
Митохондрия | steelbros.ru — сообщество стальных братюней
Митохондрия.Митохондрия — состоящая из двух мембран органелла толщиной около 0,5 мкм.

Энергетическая станция клетки; основная функция — окисление органических соединений и использование, освобождающейся при их распаде энергии в синтезе молекул атф ( универсальный источник энергии для всех биохимических процессов ).
По своему строению они представляют собой цилиндрические органеллы, встречающиеся в эукариотической клетке в количестве от нескольких сот до 1-2 тысяч и занимающие 10-20 % её внутреннего объёма. Сильно варьируют так же размеры (от 1 до 70 мкм) и форма митохондрий. При этом ширина этих частей клетки относительно постоянна (0,5-1 мкм). Способны изменять форму. в зависимости от того, в каких участках клетки в каждый конкретный момент происходит повышенное потребление энергии, митохондрии способны перемещаться по цитоплазме в зоны наибольшего энергопотребления, используя для движения структуры клеточного каркаса эукариотической клетки.
Красавица митохондрия в 3д представлении)
Альтернативой множеству разрозненных небольших митохондрий, функционирующих независимо друг от друга и снабжающих атф небольшие участки цитоплазмы, является существование длинных и разветвлённых митохондрий, каждая из которых может энергетически обеспечивать отдалённые друг от друга участки клетки. вариантом такой протяжённой системы может также являться упорядоченное пространственное объединение множества митохондрий (хондриом или митохондрион), обеспечивающее их кооперативную работу.
Особенно сложно этот тип хондриома устроен в мышцах, где группы гигантских разветвлённых митохондрий связаны друг с другом с помощью межмитохондриальных контактов (ммк). Последние образованы плотно прилегающими друг к другу наружными митохондриальными мембранами, в результате чего межмембранное пространство в этой зоне имеет повышенную электронную плотность ( много отрицательно заряженных частиц ). Особенно обильно ммк представлены в клетках сердечных мышц, где они связывают множественные отдельные митохондрии в согласованную работающую кооперативную систему.
Структура.
Наружная мембрана.
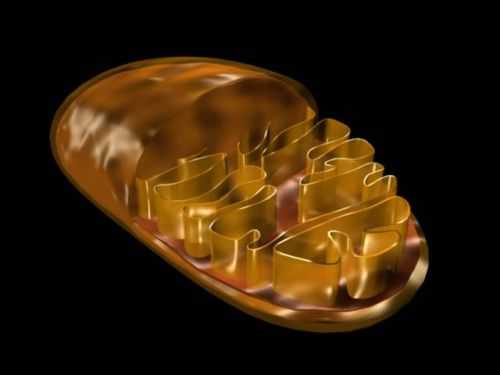
Наружная мембрана митохондрии имеет толщину около 7 нм, не образует впячиваний и складок, и замкнута сама на себя. на наружную мембрану приходится около 7 % от площади поверхности всех мембран клеточных органелл. Основная функция — отграничение митохондрии от цитоплазмы. Наружная мембрана митохондрии состоит из двойного жирового слоя ( как и у клеточной мембраны ) и пронизывающих его белков. Белки и жиры в равных пропорциях по массе.
Особую роль играет
Он формирует в наружной мембране отверстия диаметром 2-3 нм, через которые могут проникать небольшие молекулы и ионы. Крупные молекулы могут пересекать наружную мембрану только посредством активного транспорта через транспортные белки митохондриальных мембран. Наружная мембрана митохондрии может взаимодействовать с мембраной эндоплазматического ретикулума; это играет важную роль в транспортировке липидов и ионов кальция.
Внутренняя мембрана.
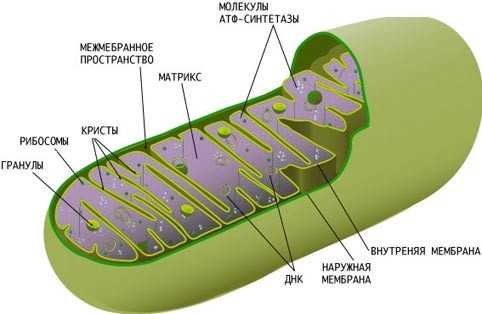
Внутренняя мембрана образует многочисленные гребневидные складки — кристы,
существенно увеличивающие площадь ее поверхности и, например, в клетках печени составляет около трети всех клеточных мембран. характерной чертой состава внутренней мембраны митохондрий является присутствие в ней кардиолопина — особый сложный жир, содержащего сразу четыре жирные кислоты и делающего мембрану абсолютно непроницаемой для протонов ( положительно заряженных частиц ).
Ещё одна особенность внутренней мембраны митохондрий — очень высокое содержание белков (до 70 % по весу), представленных транспортными белками, ферментами дыхательной цепи, а также крупными ферментами комплексами производящими атф. Внутренняя мембрана митохондрии в отличие от внешней не имеет специальных отверстий для транспорта мелких молекул и ионов; на ней, на стороне, обращенной к матриксу, располагаются особые молекулы ферменты производящие атф, состоящие из головки, ножки и основания. При прохождении через них протонов происходит создание атф.
В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты дыхательной цепи. наружная и внутренняя мембраны в некоторых местах соприкасаются, там находится специальный белок-рецептор, способствующий транспорту митохондриальных белков, закодированных в ядре, в матрикс митохондрии.
Матрикс.
Матрикс — ограниченное внутренней мембраной пространство. В матриксе ( розовом веществе ) митохондрии находятся ферментные системы окисления пирувата жирных кислот, а так же ферменты типа трикарбоновых кислот ( цикл дыхания клетки ). Кроме того, здесь же находится митохондриальная днк, рнк и собственный белоксинтезирующий аппарат митохондрии.
пируваты (соли пировиноградной кислоты)— важные химические соединения в биохимии. Они является конечным продуктом обмена веществ глюкозы в процессе ее расщепления.
