
Хромосомная теория наследственности — Википедия
Материал из Википедии — свободной энциклопедии
Обмен участками между отцовской и материнской гомологичными хромосомами в мейозе согласно теории хиазмотипии Ф. Янсенса (1909)Хромосомная теория наследственности — теория, согласно которой передача наследственной информации в ряду поколений связана с передачей хромосом, в которых в определённой и линейной последовательности расположены гены. Эта теория сформулирована в начале XX века, основной вклад в её создание внесли американский цитолог У. Саттон (англ.)русск., немецкий эмбриолог Т. Бовери и американский генетик Т. Морган со своими сотрудниками К. Бриджесом, А. Стёртевантом и Г. Мёллером[1].
В 1902-1903 годах У. Сеттон и Т. Бовери независимо друг от друга выявили параллелизм в поведении менделевских факторов наследственности (генов) и хромосом. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Экспериментальное доказательство локализации генов в хромосомах было получено позднее Т. Морганом и его сотрудниками, работавшими с плодовой мушкой
ru.wikipedia.org
Хромосомная теория наследственности | Наука
Хромосомная теория наследственности— теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, то есть преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Хромосомная теория наследственности возникла в начале 20 в. на основе клеточной теории и использовалась для изучения наследственных свойств организмов гибридологического анализа.
1.Формирование хромосомной теории Править
В 1902-1903 гг. американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бовери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911—1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов.
Пол, как и любой другой признак организма, наследственно детерминирован. Важнейшая роль в генетической детерминации пола и в поддержании закономерного соотношения полов принадлежит хромосомному аппарату.
У раздельнополых организмов (животных и двудомных растении) соотношение полов обычно составляет 1:1, то есть мужские и женские особи встречаются одинаково часто. Это соотношение совпадает с расщеплением в анализирующем скрещивании, когда одна из скрещиваемых форм является гетерозиготной (Аа), а другая — гомозиготной по рецессивным аллелям (аа). В потомстве в этом случае наблюдается расщепление в отношении 1Аа:1аа. Если пол наследуется по такому же принципу, то вполне логично было бы предположить, что один пол должен быть гомозиготным, а другой — гетерозиготным. Тогда расщепление по полу должно быть в каждом поколении равным 1:1, что и наблюдается в действительности.
При изучении хромосомных наборов самцов и самок ряда животных между ними были обнаружены некоторые различия. Как у мужских, так и у женских особей во всех клетках имеются пары одинаковых (гомологичных) хромосом, но по одной паре хромосом они различаются. Так, у самки дрозофилы имеются две палочковидные хромосомы, а у самца — одна такая же палочковидная, а вторая, парная первой, — изогнутая. Такие хромосомы, по которым самцы и самки отличаются друг от друга, называют половыми хромосомами. Те из них, которые являются парными у одного из полов, называют X-хромосомами (например, у дрозофилы и млекопитающих) или Z-хромосомами (например, у птиц). Непарная половая хромосома, имеющаяся у особей только одного пола, была названа У-хромосомой (у дрозофилы и млекопитающих) или W-хромосомой (у птиц). Хромосомы, в отношении которых между самцами и самками нет различий, называют аутосомами. Следовательно, у дрозофилы особи обоих полов имеют по шесть одинаковых аутосом плюс две половые хромосомы (ХХ у самок и XY у самцов).
Пол, имеющий различные половые хромосомы (X и У), образует гаметы двух типов (половина с X-хромосомой и половина с У-хромосомой), то есть, является гетерогаметным, а пол, содержащий в каждой клетке одинаковые половые хромосомы (X-хромосомы), — гомогаметным.
Открытие половых хромосом и установление их роли в определении пола послужило важным доводом в пользу того, что хромосомы определяют признаки организма.
Определение пола Править
От чего же зависит рождение мужских и женских особей? Рассмотрим это на примере определения пола у дрозофилы. В ходе гаметогенеза у самок образуется один тип гамет, содержащий гаплоидный набор аутосом и одну X-хромосому. Самцы образуют два типа гамет, половина из которых содержит три аутосомы и одну X-хромосому (ЗА+Х), а половина — три аутосомы и одну У-хромосому (ЗА+У). При оплодотворении яйцеклеток (ЗА+Х) сперматозоидами с X-хромосомами будут формироваться самки (6А+ХХ), а от слияния яйцеклеток со сперматозоидами, несущими У-хромосому, — самцы (6A+XY). Поскольку число мужских гамет с X- и У-хромосомами одинаково, то и количество самцов и самок тоже одинаково. В данном случае пол организма определяется в момент оплодотворения и зависит от хромосомного набора зиготы.
Сходный способ определения пола (XY-тип) присущ всем млекопитающим, в том числе и человеку, клетки которого содержат 44 аутосомы и две X-хромосомы у женщин либо XY-хромосомы у мужчин.
Таким образом, XY-тип определения пола, или тип дрозофилы и человека, — самый распространенный способ определения пола, характерный для большинства позвоночных и некоторых беспозвоночных. Х0-тип встречается у большинства прямокрылых, клопов, жуков, пауков, у которых Y-хромосомы нет вовсе, так что самец имеет генотип Х0, а самка — XX.
У всех птиц, большинства бабочек и некоторых пресмыкающихся самцы являются гомогаметным полом, а самки —- гетерогаметным (типа XY или типа ХО). Половые хромосомы у этих видов обозначают буквами Z и W, чтобы выделить таким образом данный способ определения пола; при этом набор хромосом самцов обозначают символом ZZ, а самки — символом ZW или Z0.
Доказательства того, что половые хромосомы определяют пол организма, были получены при изучении нерасхождения половых хромосом у дрозофилы. Если в одну из гамет попадут обе половые хромосом, а в другую — ни одной, то при слиянии таких гамет с нормальными могут получиться особи с набором половых хромосом ХХХ, ХО, ХХУ и др. Выяснилось, что у дрозофилы особи с набором ХО — самцы, а с набором ХХУ — самки (у человека — наоборот). Особи с набором ХХХ имеют гипертрофированные признаки женского пола (сверхсамки). (Особи со всеми этими хромосомными аберрациями у дрозофилы стерильны). В дальнейшем было доказано, что у дрозофилы пол определяется соотношением (балансом) между числом X-хромосом и числом наборов аутосом.
Наследование признаков, сцепленных с полом Править
В том случае, когда гены, контролирующие формирование того или иного признака, локализованы в аутосомах, наследование осуществляется независимо от того, кто из родителей (мать или отец) является носителем изучаемого признака. Если же гены находятся в половых хромосомах, характер наследования признаков резко изменяется. Например, у дрозофилы гены, локализованные в X-хромосоме, как правило, не имеют аллелей в У-хромосоме. По этой причине рецессивные гены в X-хромосоме гетерогаметного пола практически всегда проявляются, будучи в единственном числе.
Признаки, гены которых локализованы в половых хромосомах, называются признаками, сцепленными с полом. Явление наследования, сцепленного с полом, было открыто Т. Морганом у дрозофилы.
Х- и У-хромосомы у человека имеют гомологичный (псевдоаутосомный) участок, где локализованы гены, наследование которых не отличается от наследования аутосомных генов.
