
Химический состав прокариотной клетки. Пищевые потребности прокариот. Источники углерода.
Химический состав клеток в принципе одинаков у всех организмов. Клетки прокариот содержат от 70 до 90% воды. Основную массу сухих веществ, на долю которых приходятся остальные 10–30%, составляют белки, нуклеиновые кислоты, липиды и полисахариды. Несколько процентов сухого вещества клеток приходится на низкомолекулярные органические вещества и соли. Макромолекулы, составляющие основную массу сухих веществ клетки, — полимеры, построенные из мономерных единиц. Исключением служат липиды, не являющиеся полимерами, так как молекулы в них не соединены между собой ковалентными связями. Углеводные полимеры построены на основе повторяющихся единиц одного, двух или более типов, например, запасной полисахарид гликоген, построенный из остатков глюкозы, или пептидогликан клеточной стенки, образованный чередованием N-ацетилглюкозамина и N-ацетилмурамовой кислоты. В клетке углеводные полимеры представлены часто одним видом молекулы.
Прокариотная клетка в норме содержит примерно 2000—2500 различных белков, каждый из которых представлен 400—1000 молекулами. Количество молекул нуклеиновых кислот каждого вида определяется их функциональным назначением: ДНК — одного вида и представлена одной или несколькими копиями; количество разных молекул РНК в клетке колеблется на несколько порядков.
Пит. В-ва. Мономеры, необходимые для построения основных клеточных компонентов, могут быть синтезированы клеткой или поступать в готовом виде из среды. Чем больше готовых соединений должен получать организм извне, тем ниже уровень его биосинтетических способностей, так как химическая организация всех свободноживущих форм одинакова.
углеводы. Автотрофы – сами синтезируют всю органику из СО2. гетеротрофы – из органики или любых других в-в.
наибольшие гетеротрофы – облигатные внутриклеточные паразиты., потом идут факультативные, а птотм сапрофиты.
паразитам нужны все в-ва, тк у них многие метаболические пути редуцированы. Даже АЬФ берут у хозяина. А сапрофиты просто берут готовуую органику. АТФ сами делают, непосредственно от др не зависят.
Олиготрофы – растут при малых конц в-ва – 10мкгл углерода. А есть копиотрофы – 10 г углерода на Л среды.
Азот. В расчете на сухие вещества его содержится приблизительно 10%. Природный азот бывает в окисленной, восстановленной и молекулярной формах. Подавляющее большинство прокариот усваивают азот в восстановленной форме. Это соли аммония, мочевины, органические соединения (аминокислоты или пептиды). Окисленные формы азота, главным образом нитраты, также могут потребляться многими прокариотами. Так как азот в конструктивном клеточном метаболизме используется в форме аммиака, нитраты перед включением в органические соединения должны быть восстановлены.
Сера и фосфор. Сера входит в состав аминокислот (цистеин, метионин), витаминов и кофакторов (биотин, липоевая кислота, кофермент А и др.), а фосфор — необходимый компонент нуклеиновых кислот, фосфолипидов, коферментов. В природе сера находится в форме неорганических солей, главным образом сульфатов, в виде молекулярной (элементной) серы или входит в состав органических соединений. Большинство прокариот для биосинтетических целей потребляют серу в форме сульфата, который при этом восстанавливается до уровня сульфида. Однако некоторые группы прокариот не способны к восстановлению сульфата и нуждаются в восстановленных соединениях серы. Основной формой фосфора в природе являются фосфаты, которые и удовлетворяют потребности прокариот в этом элементе.
Металлы. Всем прокариотным организмам необходимы металлы, которые могут использоваться в форме катионов неорганических солей. Некоторые из них (магний, кальций, калий, железо) нужны в достаточно высоких концентрациях, потребность в других (цинк, марганец, натрий, молибден, медь, ванадий, никель, кобальт) невелика. Роль перечисленных выше металлов определяется тем, что они входят в состав основных клеточных метаболитов и, таким образом, участвуют в осуществлении жизненно важных функций организма.
Факторы роста. Некоторые прокариоты обнаруживают потребность в одном каком-либо органическом соединении из группы витаминов, аминокислот или азотистых оснований, которое они по каким-то причинам не могут синтезировать из используемого источника углерода. Такие органические соединения, необходимые в очень небольших количествах, получили название факторов роста. Организмы, которым в дополнение к основному источнику углерода необходим один или больше факторов роста, называют ауксотрофами, в отличие от прототрофов, синтезирующих все необходимые органические соединения из основного источника углерода.
Ауксотрофы – рганизмы, которые не способны синтезировать определенноеорганическое соединение, необходимое для роста этого организма. Ауксотрофия — характеристика подобных организмов, этот термин противоположен прототрофии.
Растворение твердого полимерного органического вещества осуществляется под действием экзоферментов — гидролаз. Организмы, обладающие такой способностью, получили название гидролитиков. Растворимые полимеры разлагаются под действием гидролаз, чтобы превратиться в транспортируемые в клетки низкомолекулярные соединения, например, под действием амилазы для крахмала или протеаз для растворимых белков. Но это не составляет трудностей, и крахмал или пектин, обычно относят к легкоусвояемым веществам. Их разлагают, соответственно, ами-лолитические и пектолитические организмы. Труднее разлагаются капсульные слизи, которые по своему назначению должны быть устойчивыми. Нерастворимые полимеры, ВОВ, должны быть переведены в растворенное состояние. В биотехнологии процесс получил название твердофазной (гетерофазной) ферментации, скорость которой, очевидно, прямо зависит от заселенной поверхности, а не концентрации ВОВ. Гидролитики разделяются на три основные группы: сахаролитическую, пептолитическую, липолитическую, что соответствует трем классам субстратов — полисахаридам, белкам, липидам. При гидролизе ВОВ должен быть обеспечен плотный контакт между поверхностью твердой частицы и гидролазами. Это достигается при минимальном расстоянии между бактериальной клеткой и поверхностью, а также за счет создания слизистого чехла, не позволяющего гидролазам рассеиваться в окружающей среде.
Диссиптотрофы – доедают за гидролитиками то, что почему то они не съели – продукты гидролиза полимеров, тк сами этого не умеют. Представл микрофлору растений.
Газотрофы — группа бактерий, использующих в качестве источника энергии восстановленные газы. СН4 окисляют метанотрофы, Н2 – водородные бактерии, СО –карбоксидобактерии, Nh4 – нитрификаторы, Н2S – серобактерии. При этом многие Г. способны окислятьвещества в очень низкой концентрации, являясь, таким образом, олиготрофами. Г. являются составнойчастью «бактериального окислительного фильтра», их сообщество располагается в водоемах на границераздела анаэробной и аэробной зон. Образование восстановленных газов происходит в анаэробной зоневодоемов при деградации органического вещества. Окисляя эти газы кислородом, Г. препятствуют их выходув атмосферу.
students-library.com
ЛЕКЦИЯ №3. СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ ПРОКАРИОТОВ
3.1.Общее строение прокариотической клетки
3.2.Поверхностные структуры клетки
3.2.1.Строение клеточной стенки
3.2.2.Капсулы, слизистые слои и чехлы3.2.3.Жгутики и механизмы движения
3.2.4.Ворсинки
3.3.Мембранные структуры клетки
3.4.Цитоплазма и ее содержимое
3.5.Клеточные включения
3.6.Генетический аппарат (слайд 3.3)
3.1. Общеестроениепрокариотическойклетки
Все микроорганизмы в зависимости от строения клетки делят на эукариоты и прокариоты. К эукариотным микроорганизмам относятся водоросли, грибы и простейшие; к прокариотам – бактерии (слайд 3.4).
Прокариотный и эукариотный типы клеточной организации являются наиболее существенной границей, разделяющей все клеточные формы жизни
(слайд 3.5).
3.2. Поверхностныеструктурыклетки
Клетка состоит из протопласта и поверхностных структур. Поверхностные структуры клетки, расположенные снаружи от ЦПМ – клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки. Протопласт
– ЦПМ вместе с цитоплазмой (слайд 3.6).
3.2.1. Строениеклеточнойстенки
Клеточная стенка – важный и обязательный структурный элемент большинства бактерий. На долю клеточной стенки приходится от 5 до 50% сухих веществ клетки (слайд 3.7).
В состав клеточной стенки входят специфические полимерные комплексы, которые не содержатся в других структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты окрашиваются по-разномуи делятся на две группы: грамположительные и грамотрицательные. Способ окраски был предложен в 1884 г. датским ученым X. Грамом, занимавшимся окрашиванием тканей
(слайд 3.8).
Микробиология с основами вирусологии. Конспект лекций | -20- |

ЛЕКЦИЯ №3. СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ ПРОКАРИОТОВ
3.2. Поверхностные структуры клетки
Клеточные стенки грамположительных и грамотрицательных бактерий резко различаются как по химическому составу, так и по ультраструктуре.
Клеточная стенка грамположительных бактерий плотно прилегает к ЦПМ. Под электронным микроскопом она выглядит как гомогенный электронно-плотный слой, толщина которого колеблется от 20 до 80 нм.
У грамположительных бактерий пептидогликан составляет основную массу вещества клеточной стенки (40-90%).Пептидогликан – это гетерополимер, состоящий из чередующихся остатковN-ацетилглюкозаминаиN-ацетилмурамовойкислоты, соединенных между собойβ-1,4-гликозидными связями (рисунок). КN-ацетилмурамовойкислоте присоединен короткий пептидный хвост, состоящий из небольшого числа (обычно4–5)аминокислот. У грамположительных бактерий обнаружено более 100 различных химических типов пептидогликана. Большинство различий относится к пептидной части его молекулы (слайд 3.9).
Две особенности пептидного хвоста:
1.наличие аминокислот в D-форме(неприродная конфигурация) и
2.высокое содержание аминокислот с двумя аминогруппами.
Вторые аминогруппы участвуют в формировании дополнительных пептидных связей между гетерополимерными цепочками.
Всостав клеточных стенок грамположительных бактерий входят тейхоевые кислоты. Это полимеры, построенные на основе рибита или глицерина, соединенных между собой фосфодиэфирными связями. Некоторые свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками D-аланина,глюкозы,N-ацетилглюкозаминаи некоторых других сахаров (слайд 3.10).
