
Меристема | Info-Farm.RU
Меристема (от греч. Meristos — делящийся) — боковая ткань растений, состоящая из недифференцированных клеток (меристематическая клеток), и находится в частях растений, где происходит рост.
Цитологические особенности меристем
Дифференцированные клетки растений в целом не могут делиться или превращаться в клетки других типов. Поэтому, деление клеток в меристемах необходим для обеспечения новых клеток для роста других тканей, образование новых органов и обеспечения структуры тела растения. По функции меристематическая клетки аналогичные стволовым клеткам животных, не дифференцируются или дифференцируются незначительно, и способные к непрерывному клеточного деления. Меристематическая клетки маленькие, а цитоплазма и ядро полностью заполняют клетку. Вакуоли чрезвычайно маленькие, а цитоплазма не содержит дифференцированных пластид (хлоропластов или хромопластов), хотя они присутствуют в рудиментарной форме (протопластиды). Меристематическая клетки плотно упакованы, почти без межклеточного пространства. Клеточная стенка — очень тонкая первичная клеточная стенка.
Физиологическая деятельность меристем требует баланса между двумя антагонистическими процессами: образованием новых тканей и поддержкой обновление популяции меристематическая клеток.
Меристема — образующая растительная ткань, из которой могут образовываться все другие ткани, меристематическая клетки длительное время сохраняют способность к делению с образованием новых неспециализированных клеток.
Клетки меристемы уплощенные, мелкие, плотно расположены друг возле друга, без межклитинникив, в центре клеток расположено ядро, вакуоли мелкие, многочисленные, клеточная оболочка первичная, имеющиеся только предшественники пластид.
Цитологические особенности меристем наиболее типично выражены в апикальных меристем. Клетки изодиаметрични многоугольники, не разделены межклетниками. Клеточные стенки тонкие, с небольшим содержанием целлюлозы. Цитоплазма густая, ядро большое, расположенное в центре. В цитоплазме большое количество рибосом и митохнодрий (происходит интенсивный синтез белков и других веществ). Многочисленные вакуоли очень мелкие.
Клетки латеральных меристем неодинаковы по размеру и форме. Это связано с отличием клеток постоянных тканей, из них образуются. Так, например, в камбии является паренхимной и прозенхимных клетки. С паренхимных инициалы образуется паренхима ведущих комплексов, с прозенхимных — собственно ведущие элементы.
Меристемы классифицируют по положению их в растении:
- Верхушечные (апикальные).
- Боковые (латеральные).
- Вставные (интеркалярный).
И по происхождению:
- первичные
- вторичные.
Первичные меристемы находятся в тесной связи с апикальными меристеме и непосредственно из них образуются, в частности, протодермы, основная меристема, прокамбий. Протодермы — однослойный слой малодифференцированных клеток, покрывающий апекс извне, в дальнейшем из нее формируется эпидерма. С основной меристемы формируется кора, а с прокамбия, клетки которого вытянуты вдоль оси стебля, формируется центральный цилиндр.
Вторичные меристемы возникают из специализированных клеток зрелых органов растений и обеспечивают их латеральный рост (феллоген, перицикл, камбий). Ткани, образуются из первичной меристемы, называются первичными, а с вторичной — вторичными. К вторичным меристем следует также отнести так называемые травматические меристемы, которые формируются в местах повреждения тела растений и обеспечивают регенерацию.
Среди перечисленных типов меристем первой в онтогенезе появляется апикальная меристема, с которой происходит дифференциация всех других тканей. Расположена она на верхушках (апекс) стебля и корня и их боковых ответвлений. Особенностью жизнедеятельности растительного организма является, в частности, сохранение образующей деятельности в апикальных меристем течение всего онтогенеза, поэтому говорят, что растения способны к неограниченному росту.
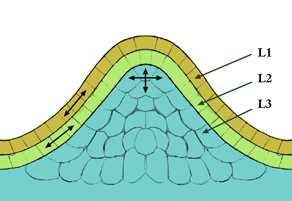
Апикальная меристемами
Апикальная меристема — представлена совокупностью клеток, расположенных на верхушках главной и боковых осей стебля и корня, за счет них органы растут в длину.
Апикальная меристемами локализуется на полюсах зародыша — кончике корня и почки. Обеспечивает рост корня и побега в длину. Апикальные меристемы первичные, они образуют конус нарастания корня и побега.
Латеральная меристема
Боковая меристема (латеральная) — клетки располагаются вдоль оси органов, обеспечивая их рост в толщину. Боковые меристемы могут быть как первичные (прокамбий, перицикл) так и вторичными (камбий, феллоген).
Латеральная меристема расположена по кругу осевых органов, образуя цилиндры, на поперечных срезах имеют вид кольца.
Первичные боковые меристемы — прокамбий, перицикл — возникают под апекс и находятся в непосредственной связи с ними.
Вторичные боковые меристемы — камбий и феллоген (пробковый камбий) — формируются позже с промеристем или постоянных тканей путем их дифференциации.
Боковые меристемы обеспечивают утолщение корня и стебля. С прокамбия и камбия образуются проводящие ткани, с феллогена — корка.
Феллоген (рус. Феллоген) — вторичная меристематическая ткань — формируется за счет периклинальных делений клеток перицикла и закладывается под эндодермой. Феллоген откладывает наружу вторичную покровную ткань — пробка. Деятельность феллогена приводит к тому, что первичная кора под давлением нарастающих вторичных тканей изолируется от центрального цилиндра, отмирает и отделяется.
Таким образом, на смену первичной формируется вторичная кора, представляет собой совокупность тканей, расположенных кнаружи от камбия.
Интеркалярный меристемами
Интеркалярный меристема (синоним — вставная меристема) — клетки располагаются между участками дифференцированных тканей, обеспечивая вставной рост.
Интеркалярный меристема, например, располагается в узлах злаков, у основания черешков, тычиночных нитей. Это остаточные первичные меристемы. Они происходят от веривкових меристем, но их преобразования в постоянные ткани задержано по сравнению с другими тканями стебля. Это нежные меристемы особенно хорошо заметны в злаков. В случае состояния хлебов они обеспечивают повышение стеблей за счет неравномерного деления клеток из нижнего и верхнего стороны соломинки.
Раневая, или травматическое меристемами
Образующиеся при ранении тканей и органов. Живые клетки, окружающие пораженные участки, дедиференциюються и начинают делиться, то есть превращаются в вторичную меристему. Раневые меристемы образуют калюс — плотную ткань биловатого или желтоватого цвета, состоящий из паренхимных клеток разного размера, расположенных не в порядке. Клетки Калюс имеют крупные ядра и относительно толстые клеточные стенки. Калюс может дать начало любой ткани или органа растения. На перефирии формируется пробка, возможна дифференциация клеток каллуса на другие ткани. В Калюсе могут закладаться дополнительные корни и почки. Калюс возникает при привиты растений, обеспечивает рост привоя и подвоя; в основе черенков. Также калюс используют для получения культуры изолированных тканей.
Изображения по теме
info-farm.ru
Описание меристемных тканей
Меристемы (греч. meristos — делимый, делитель) стоят особняком среди других тканей, поскольку состоят из живых недифференцированных клеток, способных постоянно делиться. В онтогенезе этот тип растительных тканей возникает первым в результате деления зиготы. На ранних этапах развития весь зародыш состоит из меристем. Затем из них образуются все ткани растения.
Процесс детерминации (лат. determinare — определять) происходит следующим образом. Сначала в клетке возникает полярность. Она выражается в неодинаковой организации полюсов клетки. Полярность наблюдается уже в яйцеклетке, которая находится в зародышевом мешке. Так, у нее выделяют халазный полюс и полюс, обращенный к пыльцевходу (микропиле). Полярность меристематических клеток, которые детерминируются в клетки различных тканей, определяется различными факторами окружающей среды. Такие факторы могут иметь физическую природу (температура, свет, сила притяжения Земли и т.д.) или химическую (ионы, фитогормоны и другие вещества, выделяемые микроокружением). Детерминация приводит к внутриклеточным биохимическим перестройкам, в результате чего клетка получает способность развиваться специализированно, т.е. морфологически дифференцироваться в специализированную клетку ткани, определенной детерминацией.
Следующим этапом будет рост клеток. Этот процесс проходит удивительно согласованно. При этом не происходит смещения клеточных стенок относительно друг друга и цитоплазматические связи между соседними клетками сохраняются. В результате дифференцировки клетка приобретает окончательные размеры и форму, но утрачивает способность размножаться. Вероятно, митостатическое воздействие оказывает микроокружение, так как выделение неделящихся клеток приводит к превращению ее в делящуюся клетку каллуса.
Сами меристемы состоят из клеток двух типов. Клетки первого типа называются инициальными (лат. initialis - начальный). Они способны неограниченно долго размножаться (аналогичные клетки животных называются стволовыми). При этом после деления одна дочерняя клетка сохраняет свойства инициальной, а другая, продолжая делиться, детерминируется как клетка определенной ткани и приступает к начальным этапам дифференцировки. Именно они представляют собой клетки второго типа.
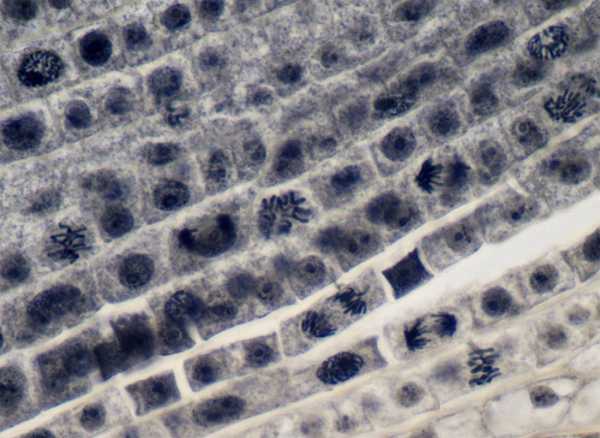
Клетки апикальной меристемы кончика корня лука
Инициальных клеток обычно бывает немного, иногда всего одна. Расположенные на апексах (верхушках осевых органов) они часто имеют форму многогранника (по граням здесь и происходят деления клетки).
Морфологически клетки меристем отличаются от других рядом признаков. Внешне они представляют собой похожие друг на друга относительно мелкие клетки с тонкими стенками, способными к растяжению. Ядро занимает центральное положение, вокруг ядра располагается сравнительно небольшое количество цитоплазмы. Характерно сильное развитие гранулярного эндоплазматического ретикулума, что свидетельствует об усиленном синтезе белков в клетке. Имеется много митохондрий, зато вакуоли мелкие, под световым микроскопом они не различимы.
Образовательные ткани в теле растения располагаются в разных местах, в связи с чем их делят на несколько групп.
Верхушечные, или апикальные, меристемы располагаются на верхушках (апексах) осевых органов — стебля или корня. С помощью этих меристем вегетативные органы растений осуществляют свой рост в длину. Если такие меристемы выделить из соответствующих органов культурных растений и вырастить на питательной среде, можно получить посадочный материал, не зараженный вирусами.
