
Царство бактерии — общая характеристика
Царство «Бактерии» состоит из бактерий и сине-зеленых водорослей, общая характеристика которых заключается в малой величине и отсутствии разделенного мембраной от цитоплазмы ядра.
Кто такие бактерии
В переводе с греческого «bakterion» – палочка. Большей частью, микробы – это невидимые невооруженным глазом одноклеточные организмы, размножающиеся делением.
Кто их открыл
Впервые увидеть мельчайших одноклеточных в самодельный микроскоп смог исследователь из Голландии, живший в 17 веке, Антони Ван Левенгук. Изучать окружающий мир через увеличительное стекло лупы он начал во время работы в галантерейном магазине.

Антони Ван Левенгук (1632 — 1723)
В дальнейшем Левенгук сосредоточился на изготовлении линз, способных к увеличению до 300 раз. В них он рассматривал мельчайшие микроорганизмы, описывая полученную информацию и перенося увиденное на бумагу.
В 1676 году Левенгук обнаружил и изложил сведения о микроскопических существах, которым дал название «анималькули».
Чем питаются
Мельчайшие микроорганизмы существовали на Земле задолго до появления человека. Они имеют повсеместное распространение, питаясь органической пищей и неорганическими веществами.
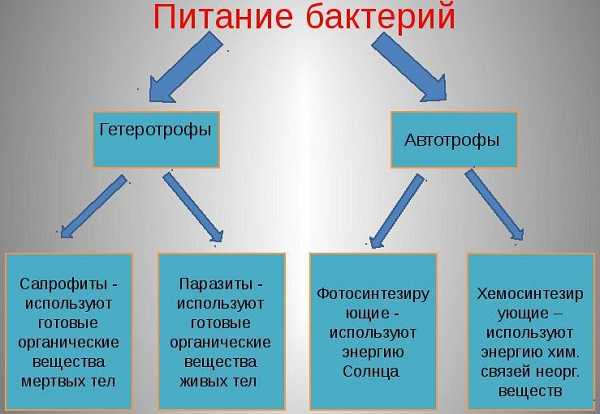
По способам усвоения питательных веществ бактерии принято делить на автотрофные и гетеротрофные. Для существования и развития гетеротрофы используют отходы жизнедеятельности, органического разложения живых организмов.
Представители бактерий
Биологами выделено около 2500 групп различных бактерий.

По форме их подразделяют на:
- кокки, имеющие шарообразные очертания;
- бациллы – в форме палочки;
- вибрионы, имеющие изгибы;
- спириллы – спиральной формы;
- стрептококки, состоящие из цепочек;
- стафилококки, образующие грозди, напоминающие виноградные.
По степени влияния на организм человека прокариотов можно разделить на:
- полезные;
- вредные.
К опасным для человека микробам относятся стафилококки и стрептококки, вызывающие гнойные заболевания.
Полезными считаются бактерии бифидо, ацидофилус, стимулирующие иммунитет и защищающие желудочно-кишечный тракт.
Как размножаются настоящие бактерии
Размножение всех видов прокариотов происходит в основном делением, с последующим ростом до исходной величины. Достигая определенного размера, взрослый микроорганизм распадается на две части.
Реже воспроизведение себе подобных одноклеточных выполняется почкованием и коньюгацией. При почковании на материнском микроорганизме вырастает до четырех новых клеток, с последующим отмиранием взрослой части.
Коньюгация считается простейшим половым процессом у одноклеточных. Чаще таким способом размножаются бактерии, обитающие в животных организмах.
Бактерии симбионты
Микроорганизмы, участвующие в пищеварении в кишечнике человека, это яркий пример бактерий симбионтов. Впервые симбиоз был открыт голландским микробиологом Мартином Виллемом Бейеринком. В 1888 году он доказал взаимовыгодное тесное сожительство одноклеточных и растений бобовых.

Обитая в корневой системе, симбионты, питаясь углеводами, снабжают растение атмосферным азотом. Таким образом, бобовые повышают плодородие, не обедняя почву.
Известно множество успешных симбиотических примеров с участием бактерий и:
- человека;
- водорослей;
- членистоногих;
- морских животных.
Микроскопические одноклеточные оказывают помощь системам человеческого организма, способствуют очищению сточных вод, участвуют в круговороте элементов и работают на достижение общих целей.
Почему бактерии выделяют в особое царство
Для этих организмов характерны мельчайшие размеры, отсутствие оформленного ядра и исключительное строение. Поэтому, несмотря на внешнее сходство, их нельзя отнести к эукариотам, обладающим оформленным клеточным ядром, ограниченным от цитоплазмы оболочкой.
Благодаря всем особенностям в XX веке ученые выделили их в отдельное царство.
Самые древние бактерии
Мельчайшие одноклеточные считаются первой зародившейся жизнью на Земле. Исследователи в 2016 году обнаружили в Гренландии сохранившиеся в погребенном состоянии цианобактерии возрастом около 3,7 миллиарда лет.

В Канаде найдены следы микроорганизмов, живших примерно 4 миллиарда лет назад в океане.
Функции бактерий
В биологии между живыми организмами и средой обитания бактерии выполняют следующие функции:
- переработка органических веществ в минеральные;
- фиксация азота.
В жизни человека одноклеточные микроорганизмы играют важную роль с первых минут рождения. Они обеспечивают сбалансированную микрофлору кишечника, оказывают влияние на иммунитет, занимаются поддержанием водно-солевого баланса.
Запасное вещество бактерий
Запасные питательные вещества у прокариота скапливаются в цитоплазме. Их накапливание происходит в благоприятных условиях, а потребляется в период голодания.
