
Водородные связи между азотистыми основаниями
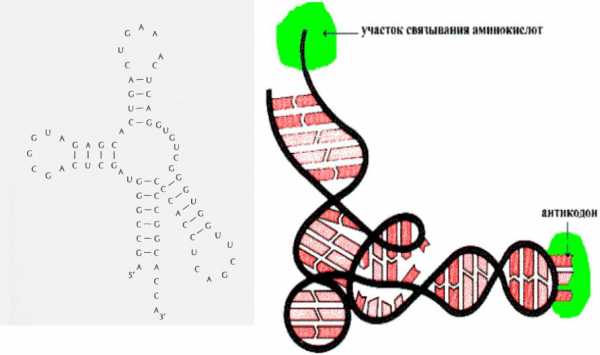
Элементы вторичной структуры РНК— стебель-
петли, петли, псевдоузлы. Двухцепочечные участки РНК

Структурный
анализ молекул РНК
показал, что они состоят не из одной длинной спирали, а из многочисленн ых коротких спиралей, расположенн ых близко друг к другу и образующих
нечто,
похожее на
третичную Вторичная структура молекулы23S-рРНК
структуру
белка.

м-РНК=и-РНК
служит посредником при передаче информации, закодированной в ДНК к рибосомам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка.
Три последовательных нуклеотида (кодон) соответствуют
одной аминокислоте.
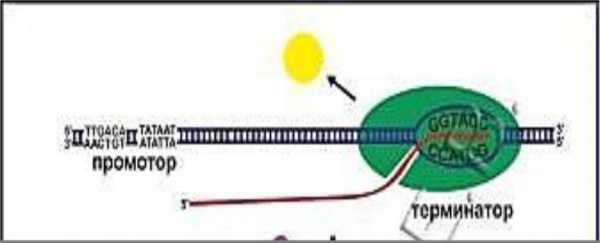
Сигналы, указывающие РНК-полимеразегде начинать транскрипцию и где останавливать
(фланкирующие последовательности нуклеотидов ДНК)
3΄-фланкер
5΄-фланкер
Промотор — сайт | Сайт | |
терминации — | ||
инициации — участок | участок ДНК, | |
ДНК предшествующий | ||
расположенный за | ||
транскрибируемому | ||
гену (5-фланкирующая | транскрибируемым | |
последовательность), | геном (3- | |
содержит пары А-Т | фланкирующая | |
| послед-сть) |
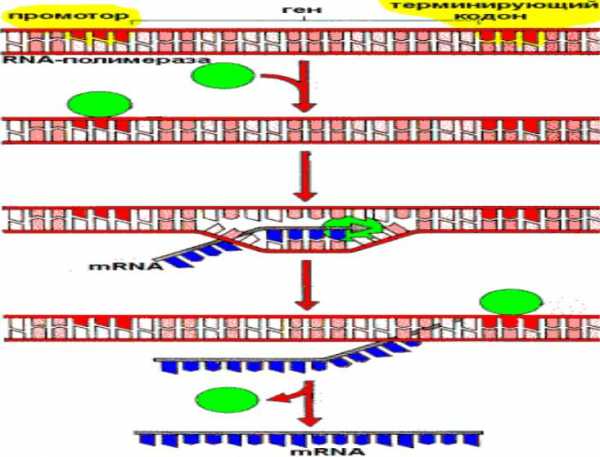
Транскрипция DNA с образованием mRNA
РНК-полимераза– основной фермент транскрипции
studfiles.net
Нуклеотидный состав оснований ДНК разных организмов
| Организмы | Среднее значение содержания ГЦ в % |
| Вирусы | 28-74 |
| Бактерии | 26-74 |
| Водоросли | 36-68 |
| Грибы | 36-54 |
| Высшие растения | 36-48 |
| Простейшие | 22-62 |
| Беспозвоночные | 34-44 |
| Позвоночные | 40-44 |
Для ДНК характерна структура трех видов — первичная, вторичная и третичная. Первичная структура ДНК заключается в том, что ДНК состоит из нуклеотидных цепей, у которых скелетную основу составляют чередующиеся сахарные и фосфатные группы, объединенные ковалентными 3′-, 5′-фосфодиэфирными, скелетными связями, а боковые группы представлены тем или иным основанием (одним из четырех) и присоединяются одна к другой молекулой сахара. Последовательно располагающиеся нуклеотиды ковалентно связаны фосфодиэфирными связями между сахарным остатком и фосфатом, и в результате этого объединены в полинук-леотидную цепь. Таким образом, первичная структура ДНК (как и РНК) определяется последовательностью нуклеотидов и характером их связей между сахарным остатком и фосфатом.
Представления о вторичной структуре ДНК (рис. 107) были сформулированы Д. Уотсоном и Ф. Криком еще в 1953 г. На основе данных об Х-дифракции молекул ДНК, структуре оснований и правил А. Чаргаффа эти представления сводятся к следующему:
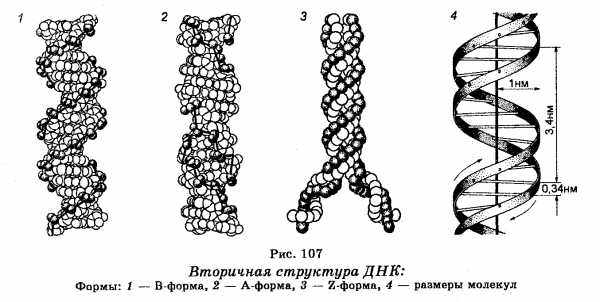
1. Молекула ДНК построена из двух скрученных направо спиралевидных полинуклеотидных цепей, причем каждый виток спирали соответствует 10 азотистым основаниям или расстоянию в 3,4 нм. Молекулы ДНК, цепи которых скручены направо, первоначально назвали В-формой.
2. Обе цепи объединены в результате закручивания одной цепи вокруг другой по общей оси. Из-за противоположной последовательности атомов в каждой цепи обе цепи инвертированы относительно одна другой, т. е. направление вдоль дуплекса есть 3′ ® 5′ для одной цепи и 5′ ® 3′ для другой.
3. Сахарофосфатные группы располагаются на внешней стороне двойной спирали, тогда как основания находятся внутри спирали под прямым углом и вдоль ее оси. Диаметр молекулы составляет 2 нм, расстояния между отдельными азотистыми основаниями в молекуле равны 0,34 нм. Таким образом, ДНК представляет собой скрученную в правостороннем направлении двойную спираль, в которой пары азотистых оснований А—Т и Г-Ц в комплементарных полинуклеотидных цепях подобны перекладинам в лестнице, а сахарофосфатные цепи являются каркасом этой лестницы.
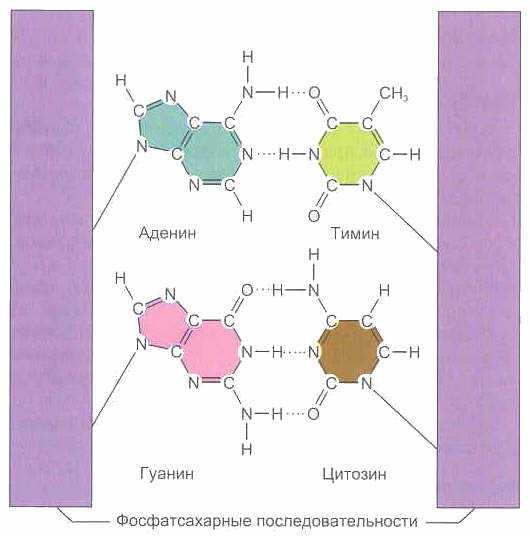
4. Цепи в молекуле не идентичны, но комплементарны и удерживаются слабыми водородными связями между азотистыми основаниями, причем спаривание азотистых оснований для связывания цепей имеет специфический характер. Водородные связи устанавливаются не просто между азотистыми основаниями цепей, а специфически между пуриновым азотистым основанием одной цепи и пиримидиновым азотистым основанием другой. В результате этого аденин одной из цепей связывается с тимином другой цепи двумя водородными связями, тогда как гуанин одной из цепей связывается с цитозином, находящимся в другой цепи, посредством трех водородных связей.