Помимо гомологичных участков, X- и У-хромосомы имеют негомологичные участки. Негомологичный участок У-хромосомы, кроме генов, определяющих мужской пол, содержит гены перепонок между пальцами ног и волосатых ушей у человека. Патологические признаки, сцепленные с негомологичным участком У-хромосомы, передаются всем сыновьям, поскольку они получают от отца У-хромосому.
Негомологичный участок X-хромосомы содержит в своем составе ряд важных для жизнедеятельности организмов генов. Поскольку у гетерогаметного пола (ХУ) X-хромосома представлена в единственном числе, то признаки, определяемые генами негомологичного участка X-хромосомы, будут проявляться даже в том случае, если они рецессивны. Такое состояние генов называется гемизиготным. Примером такого рода X-сцепленных рецессивных признаков у человека являются гемофилия, мышечная дистрофия Дюшена, атрофия зрительного нерва, дальтонизм (цветовая слепота) и др.
Гемофилия — это наследственная болезнь, при которой кровь теряет способность свертываться. Ранение, даже царапина или ушиб, могут вызвать обильные наружные или внутренние кровотечения, которые нередко заканчиваются смертью. Это заболевание встречается, за редким исключением, только у мужчин. Было установлено, что обе наиболее распространенные формы гемофилии (гемофилия А и гемофилия В) обусловлена рецессивными генами, локализованными в X-хромосоме. Гетерозиготные по данным генам женщины (носительницы) обладают нормальной или несколько пониженной свертываемостью крови.
Фенотипическое проявление гемофилии у девочек будет наблюдаться в том случае, если мать девочки является носительницей гена гемофилии, а отец — гемофиликом. Подобная закономерность наследования характерна и для других рецессивных, сцепленных с полом признаков.
Сцепленное наследование Править
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несколько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.
Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24).
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов — Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер.
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Понятие о генетической карте Править
Т. Морган и его сотрудники К. Бриджес, А. Г. Стертевант и Г. Дж. Меллер экспериментально показали, что знание явлений сцепления и кроссинговера позволяет не только установить группу сцепления генов, но и построить генетические карты хромосом, на которых указаны порядок расположения генов в хромосоме и относительные расстояния между ними.
Генетической картой хромосом называют схему взаимного расположения генов, находящихся в одной группе сцепления. Такие карты составляются для каждой пары гомологичных хромосом.
Возможность подобного картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др.), грибов (дрожжи, аспергилл), для бактерий и вирусов.
Наличие генетической карты свидетельствует о высокой степени изученности того или иного вида организма и представляет большой научный интерес. Такой организм является прекрасным объектом для проведения дальнейших экспериментальных работ, имеющих не только научное, но и практическое значение. В частности, знание генетических карт позволяет планировать работы по получению организмов с определенными сочетаниями признаков, что теперь широко используется в селекционной практике. Так, создание штаммов микроорганизмов, способных синтезировать необходимые для фармакологии и сельского хозяйства белки, гормоны и другие сложные органические вещества, возможно только на основе методов генной инженерии, которые, в свою очередь, базируются на знании генетических карт соответствующих микроорганизмов.
Генетические карты человека также могут оказаться полезными в здравоохранении и медицине. Знания о локализации гена в определенной хромосоме используются при диагностике ряда тяжелых наследственных заболеваний человека. Уже теперь появилась возможность для генной терапии, то есть для исправления структуры или функции генов.
Сравнение генетических карт разных видов живых организмов способствует также пониманию эволюционного процесса.
Основные положения хромосомной теории наследственности Править
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
- Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
- Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
- Гены расположены в хромосоме в линейной последовательности.
- Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
- Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
- Каждый биологический вид характеризуется определенным набором хромосом — кариотипом.
- Н. А. Лемеза Л. В. Камлюк Н. Д. Лисов «Пособие по биологии для поступающих в ВУЗы»
ru.science.wikia.com
Хромосомная теория наследственности
Создателем хромосомной теории (ХТ) является учёный Томас Морган. ХТ является результатом изучения наследственности на клеточном уровне.
Суть хромосомной теории:
— Материальными носителями наследственности являются хромосомы.
Основными доказательством этому является:
Цитогенетический параллелизм
Хромосомное определение пола
Сцепленное с полом наследование
Сцепление генов и кроссинговер
Основные положения хросомной теории:
Наследственные задатки (гены) локализованы в хромосомах.
Гены расположены в хромосоме в линейном порядке.
Каждый ген занимает определенный участок (локус). Аллельные гены занимают аналогичные локусы в гомологичных хромосомах.
Гены, локализованные в одной хромосоме, наследуются совместно, сцеплено (Закон Моргана) и образуют группу сцепления. Число групп сцепления равно гаплоидному числу хромосом (n).
Между гомологичными хромосомами возможен обмен участками, или рекомбинация.
Расстояние между генами измеряются в процентах кроссинговера – морганидах.
Частота кроссинговера обратно пропорциональна расстоянию между генами, а сила сцепления между генами обратно пропорциональна расстоянию между ними.
Цитогенетический параллелизм
, Дипломником Моргана Сюттоном было замечено что поведение генов по Менделю, совпадает с поведение хромосом: (ТАБЛИЦА – цитогенетический паралелизм)
Каждый организм несёт 2-а наследственных задатка, в гамету входит только 1- ин наследственный задаток из пары. При оплодотворении в зиготе и далее в организме опять 2-а наследственных задатка по каждому признаку.
Точно так же ведут себя и хромосомы, что можно предположить что гены лежат в хромосомах и наследуются вместе с ними.
Хромосомное определение пола
В 1917 году Алленом было показано что мужские и женские особи у мхов отличаются по набору хромосом. В клетках диплоидной ткани мужского организма половые хромосомы XиY, в женскомXиX. Таким образом Хромосомы определяют такой признак как пол, а следовательно могут быть материальными носителями наследственности. Позже хромосомное определение пола было показано и для других организмов, в том числе и для человека.(ТАБЛИЦА)
Сцепленное с полом наследование
Поскольку половые хромосомы различны у мужских и женских организмов, признаки, гены которых, расположены в Х или Yхромосомах, будут наследовать по-разному. Такие признаки называютсясцепленными с полом признаками.
Особенности наследования сцепленных с полом признаков
Не соблюдается 1 закон Менделя
Реципрокные скрещивания дают разный результат
Имеет место крисс-кросс (или наследование крест-накрест).
Впервые наследование связанное с признаком было обнаружено Морганом у дрозофилы.
|
W+ -красные глаза |
P: |
(C) XW+X W+ * XwY |
(C) XwXw * X W+Y | ||||
|
w – белые глаза |
Гаметы: |
XW+; |
Xw, |
Y; |
Xw; |
XW+, |
Y |
|
F1 |
(CЖ)XW+Xw – Красные глаза |
XwXW+ — Красные глаза | |||||
|
(CМ)XW+Y– Красные глаза |
XwY– Белые глаза | ||||||
|
Таким образом наследование выявленной Морганом мутация – “белые глаза” — white, характеризовалась перечисленными выше особенностями:
Такое наследование и называется «наследование крисс-кросс» | |||||||
(ТАБЛИЦА сцепленное с полом наследование)
Сцепленное с полом наследование объясняется отсутствием в Yхромосоме генов, аллельных генамXхромосоме.Yхромосома намного меньше Х хромосомы, в ней, в настоящее время, локализовано 78(?)генов, в то время как вXхромосоме их более 1098.
