
Мейоз у животных и растений. Биологический смысл мейоза
Мейоз. Это такое деление эукореотных клеток, при котором дочерние клетки получают в 2-а раза меньше информации, чем было в материнской клетке, поэтому мейоз всегда включает в себя редукционное деление, приводящее к уменьшению кол-ва генетической информации. Мейоз у животных проходит перед образованием гамет, поэтому это организмы с генетической редакцией. Мейоз у растений проходит перед образованием спор, так как для всех растений характерной чертой является чередование поколений. Бесполое поколение участвует в образовании спор и всегда имеет двойной или диплоидный набор хромосом. Так как споры образуются в ходе мейоза, то для них характерен одинарный или гаплоидный набор хромосом. Из споры прорастает половое поколение, на котором за счёт митоза формируются яйцеклетки и сперматозоиды. Они сливаются, образуют зиготу, из которой прорастает бесполое поколение.Для некоторых простейших нормой жизни является гаплоидность, поэтому мейоз проходит после образования зиготы (зиготическая редукция).Таким образом мейоз необходим для образования спор и гамет, а также для сохранения генетической стабильности вида. Типичный мейоз включает в себя 2-а деления: эквационное (2), редукционное (1)
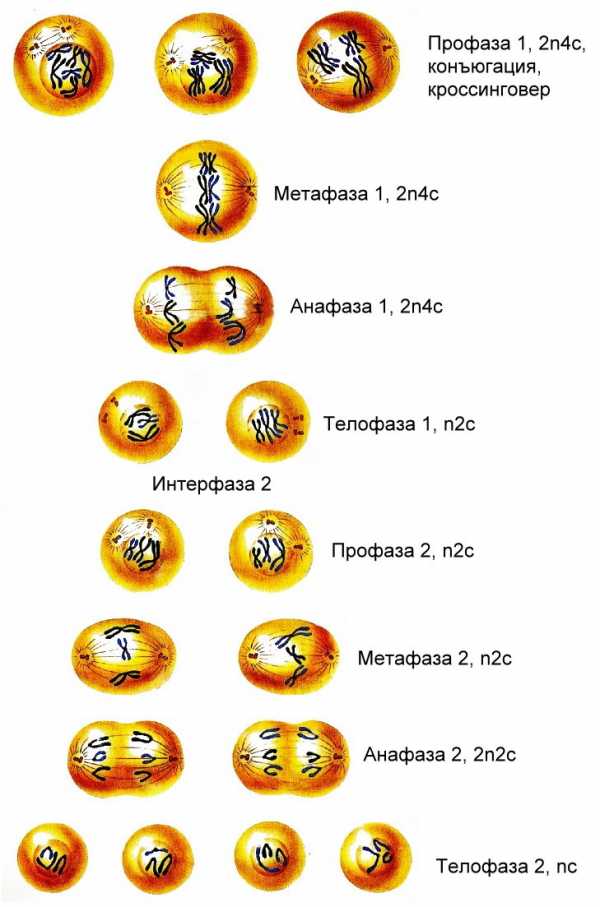
Перед редукционным делением проходит нормальная интерфаза. Между 1 и 2 делением интерфаза сильно сокращена, либо вообще отсутствует. В любом случае удвоение ДНК между этими делениями не происходит. Оба деления включают в себя все фазы. Профаза 1 Делится на несколько периодов: 1.Липтотена – в ходе этого периода ядерная оболочка сохраняется, хромосомы становятся более компактными. 2.Зиготена – начинается сближение гомологичных хромосом. Гомологичными называются хромосомы, сходные по форме, размеру, генетическому содержимому. Гомологичные хромосомы подходят друг к другу и начинается формирование пар или бивалентов. 3.Пахитена – гомологичные хромосомы объединяются в биваленты, таким образом, что находящиеся рядом сестринские хроматиды взаимодействуют друг с другом. Точка взаимодействия называется хиазмой .Кол-во хиазм может быть различным, но существуют правила. Образовавшиеся хиазмы тормозят формирование новых хиазм.(рис 1)
В
хиазмах может происходить разрыв в нитях ДНК. Обмен участками между
гомологичными хромосомами, а затем сшивание сахарофосфатного остова. Этот
процесс получил название кроссинговер. Такой обмен участками между
гомологичными хромосомами является основой комбинативной изменчивости. Таким
образом, формирование бивалентов необходимо для осуществления процесса
кроссинговера и также для нормального расхождения гомологичных хромосом к
разным полюсам клетки в ходе анафазы 1. 4.Диплотена и диокинез – гомологичные
хромосомы на этой стадии пытаются разойтись, однако их сдерживают хиазмы,
которые в данном случае смещаются к теломерным участкам хромосомы и благодаря
хиазмам биваленты сохраняются. Этот процесс получил название диокинеза.
Практически всю профазу, включая диплотену, ядерная оболочка сохраняется. К
концу профазы фрагментируются мембранные органоиды, фрагментируется оболочка
ядра, и хромосомы оказываются в цитоплазме клетки в виде бивалентов.
Формируются веретено деления, причём к каждому биваленту подходит кинетохорная
нить только от одного из полюсов. Благодаря полимеризации этой нити биваленты
начинают продвигаться по экватору клетки. Метафаза 1 В ходе метафазы биваленты
выстраиваются по экватору клетки, таким образом, что гомологичные хромосомы
одной пары лежат по разные стороны от экватора. Так как ориентация бивалентов
относительно полюсов клетки произвольна, то возможно несколько вариантов
расположения бивалентов друг относительно друга. Анафаза 1 Начинается
расхождение гомологичных хромосом к разным полюсам клетки. Так как биваленты
произвольно ориентировались относительно полюсов, то возможны различные
варианты их расхождения и, следовательно, произвольная ориентация бивалентов в
метафазе 1. Варианты расхождения гомологичных хромосом также является основой
комбинативной изменчивости. Расхождение обеспечивает основной и дополнительный
механизмы.Телофаза 1Около каждой клетки формируется полный набор
цитоплазматических органоидов и свой ядерный аппарат. После телофазы проходит
цитокинез, и образуется 2-е клетки. Причём кол-во хромосом и генетическая
информация уменьшены в 2-а раза. 2-е клетки, которые образованы в ходе
редукционного деления подвергаются эквационному делению, которое по своему
механизму является типичным митозом. Результатом эквационного деления является
образование 4-х клеток.(рис 2) В ходе анафазы 1 редукционного деления к каждому
полюсу клетки закономерно показывает только гомологичная хромосома из пары.
Поэтому после телофазы и цитокинеза каждая дочерняя клетка содержит только 1
гомологичную хромосому из пары, то есть в ходе редукционного деления кол-во
хромосом и информации в дочерних клетках уменьшается. Поскольку в метафазе 1
существовали варианты в расположении хромосом, то изучены варианты расхождения
гомологичных хромосом к разным полюсам. Это является основой комбинативной
изменчивости .
vunivere.ru
Мейоз | Student Guru
Posted in БиологияМейоз (редукционное деление) – особый тип деления клеток половых желез, результатом которого из диплоидных (2n) клеток образуются половые клетки, которые имеют гаплоидный (n) набор хромосом. У растений мейоз происходит при образовании спор – специализированных клеток бесполого размножения, характерных для водорослей, грибов, мхов, хвощей, плаунов и папоротников. У голосеменных и покрытосеменных растений мейоз происходит при образовании в пыльниках микроспор, из которых в дальнейшем развивается пыльца, и при образовании в семяпочке макроспор, одна из которых дает начало зародышевому мешку. У животных мейоз или редукционное деление происходит при образовании мужских и женских гамет — сперматозоидов и яйцеклеток.
