
Когда и при каких обстоятельствах на Земле появились первые цветковые растения?
Одно из важнейших направлений развития растительного царства — приспособление к превратностям наземной жизни. Цветковые растения являются ярчайшим выражением этой линии и доминируют на земной поверхности в данную эпоху. От полюсов до экватора нет такого участка, где возможна растительная жизнь, но не найдено покрытосеменных. Они в изобилии встречаются в долинах рек и чистых озёрах, в меньшем количестве — в солёных озёрах и морях. Однако такие водные покрытосеменные не являются примитивными формами, а возникли путём приспособления наземного предка к водной среде. К широчайшему географическому разнообразию прибавляется разнообразие форм и способов роста. Банальная ряска, покрывающая поверхность пруда, представляет собой крошечный зелёный побег с простым корешком, вертикально погруженным в воду, и с очень нечёткими листиками и частями стебля. Могучее лесное дерево столетия развивало свою сложную систему стволов и ветвей, покрытых бесчисленными веточками и листвой, а под землёй соответствующую площадь занимает мощная, хорошо развитая корневая система. Между этими двумя крайностями — бесконечные градации: водные и земные травы, ползучие, прямостоящие или карабкающиеся, кусты и деревья, гораздо большее разнообразие, чем в другом отделе семенных растений — Gymnospermae.Первые остатки покрытосеменных датируются Юрским периодом приблизительно 140 миллионов лет назад. Базируясь на современных данных, можно предположить, что предки покрытосеменных и гнетовых дивергировали в триасе (220—202 миллионов лет назад) . Остатки растений с признаками покрытосеменных появились в юре и раннем мелу (135-65 миллионов лет назад) , но это были довольно-таки малочисленные и примитивные формы. Следы широкого развития и распространения покрытосеменных появились в палеонтологической летописи в период среднего мела (около 100 миллионов лет назад) . Но уже в позднем мелу покрытосеменные оказались доминирующей формой растительной жизни, и в многих фоссилиях узнаются представители современных семейств (например, бук, дуб, клен и магнолия)
otvet.mail.ru
ПОЯВЛЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ. Жизнь в глубинах веков
ПОЯВЛЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ
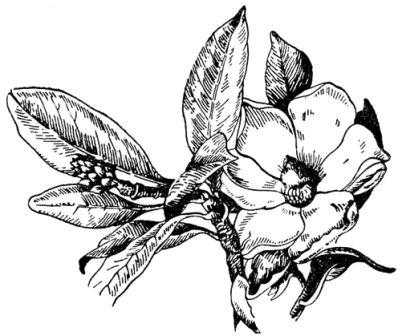
Шишка, или цветок беннеттита (слева), ицветок магнолии (часть органов растенийудалена)
В мезозойскую эру произошли два великих события в истории жизни на Земле, которые изменили и обновили органический мир Земли: появились первые млекопитающие и цветковые, или покрытосеменные, растения.Уже в юрский период леса сильно изменились: поредели папоротниковые заросли, размножились голосеменные растения и саговники. Саговники — небольшие деревья с прямыми стволами и длинными пористыми листьями. По виду они похожи на папоротники и на пальмы. Они были потомками семенных папоротников и размножались семенами. Близкими родственниками саговников были беннеттиты, расцвет которых относится к меловому периоду. Они размножались семенами, которые были собраны в шишки и напоминали цветы с древних цветковых растений — магнолий. Замечательные растения мезозоя — гинкговые. Один вид этих растений — гинкго двулопастное — дожил в Японии и Китае до наших дней. Листья гинкговых похожи на веер и расположены на верхушке. В меловом периоде в связи с похолоданием и более сухим климатом растительный покров Земли изменяется еще больше. Среди хвойных лесов нередки древние сосны и ели. В лесах из саговых пальм и других древних растений появляются тополь, ивы и другие знакомые нам деревья. Появляются настоящие цветковые, или покрытосеменные, растения. Цветоносные растения стали возникать уже в конце палеозоя, около 200 миллионов лет назад. Это были известные нам саговые, гинкговые и хвойные. Но цветы их невзрачны и имеют простое строение. Образующиеся в них семена лежат в растении «голыми», ничем не прикрытыми. Поэтому их называют голосеменными. Но и такие семена имеют преимущество перед простыми спорами. В меловой период у предков цветковых растений цветы становились более разнообразными по форме, цвету, запаху. Их семена стали помещаться в особой полости — в разрастающейся завязи цветка. Поэтому такие растения ботаники называют покрытосеменными. С появлением их растительный покров Земли стал резко изменяться, и в сравнительно короткое время эти растения широко размножились и распространились повсюду.
Цветок магнолии, одного из древних цветковых, или покрытосеменных, растений
Появление на арене жизни цветковых растений резко изменило не только растительный покров Земли, но и всю органическую жизнь, особенно на суше. Однообразные леса из папоротников и голосеменных сменились разнообразными по форме и цвету лесами и лугами из цветковых растений. Приспособляясь к изменяющимся условиям среды, цветковые растения дали многообразные деревья, кустарники и травянистые однолетние и многолетние растения. Мы уже говорили о том, что без цветковых растений едва ли может быть большой прогресс в мире животных, особенно насекомых, птиц и млекопитающих.
Следующая глава >
bio.wikireading.ru
Покрытосеменные растения появились на Земле около 130 млн лет назад. — МегаЛекции
Происхождение растений (время,
Начальные этапы развития растительного мира. Первые живые организмы появились в воде примерно 3,5—4 млрд лет назад. Простейшие одноклеточные организмы по строению были схожи с бактериями. Они ещё не имели обособленного ядра, но обладали системой обмена веществ и способностью к размножению. Постепенно запасы питательных веществ в первичном океане стали истощаться. Между клетками началась борьба за пищу. В этих условиях у некоторых клеток появился зелёный пигмент — хлорофилл, и они приспособились к использованию энергии солнечного света для превращения в пищу воды и углекислого газа. Так возник фотосинтез, то есть процесс образования органических веществ из неорганических с использованием энергии света. С появлением фотосинтеза в атмосфере стал накапливаться кислород. Состав воздуха стал постепенно приближаться к современному, то есть в основном включать азот, кислород и небольшое количество углекислого газа. Такая атмосфера способствовала развитию более совершенных форм жизни.
Появление водорослей. От древних простейших одноклеточных организмов, способных к фотосинтезу, произошли одноклеточные водоросли. Одноклеточные водоросли — родоначальники царства растений.
Важное значение для дальнейшего развития растений имело возникновение у водорослей полового размножения. Размножение половым путём способствовало изменчивости организмов и приобретению ими новых свойств, которые помогали приспособиться к новым условиям жизни.
Выход растений на сушу. Поверхность материков и дно океана со временем изменились. Поднимались новые материки, уходили под воду существовавшие раньше. Из-за колебаний земной коры на месте морей возникала суша. Изучение ископаемых остатков показывает, что растительный мир Земли тоже изменялся.
Переход растений к наземному образу жизни, по-видимому, был связан с существованием периодически заливавшихся и освобождавшихся от воды участков суши. Осушение этих участков происходило постепенно. У некоторых водорослей стали появляться приспособления к обитанию вне воды.
В это время на земном шаре был влажный и тёплый климат. Начался переход некоторых растений от водного к наземному образу жизни. У древних многоклеточных водорослей строение постепенно усложнялось, и они дали начало первым наземным растениям. Одними из первых наземных растений были росшие по берегам водоёмов риниофиты, например риния. Они существовали 420—400 млн лет назад, а потом вымерли. Размножались они спорами.
Происхождение высших споровых растений. От риниофитоподобных растений произошли древние плауны, хвощи и папоротники и, по-видимому, мхи, у которых уже были стебли, листья, корни. Это были типичные споровые растения, своего расцвета они достигли около 300 млн лет назад, когда климат был тёплым и влажным, что благоприятствовало росту и размножению папоротников, хвощей и плаунов. Однако их выход на сушу и отрыв от водной среды не были ещё окончательными. При половом размножении споровым растениям для оплодотворения необходима водная среда.
Происхождение высших растений
Развитие семенных растений. В конце каменноугольного периода климат Земли почти повсеместно стал суше и холоднее. Древовидные папоротники, хвощи и плауны постепенно вымирали. Появились примитивные голосеменные растения — потомки некоторых древних папоротниковидных .
Происхождение голосеменных от древних папоротниковидных доказывает многие черты сходства между этими растениями. Это сходство не только внешнее. Общие черты наблюдаются в строении органов: стеблей, листьев и корней. Очевидно, предками голосеменных растений были древовидные, лиановидные и травянистые семенные папоротники, впоследствии полностью вымершие. Их семена развивались на листьях, шишек ещё не было.
Условия жизни продолжали меняться. Там, где климат становился более суровым, древние голосеменные растения постепенно вымирали. Им на смену приходили более совершенные растения — сосна, ель, пихта.
Растения, размножавшиеся семенами, лучше приспособились к жизни на суше, чем растения, размножавшиеся спорами. Это связано с тем, что возможность оплодотворения у них не зависит от наличия воды во внешней среде. Особенно явно превосходство семенных растений над споровыми проявилось, когда климат стал менее влажным.
