
Биоценозы и их изменения — Энциклопедия по биологии
Исторически в природе складываются сложные многосторонние пищевые взаимоотношения, которые могут быть выражены в различных цепях питания. Каждая из них включает 5—6 звеньев. Начальное звено такой цепи питания — автотрофный организм, создающий органическое вещество своего тела из веществ неорганических с использованием солнечной (зеленые растения) или химической энергии (хемосинтезу некоторых бактерий).
Зеленое растение служит пищей гетеротрофным организмам— травоядным животным. При этом установлено, что для построения 1 кг массы своего тела травоядное животное должно съесть не меньше 10 кг растительной пищи. Травоядные животные служат пищей для хищника. И в этом случае лишь 0,1 массы всей пищи, съеденной хищником, превратится в вещество его тела.
Таким образом, даже в этой предельно короткой цепи питания обнаруживается очень важная экологическая закономерность: при переходе от одного звена цепи питания к последующему только около 10% массы всей пищи идет на построение тела организма, а 90% массы всей пищи расходуется на поддержание жизнедеятельности организма. Именно поэтому цепи питания не бывают особенно длинными.
 Рис 116. Схема основных пищевых связей в пресноводном биоценозе (стрелки направлены от потребителя к пище, волнистые линии снизу — солнечная энергия)
Рис 116. Схема основных пищевых связей в пресноводном биоценозе (стрелки направлены от потребителя к пище, волнистые линии снизу — солнечная энергия)
Очень важной характеристикой биоценоза является его способность к саморегуляции. Численность любого вида, входящего в состав биоценоза, не остается постоянной, а из года в год колеблется. Однако такие колебания в пределах данной системы биоценоза не нарушают его целостности. Поясним это следующим примером. На той или иной территории численность хищников (волк, рысь) в первую очередь определяется численностью видов, поедаемых хищниками (грызуны, птицы). Усиленное размножение хищников приводит к тому, что их жертвы уничтожаются в очень большом количестве. Численность грызунов, некоторых птиц, например куропаток, резко падает. Хищникам перестает хватать корма — снижается темп их размножения, а это приводит к снижению их численности, что сразу же отражается на жертвах хищников: они начинают усиленно размножаться. На богатой кормовой базе (много грызунов) хищники вновь быстро повышают свою численность. И хотя этот пример пределъно упрощен, из него можно понять, в чем состоит процесс саморегуляции внутри биоценоза. На рис. 117 показаны данные о сопряженном колебании численности

Таким образом, устойчивость биоценоза обеспечивается подвижным равновесием между составляющими его видами.
Кроме саморегуляции, в природе имеет место и смена биоценозов. Наглядным примером смены биоценозов может служить процесс зарастания гарей. Старый биоценоз (хвойный лес) разрушен стихийным бедствием (пожаром). Первыми поселенцами гари оказываются травянистые растения, быстро занимающие освободившуюся территорию благодаря интенсивному половому и вегетативному размножению. Вместе с травянистой растительностью появляются и быстрорастущие сеянцы светолюбивых лиственных деревьев — березы, осины. В это время условий для возобновления старого биоценоза (хвойного леса) нет: медленнорастущие, плохо приспособленные к прямому солнечному свету сеянцы хвойных не могут конкурировать с травянистыми и древесными покрытосеменными растениями. Постепенно на месте хвойного леса формируется лиственный лес. Со сменой растительности меняется и фауна: исчезают виды, для которых пищей были семена хвойных (например, белки, клесты), появляются виды насекомых, чьи гусеницы питаются листьями деревьев, а вместе с ними в лесу поселяются и насекомоядные птицы. Травянистая и кустарниковая растительность привлекает копытных, а с копытными приходят и их извечные враги — хищники. Так, старый биоценоз заменяется новым биоценозом.
Однако такой лиственный лес, окруженный хвойным лесом,— явление временное. Под пологом березового леса сеянцы хвойных деревьев находят благоприятные условия. Пройдет столетие и менее устойчивый биоценоз (березовый лес) неизбежно будет заменен более устойчивым биоценозом — хвойным лесом со всеми присущими ему обитателями.
Смена биоценоза не всегда носит обратимый характер, который только что был описан. Так, в результате процесса заболачивания может иметь место следующая необратимая смена биоценозов: пресноводный водоем → болото → лес.
При сопоставлении биоценозов на разных широтах обнаруживается географическая зональность биоценозов. Так, в направлении с севера на юг прослеживается следующая закономерная замена одного биоценоза другим: тундра → хвойный лес → лиственный лес → степь → пустыня. В горных областях имеет место вертикальная зональность биоценозов, которая в некоторой степени как бы повторяет географическую зональность. В этом случае замена биоценозов идет в таком порядке (снизу вверх): степь → лиственный лес → хвойный лес → альпийские луга.
Рис 116. Схема основных пищевых связей в пресноводном биоценозе (стрелки направлены от потребителя к пище, волнистые линии снизу — солнечная энергия)
Очень важной характеристикой биоценоза является его способность к саморегуляции. Численность любого вида, входящего в состав биоценоза, не остается постоянной, а из года в год колеблется. Однако такие колебания в пределах данной системы биоценоза не нарушают его целостности. Поясним это следующим примером. На той или иной территории численность хищников (волк, рысь) в первую очередь определяется численностью видов, поедаемых хищниками (грызуны, птицы). Усиленное размножение хищников приводит к тому, что их жертвы уничтожаются в очень большом количестве. Численность грызунов, некоторых птиц, например куропаток, резко падает. Хищникам перестает хватать корма — снижается темп их размножения, а это приводит к снижению их численности, что сразу же отражается на жертвах хищников: они начинают усиленно размножаться. На богатой кормовой базе (много грызунов) хищники вновь быстро повышают свою численность. И хотя этот пример пределъно упрощен, из него можно понять, в чем состоит процесс саморегуляции внутри биоценоза. На рис. 117 показаны данные о сопряженном колебании численности
Таким образом, устойчивость биоценоза обеспечивается подвижным равновесием между составляющими его видами.
Кроме саморегуляции, в природе имеет место и смена биоценозов. Наглядным примером смены биоценозов может служить процесс зарастания гарей. Старый биоценоз (хвойный лес) разрушен стихийным бедствием (пожаром). Первыми поселенцами гари оказываются травянистые растения, быстро занимающие освободившуюся территорию благодаря интенсивному половому и вегетативному размножению. Вместе с травянистой растительностью появляются и быстрорастущие сеянцы светолюбивых лиственных деревьев — березы, осины. В это время условий для возобновления старого биоценоза (хвойного леса) нет: медленнорастущие, плохо приспособленные к прямому солнечному свету сеянцы хвойных не могут конкурировать с травянистыми и древесными покрытосеменными растениями. Постепенно на месте хвойного леса формируется лиственный лес. Со сменой растительности меняется и фауна: исчезают виды, для которых пищей были семена хвойных (например, белки, клесты), появляются виды насекомых, чьи гусеницы питаются листьями деревьев, а вместе с ними в лесу поселяются и насекомоядные птицы. Травянистая и кустарниковая растительность привлекает копытных, а с копытными приходят и их извечные враги — хищники. Так, старый биоценоз заменяется новым биоценозом.
Однако такой лиственный лес, окруженный хвойным лесом,— явление временное. Под пологом березового леса сеянцы хвойных деревьев находят благоприятные условия. Пройдет столетие и менее устойчивый биоценоз (березовый лес) неизбежно будет заменен более устойчивым биоценозом — хвойным лесом со всеми присущими ему обитателями.
Смена биоценоза не всегда носит обратимый характер, который только что был описан. Так, в результате процесса заболачивания может иметь место следующая необратимая смена биоценозов: пресноводный водоем → болото → лес.
При сопоставлении биоценозов на разных широтах обнаруживается географическая зональность биоценозов. Так, в направлении с севера на юг прослеживается следующая закономерная замена одного биоценоза другим: тундра → хвойный лес → лиственный лес → степь → пустыня. В горных областях имеет место вертикальная зональность биоценозов, которая в некоторой степени как бы повторяет географическую зональность. В этом случае замена биоценозов идет в таком порядке (снизу вверх): степь → лиственный лес → хвойный лес → альпийские луга.
biologiya.net
Виды биоценоза | Роль видов и связей в биоценозе
Всем известно, что на определённом участке земли или водоёма уживается вместе определённое количество организмов, растений и животных. Их совокупность, а так же взаимоотношение и взаимодействие между собой и с другими абиотическими факторами принято называть биоценозом. Это слово образовано путём слияния двух латинских слов «биос» — жизнь и «ценоз» — общие. Любое биологическое сообщество состоит из таких компонентов биоцеоза как:
Следует отметить, что фитоценоз является главенствующим компонентом, определяющим зооценоз и микробиоценоз.
Происхождение понятия «биоценоз»
В конце ХІХ века немецкий учёный Карл Мёбиус изучал места проживания устриц в Северном море. В ходе изучения он установил, что эти организмы могут существовать только в конкретных условиях, которые включают глубину, скорость течения, содержание соли и температуру воды. Помимо этого он отметил, что вместе с устрицами проживают строго определённые виды морских обитателей. Так в 1877 году с выходом его книги «Устрицы и устричное хозяйство» в научной среде появился термин и понятие биоценоза.
