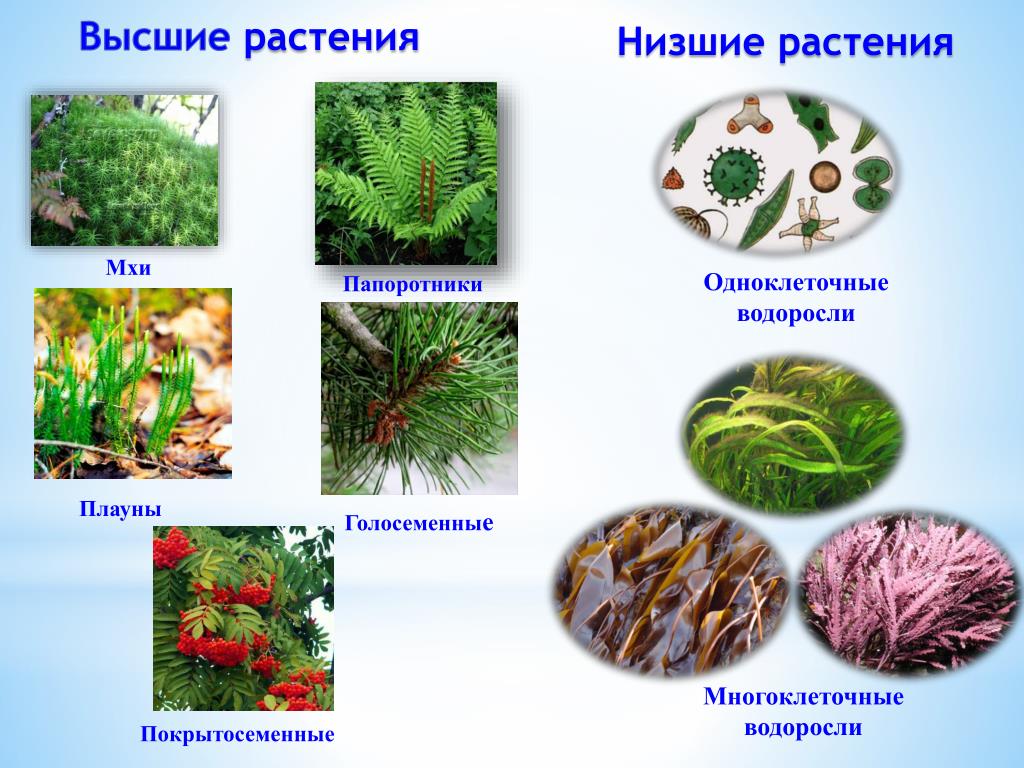 что его выживание зависит от спорофита.
что его выживание зависит от спорофита.
Царство растений | Царство Plantae Классификация и характеристики
Plantae — это царство растений, в которое входят все растения на Земле. Это многоклеточные эукариоты. Характерно, что они содержат жесткую структуру, окружающую клеточную мембрану, известную как клеточная стенка. У растений также есть пигмент зеленого цвета, известный как хлорофилл, который очень важен для фотосинтеза. Следовательно, у них автотрофный способ питания. Царство растений — это огромная группа; следовательно, королевство далее делится на подгруппы.Лучший способ изучить растения — это сначала понять, как происходит классификация в царстве Plantae. Как только вы узнаете простые вещи, вам будет легче подробно ознакомиться с каждым растением.
Биолог Уиттакер представил нам группу пяти царств, разделив все живые организмы на пять территорий — протисты, монеры, грибы, растения и животные. Чтобы узнать больше о растениях, важно знать больше о царстве Plantae или, проще говоря, о царстве растений.

Вы будете удивлены, увидев различные виды растений, которые здесь собраны. Вы можете увидеть крошечные растения, такие как водоросли, а также самые высокие растения, такие как секвойя.
Некоторые растения имеют в высшей степени красивые и привлекательные органоиды, называемые цветками, а некоторые их не содержат. У некоторых растений можно увидеть соответствующую корневую систему, систему побегов и листья, тогда как у более простых растений это только таллоидная структура. Уровень классификации основан на следующих трех критериях :1.Растительное тело: имеет ли тело хорошо дифференцированные сборки или нет.2. Сосудистая система: имеет ли растение сосудистую систему для переноса веществ или нет.
3. Развитие семян: дает ли растение цветы и семена или нет; если это так, то независимо от того, окружено оно фруктами или нет. Каждая группа растений имеет отличные и уникальные особенности, которые подходят исключительно этой группе. В то время как таллофиты являются самыми зелеными растениями, покрытосеменные растения представляют собой растения со сложным строением и хорошо развитой сосудистой и репродуктивной системами.

Голосеменные – это группа растений, имеющих эволюционное значение. Это первые растения с развитыми семенами, что является хорошо организованным механизмом продолжения группы растений в местах, удаленных от родительского растения.
Учитывая все эти особенности, царство растений было разделено на пять подгрупп. Они следующие:
1. Thallophyta
2. Bryophyta
3. Pteryidophyta
3. Gymnosperms
Thallophyta
Все растения, которые имеют короткий и хорошо дифференцированный бодибилдинг, относятся к подкатегории Thallophyta.Таллофиты обычно известны как водоросли. Популярность у них на водной основе. Некоторыми примерами являются Spirogyra, Chara, Ulothrix и т. д.Характеристики Thallophyta
 Поэтому они встречаются во влажных или мокрых местах.
Поэтому они встречаются во влажных или мокрых местах. Подразделение Thallophyta:
Подразделение Thallophyta подразделяется на две части: водоросли и грибы. Они представляют собой таллоиды, содержащие хлорофилл.
 Они автотрофы и в основном водные растения. Кстати, было замечено, что зеленые водоросли образуют симбиотические отношения с ленивцами, которые являются естественными для пышных тропических лесов Южной Америки и Центральной Америки.Мех ленивца очень грубый и легко впитывает воду. В результате процедуры с мехом ленивца представляют собой влажную и влажную среду для процветания водорослей. Взамен водоросли обеспечивают ленивца дополнительным питанием и маскировкой от хищников. Пример: Spirogyra. Это бесхлорофильные (что означает, что они не создают хлорофилл) гетеротрофные таллофиты. Иногда, чтобы преодолеть этот недостаток, грибы могут вступать в симбиотические отношения с водорослями или цианобактериями. Водоросли могут создавать пищу, так как они содержат хлорофилл, а грибы, в свою очередь, обеспечивают безопасную среду, защищающую водоросли от ультрафиолетовых лучей.Лишайник — это пример, когда два существа действуют как единое целое.
Они автотрофы и в основном водные растения. Кстати, было замечено, что зеленые водоросли образуют симбиотические отношения с ленивцами, которые являются естественными для пышных тропических лесов Южной Америки и Центральной Америки.Мех ленивца очень грубый и легко впитывает воду. В результате процедуры с мехом ленивца представляют собой влажную и влажную среду для процветания водорослей. Взамен водоросли обеспечивают ленивца дополнительным питанием и маскировкой от хищников. Пример: Spirogyra. Это бесхлорофильные (что означает, что они не создают хлорофилл) гетеротрофные таллофиты. Иногда, чтобы преодолеть этот недостаток, грибы могут вступать в симбиотические отношения с водорослями или цианобактериями. Водоросли могут создавать пищу, так как они содержат хлорофилл, а грибы, в свою очередь, обеспечивают безопасную среду, защищающую водоросли от ультрафиолетовых лучей.Лишайник — это пример, когда два существа действуют как единое целое. Bryophyta
 Но у них отсутствует сосудистое устройство для переноса веществ через тело растения. Мохообразные происходят как из наземной, так и из водной среды, поэтому называются земноводными царства растений. К этой подкатегории подходят мхи и маршантии. Единственная важная особенность мохообразных состоит в том, что у них нет настоящего сосудистого вещества.У некоторых есть ткани, которые используются для переноса воды, но не считаются истинно сосудистой тканью из-за нехватки лигнина.
Но у них отсутствует сосудистое устройство для переноса веществ через тело растения. Мохообразные происходят как из наземной, так и из водной среды, поэтому называются земноводными царства растений. К этой подкатегории подходят мхи и маршантии. Единственная важная особенность мохообразных состоит в том, что у них нет настоящего сосудистого вещества.У некоторых есть ткани, которые используются для переноса воды, но не считаются истинно сосудистой тканью из-за нехватки лигнина. Считается, что мохообразные произошли от харофитов и считаются первыми настоящими растениями, когда-либо созданными. Характеристики мохообразных • Растения этой группы не имеют корней, но содержат грубые стебли и листья.
• Они содержат «ризоиды» на месте корней, которые помогают растению закрепляться на поверхности.
• Эти корни не поглощают питательные вещества, как другие типичные корни растений.
• Мхи выделяют споры со своих листьев, которые проходят через воду и создают новые мхи на новых участках.

• Вода очень важна для роста и распространения мхов. Они могут полностью высохнуть и выжить. При соприкосновении основания с водой снова просыпаются и продолжают свой рост. Жизненный цикл мохообразных, как и у всех других наземных растений (эмбриофитов), со сменой поколений. Гаплоидная клетка гаметофит имеет фиксированное число непарных хромосом. Это дает рост диплоидному спорофиту, который, однако, имеет вдвое большее количество парных хромосом.Диплоидные зиготы образуются путем слияния гаплоидных сперматозоидов и яйцеклеток, образованных гаметофитами. Диплоидные зиготы образуют спорофит.
Pteridophyta
2. Selaginellidae (шиповидные мхи, иглы)
• Polypodiopsida
 Psilotidae: Ophioglossales (например, виноградные папоротники) и Psilotales.
Psilotidae: Ophioglossales (например, виноградные папоротники) и Psilotales. 2. Equisetidae (хвощи)
3. Polypodiidae (наиболее богатая видами группа, лептоспорангиатные папоротники)
4. Marattiidae (мараттиоидные папоротники) Pteridophyta Характеристика • Птеридофиты – первые настоящие наземные растения: предполагается, что жизнь зародилась в океаны, и в течение миллионов лет развития жизнь медленно адаптировалась к суше (земле). И одними из первых растений, которые действительно жили на земле, были папоротникообразные.
• Это растения без косточек, сосудистые криптогамы: папоротники бессемянные и производят споры. Они не должны проводить ткани для переноса воды и минералов. Вместо этого вода и минералы перетекают с поверхности растительной клетки в клетку в теле растения. Это также одно из объяснений того, почему этим растениям для жизни нужна постоянно влажная среда.
• Они показывают истинную смену поколений: у папоротников наблюдается группа спорофитов и группа гаметофитов.

• Спорофит имеет настоящие корни, листья и стебель.
• Споры, развитые в спорангиях, являются гомоспористыми: Спорангии представляют собой сборки, в которых образуются споры. Они, как правило, гомоспоровые (важность: образуется один тип спор), а также гетероспоровые (что означает: образуются два вида спор).
• Молодые листья спорофита имеют круговую вязкость: кончики листьев имеют тенденцию закручиваться внутрь, чтобы защитить уязвимые растущие части.
• Половые органы многоклеточные и покрытые оболочкой: мужские половые органы называются антеридиями, а женские половые органы называются архегониями.
Голосеменные
Голосеменные – это растения, которые имеют хорошо выраженное растительное тело, сосудистую структуру и дают семена. Название происходит от греческих слов, гимно означает голый и спермак означает семя. Семена голосеменных растений голые, что означает, что они не заключены в плод. К этому кластеру относятся постоянные вечнозеленые древесные деревья.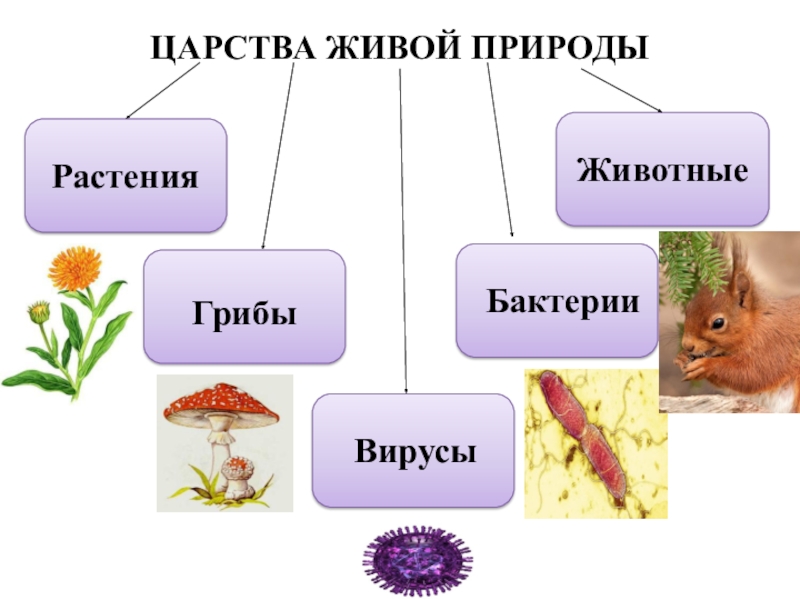 Сосна, деодар, красное дерево и т. д. — вот несколько хороших примеров.
Сосна, деодар, красное дерево и т. д. — вот несколько хороших примеров.
 У небольшого числа дополнительных видов, таких как растение Cycas, корни имеют форму специализированных корней, называемых кораллоидными корнями, и связаны с азотфиксирующими цианобактериями.Эти растения также сосудистые, с наличием как ксилемы, так и флоэмы.
У небольшого числа дополнительных видов, таких как растение Cycas, корни имеют форму специализированных корней, называемых кораллоидными корнями, и связаны с азотфиксирующими цианобактериями.Эти растения также сосудистые, с наличием как ксилемы, так и флоэмы. Покрытосеменные растения
Покрытосеменные также являются семясодержащими растениями с хорошо различимым растительным телом. Термин происходит от греческих слов: ангио: заключенный и сперма: семя. В отличие от голосеменных, семена покрытосеменных покрыты плодами. Покрытосеменные обычно известны как цветковые растения. Например, он содержит дерево манго, растение граната и т. Д. Семена развиваются из зародышевых листьев, называемых семядолями.В зависимости от количества семядолей, существующих в семенах, покрытосеменные делятся на две части: однодольные (однодольные) и двудольные (двудольные). Все покрытосеменные растения имеют типичный сосудистый пучок с материалами ксилемы и флоэмы для проведения воды, минералов и питательных веществ. Тело растения хорошо выделяется развитой корневой системой, системой побегов и листьями. Присутствуют особые структуры, называемые цветами. В этих цветках созревают мужские и женские гаметы. После опыления, когда эти цветы развились, образуются плоды с семенами внутри.Покрытосеменные растения можно увидеть в различных средах обитания и они могут быть разных размеров. Вольфи — крошечное покрытосеменное растение, в то время как австралийская рябина имеет высоту около 100 метров. Разнообразие, которое видели покрытосеменные растения, очень велико. Есть несколько растений, которые представляют собой очень высокие древесные деревья, кустарники и даже травянистые растения. Эти растения также имеют многочисленные адаптации корней, стеблей и листьев в зависимости от среды обитания, в которой они произрастают. Классификация покрытосеменных растений В зависимости от существующих типов семядолей покрытосеменные делятся на два класса.Это однодольные и двудольные растения. Двудольные покрытосеменные имеют 2 семядоли в семенах, а однодольные покрытосеменные содержат 1 семядолю.
Тело растения хорошо выделяется развитой корневой системой, системой побегов и листьями. Присутствуют особые структуры, называемые цветами. В этих цветках созревают мужские и женские гаметы. После опыления, когда эти цветы развились, образуются плоды с семенами внутри.Покрытосеменные растения можно увидеть в различных средах обитания и они могут быть разных размеров. Вольфи — крошечное покрытосеменное растение, в то время как австралийская рябина имеет высоту около 100 метров. Разнообразие, которое видели покрытосеменные растения, очень велико. Есть несколько растений, которые представляют собой очень высокие древесные деревья, кустарники и даже травянистые растения. Эти растения также имеют многочисленные адаптации корней, стеблей и листьев в зависимости от среды обитания, в которой они произрастают. Классификация покрытосеменных растений В зависимости от существующих типов семядолей покрытосеменные делятся на два класса.Это однодольные и двудольные растения. Двудольные покрытосеменные имеют 2 семядоли в семенах, а однодольные покрытосеменные содержат 1 семядолю. Однодольные растения имеют некоторые отличия, заключающиеся в наличии придаточных корней, простых листьев с похожим жилкованием и трехчленных цветков. Количество сосудистых пучков больше и замкнуто. Некоторые из примеров содержат бамбук, бананы, растения сахарного тростника, злаки, лилии и т. д. Двудольные растения имеют две (2) семядоли со стержневым корнем.Жилкование, наблюдаемое в листьях, сетчатое. Цветки могут быть четырехчленными или пятичленными. Сосудистые пучки обычно располагаются кольцом и насчитывают от 2 до 6. Примеры включают виноград, подсолнух, помидоры, одуванчики и картофель. Криптогамы и фанерогамы Царство растений было разделено на две группы — криптогамы и фанерогамы — в зависимости от их способности к развитию семян. Криптогамы — это растения, у которых нет хорошо сложенных или видимых репродуктивных органов. Они имеют скрытые репродуктивные органы и не производят семян.Таллофиты, мохообразные и папоротники относятся к криптогамам. Размножение у всех трех особей происходит посредством образования спор.
Однодольные растения имеют некоторые отличия, заключающиеся в наличии придаточных корней, простых листьев с похожим жилкованием и трехчленных цветков. Количество сосудистых пучков больше и замкнуто. Некоторые из примеров содержат бамбук, бананы, растения сахарного тростника, злаки, лилии и т. д. Двудольные растения имеют две (2) семядоли со стержневым корнем.Жилкование, наблюдаемое в листьях, сетчатое. Цветки могут быть четырехчленными или пятичленными. Сосудистые пучки обычно располагаются кольцом и насчитывают от 2 до 6. Примеры включают виноград, подсолнух, помидоры, одуванчики и картофель. Криптогамы и фанерогамы Царство растений было разделено на две группы — криптогамы и фанерогамы — в зависимости от их способности к развитию семян. Криптогамы — это растения, у которых нет хорошо сложенных или видимых репродуктивных органов. Они имеют скрытые репродуктивные органы и не производят семян.Таллофиты, мохообразные и папоротники относятся к криптогамам. Размножение у всех трех особей происходит посредством образования спор.
Растения, которые имеют видимые репродуктивные органы и могут производить семена, называются «фанерогамами». Голосеменные и покрытосеменные занимают свое место в группе фанерогамов.
Введение в Plantae
Введение в PlantaeЗеленое королевство
К Plantae относятся все наземные растения: мхи, папоротники, хвойные, цветковые растения и т. д. — удивительное разнообразие разнообразных форм.Насчитывая более 250 000 видов, они уступают в размерах только членистоногим.
Растения существуют очень давно. Впервые растения появились в ордовика, но не стал напоминают современные растения вплоть до позднего силура. К концу девона, около 360 миллионов лет назад, вокруг было большое разнообразие форм и размеров растений, в том числе крошечные ползучие растения и высокие лесные деревья.
Наиболее ярким и важным признаком растений является их зеленый цвет.
результат пигмента под названием хлорофилл.Растения используют хлорофилл для улавливания энергии света, которая способствует производству
пища – сахар, крахмал и другие углеводы. Без этих источников пищи
большая часть жизни на земле была бы невозможна. Там все еще будет
грибы и водоросли, но не было бы фруктов, овощей, злаков,
или любые животные (которые в конечном итоге тоже полагаются на растения в качестве пищи!)
Без этих источников пищи
большая часть жизни на земле была бы невозможна. Там все еще будет
грибы и водоросли, но не было бы фруктов, овощей, злаков,
или любые животные (которые в конечном итоге тоже полагаются на растения в качестве пищи!)
Еще одним важным вкладом растений является их формирование окружающей среды. Подумайте о месте без растений.Единственными такими местами на земле являются арктические пустоши, действительно засушливые пустыни и глубокие океаны. Везде, начиная с от тундры до тропического леса до пустыни, населенной растениями. На самом деле, когда мы думаем о конкретном пейзаже, это растения, которые первыми приходят на ум. Попробуйте представить себе лес без деревьев или прерию без травы. Это растения, которые производят и поддерживают земную среду, какой мы ее знаем.
Нажмите на кнопки ниже, чтобы узнать больше о Царстве растений.Вы можете углубиться в группы Plantae, выбрав Систематика!