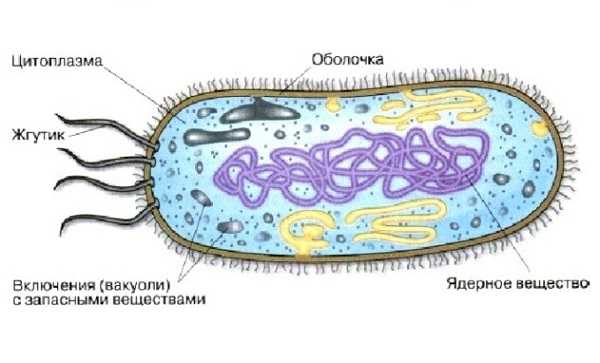
К запасным веществам бактерий относятся:
- полисахариды;
- липиды;
- полипептиды;
- полифосфаты;
- отложения серы.
Главный признак бактерий
Функцию ядра у прокариота выполняет нуклеоид.
Поэтому главным признаком бактерий является сосредоточение наследственного материала в одной хромосоме.
Почему представителей царства бактерии относят к прокариотам
Отсутствие оформленного ядра послужило причиной отнесения бактерий к прокариотным организмам.
Как бактерии переносят неблагоприятные условия
Микроскопические прокариоты способны длительное время переносить неблагоприятные условия, превращаясь в споры. Происходит потеря воды клеткой, значительное уменьшение объема и изменение формы.

Споры становятся нечувствительны к механическим, температурным и химическим воздействиям. Таким образом сохраняется свойство жизнеспособности и осуществляется эффективное расселение.
В какой среде обитают паразитические бактерии
Паразитические микробы служат возбудителями заболеваний в живых существах, питаясь за счет организма-хозяина. Средой обитания болезнетворных микроорганизмов является слюна нездорового человека, предметы, с которыми он соприкасался, воздух в помещении.
Паразитические прокариоты могут пребывать в растениях и их плодах.
Заключение
Бактерии – древнейшая форма жизни на Земле, известная задолго до появления человека. Они присутствуют повсеместно: в окружающем воздухе, воде, в поверхностном слое земной коры. Местом обитания служат растения, животные, человек.
Активное изучение одноклеточных началось в XIX веке и продолжается по сей день. Данные организмы являются основной частью повседневной жизни людей и оказывают непосредственное влияние на существование человека.
1001student.ru
Вопрос 2. Запасные вещества бактерий и их назначение — МегаЛекции
Зап.вещ.представлены полисахаридами, липидами, полипептидами, полифосфатами, отложениями серы. 1.Полисахариды:гликоген, крахмал и крахмалоподобное вещество-гранулеза(специфический запасной полисахарид клостридиев). Названные полисахариды построены из остатков глюкозы.Назначение:в неблагоприятных условиях они используются в качестве источника углерода и энергии.
2.Липиды:накапливаются в виде гранул,например,поли-b-оксимасляная к-та.У некоторых бактерий,поли-b-оксимасляная кислота составляет до 70 % сухого вещества клеток. Отложение липидов в клетке происходит в условиях, когда среда богата источником углерода и бедна азотом.
3.Полифосфаты:содержащиеся в гранулах, называемых волютиновыми, или метахроматиновыми зернами. Назначение:как источник фосфора. Полифосфаты содержат макроэргические связи и, таким образом, являются депо энергии.
4.Цианофицин-специфическое запасное вещ. цианобактерий.Они состоят из полипептида, содержащего Arg и Asp.Назначение: резерв азота, мобилизуемым при его недостатке в среде.
5.Сера:упрокариот, метаболизм которых связан с соединениями серы. Сера накапливается, когда в среде содержится сероводород, и окисляется до сульфата, когда весь сероводород среды оказывается исчерпанным. Назначение:для аэробных тионовых бактерий, окисляющих Н2S,сера служит источником энергии, а для анаэробных фотосинтезирующих серобактерий является донором электронов.
Обращает на себя внимание тот факт, что все запасные вещества представлены в виде высокомолекулярных полимерных молекул, в ряде случаев отграниченных от цитоплазмы белковой мембраной, т.е. находятся в осмотически неактивном состоянии. Это важно, так как в противном случае сосредоточение в цитоплазме большого числа молекул осмотически активных веществ оказало бы на клетку отрицательное действие.
Вопрос 3. Методы определения подвижности бактерий
Подвижность бактерий определяют микроскопией препаратов в «раздавленной» или «висячей» капле. 2)Способность к движению можно определять также после внесения культуры бактерий уколом в столбик полужидкого агара (среда Пешкова) (Неподвижные бактерии растут строго по уколу, а подвижные дают диффузный рост(«забивают» укол).
Посевом бактерий в водный конденсат скошенного столбика агара (подвижные виды переплывают из конденсата на поверхность среды и колонизируют её).
Билет № 19
Вопрос 1. Факторы патогенности бактерий.
Патогенность и вирулентность микроорганизмов
Чтобы возникла инфекционная болезнь, необходимо наличие возбудителя, обладающего патогенностью вообще и вирулентностью в частности.Патогенность микроба — видовой генетический признак, его потенциальная возможность вызвать при благоприятных условиях инфекционный процесс. По этому признаку все существующие мо подразделяют на патогенные, условно-патогенные и сапрофиты. Фактически все возбудители инфекционных болезней являются патогенными, но далеко не все из них способны вызвать инфекционную болезнь, чтобы это произошло,мо, хотя и принадлежащий к патогенному виду, должен обладать вирулентностью.Поэтому патогенностью НЕ равна вирулентности.
Факторы патогенности с токсической функцией:
Среди токсинов микробного происхождения различают:1)экзотоксины-высокоактивные яды(обычно белки-ферменты), выделяемые МО на протяжении его жизни в качестве продуктов обмена в окружающую среду (организм животного, пробирка с культурой микроба).2)эндотоксины-менее ядовитые по сравнению с экзотоксинами вещества, образующиеся в результате распада микробной клетки.Т.е эндотоксины-фрагменты или отдельные химические компоненты микробных клеток. Экзотоксины в основном образуют Гр+(возбудители ботулизма, столбняка, газовой инфекции и др.), а эндотоксины образуют Гр-(сальмонеллы, кишечная палочка, протей и др.).