Классификация биоценозов
Сегодня бытует ряд признаков, в соответствии с которыми классифицируют биоценоз. Если мы говорим о систематизации на основе размеров, то это будет:
- макробиоценоз, который изучает горные массивы, моря и океаны;
- мезобиоценоз — леса, болота, луга;
- микробиоценоз — отдельно взятый цветок, лист или пень.
Так же биоценозы могут классифицироваться в зависимости от среды обитания. Тогда будут выделены следующие типы:
- морские;
- пресноводные;
- наземные.
Самой простой систематизацией биологических сообществ является их деление на природные и искусственные биоценозы. К числу первых относятся первичные, образованные без влияния человека, а так же вторичные, на которые повлияли природные стихии. Ко второй группе относятся те, кто претерпел изменения вследствие антропогенных факторов. Давайте более детально остановимся на их особенностях.
Природные биоценозы
Природные биоценозы представляют собой объединения живых существ, созданные самой природой. Такие сообщества являются исторически сложившимися системами, которые создаются, развиваются и функционируют по своим особенным законам. Немецкий учёный В. Тишлер обозначил следующие характеристики подобных образований:
- Возникают биоценозы с готовых элементов, которыми могут являться как представители отдельных видов, так и целые комплексы;
- отдельные части сообщества могут быть заменены другими. Так один вид может быть вытеснен другим, без негативных последствий для всей системы;
- с учётом того, что в биоценозе интересы разных видов противоположны, то вся надорганизменная система основана и держится благодаря действию силы противодействия;
- каждое природное сообщество строится путём количественной регуляции одного вида другим;
- размеры любых надорганизменных систем зависят от внешних факторов.
Искусственные биологические системы
Искуственные биоценозы формируются, поддерживаются и регулируются человеком. Профессор Б.Г. Иоганзен ввёл в экологию определение антропоценоза, то есть умышленно созданной человеком природной системы. Это может быть парк, сквер, аквариум, террариум и т.д.
Среди рукотворных биоценозов выделяют агробиоценозы — это биосистемы, созданные для получения продовольствия. К ним причисляют:
- водохранилища;
- каналы;
- пруды;
- пастбища;
- поля;
- лесопосадки.
Типичной особенностью агроценоза является тот факт, что он не в состоянии существовать длительный период времени без человеческого вмешательства.
ecoportal.info
Формирование, динамика, классификация биоценозов
Отношения между организмами в биоценозе
В структуре биоценоза находят отражение и отношения между организмами, которые невероятно сложны. В первую очередь это внутривидовые и межвидовые биотические отношения всех типов и трофические. В биоценозе организмы связаны также топическими, форическими и фабрическими отношениями. При технических один вид предоставляет себя для поселения другого вида (деревья используются птицами для гнездования). Вступая в форические отношения, организм одного вида способствует перемещению в пространстве организма другого вида (перенос малярийными комарами возбудителей малярии, птицами — пухоедов, млекопитающими — клещей, блох и других паразитов). При фабрических отношениях один вид применяет другой для строительства жилья, гнезд, убежищ и т. д. (бобр создает плотины и хатки, используя растительный материал).
Структурно– функциональная единица биоценоза, включающая самостоятельно существующее зеленое растение и все организмы, связанные с ним трофическими, топическими, форическими и фабрическими отношениями, называется консорцией. Самостоятельно существующее, т. е. не нуждающееся в опоре и не поселяющееся на другом виде, автотрофное растение является центром консорции, или детерминантой, по отношению к которой связанные с ним виды именуются консортами. Консортами могут быть паразиты и полупаразиты из мира растений, грибы, эпифиты, лианы, птицы, насекомые и животные–фитофаги, исключая консументов второго порядка, поедающих растительноядных животных.
Выделяют следующие типы консорций:
– индивидуальные (одного растения),
– ценопопуляционные (популяции вида в растительном сообществе),
– региональные,
– видовые.
Отношения между организмами в биоценозе определяются также временем их нахождения в сообществе.
Они могут быть постоянными (сессильными) и временными (вагильными). Постоянство характеризует главным образом растения, так как животные в большинстве случаев пребывают в сообществе временно в течение суток, сезона или в период миграции.
По Беклемишеву, межвидовые отношения подразделяются на четыре типа: трофические, топические, форические и фабрические.
Трофические связи возникают, когда один вид питается другим (либо живыми особями, либо их останками и продуктами жизнедеятельности).

Лес представляет собой отдельный биоценоз. Фото: Scott Wylie
Топические связи характеризуют любое, физическое или химическое изменение условий обитания одного вида в результате жизнедеятельности другого. Они заключаются в создании одним видом среды для другого, в формировании субстрата, во влиянии на движение воды, воздуха, в изменении температуры, насыщении среды продуктами выделения и т. п.
Форические связи – участие одного вида в распространении другого.
Фабрические связи – когда один вид использует для строительства своих сооружений продукты выделения или останки либо даже живых особей другого вида.
Динамика биоценозов
В целом сообществу присуща суточная, сезонная (годичная) и многолетняя динамика, свойственная как растениям, так и животным. Суточная, вызываемая сменой светлой и темной части суток, у растений проявляется в интенсивности фотосинтеза, дыхания, раскрывании и закрывании цветков, у животных — в разной суточной активности (дневные, сумеречные и ночные).
Нередко животные в течение суток меняют сообщество. Так, цапля кормится на мелководьях водоемов, а гнездится и ночует в кронах деревьев, насекомые– опылители (например, пчелы) могут перелетать из лесного сообщества в луговое.
Сезонная динамика биоценоза зависит от фенологического состояния фитоценоза, видового состава и численности обитающих в нем животных. Каждый вид растительных организмов в течение вегетационного периода проходит определенные стадии развития (начало вегетации, цветение, плодоношение и отмирание). В фитоценозе, состоящем из множества видов, фазы развития растений могут совпадать и не совпадать.
Внешний вид фитоценоза, изменяющийся на протяжении года с чередованием фаз развития, называется аспектом. Как правило, аспект повторяется из года в год с неизменной последовательностью, отражая цветовую гамму растительного сообщества (весенняя яркая зелень, летнее разноцветье и осенняя пестрота лесов). Аспект именуют обычно по растениям, придающим фитоценозу наиболее заметную цветовую окраску, например голубой аспект незабудки болотной, белый аспект пушицы, бурый аспект листьев осоки и т. д.
Сезонная динамика животных представителей биоценоза связана с их размножением, жизненной активностью и миграциями. Весенний прилет и осенний отлет птиц, нерест рыб, появление молодняка, активность насекомых– опылителей на лугах, зимняя спячка медведя только ничтожно малая часть примеров сезонной динамики животного населения биоценоза.
Многолетняя динамика сообщества вызывается его повторяющимися изменениями на протяжении нескольких лет при отсутствии резкой смены видового состава. Изменения затрагивают в основном численность особей видов, образующих биоценоз. В качестве примера можно привести изменения в лесах некоторых заповедников Беларуси и России, обусловленные увеличением численности лося – основного потребителя древесно–кустарниковых кормов. За год лось съедает около 7 т кормов, причем более половины составляют побеги лиственных и хвойных пород. При увеличении плотности животного порча подроста увеличивается. Наступает период, когда в лесном насаждении почти полностью уничтожается молодое поколение древостоя. Из–за бескормицы лоси вынуждены покидать такие участки леса.
Стадии формирования биоценозов
Возникновение биоценоза начинается с появления первых организмов на лишенных жизни участках (лавовых потоках, вулканических островах, осыпях, обнажившихся горных породах, песчаных наносах и обсохших днищах водоемов). Заселение начинается со случайного заноса организмов с уже освоенных ими территорий и зависит от свойств субстрата. Этот участок для многих семян растений и проникших сюда животных может оказаться неподходящим для размножения. Нередко, особенно в гумидной зоне, первыми поселенцами оказываются представители водорослей, мхов и лишайников.
Как правило, успешно развиваются лишь немногие из занесенных растительных видов. Животные– консументы поселяются несколько позднее, так как их существование без пищи невозможно, но случайное посещение ими осваиваемых участков — довольно частое явление. Данная стадия развития биоценоза получила название пионерной. Хотя на этой стадии сообщество еще не сложилось (непостоянный видовой состав, разреженный растительный покров), оно уже оказывает влияние на абиотическую среду: начинает образовываться почва.
Пионерная стадия сменяется ненасыщенной, когда растения начинают возобновляться (семенами или вегетативным путем),_а животные размножаться. В ненасыщенном биоценозе заняты не все экологические ниши.
Постепенно темпы заселения участка возрастают за счет как увеличения числа особей пионерной растительности до образования зарослей, так и внедрения новых видов. Видовой состав такого сообщества пока неустойчив, новые виды внедряются довольно легко, хотя конкуренция начинает играть заметную роль. Эта стадия развития биоценоза группировка.
При последующем развитии сообщества происходит дифференциация растительного покрова по ярусам и синузиям, устойчивое постоянство приобретают его мозаичность, видовой состав, пищевые цепи и консорции. В конечном итоге занятыми оказываются все экологические ниши и дальнейшее вселение организмов становится возможным только после – вытеснения или уничтожения старожилов. Данная, заключительная, стадия образования биоценоза получила название насыщенной. Однако дальнейшее развитие биоценоза не прекращается и случайные отклонения в видовом составе и взаимоотношениях как между организмами, так и с окружающей средой еще могут иметь место.