Латеральные меристемы также характерны для осевых органов, где располагаются концентрически, в виде муфты. Специализированные клетки здесь располагаются как внутри (ближе к сердцевине), так и снаружи (ближе к поверхности тела). Первичные латеральные меристемы образуются из апикальных и дают начало всем тканям растения. В дальнейшем их развитие у разных форм растений происходит по-разному. У древесных они сохраняются в течение всей жизни в виде камбия, обеспечивая вторичное утолщение. У травянистых форм латеральные меристемы быстро исчезают, поэтому вторичное утолщение у них не происходит.
Интеркалярные (лат. intercalate — внедрять), или вставочные, меристемы происходят от верхушечных. Они представляют собой группы клеток, еще способных размножаться, но уже вставших на путь дифференциации. Инициальных клеток среди них нет, зато много специализированных.
Интеркалярные меристемы можно обнаружить в основании молодых листьев. Они характерны для злаков, где располагаются в нижних частях междоузлий, окруженных влагалищем листа. Эта особенность позволяет подниматься полегшим после сильного ветра злакам — изгиб побега образуется именно в указанных выше местах.
Раневые меристемы обеспечивают восстановление поврежденной части тела. Они образуются из расположенных рядом с поврежденными участками специализированных живых клеток. Регенерация начинается с дедифференциации — обратного развития от специализированных клеток к меристематическим. Вступившие в этот процесс клетки вновь обретают способность делиться. Оказавшись на поверхности, они превращаются в феллоген, который, в свою очередь, образует пробку, покрывающую поверхность раны.
В другом случае дедифференцированные клетки, делясь, образуют рыхлую паренхиматозную ткань — каллус (лат. callus — толстая кожа, мозоль). При определенных условиях из него могут формироваться органы растения. Иногда из клеток каллуса могут развиваться зародыши, которые впоследствии развиваются в самостоятельный организм (это легко наблюдать на отрезанном листе бегонии, где зародыши будут развиваться из эпидермальных клеток в области перерезанных жилок).
Первичные меристемы обладают меристематической активностью, т. е. способны к делению изначально. В ряде случаев способность к активному делению может вновь возникнуть и у клеток, уже почти утративших это свойство. Такие «вновь» возникшие меристемы называют вторичными.
Первичными меристемами являются апикальная, прокамбий и интеркалярная, вторичными — камбий и феллоген, перицикл и раневая меристема.
biofile.ru
Х…я Стенка клетки
Стенка клетки.
Клетки растений, в отличие от клеток животных, имеют хорошо развитую, обычно твердую стенку. Совокупность клеточных стенок создает скелет растения, обладающий механической прочностью. Стенка играет важную роль в поглощении и передвижении веществ. Она часто сохраняется дольше, чем протопласт, поэтому и мертвые клетки продолжают выполнять разнообразные функции.
Первичная стенка образуется при делении клетки. К концу телофазы в экваториальной плоскости клетки возникает клеточная пластинка, впоследствии превращающаяся в срединную пластинку. Она состоит в основном из пектиновых веществ. На клеточную пластинку протопласт каждой из дочерних клеток откладывает первичную стенку. Толщина ее равна 0,1 — 0,5 мкм. В состав первичной стенки входят в основном пектиновые вещества, гемицеллюлозы и целлюлоза (всего 10—12 %), а также много воды.
Построение стенки и ее рост связаны с деятельностью аппарата Гольджи и плазмалеммы. Рост первичной стенки происходит путем внедрения молекул целлюлозы, синтезируемых на наружной поверхности плазмалеммы, и аморфных пектиновых веществ, доставляемых пузырьками Гольджи. При этом поверхность первичной стенки увеличивается, а толщина остается постоянной. Увеличение поверхности первичной стенки продолжается до достижения клеткой постоянного размера.
Вторичная стенка образуется путем наложения изнутри на первичную стенку новых слоев. При этом происходит рост стенки в толщину, а объем полости клетки уменьшается. Толщина вторичной стенки зависит от специализации клетки и колеблется от 1 до 10 мкм. Наиболее мощная она у клеток, выполняющих механическую функцию. У клеток, по которым передвигается вода, вторичная стенка откладывается в виде колец, спирали или имеет неравномерную толщину.
Вторичная стенка в основном состоит из целлюлозы (до 90%) и гемицеллюлоз. Пектиновых веществ и воды в ней значительно меньше, чем в первичной стенке.
Ткань — группа клеток, сходных по строению и выполняющих одинаковые функции.
|
Ткань |
Где располагается |
Особенности строения клеток |
Значение |
|
Образовательная |
Верхушки стеблей Кончики корней Камбий Рана |
Мелкие делящиеся клетки без вакуолей |
Рост растения |
|
Покровная |
Эпидермис (кожица) Пробка Кора |
Живые и мертвые клетки с толстыми и прочными оболочками, плотно прилегают друг к другу |
Защита от неблагоприятных воздействий |
|
Механическая |
Луб Древесина |
Толстые одревесневшие оболочки |
Опора органам растения |
|
Проводящая |
Сосуды древесины Ситовидные трубки луба |
Ситовидные трубки |
Распределение веществ, поступающих в корни и образующиеся в листьях по всему организму |
|
Основная |
Мякоть листа Сердцевина стебля Корень |
Хлоропласты в клетках |
Образование и накопление питательных веществ |
|
Выделительная |
Нектарники Железы |
|
Выделение эфирных масел, воды, нектара |
Переход растений от сравнительно однообразных условий жизни в водной среде к наземным сопровождался интенсивным процессом расчленения однородного вегетативного тела на органы — стебель, листья и корень. Эти органы состоят из разнообразных по структуре клеток, которые составляют легко различимые группы. Группы однородных по структуре клеток, выполняющие одинаковую функцию и имеющие общее происхождение, называют тканями. Часто несколько тканей, имеющих одинаковое происхождение, образуют комплекс, функционирующий как единое целое.
Выделяют шесть основных групп тканей: меристематические (образовательные), покровные, основные, механические, проводящие и выделительные.
Меристематические ткани.

Растения, в отличие от животных, растут и образуют новые органы на протяжении всей жизни. Это обусловлено наличием меристематических тканей, которые локализованы в определенных местах растения.
Меристема состоит из плотно сомкнутых живых клеток. Полость такой клетки заполнена цитоплазмой, в центре располагается крупное ядро, больших вакуолей нет, клеточная стенка очень тонкая, первичная. Клетки меристемы характеризуются двумя основными свойствами: интенсивным делением и дифференциацией, то есть превращением в клетки других тканей.
Первичная и вторичная меристема. Первичная меристема возникает в самом начале развития организма. Оплодотворенная яйцеклетка делится и образует зародыш, который состоит из первичной меристемы, вторичная возникает, как правило, позднее из первичной или из клеток уже дифференцированных тканей. Из первичной меристемы образуются первичные ткани, из вторичной — вторичные.
По месту расположения различают четыре группы меристем.
Гистология (учение о тканях).
Переход растений от сравнительно однообразных условий жизни в водной среде к наземным сопровождался интенсивным процессом расчленения однородного вегетативного тела на органы — стебель, листья и корень. Эти органы состоят из разнообразных по структуре клеток, которые составляют легко различимые группы. Группы однородных по структуре клеток, выполняющие одинаковую функцию и имеющие общее происхождение, называют тканями. Часто несколько тканей, имеющих одинаковое происхождение, образуют комплекс, функционирующий как единое целое.
Выделяют шесть основных групп тканей: меристематические (образовательные), покровные, основные, механические, проводящие и выделительные.
Покровные ткани.
Главное назначение покровных тканей — предохранение растения от высыхания и других неблагоприятных воздействий внешней среды. В зависимости от происхождения различают три группы покровных тканей: эпидерму, пробку, корку.

Гистология (учение о тканях).
Переход растений от сравнительно однообразных условий жизни в водной среде к наземным сопровождался интенсивным процессом расчленения однородного вегетативного тела на органы — стебель, листья и корень. Эти органы состоят из разнообразных по структуре клеток, которые составляют легко различимые группы. Группы однородных по структуре клеток, выполняющие одинаковую функцию и имеющие общее происхождение, называют тканями. Часто несколько тканей, имеющих одинаковое происхождение, образуют комплекс, функционирующий как единое целое.
Выделяют шесть основных групп тканей: меристематические (образовательные), покровные, основные, механические, проводящие и выделительные.
Основные ткани.
Под этим названием объединяют ткани, составляющие основную массу различных органов растения. Их называют также выполняющими, основной паренхимой или просто паренхимой. Основная ткань состоит из живых паренхимных клеток с тонкими стенками. Между клетками имеются межклетники. Паренхимные клетки выполняют разнообразные функции: фотосинтез, хранение запасных продуктов, поглощение веществ и др. Выделяют следующие основные ткани.
Механические ткани.
Механические ткани в совокупности составляют остов, поддерживающий все органы растения, противодействуя их излому или разрыву. Эти ткани состоят из клеток с толстыми стенками, часто (но не всегда) одревесневающими. Во многих случаях это мертвые клетки. В осевых органах это в основном прозенхимные клетки, в листьях и плодах — паренхимные.

Проводящие ткани.
Растение имеет два полюса питания: листья, осуществляющие воздушное питание, и корни, обеспечивающие почвенное питание. В соответствии с этим существуют два основных пути для транзита питательных веществ: путь, по которому вода и минеральные соли поднимаются от корня по стеблю к листьям, и путь, по которому органические вещества из листьев направляются во все остальные органы растений, где они потребляются или откладываются в запас.
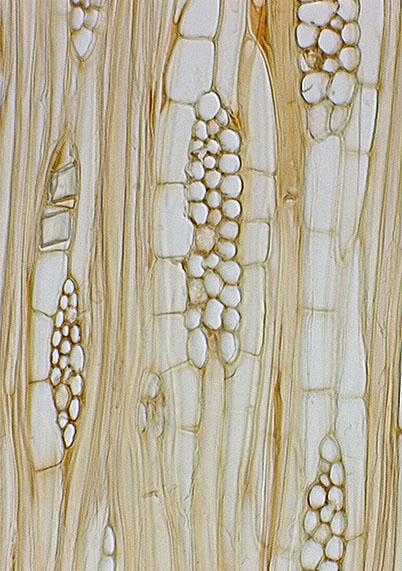
Сосуды (трахеи) и трахеиды — проводящие ткани, по которым осуществляется передвижение воды и минеральных солей. Сосуды (трахеи) — трубки, состоящие из члеников. Они дифференцируются из вертикального ряда клеток прокамбия или камбия, у которых утолщаются и одревесневают боковые стенки, отмирает содержимое, а в поперечных стенках образуются одна или несколько перфораций. Средняя длина сосудов 10 см.
Трахеиды, как и сосуды — мертвые образования, но в отличие от последних это не трубки, а прозенхимные клетки, в стенках которых имеются окаймленные поры. Длина трахеид в среднем 1 — 10 мм.
В зависимости от формы утолщений стенки сосуды и трахеиды бывают кольчатые, спиральные, сетчатые и др.. Кольчатые и спиральные сосуды имеют небольшой диаметр. Они свойственны молодым органам, .так как стенки их имеют неодревесневающие участки и способны растягиваться. Сетчатые и пористые сосуды гораздо большего диаметра, стенки их полностью одревесневают. Они обычно образуются позднее кольчатых и спиральных сосудов из камбия. Сосуды и трахеиды выполняют также и механическую функцию, придавая прочность растению. Они функционируют несколько лет, пока не происходит их закупорка окружающими живыми клетками паренхимы. Выросты последних, проникающие через поры в полость сосуда, называют тиллами.