Основные факторы патогенности:
-способность МО к колонизации(прежде всего патогенному МО нужно прикрепится к клетке,это обусловлено наличием микробных продуктов адгезинов,которык могут быть в составе бактер.клетки или в составе микроворсинок или капсул)
-их устойчивость к разным микробицидным факторам организма,
-свойства инвазивности и токсигенности
-способность к длительному персистированию.
А-Токсины-важнейшие факторы(фосфолипазы,детергенты,цитолизины) патогенности, вырабатываемые МО и реализующие основные механизмы инфекционного процесса. Роль микробных токсинов в патогенезе инфекционных болезней впервые доказали Э. Ру и А. Иерсён (1888), отделившие «ядовитое начало» возбудителя дифтерии от бактериальных клеток и сумевшие воспроизвести с его помощью клиническую картину болезни.
По степени связи с бактериальной клеткой экзотоксины разделяют на:.
Группа А экзотоксинов — токсины, секретируемые во внешнюю среду (токсин дифтерийной палочки).
Группа В экзотоксинов — токсины, частично секретируемые во внешнюю среду и частично ассоциированные с бактериальной клеткой (тетаноспазмин столбнячной палочки).
Группа С экзотоксинов — токсины, связанные с бактериальной клеткой и высвобождающиеся после её гибели {например, экзотоксины энтеробактерий).
По механизму действия экзотоксины:- цитотоксины(энтеротоксины или дерматонекротоксины), -мембранотоксины(гемолизины и лейкоцидины),-функциональные блокаторы(холероген), -эритрогенины. Нередко патогенные бактерии синтезируют несколько экзотоксинов.
Эндотоксины — интегральные компоненты клеточной стенки Гр- бактерий; большая их часть высвобождается только после гибели бактериальной клетки. Представлены комплексом протеинов, липидных и полисахаридных остатков. За проявление биологического эффекта ответственны все группировки молекулы эндотоксина.
Эндотоксинемия обычно сопровождается лихорадкой, обусловленной выбросом эндогенных пирогенов из гранулоцитов и моноцитов. При попадании значительного количества эндотоксина в кровоток возможен эндотоксиновый шок, обычно заканчивающийся смертью больного.
Б-Экзоферменты(лецитиназа, гиалуронидаза, коллагеназа и др.), нарушающие гомеостаз клеток и тканей, что приводит к их повреждению.
В-Суперантигены-токсины способные вызывать поликлональную активацию различных клонов лимфоцитов. Поликлональная активация сопровождается гиперсекрецией лимфокинов с развитием цитокинопосредованной интоксикации.
Г-Капсула
Капсула (или её менее выраженный аналог — слизистый слой) ингибирует начальные этапы защитных реакций — распознавание и поглощение.
• Капсулы «экранируют» бактериальные структуры, активирующие систему комплемента, а также структуры, распознаваемые иммунокомпетентными клетками.
• Гидрофильность капсул затрудняет их поглощение фагоцитами, а само капсульное вещество защищает бактерию от действия лизосомальных ферментов и токсичных оксидантов, выделяемых фагоцитирующими клетками.
• Большое значение имеет лёгкая отделяемость капсул или слизистого слоя от поверхности бактерий. В частности, при поглощении капсулированных бактерий (например, синегнойной палочки), последние легко «снимают с себя» капсулы и избегают прямого контакта с фагоцитом.
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
megalektsii.ru
Запасные вещества прокариот
К внутрицитоплазматическим включениям бактериальной клетки относятся и запасные вещества – полифосфаты, полисахариды, поли--оксимасляная кислота и отложения серы, В условиях голодания запасные вещества являются источником энергии и питания бактерий. Запасные вещества накапливаются в цитоплазме при выращивании бактерий на обогащенных питательных средах, при заторможенных темпах роста культуры или при переходе ее в стадию покоя. В клетках бактерий может накапливаться одно запасное вещество или несколько разных по химической природе веществ. Например, клетки Escherichia coli предпочтительнее запасают гликоген, a Chromatium okenii способен накапливать полифосфаты. Аналогом крахмала у представителей рода Clostridium является вещество гранулеза. В большинстве случаев запасные вещества отграничены от цитоплазмы белковой оболочкой или находятся в виде высокомолекулярных полимеров.
Эндоспоры и другие покоящиеся стадии бактерий.
Эндоспоры – особый тип покоящихся клеток фирмакутных, т.е. бактерий с грамположительным типом оболочки, хотя некоторые образующие споры виды могут по методу Грама окрашиваться отрицательно.
К настоящему времени описано более 15 родов бактерий, образующих эндоспоры. Эти организмы различаются как морфологически, так и по физиологии и экологии. Образование спор является приспособлением, необходимым для существования соответствующих видов, так как позволяет переносить неблагоприятные условия среды.
Одна клетка образует одну спору, увеличения числа организмов при этом не происходит, поэтому эндоспорообразование не способ размножения бактерий. Эндоспоры представляют собою стадию покоя и приспособлены к перенесению неблагоприятных условий.
Способность к образованию спор определяется наличием специальных генов, комплекс которых иногда определяют как спорулон, хотя они не оформлены в виде какой-либо определенной структуры, но несколько их групп-оперонов расположены в различных участках хромосомы.
Для начала спорообразования необходимо завершение репликации хромосомы, причем сигнал к спорообразованию должен быть получен еще в период репликации. Если репликация заканчивается в богатой среде, наступает вегетативное деление. В процессе образования споры хромосома не реплицируется, таким образом, в спору попадает одна или несколько полностью реплицированных хромосом.
Процесс спорообразования принято подразделять на 6 последовательных стадий, смена которых определяется активированием соответствующих генов. При этом один из продуктов работающего по данной стадии оперона служит индуктором активности последующих генов, выключение любого этапа делает невозможными последующие. Депрессия споровых генов до 3-й стадии спорообразования (образование проспоры) обратима, и изменение среды может привести к их репрессии и переходу клетки к вегетативному росту.
Процесс образования споры довольно продолжителен, у аэробных мезофильных бацилл он занимает около 8 ч.
