
Локализованные гены в одной хромосоме
Закономерности наследования генов, локализованных в одной хромосоме, изучены Т. Морганом и его учениками в опытах на плодовой мушке дрозофиле. Это насекомое очень удобно для генетических исследований. Мушка очень плодовита: легко разводится при температуре 25°С в лабораторных условиях, она дает потомство каждые 10—15 дней, обладает многочисленными и разнооб разными наследственными признаками и малым числом хромосом (в диплоидном наборе — 8).
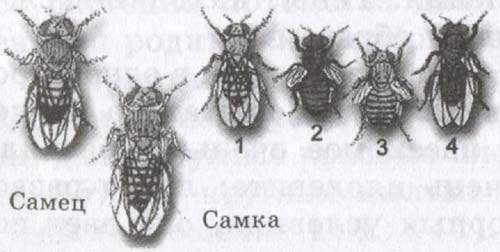
Разные наследственные формы мухи дрозофилы: 1 — серое тело — нормальные крылья; 2 — темное тело — зачаточные крылья; 3 — серое тело — зачаточные крылья; 4 — темное тело — нормальные крылья.
Последние две формы — результат перекомбинации признаков родительских форм из-за перекреста хромосом.
Статьи по теме:
Как известно из опытов, гены, локализованные в одной хромосоме, не обнаруживают независимого распределения и наследуются, в основном, совместно. Рассмотрим это на конкретном примере. Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья, с мушкой, обладающей темной окраской тела и зачаточными крыльями, то в первом поколении гибридов все мухи будут серыми с нормальными крыльями. Это гетерозиготы по двум парам аллелей (серое тело и нормальные крылья, темное тело и зачаточные крылья). Проведем анализирующее скрещивание дигетерозиготных самок (с серым телом и нормальными крыльями) с самцами, имеющими рецессивные признаки (темное тело и зачаточные крылья). В соответствии со вторым законом Менделя можно было бы ожидать получения в потомстве мух с четырьмя фенотипами: 25% с нормальными крыльями и серым телом, 25% с зачаточными крыльями и серым телом, 25% с нормальными крыльями и темным телом и 25% с зачаточными крыльями и темным телом. На самом деле на опыте мух с исходной комбинацией признаков (серое тело — нормальные крылья, темное тело — зачаточные крылья) оказывается значительно больше (в данном опыте 41,5%), чем мух с перекомбинированными признаками (серое тело — зачаточные крылья и темное тело — нормальные крылья). Последних везде будет по 8,5% каждого типа. Из этого примера видно, что гены, обусловившие признаки: серое тело — нормальные крылья и темное тело — зачаточные крылья, наследуются преимущественно вместе, или, иначе говоря, в сцепленном виде. Это сцепление является следствием локализации генов в одной и той же хромосоме. Поэтому при мейозе эти гены не расходятся, а наследуются вместе. Явление сцепления генов, локализованных в одной хромосоме, известно под названием закона Моргана.
biologylife.ru
Явление совместного наследования генов, локализованных в одной хромосоме, называют _ наследованием
Формы взаимодействия неаллельных генов как взаимодополняемость их действия или подавление одним геном неаллельного ему гена относятся к признакам
Характерное свойство мутаций — это
(*ответ*) повторное возникновение
ненаследуемость
групповой характер изменений
пределы изменчивости определяются генотипом
Хромосомную теорию наследственности сформулировал
(*ответ*) Морган
Хромосомы, одинаковые у обоих полов, называются
(*ответ*) аутосомами
Хромосомы, по которым женский и мужской пол различаются друг от друга, называются
(*ответ*) гетерохромосомами
соматическими
половыми
гаметами
Хромосомы, по которым мужской пол отличается от женского, называются _ хромосомами
(*ответ*) половыми
Хромосомы. одинаковые у обоих полов, называются
(*ответ*) аутосомами
соматическими
половыми
гаметами
Цель интенсивного сельскохозяйственного производства – это:
(*ответ*) максимальное производство продуктов питания
(*ответ*) минимальные затраты средств на единицу продукции
повсеместное разведение новых пород и сортов
синтез в промышленных условиях кормовых белков
Центры происхождения культурных растений и одомашнивания животных получили название центров
(*ответ*) доместикации
Центры происхождения культурных растений тесно связаны с районами одомашнивания животных. Такие центры получили названия
(*ответ*) доместикаций
окультуренных
деревенских
сельских
Человек создал для себя среду обитания, основанную на интелекте и развитии общественных отношений. Такая среда обитания называется
(*ответ*) искусственной
Черенок растения или почка, которые прививаются на корнесобственное растение, называется
(*ответ*) привоем
Число групп сцепления соответствуют числу хромосом в гаплоидном наборе. Так, у человека их
(*ответ*) 23
46
58
19
Эксперименты, связанные с передачей наследственных признаков из поколения в поколение, начались в _ веке
XVI
XVII
XIX
Эти мутации могут возникать вследствие утраты части хромосомы. Если в утраченный участок входят жизненно важные гены, то это приводит к гибели организма. Такие мутации получили название _ мутаций
(*ответ*) хромосомных
Этот гибрид пшеницы с рожью удачно сочетает в себе ценные признаки пшеницы и ржи, давая высокие урожаи зерна и зеленой массы. Гибрид получил название «_»
(*ответ*) тритикале
Этот метод селекции применяют к перекрестноопыляемым растениям, когда в посеве сохраняют растения только с желаемыми признаками. Такой метод называют _ отбором
(*ответ*) массовым
Явление совместного наследования генов, локализованных в одной хромосоме, называют _ наследованием
(*ответ*) сцепленным
индивидуальным
комбинированным
совместным
Явление, когда один ген определяет развитие нескольких признаков и свойств организмов, называется
(*ответ*) плейотропией www.soloby.ru
Локализация генов в одной хромосоме называется _ генов сцеплением оседанием фиксированием наследованием
Ген несет информацию о(об)
(*ответ*) синтезе молекулы белка
образовании организма
образовании органа
синтезе АТФ
Ген расположен в
(*ответ*) хромосоме
цитоплазме
ядерном соке
митохондриях
Гомогаметный пол обозначается
(*ответ*) ХХ
ХУ
УУ
УХ
Для установления генотипа фенотипически сходных организмов проводят скрещивание с
(*ответ*) гомозиготой по рецессивному признаку
гетерозиготой
гомозиготой по доминантному признаку
с любой особью
Единица расстояния между генами, находящимися в одной хромосоме, равная 1 % кроссинговера, называется
броунидой
дарвинидой
блекидой
Закон гомологических рядов наследственной изменчивости сформулировал
(*ответ*) Н.И.Вавилов
И.И.Мечников
Ч.Дарвин
В.И.Вернадский
Изменения кариотипа, вследствие которого одна из образовавшихся гамет содержит на одну хромосому больше, а другая – на одну хромосому меньше, называются
(*ответ*) анэуполиплоидией
полиплоидией
гетероплоидией
моноплоидией
Изменения структуры хромосом называются
(*ответ*) хромосомными
генными
соматическими
генеративными
Изменения, обусловленные заменой одного или нескольких нуклеотидов в пределах одного гена, называются
(*ответ*) генными
генеративными
соматическими
хромосомными
Изменчивость, обусловленная перегруппировкой хромосом в процессе полового размножения и участков хромосом в процессе кроссинговера, приводящей к изменению определенных признаков и свойств организма, называется
(*ответ*) комбинативной
модификационной
мутационной
генеративной
Количество генотипов при скрещивании Аа + Аа составляет
(*ответ*) 3
1
2
4
Количество фенотипов при скрещивании Аа + Аа в случае полного доминирования составляет
(*ответ*) 2
1
3
4
Локализация генов в одной хромосоме называется _ генов
(*ответ*) сцеплением
оседанием
фиксированием
наследованием
www.soloby.ru
Ekzamen_OTVETY_33_33_33_33_1 — Стр 7
вместо ожидаемого при дигибридном анализирующем скрещивании 1:1:1:1. Такое отклонение от независимого распределения означает, что гены, локализованные в одной хромосоме, наследуются совместно.Явление совместного наследования генов, локализованных в одной хромосоме, называется сцепленным наследованием, а локализация генов в одной хромосоме — сцеплением генов. Сцепленное наследование генов, локализованных в одной хромосоме, установил Морган.