Митохондриальная днк.
Несколько отличий от днк ядерной:
— митохондриальная днк – кольцевая, в отличии от ядерной днк, которая упакована в хромосомы.
— между различными эволюционными вариантами митохондриальной днк одного вида невозможен обмен сходными участками.
И поэтому вся молекула изменяется только путем медленного мутирования в течение тысячелетий.
— мутации кода в митохондриальных днк могут возникать независимо от ядерной днк.
Мутация ядерного кода днк возникает в основном при делении клетки, но митохондрии делятся независимо от клетки, и могут получать мутацию кода отдельно от ядерной днк.
— сама структура митохондриальной днк упрощена, т.к. многие составные процессы чтения днк утеряны.
— транспортные рнк имеют одинаковое строение. но рнк- митохондрий учавствуют только в синтезе митохондриальных белков.
Имея собственный генетический аппарат, митохондрия обладает и собственной белоксинтезирующей системой, особенностью которой в клетках животных и грибов являются очень маленькие рибосомы.
Функции.
Энергообразование.
Основной функцией митохондрий является синтез атф — универсальной формы химической энергии в любой живой клетке.
Данная молекула может образовываться двумя путями:
— путем реакции, в которых энергия освобождающаяся на определенных окислительных этапах брожения запасается в виде атф.
— благодаря энергии, выделяющейся при окислении органических веществв в процессе клеточного дыхания.
Митохондрии реализуют оба эти пути, первый из которых характерен для начальных процессов окисления и происходит в матриксе, а второй завершает процессы энергообразования и связан с кристами митохондрий.
При этом своеобразие митохондрий как энергообразующих органелл эукариотической клетки определяет именно второй путь генерации атф, получивший название «хемиосмотического сопряжения».
В целом весь процесс энергообразования в митохондриях может быть разбит на четыре основные стадии, первые две из которых протекают в матриксе, а две последние — на кристах митохондрий:
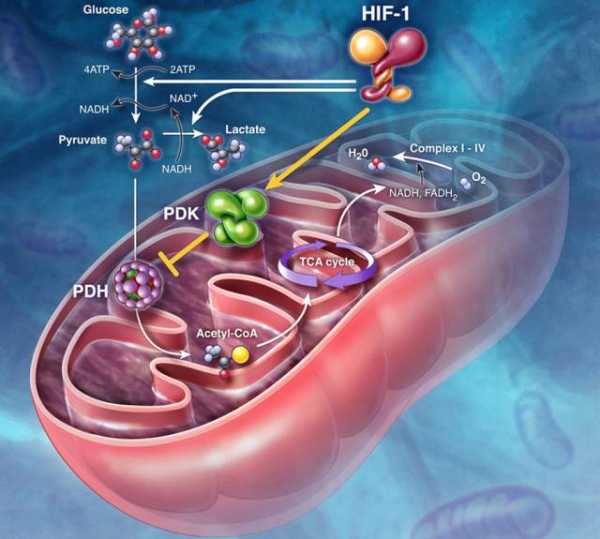
1) Превращение поступивших из цитоплазмы в митохондрию пируват ( конечный продукт расщепления глюкозы ) и жирных кислот в ацетил-коа;
ацетил-коа – важное соединение в обмене веществ, используемое во многих биохимических реакциях. его главная функция – доставлять атомы углерода (с) с ацетил-группой (ch4 co) в цикл клеточного дыхания, чтобы те были окислены с выделением энергии.
клеточное дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, жиров и аминокислот до углекислого газа и воды.
2) Окисление ацетил-соа в цикле клеточного дыхания, ведущее к образованию надн;
НАДН – кофермент, выполняет функцию переносчика электронов и водорода, которые принимает от окисляемых веществ.
3) Перенос электронов с надн на кислород по дыхательной цепи;
4) Образование атф в результате деятельности мембранного атф-создающего комплекса.
АТФ- синтетаза.
АТФ-синтетаза– станция по производству молекул АТФ.
Внимание [!!!] дальше очень заумный текст, описывающий принцип работы АТФ-синтазы, читать это и вникать только особо фанатичным личностям, желательно на светлую голову.
В структурно-функциональном плане АТФ-синтетаза состоит из двух крупных фрагментов, обозначаемых символами F1 и F0. Первый из них (фактор сопряжения F1) обращён в сторону матрикса митохондрии и заметно выступает из мембраны в виде сферического образования высотой 8 нм и шириной 10 нм. Он состоит из девяти субъединиц, представленных пятью типами белков. Полипептидные цепи трёх субъединиц α и стольких же субъединиц β уложены в похожие по строению белковые глобулы, которые вместе образуют гексамер (αβ)3, имеющий вид слегка приплюснутого шара.
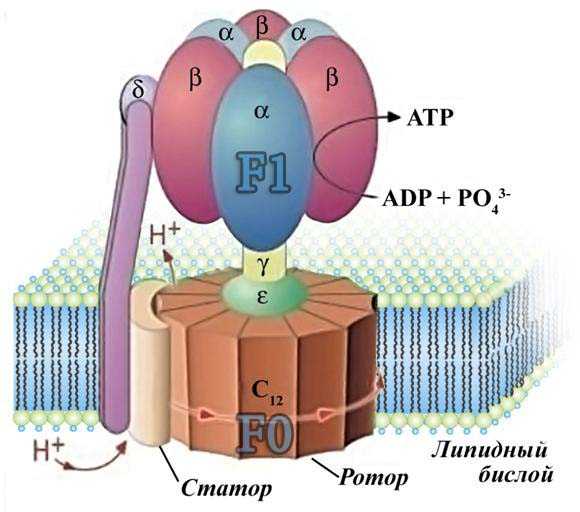
Субъединица – это структурный и функциональный компонент какой либо частицы
Полипептиды — органические соединения, содержащие от 6 до 80—90 аминокислотных остатков.