Стадия 1 характеризуется, прежде всего, необычным поведением ДНК, образующей осевой тяж вдоль клетки. У видов Bacillus осевой тяж проходит через всю клетку и включает в себя всю ДНК клетки, тогда как у некоторых видов Clostridium он образуется только частью содержащейся в клетке ДНК и располагается на одном из полюсов клетки, занимая лишь незначительную часть ее длины.
Конденсацию ДНК в относительно компактную структуру в процессе спорообразования связывают с изменением активности генома, с прекращением репликации хромосомы, транскрипции и трансляции вегетативных генов и потерей ДНК связи с рибосомами и мембраной. Показано, что на первых двух стадиях спорообразования активность генома низка.
Цитоплазма спорогенной зоны обычно не содержит включений запасных веществ.
Стадия 2 характеризуется отделением от осевого тяжа ДНК участка, соответствующего хромосоме, который движется к полюсу клетки. Затем часть цитоплазмы клетки с заключенной в нее хромосомой отделяется цитоплазматической мембраной, врастающей так же, как при клеточном делении, но здесь оболочка клетки участия в делении не принимает. В результате образуются две разделенные мембраной клетки, неравные по размерам. В это время процесс спорообразования еще обратим. Если бактерии перенести в условия, благоприятные для вегетативного роста, деление будет завершено слоями оболочки, и образуются две вегетативные клетки разной длины. Обе в дальнейшем могут делиться, хотя меньшая клетка медленнее.
События, происходящие на второй стадии спорообразования, определяются генами, по крайней мере 8 оперонов, расположенных в разных участках хромосомы.
Между 2-й и 3-й стадиями происходит врастание двойной мембраны или септы так, что в результате образуется округлая проспора, окруженная двумя мембранами. У некоторых видов в этом процессе принимает участие мезосома. Проспора в конце концов совершенно отделяется от мембраны материнской клетки и как бы свободно плавает внутри нее. На этом этапе происходит резкое изменение свойств мембран проспоры, на срезах они окрашиваются менее контрастно, чем цитоплазматическая. Считают, что в этот момент происходит изменение их транспортных свойств. Процесс спорообразования становится необратимым, причем не только для проспоры, но и для материнской клетки, которая уже не может вернуться к вегетативному росту.
Проспора остается у полюса клетки или, у некоторых видов, перемещается к ее центру. В процессе образования проспоры две окружающие ее мембраны оказываются расположенными зеркально, т.е. внешний, ранее обращенный к среде лепесток внутренней мембраны обращен наружу, а у внешней мембраны – внутрь. При этом, вероятно, эффективный транспорт веществ внутрь проспоры из цитоплазмы материнской клетки может обеспечиваться только за счет пассивной или облегченной диффузии.
После завершения образования проспоры бактерия начинает интенсивно потреблять из среды ионы Са2+, которые в конце концов могут составить до 2% от сухой массы споры и в молярном выражении превысить сумму всех других находящихся в споре катионов. Поступление Са2+ в спорангии, т.е. в спорулирующую клетку, происходит в результате энергозависимого активного транспорта, но из цитоплазмы спорангия в проспору он поступает за счет диффузии. Внутри проспоры образуется комплекс Са с дипиколиновой кислотой (ДПК), так что концентрация свободного Са2+ в проспоре оказывается низкой и не препятствует его диффузии из клетки.
На 4-й стадии между двумя мембранами проспоры начинается образование муреинового слоя. Процесс идет в 2 стадии. Сначала образуется примордиальный (зачаточный) слой, который после прорастания споры станет оболочкой проростка. Затем над ним откладывается несколько видоизмененный муреин, образующий толстый слой коры или кортекса. Синтез вещества кортекса является более кислым, чем муреин оболочки, за счет того, что многие остатки мурамовой кислоты не содержат пептидов или соединены лишь с 1–3 аминокислотами, которые не образуют сшивок и, следовательно, имеют свободные карбоксильные группы.
По мере развития проспоры происходит усиление ее способности к светопреломлению, вероятно, связанной с обогащением споры комплексом ДПК-Са. В фазово-контрастном микроскопе проспора выглядит серой. Способность к светопреломлению является характерной особенностью зрелой бактериальной споры.
На стадиях 3 и 4 проспора, выделенная из спорангия, осмотически нестабильна и нежизнеспособна.
На 5-й стадии спорообразования у бацилл начинают формироваться покровы споры. На срезах они сначала имеют вид отдельных коротких электронно-плотных линий или пластинок, расположенных в цитоплазме спорангия в 20–30 нм от наружной мембраны проспоры. Пластинки затем разрастаются и сливаются, образуя непрерывные слои вокруг споры. Внешняя мембрана проспоры становится плохо заметной на срезах, хотя, видимо, сохраняется в дальнейшем.
На 5-й стадии продолжается накопление спорой ДПК и Са, повышается устойчивость ее к органическим растворителям, например октанолу, хлороформу, спора становится светлой в фазовом контрасте. Она уже осмотически стабильна, но сохраняет термочувствительность. Выделенные из спорангия проспоры могут прорастать, при этом их прорастание индуцируется теми же индукторами, что и прорастание зрелых спор, но не наблюдается термоиндукции прорастания.
На 6-й стадии спорообразования заканчивается формирование всех характерных структур споры, она приобретает термоустойчивость. Иногда выделяют еще 7-ю стадию, на которой происходит лизис спорангия и выход споры в окружающую среду. При этом материнская клетка, или спорангий, до самого последнего момента сохраняет активность. Развитие споры определяется активностью двух геномов – материнской клетки и проспоры. Геном материнской клетки определяет синтез ДПК, белков, покровов споры, экзоспориума, выростов и газовых вакуолей у спор, содержащих эти структуры. От активности генома материнской клетки зависит образование ассоциированных со спорообразованием протеаз, антибиотиков и токсинов, а так же литических ферментов, образующихся на последнем этапе спорообразования. Определенная степень координации метаболизма материнской клетки сохраняется вплоть до ее лизиса.