53.
Условия менделирования признаков.
Менделирующие признаки у человека –
примерыМенделирующими
признаками называются те, наследование
которых про исходит по закономерностям,
установленным Г. Менделем. Менделирующие
признаки определяются одним геном
моногенно (от греч.monos-один) то есть когда
проявление признака определяется
взаимодействием аллельных генов, один
из которых доминирует (подавляет) другой.
Менделевские законы справедливы для
аутосомных генов с полной пенетрантностью
(от лат.penetrans-проникающий, достигающий)
и постоянной экспрессивностью (степенью
выраженности признака). Если гены
локализованы в половых хромосомах (за
исключением гомологичного участка в
Х- и У-хромосомах), или в одной хромосоме
сцеплено, или в ДНК органоидов, то
результаты скрещивания не будут следовать
законам Менделя. Общие законы
наследственности одинаковы для всех
эукариот. У человека также имеются
менделирующие признаки, и для него
характерны все типы их наследования:
аутосомно-доминантный, аутосомно-рецессивный,
сцепленный с половыми хромосомами (с
гомологичным участком Х- и У-хромосом).У
человека менделирующими признаками
являются, напр., альбинизм (отсутствие
пигментации, вызываемое рецессивным
геном; встречается у всех человеческих
рас с частотой 1 на 20— 30 тыс. новорожденных),
цвет глаз, характер волос (курчавые или
гладкие), групповые отличия по различным
факторам в крови (см.
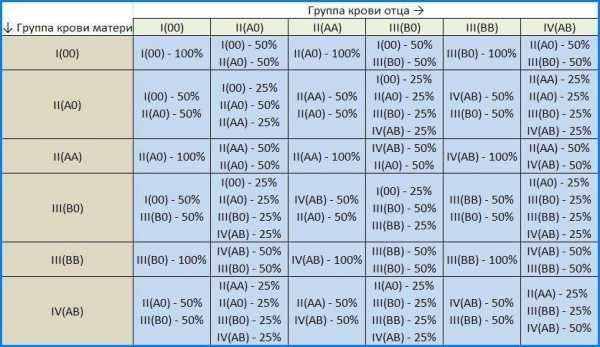
54. Множественные аллели. Наследование групп крови (АВО — системы) у человека.Множественный аллелизм — это существование в популяции более двух аллелей данного гена. В популяции оказываются не два аллельных гена, а несколько. Возникают в результате разных мутаций одного локуса. Гены множественных аллелей взаимодействуют между собой различным образом.В популяциях как гаплоидных, так и диплоидных организмов обычно существует множество аллелей для каждого гена. Это следует из сложной структуры гена — замена любого из нуклеотидов или иные мутации приводят к появлению новых аллелей. Видимо, лишь в очень редких случаях любая мутация столь сильно влияет на работу гена, а ген оказывается столь важным, что все его мутации приводят к гибели носителей. Так, для хорошо изученных у человека глобиновых генов известно несколько сотен аллелей, лишь около десятка из них приводит к серьезным патологиям.Гру́ппа кро́ви — описание индивидуальных антигенных характеристик эритроцитов, определяемое с помощью методов идентификации специфических групп углеводов и белков, включённых в мембраны эритроцитов животных.В наследовании групп крови есть несколько очевидных закономерностей:Если хоть у одного родителя группа крови I(0), в таком браке не может родиться ребёнок с IV(AB) группой крови, вне зависимости от группы второго родителя.1.Если у обоих родителей I группа крови, то у их детей может быть только I группа. 2.Если у обоих родителей II группа крови, то у их детей может быть только II или I группа. 3.Если у обоих родителей III группа крови, то у их детей может быть только III или I группа. 4.Если хоть у одного родителя группа крови IV(AB), в таком браке не может родиться ребёнок с I(0) группой крови, вне зависимости от группы второго родителя. 5.Наиболее непредсказуемо наследование ребенком группы крови при союзе родителей со II и III группами. Их дети могут иметь любую из четырёх групп крови.[30] Остальные варианты рассмотрены в таблице:
 55.
Полигенное наследование — примеры, в
том числе и у человека. это
Н. признака, контролируемого
совместно группой неаллельных
генов.
Широко распространенные болезни человека,
например, артериальная гипертензия,
ишемическая болезнь сердца, бронхиальная
астма, язвенная болезнь желудка,
наследуются полигенно. При этом тяжесть
заболевания зависит не только от
совокупного действия множества генов,
но и от провоцирующих средовых факторов.