Таблица 12
Свойства разных конформационных форм ДНК
| Свойство | Формы спиралей | |||
| А | В | С | Z | |
| Направление скрученное™ | вправо | вправо | вправо | влево |
| Диаметр молекулы | 23 А̊ | 19 А̊ | 19 А̊ | 18 А̊ |
| Количество оснований в витке | 91/3 |
Дезоксирибозные остатки пар А-Т и Г-Ц разделены одинаковыми расстояниями. Для сахарофосфатных скелетных связей характерна полярность, поскольку фосфат связывает группу 3′-ОН одной дезоксирибозы с группой 5′-ОН другой, тогда как комплементарные цепи имеют противоположную полярность.
Двойная спираль имеет упорядоченный характер, поскольку каждая связь основание-сахар имеет одинаковое расстояние от оси спирали и перевернута на 36°. Как видно, вторичная структура отражает собой форму нуклеиновой кислоты.
Исследования рентгеновской дифракции молекул ДНК показали, что количество оснований в витках закрученной направо спирали может составлять не только 10, как у В-формы, но и 11, а то и ОУз оснований. Эти формы спиралей получили название А- и С-форм. Установлено также, что в молекулах ДНК встречаются районы, цепи в которых закручены налево. Эти районы получили название Z-форм. Различия между А-, В-, С- и Z-формами приведены в табл. 12, однако степень регулярности и конформации Z-формы еще не выяснена.
Степень суперскручивания ДНК зависит от ферментов, в частности от динамического баланса между взаимоантагонистическими ферментами ДНК — гиразой, которая ответственна за суперскручивание и ДНК — топоизомеразой I, которая устраняет суперскручивание.
Третичная структура ДНК связана с трехмерной пространственной конфигурацией молекул и зависит от внутримолекулярных условий. Однако эта структура достаточно еще не изучена.
Размеры молекул ДНК обычно устанавливают определением молекулярной массы в дальтонах и длины в количестве пар оснований. Молекулярная масса пары А-Т составляет 617 дальтон, пары Г-Ц — 618 дальтон. Молекулярная масса 1000 пар азотистых оснований (1 килобаса) составляет 617 500 дальтон или 6,175´105/6,02×1023 г = 1,026×10-18 г = 1,026´10-6 пикограммов (пг), 1 пг ДНК = 9,75´105 килобасов = 0,975 х 106 килобасов.
Препараты ДНК, выделяемой из клеток с помощью обычных методов, имеют молекулярную массу порядка 1,0´107. Длина витка по оси ДНК В-формы равна 34 А̊. Расстояние между парами оснований в ДНК В-формы Е. coli равно 0,34 нм.
Для характеристики строения ДНК используют также такие физические константы, как плотность ее при центрифугировании в градиенте тяжелых металлов, а также температура плавления; первая константа отражает полидисперсность препаратов ДНК, тогда как вторая — их гетерогенность. Нагревание ДНК в растворах разрывает водородные связи между основаниями в парах и разрушает вторичную структуру ДНК, т. е. вызывает плавление ДНК. В 0,1 М раствора NaCI плавление наступает при 95°С.
Плавление ДНК есть ее денатурация. Однако замечательное свойство денатурированной ДНК заключается в том, что она способна к денатурации in vitro, т. е. способна восстанавливать двухцепочечную структуру, причем ренатурация является очень точной. Две цепи денатурированной ДНК могут ренатурировать в природную двухцепочечную спиральную форму, если их последовательности комплементарны или, другими словами, если последовательности цепей позволяют формирование пар оснований, соединенных водородными связями. Ренатурацию можно оценить и в качестве гибридизации ДНК.
Между тем способность самокомплементарных последовательностей к гибридизации и формированию двухцепочечной спирали присуща на только ДНК, но и РНК. В результате этого in vitro можно конструировать двухцепочечные гибридные структуры РНК-РНК или РНК-ДНК. Способность нуклеиновых кислот к ре-натурации имеет значение в изучении специфики отдельных последовательностей, а также в таксономии.
В зависимости от локализации ДНК в клетке различают ядерные (хромосомные) и экстраядерные (экстрахромосомные) детерминанты наследственности. Кроме того, известны транспозируемые генетические элементы (инсерционные последовательности, транспо-зоны и др.).
Похожие статьи:
poznayka.org
Водородные связи оснований — Справочник химика 21
Авторы работы [399] попытались использовать также независимый от критериев способ определения числа водородных связей, основанный на вычислении площади под первым максимумом пика функции распределения расстояния / он(й он). На условность и неоднозначность методов подобного рода для анализа структурных особенностей ансамблей /-структур уже указывалось ранее [386]. Обратим внимание, что применение критерия он приводит к числу водородных связей, равному или даже большему, чем полученное с энергетическим критерием Инв = = 8,37 кДж/моль (2 ккал/моль). Как показывает опыт численного моделирования, среди пар молекул, энергия взаимодействия которых близка к этому значению, встречаются такие, кон-Автором и сотр. [405] предложен динамический критерий водородной связи, основанный на анализе не только данной конфигурации, полученной в ходе молекулярно-динамического моделирования, но и ее предыстории и ее дальнейшей судьбы . Построение временных зависимостей энергий взаимодействия двух молекул в большинстве случаев позволяет судить о том, существует ли при данной конфигурации водородная связь, в какие моменты времени она возникла и когда была разорвана (рис. 8.2). Как и всегда при рассмотрении ансамбля /-структур, при использовании этого подхода могут возникать сомнительные случаи. Однако такие случаи редки и по степени определенности поиска водородных связей динамический критерий приближается к анализу f-структур. [c.142]
Здесь мы ограничились рассмотрением структур кристаллических оксокислот и кислых солей, хотя системы с водородными связями могут быть представлены и другими типами соединений. Два примера в табл. 8.5 иллюстрируют трехмерные системы водородных связей, основанные на простейших сетках со связанностью 4 и 6. Этот ряд можно дополнить примерами [c.46]
Компонентами нуклеиновых кислот являются основания, формулы которых приведены на рис. 1-4. Эти основания состоят из относительно гидрофобных циклов, которые содержат группы, способные образовывать водородные связи при этом получаются пары оснований, показанные на рисунке. Основание ковалентно связано с сахаром (2-дезокси-о-ри6оза в ДНК или о-рибоза в РНК), к которому присоединена фосфатная группа при поликонденсации таких веществ между мономерами возникают фос-фодиэфирные связи и образуются нуклеиновые кислоты, как показано на рис. 1-5. Когда между основаниями образуются водородные связи, полярная часть каждого из них становится менее доступной для воды поскольку гетероциклы явно гидрофобны (особенно по сравнению с сахаром и заряженным фосфатным остатком), в нуклеиновых кислотах, где присутствуют водородные связи, основания расположены таким образом, что они ми- [c.12]
Уместно вспомнить об одном обстоятельстве из истории развития теории химической связи и межмолекулярного взаимодействия. После первых расчетов энергии связи в молекулах с разными атомами (металл — металлоид) стало ясно, что эта величина мало чувствительна к принятой модели. Расчеты гетерополярных молекул с учетом или без учета поляризации, по модели твердых шаров или по любой модели, учитывающей отталкивание, почти всегда приводили к близким к эксперименту значениям энергии связи. Попытки вычислить энергию, например, водородной связи, основанные на разных моделях как электростатических, так и ковалентных, почти всегда давали вполне удовлетворительный результат. То же относится и к расчетам теплот адсорбции. Правильный порядок величины обеспечивается тем, что из эксперимента берутся две или три константы, а правильный характер всей потенциальной кривой постулирован заранее. Сама по себе полуэмпирическая потенциальная кривая, будь то кривая Леннард-Джонса или кривая, в которой коэффициент при берется по Лондону или каким-либо иным теоретическим способом, ничего не может сказать о природе сил адсорбции, так же как и кривая Морзе для двухатомной молекулы ничего не говорит о природе связи атомов в ней. [c.83]