Примеры сцепленных с полом наследований:
— гемофилия, дистрофия Дюшенна, синдром Данкана, синдром Альпорта, и др.
Есть гены, которые наоборот содержатся в Yхромосоме и отсутствуют вXхромосоме, они, следовательно, встречаются только в мужских организмах, и никогда в женских (голандрическое наследование) и передаются только сыновьям от отца.
Сцепление генов и кроссинговер
В генетике было известно такое явления как «притяжение генов»: некоторые неаллельные признаки наследовались не независимо, как должны по IIIзакону Менделя, а наследовались вместе, не давали новых комбинаций. Морган объяснил это тем, что эти гены находятся в одной хромосоме, поэтому они расходятся в дочерние клетки вместе одной группой, как бы сцеплено. Он назвал это явление –сцепленным наследованием.
Закон сцепления Моргана:
— Гены расположенные в одной хромосоме наследуются совместно, сцеплено.
Гены расположенные в одной хромосоме образуют группу сцепления. Число групп сцепления равно «n» — гаплоидному числу хромосом.
Пример:
Скрещивали гомозиготные линии мух с серым цветом тела и длинными крыльями и мух, имеющих чёрное тело и короткие крылья. Гены цвета тела и длинны крыльев – сцеплены, т.е. лежат в одной хромосоме.
|
А- серое тело а- чёрное тело B- нормальные крылья (длинные) b- зачаточные крылья |
P: |
(С Ж) AABBxaabb(CМ) | ||||
|
Серые длиннокрылые |
Чёрные короткокрылые | |||||
|
Запись в хромосомном выражении | ||||||
|
P: |
| |||||
|
Серое тело Длинные крылья |
Чёрное тело Короткое тело | |||||
|
G: | ||||||
|
F1 |
Все мухи имеют серое тело и длинные крылья | |||||
|
Т.е. в этом случае закон единообразия гибридов Iпоколения соблюдается. Однако вF2вместо ожидаемого расщепления 9:3:3:1 получилось отношение на 3 серых длиннокрылых на 1 часть чёрных короткокрылых, т.е. новых сочетаний признаков не появлялось. Морган предположил что дегетерозиготыF2— ()продуцируют (дают) гаметы не 4, а только 2 типов -и. Проведенные анализирующие скрещивания это подтвердило: | ||||||
|
P |
| |||||
|
Серое тело Длинные крылья |
Чёрное тело Короткое тело | |||||
|
G | ||||||
|
Fa | ||||||
|
Серое тело Длинные крылья |
Чёрное тело Короткие крылья | |||||
В результате в F2расщепление идёт как при моногибридном скрещивании 3:1.
|
P (F1) | ||||
|
G | ||||
|
F2 | ||||
|
Серое тело Длинные крылья |
Серое тело Длинные крылья |
Серое тело Длинные крылья |
Чёрное тело Короткие крылья | |
Кроссинговер.
В небольшом проценте случаев в F2в опытах Моргана появлялись мухи с новыми сочетаниями признаков: крылья длинные, тело черное; крылья короткие, а тело серое. Т.е. признаки «расцепились». Морган объяснил это тем, что хромосомы во время конъюгации в мейозе обмениваются генами. В результате получаются особи с новыми сочетаниями признаков, т.е. как и положено по третьему закону Менделя. Морган назвал этот обмен генами рекомбинацией.
Позже цитологи действительно подтвердили гипотезу Моргана, обнаружив обмен участками хромосом у кукурузы и у саламандры. Они назвали этот процесс кроссинговер.
Кроссинговер увеличивает разнообразие потомства в популяции.
studfiles.net
Лекция № 3 Хромосомная теория наследственности.
Лекция № 3
Хромосомная теория наследственности.
Основные положення хромосомной теории наследственности. Хромосомный анализ.
Формирование хромосомной теории. В 1902—1903 гг. американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бовери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911—1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Хромосомная теория наследственности — теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, то есть преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Хромосомная теория наследственности возникла в начале 20 в. на основе клеточной теории и использовалась для изучения наследственных свойств организмов гибридологического анализа.
Основные положения хромосомной теории наследственности.
1. Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
2. Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
3. Гены расположены в хромосоме в линейной последовательности.
4. Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
5. Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
6. Каждый биологический вид характеризуется определенным набором хромосом — кариотипом.
Сцепленное наследование
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несколько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.
Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например, у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24).
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов— Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер.
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Сцепление и кроссинговер. Из принципов генетического анализа, изложенных в предыдущих главах, с очевидностью вытекает, что независимое комбинирование признаков может осуществляться лишь при условии, что гены, определяющие эти признаки, находятся в негомологичных хромосомах. Следовательно, у каждого организма число пар признаков, по которым наблюдается независимое наследование, ограничено числом пар хромосом. С другой стороны, очевидно, что число признаков и свойств организма, контролируемых генами, чрезвычайно велико, а число пар хромосом у каждого вида относительно мало и постоянно.
Остается предположить, что в каждой хромосоме находится не один ген, а много. Если это так, то третий закон Менделя касается распределения хромосом, а не генов, т. е. его действие ограничено.
Явление сцепленного наследования. Из третьего закона Менделя следует,, что при скрещивании форм, различающихся двумя парами генов (АВ и аb), получается гибрид АaВb, образующий четыре сорта гамет АВ, Аb, аВ и аb в равных количествах.
В соответствии с этим в анализирующем скрещивании осуществляется расщепление 1 : 1 : 1 : 1, т.е. сочетания признаков, свойственные родительским формам (АВ и аb), встречаются с такой же частотой, как и новые комбинации (Аb и аВ),— по 25%. Однако по мере накопления фактов генетики все чаще стали сталкиваться с отклонениями от независимого наследования. В отдельных случаях новые комбинации признаков (Аb и аВ) в Fbсовсем отсутствовали — наблюдалось полное сцепление между генами исходных форм. Но чаще в потомстве в той или иной степени преобладали родительские сочетания признаков, а новые комбинации встречались с меньшей частотой, чем ожидается при независимом наследовании, т.е. меньше 50%. Таким образом, в данном случае гены чаще наследовались в исходном сочетании (были сцеплены), но иногда это сцепление нарушалось, давая новые комбинации.
Совместное наследование генов, ограничивающее их свободное комбинирование, Морган предложил называть сцеплением генов или сцепленным наследованием.
Кроссинговер и его генетическое доказательство. При допущении размещения в одной хромосоме более одного гена встает вопрос, могут ли аллели одного гена в гомологичной паре хромосом меняться местами, перемещаясь из одной гомологичной хромосомы в другую. Если бы такой процесс не происходил, то гены комбинировались бы только путем случайного расхождения негомологичных хромосом в мейозе, а гены, находящиеся в одной паре гомологичных хромосом, наследовались бы всегда сцепленно — группой.
Исследования Т.Моргана и его школы показали, что в гомологичной паре хромосом регулярно происходит обмен генами. Процесс обмена идентичными участками гомологичных хромосом с содержащимися в них генами называют перекрестом хромосом или кроссинговером. Кроссинговер обеспечивает новые сочетания генов, находящихся в гомологичных хромосомах. Явление кроссинговера, так же как и сцепление, оказалось общим для всех животных, растений и микроорганизмов. Наличие обмена идентичными участками между гомологичными хромосомами обеспечивает обмен или рекомбинацию генов и тем самым значительно увеличивает роль комбинативной изменчивости в эволюции. О перекресте хромосом можно судить по частоте возникновения организмов с новым сочетанием признаков. Такие организмы называют рекомбинантами.