Покрытосеменные растения появились на Земле около 130 млн лет назад.
Покрытосеменные оказались наиболее приспособленными к жизни на суше растениями. Только у покрытосеменных имеются цветки, их семена развиваются внутри плода и защищены околоплодником. Покрытосеменные быстро расселились по всей Земле и заняли все возможные места обитания. Уже более 60 млн лет покрытосеменные растения господствуют на Земле.
Приспособившись к различным условиям существования, покрытосеменные создали разнообразный растительный покров Земли из деревьев, кустарников и трав.
В современной систематике царство растений делится на три подцарства:настоящие водоросли, багрянки (красные водоросли) и высшие растения (зародышевые).
Подцарство «низшие растения» Водоросли – древнейшие Среди них встречаются как одноклеточные, микроскопических размеров растения, так и многоклеточные гиганты. Тело представлено талломом или слоевищем. Органы полового и бесполого размножения, как правило, одноклеточные. 1. Тело не поделено на органы 2. Плохо развиты проводящая, механическая и покровная ткани, а значит и небольшие размеры 3. Взрослая особь гаплоидна 4. В цикле развития преобладает спорофит, Два подцарства — Красные, или Багрянки, и Настоящие водоросли. У наст вод три отдела (Зеленые, Харовые, Бурые и др.).
Для высших растений характерна дифференцировка тела на вегетативные органы и наличие тканей, выполняющих определенные функции.
Высшие растения делят на две группы — высшие споровые (отделы Моховидные, Плауновидные, Псилотовидные, Хвощевидные, Папоротниковидные) и семенные растения (отделы Голосеменные и Покрытосеменные).
Моховидные,. Известны с карбонового периода. Предки- зеленые водоросли. Три класса: листостебельные, антоцеротовые и печеночные мхи. Листостебельные, или настоящие мхи (кукушкин лен и сфагнум). Лишены корней, Тело делится на стебель и листья. Ризоиды – аналоги корней высших растений. Размножение происходит бесполым, вегетативным и половым путем. Спорофит короткоживущий, находится на гаметофите и состоит из трех частей: спорангия (коробочки) со спорами, ножки, к которой крепится коробочка, и подошвы – места прикрепления к гаметофиту. Тесная связь спорофита и гаметофита объясняет факт групповых форм роста мхов.
Отдел Плауновидные – очень древняя группа. Имеют длинные стелющиеся дихотомически ветвящиеся побеги, густо усаженные жесткими мелкими листьям. Преобладает спорофит. (плаун булавовидный, плаун-баранец, плаун двуострый).
Отдел хвощевидные. Многолетние травы лугов, болот, лесов и полей. От корневищ отходят придаточные корни и надземные побеги, имеющие типичное членистое строение. От узлов отходят мутовки бурых чешуйчатых листьев, сросшихся в трубчатое влагалище, и мутовки боковых побегов.
Отдел папоротниковидные. Преобладание долговечного листостебельного спорофита над эфемерным примитивным гаметофитом, наличие у спорофита крупных, обычно перисто-рассеченных, сложно устроенных листьев – вай, расположение спорангиев группами (сорусами) на нижней стороне листьев.
| Основные признаки Спорофит является доминирующим поколением; гаметофит крайне редуцирован |
Голосеменные Обычно образуют шишки, на которых развиваются спорангии, споры и семена Семена не спрятаны в завязи. Они лежат на поверхности специализированных листьев, называемых семенными чешуями*, которые собраны в шишки. Не образуют плодов, поскольку не имеют завязи
Покрытосеменные. Образуют цветки, в которых развиваются спорангии, споры и семена* Семена спрятаны в завязи* После оплодотворения из завязи развиваются плоды
Семенные растения подразделяют на два отдела: Голосеменные и Покрытосеменные
Голосеменные– это древний отдел растений, которые характеризуются наличием семязачатков с последующим развитием семени, но не образуют цветков и плодов. Это главные признаки голосеменных. Отдел голосеменных растений включает 4 класса: хвойные, гинкговые, гнетовые, саговниковые. К голосеменным относятся около 800 современных видов. Преобладающее большинство видов этого отдела (более 500) составляют класс хвойных растений, появившихся на наше планете в конце карбонового периода. Они произрастают на всех континентах. Почти 95% лесов Земли состоят только из хвойных деревьев или являются смешанными.
В отделе Покрытосеменных (Цветковых) растений выделяют два класса — Двудольные и Однодольные растения. Двудольные растения появились раньше, в последствие от них произошли однодольные.
Отличительные признаки двудольных растений: У зародыша семени две семядоли. Запас питательных веществ семени хранится либо в зародыше, либо в эндосперме. Зародышевый корешок дает начало хорошо развитому главному корню. Стержневая корневая система. Листья с перистым или пальчатым жилкованием. Проводящая ткань стебля имеет выраженное кольцевое строение. Благодаря кольцевому слою камбия стебель растет в толщину. Много как древесных, так и травянистых форм. Бобовые, сложноцветные, розоцветные, пасленовые и тд
Отличительные признаки однодольных растений: У зародыша семени одна семядоля. Запас питательных веществ находится в эндосперме. Зародышевый корешок развит слабо. При прорастании от стебля отходят придаточные корни. Мочковатая корневая система. Листья с параллельным или дуговым жилкованием. Черешок листа либо почти не развит, либо является листовым влагалищем. Проводящая ткань стебля состоит из пучков. Кольца камбия как у двудольных нет. Преимущественно травы.Злаковые, лилейные.
Филогенез растений
В конце архейской эры появились простейшие организмы, способные к фотосинтезу и давшие начало бактериям и сине-зеленым водорослям. обогащают окружающую среду кислородом.
Настоящие растения появились в начале протерозойской эры — это зеленые и красные водоросли.
Кембрийский и ордовикский периоды палеозойской эры характеризуются появлением крупных многоклеточных водорослей. В это время продолжают господствовать бактерии, сине-зеленые, красные и зеленые водоросли. Некоторые древние виды водорослей вымирают.
В силурийском периоде появляются псилофиты, водная растительность в виде водорослей продолжает развиваться, древние виды водорослей вымирают.
В следующем за силуром девонском периоде палеозойской эры появляются папоротники, хвощи, плауны, грибы и мхи. Псилофиты достигли господствующего положения, в морях распространились различные отделы водорослей.
За девоном наступил каменноугольный период (карбон). В отличие от засушливого девона, карбон был влажным и теплым, что привело к значительному развитию папоротников. Влажная, теплая, богатая углекислым газом атмосфера способствовала интенсивному развитию гигантских папоротников. Их отмершие стебли попадали в анаэробные условия, не подвергались гниению, что привело к формированию залежей каменного угля (отсюда и название периода).
В карбоне возникают первичные голосеменные растения. Появление семенного размножения явилось значительным ароморфозом в мире растений. Семенное размножение дает ряд преимуществ для растений: обеспечивается лучшая защита зародыша на первых этапах жизни, что способствует более хорошему выживанию в среде обитания, утрачивается зависимость полового процесса от внешней водной среды и автономно существующее растение получает диплоидный набор хромосом, что также способствует лучшему выживанию нового организма.
Каменноугольный период характеризуется господством папоротникообразных на суше, различных водорослей в море. В этом периоде псилофиты вымирают и появляются семенные папоротники.
Палеозойская эра завершается пермским периодом, который в отличие от карбона характеризовался сухим климатом. В этих условиях гигантские папоротники стали вымирать, появились травянистые формы папоротников, хвощей и плаунов. Вымирают и семенные папоротники (в конце периода), а их место занимают древние голосеменные растения.
После палеозойской эры наступила мезозойская эра, или мезозой, состоящая из трех периодов — триасового, юрского и мелового.
В триасовом периоде, начинающем мезозой, появляются современные голосеменные растения, господствуют древние голосеменные. Окончательно вымирают гигантские папоротники, хвощи и плауны, а голосеменные достигают наивысшего расцвета.
В следующем, юрском, периоде мезозоя появляются первые покрытосеменные растения (новый значительный ароморфоз в растительном мире.
Появление покрытосеменных растений является резким повышением уровня организации в растительном мире, так как у этих растений половой процесс становится окончательно автономным, не зависящим от воды, находящейся во внешней среде. Двойное оплодотворение повысило жизнеспособность зародыша, а покровы в виде оболочек плода повысили устойчивость семян к вредному воздействию условий внешней среды и .способствовали лучшему распространению плодов и семян, расширению ареала этих растений.
В меловом периоде, завершающем мезозой, появляются древние покрытосеменные растения, господствуют современные голосеменные растения, окончательно вымирают древние голосеменные растения и некоторые виды папоротников.
В конце мелового периода усиливаются горообразовательные процессы, наступает похолодание, уменьшается поверхность воды, сокращаются ареалы околоводной растительности.
Начинается кайнозойская эра, состоящая из палеогена, неогена и антропогена. В начале кайнозоя горообразовательные процессы завершаются. Обособляются Средиземное, Черное, Каспийское и Аральское моря. Устанавливается теплый, равномерный климат. На севере преобладает хвойная, а на юге лиственная растительность. Леса сменяются на степь, полупустыню и пустыню. Происходит формирование современных растительных сообществ.