Тейхоевые кислоты ковалентно соединяются с N-ацетилмурамовойкислотой. Как полианионы тейхоевые кислоты определяют поверхностный заряд клетки. Сахарные компоненты тейхоевых кислот входят в состав рецепторов для некоторых бактериофагов и определяют возможность адсорбции фага на клеточной поверхности (слайд 3.11).
Всоставе клеточной стенки грамположительных бактерий в небольших количествах также найдены полисахариды, белки и липиды.
Клеточная стенка грамотрицательных бактерий многослойная.
Внутренний электронно-плотныйслой(2-3нм) состоит из пептидогликана. Снаружи к нему прилегает волнистый слой(8-10нм), имеющий строение, характерное для элементарных мембран – наружная мембрана. Слой пептидогликана отделен от ЦПМ периплазматическим пространством. У грамотрицательных бактерий содержание пептидогликана значительно меньше(1-10%).У большинства видов он образует одноили двухслойную структуру с довольно редкими поперечными связями между цепями (слайд
3.12).
Наружная мембрана состоит из фосфолипидов, типичных для элементарных мембран; белков; липопротеина и липополисахарида. Специфическим компонентом наружной мембраны является липополисахарид (ЛПС), занимающий около 30-40% ее поверхности и
Микробиология с основами вирусологии. Конспект лекций | -21- |

ЛЕКЦИЯ №3. СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ ПРОКАРИОТОВ
3.2. Поверхностные структуры клетки
локализованный во внешнем слое. ЛПС содержат три участка: липид А, сердцевинную часть и О-специфическуюполисахаридную цепь. ЛПС являются антигенами бактерий (слайд 3.13).
Белки-пориныпронизывают наружную мембрану насквозь и формируют гидрофильные поры, через которые осуществляется неспецифическая диффузия молекул. Минорные белки наружной мембраны представлены большим числом видов. Их основная функция – транспортная и рецепторная.
Необычные клеточные стенки прокариот. Некоторые скользящие бактерии способны в процессе перемещения по твердому субстрату менять форму клеток, что говорит об эластичности их клеточной стенки, и в первую очередь ее пептидогликанового слоя. Наиболее вероятное объяснение гибкости клеточной стенки этих бактерий – чрезвычайно слабая сшитость ее пептидогликанового компонента (слайд 3.14).
Клеточная стенка архебактерий по структуре и химическому составу резко отличается от описанных выше типов. Клеточные стенки метанобразующих архебактерий содержат пептидогликан особого химического строения. У других представителей этой группы клеточная стенка может состоять из кислого гетерополисахарида или только из белка. Архебактерии с клеточной стенкой белковой природы не окрашиваются по Граму, остальные типы архебактериальной клеточной стенки дают грамположительную реакцию.
Прокариоты без клеточной стенки. Протопласты – клетки, лишенные клеточной стенки (слайд 3.15). Получают их из грамположительных бактерий, с помощью литических ферментов: лизоцима, эндопептидаз, амидаз, гликозидаз и др.
Независимо от формы исходных клеток бактерий протопласты всегда приобретают сферическую форму. В протопластах осуществляются основные процессы жизнедеятельности: дыхание, синтез белков, нуклеиновых кислот, спорообразование. Они могут увеличиваться в размерах, фиксировать азот (у азотфиксирующих бактерий). Протопласты не способны ресинтезировать клеточную стенку, редко делятся, не адсорбируют фаги, так как рецепторы фагов локализованы в клеточной стенке. При некоторых условиях (например, в 30 %-мжелатине) в протопластах можно индуцировать регенерацию клеточных стенок и они реверсируют в исходную форму, но это происходит чрезвычайно редко. Протопласты используют в функциональной анатомии бактерий, для выделения и изучения мембранных структур, в генетике бактерий.
Сферопласты – бактериальные клетки, частично лишенные клеточной стенки (слайд 3.16). Их обнаруживают в старых культурах, в условиях несбалансированного роста, под влиянием иммунных сывороток и др. Их легче всего получать под влиянием пенициллина в гипертоническом растворе сахарозы или NaCI (осмотические стабилизаторы). Пенициллин предотвращает образование пептидогликана у растущих клеток, нарушая
Микробиология с основами вирусологии. Конспект лекций | -22- |

ЛЕКЦИЯ №3. СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ ПРОКАРИОТОВ
3.2. Поверхностные структуры клетки
процесс образования поперечных связей между пептидными цепочками муреина.
Сферопласты отличаются от протопластов тем, что адсорбируют фаги, так как частично сохраняют клеточную стенку, размножаются, легко реверсируют в исходную клеточную форму при устранении факторов, вызвавших их образование. Общими свойствами протопластов и сферопластов являются большие размеры, отсутствие клеточных мембран типа мезосом, чрезвычайная чувствительность к осмотическим условиям.
L-формыбактерий – образуются при антибиотикотерапии в условиях нарушения биосинтеза пептидогликана и полностью или частично лишены его (слайд 3.17). УL-формбактерий нарушается функция размножения при сохранении функции роста, в результате чего значительно увеличиваются размеры клеток, которые превращаются в гигантские (до 50 мкм) шаровидные, нитевидные, грушевидные сильно вакуолизированные формы.L-формыобладают метаболической активностью, способностью к делению и слиянию их элементов.L-формымедленно(1-4и более недель) растут в виде характерных колоний с врастающим в среду слегка пигментированным центром и нежным кружевным краем (яичница).
L-формыболезнетворных бактерий – патогенные. Они сохраняют способность продуцировать токсины и другие вещества, синтез которых осуществляется в цитоплазме либо в цитоплазматической мембране. Заболевания, обусловленные реверсиейL-форм,характеризуются длительностью течения, меньшей смертностью, большей инвалидностью. L- формы имеют приспособительное значение для клетки как способ переживания бактериями неблагоприятных условий (слайд 3.18).
Функции клеточной стенки прокариот (слайд 3.19):
1.Поддержание внешней формы клетки.
2.Защита от воздействий окружающей среды.
3.Защита от внутреннего осмотического давления.
4.Транспорт веществ и ионов, необходимых клетке.
5.Препятствует проникновению токсических веществ и антибиотиков.
6.Изолирует содержимое клетки от гидролитических ферментов,
7.Содержит транспортные белки и гидролитические ферменты.
8.Содержит специфические рецепторы и антигены.
9.Обеспечивает межклеточные взаимодействия при конъюгации, а также между патогенными бактериями и тканями высших организмов.
3.2.2. Капсулы, слизистыеслоиичехлы
Снаружи клеточная стенка прокариот часто бывает окружена слизистым веществом. Такие образования в зависимости от структурных особенностей получили название капсул, слизистых слоев или чехлов. Все они являются результатом биосинтеза клеткой органических полимеров
(слайд 3.20).
Микробиология с основами вирусологии. Конспект лекций | -23- |

ЛЕКЦИЯ №3. СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ ПРОКАРИОТОВ
3.2. Поверхностные структуры клетки
Капсула – это слизистое образование, обволакивающее клетку, сохраняющее связь с клеточной стенкой и имеющее аморфное строение. Если толщина образования меньше 0,2 мкм – микрокапсула, если больше 0,2 мкм – макрокапсула. Макрокапсулу можно видеть в обычный световой микроскоп при контрастном окрашивании. Наличие капсулы зависит от штамма микроорганизма и условий его культивирования. Бактерии, образующие капсулу, могут легко в результате мутации превращаться в бескапсульные формы.
Чехлы имеют несколько слоев с разным строением. Чехлы ряда бактерий, метаболизм которых связан с окислением восстановленных соединений металлов, часто инкрустированы их окислами (слайд 3.21).
Основные химические компоненты большинства капсул прокариот – гомоили гетерополисахариды. Чехлы как более сложные структуры имеют обычно и более сложный химический состав.
Функции: защищают клетку от механических повреждений, высыхания, создают дополнительный осмотический барьер, служат препятствием для проникновения фагов. Иногда могут служить источником запасных питательных веществ. С помощью слизи осуществляется связь между соседними клетками в колонии, а также прикрепление клеток к различным поверхностям. Способность определенных бактерий синтезировать эти своеобразные внеклеточные полимеры находит практическое применение: их используют в качестве заменителя плазмы крови, а также для получения синтетических пленок (слайд 3.22).
3.2.3. Жгутикиимеханизмыдвижения
Жгутики – это поверхностные структуры прокариот, определяющие способность клетки к движению в жидкой среде. Их число, размеры, расположение являются признаками, постоянными для определенного вида
(слайд 3.23).
Если жгутики находятся у полюсов или в полярной области клетки, говорят об их полярном или субполярном расположении, если – вдоль боковой поверхности, говорят о латеральном расположении. В зависимости от числа жгутиков и их локализации на поверхности клетки различают: монополярные монотрихи, монополярные политрихи или лофотрихи, биполярные политрихи или амфитрихи и перитрихи (слайд 3.24).
Обычная толщина жгутика – 10-20нм, длина – от 3 до 15 мкм. Вращение жгутика осуществляется против часовой стрелки, с частотой от 40 до 60 оборотов в секунду. Клетка вращается в противоположном направлении со значительно меньшей скоростью – порядка12–14об/мин. Скорость поступательного движения клетки для разных видов бактерий составляет от 16 до 100 мкм/с. Движение жгутика обеспечивается энергией трансмембранного электрохимического потенциала.
Жгутик состоит из трех частей: спиральной нити, «крюка» вблизи поверхности клетки и базального тельца. Нити жгутиков состоят из
Микробиология с основами вирусологии. Конспект лекций | -24- |

ЛЕКЦИЯ №3. СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ ПРОКАРИОТОВ
3.2. Поверхностные структуры клетки
специфического белка флагеллина. Крюк состоит из белка, отличающегося от флагеллина, и служит для обеспечения гибкого соединения нити с базальным телом. Базальное тело содержит 9–12различных белков (слайд
3.25).
Базальное тельце закрепляет жгутик в плазматической мембране и клеточной стенке. Оно состоит из центрального стержня и колец. Кольца Р и L локализованы в пептидогликановом слое и в наружной мембране. M- кольцо локализовано в ЦПМ, S-кольцорасполагается в периплазматическом пространстве грамотрицательных или в пептидогликановом мешке грамположительных бактерий. Предполагают, что вращение жгутика определяется вращениемM-кольца.Другие кольца базального тела неподвижны и служат для крепления стержня.