Посетите также следующие сайты для получения дополнительной информации о палеоботаника, систематика растений и эволюция:
Таксономия растений | Давайте поговорим о науке
Таксономия – это метод, используемый учеными для классификации всех живых существ, чтобы лучше понять их эволюционные отношения. Таксономия включает описания и идентификацию видов. Современная таксономия возникла в середине 1700-х годов, когда Карл Линней создал систему классификации всех живых организмов. Он сделал это, дав каждому виду латинское название, состоящее из двух частей, также известное как научное название . Линней классифицировал растения на основе их репродуктивных структур , пытаясь лучше понять эволюционные отношения между различными растениями.
Таксономия включает описания и идентификацию видов. Современная таксономия возникла в середине 1700-х годов, когда Карл Линней создал систему классификации всех живых организмов. Он сделал это, дав каждому виду латинское название, состоящее из двух частей, также известное как научное название . Линней классифицировал растения на основе их репродуктивных структур , пытаясь лучше понять эволюционные отношения между различными растениями.
Все живые существа сгруппированы в широкие категории, называемые царствами .Растения относятся к царству Plantae . Внутри королевства есть много категорий, называемых phyla (единственное число – это phylum). Царство растений включает типы Chlorophyta (зеленые водоросли), Bryophyta (мхи), Pteridophyta (папоротники), Coniferophyta (хвойные) и Magnoliophyta
Chlorophyta – Зеленые водоросли
Хотя технически это не растения, зеленые водоросли представляют собой организмы, из которых произошли наземные растения. К зеленым водорослям относятся одноклеточных жгутиконосцев (одноклеточные организмы с хвостовидными структурами), многоклеточные формы и макроскопические водоросли — все они могут фотосинтезировать. Зеленые водоросли в основном водные; чаще всего встречается в пресноводных и морских средах обитания, но также может быть найден на деревьях и скалах. Некоторые также симбиотически (живут во взаимоотношениях) с грибами, образуя так называемые лишайники . Группа зеленых водорослей, наиболее тесно связанных с наземными растениями, — это харофиты.
К зеленым водорослям относятся одноклеточных жгутиконосцев (одноклеточные организмы с хвостовидными структурами), многоклеточные формы и макроскопические водоросли — все они могут фотосинтезировать. Зеленые водоросли в основном водные; чаще всего встречается в пресноводных и морских средах обитания, но также может быть найден на деревьях и скалах. Некоторые также симбиотически (живут во взаимоотношениях) с грибами, образуя так называемые лишайники . Группа зеленых водорослей, наиболее тесно связанных с наземными растениями, — это харофиты.
Мохообразные – Мхи
Это самые примитивные настоящие растения. У мхов отсутствует сосудистая система , а это означает, что у них нет ткани (группы клеток, которые работают вместе для выполнения определенной функции/роли) для транспортировки воды по всему растению.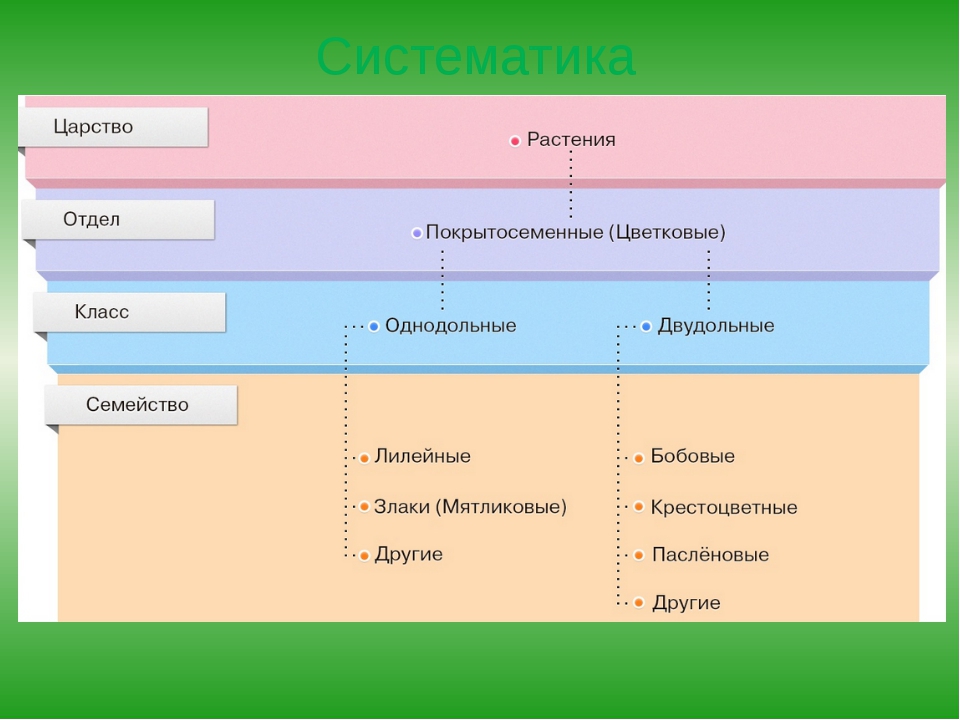 Вместо этого мхи получают воду, питательные вещества и минералы в процессе, называемом осмосом , когда вода перемещается из областей с большим количеством воды в области с меньшим количеством воды.Поскольку мохообразные не могут легко переносить воду и питательные вещества по всему большому растению, мхи растут низко над землей (обычно высотой всего несколько сантиметров). У мхов также отсутствуют корней . Вместо этого у мхов есть ризоидов , которые представляют собой корневидные нити, которые помогают прикрепить растение к земле, не поглощая питательные вещества и воду.
Вместо этого мхи получают воду, питательные вещества и минералы в процессе, называемом осмосом , когда вода перемещается из областей с большим количеством воды в области с меньшим количеством воды.Поскольку мохообразные не могут легко переносить воду и питательные вещества по всему большому растению, мхи растут низко над землей (обычно высотой всего несколько сантиметров). У мхов также отсутствуют корней . Вместо этого у мхов есть ризоидов , которые представляют собой корневидные нити, которые помогают прикрепить растение к земле, не поглощая питательные вещества и воду.
Мхи обычно живут во влажных и тенистых местах и растут группами, образуя густые, мягкие массы растительности. Мхи — одни из первых видов растений, приживающихся на каменистой почве.Подобно лишайникам, они способны разрушать горные породы, обеспечивая ранние стадии формирования почвы, что необходимо для роста крупных растений. Мхи способны поглощать воду, во много раз превышающую их вес, и помогают предотвратить эрозию почвы, улавливая дождевые осадки.
Птеридофиты – Папоротники
Все еще примитивные, но более развитые, чем мхи, папоротники имеют сосудистую систему для транспортировки воды и питательных веществ по всему растению. Сосудистая система включает такие ткани, как ксилема (транспортная ткань, которая перемещает воду по растению) и флоэма (транспортная ткань, которая перемещает питательные вещества, особенно сахар, по растению).У папоротников есть корни, которые поглощают воду и питательные вещества из почвы, которые затем переносятся к стеблям и листьям через ксилему и флоэму. Папоротники также известны своими fiddleheads – скрученными листьями молодых папоротников. Папоротники растут в самых разных местах обитания, в том числе в горах, в расщелинах скал, вокруг болот и во влажных лесах. Одним из самых распространенных папоротников является папоротник орляк.
Pteridium aquilinum , широко известный как папоротник орляк-орляк, встречается в умеренных и субтропических регионах (Источник: изображение © 2013 Ariane Batic.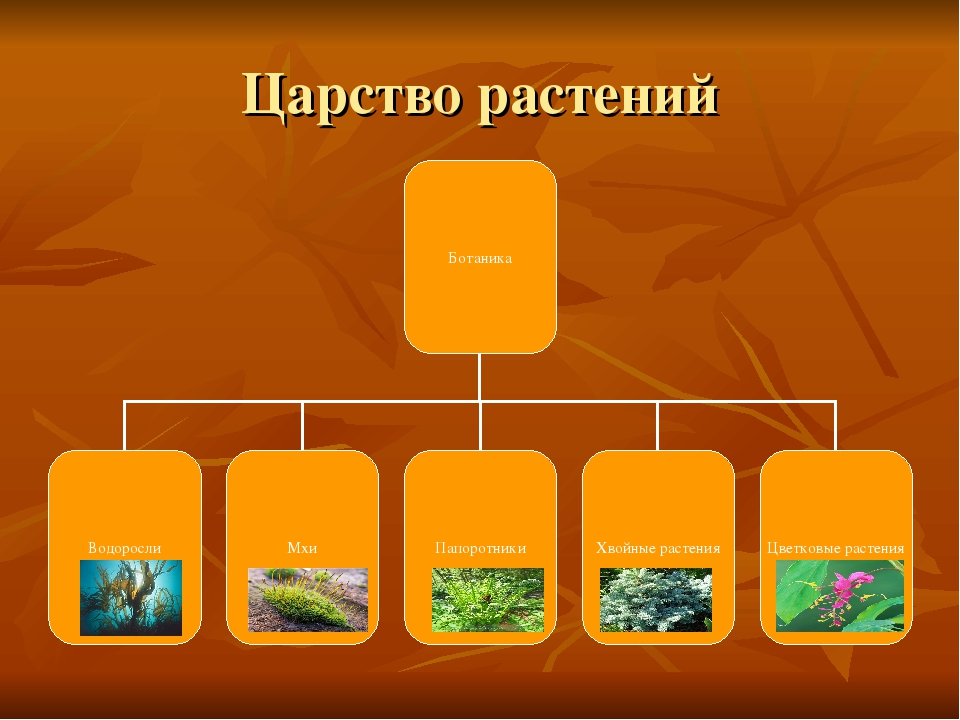 Используется с разрешения).
Используется с разрешения).
Coniferophyta – Хвойные
Голосеменные – это первая группа сосудистых растений, дающая семян (см. стр. 13). В отличие от покрытосеменных (цветковых растений), голосеменные растения имеют непокрытые семена, отсюда и их название — gymno, означает «голый» и сперматозоиды, означает «семя». Наиболее распространенным знакомым типом голосеменных растений являются хвойные ( хвойных ), к которым относятся сосны, кедры, можжевельники, ели и многие другие.
Хвойные деревья деревянистые и имеют шишек и иголок (на самом деле иглы — это просто длинные заостренные листья!). Они также, как правило, вечнозеленые , что означает, что они не теряют свои листья/хвою зимой, а вместо этого постепенно заменяют их на протяжении всей жизни. Сосновые иголки покрыты восковым налетом, что позволяет им удерживать воду в течение всей зимы, но в то же время позволяет им улавливать солнечный свет для фотосинтеза.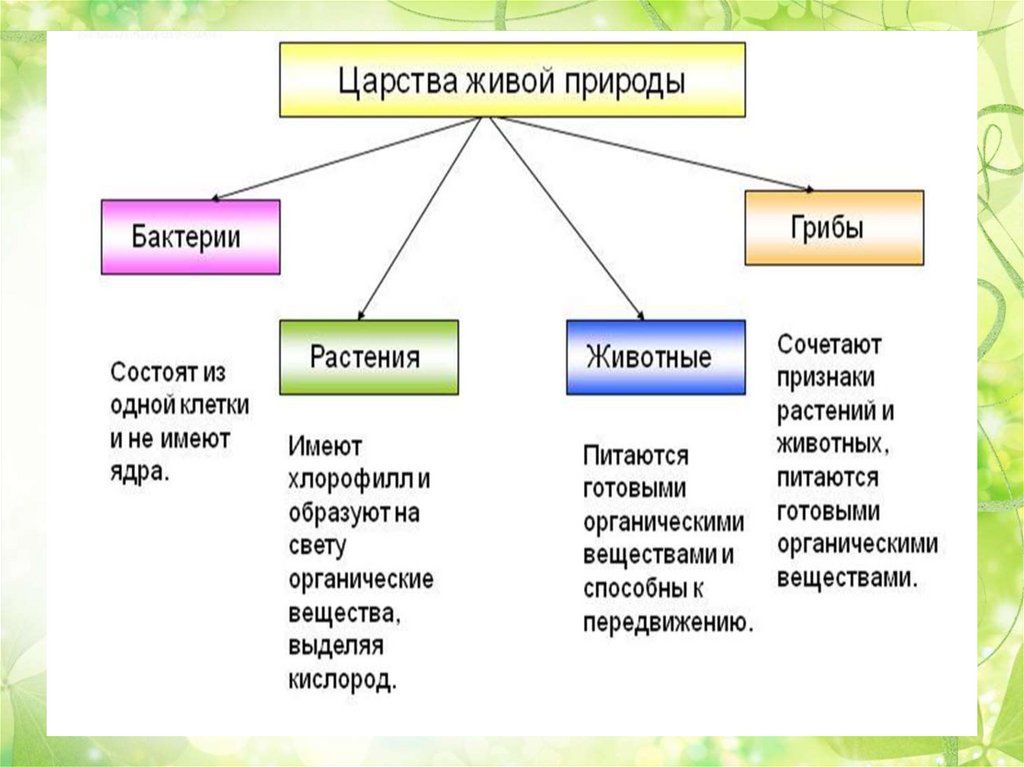 Хвойные в основном встречаются в северном полушарии.Они процветают в районах с коротким летом и длинной зимой из-за их способности удерживать воду зимой. Они также являются одними из самых высоких деревьев, некоторые виды достигают высоты 90 метров!
Хвойные в основном встречаются в северном полушарии.Они процветают в районах с коротким летом и длинной зимой из-за их способности удерживать воду зимой. Они также являются одними из самых высоких деревьев, некоторые виды достигают высоты 90 метров!
Magnoliophyta – цветковые растения
Цветковые растения ( покрытосеменные ) в типе Magnoliophyta являются наиболее развитой, наиболее разнообразной и наиболее успешной группой растений.На самом деле, цветковые растения составляют примерно 90 процентов Царства Plantae! Покрытосеменные — это сосудистые семенные растения с цветками и плодами , в которые заключены семена. Цветковые растения колонизировали почти все мыслимые места обитания на суше — от пустынь до ветреных альпийских вершин, болот и тропических лесов. Цветущие растения включают травы, лилии, розы, кактусы и большинство широколиственных деревьев.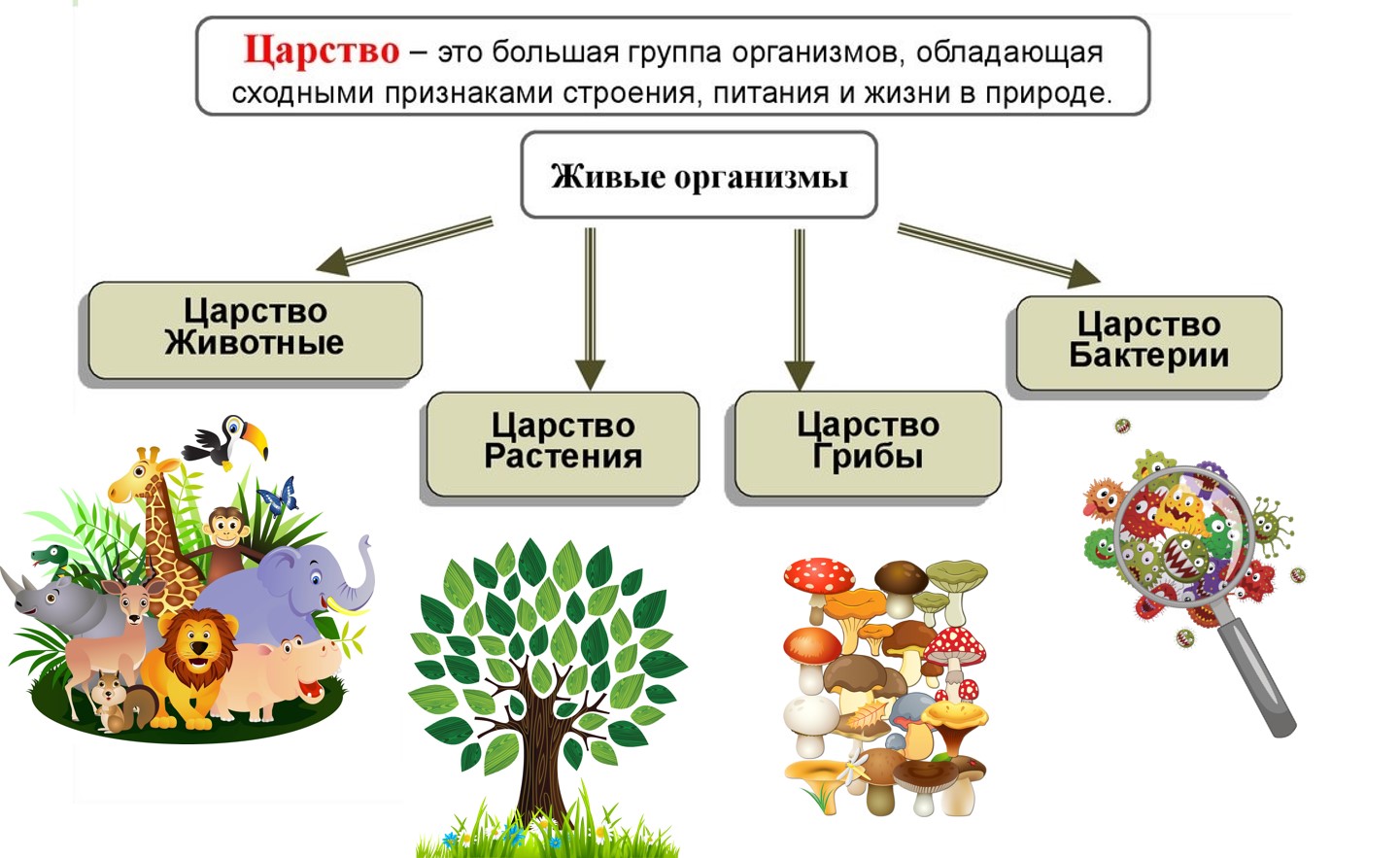
| Anacardiaceae | Melanochyla auriculata Hook.f. | Schopfbaum | Mly | NG (1980) | ||||||||||||||||||
| Annonaceae | Polyalthia Insignis (Cook.f.) Airy Shaw 56 | Schopfbaum | Bor, Phi | Kew & Wong (1988) | ||||||||||||||||||
| Apocynaceae | Calocrater preussii K.Шум. | Корневищный кустарник | CMN, CON, GAB | Lachenaud & Jongkind (2013) | ||||||||||||||||||
| Araceae | Anthurium acutifolium 90 | Наземные или эпифитные | COS, ПАН | хорватов (1991) | ||||||||||||||||||
| ароидных | антуриум аффинное Schott | эфирным | BZC, BZE, BZL | хорватов (1991) | ||||||||||||||||||
| ароидных | Annthurium AnticiCola Liebm | EPIPHYTE | MXC, MXG, MXS, MXT | Corobert (1983) | ||||||||||||||||||
| Araceae | Annthurium Angustilaminatum Engl. | Наземная | БУД | хорватов (1991) | ||||||||||||||||||
| ароидных | антуриум anorianum хорвата | Эпифит | CLM | хорватов, 1991 | ||||||||||||||||||
| ароидных | антуриум asplundii хорвата | Эпифит | ECU | CROB | CROBOT, 1991 | |||||||||||||||||
| Araceae | ||||||||||||||||||||||
| Araceae | ||||||||||||||||||||||
| Annropurpurpureum R.schultes & Maguire | TreneStrical | PERON, BOL, CLM, ECU, PER, BZN | CROBOT, 1991 | |||||||||||||||||||
| ARACEAE | Anthurium banerasense Хорватский | Наземный | ECU | Хорватский и др. , 2013 | ||||||||||||||||||
| Araceae | Anthurium barclayanum Engl. | Эпифит | ECU, PER | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум basirotundum хорват | Эпифит | PER | хорвата, 1991 | ||||||||||||||||||
| ароидных | антуриум becerrae хорват | Эпифит | PER | Хорват и др. , 2013 , 2013 | ||||||||||||||||||
| Araceae | ||||||||||||||||||||||
| Araceae | Annturium Bonplandii Gsbunting | Наземный или EpiPHYTE | FRG, Парень, SUR, VEN, CLM, BZN | CROBOT, 1991 | ||||||||||||||||||
| Araceae | Angurium Bradeanum хорватов и Grayum | эпифит | COS, NIC, PAN | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум bucayanum хорват | Наземные или эпифитные | ECU | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум campii хорвата | Наземные или эпифитные | БУД | хорватов, 1991 | ||||||||||||||||||
| ароидных | антуриум cataniapoense хорвата | эпифит | VEN, | хорватов, 1991 | ||||||||||||||||||
| ароидных | Anthurium caucavalense хорватский | 90 455 ЭпифитЦЛМ | Зона, чел.наблюдать | |||||||||||||||||||
ароидных антуриум Конколор K. Krause Krause | Эпифит PAN, CLM | хорват, 1991 | ||||||||||||||||||||
| ароидных антуриум consobrinum Schott | Эпифит COS, HON, NIC, ПАН | хорватов, 1991 | ||||||||||||||||||||
| ароидных антуриум coriaceum | G.Don Наземные или эпифитные | BZL, BZS | хорватов, 1991 | |||||||||||||||||||
| ароидных антуриум cotobrusii хорвата & Р.A.Baker | Наземные или эпифитные | PAN, COS | хорват, 1991 | |||||||||||||||||||
| ароидных антуриум cowanii хорват | Эпифит GUY | хорват, 1991 | ||||||||||||||||||||
| ароидных антуриум crassinervium (Jacq.) Schott | Наземный или эпифитный | CLM, VEN | Зона, лич. наблюдать.; Croat, 1991 | |||||||||||||||||||
