
Эволюция птиц — Википедия
Материал из Википедии — свободной энциклопедии
Эволюция птиц началась, согласно традиционным представлениям, в юрском периоде. Согласно этой версии, самым древним из известных видов птиц считается археоптерикс (Archaeopteryx lithographica). Птицы в этом случае рассматриваются как специализированные представители теропод, близкие к другим оперённым динозаврам из группы манирапторов, например дромеозавридам.
Вместе с тем ряд авторов придерживается альтернативной версии, согласно которой, первой достоверной находкой птиц является довольно спорное ископаемое протоавис из позднего триаса. Согласно этой версии, и птицы, и тероподы произошли от одной из более ранних групп архозавров.
В настоящее время выдвинуто несколько различных гипотез происхождения и эволюции птиц, но общепринятой филогении нет.
«с деревьев вниз» (arboreal; trees down)
ru.wikipedia.org
Эволюция птиц. Гипотезы происхождения птиц. Оперенные динозавры
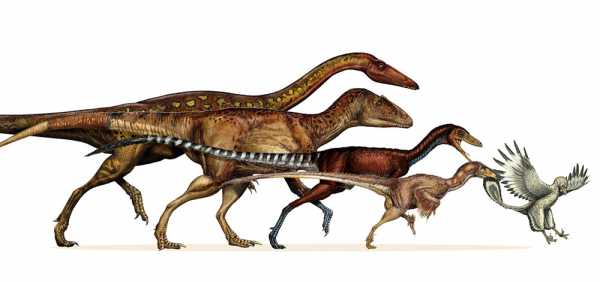
Современные птицы сильно отличаются от родственных им позвоночных. До недавнего времени их происхождение было одной из великих тайн биологии. Перья, беззубые клювы, полые и глубокие грудные кости являются лишь частью особых признаков, которых нет у других животных. Наблюдение за птицами не объясняет, как у них появились перья и способность к полету. За последние десятилетия появились новые открытия и новые методы исследования, которые позволили палеонтологам совершить несколько открытий. Найденные в районе Китая, Южной Америки окаменелости, а также образцы, хранящиеся в музеях, которые теперь можно изучать с помощью новых технологий, стали причиной возникновения очередной волны интереса к изучению истории эволюции птиц. Благодаря этому, ранее появилась теория их происхождения от мелких хищных динозавров, обитавших на Земле в конце юрского периода.

Начало исследования возникновения птиц
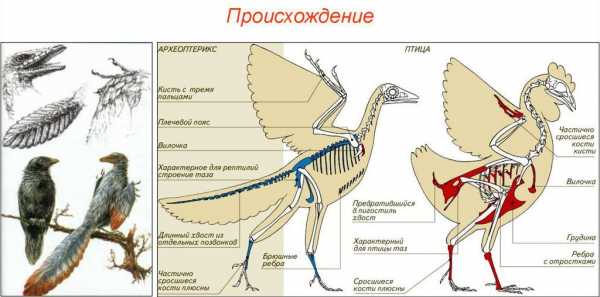
Ученые принялись размышлять об эволюции птиц, начиная с того времени, как Чарльз Дарвин изложил свою теорию эволюции в работе «О происхождении видов». В 1861 году, год спустя после публикации трактата Дарвина, древнее птичье перо было найдено в баварских известняковых отложениях, возраст которых составлял около 150 млн лет. В следующем году был найден скелет животного, у которого были птичьи крылья и перья, очень длинный костлявый хвост и челюсть с зубами. Он был найден в том же регионе. Окаменевший скелет был назван археоптериксом. Он стал первым найденным древнейшим животным с оперением. Скелетная анатомия археоптерикса давала четкие доказательства, что предками птиц были динозавры, но в 1861 году ученые еще не смогли установить эту связь. Тогда и началось изучение эволюции птиц и охота за предками современных видов пернатых.

Анатомическое строение археоптерикса
Окаменелость была обнаружена в начале 1860 годов. Долгое время знаменитая находка из поздней юры была уникальной. Она единственная могла дать какую-то информацию о том, как мог произойти эволюционный переход от рептилий к птицам, поскольку в ней сочетались признаки как птицы, так и рептилии. У строения археоптерикса было много общего со строением птиц. Например, перья вдоль передних лап, ставших крыльями. Но, в отличие от современных видов, также присутствовали зубы и костистый хвост.
Первые теории о животных юрского периода
Стоит отметить, что многие из костей животного, в том числе передних лап, плечевого пояса, таза и ног, были отчетливыми, не сросшимися, как у его потомков. Несколько лет спустя Томас Генри Хаксли стал первым ученым, кто нашел связь между строением птиц и динозаврами. Он сравнивал задние конечности гигантского динозавра с изображением страуса и отметил 35 признаков, доказывающих, что они связаны друг с другом. Хаксли представил свои результаты Геологическому обществу. Но в 1870 году в Лондоне палеонтолог Гарри Говье Сили решил оспорить гипотезу происхождения птиц и их родство с динозаврами. Сили предположил, что задние конечности страуса и динозавра могут выглядеть похожим только потому, что эти животные были большими и двуногими, а свои задние конечности использовали в сходных условиях. Кроме того, динозавры были даже больше, чем страусы, и никто из них не мог летать. У ученого возникал вопрос, как тогда могут летать птицы, если они эволюционировали от динозавра.
Теория Герхарда Хайльмана
Тайна происхождения птиц вновь заинтересовала ученых спустя приблизительно полвека. В 1916 году Герхард Хайльман, медицинский врач, интересующийся палеонтологией, опубликовал на датском языке блестящую книгу об истории происхождения птиц, которая в 1926 году была переведена на английский под названием “Происхождение птиц”. Хайльман показал, что пернатые были анатомически более похожи на динозавров-тероподов, чем на любую другую ископаемую группу. Но существовало одно неизбежное несоответствие: у тероподов явно отсутствовали ключицы, те самые, которые сливаются в вилочку у птиц.
Недостатки теории Герхарда Хайльмана
Из-за того факта, что у других рептилий имелись ключицы, Хайльман предположил, что тероподы, в частности триасовые псевдозухии, по какой-то причине потеряли их. Для него эта потеря означала, что птицы не могли развиться от этого вида динозавров. Потому он был убежден, как позже выяснилось, ошибочно, что функция ключиц, потерянная во время эволюции, не может быть восстановлена. Птицы, утверждал он, должны были развиться из более архаичной рептильной группы, у которой были ключицы.

Как ранее Сили, Хайльман пришел к выводу, что сходство между птицами и динозаврами должно просто отражать тот факт, что обе группы были двуногими. Его выводы повлияли на палеонтологические исследования и в течение длительного времени были приоритетными, хотя новая появившаяся информация и опровергла некоторые факты. Но до сих пор некоторые ученые придерживаются именно ее. Два отдельных исследования показали, что тероподы действительно имели ключицы. В 1924 году был опубликован анатомический рисунок необычного теропода с попугайным оперением. Он был назван овираптором, и его существование опровергало теорию Хайльмана.

Новые доказательства
В 1936 году Чарльз Камп из Калифорнийского университета в Беркли нашел останки небольшого теропода раннего юрского периода и хотел поставить точку в истории с ключицами. Но доказательство ошибочности теории Хайльмана было мало кем признано. Недавние исследования нашли ключицы у многих тероподов. Исходя из строения птиц в настоящее время, а также в результате ряда исследований, археоптерикс был назван промежуточным звеном в истории эволюции пернатых. Но оставался вопрос: между какими именно рептилиями? Дальнейшие ископаемые птицы найдены в слоях ниже раннего мела. Большинство из них были обнаружены с начала 1990 годов, главным образом в Китае. Между тем известно более тридцати видов птиц из мелового периода. Кроме того, было обнаружено большое число маленьких динозавров, передвигающихся на двух ногах с характерными для древних птиц чертами. Таким образом, разрыв между динозаврами и птицами благодаря изучению окаменелостей стал значительно меньше.
Теория Острома
Наконец, спустя столетия после спорной презентации Хаксли для Геологического общества Лондона, Джон Х. Остром из Йельского университета возродил идею о том, что птицы были связаны с тероподами. Он прямо предположил, что пернатые являются их прямыми потомками. В конце 1960-х годов Остром описал скелетную анатомию хищного теропода дейнониха, который был размером с человеческого подростка и обитал на Земле около 115 миллионов лет назад, в эпоху раннего мела. В дальнейших своих опубликованных работах Остром продолжил определение анатомии птиц и группы особенностей, которые они, включая археоптерикса, делили с дейнонихом и другими тероподами, но не с другими рептилиями.
На основе этих данных он пришел к выводу, что птицы происходят непосредственно от маленьких динозавров-тероподов. После выдвижения теории Остром продолжал собирать свои доказательства происхождения пернатых от тероподов. Он использовал новый метод расшифровки отношений между организмами, применяя его в музеях естественной истории в Нью-Йорке, Париже и других местах. Этот метод получил название кладистики или филогенетической систематики. Поскольку он стал стандартом для сравнительной биологии, его использование строго обосновывало выводы Острома.