Случайные отклонения в структуре биоценоза называются флуктуациями. Как правило, они обусловлены случайными или сезонными изменениями численности видов, входящих в биоценоз, в результате неблагоприятных метеорологических явлений, наводнений, землетрясений и т. д. Обильные снегопады и иней, например, приводят к изреживанию крон, и весной под пологом леса большее развитие получают злаки. Весенние заморозки и поздневесенний снежный покров не только повреждают цветущие растения, что сказывается на их плодоношении, но и нередко служат причиной массовой гибели перелетных птиц. Сильные ветры, наводнения и землетрясения вызывают нарушения в биоценозах, после которых для восстановления сообщества требуется продолжительное время.
Хотя биоценоз представляет собой довольно консервативную природную систему, однако под давлением внешних обстоятельств он может уступить место другому биоценозу. Последовательная смена во времени одних сообществ другими на определенном участке среды называется сукцессией (от. Лат successia преемственность, наследование). В результате сукцессии одно сообщество последовательно сменяется другим без возврата к исходному состоянию. К сукцессии приводит взаимодействие организмов, главным образом ранений, друг с другом и со средой.
Сукцессии подразделяются на первичные– историчные. Первичные происходят на первичносвободных от почвы Грунтах — вулканических туфовых и лавовых полях, сыпучих песках, каменистых россыпях и т. д. По мере развития фитоценоза от пионерной стадии до насыщенной почва становится плодороднее и в биологический круговорот вовлекается все больше химических элементов в возрастающем количестве. С увеличением плодородия виды растений, развивающиеся на богатых питательными веществами почвах, вытесняют менее требовательные в этом отношении виды. Одновременно сменяется и животное население. Вторичные сукцессии осуществляются на местообитаниях разрушенных сообществ, где сохранилась почвы и некоторые живые организмы. Разрушение биоценозов может вызываться стихийными природными процессами (ураганы, ливни, наводнения, оползни, продолжительные засухи, извержение вулканов и т. д.). а также изменение Организмами среды обитания (например, при зарастании водоема водная среда сменяется отложениями торфа). Вторичные сукцессии характерны для деградированных пастбищ, гарей, вырубок леса, исключенные из сельскохозяйственного пользования пашен и других угодий. а также для искусственных лесопосадок. Например, нередко под пологом средневозрастных культур сосны на супесчаных почвах начинается обильное естественное возобновление ели, которая со временем вытеснит сосну при условии, если не будут проводиться очередные сплошные рубки соснового древостоя и лесокультурные работы. На гарях с супесчаными и суглинистыми почвами пионерная растительность из иван– чая и березы бородавчатой со временем сменяется еловыми насаждениями.
В последние десятилетия особое значение в изменении растительного покрова приобрели крупномасштабные осушительные и оросительные работы. В болотных лесах, оказавшихся в зоне влияния осушительных каналов, исчезают растения– гигрофиты (ольсы осоковые, например, трансформируются в крапивные). Преобразование видового состава, включая животное население, затрагивает и лесные массивы, прилетающие к осушаемым болотам. Оросительные мелиорации, наоборот, способствуют активному проникновению растений гигрофильной и мезофильной групп на переувлаженные участки в результате скопления вод, использованных для орошения. Заметное влияние на биоценозы оказывает и промышленное загрязнение среды. Все эти изменения являются вторичными сукцессиями.
Смена одного биоценоза другим в ходе сукцессии образует сукцессионный ряд, или серию. Изучение сукцессионных рядов имеет большое значение в связи с усиливающимся антропогенным влиянием на биоценозы. Конечным результатом такого рода исследований может быть прогнозирование образования природно–антропогенных ландшафтов. Изучение вторичных сукцессии и факторов, их вызывающих, играет важную роль в решении проблем охраны и рационального использования биологических и земельных ресурсов.
Если естественное течение сукцессии не нарушается, сообщество постепенно приходит к относительно устойчивому состоянию, в котором поддерживается равновесие между организмами, а также между ними и средой, – к климаксу. Без вмешательства человека этот биоценоз может существовать неопределенно долго, например, сосняк черничный, лишайниковая тундра на песчаных почвах.
Концепция климакса детально разработана американским ботаником X. Каулсом и широко используется в зарубежной ботанико–географической литературе. Согласно данной концепции, климакс — терминальная стадия эволюции сообщества, которой соответствует почва определенного типа — педоклимакс. Сукцессии, ведущие к этой стадии, называются прогрессивными, а удаляющие биоценоз от нее, — регрессивными. Нельзя, однако, придавать понятию «климакс» абсолютное значение и полагать, что при достижении его сообщество прекращает развитие.
Биоценозы, которые, будучи нарушенными, возвращаются к своему исходному состоянию, называются коренными. На месте вырубки сосняка черничного или ельника кисличного вырастет березняк, а он, в свою очередь, опять сменится сосняком черничным или ельником кисличным. В данном случае речь идет о коренных типах леса.
Трансформированные биоценозы не возвращаются в исходное состояние. Так, осушенное и освоенное под посевы сельскохозяйственных культур низинное осоковое болото после сработки торфяной залежи и разрушения мелиоративной сети с прекращением по каким–либо причинам сельскохозяйственного использования развивается в направлении формирования березового или ольхового мелколесья. Зооценоз этого мелколесья отличается от сообщества видов животных открытого травяного болота.
Классификация биоценозов
В целях научного познания биоценозов и практического применения знаний о них сообщества организмов необходимо классифицировать по их относительной размерности и сложности организации.
Классификация призвана привести в порядок все их многообразие с помощью системы таксономических категорий, т. е. таксонов, объединяющих в данном случае группы биоценозов с той или иной степенью общности отдельных свойств и признаков, а также строения и происхождения. При этом должны быть соблюдены определенная соподчиненность простых по содержанию таксонов сложным, таксонов малой (локальной) размерности таксонам планетарной размерности, постепенное усложнение их организации. Кроме того, при классификации биоценозов следует учитывать наличие возможных границ между ними.
Особых затруднений в установлении границ не возникает в том случае, когда соседствующие биоценозы имеют четкие индикационные признаки. Например, верховое болото с багульниково– моховым покровом и низкорослым сосновым древостоем контрастирует с окружающим его сосновым лесным сообществом на песчаных почвах. Граница между лесом и лугом также явно прослеживается. Однако, поскольку условия существования сообществ сменяются более постепенно, чем сами сообщества, границы биоценозов обычно размыты. Постепенность перехода от одного фитоценоза к другому при их соседстве и смене одного фитоценоза другим во времени отражена в концепции о континууме (от лат. continuum – непрерывное) растительности, развиваемой советским геоботаником Л. Г. Раменским, американским экологом P. X. Уиттекером.
Более резко границы между сообществами выступают в тех случаях, когда эдификаторы оказывают наибольшее преобразовательное воздействие на среду, например границы между лесными массивами, образованными разными древесными породами – сосной, елью, дубом и другими. В степях, полупустынях и пустынях границы между сообществами более постепенные, так как средопреобразующая роль травянистых видов оказывается менее контрастной.
В классификации сообществ используются таксономические категории, принятые в географии растений и основанные на выделении доминант и эдификаторов, что свидетельствует о признании фитоценоза как экологического каркаса, определяющего структуру биоценоза. Построенная по доминантам и эдификаторам таксономическая система сообществ может быть выражена следующим рядом: ассоциация – группа ассоциаций формация группа формаций класс формаций тип биома — биоценотический покров.
Самой низшей таксономической категорией является ассоциация. Она представляет собой совокупность однородных микробиоценозов с одинаковыми структурой, видовым составом и сходными отношениями как между организмами, так и между ними и средой. В полевых условиях основными признаками ее выделения служат: одно и то же ярусное сложение, сходная мозаика (пятнистая, рассеянная), совпадение доминант и эдификаторов, а также относительная однородность местообитания. Название ассоциации для многоярусных сообществ состоит из родовых названий доминанты господствующего яруса (кондоминанты) и эдификаторов в каждом ярусе, например сосняк можжевельниково–мшистый, ельник березово–черничный и т. д. Наименование сложных луговых ассоциаций образуется перечислением доминант и субдоминант, причем доминанта называется последней, например едколютично–лугово–мятликовая ассоциация. Обычно луговые ассоциации обозначаются на латинском языке: Ranunculus + Роа pratensis.
Группу биоценотических ассоциаций образуют ассоциации, различающиеся составом одного из ярусов. Сосняк черничный, например, объединяет ассоциации с ярусом подлеска из можжевельника, крушины и подроста березы. Группа злаково– мелкоосоково– разнотравных ассоциаций включает луговые сообщества с набором названных групп луговых трав (злаки, мелкие осоки, разнотравье).
В биоценотическую формацию входят группы ассоциаций. Формацию выделяют по доминанте, по которой она и называется: формация сосны обыкновенной, ольхи черной, дуба черешчатого, саксаула белого, лютика едкого, полыни горькой и т. д. Это основная единица среднего ранга, широко используемая при картографировании лесной растительности.
Группа формаций — это все формации, доминанты которых принадлежат к одной и той же жизненной форме. Поскольку жизненные формы растений чрезвычайно разнообразны, объем групп формаций неоднороден: темно– хвойные, светлохвойные, листопадные, вечнозеленые, мелколиственные и широколиственные леса; крупнозлаковые, мелкозлаковые, низкозлаковые, мелкоразнотравные и другие группы луговых формаций.