О- и Н-антигены.Клетки, обладающие большой подвижностью, распространяется по всей поверхности агара в виде тонкого серого налета (Н- форма,Hauch-налет).Некоторые штаммы налета не образуют(O-форма,ohneHauch-безналета). Эти штаммы неподвижны, они лишены жгутиков. Отсюда ведет начало терминология, принятая в бактериальной серодиагностике: антигены поверхности или тела клетки (соматические) называют O- антигенами, а антигены жгутиков –Н-антигенами.
Движение спирохет осуществляется за счет вращения аксиальных фибрилл в периплазматическом пространстве между пептидогликановым слоем и наружной мембраной клеточной стенки (слайд 3.26). Количество фибрилл колеблется от 2 до 100. Один конец каждой фибриллы прикреплен вблизи полюса клетки, другой – свободный (слайд 3.27). По химическому составу аксиальные фибриллы близки с бактериальными жгутиками.
Скользящее движение. Способность к скольжению обнаружена у разных групп прокариот, как одноклеточных, так и многоклеточных (нитчатых). Скорость этого типа движения невелика: 2-11мкм/с. Общим для всех скользящих организмов является способность к выделению слизи. Кроме того, у ряда скользящих форм в составе клеточной стенки между пептидогликановым слоем и наружной мембраной обнаружен тонкий слой, состоящий из белковых фибрилл. Направление вращения является видоспецифическим признаком и зависит от направления движения белковых фибрилл.
Таксисы бактерий (слайд 3.28). Подвижные бактерии активно перемещаются в направлении, определяемом внешними факторами. Хемотаксис – движение относительно источника химического вещества. Для каждого организма все химические вещества в этом плане могут быть разделены на две группы: инертные и вызывающие таксисы (эффекторы). Среди эффекторов выделяют аттрактанты (вещества, привлекающие бактерий) и репелленты (вещества, отпугивающие бактерий). Аттрактантами могут быть сахара, аминокислоты, витамины, нуклеотиды; репеллентами – аминокислоты, спирты, фенолы, неорганические ионы и другие. Частным случаем хемотаксиса является аэротаксис (слайд 3.29).
Микробиология с основами вирусологии. Конспект лекций | -25- |
studfiles.net
Строение прокариотной клетки
СТРОЕНИЕ ПРОКАРИОТНОЙ КЛЕТКИ
В 70-х гг. обнаружены микроорганизмы, структурно относящиеся к прокариотному типу, но значительно отличающиеся химическим строением важных клеточных макромолекул и способностью осуществлять уникальные биохимические процессы. Эти необычные прокариотные организмы были названы архебактериями. Типичные прокариоты, или бактерии, получили соответственно название эубактерий (истинных бактерий). Число известных архебактерий по сравнению с эубактериями чрезвычайно мало.
Материал настоящего раздела посвящен общей характеристике прокариотных организмов (в основном эубактерий), отличающихся морфологическим и особенно физиологическим разнообразием. В основе морфологического разнообразия лежат различия в размерах и форме отдельных клеток, способах их деления, природе и наборе цитоплазматических включений, строении клеточной стенки и структур, локализованных снаружи от нее, наличии и типе дифференцированных форм, образующихся в процессе жизненного цикла.
ФОРМА ПРОКАРИОТ
До недавнего времени большинство исследователей традиционно считали, что клетки прокариот достаточно однообразны и в подавляющем большинстве имеют форму сферы, цилиндра или спирали. Они бывают одиночными, в иных случаях образуют нити или колонии. Прокариоты сферической формы, называемые кокками, могут после деления не расходиться. Если деление происходит в одной плоскости, образуются пары клеток (диплококки) или цепочки (стрептококки). В том случае, когда деление происходит относительно равномерно в трех взаимно перпендикулярных направлениях и клетки после деления остаются соединенными друг с другом, возникают пакеты правильной формы (сарцины) или колонии сферической формы. Если же деление происходит в нескольких плоскостях неравномерно, образуются клеточные скопления неправильной формы (слайд 2, 1—5). Прокариоты, имеющие форму цилиндра (палочковидные), сильно различаются по величине отношения длины клетки к ее поперечнику. Прокариоты спиралевидной формы характеризуются разным числом витков: у спирилл — от одного до нескольких витков, вибрионы выглядят наподобие изогнутых палочек, так что их можно рассматривать как неполный виток спирали (слайд 2, 6—8).
За последнее время среди прокариот обнаружены организмы, отличающиеся от описанных выше основных форм. Некоторые бактерии имеют вид кольца, замкнутого или разомкнутого в зависимости от стадии роста (слайд 2, 9). У прокариот, в основном размножающихся почкованием, описано образование клеточных выростов (простек), число которых может колебаться от 1 до 8 и более (слайд 2, 10). Из природных субстратов выделены бактерии червеобразной формы и напоминающие шестиугольную звезду (слайд 2, 11, 12). Для некоторых видов характерно слабое или довольно хорошо выраженное ветвление (слайд 2, 13). Описаны прокариоты, обладающие морфологической изменчивостью, в зависимости от условий имеющие вид палочек, кокков или обнаруживающие слабое ветвление.
Форма многоклеточных прокариот также разнообразна: это скопления различной конфигурации, чаще — нити (слайд 2, 14—16). Своеобразие бактериальным клеткам придают жгутики, имеющие различное расположение на клеточной поверхности (слайд 2, 8, 15, 17, 18). а также выделения внеклеточных веществ разной химической природы (слайд 2, 19—22).
СТРУКТУРА, ХИМИЧЕСКИЙ СОСТАВ И ФУНКЦИИ КОМПОНЕНТОВ ПРОКАРИОТНОЙ КЛЕТКИ
Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации (слайд 3). Структуры, расположенные снаружи от ЦПМ (клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки), называют обычно поверхностными структурами. Термином «клеточная оболочка» часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). ЦПМ вместе с цитоплазмой называется протопластом. Рассмотрим сначала строение, химический состав и функции поверхностных клеточных структур.
Клеточная стенка
Клеточная стенка — важный и обязательный структурный элемент подавляющего большинства прокариотных клеток, располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с окружающей средой (у клеток, не содержащих этих слоев клеточной оболочки). На долю клеточной стенки приходится от 5 до 50% сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. Концентрация солей в клетке, как правило, намного выше, чем в окружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка чисто механически защищает клетку от проникновения в нее избытка воды.
По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты, относящиеся к эубактериям, делятся на две большие группы. Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна: у так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются6. У некоторых эубактерий положительная реакция при окрашивании описанным выше способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки.
Клеточные стенки грамположительных и грамотрицательных эубактерий резко различаются как по химическому составу (табл. 1, слайд 5), так и по ультраструктуре (слайд 4).
Под электронным микроскопом клеточная стенка грамположительных эубактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм. У грамотрицательных эубактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2—3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8—10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных эубактерий получил название наружной мембраны.
Клеточная стенка грамположительных эубактерий. Основную массу клеточной стенки грамположительных эубактерий составляет специфический гетерополимер — пептидогликан. Полисахаридный остов молекулы построен из чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных между собой посредством) b-1,4-гликозидных связей. К N-ацетилмурамовой кислоте присоединен короткий пептидный хвост, состоящий из небольшого числа (обычно 4—5) аминокислот. У грамположительных эубактерий обнаружено более 100 различных химических типов пептидогликана. Большинство различий относится к пептидной части его молекулы.
Две особенности пептидного хвоста заслуживают внимания: наличие аминокислот в D-форме (неприродная конфигурация) и высокое содержание аминокислот с двумя аминогруппами. Это имеет принципиальное значение для пространственной организации пептидогликана. Обе аминогруппы этих аминокислот могут участвовать в образовании пептидных связей, причем вторые аминогруппы — в формировании дополнительных пептидных связей между гетерополимерными цепочками. В большинстве случаев в образовании пептидной связи участвует карбоксильная группа D-аланина одного тетрапептида и свободнаяаминогруппа диаминокислоты другого. Иногда связь между тетрапептидами разных гликановых цепей осуществляется с помощью других аминокислот. Нетрудно себе представить, что этим способом можно «сшить» между собой множество гетерополимерных цепей. Частота «сшивок» различна, поскольку не все пептидные хвосты участвуют в формировании межцепочечных связей. Некоторые образуют ковалентные связи с другими химическими молекулами, входящими в состав клеточной стенки, и, наконец, часть тетрапептидных хвостов находится в свободном состоянии.
Пептидогликан, окружающий протопласт грамположительных эуКроме пептидогликана в состав клеточных стенок грамположительных эубактерий входит другой уникальный класс химических соединений — тейхоевые кислоты, представляющие собой полимеры, построенные на основе рибита (пятиатомного спирта) или глицерина (трехатомного спирта), остатки которых соединены между собой фосфодиэфирными связями.
В составе клеточной стенки грамположительных эубактерий в небольших количествах также найдены полисахариды, белки и липиды. Для полисахаридов и липидов показана возможность ковалентного связывания с макромолекулами клеточной стенки, в отличие от белков, которые (у тех видов, где имеются) формируют на ее внешней поверхности отдельный слой.
Таким образом, основными компонентами клеточной стенки грамположительных эубактерий являются три типа макромолекул: пептидогликаны, тейхоевые кислоты и полисахариды, которые с помощью ковалентных связей образуют сложную структуру с весьма упорядоченной пространственной организацией.
Клеточная стенка бацилл, например Bacillus subtilis, приблизительно соответствует толщине 40 молекул пептидогликана. В целом клеточную стенку грамположительных эубактерий можно представить в виде губчатой структуры с порами диаметром примерно 1—6 нм. Возможность прохождения молекул через такую клеточную стенку определяется ее зарядом и размером пор.
Клеточная стенка грамотрицательных эубактерий. У грамотрицательных эубактерий строение клеточной стенки намного сложнее, чем у грамположительных (см. слайд4). В ее состав входит гораздо большее число макромолекул разного химического типа (см.табл. 1). Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов грамотрицательных эубактерий содержание этого гетерополимера колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями.
Химическая структура пептидогликана грамотрицательных эубактерий в основном сходна со структурой типичного пептидогликана грамположительных эубактерий. Снаружи от пептидогликана располагается дополнительный слой клеточной стенки — наружная мембрана. Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида (слайд6, А). Специфическим компонентом наружной мембраны является липополисахарид сложного молекулярного строения, занимающий около 30—40% ее поверхности и локализованный во внешнем слое (слайд6, 5).