Гаметы с хромосомами, претерпевшими кроссинговер, называют кроссоверными, а с непретерпевшими — некроссоверными. Соответственно организмы, возникшие от сочетания кроссоверных гамет гибрида с гаметами анализатора, называют кроссоверами или рекомбинантами, а возникшие за счет некроссоворных гамет гибрида — некроссоверными или нерекомбинантными.
Закон сцепления Моргана. При анализе расщепления в случае кроссинговера обращает на себя внимание определенное количественное отношение кроссоверных и некроссоверных классов. Обе исходные родительские комбинации признаков, образовавшиеся из некроссоверных гамет, оказываются в потомстве анализирующего скрещивания в равном количественном отношении. В указанном опыте с дрозофилой тех и других особей было примерно по 41,5%. В сумме некроссоверные мухи составили 83% от общего числа потомков. Два кроссоверных класса по числу особей также одинаковы, и сумма их равна 17%.
Частота кроссинговера не зависит от аллельного состояния генов, участвующих в скрещивании. Если в качестве родителя использовать мух и , то в анализирующем скрещивании кроссоверные (b+vg и bvg+) и некроссоверные (bvg и b+vg+) особи появятся с той же частотой (соответственно 17 и 83%), что и в первом случае.
Результаты этих опытов показывают, что сцепление генов реально существует, и лишь в известном проценте случаев оно нарушается вследствие кроссинговера. Отсюда и был сделан вывод, что между гомологичными хромосомами может осуществляться взаимный обмен идентичными участками, в результате чего гены, находящиеся в этих участках парных хромосом, перемещаются из одной гомологичной хромосомы в другую. Отсутствие перекреста (полное сцепление) между генами представляет исключение и известно лишь у гетерогаметного пола немногих видов, например у дрозофилы и шелкопряда.
Изученное Морганом сцепленное наследование признаков получило название закона сцепления Моргана. Поскольку рекомбинация осуществляется между генами, а сам ген кроссинговером не разделяется, его стали считать единицей кроссинговера.
Величина кроссинговера. Величина кроссинговера измеряется отношением числа кроссоверных особей к общему числу особей в потомстве от анализирующего скрещивания. Рекомбинация происходит реципрокно, т.е. между родительскими хромосомами осуществляется взаимный обмен; это обязывает подсчитывать кроссоверные классы вместе как результат одного события. Величина кроссинговера выражается в процентах. Один процент кроссинговера составляет единицу расстояния между генами.
Линейное расположение генов в хромосоме. Т. Морган предположил, что гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме; чем реже кроссинговер, тем они ближе друг к другу.
Одним из классических опытов Моргана на дрозофиле, доказывающим линейное расположение генов, был следующий. Самки, гетерозиготные по трем сцепленным рецессивным генам, определяющим желтую окраску тела y, белый цвет глаз w и вильчатые крылья bi, были скрещены с самцами, гомозиготными по этим трем генам. В потомстве было получено 1,2% мух кроссоверных, возникших от перекреста между генами у и w; 3,5% − от кроссинговера между генами w и bi и 4,7% — между у и bi.
Из этих данных с очевидностью вытекает, что процент перекреста является функцией расстояния между генами. Поскольку расстояние между крайними генами у и bi равно сумме двух расстояний между у и w, w и bi, следует предположить, что гены расположены в хромосоме последовательно, т.е. линейно.
Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов в хромосоме строго фиксировано, т. е. каждый ген занимает в хромосоме свое опрделенное место — локус.
Основным положениям хромосомной теории наследственности — парности аллелей, их редукции в мейозе и линейному расположению генов в хромосоме — соответствует однонитчатая модель хромосомы.
Одинарный и множественный перекресты. Приняв положения, что генов в хромосоме может быть много и расположены они в хромосоме в линейном порядке, а каждый ген занимает определённый локус в хромосоме, Морган допустил, что перекрест между гомологичными хромосомами может происходить одновременно в нескольких точках. Это предположение было им доказано тоже на дрозофиле, а затем полностью подтвердилось на ряде других животных, а также на растениях и микроорганизмах.
Кроссинговер, происходящий лишь в одном месте, называют одинарным, в двух точках одновременно – двойным, в трёх – тройным и т.д., т.е. он может быть множественным.
Чем дальше отстоят друг от друга в хромосоме гены, тем больше вероятность двойных перекрестов между ними. Процент рекомбинаций между двумя генами тем точнее отражает расстояние между ними, чем оно меньше, так как в случае малого расстояния уменьшается возможность двойных обменов.
Для учета двойного кроссинговера необходимо иметь дополнительный маркер, находящийся между двумя изучаемыми генами. Определение расстояния между генами осуществляют следующим образом: к сумме процентов одинарных кроссоверных классов прибавляют удвоенный процент двойных кроссинговеров. Удвоение процента двойных кроссинговеров необходимо в связи с тем, что каждый двойной кроссинговер возникает благодаря двум независимым одинарным разрывам в двух точках.
Интерференция. Установлено, что кроссинговер, происшедший в одном месте хромосомы, подавляет кроссинговер в близлежащих районах. Это явление носит название интерференции. При двойном перекресте интерференция проявляется особенно сильно в случае малых расстояний между генами. Разрывы хромосом оказываются зависимыми друг от друга. Степень этой зависимости определяется расстоянием между происходящими разрывами: по мере удаления от места разрыва возможность другого разрыва увеличивается.
Эффект интерференции измеряется отношением числа наблюдаемых двойных разрывов к числу возможных при допущении полной независимости каждого из разрывов.
Локализация гена. Если гены расположены в хромосоме линейно, а частота кроссинговера отражает расстояние между ними, то можно определить местоположение гена в хромосоме.
Прежде чем определить, положение гена, т. е. его локализацию, необходимо определить, в какой хромосоме находится данный ген. Гены, находящиеся в одной хромосоме и наследующиеся сцепленно, составляют группу сцепления. Очевидно, что количество групп сцепления у каждого вида должно соответствовать гаплоидному набору хромосом.
К настоящему времени группы сцепления определены у наиболее изученных в генетическом отношении объектов, причем во всех этих случаях обнаружено полное соответствие числа групп сцепления гаплоидному числу хромосом. Так, у кукурузы (Zea mays) гаплоидный набор хромосом и число групп сцепления составляют 10, у гороха (Pisum sativum) – 7, дрозофилы (Drosophila melanogaster) – 4, домовой мыши (Mus musculus) – 20 и т. п.
Поскольку ген занимает определенное место в группе сцепления, это позволяет устанавливать порядок расположения генов в каждой хромосоме и строить генетические карты хромосом.
Генетические карты. Генетической картой хромосом называют схему относительного расположения генов, находящихся в данной группе сцепления. Они составлены пока лишь для некоторых наиболее изученных с генетической точки зрения объектов: дрозофилы, кукурузы, томатов, мыши, нейроспоры, кишечной палочки и др.
Генетические карты составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют.