Выделительные ткани.
Растения не имеют специализированных органов выделения, но все же экскреторные вещества у них так или иначе удаляются из организма или накапливаются в особых вместилищах. Поэтому выделительные ткани можно разделить на две группы: внутренней и внешней секреции. К первым относят млечники, одиночные выделительные клетки, схизогенные и лизигенные вместилища. Продукты внутренней секреции — дубильные вещества, смолы, эфирные масла и др. Ко вторым относят железистые волоски и желёзки, расположенные на поверхности органов. Продукты внешней секреции — эфирные масла, нектар, вода и др.
Млечники — это живые клетки с цитоплазмой, множеством ядер и вакуолью, заполненной млечным соком (латексом). Стенка их состоит из целлюлозы. Различают два вида млечников: членистые и нечленистые. Членистые млечники образуются так же, как и сосуды, в результате разрушения поперечных стенок у вертикального ряда клеток, нечленистые возникают в результате разрастания специальных клеток зародыша. Это гигантские цилиндрические или разветвленные клетки. Млечники располагаются только во флоэме, или пронизывают весь орган (стебель, корень, лист). Они выполняют не только выделительную, но также проводящую и запасающую функции. Проводящая функция осуществляется тогда, когда млечники соединяют места синтеза органических веществ, например листья, с местами их потребления. О выделительной и запасающей функциях млечников можно судить по составу латекса. У разных растений состав латекса очень разнообразен. Из экскреторных веществ он часто содержит каучук, танниды, алкалоиды, смолы и др., из запасных — крахмал, сахара, белки, масло и др. Иногда в латексе имеются ферменты. Цвет латекса белый или оранжево-красный. Млечники присущи лишь некоторым группам растений, например части сложноцветных, маковым, молочайным и др.
Выделительные клетки рассеяны среди клеток других тканей. По мере накопления экскрета протопласт их отмирает, а на стенку клетки изнутри откладывается суберин, изолирующий ядовитые вещества от окружающих клеток. Выделительные клетки имеются в листьях чая, лавра, в корневищах бадана и др.
Схизогенные и лизигенные вместилища служат для накопления и длительного хранения многих конечных продуктов жизнедеятельности, выключенных из обмена веществ. Они имеют округлую или каналовидную форму и различную величину.
Схизогенные вместилища формируются из межклетников, возникающих в результате раздвигания клеток. Прилегающие к вместилищу живые клетки становятся эпителиальными и выделяют в полость вместилища экскреторные вещества. Схизогенные вместилища бывают у весьма разнообразных групп растений, но особенно характерны для голосеменных, у которых в смоляных каналах накапливается бальзам — раствор смол в эфирных маслах.
Лизигенные вместилища образуются в результате растворения группы клеток, заполненных экскреторными веществами. Они характерны для цитрусовых.
Железистые волоски образуются из клеток эпидермы. В них накапливаются и выводятся из тела растения во внешнюю среду разнообразные экскреторные вещества в газообразном, жидком и твердом виде. По строению они очень разнообразны, но сохраняют постоянство структуры для каждой группы растений.
Нектарии, или нектарники — желёзки, выделяющие на поверхность органа раствор углеводов. Сахаристые выделения нектариев цветка привлекают насекомых-опылителей.
Гидатоды — это желёзки, выделяющие наружу через устьичные аппараты воду. Замыкающие клетки таких аппаратов лишены подвижности, устьичные щели постоянно открыты. Они сосредоточены по краю листа, преимущественно на верхушках зубчиков.
Пластиды являются основными цитоплазматическими органеллами клеток автотрофных растений. Название происходит от греческого слова «plastos», что в переводе означает «вылепленный».
Главная функция пластид – синтез органических веществ, благодаря наличию собственных ДНК и РНК и структур белкового синтеза. В пластидах также содержатся пигменты, обусловливающие их цвет. Все виды данных органелл имеют сложное внутреннее строение. Снаружи пластиду покрывают две элементарные мембраны, имеется система внутренних мембран, погруженных в строму или матрикс.
Классификация пластид по окраске и выполняемой функции подразумевает деление этих органоидов на три типа: хлоропласты, лейкопласты и хромопласты. Пластиды водорослей именуются хроматофорами.
Хлоропласты – это зеленые пластиды высших растений, содержащие хлорофилл – фотосинтезирующий пигмент. Представляют собой тельца округлой формы размерами от 4 до 10 мкм. Химический состав хлоропласта: примерно 50% белка, 35% жиров, 7% пигментов, малое количество ДНК и РНК. У представителей разных групп растений комплекс пигментов, определяющих окраску и принимающих участие в фотосинтезе, отличается. Это подтипы хлорофилла и каротиноиды (ксантофилл и каротин). При рассматривании под световым микроскопом видна зернистая структура пластид – это граны. Под электронным микроскопом наблюдаются небольшие прозрачные уплощенные мешочки (цистерны, или граны), образованные белково-липидной мембраной и располагающиеся в непосредственно в строме. Причем некоторые из них сгруппированы в пачки, похожие на столбики монет (тилакоиды гран), другие, более крупные находятся между тилакоидами. Благодаря такому строению, увеличивается активная синтезирующая поверхность липидно-белково-пигментного комплекса гран, в котором на свету происходит фотосинтез.

Ткани растений – это группы клеток, которые в определенном порядке располагаются в теле растения и предназначены для выполнения различных функций в жизнедеятельности растения. Все многоклеточные организмы имеют клетки различной структуры, совокупности которых являются тканями. Степень дифференцировки клеток тканей растений возрастает от низших растений к высшим. В отличие от тканей животных, у растений процесс образования тканей из первичных клеток можно наблюдать не на зародыше, а в растущих частях тела растения. Первичные клетки растения однородны, имеют примерно равные размеры и пропорции, состоят из протоплазмы и ядра. Из этих клеток формируется первоначальная меристема. Она в свою очередь позднее делится на составляющие: первый внешний слой (протодерм), из которого образуется кожица; срединный слой (прокамбий), являющийся предшественником сосудисто-волокнистых пучков; слой основной меристемы, который находится между протодермом и прокамбием и называется основной паренхимой или основной тканью растений, из нее появляется сердцевина, часть проводящей паренхимы. Это образовательная ткань растений.
О том, как проходит сосудисто-волокнистый пучок, можно судить по нервацифи листьев. Образуется характерная сеть, причем пучки листьев соединены с пучками стеблей, которые формируют разветвленную систему, переходящую в корень. Это проводящая ткань растения. Если изучать строение этой системы, можно увидеть, что образуется сплошной скелет во всем теле растения. Он состоит из правильно связанных друг с другом пучков, хотя они соединены по-разному у разных растений. Скелет растения из проводящих волокон, по которым перемещаются питательные вещества от листьев к корню и наоборот, представляет собой механическую ткань растения.
Внутренняя структура корня относительно проста по сравнению со структурой стебля . Это связано прежде всего с отсутствием листьев и соответственно узлов и междоузлий . Вследствие этого в расположении тканей на разных уровнях наблюдаются сравнительно небольшие различия.
Уже в самом начале зоны роста масса клеток дифференцируется на три зоны: эпиблему , первичную кору и осевой цилиндр , который может быть сплошным или полым.
Снаружи молодые корневые окончания покрыты эпиблемой . Эпиблема дифференцируется из самого наружного слоя верхушечной меристемы , называемого дерматогеном . Она достигает полного развития в зоне поглощения, где ее клетки образуют корневые волоски . В зоне проведения эпиблема довольно быстро слущивается. Количество корневых волосков иногда весьма значительно. В одном из исследований сообщалось, что у четырехмесячного растения риса примерно 14 млрд. корневых волосков с суммарной длиной более 10000 км и площадью поглощения 40 кв.м.
Первичная кора обычно дифференцируется из периферийного отдела верхушечной меристемы, лежащего глубжедерматогена , — периблемы . Осевой (центральный) цилиндр формируется из внутренней части меристемы — плеромы . Здесь, используя микроскоп с большим увеличением, уже удается заметить клетки прокамбия .
Первичная кора, на которую приходится основная масса ткани молодого корня, образована паренхимными клетками и обычно дифференцируется на уровне зоны растяжения. Она рыхлая и имеет систему межклетников, по которой вдоль оси корня циркулируют газы, необходимые для дыхания и поддержки обмена веществ. У болотных и водных растений межклетники особенно обширны и вся коровая часть оказывается занятой аэренхимой . Кроме того, кора является той частью корня, через которую активно проходит радиальный (ближний) транспорт воды и растворенных солей отэпиблемы к осевому цилиндру .
В тканях коры осуществляется активный синтез метаболитов и откладываются запасные питательные вещества. Наружные клетки первичной коры, лежащие непосредственно под эпиблемой , называются экзодермой . В зоне проведения после слущивания эпиблемы экзодерма оказывается снаружи, может видоизменяться (опробковевать) и выполнять функцию защитной покровной ткани. Основная масса первичной коры (мезодерма) образована паренхимными клетками . Самый внутренний слой коры — эндодерма . Она диффенцируется в корне на уровне начала зоны всасывания. В отличие от стебля, в корне клетки эндодермы заметно отличаются морфологически от прочих клеток. Это связано с особой функциейэндодермы , выполняющей роль барьера, который контролирует передвижение веществ из коры в осевой цилиндр и обратно. На ранних этапах развития эндодерма состоит из живых, тонкостенных клеток. Позднее ее клетки приобретают некоторые характерные особенности. В частности, на их радиальных стенках появляются особые утолщения — пояски Каспари , с помощью которых перекрывается передвижение растворов вдоль клеточных стенок ( рис. 66 ).
У многих двудольных и голосеменных растений образованием поясков Каспари обычно заканчивается дифференциацияэндодермы . У однодольных в клетках эндодермы могут происходить дальнейшие изменения. В частности, на внутренней поверхности первичных оболочек клеток откладывается суберин и далее толстая вторичная целлюлозная оболочка, которая со временем одревесневает. Наружные стенки почти не утолщаются. Среди толстостенных отмирающих клетокэндодермы встречаются живые клетки с тонкими неодревесневшими стенками, несущие только пояски Каспари . Это пропускные клетки; через них осуществляется физиологическая связь между первичной корой и осевым цилиндром ( рис. 67 ).
Осевой цилиндр (стела) начинает дифференцироваться в зоне роста, вплотную к зоне деления. Формирование осевого цилиндра начинается с образования наружного его слоя — перицикла . Перицикл представляет собой образовательную ткань , длительно сохраняющую меристематическую активность. Перицикл играет роль «корнеродного» слоя, так как в нем закладываются боковые корни, которые, таким образом, имеют эндогенное происхождение. В перицикле корня некоторых растений возникают также зачатки придаточных почек. У двудольных он участвует во вторичном утолщении корня, отчасти образуя камбий и феллоген . Под перициклом закладываются клетки боковой меристемы — прокамбия , дающие начало первичной флоэме , а несколько позднее — первичной ксилеме . Элементы флоэмы и ксилемы закладываются по кругу, чередуясь друг с другом, и развиваются центростремительно, т.е. по направлению к центру корня. Однако масса элементов ксилемы дифференцируется быстрее, обгоняет флоэму и занимает центр корня. В конечном итоге на поперечном сечении корня тяж ксилемы напоминает очертания звезды с различным числом лучей, между которыми располагаются участки флоэмы. Сформировавшаяся структура проводящей ткани получила название радиального проводящего пучка.