Строение зрелой споры у разных видов соответствует одной и той же схеме. Сердцевина споры содержит белки и нуклеиновые кислоты, на их долю приходится не более 50–60% сухой массы споры, остальную ее часть составляют ДПК и другие низкомолекулярные вещества. Спора содержит так же рибосомы и различные ферменты, большинство из которых ничем не отличаются от соответствующих ферментов вегетативной клетки. Липиды вегетативной клетки и споры одинаковы, хотя количественное содержание определенных жирных кислот может быть различным.
Сердцевина споры окружена цитоплазматической мембраной, липиды которой находятся в упорядоченном кристаллическом состоянии геля. В их стабилизации, возможно, принимают участие ионы Са2+. Как уже говорилось, над мембраной находится примордиальный муреиновый слой, а затем толстый слой кортекса. Последний занимает 13–60% от объема споры, хотя образующий его муреин составляет всего 9–15% от массы споры. Это несоответствие объясняют высокой обводненностью кортекса. Даже довольно значительное его уменьшение может заметно не повлиять на свойства споры, но имеется определенный минимум массы кортекса, ниже которого спора теряет термоустойчивость
На срезах споры кортекс выглядит как массивный электронно-прозрачный слой, у покоящихся спор гомогенный, у прорастающих или разрушающихся – фибриллярный.
Как уже было отмечено, муреин кортекса является кислым за счет того, что только незначительная часть остатков мурамовой кислоты несет пептиды, часть занята только остатками L-аланина, а часть не соединена с аминокислотой и образует лактам.
Видимо, лактам, пептид и аланин чередуются в молекуле муреина. Состав пептидов кортекса может несколько отличаться от состава пептидов муреина оболочки. Длина нитей полисахаридной части муреина кортекса больше, чем у вегетативных клеток, и соответствует 80–100 дисахаридным остаткам. Синтез мурамового лактама начинается одновременно с синтезом ДПК.
Кортекс и зародышевая стенка кроме муреина содержат ряд литических ферментов, которые активируются при прорастании или при механическом нарушении целостности споры.
Снаружи кортекс ограничен внешней мембраной проспоры. Затем следуют покровы споры. Между мембраной и покровами находятся захваченные при спорообразовании участки цитоплазмы материнской клетки.
Над кортексом споры одеты слоистыми покровами, число и строение которых у разных видов бывают различны. У многих видов покровы дифференцированы на три слоя: внутренний, средний и внешний. Каждый из этих слоев в свою очередь может состоять из нескольких чередующихся электронно-плотных и электронно-прозрачных слоев. При этом внешний покров часто структурирован.
Покровы споры состоят из белка обычно с небольшой примесью углеводов и липидов, иногда содержат много фосфора.
У зрелой споры выявляются три основных слоя покровов, но они имеют комплексное строение. Внешний слой содержит белок, синтезированный одним из первых, и белок, синтезируемый на поздних стадиях спорообразования. Именно внешний покров обеспечивает устойчивость спор к лизоциму. Покровы споры обеспечивают ее устойчивость к лизоциму и другим ферментам, органическим растворителям, поверхностно активным соединениям, придают споре гидрофобность. Покровы чувствительны к щелочам, мочевине, сильным детергентам, например додецилсульфату. При нарушении целостности покровов сохраняется термоустойчивость и радиоустойчивость споры и ее способность к светопреломлению. При обработке крепкой кислотой спора разрывается и ее содержимое выходит. При этом, видимо, происходит освобождение Са2+ из его комплекса с ДПК, и спора разрывается за счет создавшегося высокого внутреннего осмотического давления.
Экзоспориум образует как бы свободный мешок, внутри которого лежит спора, контактируя с ним только в некоторых участках. Обычно в его состав входят белки, липиды, углеводы. Базальный слой экзоспориума образован белком и формируется в результате самосборки белковых молекул.
Экзоспориум, видимо, никогда не бывает полностью замкнутым, и ограниченная им цитоплазма сообщается с остальной частью цитоплазмы спорангия. Вместе с тем экзоспориум создает определенную компартментализацию цитоплазмы.
У многих видов рода Clostridium споры снабжены различными придатками и выростами. По классификации, предложенной В.И. Дудой, можно выделить 6 типов и 16 разновидностей выростов спор Clostridium. 1-й тип – трубчатые выросты, которые могут иметь капсулярный слой, латеральные разветвления или шляпки; 2-й тип – лентовидные выросты; 3-й тип – булавовидные выросты, располагающиеся на споре по одному или несколько перитрихиально; 4-й тип – фибриллярные тяжи; 5-й тип – волосовидные выросты, расположенные пучками на полюсе споры или по всей поверхности споры; 6-й тип – древовидные выросты. В составе выростов обнаружены белки, полисахариды, липиды. Предполагают, что выросты являются одним из средств пространственной организации (компартментализации) процессов в спорулирующих клетках.
Споры иногда окрашены и в массе выглядят желтыми или коричневыми. Споры не окрашиваются при использовании обычных методов, принятых в светооптической микроскопии, но без труда фиксируются методами, принятыми в электронно-микроскопических исследованиях. Характерной особенностью спор является способность к светоприломлению. Индекс светопреломления споры (около 1,55) близок к таковому сухого белка. Споры могут содержать до 79% воды, многократно высказывалось мнение, что эта вода сосредоточена только в районе кортекса, тогда как сердцевина обезвожена, имеются, однако, наблюдения, свидетельствующие о равномерном распределении воды в спорах.
Некоторые бактерии одновременно со спорами образуют параспоральные тела, которые не являются элементами спор или нормальными компонентами бактериальной клетки.
Бактериальные споры обладают уникальной устойчивостью к ряду неблагоприятных воздействий. Широко известная высокая устойчивость спор к температуре может быть различной и зависит от температуры, при которой происходит вегетативный рост соответствующего вида.
Повышенную устойчивость спор к ферментам, ядам, органическим растворителям можно в основном объяснить барьерной ролью белковых покровов споры.
Процесс прорастания споры принято подразделять на три стадии – активацию, инициацию и вырастание.