Современная флора сформировалась к началу антропогена, в конце которого появляются различные виды культурных растений, выведенных человеком. Часть растений вымирает как под воздействием внешних факторов среды, так и под влиянием человека, деятельность которого в настоящее время стала «геологической силой».
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
megalektsii.ru
Когда на Земле появились первые цветы? | | Вода
Люди склонны связывать растения с цветами, но на самом деле цветы появились очень недавно. Это пример недавнего развития форм жизни, по крайней мере, на эволюционной шкале времени.
Прежде, чем они появились, растения воспроизводились с помощью спор в течение сотен миллионов лет. На самом деле, ученые даже не знают, почему цветы развились в первую очередь. Цветы являются тонкими, хрупкими и требуют огромного количества энергии. Многие растения могут найти лучшее применение для выращивания семян или своего увеличения в высоту.
Кроме того, не цветущие растения не имеют ничего, что действительно соответствует форме цветов. Это делает цветы чем-то вроде головоломки, где они пришли как бы ниоткуда. Дарвин лихо описал рост цветов, как «презренную тайну.»
Древнейшие известные ископаемые цветковых растений относятся к меловому периоду, между 115 и 125 миллионами лет назад.
Старшее среди них, цветковое растение Potomacapnos, удивительно напоминает комплекс семейства современного мака.
Такая сложность в начале ископаемых лет показывает, что цветы, вероятно, прошли очень быстрый этап эволюции, чтобы приблизиться к их современному виду, а не было длительного развития, медленного в течение длительного периода.
Из этого трудно сделать какие-либо окончательные выводы, поскольку цветы являются хрупкими и редко встречаются в виде окаменелостей. Тем не менее, некоторые ответы найдены — они лежать с редкими отпечатками кустарников, найденных на Тихоокеанском острове Новая Каледония. И только там есть эти следы.

Амборелла trichopoda (на фото) является единственным выжившим членом Amborellales. В начале мелового периода, в Amborellales отделились от не-цветковых растений, доминирующих в окружающем пейзаже, став старейшим порядковым цветковым растением, из выживших в настоящем.
Позже, два других сохранившихся окаменелых отпечатка появились у исследователей: это были кувшинкоцветные, которые стали родоначальниками современным кувшинкам.
А ещё найдены австробэйлиецветные, из которых, по мнению учёных, произошли все остальные цветковые растения.
Поскольку Amborellales появились сначала , то современная амборелла trichopoda остается близким к оригинальным среди цветковых растений. Сравнивая его с австробэйлиецветными, мы можем получить некоторое представление о том, какие черты цветущие растения первоначально имели , а какие просто эволюционировали далее вниз по линии.
Филогенетические отношения между основными линиями цветковых растений на основе геномных последовательностей пластид. Изображенный против часовой стрелки от корня на базе дерева окружности: амборелла trichopoda, Nymphaea Odorata, Illicium floridanum, Chloranthus узколистного Пайпер лонгум, лириодендрон тюльпановый, Ceratophyllum demersum, лютик Ficaria, пеларгонии exstipulatum, подсолнечник однолетний, Юкка filamentosa, Triticum AESTIVUM, и аира атепсапиз. Новая Каледония, где проживает амборелла trichopoda, показано в фоновом режиме. Кредит: Гвен Гейдж
Роберт Янсен, профессор интегративной биологии в Университете штата Техас в Остине, отметил, что работа создает основу для всех будущих сравнительных исследований цветковых растений. «Если вы заинтересованы в понимании эволюции цветковых растений, вы не можете сделать это, если только вы понимаете их отношения «, сказал Янсен. Университет Флориды команды, во главе с Дагом и Пэм Солтис, также показали, что диверсификация цветущих растений, настолько была ошеломляющей, что исследователи называют её «Big Bang», она состоялась в сравнительно короткий период — менее пяти миллионов лет.
Цветок, который цветёт 1 раз в 3000 лет
Комнатные цветы фото и название
Растения, цветы и деревья, которые будут расти в щелочной почве
voda.molodostivivat.ru
Цветковые растения появились одновременно с динозаврами
Цветковые растения, характерные для современной земной флоры, появились не в меловом периоде, как это считалось прежде, а намного раньше. Швейцарские палеонтологи обнаружили их окаменевшую пыльцу в горных породах триасового возраста, что говорит о том, что история цветковых по меньшей мере на 100 млн лет длиннее и они начали свою эволюцию практически одновременно с первыми динозаврами.
Господствующие сегодня цветковые произошли от вымерших растений, связанных с хвойными, гинкго, цикадовыми и семенными папоротниками. Наиболее древними окаменелостями цветковых считается пыльца – ее небольшие, прочные и многочисленные зернышки найти намного легче, чем ископаемые цветы или листья. Изучение пыльцы со временем выделилось даже в отдельную науку, которая называется палинология.
Непрерывная последовательность пыльцы ископаемых цветковых начинается в раннем мелу, примерно 140 млн лет назад, и прослеживается до наших дней. На этом основании палеонтологи полагали, что первые цветковые растения появились как раз в меловом периоде. Но находка, сделанная Питером Хочули и Сюзанной Файст-Букхардт из университета Цюриха, показывает, что цветковые росли на нашей планете по меньшей мере на 100 млн лет раньше – еще в раннем триасе. Возможно, что их история уходит в прошлое еще дальше.
Хочули и Файст-Букхардт изучали два керна (столбика горных пород, получаемых при бурении геологических скважин), происходящих из северной Швейцарии. Внутри пород возрастом 252-247 млн лет они нашли пыльцевые зерна, напоминающие пыльцу самых ранних из известных науке цветковых растений. С помощью конфокальной лазерной сканирующей микроскопии им удалось получить изображения с высоким разрешением в трех измерениях сразу шести различных типов пыльцы.
Это не первая находка древнейших пыльцевых зерен, открытых Хочули и Файст-Букхардт. В 2004 году они описали достоверную пыльцу, несколько отличающуюся от цветковых, из среднетриасовых кернов, поднятых со дна Баренцева моря южнее Шпицбергена. Их новая работа посвящена растениям, росшим на 3000 км южнее. «Мы считаем, что даже самые осторожные ученые теперь признают, что цветковые растения эволюционировали задолго до мелового периода», – заявил Хочули.
Согласно современным палеоэкологическим реконструкциям, в триасовом периоде и окрестности Шпицбергена, и территория современной Швейцарии находились в субтропическом поясе. Однако климат на юге был намного суше, чем на севере, а из этого следует, что уже самые ранние цветковые отлично чувствовали себя в самых разных экологических обстановках. Судя по деталям строения пыльцы, эти растения уже опылялись насекомыми, переносившими зерна пыльцы от цветка к цветку.
Многие ученые и раньше пытались оценить возраст цветковых растений, используя для этого, в частности, «молекулярные часы», пишет PhysOrg. Но до сих пор приемлемой точности результата получить не удавалось – в зависимости от набора данных и использованных методик появление первых цветковых приходилось то на мел, то на триас. «Вот почему находка пыльцы ранних цветковых в триасовых породах является такой важной», – отметил Хочули.
