
Расположение генов в хромосомах
Задача 3. При скрещивании самки дрозофилы, дигетерозиготной по генам «А» и «В», с
рецессивным самцом в потомстве произошло расщепление по фенотипу:
АВ : Ав : аВ : ав = 25 % : 25 % : 25 % : 25 % .
При каком расположении генов в хромосомах может наблюдаться указанное фенотипическое расщепление?
Ход рассуждения: В задаче два фенотипических признака (или две пары контрастных – альтернативных признаков), каждый из которых в свою очередь имеет два варианта – доминантный и рецессивный. В представлении фенотипического расщепления буквой обозначается не ген (аллель), а именно признак. При написании генотипов такими же буквами обозначаются не сами признаки, а определяющие их аллели соответствующих генов.
Указанное в условии фенотипическое расщепление возможно в двух случаях:
— Практически: если гены расположены в разных хромосомах и расщепление происходит в соответствии с менделеевским законом независимого комбинирования признаков по типу анализирующего скрещивания. Представим схему скрещивания с изображением хромосом и указанием расположения в них аллелей генов.
Генотип самки: А__ В
а в Генотип самца: __а__ в
__а__ в
Гаметы: ♀
♂
100%
25% 25% 25% 25%
Расщепление АВ Ав аВ ав
по фенотипу:
25% 25% 25% 25%
Если соединить гаметы 4-х сортов самки с одинаковыми гаметами самца получиться потомство 4-х фенотипических классов поровну (по25%), как если бы гены находились в разных хромосомах.
— Теоретически такое же расщепление может получиться и в случае, когда оба гена сцеплены, то есть находятся в одной хромосоме, но только если расстояние между ними будет равняться 50 морганидам. Тогда и два сорта кроссоверных гамет, и два сорта некроссоверных гамет будут образовываться поровну – по 25%.
Полное сцепление аутосомных генов
Задача 4. Катаракта и полидактилия у человека обусловлены доминантными аутосомными тесно сцепленными (нет кроссинговера) генами. Однако сцепленными могут быть необязательно гены указанных аномалий, но и ген катаракты с геном
нормального строения кисти и наоборот.
Женщина унаследовала катаракту от своей матери, а полидактилию – от отца. У
её мужа оба признака нормальны.
Чего скорее можно ожидать у их детей: одновременного появления катаракты и
полидактилии, отсутствия обоих этих заболеваний или наличия только одной
аномалии – катаракты или полидактилии?
Ход рассуждения: Фактически от родителей дети получают не отдельные гены, а хромосомы – по одной из каждой пары. По условию задачи женщина унаследовала от своей матери только катаракту, а полидактилию — от отца. Следовательно, доминантные аллели, определяющие эти заболевания, находятся в разных гомологах этой пары хромосом, то есть, имеет место ТРАНС-положение. Поскольку гены обоих заболеваний проявили полное сцепление, кроссинговера не происходит, у женщины образуется два сорта гамет.
Образец оформления решения задачи:
Дано:
|
Ген ( аллель) и определяемый им признак |
Схема брака, потомство | |
|
А — Аллель катаракты (помутнение хрусталика глаза) а — Аллель нормы хрусталика В – аллель полидактилии (шестипалости) в — аллель нормального строения кистей рук Гены сцеплены полно Фенотипы в F1 = ? |
| |
Неполное сцепление аутосомных генов
Определение расстояния
Задача 5. У мух дрозофил признаки окраски тела и формы крыльев сцеплены. Темная окраска тела рецессивна по отношению к серой, а короткие крылья – к длинным. В лаборатории скрещивались серые длиннокрылые самки, гетерозиготные по обоим генам, с самцами, имеющими чёрное тело и короткие крылья. В потомстве оказалось:
|
Серых длиннокрылых особей – 1394; |
Чёрных короткокрылых – 1418; |
|
Чёрных длиннокрылых –287; |
Серых короткокрылых- 288 |
Определите расстояние между генами.
Ход рассуждения:
В потомстве наблюдается 4 фенотипически различающихся группы особей. При этом две многочисленные группы – некроссоверное потомство, а две малочисленные группы составляют особи, развившиеся в результате оплодотворения кроссоверных гамет самок сперматозоидами самцов. Это – кроссоверное потомство.
При вычислении расстояния между генами в морганидах по результатам анализирующего скрещивания используется формула:
Х
=
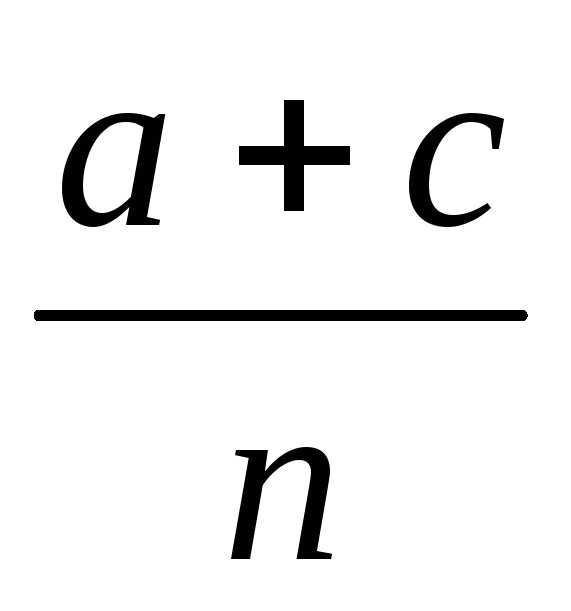 * 100%,Где а
– количество в потомстве кроссоверных
форм 1-й группы; с
– количество
кроссоверных форм 2-й группы; n
– общее
количество потомков; х — % кроссинговера
между ними. Вводим в формулу количественные
данные из условия задачи:
* 100%,Где а
– количество в потомстве кроссоверных
форм 1-й группы; с
– количество
кроссоверных форм 2-й группы; n
– общее
количество потомков; х — % кроссинговера
между ними. Вводим в формулу количественные
данные из условия задачи:
Х = ~ 17% .
Ответ: расстояние между генами составляет 17%
Определение вероятности признака у потомства
Задача 6. У человека локус резус-фактора сцеплен с локусом, определяющим форму эритроцитов, и находится от него на расстоянии 3-х морганид. Резус-положительность и эллиптоцитоз определяются доминантными аутосомными генами.
Жена гетерозиготна по обоим признакам (парам аллелей). При этом резус-положительность она унаследовала от одного родителя, эллиптоцитоз – от другого. Муж резус-отрицателен и имеет нормальные эритроциты.
Определите возможные генотипы и фенотипы у их детей (раздельно – у мальчиков и девочек). Вычислите вероятность рождения детей с каждым гено- и фенотипическим вариантом.