Для того, чтобы составить карты, необходимо изучить закономерности наследования большого числа генов. У дрозофилы, например, изучено более 500 генов, локализованных в четырех группах сцепления, у кукурузы — более 400 генов, локализованных в десяти группах сцепления и т.д. При составлении генетических карт указывается группа сцепления, полное или сокращенное название генов, расстояние в процентах от одного из концов хромосомы, принятого за нулевую точку; иногда обозначается место центромеры.
У многоклеточных организмов рекомбинация генов бывает реципрокной. У микроорганизмов она может быть односторонней. Так, у ряда бактерий, например у кишечной палочки (Escherichia coli), перенос генетической информации происходит во время конъюгации клеток. Единственная хромосома бактерии, имеющая форму замкнутого кольца, рвется во время конъюгации всегда в определенной точке и переходит из одной клетки в другую.
Длина переданного участка хромосомы зависит от длительности конъюгации. Последовательность генов в хромосоме оказывается постоянной. В силу этого расстояние между генами на такой кольцевой карте измеряется не в процентах кроссинговера, а в минутах, что отражает продолжительность конъюгации.
Цитологическое доказательство кроссинговера. После того как генетическими методами удалось установить явление кроссинговера, необходимо было получить прямое доказательство обмена участками гомологичных хромосом, сопровождающегося рекомбинацией генов. Наблюдаемые в профазе мейоза картины хиазм могут служить лишь косвенным доказательством этого явления, констатация происшедшего обмена прямым наблюдением невозможна, так как обменивающиеся участками гомологичные хромосомы обычно абсолютна одинаковы но величине и форме.
Чтобы сопоставить цитологические карты гигантских хромосом с генетическими, Бриджес предложил воспользоваться коэффициентом кроссинговера. Для этого он разделил общую длину всех хромосом слюнных желез (1180 мкм) на общую длину генетических карт (279 единиц). В среднем это отношение оказалось равным 4,2. Следовательно, каждой единице перекреста на генетической карте соответствует 4,2 мкм на цитологической карте (для хромосом слюнных желез). Зная расстояние между генами на генетической карте какой-либо хромосомы, можно сравнить относительную частоту перекреста в разных ее районах. Например, в Х-хромосоме дрозофилы гены у и ec находятся на расстоянии 5,5%, следовательно, расстояние между ними в гигантской хромосоме должно быть 4,2 мкм Х 5,5 = 23 мкм, но непосредственное измерение дает 30 мкм. Значит, в этом районе Х-хромосомы кроссинговер идет реже средней нормы.
В силу неравномерного осуществления обменов по длине хромосом гены при нанесении их на карту распределяются на ней с разной плотностью. Следовательно, распределение генов на генетических картах можно рассматривать как показатель возможности осуществления перекреста по длине хромосомы.
Механизм кроссинговера. Еще до открытия перекреста хромосом генетическими методами цитологи, изучая профазу мейоза, наблюдали явление взаимного обвивания хромосом, образования ими χ-образных фигур – хиазм (χ-греческая буква «хи»). В 1909 г. Ф.Янсенс высказал предположение, что хиазмы связаны с обменом участками хромосом. Впоследствии эти картины послужили дополнительным аргументом в пользу гипотезы генетического перекреста хромосом, выдвинутой Т.Морганом в 1911 г.
Механизм перекреста хромосом связан с поведением гомологичных хромосом в профазе I мейоза.
Кроссинговер происходит на стадии четырех хроматид и приурочен к образованию хиазм.
Если в одном биваленте произошел не один обмен, а два и более, то и этом случае образуется несколько хиазм. Поскольку в биваленте четыре хроматиды, то, очевидно, каждая из них имеет равную вероятность обменяться участками с любой другой. При этом в обмене могут участвовать две, три или четыре хроматиды.
Обмен внутри сестринских хроматид не может приводить к рекомбинациям, поскольку они генетически идентичны, и в силу этого такой обмен не имеет смысла в качестве биологического механизма комбинативной изменчивости.
Соматический (митотический) кроссинговер. Как уже говорилось, кроссинговер происходит в профазе I мейоза при образовании гамет. Однако существует соматический, или митотический, кроссинговер, который осуществляется при митотическом делении соматических клеток главным образом эмбриональных тканей.
Известно, что гомологичные хромосомы в профазе митоза обычно не конъюгируют и располагаются независимо друг от друга. Однако иногда удается наблюдать синапсис гомологичных хромосом и фигуры, похожие на хиазмы, но при этом редукции числа хромосом не наблюдается.
Гипотезы о механизме кроссинговера. По поводу механизма перекреста существует несколько гипотез, но ни одна из них не объясняет полностью фактов рекомбинации генов и наблюдаемых при этом цитологических картин.
Согласно гипотезе, предложенной Ф.Янсенсом и развитой К.Дарлингтоном, в процессе синапсиса гомологичных хромосом в биваленте создается динамическое напряжение, возникающее в связи со спирализацией хромосомных нитей, а также при взаимном обвивании гомологов в биваленте. В силу этого напряжения одна из четырех хроматид рвется. Разрыв, нарушая равновесие в биваленте, приводит к компенсирующему разрыву в строго идентичной точке какой-либо другой хроматиды этого же бивалента. Затем происходит реципрокное воссоединение разорванных концов, приводящее к кроссинговеру. Согласно этой гипотезе хиазмы непосредственно связаны с кроссинговером.
По гипотезе К.Сакса хиазмы не являются результатом кроссинговера: сначала образуются хиазмы, а затем происходит обмен. При расхождении хромосом к полюсам вследствие механического напряжения в местах хиазм происходят разрывы и обмен соответствующими участками. После обмена хиазма исчезает.
Смысл другой гипотезы, предложенной Д.Беллингом и модернизированной И.Ледербергом, заключается в том, что процесс репликации ДНК может реципрокно переключаться с одной нити на другую; воспроизведение, начавшись на одной матрице, с какой-то точки переключается на матричную нить ДНК.
Факторы, влияющие на перекрест хромосом. На кроссинговер влияет множество факторов как генетической природы, так и внешней среды. Поэтому в реальном эксперименте о частоте кроссинговера можно говорить, имея в виду все те условия, в которых она была определена. Кроссинговер практически отсутствует между гетероморфными Х— и Y-хромосомами. Если бы он происходил, то хромосомный механизм определения пола постоянно разрушался бы. Блокирование кроссинговера между этими хромосомами связано не только с различием в их величине (оно наблюдается не всегда), но и обусловлено Y-специфичными нуклеотидными последовательностями. Обязательное условие синапса хромосом (или их участков) — гомология нуклеотидных последовательностей.
Для абсолютного большинства высших эукариот характерна примерно одинаковая частота кроссинговера как у гомогаметного, так и гетерогаметного полов. Однако есть виды, у которых кроссинговер отсутствует у особей гетерогаметного пола, в то время как у особей гомогаметного пола он протекает нормально. Такая ситуация наблюдается у гетерогаметных самцов дрозофилы и самок шелкопряда. Существенно, что частота митотического кроссинговера у этих видов у самцов и самок практически одинакова, что указывает на различные элементы контроля отдельных этапов генетической рекомбинации в половых и соматических клетках. В гетерохроматических районах, в частности прицентромерных, частота кроссинговера снижена, и поэтому истинное расстояние между генами в этих участках может быть изменено.