Исследования 1970-х годов
В 1970-х годах ученые продолжали интересоваться некоторыми общими уникальными особенностями археоптерикса, которые роднят его с некоторыми животными юрского периода. В их числе оказались те самые малые плотоядные динозавры, передвигающиеся на двух ногах, которые получили название тероподов. Внимательно изучив эти черты, ученые вновь выдвинули теорию, что, возможно, они были предками птиц. Ведущие палеонтологи построили эволюционные деревья, и после этого оказались еще более убеждены в своей правоте.
Что такое кладограмма
Эти деревья, иначе известные как кладограмма, являются современным золотым стандартом в анализе эволюционных отношений между животными. Сам метод получил название кладистика. Практикующие кладистики определяют эволюционную историю группы животных, изучая различные виды признаков. В процессе эволюции у какого-то животного может появится новая, генетически определенная черта, которая будет передана его потомкам. Следовательно, палеонтологи могут заключить, что две группы, однозначно разделяющие набор таких новых или полученных черт, более тесно связаны друг с другом, чем с животными, которые этих черт лишены. Узлы или точки ветвления на кладограмме указывают на появление линии с новым набором производных признаков. В результате, птицы показались ученым просто веткой на древе жизни динозавров. В работах Дж. Острома в середине 1970-х годов тероподы получили статус наиболее вероятной группы для предков птиц. Поэтому сегодня идея о том, что птицы являются пернатыми динозаврами, практически не оспаривается среди эволюционных теоретиков.
Особенности метода кладистики
Традиционные методы группировки организмов по принципу сходства и различия могут исключить вид из группы только потому, что он имеет черты, которые не были найдены у других членов. В противоположность этому, группы кладистики были основаны исключительно на определенных группах общих черт, которые особенно информативны. Этот метод начинается с дарвиновской заповеди о том, что эволюция продолжается, когда появляется новая наследственная черта в некоторых организмах, которая и передается генетически его потомкам. Предписание указывает, что две группы животных, разделяющие такие новые черты, более тесно связаны друг с другом, чем те, что разделяют только оригинальные, но не производные, переданные по наследству. Определяя общие производные черты, практики-кладистики могут определять отношения среди изучаемых организмов. Результаты таких анализов, которые обычно исследуются, могут быть представлены в виде кладограммы. Древовидная диаграмма, изображающая порядок, в котором появлялись новые характеристики и новые существа, отражают тот порядок, в котором происходила эволюция.
Причина появления перьев у птиц
По мере эволюции птиц и их удаления от динозавров, названных тероподами, множество их особенностей изменялись и совершенствовались. Если присмотреться внимательнее, становится совершенно очевидно, что набор этих характерных признаков долго развивался и служил для выполнения определенной функции. Например, появление оперения у маленького теропода было связано с необходимостью изоляции определенных участков кожи и сначала больше напоминали волосы. На первых птичьих перьях, возможно, присутствовали различные цветовые узоры. Есть версия, что они были предназначены для камуфляжа, распознавания своего вида и других функций.
Сравнение анатомических структур птиц и динозавров
Сравнительная анатомия птиц и тероподов помогла связать их друг с другом. Она также выявила некоторые способы изменения этих характеристик, поскольку динозавры стали более близки к птицам, а птицы оказались более современными. Например, в области таза лобковая кость, первоначально направленная вперед, позже смещается вертикально или назад. В передних лапах относительные пропорции костей оставались довольно постоянными у ранних птиц, но запястье изменилось. У некоторых видов кость в запястье приняла форму полумесяца. В результате ее форма способствовала возможности взлета. Широкая бумерангообразная вилочка у первых оперенных динозавров стала тоньше и образовала более глубокую дугу, когда эта особенность стала необходима для полета.
Общие признаки динозавров и птиц
Исследования Готье, проведенные в последнее время, демонстрируют, что многие функции, которые традиционно считаются принадлежащими только птицам, фактически появились до них, у их предков — тероподов. Многие из этих свойств помогли своим первоначальным обладателям выжить. Эти же черты и некоторые другие были в конечном итоге использованы или преобразованы для полета и жизни на деревьях. Птичьи характеристики тероподов, которые эволюционировали до птиц, не проявлялись сразу, а некоторые присутствовали до того, как появились сами тероподы. Они были еще у динозавров, существовавших ранее. Например, непосредственный предок тероподов был двуногими и передвигался как птица. Он был маленьким и плотоядным. У него были передние лапы, как и у ранних птиц. Кроме того, второй, а не третий палец, как у других рептилий, был длиннее. У предков динозавров голеностопный сустав стал шарнирным, а кости стопы — удлиненными. Многие из изменений в строении ног, как полагают, повлияли на увеличение длины шага и скорость бега. Это свойство в один прекрасный день поможет птичьим тероподам взлететь.
Самая большая в мире летающая птица и самая маленькая
Гигантские птицы обитали на Земле свыше 6 млн лет назад. Самой большой в мире летающей птицей был аргентавис. Он обитал в Аргентине и относился к семейству соколиных. Взрослая птица достигала 2 метров в высоту, а ее череп был примерно полметра в длину. Огромные крылья в размахе были как трехэтажный дом. Самая маленькая птица существует и сейчас. Это широко известный вид — колибри-пчелка. Длина его тела обычно не превышает 6 см. Место обитания колибри — степные районы бразильской провинции Минас-Жерайс. Самая маленькая птица питается нектаром цветов и очень быстро машет крыльями, как пчела, потому и получила такое название.
fb.ru
Эволюция птиц
Эволюция птиц
Изучение эволюции птиц сильно осложнено тем, что их достаточно хрупкие кости плохо сохранились. Вместе с тем, учёные сходятся во мнении, что предками птиц (а также крокодилов и динозавров) были мезозойские рептилии из группы текодонтов – псевдозухии. По-видимому, некоторые из этих животных постепенно приспособились к жизни на деревьях. Чешуйки, покрывавшие передние конечности, сильно разрослись и превратились в перья. Таким образом предки птиц сначала получили возможность перепрыгивать с ветки на ветку, затем планировать и, наконец, завоевали воздушную среду.

1 |
Отпечаток археоптерикса на камне |
Один из «кандидатов» на звание первой птицы – это ископаемое юрского периода – археоптерикс. Это небольшое животное размером с ворону имело перья, но наличие зубов и брюшных рёбер, длинный хвост, пальцы с когтями, отсутствие киля и воздухоносных полостей в костях сближают его с пресмыкающимися. Возможно, археоптерикс не мог поддерживать температуру своего тела на постоянном уровне. Археоптериксы жили на деревьях; по-видимому, они умели лишь планировать. Эти животные настолько отличаются от современных птиц, что их выделяют в отдельный подкласс ящерохвостых птиц, противопоставляя всем остальным птицам.
Остатки настоящих (веерохвостых) птиц были обнаружены в отложениях мелового периода. Первые из них – гесперорнисы и ихтиорнисы – ещё имели зубы, однако впоследствии зубы были утрачены, а челюсти образовали клюв.
В настоящее время появилось много свидетельств о том, что археоптерикс – тупиковая ветвь эволюции, а развитие ящерохвостых и веерохвостых птиц шло параллельно друг другу.
В заключение этого параграфа перечислим основные сходства и различия птиц с пресмыкающимися.
Сходства:
— роговые образования на поверхности кожи;
— сухая кожа, почти лишённая желёз;
— схожесть строения скелета;
— два круга кровообращения;
— органы слуха представлены внутренним и средним ухом;
— наличие клоаки;
— внутреннее оплодотворение;
— размножение яйцами и сходство в строении яйца;
— сходство эмбрионального развития.
Различия:
— передняя конечность превратилась в крыло; полёт стал основным способом передвижения птиц;
— у птиц высокая и постоянная температура тела, не зависящая от внешней среды; рептилии в холодное время года впадают в спячку;
— четырёхкамерное сердце, в котором происходит полное разделение артериальной крови и венозной;
— срастание многих костей у птиц; наличие цевки;
— наличие воздушных мешков у птиц;
— более высокий уровень развития центральной нервной системы у птиц;
— построение птицами гнёзд, высиживание яиц и выкармливание птенцов.
www.ebio.ru
|
Поиск Лекций
Мезозойским птицам и другим крылатым динозаврам
МЭТТЬЮ МАРТЫНЮК
Справочник по Мезозойским птицам и другим крылатым динозаврам
Мэттью Мартынюк
Vernon, NJ: Pan Aves
© Мэттью Мартынюк Опубликовано Pan Aves Вернон, Нью-Джерси www.panaves.com
ISBN-13: 978-0-9885965-1-1 Содержание Благодарности Вступление Кто такие «Птицы» Происхождение и эволюция птиц Предки птиц Происхождение перьев Первые птицы Разнообразие мезозойских птиц Эволюция полёта Реконструкция мезозойских птиц Перья и крылья Клювы и зубы Цвет перьев Справочник по мезозойским птицам Об этом справочнике Базальные ценагнатиформы Ценагнатоиды Базальные эуманирапторы и дейнонихозавры Микрорапторы Эудромеозавры Уненлагиины Троодонтиды Базальные авиалы Базальные энанциорнисы Эоэнанциорнитиформы Катайорнитиформы Энанциорнитиформы Базальные эуорнитины Патагоптеригиформы Прогрессивные эуорнитины и базальные килегрудые Птицы Гесперорнисы Приложение A: Исключённые виды Приложение C: Эволюционная линеевская классификация Глоссарий Список литературы
На обложке: Boluochia zhengi На титульном листе: Confuciusornis sanctus
Благодарности
Написание этой книги не было бы возможным без огромного объёма данных, справочного материала, поддержки и вдохновения. Особенно, я должен поблагодарить:
Джона Конвея, за его участие в подготовки этого проекта и за вдохновение на широкий масштаб охвата всех мезозойских птиц.