Глобула
Гексамер – соединение содержащее 6 субъедениц.
Подобно плотно уложенным долькам апельсина, последовательно расположенные субъединицы α и β образуют структуру, характеризующуюся симметричность относительно угла поворота 120°. В центре этого гексамера находится субъединица γ, которая образована двумя протяжёнными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. При этом нижняя часть субъединицы γ выступает из шара на 3 нм в сторону мембранного комплекса F0. Также внутри гексамера находится минорная субъединица ε, связанная с γ. Последняя (девятая) субъединица обозначается символом δ и расположена на внешней стороне F1.
Минорная – одиночная субъеденица.
Мембранная часть АТФ-синтетазы, представляет собой водо-отталкивающий белковый комплекс, пронизывающий мембрану насквозь и имеющий внутри себя два полуканала для прохождения протонов водорода. Всего в состав комплекса F0 входит одна белковая субъединица типа
Таким образом, в молекуле АТФазы можно выделить две группы белковых субъединиц, которые могут быть уподоблены двум деталям мотора: ротору и статору.
«Статор» неподвижен относительно мембраны и включает в себя шарообразный гексамер (αβ)3, находящуюся на его поверхности и субъединицу δ, а также субъединицы a и b мембранного комплекса F0.
Подвижный относительно этой конструкции «ротор» состоит из субъединиц γ и ε, которые, заметно выступая из комплекса (αβ)3, соединяются с погружённым в мембрану кольцом из субъединиц c.
Способность синтезировать АТФ — свойство единого комплекса F0F1, объедененного с переносом протонов водорода через F0 к F1, в последнем из которых как раз и расположены центры реакции, осуществляющие преобразование АДФ и фосфата в молекулу АТФ. Движущей же силой для работы АТФ-синтетазы является протонный ( положительно заряженный ) потенциал, создаваемый на внутренней мембране митохондрий в результате работы цепи электронного (отрицательно заряженного) транспорта.
Сила, приводящая в движение «ротор» АТФ-синтетазы, возникает при достижении разности потенциалов между наружной и внутренней сторонами мембраны > 220 10−3 Вольт и обеспечивается потоком протонов, протекающих через специальный канал в F0, расположенный на границе между субъединицами a и c. При этом путь переноса протонов включает в себя следующие структурные элементы:
1) Два расположенных на разных осях «полуканала», первый из которых обеспечивает поступление протонов из межмембранного пространства к существенно важным функциональным группам F0, а другой обеспечивает их выход в матрикс митохондрии;
2) Кольцо из субъединиц c, каждая из которых в своей центральной части содержит протонируемую карбоксильную группу ( COOH ), способную присоединять H+ из межмембранного пространства и отдавать их через соответствующие протонные каналы. В результате периодических смещений субъединиц с, обусловленных потоком протонов через протонный канал происходит поворот субъединицы γ, погружённой в кольцо из субъединиц с.
Таким образом, объединяющая активность АТФ-синтетазы непосредственно связана с вращением её «ротора», при котором поворот субъединицы γ вызывает одновременное изменение конформации всех трёх объединяющих субъединиц β, что в конечном счёте и обеспечивает работу фермента. При этом в случае образования АТФ «ротор» крутится по часовой стрелке со скоростью четыре оборота в секунду, а само подобное вращение происходит точными скачками по 120°, каждый из которых сопровождается образованием одной молекулы АТФ.
Работа АТФ-синтетазы связана с механическими движениями её отдельных частей, что позволило отнести этот процесс к особому типу явлений, названных «вращательным катализом». Подобно тому, как электрический ток в обмотке электродвигателя приводит в движение ротор относительно статора, направленный перенос протонов через АТФ-синтетазу вызывает вращение отдельных субъединиц фактора сопряжения F1 относительно других субъединиц ферментного комплекса, в результате чего это уникальное энергообразующее устройство совершает химическую работу — синтезирует молекулы АТФ. В дальнейшем АТФ поступает в цитоплазму клетки, где расходуется на самые разнообразные энергозависимые процессы. Подобный перенос осуществляется специальным встроенным в мембрану митохондрий ферментом АТФ/АДФ-транслоказой.
АДФ-транслоказа – пронизывающий внутреннюю мембрану белок, который обменивает вновь синтезированную АТФ на цитоплазматическую АДФ, что гарантирует сохранность фонда внутри митохондрий.
Митохондрии и наследственность.
ДНК митохондрий наследуются почти исключительно по материнской линии. Каждая митохондрия имеет несколько участков нуклеотидов в ДНК, идентичных во всех митохондриях (то есть в клетке много копий митохондриальных ДНК), что очень важно для митохондрий, неспособных восстанавливать ДНК от повреждений (наблюдается высокая частота мутаций). Мутации в митохондриальной ДНК являются причиной целого ряда наследственных заболеваний человека.
3д модель
Дисковери
С англ озвучкой
Немного о дыхании клетки и митохондрии на зарубежном языке
Структура строения
Просто анимация
http://www.youtube.com/watch?v=FFSdi3EJPys&feature=player_embedded
steelbros.ru
33. Митохондрии. Организация потока энергии в клетке.