Класс формаций образуется всеми группами формаций, доминанты которых имеют экологически близкие жизненные формы, например хвойные леса (с игольчатой пластинкой), лиственные леса и т.д.
Тип биома (биоценотический тип) объединяет классы формаций. Типами биома являются тундра, лесотундра, тайга, луга, степи, пустыни, прерии, влажные тропические леса и т. д.
Биоценотический покров представляет собой высшую таксономическую единицу, включающую все типы биомов суши.
В ботанико–географической литературе встречаются и другие классификации фитоценозов. Для водной среды, в которой роль растительности ограничена, выделение таксономических категорий биоценозов основывается на животном населении.
Каждый биогеоценоз имеет свойственную ему пространственную структуру, которая в вертикальном направлении выражается в ярусности, а в горизонтальном в синузиях. Непрекращающиеся взаимодействия и взаимоизменения компонентов биогеоценоза (атмосферы, почвы и горной породы, воды, животного и растительного мира и микроорганизмов) обусловливают его непрерывное развитие, что приводит к смене одних биогеоценозов другими – к сукцессиям. В конечном итоге разрушение одних сообществ и созидание новых определяет непрерывное развитие биогеоценотического покрова Земли. Со временем непрерывное изменение отдельного биогеоценоза замедляется, так как ослабевает процесс вселения новых организмов и наступает климаксовая стадия.
Саморазвитие биогеоценоза, определяемое внутренними (эндогенными) процессами, нарушается внешними (экзогенными) воздействиями, в результате чего возникают новые сукцессионные ряды. К числу важнейших экзогенных факторов относится деятельность человека, но сам человек не входит в число компонентов биогеоценоза.
Биогеоценозы представляют собой элементарные ячейки биогеосферы (биогеоценотического покрова) – оболочки Земли, в которой сконцентрировано живое вещество планеты. Биогеосфера является единственной оболочкой Земли, в которой возможны постоянное нахождение и нормальная всесторонняя деятельность человека.
biofile.ru
Учебное пособие для 11-го класса
[ Содержание учебника Общая биология: Учебное пособие для 11-го класса ]
Вернуться к графической версии учебника…
§ 5. Биоценоз. Многообразие биоценозов
Понятие биоценоза. Живые организмы встречаются на Земле не в любых случайных сочетаниях, как независимые особи, а образуют закономерные комплексы (сообщества). Впервые на возможность выделения таких сообществ обратил внимание немецкий биолог Карл Август Мёбиус (1825— 1908). В 1877 г. он предложил для обозначения комплекса живых организмов, постоянно встречающихся вместе, при наличии одинаковых условий существования, термин биоценоз (от греч. bios — жизнь и koinos — общий, делать что-либо общим).
Биоценоз — это исторически сложившаяся группировка растений, животных, грибов и микроорганизмов, населяющих относительно однородное жизненное пространство (участок суши или водоема) (рис. 2.1).
Итак, каждый биоценоз состоит из определенной совокупности живых организмов, относящихся к разным видам. Но мы знаем, что особи одного вида объединяются в природные системы, которые называются популяциями. Поэтому биоценоз может быть определен также и как совокупность популяций всех видов живых организмов, заселяющих общие места обитания.
В состав биоценоза входят совокупность растений на определенной территории — фитоценоз (от греч.phyton — растение), совокупность животных, проживающих в пределах фитоценоза, — зооценоз (от греч.zoon — животное), микробоценоз — совокупность микроорганизмов, населяющих почву, и микоценоз (от греч. mykes — гриб) — совокупность грибов. Примерами биоценозов являются лиственный, еловый, сосновый или смешанный лес, луг, болото и т.д.
Каждый биоценоз развивается в пределах однородного пространства, которое характеризуется определенным сочетанием абиотических факторов, таких как количество приходящей солнечной радиации, температура, влажность, химический и механический состав почвы, ее кислотность, рельеф местности и др. Такое однородное пространство (часть абиотической среды), занимаемое биоценозом, называется биотоп.Это может быть какой-либо участок суши или водоема, берег моря или склон горы. Биотоп — это неорганическая среда, которая является необходимым условием существования биоценоза. Между биоценозом и биотопом существует тесное взаимодействие.
Масштабы биоценозов могут быть различны — от сообществ подушек лишайников на стволах деревьев, моховых кочек на болоте или разлагающегося пня до населения целых ландшафтов. Так, на суше можно выделить биоценоз суходольного (незаливаемого водой) луга, биоценоз сосняка-беломошника, биоценоз ковыльной степи, биоценоз пшеничного поля и т .д.
В водной среде биоценозы обычно выделяют в соответствии с экологическими подразделениями водоемов — биоценоз прибрежных песчанистых или
илистых грунтов, биоценоз приливной зоны моря, биоценоз крупных водных растений прибрежной зоны озера, биоценоз пресного водоема и т.д. (рис. 2.2).
В конкретный биоценоз включаются не только организмы, постоянно обитающие на определенной территории, но и те, которые оказывают существенное воздействие на его жизнь, хоть и обитают в других биоценозах.
Например, многие насекомые размножаются в водоемах, где являются важным источником питания рыб и некоторых других животных. В молодом возрасте они входят в состав водного биоценоза, а во взрослом состоянии ведут наземный образ жизни, т.е. выступают как элементы сухопутных биоценозов. Зайцы могут питаться на лугу, а обитать в лесу. То же касается и многих видов лесных птиц, которые ищут себе пропитание не только в лесу, а и на прилегающих лугах или болотах.
Видовая структура биоценоза. Каждый биоценоз можно описать, основываясь на совокупности составляющих его видов. Видовое разнообразие различных биоценозов разное, что обусловлено их разным географическим положением. Установлено: оно уменьшается по направлению от тропиков в сторону высоких широт, что объясняется ухудшением условий жизни организмов.
Например, во влажных тропических лесах Малайзии на 1 га леса можно насчитать до 200 видов древесных пород. Биоценоз соснового леса в условиях Беларуси может включать максимум до десяти видов деревьев на 1 га, а на севере таежной области на такой же площади присутствует 2—5 видов. Наиболее бедными биоценозами по набору видов являются альпийские и арктические пустыни, самыми богатыми — тропические леса.
Если какой-либо вид растения (или животного) количественно преобладает в сообществе (имеет большую биомассу, продуктивность или численность), то такой вид называется доминантным, или доминирующим.
Доминантные виды есть в любом биоценозе. В дубраве это могучие дубы. Используя основную долю солнечной энергии и наращивая наибольшую биомассу, они затеняют почву, ослабляют движение воздуха и создают особые условия для жизни других обитателей леса.
Однако кроме дубов в дубраве проживает большое количество других организмов. Например, дождевые черви, живущие здесь, улучшают физические и химические свойства почвы, пропуская через пищеварительную систему частицы отмерших растений и опавших листьев. Дуб и червь вносят свой особый вклад в жизнедеятельность биоценоза, однако роль дуба здесь определяющая, поскольку вся жизнь дубового леса обусловливается этой древесной породой и связанными с ней растениями. Поэтому именно дуб является доминирующим видом в таком лесу.
Пространственная структура биоценоза. Виды распределяются в пространстве в соответствии с их потребностями и условиями местообитания. Такое распределение в пространстве видов, составляющих биоценоз, называется пространственной структурой биоценоза. Различают вертикальную и горизонтальную структуру биоценоза.
Вертикальная структура биоценоза образована отдельными его элементами, особыми слоями, которые называются ярусами. Ярус — совместно произрастающие группы видов растении, различающиеся по высоте и положению в биоценозе ассимилирующих органов (листья, стебли, подземные органы — клубни, корневища, луковицы и т.п.). Как правило, разные ярусы образованы разными жизненными формами (деревьями, кустарниками, кустарничками, травами, мхами). Наиболее четко ярусность выражена в лесных биоценозах (рис. 2.3). Так, первый ярус здесь обычно формируют самые большие деревья с высоко расположенной листвой, которая хорошо освещается солнцем. Неиспользованный свет может поглощаться деревьями поменьше, образующими второй, подпологовый, ярус. Около 10 % солнечной радиации перехватывается ярусом подлеска, который образуют различные кустарники, и только от 1 до 5 % — растениями травяного покрова (травяно-кустарничковый ярус).
Напочвенный слой мхов и лишайников формирует мохово-лишайнико-вый ярус. Итак, схематично в лесном биоценозе выделяется 5 ярусов.
Подобно распределению растительности разные виды животных в биоценозах также занимают определенные уровни (рис. 2.4). В почве живут почвенные черви, микроорганизмы, землеройные животные. В листовом опаде, на поверхности почвы живут различные многоножки, жужелицыклещи и другие мелкие животные. В верхнем пологе леса гнездятся птицы, причем одни могут питаться и гнездиться ниже верхнего яруса, другие — в кустарниках, а третьи — возле самой земли. Крупные млекопитающие обитают в нижних ярусах.
Ярусность наблюдается также в биоценозах океанов и морей. Разные виды планктона держатся на разной глубине, в зависимости от освещения, а разные виды рыб — в зависимости от того, где они находят себе пропитание.
Живые организмы распределены в пространстве неравномерно. Обычно они составляют группировки, что является приспособительным фактором в их жизни. Такие группировки организмов определяютгоризонтальную структуру биоценоза.