Для мейоза характерны те же процессы и стадии, которые наблюдаются при митотическом делении клеток, однако они значительно растянуты во времени. Перед началом мейоза в конце интерфазы происходит процесс удвоения генетического материала и синтез необходимых для деления белков. Сам мейоз протекает в два этапа (мейоз I и мейоз II ), которые быстро сменяют друг друга практически без интерфазы, причем, каждый из этих этапов, как и митоз, делится, в свою очередь, на 4 фазы – профаза I, метафаза I, анафаза I, телофаза I и профаза II, метафаза II, анафаза II и телофаза II. В результате мейоза I число хромосом в клетке уменьшается вдвое (собственно редукционное деление), а при мейозе II — сохраняется гаплоидный набор хромосом. В процессе оплодотворения гаплоидные ядра половых клеток сливаются, в результате чего зигота получает двойной набор хромосом.
Таким образом за счет мейоза при половом размножении поддерживается постоянство хромосомного набора и количества ДНК в клетках животных и растений.
Перейти к оглавлению.
You can leave a response, or trackback from your own site.
www.studentguru.ru
Мейоз как основа полового размножения
Мейозом называется особый способ деления эукариотических клеток, при котором исходное число хромосом уменьшается в 2 раза (от древнегреч. «мейон» – меньше – и от «мейозис» – уменьшение).
Отдельные фазы мейоза у животных описал В. Флемминг (1882), а у растений – Э.Страсбургер (1888), а затем российский ученый В.И. Беляев. В это же время (1887) А. Вайсман теоретически обосновал необходимость мейоза как механизма поддержания постоянного числа хромосом. Первое подробное описание мейоза в ооцитах кролика дал Уиниуортер (1900).
Хотя мейоз открыт более 100 лет назад, но изучение мейоза продолжается до сих пор. Интерес к мейозу резко возрос в конце 60-х годов, когда выяснилось, что одни и те же контролируемые генами ферменты могут принимать участие во многих процессах, связанных с ДНК. В последнее время ряд биологов развивают оригинальную идею: мейоз у высших организмов служит гарантом стабильности генетического материала, ибо в процессе мейоза, когда пары хромосом-гомологов тесно соприкасаются, происходит проверка нитей ДНК на точность и восстановление повреждений, затрагивающих сразу обе нити. Изучение мейоза связало методы и интересы двух наук: цитологии и генетики. Это привело к рождению новой ветки знания — цитогенетики, тесно соприкасающейся ныне с молекулярной биологией и генной инженерией.
Биологическое значение мейоза заключается в следующих процессах:
1.Благодаря редукции числа хромосом в результате мейоза в ряду поколений при половом размножении обеспечивается постоянство числа хромосом.
2.Независимое распределение хромосом в анафазе первого деления обеспечивает рекомбинацию генов, относящихся к разным группам сцепления (находящихся в разных хромосомах). Мейотическое распределение хромосом по дочерним клеткам называется сегрегацией хромосом.
3.Кроссинговер в профазе I мейоза обеспечивает рекомбинацию генов, относящихся к одной группе сцепления (находящихся в одной хромосоме).
4. Случайное сочетание гамет при оплодотворении вместе с вышеперечисленными процессами способствует генетической изменчивости.
5. В процессе мейоза происходит еще одно существенное явление. Это процесс активации синтеза РНК (или транскрипционной деятельности хромосом) в ходе профазы (диплотены), связанный с формированием хромосом типа «ламповых щеток» (обнаружены у животных и некоторых растений).
Эта реверсия профазы к интерфазному состоянию (при митозе только в интерфазе идет синтез и-РНК) является специфической характеристикой мейоза как особого типа деления клеток.
Следует отметить, что у простейших наблюдается значительное разнообразие процессов мейоза.
В соответствии с положением в жизненном цикле различают три типа мейоза:
Зиготный (исходный) мейоз происходит в зиготе, т.е. непосредственно после оплодотворения. Он характерен для организмов, в жизненном цикле которых преобладает гаплоидная фаза (аскомицеты, бизидиомицеты, некоторые водоросли, споровики и др.).
Гаметный (терминальный) мейоз происходит во время формирования гамет. Он наблюдается у многоклеточных животных (в т.ч. у человека), а также среди простейших и некоторых низших растений, в жизненном цикле которых преобладает диплоидная фаза.
Промежуточный (споровый) мейоз протекает во время спорообразования у высших растений, включаясь между стадиями спорофита (растения) и гаметофита (пыльца, зародышевый мешок).
Таким образом, мейоз — это форма ядерного деления, сопровождающаяся уменьшением числа хромосом с диплоидного до гаплоидного и изменением генетического материала. Результат мейоза — образование клеток с гаплоидным набором хромосом (половых клеток).
Продолжительность мейоза может отличаться в зависимости от вида растений и животных (табл. 1).
Таблица 1. Продолжительность мейоза у различных видов растений
| Вид | 2n | Мейотический цикл, час |
| Secate sereate (рожь культурная) | 51,2 | |
| Allium сера (лук репчатый) | 96,0 | |
| Tradescantia paludosa | 126,0 | |
| Lilium herry | 170,0 | |
| Lilium longiflorum | 192,0 | |
| Trillium erectum (трилиум прямостоячий) | 274,0 |
Типичный мейоз состоит из двух последовательных клеточных делений, которые соответственно называются мейоз I и мейоз II. В первом делении происходит уменьшение числа хромосом в два раза, поэтому первое мейотическое деление называют редукционным, реже – гетеротипным. Во втором делении число хромосом не изменяется; такое деление называют эквационным (уравнивающим), реже – гомеотипным. Выражения «мейоз» и «редукционное деление» часто используют как синонимы.
Исходное число хромосом в мейоцитах (клетках, вступающих в мейоз) называется диплоидным хромосомным числом (2n) Число хромосом в клетках, образовавшихся в результате мейоза, называется гаплоидным хромосомным числом (n). Минимальное число хромосом в клетке называется основным числом (x). Основному числу хромосом в клетке соответствует и минимальный объем генетической информации (минимальный объем ДНК), который называется геном.
Количество геномов в клетке называется геномным числом (n). У большинства многоклеточных животных, у всех голосеменных и многих покрытосеменных растений понятие гаплоидности–диплоидности и понятие геномного числа совпадают. Например, у человека n=x=23 и 2n=2x=46.
Морфология мейоза — характеристика фаз
Интерфаза
Предмейотическая интерфаза отличается от обычной интерфазы тем, что процесс репликации ДНК не доходит до конца: примерно 0,2…0,4 % ДНК остается неудвоенной. Таким образом, деление клетки начинается на синтетической стадии клеточного цикла. Поэтому мейоз образно называют преждевременным митозом. Однако в целом, можно считать, что в диплоидной клетке (2n) содержание ДНК составляет 4с.
При наличии центриолей происходит их удвоение таким образом, что в клетке имеется две диплосомы, каждая из которых содержит пару центриолей.
Первое деление мейоза
ДНК прошла репликацию. Начитается профаза I, самая продолжительная стадия мейоза.
Стадия профазы I подразделяется на следующие стадии:
лептотена — стадия тонких нитей;
зиготена — стадия двойных нитей;
пахитена — стадия толстых нитей;
диплотена — кроссинговер;
диакинез — исчезновение ядерной оболочки и ядрышка.
В ранней профазе (лептотене) происходит подготовка к конъюгации хромосом. Хромосомы уже удвоены, но сестринские хроматиды в них еще неразличимы. Хромосомы начинают упаковываться (спирализоваться).