Микки Мортимер, чья база данных по тероподам, обеспечила неоценимыми сведениями по имеющимся данным.
Скотта Хартмана, Джейми Хэддена, Вилли Синконена и других художников, чьи подробные скелетные реконструкции, обеспечили замечательным справочным материалом для некоторых моих реконструкций.
Роджера Тори Петерсона, чей полевой определитель и определительная система, повлияли на содержание данного и многих других справочников по определению птиц.
Все иллюстрации созданы при помощи Adobe Photoshop CS5 и Adobe Illustrator CS5 с Wacom Graphire 3 и WACOM Bamboo Capture. Вёрстка произведена с использованием Adobe In Design CS5. Все иллюстрации и диаграммы выполнены Мэттью Мартынюк. Силуэт человека, используемый в масштабных схемах, заимствован из Пластинок «Пионера», созданных NASA и которые являются общественным достоянием.
Вступление
Кто такие «Птицы»
Доисторические птицы в общественном сознании, зачастую остаются в тени своих более крупных родственников — динозавров; Первая окаменелость Archaeopteryx и, фактически, первая из найденных ископаемых мезозойских птиц, в практически любом определении этого термина (см. ниже), была описана Германом фон Мейером в 1861 году. Состоящая из одного окаменевшего пера, найденного в известняковых карьерах Зольнхофена в Баварии, эта историческая находка в настоящее время находится в коллекции музея Гумбольдта в Берлине. Вскоре последовали находки полных скелетов, а открытые виды значительно отличались от современных птиц. Archaeopteryx lithographica («древнее крыло из литографского камня», имеется ввиду известняк, в котором сохранился литографический отпечаток) имел длинный хвост, как у рептилии, многочисленные мелкие зубы и когтистые пальцы, на фоне других примитивных черт, клюв отсутствовал. Последующие открытия мелких динозавров демонстрируют поразительное сходство с Archaeopteryx, что привело к возникновению гипотезы, что динозавры и птицы были близкими родственниками. Эта гипотеза будет укрепляться с открытием таких видов, как Deinonychus antirrhopus, который настолько был похож на Archaeopteryx, что некоторые учёные разместили их в одном биологическом «семействе». Сегодня подавляющее большинство ученых придерживаются мнения, что птицы являются прямыми потомками динозавров, и, следовательно, считаются представителями подгруппы в надотряде Dinosauria. Птицы являются такими же динозаврами, как и неуклюжие завроподы с длинной шеей: ни та, ни другая группа не считалась относящейся к динозаврам, в соответствии с изначальным определением этого термина, однако были отнесены к ним, благодаря последующим открытиям. За последние несколько десятилетий, появлялись и исчезали претенденты на звание Archaeopteryx – «первая птица». Самый печально известный из них, всё ещё сомнительный экземпляр — Protoavis texensis, который, вероятно, является химерой, состоящей из 10 останков различных рептилий, включая динозавров и пролацертиформов. Кроме того, сообщалось о множестве окаменевших следов, очень похожих на птичьи, из позднего триаса и ранней юры, на которых присутствовал отпечаток заднего пальца (первого пальца стопы, который противопоставлен остальным). Однако есть некоторые сомнения в том, действительно ли они принадлежат птицам, а не более примитивным тероподам, или даже двуногим рептилиям, связанным с крокодилами. И не исключено, что возраст некоторых следов был неверно определён. Несмотря на сомнительную природу этих претендентов, Archaeopteryx сохранил свой титул, в значительной степени, из-за характера самой классификации. Исторически так сложилось, что термин «птица» стал прочно ассоциироваться с группой Aves, названной Карлом Линнем (по праву считается отцом биологической классификации) в 1758 году. Линней назначил класс Aves (что означает «птица» на латинском языке), одним из самых многочисленных среди наземных позвоночных (тип хордовые), с высоким рангом в своей знаменитой таксономической системе (обычная иерархическая структура: царство, тип, класс, отряд, семейство, род и вид), младшим по статусу, по отношению к млекопитающим (класс Mammalia), но старшим по отношению к рептилиям (класс Reptilia). После столетий использования, система классификации Линнея начала показывать свой возраст. К 1960-м годам, биологи становятся всё более недовольными различными несовместимостями между системой Линнея и эволюционной теорией. Тем не менее, Линней разработал свою систему до того, как Дарвин продемонстрировал общее происхождение жизни. В течение следующих нескольких десятилетий, многие ученые, особенно те, которые работают в области палеонтологии позвоночных, в значительной мере будут отказываться от системы Линнея в пользу системы, основанной на кладах (группы, содержащие определённые виды, и их общего эволюционного предка, и всех других потомков этого предка, независимо от того, насколько они видоизменились по сравнению с изначальной формой). Использование клад, вместо упорядоченных классов, направлено на решение ещё одного недостатка системы Линнея: отсутствие у неё конкретных определений. Линнеевские классы были определены неясным набором признаков (диагнозов) и не имели никакого набора определений, что зачастую делает их использование невозможным. С другой стороны, клады должны иметь строгие определения и определяться не самими признаками, а эволюционным родством, выявленным при тщательном анализе этих признаков. В филогенетической системе, Mammalia — не подразумевает теплокровных позвоночных с волосяным покровом, которые кормят своих детенышей молоком, а скорее группу позвоночных, которая состоит из плацентарных, однопроходных и сумчатых, их общего предка и всех других потомков этого предка, известных в настоящее время, или которые будут обнаружены позже. Применение этого метода к птицам, оказалось спорным. Изначально, такие учёные как Жак Готье (отец филогенетической системы), определили класс Aves, по схожему принципу, как и класс Mammalia – «вершинной группой», т.е. группой, содержащей все группы современных птиц и любые близкородственные им виды, а не более отдалённые доисторические эволюционные линии. Подобное использование класса Aves, способствует возможности использования системы Линнея (он не знал ни одной доисторической птицы), но, также, и исключает такие группы ископаемых птиц, как Archaeopteryx и даже те, которые очень похожи на современных птиц, такие как Ichthyornis. Впоследствии, были предприняты попытки переопределения класса Aves, в силу исторических причин, включив в него Archaeopteryx, всё-таки он всегда считался первой птицей. Критики назвали эти попытки необоснованными: почему следует включать именно Archaeopteryx, а не его ближайших родственников (например, Microraptor), которые, в некоторых отношениях, были больше похожи на современных птиц? На момент написания этой книги, была создана организация для управления названиями клад, известная как Международное Сообщество Филогенетической Номенклатуры и, вполне вероятно, что она официально примет использование вершинной группы. Когда это использование будет официально принято, большинство мезозойских птиц окажется за пределами класса Aves. Потребуется лишь семантическое изменение, и настоящие «птицы» — окажутся ограниченной группой только современных птиц. Нептичьи представители линии, ведущей к современным птицам, такие как Ichthyornis, будут считаться «стволовыми птицами» (и, по иронии судьбы, также и родом «нептичьих динозавров», т.к. все птицы включены в кладу Dinosauria). Так кто же такие «птицы»? Поскольку этот термин не является научным и принадлежит обиходной речи, он допускает широкую интерпретацию и не требует особой научной точности. Наиболее распространённые определения птиц связаны с такими ключевыми признаками, как откладывание яиц, оперение и полёт. Если подобный подход будет принят, то Archaeopteryx по-прежнему может считаться птицей, в зависимости от его спорного стиля полёта (хотя, по крайней мере, он почти наверняка был планирующими, т.е. способным к пассивному планированию). Microraptor, вероятно, также соответствует этому определению «птиц». Конечно же, не все птицы способны летать, среди современных представителей, есть также и наземные не летающие виды. Многие эволюционные линии мезозойских птиц утратили способность к полёту, или сформировались до возникновения у них полёта. Таким образом, динозавры, которые могут быть включены по одному из первых определений, предложенному Аланом Чаригом в 1985-м году, для клады Aves, описаны в этой книге. Определение Чарига класса Aves, объединяет его по признаку наличия перьев. В то время он не уточнил, что он подразумевал под перьями, для этого, в этой книге я использую осторожную интерпретацию: перья с полностью современной анатомией, состоят из стержня (средний «ствол пера»), а опахало состоит из бородок, соединённых друг с другом бородочками (см. схему на стр. 36). Это такая же характеристика, которая, в первую очередь, использовалась для классификации Archaeopteryx как птицы, таким образом, она кажется наиболее подходящей для использования в этой работе. Подобное определение эквивалентно современной кладе Aviremigia, т.е. всем крылатым динозаврам. По своей природе, перья не фоссилизируются достаточно хорошо и часто, так что, безусловно, некоторые виды, или даже целые эволюционные линии, которые являлись «птицами» по приведённому выше определению, здесь исключены, поскольку у нас нет достаточной информации об их оперении. Не смотря на то, что некоторые примитивные целурозавры, такие как компсогнатиды и дейнодонтоиды, имели «перья», они, очевидно, полностью состояли из нитевидных и/или пуховых перьев, не имеющих опахала. Оперение орнитомимозавров тоже известно как пуховое и, не смотря на то, что у взрослых особей перья, очевидно, имели ствол, по крайней мере, те, которые располагались на их передних конечностях, хотя точная анатомия этих структур, на данный момент неизвестна. Среди манирапторов, у каждого экземпляра альваресзавров и сегнозавров сохранились перья, но и отсутствуют доказательства того, что эти перья имели опахала (возможно, только из-за плохого качества сохранности). Один любопытный вид, известный только по перьям, Praeornis sharovi, имел стержни (стволы) с бородками, но настоящие бородочки отсутствовали, имелась только странная гребнистая ткань, выстилающая каждую бородку (возможно, эволюционный предшественник бородочек), и поэтому является ближайшим другим примером, используемым здесь в определении «перо». Все эти виды не включены в данную книгу, хотя будущие открытия могут показать, что у них всё-таки были перья современного типа.