При
скрещивании сорта пшеницы,
зерна которой имели темно-красную
окраску, с сортом, имеющим белые зерна,
гибриды первого поколения имели красную
окраску более светлых тонов. Во втором
поколении получилось такое соотношение
по фенотипу: на 63 окрашенных зерна с
различными оттенками красного цвета
приходилась 1 белое зерно (неокрашенное).
Эти результаты были объяснены Нильсоном-Эле
следующим образом. Темно-красная окраска
зерен пшеницы обусловлена действием
трех пар доминантных генов, а белая —
трех пар рецессивных, при этом по мере
увеличения числа доминантных генов
окраска становится более интенсивной.
Обозначим доминантные аллели трех
генов, локализованных в разных хромосомах,
прописными буквами А1 А2 А3 а рецессивные
— строчными а1 а1 а3, тогда генотипы
исходных форм будут: А1А1 А2А2 А3А3 x а1я1
а2а2 a33a.Окраска
зерен у гибридов первого
поколения A1a1 A2a2 A3a3 при наличии трех
доминантных аллелей будет промежуточной
светло-красной. При скрещивании гибридов
первого поколения A1a1 A2a2 A3a3 x A1a1 A2a2 A3a3 у
каждого из гибридов образуется по 8
типов гамет, поэтому во втором поколении
ожидается расщепление в 64-х долях (8 х
8). Среди 63/64 растений с окрашенными
зернами интенсивность окраски усиливается
по мере увеличения числа доминантных
аллелей различных генов в генотипе.
Видимо, каждый доминантный ген способствует
увеличению количества синтезированного
пигмента, и в этом смысле такой признак
можно отнести к количественным
55.
Полигенное наследование — примеры, в
том числе и у человека. это
Н. признака, контролируемого
совместно группой неаллельных
генов.
Широко распространенные болезни человека,
например, артериальная гипертензия,
ишемическая болезнь сердца, бронхиальная
астма, язвенная болезнь желудка,
наследуются полигенно. При этом тяжесть
заболевания зависит не только от
совокупного действия множества генов,
но и от провоцирующих средовых факторов.
При
скрещивании сорта пшеницы,
зерна которой имели темно-красную
окраску, с сортом, имеющим белые зерна,
гибриды первого поколения имели красную
окраску более светлых тонов. Во втором
поколении получилось такое соотношение
по фенотипу: на 63 окрашенных зерна с
различными оттенками красного цвета
приходилась 1 белое зерно (неокрашенное).
Эти результаты были объяснены Нильсоном-Эле
следующим образом. Темно-красная окраска
зерен пшеницы обусловлена действием
трех пар доминантных генов, а белая —
трех пар рецессивных, при этом по мере
увеличения числа доминантных генов
окраска становится более интенсивной.
Обозначим доминантные аллели трех
генов, локализованных в разных хромосомах,
прописными буквами А1 А2 А3 а рецессивные
— строчными а1 а1 а3, тогда генотипы
исходных форм будут: А1А1 А2А2 А3А3 x а1я1
а2а2 a33a.Окраска
зерен у гибридов первого
поколения A1a1 A2a2 A3a3 при наличии трех
доминантных аллелей будет промежуточной
светло-красной. При скрещивании гибридов
первого поколения A1a1 A2a2 A3a3 x A1a1 A2a2 A3a3 у
каждого из гибридов образуется по 8
типов гамет, поэтому во втором поколении
ожидается расщепление в 64-х долях (8 х
8). Среди 63/64 растений с окрашенными
зернами интенсивность окраски усиливается
по мере увеличения числа доминантных
аллелей различных генов в генотипе.
Видимо, каждый доминантный ген способствует
увеличению количества синтезированного
пигмента, и в этом смысле такой признак
можно отнести к количественным
56. Взаимодействие неаллельных генов — комплементарность, эпистаз, плейотропия — примеры.Комплементарное действие генов наиболее четко проявляется, когда скрещиваются две белые формы некоторых животных (кур) или растений (душистого горошка, белого клевера, кукурузы), а в потомстве появляются окрашенные формы. При скрещивании двух рас душистого горошка с белыми цветками (Lathyrus odoratus) в FI формируются растения с пурпурной окраской. При самоопылении этих растений в р2 наблюдается отклонение от менделевского расщепления: 9/ie растений имеют цветки с пурпурной окраской, тогда как 7/ie- с белой. В этом случае сущность взаимодействия генов, так же как при образовании ореховидного гребня у кур, состоит в том, что окрашенные цветки появляются при взаимодействии двух неаллельных доминантных генов А и В.Генотипы растений с неокрашенными цветками могут быть AAbb или ааВВ; если такие растения скрещиваются, то генотип растений FI будет АаВЬ. Это значит, что все растения этого поколения будут иметь пурпурные цветки. У кукурузы установлено, что для возникновения окрашенных семян необходим антоциан, который образуется только в присутствии двух доминантных генов А и В.Эпистатическое действие генов. Эпистаз — подавление действия одного гена другим, не аллельным, геном. Ген-подавитель, или супрессор, действует на подавляемый гипостатический ген по принципу, близкому к доминантности — рецессивности. Разница тут состоит в том, что эпистатический и гипостатический гены не являются ал-лельными, т. е. занимают различные локусы в гомологичных или негомологичных хромосомах. Эпистаз широко распространен в природе, в некоторых случаях изучены биохимические механизмы эпистатических взаимодействий. Так, у домашних птиц, в частности у кур, имеется зпистатическая система из двух генов, влияющих на окраску оперения. Эпистатический ген сам по себе не влияет на окраску пера. В то же время ген С в доминантной форме определяет нормальную продукцию пигмента. Однако белые леггорны с генотипом ССП не имеют пигмента в результате действия эпистатического гена. Есть и другие белые породы кур, окраска которых определяется другим генотипом. Так, оперение белых виандотов обусловлено тем, что они гомозиготны по рецессивным генам ее и ii, тур.-и по гипостатическому, и по эпистатическому генам. Расщепление при эпистазе, которое является результатом дигибридного скрещивания, отличается и от классического менделевского, и от того, которое наблюдается при комп- лементарностиАнализ данных на решетке Пеннета показывает, что только у 3/i6 потомства будет присутствовать ген С в доминантной форме и одновременно эпистатический ген в рецессивной форме i. Соотношение белых и пестрых птиц составит 13:3. Этот тип взаимодействия неаллельных генов называют доминантным эпистазом. В отличие от него при рецессивном эпистазе рецессивная аллель одного гена, будучи в гомозиготном состоянии, подавляет проявление доминантной или рецессивной аллели другого гена, В этом случае вместо ожидаемого при дигибридном скрещивании расщепления 9:3:3:1 получается отношение 9:7.Полимерия. До сих пор рассматривалось действие генов, определяющих контрастирующие, альтернативные признаки. С такого рода генами имел дело Мендель, экспериментируя на горохе. Однако уже с первого десятилетия XX в. известны гены, одинаково влияющие на признаки. Их действие может быть тождественным, но чаще бывает аддитивным, т. е. суммирующимся. В первом случае для развития признака достаточно присутствия одной из аллелей у дигетерозиготы в доминантной форме, тогда как аддитивное действие выражается в степени проявления признаков в зависимости от числа доминантных аллелей. Так, при аддитивном действии фенотип будет более выражен при генотипе ААВВ, чем при АаВЬ.