Митохондрии -двухмембранные органеллы эукариотических клеток, обладающие собственной ДНК(испыт повреждения, тк нах в матриксе где много биоокислителей-митохондриальн.болезни) и выполняющие функцию синтеза АTФ.Митохондрии расположены около мест высокого потребления АТФ(в скелетных мышцах — вблизи миофибрилл. В сперматозоидах — футляр вокруг оси жгутика. В аксонах нервных кл-около синапсов, где происходит процесс передачи нервного импульса)
Поток энергии обеспечивается механизмами энергоснабжения —брожением, фото- или хемосинтезом, дыханием. Центральная роль в биоэнергетике клеток животных принадлежит дыхательному обмену. Он включает реакции расщепления глюкозы, жирных кислот, аминокислот, а также использование выделяемой энергии для образования «топлива» в виде аденозинтрифосфата (АТФ). Особое место в дыхательном обмене принадлежит митохондриям, выполняющим функцию окислительного фосфорилирования. Недоокисленные продукты гликолиза (пируват) поступают в матрикс митохондрий, где в условиях полного окисления, сопряженного с фосфорилированием АДФ до АТФ, отдают для нужд клетки оставшуюся в их химических связях энергию. Взаимосвязь внутриклеточного обмена белков, жиров и углеводов обеспечивает цикл Кребса.Ацетил-Коа взаимод с оксалоацетатом=цитрат. Далее в ходе 7-ми последовательных реакц 2 атома «С» удаляются в идее СО2, в конце концов регенерируется оксалоацетат. Высвобаждаются: 1АТФ и 1ФАДН2 с 3НАДН2, кот используются в окислительном фосфорилировании для синтеза АТФ путем перекачки протонов и электоронов. Окислительное фосфорилирование-внутр мембр митохондрий, где нах дых цепь(НАДН-дегидрогеназный компл, компл цитохромов b-c1,цитохромоксидазный компл) и комплекс АТФ-ситетазы. Электроны перемещ от одного компл к другому, достигая кислорода, имеющ наибольшее сродство к электрону. Переход на более низкие уровни сопровождается выделением энергии, использ для перекачки протонов в межмембранное пространство. Так возникает протонный градиент(мембр потенциал+градиент рН). При достижении определ разности потенциалов, АТФ-синтетаза начинает транспортировать протоны в матрикс, превращая АДФ и неорганич фосфат в энергию АТФ. ИТОГ: 34+2+2=38 АТФ Суть цикла: Запасение энергии в виде высокоэнергетическоемких электронов, входящих в соста атомов водорода, связ с НАД и ФАД, в электрон транспортную сеть. Суммарное уравнение может быть записано в следующем виде: Весь водород из молекулы глюкозы оказывается в конечном счете у переносчиков (НАД и ФАД). Весь углерод теряется в виде С02.
34. Образование атф в митохондриях.
См. 31 вопрос
35. Образование атф в клетке. Хемиосмотическая гипотеза Митчела.
См. 31 вопрос
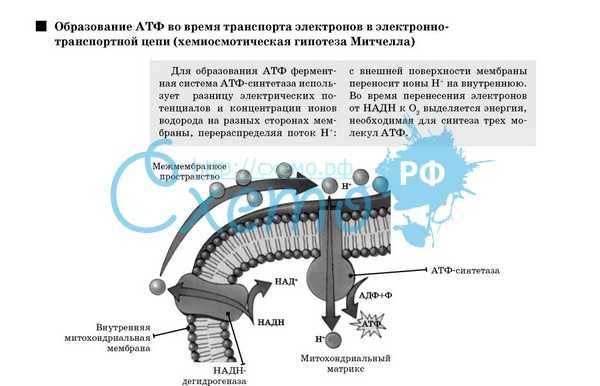
Хемиосмотическая теория:
Для образования АТФ ферментная система АТФ-синтетаза использует разницу электрохимических потенциалов и концентрации ионов водорода на разных сторонах мембраны, перераспределяя поток водорода: с внешней мембраны переносит ионы водорода на внутреннюю. Во время перенесения электронов от НАДН к кислороду выделяется энергия, необходимая для синтеза трех молекул АТФ.
studfiles.net
10. Митохондрии. Атф.
Митохондрии — микроскопические двумембранные полуавтономные органоиды общего назначения, обеспечивающие клетку энергией, получаемой благодаря процессам окисления и запасаемой в виде фосфатных связей АТФ. Митохондрии также участвуют в биосинтезе стероидов, окислении жирных кислот и синтезе нуклеиновых кислот. Присутствуют во всех эукариотических клетках. В прокариотических клетках митохондрий нет, их функцию выполняют мезосомы — впячивания наружной цитоплазматической мембраны внутрь клетки.
Митохондрии могут иметь эллиптическую, сферическую, палочковидную, нитевидную и др. формы, которые могут изменяться в течение определенного времени. Количество митохондрий в клетках, выполняющих различные функции, варьирует в широких пределах — от 50 и достигая в наиболее активных клетках 500-5000. Их больше там, где интенсивны синтетические процессы (печень) или велики затраты энергии (мышечные клетки). В клетках печени (гепатоцитах) их число составляет 800. а занимаемый ими объем равен примерно 20% объема цитоплазмы. Размеры митохондрий составляют от 0,2 до 1-2 мкм в диаметре и от 2 до 5-7 (10) мкм в длину. На светооптическом уровне митохондрии выявляются в цитоплазме специальными методами и имеют вид мелких зерен и нитей (что обусловило их название — от греч. mitos — нить и chondros — зерно).
В цитоплазме митохондрии могут располагаться диффузно, однако обычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл) органелл движения (аксонем спермия, ресничек), компонентов синтетического аппарата (цистерн ЭПС). Согласно одной из гипотез, все митохондрии клетки связаны друг с другом и образуют трехмерную сеть.
Митохондрия окружена двумя мембранами — наружной и внутренней, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны — кристы.
Наружная митохондриальная мембрана гладкая, по химическому составу сходна с наружной цитоплазматической мембраной и обладает высокой проницаемостью для молекул массой до 10 килодальтон, проникающих из цитозоля в межмембранное пространство. По своему составу она похожа на плазмалемму, 25% составляют белки, 75% липиды. Среди липидов присутствует холестерол. Наружная мембранаа содержит много молекул специализированных транспортных белков (например, поринов), которые формируют широкие гидрофильные каналы и обеспечивают ее высокую проницаемость, а также небольшое количество ферментных систем. На ней находятся рецепторы, распознающие белки, которые переносятся через обе митохондриальные мембраны в особых точках их контакта — зонах слипания.