Внизу: Эволюция современных птиц из пернатых динозавров. Слева направо: Dilong paradoxus (Deinodontoidea), Nqwebasaurus thawzi (Ornithomimosauria), Haplocheirus sollers (Alvarezsauria), Yixianosaurus longimanus, Xiaotingia zhengi (Deinonychosauria), Archaeopteryx lithographica, Confuciusornis sanctus (Confuciusornithiformes), Bohaiornis guoi (Enantiornithes), Apsaravis ukhaana, Ichthyornis anceps
Происхождение птиц и эволюция
Предки птиц
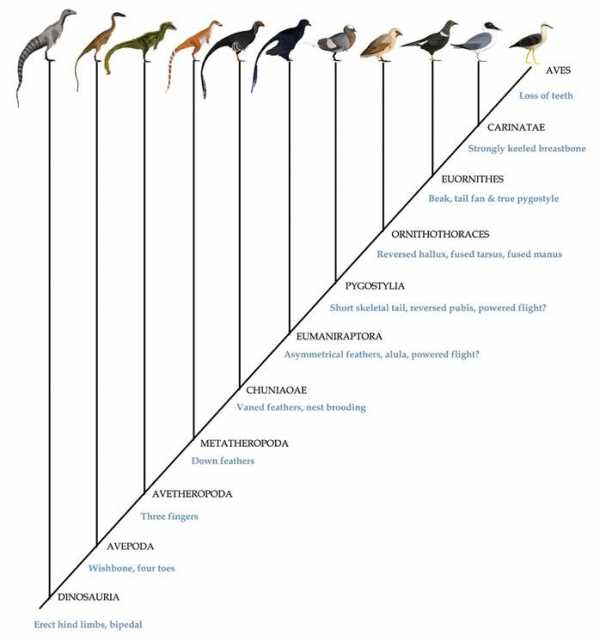
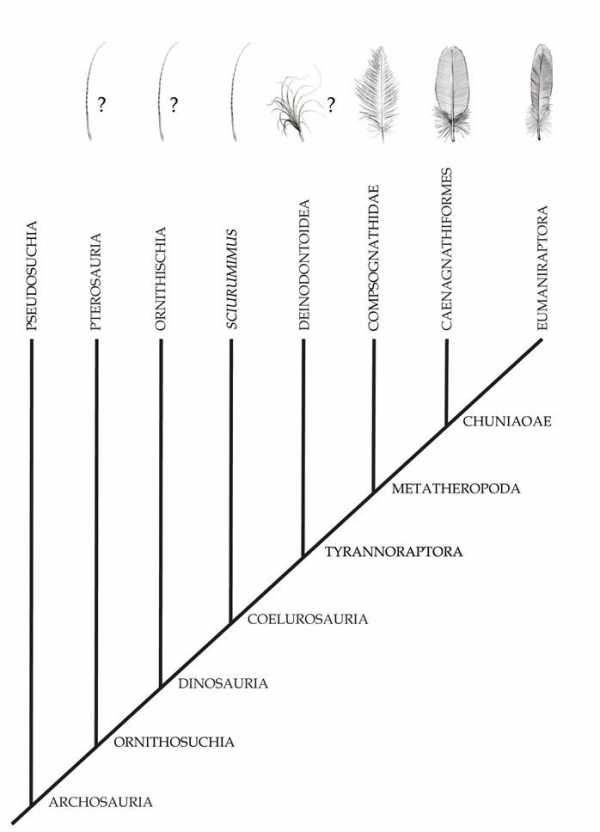
В 1970-х годах большинство ученых пришло к общему мнению, что птицы произошли непосредственно от особой группы двуногих хищных или всеядных динозавров, известной как целурозавры — Coelurosauria. И действительно, анатомическую связь переходных форм от ящероподобных ранних диапсидных рептилий до современных птиц, можно проследить по всей динозавровой линии. Хотя до сих пор существуют и другие взгляды на эту теорию, их приверженцы так и не смогли предложить альтернативные модели происхождения птиц и подтвердить свои позиции корректными и повторяемыми исследованиями. Все исследователи сходятся во мнении, что ближайшими родственниками птиц среди современных животных являются крокодилы. В большинстве случаев, эта точка зрения поддерживалась как ископаемыми свидетельствами, так и генетическими доказательствами, утверждающими, что крокодилы более тесно связаны с птицами, чем с ящерицами. Последний общий предок птиц и крокодилов (из группы архозавров) жил, вероятно, в начале триасового периода, на заре мезозойской эры. Вскоре после этого архозавры разделились на две разные ветви — крокодилов (Pseudosuchia, не совсем верно именуемые «ложными крокодилами») и птиц (Ornithosuchia, или «птичьи крокодилы»). Хотя трудно представить себе две группы животных, больше отличающиеся друг от друга с точки зрения анатомии, но предков птиц и крокодилов действительно объединяли некоторые важные общие характеристики. Например, и те, и другие обладают четырехкамерным сердцем, и вполне вероятно, что такое же было и у их общего триасового предка. Этот предок, возможно, строил гнезда из растительности, а не зарывал яйца в землю, и в некоторой степени демонстрировал заботу о потомстве – ведь даже современные крокодилы защищают свой молодняк от хищников. Однако внешне этот предок был больше крокодилом, чем птицей. Походка современных крокодилов, волочащих свое тело по земле, в значительной мере обусловлена их водной специализацией, но предок птиц и крокодилов, вероятно, тоже был ползающим, в прямом смысле слова пресмыкающимся четвероногим. Следующим крупным ответвлением в линии ведущей к птицам, стал раскол между их родоначальниками и птерозаврами – очень разнообразными мезозойскими позвоночными, включающими в себя птеродактилей. Общий предок обеих линий, входивший в группу Ornithodira, похоже, все еще перемещался полуползающей походкой и откладывал яйца с кожистой оболочкой, а не с твердой и хрупкой скорлупой, как современные птицы. Интересно, что все известные птерозавры имели плотный покров из напоминающих мех нитей – пикнофибр. Вполне возможно, что он имелся и у общего предка птиц и птерозавров, представляя одну из ранних стадий (стадию 1, описанную ниже) эволюции перьев. Однако прежде чем мы сможем проверить эту гипотезу, ученым предстоит больше узнать о коже орнитодир. У основания линии орнитодир и непосредственно в подгруппе Dinosauromorpha, предки птиц освоили ходьбу с вертикальным положением тела. Вероятно, на первых порах это был просто новый способ быстро передвигаться, подобно тому, как при быстрой ходьбе крокодил приподнимается на своих лапах, которые принимают почти вертикальное положение. Ранние орнитодиры, будучи еще четвероногими, обзавелись очень длинными задними лапами и научились бегать только с их помощью. У чуть более прогрессивных Dinosauriformes все четыре лапы оказались подведены под тело и располагались вертикально, а у появившихся вслед за тем Dinosauria задние конечности уже входили в специальные углубления таза (вертлужные впадины), удерживающие их постоянно в вертикальном положении и предотвращающие расхождение в разные стороны. Исключение составляли динозавры, специально адаптировавшие свои конечности к плаванию или лазанию. Первый динозавр, общий предок птиц и орнитиший (вот еще одно неудачное название, дословно означающее «птицетазовый»), был всеядным и двуногим, с пятью пальцами на каждой лапе (на передних имелось еще и по три когтя). Он, вероятно, жил в начале позднего триаса. Вскоре после расхождения линий птиц и птицетазовых динозавров эузауришии — общие предки птиц и динозавров-зауропод – обзавелись дыхательной системой птичьего типа, включающей в себя кроме легких еще и так называемые легочные мешки — проникающие в кости и делающие их более легкими и «воздушными» выросты легких. С появлением группы Theropoda птичьи предки полностью перешли к двуногости и бегающему образу жизни. Вскоре после этого их задние (или теперь уже нижние) лапы изменились таким образом, что стопа стала состоять только из трех основных пальцев. Маленький четвертый палец сместился далеко на лодыжку, а пятый полностью исчез. Этот маленький четвертый палец у нескольких групп теропод со временем будет использоваться для захвата добычи или усаживания на ветку. На передних лапах два лишенных когтей пальца также будут постепенно редуцироваться, и ко времени появления группы Avetheropoda пальцев останется только три. В какой-то момент, скорее всего уже после появления Avetheropoda, линия птиц обзавелась одной из самых важных своих особенностей – оперением. Первые перья были, вероятно, простыми волосоподобными нитями из бета-кератина. Один из самых ранних их примеров описан у примитивного целурозавра Sciurumimus (важно отметить, что это не совсем те «перья», что известны у птерозавров и некоторых птицетазовых динозавров). Самые ранние пуховые перья найдены у группы Tyrannoraptora, причем одновременно у обоих примитивных ее представителей – дейнодонтоидов и компсогнатид. Скорее