У большинства двудольных «звезда» ксилемы бывает ди-, три-, тетра- или пентархной, т.е. имеет соответственно 2, 3, 4 или 5 лучей. У однодольных она, как правило, многолучевая, или полиархная ( рис. 68 ).
Сердцевина нетипична для корня, но иногда заметна в центре в виде небольшого участка механической ткани или тонкостенных клеток, возникающих из прокамбия .
У большинства семенных растений боковые корни берут начало в перицикле . По мере увеличения размеров молодого бокового корня он проходит через первичную кору, возможно, секретируя при этом ферменты, разрушающие коровые клетки. Уже на ранних стадиях эндогенного развития корневой зачаток формирует корневой чехлик , апикальную и первичные меристемы. Центральные цилиндры главного и молодого боков корней первоначально не связаны между собой, но позднее соединяются за счет дифференциации в элементы ксилемы и флоэмы лежащих между нимипаренхимных клеток .
Описанное строение корня получило название первичного ( рис. 67 ). У однодольных и папоротников первичная структура корня сохраняется в течение всей жизни и вторичные ткани не возникают.
Корни некоторых строений имеют склонность к метаморфозу.
Видоизменения корней:
Корнеплод — утолщённый главный корень. В образовании корнеплода участвуют главный корень и нижняя часть стебля. Большинство корнеплодных растений двулетние. Корнеплоды состоят в основном из запасающей основной ткани (репа, морковь, петрушка).
Корневые клубни (корневые шишки) образуются в результате утолщения боковых и придаточных корней. С их помощью растение цветёт быстрее.
Корни-зацепки — своеобразные придаточные корни. При помощи этих корней растение «приклеивается» к любой опоре.
Ходульные корни — выполняют роль опоры.
Досковидные корни представляют собой боковые корни, проходящие у самой поверхности почвы или над ней, образующие треугольные вертикальные выросты, примыкающие к стволу. Характерны для крупных деревьев тропического дождевого леса.
Воздушные корни — боковые корни, растут в надземной части. Поглощают дождевую воду и кислород из воздуха. Образуются у многих тропических растений в условиях недостатка минеральных солей в почве тропического леса.
Микориза — сожительство корней высших растений с гифами грибов. При таком взаимовыгодном сожительстве, называемом симбиозом, растение получает от гриба воду с растворёнными в ней питательными веществами, а гриб — органические вещества. Микориза характерна для корней многих высших растений, особенно древесных. Грибные гифы, оплетающие толстые одревесневшие корни деревьев и кустарников, выполняют функции корневых волосков.
Бактериальные клубеньки на корнях высших растений — сожительство высших растений с азотфиксирующими бактериями — представляют собой видоизменённые боковые корни, приспособленные к симбиозу с бактериями. Бактерии проникают через корневые волоски внутрь молодых корней и вызывают у них образование клубеньков. При таком симбиотическом сожительстве бактерии переводят азот, содержащийся в воздухе, в минеральную форму, доступную для растений. А растения, в свою очередь, предоставляют бактериям особое местообитание, в котором отсутствует конкуренция с другими видами почвенных бактерий. Бактерии также используют вещества, находящиеся в корнях высшего растения. Чаще других бактериальные клубеньки образуются на корнях растений семейства Бобовые. В связи с этой особенностью семена бобовых богаты белком, а представителей семейства широко используют в севообороте для обогащения почвы азотом.
Дыхательные корни — у тропических растений — выполняют функцию дополнительного дыхания.
Части корня
Точка роста. Участок непосредственно за кончиком корня, где клетки делятся, обеспечивая новый рост.
Зона растяжения. Участок новых клеток, образованных в точке роста и расположенных непосредственно за ней. Клетки, пока их клеточные стенки не станут жёсткими, растягиваются в длину при всасывании воды. Это растяжение толкает кончик корня дальше в почву.
Слой корневых волосков. Наиболее молодая часть эпидермиса или внешней кожицы корня. Это зона, где образуютсякорневые волоски. Она расположена непосредственно за зоной растяжения. Как только стенки вытянувшихся клеток становятся жёстче, самые крайние из них превращаются в слой корневых волосков. Более старая часть этого слоя (выше по корню) медленно отделяется, замещаясь слоем жёстких клеток, входящих в экзодерму (самый наружный слой коры).
Корневые волоски. Длинные выросты клеток слоя корневых волосков. Они всасывают воду и минеральные вещества.
Корневой чехлик. Слой клеток, защищающих кончик корня при росте.
studfiles.net
Ответы@Mail.Ru: биология. клетка. ткани растений.
Клетка — это простейшая и обязательная единица живого, это его элемент, основа строения, развития и всей жизнедеятельности организма. Ткани все растений состоят только из клеток, в них нет межклеточного вещества. Ткани высших растений это: Основная ткань. Паренхима выполняет соединительную, запасающую, механическую, проводящую, защитную, секретирующую, фотосинтезирующую функции и функцию деления. У этой ткани толстая первичная клеточная стенка, а вторичная клеточная стенка без лигнина. Паренхима иногда может превращаться в меристему! Хлоренхима — фотосинтезирующая паренхима. Механическая. Механические ткани делятся на колленхиму и склеренхиму. Они придают растению прочность. Колленхима всегда находится в состоянии тургора, благодаря которому растения могут передвигать свои части. Она твердая, но упругая. Склеренхима — это мертвая ткань растений, у нее толстые вторичные клеточные стенки. Эта ткань делится по структуре на волокна, обеспечивающие прочность, и склереиды — защита от удара. Склереиды и колленхима — мертвые ткани. Колленхима очень часто находится под фотосинтезирующей паренхимой. Образовательная. (Меристема) Клетки меристемы постоянно делятся. У нее нет цветных пластидов, следовательно она прозрачная. Меристема бывает первичная (апикальная, прокамбия, интеркалярная) и вторичная (камбий, перецикл, феллоген, раневая меристема) . Проводящая. Проводящие ткани обеспечивают быстрый транспорт воды по растению. Проводящие ткани делятся на ксилему и флоэму . Ксилема состоит из: 1.Лучевая паренхима (живые клетки) . 2.Трахеиды. (мертвые клетки) 3.Членики сосудов. (мертвые клетки) 4.Волокна либриформа. 5.Запасающая паренхима. Она выполняет проводящую и механическую функции. Флоэма состоит из: 1.Лучевая паренхима флоэмы. 2.Ситовидные клетки. 3.Ситовидные труки. 4.Клетки спутники. Клетки флоэмы живые — в них сохраняется цитоплазма, но утрачивается ядро. У клеток зтой ткани есть клетки спутники, которые помогают «выжить» проводящим клеткам флоэмы. Они «качают» некоторые химические вещества через себя. Флоэму часто называют лубом. Покровная Покровная ткань находится на границе организма и внешней среды и выполняет защитную функцию и функцию обеспечения транспорта. У покровной ткани утолщенная клеточная стенка и на ее поверхности есть слой воска или кутина (кутикула) . Клетки, которые выполняют всасывание не умеют кутикулу и имеют максимально тонкую стенку. Устьица — образования, служащие для газо-обмена. Покровные ткани делятся на первичные (эпидерма, ризодерма) и вторичные (перидерма) . Перидерма состоит из феллемы (пробка) , феллогена и феллогена. Эпидерма покрывает органы растений, а ризодерма покрывает корень. Феллодерма выполняет запасающую функцию, она похожа на паренхиму. На поверхности покровных тканей есть чечевички — места, где перидерма лежит рыхло, они служат для газообмена.
touch.otvet.mail.ru
Меристема
(греч. «меристос» — делимый) — образовательная ткань, которая появляется по мере деления зиготы. Она формирует тело зародыша, а по мере роста растения перемещается во все его точки роста — верхушки корней, стеблей, в основания междоузлий и листьев. Такие меристемы называются первичными. По положению на растении они подразделяются на верхушечные (апикальные), боковые (латеральные) и вставочные (интеркалярные). Клетки меристем некрупные, с плотной цитоплазмой, без вакуолей, с относительно большим ядром, занимающим центральное положение. Основным свойством меристем является способность, делиться путем митоза и дифференцироваться, т. е. преобразовываться в любые другие ткани. Меристемы могут возникать и из уже имеющихся тканей, например из основных тканей или из первичных меристем. Такие меристемы называют вторичными; к ним относятся камбий, феллоген (пробковый камбий), раневые меристемы. Наиболее существенным различием между меристемами является направление деления клеток по отношению к поверхности органа: у первичных меристем клетки делятся в поперечном, радиальном и тангенциальном (параллельном поверхности) направлениях, а у вторичных — только в тангенциальном. Это отражается на порядке расположения образованных ими клеток — клетки, образованные первичной меристемой, лежат беспорядочно, а образованные вторичной лежат четкими рядами, одна клетка над другой, что особенно четко видно на поперечных срезах корня и стебля. Вторичные меристемы свойственны только двудольным покрытосеменным и голосеменным растениям, первичные же — универсальны. Органы, где функционируют вторичные меристемы, приобретают вторичное строение, а те органы, где функционируют первичные меристемы, имеют только первичное строение. Лист, цветок имеют только первичное строение, даже если они находятся на двудольном растении.
Источник: Т. Л. Богданова «Пособие для поступающих в вузы»
xn--90aeobapscbe.xn--p1ai
2.5. Клеточная стенка
Клеточная стенка (клеточная оболочка) – характерный признак растительной клетки, отличающий ее от клетки животной. Клеточная стенка придает клетке определенную форму. Культивируемые на специальных питательных средах клетки растений, у которых ферментативным путем удаляется стенка, всегда принимают сферическую форму. Клеточная стенка придает клетке прочность и защищает протопласт, она уравновешивает тургорное давление и препятствует, таким образом, разрыву плазмалеммы. Совокупность клеточных стенок образует внутренний скелет, поддерживающий тело растения и придающий ему механическую прочность.
Клеточная стенка бесцветна и прозрачна, легко пропускает солнечный свет. Обычно стенки пропитаны водой. По системе клеточных стенок осуществляется транспорт воды и растворенных в ней низкомолекулярных соединений (транспорт по апопласту).
Клеточная стенка состоит в основном из полисахаридов, которые можно подразделить на скелетные вещества и вещества матрикса.
Скелетным веществом клеточной стенки растений является целлюлоза (клетчатка), представляющая собой бета-1,4-D-глюкан. Это самое распространенное органическое вещество биосферы. Молекулы целлюлозы представляют собой очень длинные неразветвленные цепи, они располагаются параллельно друг другу группами по нескольку десятков и скреплены многочисленными водородными связями. В результате образуются микрофибриллы, которые создают структурный каркас стенки и обусловливают ее прочность. Микрофибриллы целлюлозы видны только в электронный микроскоп, их диаметр равен 10-30 нм, длина достигает нескольких мкм.
Целлюлоза нерастворима и не набухает в воде. Она очень инертна в химическом отношении, не растворяется в органических растворителях, концентрированных щелочах и разведенных кислотах. Микрофибриллы целлюлозы эластичны и очень прочны на разрыв (сходны со сталью). Эти свойства определяют широкое применение целлюлозы и ее продуктов. Мировая продукция хлопкового волокна, состоящего почти целиком из целлюлозы, составляет 1,5•107 тонн в год. Из целлюлозы получают бездымный порох, ацетатный шелк и вискозу, целлофан, бумагу. Качественную реакцию на целлюлозу проводят с реактивом хлор-цинк-йод, целлюлозная клеточная стенка окрашивается в сине-фиолетовый цвет.