Решение:
Поскольку жена доминантные аллели обоих генов получила от разных родителей, имеет место исходное – ТРАНС-положение их в хромосоме.
|
R- Rh+ r Rh- А- элиптоцитоз а — норма R A 3 морганиды F —? |
Rh+, элиптоцитоз Rh-, норма Р♀ R r х ♂ r r а А ХХ а а ХY F
Ответ: Вероятные генотипы и фенотипы девочек (верхний ряд) и мальчиков (нижний ряд). По 24, 25% будут девочки и мальчики с Rh+ с нормальными эритроцитами, а также в таком же % соотношении с Rh— и больные. С перекомбинацией сочетаний (за счет образования кроссоверных гамет) будут как мальчики, так и девочки с вероятностью 0,75% (Rh+ больные и Rh-здоровые). |
Примечания:
1) решение строится на основе теории вероятности: если события происходят независимо и каждое событие имеет свою вероятность наступления, то можно определить вероятность того, что эти события произойдут одновременно. Для этого нужно перемножить вероятности этих событий.
2) все расчеты производятся в долях единицы (простых или десятичных). Умножать проценты на проценты – нонсенс! Только конечный результат, из чисто психологических соображений – для удобства восприятия, может быть представлен в процентах.
Ответ: сформулирован таблице.
Биология и генетика пола, сцепление с полом
Образование гамет
Задача 7. У дрозофилы, млекопитающих и человека гомогаметен – женский пол, а
гетерогаметен – мужской. У птиц гомогаметен мужской пол, а гетерогаметен –
женский.
Сколько типов гамет, различающихся по половым хромосомам,
образуется при гаметогенезе:
а) у самки дрозофилы? б) у самца дрозофилы?
в) у курицы? г) у петуха?
д) у женщины? е) у мужчины?
Ход рассуждения:
Если женский пол гомогаметен,, а мужской гетерогаметен, половые хромосомы обозначаются символами «Х» и «Y»
Если же, напротив, гетерогаметен женский пол, а мужской гомогаметен, то половые хромосомы обозначаются символами «Z» и «W»
а) ♀ дрозофилы ХХ
Гаметы: одного типа -Х
б) ♂самец дрозофилы — ХY
Гаметы: двух типов Х и Y
в) курица ♀ZW
Гаметы: двух типов: Z и W
г) петух ♂ — ZZ
Гаметы: одного типа -Z
Наследование пола и признаков, зависимых от пола
Задача 8. Известно, что «трехшерстные» кошки – всегда самки. Это обусловлено тем, что
гены черного и рыжего цвета их шерсти аллельны, находятся в Х- хромосоме, при этом взаимодействуют друг с другом по типу кодоминирования. В результате кодоминировани и формируется сочетание рыжего и черного цвета – «трехшерстность».
Какова вероятность получения в потомстве трехшерстных котят от скрещивания трехшерстной кошки с черным котом?
Образец решения и оформления:
Дано:
|
Ген (аллель) и определяемый признак |
Схема скрещивания, потомство |
|
|
трехшерстная черный
Р.
♀
Гаметы: |
 —
аллель черного цвета шерсти
—
аллель черного цвета шерсти —
аллель рыжего цвета шерсти
—
аллель рыжего цвета шерсти 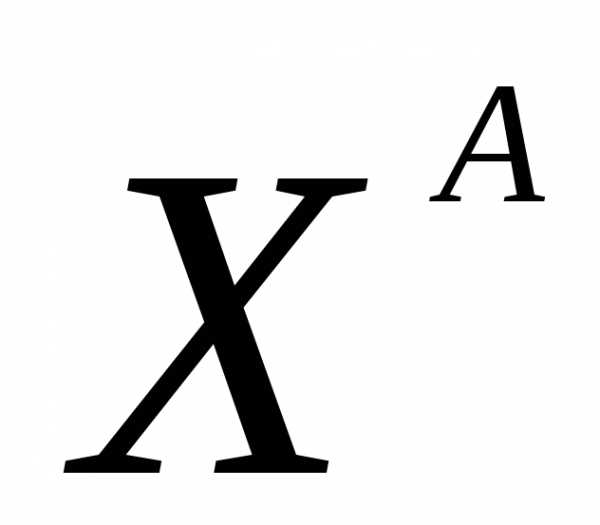
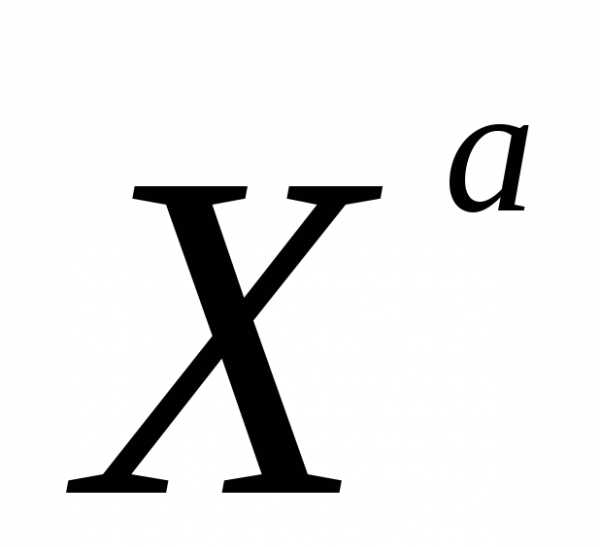 —
трёхшерстность (кодоминирование)
—
трёхшерстность (кодоминирование)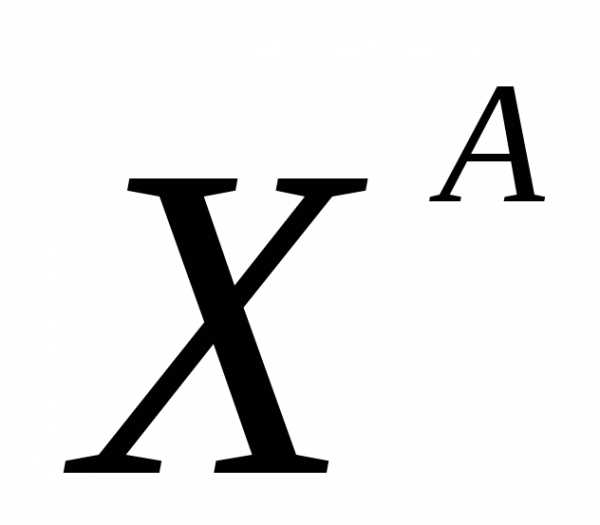
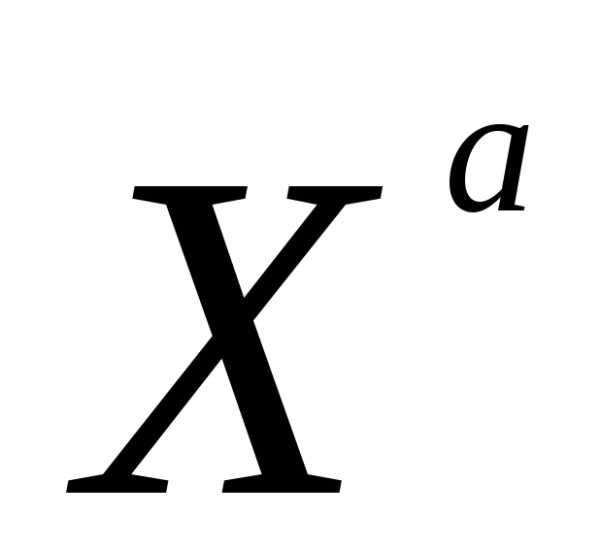 ♂
♂


studfiles.net
29. Взаимодействие аллельных генов
Взаимодействие между аллельными генами осуществляется в виде трех форм: полное доминирование, неполное доминирование и независимое проявление (кодоминирование).