всего, это означает, что они появились чуть раньше среднеюрского времени, когда жил общий предок двух этих групп. Образцы орнитомимозавров Dromiceiomimus, обособившихся после компсогнатид, но перед манирапторами, сохранили вероятные остатки контурных перьев, покрывавших передние конечности и превращавших их в ранние крылья. У чуть более эволюционно продвинутой группы Segnosauria также известны остатки длинных перьев на крыльях, но они имеют необычную нитевидную форму. Новые находки образцов окаменелостей хорошей сохранности помогут нам более точно восстановить природу динозавровых перьев. В средней юре у общего предка птиц и динозавровой группы Caenagnathiformes появились настоящие контурные перья современного типа. У этой же группы (Chuniaoae) впервые в летописи окаменелостей встречаются гнезда для высиживания яиц. Вероятно, это новшество связано с превосходными теплорегулирующими качествами перьев. Аэродинамические, асимметричные перья появляются у следующей группы – Eumaniraptora, причем их перья покрывают первый палец кисти, формируя
Сверху: последовательность приобретения основных птичьих признаков в группе Dinosauria
крылышко. Данные адаптации убедительно свидетельствуют о том, что к этому моменту птицы начали предпринимать первые шаги в направлении полета. Очевидно, что дальнейшие изменения в анатомии птиц также были направлены в сторону повышения аэродинамических свойств. Утрата длинного хвоста, например, привела к появлению группы Pygostylia, после чего развитие адаптаций к полету приобретает взрывной характер. В группе Ornithothoraces анатомия плеча меняется, делая возможным расправление крыльев в вертикальной плоскости, необходимое для полностью активного полета. Большой и малый палец передних конечностей тем временем слились воедино, также как и кости предплюсны. Первый палец, который произошёл от рудиментарного отростка для захвата добычи и лазанья у манирапторов, превратился в противопоставленный большой палец стопы у птицегрудых (Ornithothoraces), который позволил птицам хвататься за мелкие ветки. У Euornithes место предковой зубастой пасти занял первый настоящий птичий клюв, а перья хвоста превратились в раскрываемую веерообразную конструкцию, крепящуюся к настоящему пигостилю. Дальнейшее развитие анатомии крыла дало возможность взлетать с земли и воды, освобождая от необходимости предварительно забираться на деревья. Carinatae заметно уменьшили расположенные на крыльях когти, хотя небольшого размера коготки до сих пор встречаются у многих современных птиц. Киль, впервые развившийся в нижней части грудины еще у Ornithothoraces, у Carinatae заметно увеличился, демонстрируя переход к полету современного типа — за счет сильных грудных мышц. И, наконец, общий предок всех современных птиц лишился зубов, сформировав клюв, характерный для представителей класса Aves.
Сверху: Juravenator starki, ранний целурозавр, покрытый чешуёй и простыми (стадия I или II ) перьями. Происхождение перьев
Детали, касающиеся того, как именно впервые появились перья, всегда были спорными и до сих пор не совсем ясны. Тем не менее, гипотезы, основанные на стадиях развития перьев современных птиц (например, предложенная Ричардом Прумом; Prum 1999), похоже, в то же время довольно хорошо соответствуют летописи окаменелостей. Согласно Пруму, первые перья (стадия I эволюции перьев) были простыми стержнеподобными структурами, появившимися из фолликулов в коже, подобно тому, как появляются из кожных фолликулов чешуи и волосы. Эти простые стерженьки, часто относимые к «протоперьям», были моноволокнистыми и не имели разветвленной структуры, эквивалентной бородкам, которые формируют главную часть перьев современных птиц. Наличие стадии I в летописи окаменелостей неясно. Самые ранние вероятные примеры предоставляют позднеюрские целурозавры или мегалозавр рода Sciurumimus. Длинные, несколько уплощенные «широкие удлиненные нитевидные перья» (elongate broad filamentous feathers – EBFFs) некоторых примитивных пернатых динозавров, в том числе сегнозавра Beipiaosaurus inexpectus, могут представлять собой либо стадию I, либо новые структуры, развившиеся из более продвинутых типов перьев, или просто разрушенные перья с опахалом. Кроме того, стадию I могут представлять необычные моноволокнистые стерженьки цератопса Psittacosaurus и гетеродонтозавра Tianyulong. Если это так, то первые перья должны были появиться во время или до происхождения Dinosauria. Возможно даже, что обнаруженные у птерозавров «волоски» — пуховидные волокна — эволюционировали от стадии I перьев. Если это так, перья могут иметь истоки в начале триасового периода в основании группы Ornithosuchia, вскоре после разделения предков птиц и предков крокодилов. Стадия II эволюции перьев, появилась, когда воротничок перьевого фолликула (эквивалентный очину перьев современных птиц, который также используется в качестве стержня в перьевых ручках) дал начало нескольким нитям вместо образования одного длинного пера. Этот тип пера все еще существует в виде пуха, простейшая конструкция которого состоит из короткого очина и многочисленных длинных, мягких нитей, формирующих пучок, а не опахало. Все нити (называемые бородками) в пухе крепятся к центральному очину, а не к центральному стержню. Пух был обнаружен у компсогнатид, таких как Sinosauropteryx prima. Более примитивные тероподы, такие как Dilong и Yutyrannus, могли иметь пух II стадии, но сохранность соответствующих окаменелостей слишком плохая, чтобы бы уверенно это утверждать. По крайней мере, мы можем предположить, что пух появился в основании продвинутой группы теропод Coelurosauria или вблизи него, и присутствует у всех более продвинутых теропод, в том числе у современных птиц. Следующий этап в эволюции перьев по Пруму неясен. Основой стадия III мог быть пух с добавлением маленьких, микроскопических ветвящихся структур на их бородках (называемых бородочками). С другой стороны, на следующем этапе, возможно,
Сверху: Sinosauropteryx prima, компсогнатид, демонстрирующий пух III стадии.