Активация означает готовность споры к прорастанию, при этом сохраняется ее устойчивость к температуре, способность к светопреломлению и др. Активация происходит в процессе старения спор, и тогда она необратима. Активация в сильной степени зависит от температуры. Активация может быть достигнута снижением величин рН, воздействиями редуцирующих агентов, в этих же случаях возможна деактивация.
Инициация прорастания необратима и длится в течение нескольких минут, при этом наблюдается определенная последовательность событий. Сначала снижается устойчивость споры к прогреванию, выход в среду ДПК-Са, спора становится темной в фазовом контрасте, хорошо окрашивается и, наконец, в среду начинает освобождаться глюкозамин и ДАП из растворяющегося кортекса. Происходит снижение светорассеяния суспензий спор. Процесс инициации зависит от температуры, влажности, величины рН раствора, но может происходить в условиях, исключающих вегетативный рост. Инициация нечувствительна к действию антиметаболитов, не зависит от присутствия источников азота, фосфора. Она может быть вызвана неспецифическими механическими воздействиями, обработкой спор ультразвуком, растиранием со стеклянными бусами, воздействием давлением несколько сот атмосфер. Подобная индукция инициации не является естественной и, вероятно, не имеет существенного биологического значения. Большой интерес представляет индукция инициации, вызываемая специфическими индукторами. Индукторами могут быть различные органические и неорганические вещества, очень часто некоторые аминокислоты.
Оказалось, что при спорообразовании в проспоре накапливается белок с молекулярной массой 46000, который в процессе созревания споры превращается в белок с молекулярной массой 4100, и, наконец, при прорастании споры в белок с молекулярной массой 40000, который и обнаруживает активность. Превращение белка с массой 41000 в белок с массой 40000 не требует затраты энергии, но дальнейшая деградация специфической протеазы – энергозависимый процесс.
Вырастание – это процесс активного роста, однако имеются некоторые отличия процессов, происходящих при вырастании и вегетативном росте. При вырастании идет активный синтез белка и РНК, но репликация ДНК начинается только через 1–2 ч после начала прорастания споры. Сначала происходят процессы репарации повреждений ДНК, произошедших в период покоя споры.
В процессе прорастания споры происходит лизис ее оболочек или их разрыв и выход проростка из оболочек. Тот или иной способ прорастания характерен для данного вида.
Образование эндоспор у некоторых видов может быть связано с прохождением бактерией специализированного жизненного цикла. Так, эндоспоры, одновременно являющиеся инфицирующей стадией организма, образуют бактериальный паразит нематод. Округлая спора находится внутри экзоспориума, имеющего чашевидную форму, и окружена кольцом фибриллярного материала. Плоская сторона этого комплекса прикрепляется к кутикуле нематоды, после чего спора прорастает в трубке, проходящей внутрь тела животного, где формируется примитивный мицелий. Отдельные клетки мицелия, сильно разрастаясь, превращаются в спорангии, в которых формируются споры. После лизиса зараженной нематоды споры оказываются в почве, где заражают новых животных. Из одной умершей нематоды может освободиться до 2 млн. спор.
По расположению в клетке выделяют типы спор. Бациллярный –спора располагается в центре клетки, клостридиальный – спора смещена от центра к одному из полюсов, плектридиальный – спора располагается полярно, приобретая вид барабанной палочки.
Кроме эндоспор у бактерий могут обнаруживаться также другие типы покоящихся стадий. Цианофиты, спирохеты, метилотрофные бактерии способны образовать цисты, содержащие вдвое больше липидов, чем вегетативные клетки. Некоторые фотосинтезирующие и метанолокисляющие бактерии образуют экзоспоры. Одна вегетативная клетка способна сформировать до 4 экзоспор.
studfiles.net
16. Морфология бактерий. Запасные включения бактериальной клетки.
Цитоплазматические включения разнообразны. Это могут быть питательные вещества. Откладываются в клетке, когда она развивается в условиях избытка питательных веществ в среде, и потребляются, когда клетки попадают в условия голодания.
|
Запасное вещество |
Структурные характеристики |
Химический состав |
Функции |
Распространение |
|
Гранулы гликогена (а-гранулы) |
Сферической формы, диаметр 20–100 нм |
Высокомолекулярные полимеры глюкозы |
Источник углерода и энергии |
Широко распространенный тип запасных веществ |
|
Гранулы поли-p-оксимасляной кислоты (до 50 % сухого вещества клетки) |
Диаметр 100–1000 нм; окружены однослойной белковой мебраной 2–3 нм толщиной |
98 % полимера поли-3- оксимасляной кислоты, 2 % белка |
Источник углерода и энергии |
Широко распространены только у прокариот |
|
Гранулы или капли жира |
В жидком состоянии в виде капель |
Триацилглицериды |
Источник углерода и энергии |
Широко распространенный тип запасных веществ |
|
Гранулы полифосфата |
Диаметр 500 нм, зависит от объекта и условий роста |
Линейные полимеры ортофосфата |
Источник фосфора и энергии |
Распространенный тип запасных гранул |
|
Запасное вещество |
Структурные характеристики |
Химический состав |
Функции |
Распространение |
|
Цианофициновые гранулы |
Размер и форма различны; могут достигать в диаметре 500 нм |
Полипептид, содержащий аргинин и аспарагиновую кислоту (1: 1), М=(25–100)·103Да |
Источник азота |
Обнаружены у многих видов цианобактерий |
|
Гранулы серы |
Диаметр 100–800 нм; окружены мембраной |
Включения жидкой серы |
Донор электронов или источник энергии |
Пурпурные серобактерии, бесцветные бактерии, окисляющие h3S |
|
Углеводородные гранулы |
Диаметр 200–300 нм; окружены белковой оболочкой 2–4 нм толщиной |
Углеводороды того же типа, что и в среде |
Источник углерода и энергии |
Представители родов Arthrobacter, Acinetobacter, Мусоbacterium, Nocardia и другие прокариоты, использующие углеводороды |
*Аэросомы – газовые вакуоли, имеющиеся у 15 таксономических групп почвенных и водных бактерий в цитоплазме. Аэросомы – это сложно организованные структуры, состоящие из множества регулярно расположенных газовых пузырьков, имеющих форму вытянутого цилиндра с заостренными концами длиной 200-1200 и диаметром 65-115 нм. Пузырьки окружены белковой мембраной и заполнены азотом, количество их может достигать 40–60 шт. Мембрана аэросом проницаема для газов, но не для воды. Регулируя степень наполнения вакуолей газом, клетка осуществляет механическое передвижение снизу вверх в водоемах или капиллярах почвы. Аэросомы присущи безжгутиковым формам и являются альтернативным видом движения.