Помимо слоев клеточной стенки, типичных для большинства грамотрицательных эубактерий, у некоторых представителей этой группы обнаружены дополнительные слои разной электронной плотности, располагающиеся с внешней стороны от наружной клеточной мембраны. Однако до настоящего времени не ясно, относятся ли они к клеточной стенке, являясь результатом ее последующего усложнения, или же представляют собой структурные элементы многослойного чехла.
Необычные клеточные стенки прокариот. Некоторые скользящие бактерии (миксобактерии, флексибактерии) способны в процессе перемещения по твердому субстрату периодически менять форму клеток, например путем изгибания, что говорит об эластичности их клеточной стенки, и в первую очередь ее пептидогликанового слоя. Электронно-микроскопическое изучение, однако, обнаружило у них клеточную стенку, типичную для грамотрицательных эубактерий. Наиболее вероятное объяснение гибкости клеточной стенки этих бактерий — чрезвычайно низкая сшитость ее пептидогликанового компонента.
Наконец, обнаружены прокариоты, клеточная стенка которых по структуре и химическому составу резко отличается от описанных выше типов. Они принадлежат к группе архебактерий. Клеточные стенки метанобразующих архебактерий содержат пептидогликан особого химического строения. У других представителей этой группы клеточная стенка состоит исключительно из кислого гетерополисахарида, а у некоторых экстремально галофильных, метанобразующих и ацидотермофильных архебактерий — только из белка. Архебактерии с клеточной стенкой белковой природы не окрашиваются по Граму, остальные типы архебактериальной клеточной стенки дают грамположительную реакцию.
Капсулы, слизистые слои и чехлы
Снаружи клеточная стенка прокариот часто бывает окружена слизистым веществом. Такие образования в зависимости от структурных особенностей получили название капсул, слизистых слоев или чехлов. Все они являются результатом биосинтеза прокариотами органических полимеров и отложения их вокруг клеток.
Под капсулой понимают слизистое образование, обволакивающее клетку, сохраняющее связь с клеточной стенкой и имеющее аморфное строение (см. слайд2, 19,4, 2). Если толщина образования меньше 0,2 мкм и, следовательно, оно может быть обнаружено только с помощью электронного микроскопа, говорят о микрокапсуле. Если больше 0,2 мкм, говорят о макрокапсуле. Последнюю можно видеть в обычный световой микроскоп. Для этого препарат просматривают в капле туши, которая не в состоянии проникнуть в капсулу. На темном фоне выделяются клетки, окруженные светлыми зонами. Если же слизистое вещество имеет аморфный, бесструктурный вид и легко отделяется от поверхности прокариотной клетки, говорят о слизистых слоях, окружающих клетку (см.слайд3, 3).
В отличие от капсул чехлы имеют тонкую структуру. Нередко в них обнаруживают несколько слоев с разным строением (см. слайд3, 4). Чехлы ряда бактерий, метаболизм которых связан с окислением восстановленных соединений металлов, часто инкрустированы их окислами. Между этими структурами у прокариот обнаружено много переходных форм, так что иногда нельзя четко отграничивать капсулу от слизистых клеточных выделений или капсулу от чехла.
Наличие капсулы зависит от штамма микроорганизма и условий его культивирования. Бактерии, образующие капсулу, могут легко в результате мутации превращаться в бескапсульные формы, что не приводит к какому-либо нарушению клеточной активности, поэтому капсулы нельзя рассматривать как обязательный структурный компонент прокариотной клетки.
Жгутики и механизмы движения
На клеточной поверхности многих прокариот имеются структуры, определяющие способность клетки к движению в жидкой среде. Это — жгутики. Их число, размеры, расположение, как правило, являются признаками, постоянными для определенного вида, и поэтому учитываются при систематике прокариот. Однако накапливаются данные о том, что количество и расположение жгутиков у одного и того же вида могут в значительной степени определяться условиями культивирования и стадией жизненного цикла, и, следовательно, не стоит переоценивать таксономическое значение этого признака.
Если жгутики находятся у полюсов или в полярной области клетки, говорят об их полярном или субполярном расположении, если — вдоль боковой поверхности, говорят о латеральном расположении. В зависимости от числа жгутиков и их локализации на поверхности клетки различают монополярные монотрихи (один жгутик прикреплен к одному полюсу клетки; см. слайд2, 8), монополярные политрихи (пучок жгутиков расположен на одном полюсе клетки), биполярные политрихи (на каждом полюсе — по пучку жгутиков; см.слайд2, 17) и перитрихи (многочисленные жгутики расположены по всей поверхности клетки или вдоль ее боковой поверхности; см.слайд2, 18). В последнем случае число жгутиков может достигать 1000 на клетку.
Обычная толщина жгутика — 10—20 нм, длина — от 3 до 15 мкм. У некоторых бактерий длина жгутика может на порядок превышать диаметр клетки. Как правило, полярные жгутики более толстые, чем перитрихиальные. Жгутик представляет собой относительную жесткую спираль, обычно закрученную против часовой стрелки. Вращение жгутика также осуществляется против часовой стрелки с частотой от 40 до 60 об/с, что вызывает вращение клетки, но в противоположном направлении. Поскольку клетка намного массивнее жгутика, она вращается со значительно меньшей скоростью — порядка 12—14 об/мин. Вращательное движение жгутика преобразуется также в поступательное движение клетки, скорость которого в жидкой среде для разных видов бактерий составляет от 16 до 100 мкм/с.
Изучение строения жгутика под электронным микроскопом обнаружило, что он состоит из трех частей (слайд7). Основную массу жгутика составляет длинная спиральная нить (фибрилла), у поверхности клеточной стенки переходящая в утолщенную изогнутую структуру—крюк. Нить с помощью крюка прикреплена к базальному телу, вмонтированному в ЦПМ и клеточную стенку. У большинства прокариот нить состоит только из одного типа белка — флагеллина. Белковые субъединицы уложены в виде спирали, внутри которой проходит полый канал. Наращивание жгутика происходит с дистального конца, куда субъединицы поступают по внутреннему каналу. У некоторых видов жгутик снаружи дополнительно покрыт чехлом особого химического строения или же являющимся продолжением клеточной стенки и, вероятно, построенным из того же материала.
Крюк (толщина 20—45 нм) состоит из белка, отличающегося от флагеллина, и служит для обеспечения гибкого соединения нити с базальным телом. Базальное тело содержит 9—12 различных белков и представляет собой систему из двух или четырех колец, нанизанных на стержень, являющийся продолжением крюка. Два внутренних кольца (M и S) — обязательные составные части базального тела, в то время как наружные кольца (Р и L) отсутствуют у грамположительных эубактерий и, следовательно, не необходимы для движения. M-кольцо локализовано в ЦПМ, S-кольцо располагается в периплазматическом пространстве грамотрицательных или в пептидогликановом мешке грамположительных эубактерий.
Большие успехи достигнуты в расшифровке механизма движения прокариот, имеющих жгутики. Если в клетке много жгутиков. все они при движении собираются в пучок, вращаясь в одном направлении (слайд8). Вращение жгутиков передается клетке, начинающей вращаться в противоположном направлении, и обеспечивает эффективное движение (плавание) в жидкой среде и более медленное перемещение по поверхности твердых сред.
Необычная локализация структур, ответственных за движение, описана у спирохет (слайд9). Трехслойная структура, окружающая клетку и называемая у спирохет наружным чехлом, аналогична наружной мембране клеточной стенки грамотрицательных эубактерий. Этот чехол окружает так называемый протоплазматический цилиндр, состоящий из пептидогликанового слоя клеточной стенки, ЦПМ и цитоплазматического содержимого. Протоплазматический цилиндр обвивается пучком нитчатых структур — аксиальных фибрилл. Число их колеблется от 2 до 100. Один конец каждой аксиальной фибриллы прикреплен вблизи полюса протоплазматического цилиндра, другой— свободный. Клетка содержит по два набора фибрилл, прикрепленных субполярно у каждого клеточного конца. Так как каждая аксиальная фибрилла тянется почти вдоль всей длины клетки, пучки фибрилл, прикрепленных у разных полюсов, в центральной части перекрываются.
Механизм скользящего движения не ясен. Согласно гипотезе реактивного движения оно обусловлено выделением слизи через многочисленные слизевые поры в клеточной стенке, в результате чего клетка отталкивается от субстрата в направлении, противоположном направлению выделения слизи. Однако анализ этой модели привел к заключению, что для обеспечения скольжения по «реактивному» механизму клетке необходимо в течение 1 с выделять такой объем слизи, который во много раз превосходит ее цитоплазматическое содержимое.
По другой гипотезе, получившей распространение в последние годы, скользящее движение связано с особенностями строения клеточной стенки подвижных безжгутиковых форм — наличием белкового слоя, состоящего из упорядоченно расположенных фибрилл, аналогичных нитям жгутиков, с той разницей, что находятся фибриллы «внутри» клеточной стенки. У некоторых скользящих бактерий описаны структуры, весьма напоминающие базальные тела жгутиковых форм. Вращательное движение фибрилл, «запускаемое» этими структурами, приводит к появлению на поверхности клетки так называемой «бегущей волны», т. е. движущихся микроскопических выпуклостей клеточной стенки, в результате чего клетка отталкивается от твердого или вязкого субстрата. На скольжение расходуется около 5% энергии от общего объема клеточных энергетических затрат. Скользящее движение в разных группах бактерий обеспечивается энергией в форме АТФ или DmH+.
Необходимость для скольжения слизи пока не ясна. Скольжение может происходить в среде подходящей консистенции без какого-либо выделения слизи. Более того, выделение больших количеств слизи, как правило, затрудняет движение клетки и приводит к потере ею подвижности. Согласно гипотезе «бегущей волны» выделение слизи не является абсолютно необходимым для скольжения, но облегчает в определенных условиях отталкивание клетки от субстрата.
Фототаксис, т. е. движение к свету или от него, свойствен прежде всего фототрофным бактериям. Способность перемещаться по силовым линиям магнитного поля Земли или магнита — магнитотаксис — обнаружен у разных бактерий, обитающих в пресной и морской воде. В клетках этих бактерий найдены непрозрачные частицы определенной геометрической формы — магнитосомы, заполненные железом в форме магнетита (Fe3O4) и выполняющие функцию магнитной стрелки. На долю магнетита может приходиться до 4% сухого вещества бактерий. В северном полушарии такие магниточувствительные бактерии плывут в направлении северного полюса Земли, в южном — в направлении южного. У ряда бактерий обнаружен вискозитаксис — способность реагировать на изменение вязкости раствора и перемещаться в направлении ее увеличения или уменьшения.