У грибов скелетным веществом клеточной стенки является хитин – полисахарид, построенный из остатков глюкозамина. Хитин еще более прочен, чем целлюлоза.
Микрофибриллы погружены в аморфный матрикс, обычно представляющий собой насыщенный водой пластичный гель. Матрикс является сложной смесью полисахаридов, молекулы которых состоят из остатков нескольких различных сахаров и представляют собой более короткие, чем у целлюлозы, и разветвленные цепи. Матричные полисахариды определяют такие свойства клеточной стенки, как сильная набухаемость, высокая проницаемость для воды и растворенных в ней низкомолекулярных соединений, катионообменные свойства. Полисахариды матрикса делят на две группы – пектиновые вещества и гемицеллюлозы.
Пектиновые вещества сильно набухают или растворяются в воде. Они легко разрушаются под действием щелочей и кислот. Простейшими представителями пектиновых веществ являются растворимые в воде пектовые кислоты – продукты полимеризации альфа-D-галактуроновой кислоты (до 100 единиц), связанных 1,4-связями в линейные цепи (альфа-1,4-D-галактуронан). Пектиновые кислоты (пектины) – это более высокомолекулярные (100-200 единиц) полимерные соединения альфа-D-галактуроновой кислоты, в которых карбоксильные группы частично метилированы. Пектаты и пектинаты – кальциевые и магниевые соли пектовых и пектиновых кислот. Пектиновые кислоты, пектаты и пектинаты растворимы в воде в присутствии сахаров и органических кислот с образованием плотных гелей.
В клеточных стенках растений в основном присутствуют протопектины – высокомолекулярные полимеры метоксилированной полигалактуроновой кислоты с арабинанами и галактанами, у двудольных растений в состав цепей галактуронана входит небольшое количество рамнозы. Протопектины нерастворимы в воде.
Гемицеллюлозы представляют собой разветвленные цепи, построенные из остатков нейтральных сахаров, чаще встречаются глюкоза, галактоза, манноза, ксилоза; степень полимеризации 50-300. Гемицеллюлозы химически более устойчивы, чем пектиновые вещества, они труднее гидролизуются и слабее набухают в воде. Гемицеллюлозы могут откладываться в стенках клеток семян в качестве запасных веществ (финиковая пальма, хурма). Пектиновые вещества и гемицеллюлозы связаны взаимными переходами. Помимо полисахаридов, в матриксе клеточных стенок присутствует особый структурный белок. Он связан с остатками сахара арабинозы и поэтому является гликопротеидом.
Матричные полисахариды не просто заполняют промежутки между целлюлозными микрофибриллами. Их цепи располагаются упорядоченно и образуют многочисленные связи как друг с другом, так и с микрофибриллами, что значительно повышает прочность клеточной стенки.
Клеточные стенки растений часто подвергаются химическим видоизменениям. Одревеснение, или лигнификация происходит в том случае, если в матриксе откладывается лигнин – полимерное соединение фенольной природы, нерастворимое в воде. Одревесневшая клеточная стенка теряет эластичность, резко повышается ее твердость и прочность на сжатие, снижается проницаемость для воды. Реактивами на лигнин являются: 1) флороглюцин и концентрированная хлористоводородная или серная кислота (одревесневшие стенки приобретают вишнево-красную окраску) и 2) сульфат анилина, под действием которого одревесневшие стенки становятся лимонно-желтыми. Лигнификация характерна для стенок клеток проводящей ткани ксилемы (древесины) и механической ткани склеренхимы.
Опробковение, или суберинизация происходит в результате отложения с внутренней стороны клеточной стенки гидрофобных полимеров – суберина и воска. Суберин представляет собой смесь эфиров полимерных жирных кислот. Мономерами воска являются жирные спирты и восковые эфиры. Воск легко извлекается органическими растворителями и быстро плавится, образует кристаллы. Суберин – аморфное соединение, не плавится и не растворяется в органических растворителях. Суберин и воск, образуя чередующиеся параллельные слои, выстилают всю полость клетки с внутренней стороны в виде пленки. Субериновая пленка практически непроницаема для воды и для газов, поэтому после ее образования клетка обычно отмирает. Опробковение характерно для стенок клеток покровной ткани пробки. Реактивом на опробковевшую клеточную стенку является судан III, окраска оранжево-красная.
Кутинизации подвергаются наружные стенки клеток покровной ткани эпидермы. Кутин и воск откладываются чередующимися слоями на наружной поверхности клеточной стенки в виде пленки – кутикулы. Кутин представляет собой жироподобное полимерное соединение, близкое по химической природе и свойствам суберину. Кутикула предохраняет растение от излишнего испарения воды с поверхности растения. Окрасить ее можно реактивом судан III в оранжево-красный цвет.
Минерализация клеточной стенки происходит вследствие отложения в матриксе большого количества минеральных веществ, чаще всего кремнезема (оксида кремния), реже оксалата и карбоната кальция. Минеральные вещества придают стенке твердость и хрупкость. Отложение кремнезема характерно для клеток эпидермы хвощей, осок и злаков. Приобретенная в результате окремнения жесткость стеблей и листьев служит защитным средством против улиток, а также значительно снижает поедаемость и кормовую ценность растений.
У некоторых специализированных клеток наблюдается ослизнение клеточной стенки. При этом вместо целлюлозной вторичной стенки происходит отложение аморфных, сильно гидратированных кислых полисахаридов в виде слизей и камедей, близких по химической природе к пектиновым веществам. Слизи хорошо растворяются в воде с образованием слизистых растворов. Камеди клейкие, вытягиваются в нити. В сухом виде они имеют роговую консистенцию. При отложении слизи протопласт постепенно оттесняется к центру клетки, его объем и объем вакуоли постепенно уменьшаются. В конце концов, полость клетки может целиком заполниться слизью, и клетка отмирает. В некоторых случаях слизь может проходить через первичную клеточную стенку на поверхность. В синтезе и секреции слизи основное участие принимает аппарат Гольджи.
Выделяемая растительными клетками слизь выполняет различные функции. Так, слизь корневого чехлика служит в качестве смазки, облегчающей рост кончика корня в почве. Слизевые железки насекомоядных растений (росянка) выделяют ловчую слизь, к которой приклеиваются насекомые. Слизь, выделяемая наружными клетками семенной кожуры (лен, айва, подорожники), закрепляет семя на поверхности почвы и защищает проросток от высыхания. Слизь окрашивается реактивом метиленовый синий в голубой цвет.
Выделение камедей обычно происходит при поранении растений. Например, камедетечение из пораненных участков стволов и ветвей часто наблюдается у вишни и сливы. Вишневый клей представляет собой застывшую камедь. Камедь выполняет защитную функцию, закрывая рану с поверхности. Образуются камеди в основном у древесных растений из семейств бобовых (акации, трагакантовые астрагалы) и розоцветных подсемейства сливовых (вишня, слива, абрикос). Камеди и слизи используются в медицине.
Клеточная стенка является продуктом жизнедеятельности протопласта. Полисахариды матрикса, гликопротеид стенки, лигнин и слизи образуются в аппарате Гольджи. Синтез целлюлозы, образование и ориентация микрофибрилл осуществляются плазмалеммой. Большая роль в ориентации микрофибрилл принадлежит микротрубочкам, которые располагаются параллельно откладывающимся микрофибриллам вблизи плазмалеммы. Если микротрубочки разрушить, образуются только изодиаметрические клетки.
Образование клеточной стенки начинается во время деления клетки. В плоскости деления образуется клеточная пластинка, единый слой, общий для двух дочерних клеток. Она состоит из пектиновых веществ, имеющих полужидкую консистенцию; целлюлоза отсутствует. Во взрослой клетке клеточная пластинка сохраняется, но претерпевает изменения, поэтому ее называют срединной, или межклеточной пластинкой (межклеточным веществом) (рис. 2.16 ). Срединная пластинка обычно очень тонка и почти неразличима.
Сразу после образования клеточной пластинки протопласты дочерних клеток начинают откладывать собственную клеточную стенку. Она откладывается изнутри как на поверхности клеточной пластинки, так и на поверхности других клеточных стенок, принадлежавших ранее материнской клетке. После деления клетка вступает в фазу роста растяжением, который обусловлен интенсивным осмотическим поглощением клеткой воды, связанным с образованием и ростом центральной вакуоли. Тургорное давление начинает растягивать стенку, но она не рвется благодаря тому, что в нее постоянно откладываются новые порции микрофибрилл и веществ матрикса. Отложение новых порций материала происходит равномерно по всей поверхности протопласта, поэтому толщина клеточной стенки не уменьшается.
Стенки делящихся и растущих клеток называют первичными. Они содержат много (60-90%) воды. В сухом веществе преобладают матричные полисахариды (60-70%), содержание целлюлозы не превышает 30%, лигнин отсутствует. Толщина первичной стенки очень невелика (0,1-0,5 мкм).
Для многих клеток отложение клеточной стенки прекращается одновременно с прекращением роста клетки. Такие клетки окружены тонкой первичной стенкой до конца жизни ( рис. 2.16).
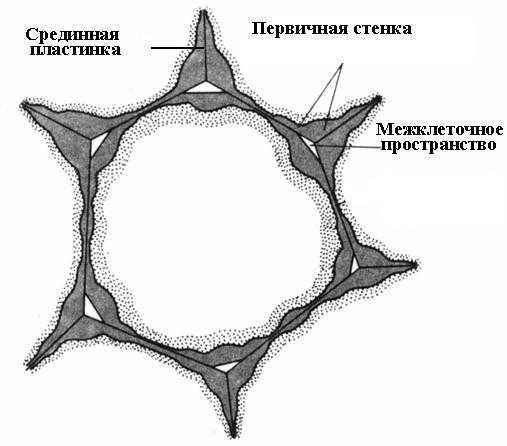
Рис. 2.16. Паренхимная клетка с первичной стенкой.
У других клеток отложение стенки продолжается и по достижении клеткой окончательного размера. При этом толщина стенки увеличивается, а объем, занимаемый полостью клетки, сокращается. Такой процесс носит название вторичного утолщения стенки, а саму стенку называют вторичной (рис. 2.17 ).
Вторичная стенка может рассматриваться как дополнительная, выполняющая главным образом механическую, опорную функцию. Именно вторичная стенка ответственна за свойства древесины, текстильного волокна, бумаги. Вторичная стенка содержит значительно меньше воды, чем первичная; в ней преобладают микрофибриллы целлюлозы (40-50% от массы сухого вещества), которые располагаются параллельно друг другу. Из полисахаридов матрикса характерны гемицеллюлозы (20-30%), пектиновых веществ очень мало. Вторичные клеточные стенки, как правило, подвергаются одревеснению. В неодревесневших вторичных стенках (лубяные волокна льна, волоски хлопчатника) содержание целлюлозы может достигать 95%. Большое содержание и строго упорядоченная ориентация микрофибрилл определяют высокие механические свойства вторичных стенок. Часто клетки, имеющие вторичную одревесневшую клеточную стенку, после завершения вторичного утолщения отмирают.