Во многих случаях, когда существует необычно сильная связь между молекулами или между разными группами в одной и той же молекуле, эксперимент показывает, что в связи участвует атом водорода. Речь идет о так называемой водородной связи. Основанием для этого служат четыре источника данных [c.450]
Величина 5 характеризует среднее число нуклеотидных пар, связанных в упорядоченную биспиральную структуру, а А — среднюю длину такого упорядоченного участка спирали. Таким образом, эти величины дают сведения о размерах областей цепи ДНК, на которые распространяется дальний порядок в ориентации плоскостей оснований, ответственных за отрицательную анизотропию цепи и удерживаемых внутримолекулярными водородными связями. Упорядоченные структурные элементы связаны между собой участками, где вторичная структура молекулы (водородные связи оснований) оказывается почему-либо ослабленной, что обеспечивает некоторую гибкость цепи (рис. 311) [258]. В то же время динамооптические свойства ДНК явно не соответствуют модели прямого стержня, ибо в этом случае число мономеров в сегменте 5 просто было бы равно степени полимеризации и анизотропия молекулы, например, для образца, представленного в табл. 40, соответственно должна была бы в М/Ме = 50 раз превосходить величину, найденную экспериментально. Гидродинамические свойства растворов ДНК также лучше согласуются с моле- [c.478]
Соединенные водородными связями основания никогда не бывают идентичными, причем аденин всегда соединен с тими-ном, а гуанин — с цитозином. Истинная причина такого выбора или предпочтения одного соединения другому точно еще не установлена, но поскольку они соединяются при помощи водородной связи, то этот выбор может иметь близкое отношение к стерическим условиям и к природе самой водородной связи. [c.90]
Альтернативное описание ковалентного вклада в водородную связь, основанное на методе молекулярных орбиталей, по мнению автора, более удовлетворительно. Одна из проблем водородной связи заключается в том, что для ее образования необходимо, чтобы атом водорода, обладающий единственной 15-орбиталью, способной к образованию связи, взаимодействовал с двумя атомами. В резонансной модели это объяснялось несвязывающим резонансом между структурами а, с ж е. ]У1етод молекулярных орбиталей дает вполне обоснованное объяснение связывания атома водорода, которое по существу эквивалентно объяснению образования донорно-акцепторпых (с переносом заряда) комплексов и пятивалентного переходного состояния в 5 Jv2-замещении при атоме углерода без привлечения орбиталей с высокой энергией [1[. В соответствии с этой моделью молекулярные орби- [c.270]
В работе предяожевн алектростатическая модель воды и теория водородной связи, основанная на учете электростатического взаимодействия дробных зарядов во взаинодействуицих молекулах водн . [c.675]
Антипараллельность цепей имеет решающее значение при спаривании оснований в одноцепочечных молекулах (таких, как тРНК). Параллельное спаривание не может обеспечить образование щпилек. Двойные спирали, изображенные на рис. 3.12, 3.13 и 3.14, имеют второй набор осей псевдосимметрии С , проходящих между соседними парами связанных водородными связями оснований. Для самокомплементарных последовательностей, таких, как СрС или АрТ (или палиндромных последовательностей, аналогичных приведенным на рис. 3.5), оси, проходящие между двумя центральными парами, являются истинными осями симметрии, поскольку поворот на 180° не изменяет положения остова и переводит пары оснований в идентичные положения. [c.173]
Макромолекула ДНК состоит из двух нитей, закрученных одна вокруг другой так, что образуется так называемая «двойная спираль» (рис. 2.2). Нити состоят из «остова» — чередующихся фосфатных и сахарных групп — и прикрепленного к каждой сахарной группе азотистого основания. Они соединены между собой водородными связями оснований. Если двойную спираль раскрутить, то она будет напоминать лестницу (рис. 2.2, а(, где вертикальные части состоят из сахарофосфатного остова (рис. 2.2, б и г), а перекладины — из комбинации пар оснований, соединенных водородньши связями (рис. 2.2, д). Существуют четыре типа оснований два пиримидиновых (тимин и цитозин) и два пуриновых (гуанин и аденин). Число молекул аденина в любом образце ДНК всегда равно числу молекуп тимина, и соответственно чиспо молекул цитозина равно числу молекул гуанина. Эти пары оснований расположены в спирали друг против друга (рис. 2.2, бив), что делает нити комплементарными одна другой. Издавна существовали проблемы, связанные с супер-спирализацией ДНК и с разделением двойной спирали — такую модель строения ДНК в свое время предложили Уотсон и Крик. Недавно Сазисе- [c.31]
Два вида сил удерживают две полидезоксирибонуклеотидные цепи в биспиральной молекуле ДНК. Во-первых, это водородные связи между комплементарными азотистыми основаниями, обращенными внутрь двойной спирали ДНК (см. рис. 66). Образуя водородные связи, основания находятся в плоскости, перпендикулярной продольной оси В-формы ДНК, так что эти связи действуют в поперечном направлении. Во-вторых, это силы гидрофобных взаимодействий между азотистыми основаниями, собранными в стопку вдоль молекулы ДНК при такой упаковке оснований в водной среде возникают силы, препятствующие контактам неполярных (гидрофобных) оснований с молекулами воды, вследствие чего основания сближаются, а стопкообразная упаковка упрочняется вследствие межплоскостных взаимодействий их друг [c.208]
При сравнительном изучении поведения ДНК, синтетических полинуклеотидов и природных РНК при нагревании в растворе Доти и др. [469] иришди 11 выводу, что при определенных условиях Р1-1К содержит многочисленные 1гезначительпой длины спиральные участки, образованные спаренными посредством водородных связей основаниями, перемежающиеся с одноцепочечпыми областями. Молекулы РИК при этом имеют более или менее компактную структуру. Степень спирализации молекулы РНК при стандартных условиях в некоторой степени зависит от состава оснований. [c.65]
chem21.info
6.11.2012 Г лекция молекулярные основы наследственности
Материальными носителями наследственности являются хромосомы и гены. В 1879 году Флеминг обнаружил в ядре темно окрашивающиеся структуры, которые он назвал хроматином. Зоолог Вейсман в книге «Зародышевая плазма. Теория наследственности» привел аргументы, что наследственное вещество находится в ядре.
В 1902 году Бовери и Сеттон установили, что менделеевские закономерности наследования точно соответствуют закономерностям поведения хромосом. Таким образом, основное назначение хромосом – передача точной информации от поколения к поколению. Врач Мишер (1868 год) используя гнойный материал после операционных повязок, выделил из ядер погибших лейкоцитов вещество, которое назвал нуклеин. В 1914 году доцент Петербургского университета Щепотьев доказал, что это кислоты – нуклеиновые кислоты. Кольцов представляет гипотезу об удвоении (редупликации) хромосом. Работа Эвери и Гриффита раскрыло химическую основу ДНК — наличие азотистых оснований. Одной из главных достижений позволивших построить модель ДНК было правило Эрвина Чаргаффа о комплементарности пуриновых и пиримединовых азотистых оснований. Комплементарность – это способность образовывать водородные связи. По этому правилу количество аденина + гуанина = количеству цитозина + тимина.
Прямым доказательством генетической роли ДНК способность ДНК трансформировать пневмококки одного типа в другой – работа Эвери 1944 года по размножению бактериальных вирусов – фагов.
Рентгеноструктурный анализ (методика дифракции рентгеновских лучей) и методы кристаллографии позволили Максу Перутцу и Морису Уилкинсу практически создать модель ДНК. Однако первенство в создании модели ДНК и Нобелевская премия были присуждены Френсису Крику и Джеймсу Уотсону, которые 25 апреля 1953 года в журнале «Нейчур» опубликовали небольшую статью (900 слов) «Молекулярная структура нуклеиновых кислот» о модели ДНК.
Молекула ДНК являются линейными макромолекулами, представляющими собой длинные двойные цепи (тяжи) полимеров, составленных из мономеров, получивших название нуклеотидов (малых органических молекул) и являющихся строительными блоками ДНК.