За чувствительность бактерий к градиентам определенных факторов ответственны специфические рецепторы. Изучение хемотаксиса у Escherichia coli позволило обнаружить свыше 30 различных хеморецепторов, представляющих собой белки, синтезируемые независимо от присутствия индуктора или только в результате индукции. Рецептор реагирует на эффектор и передает сигнал по определенному пути, конкретный механизм которого неизвестен, на «мотор» жгутика. У бактерий с перитрихиальным жгутикованием выявлены два вида двигательного поведения: прямолинейное движение и кувырканье, т. е. периодические и случайные изменения направления движения. Если бактерия перемещается в сторону оптимальной концентрации аттрактанта, ее прямолинейное движение, ориентированное по отношению к химическому веществу, становится более длительным, а частота кувырканий более низкой, что позволяет ей в конечном итоге перемещаться в нужном направлении.
Ворсинки
К поверхностным структурам бактериальной клетки относятся также ворсинки (фимбрии, пили) (см. слайд3, 6). Их насчитывается от нескольких единиц до нескольких тысяч на клетку. Эти структуры не имеют отношения к движению бактерий и обнаружены у подвижных и неподвижных форм. Ворсинки построены из одного вида белка — пилина — и представляют собой прямые белковые цилиндры, отходящие от поверхности клетки. Они, как правило, тоньше жгутиков (диаметр — 5—10 нм, длина 0,2–2,0 мкм), расположены перитрихиально или полярно. Больше всего сведений имеется о ворсинках Е. coli. У этой бактерии описаны ворсинки общего типа и половые.
Ворсинки общего типа придают бактериям свойство гидрофобности, обеспечивают их прикрепление к клеткам растений, грибов и неорганическим частицам, принимают участие в транспорте метаболитов. Через ворсинки в клетку могут проникать вирусы.
Наиболее хорошо изучены половые ворсинки, или F-пили, принимающие участие в половом процессе бактерий. F-пили необходимы клетке-донору для обеспечения контакта между ней и реципиентом и в качестве конъюгационного тоннеля, по которому происходит передача ДНК. Ворсинки нельзя считать обязательной клеточной структурой, так как и без них бактерии хорошо растут и размножаются.
Мембраны
Содержимое клетки отделяется от клеточной стенки цитоплазматической мембраной (ЦПМ) — обязательным структурным элементом любой клетки, нарушение целостности которого приводит к потере клеткой жизнеспособности. На долю ЦПМ приходится 8—15% сухого вещества клеток. У большинства прокариотных клеток ЦПМ — единственная мембрана. В клетках фототрофных и ряда хемотрофных прокариот содержатся также мембранные структуры, располагающиеся в цитоплазме и получившие название внутрицитоплазматических мембран. Их происхождение и функции будут рассмотрены ниже.
Химический состав мембран. ЦПМ — белково-липидный комплекс, в котором белки составляют 50–75%, липиды — от 15 до 45%. Кроме того, в составе мембран обнаружено небольшое количество углеводов. Как правило, липиды и белки составляют 95% и больше вещества мембран. Главным липидным компонентом бактериальных мембран являются фосфолипиды — производные 3-фосфоглицерина. Хотя у прокариот найдено множество различных фосфолипидов, набор их в значительной степени родо- и даже видоспецифичен. Широко представлены в бактериальных мембранах различные гликолипиды. Стерины отсутствуют у подавляющего большинства прокариот, за исключением представителей группы микоплазм и некоторых бактерий. Так, в ЦПМ Acholeplasma содержится 10–30% холестерина, поглощаемого из внешней среды, от общего содержания мембранных липидов. Из других групп липидов в мембранах прокариот обнаружены каротиноиды, хиноны, углеводороды.
Все липиды эубактерий — производные глицерина — содержат один или несколько остатков жирных кислот, состав которых весьма своеобразен (слайд10). В основном это насыщенные или мононенасыщенные жирные кислоты с 16—18 углеродными атомами. Полиненасыщенные жирные кислоты у эубактерий отсутствуют. Исключение составляют цианобактерии, у разных видов которых найдены полиненасыщенные жирные кислоты типа C16:2, C18:2, C18:3, C15:4. Помимо обычных жирных кислот, т. е. обнаруживаемых и в клетках эукариот, в составе мембранных липидов эубактерий находят и кислоты, не встречающиеся, как правило, в мембранах эукариот. Это циклопропановые жирные кислоты, содержащие одно или больше трехчленных колец, присоединенных вдоль углеводородной цепи. Другие, редко встречающиеся и обнаруженные практически только у эубактерий кислоты — это разветвленные жирные кислоты с 15—17 углеродными атомами.
Структура мембран. Мембранные липиды всех эубактерий и части архебактерий образуют бислои, в которых гидрофильные «головы» молекул обращены наружу, а гидрофобные «хвосты» погружены в толщу мембраны (слайд11). Углеводородные цепи, прилегающие к гидрофильным «головам», довольно жестко фиксированы, а более удаленные части «хвостов» обладают достаточной гибкостью. У некоторых архебактерий (ряд метаногенов, термоацидофилы) мембранные липиды, в состав которых входит C40-спирт, формируют монослойную мембрану, по толщине равную бислойной. Монослойные липидные мембраны обладают большей жесткостью сравнительно с бислойной. При «биологических» температурах мембранные липиды находятся в жидкостно-кристаллическом состоянии, характеризующемся частичной упорядоченностью структуры. При понижении температуры они переходят в квазикристаллическое состояние. Чем более ненасыщены и разветвлены остатки жирных кислот или чем большее число циклических группировок они содержат, тем ниже температура перехода из жидкостно-кристаллического состояния в квазикристаллическое.
studfiles.net
Биология для студентов — 06. Строение и химический состав прокариотической клетки
Когда изучали бактерий, оказалось, что они также делятся на две сильно отличающиеся группы. Соответственно, их пришлось разделить на два царства:
- Эубактерии (собственно бактерии),
- Архебактерии (другое название – Археи).
Последние также не имеют ядра, но по структуре сильно отличаются от бактерий. Такое деление возникло недавно. В 1990 году вышла посвященная этой теме публикация. Деление было сделано на основе последовательности рибосомной РНК.
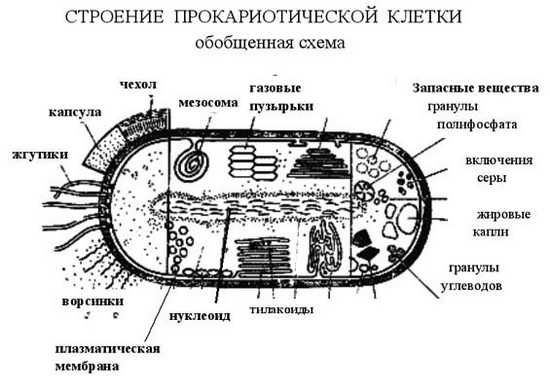
У прокариотических клеток есть цитоплазматическая мембрана, также, как и эукариотических. У бактерий мембрана двуслойная (липидный бислой), у архей мембрана довольно часто бывает однослойной. Мембрана архей состоит из веществ, отличных от тех, из которых состоит мембрана бактерий. Поверхность клеток может быть покрыта капсулой, чехлом или слизью. У них могут быть жгутики и ворсинки.
Клеточное ядро, такое как у эукариот, у прокариот отсутствует. ДНК находится внутри клетки, упорядоченно свернутая и поддерживаемая белками. Этот ДНК-белковый комплекс называется нуклеоид. У эубактерий белки, которые поддерживают, ДНК отличаются от гистонов, которые образуют нуклеосомы (у эукариот). А у архибактерий гистоны есть, и этим они похожи на эукариот. Энергетические процессы у прокариотов идут в цитоплазме и на специальных структурах — мезосомах (выростах клеточной мембраны, которые закручены в спираль для увеличения площади поверхности, на которой происходит синтез АТФ). Внутри клетки могут находиться газовые пузырьки, запасные вещества в виде гранул полифосфатов, гранул углеводов, жировых капель. Могут присутствовать включения серы (образующейся, например, в результате бескислородного фотосинтеза). У фотосинтетических бактерий имеются складчатые структуры, называемые тилакоидами, на которых идет фотосинтез. Таким образом, у прокариот, в принципе, имеются те же самые элементы, но без перегородок, без внутренних мембран. Те перегородки, которые имеются, являются выростами клеточной мембраны.
Самая маленькая бактерия – это паразитическая микоплазма (она живет внутри клеток эукариот). Она имеет размер 0,1 мкм. Самые большие представители прокариот видны невооруженным глазом (граница видимости – 70-80 мкм). Эта спирохета имеет длину 250 мкм. Типичный же представитель прокариот имеет размер 0,5 мкм в ширину и 2 мкм в ширину. Хотя и археи, и бактерии относятся к прокариотическим (безядерным) организмам, строение их клеток имеет некоторые существенные отличия: бактерии имеют липидный бислой (когда гидрофобные концы погружены в мембрану, а заряженные головки торчат с двух сторон наружу), а археи могут иметь монослойную мембрану (заряженные головки имеются с двух сторон, а внутри единая целая молекула; эта структура может быть более жесткой, чем бислой). Бактерии и археи отличаются строением и размером РНК-полимеры. В состав бактериальных РНК-полимераз входит 4-8 белковых субъединиц, в сотав эукариотических РНК-полимераз входит 10-14 белковых субъединиц, а у архей размер промежуточный: 5-11 субъединиц. Рибосомы бактерий меньше рибосом эукариот и меньше, чем рибосомы архей (которые также имеют промежуточные размеры).
Химический состав клеток в принципе одинаков у всех организмов. Клетки прокариот содержат от 70 до 90% воды. Основную массу сухих веществ, на долю которых приходятся остальные 10-30%, составляют белки, нуклеиновые кислоты, липиды и полисахариды. Несколько процентов сухого вещества клеток приходится на низкомолекулярные органические вещества и соли.
Макромолекулы, составляющие основную массу сухих веществ клетки, — полимеры, построенные из мономерных единиц. Исключением служат липиды, не являющиеся полимерами, так как молекулы в них не соединены между собой ковалентными связями. Углеводные полимеры построены на основе повторяющихся единиц одного, двух или более типов, например, запасной полисахарид гликоген, построенный из остатков глюкозы, или пептидогликан клеточной стенки, образованный чередованием N-ацетилглюкозамина и N- ацетилмурамовой кислоты. В клетке углеводные полимеры представлены часто одним видом молекулы.