Расчлененность в горизонтальном направлении — мозаичность — свойственна практически всем биоценозам. Примеров такого распределения можно привести множество. Огромными косяками передвигаются с места на место многие виды рыб. В большие стаи собираются водоплавающие и воробьиные птицы, готовящиеся к дальним перелетам. Североамериканские олени карибу в условиях тундры образуют огромные стада. В южноамериканских тропиках группы муравьев, вооруженные могучими челюстями и жалами, выстраиваются фронтом 20-метровой ширины и идут в атаку, истребляя всех, кто замешкался и не в силах спастись бегством.
Такие же примеры можно привести и для растений: пятнистое размещение особей клевера на лугу, пятна мхов и лишайников, скопление кустарничков брусники в сосновом лесу, обширные пятна кислицы в еловом лесу, земляничные поляны на светлых опушках.
Наличие мозаичности имеет важное значение для жизни сообщества. Мозаичность позволяет более полно использовать различные типы микроместообитаний. Особям, образующим группировки, свойственна высокая выживаемость, они наиболее эффективно используют пищевые ресурсы. Это ведет к увеличению численности и разнообразию видов в биоценозе, способствует его устойчивости и жизнеспособности.
Отношения организмов в биоценозах. Особи разных видов существуют в биоценозах не изолированно; они вступают между собой в разнообразные прямые и косвенные отношения. Прямые отношения разделяют на четыре типа: трофические, топические, форические, фабрические.
Трофические отношения возникают тогда, когда один вид в биоценозе питается другим (либо мертвыми остатками особей этого вида, либо продуктами их жизнедеятельности). Божья коровка, питающаяся тлей, корова на лугу, поедающая сочную траву, волк, охотящийся на зайца, — это все примеры прямых трофических связей между видами.
Топические отношения характеризуют изменение условий обитания одного вида в результате жизнедеятельности другого. Ель, затеняя почву, вытесняет светолюбивые виды из-под своей кроны, ракообразные поселяются на коже китов, мхи и лишайники располагаются на коре деревьев. Все эти организмы связаны друг с другом топическими связями.
Форические отношения — участие одного вида в распространении другого. В этой роли обычно выступают животные, переносящие семена, споры, пыльцу растений. Так, обладающие цепляющимися шипами семена лопуха или череды могут захватываться шерстью крупных млекопитающих и переноситься на большие расстояния.
Фабрические отношения — тип связей, при которых особи одного вида используют для своих сооружений продукты выделения, мертвые остатки либо даже живых особей другого вида. Например, птицы строят гнезда из сухих веточек, травы, шерсти млекопитающих и т.п. Личинки ручейников для строительства своих домиков используют кусочки корыпесчинки, обломки раковин или же сами раковины с живыми моллюсками мелких видов.
Примером косвенных межвидовых отношений могут служить следующие. Насекомоядные птицы не питаются растениями, но поедают многих насекомых, которые кормятся листьями или опыляют цветки. Поедая насекомых-опылителей, птицы косвенным образом воздействуют на число производимых деревом плодов, т.е. на количество пищи, доступной животным, которые питаются плодами и проростками, на хищников и паразитов этих животных и т.д.
Из всех типов биотических отношений между видами в биоценозе наибольшее значение имеют топические и трофические связи, поскольку они удерживают друг возле друга организмы разных видов, объединяя их в достаточно стабильные сообщества разного масштаба.
По размеру биоценозы могут быть разными — от мелких (кочка на болоте, муравейник, подушки лишайников на стволах деревьев, небольшой пруд) до очень больших (биоценоз леса, луга, озера, болота, ковыльной степи).
Биоценозы чаще всего не имеют четких границ. В природе они переходят друг в друга постепенно, благодаря чему невозможно определить, где кончается один биоценоз и начинается другой. Например, биоценоз сухого леса постепенно переходит в биоценоз увлажненного луга, который сменяется болотом. Визуально мы можем отграничить лесной биоценоз от лугового и болотного, но сказать четко, где проходит линия границы, мы не в состоянии. В подавляющем большинстве случаев мы будем иметь дело со своеобразной переходной полосой различной ширины и длины, потому что жесткие, резкие границы в природе — редкое исключение. Такая переходная полоса (или зона) между смежными физиономически различимыми сообществами называется экотоном.
Исторически сложившиеся группировки совместно обитающих и взаимосвязанных организмов разных видов называются биоценозами. В состав биоценоза входят фитоценоз, зооценоз, микоценоз и мик-робоценоз. Каждый биоценоз характеризуется видовой и пространственной (вертикальной и горизонтальной) структурой и различными биотическими взаимоотношениями организмов.
studfiles.net
Сукцессии (смена биоценозов и экосистем)
Любая экосистема, приспосабливаясь к изменениям внешней среды, находится в состоянии динамики. Эта динамика может касаться отдельных звеньев экосистем (организмов, популяций, трофических групп), так и всей системы в целом. При этом динамика может быть связана, с одной стороны, с адаптациями к факторам, которые являются внешними по отношению к системе, а с другой — к факторам, которые создаёт и изменяет сама экосистема.
Самый простой тип динамики — суточный. Он связан с изменениями в фотосинтезе и транспирации (испарение воды) растений. В ещё большей мере эти изменения связаны с поведением животного населения. Одни из них более активны днём, другие — в сумерки, третьи — ночью. Аналогичные примеры можно привести по отношению к сезонным явлениям, с которыми ещё больше связана активность жизнедеятельности организмов. Не остаются неизменными экосистемы и в многолетнем ряду. Если в качестве примера взять лес или луг, то не трудно заметить, что в разные годы этим экосистемам свойственны свои особенности. В одни годы мы можем наблюдать увеличение численности одних видов (на лугах, например, бывают «клеверные» годы, годы с резким увеличением злаков и других видов или групп видов). Из этого следует, что каждый вид индивидуален по своим требованиям к среде, и её изменения для одних видов благоприятны, а на другие, наоборот, оказывают угнетающее влияние. Сказывается также и периодичность в интенсивности размножения.
Эти изменения в одних случаях могут в какой-то мере повторяться, в других же имеют место изменения, которые на фоне периодически повторяющейся динамики имеют однонаправленность, поступательный характер и обусловливают развитие экосистемы в определённом направлении. Периодически повторяющуюся динамику называют циклическими изменениями, или флюктуациями, а направленную динамику именуют поступательной или развитием экосистем. Для последнего вида динамики характерным является либо внедрение в экосистемы новых видов, либо смена одних видов другими. В конечном счёте происходят смены биоценозов и экосистем в целом. Этот процесс называют сукцессией (от лат.»сукцессио» — преемственность, наследование). Если сукцессия обуславливается в основном внешними по отношению к системе факторами, то такие смены называют экзогенетическими, или экзодинамическими (от греч. «эндон» — внутри).
Экзогенетические смены (сукцессии) могут быть вызваны изменением климата в одном направлении, например, в сторону потепления или похолодания, иссушением почв, например, в результате осушения или понижения уровней грунтовых вод по другим причинам. Такие смены могут длиться столетиями и тысячелетиями и их называют вековыми сукцессиями.
Ход эндодинамических сукцессий рассмотрим на примере наземных экосистем. Если взять участок земной поверхности, например, заброшенные пахотные земли в различных географических районах (в лесной, степной зонах либо среди тропических лесов и тому подобное), то для всех этих объектов будут характерны как общие, так и специфические изменения в экосистемах.
В качестве общих закономерностей будет иметь место заселение живыми организмами, увеличение их видового разнообразия, постепенное обогащение почвы органическим веществом, возрастание их плодородия, усиление связей между различными видами или трофическими группами организмов, уменьшение числа свободных экологических ниш, постепенное формирование всё более сложных биоценозов и экосистем, повышение их продуктивности. Более мелкие виды организмов, особенно растительных, при этом, как правило, сменяются более крупными интенсифицируются процессы круговорота веществ и тому подобное. В каждом случае при этом можно выделить последовательные стадии сукцессий, под которыми понимается смена одних экосистем другими, а сукцессионные ряды заканчиваются относительно мало изменяющимися экосистемами. Их называют климаксными (от греч. климакс — лестница), коренными, или узловыми.
Специфические закономерности сукцессий заключаются прежде всего в том, что в каждой из них, как и каждой стадии, присущ тот набор видов, которые, во-первых, характерны для данного региона, а, во-вторых, наиболее приспособленного ряда. Различными будут и завершающие (климаксные) сообщества (экосистемы).
Американский эколог Клементс, наиболее полно разработавший учение о сукцессиях, считает, что в любом обширном географическом районе, который по масштабам можно примерно приравнять к природной зоне (лесная, степная, пустынная и тому подобное), каждый ряд завершается одной и той же климаксной экосистемой (моноклимаксом). Такой климакс был назван климатическим. Это, однако, не значит, что для любого участка географической зоны (моноклимакса) характерен один и тот же набор видов. Видовой состав климаксных экосистем может существенно различаться. Общим является лишь то, что эти экосистемы объединяет сходство видов-эдификаторов, то есть тех, которые в наибольшей мере создают среду обитания. Например, для степных экосистем эдификаторами являются плотнокустовые злаки (ковыль и типчак). Для тропических лесов в качестве эдификаторов выступает большое количество древесных видов, создающих сильное затенение для других видов своим пологом.