У всех живых существ макромолекулы ДНК построены по одному и тому же плану. Они слагаются в основном из одних и тех же нуклеотидов, каждый из которых содержит по одной молекуле фосфорной кислоты и сахара, а также одно из четырех азотистых оснований — аденин, гуанин, цитозин или тимин. Аденин и гуанин являются пуриновыми основаниями, тогда как тимин и цитозин — пиримидиновыми. Пурины и пиримидины называют основаниями по той причине, что в кислой среде они способны присоединять к себе ион Н+. Пиримидины являются производными шестичленного пиримидинового кольца, тогда как пурины представляют основания, у которых второе пятичленное кольцо слито с шестичленным кольцом.
Сахаром в ДНК является 2-дезокси-D-рибоза, отличающаяся от глюкозы тем, что в ее молекуле не 6, а 5 атомов углерода, т. е. является пятиуглеродным сахаром (пентозой). Особенностью этого сахара является также то, что он имеет атом водорода (Н), присоединенный к одному (специфическому) из атомов углерода, но не гидроксильную группу. Следовательно, этот сахар представляет собой дезоксирибозу, т. к. он является рибозой, лишенной кислорода.
Сахарофосфат соединяется с азотистым основанием посредством β-гликозидной связи. Основание прикрепляется к I положению де-зоксирибозы. Структура, образованная соединением азотистого основания и сахара, носит название нуклеозида. Таким образом, химическими группами, которые образуют ДНК, являются пуриновые и пиримидиновые азотистые основания (аденин, гуанин, тимин и цитозин), сахар (2-дезокси-D-рибоза) и фосфорная кислота.
РНК характеризуется такой же структурой, как и ДНК. Однако в отличие от ДНК в РНК сахаром является рибоза с кислородом, представляющая собой сахар с 5 атомами углерода, к одному из которых прикреплена 2′-гидроксильная группа (-ОН). Кроме того, в РНК тимин не имеет метильной группы и является урацилом, т. е, в РНК тимин заменен на урацил, также являющийся пиримидиновым основанием.
Нуклеиновые кислоты называют кислотами по той причине, что их фосфатные группы освобождают в растворах ионы водорода.
Для состава ДНК характерны закономерности, известные в качестве правил А. Чаргаффа, а именно:
1. Сумма нуклеотидов, содержащих пуриновые азотистые основания, равна сумме нуклеотидов, содержащих пиримидиновые азотистые основания
2. Содержание аденина равно содержанию тимина, а гуанина — цитозину
3. ДНК из разных источников может иметь различия, обусловленные в одних случаях преобладанием аденина над гуанином и тимина над цитозином (А + Т > Г + Ц), в других случаях — преобладанием гуанина и цитозина над аденином и тимином (Г + Ц > А + Т).
Для ДНК характерна структура трех видов — первичная, вторичная и третичная. Первичная структура ДНК заключается в том, что ДНК состоит из нуклеотидных цепей, у которых скелетную основу составляют чередующиеся сахарные и фосфатные группы, объединенные ковалентными 3′-, 5′-фосфодиэфирными, скелетными связями, а боковые группы представлены тем или иным основанием (одним из четырех) и присоединяются одна к другой молекулой сахара. Последовательно располагающиеся нуклеотиды ковалентно связаны фосфодиэфирными связями между сахарным остатком и фосфатом, и в результате этого объединены в полинуклеотидную цепь. Таким образом, первичная структура ДНК (как и РНК) определяется последовательностью нуклеотидов и характером их связей между сахарным остатком и фосфатом.
Представления о вторичной структуре ДНК были сформулированы Д. Уотсоном и Ф. Криком еще в 1953 г. На основе данных об Х-дифракции молекул ДНК, структуре оснований и правил А. Чаргаффа эти представления сводятся к следующему:
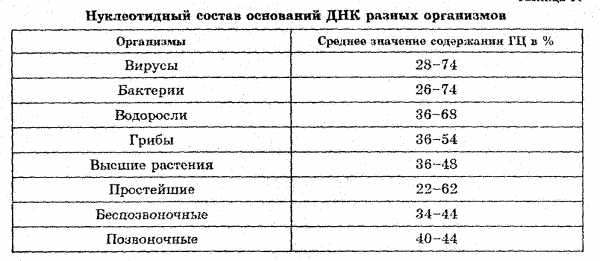
1. По Уотсону и Крику (1953 год) молекула ДНК (форма В) построена из двух скрученных направо спиралевидных полинуклеотидных цепей, причем каждый виток спирали соответствует 10 азотистым основаниям или расстоянию в 3,4 нм. Молекулы ДНК, цепи которых скручены направо, первоначально назвали В-формой.
2. Обе цепи объединены в результате закручивания одной цепи вокруг другой по общей оси. Из-за противоположной последовательности атомов в каждой цепи обе цепи инвертированы относительно одна другой, т. е. направление вдоль дуплекса есть 3’→5′ для одной цепи и 5’~» 3′ для другой.
3. Сахарофосфатные группы располагаются на внешней стороне двойной спирали, тогда как основания находятся внутри спирали под прямым углом и вдоль ее оси. Диаметр молекулы составляет 2 нм, расстояния между отдельными азотистыми основаниями в молекуле равны 0,34 нм. Таким образом, ДНК представляет собой скрученную в правостороннем направлении двойную спираль, в которой пары азотистых оснований А-Т и Г-Ц в комплементарных полинуклеотидных цепях подобны перекладинам в лестнице, а сахарофосфатные цепи являются каркасом этой лестницы.
4. Цепи в молекуле не идентичны, но комплементарны и удерживаются слабыми водородными связями между азотистыми основаниями, причем спаривание азотистых оснований для связывания цепей имеет специфический характер. Водородные связи устанавливаются не просто между азотистыми основаниями цепей, а специфически между пуриновым азотистым основанием одной цепи и пиримидиновым азотистым основанием другой. В результате этого аденин одной из цепей связывается с тимином другой цепи двумя водородными связями, тогда как гуанин одной из цепей связывается с цитозином, находящимся в другой цепи, посредством трех водородных связей.
studfiles.net
Азотистые основания — это… Что такое Азотистые основания?
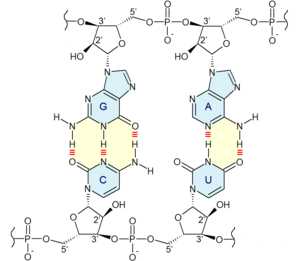 Фрагмент комплементарного участка РНК. Азотистые основания закрашены синим цветом
Фрагмент комплементарного участка РНК. Азотистые основания закрашены синим цветом
Азо́тистые основа́ния — гетероциклические органические соединения, производные пиримидина и пурина, входящие в состав нуклеиновых кислот. Для сокращенного обозначения пользуются большими латинскими буквами. К азотистым основаниям относят аденин (A), гуанин (G), цитозин (C), которые входят в состав как ДНК, так и РНК. Тимин (T) входит в состав только ДНК, а урацил (U) встречается только в РНК.[1]
Аденин и гуанин являются производными пурина, а цитозин, урацил и тимин — производными пиримидина.
Тимин, который присутствует только в ДНК, и урацил, который встречается только в РНК, обладают сходной химической структурой. Урацил отличается от тимина отсутствием метильной группы у 5-го атома углерода.
Азотистые основания, соединяясь ковалентной связью с 1′ атомом рибозы или дезоксирибозы, образуют N-гликозиды, которые называют нуклеозиды. Нуклеозиды, в которых к 5′-гидроксильной группе сахара присоединены одна или несколько фосфатных групп, называются нуклеотидами. Эти соединения являются строительными блоками молекул нуклеиновых кислот — ДНК и РНК.
После образования молекулы нуклеиновой кислоты входящие в её состав азотистые основания могут вступать в различные химические реакции под действием ферментов а также факторов внешней среды. Таким образом, нуклеиновые кислоты часто содержат модифицированные азотистые основания. Типичной модификацией такого рода является метилирование.[2]
Структура
В таблице приведена структура главных азотистых оснований.