57. Сцепление генов. Основные положения хромосомной теории наследственности. Полное и неполное сцепление, кроссинговер.Сцепленное наследование — феномен скоррелированного наследования определенных состояний генов, расположенных в одной хромосоме.Полной корреляция не бывает из-за мейотического кроссинговера, так как сцепленные гены могут разойтись по разным гаметам. Кроссинговер наблюдается в виде расцепления у потомства тех аллелей генов и, соответственно, состояний признаков, которые были сцеплены у родителей.Наблюдения, проведенные Томасом Морганом, показали, что вероятность кроссинговера между различными парами генов разная, и появилась идея создать генные карты на основании частот кроссинговера между разными генами. Первая генная карта была построена студентом Моргана, Альфредом Стёртевантом (англ.) в 1913 году на материале Drosophila melanogaster.Расстояние между генами, расположенными в одной хромосоме, определяется по проценту кроссинговера между ними и прямо пропорционально ему. За единицу расстояния принят 1% кроссинговера (1 морганида или 1 сантиморганида). Чем дальше гены находятся друг от друга в хромосоме, тем чаще между ними будет происходить кроссинговер. Максимальное расстояние между генами, расположенными в одной хромосоме, может быть равно 49 сантиморганидам.Основные положения хромосомной теории наследственностиАнализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален. Аллельные гены занимают одинаковые локусыв гомологичных хромосомах. Гены расположены в хромосоме в линейной последовательности. Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола). Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами). Каждый биологический вид характеризуется определенным набором хромосом —кариотипом.Кроссинго́вер (другое название в биологии перекрёст) — явление обмена участками гомологичных хромосом во время конъюгации при мейозе. Помимо мейотического описан также митотический кроссинговер.Во время пахитены (стадия толстых нитей), гомологичные хромосомы находятся в состоянии конъюгации длительный период: у дрозофилы — четверо суток, у человека больше двух недель. Все это время отдельные участки хромосом находятся в очень тесном соприкосновении. Если в таком участке произойдет разрыв цепочек ДНК одновременно в двух хроматидах, принадлежащих разным гомологам, то при восстановлении разрыва может получиться так, что ДНК одного гомолога окажется соединенной с ДНК другой, гомологичной хромосомы. Этот процесс носит -название кроссинговера (англ. crossing-over — перекрест).Кроссинговер — взаимный обмен гомологичными участками хромосом между гомологичными (парными) хромосомами исходных гаплоидных наборов. Таким образом, особи имеют новые, различающиеся между собой генотипы. При этом достигается перекомбинация наследственных свойств родителей, что увеличивает изменчивость и дает более богатый материал для естественного отбора. Как правило, кроссинговер происходит только у женских организмов, однако, встречаются случаи кроссинговера и у гермафродитов.Гены перемешиваются благодаря слиянию гамет двух различных особей, однако генетические изменения осуществляются не только этим путем. Никакие два потомка одних и тех же родителей (если только это не идентичные близнецы) не будут абсолютно одинаковыми. Во время мейоза осуществляются два различных вида пересортировки генов. Один вид пересортировки — это результат случайного распределения разных материнских и отцовских гомологов между дочерними клетками при первом делении мейоза, каждая гамета получает свою, отличную от других выборку материнских и отцовских хромосом. Из этого следует, что клетки любой особи могут в принципе образовать 2 в степени n генетически различающихся гамет, где n — гаплоидное число хромосом. Однако на самом деле число возможных гамет неизмеримо больше из-за кросинговера (перекреста) — процесса, происходящего во время длительной профазы первого деления мейоза , когда гомологичные хромосомы обмениваются участками. У человека в каждой паре гомологичных хромосом кроссинговер происходит в среднем в 2 — 3 точках .При кроссинговере происходит разрыв двойной спирали ДНК в одной материнской и одной отцовской хроматиде, а затем получившиеся отрезки воссоединяются «наперекрест» (процесс генетической рекомбинации). Рекомбинация происходит в профазе первого деления мейоза, когда две сестринские хроматиды так тесно сближены друг с другом, что их невозможно увидеть в отдельности. Гораздо позже в этой растянутой профазе становятся ясно различимы две отдельные хроматиды каждой хромосомы. В это время видно, что они связаны своими центромерами и тесно сближены по всей длине. Два гомолога остаются связанными в тех точках, где произошел кроссинговер между отцовской и материнской хроматидами. В каждой такой точке, которую называют хиазмой , две из четырех хроматид перекрещиваются Таким образом, это морфологический результат произошедшего кроссинговера, который сам по себе недоступен для наблюдения.На этой стадии мейоза гомологи в каждой паре (или бивалент) остаются связанными друг с другом по меньшей мере одной хиазмой. Во многих бивалентах бывает большее число хиазм, так как возможны множественные перекресты между гомологами.Поскольку кроссинговер вносит возмущения в картину сцепленного наследования, его удалось использовать для картирования «групп сцепления» (хромосом). Возможность картирования была основана на предположении о том, что, чем чаще наблюдается кроссинговер между двумя генами, тем дальше друг от друга расположены эти гены в группе сцепления и тем чаще будут наблюдаться отклонения от сцепленного наследования. Первые карты хромосом были построены в 1913 г. для классического экспериментального объекта плодовой мушки Drosophila melanogaster Альфредом Стёртевантом, учеником и сотрудником Томаса Ханта Моргана.