Обнаружены гены, выполняющие роль запирателей кроссинговера, но есть также гены, повышающие его частоту. Они иногда могут индуцировать заметное число кроссоверов у самцов дрозофилы. В качестве запирателей кроссинговера могут выступать также хромосомные перестройки, в частности инверсии. Они нарушают нормальную конъюгацию хромосом в зиготене.
Обнаружено, что на частоту кроссинговера влияют возраст организма, а также экзогенные факторы: температура, радиация, концентрация солей, химические мутагены, лекарства, гормоны. При большинстве указанных воздействий частота кроссинговера повышается.
В целом кроссинговер представляет собой один из регулярных генетических процессов, контролируемых многими генами как непосредственно, так и через физиологическое состояние мейотических или митотических клеток. Частота различных типов рекомбинаций (мейотический, митотический кроссинговер и сестринские, хроматидные обмены) может служить мерой действия мутагенов, канцерогенов, антибиотиков и др.
Законы наследования Моргана и вытекающие из них принципы наследственности. Огромную роль в создании и развитии генетики сыграли работы Т.Моргана. Он автор хромосомной теории наследственности. Им были открыты законы наследования: наследование признаков, сцепленных с полом, сцепленное наследование.
Из этих законов вытекает следующие принципы наследственности:
1. Фактор-ген есть определённый локус хромосомы.
2. Аллели гена расположены в идентичных локусах гомологичных хромосом.
3. Гены расположены в хромосоме линейно.
4. Кроссинговер – регулярный процесс обмена генами между гомологичными хромосомами.
Мобильные элементы генома. В 1948 г. американская исследовательница Мак-Клинток открыла у кукурузы гены перемещающиеся из одного участка хромосомы в другой и назвала феномен транспозицией, а сами гены контролириующими элементами (КЭ). 1.Эти элементы могут перемещаться из одного сайта в другой; 2. их встраивание в данный район влияет на активность генов расположенных рядом; 3. утрата КЭ в данном локусе превращает прежде мутабильный локус в стабильный; 4. в сайтах, в которых присутствуют КЭ, могут возникать делеции, транслокации, транспозиции, инверсии, а также разрывы хромосом. В 1983 г. за открытие мобильных генетических элементов Нобелевская премия была присуждена Барбаре Мак-Клинток.
Наличие мобильных элементов в геномах имеет разнообразные последствия:
1. Перемещения и внедрение мобильных элементов в гены может вызывать мутации;
2. Изменение состояния активности генов;
3. Формирование хромосомных перестроек;
4. Формирование теломер.
5. Участие в горизонтальном переносе генов;
6. Транспозоны на основе Р-элемента используют для трансформации у эукариот, клонирования генов, поиска энхансеров и т.д.
У прокариот существуют три типа мобильных элементов – IS-элементы (инсерции), транспозоны, и некоторые бактериофаги. IS-элементы встраиваются в любой участок ДНК, часто вызывают мутации, разрушая кодирующие или регуляторные последовательности, влияют на экспрессию соседних генов. Бактериофаг может вызывать мутации в результате встраивания.
kursak.net
Хромосомная теория наследственности и её положения | Биология. Реферат, доклад, сообщение, краткое содержание, лекция, шпаргалка, конспект, ГДЗ, тест
Раздел: Наследственность
Считается, что формирование хромосомной теории наследственности стало следующим этапом развития клеточной теории. Главные положения хромосомной теории сейчас рассматривают как постулаты современной клеточной теории.
Вспомните мушиную комнату, о которой шла речь при изучении истории генетики. В ней проводились опыты, ставшие основой одного из самых важных открытий в биологии — хромосомной теории наследственности. Оно было сформулировано ещё в начале XX ст. и остаётся актуальным поныне.
Ещё до исследований Моргана, сразу же после переоткрытия законов Менделя учёные заметили, что при формировании половых клеток хромосомы ведут себя, как гены. Но именно Морган через 10 лет после переоткрытия законов наследования доказал, что гены находятся в хромосомах. Именно он сформулировал главное положение хромосомной теории наследственности: гены находятся в хромосомах и расположены там в линейном порядке.
Современные молекулярно-генетические исследования подтвердили эти положения. Реализация программы «Геном человека», в рамках которой исследовалось, какие именно гены расположены в каких именно хромосомах (рис. 39), дала следующие результаты. Было установлено, что каждая хромосома содержит от нескольких сотен до нескольких тысяч структурных генов — всего 32 135 генов. Причём количество генов в хромосоме часто не совпадает с её размерами. Например, 5-я огромная субметацентрическая хромосома несёт только 609 генов, а самая маленькая акроцентрическая 22-я хромосома имеет 1092 гена.
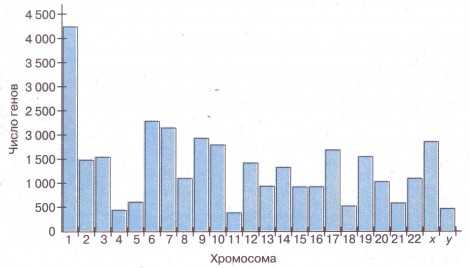 |
| Рис. 39. Число генов в каждой из хромосом человека |
Со времён Моргана хромосомная теория постоянно развивалась и в современном виде включает несколько положений.
- гены находятся в хромосомах;
- каждый ген занимает определённое место в хромосоме;
- гены в хромосомах расположены в линейном порядке;
- каждая хромосома представляет собой группу сцепления;
- число групп сцепления равно гаплоидному числу хромосом;
- между гомологичными хромосомами происходит обмен аллельными генами;
- расстояние между генами пропорционально проценту кроссинговера между ними. Материал с сайта http://worldofschool.ru
Одним из самых важных открытий биологии XX столетия является формулирование хромосомного закона наследственности. Согласно этому закону все гены размещаются в хромосомах в линейном порядке. В соответствии с законами Менделя независимо наследуются, только гены, расположенные в разных хромосомах. Гены, находящиеся в одной хромосоме, наследуются вместе. Однако со временем выяснилось, что и здесь есть своё исключение — кроссинговер, который представляет собой обмен участками гомологических хромосом в процессе их конъюгации.
На этой странице материал по темам:-
Географическое положение генов в хромосоме
Хромосомная теория наследственности конспект
Краткое содержание о хромосомном теории
В чём суть наследственности
Хромосомная теория наследственности кратко
-
В чём суть хромосомной теории наследственности?
-
Назовите основные положения хромосомной теории наследственности.
worldofschool.ru
Хромосомная теория наследственности
После того, как в генетике утвердилось понятие о наследственных факторах, были проведены исследования с целью определить, с какими клеточными структурами они связаны.
Факты, установленные генетическими и цитологическими работами еще в начале текущего столетия, показывали, что носители наследственных факторов (генов) — хромосомы.
В результате дальнейшего развития генетики появилась хромосомная теория наследственности. Создатель ее — американский генетик Т. Морган.
Ученый проводил исследования на плодовой мушке дрозофиле, которую можно легко разводить в пробирках. У этой мушки очень короткий цикл развития: в течение двух недель из оплодотворенного яйца через промежуточные стадии личинки и куколки развивается взрослая особь, способная сразу же давать потомство. Одна оплодотворенная самка может дать несколько сот новых насекомых.
Дрозофила имеет большое число хорошо отличимых признаков, наследование которых легко наблюдать при различных видах скрещиваний. В соматических клетках у нее всего четыре пары хромосом.