| Araceae | Anthurium crenatum (L.) Кунт | Эпифит | ДОМ, ЛИ, ПУЭ | Зона, чел. наблюдать.; Croat, 1991 наблюдать.; Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium cubense Engl. | Эпифит | MXT, COS, GUA, NIC, PAN, CUB, VEN, CLM | Зона, чел. наблюдать.; CROBOT, 1991 | ||||||||||||||||||
| ARACEAE | ||||||||||||||||||||||
| Anthurium Curtispadix Corobhyte | EPIPHYTE | ECU | CROBOT, 1991 | |||||||||||||||||||
| Araceae | Anthurium Dombeyanum Brongn.ex Schott | Наземные | ECU, PER | Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium eggersii Engl. | Эпифит | ЭКЮ | Зона, чел. наблюдать | ||||||||||||||||||
| Araceae | Anthurium ernestii Engl. | Epiphyte | BOL, CLM, ECU, PER, BZN | Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium eximium Engl. | Эпифит COS | хорват, 1991 | |||||||||||||||||||
ароидных антуриум fatoense K. Krause Krause | Эпифит PAN, COS | хорват, 1991 | ||||||||||||||||||||
| ароидных антуриум fendleri Schott | Эпифит | ПАН, КЛМ, ВЕН | Зона, чел. наблюдать.; Хорват, 1991 | |||||||||||||||||||
| ароидных | антуриум galactospadix хорвата | эпифит | CLM, PER, BZN | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум glaucospadix хорват | сушей или эпифит | CLM , | Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium hacumense Engl. | Эпифитных COS, PAN | ||||||||||||||||||||
| ароидных антуриум halmoorei хорват | Эпифитных MXS | хорват, 1991 | ||||||||||||||||||||
| ароидных антуриум harlingianum хорват | Эпифитных | ЭБУ, CLM | хорватов, 1991 | |||||||||||||||||||
ароидных антуриум harrisii (Грэхэм) G. Don Don | Наземные или эпифитные | BZL хорватов, 1991 | ||||||||||||||||||||
| ароидных антуриум Holmnielsenii CROBIPH | EPIPHYTIC | ECU | CROBOT | CROBOT, 1991 | ||||||||||||||||||
| ARACEAE | Annthurium Hookeri Kunth | EPIPHYTE | NLA, TRT, VNA, Win, Frg, Guy, Sur, Ven | Zona, Ven | чел.наблюдать | |||||||||||||||||
| Araceae | Anthurium imazaense Croat | Epiphyte | PER | Croat et al. , 2013 | ||||||||||||||||||
| Araceae | Anthurium Imperiale Miq. ex Schott | Эпифит | ? | Зона, чел. наблюдать | ||||||||||||||||||
| Araceae | Anthurium Iramirezae Bunting | EpiPHYTE | EPIPHYTE | VEPIPHYTE | CROBOT | |||||||||||||||||
| Araceae | Anthurium Jenmanii Engl. | Наземный или EPIPHYTE | TRT, FRG, GUY, SUR, VEN, BZN | CROBOT, 1991 | ||||||||||||||||||
| Araceae | Anthurium Johnsoniae Crobout | TRORESTRIAL | CLM | CROBOT, 1991 | ||||||||||||||||||
Anthurium latissimum Engl. | Наземные или эпифитные | НА | хорвата, 1991 | |||||||||||||||||||
| ароидных | антуриум lennartii хорвата | Эпифит | БУД | хорватов, 1991 | ||||||||||||||||||
| ароидных | антуриум leonianum Sodiro | Наземные | ECU | Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium linguifolium Engl. | Наземная | ECU | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум llewelynii хорват | Суша | PER | хорвата, 1991 | ||||||||||||||||||
| Araceae | антуриум loretense хорват | Эпифит | НА | хорвата, 1991 | ||||||||||||||||||
| ароидных | антуриума luteynii хорват | наземного или эпифит | ПАН | хорвата, 1991 | ||||||||||||||||||
| ароидных | Антуриума machetioides Matuda | наземных или эпифит | MXS | Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium magrewii Croat | ? | ЭБУ | хорват и др. , 2013 , 2013 | ||||||||||||||||||
| ароидных | антуриум manabianum хорват | Наземная | ECU | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум manuanum хорват | Эпифит | PER | хорвата, 1991 | ||||||||||||||||||
| Araceae | Anthurium martianum K.Koch & Kolb. | Наземный или эпифит | GUY, SUR | Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium napaeum Engl. | Наземная | ECU | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум narinoense хорват | Эпифит | CLM | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум nizandense Matuda | Наземная | MXS | CROBS | CROBOT, 1991 | |||||||||||||||||
| ARACEAE | Angurium ObscuriNervium CrobiT | TRORESTRIAL или EPIPHYTE | ECU | CROBOT, 1991 | ||||||||||||||||||
| Araceae | Anthurium Ottonis K. Krause Krause | Наземные | BOL | Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium oxycarpum Poepp. | Наземные или эпифитные | CLM, ЭБУ, PER, БОЛ, BZN | хорватов, 1991 | ||||||||||||||||||
| ароидных | антуриум oxyphyllum Sodiro | Эпифит | БУД | хорватов, 1991 | ||||||||||||||||||
| ароидных | антуриум pachylaminum хорват | Эпифит | PER | хорвата, 1991 | ||||||||||||||||||
| ароидных | антуриум palenquense хорват | Наземная | ECU | хорвата, 1991 | ||||||||||||||||||
| ароидных | антуриум pallatangense англ. | Наземный или EPIPHYTE | ECU | CROBOT, 1991 | ||||||||||||||||||
| ARACEAE | Annthurium Paloraense Corobhte | EPIPHYTE | ECU | Croat et al. , 2013 | ||||||||||||||||||
| Araceae | Anthurium paraguayense Engl. | Наземный или EPIPHYTE | BOL, PER, BZC, AGE, PAR | CROBOT, 1991 | ||||||||||||||||||
| Araceae | Annthurium Punning Crobout | Trustrestrical | ECU, за | Corob, 1991 | ||||||||||||||||||
| Araceae | Anthurium plowmanii Croat | Epiphyte | BOL, PER, BZC, BZN, PAR | Зона, чел.наблюдать.; Хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум pranceanum хорват | Наземная | BZN | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум purpureospathum хорват | Наземные или эпифитные | PAN | хорват, 1991 | ||||||||||||||||||
| Araceae | Anthurium reflexinervium Croat | Epiphyte | PER | Zona, перс.наблюдать.; Хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум remotigeniculatum хорват | Наземная | PER | хорвата, 1991 | ||||||||||||||||||
| Araceae | антуриум salvadorense хорват | Суша или эпифит | ELS, GUA | Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium salvinii Hemsl. | Epiphyte | MXT, COS, GUA, HON, NIC, PAN, CLM | Зона, чел.наблюдать.; Хорватов, 1991 | ||||||||||||||||||
| ароидных | антуриум santiagoense хорватские | Наземные | ECU | хорвата, 1991 | ||||||||||||||||||
| ароидных | антуриум sarukhanianum хорвата & Haager | Эпифит | MXS, MXT | Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium schlechtendalii Kunth | Epiphyte | MXG, MXS, MXT, BLZ, GUA, HON, NIC 5наблюдать.; Хорватов, 1991 | |||||||||||||||||||
| ароидных антуриум simpsonii | хорвата Эпифит | НА хорватов, 1991 | ||||||||||||||||||||
| ароидных антуриум solitarium (Vell.Conc.) Шотт | Эпифит BZC , BZE, BZL | хорват, 1991 | ||||||||||||||||||||
| ароидных | антуриум solomonii хорват | Эпифит | BOL | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум soukupii хорват | Наземная | BOL , ECU | CROBOT, 1991 | ||||||||||||||||||
| ARACEAE | Anthurium Sparreorum Crobibhyte | EPIPHYTE | ECU | CROBOT, 1991 | ||||||||||||||||||
| Araceae | Annthurium Spathiphylum N. Э.Бр. Э.Бр. | EPIPHYTE | NIC, COS, CON | CROB | ||||||||||||||||||
| Araceae | Superbum Madison | EPIPHYTE | ECU | ZONA, PES. наблюдать.; Croat, 1991 | ||||||||||||||||||
| Araceae | Anthurium tarapotense Engl. | Наземная | PER | хорвата, 1991 | ||||||||||||||||||
| Araceae | антуриум tenaense хорват | эпифит | ECU | хорват, 1991 | ||||||||||||||||||
| ароидных | антуриум trujilloi хорват | Эпифит | CLM | Croat и др., 2013 | ||||||||||||||||||
| ароидных антуриум upalaense хорватов и RABaker | Эпифит COS, NIC | хорват, 1991 | ||||||||||||||||||||
| ароидных антуриум validifolium K.Krause | Эпифит Pan, Cos | Corob | Corober, 1991 | |||||||||||||||||||
| Araceae | Annturium Vaupesianum CROBHT | EPIPHYTE | CLM | CLM | CROBOT, 1991 | |||||||||||||||||
| Araceae | Anthurium Venosum Griseb. | Эпифит | КУБ, ДЖЕМ | Зона, чел. наблюдать.; Хорватов, 1991 | ||||||||||||||||||
| ароидных антуриум vinillense Бантинг | Наземная VEN | хорватов, 1991 | ||||||||||||||||||||
| ароидных антуриум wagenerianum K.Koch & BOUCHE | Наземная НОА, VEN | CROBOT, 1991 | ||||||||||||||||||||
| ARACEAE | ARACEAE | Annthurium Willifordii Corob | | PER | Corob | | Aracae | Culcasia dinklagei Engl. | Herb | GUI, IVO, LBR, CAF, CMN, CON, GAB, ZAI | Lachenaud & Jongkind, 2013 | Araceae | Cul. & K.kruse | Herb | CMN, GAB | CMN, GAB | Lachenaud & Jongkind, 2013 | Araceae | PhiloDendron Acutifolium K.kruse | Trenshial или EpiPHYTE | ECU, PER | ZONA, ЧАСТЬ. |  наблюдать наблюдать |
| Araceae | PhiloDendron Insigne SCHOTT | EPIPHYTE | FRG, GUE, SUR, VEN, CLM, ECU, PER, BZE, BZL, BZN | Gibernau et al. , 2007 | ||||||||||||||||||
| Araceae | Philodendron martianum Engl. | Эпифит | БЗЭ, БЗЛ, БЗС | Зона, чел. наблюдать | ||||||||||||||||||
| Araceae | Philodendron melinonii Brongn.Ex Regel | EPIPHYTE | EPIPHYTE | FRG, Парень, SUR, VEN, BZN | Endroid | |||||||||||||||||
| Araceae | Pichinia DISTICHA S.Y.WONG & P.PIPHYTE | EPIPHYTE | BOR | BOYCE et al. , 2010 | ||||||||||||||||||
| Araceae | ||||||||||||||||||||||
| Araceae | Rhaphidophora Angustata SCHOTT | MLY | Mly, сумма | Boyce, 1999 | ||||||||||||||||||
| Araceae | Scindapsus Beccarii Engl. | Epiphyte | BOR | Boyce, 1995 | ||||||||||||||||||
| Araceae | Scindapsus geniculatus Engl. | Эпифит | БОР | Бойс и др. , 2010 | ||||||||||||||||||
| Araceae | Spathiphyllum wendlandii Schott | Розетка травы | COS | Weisshofen , 2008 | ||||||||||||||||||
| Arecaceae | Asterogyne guianensis Granv.и Эй Джей Хенд. | Palm | FRG | de Granville & Henderson, 1988 | ||||||||||||||||||
| Arecaceae | Asterogyne martiana (H.Wendl.) H.Wendl. ex Drude | Palm | BLZ, COS, GUA, HON, NIC, PAN, CLM, ECU | Raich, 1983; Weissenhofer и др. , 2008 г.; Hübner, 2013 | ||||||||||||||||||
| Arecaceae | Asterogyne Ramosa (H.E.moore) WESS.BOER (H.E.MOORE) Palm | Palm | Palm | English | ||||||||||||||||||
| Arecaceae | Asterogyne Spicata (H.E.Moore) Wess.Boer | Palm | VEN | Inferred | ||||||||||||||||||
| Arecaceae | Asterogyne yaracuyense A. J.Hend. и Стейерм. J.Hend. и Стейерм. | Palm | VEN | Inferred | ||||||||||||||||||
| Arecaceae | Astrocarium acaule Mart. | Palm | GUY, VEN, CLM, BZN | Предполагается; см. Kahn, 2008 | ||||||||||||||||||
| Arecaceae | Astrocarium ciliatum F.Kahn & B.Millan | Palm | CLM, PER, BZN | Предположительно; см. Kahn, 2008 | ||||||||||||||||||
| Arecaceae | Astrocaryum farinosum Barb.Rodr. | Ладонь | GUY, SUR, BZN | Предположительно; см. Kahn, 2008 | ||||||||||||||||||
| Arecaceae | Astrocaryum giganteum Barb.Rodr. | Ладонь | БЗН | Предположительно; см. Kahn, 2008 | ||||||||||||||||||
| Arecaceae | Astrocaryum javarense (Trail) Drude | Palm | PER, BZN | Inferred; см. Kahn, 2008 | ||||||||||||||||||
| Arecaceae | Astrocarium malybo H.Крас. | Palm | CLM | Предполагается; см. Kahn, 2008 Kahn, 2008 | ||||||||||||||||||
| Arecaceae | Astrocarium mexicanum Liebm. бывший Март. | Palm | MXG, MXS, MXT, BLZ, ELS, GUA, HON, NIC | Alvarez-Sánchez & Guervara, 1999 | ||||||||||||||||||
| Arecaceae | Astrocary Minus Trail | Palm | FRG, BZN | предполагаемый; см. Kahn, 2008 | ||||||||||||||||||
| Arecaceae | Astrocaryum paramaca Mart. | Palm | Palm | FRG, SUR, BZN | De Granville, 1977 | |||||||||||||||||