Полное доминирование – когда один доминантный аллель полностью подавляет проявление рецессивного аллеля, например, желтая окраска горошин доминирует над зеленой.
Неполное доминирование наблюдается в том случае, когда один ген из пары аллелей не обеспечивает образование в достаточном для нормального проявления признака его белкового продукта. При этой форме взаимодействия генов все гетерозиготы и гомозиготы значительно отличаются по фенотипу друг от друга. Примером расщепления при неполном доминировании может служить наследование окраски цветков Ночной красавицы.
При скрещивании растений с красными цветками (АА) и растений с белыми (аа) гибриды F1 имеют розовые цветки (Аа). Таким образом, имеет место неполное доминирование; в F2 наблюдается расщепление 1 : 2 : 1 как по фенотипу, так и по генотипу.
Кроме полного и неполного доминирования известны случаи отсутствия доминантно-рецессивных отношений или кодоминирования. При кодоминировании у гетерозиготных организмов каждый из аллельных генов вызывает формирование в фенотипе контролируемого им признака.
Примером этой формы взаимодействия аллелей служит наследование групп крови человека по системе АВ0, детерминируемых геном I. Существует три аллеля этого гена Io, Ia, Ib, определяющие антигены групп крови. Наследование групп крови иллюстрирует также явление множественного аллелизма: в генофондах популяций человека ген I существует в виде трех разных аллелей, которые комбинируются у отдельных индивидуумов только попарно. До этого примера мы говорили о генах, существующих только в двух разных аллельных формах. Однако многие гены состоят из сотен пар нуклеотидов, так что мутации могут проходить во многих участках гена и порождать множество различных его аллельных форм. Так как в каждой из гомологичной хромосом имеется по одному аллельному гену, то, разумеется, диплоидный организм имеет не более двух из серии аллелей генофонда популяции.
30. Неаллельные взаимодействия генов
Неаллельные гены— это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой.
При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы и взаимодействия неаллельных генов:
комплемментарность;
эпистаз;
полимерия.
Комплементарное(дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9:6:1, 9:3:4, 9:7, иногда 9:3:3:1. Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлинённую. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдёт расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлинённые.
Эпистаз— взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3, 7:6:3. Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7, 13:3.
Полимерия — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 но фенотипу происходит в соотношении 1:4:6:4:1.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1.
Пример: цвет кожи у людей, который зависит от четырёх генов.
studfiles.net
Неаллельные гены. Наследование признаков, сцепленных с полом
Неаллельные гены. Наследование признаков, сцепленных с полом
Неаллельные гены – это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки.
1. Комплементарное (дополнительное) действие генов – это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9: 6: 1, 9: 3: 4, 9: 7, иногда 9: 3: 3: 1.
2. Эпистаз – взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый – гипостатичным.
Если эпистатичный ген не имеет собственного фено-типического проявления, то он называется ингибитором и обозначается буквой I.
Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным.
3. Полимерия – взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локу-са имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным.
Пол организма – это совокупность признаков и анатомических структур, обеспечивающих половой путь размножения и передачу наследственной информации.
В кариотипе человека содержится 44 аутосомы и 2 половых хромосомы – Х и Y За развитие женского пола у человека отвечают две Х-хромосомы, т. е. женский пол гомогаметен. Развитие мужского пола определяется наличием Х– и Y-хромосом, т. е. мужской пол гетерогаметен.
Признаки, сцепленные с полом – это признаки, которые кодируются генами, находящимися на половых хромосомах. У человека признаки, кодируемые генами Х-хромосомы, могут проявляться у представителей обоих полов, а кодируемые генами Y-хромосо-мы – только у мужчин.
Различают Х-сцепленное и Y-сцепленное (голандри-ческое) наследование.
Так как Х-хромосома присутствует в кариотипе каждого человека, то и признаки, наследуемые сцеплено с Х-хромосомой, проявляются у представителей обоих полов. Женщины получают эти гены от обоих родителей и через свои гаметы передают их потомкам. Мужчины получают Х-хромосому от матери и передают ее своему потомству женского пола.
Различают Х-сцепленное доминантное и Х-сцеплен-ное рецессивное наследование. У человека Х-сцеп-ленный доминантный признак передается матерью всему потомству. Мужчина передает свой Х-сцеплен-ный доминантный признак лишь своим дочерям.
Y-сцепленные гены присутствуют в генотипе только мужчин и передаются из поколения в поколение от отца к сыну.
cribs.me
Неаллельные гены. Наследственность.
Неаллельные гены
Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки.
Неаллельные гены также могут взаимодействовать между собой. При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы взаимодействия неаллельных генов:
1) комплементарность;
2) эпистаз;
3) полимерия.
Комплементарное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9 : 6 : 1, 9 : 3 : 4, 9 : 7, иногда 9 : 3 : 3 : 1.
Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлиненную. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдет расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлиненные.
Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным.
Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I.
Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (B, b) подавляется доминантным эпистатичным геном (I > B, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12 : 3 : 1, 13 : 3, 7 : 6 : 3.
Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > B, b). Расщепление по фенотипу может идти в соотношении 9 : 3 : 4, 9 : 7, 13 : 3.
Полимерия — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 по фенотипу происходит в соотношении 1 : 4 : 6 : 4 : 1.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15 : 1.
Источник: Н. С. Курбатова, Е. А. Козлова «Конспект лекций по общей биологии»
xn--90aeobapscbe.xn--p1ai
6. Частота кроссинговера и линейное расположение генов в хромосоме. Цитологические доказательства кроссинговера
А. Стертевант предположил, что частота кроссинговера показывает относительное расстояние между генами: чем чаще осуществляется кроссинговер, тем дальше отстоят гены друг от друга в хромосоме. Чем реже кроссинговер, тем они ближе. Таким образом он предложил строить линейные карты расположения генов.
В России для обозначения частоты кроссинговера, равной 1%, использовали термин «морганида» в честь Т.Х. Моргана. В США долгое время использовали термин «единица карты» (map unit — m.u.). Начиная с 80-х годов почти повсеместно как в русскоязычной, так и англоязычной литературе используют термин «сантиморган» (centiMorgan или сМ).