Для лесной зоны северных и срединных регионов Евразии основными эдификаторами выступает ель и пихта. Они из набора всех древесных видов в наибольшей степени изменяют условия местопроизрастания: сильно затеняют подпологовое пространство, создают кислую среду почв и обуславливают процессы их оподзоливания (растворение и вымывание из приповерхностного слоя практически всех минералов, кроме кварца), которые не отстают от них в росте и способны первыми захватить пространство. При сочетании таких условий возможно формирование климаксных смешанных елово-лиственных (пихтово-лиственных), чаще всего с берёзой и осиной, лесов. Последнее наиболее характерно для зоны смешанных лесов. Для таежной (более северной) зоны более типичны климаксные леса с явным преобладанием только эдификаторов (ель, пихта).
Однако прежде чем сформируется климаксное сообщество (экосистема), ему предшествует, как отмечалось выше, ряд промежуточных стадий или серий. Так, на исходно безжизненном субстрате здесь сначала появляются организмы-пионеры, например, корковые водоросли, накипные лишайники. Они несколько обогащают субстрат органическими и доступными для усвоения растениями веществами. За ними следуют отдельные травянистые растения, обычно способные быстро осваивать бедный субстрат. Эта стадия сменяется полукустарниками и кустарниками, а ей на смену приходят лиственные виды деревьев (чаще всего берёза, осина, ива). Последние характеризуются быстрым ростом, но, отличаясь высоким светолюбием, быстро изреживаются (к 40-50-летнему возрасту). В результате этого под их пологом создаются условия для поселения теневыносливой ели, которая постепенно догоняет в росте стареющие лиственные виды деревьев и выходит в первый ярус. На данной стадии и образуется климаксное смешанное елово-лиственное сообщество или чисто еловый лес со свойственным им набором других видов растений и животных.
Названия экосистем, биоценозов. Значительное разнообразие (богатство почв, увлажнение) в пределах формирования сходных климаксных сообществ обуславливает существенное различие продуктивности отдельных экосистем и сопутсвующих эдификаторам видов растений и животных. Обычно степень благоприятности условий местопроизрастания оценивается либо по результатам прямого определения значений факторов, либо по растениям-индикаторам.
Так, для лесной зоны кислица указывает на условия увлажнения, близкие к оптимальным, и значительное богаство почв питальными минеральными веществами; черника — на несколько избыточное увлажнение и некоторый дефицит элементов минерального питания; брусника — на дефицит увлажнения и почвенного плодородия; мхи (кукушкин лён и ,особенно, сфагнум) — на чрезмерно избыточное увлажнение, дефицит минеральных веществ, недостаток кислорода для дыхания корней и наличие процессов торфообразования. Наряду с индикаторами меняется состав и других видов, произрастающих под пологом эдификаторов.
По растениям-эдификаторам и растения-индикаторам называют биоценозы (экосистемы). Лесоводы их определяют как типы леса (например, ельники-кисличники, ельники-черничники, ельнико-сфагновые и другие). По такому же принципу классифицируются и называются другие растительные сообщества (не лесные) и экосистемы в целом. Но в этом случае они носят название не типов, а ассоциаций растений, по которым называются экосистемы. Это элементарные единицы относительно однородного по видовому составу и другим признакам растительного покрова. Например, для степей выделяются типчаково-ковыльные, злакотравные и тому подобные экосистемы (биогеоценозы).
Наряду с теорией моноклимакса существует точка зрения, в соответствии с которой в одном и том же географическом районе может формироваться несколько завершающих (климаксных) экосистем. Например, в лесной зоне, наряду с еловыми и елово-лиственными лесами в качестве климаксных рассматриваются также луговые экосистемы, сосновые леса. Однако сторонники моноклимакса считают, что луга в лесной зоне могут длительно существовать только в результате их использования (скашивания, выпаса). При прекращении таких воздействий на смену им неизбежно придут лесные сообщества. Что касается сосновых лесов, то длительное существование их связывается с тем, что они занимают обычно крайне бедные (например, песчаные, щебнистые, сильно заболоченные) места обитания, где ель (более сильный эдификатор) не может внедряться и существовать вследствие более значительной требовательности к почвенному плодородию. Однако с течением времени и по мере накопления в почве органических веществ и необходимых для жизни минеральных элементов и эти сосновые места обитания, с точки зрения сторонников моноклимакса, будут заняты еловыми лесами, как обладающими более сильной эдификаторной способностью.
Причина сукцессий (частных).
Сукцессионные смены обычно связывают с тем, что существующая экосистема (сообщество) создает неблагоприятные условия для наполняющих ее организмов (почвоутомление, неполный круговорот веществ, самоотравление продуктами выделения или разложения и т.п.). Такие явления реальны, но не объясняют всех случаев смен экосистем. Например, в северных лесах внедрение под полог лиственных древесных сообществ ели связано прежде всего с тем, что последняя использует биологические свойства первых по слабому притенению почвы. Сами же почвенные условия остаются не только благоприятными для лиственных древостоев, но и постепенно улучшаются для них ( идет накопление питательных веществ, уменьшается кислотность и т.п.). Следовательно, здесь нет оснований говорить о самоотравлении или других подобных причинах смен.
Не подтверждается безоговорочно и точка зрения о том, что появление ели под пологом лиственных лесов и древостоев связано с тем, что в молодом возрасте этот вид требует затенения. Известно, например, что ель и в молодом возрасте прекрасно растет при полном освещении (значительно лучше, чем под пологом других древесных видов). Об этом, в частности, свидетельствуют многочисленные примеры создания культурных фитоценозов ели (посадкой молодых растений или посевом семян) на открытых площадях.
Наряду с природными факторами причинами динамики экосистем все чаще выступает человек. К настоящему времени им разрушено большинство коренных (климаксных) экосистем. Например, степи почти полностью распаханы (сохранились только на заповедных участках). Преобладающие площади лесов представлены переходными (временными) экосистемами из лиственных древесных пород (береза, осина, реже ива, ольха и другие). Эти леса обычно называют производными, или вторичными. Они, как отмечалось выше, являются промежуточными стадиями сукцессий. К сменам экосистем ведут также такие виды деятельности человека, как осушение болот, чрезмерные нагрузки на леса. Например, в результате отдыха населения (рекреации), химических загрязнений среды, усиленного выпаса скота, пожаров и т.п. Антропогенные воздействия часто ведут к упрощению экосистем. Такие явления обычно называют дегрессиями. Различают, например, пастбищные, рекреационные и другие дегрессии. Смены такого типа обычно завершаются не климаксными экосистемами, для которых характерно усложение структуры, а стадиями катоценоза, которые нередко заканчиваются полным распадом экосистем. Климаксные экосистемы обычно чувствительны к различным вмешательствам в их жизнь. К подобным воздействиям, кроме хвойных лесов, чувствительны и другие коренные сообщества, например, дубовые леса. Это одна из причин катастрофической гибели дубрав в современный период и замена их, как и хвойных лесов, менее ценными, но более устойчивыми временными экосистемами из березы, осины, кустарников или трав. Последнее особенно типично при разрушении степных и лесостепных дубрав.
Виды сукцессий.
Сукцессии, с которыми мы познакомились на примере лесной зоны, называют первичными по той причине, что они начинаются с исходно безжизненного пространства (субстрата). Кроме отвалов горных пород, такие сукцессии могут начинаться на песчаных обнажениях, продуктах извержения вулканов (застывшая лава, отложения пепла) и т.п.
Наряду с первичными выделяют вторичные сукцессии. Последние отличаются от первичных тем, что они начинаются обычно не с нулевых значений, а возникают на месте нарушенных или разрушенных экосистем. Например, после вырубок лесов, лесных пожаров, при израстании площадей, находившихся под сельскохозяйственными угодьями. Основное отличие этих сукцессий заключается в том, что они протекают несравненно быстрее первичных, так как начинаются с промежуточной стадии (трав, кустарников или древесных растений-пионеров) и на фоне более богатых почв. Конечно, вторичная сукцессия возможна только в тех случаях, если человек не будет оказывать сильное и постоянное влияние на развивающиеся экосистемы. В последнем случае, как отмечалось выше, процесс пойдет по схеме дегрессий и завершится стадией катоценоза и опустынивания территорий.
Различают также автотрофные и гетеротрофные сукцессии. Рассмотренные выше примеры сукцессий относятся к автотрофным, поскольку все они протекают в экосистемах, где центральным звеном является растительный покров. С его развитием связаны смены гетеротрофных компонентов. Такие сукцессии потенциально бессмертны, поскольку все время пополняются энергией и веществом, образующимися или фиксирующимися в организмах в процессе фотосинтеза либо хемосинтеза. Завершаются они, как отмечалось, климаксной стадией развития экосистем.
К гетеротрофным относятся те сукцессии, которые протекают в субстратах, где отстутствуют живые растения (продуценты), а участвуют только животные (гетеротрофы) или мертвые растения. Этот вид сукцессий имеет место только до тех пор, пока присутствует запас готового органического вещества, в котором сменяются различные виды организмов-разрушителей. По мере разрушения органического вещества и высвобождения из него энергии сукцессионный ряд заканчивается, система распадается. Таким образом, эта сукцессия по природе своей деструктивна. Примерами гетеротрофных являются сукцессии, имеющие место, например, при разложении мертвого дерева или трупа животного. Так, при разложении мертвого дерева можно выделить несколько стадий смен гетеротрофов. Первыми на мертвом, чаще ослабленном дереве, поселяются насекомые-короеды. Далее их сменяют насекомые, питающиеся древесиной (ксилофаги). К ним относятся личинки усачей, златок и других. Одновременно идут смены грибного населения. Они имеют примерно следующую последовательность: грибы-пионеры (обычно окрашивают древесину в разные цвета), грибы-деструкторы, способствующие появлению мягкой гнили, и грибы-гумификаторы, превращающие часть гнилой древесины в гумус. На всех стадиях сукцессий присутствуют также бактерии. В конечном счете органическое вещество в основной массе разлагается до конечных продуктов: минеральных веществ и углекислого газа. Гетеротрофные сукцессии широко осуществляются при разложении детрита (в лесах он представлен лесной подстилкой). Они протекают также в экскрементах животных, в загрязненных водах, в частности, интенсивно идут при биологической очистке вод с использованием активного ила, насыщенного большим количеством организмов.