Внутренняя мембрана имеет выросты внутрь — гребни или кристы, делящие матрикс митохондрии на отсеки. Кристы увеличивают площадь поверхности внутренней мембраны. Таким образом, внутреняя митохондриальная мембрана по площади превосходит наружную. Кристы расположены перпендикулярно или продольно длине митохондрии. Кристы по форме могут быть везикулярные, тубулярные или ламеллярные.
Химический состав внутренней мембраны митохондрий сходен с мембранами прокариот (например, в ней присутствует особый липид — кардиодипин и отсутствует холестерол). Во внутренней митохондриальной мембране преобладают белки, составляющие 75%. Во внутреннюю мембрану встроены белки трех типов (а) белки электрон-транспортной цепи (дыхательной цепи) — НАД’Н-дегидрогеназа и ФАД’Н дегидрогеназа — и другие транспортные белки, (б) грибовидные тельца АТФ-синтетазы (головки которых обращены в сторону матрикса) и (в) часть ферментов цикла Кребса (сукцинатдегидрогеназа). Внутренняя митохондриальная мембрана отличается чрезвычайно низкой проницаемостью, транспорт веществ осуществляется через контактные сайты. Низкая проницаемость внутренней мембраны для мелких ионов из-за высокого содержания фосфолипида
Митохондрии — полуавтономные органоиды клетки, т.к. содержат собственную ДНК, полуавтономную систему репликации, транскрипции и собственный белоксинтезируюший аппарат — полуавтономную систему трансляции (рибосомы 70S типа и т-РНК). Благодаря этому митохондрии синтезируют часть собственных белков. Митохондрии могут делиться независимо от деления клетки. Если из клетки удалить все митохондрии, то новые в ней не появятся. Согласно теории эндосимбиоза митохондрии произошли от аэробных прокариотических клеток, которые попали в клетку хозяина, но не переварились, вступили на путь глубокого симбиоза и постепенно, утратив автономность, превратились в митохондрии.
Митохондрии — полуавтономные органоиды, что выражается следующими признаками:
1) наличие собственного генетического материала (нити ДНК), что позволяет осуществлять синтез белка, а также позволяет самостоятельно делиться независимо от клетки;
2) наличие двойной мембраны;
3) пластиды и митохондрии способны синтезировать АТФ (для хлоропластов источник энергии — свет, в митохондриях АТФ образуется в результате окисления органических веществ).
Функции митохондрий:
1) Энергетическая — синтез АТФ (отсюда эти органоиды и получили название «энергетических станций клетки»):
При аэробном дыхание на кристах происходит окислительное фосфорилирование (образование АТФ из АДФ и неорганического фосфата за счет энергии, освободившейся при окислении органических веществ) и перенос электронов по электрон-транспортной цепи. На внутренней мембране митохондрии расположены ферменты, участвующие в клеточном дыхании;
2) участие в биосинтезе многих соединений (в митохондриях синтезируются некоторые аминокислоты, стероиды (стероидогенез), синтезируется часть собственных белков), а также накопление ионов (Са2+), гликопротеидов, белков, липидов;
3) окисление жирных кислот;
4) генетическая — синтез нуклеиновых кислот (идут процессы репликации и транскрипции). Митохондриальная ДНК обеспечивает цитоплазматическую наследственность.
АТФ
АТФ была открыта в 1929 году немецким химиком Ломанном. В 1935 году Владимир Энгельгардт обратил внимание на то, что мышечные сокращения невозможны без наличия АТФ. В период с 1939 под 1941 г. лауреат Нобелевской премии Фриц Липман доказал, что основным источником энергии для метаболической реакции является АТФ, и ввел в обращение термин «энергетически богатые фосфатные связи». Кардинальные изменения в изучении действия АТФ на организм произошли в середине 70-х годов, когда было обнаружено наличие специфических рецепторов на наружной поверхности клеточных мембран, чувствительных к молекуле АТФ. С тех пор интенсивно изучается триггерное (регуляторное) действие АТФ на различные функции организма
Аденозинтрифосфорная кислота (АТФ, аденинтрифосфорная кислота) — нуклеотид, играет исключительно важную роль в обмене энергии и веществ в организмах; в первую очередь соединение известно как универсальный источник энергии для всех биохимических процессов, протекающих в живых системах.
Химически АТФ представляет собой трифосфорный эфир аденозина, который является производным аденина и рибозы.
Пуриновое азотистое основание — аденин — соединяется β-N-гликозидной сзязью с 5′-углеродом рибозы, к которой последовательно присоединяются три молекулы фосфорной кислоты, обозначаемые соответственно буквами: α, β и γ.
АТФ относится к так называемым макроэргическим соединениям, то есть к химическим соединениям, содержащим связи, при гидролизе которых происходит освобождение значительного количества энергии. Гидролиз фосфоэфирных связей молекулы АТФ, сопровождаемый отщеплением 1 или 2 остатков фосфорной кислоты, приводит к выделению, по различным данным, от 40 до 60 кДж/моль.
АТФ + H2O → AДФ + H3PO4 + энергия
АТФ + H2O → AМФ + H4P2O7 + энергия
Высвобождённая энергия используется в разнообразных процессах, протекающих с затратой энергии
функции
1)Главная — энергетическая. АТФ служит непосредственным источником энергии для множества энергозатратных биохимических и физиологических процессов.
2) синтез нуклеиновых кислот.
3) регуляция множества биохимических процессов. АТФ, присоединяясь к регуляторным центрам ферментов, усиливает или подавляет их активность.
непосредственный предшественник синтеза циклоаденозинмонофосфата — вторичного посредника передачи в клетку гормонального сигнала.