| Arecaceae | Astrocaryum Sciophilum (Miq.) Palle | Palm | FRG, парень, сюр, BZN | De Granville, 1977 | ||||||||||||||||||
| Arecaceae | Astrocaryum sociale Barb.Rodr. | Пальма | BZN | de Vasconcelos, 1990 | ||||||||||||||||||
| Arecaceae | Attalea microcarpa Mart. | Palm | FRG, GUY, SUR, VEN, CLM, PER, BZN | de Granville, 1992 | ||||||||||||||||||
| Arecaceae | Attalea spectabilis | Пальма | BZN | de Vasconcelos, 1990 | ||||||||||||||||||
| Arecaceae | Calamus geniculatus Griff. | Палм | ТХА, МЛЙ, СУММ | Зона, чел. наблюдать | ||||||||||||||||||
| Arecaceae | Calamus macrophyllus (Becc.) W.J.Baker | Palm | THA, MLY | Rickson & Rickson, 1986 | ||||||||||||||||||
| Arecaceae | Calamus verticillaris ris. | Palm | THA, MLY, SUM | Rickson & Rickson, 1986 | ||||||||||||||||||
| Arecaceae | Calyptrogyne ghiesbreghtiana 9000 (H.Wendl.Wendl.L.L. | Palm | MXG, MXT, BLZ, COS, GUA, HON, NIC, PAN | Weissenhofer и др., 2008 | ||||||||||||||||||
| Arecaceae | Calyptrogyne Kunorum Denevers | Palm | PAN | De Nevers, 1995 | ||||||||||||||||||
| Arecaceae | Chamaedorea Enernesti-Augusti H.Wendl. | Palm | MXG, MXS, MXT, BLZ, GUA, HON | Зона, чел. наблюдать | ||||||||||||||||||
| Arecaceae | Chamaedorea oreophila Mart. | Пальма | MXG, MXS | Hodel, 1992 | ||||||||||||||||||
| Arecaceae | Chamaedorea Rigida H.Вендл. | Палм | MXS | Hodel, 1992 | ||||||||||||||||||
| Arecaceae | Dypsis marojejyi Beentje | Палм | MDG | Dransfield & Beentje, 1995 | ||||||||||||||||||
| Arecaceae | Dypsis perrieri (Юм. ) Beentje & J.Dransf. | Palm | MDG | Dransfield & Beentje, 1995 | ||||||||||||||||||
| Arecaceae | Dypsis turkii J.Трансф. | Palm | MDG | MDG | Dransfield, 2003 | |||||||||||||||||
| Arecaceae | Arecaceae | ELAEIS OLEIFERA (Kunth) Cortés | Palm | Palm | COS, HON, NIC, PAN, FRG, SUR, CLM, ECU, PER, BZN | Weissenhofer и др. , 2008 | , 2008||||||||||||||||
| Arecaceae | Eleidoxa Conferta (GRIFF.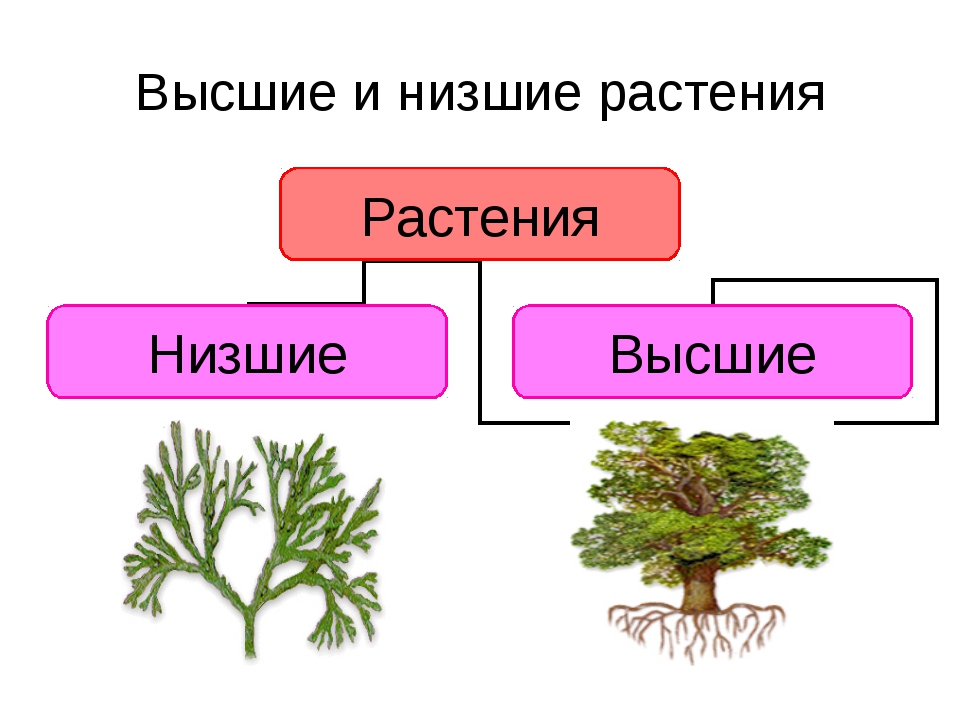 ) Brett ) Brett | Palm | Tha, Bor, Mly, сумма | Выбор | ||||||||||||||||||
| Arecaceae | Eugeissona Brachystachys Ridl. | Palm | MLY | Inferred | ||||||||||||||||||
| Arecaceae | Eugeissona insignis Becc. | Palm | BOR | Inferred | ||||||||||||||||||
| Arecaceae | Eugeissona minor Becc. | Palm | BOR | Holbrook и др. , 1985 | ||||||||||||||||||
| Arecaceae | Eugeissona tristis Griff. | Пальмовый | THA, MLY | Предполагаемый | ||||||||||||||||||
| Arecaceae | Eugeissona utilis Becc. | Palm | BOR | Inferred | ||||||||||||||||||
| Arecaceae | Geonoma brongniartii Mart. | Palm | VEN, BOL, CLM, ECU, PER, BZN | J. Roncal, чел. ком. | ||||||||||||||||||
| Arecaceae | Geonoma calyptrogynoidea Burret | Palm | PAN, CLM, ECU | J.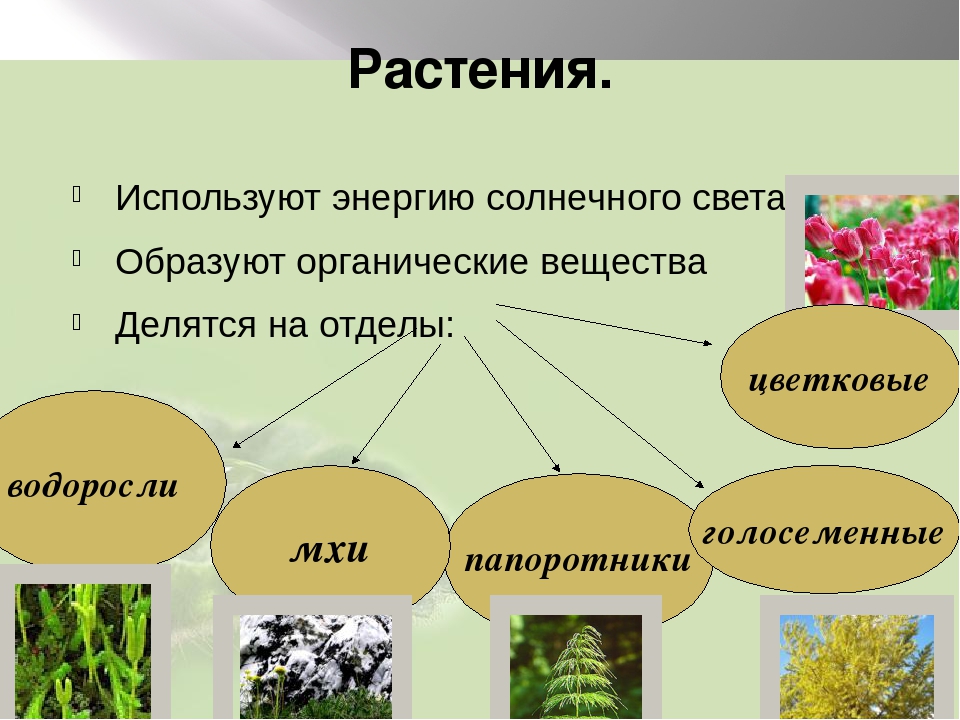 Roncal, перс. ком. Roncal, перс. ком. | ||||||||||||||||||
| Arecaceae | Geonoma camana Trail | Palm | CLM, ECU, PER, BZN | J.Ронкаль, перс. ком. | ||||||||||||||||||
| Arecaceae | Geonoma chlamydostachys Galeano | Palm | CLM | J. Roncal, перс. ком. | ||||||||||||||||||
| Arecaceae | Geonoma cuneata H.Wendl. ex Spruce | Palm | COS, NIC, PAN, VEN, CLM, ECU | Weissenhofer и др. , 2008 | ||||||||||||||||||
| Arecaceae | Geonoma epetiolata H.E.Moore | Palm | COS, PAN, | 9045наблюдать.; Blanco & Martén-Rodríguez, 2007|||||||||||||||||||
| Arecaceae | Geonoma frontinensis Burret | Palm | CLM | J. ком. | ||||||||||||||||||
| Arecaceae | Geonoma interrupta (Ruiz & Pav.) Mart. | Palm | MXG, MXS, MXT, BLZ, COS, GUA, HON, NIC, PAN, VEN, BOL, CLM, ECU, PER | J. Roncal, перс. ком. Roncal, перс. ком. | ||||||||||||||||||
| Arecaceae | Geonoma macrostachys Mart. | Палм | ВЕН, БОЛ, КЛМ, ЭБУ, ПЕР, БЗН | Зона, чел. наблюдать | ||||||||||||||||||
| Arecaceae | Geonoma oldemanii Granv. | Пальма | ФРГ, БЗН | предполагаемая | ||||||||||||||||||
| Arecaceae | Geonoma poeppigiana Mart. | Palm | CLM, ECU, PER, BZN | J. Roncal, перс. ком. | ||||||||||||||||||
| Arecaceae | Geonoma triglochin Burret | Palm | BOL, CLM, ECU, PER | J.Ронкаль, перс. ком. | ||||||||||||||||||
| Arecaceae | Geonoma undata Klotzsch | Palm | MXT, COS, GUA, HON, NIC, PANCU LEE, WIN, FRG, GUY, CLMPER, BOLUR, SOLUR, VEN | Ж. Ронкаль, чел. ком. | ||||||||||||||||||
| Arecaceae | Heterospathe brevicaulis Fernando | Пальма | PHI | Предполагается; См. Fernando, 1990 Fernando, 1990 | ||||||||||||||||||
| Arecaceae | гетероприосэсфер | Palm | PHI | Fernando & Sotalbo, 2001 | ||||||||||||||||||
| Arecaceae | Iguanura Wallichiana (MART.) Бент. & Крюк.ф. бывший Бек. | Palm | Tha, Mly, Sum | Kew & Wong, 1988 | ||||||||||||||||||
| Arecaceae | Arecaceae | Johannesteiksmannia Altifrons (RCHB.F. & ZOLL.) HEMOOORE | Palm | Tha, Bor, Mly, СУММА | Зона, чел. наблюдать | |||||||||||||||||
| Arecaceae | Johannesteijsmannia lanceolata J.Dransf. | Palm | MLY | Inferred | ||||||||||||||||||
| Arecaceae | Johannesteijsmannia magnifica J.Трансф. | Palm | MLY | Inferred | ||||||||||||||||||
| Arecaceae | Johannesteijsmannia perakensis J.Dransf. | Palm | MLY | Inferred | ||||||||||||||||||
| Arecaceae | Licuala acutifida Mart. | Palm | MLY | Предполагается; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala ahlidurii Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala atrovirens Saw | Palm | BOR | Предполагается; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala Bayana Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala bidentata Becc. | Ладонь | БОР | Предположительно; Смотреть, 2012 | ||||||||||||||||||
| Arecaceae | Licala Bifida Heatubun & Barfod | PalmNWG | Heatubun & Barfod, 2008 | |||||||||||||||||||
| Arecaceae | Licala Bintulensis BECC. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala Borneensis Becc.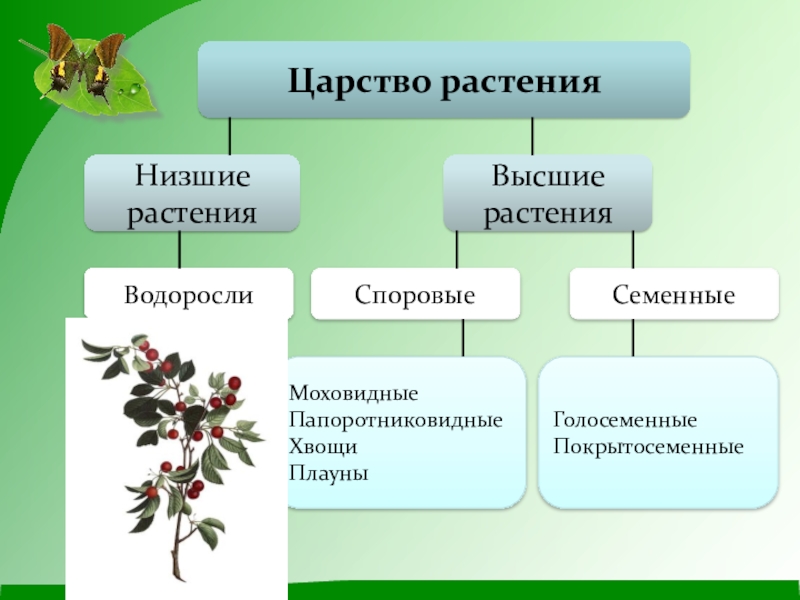 | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala bruneiana Saw | Palm | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala burretii Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala cameranensis Пила | Пальма | MLY | Предполагается; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala campestris Saw | Palm | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala chaiana Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala chaiana Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala cordata Becc. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala Corni Furtado | Palm | MLY | Zona, pers. наблюдать | ||||||||||||||||||
| Arecaceae | Licuala densiflora Becc. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala egregia Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala elliptica Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala exigua Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala ferruginea Becc. | Palm | MLY | Предполагается; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala flammula Пила | Пальма | BOR | Предполагается; см. Saw, 2012 Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala fractiflexa Saw | Palm | MLY | Zona, pers. наблюдать | ||||||||||||||||||
| Arecaceae | Licuala glabra Griff. | Palm | THA, BOR, MLY | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala hallieriana Becc. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala intermedia Saw | Palm | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala kamarudinii Пила | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala kemamanensis Furtado | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala khoonmengii Saw | Palm | MLY | Inferred; см. Saw, 1997 Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala kiahii Furtado | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala kingiana Becc. | Palm | MLY | Предполагается; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala kuchingensis Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala kunstleri Becc. | Palm | THA, MLY | Предположительно; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala lambii Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala lanata J.Трансф. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala lanuginosa Ridl. | Palm | MLY | Предполагается; см.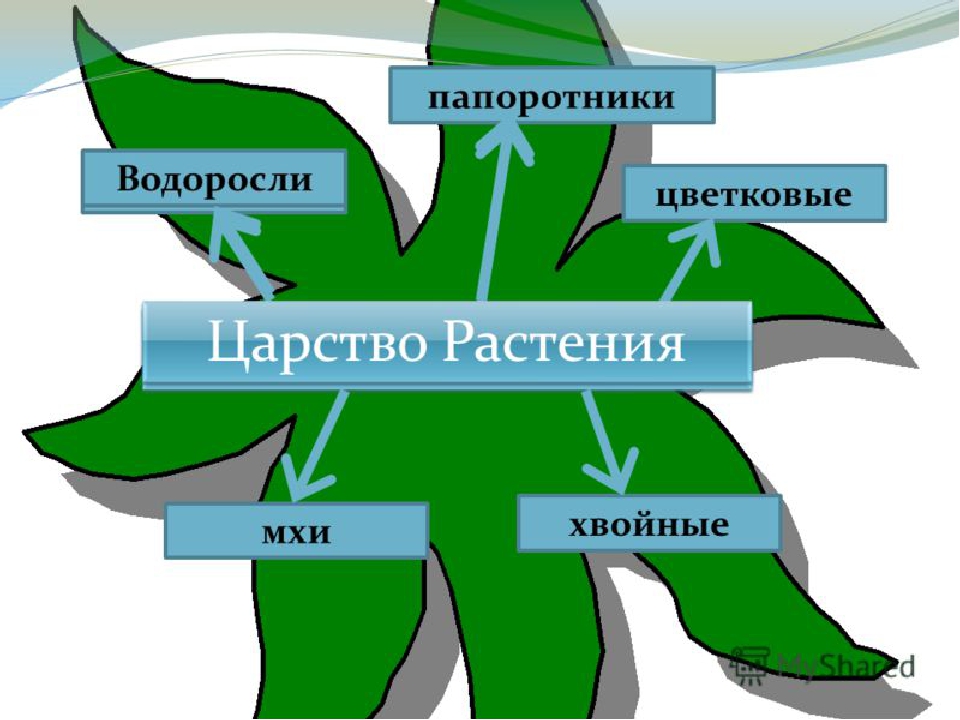 Saw, 1997 Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala leopoldii Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala leucocarpa Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala longicalycata Furtado | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala longipes Griff. | Palm | MLY, SUM | Предполагаемый; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala maculata Saw | Palm | BOR | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala malajana Becc. | Palm | THA, MLY | Предположительно; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala mattanensis Becc. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala meijeri Saw | Palm | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala micholitzii Ridl. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala mirabilis Furtado | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala miriensis Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala Modesta Becc. | Palm | THA, MLY | Предположительно; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala moyseyi Furtado | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala mukahensis Пила | Пальма | BOR | Предполагается; см. Saw, 2012 Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala mustapana Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala orbicularis Becc. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala oryzoides Saw | Palm | BOR | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala pahangensis Furtado | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala palas Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala paludosa Griff. | Palm | AND, CBD, THA, VIE, BOR, MLY, SUM | Предполагаемый; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala patens Ridl. | Palm | MLY | Предполагается; см.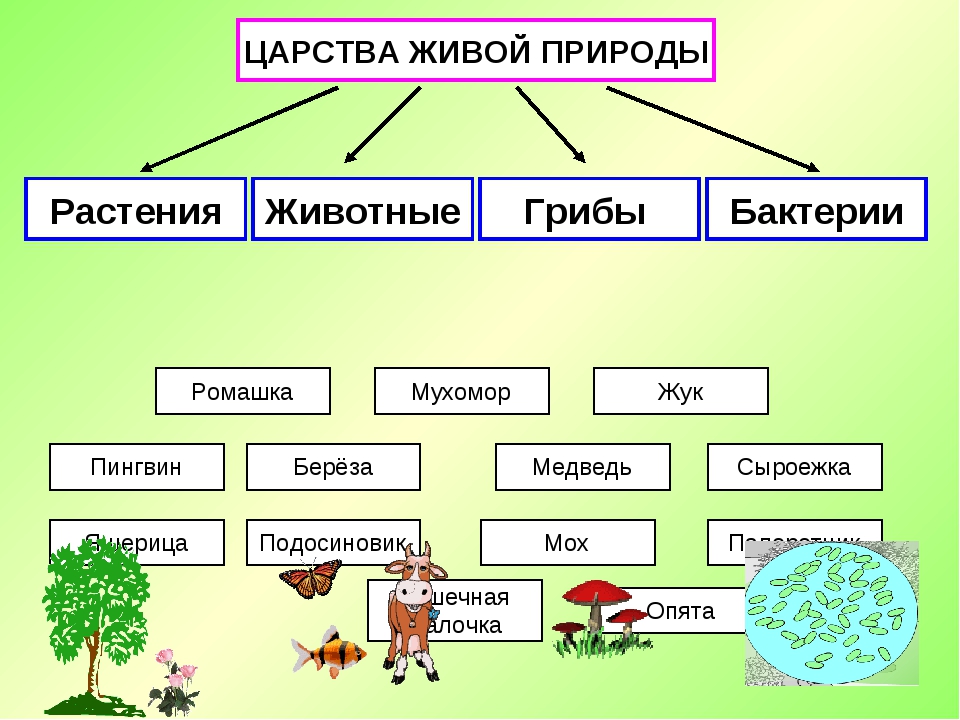 Saw, 1997 Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala petiolulata Becc. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala pilosa Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala pseudovalida Saw | Palm | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala punctulata Burret | Palm | BOR | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala pusilla Becc. | Palm | THA, MLY | Предположительно; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala reptans Becc | Palm | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala ridleyana Becc. | Palm | MLY | Предполагается; см. Saw, 1997 Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala rubiginosa Пила | Пальма | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala ruthiae Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala sabahana Saw | Palm | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala sallehana Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala sar wakesis Becc. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala scortechinii Becc. | Palm | MLY | Предполагается; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala spathellifera Becc. | Ладонь | БОР | Предположительно; см. Saw, 2012 Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala spicata Becc. | Ладонь | БОР | Предположительно; Смотри, 2012 | ||||||||||||||||||