Общие закономерности сукцессионного процесса.
Для любой сукцессии, особенно первичной, характерны следующие общие закономерности протекания процесса:
1. На начальных стадиях видовое разнообразие незначительно, продуктивность и биомасса малы. но по мере развития сукцессии эти показатели возрастают.
2. С развитием сукцессионного ряда увеличиваются взаимосвязи между организмами. Особенно возрастает количество и роль симбиотических отношений. Полнее осваивается среда обитания, усложняются цепи и сети питания.
3. Уменьшается количество свободных экологических ниш, и в климаксном сообществе они либо отсутствуют, либо находятся в минимуме. В связи с этим по мере развития сукцессий уменьшается вероятность вспышек численности отдельных видов.
4. Интенсифицируются процессы круговорота веществ, поток энергии и дыхание экосистем.
5. Скорость суцессионного процесса в большей мере зависит от продолжительности жизни организмов, играющих основную роль в сложении и функционировании экосистем. В этом отношении наиболее продолжительные сукцессии в лесных экосистемах. Короче они в экосистемах, где автотрофное звено представлено травянистыми растениями, и еще быстрее протекают в водных экосистемах.
6. Неизменяемость завершающих (климаксных) стадий сукцессий относительна. Динамические процессы при этом не приостанавливаются, а лишь замедляются. Продолжаются динамические процессы, обуславливаемые изменениями среды обитания, сменой поколений организмов и другими явлениями. Относительно большой удельный вес занимают динамические процессы циклического (флуктуационного) плана.
7. В зрелой стадии климаксного сообщества биомасса обычно достигает максимальных или близких к максимальным значений. Неоднозначна продуктивность отдельных сообществ на стадии климакса.
Обычно считается, что по мере развития сукцессионного процесса продуктивность увеличивается и достигает максимума на промежуточных стадиях, а затем в климаксном сообществе резко уменьшается. Последнее связывают, во-первых, с тем, что в это время максимум первичной продукции потребляется консументами, а, во-вторых, экосистема развивает чрезвычайно большую массу ассимиляционного аппарата, что ведет к дефициту освещенности, следствием чего является снижение интенсивности фотосинтеза при одновременном возрастании потерь продуктов ассимиляции на дыхание самих автотрофов.
Эти положения нельзя распространять на все климаксные сообщества. Например, нет реальных предпосылок для увеличения численности гетеротрофов в хвойных лесах по сравнению с лиственными. Скорее, в последних больше потребителей зеленой продукции и, вероятнее, вспышки численности отдельных видов-фитофагов, например, насекомых.
Нет также ни теоретических предпосылок, ни фактических данных, которые бы свидетельствовали, что в зрелой климаксной системе, например в еловых лесах, масса хвои достигает чрезмерно высоких значений. Это противоречит принципам адаптации к увеличению биогенной геохимической энергии организмами как условию их выживания (второй биогеохимический принцип В.И. Вернадского). Весь опыт лесоводства также свидетельствует о наиболее высокой продуктивности климаксных лесных сообществ (применительно к лесной зоне хвойных или смешанных хвойно-лиственных лесов). В противном случае, с точки зрения получения продукции (древесины), неизбежен вывод о нецелесообразности ориентации на выращивание и сохранение климаксных стадий лесов.
Применительно к другим экосистемам, например луговым, можно согласиться с тем, что возможности получения продукции на климаксной стадии уменьшаются, однако не потому, что сокращается ее нарастание (прирост, продуктивность), а по той причине, что более значительная часть ее отчуждается гетеротрофами в результате образования устойчивых цепей выедания.
Другими словами, продуктивность экосистем на климаксных стадиях сукцессий высока. Как правило, максимальна вследствие более полного освоения пространства. Однако возможности снятия человеком первичной продукции лимитируются (иногда до нулевых значений) вследствие включения ее в цепи питания.
www.examen.ru
Биоценозы и их изменения
Исторически в природе складываются сложные многосторонние пищевые взаимоотношения, которые могут быть выражены в различных цепях питания. Каждая из них включает 5-6 звеньев. Начальное звено такой цепи питания — автотрофный организм, создающий органическое вещество своего тела из веществ неорганических с использованием солнечной (зеленые растения) или химической энергии (хемосинтезу некоторых бактерий).
Зеленое растение служит пищей гетеротрофным организмам — травоядным животным. При этом установлено, что для построения 1 кг массы своего тела травоядное животное должно съесть не меньше 10 кг растительной пищи. Травоядные животные служат пищей для хищника. И в этом случае лишь 0,1 массы всей пищи, съеденной хищником, превратится в вещество его тела.
Таким образом, даже в этой предельно короткой цепи питания обнаруживается очень важная экологическая закономерность: при переходе от одного звена цепи питания к последующему только около 10% массы всей пищи идет на построение тела организма, а 90% массы всей пищи расходуется на поддержание жизнедеятельности организма. Именно поэтому цепи питания не бывают особенно длинными.
В каждой местности цепи питания и другие взаимоотношения между организмами создаются по-своему, своим комплексом видов, которые вместе с факторами абиотической среды составляют относительно устойчивую единую систему. Такие системы в экологии получили название биоценозов. Лес, луг, степь, тундра, тайга, пресноводный водоем — все это будут различные биоценозы. На рисунке в качестве примера приведены цепи питания и их сложные переплетения, которые характеризуют пресноводный биоценоз.
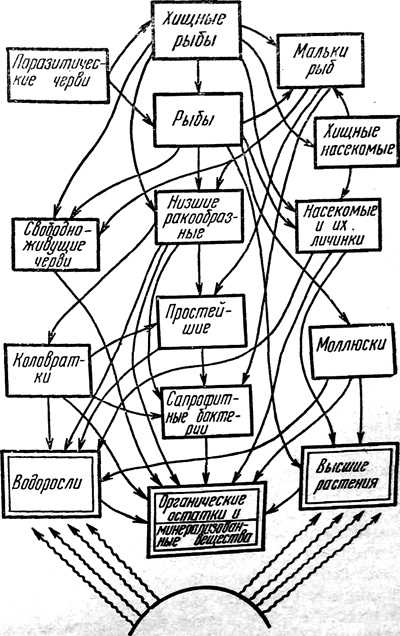
Очень важной характеристикой биоценоза является его способность к саморегуляции. Численность любого вида, входящего в состав биоценоза, не остается постоянной, а из года в год колеблется. Однако такие колебания в пределах данной системы биоценоза не нарушают его целостности. Поясним это следующим примером. На той или иной территории численность хищников (волк, рысь) в первую очередь определяется численностью видов, поедаемых хищниками (грызуны, птицы). Усиленное размножение хищников приводит к тому, что их жертвы уничтожаются в очень большом количестве. Численность грызунов, некоторых птиц, например куропаток, резко падает. Хищникам перестает хватать корма — снижается темп их размножения, а это приводит к снижению их численности, что сразу же отражается на жертвах хищников: они начинают усиленно размножаться. На богатой кормовой базе (много грызунов) хищники вновь быстро повышают свою численность. И хотя этот пример пределъно упрощен, из него можно понять, в чем состоит процесс саморегуляции внутри биоценоза. На рисунке ниже показаны данные о сопряженном колебании численности рыси и ее жертвы — зайца. Из рисунка видно, как подъемы и падения численности хищника закономерно следуют за подъемами и падениями численности жертвы. Но при этом ни один из двух видов не исчезает полностью.

Таким образом, устойчивость биоценоза обеспечивается подвижным равновесием между составляющими его видами.
Кроме саморегуляции, в природе имеет место и смена биоценозов. Наглядным примером смены биоценозов может служить процесс зарастания гарей. Старый биоценоз (хвойный лес) разрушен стихийным бедствием (пожаром). Первыми поселенцами гари оказываются травянистые растения, быстро занимающие освободившуюся территорию благодаря интенсивному половому и вегетативному размножению. Вместе с травянистой растительностью появляются и быстрорастущие сеянцы светолюбивых лиственных деревьев — березы, осины. В это время условий для возобновления старого биоценоза (хвойного леса) нет: медленнорастущие, плохо приспособленные к прямому солнечному свету сеянцы хвойных не могут конкурировать с травянистыми и древесными покрытосеменными растениями. Постепенно на месте хвойного леса формируется лиственный лес. Со сменой растительности меняется и фауна: исчезают виды, для которых пищей были семена хвойных (например, белки, клесты), появляются виды насекомых, чьи гусеницы питаются листьями деревьев, а вместе с ними в лесу поселяются и насекомоядные птицы. Травянистая и кустарниковая растительность привлекает копытных, а с копытными приходят и их извечные враги — хищники. Так, старый биоценоз заменяется новым биоценозом.