Примечания
- ↑ Кнорре Д. Г., Мызина С. Д. Биологическая химия. — 3. — Москва: Высшая школа, 2000. — 479 с. — 7000 экз. — ISBN 5060037207
- ↑ Альбертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. Молекулярная биология клетки: в трех томах. — 2. — Москва: Мир, 1994. — Т. 1. — 517 с. — 10 000 экз. — ISBN 5030019855
Литература
- Кнорре Д. Г., Мызина С. Д. Биологическая химия. — 3. — Москва: Высшая школа, 2000. — 479 с. — 7000 экз. — ISBN 5060037207
- Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: в двух томах. — Москва: Мир, 2004. — Т. 1. — 381 с. — 2000 экз. — ISBN 5030036008
- Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: в двух томах. — Москва: Мир, 2004. — Т. 2. — 414 с. — 2000 экз. — ISBN 5030036016
dikc.academic.ru
Дезоксирибонуклеиновая кислота (ДНК) | Student Guru
Дезоксирибонуклеиновая кислота или ДНК является носителем генетической информации. Главным образом, ДНК в клетках сосредоточена в ядре. Это основной компонент хромосом. У эукариот ДНК также обнаруживается еще в митохондриях и пластидах. ДНК состоит из мононуклеотидов, ковалентно связанных между собой, представляя собой длинный неразветвленный полимер. Мононуклеотиды, входящие в состав ДНК, состоят из дезоксирибозы, одного из 4-х азотистых оснований (аденин, гуанин, цитозин и тимин), и остатка фосфорной кислоты. Количество этих мононуклеотидов очень велико. Например, в клетках прокариот, содержащих одну единственную хромосому, ДНК представляет собой одну макромолекулу с молекулярной массой более 2 x 109.
Мононуклеотиды одной цепи ДНК соединяются последовательно друг с другом благодаря образованию ковалентных фосфодиэфирных связей между ОН-группой дезоксирибозы одного мононуклеотида и остатком фосфорной кислоты другого. С одной стороны от образовавшегося остова одной цепи ДНК располагаются азотистые основания. Их можно сравнить, с четырьмя разными бусинками надетых на одну нить, т.к. они как бы нанизаны на сахарофосфатную цепь.
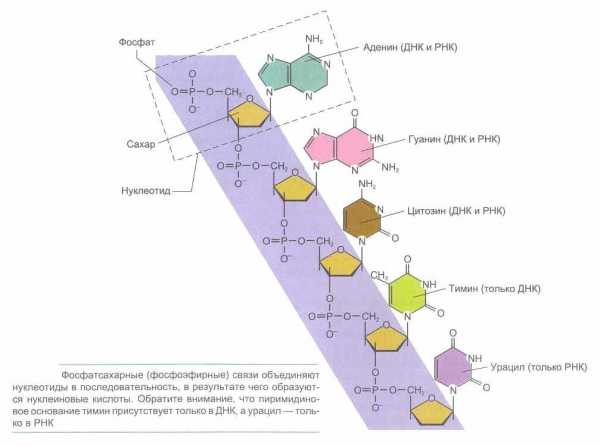
Структура ДНК и РНК
Возникает вопрос, как эта длинная полинуклеотидная цепь может кодировать программу развития клетки или даже целого организма? Ответ на этот вопрос можно получить, поняв, как образуется пространственная структура ДНК. Структура этой молекулы была расшифрована и описана Дж. Уотсоном и Ф. Криком в 1953 году.
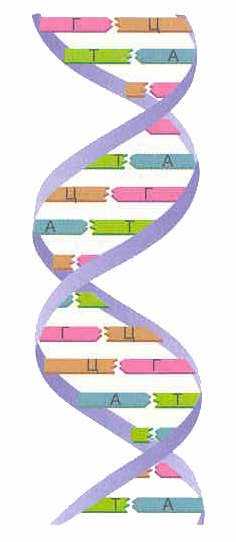
Молекула ДНК представляет собой две параллельные нити.
Молекулы ДНК представляют собой две нити, которые располагаются параллельно друг другу и формируют правозакрученную спираль. Ширина этой спирали составляет около 2 нм, зато ее длина может достигать сотен тысяч нанометров. Уотсоном и Криком предложили модель ДНК, согласно которой все основания ДНК расположены внутри спирали, снаружи находится сахарофосфатный остов. Таким образом, основания одной цепи максимально сближены с основаниями другой,
поэтому между ними формируются водородные связи. Структура спирали ДНК такова, что полинуклеотидные цепи, которые входят в ее состав, могут быть разделены только после ее раскручивания.
Благодаря максимальной сближенности двух цепей ДНК в ее составе содержится одинаковое количество азотистых оснований одного типа (аденин и гуанин) и азотистых оснований другого типа (тимин и цитозин), т. е. справедлива формула: А+Г=Т+Ц. Это объясняется размерами азотистых оснований, а именно, длина структур, которые образуются благодаря возникновению водородной связи между парами аденин-тимин и гуанин-цитозин, приблизительно составляет 1,1 нм. Суммарные размеры этих пар соответствуют размерам внутренней части спирали ДНК. Для формирования спирали пара Ц-Т была бы слишком мала, а пара А-Г, наоборот, слишком велика. Т.е., азотистое основание первой цепи ДНК, определяет основание, которое располагается в том же самом месте другой цепи ДНК. Строгое соответствие нуклеотидов, расположенных в молекуле ДНК в парных цепочках параллельно друг другу, назвали комплементарностью (дополнительностью). Точное воспроизведение или репликация генетической информации возможна именно благодаря этой особенности молекулы ДНК.
В ДНК биологическая информация записана таким образом, что она может в точности копироваться и передаваться клеткам-потомкам. До деления клетки в ней происходит репликация (самоудвоение) ДНК. Поскольку каждая цепь содержит последовательность нуклеотидов, комплементарную последовательности цепипартнера, то на самом деле они несут одинаковую генетическую информацию. Если разделить цепи и использовать каждую из них в качестве шаблона (матрицы)для построения второй цепи, то получится две новых идентичных цепи ДНК. Именно таким образом и происходит удвоение ДНК в клетке.
Фосфатсахарные последовательности
Водородные связи между азотистыми основаниями соседних цепей ДНК разрываются. Эти цепи разделяются. Затем происходит синтез двух новых молекул ДНК (дочерних) с использованием в виде матриц их родительских цепей. Эти реакции стали называть реакциями матричного синтеза.
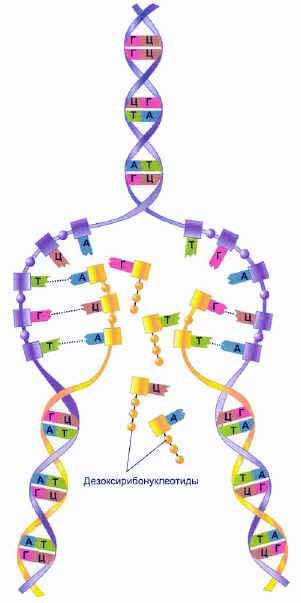
Процесс репликации ДНК
Подробнее про репликацию ДНК можно прочесть в разделе «Репликация ДНК«.
Перейти к оглавлению.
from your own site.