| Arecaceae | Licuala Spinosa Wurmb | Palm | Palm | ЧH, запрет и CBD, MYA, NCB, THA, VIE, BOR, JAW, MLY, PHI, SUM | см. Saw, 2012 | |||||||||||||||||
| Arecaceae | Licuala stongensis Пила | Пальма | MLY | Предполагается; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala tenuissima Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala terengganuensis Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala thoana Saw & J.Трансф. | Palm | MLY | Предполагается; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala tiomanensis Furtado | Palm | MLY | Inferred; см. Saw, 1997 Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala triphylla Griff. | Palm | THA, BOR, MLY, SUM | Предполагаемый; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala valida Becc. | Ладонь | БОР | Предположительно; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Licuala whitmorei Saw | Palm | MLY | Inferred; см. Saw, 1997 | ||||||||||||||||||
| Arecaceae | Licuala yiiana Saw | Palm | BOR | Предполагается; см. Saw, 2012 | ||||||||||||||||||
| Arecaceae | Marojejya darianii J.Dransf. и Н.W.Uhl | Палм MDG | Dransfield & Beentje, 1995 | |||||||||||||||||||
| Arecaceae марудзезия замечательная Гумберта | Вербное ЦРТ | Dransfield & Beentje, 1995 | ||||||||||||||||||||
| Arecaceae Masoala kona Beentje | Palm | MDG | Dransfield & Beentje, 1995 | |||||||||||||||||||
| Arecaceae | Masoala madagascariensis | Palm | MDG | Dransfield & Beentje, 1995 | ||||||||||||||||||
| Arecaceae | Oenocarpus mapora H. Karst. Karst. | Palm | COS, PAN, VEN, BOL, CLM, ECU, PER, BZN | Weissenhofer и др. , 2008 | , 2008||||||||||||||||||
| Arecaceae | Phytelephas SEANANY O.F.COOK | Palm | Palm | Palm | Bernal & Balslev, 1996 | |||||||||||||||||
| Arecaceae | Prestoea Decurrens (H.Вендл. ex Burret) H.E.Moore | Palm | COS, NIC, PAN, CLM, ECU | Weissenhofer и др. , 2008 | ||||||||||||||||||
| Arecaceae | Raphia taedigera (Mart.) Mart. | Palm | NGA, CMN, COS, NIC, PAN, CLM, BZN | Weissenhofer и др. , 2008 | ||||||||||||||||||
| Arecaceae | Ravenea betjei Rakotoarin. & J.Dransf. | Palm | MDG | Rakotoarinivo & Dransfield, 2010 | ||||||||||||||||||
| Arecaceae | Ravenea declivium J.Трансф. & Ракотоарин. | Палм | MDG | Dransfield & Rakotoarinivo, 2012 | ||||||||||||||||||
| Arecaceae | Ravenea dransfieldii Beentje | Вербное | ЦРТ | Dransfield & Beentje, 1995 | ||||||||||||||||||
| Arecaceae | Ravenea hypoleuca Rakotoarin . & J.Dransf. & J.Dransf. | Палм | MDG | Rakotoarinivo & Dransfield, 2010 | ||||||||||||||||||
| Arecaceae | Ravenea louvelii Beentje | Вербное | ЦРТ | Dransfield & Beentje, 1995 | ||||||||||||||||||
| Arecaceae | Sabinaria величественная Galeano & Bernal | Palm | Palm | CLM | CLM | Galeano & Bernal, 2013 | ||||||||||||||||
| Arecaceae | Salacca Aceensis Mogea & Zumaidar | Palm | Sum, | . см. Zumaidar et al. , 2014 | ||||||||||||||||||
| Arecaceae | Salacca Bakeriana J.Dransf. | Ладонь | БОР | Предположительно; см. Dransfield, 2009 | ||||||||||||||||||
| Arecaceae | Salacca clemensiana Becc. | Palm | BOR, PHI | Предполагается; см. Mogea, 1984 | ||||||||||||||||||
| Arecaceae | Salacca dransfieldiana Mogea | Пальма | BOR | Предполагается; см.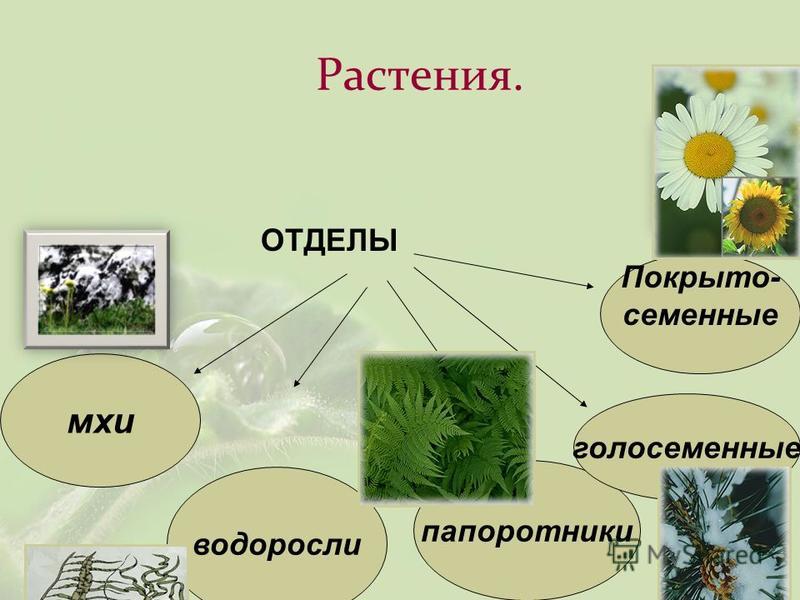 Mogea, 1980, 1984 Mogea, 1980, 1984 | ||||||||||||||||||
| Arecaceae | Salacca flabellata Furtado | Palm | MLY | Inferred; см. Mogea, 1980, 1984 | ||||||||||||||||||
| Arecaceae | Salacca magnifica Mogea | Palm | BOR | Предполагается; см. Mogea, 1980, 1984 | ||||||||||||||||||
| Arecaceae | Salacca minuta Mogea | Пальма | MLY | Предполагается; см. Mogea, 1984 | ||||||||||||||||||
| Arecaceae | Salacca multiflora Mogea | Пальма | MLY | Предполагается; см. Mogea, 1984 | ||||||||||||||||||
| Arecaceae | Salacca sar wakesis Mogea | Пальма | BOR | Предполагается; см. Mogea, 1980 | ||||||||||||||||||
| Arecaceae | Sclerosperma mannii H.Вендл. | Palm | LBR, NGA, CAB, CMN, CON, EQG, GAB, GGI, ZAI, ANG | van Valkenburg и др. , 2008 | ||||||||||||||||||
| Arecaceae | Sclerosperma profiziana Valk. & Сандер. & Сандер. | Palm | GHA, NGA?, CON, ZAI, ANG | van Valkenburg и др. , 2008 | ||||||||||||||||||
| Arecaceae | Sclerosperma walkeri A.Chev. | Palm | GAB, ZAI | van Valkenburg и др. , 2008 | ||||||||||||||||||
| Arecaceae | Sommieria leucophylla Becc. | Пальма | NWG | Heatubun, 2002 | ||||||||||||||||||
| Arecaceae | Welfia regia H.Wendl. | Palm | COS, HON, NIC, PAN, CLM, ECU | Weissenhofer и др. , 2008 | ||||||||||||||||||
| Asparagaceae | Dracaena adamii Hepper | Schopfbaum | GHA, IVO, LBR | 2 Hawthorne | 6 | Lachenaud & Jongkind, 2013|||||||||||||||||
| Aspleniaceae | Asplenium antiquum Makino | Эпифит | CHC, CHS, JAP, KOR, TAI 904; см. Murakami et al., 1999 | |||||||||||||||||||
| Aspleniaceae асплениум antrophyoides Христос | литофит CHC, CHS, ЛАО, ТНА, VIE | Holttum, 1974 | ||||||||||||||||||||
Aspleniaceae КОСТЕНЕЦ ЮЖНОАЗИАТСКИЙ (J. Sm. ) Крюк. Sm. ) Крюк. | Эпифит | QLD | Christenhusz, чел. наблюдать | |||||||||||||||||||
| Aspleniaceae | Asplenium carinatum Alderw. | литофит | СУП | Holttum, 1974 | ||||||||||||||||||
| Aspleniaceae | Asplenium colubrinum Христос | Эпифит | PHI | Holttum, 1974 | ||||||||||||||||||
| Aspleniaceae | Asplenium cymbifolium Христос | Эпифит | сумма, BOR, PHI, NWG, SAM | Holttum, 1974 | ||||||||||||||||||
| Aspleniaceae | Asplenium dregeanum Kunze | EpiPHYTE | GHA, GUI, NGA, BUR, CMN, EQG, GAB, KEN, TAN, UGA, MLW, ZIM, MAU, MDG | Pócs, 1980 | ||||||||||||||||||
| Aspleniaceae | Asplenium goudeyi D.L.Jones | Эпифит | NFK | Предполагаемый; см. Murakami et al. , 1999 | ||||||||||||||||||
| Aspleniaceae | Asplenium grevillei Стенка. бывший Крюк. & Грев. бывший Крюк. & Грев. | Эпифит | IND, SRL, VIE | Holttum, 1974 | ||||||||||||||||||
| Aspleniaceae | Asplenium holstii Hieron. | Эпифит | IVO, CON, ZAI, KEN, TAN, UGA, MLW, MOZ, ZIM | Christenhusz, чел.наблюдать | ||||||||||||||||||
| Aspleniaceae | Aspleenium Humbertii Tardieu | EPIPHYTE | CHC, ЧH, VIE | Holttum, 1974 | ||||||||||||||||||
| Aspleniaceae | Asplenium Musifolium Messt. | EpiPHYTE | PHI | PHI | Copeland, 1907 | |||||||||||||||||
| Aspleniaceae | Asplenium NIDUS L. | EPIPHYTE | MDG, CHC, CHS, ЧH, JAP, TAI, IND, SRL, MYA, THA, Лаос , VIE, MYA, QLD, | Zona & Christenhusz, чел.наблюдать | ||||||||||||||||||
| Aspleniaceae Asplenium pacificum Holttum | Эпифит — | Holttum, 1974 | ||||||||||||||||||||
Aspleniaceae Asplenium phyllitidis D. Don Don | эпифит CHS, CHC, СНН, BAN, IND, EHM, NEP, MYA, THA, MLY, JAW, NWG | Copeland, 1907; Holttum, 1974 | ||||||||||||||||||||
| Aspleniaceae | Asplenium Serratum L. | EpiPHYTE | JAW, NWC, FLA, MXG, MXS, MXT, BLZ, GUA, HON, NIC, PAN, COS, CUB, DOM, JAM , LEE, TRT, CLM, FRG, GUY, SUR, VEN, ECU, PER, BOL, BZC, BZL, BZN, BZS, AGE, PAR | Zuquim и др., 2008 г.; Зона, чел. наблюдать | ||||||||||||||||||
| Aspleniaceae | Asplenium setoi N.Murak. & Сериз. | Эпифит | NNS, OGA | Предполагаемый; см. Murakami et al. , 1999 | ||||||||||||||||||
| Aspleniaceae | Asplenium simonsianum Hook. | Эпифит | ASS, BAN, EHM | Holttum, 1974 | ||||||||||||||||||
| Aspleniaceae | Asplenium stuebelianum Hieron. | Epiphyte | BOL, CLM, ECU, PER, BZN, PAR | Zuquim и др. , 2008 | ||||||||||||||||||
| Aspleniaceae | Blechnum discolor Keyserl. | Розетка травы | NZN, NZS, NSW, SOA | Dearden & Wardle, 2008 | ||||||||||||||||||
| Asteliaceae | Collospermum hastatum (Collospermum hastatum) | Эпифит | NZN, NZS | Derraik, 2005 | ||||||||||||||||||
| Bignoniaceae | Colea resupinata M.Л.Зжхра | Шопфбаум | ЦРТ | Предположительно; см. Zjhra, 2006 | ||||||||||||||||||
| Bignoniaceae | Colea rosea M.L.Zjhra | Schopfbaum | ЦРТ | Предположительно; см. Zjhra, 2006 | ||||||||||||||||||
| Bignoniaceae | Colea sytsmae M.L.Zjhra | Schopfbaum | ЦРТ | 904; см. Zjhra, 2006|||||||||||||||||||
| Bignoniaceae | Colea unifoliolata Callm.& Phillipson | Schopfbaum | MDG | Inferred | ||||||||||||||||||
| Bignoniaceae | Rhodocolea magnifica Callm. & Филлипсон | Schopfbaum ЦРТ | ||||||||||||||||||||
| калофилловые маммея megaphylla (Leroy) PFStevens | Schopfbaum ЦРТ | Стивенс, 2005 | ||||||||||||||||||||
калофилловые маммея zeereae Р . F.stevens F.stevens | Schopfbaum | MDG | MDG | STEVENS, 2005 | ||||||||||||||||||
| Capparaceae | Quadrella Antonensis (Woodson) Ildis & Cornejo | разветвленные Schopfbaum | Pan | Dressler, 1985; Iltis & Cornejo, 2010 | ||||||||||||||||||
| Capparaceae | Quadrella dressleri Cornejo & Iltis | Schopfbaum | PAN | Dressler; Cornejo & Iiltis, 2010|||||||||||||||||||
| CyasteaceeCeae | Cyathea Choricarpa (Maxon) Drom | розетка Herb | COS, NIC, PAN, CLM, ECU | Kaczmarczyk, 2002 | ||||||||||||||||||
| Cyclanthaceae | Asplundia Alata Harling | Розетка травы | COS, PAN, CLM, ECU, PER | Weissenhofer et al., 2008 | ||||||||||||||||||
| Cyclanthaceae | Asplundia albicarpa Hammel | Наземный или эпифитный | COS, PAN, CLM, 5 Zona 5 per. наблюдать | |||||||||||||||||||
| Cyclanthaceae | Asplundia brachyphylla Харлинг | Розетка | ФРГ, ГАЙ, ЮАР | Зона, чел.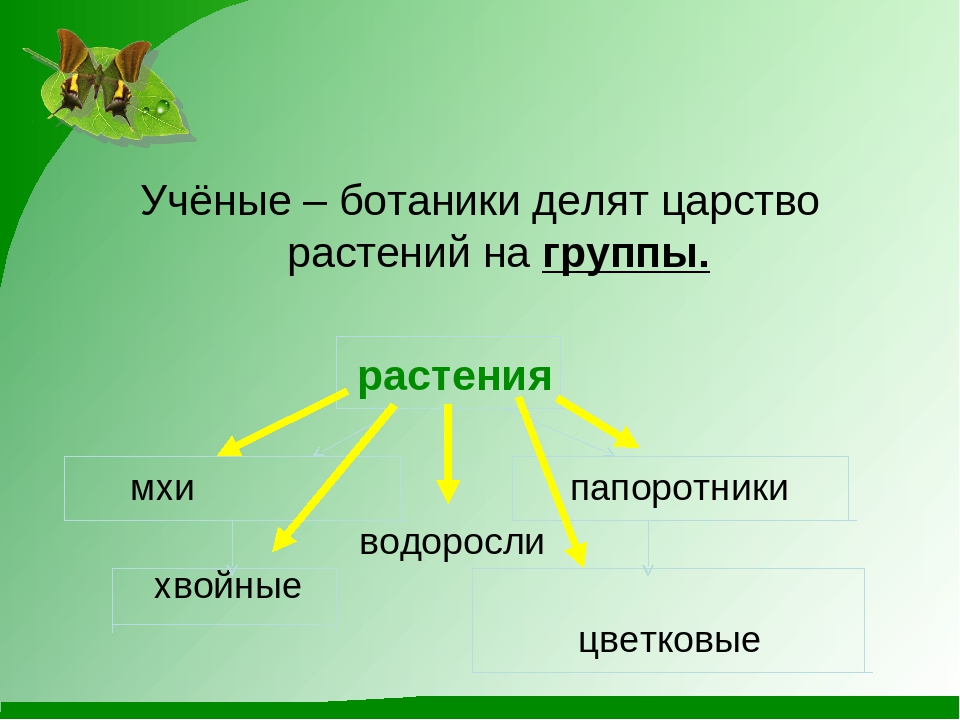 наблюдать наблюдать | ||||||||||||||||||
| Cyclanthaceae | Asplundia pittieri (Woodson) Harling | Розетка травы | COS, PAN, CLM | Weiss05hofer 9 0al.05hofer 0al. , 2008 | ||||||||||||||||||
| Cyclanthaceae | Ludovia lancifolia Brongn. | EPIPHYTE | PAN, FRG, GUY, SUR, VEN, CLM, ECU, PER, BZN | Freiberg, 1999 | ||||||||||||||||||
| Sphaeradenia AcutitePala Harling | розетка Harb | COS, NIC, PAN , CLM | Weissenhofer и др. , 2008 | |||||||||||||||||||
| Cyperaceae | Becquerelia cymosa Brongn. | розетка Herb | COS, NIC, PAN, TRT, FRG, GUE, SUR, VEN, CLM, ECU, PER, BZC, BZE, BZL, BZS | HÜBNER, 2013 | ||||||||||||||||||
| Cyperaceae | Diplasia Karatifolia Богатый. экс перс. | Розетка | COS, PAN, TRT, FRG, GUY, SUR, VEN, BOL, CLM, ECU, PER, BZN | Weissenhofer и др.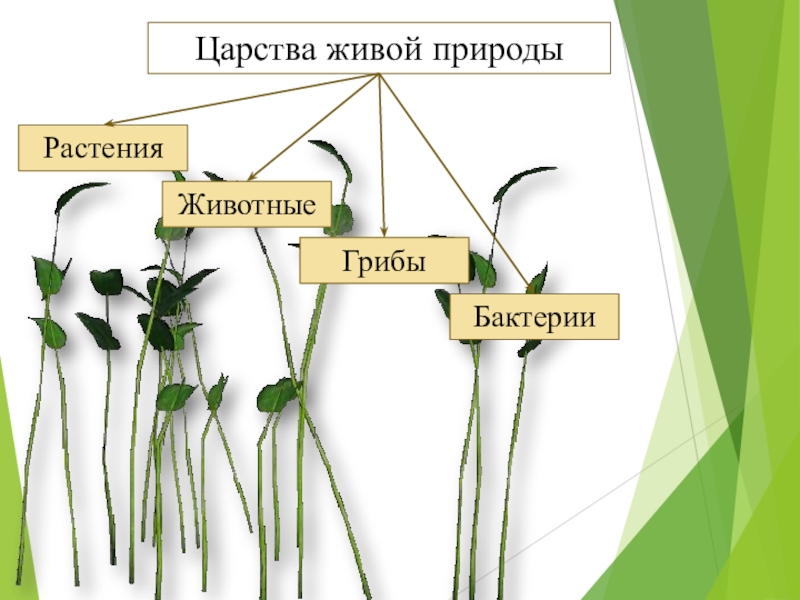 , 2008 , 2008 | ||||||||||||||||||
| Euphorbiaceae | Agrosstistachys Borneensis Becc. | Schopfbaum | IND, SRL, Tha, Vie, Bor, Mly, Phi, Sum, NWG | NG, 1980 | NG, 1980 | |||||||||||||||||
| Euphorbiaceae | эритрококка Membranacea (Müll.arg.) Prinain | Schopfbaum | НГА, CMN, GAB | 1992 | Blanc||||||||||||||||||
| молочайные Trigonostemon detritiferus R.Milne | Schopfbaum БОР | Милна, 1994 | ||||||||||||||||||||
| молочайные Trigonostemon wetriifolius AiryShaw & Нг | Schopfbaum | MLY | Ng, 1980 | |||||||||||||||||||
| Gelsemiaceae | Mostuea brunonis Didr. | Shrub | CMN, CON, GAB, TAN, MLW, MDG | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Gesneriaceae | CODONOBOEA PLATYPUS (CBCLARKE) CLIMLIC | Shrub | Mly | Kew & Wong, 1988 | ||||||||||||||||||