Срединная пластинка склеивает соседние клетки. Если ее растворить, стенки клеток теряют связь друг с другом и разъединяются. Этот процесс называется мацерация. Довольно обычна естественная мацерация, при которой пектиновые вещества срединной пластинки переводятся в растворимое состояние с помощью фермента пектиназы и затем вымываются водой (перезрелые плоды груши, дыни, персика, банана). Часто наблюдается частичная мацерация, при которой срединная пластинка растворяется не по всей поверхности, а лишь в углах клеток. Вследствие тургорного давления соседние клетки в этих местах округляются, в результате чего образуются межклетники (рис. 2.16 ). Межклетники образуют единую разветвленную сеть, которая заполняется парами воды и газами. Таким образом, межклетники улучшают газообмен клеток.
Характерная особенность вторичной стенки – ее неравномерное отложение поверх первичной стенки, в результате чего во вторичной стенке остаются неутолщенные участки – поры. Если вторичная стенка не достигает большой толщины, поры выглядят как мелкие углубления. У клеток с мощной вторичной стенкой поры в разрезе имеют вид радиальных каналов, идущих от полости клетки до первичной стенки. По форме порового канала различают поры двух типов – простые и окаймленные (рис. 2.17 ).
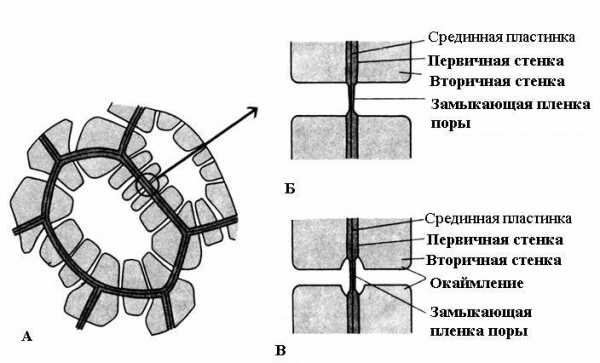
Рис. 2.17. Типы пор : А – клетки с вторичными стенками и многочисленными простыми порами; Б – пара простых пор; В – пара окаймленных пор.
У простых пор диаметр порового канала по всей длине одинаковый и имеет форму узкого цилиндра. Простые поры характерны для паренхимных клеток, лубяных и древесинных волокон.
Поры в двух смежных клетках, как правило, возникают друг против друга. Эти общие поры имеют вид одного канала, разделенного тонкой перегородкой из срединной пластинки и первичной стенки. Такая совокупность двух пор смежных стенок соседних клеток носит название пары пор и функционирует как одно целое. Разделяющий их канал участок стенки называется замыкающей пленкой поры, или поровой мембраной. В живых клетках замыкающая пленка поры пронизана многочисленными плазмодесмами (рис. 2.18 ).
Плазмодесмы присущи только растительным клеткам. Они представляют собой тяжи цитоплазмы, пересекающие стенку смежных клеток. Число плазмодесм в одной клетке очень велико – от нескольких сотен до десятков тысяч, обычно плазмодесмы собраны в группы. Диаметр плазмодесменного канала составляет 30-60 нм. Его стенки выстланы плазмалеммой, непрерывной с плазмалеммой смежных клеток. В центре плазмодесмы проходит мембранный цилиндр – центральный стержень плазмодесмы, непрерывный с мембранами элементов эндоплазматической сети обеих клеток. Между центральным стержнем и плазмалеммой в канале находится гиалоплазма, непрерывная с гиалоплазмой смежных клеток.
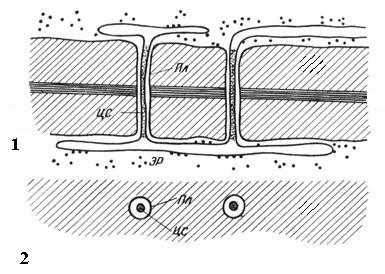
Рис. 2.18. Плазмодесмы под электронным микроскопом (схема ): 1 – на продольном срезе; 2 – на поперечном срезе; Пл – плазмалемма; ЦС – центральный стержень плазмодесмы; ЭР – элемент эндоплазматического ретикулума.
Таким образом, протопласты клеток не полностью изолированы друг от друга, а сообщаются по каналам плазмодесм. По ним происходит межклеточный транспорт ионов и мелких молекул, а также передаются гормональные стимулы. Посредством плазмодесм протопласты клеток в растительном организме образуют единое целое, называемое симпластом, а транспорт веществ через плазмодесмы получил название симпластического в отличие от апопластического транспорта по клеточным стенкам и межклетникам.
У окаймленных пор (рис. 2.17 ) канал резко суживается в процессе отложения клеточной стенки, поэтому внутреннее отверстие поры, выходящее в полость клетки, гораздо уже, чем наружное, упирающееся в первичную стенку. Окаймленные поры характерны для рано отмирающих клеток водопроводящих элементов древесины. У них поровый канал по направлению к замыкающей пленке воронковидно расширяется, а вторичная стенка нависает в виде валика над расширенной частью канала, образуя камеру поры. Название окаймленной поры происходит оттого, что при рассмотрении с поверхности внутреннее отверстие имеет вид маленького круга или узкой щели, тогда как наружное отверстие как бы окаймляет внутреннее в виде круга большего диаметра или более широкой щели.
Поры облегчают транспорт воды и растворенных веществ от клетки к клетке, в то же время не снижая прочности клеточной стенки.
studfiles.net
Ткани
Категория: Анатомия растений
Ткани
Тело высших растений состоит из разнородных клеток, специализированных на выполнении различных функций. Физиологические отправления клеток определяют их структуру. Комплексы клеток, сходных по функциям, а большей частью и по строению, имеющих одинаковое происхождение и определенную локализацию в теле растения, называют тканями.
Распределение тканей в органах растений и их структура тесно связаны с выполнением ими определенных физиологических функций.
Ткани, состоящие из живых тонкостенных, интенсивно делящихся клеток, называются образовательными или меристемами. Верхушечные (апикальные) меристемы расположены на верхушках стеблей и в окончаниях корней. Они обусловливают рост этих органов в длину.
Рассмотрение апикальных меристем побега не входит в задачи настоящего курса.
К боковым меристемам относятся камбий и фелло-ген. Камбий обеспечивает утолщение стебля и корня. Феллоген образует пробку.
Ткани, возникающие в результате роста и дифференциации клеток — производных меристем, называют п о-стоянными. Распространенная в настоящее время классификация постоянных тканей основана на их ана-томо-физиологических особенностях.
Покровные ткани защищают внутренние ткани растений от прямого влияния внешней среды, регулируют испарение и газообмен. К ним относятся эпидермис и пробка, состоящие из плотно соединенных клеток. В корнях однодольных и молодых корнях некоторых двудольных растений функцию покровной ткани выполняет экзодерма.
М еханические ткани обусловливают прочность растения. Стенки клеток, слагающих эти ткани, утолщены. К механическим тканям относятся колленхим а, состоящая из паренхимных или несколько удлиненных клеток с неравномерно утолщенными целлюлозными стенками, и склеренхима, клетки которой имеют равномерно утолщенные одревесневшие стенки. Склеренхима может быть представлена волокнами и склереидами изодиаметрической (каменистые клетки), ветвистой и звездчатой форм.
Проводящие ткани обеспечивают проведение воды, почвенных растворов и продуктов ассимиляции, вырабатываемых листьями. Ткань, проводящую воду, называют ксилемой или древесиной (особенно у древесных растений), а ткань, проводящую органические вещества, образованные растением в процессе фотосинтеза,— флоэмой или лубом. В состав ксилемы и флоэмы наряду с собственно проводящими элементами большей частью входят механические и запасающие клетки, поэтому эти ткани называют сложными. Проводящие ткани по происхождению могут быть первичными и вторичными. Первичные возникают из прокамбия — меристематической ткани, закладывающейся на ранних этапах онтогенеза растения. Вторичные ткани образуют камбий, который дифференцируется из прокамбия.
Обычно флоэма и ксилема располагаются рядом, составляя проводящий пучок. В зависимости от взаимного расположения ксилемы и флоэмы различают несколько типов пучков.
Коллатеральные (бокобочные), состоящие из одного тяжа флоэмы, к которому плотно примыкает тяж ксилемы. В стебле флоэма обращена к периферии, в листьях — к нижней стороне пластинки.
Биколлатеральные (двубокобочные), в которых ксилема находится между двумя тяжами элементов флоэмы, наружным и внутренним.
Концентрические, у которых либо ксилема со всех сторон окружает флоэму, либо флоэма окружает ксилему (амфикрибральный пучок).
Сложные радиальные, в которых тяжи флоэмы чередуются с радиальными тяжами ксилемы, образующими на поперечном срезе более или менее звездчатую фигуру. Эти пучки характерны для корней. Пучки, состоящие из какой-либо одной ткани — флоэмы или ксилемы, называют простыми или неполными. Ими заканчиваются, например, мелкие ответвления жилок в листовых пластинках.
Если флоэма и ксилема разделены камбием, в результате деятельности которого возникают вторичные проводящие ткани, пучок называют открытым, т. е. способным к дальнейшему образованию клеток. Открытые пучки свойственны стеблям и корням двудольных и голосеменных растений, они могут быть коллатеральными и биколлатеральными. Закрытые проводящие пучки, не имеющие камбия, характерны для стеблей и корней папоротникообразных и однодольных растений и для большинства листьев. Закрытые пучки представлены концентрическими, коллатеральными и радиальными пучками. Проводящие пучки нередко армированы склеренхимой, которая окружает пучок со всех сторон либо образует тяжи со стороны флоэмы или древесины. Такие пучки нередко называют сосудисто-волокнистыми.
Ткани, состоящие из однородных паренхимных клеток, которые заполняют пространства между другими тканями, называют основными. Живые клетки основной паренхимы находятся в состоянии тургора и в контакте с другими тканями увеличивают механическую прочность растения. Основная паренхима может специализироваться на выполнении различных функций. К системе основных тканей относятся запасающие, в которых откладывается запас питательных веществ; ассимиляционные, в которых происходит фотосинтез; выделительные, содержащие продукты отброса; ткани поглощения воды и почвенных растворов. К последним относится эпиблема или ризодер-м а. Она расположена на самой периферии молодого корня и наряду со своей основной функцией играет роль покровной ткани.
В растениях выделяют систему проветривания, которая представлена устьицами, расположенными в эпидермисе, чечевичками, находящимися в перидерме, воздухоносными полостями и межклетниками.
Изучение строения тканей целесообразно начать с рассмотрения покровных, механических и проводящих.
Разнообразие тканей в стебле тыквы обыкновенной (Cucurbita реро L.)
Кусочки стеблей длиной 2—3 см, вырезанные из междоузлий с небольшой внутренней воздушной полостью, фиксируют спиртом в конце вегетационного периода (в августе — сентябре).

Рис. 1. Схема поперечного среза стебля тыквы: эп — эпидермис, кол — колленхима, п — паренхима, скл — склеренхима, фл — флоэма, кмб — камбий, кс — ксилема, в. п. — воздушная полость
Стебель тыквы в очертании округлый или округло-пятиугольной, с пятилучевой воздушной полостью (рис. 1). Между лучами полости расположены пять крупных проводящих пучков, которые хорошо видны невооруженным глазом. Против лучей полости, немного ближе к периферии стебля, находится второе кольцо из пяти таких же, но более мелких пучков.