58. Хромосомное определение пола. Наследование признаков, сцепленных с полом (на примере человека).У животных, растений и человека хромосомный механизм является начальным механизмом, определяющим пол. Согласно хромосомной теории, пол организма определяется половыми хромосомамив момент оплодотворения.XY определение полаУ одного пола ядра всех соматических клеток содержат диплоидный набораутосом2А и две одинаковыеполовые хромосомы(XX). Поэтому всегаметыэтого пола содержат по одной X-хромосоме. Это гомогаметный пол. У другого пола в каждой соматической клетке, помимо диплоидного набора аутосом 2А, содержатся две разные половые хромосомы Х и Y. Поэтому у него два вида гамет: X- и Y-несущие. Это гетерогаметный пол. У большинства видов животных и растений гомогаметен женский пол, а гетерогаметен мужской. Сюда относятся млекопитающие, большинство насекомых, многие рыбы, растения и др. К видам с мужской гетерогаметностью относятся и плодовые мушки родаDrosophila. Бывает два вида XY-определения пола. Один из них — как у человека: пол зависит от наличия Y-хромосомы (если она есть, генотип самца, если нет — самки). Второй — как у представителей рода Drosophila: пол определяется по соотношению числа X-хромосом и числа аутосом.Наследование, сцепленное с полом — наследование какого-либо гена, находящегося в половых хромосомах. Наследование признаков, проявляющихся только у особей одного пола , но не определяемых генами, находящимися в половых хромосомах, называется наследованием, ограниченным полом.Наследованием, сцепленным с X-хромосомой, называют наследование генов в случае, когда мужской пол гетерогаметен и характеризуется наличием Y-хромосомы (XY), а особи женского пола гомогаметны и имеют две X-хромосомы (XX). Таким типом наследования обладают все млекопитающие (в т.ч. человек), большинство насекомых и пресмыкающихся.
59. Понятие о гене, особенности его строения в клетках прокариот.Ген — структурная и функциональная единица наследственности, контролирующая развитие определенного признака или свойств.Ген — материальный носитель наследственной информации, совокупность которых родители передают потомкам во время размножения. В настоящее время, в молекулярной биологииустановлено, что гены — это участкиДНК, несущие какую-либо целостную информацию — о строении одноймолекулыбелкаили одной молекулыРНК. Эти и другие функциональные молекулы определяют рост и функционированиеорганизма.В то же время, каждый ген характеризуется рядом специфическихрегуляторных последовательностей ДНК, таких какпромоторы, которые принимают непосредственное участие в регулировании проявления гена. Регуляторные последовательности могут находиться как в непосредственной близости ототкрытой рамки считывания, кодирующей белок, или начала последовательности РНК, как в случае с промоторами (так называемыеcis-регуляторные элементы, англ.cis-regulatory elements), так и на расстоянии многих миллионов пар оснований (нуклеотидов), как в случае сэнхансерамиисупрессорами(иногда классифицируемые какtrans-регуляторные элементы, англ.trans-regulatory elements). Таким образом, понятие гена не ограничено только кодирующим участком ДНК, а представляет собой более широкую концепцию, включающую в себя и регуляторные последовательности.Изначально термин ген появился как теоретическая единица передачи дискретной наследственной информации. История биологии помнит споры о том, какие молекулымогут являться носителями наследственной информации. Большинство исследователей считали, что такими носителями могут быть толькобелки, так как их строение (20аминокислот) позволяет создать больше вариантов, чем строение ДНК, которое составлено всего из четырёх видовнуклеотидов. Позже было экспериментально доказано, что именно ДНК включает в себя наследственную информацию, что было выражено в видецентральной догмы молекулярной биологии.Гены могут подвергатьсямутациям— случайным или целенаправленным изменениям последовательности нуклеотидов в цепи ДНК. Мутации могут приводить к изменению последовательности, а следовательно изменению биологических характеристик белка или РНК, которые, в свою очередь, могут иметь результатом общее или локальное изменённое или анормальное функционирование организма. Такие мутации в ряде случаев являются патогенными, так как их результатом являетсязаболевание, или летальными наэмбриональномуровне. Однако, далеко не все изменения последовательности нуклеотидов приводят к изменению последовательности белка (благодаря эффекту вырожденностигенетического кода) или к существенному изменению последовательности и не являются патогенными. В частности, геном человека характеризуетсяоднонуклеотидными полиморфизмамиивариациями числа копий(англ.copy number variations), такими как делецииидупликации, которые составляют около 1 % всей нуклеотидной последовательности человека[1]. Однонуклеотидные полиморфизмы, в частности, определяют различные аллелиодного гена.Свойства генов:дискретность— несмешиваемость генов;стабильность— способность сохранять структуру;лабильность— способность многократно мутировать;множественный аллелизм— многие гены существуют в популяции во множестве молекулярных форм;аллельность— в генотипе диплоидных организмов только две формы гена;специфичность— каждый ген кодирует свой продукт;плейотропия— множественный эффект гена;экспрессивность— степень выраженности гена в признаке;пенетрантность— частота проявления гена в фенотипе;амплификация— увеличение количества копий гена..Классификация генов: 1) Структурные гены — уникальные компоненты генома, представляющие единственную последовательность, кодирующую определенный белок или некоторые виды РНК(тРНК, РРНК, м/яРНК) 2) Функциональные гены — регулируют работу структурных геновРазличные организмы резко отличаются по количеству ДНК, составляющей их геномы. У вирусов в зависимости от их величины и сложности размер генома колеблется от нескольких тысяч до сотен пар нуклеотидов. Гены в таких просто устроенных геномах расположены один за другим и занимают до 100% длины соответствующей нуклеиновой кислоты(РНК и ДНК). Для многих вирусов становлена полная нуклеотидная последовательность ДНК. У бактерий размер генома значительно больше. У кишечной палочки единственная нить ДНК – бактериальная хромосома состоит из 4,2х106(6 степень) пар нуклеотидов. Более половины этого количества состоит из структурных генов, т.е. генов, кодирующих определенные белки. Остальную часть бактериальной хромосомы составляют неспособные транскрибироваться нуклеотидные последовательности, функция которых не вполне ясна. Подавляющее большинство бактериальных генов уникальны, т.е. представлены в геноме один раз. Исключение составляют гены транспортных и рибосомальных РНК, которые могут повторяться десятки раз. Геном эукариот, особенно высших, резко превышает по размерам геном прокариот и достигает, как отмечалось, сотен миллионов и миллиардов пар нуклеотидов. Количество структурных генов при этом возрастает не очень сильно. Количество ДНК в геноме человека достаточно для образования примерно 2 млн. структурных генов. Реально имеющееся число оценивается как 50-100 тыс. генов, т.е. в 20-40 раз меньше того, что могло бы кодироваться геномом такого размера. Следовательно, приходится констатировать избыточность генома эукариот. Причины избыточности в настоящее время в значительной степени прояснились: во-первых, некоторые гены и последовательности нуклеотидов многократно повторены, во-вторых, в геноме существует много генетических элементов, имеющих регуляторную функцию, в-третьих, часть ДНК вообще не содержит генов
|
Согласно
современным представлениям, ген,
кодирующий синтез определенного
белка, у эукариот состоит из нескольких
обязательных элементов. Прежде всего
это обширная регуляторная зона,
оказывающая сильное влияние на
активность гена в той или иной ткани
организма на определенной стадии его
индивидуального развития. Далее
расположен непосредственно примыкающий
к кодирующим элементам гена промотор
– последовательность ДНКдлиной до 80-100 пар нуклеотидов,
ответственная за связывание
РНК-полимеразы, осуществляющейтранскрипциюданного гена. Вслед за промотором
лежит структурная часть гена, заключающая
в себе информацию о первичной структуре
соответствующегобелка.