| Gesneriaceae | Cyrtandra Erectipila BLBURT | Shrub | BOR | Kew & Wong, 1988 | ||||||||||||||||||
| LecythiDaceae | Barringtonia Colori Kew & K. M.Wong M.Wong | Shopfbaum MLY | Kiew & Wong, 1988 | |||||||||||||||||||
| Lecythidaceae Grias angustipetala Cornejo & Samori | Разветвленная schopfbaum | ECU КОРНЕХО & Mori, 2012 | ||||||||||||||||||||
| Lecythidaceae | Grias cauliflora L. | Schopfbaum | BLZ, COS, GUA, HON, NIC, PAN, JAM, CLM | Предполагается; см. Prance & Mori, 1979 | ||||||||||||||||||
| Lecythidaceae | Grias colombiana Cuatrec. | Шопфбаум | CLM | Предполагаемый; см Prance & Mori, 1979 | ||||||||||||||||||
| Lecythidaceae Grias ecuadorica Cornejo & Samori | Schopfbaum ECU | КОРНЕХО & Mori, 2012 | ||||||||||||||||||||
| Lecythidaceae Grias haughtii R.Kunth | Шопфбаум | CLM | Предполагаемый; см. Prance & Mori, 1979 | |||||||||||||||||||
| Lecythidaceae | Grias longirachis S. A.Mori & J.L.Clark A.Mori & J.L.Clark | Schopfbaum | ECU | Clark & Mori, 2000 | ||||||||||||||||||
| Lecythidaceae | Grias multinerva 9000 | Шопфбаум | ЭБУ | Расчетный; см. Prance & Mori, 1979 | ||||||||||||||||||
| Lecythidaceae | Grias neuberthii J.F.Macbr. | Schopfbaum | PER, CLM, ECU | Предположительно; см. Prance & Mori, 1979 | ||||||||||||||||||
| Lecythidaceae | Grias peruviana Miers | Schopfbaum | ECU, PER | Предполагается; см. Prance & Mori, 1979 | ||||||||||||||||||
| Lecythidaceae | Grias purpuripetala S.А. Мори и Х. Д. Гарсия-Гонц. | Шопфбаум | CLM | Мори и др. , 2010 | ||||||||||||||||||
| Lecythidaceae Grias subbullata КОРНЕХО & Samori | Schopfbaum ECU | Cornejo & Мори, 2011 | ||||||||||||||||||||
| Lecythidaceae Grias theobromicarpa Cornejo & Samori | Schopfbaum | ECU | Cornejo & Mori, 2010 | |||||||||||||||||||
| Lecythidaceae | Gustavia grandibracteata Croat & S.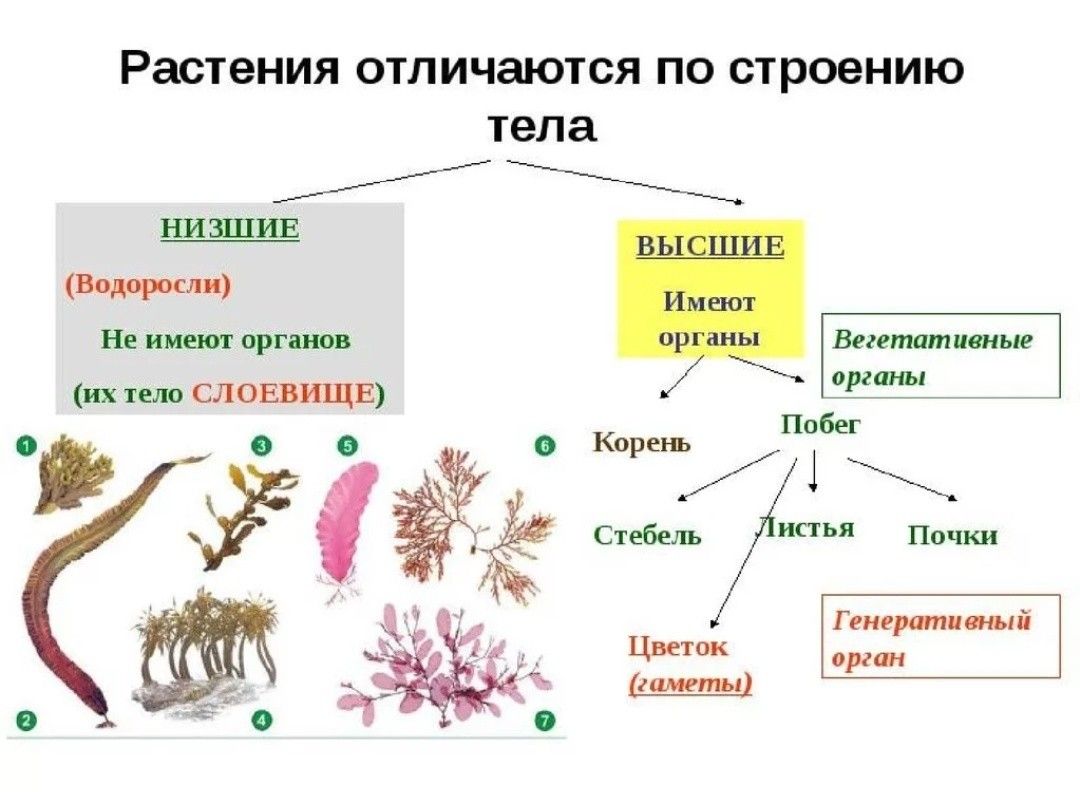 А. Мори А. Мори | Шопфбаум разветвленный | PAN, CLM | Предполагаемый; см. Prance & Mori, 1979 | ||||||||||||||||||
| Lecythidaceae | Gustavia monocaulis S.A.Mori | Schopfbaum разветвленный | PAN | 904 см. Prance & Mori, 1979|||||||||||||||||||
| Lecythidaceae | Gustavia pubescens Ruiz & Pav. бывший О.Берг. | Шопфбаум | ЭБУ | Расчетный; см. Prance & Mori, 1979 | ||||||||||||||||||
| Lecythidaceae | Gustavia sessilis S.А. Мори | Шопфбаум | CLM | Предположительно; см. Prance & Mori, 1979 | ||||||||||||||||||
| Malvaceae | Cola semecarpophylla K.Schum. | Schopfbaum | CMN | Kenfack и др. , 2007 | , 2007||||||||||||||||||
| Malvaceae | Overaribea Pumila W.S.S.Alverson | Schopfbaum | COS | COS | Alverson, 1984 | |||||||||||||||||
| Malvaceae | Scaphopetalum Mannii мачты. | Schopfbaum | GAB | GAB | Lachenaud & Jongkind, 2013 | |||||||||||||||||
| MORACEAE | FICUS PSEUDOPALMA BLANCO | SCHOPFBAUM | PHI | ZONA, PES. наблюдать | ||||||||||||||||||
| Moraceae | Scyphosyce manniana Baill. | Разветвленные Schopfbaum | CMN, GAB | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Nepenthaceae | Nepenthes Ampullaria гнездо | Bor | Bor | Moran et al. , 2003 | ||||||||||||||||||
| Nepenthaceae | Nepenthes lowii Hook.f. | Кувшин | БОР | Clarke и др. , 2009 | ||||||||||||||||||
| Nepenthaceae | Nepenthes rajah Hook.f. | Питчерная установка | БОР | Колодцы и др. , 2011 | , 2011||||||||||||||||||
| Ochnaceae | Gomphia Amplectens STAPF | Schopfbaum | LBR | Hawtherne, 2013 | ||||||||||||||||||
| Ochnaceae | Gomphia Duparquetiana Baill. | Schopfbaum | EQG, GAB | Lachenaud & JongInd, 2013 | ||||||||||||||||||
| Ochnaceae | Gomphia SubCordata STAPF | Schopfbaum | IVO, LBR, SIE | . см. Poorter et al. , 2004 | ||||||||||||||||||
| Orchidaceae | Acriopsis liliifolia (J.Koenig) Seidenf. | Эпифит | EHM, AND, CBD, LAO, MYA, THA, VIE, BOR, JAW, LSI, MLY, MOL, PHI, SUL, SUM, NWG, SOL, QLD, CRL | Зона, перс.наблюдать | ||||||||||||||||||
| Orchidaceae | Ansellia africana Lindl. | Epiphyte | BEN, GHA, GNB, IVO, LBR, NGA, SIE, BUR, CAF, CMN, CON, EQG, GAB, GGI, RWA, ZAI, SUD, KEN, TAN, UGA, ANG, MLW, MOZ , ЗАМ, ЗИМ, БОТ, НАМ, НАТ, ЮЗЗ, ТВЛ | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Bulbophyllum beccarii Rchb.f. | Эпифит | БОР | Зона, чел. наблюдать | ||||||||||||||||||
| Орхидные | Catasetum atratum Lindl. | Эпифит | БЗЛ, БЗН, БЗС | Зона, чел. наблюдать.; Holst, 1999 | ||||||||||||||||||
| Orchidaceae | Catasetum cernuum (Lindl.) Rchb.f. | Эпифит | ТРТ, ВЭН, БЗЛ, БЗС | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Catasetum fimbriatum (C.Morren) Lindl. | Epiphyte | VEN, BOL, BZC, BZL, BZS, AGE, AGW, PAR | Holst, 1999; Педросо-де-Мораес и др. , 2012 | ||||||||||||||||||
| Orchidaceae | Catasetum globiflorum Hook. | Эпифит | БЗЛ | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Catasetum integerrimum Hook. | EpiPHYTE | MXC, MXE, MXG, MXS, MXT, BLZ, COS, ELS, GUA, HON, NIC, COUS | Holst, 1999 | ||||||||||||||||||
| Orchidaceae | Catasetum Lanciferum Lindl. | Эпифит | BZL | Holst, 1999 | ||||||||||||||||||
| Orchidaceae | Catasetum macrocarpum Rich. ex Kunth ex Kunth | Epiphyte | TRT, FRG, GUY, SUR, VEN, CLM, BZC, BZE, BZL, BZN, AGE, AGW | Зона, чел. наблюдать | ||||||||||||||||||
| Орхидных Catasetum osculatum Ласердо & VPCastro | Эпифита BZC, BZN | Холста, 1999 | ||||||||||||||||||||
| Орхидных Catasetum rodigasianum Рольф | Эпифита BZS | Зона, чел. наблюдать | ||||||||||||||||||||
| Орхидные | Catasetum saccatum Lindl. | Эпифит | ГАЙ, ВЕН, БОЛ, КЛМ, ЭБУ, ПЕР, БЗН | Зона, чел. наблюдать | ||||||||||||||||||
| Орхидных | Catasetum spitzii Hoehne | Эпифита | BZC, BZN | Холста, 1999 | ||||||||||||||||||
| Орхидных | Catasetum uncatum Рольфа | Эпифита | BZE | Холста, 1999 | ||||||||||||||||||
| Orchidaceae | Catasetum viridiflavum Hook. | Эпифит | HON, PAN | Holst, 1999 | ||||||||||||||||||
| Orchidaceae | Clowesia rosea Lindl. | Эпифит | MXS | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Coryanthes picturata Rchb.f. | Эпифит | MXG, MXS, MXT, BLZ, COS, GUA, HON, NIC, PAN | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Coryanthes speciosa Hook. | Эпифит | ТРТ, ФРГ, ГАЙ, СУР, ВЕН, ПЕР, БЗЦ, БЗЭ, БЗЛ, БЗН, БЗС | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Cymbidium aloifolium (L.) Sw. | Эпифит | CHC, CHS, ASS, BAN, EHM, IND, NEP, SRL, AND, CBD, LAO, MYA, THA, VIE, JAW, MLY, SUM | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Cymbidium finlaysonianum Lindl. | Эпифит | CBD, THA, VIE, BOR, JAW, MLY, MOL, PHI, SUL, SUM | N.Юи, чел. ком. | ||||||||||||||||||
| Orchidaceae | Cyrtopodium gigas (Vell.) Hoehne | Эпифит | BZC, BZE, BZL, BZN, BZS | 9045 наблюдать|||||||||||||||||||
| Orchidaceae | Cyrtopodium punctatum (L. ) Lindl. ) Lindl. | Эпифит | ФЛА, КАЙ, КУБ, ДОМ, ХАЙ, ПУЭ, ТРТ, ВЕН | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Gongora bufonia Lindl. | Эпифит | БЗЭ, БЗЛ, БЗС | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Grammangis ellisii (Lindl.) Rchb.f. | Эпифит | ЦРТ | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Grammatophyllum multiflorum Lindl. var. тигринум Линдл. | Эпифит | ФИ, | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Grammatophyllum scriptum (L.) Blume | Epiphyte | BOR, LSI, MOL, PHI, SUL, BIS, NWG, SOL, FIJ, SCZ | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Grammatophyllum speciosum Blume | Эпифит | LAO, MYA, THA, VIE, BOR, JAW, MLY, SUL, SUM 9045. наблюдать | |||||||||||||||||||
| Orchidaceae | Graphorkis lurida (Шв. ) Kuntze ) Kuntze | Эпифит | BEN, GHA, GNB, GUI, IVO, LBR, NGA, CONGAF, EQGN, BUR, SIE, TO , GAB, GGI, RWA, ZAI, ETH, TAN, UGA, ANG, ZAM | Зона, чел.наблюдать | ||||||||||||||||||
| Orchidaceae | Stanhopea cirrhata Lindl. | Epiphyte | COS, HON, NIC, PAN | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Stanhopea grandiflora (Lodd.) Lindl. | Эпифит | ТРТ, ФРГ, ГАЙ, СУР, ВЕН, ЦЛМ, ПЕР, БЗЭ, БЗЛ, БЗН | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Stanhopea lietzei (Regel) Schltr. | Эпифит | БЗЭ, БЗЛ, БЗС | Зона, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Stanhopea tigrina Bateman ex Lindl. var. nigroviolacea C.Morren | Epiphyte | MXE, MXG | Zona, чел. наблюдать | ||||||||||||||||||
| Orchidaceae | Thecostele alata (Roxb.) C.S.P.Parish & Rchb.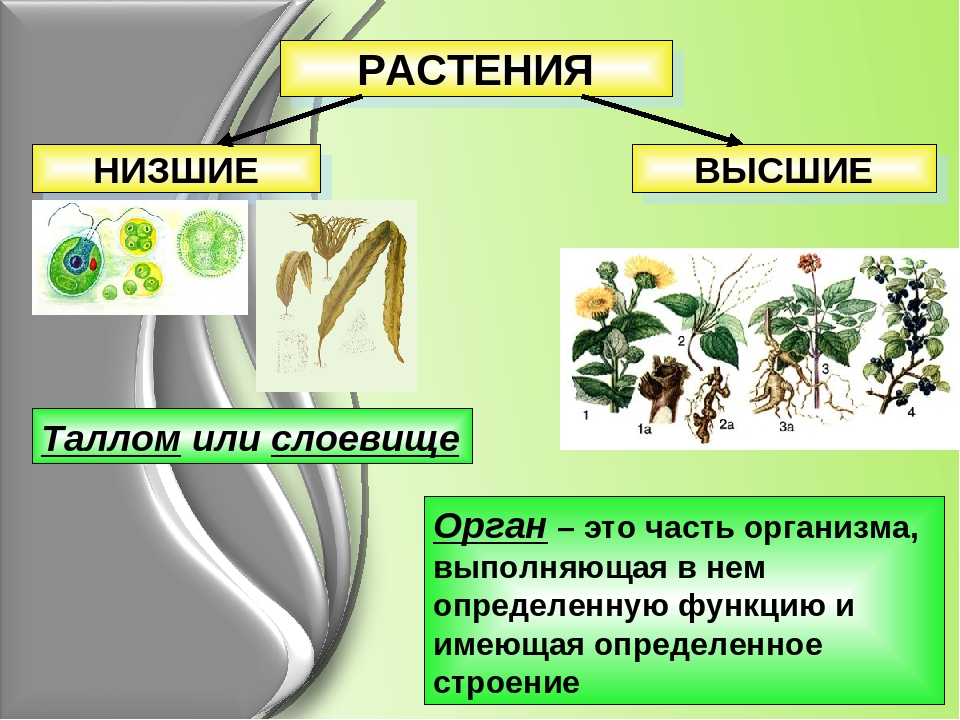 f. f. | Эпифит | БАН, ЛАО, МЬЯ, ТХА, ВИЭ, БОР, ЧЕЛЮСТЬ, МЛИ, ФИ, СУММ | Зона, чел.наблюдать | ||||||||||||||||||
| Pandanaceae | Benstonea alticola (Holttum & H. St. John) Callm. & Буэрки | Эпифит | БОР, МЛУ, | Каллмандер, чел. ком. | ||||||||||||||||||
| Pandanaceae | Benstonea dumetorum (Holttum & H. St. John) Callm. и буерки | Кустарник | МЛУ | Коллмандер, чел. ком. | ||||||||||||||||||
| Pandanaceae | Benstonea epiphytica (Martelli) Callm.& Buerki | Epiphyte | BOR, MLY, SUM | Ng, 1980 | ||||||||||||||||||
| Pandanaceae | Benstonea inquilina (B.C. & Буэрки | Эпифит | БОР | Коллмандер, чел. ком. | ||||||||||||||||||
| Pandanaceae | Benstonea leptatophila (BC Stone) Callm. и буерки | Кустарник | БОР | Коллмандер, чел. ком. | ||||||||||||||||||
| Pandanaceae | Benstonea nana (Martelli) Callm. и буерки и буерки | Кустарник | ТХА, МЛН | Каллмандер, чел. ком. | ||||||||||||||||||
| Pandanaceae | Benstonea pachyphylla (Merr.) Callm. & Buerki | Schopfbaum | BOR | Callmander, чел. ком. | ||||||||||||||||||
| Pandanaceae | Benstonea pectinata (Martelli) Callm. и буерки | Кустарник | БОР, СУММ | Callmander, чел. ком. | ||||||||||||||||||
| Pandanaceae | Benstonea Saint-johnii (B.К. Стоун) Callm. и буерки | Кустарник | МЛУ | Коллмандер, чел. ком. | ||||||||||||||||||
| Pandanaceae | Benstonea thomissopylla (B.C. Stone) Callm. и буерки | Кустарник | БОР | Коллмандер, чел. ком. | ||||||||||||||||||
| Pandanaceae | Pandanus sechellarum Balf.f. | Шопфбаум разветвленный | SEY | Christenhusz, чел. наблюдать | ||||||||||||||||||
| Polypodiaceae | Campyloneurum phyllitidis (L. ) C.Presl ) C.Presl | Epiphyte | FLA, MXC, MXE, MXG, MXS, MXT, BLZ, GUA, HON, NIC, PAN, COS, BAH, CUB, DOM, JAM, LEE, TRT, CLM, FRG, SUR , VEN, ECU, PER, BOL, BZC, BZL, BZN, BZS, AGE | Zuquim и др. , 2008 г.; Зона, чел. Операционные системы. | ||||||||||||||||||
| Polypodiaceae | Drynaria aglaomorpha Christenh | Эпифит | PHI | Copeland 1905, Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria bonii Христос | эпифит | THA, VIE, MLY | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria brooksii (Copel.) Кристен. | Epiphyte | BOR | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria cornucopia (Copel.) Alderw | Epiphyte | PHI | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria coronans J.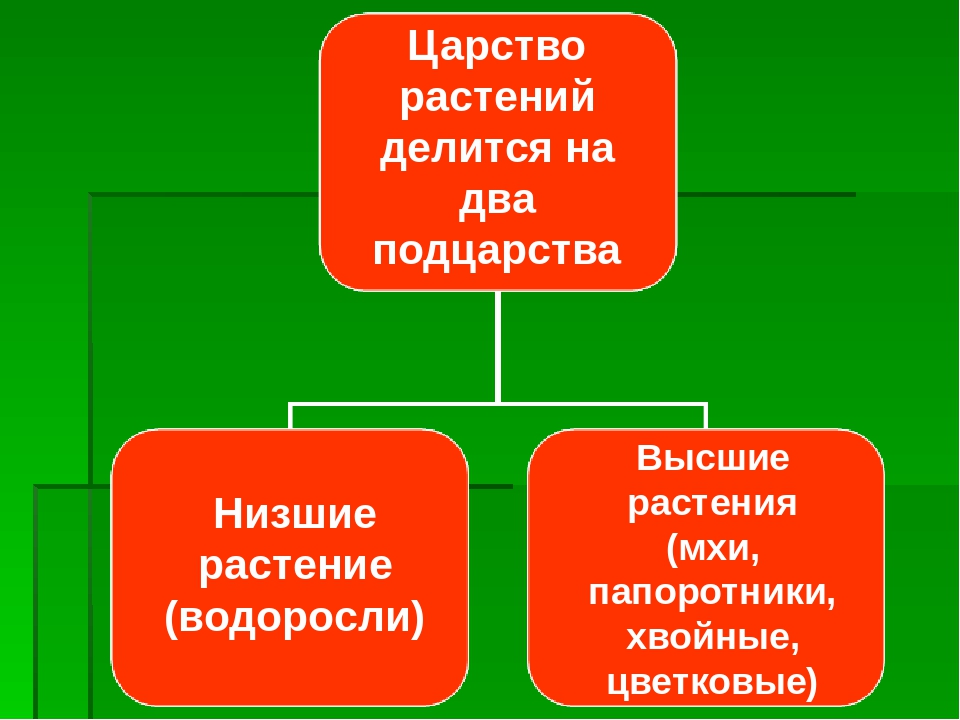 Sm. Sm. | Эпифит | ЭУ, КБР, VIE, MYA, ТНА, MLY | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria delavayi Христос | Эпифит | CHC | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria descensa Copel. | Epiphyte | PHI | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria drynarioides (Hook.) Christenh. | EPIPHYTE | , MLY, SUL, SUL, PHI, NWG, BIS, SOL, SUL, PHI, NWG, BIS, SOL | JANSSEN & SCHNEIDER, 2005 | ||||||||||||||||||