В отличие от профазы митоза, где хромосомы расположены по мембране ядра конец в конец и, упаковываясь, притягиваются к мембране, лептотенные хромосомы своими теломерными участками (концами) располагаются в одном из полюсов ядра, образуя фигуру «букета» у животных и сжатие в клубок «синезис» — у растений. Такое расположение или ориентации в ядре позволяет хромосомам быстрее и легче осуществлять конъюгацию гомологичных локусов хромосом (рис. 1).
Центральное событие — таинственный процесс узнавания гомологичных хромосом и их попарное сближение друг с другом происходит в зиготене профазы I. При конъюгации (сближении) гомологичных хромосом происходит образование пар — бивалентов и хромосомы заметно укорачиваются. С этого момента начинается формирование синаптонемного комплекса (СК). Формирование синаптонемного комплекса и синопсис хромосом — синонимы.
Рис. 1. Стадия профазы
В ходе следующей стадии профазы I – пахитене между гомологичными хромосомами усивается тесное соприкосновение, которое и называется синапсисом (от греч. synopsis — соединение, связь). Хромосомы в этой стадии сильно спирализованы, что делает возможным наблюдение их под микроскопом.
В ходе синапсиса гомологи переплетаются, т.е. конъюгируют. Конъюгирующие биваленты связаны хиазмами. Каждый бивалент состоит из двух хромосом и четырех хроматид, где каждая хромосома пришла от своего родителя. При образовании синапсиса (СК), происходит обмен участками между гомологичными хроматидами. Этот процесс, называемый кроссинговером, приводит к тому, что хроматиды теперь имеют иной состав генов.
Синаптонемный комплекс (СК) в пахитене достигает наибольшего развития и в этот период представляет собой лентовидную структуру, располагающуюся в пространстве между параллельно лежащими гомологичными хромосомами. СК состоит из двух параллельных латеральных элементов, сформированных плотно уложенными белками и менее плотного центрального элемента, протягивающегося между ними (рис. 2).
Рис. 2. Схема синаптонемного комлекса
Каждый латеральный элемент формируется парой сестринских хроматид в виде продольной оси лептотенной хромосомы и до того, как становится частью СК, носит название осевого элемента. Боковые петли хроматина лежат вне СК, окружая его со всех сторон.
Развитие СК в процессе мейоза:
лептотена—структура хромосом, вступивших в лептотену, сразу же оказывается необычной: в каждом гомологе наблюдается продольный тяж, идущий по оси хромосом на всем ее протяжении;
зиготена — на этой стадии осевые тяжи гомологов сближаются, при этом концы осевых тяжей, прикрепленных к ядерной мембране, как бы скользят по ее внутренней поверхности навстречу друг к другу;
пахитена. Наибольшее развитие СК достигает в пахитене, когда все элементы его приобретают максимальную плотность, а хроматин — вид плотной сплошной «шубы» вокруг него.
Функции СК:
1.Полностью развитый синаптонемный комплекс необходим для нормального удержания гомологов в биваленте так долго, как это необходимо для осуществления кроссинговера и закладки хиазм. Хромосомы соединяются с помощью синаптонемного комплекса на некоторое время (от 2 ч у дрожжей до 2–3 сут. у человека), в течение которого между гомологичными хромосомами совершается обмен гомологичными участками ДНК — кроссинговер (от англ, crossing over — образование перекреста).
2.Предотвращение слишком прочного соединения гомологов и удержание их на определенном расстоянии, сохранение их индивидуальности, создание возможности оттолкнуться в диплотене и разойтись в анафазе.
Процесс кроссинговера связан с работой определенных ферментов, которые при образовании хиазм между сестринскими хроматидами, «разрезают» их в месте перекреста с последующим воссоединением образовавшихся фрагментов. В большинстве случаев указанные процессы не приводят к каким-либо нарушениям в генетической структуре гомологичных хромосом, т.е. происходит правильное соединение фрагментов хроматид и восстановление их первоначального строения.
Однако, возможен и другой (более редкий) вариант событий, который связан с ошибочным воссоединением фрагментов разрезанных структур. При этом происходит взаимный обмен участками генетического материала между конъюгирующими хроматидами (генетическая рекомбинация).
На рис. 3 приведена упрощенная схема некоторых возможных вариантов одиночного либо двойного кроссинговера с участием двух хроматид из пары гомологичных хромосом. Необходимо подчеркнуть, что кроссинговер представляет собой случайное событие, которое с той или иной вероятностью может возникнуть на любом участке (либо на двух и большем числе участков) гомологичных хромосом. Следовательно, на этапе созревания гамет эукариотического организма в профазе первого деления мейоза действует универсальный принцип случайного (свободного) комбинирования (рекомбинации) генетического материала гомологичных хромосом.
В цитологических исследованиях синапсиса в последние два десятилетия важную роль играет метод распластывания профазных мейотических клеток животных и растений под действием гипотонического раствора. Метод вошел в цитогенетику после работ Мозеса и сыграл такую же роль, какую в свое время сыграл метод приготовления «давленых» препаратов для исследования метафазных хромосом, избавив цитогенетиков от микротомных срезов.
Метод Мозеса и его модификации стали более удобными, чем анализ СК на ультратонких срезах. Этот метод был положен в основу исследований мейоза и постепенно охватил вопросы генного контроля мейоза у животных и растений.
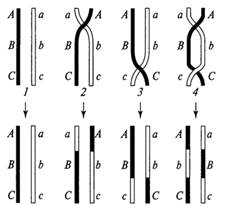
Рис. 3. Отдельные варианты одиночного и двойного кроссинговера с участием двух хроматид: 1 исходные хроматиды и вариант без кроссинговера; 2 одиночный кроссинговер на участке А В и кроссоверные хроматиды; 3 одиночный кроссинговер на участке В-С и кроссоверные хроматиды; 4 двойной кроссинговер и кроссоверные хроматиды нескольких разных участках на основе гомологичности генетического материала этих участков. Полагают, что с каждой стороны в процессе конъюгации могут участвовать либо одна из двух сестринских хроматид соответствующей хромосомы либо обе хроматиды.
В диппотене гомологичные хромосомы после спаривания и кроссинговера начинают отталкиваться друг от друга. Процесс отталкивания начинается с центромер. Расхождению гомологов препятствуют хиазмы — место соединения несестринских хроматид, возникших в результате перекреста. По мере расхождения хроматид некоторые хиазмы смещаются к концу плеча хромосомы. Обычно перекрестов бывает несколько, и чем длиннее хромосомы, тем их больше, поэтому в диплотене, как правило, несколько хиазм в одном биваленте.
В стадии диакинеза происходит уменьшение числа хиазм. Биваленты располагаются по периферии ядра. Ядрышко растворяется, мембрана разрушается и начинается переход к метафазе I. На протяжении всей профазы сохраняется ядрышко и ядерная оболочка. Перед профазой в период синтетического периода интерфазы происходит репликация ДНК и репродукция хромосом. Однако полностью этот синтез не заканчивается: ДНК синтезируется на 99,8%, а белки — на 75%. Синтез ДНК заканчивается в пахитене, белков — в диплотене.
В метафазе I становится заметной веретеновидная структура, образуемая микротрубочками. В ходе мейоза к центромерам хромосом каждого бивалента прикрепляются отдельные микрокрубочки. Затем пары хромосом перемещаются в экваториальную плоскость клетки, где выстраиваются в случайном порядке. Центромеры гомологичных хромосом располагаются в противоположных сторонах от экваториальной плоскости; в метафазе митоза, напротив, центромеры отдельных хромосом располагаются в экваториальной плоскости.