медиатор в синапсах
пути синтеза:
В организме АТФ синтезируется из АДФ, используя энергию окисляющихся веществ:
АДФ + H3PO4 + энергия → AТФ + H2O.
Фосфорилирование АДФ возможно двумя способами: субстратное фосфорилирование и окислительное фосфорилирование. Основная масса АТФ образуется на мембранах в митохондриях путём окислительного фосфорилирования ферментом H-зависимой АТФ-синтетазой. Субстратное фосфорилирование АДФ не требует участия мембран, оно происходит в процессе гликолиза или путём переноса фосфатной группы с других макроэргических соединений.
Реакции фосфорилирования АДФ и последующего использования АТФ в качестве источника энергии образуют циклический процесс, составляющий суть энергетического обмена.
В организме АТФ является одним из самых часто обновляемых веществ. В течение суток одна молекула АТФ проходит в среднем 2000—3000 циклов ресинтеза (человеческий организм синтезирует около 40 кг в день), то есть запаса АТФ в организме практически не создаётся, и для нормальной жизнедеятельности необходимо постоянно синтезировать новые молекулы АТФ.
studfiles.net
Как вырабатывается энергия в митохондриях?
Митохо́ндрия (от греч. μίτος — нить и χόνδρος — зёрнышко, крупинка) — двумембранная гранулярная или нитевидная органелла толщиной около 0,5 мкм. Характерна для большинства эукариотических клеток как автотрофов (фотосинтезирующие растения) , так и гетеротрофов (грибы, животные) . Энергетическая станция клетки; основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии в синтезе молекул АТФ, который происходит за счёт движения электрона по электронно-транспортной цепи белков внутренней мембраны.Весь процесс энергообразования в митохондриях может быть разбит на 4 основные стадии, первые две протекают в матриксе, а еще две — на кристах митохондрий:
——Превращение поступивших из цитоплазмы пирувата и жирных кислот в ацетил-СоА;
——Окисление ацетил-СоА в цикле Кребса, ведущее к образованию НАДН;
——Перенос электронов с НАДН на кислород по дыхательной цепи;
——Образование АТФ в результате деятельности мембранного АТФ-синтетазного комплекса.

В цитоплазме шестиуглеродная молекула глюкозы частично окисляется до двух трёхуглеродных молекул пирувата с образованием двух молекул АТФ. Пируват переносится из цитозоля через наружную и внутреннюю мембраны в матрикс, где первоначально превращается в ацетил-СоА. Процесс катализируется крупным пируватдегидрогеназным комплексом, имеющим размер, сопоставимый с размером рибосомы, и состоящим из трёх ферментов, пяти коферментов и двух регуляторных белков. Точно так же жирные кислоты, полученные при расщеплении нерастворимых триглицеридов в цитоплазме, переносятся в митохондриальный матрикс в виде ацетил-СоА-производных.
На следующем этапе, также протекающем в матриксе митохондрии, ацетил-СоА полностью окисляется в цикле Кребса. Здесь задействованы 4 фермента, за каждый цикл обеспечивающие укорочение углеводородной цепи на два атома углерода, которые в дальнейшем превращаются в СО2. При этом образуется одна молекула ГТФ, а также НАДН — высокоэнергетического промежуточного соединения, которое легко отдаёт электроны в цепь переноса электронов на кристах митохондрий.
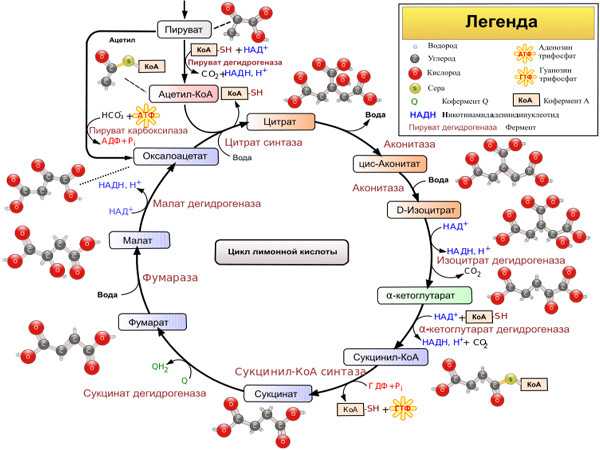
Цикл Кре́бса или Ци́кл трикарбо́новых кисло́т — центральная часть общего пути катаболизма, циклический биохимический аэробный процесс, в ходе которого происходит превращение двух- и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO2. Освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая участие в синтезе АТФ.
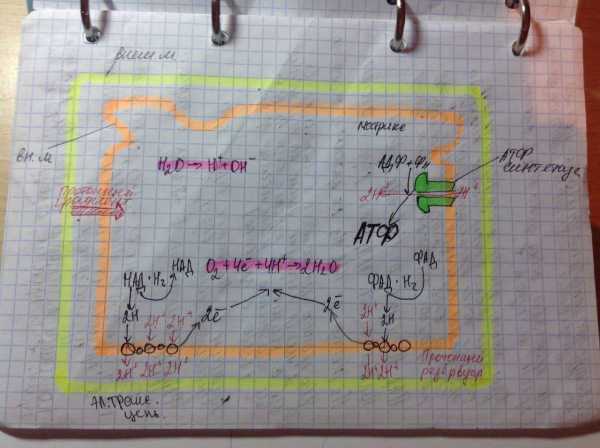
Дальнейшие процессы энергообразования в митохондрии происходят на её кристах и связаны с переносом электронов от НАДН к кислороду. В соответствии с тем, что потребление кислорода в качестве окислителя обычно называют «внутриклеточным дыханием» , электронно-транспортную цепь ферментов, осуществляющих последовательный перенос электронов от НАДН к кислороду, часто называют «дыхательной цепью» .
Трансформация энергии окисления осуществляется ферментами, расположенными на кристах митохондрий и осуществляющими векторный перенос протонов водорода из матрикса митохондрии в межмембранное пространство.