В начале 30-х годов К. Штерн получил линии дрозофилы, имеющие половые хромосомы, отличимые друг от друга на цитологическом уровне. У самки на одну из Х-хромосом был перенесен небольшой фрагмент Y—хромосомы, что придало ей специфическую Г-образную форму, легко различимую под микроскопом. (Рис.).
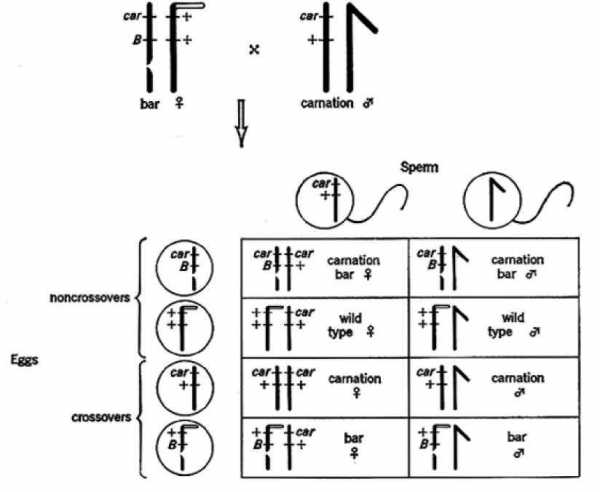
Схема опыта по цитологическому доказательству кроссинговера на D. melanogaster
Были получены самки, гетерозиготные по двум указанным морфологически различным X—хромосомам и одновременно по двум генам Ваг (В) и carnation {car).
Цитологический анализ 374 препаратов самок показал, что в 369 случаях кариотип соответствовал ожидаемому. Все четыре класса самок имели по одной нормальной, т.е. палочковидной Х-хромосоме, полученной от отца. Кроссоверные (т.е. В саг+ по фенотипу) самки содержали двуплечую Г-образную Х-хромосому.
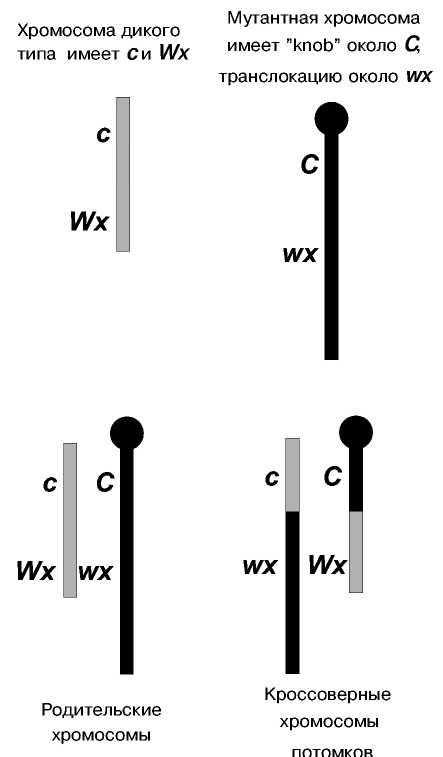
Схема кроссинговера в IX паре хромосом кукурузы в опытах Крейтона и МакКлинток
Эти результаты подтвердили гипотезу Т.Х. Моргана и его сотрудников о том, что кроссинговер представляет собой обмен участками гомологичных хромосом и что гены действительно локализованы в хромосомах.
Аналогичные результаты были получены на кукурузе (Рис.).
Г. Крейтон и Б. МакКлинток получили линию, в которой хромосомы IX пары у кукурузы оказались цитологически различными (гетероморфными). Одна была нормальной и несла гены с (неокрашенный эндосперм) и Wx (крахмалистый эндосперм), другая несла knob (утолщение) одного плеча, другое плечо было длиннее. Эта хромосома была помечена генами С (окрашенный эндосперм) и wx (восковидный эндосперм).
В этом скрещивании обнаружили, что кроссоверные зерна неизменно содержали IX хромосому с обменявшимися участками: хромосому нормальной длины, но с утолщением, или хромосому без утолщения, но удлиненную.
7.Митотический (соматический) кроссинговер. Факторы, влияющие на кроссинговер
До сих пор мы рассматривали обмены участками хромосом, происходящие в ходе мейоза. Однако, как выяснилось, кроссинговер может происходить и в соматических клетках в ходе митоза.
Гомологичные хромосомы в интерфазе конъюгируют и входят в митотическое деление спаренными. Соматический кроссинговер может быть обнаружен, если он осуществляется на стадии четырех хроматид (Рис.).
На частоту кроссинговера влияют:
а) внешние условия (температура и др.),
б) стадии развития (возраст),
в) пол,
г) генотип (определенные гены или структурные изменения хромосом).
Кроссинговер обнаружен почти у всех изученных видов животных и растений, однако формирование обменов хромосом зависит от пола, например, у самцов дрозофилы и самок шелкопряда (обапола гетерогаметные) кроссинговер не происходит.
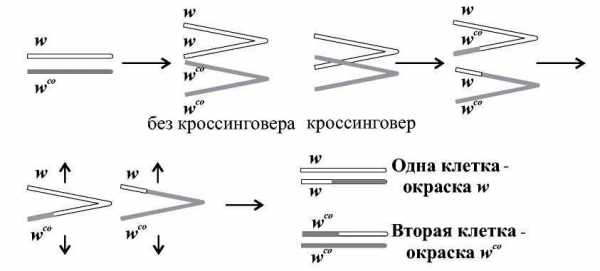
Схема митотического кроссинговера
Острое воздействие высокой температурой, например, тепловой шок при 35°С на стадии 12 час куколки дрозофилы приводит к огромному увеличению частоты обменов. На участке между генами It и stw при 25°С нормальной для дрозофилы температуры — выявлены низкая частота обменов 0,05%. После теплового шока она возрастает более чем в 30 раз (т.е. достигает значения 1,7%).
Обнаружены и другие факторы, влияющие на частоты перекреста, такие как дополнительные хромосомы в геноме, хромосомные перестройки и т.д.
studfiles.net
Генетика
Взаимодействие неаллельных генов
Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой.
При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов.
Выделяют три формы и взаимодействия неаллельных генов:
Комплиментарность
Комплементарное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9:6:1, 9:3:4, 9:7, иногда 9:3:3:1.
Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлинённую. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдёт расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлинённые.
Эпистаз
Подавление (ингибирование) действия одной аллельной пары генов геном другой, не аллельной им пары, называется эпистазом. Различают доминантный и рецессивный эпистаз. Если обычное аллельное доминирование можно представить в виде формулы А>а, То явление эпистаза выразится формулой А>В (доминантный эпистаз) или А>В (рецессивный эпистаз), когда доминантный или рецессивный ген одной аллельной пары не допускает проявления генов другой аллельной пары.