Полимерные молекулы белков и нуклеиновых кислот синтезируются на матрице, которая и определяет последовательность составляющих их мономеров. Возможности для синтеза разнообразных по функциям и структуре клеточных метаболитов реализуются на стадии сборки полимеров путем различных сочетаний исходных строительных блоков. В основе огромного числа видо-и функционально специфических белков лежат комбинации из 20 аминокислот, а чтобы зашифровать весь объем генетической информации одной клетки или многоклеточного организма оказалось достаточным комбинации из 4 нуклеотидов. Прокариотная клетка в норме содержит примерно 2000-2500 различных белков, каждый из которых представлен 400-1000 молекулами. Количество молекул нуклеиновых кислот каждого вида определяется их функциональным назначением:
- ДНК- одного вида и представлена одной или несколькими копиями;
- количество разных молекул РНКв клетке колеблется на несколько порядков.
vseobiology.ru
Химический состав прокариотной клетки
Химический состав прокариотной клетки
Химический состав клеток в принципе одинаков у всех организмов. Клетки прокариот содержат от 70 до 90% воды. Основную массу сухих веществ, на долю которых приходятся остальные 10-30%, составляют белки , нуклеиновые кислоты , липиды и полисахариды . Несколько процентов сухого вещества клеток приходится на низкомолекулярные органические вещества и соли ( табл. 9 ).
Макромолекулы, составляющие основную массу сухих веществ клетки, — полимеры, построенные из мономерных единиц. Исключением служат липиды, не являющиеся полимерами, так как молекулы в них не соединены между собой ковалентными связями. Углеводные полимеры построены на основе повторяющихся единиц одного, двух или более типов, например, запасной полисахарид гликоген , построенный из остатков глюкозы, или пептидогликан клеточной стенки, образованный чередованием N-ацетилглюкозамина и N- ацетилмурамовой кислоты. В клетке углеводные полимеры представлены часто одним видом молекулы ( табл. 9 ).
Полимерные молекулы белков и нуклеиновых кислот синтезируются на матрице, которая и определяет последовательность составляющих их мономеров. Возможности для синтеза разнообразных по функциям и структуре клеточных метаболитов реализуются на стадии сборки полимеров путем различных сочетаний исходных строительных блоков. В основе огромного числа видо-и функционально специфических белков лежат комбинации из 20 аминокислот , а чтобы зашифровать весь объем генетической информации одной клетки или многоклеточного организма оказалось достаточным комбинации из 4 нуклеотидов . Прокариотная клетка в норме содержит примерно 2000-2500 различных белков, каждый из которых представлен 400-1000 молекулами. Количество молекул нуклеиновых кислот каждого вида определяется их функциональным назначением: ДНК — одного вида и представлена одной или несколькими копиями; количество разных молекул РНК в клетке колеблется на несколько порядков.
Ссылки:
medbiol.ru
Структурно-функциональная организация клеток прокариот.
Оболочка и надоболочечные структуры клеток прокариот
1. Структуры прокариотной клетки.
2. Основные вещества клеточной стенки прокариот.
3. Строение клеточной стенки грациликутных (Г—) и фирмакутных (Г +) бактериях.
4. Отличительные черты строения оболочек микобактерий.
5. Протопласты, сферопласты, L – формы.
6. Надоболочечные структуры клеток: жгутики, пили. Движение бактерий.
Структуры прокариотной клетки
Клетка прокариот имеет все структурные компоненты, присущие любой клетке. Среди структур бактериальной клетки различают:
основные структуры – клеточную стенку, цитоплазматическую мембрану, цитоплазму с различными цитоплазматическими включениями и нуклеоид;
временные структуры – капсулу, жгутики, фимбрии, у некоторых представителей бактерий – эндоспоры.
Клеточная стенка. Клеточная стенка является обязательным структурным элементом бактериальной клетки, исключение составляют микоплазмы и L-формы бактерий. Обладая высокой степенью эластичности и упругости, она выдерживает внутриклеточное давление. Клеточная стенка служит механическим барьером между протоплазмой и внешней средой, придает клеткам определенную форму, определяет способность удерживания или вымывания красителей, дает возможность клетке существовать в гипотонических растворах.

Рис. 1. Схематическое строение прокариотной клетки. А – поверхностные структуры: 1 – клеточная стенка; 2 – капсула, или слизистый слой; 3 – жгутики; 4 – фимбрии; Б – цитоплазматические структуры: 5 – цитоплазматическая мембрана; 6 – цитоплазма; 7 – нуклеоид; 8 – рибосомы; 9 –хроматофоры; 10 – везикулы; 11 – пластинчатые тилакоиды; 12 – трубчатые тилакоиды; 13 – мезосома; 14 – аэросомы; 15 – ламеллярные структуры; 16 – карбоксисомы; В – запасные вещества: 17 – полифосфаты; 18 – полисахариды; 19 – поли--оксимасляная кислота; 20 – включения серы; 21, 22 – жировые капли (Г. Шлегель, 1972)
Поверхностные макромолекулы внешней стороны клеточной стенки выполняют разнообразные функции, такие, как:
– специфическая рецепторная для фагов и колицинов;
– антигенная роль;
– функция межклеточного взаимодействия при коньюгации и при взаимодействии с клетками тканей высших организмов.
Клеточная стенка грамотрицательных (грациликутных) значительно расширяет круг функций, так как осуществляет роль дополнительного клеточного барьера, имеет дополнительные специфические и неспецифические каналы (диффузные поры), препятствует проникновению в клетку токсических веществ.
Без специальных методов окраски рассмотреть клеточную стенку бактерий под световым микроскопом не удается. На долю клеточной стенки приходится от 5 до 50% сухой массы клетки. Толщина данной структуры составляет 10–80 нм. Клеточной стенкой или клеточной оболочкой называют структуры, расположенные над цитоплазматической мембраной. Различают также поверхностные дополнительные структуры (капсулы, жгутики, фимбрии и т.п.).
По химическому составу клеточная стенка прокариот коренным образом отличается от оболочки клеток эукариот. Основным компонентом клеточной стенки бактерий является муреин, относящийся к классу пептидогликанов. Муреин – гетерополимер, образованный чередующимися остатками N-ацетил-N-глюкозамина и N-ацетилмурамовой кислоты, соединенными -1,4-гликозидными связями. N-ацетилмурамовая кислота соединена с пептидом, в состав которого входят 4–6 различных аминокислот. Основу пептидного компонента муреина составляют тетрапептиды, образованные обычно L-аланином, D-глутаминовой кислотой, специфической для прокариот мезодиаминопимелиновой кислотой (мезо- ДАП) и D-аланином. У некоторых бактерий мезодиаминопимелиновая кислота замещена на L-лизин, либо L- или D-орнитин, либо на 2,4-диаминомасляную кислоту.
По содержанию муреина и специфике дополнительных компонентов, включенных в муреиновую сеть, все бактерии подразделяются на две группы: грамположительные (фирмакутные) и грамотрицательные (грациликутные).
В 1884 г. X. Грам предложил метод окраски бактерий, вошедший в практику микробиологии как один из диагностических признаков. Метод основан на различной способности микробов удерживать красители трифенилметанового ряда – кристаллвиолет и генцианвиолет в клетке. При окрашивании фиксированных мазков раствором фиолетового красителя и закреплении его раствором йода в калий-йод, грамположительные бактерии образуют стойкое соединение красителя с йодом и при последующей обработке мазков спиртом или ацетоном не раскрашиваются, сохраняя фиолетовую окраску. Грамотрицательные бактерии не образуют стойкого соединения красителя с йодом, полностью обесцвечиваются спиртом или ацетоном, и мазки их подлежат дополнительной окраске, обычно водным раствором фуксина. Способность микроорганизмов окрашиваться по методу Грама или утрачивать окраску объясняется спецификой химического состава и ультраструктуры их клеточной стенки.
Клеточная стенка грамположительных бактерий достаточно массивна, толщина ее достигает 20–80 нм. Она имеет гомогенную губчатую структуру, пронизанную порами, и плотно прилегает к цитоплазматической мембране. Муреин в клеточной стенке грамположительных бактерий составляет от 50 до 90% ее сухой массы. С муреином связаны тейхоевые кислоты. Они представляют собой полимеры трехатомного спирта глицерина или пятиатомного спирта рибита, остатки которых соединены фосфодиэфирными связями. Одна молекула тейхоевой кислоты обычно включает от 7 до 15 спиртовых остатков. Свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками D-аланина, глюкозы, N-ацетилглюкозамина или N-ацетил-галактозамина. Глицеринтейхоевые кислоты нередко связаны с липидом, находящимся в цитоплазматической мембране. Тейхоевые кислоты оказывают влияние на катионный обмен клетки. У некоторых бактерий они принимают участие в регуляции активности автолитических ферментов – гидролаз, способных разрушать собственную оболочку. В составе клеточной стенки грамположительных бактерий в небольшом количестве обнаружены полисахариды, белки и липиды. Оболочка фирмакутных – эффективный ионообменник с высокой поглотительной способностью и может содержать большое количество катионов. Поглотительная способность оболочек фирмакутных сопоставима с лучшими промышленными катионитами. У большинства фирмакутных в составе оболочек присутствуют дополнительные структурные компоненты, образованные полисахаридами, белками или гликопротеидами. Нередко одиночно расположенные молекулы белков образуют дополнительную структуру на поверхности основного слоя клеточной стенки. Например, молекулы белка А стафилококков формируют одномолекулярную антипараллельную структуру, обладающую высокой биологической активностью. Белок А вызывает агглютинацию эритроцитов, угнетает фагоцитоз, комплемент, препятствует трансформации лимфоцитов, является иммунодепрессантом. У стрептококков группы А белок М образует ворсинки из спаренных молекул, связанных с муреином оболочки. Белок М обеспечивает связь в цепочке кокков, а также взаимодействие с клетками эпителия. По своему строению белок М очень близок к мышечному белку позвоночных тропомиозину, поэтому организм не вырабатывает против него антитела. Белок М является иммунологическим мимикрантом.
В наиболее сложно устроенных оболочках фирмакутных на срезах выявляются два электронно плотных слоя, разделенных слоем с меньшей электронной плотностью для электронов, пронизанных фибриллярными мостиками, имеющие на поверхности структуры белков.