бородки начали расти по спирали вдоль центральной нити, или стержня, образуя продвинутый тип пуха, известный как пуховое перо. Первым могли развиться бородочки или пуховое перо, но они могли эволюционировать и одновременно. Ископаемые доказательства не дают информации по этому вопросу, так как ни пух с бородочками, ни пуховые перья не были окончательно определены в окаменелостях не-авиремигий. Тем не менее, некоторые потенциальные ископаемые перья, классифицированных как Praeornis sharovi, кажется, демонстрируют центральный стержень и толстые бородки без различимых бородочек, вместо бородочек на бородках были гребни. Это может быть ранней формой пухового пера без бородочек или производным от него. Тем не менее, идентификация перьев Praeornis спорна, и некоторые исследователи даже предположили, что они являются совсем не перьями, а листьями саговника, хотя химические исследования, похоже, указывают, что они все же животного происхождения. Стадия III эволюции перьев была обнаружена у компсогнатида Sinosauropteryx prima. Этот вид, вероятно, имел относительно длинные стержни с длинными бородками (без бородочек) на всем протяжении. Стадия III также может присутствовать у орнитомимозавров, таких как Dromiceiommus brevitertius, у которых молодые особи, вероятно, были покрыты пухом II или III стадии, а у взрослых, вероятно, были примитивные крылоподобные передние конечности с точками прикрепления для контурных кроющих и маховых перьев, но было ли у них опахало (что делает их стадией IV) в настоящее время неизвестно (Zelenitsky & al. 2012). Стадия IV эволюции перьев представляет истинные перья современных птиц. Первичные бородки и простые вторичные бородочки пухового пера развили другой уровень ветвления в виде третичных крючочков. Они позволяют бородочкам сцепляться между собой и соединять бородки в единое опахало. Стадия IV впервые отмечена у ценагнатиформ и базальных эуманирапторов. Перья с опахалом аэродинамические и могут использоваться для полета, особенно некоторые более сложные их производные с асимметричным опахалом (когда бородки с одной стороны стержня длиннее, чем с другой). Прум называет перья с асимметричным опахалом стадией V. Стадия V, или маховые перья, впервые появляется у орнитодесмид и других эуманирапторов, в том числе Archaeopteryx. Схема на странице 22 демонстрирует приблизительный внешний вид различных этапов эволюции перьев на генеалогическом дереве архозавров. Тем не менее, следует отметить, что из этих основных структур часто развиваются новые типы перьев, а во многих случаях современный тип перьев утрачивается у нелетающих групп. Например, среди современных бескилевых (страусы, эму, киви и др.), перья с опахалом были утрачены. Маховые и рулевые перья этих птиц возвратились к стадии III, пуховым перьям с центральным стержнем, но без бородочек. Это также относится и к известным перьям гесперорнисов. У современных нелетающих птиц, таких как попугай какапо (который все же сохраняет некоторую способность к планированию), перья крыла вернулись со стадии V на стадию IV с симметричным опахалом, и напоминают перья каудиптерид или Anchiornis. Вполне возможно, что через некоторое время эти перья полностью потеряют свои бородочки, так как планирование становится все менее важной частью образа жизни птицы. Эта дегенерация маховых перьев, кажется, общая тенденция у птиц при утрате способности к полету, вероятно, потому, что уход за перьями с опахалом более дорогостоящ, чем за пухом или пуховыми перьями. Новые типы перьев, которые развились из стадий эвролюции перьев, указанных выше, включают EBFFs сегнозавров, состоящие из длинных, плоских, моноволокнистых стерженьков; ленточные перья хвоста так называемых «противоптиц» (энанциорнисов), вероятно, развившиеся путем расширения и уплощения стержня и утраты опахала на большей части длины; нитевидные перья, небольшие перья с бородками, расположенными на кончике тонкого стержня, которые могут функционировать как сенсорные органы; щетинки, которые, в противоположность нитевидным перьям, имеют бородки только у основания и голый, тонкий усовидный стержень на конце; порошковый пух, который никогда не линяет, но постоянно растет и крошится на концах бородок, создавая пудру, используемую при чистке и для защиты от намокания; а также демонстрационные перья, как у некоторых райских птиц, в которых бородки слились на большей части длины стержня, создавая цельную пластинку. Хотя большинство из них, как известно, присутствует только у современных птиц, некоторые эволюционировали более одного раза, и не исключено, что некоторые мезозойские птицы приобрели аналогичные структуры, которые ещё будут найдены в летописи окаменелостей.
Напротив: Иллюстрация Epidexipteryx hui, являющегося частью фауны Даухугоу.
Первые птицы
С момента первого открытия Archaeopteryx lithographica в 1861-м году, этот вид считался самой первой (то есть самой ранней из известных) птицей, классифицированной на основании присутствия настоящих перьев. В течение многих десятилетий он оставался не только самой ранней, но и единственной известной мезозойской окаменелостью имевшей отпечатки перьев (в раннем экземпляре Parahesperornis тоже сохранились следы перьев, но, как и следовало ожидать, более «прогрессивной» группы). К тому времени, когда в 1990-х годах было обнаружено ещё больше окаменелостей мезозойского возраста, содержащих отпечатки перьев, в кладистике было всё чтобы заменить классификацию Линнея в палеонтологии. Поэтому, когда было обнаружено больше видов с перьями и крыльями, они не рассматривались как птицы, а скорее пернатые, нептичьи динозавры. Такие виды динозавров, как Sinornithosaurus millenii и Microraptor zhaoianus несмотря на наличие оперённых крыльев и, возможно, некоторых форм полёта — не считаются птицами, поскольку они оказываются в непосредственной близости от клады, содержащей современных птиц и Archaeopteryx. В любом случае, эти окаменелости из формаций Йиксиань и Juifotang, меловых отложений Китая, позднее юрского Archaeopteryx. Были ли Archaeopteryx птицами, или нет, но они по-прежнему оставались самыми ранними животными с настоящими перьями. Открытие пернатых динозавров в китайских отложениях Даухугоу — взмутило воду. На сегодняшний день известно три вида из Даухугоу, имевшие перья: Pedopenna dau-hugouensis, Scansoriopteryx heilmanni, и Epidexipteryx hui. Даухугоу датируется более ранним возрастом, чем Йиксиань, но насколько более ранним — вопрос спорный. Средний возраст, найденный в опубликованной научной литературе, датируется поздней юрой, соответственно, являясь одинаковым, или чуть более ранним, чем у Archaeopteryx. Некоторые исследования указывают на среднеюрский возраст, что делает датировку значительно более ранней, чем у Archaeopteryx. Виды из Даухугоу уступили свои титулы первых пернатых животных в 2009-м году, когда окаменелости, содержащие отпечатки перьев были описаны из ещё более ранней китайской формации — Тяоджишан, датируемой между 161 и 155 миллионами лет назад, по крайней мере, на пять миллионов лет старше, чем Archaeopteryx. Первые из них, которые были представлены общественности — Anchiornis huxleyi и Xiaotingia zhengi, оба тесно связаны с Archaeopteryx. На подобие Pedopenna из Даухугоу и Microraptor из Jiufotang, Anchiornis обладал не только хорошо развитыми крыльями, но и крылоподобной структурой перьев на задних конечностях, весомо свидетельствуя о том, что в итоге птицы произошли от скользящих по воздуху, четырёхкрылых форм. На момент написания этой книги, Anchiornis huxleyi и Xiaotingia zhengi — являлись первыми «птицами», в широком понимании этого слова. Как отмечалось выше, многие современные исследователи ограничивают Aves как вершинную группу, и в этом смысле, самыми ранними из известных настоящих птиц в действительности были значительно более поздние, чем Archaeopteryx. Самая ранняя из известных настоящих птиц, которую с большой степенью уверенности можно отнести к таковым — Austinornis lentus из позднего мела Техаса, 85 миллионов лет назад. Однако молекулярные исследования указывают на то, что настоящие птицы произошли ближе к 130 миллионам лет назад (около 20 миллионов лет после Archaeopteryx) и, по крайней мере, одна потенциальная птица известна из того же временного промежутка во Франции: Gallornis straelani, хотя это определение далеко небесспорно.
Рекомендуемые страницы: |
|



poisk-ru.ru
Эволюция птиц — Википедия (с комментариями)
Материал из Википедии — свободной энциклопедии


 Эволюция птиц началась, согласно традиционным представлениям, в юрском периоде. Согласно этой версии, самым древним из известных видов птиц считается археоптерикс (Archaeopteryx lithographica). Птицы в этом случае рассматриваются как специализированные представители теропод, близкие к другим оперённым динозаврам из группы манирапторов, например, дромеозавридам.
Эволюция птиц началась, согласно традиционным представлениям, в юрском периоде. Согласно этой версии, самым древним из известных видов птиц считается археоптерикс (Archaeopteryx lithographica). Птицы в этом случае рассматриваются как специализированные представители теропод, близкие к другим оперённым динозаврам из группы манирапторов, например, дромеозавридам.
Вместе с тем, ряд авторов придерживается альтернативной версии, согласно которой первой достоверной находкой птиц является довольно спорное ископаемое протоавис из позднего триаса. Согласно этой версии, и птицы, и тероподы произошли от одной из более ранних групп архозавров.
В настоящее время выдвинуто несколько различных гипотез происхождения и эволюции птиц, но общепринятой филогении нет.
Классические гипотезы
«с деревьев вниз» (arboreal; trees down)
- Текодонтная гипотеза
Эту гипотезу предложил в начале XX века Герхард Хайльман (англ.)русск., основываясь на детальном обзоре биологии птиц (включая археоптерикса) и рептилий. По его мнению, наиболее близкими родственниками птиц следует считать триасовых псевдозухий (из надотряда текодонтов). Текодонтная гипотеза была общепринятой в середине XX столетия и поддерживается сегодня рядом специалистов. Однако вплоть до настоящего времени среди псевдозухий не удалось обнаружить форм, филогенетически близких к археоптериксу или другим ранним птицам.