Гены, подавляющие действие других, не аллельных им генов, называются Эпистатичными, А подавляемые — Гипостатичными. Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию. При эпистазе фермент, образующийся под контролем одного гена, полностью подавляет или нейтрализует действие фермента, контролируемого другим геном.
Разберем эпистатическое действие генов на примере наследования окраски зерна у овса (рис. 28). У этой культуры были установлены доминантные гены, определяющие черную и серую окраску зерна. Обозначим один из них буквой А, А второй — В. При этом можно представить себе скрещивание, в котором родительские формы имели генотипы A Abb (черносемянный) и АаВВ (серосемянный). В генотипе растения первого поколения (АаВB) Содержатся доминантные гены и черной окраски А, И серой окраски В. Так как ген А Эпистатичен по отношению к гену В, Он не дает ему проявиться, и все гибриды F1 Будут черносемянными. В F1 Произойдет расщепление в отношении 12 черных : 3 серых: 1 белый. Такой результат расщепления легко понять, если представить себе отношение 12:3:1 как видоизменение типичного для дигибридных скрещиваний отношения 9:3:3:1.
В девяти сочетаниях присутствуют оба доминантных гена А И В, Но ген серой окраски В Не может проявляться, и они дают черносемянные растения. В трех сочетаниях (AAbbAabb, Aabb) Ген черной окраски семян А Также обусловит развитие черносемянных растений. Эта группа по фенотипу будет совершенно сходна с первой, и, следовательно, из каждых 16 растений 12 будут черносемянными. В трех сочетаниях (ааВВ, ааВB, ааВB) Доминантный ген В При отсутствии эпистатичного гена А Может проявить доминантное действие по отношению к своему рецессивному аллелю b, И разовьются растения с серыми семенами. Один генотип (Aabb) Представляет собой новую комбинацию, в которой проявится белая окраска зерна, так как отсутствуют оба доминантных гена.
Полимерия
Полимерия — взаимодействие неаллельных множественных генов, однонаправленно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс.
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммарного действия нескольких генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление в F2 по фенотипу при дигибридном скрещивании происходит в соотношении 1:4:6:4:1, а в целом соответствует третьей, пятой (при дигибридном скрещивании), седьмой (при тригибридном скрещивании) и т.п. строчкам в треугольнике Паскаля.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление в F2 по фенотипу при дигибридном скрещивании — 15:1.
Пример полимерии — наследование цвета кожи у людей, который зависит (в первом приближении) от четырёх генов с кумулятивным эффектом.
genetics.prep74.ru
Резус-конфликт
Может возникнуть при беременности резус-отрицательной женщины резус-положительным плодом (резус-фактор от отца). При попадании эритроцитов плода в кровоток матери, против резус-фактора у нее образуются антирезусные антитела. В норме кровоток матери и плода смешивается только во время родов, поэтому теоретически возможным резус-конфликт считается во вторую и последующие беременности резус-положительным плодом. Практически в современных условиях часто происходит повышение проницаемости сосудов плаценты, различные патологии беременности, приводящие к попаданию эритроцитов плода в кровь матери и во время первой беременности. Поэтому антирезусные антитела необходимо определять при любой беременности у резус-отрицательной женщины начиная с 8 недель (время образования резус-фактора у плода). Для предотвращения их образования во время родов, в течение 72 часов после любого окончания беременности срока более 8 недель вводят антирезусный иммуноглобулин.
Вопрос № 31.
ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ.
а) Понятие о неаллельных генах. Их локализация в хромосомах.
Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой.
б) Эпистаз и гипостаз, как отражение взаимодействия неаллельных генов (пример).
Эпистаз – такая форма взаимодействия неаллельных генов между собой, при которой гены одной аллельной пары подавляют действие генов другой аллельной пары. Ген, подавляющий действие другого гена называется эпистатическим геном (супрессором), а подавляемый им ген носит название гипостатического. Выделяют доминантный и рецессивный эпистаз. Доминантный эпистаз – эпистатическое действие оказывает доминантный ген в гомо- или гетерозиготном состоянии (например, окраска оперения у кур). Рецессивный эпистаз – эпистатическое действие оказывает рецессивный ген только в гомозиготном состоянии (например, окраска шерсти у домовой мыши). При доминантном эпистазе при скрещивании двух гомозиготных особей, отличающихся по двум парам неаллельных генов, расщепление по фенотипу среди гибридов второго поколения будет выражаться соотношением 12:3:1 или 13:3. При рецессивном эпистазе это соотношение 9:3:4. Примером доминантного эпистаза может служить наследование окраски оперения у кур. У большинства пород кур (плимутрок, виандот) доминантный ген С обусловливает развитие пигментированного оперения, а его аллель с — белого (альбинизм). У породы леггорн существует эпистатический ген I, подавляющий действие доминантного гена С. Его аллель — i не влияет на проявление гена С. Леггорны с генотипом IICC имеют белое оперение в связи с эпистазом гена I над геном С. Виандоты с генотипом iicc имеют белое оперение вследствие отсутствия доминантного аллеля пигментации — С. Проведено скрещивание гомозиготного белого леггорна (IICC) с белым виандотом (iicc). Потомство от этого скрещивания все оказалось белым (IiCc). Далее при скрещивании гибридов первого поколения между собой в F2 наблюдалось следующее расщепление: 13/16 белых кур (I-C-, Icc, iicc) и 3/16 кур с пигментированным оперением (iiC-). В целом насле- дование окраски оперения у кур
Явление рецессивного эпистаза можно проиллюстрировать на приме- ре наследования пигментации шерсти у мышей. Доминантный ген А определяет зональное распределение пигмента по длине волоса у мышей, а его аллель а не влияет на зональное распределение пигмента. Доминантный ген С детерминирует синтез пигмента, а его аллель с определяет отсутствие пигмента (альбинизм) и не дает проявиться гену А (а) т.е. является эпистатическим геном. Произведено скрещивание гомозиготной черной особи (CCаа) с гомо- зиготной белой особью (ссАА). Потомство от этого скрещивания оказалось серым (СсАа). Далее при скрещивании гибридов первого поколения между собой в F2 отмечено следующее расщепление: 9/16 серых мышей (С-А-), 3/16 черных (С-аа) и 4/16 белых (ссА-, ссаа).
в) Комплементарность (определение, пример).
Комплементарность – это такой тип взаимодействия неаллельных генов, при котором гены из разных аллельных пар дополняют действие друг друга, приводя к формированию нового признака. Выделяют доминантную или рецессивную комплементарность (т.е. формирование новых признаков связано с взаимодействием доминантных либо только рецессивных аллелей). Например, серая окраска шерсти у мышей контролируется двумя генами (А и В). Ген А детерминирует синтез пигмента, однако как гомозиготы (АА), так и гетерозиготы (Аа) — альбиносы. Другой ген В обеспечивает скопления пигмента преимущественно у основания и на кончиках волос. Таким образом, при комплементарном взаимодействии генов расщепление по фенотипу в потомстве при скрещивании двух дигетерозиготных организмов будет составлять 9:7 (доминантная комплементарность), 9:3:3:1 или 9:6:1 (доминантная и рецессивная комплементарность). Примером комплементарного взаимодействия генов у человека может быть синтез защитного белка — интерферона. Его образование в организме связано с комплементарным взаимодействием двух неаллельних генов, расположенных в разных хромосомах.