Эта область для большинства генов
эукариот существенно короче регуляторной
зоны, однако ее длина может измеряться
тысячами пар нуклеотидов.
Важная
особенность эукариотических генов –
их прерывность. Это значит, что область
гена, кодирующая белок, состоит из
нуклеотидных последовательностей
двух типов. Одни – экзоны – это участкиДНК,
которые несут информацию и строениибелкаи
входят в состав соответствующих РНК
и белка. Другие – интроны – не кодируют
структурубелкаи в состав зрелой молекулы и-РНК не
входят, хотя и транскрибируются.
Процесс вырезания интронов – «ненужных»
участков молекулыРНКи сращивания экзонов при образовании
и-РНК осуществляется специальными
ферментами и получил название Сплайсинг
(сшивание, сращивание). |
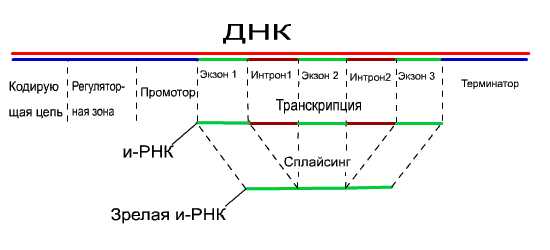 Экзоны обычно соединяются вместе в
том же порядке, в котором они распологаются
в ДНК.
Однако не абсолютно все гены эукариот
прерывисты. Иначе говоря, у некоторых
генов, подобно бактериальным, наблюдается
полное соответствие нуклеотидов
последовательности первичной структуре
кодируемых имибелков.
Таким образом, ген эукариот во многом
похож на оперон прокариот, хотя и
отличается от него более сложной и
протяженной регуляторной зоной, а
также тем, что он кодирует обычно
только одинбелок,
а не несколько, как оперон у бактерии.
Экзоны обычно соединяются вместе в
том же порядке, в котором они распологаются
в ДНК.
Однако не абсолютно все гены эукариот
прерывисты. Иначе говоря, у некоторых
генов, подобно бактериальным, наблюдается
полное соответствие нуклеотидов
последовательности первичной структуре
кодируемых имибелков.
Таким образом, ген эукариот во многом
похож на оперон прокариот, хотя и
отличается от него более сложной и
протяженной регуляторной зоной, а
также тем, что он кодирует обычно
только одинбелок,
а не несколько, как оперон у бактерии.60. Генетический код и его свойства. Структурная и функциональная классификация генов. ПримерыГенети́ческий код — это свойственный всем живым организмамспособкодированияаминокислотнойпоследовательностибелковпри помощи последовательностинуклеотидов.ВДНКиспользуется четыре нуклеотида —аденин(А),гуанин(G),цитозин(С),тимин(T), которые в русскоязычной литературе обозначаются буквами А, Г, Ц и Т. Эти буквы составляюталфавитгенетическогокода. В РНК используются те же нуклеотиды, за исключением тимина, который заменён похожим нуклеотидом —урацилом, который обозначается буквой U (У в русскоязычной литературе). В молекулахДНКиРНКнуклеотиды выстраиваются в цепочки и, таким образом, получаются последовательности генетических букв.Для построения белков в природе используется 20 различныхаминокислот. Каждый белок представляет собой цепочку или несколько цепочек аминокислот в строго определённой последовательности. Эта последовательность определяет строение белка, а следовательно все его биологические свойства. Набор аминокислот также универсален для почти всех живых организмов.Реализация генетической информациив живых клетках (то есть синтез белка, кодируемогогеном) осуществляется при помощи двух матричных процессов:транскрипции(то есть синтезаиРНКна матрицеДНК) итрансляциигенетического кода в аминокислотную последовательность (синтез полипептидной цепи на матрицеиРНК). Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов. Набор из трёх нуклеотидов называетсятриплетом. Свойства генетического кодаТриплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно. Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов. (Не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки). Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте. (Свойство не является универсальным. Кодон UGA уEuplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин)[1] Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов. Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусовдочеловека(на этом основаны методыгенной инженерии) (Из этого свойства также есть ряд исключений, см. таблицу в разделе «Вариации стандартного генетического кода» в данной статье). 61. Избыточность ДНК эукариот. Структурная и функциональная классификация генов.Количество ДНК в хромосомах велико и возрастает по мере усложнения организмов. Для эукариот также характерна избыточность генов. Так, у человека геном содержит число нуклеотидных пар, достаточное для образования более 2 млн. структурных генов, в то время как у человека имеется по данным 2000 года 31 тыс. всех генов.Ген — структурная и функциональная единица наследственности, контролирующая развитие определённого признака или свойства. Совокупность генов родители передают потомкам во время размножения. Однако перенос генов от родителей к потомкам не является единственным способом передачи генов.труктурные гены — уникальные компоненты генома, представляющие единственную последовательность, кодирующую определённый белок или некоторые виды РНК. (См. также статью гены домашнего хозяйства).Функциональные гены — регулируют работу структурных генов
studfiles.net
4. Особенности локализации генов в половых хромосомах
Анализ локализации генов в половых хромосомах показал что Х- и У-хромосомы отличаются друг от друга по генетическому составу и по большинству генов они не гомологичны друг другу.
Лишь небольшие участки X— и У-хромосом являются гомологичными. В этих участках расположены аллельные гены, и их наследование осуществляется в соответствии с менделеевскими закономерностями наследования. У человека есть аллергическое заболевание – геморрагический диатез, вызываемый рецессивным геном. Аллели этого гены находятся в Х и У хромосоме.