- Гипотеза Уокера (архозавроморфная)
Уокер обосновывал в 70-х годах XX века родство археоптерикса и всех птиц с триасовыми архозаврами из подотряда сфенозухий, родственных современным крокодилам. Аргументы Уокера неоднократно критиковались, и сам автор впоследствии признал гипотезу несостоятельной.
До недавнего времени обе гипотезы постепенно теряли сторонников. Однако после открытия четырёхкрылого оперённого динозавра в Китае, предсказанного ещё Бибом в 1915 году[1] под именем «тетраптерикс» (Tertrapteryx), группа гипотез «с деревьев вниз» вновь переживает расцвет. Что любопытно, подтвердить гипотезу Биба в 1926 году пытался Хейльман[2], исследовавший ноги птенцов горлиц и яканы, однако его исследования были тогда восприняты с большим скепсисом.
Опубликованное в июле 2014 года в журнале Journal of Ornithology исследование позволило отнести Scansoriopteryx к древним птицам, потомкам архозавров. У Scansoriopteryx обнаружили характерные для птиц анатомические особенности: удлиненные передние конечности, кости запястий в форме полумесяца, лапы, приспособленные для сидения на ветках. Перья, расположенные на передних и задних конечностях свидетельствуют о том, что он мог планировать в воздухе, перелетая между деревьями, а это, в свою очередь означает что полет возник не «снизу вверх», когда наземные динозавры-тероподы начали летать, а «сверху вниз», когда архозавры нарастили перья, позволившие им короткое время держаться в воздухе. Факт отнесения Scansoriopteryx к птицам вновь ставит вопрос о том, что птицы не являются потомками динозавров.[3]
«с земли вверх» (cursorial; ground up)
- Тероподная гипотеза
Гипотеза первоначально была выдвинута Томасом Генри Хаксли в XIX веке. В 70-х годах XX века её возродил Джон Остром. Основываясь на сравнении пересмотренной им остеологии археоптерикса и тероподных динозавров (в особенности описанного им дейнониха), Остром заключил, что ближайшими родственниками археоптерикса являются целурозавры из семейства дромеозаврид.
Впоследствии выводы Острома были подтверждены кладистическими исследованиями, обнаружением экземпляров целурозавров с отпечатками перьев, сравнением микроструктуры яичной скорлупы и репродуктивной биологии целурозавров и птиц[4][5][6]. Сегодня тероподная гипотеза принимается большинством палеонтологов.
Современная филогения птиц и наиболее близких к ним групп тероподов выглядит следующим образом:
Однако после открытия четырёхкрылого оперённого динозавра в Китае основная идея гипотезы — «с земли вверх» — стала всё чаще подвергаться острой критике. При этом надо заметить, что остальные находки оперённых динозавров в Китае довольно хорошо вписываются в концепцию «с земли вверх».
Другие гипотезы
После открытия протоависа Санкаром Чаттерджи (en:Sankar Chatterjee) в 1984 году появился ряд новых гипотез:
- По мнению самого Чаттерджи, древнейшей птицей является протоавис, а не археоптерикс. При этом ближайшими родственниками птиц являются триасовые цератозавры — группа тероподных динозавров, обладавшая рядом существенных особенностей, характерных для птиц (близкое к гетероцильному строение шейных позвонков, сплав тазовых костей и др.). Эти особенности скелета цератозавров традиционно расцениваются как результат конвергенции. При этом полёт появился в результате планирования дромерозаврид с деревьев.
- Гипотеза дифилетического происхождения птиц, разрабатываемая Е. Н. Курочкиным частично основывается на изучении протоависа. Согласно этой гипотезе, веерохвостые птицы произошли от некоторой группы юрских архозавров, близким родственником которых является протоавис. Ящерохвостые птицы (например, археоптерикс и энанциорнисы) являются потомками тероподов. При этом предки птицы обитали не на настоящих деревьях, а в кустарниках, на которые они запрыгивали, используя силу ног, а спрыгивали, замедляя прыжок быстрыми махами оперённых передних конечностей. Ключевым в этой гипотезе является отсутствие стадии планирующего полёта, которая неоднократно критиковалась сторонниками гипотезы «с земли верх», согласно мнению которых планирующий полёт самодостаточен и не способен перейти в машущий. Тем не менее, согласно Курочкину, планирующий полёт всё же имел место, но относился не к настоящим птицам, а к тупиковой ветви развития, представленной археоптериксом. Другим ключевым моментом в гипотезе Курочкина является то, что в отличие от авторов классических гипотез «с деревьев вниз» он считает, что передние конечности при лазаньи играли вспомогательную роль, и не обхватывали ствол дерева, а цеплялись за мелкие соседние ветви, помогая балансировать. Основная роль при лазаньи в гипотезе Курочкина отводится сильным задним конечностям. Это является вторым ответом критикам гипотезы «с деревьев вниз», по мнению которых лазанье по деревьям с обхватом передними конечностями ствола дерева несовместимо с машущим полётом.
- Гипотеза Диала — выдвинута в 2003 году, на основе наблюдения птенцов кекликов, взбиравшихся по наклонным поверхностям (вплоть до почти вертикальных), помогая себе взмахами крыльев.
- Гипотеза Гарнера «атакующий проавис» («prouncing proavis») — выдвинута в 1999 году, отвергает как идею «с земли вверх», так и «с деревьев вниз». Согласно этой гипотезе, полёт появился в результате прыжков на добычу из засады с возвышения, а появление перьев служило повышению точности прыжка. В подтверждение гипотезы Гарнер приводит кладистическую линию Sinosauropteryx-Protarchaeopteryx-Caudipteryx-Archaeopteryx-Iberomesornis-Enantiornithes-Ornithurae, годом ранее построенную Джи.
- Гипотеза Петерса — выдвинута в 2002 году как развитие идеи Гарнера. В качестве одного из доказательств своей гипотезы Петерс раскритиковал как классическую гипотезу «с деревьев вниз», так и классическую «с земли вверх». Так, например, по его мнению, приспособление передних конечностей к лазанью противоречит приспособлению к полёту.
- Гипотеза Лакасы-Руиса (Lacasa-Ruiz, 1993) и Лопарёва (1996) — изначально выдвинута Лакасой-Руисом в 1993 году, а затем доработана в 1996 году С. А. Лопарёвым, который в качестве доказательства привёл сравнительный анализ строения таза птиц, а также онтогенез современных птиц. Согласно гипотезе, предки птиц обитали на заросшем мелководье, и перья на передних конечностях изначально служили для создания тени с целью привлечения рыбы (так охотятся некоторые болотные птицы). При спасении от хищников махи передними конечностями облегчали бег по заросшему болотистому мелководью, что постепенно перешло в использование экранного эффекта, затем в транзитный полёт (cruising flight).
- Гипотеза Савельева — выдвинута в 2006. Согласно этой гипотезе, птицы изначально обитали на мелководье, а затем научились нырять, используя передние конечности подобно оляпке и пингвину, что явилось преадаптацией к полёту.
- Гипотеза Острома (Ostrom) — выдвинута в 1976 году, и согласно ей перья на передних конечностях изначально использовались в качестве ловчих сетей для насекомых. Гипотеза неоднократно критиковалась, и в 1997 году Остром от неё отказался.
Найденные ископаемые
Эволюционное происхождение перьев можно проследить начиная с карнотавра, покрытого многочисленными бугорками[7], похожими на фолликулы. Следующий этап виден у синозавроптерикса и у Dilong paradoxus, покрытых волокнистым пухом[7]. И наконец у каудиптерикса, протархеоптерикса, синорнитозавра и микрораптора можно наблюдать настоящие перья[7].
См. также
Напишите отзыв о статье «Эволюция птиц»
Примечания
- ↑ Beebe W.H. «Zoologia» 1915.V.2 p.38-52
- ↑ Heilmann G. «The Origin of Birds» L.1926
- ↑ [lenta.ru/news/2014/07/12/dinobird/ Lenta.ru: Наука и техника: Наука: Птицы оказались «братьями» динозавров]
- ↑ Gauthier J. «Saurischian Monophyly and the origin of birds», in K. Padian: The Origin of Birds and the Evolution of Flight. 1986. Mem. California Acad. Sci 8, 1-55
- ↑ Padian K., Chiappe L. The origin and early evolution of birds. Biol. Rev., 1998, v. 73, p. 1-42
- ↑ Chiappe L. The Closest Relatives of Birds. Ornitología Neotropical, 2004, v. 15, p. 101—116
- ↑ 1 2 3 «Dinosaurs. A Concise Natural History.» David E. Fastovsky (University of Rhode Island) and and David B. Weishampel (The Johns Hopkins University), With illustrations by John Sibbick, Cambridge University Press 2009, ISBN 978-0-511-47941-0 (e-book), ISBN 978-0-521-88996-4 (в твёрдой обложке), ISBN 978-0-521-71902-5 (в мягкой обложке), часть III: «Saurischia: meat, might, and magnitude», глава 9 «Theropoda I: nature red in tooth and claw», раздел «Theropod lives and lifestyles», подраздел «The skinny on skin», Страница 199
Ссылки
- [macroevolution.narod.ru/kurzanov.htm Курзанов С. М. «Авимимиды и проблема происхождения птиц»]
- [macroevolution.narod.ru/kurochkin.htm Е. Н. Курочкин «Новые идеи о происхождении и ранней эволюции птиц»]
- [macroevolution.narod.ru/kurochkin2.htm Е. Н. Курочкин «О четырёхкрылом динозавре и происхождении птиц.»]