г) Плейотропия (определение, пример)
Плейотропное действие генов — это зависимость нескольких признаков от одного гена, то есть множественное действие одного гена. У дрозофилы ген белого цвета глаз одновременно влияет на цвет тела, длины, крыльев, строение полового аппарата, снижает плодовитость, уменьшает продолжительность жизни. У человека известна наследственная болезнь — арахнодактилия («паучьи пальцы»-очень тонкие и длинные пальцы), или болезнь Марфана. Ген, отвечающий за эту болезнь, вызывает нарушение развития соединительной ткани и одновременно влияет на развитие нескольких признаков: нарушение строения хрусталика глаза, аномалии в сердечно-сосудистой системе. Плейотропное действие гена может быть первичным и вторичным. При первичной плейотропии ген проявляет свой множественный эффект. Например, при болезни Хартнупа мутация гена приводит к нарушению всасывания аминокислоты триптофана в кишечнике и его реабсорбции в почечных канальцах. При этом поражаются одновременно мембраны эпителиальных клеток кишечника и почечных канальцев с расстройствами пищеварительной и выделительной систем. При вторичной плейотропии есть один первичный фенотипний проявление гена, вслед за которым развивается ступенчатый процесс вторичных изменений, приводящих к множественным эффектам. Так, при серповидно клеточной анемии у гомозигот наблюдается несколько патологических признаков: анемия, увеличенная селезенка, поражение кожи, сердца, почек и мозга. Поэтому гомозиготы с геном серповидно клеточной анемии гибнут, как правило, в детском возрасте. Все эти фенотипные проявления гена составляют иерархию вторичных проявлений. Первопричиной, непосредственным фенотипним проявлением дефектного гена является аномальный гемоглобин и эритроциты серповидной формы. Вследствие этого происходят последовательно другие патологические процессы: слипание и разрушение эритроцитов, анемия, дефекты в почках, сердце, мозге — эти патологические признаки вторичны. При плейотропии, ген, воздействуя на какой то один основнй признак, может также менять, модифицировать проявление других генов, в связи с чем введено понятие о генах-модификаторах. Последние усиливают или ослабляют развитие признаков, кодируемых «основным» геном.
д) ?Мультифакториальный принцип формирования фенотипа как выражение диалектического единства генетических и средовых факторов.
Показателями зависимости функционирования наследственных задатков от характеристик генотипа является пенетрантность и экспрессивность. Пенентрантность – вероятность проявления генов, явление появления или отсутствия признака у организмов, одинаковых по генотипу. Пенетрантность значительно колеблется как среди доминантных, так и среди рецессивных генов. Наряду с генами, фенотип которых появляется только при сочетании определенных условий и достаточно редких внешних условий (высокая пенетрантность), у человека есть гены, фенотипное проявление которых происходит при любых соединениях внешних условий (низкая пенетрантность). Пенетрантностью измеряется процентом организмов с фенотипным признаком от общего количества обследованных носителей соответствующих аллелей. Если ген полностью, независимо от окружающей среды, определяет фенотипное проявление, то он имеет пенетрантность 100 процентов. Однако некоторые доминантные гены проявляются менее регулярно. Так, полидактилия имеет четкое вертикальное наследования, но бывают пропуски поколений. Доминантная аномалия — преждевременное половое созревание — присуще только мужчинам, однако иногда может передаться заболевания от человека, который не страдал этой патологией. Пенетрантностью указывает, в каком проценте носителей гена оказывается соответствующий фенотип. Итак, пенетрантность зависит от генов, от среды, от того и другого. Таким образом, это не константное свойство гена, а функция генов в конкретных условиях среды. Расчет пенентрантности = число особей с фенотипическим проявление признака : общее число особей с геном. Пенентрантность врожденного вывиха бедра 25% Экспрессивность – степень проявления (вырожденности) признака. изменение количественного проявления признака у разных особей-носителей соответствующего аллелей. При доминантных наследственных заболеваниях экспрессивность может колебаться. В одной и той же семье могут проявляться наследственные болезни от легких, едва заметных, до тяжелых: различные формы гипертонии, шизофрении, сахарного диабета и т.д. Рецессивные наследственные заболевания в пределах семьи проявляются однотипно и имеют незначительные колебания экспрессивности. Может быть вызвана как генетическими факторами, так и окружающей средой.
Вопрос № 32.
НАСЛЕДОВАНИЕ ПОЛА.
а) Роль хромосом в детерминации пола.
У растений и животных хромосомный механизм определения пола является наиболее распространённым. Согласно хромосомной теории, пол организма определяется половыми хромосомами (гоносомами). В зависимости от того, какой пол является гетерогаметным, выделяют следующие типы хромосомной детерминации:
самки гомогаметны, самцы гетерогаметны (Drosophila-тип)
самки гетерогаметны, самцы гомогаметны (Abraxas-тип)
У особей гомогаметного пола ядра всех соматических клеток содержат диплоидный набор аутосом и две одинаковые половые хромосомы, которые обозначаются как XX (ZZ). Организмы такого пола продуцируют гаметы только одного класса — содержащие по одной X (Z) хромосоме.
У особей же гетерогаметного пола в каждой соматической клетке, помимо диплоидного набора аутосом, содержатся либо две разнокачественные половые хромосомы, обозначаемые как Х и Y (Z и W), либо только одна — X (Z) (тогда количество хромосом получается нечётным). Соответственно у особей такого пола образуются два класса гамет: либо несущие X/Z-хромосомы и Y/W-хромосомы, либо несущие X/Z-хромосомы и не несущие никаких половых хромосом.
У большинства видов животных и растений гомогаметен женский пол, а гетерогаметен мужской. Сюда относятся млекопитающие, большинство насекомых, некоторые рыбы, растения и др.
Существует два основных типа XY-детерминации пола:
Как у человека: пол зависит от наличия Y-хромосомы (если она есть, проявляется фенотип самца, если нет — самки). Так, при мутациях, связанных с изменением числа половых хромосом, особи, имеющие в генотипе комбинации XY, XXY, XYY, XXYY и т. п. будут обладать мужским фенотипом различной степени выраженности, а имеющие комбинации XX, X, XXX, XXXX и т. п. — женским.
Как у представителей рода мушек-дрозофил: пол определяется по соотношению числа X-хромосом и числа аутосом, наличие же Y-хромосомы никак не влияет на детерминацию пола, но самцы без неё стерильны, так как в ней находятся гены, ответственные за сперматогенез.