Завершающим этапом функционирования митохондрии является генерация АТФ, осуществляемая встроенным во внутреннюю мембрану специальным макромолекулярным комплексом.
Удачи Вам!
otvet.mail.ru
синтез ДНК, синтез АТФ, синтез углеводов или РНК ???
Митохо́ндрия (от греч. μίτος — нить и χόνδρος — зёрнышко, крупинка) — двумембранная гранулярная или нитевидная органелла толщиной около 0,5 мкм. Характерна для большинства эукариотических клеток как автотрофов (фотосинтезирующие растения) , так и гетеротрофов (грибы, животные) . Энергетическая станция клетки; основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии в синтезе молекул АТФ, который происходит за счёт движения электрона по электронно-транспортной цепи белков внутренней мембраны. Количество митохондрий в клетках различных организмов существенно отличается: так, одноклеточные зелёные водоросли (эвглена, хлорелла, политомелла) и трипаносомы имеют лишь одну гигантскую митохондрию, тогда как ооцит и амёба Chaos chaos содержат 300 000 и 500 000 митохондрий соответственно; у кишечных анаэробных энтамёб и некоторых других паразитических простейших митохондрии отсутствуют.Основной функцией митохондрий является синтез АТФ — универсальной формы химической энергии в любой живой клетке. Как и у прокариот, данная молекула может образовываться двумя путями: в результате субстратного фосфорилирования в жидкой фазе (например, при гликолизе) или в процессе мембранного фосфорилирования, связанного с использованием энергии трансмембранного электрохимического градиента (англ.) русск. протонов (ионов водорода) . Митохондрии реализуют оба эти пути, первый из которых характерен для начальных процессов окисления субстрата и происходит в матриксе, а второй завершает процессы энергообразования и связан с кристами митохондрий. При этом своеобразие митохондрий как энергообразующих органелл эукариотической клетки определяет именно второй путь генерации АТФ, получивший название «хемиосмотического сопряжения» . По сути это последовательное превращение химической энергии восстанавливающих эквивалентов НАДН в электрохимический протонный градиент ΔμН+ по обе стороны внутренней мембраны митохондрии, что приводит в действие мембранно-связанную АТФ-синтетазу и завершается образованием макроэргической связи в молекуле АТФ.
otvet.mail.ru
Митохондрии: общие сведения
Митохондрии: общие сведения
Митохондрия (mitochondrion): ДНК-содержащая цитоплазматическая органелла эукариот, вырабатывающая АТФ.
Митохондрии или хондриосомы (от греч. mitos — нить, chondrion — зернышко, soma — тельце) представляют собой гранулярные или нитевидные органоиды ( рис. 1, а ). Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. В таких клетках митохондрии могут двигаться, перемещаться, сливаться друг с другом. Особенно хорошо митохондрии выявляются на препаратах, окрашенных различными способами. Размеры митохондрий непостоянны у разных видов, так же изменчива их форма. Все же у большинства клеток толщина этих структур относительно постоянна (около 0,5 мкм), но длина колеблется, достигая у нитчатых форм 7-60 мкм.
Митохондрии независимо от их величины и формы имеют универсальное строение, их ультраструктура однообразна. Митохондрии ограничены двумя мембранами ( рис. 1, б ), у них четыре субкомпартмента: митохондриальный матрикс , внутренняя мембрана , мембранное пространство и внешняя мембрана , обращенная к цитозолю. Внешняя мембрана отделяет ее от остальной цитоплазмы. Толщина внешней мембраны около 7 нм, она не связана ни с какими другими мембранами цитоплазмы и замкнута сама на себя, так что представляет собой мембранный мешок. Наружную мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс, или митоплазму . Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные выпячивания (складки) внутрь митохондрий. Такие выпячивания ( кристы, рис. 27 ) чаще всего имеют вид плоских гребней. Митохондрии осуществляют синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования АДФ.
Митохондрии специализируются на синтезе АТФ путем транспорта электронов и окислительного фосфорилирования. (рис 21-1). Хотя они имеют свою собственную ДНК и аппарат белкового синтеза, большинство их белков кодируется клеточной ДНК и поступает из цитозоля. Более того, каждый поступивший в органеллу белок должен достичь определенного субкомпартмента, в котором он функционирует.
Митохондрии — это «энергетические станции» эукариотических клеток. В кристы встроены ферменты, участвующие в преобразовании энергии питательных веществ, поступающих в клетку извне, в энергию молекул АТФ. АТФ — «универсальная валюта», которой клетки расплачиваются за все свои энергетические расходы. Складчатость внутренней мембраны увеличивает поверхность, на которой размещаются ферменты, синтезирующие АТФ. Количество крист в митохондрии и количество самих митохондрий в клетке тем больше, чем больше энергетических трат осуществляет данная клетка. В летательных мышцах насекомых каждая клетка содержит несколько тысяч митохондрий. Меняется их количество и в процессе индивидуального развития (онтогенеза): в молодых эмбриональных клетках они более многочисленны, чем в клетках стареющих. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ, образующейся в митохондриях.
Расстояние между мембранами в кристе составляет около 10-20 нм. У простейших, одноклеточных водорослей в некоторых клетках растений и животных выросты внутренней мембраны имеют вид трубочек диаметром около 50 нм. Это так называемые трубчатые кристы.
Митохондриальный матрикс гомогенен и имеет более плотную консистенцию, чем окружающая митохондрию гиалоплазма. В матриксе выявляются тонкие нити ДНК и РНК, а также митохондриальные рибосомы, на которых синтезируются некоторые митохондриальные белки. С помощью электронного микроскопа на внутренней мембране и кристах со стороны матрикса можно увидеть грибовидные образования — АТФ-сомы. Это ферменты, образующие молекулы АТФ. Их может быть до 400 на 1 мкм.