Статья «Angiosperm-like pollen and Afropollis from the Middle Triassic (Anisian) of the Germanic Basin (Northern Switzerland)» доступна на сайте Frontiers
www.paleonews.ru
|
^ ХАРАКТЕРИСТИКА СЕМЕЙСТВ КЛАССА ОДНОДОЛЬНЫЕ 4.1. Подкласс Лилииды 4.1.1. Семейство ЛилейныеСемейство Лилейные (Liliaceae) является основным и самым примитивным в древнем порядке Лилиецветные (Liliales), в котором типичные признаки класса Однодольные выражены наиболее отчетливо. Систематика лилиецветных не доведена до логического завершения. Поэтому в научной и учебной литературе нет единого мнения и о видовом составе семейства Лилейные. Лилейные включает около 200 родов и до 4000 видов многолетних растений, произрастающих в разных климатических зонах земного шара. Однако наибольшее распространение лилейные имеют во внетропических областях Европы, Азии, Африки и Северной Америки. Они особенно многочисленны в субтропиках с периодическими засухами, а также в степях и полупустынях. В горных районах лилейные выходят на уровень альпийских лугов. В естественной флоре Беларуси встречается 25 видов из 15 родов, но в овощеводстве и декоративном озеленении используется немало сортов интродуцентов. В семействе преобладают травянистые растения. Однако в тропиках встречаются древовидные: драцена (Dracaena drago), юкка карнерозанская (Yucca carnerosana), алоэ Байнеса (Aloe bainesii) и «травянистое дерево» – кингия австралийская (Kingia australis) и др. Реже встречаются лианы (мирзифиллум – Myrsiphyllum). Комнатным листовым суккулентом является алоэ древовидное (A. arborescens), в естественных местообитаниях оно формирует габитус древесного растения. Для большинства лилейных свойственно образование характерных для эфемероидов подземных луковиц (лук – Allium, лилия – Lilium, тюльпан – Tulipa), клубнелуковиц (безвременник – Colchicum) или корневищ (чемерица – Veratrum, ландыш – Convallaria, спаржа – Asparagus). Эти видоизменения побегов обеспечивают сохранение почек возобновления во время летней жары и зимней стужи. Надземный стебель прямой, реже вьющийся, облиственный или безлистный, именуемый цветоносной стрелкой. Листья более или менее мясистые, сидячие; узкие, реже широкие; цельнокрайние с параллельным или дуговым жилкованием. Листорасположение очередное, у вороньего глаза (Paris) 4 – 10 листьев образуют мутовку на стебле. Цветки одиночные, как у безвременника, или многочисленные, например, у ландыша. Соцветия самые разнообразные: метельчатое (чемерица), кистевидное (эремурус – Eremurus), иногда зонтик (лук). Цветки актиноморфные, обоеполые. Околоцветник простой, чаще из шести свободных листочков или сросшихся в длинную трубку. Околоцветник обычно венчиковидный, реже, как у вароньего глаза, из восьми листочков околоцветника четыре образуют внешний чашечковидный круг. Андроцей состоит из 6, иногда 4 или 8 тычинок, расположенных в два круга. Гинецей синкарпный из трех плодолистиков. Рыльце обычно сидячее, как у тюльпана. Завязь верхняя, очень редко полунижняя; трехгнездная, реже одно- или четырехгнездная, с многочисленными семязачатками. Формула цветка:Р3+3А3+3G(3) илиР(3+3)А3+3G(3). Плод 3-гнездная коробочка (лук) или ягода (ландыш). Семена с эндоспермом. Растения опыляются в большинстве случаев насекомыми, редко – ветром, в тропиках – птицами. Лилейные имеют большое хозяйственное значение. Ценными овощными культурами являются многочисленные виды лука и спаржи. Род Лук (Allium) представлен луковичными или корневищными растениями. Луковицы простые (лук репчатый – A. cepa) и сложные (чеснок – A. sativum). Листья либо плоские, как у лука поррея (A. porrum) и чеснока, либо трубчатые, как у лука репчатого и лука дудчатого (A. fistulosum). Соцветие – зонтик, покрытый до распускания пленчатой оберткой, расположен на цветоносной стрелке и включает до 2000 цветков. Цветки мелкие, обоеполые на длинных цветоножках; зеленоватой, белой, фиолетовой, розовой, желтой и других окрасок. Иногда в соцветии развиваются луковички – выводковые почки. Плод – трехгранная, трехгнездная коробочка. Опыляется пчелами и мухами. В Беларуси встречается 6 дикорастущих видов. В культуре наиболее широко распространен лук репчатый, выходец из Центральной Азии. Луковицы содержат 2 – 15 % сахаров, 12 – 16 мг% эфирного лукового масла, обладающего бактерицидным действием, инулин, фитин, сапонины, витамин С, соли калия, фосфора, железа. Большим набором витаминов отличаются листья. Сорта подразделяются на острые (Бессоновский), полуострые (Стригуновский) и сладкие (Ката и др.). Среди многочисленных сортов в Беларуси рекомендованы для производства Ветразь, Дыямент и Супра. Повсеместно в средней и центральной полосе Европы, на Кавказе, в азиатском регионе возделывается чеснок. Луковица состоит из многочисленных почек-зубков, сидящих на плоском донце. Листья плоские. Каждый последующий лист прорастает внутри трубки предыдущего и выходит выше него. В результате формируется ложный стебель высотой до 50 см. В соцветии часто образуются почки-детки. Плоды и семена, как правило, не образуются. В зубках чеснока содержится около 35% сухого вещества, до 27% углеводов, 8% белка, до 30 мг% витамина С, инулин и фитонциды. Наряду с луком репчатым широко используется в народной медицине. Ценным в Беларуси является озимый сорт Вiтажэнец. Пищевое значение имеют также лук поррей (A. porrum), лук батун (A. fistulosum), лук шаллот (A. ascalonicum), лук слизун (A. nutans). Из дикорастущих видов используются лук победный (Allium victorialis) и лук медвежий, или черемша (Allium ursinum). Деликатесной овощной культурой является спаржа (Asparagus). Это корневищные растения с сильно разветвленным стеблем. Листья очень мелкие, пленчато-чешуйчатые, при основании с мелкими колючками. Из пазух листьев выходят обычно многочисленные зеленые, большей частью игловидные веточки – кладодии, заменяющие листья. Растения двудомные раздельнополые. Околоцветник из 6 сросшихся листочков, тычинок 6, плод – ягода. В пищу употребляются отваренные молодые нежные этиолированные побеги, богатые углеводами, витаминами, аминокислотами. Невозможно переоценить декоративное значение лилейных. Важнейшими из них являются лилии (Lilium): лилия белая (L. candidum), желтоцветная лилия однобратственная (L. monodelphum), лилия королевская (L. regale), лилия кудрявая, или мартагон (L. martagon). Они имеют крупные нарядные цветы со стойким приятным запахом. Тюльпаны выращиваются не только в садах и парках. Они являются объектом цветочной индустрии в Нидерландах и других странах мира. Создано более 10 тысяч сортов, многие из которых относятся к виду Тюльпан Геснера (Tulipa gesneriana). Эффектными декоративными растениями являются эремурусы (Eremurus), у которых стрелка достигает 2 м высотой и в одном кистевидном соцветии собрано до 800 цветков красной и других окрасок. Из других декоративных лилейных можно отметить безвременник великолепный (Colchicum speciosum), краснодев желтый (Hemerocallis flava), рябчик Эдуарда (Fritillaria eduardii), кандык сибирский (Erythronium sibiricum), гиацинты (Hyacinthus), миниатюрные весенние пролески (Scilla), мускари (Muscari). К лекарственным относятся купена лекарственная (Polygonatum officinale), ландыш майский (Convallaria majalis), алоэ древовидное (Aloe arborescens) и др. Колхицин, добываемый из безвременника осеннего (Colchicum autumnale), используется в генетике для удвоения числа хромосом. В лесах и среди кустарников довольно часто встречаются корневищные ядовитые растения, одновременно и лекарственные: ландыш, купена, вороний глаз и другие. К ядовитым относятся также лук луговой (A. angulossum) и чемерица Лобеля (V. lobelianum). Они вызывают нервное возбуждение и нарушение сердечной деятельности. В Красную книгу Республики Беларусь занесено 8 видов охраняемых растений семейства Лилейные. Это тофильдия чашечковая (Tofieldia calyculata), лук медвежий (Allium ursinum), лук скорода (Allium schoenoprasum), безвременник осенний (Colchicum autumnale), гусиный лук луговой (Gagea pratensis), гусиный лук покрывальцевый (Gagea spathacea), тюльпан лесной (Tulipa sylvestris), лилия кудреватая (Lilium martagon).
Fritillaria ruthenica Wikstr. Семейство Лилейные Liliaceae.