*Хлоросомы – структуры, присущие фотосинтезирующим зеленым бактериям, локализующие пигменты – хлорофиллы с, d и е, поглощающие кванты света и передающие их в реакционные центры клетки. Хлоросомы имеют форму продолговатых пузырьков длиной 90-120 и шириной 25-70 нм, окруженных однослойной белковой мембраной толщиной 2-3 нм. Хлоросомы расположены вблизи от ЦПМ.
*Фикобилисомы – структуры цианобактерий, расположенные правильными рядами на внешних поверхностях фотосинтетических мембран, локализуют фикобилипротеины, имеют вид гранул диаметром 28-55 нм.
* Карбоксисомы – полиэдральные тела клеток некоторых фототрофных и хемолитотрофных эубактерий. Карбоксисомы имеют форму многогранника с 4-6 сторонами и диаметром 90-500 нм.
Структуры белковой природы толщиной 3 нм заполнены гранулярным содержимым, включающим рибулозодифосфаткарбоксилазу, принимают участие в восстановительном пентозофосфатном цикле.
Карбоксисомы обеспечивают защиту и консервирование ключевого фермента рибулезофосфатного метаболического пути, предотвращая его разрушение внутриклеточными протеазами.
* Магнитосомы – цитоплазматические включения магнитных бактерий, заполненные железом в форме магнетита (Fe3O4) и выполняющие функцию магнитной стрелки. На долю магнетита может приходиться до 4 % сухого вещества таких бактерий. Магнитосомы для бактерий имеют приспособительное значение.
studfiles.net
Ответы@Mail.Ru: царство бактерий общая характеристика
Царство бактерий. Общая характеристика.Известно около 2500 видов. Имеют клеточное строение, но не имеют ядра, отделенного мембраной от цитоплазмы. Бактерии по форме бывают: шаровидные (кокки) , палочковидные (бациллы) , изогнутые (вибрионы) , спиральные (спириллы) , в виде цепочки (стрептококки) , в виде гроздей (стафилококки) .
Большинство не содержит хлорофилла и питается готовыми органическими веществами – гетеротрофно. По способу добычи пищи гетеротрофные делятся на три группы: паразиты, сапрофиты и симбионты.
Освоили все среды обитания. Живут практически везде: в почве, в пыли, в воздухе, в воде, на теле животных, внутри живых организмов.
Размножаются каждые 20–30 минут.
Имеют очень важное значение для человека.
1)При жизнедеятельности почвенных бактерий происходит образование гумуса, который представляет собой разложившееся органическое вещество, содержащее все необходимые вещества для жизни растений.
2)Для очистки сточных вод применяются микроорганизмы, которые в короткие сроки могут перевести большинство органических соединений в неорганические.
3)В кишечнике многих животных и человека обитает микрофлора, которая способна переваривать потребляемую организмом пищу и синтезируют витамины (бактерии симбионты) .
4)Путем брожения человек может получать различные вещества, например, уксусная кислота, силос, спирт, кисломолочные продукты.
5)Производство антибиотиков. Эти вещества выделяются некоторыми бактериями и грибами. Они вызывают угнетение жизнедеятельности других бактерий.
6)Производство кормового белка.
7)Производство ферментов и генная инженерия. Возможность промышленно производить инсулин, получать спирты, органические кислоты, полимерные вещества.
8)Биологические методы борьбы с вредителями, различные бактерии могут заражать и вызывать гибель вредителей сельского хозяйства.
http://ebiology.ru/botanika/carstvo-bakterij-obshhaya-xarakteristika/
[ссылка заблокирована по решению администрации проекта]
otvet.mail.ru
16. Морфология бактерий. Запасные включения бактериальной клетки.
Цитоплазматические включения разнообразны. Это могут быть питательные вещества. Откладываются в клетке, когда она развивается в условиях избытка питательных веществ в среде, и потребляются, когда клетки попадают в условия голодания.
|
Запасное вещество |
Структурные характеристики |
Химический состав |
Функции |
Распространение |
|
Гранулы гликогена (а-гранулы) |
Сферической формы, диаметр 20–100 нм |
Высокомолекулярные полимеры глюкозы |
Источник углерода и энергии |
Широко распространенный тип запасных веществ |
|
Гранулы поли-p-оксимасляной кислоты (до 50 % сухого вещества клетки) |
Диаметр 100–1000 нм; окружены однослойной белковой мебраной 2–3 нм толщиной |
98 % полимера поли-3- оксимасляной кислоты, 2 % белка |
Источник углерода и энергии |
Широко распространены только у прокариот |
|
Гранулы или капли жира |
В жидком состоянии в виде капель |
Триацилглицериды |
Источник углерода и энергии |
Широко распространенный тип запасных веществ |
|
Гранулы полифосфата |
Диаметр 500 нм, зависит от объекта и условий роста |
Линейные полимеры ортофосфата |
Источник фосфора и энергии |
Распространенный тип запасных гранул |
|
Запасное вещество |
Структурные характеристики |
Химический состав |
Функции |
Распространение |
|
Цианофициновые гранулы |
Размер и форма различны; могут достигать в диаметре 500 нм |
Полипептид, содержащий аргинин и аспарагиновую кислоту (1: 1), М=(25–100)·103Да |
Источник азота |
Обнаружены у многих видов цианобактерий |
|
Гранулы серы |
Диаметр 100–800 нм; окружены мембраной |
Включения жидкой серы |
Донор электронов или источник энергии |
Пурпурные серобактерии, бесцветные бактерии, окисляющие h3S |
|
Углеводородные гранулы |
Диаметр 200–300 нм; окружены белковой оболочкой 2–4 нм толщиной |
Углеводороды того же типа, что и в среде |
Источник углерода и энергии |
Представители родов Arthrobacter, Acinetobacter, Мусоbacterium, Nocardia и другие прокариоты, использующие углеводороды |
*Аэросомы – газовые вакуоли, имеющиеся у 15 таксономических групп почвенных и водных бактерий в цитоплазме. Аэросомы – это сложно организованные структуры, состоящие из множества регулярно расположенных газовых пузырьков, имеющих форму вытянутого цилиндра с заостренными концами длиной 200-1200 и диаметром 65-115 нм. Пузырьки окружены белковой мембраной и заполнены азотом, количество их может достигать 40–60 шт. Мембрана аэросом проницаема для газов, но не для воды. Регулируя степень наполнения вакуолей газом, клетка осуществляет механическое передвижение снизу вверх в водоемах или капиллярах почвы. Аэросомы присущи безжгутиковым формам и являются альтернативным видом движения.