Внутреннее строение стебля изучают на продольном и поперечном срезах. Поперечный срез должен захватИть не менее половины сечения стебля. Если такой срез не получается достаточно тонким, то для работы с большим увеличением микроскопа следует приготовить еще один как можно более тонкий срез, на котором должны быть наружная часть стебля и хотя бы один крупный проводящий пучок. Продольный радиальный срез, проходящий посередине крупного пучка, лучше делать с междоузлий толстых стеблей. Перед изготовлением среза стебель разрезают вдоль по диаметру. Ткани, расположенные снаружи от пучка, можно удалить. Чтобы срезы не были очень длинными, поверхность, с которой их делают, подсекают бритвой на расстоянии 0,5 см от верхнего края кусочка. Наиболее удачные срезы кладут в раствор иода в водном растворе иоди-стого калия и накрывают покровным стеклом. Некоторые срезы можно последовательно обрабатывать фло-роглюцином и соляной кислотой и рассматривать их в глицерине.
Общий план расположения тканей изучают на поперечном срезе стебля при малом увеличении микроскопа, строение отдельных тканей рассматривают при большом увеличении на поперечном и продольном срезах.
Стебель покрыт эпидермисом, на поверхности которого хорошо видна тонкая светлая пленка — кутикула. Эпидермис состоит из одного слоя плотно сомкнутых живых клеток с утолщенной наружной стенкой. Некоторые клетки образуют многоклеточные волоски.
Под эпидермисом расположены участки механической ткани — колленхимы. Наиболее крупные из них находятся в ребрах стебля. Колленхима состоит из живых многоугольных клеток с неравномерно утолщенными стенками. Стенки утолщены в углах, т. е. в местах соединения нескольких клеток. Такая колленхима называется уголковой (рис. 2, А). На поперечном срезе утолщения имеют вид треугольников, если соединяются три клетки, или ромбов, если соединяются четыре клетки. Утолщенные оболочки вследствие сильного преломления света выглядят блестящими, они хорошо заметны вокруг округлых, более гемных, заполненных содержимым полостей клеток. На продольном срезе клетки колленхимы удлинены.
По времени возникновения колленхима —самая ранняя механическая ткань, оболочки ее клеток эластичны, поэтому колленхима может растягиваться вместе с ростом стебля. Механическую роль эта ткань выполняет только в состоянии тургора, так как неутолщенные места оболочек при подвядании спадаются.
Между участками колленхимы, непосредственно под ней и между другими специализированными тканями и проводящими пучками расположены тонкостенные живые, почти округлые клетки с небольшими межклетниками. Это паренхима. Размеры клеток паренхимы в разных частях стебля неодинаковы.
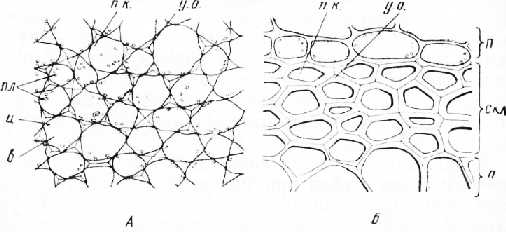
Рис. 2. Механические ткани стебля тыквы. А — уголковая колленхима; Б — склеренхима: л — паренхимные клетки, у. о. — утолщения оболочки, пл — пластиды, Ч — цитоплазма, в — вакуоль, скл — склеренхима, п. к. — полости клеток
В некоторых паренхимных клетках откладывается запасной крахмал. В периферических слоях стебля клетки основной паренхимы содержат хлороплаеты и поэтому выполняют ассимиляционную функцию.
Паренхима, подстилающая участки колленхимы, граничит с несколькими рядами плотно сомкнутых многоугольных клеток с равномерно утолщенными одревесневшими стенками. Это склеренхима (рис. 2, Б). Оболочки ее клеток после взаимодействия с иодом приобретают ярко-оранжевый цвет, а после флороглюциновой реакции они становятся малиново-красными. На поперечном срезе склеренхима располагается кольцом. В старых стеблях клетки склеренхимы мертвые, в более молодых стеблях в ее клетках видны остатки содержимого. Склеренхима представлена сильно вытянутыми, заостренными на концах волокнами, которые хорошо видны на продольном срезе.
Склеренхима выполняет механическую функцию. Она образуется позднее колленхимы. Одревеснение стенок склеренхимных волокон начинается по окончании роста стебля в длину. Так как одревесневшие стенки теряют эластичность, при утолщении стебля кольцо склеренхимы нередко разрывается. Разрывы заполняются парен-химной тканью.
Расположение механических тканей — колленхимы и склеренхимы — на периферии стебля повышает сопротивление стебля изгибу, сжатию и растяжению, которым подвергаются главным образом наружные зоны органа.
Кроме наиболее распространенной уголковой колленхимы у растений нередко встречается колленхима иного строения. Так, в стеблях с сильным вторичным утолщением развивается колленхима, у которой утолщены тангентальные стенки клеток, т. е. стенки, параллельные поверхности органа. Радиальные стенки остаются тонкими. Такую колленхиму называют пластинчатой (рис. 28, А). Ее можно видеть в стебле подсолнечника, в побегах многих древесных растений.
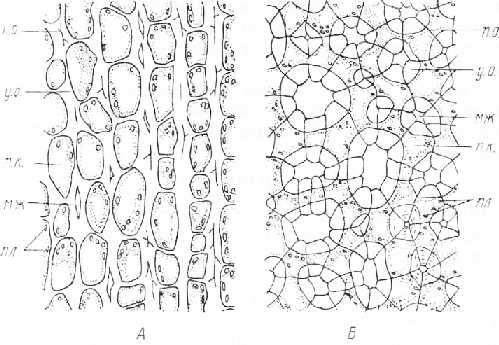
Рис. 3. Поперечный срез колленхимы. А — пластинчатая; Б — рыхлая: п. о. первичная оболочка, у. о.—утолщенная оболочка, п. к. — полости клеток с содержимым, пл — пластиды, мж — межклетники
Колленхима, в которой есть межклетники, называется рыхлой (рис. 28, Б). В ней сильно утолщены стенки, ограничивающие межклетник. Утолщенные участки оболочек имеют вид колец. На срезах, обработанных раствором иода в водном растворе иодистого калия, межклетники хорошо отличаются от полостей клеток отсутствием содержимого. Рыхлая колленхима характерна для черешков ревеня (Rheum), стеблей горца (Polygonum), мари белой (Chenopodium album L.)
Строение проводящих пучков в стебле тыквы (Cucurbita реро L.)
Все десять пучков стебля тыквы устроены одинаково, поэтому можно ограничиться рассмотрением одного крупного пучка.
В средней части пучка расположена ксилема, или древесина. В ней даже невооруженным глазом хорошо заметны крупные округлые полости клеток с одревесневшими оболочками. Это сосуды, или трахеи, — элементы, проводящие воду.
Наружная и внутренняя части пучка состоят из довольно мелких неодревесневших элементов, совокупность которых составляет флоэму, или луб. Между наружной флоэмой и ксилемой лежат живые тонкостенные клетки образовательной ткани — камбия. Такой пучок называется открытым биколлатераль-н ы м.
Ознакомившись с общим планом расположения проводящих тканей в пучке, следует перейти к их детальному изучению при большом (400—600 раз) увеличении микроскопа.
Флоэма
Флоэма, или луб, представляет собой сложную ткань, в состав которой наряду с проводящими элементами могут входить клетки паренхимы и флоэмные волокна (у тыквы волокон нет).
Проводящие элементы флоэмы — ситовидные трубки — образуются из удлиненных, составляющих вертикальный ряд клеток, принадлежащих прокамбию или возникших вследствие деления клеток камбия. Каждая клетка этого ряда делится продольной перегородкой. Одна из сестринских клеток разрастается в ширину, в ее поперечных стенках появляются мелкие сквозные отверстия — перфорации, или прободения, через которые проходят тяжи цитоплазмы, соединяющие протопласты соседних по вертикали клеток.
Перегородки со сквозными отверстиями получили название ситовидных пластинок. Вертикальный ряд клеток (члеников) с ситовидными пластинками составляет ситовидную трубку. Оболочки ситовидных трубок неодревесневшие, сравнительно тонкие. Содержимое представлено постенным слоем цитоплазмы и слизистым клеточным соком. В функционирующих трубках ядра обычно разрушаются.
Вторая клетка, отделявшаяся от материнской одновременно с клеткой-члеником ситовидной трубки, остается узкой: чаще всего она делится несколько раз поперечными перегородками, образуя тяж сопровождающих клеток, или клеток-спутниц. В течение всей жизни сопровождающие клетки сохраняют ядро и густую цитоплазму.
Для полного представления о строении флоэмы необходимо рассмотреть не только поперечные, но и продольные срезы. Структура наружной и внутренней флоэмы у тыквы одинакова. На поперечном срезе флоэмы ситовидные трубки наиболее широкопросветные, в очертании округлые или округло-многоугольные. Если ситовидные трубки перерезаны на уровне ситовидной пластинки, то на поперечном срезе эта пластинка со сквозными отверстиями видна в плане, а сопровождающие клетка большей частью не видны. Если ситовидная трубка перерезана посередине членика, то тогда очень хорошо видны плотно примыкающие к ситовидным трубкам мелкие сопровождающие клетки с густым зернистым содержимым.
Ситовидные пластинки в ситовидных трубках могут быть горизонтальными или слегка наклонными. На строго радиальном срезе стебля тыквы ситовидные пластинки обычно расположены почти на одном уровне, так как членики ситовидных трубок, находящихся в одном радиальном ряду, являются производными одной камбиальной клетки. В перерезанных ситовидных пластинках видны тонкие сквозные каналы. На боковых стенках ситовидных трубок встречаются небольшие округлые ситовидные поля, через которые проходят тончайшие цитоплазматические тяжи, соединяющие протопласты двух рядом расположенных трубок. Полости ситовидных трубок обычно заполнены свернувшимся при фиксации содержимым. В нем много белков, поэтому после обработки срезов йодным раствором содержимое становится желто-оранжевым. На продольном срезе содержимое ситовидных трубок имеет вид тяжей, пересекающих клетки и расширяющихся у ситовидных пластинок.
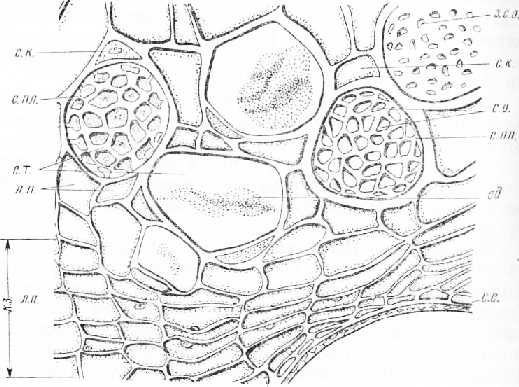
Рис. 4. Поперечный срез флоэмы и камбиальной зоны стебля тыквы: с. т. — ситовидные трубки, с. пл. — ситовидная пластинка, с. «. — сопровождающая клетка, л. л.—лубяная паренхима, с. о. — ситовидные отверстия, з. с. о. — закупоренные каллозой ситовидные отверстия, сд — содержимое ситовидной трубки, к. з. — камбиальная зона, с. с. — стенка сосуда
Если срез прошел косо по отношению к продольной оси стебля, то ситовидные пластинки имеют вид эллипса с овальными сквозными отверстиями, а перерезанные тяжи свернувшегося содержимого заметны как темные сгустки лишь у ситовидных пластинок.