Нарциссы Нарциссы
Лук жёлтый
Лилия Вороний
глаз
4.1.2. Семейство ОсоковыеСемейство Осоковые (Сурегасеае) является единственным представителем порядка Осокоцветные (Cyperales). Оно включает около 100 родов и свыше 4000 видов. Семейство Осоковые является космополитным. Оно широко распространено от экватора до высоких широт обоих полушарий. Хотя большинство видов осоковых сосредоточено в тропическом поясе, многие осоковые в холодном и умеренном поясе являются неотъемлемым компонентом луговой и болотной растительности. Во флоре Беларуси их насчитывается 93 вида из 15 родов. Подавляющее большинство осоковых является гигрофитами, произрастающими в избыточно влажных местах (камыш лесной – Scirpus sylvaticus). Вместе с тем осоковые встречаются в умеренно влажных лесах (осока лесная – Carex sylvatica, осока волосистая – C. pilosa), в степях и на сухих склонах (осока низкая – C. humilis), песчаных пустынях (осока вздутая – C. physodes). Осоковые – это большей частью многолетние корневищные, нередко очень крупные травы (камыш – Scirpus, папирус – Cyperus papirus) высотой до 1,5 – 5 м и диаметром до 7 см. В экваториальной Западной Африке встречается почти древовидный микродракоидес чешуйчатый (Microdracoides squamosa). На камнях и скалах у водопадов, и даже как эпифиты на коре деревьев, в Южной Америке произрастают кустарничковые цефалокарпусы (Cephalocarpus). Среди осоковых имеются и лианы, как, например, произрастающая во влажных тропических лесах склерия режущая (Scleria secans). Сравнительно немногие представители из родов сыть и камыш являются однолетниками. Корневая система образуется придаточными корнями, отходящими от нижней части стебля или от узлов корневищ. Стебли трехгранные (осоки), реже цилиндрические (камыш) или почти плоские, обычно выполнены паренхимной тканью и слабо дифференцированы на узлы и междоузлия. Узлы обычно сильно сближены у основания стебля, иногда расположены по всей его длине (склерия, меч-трава – Cladium). Стебли, как правило, олиственные, однако некоторые растения (болотница – Eleocharis) имеют безлистый стебель, у основания закрытый листовым влагалищем. Листья линейные или линейно-ланцетные с длинным замкнутым и редко открытым (колеохлоа – Coleochloa) влагалищем. В месте перехода влагалища в листовую пластинку может быть (осока, колеохлоа) язычок в виде узкой пластины или ресничек. У большинства осоковых листовые пластинки в поперечнике билатеральные, трехгранные или почти цилиндрические; с шероховатыми или острорежущими краями благодаря наличию зубчиков, минерализованных солями кальция и кремнеземом. Листорасположение, как правило, очередное трехрядное, реже двухрядное. Цветки осоковых собраны в разнообразные колоски, расположенные в пазухах листовидных прицветных листьев. Колоски образуют сложные соцветия – колосовидное, метельчатое, зонтиковидное, кистевидное или головчатое. Цветки мелкие, невзрачные, обоеполые или однополые. На основании строения колосков и цветков семейство Осоковые разделяют на 3 подсемейства –Сытевые (Cyperoideae), Ринхоспоровые (Rhynchosporoideae)и собственноОсоковые (Caricoideae).Сытевые характеризуются обоеполыми цветками, с околоцветником или без него, колоски обычно многоцветковые. Ринхоспоровые – обитатели теплых стран, имеют обычно обоеполые цветки, колоски мелкие, 1–3- цветковые. Подсемейство Осоковые отличается однопо-лыми цветками, ни имеющими околоцветника. Околоцветник или полностью атрофирован (сыть, осока), или сильно редуцирован и состоит из 6 (у камыша из 3) чешуй, или 3 – 14 (обычно 6) мелкозубчатых щетинок, или из 6 и более шелковистых волосков. В обоеполых и мужских цветках имеется 3 тычинки, очень редко 1 – 2, 6, 12, расположенные в один круг. Тычинки имеют длинные поникающие нити и удлиненные, вскрывающиеся пыльники. В процессе микроспорогенеза тетрада микроспор редуцируется. Она содержит лишь одно пыльцевое зерно, покрытое кроме интины и экзины оболочкой материнской клетки тетрады. В обоеполом и женском цветке гинецей состоит из 2 – 3 сросшихся плодолистиков. Гинецей может быть заключен в мешочек, состоящий из 2 сросшихся прицветников. Завязь верхняя, одногнездная, с одним семязачатком. Столбик пестика длинный, с 2 – 3 длинными рыльцевыми ветвями. Формулы цветков разнообразны. Например, у обоеполого цветка пушицы влагалищной – ↑ P∞A3G(3). У осоки пузырчатой женский цветок имеет формулу♀ ↑ P0A0G(3), а мужской –♂ ↑ P0A3G0. Плод ореховидный, нераскрывающийся, трехгранный, слегка двояковыпуклый или шаровидный с твердым перикарпием. Семена с маленьким зародышем, окруженным хорошо развитым крахмалистым или маслянистым эндоспермом. Строение соцветия является важным систематическим признаком, по которому осоки подразделяются на равноколосые и разноколосые. У равноколосых (осока лисья – C. vulpina) в верхней части колоса находятся мужские цветки, а в нижней – женские или наоборот (осока раздвинутая – C. remota). Разноколосые осоки имеют разобщенные мужские и женские колосовидные соцветия. При этом растения бывают однодомными (осока малоцветковая – C. pauciflora), когда мужские и женские колосья находятся на одном растении, и редко двудомными (осока двудомная – C. dioica), когда разные колосья расположены на разных растениях. Осоковые являются ветроопыляемыми растениями. Их размножение осуществляется семенами, которые распространяются ветром, водой, птицами, а также вегетативно – корневищами. В естественных фитоценозах осоковые являются местом гнездования птиц и обитания диких животных (камыш, пушица, осока, сыть, меч-трава), местом аккумуляции пресной воды (осока береговая – C. riparia). Пушица влагалищная (Eriophorum vaginatum), камыш озерный (Scirpus lacustris) наряду с другими видами принимают участие в торфообразовании. Длиннокорневищные виды, например осока песчаная (C. arenaria), являются хорошими закрепителями песков. Хозяйственное значение осок сравнительно невелико. В Древнем Египте сердцевина стеблей папируса (Cyperus papyrus) использовалась для изготовления писчего материала. Её разрезали на продольные полоски, складывали в два слоя – один поперек другого и сжимали до естественного их склеивания. Высушенные полоски полировали кусочками слоновой кости и склеивали в свитки шириной 20 – 30 см и длиной до 40 м. Клубневидные образования на корневищах болотницы сладкой (Eleocharis dulcis), сыти съедобной (чуфа, или земляной миндаль – Cyperus esculentus), клубнекамыша (Bolboschoenus) и сочные корневища тропических камышей и папируса используются в пищу. В частности чуфа возделывается в Испании и Италии ради небольших клубней, длиной до 3 см, которые содержат до 50% углеводов, 25% масла, 9% белка. Масло по качеству почти не уступает оливковому и ореховому. Камыш, фимбристилис (Fimbristyllis), папирус, ринхоспора (Rhynchospora) применяются в качестве строительного материала и для изготовления плетеных изделий. Камыш лесной, пушица влагалищная, осока вздутая и другие, виды кобрезии (Kobresia) используются на корм домашним и диким животным. Лекарственное значение имеют антибиотики болотницы сладкой и других осоковых. К ядовитым относится пушица узколистная (Eriophorum angustifolium), которая вызывает у животных поносы. Комнатным и оранжерейным декоративным растением является сыть очереднолистная (Cyperus alternifollius). Флора Беларуси богата осоками. Из 68 видов наиболее распространенными являются осока дернистая (C. cespitosa), осока сероватая (C. cinerea), осока заячья (C. ovalis), осока вздутая (C. rostpata), осока пузырчатая (C. vesicaria) и др. Многочисленны виды ситняга, или болотницы (Eleocharis) – 6 видов, пушицы (Eriophorum) – 4 вида. Из камышей (Scirpus) широкое распространение имеют камыш озерный (S. lacustris) и камыш лесной (S. sylvaticus). На территории Беларуси произрастают редкие и исчезающие виды осоковых. В Красную книгу Республики Беларусь занесены меч-трава обыкновенная (Cladium mariscus), осока Дэвелла (C. davalliana), осока Буксбаума (C. buxbaumii), осока волосовидная (C.capillaris), осока птиценожковая (C. ornithopoda), осока приземистая (C. supina), осока повислая (C. flacca), осока Отрубы (C. otrubae), осока малоцветковая (C. pauciflora), осока заливная (C. paupercula), осока корневищная (C. rhizina), осока теневая (C. umbrosa). Литература: 1, 5 (с. 292 – 310), 6 (с. 508 – 510), 8 – 12, 14 (с. 443 – 447), 15 (с. 477 – 483), 17.