В метафазе I биваленты располагаются в центре клетки, в зоне экваториальной пластинки (рис. 4).
Рис. 4. Стадии мейоза: профаза I — метафаза I
Анафаза начинается с расхождения гомологичных хромосом и движения их в направлении полюсов. У хромосом без центромера крепления не может существовать. В анафазе митоза центромеры делятся и идентичные хроматиды расходятся. В анафазе I мейоза центромеры не делятся, хроматиды остаются вместе, а разъединяются гомологичные хромосомы. Однако из-за обмена фрагментами в результате кроссинговера хроматиды не идентичны, как в начале мейоза. В анафазе I конъюгирующие гомологи расходятся к полюсам.
В дочерних клетках число хромосом вдвое меньше (гаплоидный набор), при этом масса ДНК уменьшается также вдвое и хромосомы остаются дихроматидными. Точное расхождение гомологичных пар к противоположным полюсам лежит в основе редукции их числа.
В телофазе I происходит сосредоточение хромосом у полюсов, некоторая их деконденсация, за счет чего спирализация хромосом ослабевает, они удлиняются и снова становятся неразличимыми (рис. 5). По мере того как телофаза постепенно переходит в интерфазу, из эндоплазматического ретикулума возникает ядерная оболочка (в том числе и из фрагментов оболочки ядра материнской клетки), а также клеточная перегородка. Наконец вновь образуется ядрышко и возобновляется синтез белка.
Рис. 5. Стадии мейоза: анафаза I — телофаза I
В интеркинезе образуются ядра, в каждой из которых находится n дихроматидных хромосом.
Особенность второго деления мейоза состоит, прежде всего, в том, что в интерфазе II не происходит удвоения хроматина, поэтому каждая клетка, вступающая в профазу II, сохраняет прежнее соотношение n2с.
Второе деление мейоза
В период второго деления мейоза сестринские хроматиды каждой хромосомы расходятся к полюсам. Поскольку в профазе I мог произойти кроссинговер и сестринские хроматиды могли стать неидентичными, то принято говорить, что второе деление протекает по типу митоза, однако это не настоящий митоз, при котором в норме дочерние клетки содержат хромосомы идентичные по форме и набору генов.
В начале второго мейотического деления хроматиды все еще связаны центромерами. Это деление похоже на митоз: если в телофазе I образовалась ядерная оболочка, то теперь она разрушается, и к концу короткой профазы II исчезает ядрышко.
Рис. 6. Стадии мейоза: профаза II—метафаза II
В метафазе II снова можно увидеть веретено и хромосомы, состоящие из двух хроматид. Хромосомы прикрепляются центромерами к нитям веретена и выстраиваются в экваториальной плоскости (рис. 6). В анафазе II центромеры делятся и расходятся, а сестринские хроматиды, ставшие теперь хромосомами, движутся к противоположным полюсам. В телофазе II образуются новые ядерные оболочки и ядрышки, сжатие хромосом ослабевает и в интерфазном ядре они становятся невидимыми (рис. 7).
Рис. 7. Стадии мейоза: анафаза II — телофаза II
Завершается мейоз формированием гаплоидных клеток — гаметы, тетрады спор — потомков исходной клетки с редукционным вдвое (гаплоидным) набором хромосом и гаплоидной массой ДНК (исходная клетка 2n, 4с, — споры, гаметы — n, с).
Общая схема распределения хромосом гомологичной пары и содержащихся в них двух пар различающихся аллельных генов во время двух делений мейоза приведена на рис.8. Как видно из этой схемы, возможны два принципиально разных варианта такого распределения. Первый (более вероятный) вариант связан с образованием двух типов генетически различающихся гамет с хромосомами, не претерпевшими кроссинговеров на участках, где локализованы рассматриваемые гены. Такие гаметы принято называть некроссоверными. При втором (менее вероятном) варианте наряду с некроссоверными возникают также кроссоверные гаметы как результат генетического обмена (генетической рекомбинации) в участках гомологичных хромосом, расположенных между локусами двух неаллельных генов.
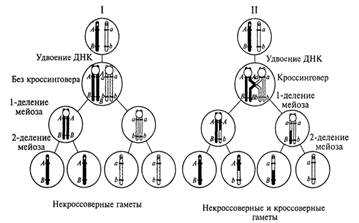
Рис. 8. Два варианта распределения хромосом гомологичной пары и содержащихся в них неаллельных генов как результат двух делений мейоза
Похожие статьи:
poznayka.org
Мейоз | Фазы мейоза
Мейоз(греч. meiosis – уменьшение, убывание) или редукционное деление. В результате мейоза происходит уменьшение числа хромосом, т.е. из диплоидного набора хромосом (2п) образуется гаплоидный (n).
Мейоз состоит из 2-х последовательных делений:
I деление называется редукционное или уменьшительное.
II деление называется эквационное или уравнительное, т.е. идет по типу митоза (значит число хромосом в материнской и дочерних клетках остается прежним).
Биологический смысл мейоза заключается в том, что из одной материнской клетки с диплоидным набором хромосом образуется четыре гаплоидные клетки, таким образом количество хромосом уменьшается в два раза, а количество ДНК в четыре раза. В результате такого деления образуются половые клетки (гаметы) у животных и споры у растений.
Фазы называются также как и в митозе, а перед началом мейоза клетка также проходит интерфазу.
Профаза I – самая продолжительная фаза и ее условно делят на 5 стадий:
1) Лептонема (лептотена) – или стадия тонких нитей. Идет спирализация хромосом, хромосома состоит из 2-х хроматид, на еще тонких нитях хроматид видны утолщения или сгустки хроматина, которые называются – хромомерами.
2) Зигонема (зиготена, греч. сливающиеся нити) — стадия парных нитей. На этой стадии попарно сближаются гомологичные хромосомы (одинаковые по форме величине), они притягиваются и прикладываются друг к другу по всей длине, т.е. коньюгируют в области хромомеров. Это похоже на замок «молния». Пару гомологичных хромосом называют биваленты. Число бивалентов равно гаплоидному набору хромосом.
3) Пахинема (пахитена, греч. толстая) – стадия толстых нитей. Идет дальнейшая спирализация хромосом. Затем каждая гомологичная хромосома расщепляется в продольном направлении и становится хорошо видно, что каждая хромосома состоит из двух хроматид такие структуры называют тетрадами, т.е. 4 хроматиды. В это время идет кроссинговер, т.е. обмен гомологичными участками хроматид.
4) Диплонема (диплотена) – стадия двойных нитей. Гомологичные хромосомы начинают отталкиваться, отходят друг от друга, но сохраняют взаимосвязь при помощи мостиков – хиазм, это места где произойдет кроссинговер. В каждом соединении хроматид (т.е. хиазме), осуществляется обмен участками хроматид. Хромосомы спирализуются и укорачиваются.
5) Диакинез – стадия обособленных двойных нитей. На этой стадии хромосомы полностью уплотнены и интенсивно окрашиваются. Ядерная оболочка и ядрышки разрушаются. Центриоли перемещаются к полюсам клетки и образуют нити веретена деления. Хромосомный набор профазы I составляет — 2n4c.
Таким образом, в профазу I происходит:
1. конъюгация гомологичных хромосом;
2. образование бивалентов или тетрад;
3. кроссинговер.
В зависимости от конъюгирования хроматид могут быть различные виды кроссинговера: 1 – правильный или неправильный; 2 – равный или неравный; 3 – цитологический или эффективный; 4 – единичный или множественный.