По мере старения ситовидных трубок ситовидные прободения постепенно закупориваются особым веществом — каллозой, при этом отверстия сужаются и на поперечном срезе фпоэмы имеют вид небольших точек. К концу вегетационного периода каллоза образует мозолистые тела, которые представляют собой сильно преломляющие свет блестящие утолщения на обеих сторонах ситовидной пластинки.
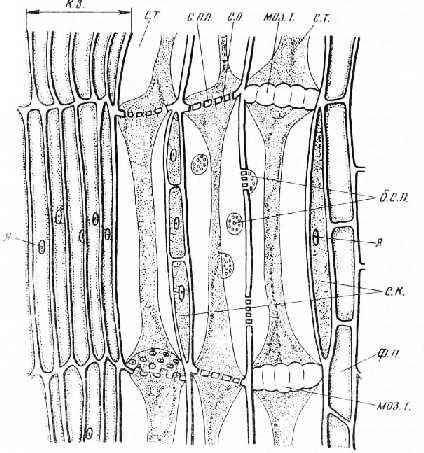
Рис. 5. Продольный срез флоэмы и камбиальной зоны стебля тыквы: с. т. — ситовидные трубки с тяжами содержимого, с. пл. — ситовидные пластинки, с. о.— ситовидные отверстия, б. с. п.— ситовидные поля на боковых стенках, с. к. — сопровождающие клетки, ф. п. — флоэмная паренхима, моз. т. — мозолистое тело, к. з. — камбиальная зона, я — ядро
Мозолистые тела особенно хорошо заметны на продольных срезах старой флоэмы, наиболее удаленной от камбия.
Вдоль членика ситовидной трубки, плотно прилегая к нему, расположена одна длинная или тяж из нескольких коротких сопровождающих клеток.
Заостренные концы краевых клеток тяжа находятся на уровне ситовидных пластинок, что указывает на общность происхождения сопровождающих клеток и члеников ситовидных трубок.
Живые паренхимные клетки, входящие в состав флоэмы, отличаются от члеников ситовидных трубок меньшими размерами и отсутствием сит, но они значительно крупнее сопровождающих клеток, сильнее вакуо-лизированы и кажутся более светлыми.
Камбиальная зона
В проводящем пучке между наружной флоэмой и ксилемой находятся живые тонкостенные клетки, составляющие камбиальную зону. Клетки расположены правильными радиальными рядами.
Все клетки этой зоны возникают в результате деления клеток однорядного слоя образовательной ткани — камбия.
Клетки камбия делятся продольными тангентальны-ми (т. е. параллельными поверхности органа) перегородками, откладывая клетки наружу, в сторону флоэмы, и внутрь, в сторону ксилемы.
Чем интенсивнее деление камбия, тем шире камбиальная зона.
На поперечном срезе клетки этой зоны имеют почти прямоугольные очертания. Тангентальные стенки клеток, находящихся в соседних радиальных рядах, обычно не совпадают.
На продольном срезе (рис. 30) клетки расположены ярусами. Они вытянуты в длину и имеют слабо заостренные или слегка закругленные окончания, находящиеся на одном уровне. Продольные стенки клеток одного яруса вклиниваются между стенками клеток соседнего по вертикали яруса. В клетке видны цитоплазма и ядро.
Периферические клетки камбиальной зоны впоследствии дифференцируются в элементы вторичной флоэмы. На удачно сделанных продольных радиальных срезах можно видеть, что длина клеток камбиальной зоны почти равна длине члеников ситовидных трубок. Из внутренних клеток камбиальной зоны формируются элементы вторичной ксилемы.
Ксилема
Ксилема (древесина) состоит из сосудов (трахей) с довольно толстой одревесневшей оболочкой. Наиболее широкопросветные сосуды видны даже невооруженным глазом. Между сосудами расположены мелкие клетки, в очертании многоугольные.
Задание.
1. При малом увеличении микроскопа зарисовать схему строения поперечного среза стебля тыквы, отметив на ней эпидермис, участки колленхимы, паренхиму, кольцо склеренхимы, биколлатеральные пучки с их тканями и центральную воздушную полость. При зарисовке схемы необходимо соблюдать масштаб. Лучше всего начинать схему с очертаний воздушной полости и контуров проводящих пучков. Это можно сделать, рассматривая срез под лупой. Очертания остальных тканей отмечают, рассматривая срез при малом увеличении микроскопа, передвигая препарат. Чтобы схема была наглядной, для каждой ткани выбирают условные обозначения (точки, одностороннюю или двустороннюю штриховку и т. п.). Можно пользоваться и цветными карандашами. Основную ткань не закрашивают, флоэму можно обозначить синим, ксилему — красным цветом, кружками в ней показывают наиболее крупные сосуды. Схема должна быть снабжена пояснительными надписями. Линии, выносящие надписи, желательно делать горизонтальными; если это почему-либо невозможно, нужно стараться не перекрещивать линии.
2. При большом увеличении микроскопа с поперечного среза детально зарисовать клетки эпидермиса, колленхимы, склеренхимы, флоэмы и камбиальной зоны. Во флоэме изобразить перерезанную ситовидную трубку с внутренним содержимым, ситовидную пластинку в плане, сопровождающие и паренхимные клетки.
3. С продольного среза зарисовать при большом увеличении микроскопа: а) ситовидную трубку с сопровождающими клетками, отметив в ней перерезанную ситовидную пластинку, тяжи свернувшегося содержимого, мозолистое тело, ситовидные поля на боковых стенках; б) клетки камбиальной зоны.
Строение проводящих элементов ксилемы в стебле подсолнечника однолетнего (Helianthus annuus L.)
Ксилема, или древесина, так же как и флоэма, представляет собой сложную ткань, состоящую не только из элементов, проводящих воду с растворенными в ней минеральными веществами, но и из клеток, выполняющих механическую и запасающую функции.
К проводящим элементам ксилемы относятся сосуды, или трахеи, и трахеиды. Трахеиды — это длинные прозенхимные клетки с сильно скошенными концами и многочисленными окаймленными порами на боковых стенках. Сосуды — полые трубки, представляющие собой вертикальный ряд удлиненных клеток (члеников). Смежные стенки, отделяющие одну клетку вертикального ряда от другой, в процессе развития сосудов разрушаются, на их месте возникают сквозные отверстия — перфорации. Боковые стенки члеников утолщаются и одревесневают, протопласты клеток отмирают.
Особенности строения проводящих элементов лучше всего видны на продольных срезах.
Кусочки стебля подсолнечника толщиной около 1 см, вырезанные из междоузлий и фиксированные спиртом, разрезают вдоль по радиусам и очищают от паренхимных клеток сердцевины. Затем делают тонкие радиальные срезы твердой части стебля или серию тангентальных срезов, внутренней зоны древесины, граничащей с сердцевиной. Срезы сначала обрабатывают спиртовым раствором флороглюцина, а затем крепкой соляной кислотой и заключают в глицерин. Их рассматривают при большом увеличении микроскопа.
Сосуды, расположенные близ сердцевины, имеют тонкую первичную целлюлозную оболочку и одревесневшую вторичную оболочку в виде отдельных колец. После проведения реакции на одревеснение кольца хорошо заметны благодаря красной окраске. Эти сосуды называют кольчатыми.
Рядом с ними, ближе к периферии стебля, находятся сосуды со вторичным утолщением оболочки в виде одинарной или двойной спирали. Это спиральные сосуды. Завитки спиралей отдельных клеток-члеников, слагающих сосуд, соединяясь концами, образуют единую
сплошную спираль. На препаратах спиральные и кольчатые сосуды могут быть разрезаны вдоль, тогда их утолщения имеют вид полуколец. Иногда в одном сосуде спиральные утолщения чередуются с кольчатыми. Этот промежуточный тип сосудов называют кольчато-спиральным.
Рис. 6. Сосуды на продольном срезе стебля подсолнечника: с. к. с. — спирально-кольчатый сосуд, с. с. — спиральный сосуд, с. с. р. — спиральный сосуд в разрезе, л. с. — лестничный сосуд, пор. с. — пористый сосуд, п. о. — первичная оболочка, в. о. — вторичная оболочка, о. п. — окаймленные поры, г. чл. — граница членика сосуда
Кольчатые, спиральные и спирально-кольчатые сосуды возникают в онтогенезе стебля очень рано, когда рост органа в длину еще не закончен. Они формируются из клеток прокамбия и принадлежат протоксилеме. По мере роста стебля неутолщенная первичная оболочка этих сосудов растягивается, а кольчатые утолщения и завитки спиральных утолщений раздвигаются. Впоследствии под давлением соседних клеток неутолщенные стенки кольчатых сосудов сдавливаются, их диаметр в этих местах уменьшается.
К периферии от спиральных сосудов расположены сосуды с более сильно утолщенной оболочкой, не способной к растяжению. Те из них, которые развиваются из прокамбия, относятся к метаксилеме. Протоксилема и метаксилема составляют первичную ксилему. Сосуды, образующиеся позднее, в результате деятельности камбия, входят в состав вторичной ксилемы. Сосуды метаксилемы и вторичной ксилемы имеют лестничные и сетчатые утолщения боковых стенок.
Формированию лестничного сосуда предшествует образование спиральных утолщений вторичной оболочки, причем отдельные обороты спиралей настолько сближены, что в некоторых местах они соединяются. Узкие пространства между несоединенными участками спиральных утолщений представляют собой поры. В плане они имеют вид вытянутых по ширине сосуда продолговатых или почти щелевидных участков, расположенных вертикальными рядами, как ступени в лестнице. Такой тип поровости стенок сосуда называется лестничным.
При сетчатом утолщении поры в стенках сосуда округлые или овальные в очертании и расположены в один или несколько рядов. В стебле подсолнечника эти сосуды находятся снаружи от лестничных. Многочисленные, очень мелкие, плотно примыкающие одна к другой поры можно было видеть в сосудах тыквы.
Строение пор сосудов отличается от строения простых пор в оболочках паренхимных клеток наличием окаймления. Окаймлением называется приподнятость вторичных слоев оболочки над поровым каналом (подробнее о строении окаймленных пор см. в разделе «Древесина хвойных растений»). Поры на продольных стенках сосудов служат для проведения воды в горизонтальном направлении. Если сосуд в поперечном сечении многоугольный, то на продольном срезе на его стенке заметны вертикальные полосы.
Большая длина как следствие разрушения поперечных стенок между клетками, утолщенные одревесневшие боковые стенки, обеспечивающие прочность, отсутствие живого содержимого характеризует сосуды как наиболее высокоорганизованный тип проводящих элементов, наилучшим образом выполняющих функцию проведения воды.
Кроме сосудов в состав ксилемы входят живые тонкостенные паренхимные клетки, располагающиеся между сосудами, а также сильно одревесневшие волокна, сходные по строению с волокнами склеренхимы.
Задание. При большом увеличении микроскопа зарисовать кольчатые, спиральные, лестничные и пористые сосуды. Обратить внимание на характер утолщений стенок, очертание и расположение пор, отметить перфорацию.
Анатомия растений — Ткани
gardenweb.ru