| PolyPodiaceae | Drynaria Fortunei (Kunze EX Mett.) J.SM. (Kunze)Moore | Epiphyte | THA, MLY, SUM, SUL, PHI, NWG, BIS, SOL | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | 90.455 Dry involderariaEpiphyte | MYA, MLY | Janssen & Schneider, 2005 | |||||||||||||||||||
| Polypodiaceae | Drynaria laurentii (Christ) Hieron. | Эпифит | GAB, LBR | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria meyeniana (Schott) | Эпифит | PHI | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria mollis Bedd. | Epiphyte | WHM | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria novoguineensis (Christenh.Brause) | Эпифит | NWG | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria pleuridioides C.Presl | Epiphyte | JAW, PHI | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria propinqua (Wall.)d ex. | Эпифит | WHM, NEP | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria quercifolia (L. | Эпифит | CHC, CHS, CHH, TAI, EHM, IND, THA, VIE, BOR, JAW, MOL, SUM, MLY, PHI, SUL, NWG, BIS, SOL, QLD | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria hardula (Sw. ) Постель. ) Постель. | Наземный или EpiPhyte | Tha, Vie, Bor, Jaw, Mol, Sum, Mly, Phi, Sul, NWG, BIS, SOL, QLD | Janssen & Schneider, 2005 | ||||||||||||||||||
| PolyPodiaceae | Drynaria Roosii Nakaike | Эпифит | CHS, | Прогнозные | ||||||||||||||||||
| Polypodiaceae | Drynaria Синика Дильса | Эпифит | CHC | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria sparsisora (Desv.) T.moore | EPIPHYTE | THA, VIE, BOR, JAW, MOL, SUM, MLY, PHI, SUL, NWG, BIS, SOL, QLD | JANSSEN & SCHNEIDER, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria volkensii Гиерон. | EPIPHYTE | CMN, RWA, ZAI, GGI, ETH, SUD, KEN, TAN, MLW | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Dromnaria Willdenowiii (BRORY) T.MOORE | EPIPHYTE | SUD, MDG, MAU | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Drynaria x leporella K. I.Goebel I.Goebel | Epiphyte | – | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Elaphoglossum aemulum 90.006 (BrackenKackerulfum aemulum) | Эпифит ГАУ | ||||||||||||||||||||
| Polypodiaceae Elaphoglossum alatum T.Moore | Эпифит FIJ, ГАУ | |||||||||||||||||||||
| Polypodiaceae Elaphoglossum blumeanum (Fée), J.См. | Эпифит | JAW, MLY, PHI | Предполагаемый | |||||||||||||||||||
| Polypodiaceae | Elaphoglossum crassicaule Copel. | EPIPHYTE | HAW | Вывод | Вывод | |||||||||||||||||
| Polypodiaceae | Elaphoglossum Eximium (Mett.) Христос | MXS, MXT, COS, ELS, GUA, HON, NIC, PAN, GUY, VE, PER CLM, ECU, PER | Christenhusz, чел. наблюдать | |||||||||||||||||||
| PolyPodiaceae | Elapholossum Luridum (Fée) Христос | EpiPhyte | Pan, Trt, Frg, Sur, Ven, Bol, CLM, ECU, PER, BZL, BZS | ZUQUIM et al. , 2008 г.; Кристенхуш, перс. наблюдать , 2008 г.; Кристенхуш, перс. наблюдать | ||||||||||||||||||
| Polypodiaceae | Elaphoglossum macrophyllum (Klotzsch) Christ | Epiphyte | FRG, GUY, VEN, CLM, BZL, BZS 6 per.huzs 6 наблюдать | |||||||||||||||||||
| Polypodiaceae | Elaphoglossumpalaceum (Hook. & Grev.) Sledge | Epiphyte | MXC, MXG, MXS, COS, VDU ELS, JAM, EOM, EOM, GANUA, HON PER, HAW | Christenhusz, чел.наблюдать | ||||||||||||||||||
| Polypodiaceae | Elaphoglossum parvisquameum Skottsb. | EPIPHYTE | HAW | Вывод | Вывод | |||||||||||||||||
| Polypodiaceae | Elapholossum Plumosum (FEE) T.moore | EpiPhyte | COS, PAN, LEE, CLM, VEN, TRT, FRG, GUY, SUR, BOL , CLM, ECU, PER, BZC, BZL, BZN, BZS | Zuquim и др. , 2008 | ||||||||||||||||||
| Polypodiaceae | Grammitis punctata Raddi | Epiphyte | BZL, BZS | Bishop Copeland, 1907 | ||||||||||||||||||
| Polypodiaceae | Loxogramme lanceolata (Шв. )C.Presl )C.Presl | Epiphyte | GHA, GUI, IVO, LBR, NGA, SIE, TOG, CAF, CMN, CON, EQG, GAB, GGI, ZAI, ETH, SOC, SOM, SUD, KEN, TAN, UGA , ANG, CPP, LES, NAT, SWZ, TVL, COM, MAU, MDG, REU, YEM | Christenhusz, чел. наблюдать | ||||||||||||||||||
| Polypodiaceae | Microsorum linguiforme (Mett.) Copel. | EPIPHYTE | BOR, SUM, SOL, MOL, PHI, NWG, BIS, SOL, FIJ | JANSSEN & SCHNEIDER, 2005 | ||||||||||||||||||
| PolyPodiaceae | Musicifolium (Blume) Copel. | Epiphyte | MYA, THA, MLY, SUM, JAW, BOR, PHI, NWG | Janssen & Schneider, 2005 | ||||||||||||||||||
| Polypodiaceae | Epiphyte | BEN, GHA, GNB, GUI, IVO, LBR, NGA, SEN, SIE, TOG, BUR, CAF, CMN, CON, EQG, GAB, GGI, RWA, ZAI, ETH, TAN, UGA, ANG , ZAM, MDG, COM, CHC, CHS, CHH, TAI, EHM, IND, THA, VIE, BOR, JAW, MOL, SUM, MLY, PHI, SUL, NWG, BIS, SOL, QLD, FIJ | Christenhusz, чел.наблюдать | |||||||||||||||||||
| Polypodiaceae | Platycerium alcicorne (Willem. ) Desv. ) Desv. | Эпифит | КЕН, ТАН, МОЗ, ЗИМ, КОМ, ЦРТ, МАУ, РЭУ, СЭЙ | Вердкур, 2001; Кристенхуш, перс. наблюдать | ||||||||||||||||||
| Polypodiaceae | Platycerium andinum Baker | Epiphyte | BOL, PER | Janssen & Schneider, 2005; Кристенхуш, перс. наблюдать | ||||||||||||||||||
| Polypodiaceae | Platycerium bifurcatum (Cav.) С.Хр. | Epiphyte | JAW, MOL, LSI, QLD | Holttum, 1938 | ||||||||||||||||||
| Polypodiaceae | Platycerium coronarium (D. | Эпифит | MLY, BOR, JAW, LSI, PHI | Holttum, 1938 | ||||||||||||||||||
| Polypodiaceae | Platycerium слона 9006 | EPIPHYTE | SEN, GUI, CON, BUR, RWA, CAF, SUD, ETH, KEN, UGA, TAN, ANG, MLW, MOZ | Verdcourt, 2001 | ||||||||||||||||||
| Polypodiaceae | Platycrium Ellisii Baker | Epiphyte | MDG | Holttum, 1938 | ||||||||||||||||||
| Polypodiaceae | Platycerium grande (A. Канн. ex Fée) J.Sm. Канн. ex Fée) J.Sm. | Эпифит ФИ, СУП, МРН | ||||||||||||||||||||
| Polypodiaceae Platycerium hillii T.Moore | Эпифит QLD, НРГ | |||||||||||||||||||||
| Polypodiaceae Platycerium holttumii Joncheere & Hennipman | EpiPhyte | Mly, Tha, Vie, Lao, CBD | Вывоз | |||||||||||||||||||
| PolyPodiaceae | Platycrium Madagascariense Baker | EPIPHYTE | MDG | Zona, Pers.наблюдать.; Holttum, 1938 | ||||||||||||||||||
| Polypodiaceae | Platycerium quadridichotomum (Bonap.) Tardieu | Эпифит | MDG | Holttum, 1938 | ||||||||||||||||||
| Polypodiaceae | Platycerium ridleyi Христос | Эпифит | MLY, БОР , SUM, THA | Предполагаемый | ||||||||||||||||||
| Polypodiaceae | Platycerium Stemaria (P.Beauv.) Desv. | Эпифита | ГСГ, СИУ, ЯГРЫ, АКР, CON, ГГ, GAB, УЗА | Verdcourt, 2001 | ||||||||||||||||||
| Polypodiaceae | Platycerium superbum Joncheere & Hennipman | Эпифит | QLD, Новый Южный Уэльс | Christenhusz , чел. наблюдать наблюдать | ||||||||||||||||||
| Polypodiaceae | Platycerium veitchii (Underw.) C.Chr. | Эпифит | QLD | Предполагаемый | ||||||||||||||||||
| Polypodiaceae | Platycerium wallichii Hook. | Epiphyte | MYA | Holttum, 1938 | ||||||||||||||||||
| Polypodiaceae | Platycerium wandae Racib. | Эпифит | MOL, НРГ, Новый Южный Уэльс | Прогнозные | ||||||||||||||||||
| Primulaceae | Ardisia bracteata Бейкер | Разветвленная schopfbaum | GAB | Lachenaud & Йонгкинд, 2013 | ||||||||||||||||||
| Primulaceae | Ardisia hallei Taton | Schopfbaum разветвленный | GAB | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Primulaceae | Ardisia mayumbensis (R.D.Good) Taton | Branched schopfbaum | CMN, CON, GAB | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Primulaceae | Clavija bija | Шопфбаум с разветвлениями | COS, PAN, CLM | Предполагаемый; см. Ståhl, 2010 Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija caloneura Mart. бывший Мик. | Шопфбаум разветвленный | BZE, BZL, | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija cardenasii Rusby | Schopfbaum разветвленный | PER, BOL | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija cauliflora Regel | Schopfbaum | CLM | Inferred; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija clavata Decne. | Schopfbaum | VEN | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija colombiana B.Ståhl | Schopfbaum разветвленный | CLM | In см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija costaricana Pittier | Schopfbaum | COS, NIC, PAN, CLM | 90 , 2008 г.|||||||||||||||||||
| Primulaceae | Clavija domingensis Urb.& Экман | Шопфбаум | HAI, | Предположительно; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija eggersiana Mez | Schopfbaum | ECU | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija elliptica Mez | Schopfbaum разветвленный | PER, BZN | Предположительно; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija engelsii Mez | ? | CLM или VEN | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija euerganea J.Ф. Макбр. | Шопфбаум с разветвлениями | ECU, PER | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija fernandezii Philipson | Schopfbaum разветвленный | CLM | Предполагается; см. Ståhl, 2010 Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija fusca B.Ståhl | Schopfbaum разветвленный | PAN, CLM | 904; см. Ståhl, 2010|||||||||||||||||||
| Primulaceae | Clavija grandis Decne. | Шопфбаум | CLM | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija harlingii B.Ståhl | Разветвленный schopfbaum | CLM, ECU, PER | PER PER см. Ståhl, 2010|||||||||||||||||||
| Primulaceae | Clavija hookeri DC. | Шопфбаум разветвленный | PER | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija imatacae B.Ståhl | Разветвленный Schopfbaum | GUY, VEN | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija imazae B.Ståhl | Schopfbaum | PER | Inferred; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija jelskiy Szyszył.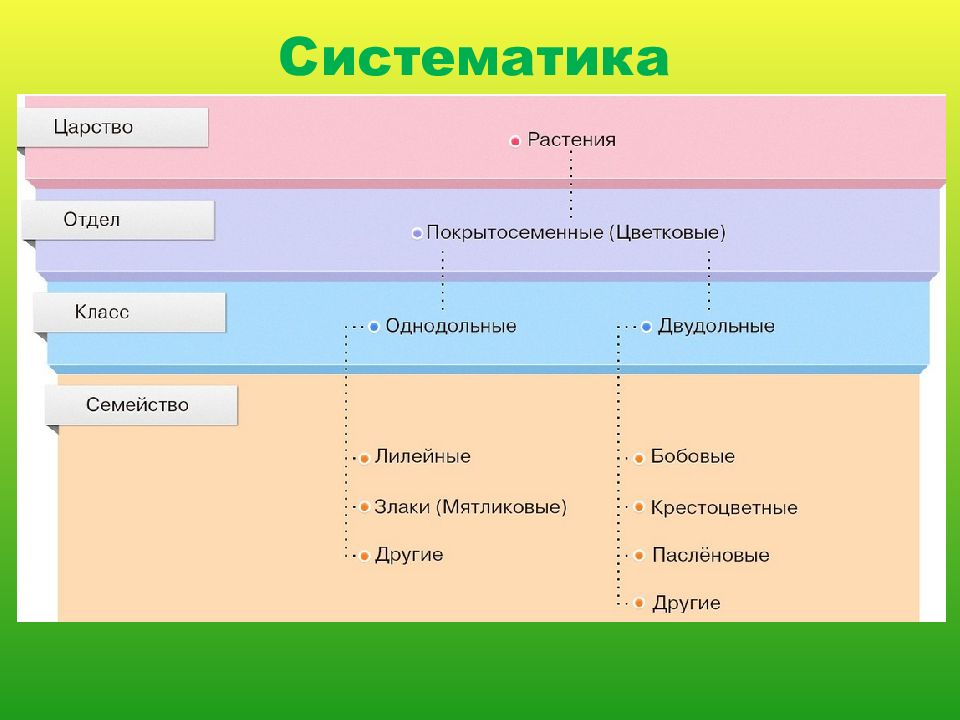 | Шопфбаум разветвленный | PER | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija kalbreyeri (Kuntze) Mez | Schopfbaum | CLM | Inferred; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija lancifolia Desf. | Шопфбаум разветвленный | SUR, GUY, FRG, VEN, BOL, BZE, BZN | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija laplanadae B.Ståhl | Schopfbaum разветвленный | CLM, ECU | Предположительно 904; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija latifolia (Willd. ex Roem. & Schult.) K.Koch | Branched schopfbaum | CLM 5 см. Ståhl, 2010 | |||||||||||||||||||
| Primulaceae | Clavija lehmannii Mez | Schopfbaum разветвленный | CLM | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija leucocraspeda B. Ståhl Ståhl | Разветвленный Schopfbaum | PER | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija longifolia Ruiz & Pav. | Шопфбаум | PER | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija macrocarpa Ruiz & Pav. | Шопфбаум | PER | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija macrophylla (Link ex Roem.& Schult.) Miq. | Шопфбаум разветвленный | ГАЙ, БЗЭ, БЗН | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija membranacea Mez | Schopfbaum разветвленный | CLM, ECU | Предположительно; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija mezii Pittier | Schopfbaum | PAN, CLM | Inferred; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija minor B.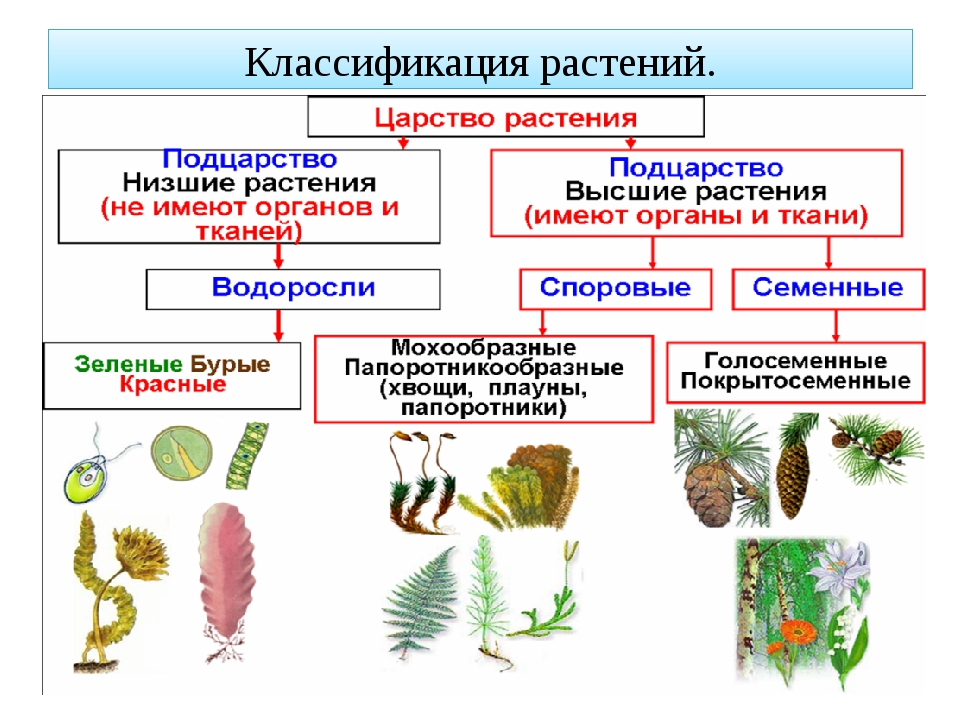 Ståhl Ståhl | Разветвленный Schopfbaum | CLM, VEN | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija myrmeciocarpa B.Ståhl | Schopfbaum разветвленный | PER | 904; см. Ståhl, 2010|||||||||||||||||||