Вследствие названных особенностей дрозофила оказалась очень удобным объектом для генетических исследований. На основе опытов с ней были разработаны многие важнейшие вопросы общей генетики.
Многочисленные лабораторные опыты, проведенные Т. Морганом, показали, что гены, находящиеся в одной хромосоме, наследуются, как правило, совместно, то есть оказываются сцепленными и поэтому не подчиняются установленному Г. Менделем правилу независимого комбинирования.
В одном из опытов Т. Моргана дрозофила, имевшая серую окраску тела и длинные крылья, была скрещена с особью, имевшей черную окраску тела и рудиментарные (укороченные) крылья. Первое поколение мух было с серым телом и длинными крыльями. При скрещивании этих гибридов между собой в F2 не произошло независимого распределения признаков по двум аллельным парам (серое тело — черное тело, длинные крылья — рудиментарные крылья) в отношении 9:3:3:1. Среди гибридов F1 преобладающее число особей унаследовало такую же комбинацию признаков, какой она была у родительских форм (серые длиннокрылые и черные короткокрылые), и лишь очень небольшая часть мух была с перекомбинированными признаками (серые короткокрылые и черные длиннокрылые). Этот пример показывает, что гены, обусловливающие признаки серого тела и длинных крыльев и черного тела и коротких крыльев, наследуются преимущественно вместе.
На основании этого и большого числа подобных опытов Т. Морган пришел к выводу, что материальная основа сцепления генов — хромосома. Каждая из хромосом по своей длине неоднородна, она состоит из отдельных элементарных наследственных единиц — генов. У любого вида организмов их всегда во много раз больше, чем хромосом. Следовательно, в каждой хромосоме находится определенное число генов, которые наследуются совместно, образуя, так называемые группы сцепления. Число групп сцепления соответствует числу пар гомологичных хромосом.
Изучая явление сцепления генов, Т. Морган и его ученики установили, что сцепление почти никогда не бывает полным.
В разбираемом примере оно также не было полным, поскольку в небольшом числе случаев отмечена перекомбинация генов. Если гены разных аллельных пар лежат в одной и той же хромосоме, то есть сцеплены, то единственной причиной их перекомбинации может быть процесс конъюгации гомологичных хромосом в профазе мейоза. Во время конъюгации парные хромосомы сближаются и прикладываются одна к другой гомологичными участками, образуя биваленты (четверки хроматид).
В это время между хроматидами может происходить обмен гомологичными участками. Этот процесс получил название перекреста хромосом или кроссинговера (от англ. кроссинг — перекрещивание).
Показана схема перекреста хромосом и рекомбинации находящихся в них генов. Две парные хромосомы в результате перекреста и последующего разрыва обмениваются участками. Два гена А к В, расположенные первоначально в одной хромосоме, в результате кроссинговера оказываются в разных хромосомах и попадают в разные гаметы.
Гаметы с хромосомами, претерпевшими кроссинговер, называются кроссоверными, а гаметы, образованные хромосомами без кроссинговера, — некроссоверными. Соответственно этому и особи, возникшие с участием кроссоверных гамет, называются кроссоверными, или рекомбинантными, а образованные без них — некроссоверными, или нерекомбинантными.
Рекомбинация генов в процессе скрещивания приводит к новообразованиям. Возникают гибридные формы, представляющие исходный материал для отбора и создания новых сортов растений и пород животных. Образование гибридных форм в природе дает материал для естественного отбора, поэтому имеет важнейшее значение в эволюции живых организмов.
Таким образом, перекомбинация генов в процессе мейоза осуществляется двумя путями — случайным расхождением негомологичных хромосом (правило независимого комбинирования по Г. Менделю) и процессом перекреста гомологичных хромосом (явление кроссинговера, установленное Т. Морганом).
В итоге разбора основных положений хромосомной теории наследственности можно сделать следующие выводы.
- Гены находятся в хромосомах, расположены линейно и образуют группу сцепления.
- Гены, локализованные в одной хромосоме, наследуются сцепленно; сила этого сцепления зависит от расстояния между генами.
- Между гомологичными хромосомами наблюдается перекрест, в результате которого происходит рекомбинация генов, имеющая важное значение как источник материала для естественного и искусственного отбора.
- Сцепление генов и их рекомбинация в результате перекреста — закономерные биологические явления, в которых выражается единство процессов наследственности и изменчивости организмов.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
www.activestudy.info
Хромосомная теория наследственности — это… Что такое Хромосомная теория наследственности?
Хромосомная теория наследственности[1] — теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, то есть преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Хромосомная теория наследственности возникла в начале 20 в. на основе клеточной теории и использовалась для изучения наследственных свойств организмов гибридологического анализа.
Формирование хромосомной теории
В 1902-1903 гг. американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бовери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911—1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов.
Генетика пола
Пол, как и любой другой признак организма, наследственно детерминирован. Важнейшая роль в генетической детерминации пола и в поддержании закономерного соотношения полов принадлежит хромосомному аппарату.
У раздельнополых организмов (животных и двудомных растении) соотношение полов обычно составляет 1:1, то есть мужские и женские особи встречаются одинаково часто. Это соотношение совпадает с расщеплением в анализирующем скрещивании, когда одна из скрещиваемых форм является гетерозиготной (Аа), а другая — гомозиготной по рецессивным аллелям (аа). В потомстве в этом случае наблюдается расщепление в отношении 1Аа:1аа. Если пол наследуется по такому же принципу, то вполне логично было бы предположить, что один пол должен быть гомозиготным, а другой — гетерозиготным. Тогда расщепление по полу должно быть в каждом поколении равным 1:1, что и наблюдается в действительности.
При изучении хромосомных наборов самцов и самок ряда животных между ними были обнаружены некоторые различия. Как у мужских, так и у женских особей во всех клетках имеются пары одинаковых (гомологичных) хромосом, но по одной паре хромосом они различаются. Так, у самки дрозофилы имеются две палочковидные хромосомы, а у самца — одна такая же палочковидная, а вторая, парная первой, — изогнутая. Такие хромосомы, по которым самцы и самки отличаются друг от друга, называют половыми хромосомами. Те из них, которые являются парными у одного из полов, называют X-хромосомами (например, у дрозофилы и млекопитающих) или Z-хромосомами (например, у птиц). Непарная половая хромосома, имеющаяся у особей только одного пола, была названа У-хромосомой (у дрозофилы и млекопитающих) или W-хромосомой (у птиц). Хромосомы, в отношении которых между самцами и самками нет различий, называют аутосомами. Следовательно, у дрозофилы особи обоих полов имеют по шесть одинаковых аутосом плюс две половые хромосомы (ХХ у самок и XY у самцов).
Пол, имеющий различные половые хромосомы (X и У), образует гаметы двух типов (половина с X-хромосомой и половина с У-хромосомой), то есть, является гетерогаметным, а пол, содержащий в каждой клетке одинаковые половые хромосомы (X-хромосомы), — гомогаметным.
Открытие половых хромосом и установление их роли в определении пола послужило важным доводом в пользу того, что хромосомы определяют признаки организма.