У многих других организмов (птицы, некоторые рептилии и рыбы, бабочки, ручейники, из растений — земляника) наблюдается обратная картина — гомогаметен мужской пол (имеет две гомологичные хромосомы Z), а гетерогаметен женский (имеет одну Z-хромосому и одну состоящую в основном из гетерохроматина и потому генетически инертную W-хромосому).
Вероятно, исходным для бабочек механизмом определения пола был механизм ZO самка/ZZ самец [1]. Затем, путем хромосомных перестроек, возникла система определения пола WZ самка/ZZ самец, характерная для 98 % видов бабочек. У видов с системой Z/ZZ определение пола зависит от соотношения числа пар половых хромосом и аутосом, но у тутового шелкопряда (система WZ/ZZ) обнаружен отвечаюший за развитие женского пола ген Fem в W-хромосоме.
При этом механизме определения пола один из полов (гомогаметный) обладает двумя X-хромосомами, в то время, как второй (гетерогаметный) только одной. При этом пол определяется так же, как и у дрозофил: по соотношению числа X-хромосом и аутосом. Этот механизм определения пола обнаружен у некоторых насекомых (клопов, бабочек и др.) и круглых червей. У нематоды Caenorhabditis elegans при наборе половых хромосом XX формируется гермафродит, а при наборе ХО — самец.
б) Гомо- и гетерогаметный пол.
Пол гомогаметный – Характеризующийся наличием двух идентичных половых хромосом и формированием однотипных гамет (у человека – женский).
Гетерогаметный пол — это генетически определённый пол, соответствующий наличию в клетках организма двух разных половых хромосом или одной, в удвоенной дозе приводящей к формированию альтернативного пола. Особи гетерогаметного пола дают две группы гамет (по содержанию разных половых хромосом).
У животных с гетерогаметным мужским полом для обозначения половых хромосом используются буквы X и Y. Особи, в норме несущие пару половых хромосом X и Y или одну хромосому X, — самцы, а несущие две хромосомы X — самки. К этой группе относятся млекопитающие, большинство видов насекомых и многие другие группы.
В случае, если гетерогаметный пол — женский, то используются другие обозначения для половых хромосом — Z и W. Особи с генотипом ZZ — самцы; ZW — самки. Гетерогаметный женский пол характерен для чешуекрылых насекомых, пресмыкающихся и птиц.
в) Типы хромосомной детерминации пола у различных биологических видов.
(Смотри в вопрос № 32 (а))
г) Наследование признаков, сцепленных с полом.
Признаки, наследуемые через половые X- и Y- хромосомы, получили название сцепленных с полом. У человека признаки, наследуемые через Y-хромосому, могут быть только у лиц мужского пола, а наследуемые через X-хромосому, — у лиц как одного, так и другого пола. Особь женского пола может быть как гомо-, так и гетерозиготной по генам, локализованным в X-хромосоме. А рецессивные аллели генов у нее проявляются только в гомозиготном состоянии. Поскольку у особей мужского пола только одна X-хромосома, все локализованные в ней гены, даже рецессивные, сразу же проявляются в фенотипе. Такой организм часто называют гомозиготным.
У человека некоторые патологические состояния сцеплены с полом. К ним относится, например, гемофилия. Аллель гена, контролирующий нормальную свертываемость крови (Н), и его аллельная пара — ген гемофилии h находятся в X-хромосоме. Аллель Н доминантен, аллель h — рецессивен, поэтому если женщина гетерозиготна по этому гену (XHXh), гемофилия у нее не проявляется. У мужчины только одна X-хромосома. Следовательно, если у него в X-хромосоме находится аллель Н, то он и проявляется.
Если же X-хромосома мужчины имеет аллель h, то мужчина страдает гемофилией: X-хромосома не несет генов, определяющих механизмы нормального свертывания крови.
Естественно, что рецессивный аллель гемофилии в гетерозиготном состоянии находится у женщин даже в течение нескольких поколений, пока снова не проявляется у кого-либо из лиц мужского пола. Женщина, страдающая гемофилией, может родиться лишь от брака женщины, гетерозиготной по гемофилии, с мужчиной, страдающим гемофилией. Ввиду редкости этого заболевания, такое сочетание маловероятно.
Аналогичным образом наследуется дальтонизм, то есть такая аномалия зрения, когда человек путает цвета, чаще всего красный с зеленым. Нормальное цветовосприятие обусловлено доминантным аллелем, локализованным в X-хромосоме. Его рецессивная аллельная пара в гомо- и гетерозиготном состоянии приводит к развитию дальтонизма.
Отсюда понятно, почему дальтонизм чаще встречается у мужчин, чем у женщин: у мужчин только одна X-хромосома, и если в ней находится рецессивный аллель, детерминирующий дальтонизм, он обязательно проявляется. У женщины две X хромосомы: она может быть как гетерозиготной, так и гомозиготной по этому гену, но в последнем случае будет страдать дальтонизмом.
д) Голандрические признаки.
Голандрические признаки детерминируются генами, расположенными в негомологичном участке Y-хромосомы (С), и проявляются фенотипически только у мужчин и передаются от отца ко всем сыновьям. Таких генов описано 6 (ихтиоз, волосатость ушей, перепонка между пальцами ног и др.) Примеры признаков: гипертрихоз ушных раковин, избыточный рост волос на средних фалангах пальцев кистей, азооспермия. Передача признака от отца всем сыновьям и только сыновьям. Дочери никогда не наследуют признак от отца.
Вопрос № 33.
НАСЛЕДСТВЕННОСТЬ И ИЗМЕНЧИВОСТЬ.
а) Ген – функциональная единица наследственного материала. Его свойства.
Ген — структурная и функциональная единица наследственности, контролирующая развитие определенного признака или свойства. Совокупность генов родители передают потомкам во время размножения.
В настоящее время, в молекулярной биологии установлено, что гены — это участки ДНК, несущие какую-либо целостную информацию — о строении одной молекулы белка или одной молекулы РНК. Эти и другие функциональные молекулы определяют развитие, рост и функционированиеорганизма.
В то же время, каждый ген характеризуется рядом специфических регуляторных последовательностей ДНК, таких как промоторы, которые принимают непосредственное участие в регулировании проявления гена.
Свойства гена:
дискретность — несмешиваемость генов;
стабильность — способность сохранять структуру;
лабильность — способность многократно мутировать;
множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
аллельность — в генотипе диплоидных организмов только две формы гена;
специфичность — каждый ген кодирует свой признак;
плейотропия — множественный эффект гена;
экспрессивность — степень выраженности гена в признаке;
пенетрантность — частота проявления гена в фенотипе;
амплификация — увеличение количества копий гена.
б) Классификация генов.
По месту локализации генов в структурах клетки различают: расположенные в хромосомах ядра, ядерные гены и цитоплазматические гены, локализация которых связана с хлоропластами и митохондриями.