www.studentguru.ru
Пуриновые основания водородные связи — Справочник химика 21
Потеря трансформирующей активности может происходить не только при критических температурах, но также при длительном нагревании при температурах, близких к критическим. В то время как разные маркеры в одном и том же образце имеют различные температуры, при которых происходит нарушение структуры, зависимость процессов инактивации от температуры при температурах, близких к критической, примерно одна и та же для всех маркеров, если только при этом не происходит их деградации. Инактивация при температурах, близких к критическим, протекает как простой процесс, она соответствует реакции первого порядка и происходит, по-видимому, лишь для пар оснований, водородные связи у которых разрушились при нагревании поэтому при температурах, приближающихся к критическим, происходит аномально быстрое увеличение скорости [257]. Процесс сильно ускоряется при pH 5,4, а более крупные генетические участки более чувствительны к инактивации при субкритических температурах [258]. Почти наверняка при этом происходит разрыв гликозидной связи в пуриновых дезоксинуклеозидах [257, 258[. Зная скорость разрушения пуриновых дезоксинуклеозидов, удалось рассчитать, что маркер, определяющий устойчивость к стрептомицину, состоит примерно из 430 пар оснований [257]. [c.581]Таково расположение боковых групп в плоскости, нормальной к оси спирали. Все пуриновые основания попарно связаны водородными связями. Образование комплексов пуриновых полинуклеотидов друг с другом и с пиримидиновыми полинуклеотидами [c.230]
Ф — фосфат С — сахар А — аденин Г — гуанин-пуриновые основания Ц цитозин Т — тимин-пиримидиновые основания . .. — водородные связи [c.32]
В тот вечер Фрэнсис и Гриффит недолго занимались пережевыванием избитых гипотез. Оба понимали, что сейчас важно установить природу этих сил притяжения. Фрэнсис убежденно доказывал, что специфические водородные связи не могут быть решением проблемы. Они не могут обеспечить необходимую строгую специфичность, потому что, как нам не раз говорили наши приятели-химики, атомы водорода в пуриновых и пиримидиновых основаниях не имеют определенного местоположения, а случайным образом перемещаются с одного места на другое. Фрэнсис предполагал, что вместо них в копировании ДНК участвуют специфические силы притяжения между плоскими поверхностями оснований. [c.75]
Водородные связи соединяют аденин всегда только с тимином, а гуанин — только с цитозином. Эта исключительность выбора объясняется геометрическими причинами молекулы пуриновых оснований более объемисты, чем пиримидиновых, оба основания пары лежат в одной плоскости, причем диаметр спирали ДНК равен 1.8 нм. В этих условиях допустимы только парные сочетания пиримидиновых и пуриновых оснований. Выбор именно тех пар, которые указаны выше, обусловлен еще и повышенной прочностью водородных связей в парах А—Т и Г—Ц (рис. IV.3). [c.351]
Особенностью т-РНК является то, что на одном конце цепочки, содержащей всего 80 нуклеотидов, всегда помещается группа из трех частиц двух цитозина и одной аденина на другом конце находится гуанин. Водородные связи между основаниями обусловливают скручивание отдельных участков цепи в двойную спираль. Свободные нуклеотиды взаимодействуют с матрицей, на которой закрепляется совокупность аминокислот во время синтеза белка. Существование таких свободных нуклеотидов, возможно, связано с наличием в т-РНК пуриновых или пиримидиновых оснований, [c.391]
В каждой молекуле ДНК существует точное соответствие между числом аденина и тимина, с одной стороны, и гуанина и цитозина —с другой. Установлено, что углевод-фосфатная спираль ориентируется таким образом, что пиримидиновые и пуриновые основания направлены внутрь спирали. Водородные связи между основаниями, расположенными на двух молекулах друг напротив друга, способствуют укреплению двойной спирали точное спаривание маленького пиримидинового основания с большим пуриновым приводит к их равному соотношению в молекуле ДНК. Хотя водородные связи, несомненно, участвуют в стабилизации двойной спирали, прочность связывания двух цепей слишком велика, чтобы ее можно было объяснить исключительно этим взаимодействием. [c.321]
Для поддержания регулярности спирали необходимо, чтобы против остатка пуринового основания в одной цепи находился остаток пиримидинового основания в другой. Дж. Уотсон и Ф. Крик нашли, что это требование выполняется только в то.м случае, если А образует стабилизированную водородными связями пару с Т, а С с С (см. следующий раздел). [c.22]
Выяснение молекулярной структуры генетического материала -ДНК — без сомнения стало одним из самых замечательных научных достижений XX в. Уотсон и Крик описали свое открытие так Мы хотим предложить структуру соли дезоксирибонуклеиновой кислоты (ДНК). Эта структура обладает весьма необычными свойствами, представляющими большой биологический интерес… Она образована двумя спиральными цепочками, закрученными вокруг общей оси… Обе спирали правые, но… последовательности атомов в них взаимно противоположны… Весьма интересен способ, с помощью которого цепочки удерживаются вместе… Пуриновые и пиримидиновые основания образуют пары, при этом пуриновое основание одной цепи соединяется водородными связями с пиримидиновым основанием другой… Если одно из оснований пары — это аденин, то… вторым основанием должен [c.45]
Установление предпочтительной таутомерной формы имеет большое значение для пуриновых и пиримидиновых оснований, которые входят в состав ДНК и РНК, поскольку водородные связи с участием карбонильного атома кислорода обеспечивают возможность спаривания. [c.24]
Физико-химические свойства ДНК. Различные факторы, нарушающие водородные связи (повышение температуры выше 80 °С, изменение pH и ионной силы, действие мочевины и др.), вызывают денатурацию ДНК, т. е. изменение пространственного расположения цепей ДНК без разрыва ковалентных связей. Двойная спираль ДНК при денатурации полностью или частично разделяется на составляющие-ее цепи. Денатурация ДНК сопровождается усилением оптического поглощения в УФ-области пуриновых и пиримидиновых оснований. Это явление называют гиперхромным эффектом. При денатурации [c.183]
Вторичная структура молекулы ДНК, по Уотсону и Крику, представляет собой а-спираль, состоящую из двух полинуклеотид ных цепей, закрученных одна вокруг другой и вокруг общей для обеих цепей оси (рис. 92). Эти цепи связаны водородными связями между молекулами пиримидиновых и пуриновых оснований. Причем было установлено, что такого рода прочные связи образуются лишь между, специфическими парами аденин — тимин, гуанин — цитозин [c.557]
Вариация структуры нуклеиновых кислот происходит за счет вариации последовательности гетероциклических оснований в их боковой части В состав ДНК входят в основном фрагменты аденина, гуанина, цитозина и тимина, РНК — фрагменты аденина, гуанина, цитозина и урацила Вторичная структура нуклеиновых кислот, представляющая собой двойную спираль переплетающихся двух полимерных цепей ДНК, двуспиральных фрагментов РНК, одноцепочечные участки РНК, обязана своим образованием возникновению водородных связей между пиримидиновыми и пуриновыми основаниями Это крупнейшее открытие XX века, сделанное Дж Уотсоном и Ф Криком в 1953 г (Нобелевская премия 1962 г ), стало возможным благодаря интеграции различных биологических, химических и физических методов исследования [c.928]
Цепи выстраиваются в противоположных направлениях и удерживаются вместе водородными связями, образующимися между пуриновыми и пиримидиновыми основаниями. Водородные связи образуются лишь между определенными основаниями А=Т (соединены двумя водородными связями) Г=Ц (соединены тремя водородными связями). Такие пары оснований называются комп-лементарны-гли парами (рис. 26). [c.663]
Цепи выстраиваются в противоположных направлениях и удерживаются вместе водородными связями, образующимися между пуриновыми и пиримидиновыми основаниями. Водородные связи образуются лищь между определенными основаниями А = Т (соединены двумя водородными связями) Г = Ц 37. Схемати-(соеинены тремя водородаымисвязя-ми). Такие пары основании называются днк (обозначения комплементарными парами (рис. 38). 8 — остаток пен- [c.721]