^ 4.1.3. Семейство МятликовыеСемейство Мятликовые (Роасеае), или Злаковые (Gramineae), относится к порядку Мятликоцветные (Poales). Это одно из наиболее крупных семейств цветковых растений, насчитывающее до 700 родов и 10000 видов. В естественной флоре Беларуси отмечено около 150 видов из 70 родов. Мятликовые равномерно распространены по всей суше Земного шара. В тропическом поясе произрастает примерно столько же их видов, сколько в странах с умеренным климатом, а в Арктических широтах они по видовому составу занимают первое место среди других семейств. Мятликовые играют важнейшую роль в формировании растительного покрова лугов, степей, прерий и саванн. Для равнинных лугов характерны мятлик (Poa), овсяница (Festuca), тимофеевка (Phleum), кострец (Bromopsis), трясунка (Briza). В степях распространены ковыль (Stipa), типчак (Festuca valesiaca), житняк (Agropyron). В прериях первостепенная роль принадлежит хлорису (Chloris), бизоновой траве (Buchloё dactyloides). На подвижных песках обитают селин (Stipagrostis) и песколюбка (Ammophila). Велики роль и разнообразие мятликовых в лесах и на торфяниках. Широкое распространение мятликовых связано со строением их вегетативных и репродуктивных органов, физиологическими и биохимическими особенностями, которые позволяют растениям иметь очень высокую приспособляемость к разнообразным экологическим условиям. В семействе встречаются типичные мезофиты – это почти все культивируемые злаки: рожь (Secale), пшеница (Triticum), ячмень (Hordeum), кукуруза (Mais), овес (Avena) и другие, гигрофиты – рис (Oriza), манник (Glyceria), а также ксерофиты – белоус торчащий (Nardus stricta), овсяница овечья (Festuca ovina), ковыли, житняки и др. А тростник обыкновенный (Phragmites australis) может произрастать и на болотах различных типов, и на горных склонах. Представители семейства – это многолетние (овсяница луговая – Festuca pratensis), двулетние (лисохвост коленчатый – Alopecurus geniculatus) и однолетние травы (канареечник канарский – Phalaris canariensis, полевые культурные злаки), реже древовидные растения (бамбук – Bambusa). У близкородственной бамбуку динохлои (Dinochloa) известны колючие лианообразные виды. Корневая система у мятликовых мочковатая при семенном и вегетативном размножении. Главный корень образуется из зародышевого корешка, но рано завершает свое развитие. Ему на смену развиваются многочисленные, хорошо развитые придаточные корни. Стебель – соломина с выраженными узлами и полыми или заполненными сердцевиной междоузлиями. Признаки анатомического строения стебля используются в систематике злаков. Так, для большинства внетропических видов (овсяница, пшеница, рожь) характерны междоузлия с широкой полостью и расположением проводящих пучков в два круга, причем более мелкие пучки находятся во внешнем круге и обслуживают остатки первичной коры. У тропических видов (кукуруза, просо, сорго) междоузлия с маленькой полостью или заполнены паренхимой. В основаниях междоузлий локализована интеркалярная меристема. Здесь же могут закладываться зачаточные бугорки боковых пазушных почек. Однако зачатки надземные почек не развиваются и боковые побеги из них не образуются. Поэтому для мятликовых характерно кущение, т.е. формирование боковых побегов из почек подземной части стебля. Кущение может быть плотным (белоус торчащий – Nardus stricta), рыхлым (пшеница мягкая – Triticum aestivum) и корневищным (пырей ползучий – Elytrigia repens). Листья состоят из охватывающего стебель влагалища, которое обычно открытое, линейной листовой пластинки, язычка – перепончатого выроста, расположенного в месте соединения влагалища и листовой пластинки. Иногда язычок отсутствует или вместо него развивается ряд волосков. У некоторых злаков (ячмень – Hordeum) края влагалища в месте отгиба образуют двусторонние линейные выросты – ушки. В эпидермисе листа имеются моторные клетки, обеспечивающие сворачивание листа в жаркую пору. У многих форм вокруг проводящих пучков располагается паренхимная обкладка, участвующая в фотосинтезе. Листорасположение двухрядное очередное. Многие мятликовые являются монокарпическими растениями, т.е. цветут и плодоносят один раз в жизни (культурные полевые злаки, бамбуки с одревесневающими стеблями). Большинство видов имеют обоеполые цветки. В случае раздельнополости цветков растения могут быть как однодомными (кукуруза – Zea), так и двудомными (пампасская трава – Cortaderia selloana). Цветки с пленчатым околоцветником собраны в элементарные соцветия – колоски, которые в свою очередь образуют сложные соцветия: колос (рожь, пшеница, ячмень), сложную кисть, или метелку (овес – Avena, просо – Panicum), султан (тимофеевка – Phleum, лисохвост – Alopecurus). У кукурузы женские цветки собраны в початок, а мужские – в метелку. У основания колоска располагаются колосковые чешуи. У плевела (Lolium) она одна. У пшеницы, ржи и многих других растений колосок несет две колосковые чешуи – верхнюю и нижнюю; у проса, риса (Oryza), душистого колоска (Anthoxanthum) и других их более двух. В строении колосковых чешуй выделяют киль, килевой зубец и плечо, размеры и форма которых учитываются при определении видов и сортов злаков. Колосок состоит из оси, на которой находится либо один (ячмень), либо несколько цветков (пшеница), расположенных поочередно двумя рядами. Цветки в колосках образуются в пазухах нижних цветковых чешуй листового происхождения. Килевые зубцы нижних чешуй могут быть короткими или развиваются в ость у остистых форм. У персидской пшеницы (Triticum persicum) ости образуются как на цветковой, так и на колосковой чешуе. Напротив нижней чешуи в цветке находится двухкилевая верхняя цветковая чешуя, образованная двумя сросшимися листочками наружного круга околоцветника. Внутренний круг околоцветника обычно представлен двумя пленочками, или лодикулами. У бамбука и ковыля их три. Во время цветения в лодикулы активно поступает вода; они увеличиваются в размерах и раздвигают нижнюю и верхнюю цветочные чешуи, обеспечивая раскрывание цветков. У большинства мятликовых в цветке три тычинки, но их может быть шесть (рис, бамбук, сахарный тростник – Saccharum officinarum), две у душистого колоска и даже одна (цинна – Cinna). Пестик образован двумя сросшимися плодолистиками, столбик с двураздельным перистым рыльцем. Завязь верхняя. Формула цветка: ↑Р(2)+2А3G(2). Плод сухой, невскрывающийся, односеменной, называется зерновкой и имеет тонкий околоплодник плотно прилегающий к семенной оболочке. Нередко у пленчатых видов пшеницы, ячменя, овса и других растений при созревании зерновки ее околоплодник слипается с плотно прилегающими к нему цветковыми чешуями. В отличие от них у голозерных форм (рожь, пшеница мягкая и др.) семена не слипаются с чешуями. Зародыш семени имеет одну развитую семядолю – щиток, который граничит с эндоспермом. Вторая семядоля – эпибласт, редуцирована. Зародышевая почечка окружена листовидным образованием, именуемым колеоптиле, которое защищает всходы при выходе их на поверхность почвы. Проростковые корни защищены корневым влагалищем, или колеоризой. Эндосперм семени хорошо развит, богат крахмалом и белком. Семена имеют разную продолжительность послеуборочного дозревания. У озимых форм она короткая, всходы могут давать даже свежеубранные семена, у яровых – длинная, до нескольких месяцев. Некоторые виды, включая отдельные сорта пшеницы, являются «двуручками», т.е. могут давать всходы как при осеннем, так и при весеннем посеве. Большинство злаков является ветроопыляемыми растениями. Это кукуруза, рожь, сорго, суданская трава, тимофеевка, кострец, овсяница и другие. К факультативным, т.е. необязательным, самоопылителям относится пшеница, эгилопс (Aegilops) и др. Для хазмогамных (открытоцветущих) ветроопыляемых злаков большое значение имеет суточная ритмика цветения растений. Совпадение времени раскрывания цветков обеспечивает успех перекрестного опыления в популяции. Более строгое, аблигатное, самоопыление характерно для клейстогамных (закрытоцветущих) ячменя и овса. Апомиктическое размножение, при котором семена образуются без слияния половых клеток, встречается у тропических просовых и сорговых злаков. Реже у мятликовых наблюдается вивипария. Так, у арктических видов мятлика (Poa), овсяницы (Festuca), щучки (Deschampsia) размножение происходит луковичкообразными почками, которые образуются в соцветии из колосков. С учетом многообразия анатомо-морфологических признаков вегетативных и репродуктивных органов семейство Мятликовые подразделяется на подсемейства. В многотомнике «Жизнь растений» [5] выделяется шесть подсемейств: Бамбуковые (Bambucoideae), Рисовые (Oryzoideae), Мятликовые (Pooideae), Тростниковые (Arundinoideae), Полевичковые (Eragrostideae), Просовые (Panicoideae).^ Подсемейство Бамбуковыеобразовано высокорослыми тропи-ческими и субтропическими одревесневающими (арундинария – Arundinaria, бамбук, дендрокаламус – Dendrocalamus) и травянистыми растениями (маклюролира – Maclurolyra, пирезия – Piresia). Бамбуковые широко распространены в Африке, Центральной и Южной Америке, Юго-Восточной Азии, где имеют пищевое и техническое значение.^ Подсемейство Рисовыеобъединяет травянистые одно- и многолетние самоопыляющиеся растения. Для них характерны сплюснутые с боков колоски с одним развитым цветком. Колосковых чешуй четыре, лодикул две, тычинок шесть. Соцветие – однобокая, раскидистая или сжатая метелка. Важнейшей культурой в мировом земледелии является рис посевной (Oryza sativa), используемый для изготовления крупы, муки, спиртных напитков, бумаги и плетеных изделий. Кподсемейству Мятликовыепринадлежит большинство внетропических травянистых одно- и многолетних злаков. Многие из них имеют пленчатые язычки листьев. Одно- и многоцветковые колоски собраны в сложный колос (пшеница), метелку (овес), султан (тимофеевка). Колосковых чешуй и лодикул две, тычинок три. Сюда относятся хлебные, крупяные, а также кормовые травы. Род Пшеница (Triticum) насчитывает около 30 видов. В культуре наиболее распространены два вида: пшеница мягкая (Т. aestivum), которая отличается полым стеблем под соцветием и рыхлым длинным колосом с растопыренными остями, семенами с мучнистым эндоспермом, а также пшеница твердая (Т. durum), у которой стебель под соцветием выполненный, колос сжат с боков, плотный, ости прямые, семя со стекловидным эндоспермом. Ценными в условиях Беларуси сортами мягкой озимой пшеницы являются Завет, Спектр, Прэм’ера и др. Зерно пшеницы содержит много клейковинообразующих белков и поэтому является основной хлебной культурой. Используется также для изготовления круп, крахмала, спирта. У ржи посевной (Secale cereale) соцветие – неломкий остистый сложный колос, колоски двухцветковые, часто с зачатком третьего цветка. Отличается высокой зимостойкостью и неприхотливостью к почвам, поэтому