| Primulaceae | Clavija ignorega B.Ståhl | Schopfbaum разветвленный | CLM | Inferred; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija nutans (Vell.) B.Ståhl | Разветвленный schopfbaum | BOL, BZC, BZL, BZS, PAR | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija obtusifolia B.Ståhl | Schopfbaum разветвленный | PER | Предположительно; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija ornata D.Don | Schopfbaum | CLM, VEN, ECU | Предположительно; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija parvula Mez | Schopfbaum разветвленный | ECU | Предполагается; см. Ståhl, 2010 Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija peruviana B.Ståhl | Разветвленный Schopfbaum | PER | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija plumbea B.Ståhl | Schopfbaum | CLM | Inferred; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija poeppigii Mez | Schopfbaum разветвленный | PER, BOL, BZN | 6 Inferred; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija procera B.Ståhl | Разветвленный Schopfbaum | ECU, PER | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija pubens D’Arcy | Schopfbaum разветвленный | PAN | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija pungens (Willd. ex Roem. & Schult.) Decne. | Разветвленный шопфбаум | ECU | Предполагаемый; см. Ståhl, 2010 Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija repanda B.Ståhl | Разветвленный Schopfbaum | ECU | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija rodekiana Linden & André | Schopfbaum разветвленный | CLM | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija Santae-Martae B.Ståhl | Schopfbaum | CLM | Inferred; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija serratifolia Mez | ? | CLM | Предполагается; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija spinosa (Vell.) Mez | Разветвленный Schopfbaum | BZL | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija subandina B.Ståhl | Schopfbaum | ECU | Inferred; см. Ståhl, 2010 Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija tarapotana Mez | Schopfbaum | BOL, PER, BZN | 904; см. Ståhl, 2010|||||||||||||||||||
| Primulaceae | Clavija umbrosa (Linden) Regel | Schopfbaum | BZN | Inferred; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija venosa B.Ståhl | Разветвленный Schopfbaum | ECU, PER | Предполагаемый; см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija wallnoeferi B.Ståhl | Schopfbaum разветвленный | PER | In см. Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Clavija weberbaueri Mez | Разветвленный schopfbaum | CLM, ECU, PER, BOL, BZC, BZN см. Ståhl, 2010 | |||||||||||||||||||
| Primulaceae | Cybianthus multiflorus (A.C.SM.) G.agostini | Schopfbaum | D’Arcy | D’Arcy | ||||||||||||||||||
| Primulaceae | Cybianthus Schlimii (Cook.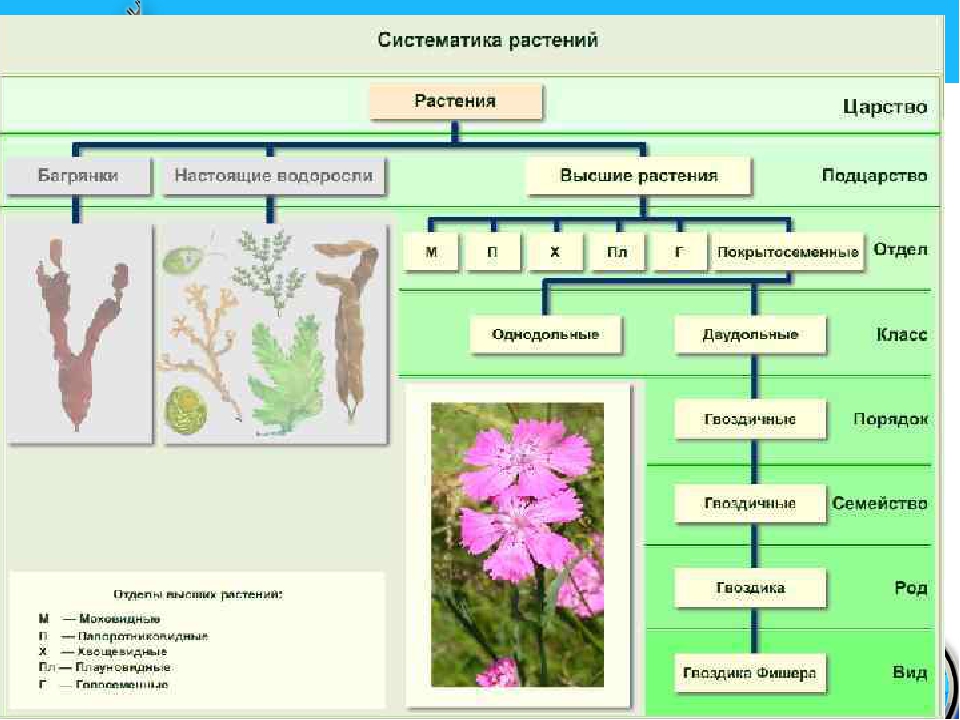 f.f.) G.Agostini f.f.) G.Agostini | Schopfbaum | NIC, COS, COS PAN, CLM, ECU, PER, BOL, BZC, BZN | Weissenhofer и др. , 2008 | ||||||||||||||||||
| Primulaceae | Cybianthus Simplex (Cook.f.) G.agostini | Schopfbaum | BZN, COS, CLM, ECU | D’Arcy, 1973 | ||||||||||||||||||
| Primulaceae | Cybianthus spectabilis (Standl.) G.Agostini | Schopfbaum COS, PAN | Д’Арси, 1973 | |||||||||||||||||||
| Primulaceae Fittingia метельчатая WNTakeuchi | Разветвленная schopfbaum | НРГ Takeuchi, 2008 | ||||||||||||||||||||
| Primulaceae | Hymenandra pittieri (Mez) Pipoly & Ricketson | Schopfbaum разветвленный | COS | Weissenhofer et al. , 2008 | ||||||||||||||||||
| Primulaceae | Theophrasta americana L. | Schopfbaum | DOM, HAI | Ståhl, 2010 | ||||||||||||||||||
| Primulaceae | Theophrasta jussieui Lindl.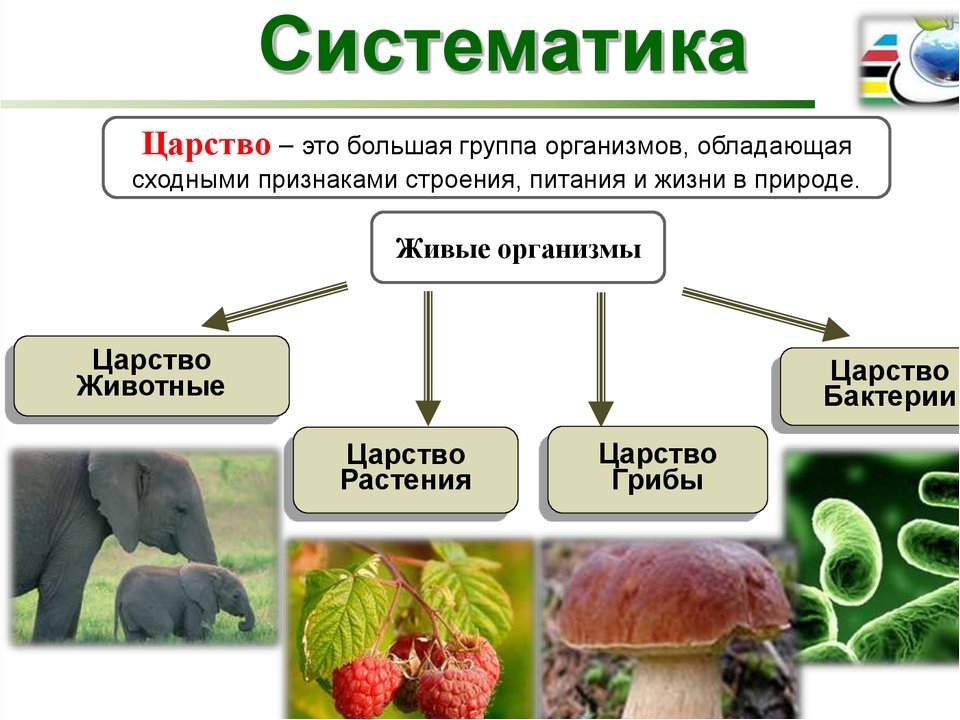 | Шопфбаум | HAI | Предполагаемый; Смотреть Stål, 2010 | ||||||||||||||||||
| Rubiaceae | Bertiera BiCarpellata (K.schum.) N.hallé | Schopfbaum | CAF, CMN, Con, Eqg, Gab, Zai | Blanc, 1992 | ||||||||||||||||||
| Rubiaceae | Bertiera Fimbriata Hepper | Schopfbaum | GUI, IVO, LBR | Hawtherne, 2013 | ||||||||||||||||||
| Rubiaceae | Chassalia Ischnophylla (K.Шуппер.) Hepper | Schopfbaum | IVO, NGA, CMN, CON, EQG, GAB | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| RUBIACEAE | Chazaliella Letozeyi Robbr. | Schopfbaum | CMN, GAB | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Rubiaceae | Chazaliella viridicalyx . | Schopfbaum | CAB, CMN, CON, EQG, GAB, ZAI | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Rubiaceae | Coffea. шев. шев. | Schopfbaum | IVO, LBR, SIE | Hawthorne, 2013 | ||||||||||||||||||
| Rubiaceae | Coffea magnistipula Stoff. & Ограб. | Schopfbaum | CMN, GAB | Cheek и др. , 2008 г.; Штоффелен и др. , 1997 | ||||||||||||||||||
| мареновые | Coffea mapiana Sonke | Schopfbaum | CMN | Hawthorne, 2013 | ||||||||||||||||||
| мареновые | Ixora scortechinii King & Gamble | Schopfbaum | ТНА, MLY | Hawthorne, 2013 | ||||||||||||||||||
| Rubiaceae | Ixora synactica DeBlock | Schopfbaum | CMN | Hawthorne 3; Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Rubiaceae | Morinda intermedia (Setch.и Кристоф.) Разафим. & B.Bremer | Schopfbaum | SAM | Предполагается; см. Setchell & Christophersen, 1935 | ||||||||||||||||||
| Rubiaceae | Morinda mayorii (Setch.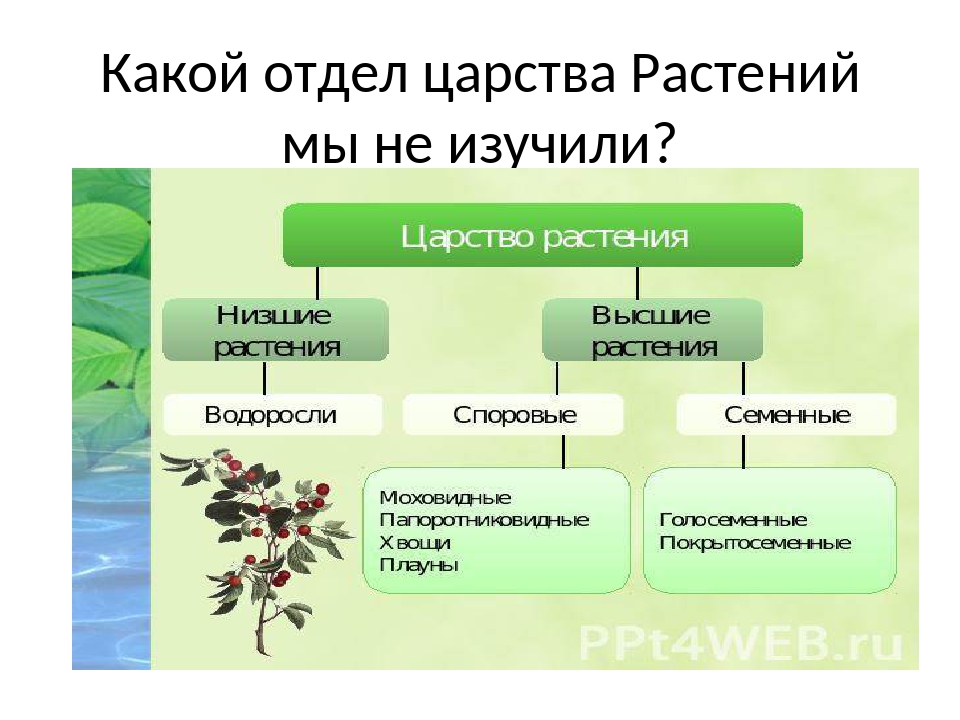 ) Razafim. & B.Bremer ) Razafim. & B.Bremer | Schopfbaum | SAM | Предполагается; см. Setchell & Christophersen, 1935 | ||||||||||||||||||
| Rubiaceae | Morinda multinervis (Setch. & Christoph.) Razafim. и Б.Бремер | Шопфбаум | СЭМ | Предположительно; см. Setchell & Christophersen, 1935 | ||||||||||||||||||
| Rubiaceae | Morinda pacifica (Reinecke) Razafim. & B.Bremer | Schopfbaum | SAM | Zona, чел. наблюдать | ||||||||||||||||||
| Rubiaceae | Morinda ramosa (Lauterb.) Razafim. & B.Bremer | Schopfbaum | SAM | Предполагается; см. Setchell & Christophersen, 1935 | ||||||||||||||||||
| Rubiaceae | Oxyanthus brevicaulis K.Krause | Schopfbaum | CMN, CON, GAB, ZAI | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Rubiaceae | Pavetta ankasensis | Разветвленный шопфбаум | GHA | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Rubiaceae | Pavetta pseudodigitata W. D.H.H.D.H. D.H.H.D.H. | Schopfbaum | IVO | Inferred | ||||||||||||||||||
| Rubiaceae | Pavetta sapoensis W.Д.Хот. | Schopfbaum | LBR | Inferred | ||||||||||||||||||
| Rubiaceae | Pentagonia macrophylla Benth. | Schopfbaum | COS | Weissenhofer и др. , 2008 | ||||||||||||||||||
| Rubiaceae | Pentagonia wendlandii Hook.f. | Schopfbaum | COS | Weissenhofer и др. , 2008 | ||||||||||||||||||
| Rubiaceae | Psychotria alfaroana Standl. | Schopfbaum | COS | Weissenhofer и др. , 2008 | ||||||||||||||||||
| мареновые Psychotria blydeniae O.Lachenaud & Jongkind | Schopfbaum Иво, LBR | Lachenaud & Jongkind, 2013 | ||||||||||||||||||||
| мареновые Psychotria bukaensis DeWild. | Schopfbaum | CAB, CON, GAB, ZAI | Lachenaud & Jongkind, 2013 | |||||||||||||||||||
| Rubiaceae | Psychotria chariana В.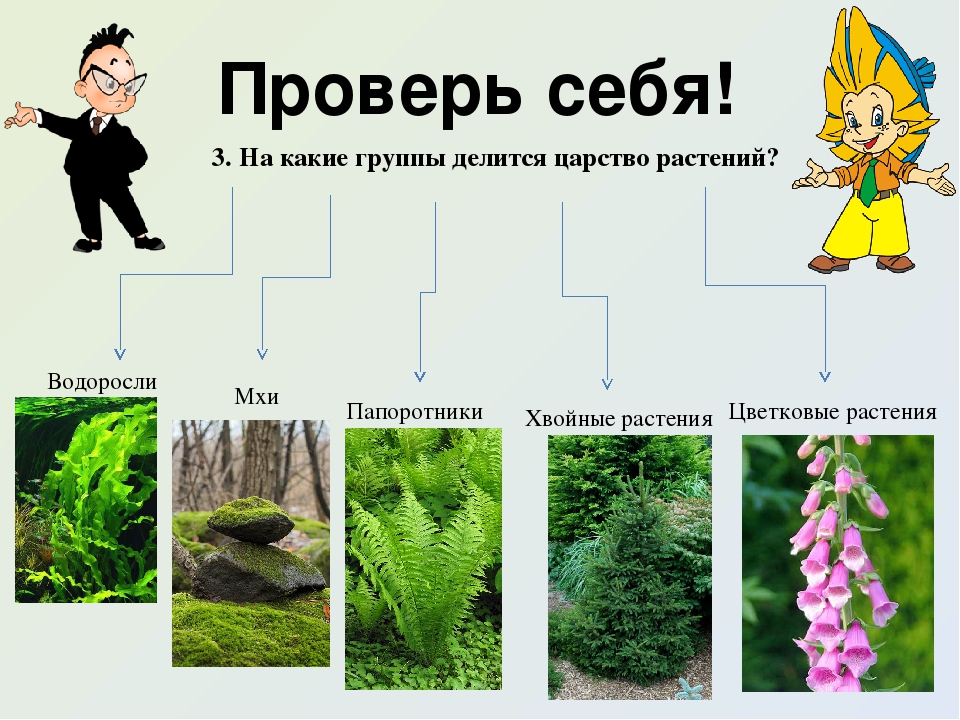 Хэм. Хэм. | Schopfbaum | COS | Weissenhofer и др. , 2008 | ||||||||||||||||||
| мареновые Psychotria dressleri (Dwyer) CWHam | Schopfbaum PAN, CLM | Dressler, 1985 | ||||||||||||||||||||
| мареновые Psychotria тысяч гектаров Hiern | Schopfbaum CMN , GAB | Lachenaud & Jongkind, 2013 | ||||||||||||||||||||
| Rubiaceae | Psychotria insueta (Dwyer) C.W.Ham | Schopfbaum PAN | Dressler 1985 | |||||||||||||||||||
| мареновые Psychotria konguensis Hiern | Schopfbaum САВ, CMN, CON, EQG, ГСЗ, ZAI | Lachenaud & Jongkind, 2013 | ||||||||||||||||||||
| Rubiaceae | Psychotria kupensis Cheek | Schopfbaum | CMN | Cheek et al. , 2008 | ||||||||||||||||||
| Rubiaceae | Psychotria micheliae (J.-G.Adam) Jongkind & Hawtherne | Schopfbaum | IVO, LBR | LACHENAUD & Jongkind, 2013 | ||||||||||||||||||
| RUBIACEAE | Psychotria Micheliana J.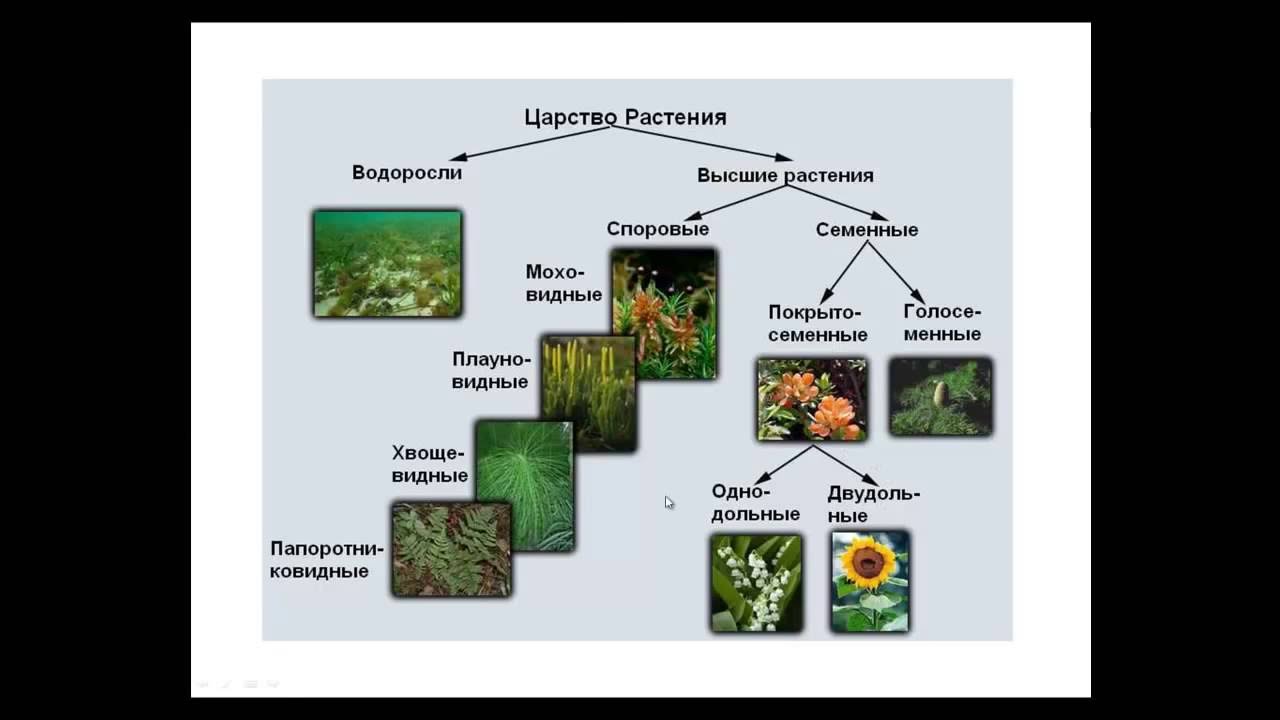 -G.Adam -G.Adam | Schopfbaum | IVO, LBR | Lachenaud & Jongkind, 2013 | ||||||||||||||||||
| Rubiaceae | Psychotria peduncularis (Salisb.) Styerm. var. suaveolens (Schweinf. ex Hiern) Verdc. | Schopfbaum | GHA, GUI, IVO, NGA, SIE, BUR, CMN, CON, GAB, RWA, ZAI, SUD, KEN, TAN, UGA, MLW, ZAM | Blanc, 1992 | ||||||||||||||||||
| Schopfbaum | LBR | Lachenaud & Jongkind, 2013 | ||||||||||||||||||||
| мареновые | Sabicea hierniana Wernham | Schopfbaum | CMN, CON, GAB | Halle, 1961 | ||||||||||||||||||
| мареновые | Stipularia africana P.Beauv. | Schopfbaum | Ben, GHA, IVO, LBR, NGA, SIE, CAF, CMN, CON, GAB, ZAI, ANG, ZAM | Hallé, 1961 | ||||||||||||||||||
| RUBIACEAE | Tricalysia Amplexizicaulis Robbr. | SchopFBAUM | CMN, GAB | HaB | HaBthorne, 2013 | |||||||||||||||||
| Rutaceae | Adiscanthus Fusciflorus Ducke | Schopfbaum | на | D’Arcy, 1973; Kallunki, 1992 | ||||||||||||||||||
| Rutaceae | Erythrochiton brasiliensis Nees & Mart. | Schopfbaum | VEN, GUY, FRG, SUR, BOL, CLM, PER, BZE, BZL, BZN | Зона, чел. наблюдать | ||||||||||||||||||
| Rutaceae | Erythrochiton fallax Kallunki | Schopfbaum | CLM, PER, ECU, BOL | Предполагается; см. Kallunki, 1992 | ||||||||||||||||||
| Rutaceae | Erythrochiton giganteus Kaastra & A.Х. Джентри | Шопфбаум | ЭКЮ | Расчетный; см. Kallunki, 1992 | ||||||||||||||||||
| Rutaceae | Erythrochiton gymnanthus Kallunki | Schopfbaum разветвленный | COS | 9045, 5; Weissenhofer & Weber, 2000|||||||||||||||||||
| Rutaceae | Erythrochiton hypophyllanthus Planch. & Linden | Schopfbaum | CLM | Предположительно; см. Kallunki, 1992 | ||||||||||||||||||
| Rutaceae | Erythrochiton odontoglossus Kallunki | Schopfbaum | ECU, PER | 6 Предположительно 90; см. Kallunki, 1992 | ||||||||||||||||||
| Rutaceae | Erythrochiton trichanthus Kallunki | Schopfbaum | PER | Предположительно; см. Kallunki, 1992 Kallunki, 1992 | ||||||||||||||||||
| Rutaceae | Leptothyrsa sprucei Benth.& Крюк.ф. | Schopfbaum | CLM, на | CLM, за | вывод | |||||||||||||||||
| Rutaceae | Toxosiphon Carinatus (Little) Kallunki | Schopfbaum | ECU, за | . см. Kallunki, 1992 | ||||||||||||||||||
| Rutaceae | Toxosiphon lindenii (Planch.) Baill. | Шопфбаум | MXT, GUA, NIC, PAN, COS | Предполагаемый; см. Kallunki, 1992 | ||||||||||||||||||
| Rutaceae | Toxosiphon macropodus (K.Краузе) Каллунки | Шопфбаум | ЭКЮ, PER, BOL, BZN | Предполагаемый; см. Kallunki, 1992 | ||||||||||||||||||
| Sabiaceae | Meliosma donnellsmithii Urb. | Schopfbaum | COS | Weissenhofer и др. , 2008 | ||||||||||||||||||
| Salicaceae | Phyllobotryon spathulatum Müll.Arg. | Шопфбаум | CMN, EQG, GAB | Бланк, 1992; Либала и др.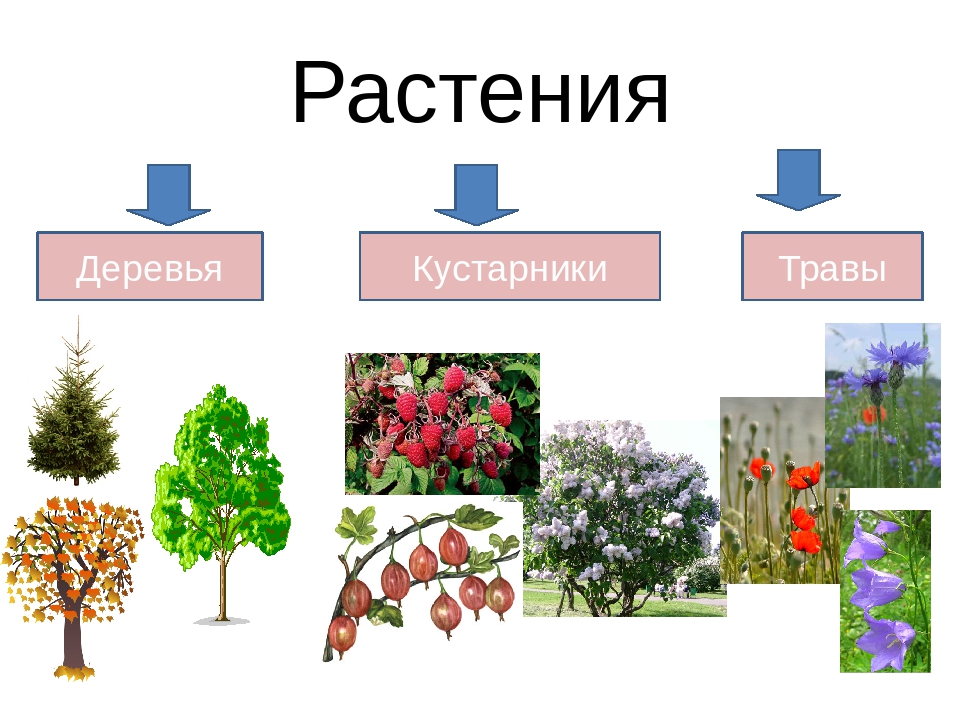

|
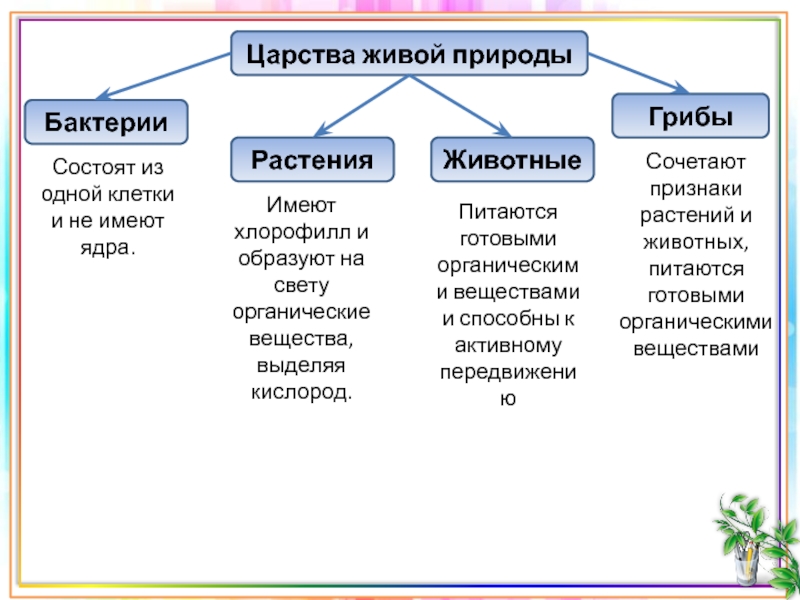 ; Ståhl, 2010
; Ståhl, 2010