Однако такой лиственный лес, окруженный хвойным лесом, — явление временное. Под пологом березового леса сеянцы хвойных деревьев находят благоприятные условия. Пройдет столетие и менее устойчивый биоценоз (березовый лес) неизбежно будет заменен более устойчивым биоценозом — хвойным лесом со всеми присущими ему обитателями.
Смена биоценоза не всегда носит обратимый характер, который только что был описан. Так, в результате процесса заболачивания может иметь место следующая необратимая смена биоценозов: пресноводный водоем → болото → лес.
При сопоставлении биоценозов на разных широтах обнаруживается географическая зональность биоценозов. Так, в направлении с севера на юг прослеживается следующая закономерная замена одного биоценоза другим: тундра → хвойный лес → лиственный лес → степь → пустыня. В горных областях имеет место вертикальная зональность биоценозов, которая в некоторой степени как бы повторяет географическую зональность. В этом случае замена биоценозов идет в таком порядке (снизу вверх): степь → лиственный лес → хвойный лес → альпийские луга.
jbio.ru
Стадии формирования биоценоза
Количество просмотров публикации Стадии формирования биоценоза — 724
Динамика биоценоза
В целом сообществу присуща суточная, сезонная (годичная) и многолетняя динамика, свойственная как растениям, так и животным. Суточная, вызываемая сменой светлой и темной части суток, у растений проявляется в интенсивности фотосинтеза, дыхания, раскрывании и закрывании цветков, у животных — в разной суточной активности (дневные, сумеречные и ночные).
Нередко животные в течение суток меняют сообщество. Так, цапля кормится на мелководьях водоемов, а гнездится и ночует в кронах деревьев, насекомые– опылители (к примеру, пчелы) могут перелетать из лесного сообщества в луговое.
Сезонная динамика биоценоза зависит от фенологического состояния фитоценоза, видового состава и численности обитающих в нем животных. Каждый вид растительных организмов в течение вегетационного периода проходит определенные стадии развития (начало вегетации, цветение, плодоношение и отмирание). В фитоценозе, состоящем из множества видов, фазы развития растений могут совпадать и не совпадать.
Внешний вид фитоценоза, изменяющийся на протяжении года с чередованием фаз развития, принято называть аспектом. Как правило, аспект повторяется из года в год с неизменной последовательностью, отражая цветовую гамму растительного сообщества (весенняя яркая зелень, летнее разноцветье и осенняя пестрота лесов). Аспект именуют обычно по растениям, придающим фитоценозу наиболее заметную цветовую окраску, к примеру голубой аспект незабудки болотной, белый аспект пушицы, бурый аспект листьев осоки и т. д.
Сезонная динамика животных представителей биоценоза связана с их размножением, жизненной активностью и миграциями. Весенний прилет и осенний отлет птиц, нерест рыб, появление молодняка, активность насекомых– опылителей на лугах, зимняя спячка медведя только ничтожно малая часть примеров сезонной динамики животного населения биоценоза.
Многолетняя динамика сообщества вызывается его повторяющимися изменениями на протяжении нескольких лет при отсутствии резкой смены видового состава. Изменения затрагивают в основном численность особей видов, образующих биоценоз. В качестве примера можно привести изменения в лесах некоторых заповедников Беларуси и России, обусловленные увеличением численности лося – основного потребителя древесно – кустарниковых кормов [2].
Возникновение биоценоза начинается с появления первых организмов на лишенных жизни участках (лавовых потоках, вулканических островах, осыпях, обнажившихся горных породах, песчаных наносах и обсохших днищах водоемов). Заселение начинается со случайного заноса организмов с уже освоенных ими территорий и зависит от свойств субстрата. Этот участок для многих семян растений и проникших сюда животных может оказаться неподходящим для размножения. Нередко, особенно в гумидной зоне, первыми поселенцами оказываются представители водорослей, мхов и лишайников.
Как правило, успешно развиваются лишь немногие из занесенных растительных видов. Животные– консументы поселяются несколько позднее, так как их существование без пищи невозможно, но случайное посещение ими осваиваемых участков — довольно частое явление. Данная стадия развития биоценоза получила название пионерной. Хотя на этой стадии сообщество еще не сложилось (непостоянный видовой состав, разреженный растительный покров), оно уже оказывает влияние на абиотическую среду: начинает образовываться почва.
Пионерная стадия сменяется ненасыщенной, когда растения начинают возобновляться (семенами или вегетативным путем), а животные размножаться.
Постепенно темпы заселения участка возрастают за счёт как увеличения числа особей пионерной растительности до образования зарослей, так и внедрения новых видов. Видовой состав такого сообщества пока неустойчив, новые виды внедряются довольно легко, хотя конкуренция начинает играть заметную роль. Эта стадия развития биоценоза группировка.
При последующем развитии сообщества происходит дифференциация растительного покрова по ярусам и синузиям, устойчивое постоянство приобретают его мозаичность, видовой состав, пищевые цепи и консорции. В конечном итоге занятыми оказываются все экологические ниши и дальнейшее вселение организмов становится возможным только после – вытеснения или уничтожения старожилов. Данная, заключительная, стадия образования биоценоза получила название насыщенной. При этом дальнейшее развитие биоценоза не прекращается и случайные отклонения в видовом составе и взаимоотношениях как между организмами, так и с окружающей средой еще могут иметь место.
Случайные отклонения в структуре биоценоза называются флуктуациями. Как правило, они обусловлены случайными или сезонными изменениями численности видов, входящих в биоценоз, в результате неблагоприятных метеорологических явлений, наводнений, землетрясений и т. д. Обильные снегопады и иней, к примеру, приводят к изреживанию крон, и весной под пологом леса большее развитие получают злаки.
Хотя биоценоз представляет собой довольно консервативную природную систему, однако под давлением внешних обстоятельств он может уступить место другому биоценозу. Последовательная смена во времени одних сообществ другими на определенном участке среды принято называть сукцессией (от. Лат successia преемственность, наследование). В результате сукцессии одно сообщество последовательно сменяется другим без возврата к исходному состоянию. К сукцессии приводит взаимодействие организмов, главным образом ранений, друг с другом и со средой.
Сукцессии подразделяются на первичные и вторичные. Первичные происходят на первичносвободных от почвы Грунтах — вулканических туфовых и лавовых полях, сыпучих песках, каменистых россыпях и т. д. По мере развития фитоценоза от пионерной стадии до насыщенной почва становится плодороднее и в биологический круговорот вовлекается все больше химических элементов в возрастающем количестве. С увеличением плодородия виды растений, развивающиеся на богатых питательными веществами почвах, вытесняют менее требовательные в данном отношении виды. Одновременно сменяется и животное население. Вторичные сукцессии реализуются на местообитаниях разрушенных сообществ, где сохранилась почвы и некоторые живые организмы. Разрушение биоценозов может вызываться стихийными природными процессами (ураганы, ливни, наводнения, оползни, продолжительные засухи, извержение вулканов и т. д.). а также изменение организмами среды обитания (к примеру, при зарастании водоема водная среда сменяется отложениями торфа). Вторичные сукцессии характерны для деградированных пастбищ, гарей, вырубок леса, исключенные из сельскохозяйственного пользования пашен и других угодий. а также для искусственных лесопосадок. К примеру, гарях с супесчаными и суглинистыми почвами пионерная растительность из иван– чая и березы бородавчатой со временем сменяется еловыми насаждениями.
Смена одного биоценоза другим в ходе сукцессии образует сукцессионный ряд, или серию. Изучение сукцессионных рядов имеет большое значение в связи с усиливающимся антропогенным влиянием на биоценозы. Конечным результатом такого рода исследований должна быть прогнозирование образования природно– антропогенных ландшафтов. Изучение вторичных сукцессии и факторов, их вызывающих, играет важную роль в решении проблем охраны и рационального использования биологических и земельных ресурсов.
В случае если естественное течение сукцессии не нарушается, сообщество постепенно приходит к относительно устойчивому состоянию, в котором поддерживается равновесие между организмами, а также между ними и средой, – к климаксу. Без вмешательства человека данный биоценоз может существовать неопределенно долго, к примеру, сосняк черничный, лишайниковая тундра на песчаных почвах.
Концепция климакса детально разработана американским ботаником X. Каулсом и широко используется в зарубежной ботанико– географической литературе. Согласно данной концепции, климакс — терминальная стадия эволюции сообщества, которой соответствует почва определенного типа — педоклимакс. Сукцессии, ведущие к этой стадии, называются прогрессивными, а удаляющие биоценоз от нее,— регрессивными. Нельзя, однако, придавать понятию ʼʼклимаксʼʼ абсолютное значение и полагать, что при достижении его сообщество прекращает развитие.
Биоценозы, которые, будучи нарушенными, возвращаются к своему исходному состоянию, называются коренными. На месте вырубки сосняка черничного или ельника кисличного вырастет березняк, а он, в свою очередь, опять сменится сосняком черничным или ельником кисличным. В данном случае речь идет о коренных типах леса.
Трансформированные биоценозы не возвращаются в исходное состояние. Так, осушенное и освоенное под посевы сельскохозяйственных культур низинное осоковое болото после сработки торфяной залежи и разрушения мелиоративной сети с прекращением по каким– либо причинам сельскохозяйственного использования развивается в направлении формирования березового или ольхового мелколесья. Зооценоз этого мелколесья отличается от сообщества видов животных открытого травяного болота [4].
referatwork.ru