Клеточная стенка грамотрицательных (грациликутных) бактерий многослойна, отличается структурной и функциональной сложностью, толщина ее составляет 14–17 нм. По химическому составу она более разнообразна. Эта группа прокариот отражает один из прогрессивных путей эволюции бактерий. Для них характерно наличие пространства между ЦПМ и клеточной стенкой, появление дополнительной внешней мембраны с большей структурной сложностью, меньшая доля муреина, входящего в состав муреинового мешка – саккулы.
Внешний слой клеточной стенки – наружная мембрана образована фосфолипидами, липополисахаридами – ЛПС, липопротеидами и белками. Основной фракцией наружной мембраны являются липиды, составляющие в среднем 22% сухой массы клеточной стенки.
По строению наружная мембрана имеет типичную трехслойную организацию, характерную для элементарных мембран. Наружная мембрана отличается большей жесткостью и более сложным химическим составом. Основу внешней мембраны составляют липиды (ПЛС, липопротеины, фосфолипиды) и белки. ПЛС составляет около 30–40% поверхности мембраны, локализован во внешнем лепестке мембраны. ЛПС – вещество с высокой антигенной активностью, О-антиген или соматический бактериальный антиген, вызывает выработку антител иммунной системой макроорганизма. ЛПС сальмонелл состоит из липида А и гетерополисахаридной структуры, в которой выделяют О-антигенные цепи и ядро. О-антигенные или О-специфические цепи состоят из повторов олигосахаридов или гетерополисахаридов уникальной структуры. Эти цепи выступают над клеточной стенкой, образуя ворсинки рецепторного назначения, активно взаимодействующие с окружающим миром. О-антигенные цепи клубеньковых бактерий находят корни бобовых симбионтов.
О-антигенные цепи определяют устойчивость бактерий к действию фагоцитов, они часто мутируют и таким образом приобретают устойчивость к действию специфических антител, что дает преимущества патогенным бактериям.
Ядро структуры ЛПС является более стабильной структурой. Иногда выделяют внешнюю и внутреннюю части. Внешняя часть состоит из гексоз разного типа (Д-глюкозы. Д-галактозы, N-ацетилглюкозоамин). Внутренняя часть состоит из гептоз, характерных ЛПС – 2-кето-3дезокси-Д-манно-октоната (КДО) и других гептоз. Формирование полной структуры ЛПС у бактериального организма определяет некоторые культуральные свойства. Бактерии, имеющие полную структуру ЛПС образуют колонии с блестящей поверхностью, а с нарушением в структуре ЛПС – колонии с матовой поверхностью, обладают рядом дефектов внешней мембраны, часто отличаются селективными преимуществами по сравнению с нормой по ЛПС.
Липид А является мембранным носителем для ЛПС и представляет собой фосфолипид. Центральную часть липида А составляет дисахарид, состоящий из двух фосфорилированных Д-глюкозоаминовых остатков. Глюкозоамины ацилированы жирнокислотными остатками.
ЛПС большинства бактерий токсичен для животного организма, то есть является эндотоксином. ЛПС вызывает лихорадку, лейкоцитоз, токсический шок и даже смерть.
Липопротеин Брауна – основной компонент внешней мембраны, имеет молекулярную массу около 7000, состоит из 58 аминокислот. В липопротеине повторяется 15 аминокислот. Первичная структура расшифрована Брауном, предполагается эволюционное закрепление дупликации гена. Отсутствие или нарушения в строении липопротеина влияют на функции внешней мембраны.
Белки основы внешней мембраны или порины составляют до 80% белков мембраны, выполняют функции формирования гидрофильных пор. Известно более 10 видов белков основы, но одновременно в клеточной оболочке бывает 2–3, но не более 5 видов. Молекулы поринов формируют тримеры с образованием гексагональной сетки матрикса мембраны. Предположительно три молекулы порина образуют водную пору диаметром 2 нм. Каждая молекула порина стабилизирована тремя молекулами липопротеина, одна из каторых ковалентно соединена с муренином. Белковый матрикс оболочек грациликутных определяет прочность и форму клеток. Белки основы выполняют важную роль в транспорте элементов питания, служат рецепторами для фагов и колицинов. Белки порины играют большее значение для прокариот естесственных мест обитания, чем для культивируемых.
Вторую группу белков составляют минорные белки внешней мембраны. Они выполняют транспортные и рецепторные функции. Полифункциональность – основное характерное свойство белков мембраны. Белки определяют культуральные свойства бактерий.
Внешняя мембрана содержит двухвалентные катионы, которые удаляют с помощью ЭДТА или воды. Внешняя мембрана обладает трансмембранным потенциалом, она отрицательно заряжена со стороны периплазмы. Величина потенциала зависит от катионного состава среды; обычно оценивают в 20–30 Мв.
Периплазматическое пространство составляет около 10 нм, объем зависит от условий среды. Периплазма содержит муреиновый слой (саккулу) и раствор белков и олигосахаридов. Толщина муреинового слоя 1,6–3 нм, что соответствует одному-трем слоям муреина. Белки периплазмы двух типов. Встречаются гидролитические ферменты и транспортные белки. Возможно, что транспортные белки периплазмы ассоциированы с белками ЦПМ, которые обуславливают транспорт субстратов. Олигосахариды играют решающую роль в осморегуляции клетки.
У некоторых видов грациликутных на внешней мембране имеются белковые молекулы, образующие сетчатую структуру, несущие дополнительные зашитные функции.
Отличительные черты клеточных оболочек микобактерий
Микобактерии характеризуются усложненными структурами оболочек. Относятся к фирмакутным бактериям. В оболочках выделяют 4 компонента:
скелет оболочек, образованный муреином, связанным ковалентно с миколатом арабиногалактана;
2) свободные липиды;
3) пептиды;
4) глюкан.
Муреин микобактерий отличается рядом особенностей. Вместо ацетилмурамовой кислоты муреин содержит N-гликолилмурамовую кислоту. Свободные карбоксильные группы аминокислот амидированы, встречаются необычные связи типа мезо-ДАП – мезо-ДАП. Характерным компонентом оболочек микобактерий являются миколовые кислоты.
Миколовые кислоты это – разветвленные – гидроксикислоты. По количеству атомов углерода различают типы кислот. Коринеформные – с 28–36 С, нокардиальные – с 40–60 атомами С, микобактериальные – с 60–90 атомами С.
Миколовые кислоты придают клеточной поверхности гидрофобность и кислотоустойчивость, способность к использованию гидрофобных субстратов (парафинов нефти), то есть определяют механизм важного адаптационного процесса. Компонентом миколовых кислот является миколат арабиногалактана, терминальные ветви которого определяют иммунные свойства. Патогенные микобактерии содержат корд-фактор, 6,6-димиколаттреголозы, обладающий высокой биологической активностью.
Клеточная стенка бактерий выполняет ряд функций. Она является наружным барьером клетки, устанавливающим контакт микроорганизма со средой. Обладая высокой степенью прочности, она выдерживает внутреннее давление протопласта в гипотоническом растворе.
Исключение составляют галофильные бактерии, клеточные стенки которых лишены муреина. Плотность (ригидность) муреинового каркаса клеточной стенки определяет форму микроба. Поскольку на поверхности клеточной стенки бактерий находятся фагоспецифичные рецепторы, следовательно, именно она определяет адсорбцию фага.
Структурную дифференциацию клеточной стенки грамотрицательных бактерий следует рассматривать как более высокую ступень эволюционного развития прокариот. Наружная мембрана клеточной стенки грамотрицательных бактерий выполняет роль дополнительного барьера, поэтому грамотрицательные бактерии оказываются более устойчивыми к действию различных химических веществ, ферментов и антибиотиков.
Протопласты, сферопласты, L-формы
Действие некоторых антибиотиков (группы пенициллина и циклосерина), подавляющих синтез клеточной стенки бактерий, а также литических ферментов, разрушающих муреин, приводит к образованию протопластов или сферопластов. У последних, в отличие от протопластов, клеточная стенка разрушена не полностью.
Протопласты и сферопласты сохраняют обмен веществ, способность к росту, а иногда и к делению. При снятии действующего фактора они превращаются (реверсируют) в нормальные клетки, либо трансформируются в так называемые L-формы, либо погибают.
L-формами бактерий принято называть бактерии, лишенные клеточной стенки, но сохранившие способность к развитию. Морфологически L-формы бактерий полиморфны. Они представлены шаровидными или неправильной формы телами, часто вакуолизированными, размером от мельчайших (0,2–1 мкм в диаметре) до гигантских (от 5 до 50 мкм). Среди L-форм бактерий различают стабильные, которые почти никогда не реверсируют в исходный вид, и нестабильные, восстанавливающие клеточную стенку после прекращения действия трансформирующего агента.
Образование L-форм патогенных бактерий в организме больного человека можно рассматривать как одну из причин хронических рецидивирующих заболеваний. При неправильном приеме антибиотиков, способствующих образованию L-форм бактерий, может наступить улучшение состояния больного. Однако после прекращения приема лечебного препарата наступает реверсия L-форм в бактерии исходного вида с восстановлением их вирулентности, что приводит к рецидиву болезни. Свое название формы бактерий получили от имени Листера – известного английского врача, применившего впервые методы антисептики.
Жгутики. Движение бактерий. Жгутики встречаются во всех группах прокариот как постоянный или временный компонент. Рассмотреть жгутики можно только в электронном микроскопе после негативного контрастирования или оттенения их тяжелыми металлами. В световом микроскопе без специальных методов обработки отдельные жгутики не видны.
Жгутик состоит из нити, крюка и базальной структуры. Длина его обычно превышает длину клетки в 3 раза и в среднем составляет 10–20 мкм. Нить жгутика представлена полым цилиндром толщиной 12–20 нм, иногда нить окружена чехлом, и тогда ее толщина может достигать 35 нм. По химическому составу нить жгутика образована белком флагеллином. Аминокислотный состав флагеллина у разных видов бактерий может несколько варьировать. Содержание его нередко достигает 2% сухой массы клетки.
Концы нити жгутика неравноценны. Дистальный конец, удаленный от клетки, имеет Т-форму; проксимальный конец, обращенный к клетке, – Н-форму. Рост жгутика происходит с Т-конца. Считают, что молекулы флагеллина проходят по цилиндру нити и пристраиваются к Т-концу. Скорость роста нити жгутика у разных бактерий разная, в среднем она составляет 0,3–0,5 мкм/с. Полноценный жгутик Escherichia coli отрастает за 10 мин.