Значительная часть генов более крупной Х-хромосомы не имеет аллельных генов в
У-хромосоме. В ней содержится небольшая группа генов, отсутствующих в Х-хромосоме. Гены, локализованные в негомологичных участках половых хромосом, наследуется сцеплено с Х-хромосомой или сцеплено с У-хромосомой.
Для генов сцепленных с У-хромосомой, характерно наследование только по мужской линии (от отца к сыновьям, затем к внукам – голандрическоенаследование). Для генов, сцепленных с Х-хромосомой, характер наследования зависит от того, является ли данный аллель доминантным или рецессивными. ХА — сцепленный доминантный аллель значительно чаще проявляется у особей женского пола, так как они гомогаметны (ХХ) и могут получить его с Х-хромосомой отца и матери. Ха — сцепленный рецессивный аллель чаще проявляется у представителей гетерогаметного мужского пола, так как У-хромосома соответствующего локуса не имеет и рецессивный аллель Х — хромосомы проявляется, будучи в единственном числе.Состояние генотипа, при котором вследствие негомологичности половых хромосом за развитие признака отвечает один аллель, называется гемизиготным. Следовательно, особи мужского пола гемизиготны по генам, локализованы в негомологичных участкахXи У — хромосомах – (гемизиготный генотип: ХАУ, Ха У, ХУГ) развитие соответствующих признаков зависит у них от того, какие аллели несутX- и У — хромосомы.
-
Наследование признаков, сцепленных с полом у человека.
Более 200 генов человека, локализованы в Х-хромосоме.
По рецессивному типу у человека сцеплено с Ха— хромосомой наследуется такие заболевания как:
— дальтонизм (цветовая слепота),
— гемофилия(несвертываемость крови),
— ангидрозная эктодермальная дисплазия(отсутствие потовых желез),
— миопатия — мышечная дистрофия Дюшена и Беккера,
— ювенильная глаукома (нарушение оттока внутриглазной жидкости),
— пигментный ретинит (сужение поля зрения),
— атрофия зрительного нерва,
— ангиокератома(заболевание кожи),
— глазной альбинизм,
— гемералопия– ночная, или куриная слепота,
— одна из форм ихтиоза и глухонемоты,
— агаммаглобулинемии(пониженная устойчивость к бактериальным заболеваниям)
— Эктодермальная дисплазия. Симптомокомплекс эктодермальной дисплазии проявляется во врожденном поражении всех структур эктодермы: кожи, волос, ногтей, частичным или полным отсутствием зачатков зубов как в период прикуса молочных, так и постоянных зубов и т.д. Наиболее тяжелыми поражениями структур кожи, слизистых, волос, полной или выраженной первичной частичной адентией всех групп зубов молочного и постоянного прикуса проявляется ангидротическая группа
— гипосозревание — наследственная гипоплазия эмали, связанная с нарушением ее
созревания.
— свыше 60 генов определяющих синдромы умственной отсталости.
Так как эти признаки наследуются по рецессивному типу, у мужчин они встречаются гораздо чаще, чем у женщин. Мужчины гемизиготныХа У по данным локусам. При наличии единичного аллеля дальтонизма или гемофилии в их единственной Х-хромосоме, соответствующие заболевания проявляются. У женщин дальтонизм или гемофилия могут проявиться только в гомозиготном состоянии: при наличии рецессивных аллелей в обеих Х – хромосомах ХаХа. Сыновья наследуют эти признаки от матерей (100% сыновей, если мать гомозиготна, и 50% сыновей, если она гетерозиготна). Для проявления дальтонизма или гемофилии у дочерей необходимо наличие соответствующего признака у отца — Ха У, в то время как мать должна быть гетерозиготной по данному признаку ХАХа. Признак проявится у 50% дочерейТаким образом, у родителей с нормальным цветовым зрением или нормальной свертываемости крови, дальтонизм или гемофилия может проявиться только у сыновей, которые наследуют соответствующий аллель от гетерозиготной матери – носительницы патологического гена.
К доминантным сцепленным с Х-хромосомой признакам относятся–
— рахит,резистентный к витаминуD(не поддающейся лечению витаминомD),
— анемия примахиновая или фавизм(при приёме фенацетина, сульфамидных препаратов происходит гемолиз эритроцитов),
— гипоплазия эмали зубов истончение эмали, сопровождается изменения цвета зубов.
studfiles.net
4. Особенности локализации генов в половых хромосомах
Анализ локализации генов в половых хромосомах показал что Х- и У-хромосомы отличаются друг от друга по генетическому составу и по большинству генов они не гомологичны друг другу.
Лишь небольшие участки X— и У-хромосом являются гомологичными. В этих участках расположены аллельные гены, и их наследование осуществляется в соответствии с менделеевскими закономерностями наследования. У человека есть аллергическое заболевание – геморрагический диатез, вызываемый рецессивным геном. Аллели этого гены находятся в Х и У хромосоме.
Значительная часть генов более крупной Х-хромосомы не имеет аллельных генов в
У-хромосоме. В ней содержится небольшая группа генов, отсутствующих в Х-хромосоме. Гены, локализованные в негомологичных участках половых хромосом, наследуется сцеплено с Х-хромосомой или сцеплено с У-хромосомой.
Для генов сцепленных с У-хромосомой, характерно наследование только по мужской линии (от отца к сыновьям, затем к внукам – голандрическоенаследование). Для генов, сцепленных с Х-хромосомой, характер наследования зависит от того, является ли данный аллель доминантным или рецессивными. ХА — сцепленный доминантный аллель значительно чаще проявляется у особей женского пола, так как они гомогаметны (ХХ) и могут получить его с Х-хромосомой отца и матери. Ха — сцепленный рецессивный аллель чаще проявляется у представителей гетерогаметного мужского пола, так как У-хромосома соответствующего локуса не имеет и рецессивный аллель Х — хромосомы проявляется, будучи в единственном числе.Состояние генотипа, при котором вследствие негомологичности половых хромосом за развитие признака отвечает один аллель, называется гемизиготным. Следовательно, особи мужского пола гемизиготны по генам, локализованы в негомологичных участкахXи У — хромосомах – (гемизиготный генотип: ХАУ, Ха У, ХУГ) развитие соответствующих признаков зависит у них от того, какие аллели несутX- и У — хромосомы.