широко возделывается в Беларуси. Ценными сортами являются Талисман и Зарница. Используется на пищевые и кормовые цели. В результате межродовой гибридизации пшеницы и ржи получены амфидиплоиды тритикале, сорта которого имеют хорошую перспективу использования в сельском хозяйстве. Род Ячмень (Hordeum) имеет плотный ломкий остистый колос, колоски одноцветковые, сидят по три на уступе членика колосового стержня. Листья с хорошо развитыми ушками. У ячменя обыкновенного (Н. vulgare) колос широкий, 4 – 6-гранный, все колоски плодущие. У ячменя двурядного (Н.distichon) колос линейный, плоский, средний колосок плодущий, а боковые бесплодные. Высокой продуктивностью отличаются сорта ярового ячменя Стратус, Якуб, Сонор. Из зерна делают крупы, пиво, корма для животных. Овес посевной (Avena sativa) однолетник, листья с длинным язычком, соцветие метелка, колоски крупные, 2 – 3-цветковые. Имеются пленчатые и голозерные формы. Овсяная крупа ценится как диетический продукт. Зерно используется на корм животным. Луговыми травами подсемейства Мятликовые являются кострец, мятлик, овсяница, тимофеевка и другие, для степей характерны ковыли. Дляподсемейства Тростниковые характерны высокорослые, многолетние травянистые растения с многоузловыми выполненными стеблями, сильно развитыми корневищами и многоцветковыми колосками. Их можно встретить на лесных полянах и болотистых лугах. Наибольшим видовым многообразием выделяются страны с теплым климатом. К тростниковым относятся тростник обыкновенный (Phragmites australis), пампасская трава (Cortaderia) Вподсемействе Полевичковыепреобладают травянистые растения, выходцы из засушливых районов тропиков. В южных регионах Беларуси изредка встречаются сорные виды полевички (Eragrostis), на солончаках Северной Африки обычна прибрежница (Aeluropus), в североамериканских прериях – дистихлис (Distichlis).^ Подсемейство Просовыеявляется наиболее высокоспециализированным в семействе. Колоски обычно двуцветковые, расположены по одному или группами по 2-3 на колосовидных веточках. В подсемейство входят просо посевное (Panicum miliaceum), ежовник, или куриное просо (Echinochloa), ежовник хлебный (E. frumentaceae), сорго (Sorghum), сахарный тростник (Saccharum officinarum), а также важнейшая пищевая и кормовая культура мирового земледелия – кукуруза (Zea mays). Зерно кукурузы перерабатывают на крупу, муку, крахмал, патоку, спирт и другие продукты. В условиях Беларуси для полного созревания зерна не хватает тепла, поэтому кукурузу выращивают на корм скоту. Для этих целей используются гетерозисные гибриды кукурузы Бемо 182 СВ, Полесский 212 СВ, Краснодарский 194 МВ и многие другие. Семейство Мятликовые имеет исключительно большое хозяйственное значение. Им принадлежит выдающаяся роль в формировании оседлого образа жизни многих народов мира в связи с переходом от сбора семян злаковых растений к их выращиванию и хлебопечению. Основными хлебными культурами в Беларуси являются мягкая пшеница и рожь. Крупу получают из ячменя, овса, проса, риса, кукурузы. Зубровка (Hierochloё) и душистый колосок (Anthoxanthum) содержат ароматические вещества, используемые в парфюмерии, пищевой промышленности и медицине. К ядовитым относится манник водный (Glyceria aquatica), который в свежем виде вызывает у животных судороги и нарушение сердечной деятельности. Многие злаковые травы являются основными компонентами естественных сенокосов и пастбищ и введены в культуру. Это тимофеевка луговая (Phleum pratensis), ежа сборная (Dactylis glomerata), овсяница луговая (Festuca pratensis), полевица гигантская (Agrostis gigantea), костер безостый (Bromus inermis), плевел многолетний (Lolium perenne), лисохвост луговой (Alopecurus pratensis), мятлик луговой (Роа pratensis), райграс высокий (Arrhenatherum elatius) и др. Овсяница красная (F. rubra), райграс пастбищный (Lolium perenne) и другие применяются в декоративном садоводстве в качестве газонной травы. Для закрепления подвижных песков используют песколюбку песчаную (Ammophila arenaria) и колосняк песчаный (Leymus arenarius). Многие злаки могут использоваться в качестве строительного материала. В тропических странах в качестве водопроводных труб применяют прочные и легкие стебли бамбуковых. Бумагу высокого качества изготавливают из ковыля и риса. Среди злаков имеется немало сорных растений. Это пырей ползучий (Elytrigia repens), костер ржаной (Bromus secalinus), ежовник (куриное просо), мятлик однолетний (Poa annua), некоторые виды щетинника (Setaria). Сорняками на лугах являются малоценные в кормовом отношении луговик дернистый, или щучка (Deschampsia cespitosa), и белоус торчащий (Nardus stricta). ЛИТЕРАТУРА 1. Травянистые растения СССР. Т. 1. / Ю.Е. Алексеев, В.Н. Вехов, Г.П. Гапочка и др. М.: Мысль, 1971. 487 с.: ил. 2. Травянистые растения СССР. Т. 2. / Ю.Е. Алексеев, В.Н. Вехов, Г.П. Гапочка и др. М.: Мысль, 1971. 309 с.: ил. 3. Жизнь растений. В 6 т. / Гл. ред. А.Л. Тахтаджян. Т. 5 (1). Цветковые растения / под ред. А.Л. Тахтаджяна. М.: Просвещение, 1980. 432 с.: ил. 4. Жизнь растений: В 6 т. / Гл. ред. А.Л. Тахтаджян. Т. 5 (2). Цветковые растения / под ред. А.Л. Тахтаджяна. М.: Просвещение, 1981. 512 с.: ил. 5. Жизнь растений: В 6 т. / Гл. ред. А.Л. Тахтаджян. Т. 6. Цветковые растения / под ред. А.Л. Тахтаджяна. М.: Просвещение, 1982. 544 с.: ил. |

 Лилейник
красоднев
Лилейник
красоднев РЯБЧИК
РУССКИЙ Красная Книга Челябинской
области
РЯБЧИК
РУССКИЙ Красная Книга Челябинской
области Ландыш
Ландыш
 ^
^


studfiles.net
Цветковые растения — Википедия
Цветко́вые расте́ния, или Покры́тосеменны́е, устар. Скрытносемянные[1] (лат. Magnoliophyta, или Angiospermae от др.-греч. ἀγγεῖον — сосуд, σπέρμα — семя) — отдел высших растений, отличительной особенностью которых является наличие цветка в качестве органа полового размножения и замкнутого вместилища у семяпочки (а затем и у происшедшего из неё семени, откуда и появилось название покрытосеменные). Ещё одна существенная особенность цветковых растений — двойное оплодотворение.
Покрытосеменные, наряду с голосеменными (Gymnospermae) составляют одну из двух групп семенных растений (Spermatophytae).
В 1999 году Амборелловые (Amborellaceae) были идентифицированы как базальная группа цветковых растений[2].
Объём таксона
По числу видов цветковые растения превосходят все остальные группы высших растений.
В литературе приводятся разные данные о числе современных видов цветковых растений. В 2009 году была опубликована работа австралийского учёного Чапмена (A. D. Chapman) «Numbers of Living Species in Australia and the World», в которой он даёт обзор мнений по этому вопросу и делает вывод, что по состоянию на 2009 год можно оценить общее число описанных современных видов цветковых растений примерно в 269 тысяч, а общее число современных видов цветковых растений на нашей планете — примерно в 350 тысяч[3].
Число видов покрытосеменных, по данным Angiosperm Phylogeny Website на февраль 2010 года, составляет 271—272 тысячи, число родов — 13350—13400[4]. В базе данных The Plant List (версия 1.1, 2013) содержится 304 419 признанных (то есть со статусом Accepted) видовых названий цветковых растений, принадлежащих к 405 семействам и 14 559 родам, при этом общее количество видов цветковых растений оценивается в 352 000[5].
Число семейств и порядков отличается в зависимости от классификаций; Система классификации APG III (2009) выделяет 414 семейств, которые, в свою очередь, объединены в 59 порядков.
Видео по теме
Морфологические особенности
Важнейшая особенность цветковых растений — наличие специализированного генеративного органа — цветка, берущего на себя функции полового размножения и привлечения агентов опыления. Цветковые растения заключают свои семязачатки (семяпочки) в полость завязи, которая образована срастанием открытого плодолистика. Стенки завязи после оплодотворения разрастаются и видоизменяются, давая образование под названием плод.
В другой группе семенных растений, у голосеменных (Pinophyta, или Gymnospermae), семязачаток не скрыт от опыления, а семена не заключены в истинный плод, но иногда семя могут покрывать мясистые структуры, например, у представителей рода Тис.
Происхождение
Амборелла волосистоножковая (Amborella trichopoda), эндемик Новой Каледонии, одно из наиболее древних цветковых растений, сохранившихся до наших дней. Отделившись от других цветковых растений 130 миллионов лет назад, амборелла с тех пор изменилась весьма незначительно, что можно объяснить относительной изолированностью экосистемы Новой КаледонииПервые остатки покрытосеменных датируются меловым периодом приблизительно 140 миллионов лет назад. Предки покрытосеменных и гнетовых дивергировали в триасе (220—202 миллионов лет назад). Первые отпечатки растений с признаками покрытосеменных обнаружены в пластах юрского и раннего мелового периодов (135—65 миллионов лет назад), но это были довольно-таки малочисленные и примитивные формы. Древнейшими покрытосеменными являются растения из группы нимфейных[6]. Следы широкого развития и распространения покрытосеменных появились в палеонтологической летописи в период среднего мела (около 100 миллионов лет назад). Но уже в позднем мелу покрытосеменные оказались доминирующей формой растительной жизни, и во многих фоссилиях узнаются представители современных семейств (например, бук, дуб, клён и магнолия).
Одно из важнейших направлений эволюции растительного царства — приспособление к изменчивым условиям наземной жизни. Цветковые растения являются ярчайшим выражением этой линии и доминируют на земной поверхности в данную эпоху.