Нить жгутика крепится к крюку, представляющему собой изогнутый белковый цилиндр длиной 30–90 нм. Проксимальная часть крюка соединяется с палочкой, или осью, базальной структуры жгутика. Базальная структура жгутика сложная и состоит из двух или четырех колец, цилиндра и палочки-оси, на которой крепятся кольца и основание крюка. У грамотрицательных бактерий в базальной структуре жгутика имеются 4 кольца, из них 2 дистальных кольца крепятся в наружной мембране и муреиновом слое клеточной стенки, проксимальные кольца локализованы: одно – в периплазматическом пространстве, другое – в цитоплазматической мембране. В базальной структуре жгутиков грамположительных бактерий имеются только 2 кольца, одно из которых крепится в цитоплазматической мембране, второе – в муреине клеточной стенки. По расположению и числу жгутиков на поверхности клетки бактерии подразделяются на: монополярные монотрихи – клетка несет один жгутик на одном из полюсов; монополярные политрихи – клетка несет пучок жгутиков на одном полюсе; биполярные монотрихи – клетка несет по одному жгутику на обоих полюсах; биполярные политрихи – клетка несет по пучку жгутиков на обоих полюсах; перетрихи – масса жгутиков располагается по всей поверхности клетки. Иногда жгутики располагаются латерально – на боковой стороне клетки. Число жгутиков (от 1–2 до 50–100 и более) и их расположение на поверхности клетки постоянны для каждого вида бактерий и являются диагностическими признаками микробов.
По функции жгутики – органы движения. Они типичны для плавающих палочковидных и извитых форм бактерий и лишь в единичных случаях встречаются у кокков. Жгутики бактерий по характеру работы подобны корабельному винту. Если клетка несет много жгутиков, они при движении собираются в пучок. Один жгутик или пучок жгутиков, быстро вращаясь против часовой стрелки, вызывает вращение клетки только в противоположном направлении и с меньшей скоростью.
Предполагается, что импульс вращения жгутики получают от вращения проксимального кольца базальной структуры, закрепленного в цитоплазматической мембране. Скорость поступательного движения клетки зависит от вида микроорганизма и условий окружающей среды. Нередко клетка проходит в 1 с расстояние в 20–60 мкм, т.е. в 20–50 раз больше длины собственного тела. К спринтерам среди бактерий относится холерный вибрион, его скорость равна 200 мкм/с.
Жгутики могут изменять направление вращения, и в этом случае бактерия останавливается и начинает кувыркаться. Вращение жгутиков у бактерий обеспечивается энергией трансмембранной протондвижущей силы. Эта сила создается при дыхании за счет выброса протонов в дыхательной цепи при переносе их через цитоплазматическую мембрану. Скорость вращения жгутиков зависит от величины мембранного потенциала.
Своеобразный тип движения характерен для спирохет. Тело спирохет состоит из протоплазматического цилиндра, окруженного цитоплазматической мембраной, и внешнего чехла, представленного муреиновым слоем и наружной мембраной. Вокруг протоплазматического цилиндра в периплазматическом пространстве перевиты пучки нитчатых структур –аксиальных фибрилл. Число их варьирует у разных спирохет от 2 до 100 и более. Одним своим концом аксиальные фибриллы закрепляются на полюсах протоплазматического цилиндра, второй конец их свободен. В центральной части тела спирохет аксиальные фибриллы перекрываются.
Своеобразное движение спирохет осуществляется за счет вращения аксиальных фибрилл, вызывающего эластичную волну на поверхности внешнего чехла. Спирохеты способны вращаться вокруг оси спирали, изгибаться и осуществлять передвижение по винтовому или волнообразному пути. Аксиальные фибриллы спирохет, являясь внутриклеточными структурами, выполняют функцию бактериальных жгутиков.
У некоторых миксобактерий, микоплазм, нитчатых серобактерий и цианобактерий установлен скользящий тип движения с очень незначительной скоростью порядка 2–10 мкм/с. Механизм скользящего движения пока неясен. У ряда нитчатых цианобактерий между муреиновым слоем и наружной мембраной в клеточной стенке обнаружен тончайший непрерывный слой фибрилл. Предполагается, что функция фибрилл аналогична функции жгутиков, только фибриллы находятся в клеточной стенке цианобактерий и не выходят на поверхность. Скользящее движение нитчатых бактерий обычно сопровождается и вращением клетки.
Направленное передвижение бактерий в ответ на действие раздражителя получило название таксиса. Таксис может быть положительным или отрицательным. Принято различать хемотаксис, аэротаксис, фототаксис, в зависимости от действующего раздражителя. Фототрофные бактерии способны различать самые незначительные изменения освещенности среды, всего на 1 0,7%, т.е. почти так же, как человеческий глаз (0,4%). Обладая высокой чувствительностью к действию различных факторов окружающей среды, микроорганизмы за короткое время локализуются в оптимальной зоне обитания.
Фимбрии. Под электронным микроскопом на поверхности бактериальных клеток открыты тонкие нитевидные структуры, получившие название фимбрий или пилей. По размерам они короче и тоньше жгутиков. Длина их составляет 0,3–0,4 мкм при диаметре 5–10 нм. Фимбрий – прямые полые цилиндры, отходящие от цитоплазматической мембраны. Они образованы специфическим белком пилином. Число фимбрий на поверхности клетки колеблется от одной-двух до нескольких тысяч. Фимбрии имеют кокковидные, палочковидные, подвижные и неподвижные бактерии. Функции фимбрий не связаны с движением клетки.
По морфологии, антигенным свойствам и выполняемым функциям принято различать несколько типов фимбрий.
Фимбрии первого типа выполняют функцию прикрепления клетки к поверхности субстрата или сцепления клеток при образовании зооглеи. Возможно, они принимают участие в проведении крупномолекулярных соединений питательного субстрата в цитоплазму.
Фимбрии второго типа – половые фимбрии, или F-пили. Они образованы белком с молекулярной массой 11800, длина их составляет около 1,1 мкм при толщине 8,5–9,5 нм. Эти пили образуются на клетках-донорах в количестве 1–2, в анаэробных условиях иногда до 5. При конъюгации конец половой фимбрии клетки-донора прикрепляется к белку наружной мембраны клетки-реципиента. Считается, что по цитоплазматическому мостику через канал F-пили или непосредственно по ее каналу происходит передача материала ДНК из клетки-донора в клетку-реципиент. Половые пили способна отрастать за 4–5 мин н также быстро сбрасываться клеткой, что свидетельствует об активности данной структуры.
studfiles.net
Химический состав клеточной стенки прокариот
В состав клеточной стенки прокариот входят семь различных групп химических веществ, при этом пептидогликан присутствует-только в клеточной стенке, придавая ей необходимую жесткость. [c.26]Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации (рис. 4). Структуры, расположенные снаружи от ЦПМ (клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки), называют обычно поверхностными структурами. Термином клеточная оболочка часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). ЦПМ вместе с цитоплазмой называется протопластом. Рассмотрим сначала строение, химический состав и функции поверхностных клеточных структур. [c.27]
У грамотрицательных прокариот строение клеточной стенки намного сложнее, чем у грамположительных (рис. 1.4). В ее состав входит гораздо большее число макромолекул разного химического типа. [c.16]
По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты, относящиеся к эубак-териям, делятся на две большие фуппы. Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна у так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются. У некоторых эубактерий положительная реакция при окрашивании описанным выше способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки. [c.28]
Клеточная стенка. Клеточная стенка — важный и обязательный структурный элемент прокариотной клетки (исключение миконлазмы и Ь-формы), располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с окружающей средой (у клеток, не содержащих этих слоев клеточной оболочки). На долю клеточной стенки приходится от 5 до 50 % сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. Концентрация солей в клетке, как правило, намного выше, чем в окружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка чисто механически защищает клетку от проникновения в нее избытка воды. По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. [c.12]
ХИМИЧЕСКИЙ СОСТАВ И СТРОЕКИЕ КЛЕТОЧНОЙ СТЕНКИ МИКРООРГАНИЗМОВ Химический состав клеточной стекни прокариот [c.10]
Кроме пептидогликана в состав клеточных стенок грамположительных прокариот входит другой уникальный класс химических соединений — тейхоевые кислоты, представляющие собой полимеры, построенные на основе рибита (пятиатомного спирта) или глицерина (трехатомного спирта), остатки которых соединены между собой фосфодиэфирны-ми связями (рис. 8). Свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками В-аланина, глюкозы, М-ацетилглюкозамина и некоторых других сахаров. Тейхоевые кислоты ковалентно могут соединяться с Ы-ацетилмурамовой кислотой (см. рис. 6). Поскольку это длинные линейные молекулы, они могут пронизывать весь пептидогликановый слой, достигая внешней поверхности клеточной стенки. В этом случае, вероятно, они являются основными антигенами грамположительных прокариот. [c.28]
Капсулы, слизистые образования и чехлы могут содержать компоненты, одинаковые с клеточной стенкой, однако их химические составы не идентичны. Как правило, химический состав капсул, образуемых бактериями, родо- или видоспецифичен. Основные химические компоненты большинства капсул прокариот — полисахариды гомо- или гетерополимерной природы. Исключение составляет капсула некоторых видов Ba illus, построенная из полипептида, являющегося полимером Л-глутаминовой кислоты. Для ряда бактерий показана способность синтезировать и вьщелять в окружающую среду волокна целлюлозы. [c.38]
Важным шагом в развитии систематики прокариот явилось использование признаков, даюших информацию о химическом строении клетки состав оснований ДНК, ДНК—ДНК- и ДНК— РНК-гомологии, аминокислотная последовательность белков, строение рибосом, компонентов клеточной стенки и т.д. [c.157]
Клеточная стенка грамотрицательных прокариот, У грамотрицательных прокариот строение клеточной стенки намного сложнее, чем у грамположительных (см. рис. 5). В ее состав входит гораздо большее число макромолекул разного химического типа (см. табл. 3). Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов грамотрицательных прокариот содержание этого гетерополимера колеблется в широких пределах (1—10% и больше от веществ клеточной стенки). Предполагается, что у большинства видов грамотрицательных прокариот он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями (рис. 9). [c.29]
chem21.info