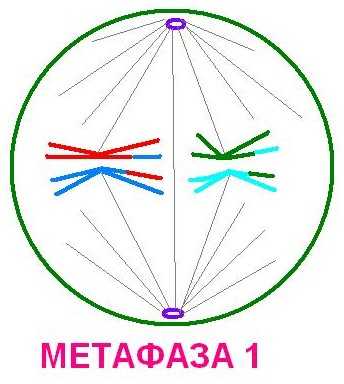
Метафаза I – спирализация хромосом достигает максимума. Биваленты выстраиваются вдоль экватора клетки, образуя метафазную пластинку. К центромерам гомологичных хромосом крепятся нити веретена деления. Биваленты оказываются соединенными с разными полюсами клетки.
Хромосомный набор метафазы I составляет — 2n4c.
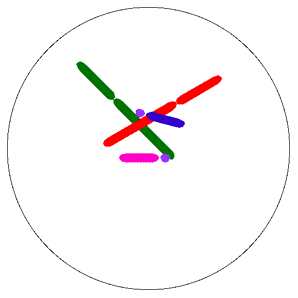
Анафаза I – центромеры хромосом не делятся, фаза начинается с деления хиазм. К полюсам клетки расходятся целые хромосомы, а не хроматиды. В дочерние клетки попадает только по одной из пары гомологичных хромосом, т.е. идет их случайное перераспределение. На каждом полюсе, оказывается, по набору хромосом — 1п2с, а в целом хромосомный набор анафазы I составляет — 2n4c.
Телофаза I – по полюсам клетки находится целые хромосомы, состоящие из 2-х хроматид, но количество их стало в 2 раза меньше. У животных и некоторых растений хроматиды деспирализуются. Вокруг них на каждом полюсе формируется ядерная мембрана.
Затем идет цитокинез
. Хромосомный набор образовавшихся после первого деления клеток составляет — n2c.
Между I и II делениями нет S-периода и не идет репликация ДНК, т.к. хромосомы уже удвоены и состоят из сестринских хроматид, поэтому интерфазу II называют интеркинезом – т.е. происходит перемещение между двумя делениями.
Профаза II – очень короткая и идет без особых изменений, если в телофазу I не образуется ядерная оболочка, то сразу образуются нити веретена деления.
Метафаза II – хромосомы выстраиваются вдоль экватора. Нити веретена деления крепятся к центромерам хромосом.
Хромосомный набор метафазы II составляет — n2c.
Анафаза II – центромеры делятся и нити веретена деления разводят хроматиды к разным полюсам. Сестринские хроматиды называются дочерними хромосомами(или материнские хроматиды это и будут дочерние хромосомы).
Хромосомный набор анафазы II составляет — 2n2c.
Телофаза II – хромосомы деспирализуются, растягиваются и после этого плохо различимы. Образуются ядерные оболочки, ядрышки. Телофаза II завершается цитокинезом.
Хромосомный набор после телофазы II составляет – nc.
Схема мейотического деления
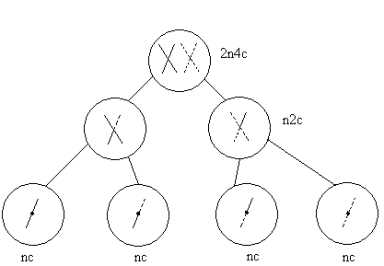
buzani.ru
Био: Мейоз
Мейоз — способ деление эукариотических клеток, при котором из одной диплоидной формируется 4 гаплоидные. В результате мейоза число хромосом уменьшается в 2 раза. поэтому его еще называют редукционным делением (правильнее редукционным называть только первое деление мейоза, а второе — эквационное). Мейозом образуются половые клетки животных и споры высших растений (из которых развиваются гаметофиты — половое поколение, образующее гаметы путем митоза).При мейозе происходит два быстро следующих друг за другом деления, каждое из которых состоит из 4 уже известных нам фаз: профазы, метафазы, анафазы и телофазы; между двумя делениями может быть короткая интерфаза, но никогда не происходит репликации ДНК.
Ход мейоза.
Перед началом мейоза (как и перед началом митоза) происходит удвоение наследственной информации клетки, т.е. ДНК реплицируется, и хромосомный набор имеет формулу 2n4с.первое деление мейоза — редукционное.
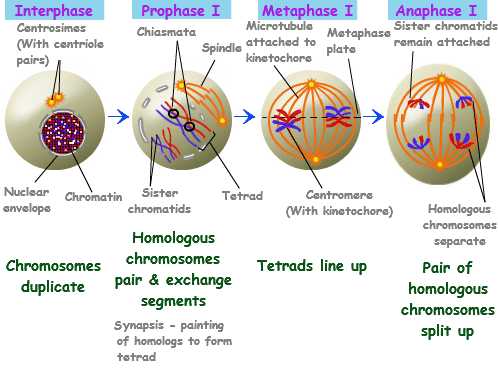
Профаза 1. 2n4c . Самая длительная фаза мейоза. Ее часто делят на пять стадий (лептотена, зиготена, пахитена, диплотена и диакинез). Хромосомы укорачиваются и становятся видимыми как обособленные структуры. Гомологичные хромосомы, происходящие из ядер материнской и отцовской гамет, приближаются одна к другой и конъюгируют. Они одинаковой длины, их центромеры занимают одинаковое положение, и они обычно содержат одинаковое число генов, расположенных в одной и той же линейной последовательности. Пары конъюгирующих гомологичных хромосом называют бивалентами. Биваленты укорачиваются и утолщаются, становятся ясно видны.
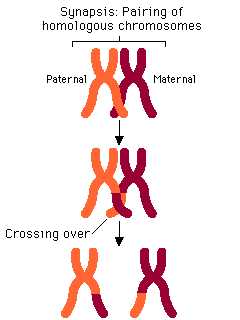
Гомологичные хромосомы, составляющие бивалент, частично разделяются, становится видно, что каждая состоит из двух хроматид. Хромосомы остаются соединенными в нескольких точках – хиазмах. В каждой хиазме происходит обмен участками хроматид в результате разрывов и соединений, в которых участвуют две из имеющихся в хиазме четырех нитей. В результате гены из одной хромосомы оказываются связанными с генами другой хромосомы, что приводит к новым генным комбинациям в образующихся хроматидах. Этот процесс называется кроссинговер. После кроссинговера гомологичные хромосомы не расходятся, а остаются прочно связанными. В клетке центриоли мигрируют к полюсам, ядрышки и ядерная мембрана разрушаются, образуются нити веретена деления.
Метафаза 1. 2n4c. Биваленты выстраиваются в экваториальной плоскости, образуя метафазную пластинку. Их центромеры ведут себя как единые структуры и организуют прикрепленные к ним нити веретена деления. В результате тянущего усилия каждый бивалент оказывается на экваторе, а обе его центромеры равноудалены от экватора (снизу и сверху).
Анафаза 1. 1n2c * 2 (к каждому полюсу клетки!) Имеющиеся у каждого бивалента две центромеры еще не делятся, но сестринские хроматиды уже не примыкают одна к другой. Нити веретена тянут центромеры, каждая из которых связана с двумя хроматидами, к противоположным полюсам веретена. В результате хромосомы разделяются на два гаплоидных набора, попадающих в дочерние клетки. Гомологичные хромосомы каждой пары расходятся к полюсам независимо от хромосом другой пары.
Телофаза 1. 1n2c в каждой образующейся клетке. Расхождение гомологичных центромер и связанных с ними хроматид к противоположным полюсам означает завершение первого деления мейоза. Число хромосом в одном наборе стало вдвое меньше, но находящиеся на каждом полюсе хромосомы состоят из двух хроматид. Вследствие кроссинговера эти хроматиды генетически неидентичны. Веретена и их нити обычно исчезают. Иногда после этой фазы хромосомы деспирализуются и возникает ядерная оболочка. Затем происходит деление цитоплазмы. Иногда не наблюдается этой фазы, и клетка переходит от анафазы1 к профазе2.