К широчайшему географическому разнообразию прибавляется разнообразие форм и способов роста. Банальная ряска, покрывающая поверхность пруда, представляет собой крошечный зелёный побег с простым корешком, вертикально погружённым в воду, и с очень нечёткими листиками и частями стебля. Могучее лесное дерево столетия развивало свою сложную систему стволов и ветвей, покрытых бесчисленными веточками и листвой, а под землёй соответствующую площадь занимает мощная, хорошо развитая корневая система. Между этими двумя крайностями — бесконечные градации: водные и земные травы, ползучие, прямостоящие или карабкающиеся, кусты и деревья, гораздо большее разнообразие, чем в другом отделе семенных растений — Gymnospermae.
Известны многочисленные водные покрытосеменные растения, они в изобилии встречаются в долинах рек и чистых озёрах, в меньшем количестве — в солёных озёрах и морях. Однако такие водные покрытосеменные не являются примитивными формами, а возникли путём приспособления наземного предка к водной среде.
Систематическое положение
Цветковые растения обычно рассматриваются как отдел. Так как эта систематическая категория более высокого ранга, чем семейство, есть определённая свобода в выборе названия. Статья 16 Международного Кодекса Ботанической Номенклатуры позволяет использовать как и традиционные исторические названия, так и название, образованное от рода. Официальное униноминальное название этого таксона — Magnoliophyta, от названия рода Magnolia. Но традиционно укоренились такие имена, как Angiospermae и Anthophyta (цветковые растения).
Классификация
Информация в этой статье или некоторых её разделах устарела. Вы можете помочь проекту, обновив её и убрав после этого данный шаблон. |
История понятия
Ботанический термин «Angiospermae» предложил Пауль Герман в 1690 году, термин был составлен из греческих слов ἀγγεῖον (вместилище, сосуд, полость) и σπέρμα (семя). Так Герман называл один из главнейших отделов растительного царства, включавший в себя растения, обладающие заключёнными в капсулы семенами. Gymnospermae же по Герману являлись цветковыми растениями, чей шизокарпичный или единственный цельный плод считались семенами с отсутствующими покровами. Сам термин и его антоним были подхвачены Карлом Линнеем, который использовал их в похожем, но более ограниченном смысле — для названий порядков своего класса Двусильных (Didynamia). В своём современном значении эти термины начали использоваться после того как Роберт Броун в 1827 году установил существование истинно голых семязачатков у Cycadeae и Coniferae, присвоив им название «Gymnospermae». С этого времени термин «Angiospermae» стал использоваться различными авторами, иногда с разными вариациями, для обозначения одной из подгрупп в пределах двудольных растений (разделение «явнобрачных» растений на бессемядольные, однодольные и двудольные получило широкое распространение несколько раньше).
Однако, после того как Вильгельм Хофмейстер описал процессы, происходящие в зародышевом мешке цветковых растений (1851 год), и сопоставил их с оплодотворением тайнобрачных, стало ясно, что Gymnospermae представляют собой группу, совершенно отличную от Angiospermae. В результате понятие «покрытосеменные» постепенно стали рассматривать как синоним понятия «цветковые», и, соответственно, двудольные (Magnoliopsida, или Dicotyledones) и однодольные (Liliopsida, или Monocotyledones) — как подгруппы в составе Angiospermae. В этом значении понятие «покрытосеменные» (Angiospermae) используется и по сей день.
Благодаря постоянному пересмотру взглядов на родство цветковых растений, внутренняя систематика этой группы подвергалась и подвергается изменениям. Две широко используемые, хотя и несколько устаревшие, системы цветковых растений — система Тахтаджяна и система Кронквиста, не отражают филогению таксона. Таким образом, классификация цветковых растений сейчас активно дорабатывается и исправляется.
Отдел цветковых растений традиционно подразделяется на 2 класса — Magnoliopsida (двудольные) от названия рода Magnolia и Liliopsida (однодольные) от названия рода Lilium. Используются также и традиционные названия этих таксонов — Dicotyledones и Monocotyledones. Происхождение этих названий связано с тем, что представители Dicotyledones обычно имеют две семядоли в семени (у некоторых видов может быть одна, три или четыре), тогда как у представителей Monocotyledones семядоля всегда одна.
Имеется целый ряд современных систем классификации цветковых растений. Классы двудольных и однодольных могут подразделяться на подклассы, те, в свою очередь, на порядки (иногда объединяемые в надпорядки), семейства, роды и виды со всеми промежуточными категориями.
Класс Двудольные
Информация в этой статье или некоторых её разделах устарела. Вы можете помочь проекту, обновив её и убрав после этого данный шаблон. |
В классе Двудольные (Magnoliopsida, или Dicotyledones) описано 6 подклассов, 128 порядков, 418 семейств, приблизительно 10 000 родов и около 199 000 видов[7] растений.
Выделяют подклассы (клады) [источник не указан 3059 дней]:
Класс Однодольные
Информация в этой статье или некоторых её разделах устарела. Вы можете помочь проекту, обновив её и убрав после этого данный шаблон. |
В класс однодольных растений (Liliopsida, или Monocotyledones) включаются 5 подклассов, 37 порядков, около 125 семейств, более 3000 родов и около 59 000 видов[8].
Выделяют подклассы[источник не указан 3059 дней]:
Филогения
Предполагается, что Однодольные эволюционно более прогрессивны, чем двудольные[источник не указан 1122 дня].
Реликтовые представители покрытосеменных
Некоторые из наиболее примитивных представителей ныне живущих двудольных сохранили архаичное строение проводящей системы, тычинок, пыльцевых зёрен, плодолистиков и других органов.
Часть видов семейства Винтеровые (Winteraceae), роды троходендрон (Trochodendron) и тетрацентрон (Tetracentron), по строению проводящей системы мало чем отличаются от примитивных представителей голосеменных. У них отсутствуют проводящие сосуды во всех органах, их роль выполняют трахеиды.
Другие демонстрируют архаичное строение цветка, в частности примитивные строение тычинок; особенно заметное у таких растений, как дегенерия (Degeneria vitiensis), гальбулимима (Galbulimima) и ряда представителей магнолиевых и винтеровых[9].
Настоящими «живыми ископаемыми» являются представители родов амборелла (Amborella), дегенерия (Degeneria), австробэйлия (Austrobaileya), буббия (Bubbia), эвпоматия (Eupomatia)[10].
См. также
Примечания
- ↑ Бекетов А. Н. Голосеменные // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- ↑ Ботаника. Учебник для вузов: в 4 тт. = Lehrbuch der Botanik für Hochschulen. Begründet von E. Strasburger, F. Noll, H. Schenck, A. F. W. Schimper. / 35. Auflage neubearbeitet von Peter Sitte, Elmar W. Weiler, Joachim W. Kadereit, Andreas Bresinsky, Christian Körner / П. Зитте, Э. В. Вайлер, Й. В. Кадерайт, А. Брезински, К. Кёрнер; на основе учебника Э. Страсбургера [и др.]; пер. с нем. Е. Б. Поспеловой, К. Л. Тарасова, Н. В. Хмелевской. — М.: Издательский центр «Академия», 2007. — Т. 3. Эволюция и систематика / под ред. А. К. Тимонина, И. И. Сидоровой. — С. 15. — 576 с. — 3000 экз. — ISBN 978-5-7695-2741-8 (рус.). ISBN 978-5-7695-2746-3 (Т. 3) (рус.), ISBN 3-8274-1010-X (Elsevier GmbH) — УДК 58(075.8)
- ↑ A. D. Chapman. Numbers of Living Species in Australia and the World. 2nd edition (англ.) (Проверено 29 мая 2010)
- ↑ Angiosperm Phylogeny Website (англ.). Число родов и видов приведено отдельно для каждого порядка. См. также Семейства покрытосеменных (APG III).
- ↑ The Angiosperms (Flowering plants) // The Plant List (2013). Version 1.1. (англ.) (лат.) (Проверено 25 июля 2015)
- ↑ Происхождение основных групп наземных растений
- ↑ Hamilton, Alan & Hamilton, Patrick (2006), Plant conservation : an ecosystem approach, L.: Earthscan, сс. 2, ISBN 978-1-84407-083-1, <https://books.google.com/?id=P6m0OTheY8sC&printsec=frontcover#PPA2,M1>
- ↑ http://cmsdata.iucn.org/downloads/2008rl_stats_tables_all.xls
- ↑ Фёдоров А. А., Тахтаджян А. Л. и др. Жизнь растений. Том 5(1). Цветковые растения — М.: Просвещение, 1980. — с. 117—127.
- ↑ Смирнов А. В. Мир растений. Рассказы о саксауле, селитрянке, баобабе, березах, кактусах, капусте, банксиях, молочаях и многих других широко известных и редких цветковых растениях. — М.: Молодая гвардия, 1979. — с. 14-18
Литература
- Abstract • Полный текст: HTML, PDF (англ.) (Проверено 24 мая 2010)
Ссылки
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| •• Angiosperm Phylogeny Group (в том числе Бремер Б., Бремер К., Ривил Д., Чейз М.) • APG I • APG II (+список) • APG III (+список) •• |
wikipedia.green