Определение пола
От чего же зависит рождение мужских и женских особей? Рассмотрим это на примере определения пола у дрозофилы. В ходе гаметогенеза у самок образуется один тип гамет, содержащий гаплоидный набор аутосом и одну X-хромосому. Самцы образуют два типа гамет, половина из которых содержит три аутосомы и одну X-хромосому (ЗА+Х), а половина — три аутосомы и одну У-хромосому (ЗА+У). При оплодотворении яйцеклеток (ЗА+Х) сперматозоидами с X-хромосомами будут формироваться самки (6А+ХХ), а от слияния яйцеклеток со сперматозоидами, несущими У-хромосому, — самцы (6A+XY). Поскольку число мужских гамет с X- и У-хромосомами одинаково, то и количество самцов и самок тоже одинаково. В данном случае пол организма определяется в момент оплодотворения и зависит от хромосомного набора зиготы.
Сходный способ определения пола (XY-тип) присущ всем млекопитающим, в том числе и человеку, клетки которого содержат 44 аутосомы и две X-хромосомы у женщин либо XY-хромосомы у мужчин.
Таким образом, XY-тип определения пола, или тип дрозофилы и человека, — самый распространенный способ определения пола, характерный для большинства позвоночных и некоторых беспозвоночных. Х0-тип встречается у большинства прямокрылых, клопов, жуков, пауков, у которых Y-хромосомы нет вовсе, так что самец имеет генотип Х0, а самка — XX.
У всех птиц, большинства бабочек и некоторых пресмыкающихся самцы являются гомогаметным полом, а самки —- гетерогаметным (типа XY или типа ХО). Половые хромосомы у этих видов обозначают буквами Z и W, чтобы выделить таким образом данный способ определения пола; при этом набор хромосом самцов обозначают символом ZZ, а самки — символом ZW или Z0.
Доказательства того, что половые хромосомы определяют пол организма, были получены при изучении нерасхождения половых хромосом у дрозофилы. Если в одну из гамет попадут обе половые хромосом, а в другую — ни одной, то при слиянии таких гамет с нормальными могут получиться особи с набором половых хромосом ХХХ, ХО, ХХУ и др. Выяснилось, что у дрозофилы особи с набором ХО — самцы, а с набором ХХУ — самки (у человека — наоборот). Особи с набором ХХХ имеют гипертрофированные признаки женского пола (сверхсамки). (Особи со всеми этими хромосомными аберрациями у дрозофилы стерильны). В дальнейшем было доказано, что у дрозофилы пол определяется соотношением (балансом) между числом X-хромосом и числом наборов аутосом.
Наследование признаков, сцепленных с полом
В том случае, когда гены, контролирующие формирование того или иного признака, локализованы в аутосомах, наследование осуществляется независимо от того, кто из родителей (мать или отец) является носителем изучаемого признака. Если же гены находятся в половых хромосомах, характер наследования признаков резко изменяется. Например, у дрозофилы гены, локализованные в X-хромосоме, как правило, не имеют аллелей в У-хромосоме. По этой причине рецессивные гены в X-хромосоме гетерогаметного пола практически всегда проявляются, будучи в единственном числе.
Признаки, гены которых локализованы в половых хромосомах, называются признаками, сцепленными с полом. Явление наследования, сцепленного с полом, было открыто Т. Морганом у дрозофилы.
Х- и У-хромосомы у человека имеют гомологичный (псевдоаутосомный) участок, где локализованы гены, наследование которых не отличается от наследования аутосомных генов.
Помимо гомологичных участков, X- и У-хромосомы имеют негомологичные участки. Негомологичный участок У-хромосомы, кроме генов, определяющих мужской пол, содержит гены перепонок между пальцами ног и волосатых ушей у человека. Патологические признаки, сцепленные с негомологичным участком У-хромосомы, передаются всем сыновьям, поскольку они получают от отца У-хромосому.
Негомологичный участок X-хромосомы содержит в своем составе ряд важных для жизнедеятельности организмов генов. Поскольку у гетерогаметного пола (ХУ) X-хромосома представлена в единственном числе, то признаки, определяемые генами негомологичного участка X-хромосомы, будут проявляться даже в том случае, если они рецессивны. Такое состояние генов называется гемизиготным. Примером такого рода X-сцепленных рецессивных признаков у человека являются гемофилия, мышечная дистрофия Дюшена, атрофия зрительного нерва, дальтонизм (цветовая слепота) и др.
Гемофилия — это наследственная болезнь, при которой кровь теряет способность свертываться. Ранение, даже царапина или ушиб, могут вызвать обильные наружные или внутренние кровотечения, которые нередко заканчиваются смертью. Это заболевание встречается, за редким исключением, только у мужчин. Было установлено, что обе наиболее распространенные формы гемофилии (гемофилия А и гемофилия В) обусловлена рецессивными генами, локализованными в X-хромосоме. Гетерозиготные по данным генам женщины (носительницы) обладают нормальной или несколько пониженной свертываемостью крови.
Фенотипическое проявление гемофилии у девочек будет наблюдаться в том случае, если мать девочки является носительницей гена гемофилии, а отец — гемофиликом. Подобная закономерность наследования характерна и для других рецессивных, сцепленных с полом признаков.
Сцепленное наследование
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несколько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.
Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например, у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24).
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов — Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер.
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Понятие о генетической карте
Т. Морган и его сотрудники К. Бриджес, А. Г. Стертевант и Г. Дж. Меллер экспериментально показали, что знание явлений сцепления и кроссинговера позволяет не только установить группу сцепления генов, но и построить генетические карты хромосом, на которых указаны порядок расположения генов в хромосоме и относительные расстояния между ними.
Генетической картой хромосом называют схему взаимного расположения генов, находящихся в одной группе сцепления. Такие карты составляются для каждой пары гомологичных хромосом.
Возможность подобного картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др.), грибов (дрожжи, аспергилл), для бактерий и вирусов.
Наличие генетической карты свидетельствует о высокой степени изученности того или иного вида организма и представляет большой научный интерес. Такой организм является прекрасным объектом для проведения дальнейших экспериментальных работ, имеющих не только научное, но и практическое значение. В частности, знание генетических карт позволяет планировать работы по получению организмов с определенными сочетаниями признаков, что теперь широко используется в селекционной практике. Так, создание штаммов микроорганизмов, способных синтезировать необходимые для фармакологии и сельского хозяйства белки, гормоны и другие сложные органические вещества, возможно только на основе методов генной инженерии, которые, в свою очередь, базируются на знании генетических карт соответствующих микроорганизмов.
Генетические карты человека также могут оказаться полезными в здравоохранении и медицине. Знания о локализации гена в определенной хромосоме используются при диагностике ряда тяжелых наследственных заболеваний человека. Уже теперь появилась возможность для генной терапии, то есть для исправления структуры или функции генов.
Сравнение генетических карт разных видов живых организмов способствует также пониманию эволюционного процесса.
Основные положения хромосомной теории наследственности
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
- Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
- Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
- Гены расположены в хромосоме в линейной последовательности.
- Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
- Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
- Каждый биологический вид характеризуется определенным набором хромосом — кариотипом.
Источники
- Н. А. Лемеза Л. В. Камлюк Н. Д. Лисов «Пособие по биологии для поступающих в ВУЗы»
Примечания
- ↑ 2006-2008 г. © Научно-Образовательный портал «Вся Биология» Копирование материалов с данного сайта разрешено, при условии наличия ссылки на ресурс «Вся Биология» [1]
dic.academic.ru