Второе деление мейоза — эквационное.
Интерфаза 2.1n2c в каждой клетке. Эта стадия обычна только в животных клетках; продолжительность варьирует. Фаза S отсутствует, и дальнейшей репликации ДНК не происходит. Синтезируются необходимые вещества, главным образом, АТФ.
Профаза 2. 1n2c. Если не было телофазы1, то этой стадии тоже нет (обратные процессы). Ядерные мембраны и ядрышки разрушаются, хроматиды укорачиваются и утолщаются. Центриоли перемещаются к противоположным полюсам, появляются нити веретена. Хроматиды располагаются таким образом, что их длинные оси перпендикулярны оси веретена первого деления мейоза.
Метафаза 2. 1n2c. Центромеры ведут себя как двойные структуры. Они организуют нити веретена, направленные к обоим полюсам, и таким образом выстраиваются по экватору веретена.
Анафаза 2. 1n1c * 2 (к каждому полюсу клетки!) . Центромеры делятся, и нити веретена деления растаскивают их к противоположным полюсам. Центромеры тянут за собой отделившиеся друг от друга хроматиды, которые теперь называются хромосомами.
Телофаза 2. 1n1c (в каждой клетке). Сходна с телофазой митоза. Хромосомы деспирализуются, растягиваются и после этого плохо различимы. Нити веретена исчезают, а центриоли реплицируются. Вокруг каждого ядра, которое содержит теперь гаплоидное число хромосом, образуется ядерная мембрана. Далее следует деление цитоплазмы. Образуется 4 дочерние клетки.
 Значение мейоза:
1. Половое
размножение. Предотвращение удвоения
числа хромосом в каждом последующем поколении.
2. Генетическая
изменчивость. Мейоз создает возможности для возникновения в гаметах новых
генных комбинаций.
Значение мейоза:
1. Половое
размножение. Предотвращение удвоения
числа хромосом в каждом последующем поколении.
2. Генетическая
изменчивость. Мейоз создает возможности для возникновения в гаметах новых
генных комбинаций.
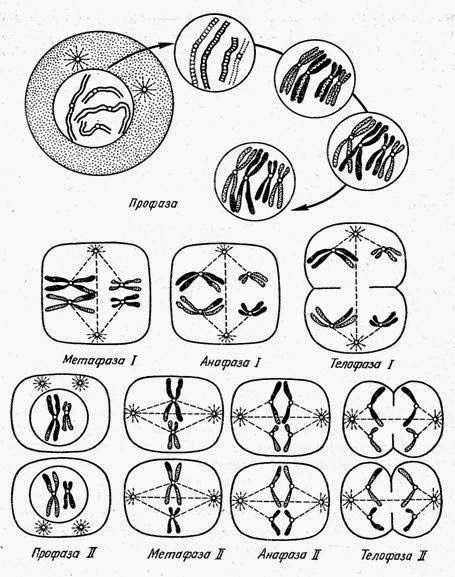
Сходства митоза и мейоза:
Черты отличия митоза и мейоза
|
1. Одно деление;
2. В метафазе по экватору выстраиваются удвоенные
хромосомы;
3. Конъюгация и кроссинговер отсутствуют;
4. Между делениями имеется интерфаза, в которую
происходит удвоение ДНК;
5. Образуются две диплоидные клетки.
Видео на тему: |
1. Два быстро следующих друг за другом деления;
2. По экватору выстраиваются пары гомологичных
хромосом;
3. Гомологичные хромосомы конъюгируют, происходит
кроссинговер;
4. Между двумя делениями мейоза нет интерфазы
(короткая) и не происходит удвоение ДНК;
5. Образуются четыре гаплоидные клетки. |
myblog-bio.blogspot.com
10. Гаметогенез и развитие растений. Размножение организмов
10. Гаметогенез и развитие растений
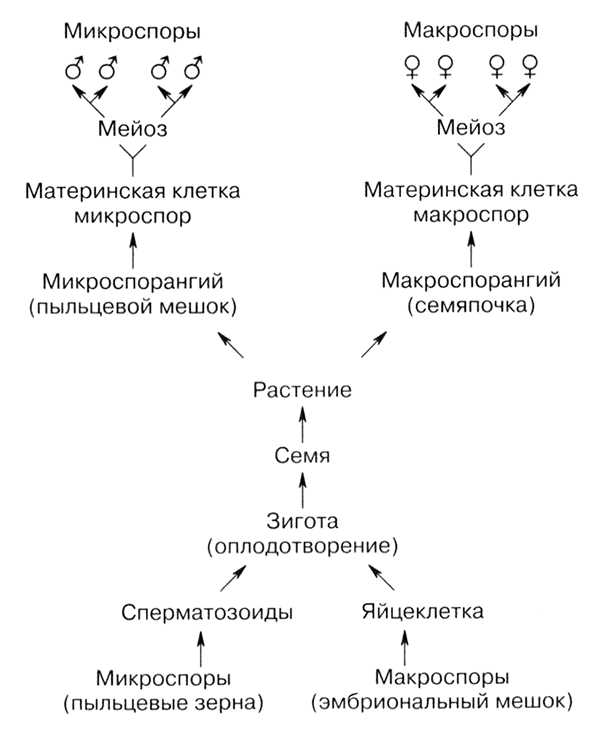
Мейоз в жизненном цикле растений. У растений гаметогенез и размножение протекают иначе, чем у животных. Процесс мейоза происходит у них не на стадии образования гамет, а на стадии образования спор. Кроме того, у растений наблюдается чередование поколений с диплоидным (2n) и гаплоидным (n) набором хромосом.
Поколение с гаплоидным набором хромосом называется гаметофитом. На гаметофите образуются гаметы в процессе митоза. Поколение с диплоидным набором хромосом называется спорофитом, и на нем образуются споры в процессе мейоза. Гаметофит развивается из гаплоидных спор, а спорофит — из диплоидной зиготы, образующейся в результате оплодотворения.
Смена поколений идет по схеме: зигота (2n) —> спорофит (2n) —> мейоз —> споры (n) —> гаметофит (n) —> митоз —> гаметы (n) —>оплодотворение —> зигота (2n)
В цикле развития мейоз всегда происходит один раз. В зависимости от периода жизни спорофита и гаметофита взрослое растение может быть гаплоидным или диплоидным.
Размножение и развитие зеленых водорослей
У низших растений преобладающим поколением является гаметофит. Он размножается бесполым путем, образуя клетки, из которых развиваются взрослые особи. В определенный период на гаметофите образуются гаметы, разные или одинаковые по величине.
После слияния гамет образуется зигота, которая сразу же делится мейозом и образует споры, дающие начало новым гаметофитам. Таким образом, в жизненном цикле зеленых водорослей диплоидное поколение представлено только одной клеткой — зиготой.
Размножение и развитие высших споровых растений
У мхов, папоротников, плаунов и хвощей размножение происходит спорами.