По функциональному значению различают: структурные гены, характеризующиеся уникальными последовательностями нуклеотидов, кодирующих свои белковые продукты, которые можно идентифицировать с помощью мутаций, нарушающих функцию белка, и регуляторные гены — последовательности нуклеотидов, не кодирующие специфические белки, а осуществляющие регуляцию действия гена (ингибирование, повышение активности и др.).
По влиянию на физиологические процессы в клетке различают: летальные, условно летальные, супервитальные гены, гены-мутаторы, гены-антимутаторы и др.
Следует отметить, что любые биохимические и биологические процессы в организме находятся под генным контролем. Так, деление клеток (митоз, мейоз) контролируется несколькими десятками генов; группы генов осуществляют контроль восстановления генетических повреждений ДНК (репарация). Онкогены и гены — супрессоры опухолей участвуют в процессах нормального деления клеток. Индивидуальное развитие организма (онтогенез) контролируется многими сотнями генов. Мутации в генах приводят к измененному синтезу белковых продуктов и нарушению биохимических или физиологических процессов.
Гомеозисные мутации у дрозофилы позволили открыть существование генов, нормальной функцией которых является выбор или поддержание определенного пути эмбрионального развития, по которому следуют клетки. Каждый путь развития характеризуется экспрессией определенного набора генов, действие которых приводит к появлению конечного результата: глаза, голова грудь, брюшко, крыло, ноги и т. д. Исследования генов комплекса bithorax дрозофилы американским генетиком Льюисом показали, что это гигантский кластер тесно сцепленных генов, функция которых необходима для нормальной сегментации груди (thorax) и брюшка (abdomen). Подобные гены получили название гомеобоксных. Гомеобоксные гены расположены в ДНК группами и проявляют свое действие строго последовательно. Такие гены обнаружены и у млекопитающих, и они имеют высокую гомологию (сходство).
в) Локализация генов в хромосомах.
Можно предполагать, что у человека имеется около 1 млн генов, однако тип наследования установлен не более чем у нескольких сотен из них.
По характеру наследования признаков человек имеет ряд особенностей, отличающих его от других организмов:
преобладают гены с неполным доминированием — важный фактор в проявлении многих доминантных и рецессивных генов;
многие признаки обусловлены полимерными генами;
установлены все 24 теоретически возможные группы сцепления генов. Из них 22 локализованы в аутосомах, в каждой из которых содержится по несколько генов.
более 100 генов локализовано в половых хромосомах. По наследованию через половые хромосомы человек также отличается от других животных. У млекопитающих, в том числе и у человека, Х- и Y-хромосомы имеют гомологический участок, в котором происходит их синопсис и возможен кроссинговер.
В зависимости от того, в каких участках половых хромосом человека находятся локализованные в них гены, их можно разделить на 3 группы:
гены, локализованные в той части Х-хромосомы, которая не имеет гомологического участка в Y-хромосоме. Они полностью сцеплены с полом и передаются исключительно через Х-хромосому. Доминантные гены из этого участка одинаково проявляются у обоих полов, рецессивные же — только в гомозиготном состоянии у женщин и только в гетерозиготном — у мужчин. К их числу относятся: — гены гемофилии; — дальтонизма; — атрофии зрительного нерва и т. д. — группа генов, расположенных в непарном участке Y-хромосомы. Они могут встречаться только у лиц мужского пола и передаются от отца к сыну. К ним относятся: — гены волосатости ушей; — ихтиоза; — перепончатых пальцев на ногах; — гены, расположенные в парном сегменте половых хромосом, т. е. гомологичном для Х- и Y-хромосомы. Их называют неполно или частично сцепленными с полом. Они могут передаваться как с Х-, так и с Y-хромосомой и переходить из одной в другую в результате кроссинговера. Если такие гены находятся в Х-хромосоме, то в ряду поколений они ведут себя как сцепленные с полом, т. е. рецессивные гены у женщин проявляются лишь в гомозиготном состоянии, а у мужчин — в гетерозиготном. Однако в результате кроссинговера эти гены могут попасть в Y-хромосому и составить с ней одну группу сцепления. Тогда они будут вести себя как сцепленные с полом и передаваться от отца только к сыновьям до тех пор, пока не произойдет кроссинговер. К этой группе относится большинство генов, локализованных в половых хромосомах. Таковы, например, гены общей цветной слепоты.
г) Структурно-функциональные уровни организации наследственного материала у про- и эукариот.
Структурно-функциональными уровнями организации наследственного материала являются генный, хромосомный и геномный.
Ген — элементарная структура генного уровня организации. Так как гены относительно независимы друг от друга, возможно дискретное
(раздельное) и независимое наследование (по третьему закону Менделя) и изменение отдельных признаков вследствие генных мутаций.
Гены эукариот находятся в хромосомах, образуя хромосомный уровень организации наследственного материала. Все гены одной хромосомы составляют группу сцепления и передаются вместе с этой хромосомой. На этом уровне происходит перекомбинация генов родителей у потомков при половом размножении и изменения структуры отдельных хромосом.
Набор генов, получаемых потомком от родителей, составляет его генотип. Геном — это гены гаплоидного набора хромосом. Действие генов в разных генотипах проявляется по-разному. Взаимодействуют между собой гены как одной хромосомы, так и разных хромосом. Нарушение набора хромосом приводит к геномным мутациям.
д) Геном – генетическая система клетки.
Геном – полная генетическая система клетки, определяющая характер онтогенетического развития организма и наследственную передачу в ряду поколений всех его структурных и функциональных признаков.
Геном — совокупность всех генов организма; его полный хромосомный набор.
Термин «геном» был предложен Гансом Винклером в 1920 г. для описания совокупности генов, заключённых в гаплоидном наборе хромосом организмов одного биологического вида. Первоначальный смысл этого термина указывал на то, что понятие генома в отличие от генотипа является генетической характеристикой вида в целом, а не отдельной особи. С развитием молекулярной генетики значение данного термина изменилось. Известно, что ДНК, которая является носителем генетической информации у большинства организмов и, следовательно, составляет основу генома, включает в себя не только гены в современном смысле этого слова. Большая часть ДНК эукариотических клеток представлена некодирующими («избыточными») последовательностями нуклеотидов, которые не заключают в себе информации о белках и РНК.
Генетическая информация в клетках содержится не только в хромосомах ядра, но и во внехромосомных молекулах ДНК. У бактерий к таким ДНК относятся плазмиды и некоторые умеренные вирусы, в клетках эукариот — это ДНК митохондрий, хлоропластов и других органоидов клеток. Объёмы генетической информации, заключённой в клетках зародышевой линии (предшественники половых клеток и сами гаметы) и соматических клетках, в ряде случаев существенно различаются. В онтогенезе соматические клетки могут утрачивать часть генетической информации клеток зародышевой линии, амплифицировать группы последовательностей и (или) значительно перестраивать исходные гены.
studfiles.net