-
Наследование признаков, сцепленных с полом у человека.
Более 200 генов человека, локализованы в Х-хромосоме.
По рецессивному типу у человека сцеплено с Ха— хромосомой наследуется такие заболевания как:
— дальтонизм (цветовая слепота),
— гемофилия(несвертываемость крови),
— ангидрозная эктодермальная дисплазия(отсутствие потовых желез),
— миопатия — мышечная дистрофия Дюшена и Беккера,
— ювенильная глаукома (нарушение оттока внутриглазной жидкости),
— пигментный ретинит (сужение поля зрения),
— атрофия зрительного нерва,
— ангиокератома(заболевание кожи),
— глазной альбинизм,
— гемералопия– ночная, или куриная слепота,
— одна из форм ихтиоза и глухонемоты,
— агаммаглобулинемии(пониженная устойчивость к бактериальным заболеваниям)
— Эктодермальная дисплазия. Симптомокомплекс эктодермальной дисплазии проявляется во врожденном поражении всех структур эктодермы: кожи, волос, ногтей, частичным или полным отсутствием зачатков зубов как в период прикуса молочных, так и постоянных зубов и т.д. Наиболее тяжелыми поражениями структур кожи, слизистых, волос, полной или выраженной первичной частичной адентией всех групп зубов молочного и постоянного прикуса проявляется ангидротическая группа
— гипосозревание — наследственная гипоплазия эмали, связанная с нарушением ее
созревания.
— свыше 60 генов определяющих синдромы умственной отсталости.
Так как эти признаки наследуются по рецессивному типу, у мужчин они встречаются гораздо чаще, чем у женщин. Мужчины гемизиготныХа У по данным локусам. При наличии единичного аллеля дальтонизма или гемофилии в их единственной Х-хромосоме, соответствующие заболевания проявляются. У женщин дальтонизм или гемофилия могут проявиться только в гомозиготном состоянии: при наличии рецессивных аллелей в обеих Х – хромосомах ХаХа. Сыновья наследуют эти признаки от матерей (100% сыновей, если мать гомозиготна, и 50% сыновей, если она гетерозиготна). Для проявления дальтонизма или гемофилии у дочерей необходимо наличие соответствующего признака у отца — Ха У, в то время как мать должна быть гетерозиготной по данному признаку ХАХа. Признак проявится у 50% дочерейТаким образом, у родителей с нормальным цветовым зрением или нормальной свертываемости крови, дальтонизм или гемофилия может проявиться только у сыновей, которые наследуют соответствующий аллель от гетерозиготной матери – носительницы патологического гена.
К доминантным сцепленным с Х-хромосомой признакам относятся–
— рахит,резистентный к витаминуD(не поддающейся лечению витаминомD),
— анемия примахиновая или фавизм(при приёме фенацетина, сульфамидных препаратов происходит гемолиз эритроцитов),
— гипоплазия эмали зубов истончение эмали, сопровождается изменения цвета зубов.
studfiles.net
Ответы@Mail.Ru: почему проявляются в виде признака рецессивные гены, локализованные в X
Нет пары в У -хромосоме.
Так вопрос НЕ о сцепленности с полом! ! В результате мейоза образуются гаметы, либо Х и Х (у женщин) , либо Х и У (у мужчин) . ВОЗМОЖЕН хромосомный набор ХХХ (девочка) , ХХУ (мальчик) , ХО (девочка) , УО (нежизнеспособен) . У человека известны рецессивные сцепленные с полом признаки, такие как гемофилия, дальтонизм, мышечная дистрофия и др. В Х – хромосоме есть ген, определяющий признак свертывания крови. Рецессивный ген вызывает развитие гемофилии. У женщин две ХХ – хромосомы. Рецессивный признак (гемофилия) проявляется в том случае, если гены, отвечающие за него, находятся в двух Х — хромосомах. Если организм гетерозиготен по этим генам, то гемофилия не проявится. Функциональную активность целого ряда других генов определяет гормональная деятельность организма. Например, ген определяющий облысение, типичное для мужчин, локализован в аутосоме, и его проявление зависит от мужских половых гормонов. У мужчин этот ген действует как доминантный, а у женщин как рецессивный. Если у женщин этот ген в гетерозиготном состоянии, то признак не проявляется. Даже в гомозиготном состоянии у женщин этот признак слабее выражен, чем у мужчин. Взаимозаменяемые гены двух хромосом, определяющие один и тот же признак, называются аллельными, или аллелями. Если оба аллеля одинаковы, то организм называется гомозиготным по данному гену, если аллели различны, — гетерозиготным по данному гену. Если в фенотипе проявляются оба аллельных гена, например, в группе крови АВ (IV), эти гены называются кодоминантными. Если аллель обусловливает преимущественное проявление признака, он называется доминантным, а другой, подавленный, аллель — рецессивным. Например, при генотипе группы крови АО непроявляющийся аллель 0 рецессивен, а проявляющийся аллель доминантен, и группа крови будет А (II). В гетерозиготном организме доминантным может оказаться ген, определяющий то или иное заболевание. Тогда, несмотря на наличие второго нормального аллеля, заболевание проявится. Доминантные заболевания, как правило, проявляются в гетерозиготном организме. При гомозиготности по доминантным патологическим генам плод обычно погибает на ранних стадиях развития из-за больших дефектов метаболизма и строения тела, поэтому в популяции такие лица обычно не встречаются. Если ген, определяющий болезнь, окажется рецессивным, то он может проявить свое действие только в гомозиготном состоянии. Рецессивные заболевания протекают тяжелее, чем доминантные. У мужчин гены, локализованные в непарных половых Х- и У-хромосомах, представлены одинарным набором, а у женщин парные Х-хромосомы и локализованные в них гены представлены двойным набором. Поэтому в женском организме патологические рецессивные гены, локализованные в Х-хромосоме, проявляются только в гомозиготном состоянии. У мужчин в этом случае патологический рецессивный ген проявит свое действие. По этой причине мать может передать патологический ген Х-хромосомы сыновьям, сама не страдая этим заболеванием, если во второй Х-хромосоме находится нормальный аллель данного гена. Из вышесказанного можно ответить на вопрос : если в организме две гаметы с рецессивными генами — для гомозигот, или Х-хромосома несёт рецессивный признак — для гетерозигот. Относительно женщин и дальтонизма : МОЯ ТЁЩА ЦВЕТА НЕ РАЗЛИЧАЕТ! Уникум!
touch.otvet.mail.ru