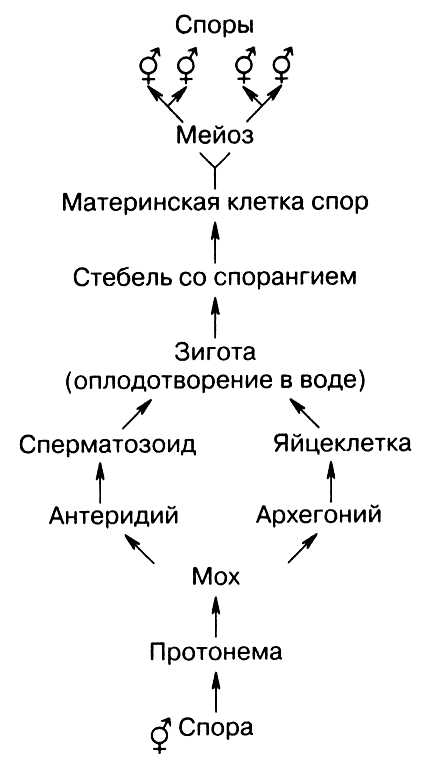
У мхов взрослым растением является гаметофит (n) — половое поколение, которое развивается при прорастании споры (рис. 19).
Рис. 19. Схема образования (А) и развития (Б) зародышевого мешка: 1 — антиподы; 2 — два центральных ядра; 3 — синергиды; 4 — яйцеклетка

Это листостебельное растение, на побегах которого развиваются органы полового размножения — антеридии (мужские органы) и архегонии (женские органы). В антеридиях в процессе митоза образуются сперматозоиды, а в архегониях — яйцеклетка (как правило, одна). При наличии воды сперматозоиды проникают в архегонии и оплодотворяют яйцеклетку, при этом образуется зигота (2n). Из зиготы на гаметофите развивается спорофит (2n) в виде коробочки на ножке — спорогона. Спорофит не способен к самостоятельному существованию и питается за счет питательных веществ гаметофита. В спорангиях (коробочке) в результате мейоза образуются споры. Споры после созревания высыпаются и во влажной среде прорастают, давая начало новым гаметофитам.
Схема жизненного цикла мхов
У папоротников, плаунов и хвощей, наоборот, взрослым растением является спорофит, на котором в специальных органах — спорангиях в результате мейоза образуются споры (n). Споры после созревания высыпаются и прорастают (рис. 20).
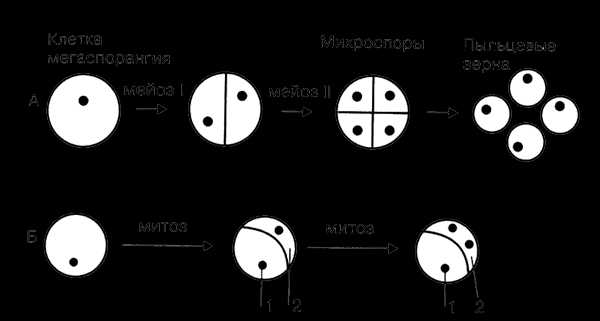
Рис. 20. Схема образования (А) и развития (Б) пыльцевого зерна: 1 — вегетативная клетка; 2 — генеративная клетка
При прорастании споры развивается половое поколение — гаметофит, который представлен небольшим заростком и существует очень недолго. На заростке развиваются органы полового размножения — антеридии и архегонии, в которых в результате митоза развиваются гаметы. В антеридиях образуются сперматозоиды, а в архегониях созревает яйцеклетка.
Схема жизненного цикла папоротников
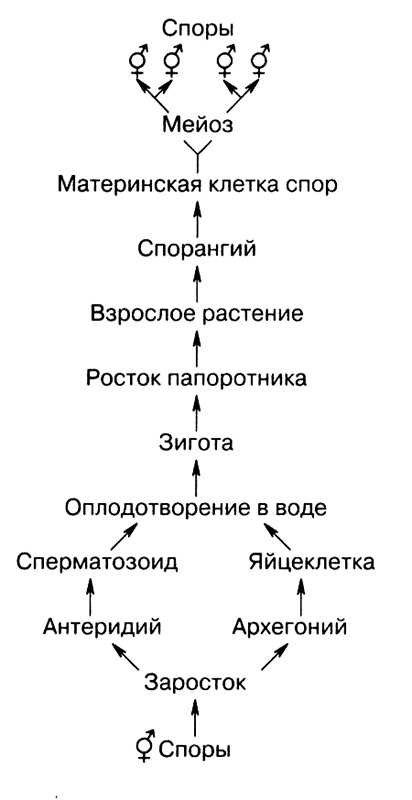
При наличии капельно-жидкой влаги сперматозоиды проникают в архегонии и оплодотворяют яйцеклетку, при этом образуется зигота. Из зиготы развивается зародыш, а далее молодое растение — спорофит.
Размножение и развитие семенных растений
У семенных растений размножение происходит семенами. Преобладающим поколением является спорофит, а гаметофит сильно редуцирован, развивается в спорофите и представлен лишь несколькими клетками.
Схема жизненного цикла голосеменных (на примере сосны)
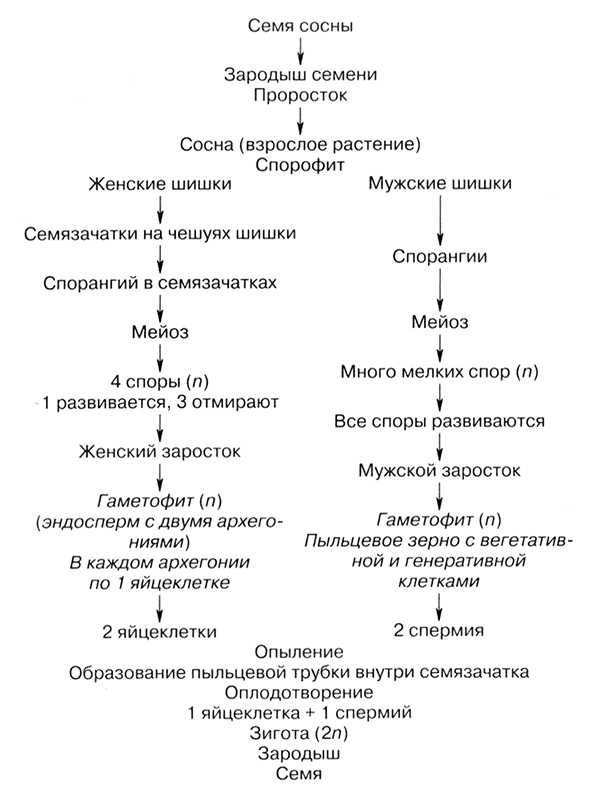
Процесс развития семенных растений рассмотрим более подробно на примере цветковых растений. Взрослое растение имеет диплоидный набор хромосом и является спорофитом. Оно развивается из семени.
Репродуктивным органом является цветок. В цветке образуются женский орган — пестик и мужской — тычинки.
В завязи пестика в семязачатках из спорогенной ткани в результате мейоза образуются 4 споры (n), причем одна из них, крупная — мегаспора развивается в женский заросток — гаметофит; три другие, более мелкие, отмирают.
Мегаспора трижды делится митозом, и образуется восьмиядерный зародышевый мешок. Восемь ядер распределяются следующим образом. Ближнее к пыльцевходу крупное ядро — яйцеклетка, рядом два ядра помельче — две сопутствующие клетки — синергиды. На противоположном полюсе мешка располагаются три ядра — антиподы, а в центре располагаются два центральных ядра. Все ядра гаплоидные. Таким образом, женский гаметофит представлен восьмиядерным зародышевым мешком.
В тычинках, в пыльцевых мешках из ткани микроспорангия в результате мейоза образуется много мелких микроспор (n). Все споры развиваются и дают начало мужскому заростку — гаметофиту. Спора делится митозом и образует вегетативную и генеративную клетки. Ядро генеративной клетки делится еще раз, и образуется два спермия. Вегетативная и генеративная клетки покрываются оболочкой, образуется пыльцевое зерно. Таким образом, мужской гаметофит представлен двумя клетками.