ДНК-РНК-гибрида также удерживаются вместе водородными связями. Водородные связи возникают между пуриновыми и пиримидиновыми основаниями двух цепей очень специфическим образом. Другими словами, основания одной цепи совмещены с основаниями второй цепи таким образом, что пуриновое основание ео.а, связ водородными связями с пириьГидиновым осн а-нием и наоборот. Говоря более точно, аденин и тймин»(урацил) способны связываться друг с другом двумя водородными мостиками, образуя пару. Точно так же спариваются гуанин и цитозин, способные образовать три водородные связи друг с другом. Такое специфическое образбваТше водородных связей [уотсон-криковские пары) важно при репликации, так как две цепи [c.114]Синтез (репликация) ДНК должен происходить таким образом, чтобы образовались две новые цепи двухтяжевой ДНК с той же самой последовательностью оснований, т. е. той же генетической информацией, что и родительская. Благодаря такому процессу из данной родительской клетки возникают две дочерние. Репликация становится возможной потому, что двухтяжевая родительская ДНК разделяется на отдельные нити, из которых каждая служит матрицей для синтеза новой спирали. Если бы две цепи были ковалентно связаны, энергия, необходимая для разделения цепей, была бы весьма значительной. Сохранение последовательности оснований в процессе репликации происходит благодаря высокой специфичности при образовании водородных связей между пуриновыми и пиримидиновыми основаниями. Так что, например, аденин на одной цепи двойной спирали всегда будет находиться напротив и образовывать водородные связи с тимином во второй цепи. При разделении цепей аденин из одной цепи всегда будет взаимодействовать с тимином в процессе синтеза новой комплементарной цепи. Аналогичным образом тимин, который находился напротив аденина в родительской двойной спирали, после разделения цепей будет взаимодействовать в процессе синтеза новой комплементарной цепи с аденином. Следовательно, на каждой из разделенных цепей родительской двойной спирали, как на матрице, синтезируются две новые цепи двухспиральмой ДНК, обладающие совершенно одинаковой последовательностью оснований с родительской молекулой. Такой механизм синтеза ДНК называется полуконсервативным механизмом репликации, поскольку исходная двойная спираль наполовину сохраняется (рис. 3.9), т, е, каждая из двух образовавшихся двойных спиралей содержит одну цепь из родительской молекулы. [c.148]
При изучении ДНК рентгенографическим методом было установ-лерю, что макромолекулы имеют строго регулярное строение, а химическое исследование показало, что число пиримидиновых оснований всегда равно числу пуриновых аденина всегда столько, сколько тимина цитозина столько же, сколько гуанина. Объяснение этим фактам дали Д. Уотсон и Ф. Крик в своей модели двойной спирали (1953 г.). Двойная спираль (рис. 25) похожа на винтовую лестницу. Две стойки этой лестницы образованы основной цепью, состоящей из углеводных и фосфатных остатков, азотистые основания образуют как бы ступеньки лестницы. Азотистое основание одной поли нуклеотидной цепи связано водородными связями с азотистым основанием другой цепи [c.352]
Мы рассмотрели только фрагмент одной цепи ДНК-Однако по данным рентгеноструктурного анализа макромолекулы ДНК, а также РНК представляют собой две взаимосвязанные спиральные цепи, фрагмент которой показан на рисунке 5. Обе спиральные цепи связаны водородными связями, возникающими в результате взаимодействия радикала гетероциклического основания одной цепи с радикалом гетероциклического основания другой цепи. Основания, образующие пары, связанные водородными связями, называются комплемёнтарными или взаимно дополняющими. При образовании таких взаимно связанных пар соблюдается следующая закономерность одно основание обязательно должно быть пуриновое, а другое— пиримидиновое. Причем тимин непременно находится в паре с аденином, а цитозин— с гуанином. [c.23]
На основании рентгеноструктурного анализа и ранее полученных данных о строении нуклеотидов и нуклеиновых кислот Уотсон и Крик предложили для ДНК структурную модель, согласно которой макромолекула ДНК имеет форму спирали, причем в спираль закручены одновременно две молекулы ДНК (двухцепочечная спиральная структура). Эта двойная спираль имеет одну общую ось и построена так, что основания обеих цепей расположены внутри спирали, а углеводные остатки с фосфатными группами — снаружи спирали (рис. 51, 52). При этом основания одной молекулярной цепи с основаниями другой цепи образуют строго фиксированные пары, соединенные друг с другом водородными связями. Симметричное построение спирали требует постоянства межспиральных расстояний, а это возможно лишь в том случае, если размеры пар оснований, расположенных друг против друга, будут одинаковыми. Такому условию отвечают пары, построенные из одного пуринового и одного пиримидинового основания аденин — тимин и цитозин — гуанин, что обеспечивает и максимальное число водородных связей в спирали [c.362]
Этот процесс представляет собой наиболее удивительный и яркий пример более общего явления — достижения биологической специфичности путем взаимодействия комплементарных структур, аналогичных тем, которые обусловливают связывание антител и антигенов (разд. 15.5). Объединение пуриновых и пиримидиновых оснований в пары путем образования двух или трех водородных связей происходит в соответствии с принципом комплементарности. Взаимное положение оснований в углеводнофосфатном скелете ДНК таково, что только пуриновое основание одной цепи и пиримидиновое основание другой цепи могут образовать между собою водородную связь. В принципе возможны и неправильные пары — АС и ОТ. Однако, как видно из рис. 15.20, между А и С не могут образоваться водородные связи при заданном положении оснований в цепи ДНК. Между О и Т могла бы возникнуть одна водородная связь, но в действительности средние атомы водорода (связанные с атомами азота колец как у О, так и у Т) создают пространственные затруднения, которые удерживают О и Т достаточно далеко друг от друга, препятствуя образованию такой связи. Энергия водородной связи составляет примерно 20 кДхпиримидинового основания дает энергетический выигрыш в 40 или 60 кДж-моль . [c.458]
Специфич. наборы водородных связей между пиримидиновыми и пуриновыми основаниями в комплементарных участках цепей (см. Комплементарность), а также меж-плоскостные взаимод. между соседними основаниями в цепи определяют формирование и стабилизацию вторичной и третичной структуры нуклеиновых к-т. Последовательность пуриновых и пиримидиновых оснований в полинуклеотидной цепи определяет генетич информацию ДНК и матричных РНК. Модификация Пов полинуклеотидах под воздействием мутагенов может приводить к изменению информац. смысла (точковой мутации). [c.530]
Нуклеиновые кислоты содержатся в каждой живой клетке. Они принимают решающее участие в биосинтезе белка и ответственны за передачу генетической информации. В настоящее время уже многое стало известно о способе передачи такой информации, которая осуществляется вторичной структурой ДНК, имеющей вид спирали из двух витков дезоксирибозофосфатной цепи, связанных с помощью водородных связей. Водородные связи соединяют остаток аденина из одного витка спирали с торчащим напротив остатком тимина второго витка, а также остаток цитозина одного витка с остатком гуанина другого. Такой порядок связывания двух дезоксирибозофосфатных цепей строго специфичен водородная связь не может образоваться между аденином одной цепи и гуанином или цитозином другой. Не может она возникнуть и между цитозином одной цепи и тимином или аденином другой и т. д. Такая специфичность определяется строением пуриновых и пиримидиновых оснований или их взаимным расположением, а возможно, и тем и другим. Приведенная схема иллюстрирует условия образования водородных связей [c.355]
РИС. 2-25. Внешние очертания пуриновых и пиримидиновых оснований нуклеиновых кислот. Изображены поверхности, определяемые вандерваальсовыми радиусами отмечены также некоторые из возможных направлений, вдоль которых могут быть образованы водородные связи. Толстыми стрелками указаны водородные связи, соответствующие схеме спаривания оснований по Уотсону и Крику. [c.135]
На рис. 2-21 показана структура четырех пуриновых и пиримидиновых оснований, образующих большинство обычных нуклеотидных ручек . Контуры представляют поверхности контакта, задаваемые ван-дерваальсовыми радиусами, а стрелки показывают некоторые из направлений, в которых могут образовываться водородные связи с соседними группами. Отличительные особенности четырех оснований как с точки зрения геометрической формы, так и сточки зрения возможного расположения водородных связей сразу становятся очевидными. При [c.189]
В создании и поддержании третичной структуры тРНК реализуется много неканонических (не Уотсон— Криковских) взаимодействий между основаниями цепи. Прежде всего, угол L-образной молекулы тРНК крепится как межплоскостными взаимодействиями, так и взаимодействиями через водородные связи между дигидроуридиловой петлей и Т-петлей. Взаимодействие между инвариантами G19 и С56 —Уотсон —Кри-ковского типа (см. рис. 19, б), но взаимодействие между инвариантами G18 и FSS очень своеобразно и включает водородные связи атома О при С4 пиримидинового кольца F как с N1, так и с N при С2 пуринового кольца G (рис. 23, а см. цветную вклейку). Кроме того, имеется необычное сильное межплоскостное взаимодействие между тремя гуанози-новыми остатками в том же углу G57 оказывается вставленным (интеркалированным) между G18 и G19. Более того, G57 через N при С2 взаимодействует водородными связями с рибозами G18 и G19, а через N7 — водородной связью с рибозой 55. [c.38]