Немногие белки, которые кодируются собственным геномом митохондрий, расположены в основном во внутренней мембране. Они обычно образуют субъединицы белковых комплексов, другие компоненты которых кодируются ядерными генами и поступают из цитозоля. Образование таких гибридных агрегатов требует сбалансирования синтеза этих двух типов субъединиц; каким образом координируется синтез белка на рибосомах разных типов, разделенных двумя мембранами, остается загадкой.
Обычно митохондрии располагаются в местах, где необходима энергия для любых жизненных процессов. Возник вопрос, каким образом транспортируется в клетке энергия — путем ли диффузии АТФ и нет ли в клетках структур, исполняющих роль электрических проводников, которые могли бы энергетически объединять отдаленные друг от друга участки клетки. Гипотеза заключается в том, что разность потенциалов в определенной области мембраны митохондрий передается вдоль нее и превращается в работу в другой области той же мембраны [ Скулачев В.П., 1989 ].
Как представлялось, подходящими кандидатами на эту же роль могли быть мембраны самих митохондрий. Кроме того, исследователей интересовали взаимодействие в клетке множественных митохондрий друг с другом, работа всего ансамбля митохондрий, всего хондриома — совокупности всех митохондрий.
Митохондрии характерны за малым исключением для всех эукариотических клеток как аутотрофных (фотосинтезирующие растения), так и гетеротрофных (животные, грибы) организмов. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии в синтезе молекул АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки.
Хондриом клетки (совокупность митохондрий) может иметь различную композицию в зависимости от энергетических потребностей клетки. В простейшем (и чаще встречающемся) случае он может быть представлен множеством разрозненных небольших митохондрий, функционирующих независимо друг от друга и снабжающих АТФ небольшие участки цитоплазмы. В других случях длинные и разветвленные митохондрии могут энергетически обеспечивать отдаленные друг от друга участки клетки. Вариантом такой протяженной системы может быть хондриом типа митохондриального ретикулума, который встречается как у одноклеточных, так и у многоклеточных организмов. Особенно сложно этот вид хондриома выражен в скелетных мышцах млекопитающих, где группы гигантских разветвленных митохондрий связаны друг с другом с помощью специальных контактов ММК . Наличие ММК характерно для хондриомов сократимых структур. Особенно обильно ММК представлены в клетках сердечных мышц, где они функционально связывают множественные отдельные митохондрии в единую разветвленную цепь.
Ссылки:
medbiol.ru
Функции митохондрий — Науколандия
Функции митохондрий как органелл аэробных эукариотических клеток – синтез молекул АТФ (аденозинтрифосфата) из АДФ. Поскольку АТФ является универсальным источником энергии для всех процессов в клетке, идущих с потреблением энергии, то говорят, что митохондрии выполняют функцию энергообеспечения, или энергообразования.
Из цитоплазмы в митохондрии поступают промежуточные продукты окисления органических веществ, кислород, АДФ, фосфорная кислота. Обратно выделяются углекислый газ, вода и молекулы АТФ.
Молекулы АТФ образуются не только в митохондриях. Небольшое их количество синтезируется в цитоплазме в процессе гликолиза, который наблюдается во всех клетках живого. В результате гликолиза молекула глюкозы разлагается на две молекулы пирувата. У аэробных прокариот далее он окисляется в присутствии кислорода на впячиваниях цитоплазматической мембраны. У эукариот же пируват поступает в митохондрии.
Здесь пируват, отдает свою ацетильную группу, содержащую два атома углерода, коферменту А. При этом выделяется первая молекула CO2. Кофермент А превращается в ацетил-кофермент-А (ацетил-КоА).
Ацетил-КоА получается не только из пирувата, но и жирных кислот, а также аминокислот. Так что не важно, какое исходное органическое вещество будет «сжигаться» в митохондриях для выработки энергии. Их функционирование в любом случае универсально.
В матриксе митохондрий ацетил-КоА вступает в цикл Кребса, или цикл трикарбоновых кислот, где ацетильная группа окисляется и разлагается до еще двух молекул CO2. Ее атомы водорода присоединяются к коферментам НАД+ и ФАД+, с образованием их восстановленных форм — НАД · H + Н+ и ФАД · H + Н+. Именно их последующее окисление приведет к синтезу АТФ.
Хотя в цикле Кребса кислород не используется, при его отсутствии митохондрия перестает выполнять свою функцию уже на этом этапе, так как накапливаются продукты реакции.
На кристах митохондрий происходит разделение электронов и протонов водорода. Электроны от НАД и ФАД передаются по мембране через цепь ферментов и коферментов, которую называют дыхательной цепью. Протоны же в начале пути перебрасываются в межмембранное пространство, на внешнюю сторону крист.
Электроны в конечном итоге передаются на молекулу кислорода, она превращается в отрицательно заряженный ион. Между внешней и внутренней сторонами крист создается электрический потенциал, так как одна заряжена положительно, а другая – отрицательно. Когда достигается критическое значение H+ устремляются через каналы АТФ-синтетазы и другие ферменты на внутреннюю сторону, где соединяются с O2— с образованием воды.
АТФ-синтетаза – это фермент, синтезирующий АТФ. В митохондриях он встроен в мембрану крист и использует энергию перемещающихся протонов для фосфорилирования АДФ.
Цикл Кребса и дыхательная цепь — это сложные многоступенчатые процессы, обеспечивающиеся целым рядом ферментов и коферментов. Каждый требует отдельного рассмотрения. В общих чертах же функции митохондрий сводятся к синтезу ацетил-КоА, использованию атомов водорода ацетильной группы для восстановления НАД и ФАД, раздельному переносу электронов и протонов водорода на кислород, использованию энергии электрохимического градиента протонов для синтеза АТФ.
Связанные статьи: Строение митохондрии, Этапы энергетического обмена
scienceland.info