При попадании пыльцы на рыльце пестика вегетативная клетка начинает прорастать, образуя пыльцевую трубку и продвигая генеративную клетку к пыльцевходу. Два спермия через пыльцевход проникают в зародышевый мешок. Один спермий сливается с яйцеклеткой, и образуется зигота (2n), из которой развивается зародыш семени. Другой спермий сливается с двумя ядрами центральной клетки, в результате чего образуется эндосперм (3n) семени, в котором запасаются питательные вещества.
Схема жизненного цикла покрытосеменных растений
Этот процесс называется двойным оплодотворением. Он был открыт русским ученым С. П. Навашиным. В результате двойного оплодотворения в семязачатке образуется семя, а из покрова семязачатка — семенная кожура. Вокруг семени из завязи и других частей цветка образуется плод (рис. 21).
Рис. 21. Схема двойного оплодотворения цветковых растений: 1 — пыльца; 2 — пыльцевая трубка вегетативной клетки; 3 — два спермия; 4 — яйцеклетка; 5 — два центральных ядра; 6 — семязачаток; 7 — эндосперм семени; 8 — зародыш; 9 — кожура семени из покрова семязачатка

У растений, при переходе от низших к высшим, наблюдается постепенное увеличение срока жизни спорофита. Начиная с папоротникообразных, у всех растений взрослый организм представлен спорофитом, а гаметофит претерпевает постепенно редукцию до одной или нескольких клеток.
Вопросы для самоконтроля
1. В чем заключается особенность жизненного цикла растений по сравнению с животными?
2. Как происходит чередование поколений у растений?
3. Что такое гаметофит и спорофит? В чем их отличие?
4. Какое поколение является господствующим у водорослей и мхов, папоротникообразных и семенных растений?
5. Как изменяется гаметофит от низших растений к высшим?
6. В чем их сходство и отличие гаметофита цветковых растений и мхов?
7. Какой процесс называется двойным оплодотворением? В чем его особенность?
Поделитесь на страничкеСледующая глава >
bio.wikireading.ru
Мейоз | Cell Biology.ru
Основные события мейоза
Продолжительность мейоза
Премейотическая интерфаза
Профаза I
Лептотена
Зиготена
Пахитена
Диплотена
Диакинез
Метафаза I
Анафаза
Телофаза
Интеркинез
Второе деление мейоза
Гаметогенез
Основные события мейоза
Мейоз – тип митоза, или редукционное деление, при котором из одной клетки образуется четыре, каждая из которых имеет вдвое меньше хромосом чем исходная; т.е. число хромосом уменьшается с диплоидного (2n) до гаплоидного (n). Мейоз происходит при образовании гамет — гаметный, при образовании спор — споровый и мейоз может быть зиготным. Смысл гаметного мейоза сводится к образованию половых клеток, при слиянии которых, восстанавливается число хромосом (до диплоидного) характерное для соматических клеток данного вида. Споровый тип мейоза происходит у растений для которых характерно чередование поколений — гаплоидного, размножающегося бесполым путем и диплоидного, размножающегося половым путем (см. обзор Размножение).
рис.1 Принципиальная схема мейоза. В соматических диплоидных клетках содержат две гомологичные хромосомы, одна отцовская и одна материнская. Они удваиваются в S-фазе клеточного цикла, образуя две пары сестринских хроматид. Хромосомы сближаются и между ними происходит кроссинговер — обмен участками между материнской и отцовской парами хроматид с образованием хроматид содержащих отцовские и материнские гены. Хромосомы конденсируются, выстраиваются и расходятся. Затем происходит второе деление мейоза.Стадии мейоза 2n—>S—>4n—>2x2n—>4x1n
Мультипликация, демонстрирующая основные события мейоза: meiosis.mpg [2,65 Mb]
Мейоз разделяют на ряд стадий, которые можно различить в световой микроскоп.
Первое деление мейоза
Профаза I мейоза разделяетя на ряд стадий.
лептотена (стадия тонких нитей) начинается спирализация х-м
зиготена (стадия сливающихся нитей),сближение и начало конъюгации гомологичных х-м, кот объединяются в бивалент
пахитена (стадия толстых нитей) м-у гомологичными х-мами осуществляется кроссинговер
диплотена (стадия двойных нитей) отталкивание гомологичных х-м, кот отделяются др от др в области центромер, но остаются связанными в областях прошедшего кроссинговера – хиазмах
диакинез (стадия обособления двойных нитей) гомологичные х-мы удерживаются в месте лишь в отдельных точках хиазм – уменьшение числа хиазм, компактность бивалентов
Метафаза I — завершается формирование веретена деления, его нити прикрепляются к центромерам хромосом, в результате чего биваленты устанавливаются в плоскости экватора веретена деления, образуя экваториальную пластинку.
Анафаза I — связи в бивалентах ослабляются и гомологичные хромосомы отходят друг от друга, направляясь к противоположным полюсам веретена деления. К каждому полюсу подходит гаплоидный набор хромосом, состоящий из двух хроматид.
Телофаза I — у полюсов веретена деления собирается одинарный гаплоидный набор хромосом, каждая из них содержит удвоенное количество ДНК (n2c).
Интеркинез — временной промежуток между первым и вторым делениями мейоза. Не всегда обязателен.
Второе мейотическое отделение (эквационное) протекает как митоз, только клетки, вступающие в него, несут гаплоидный набор хромосом.
ПРОДОЛЖИТЕЛЬНОСТЬ МЕЙОЗА
Продолжительность мейоза напрямую зависит от количества ДНК в ядре. Также продолжительность мейоза зависит от структуры хромосомной организации и особенностей данного вида.
| Вид | 2n | Время, ч | ДНК на клетку (пг) |
| Antirrhinum majus | 16 | 24.0 | 5.5 |
| Haploppapus gracilis | 4 | 36.0 | 5.5 |
| Secale cereale | 14 | 51.2 | 28.7 |
| Allium cepa | 16 | 96.0 | 54.0 |
| Tradescantia paludosa | 12 | 126.0 | 59.0 |
| Tulbaghia violacea | 12 | 130.0 | 58.5 |
| Lilium henryi | 24 | 170.0 | 100.0 |
| Lilium longiflorum | 24 | 192.0 | 106.0 |
| Trillium erectum | 10 | 274.0 | 120.0 |
ПРЕМЕЙОТИЧЕСКАЯ ИНТЕРФАЗА
ПРОФАЗА I
Лептотена
(стадия тонких нитей) начинается спирализация х-м
Зиготена
Происходит начало синапсисиса — спаривание гомологичных хромосмом с образованием бивалента, или синаптонемального комплекса, в котором хромосомы выровнены и соединены.
Во время зиготены образуется zDNA составляющая 0,3, 0,4% от всей ДНК. zDNA GC-обогащена и находится в блоках длиной 104 пн разбросанных вдоль хромосом. Добавление ингибиторов синтеза ДНК в зиготене, предотвращают синапсис.
Пахитена
Хромосомы конденсируются
Синаптонемальный комплекс
Мейоз проходит и в отсутствии синаптонемального комплекса, но без рекомбинации.
Диплотена
Диакинез
МЕТАФАЗА I
АНАФАЗА I
ТЕЛОФАЗА I
ИНТЕРКИНЕЗ
ПРОФАЗА II
МЕТАФАЗА II
АНАФАЗА II
ТЕЛОФАЗА II
ГАМЕТОГЕНЕЗ
СИНАПСИС
КРОССИНГОВЕР
Alberts, 2003
Alberts, 2003
Alberts,2003
www.cellbiol.ru