*Хлоросомы – структуры, присущие фотосинтезирующим зеленым бактериям, локализующие пигменты – хлорофиллы с, d и е, поглощающие кванты света и передающие их в реакционные центры клетки. Хлоросомы имеют форму продолговатых пузырьков длиной 90-120 и шириной 25-70 нм, окруженных однослойной белковой мембраной толщиной 2-3 нм. Хлоросомы расположены вблизи от ЦПМ.
*Фикобилисомы – структуры цианобактерий, расположенные правильными рядами на внешних поверхностях фотосинтетических мембран, локализуют фикобилипротеины, имеют вид гранул диаметром 28-55 нм.
* Карбоксисомы – полиэдральные тела клеток некоторых фототрофных и хемолитотрофных эубактерий. Карбоксисомы имеют форму многогранника с 4-6 сторонами и диаметром 90-500 нм.
Структуры белковой природы толщиной 3 нм заполнены гранулярным содержимым, включающим рибулозодифосфаткарбоксилазу, принимают участие в восстановительном пентозофосфатном цикле.
Карбоксисомы обеспечивают защиту и консервирование ключевого фермента рибулезофосфатного метаболического пути, предотвращая его разрушение внутриклеточными протеазами.
* Магнитосомы – цитоплазматические включения магнитных бактерий, заполненные железом в форме магнетита (Fe3O4) и выполняющие функцию магнитной стрелки. На долю магнетита может приходиться до 4 % сухого вещества таких бактерий. Магнитосомы для бактерий имеют приспособительное значение.
studfiles.net
Запасные гранулы бактерии
Запасные гранулысодержат временный избыток метаболитов; наличие и количество гранул изменяется в зависимости от вида бактерий и их метаболической активности. В виде гранул могут запасаться полисахариды (крахмал, гликоген, гранулёза), жиры (триглицериды, сходные с жирами высших животных, запасаются у дрожжей рода Candida), воска — у микобактерий и нокардий; полимеры р-оксимасляной кислоты (например, в клетках Bacillus megaterium), полифосфаты (волютин) у Spirillum volutans и Corynebacterium diphtheriae, сера (у бактерий, окисляющих сульфиды), кристаллизованные белки (например, токсичный для насекомых протоксин у Bacillus thuringiensis).
Физиология бактерий. Питание бактерий. Тип питания бактерии. Голозои. Голофиты. Вода. Значимость воды для бактерий.
Жизнь бактерий, как и других живых существ, в упрощенном виде сводится к таким последовательным событиям: собственное воспроизведение — обеспечение жизненных функций -> воспроизведение потомства. Нормальная реализация этого круговорота жизни для любого вида бактерий возможна лишь при развитии адекватных ответных реакций на любые воздействия окружающей среды, что во многом обеспечивается генетической информацией организма.
Жизнь бактерий, равно как и всех живых существ, характеризуется ростом и размножением, то есть увеличением живой массы отдельной особи и популяции в целом за счет ассимиляции веществ, находящихся вне клетки. В данном аспекте нет никакой существенной разницы между питанием паразитической бактерии или автотрофной бактерии, использующей, подобно высшим растениям, углекислоту и минеральные вещества. Во всех случаях процесс питания сводится к усвоению экзогенных субстратов и превращению их (через каскад сложных реакций) в вещества, необходимые для нормальной жизнедеятельности бактерий.
Питание бактерий
Пищейпринято называть любое вещество, которое, попав в организм, служит источником энергии или пластическим материалом для синтеза молекул, используемых для нужд организма. Большинство животных, включая человека, способно заглатывать и переваривать плотные частички пищи в основном за счёт их гидролиза. Подобныйтип питанияизвестен какголозойный, а организмы —голозои[от греч. hobs, полноценный, + zoikos, относящийся к животным]. Бактерии не способны захватывать твердофазные объекты, поэтому утилизируют питательные вещества в виде относительно простых молекул из водных растворов. Подобныйтип питания, присущий также всем растениям, известен какголофитный, то есть бактерии —голофиты[от греч. hobs, полноценный, + phytikos, относящийся к растениям]. Тем не менее многие бактерии способны утилизировать твёрдую пищу с помощью так называемого внешнего питания, реализуемого вне клеток, то есть бактериям также присущ иголозойный тип питания.
Для этого они имеют мощный ферментативный потенциал, хотя иногда секретируемые ферментымогут полностью инактивироваться в результате разведения, под действием конвеционных токов и других факторов. Контакт пищеварительных ферментов с экзогенным субстратом приводит к образованию низкомолекулярных продуктов, проникающих через клеточную стенку в цитоплазму. Начиная с этого момента, процессы их усвоения (метаболизма) в растительных и животных клетках протекают удивительно сходно.
Клеткивсех живых существ, от самых примитивных форм до высокоразвитых животных и растений, не только состоят из одних и тех же веществ, но и используют одни и те же механизмы для получения энергии и для роста. По сравнению с этим фундаментальным биохимическим единством существующие различия и отклонения кажутся незначительными.
studfiles.net