В многоклеточных организмах среднее число регуляторных сайтов для одного гена минимум равно пяти положительные регуляторные белки связываются со своими специфическими последовательностями в структуре ДНК (вероятнее всего, посредством водородных связей между амидной группой Глн или Асн и пуриновыми и пиримидиновыми основаниями нуклеотидов). Следует указать еще на один момент, почему эукариотическая клетка использует положительные механизмы регуляции экспрессии генов. Подсчитано, что в геноме человека содержится около 100000 генов, соответственно каждая клетка при отрицательном механизме регуляции могла бы синтезировать 100000 разных репрессоров, причем в достаточных количествах. При положительном механизме регуляции большинство генов в принципе неактивно, соответственно молекула РНК-полимеразы не связывается с промотором и клетка синтезирует ограниченный и избирательный круг активаторных белков, необходимых для инициации транскрипции. [c.538]
Итак, регуляция активных генов осуществляется с помощью различных регуляторных белков-репрессоров и активаторов транскрипции. С физической точки зрения наиболее интересным свойством этих белков является их способность у.чнавать специфические нуклеотидные последовательности ДНК. Установлено, что в комплексе с регуляторными белками сохраняется обычная -подобная конформация ДНК. Узнавание белками их специфических связывающих мест на ДНК основывается на прямом чтении белком последовательности оснований в узкой и/или широкой бороздках ДНК. Специфичность связывания обеспечивается образованием большого числа водородных связен и других слабых взаимодействий между функциональными группами белка и основаниями ДНК. Одна и та же последовательность оснований может быть прочитана как со стороны узкой, так и со стороны широкой бороздки ДНК. Однако характер и пространственное расположение функциональных групп оснований — потенциальных доноров и акцепторов водородных связей— в узкой и широкой бороздках ДНК значительно отличаются. Поэтому часто говорят о двух каналах передачи информации. В узкой бороздке ДНК атомы 02 пиримидинов и N3 пуринов могут служить в качестве акцепторов водородных связей, в то время как 2-аминогруипа гуанина часто является донором водородной связи. Важной особенностью структуры ДНК является пространственная эквивалентность положений всех этих акцепторных групп для пуриновых и пиримидиновых оснований, находящихся в одной и той же полинуклеотидной цепи. Кроме того, атомы N3 пурина и 02 пиримидина в каждой паре оснований связаны осью симметрии второго порядка. Поэтому при чтении текста со стороны узкой бороздки ДНК АТ- и ГЦ-пары легко узнать, в то время как АТ- и ТА-пары различить трудно, так как оии несут геометрически эквивалентные группы сходной химической природы. [c.290]
В широкой бороздке ДНК атомы N7 аденина и гуанина занимают эквивалентные положения, и водородные связи с этими атомами позволяют отличить пуриновые основания от пиримидиновых. Другим важным свойством является то, что как доноры, так и акцепторы водородных связей (аминогруппы аденина и цитозина, атомы 04 тимина и 06 гуанина соответственно) попарно занимают весьма близкие, хотя и неидентичные положения. Поэтому при взаимодействии гипотетической пары до-норной и акцепторных групп белка с упомянутыми выше группами оснований выполняются соотношения А Ц и Т Г. Это вырождение можно снять, если образуется дополнительная водородная свяэь с Н7-атомом пурина. [c.290]
Особенностью ДНК является эквивалентность количеств пуриновых оснований количествам пиримидиновых оснований, т.е. отношения аденин тимин, гуанин цитозин (+ ме-тилцитозин, если он имеется) всегда равны единице. Это соотношение объясняется тем, что пространственная структура ДНК создаётся за счёт образования водородных связей между остатками нуклеиновых оснований, причём аденин связывается только с ТИМИНОМ, а гуанин — только с цитозином [c.111]
Согласно модели Уотсона — Крика молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали, имеющей диаметр 1,8— 2,0 нм. Две полинуклеотидные цепи антипараллельны друг другу, т. е. направления образования фосфодиэфирных связей в них противоположны в одной цепи 5 —3, в другой — 3 —5. Пуриновые и пиримидиновые основания направлены внутрь спирали. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи возникают водородные связи. Эти основания составляют комплементарные пары. [c.444]
Что касается комплекса с келлином, то предположение о существовании этого растворимого молекулярного соединения сделано ввиду солюбилизации келлина при помощи к,эфеина или диоксипропилтео-филлина 500 выводу о возникновении молекулярного соединения J3 -нафтола с кофеином пришли на основании торможения реакции азосочетания -нафтола в присутствии кофеина 01 в последних двух случаях, мы, по- видимому, имеем дело с -комплексами,хотя для 3-нафтола возможно образование и водородной связи. Дальнейшие примеры молекулярных соединений пуриновых алкалоидов при помощи спектров установлено образование комплекса кофеина с бензойной кислотой. Высказано предположение о связывании в данном случае компонентов молекулярного соединения электростатическими силами мевдг карбоксилом бензойной кислоты и азотом в положении 7 [c.148]
Азотистые основания, входящие в состав нуклеотидов цепочки ДНК, объединены водородными связями так, что аденин всегда спарен с тимином, гуанин с цитозином содержание аденина с тимином, а гуанина с цитозином — эквимоле-кулярны, иначе говоря А = Т и Г = Ц. Отношение пуриновых оснований к пирими- [c.29]
В твердой форме эта кпслота обладает кристаллическим строением (что подтверждается четкой картиной диффракцни рентгеновских лучей), высокой плотностью и может быть вытянута в нити. Несколько лет назад Астбери [114] предложил для дезоксирибонуклеиновой кислоты плотно упакованную структуру, в которой остатки дезоксирибозы и основания находятся в слоях, разделенных фосфатными связями. Сравнительно недавно Уотсон и Крик [115] предложили модель двойной спирали, в которой две спирали переплетаются таким образом, что последовательность остатков в одной спирали противоположна их пос-ледовятрльности в другой. Основания могут быть расположены в такой структуре только определенными парами, по одному на каждой спипяли при этом дня пуриновых основания слишком велики, чтобы пара нз ннх могла разместиться в этой структуре, а два пиримидиновых основания слишком малы. При наличии определенных доказательств было принято, что противоположным компонентом в паре оснований для аденина может быть только тимин, а для гуанина — цитозин. Предполагается, что водородные связи между парами оснований обеспечивают стабильность спиральной структуры дезоксирибонуклеиновой кислоты. Предполагаемое строение ее показано на рис. 46. [c.250]
Упорядоченные структуры нуклеиновых кислот включают более, чем одну цепную молекулу. Структура дезоксирибонуклеиновой кислоты (ДНК), предложенная Уотсоном и Криком [22] и Уилкинсом [23], включает две взапмнопереплетен-ные спиральные цепочки, напоминающие винтовую лестницу. Ступеньки этой лестницы , делающие структуру стабильной, образованы водородными связями, которые соединяют комплементарные пуриновые и пиримидиновые основания. [c.24]
В работах [63—65] были измерены температуры плавления других биологически важных макромолекул, синтетических полинуклеотидов и природных нуклеиновых кислот. В упорядоченном состоянии молекула дезоксирибонуклеиновой кислоты состоит из двух спирально переплетенных цепей. Кристаллографическая структура, определенная Криком и Уотсоном [66], допускает только один способ образования пар гетероциклическими основаниями, входящими в состав каждой из этих цепей. Анализ состава нуклеиновых кислот показывает, что концентрация пуриновых оснований равна концентрации пиримидиновых оснований поэтому образование пар через водородную связь, по статистическим соображениям, возможно только между адени-ном (А) и ТИМИНОМ (Т), и между гуанином (Г) и цитозином (Ц). При плавлении цепи разделяются и переходят в беспорядочно свернутое состояние. [c.134]
В свете последних исследований для нуклеиновых кислот принимается сдвоенная спиральная структура, причем фосфатные группировкн и органические основания (пуриновые или пиримидиновые) расположеги, соответственно с внешней и внутренней стороны спирали и связаны водородными связями (рис. 170) только между оиределенными парами органических оснований (аденин — тимии или гуанин — цитозин в дезоксирибонуклеиновой кислоте). [c.242]
chem21.info