- [macroevolution.narod.ru/peria/peria.html Р. Прам, А. Баш. «Динозавры или птицы: кто оперился первым?» (В мире науки, № 7, 2003)]
Литература
- Курочкин Е. Н., Богданович И. А. К проблеме происхождения птиц: компромиссный и системный подходы. Известия РАН, Серия Биологическая, 2008 № 1 с.15-17, УДК 568.2(591.174)
- Chatterjee S. The Rise of Birds. — Baltimore and London, Johns Hopkins University Press. 1997
- Chatterjee S. Protoavis and the early evolution of birds. — Palaeontographica, Abt. A, Bd. 254, Lfg., 1999, 1-3, p. 1-100
- Chatterjee, S. 1987. Skull of Protoavis and Early Evolution of Birds. Journal of Vertebrate Paleontology 7(3)(Suppl.): 14A.
- Курочкин Е. Н. Параллельная эволюция тероподных динозавров и птиц. — Зоологический журнал 2006, том 85, № 3.
- Kevin Padian and Luis M. Chiappe. The Origin of Birds and Their Flight. SCIENTIFIC AMERICAN, FEBRUARY 1998
Отрывок, характеризующий Эволюция птиц
– Власть ваша! – сказал Дрон печально.– Эй, Дрон, оставь! – повторил Алпатыч, вынимая руку из за пазухи и торжественным жестом указывая ею на пол под ноги Дрона. – Я не то, что тебя насквозь, я под тобой на три аршина все насквозь вижу, – сказал он, вглядываясь в пол под ноги Дрона.
Дрон смутился, бегло взглянул на Алпатыча и опять опустил глаза.
– Ты вздор то оставь и народу скажи, чтобы собирались из домов идти в Москву и готовили подводы завтра к утру под княжнин обоз, да сам на сходку не ходи. Слышишь?
Дрон вдруг упал в ноги.
– Яков Алпатыч, уволь! Возьми от меня ключи, уволь ради Христа.
– Оставь! – сказал Алпатыч строго. – Под тобой насквозь на три аршина вижу, – повторил он, зная, что его мастерство ходить за пчелами, знание того, когда сеять овес, и то, что он двадцать лет умел угодить старому князю, давно приобрели ему славу колдуна и что способность видеть на три аршина под человеком приписывается колдунам.
Дрон встал и хотел что то сказать, но Алпатыч перебил его:
– Что вы это вздумали? А?.. Что ж вы думаете? А?
– Что мне с народом делать? – сказал Дрон. – Взбуровило совсем. Я и то им говорю…
– То то говорю, – сказал Алпатыч. – Пьют? – коротко спросил он.
– Весь взбуровился, Яков Алпатыч: другую бочку привезли.
– Так ты слушай. Я к исправнику поеду, а ты народу повести, и чтоб они это бросили, и чтоб подводы были.
– Слушаю, – отвечал Дрон.
Больше Яков Алпатыч не настаивал. Он долго управлял народом и знал, что главное средство для того, чтобы люди повиновались, состоит в том, чтобы не показывать им сомнения в том, что они могут не повиноваться. Добившись от Дрона покорного «слушаю с», Яков Алпатыч удовлетворился этим, хотя он не только сомневался, но почти был уверен в том, что подводы без помощи воинской команды не будут доставлены.
И действительно, к вечеру подводы не были собраны. На деревне у кабака была опять сходка, и на сходке положено было угнать лошадей в лес и не выдавать подвод. Ничего не говоря об этом княжне, Алпатыч велел сложить с пришедших из Лысых Гор свою собственную кладь и приготовить этих лошадей под кареты княжны, а сам поехал к начальству.
Х
После похорон отца княжна Марья заперлась в своей комнате и никого не впускала к себе. К двери подошла девушка сказать, что Алпатыч пришел спросить приказания об отъезде. (Это было еще до разговора Алпатыча с Дроном.) Княжна Марья приподнялась с дивана, на котором она лежала, и сквозь затворенную дверь проговорила, что она никуда и никогда не поедет и просит, чтобы ее оставили в покое.
Окна комнаты, в которой лежала княжна Марья, были на запад. Она лежала на диване лицом к стене и, перебирая пальцами пуговицы на кожаной подушке, видела только эту подушку, и неясные мысли ее были сосредоточены на одном: она думала о невозвратимости смерти и о той своей душевной мерзости, которой она не знала до сих пор и которая выказалась во время болезни ее отца. Она хотела, но не смела молиться, не смела в том душевном состоянии, в котором она находилась, обращаться к богу. Она долго лежала в этом положении.
Солнце зашло на другую сторону дома и косыми вечерними лучами в открытые окна осветило комнату и часть сафьянной подушки, на которую смотрела княжна Марья. Ход мыслей ее вдруг приостановился. Она бессознательно приподнялась, оправила волоса, встала и подошла к окну, невольно вдыхая в себя прохладу ясного, но ветреного вечера.
«Да, теперь тебе удобно любоваться вечером! Его уж нет, и никто тебе не помешает», – сказала она себе, и, опустившись на стул, она упала головой на подоконник.
Кто то нежным и тихим голосом назвал ее со стороны сада и поцеловал в голову. Она оглянулась. Это была m lle Bourienne, в черном платье и плерезах. Она тихо подошла к княжне Марье, со вздохом поцеловала ее и тотчас же заплакала. Княжна Марья оглянулась на нее. Все прежние столкновения с нею, ревность к ней, вспомнились княжне Марье; вспомнилось и то, как он последнее время изменился к m lle Bourienne, не мог ее видеть, и, стало быть, как несправедливы были те упреки, которые княжна Марья в душе своей делала ей. «Да и мне ли, мне ли, желавшей его смерти, осуждать кого нибудь! – подумала она.
Княжне Марье живо представилось положение m lle Bourienne, в последнее время отдаленной от ее общества, но вместе с тем зависящей от нее и живущей в чужом доме. И ей стало жалко ее. Она кротко вопросительно посмотрела на нее и протянула ей руку. M lle Bourienne тотчас заплакала, стала целовать ее руку и говорить о горе, постигшем княжну, делая себя участницей этого горя. Она говорила о том, что единственное утешение в ее горе есть то, что княжна позволила ей разделить его с нею. Она говорила, что все бывшие недоразумения должны уничтожиться перед великим горем, что она чувствует себя чистой перед всеми и что он оттуда видит ее любовь и благодарность. Княжна слушала ее, не понимая ее слов, но изредка взглядывая на нее и вслушиваясь в звуки ее голоса.
– Ваше положение вдвойне ужасно, милая княжна, – помолчав немного, сказала m lle Bourienne. – Я понимаю, что вы не могли и не можете думать о себе; но я моей любовью к вам обязана это сделать… Алпатыч был у вас? Говорил он с вами об отъезде? – спросила она.
Княжна Марья не отвечала. Она не понимала, куда и кто должен был ехать. «Разве можно было что нибудь предпринимать теперь, думать о чем нибудь? Разве не все равно? Она не отвечала.
– Вы знаете ли, chere Marie, – сказала m lle Bourienne, – знаете ли, что мы в опасности, что мы окружены французами; ехать теперь опасно. Ежели мы поедем, мы почти наверное попадем в плен, и бог знает…
Княжна Марья смотрела на свою подругу, не понимая того, что она говорила.
– Ах, ежели бы кто нибудь знал, как мне все все равно теперь, – сказала она. – Разумеется, я ни за что не желала бы уехать от него… Алпатыч мне говорил что то об отъезде… Поговорите с ним, я ничего, ничего не могу и не хочу…
– Я говорила с ним. Он надеется, что мы успеем уехать завтра; но я думаю, что теперь лучше бы было остаться здесь, – сказала m lle Bourienne. – Потому что, согласитесь, chere Marie, попасть в руки солдат или бунтующих мужиков на дороге – было бы ужасно. – M lle Bourienne достала из ридикюля объявление на нерусской необыкновенной бумаге французского генерала Рамо о том, чтобы жители не покидали своих домов, что им оказано будет должное покровительство французскими властями, и подала ее княжне.
– Я думаю, что лучше обратиться к этому генералу, – сказала m lle Bourienne, – и я уверена, что вам будет оказано должное уважение.
Княжна Марья читала бумагу, и сухие рыдания задергали ее лицо.
– Через кого вы получили это? – сказала она.
– Вероятно, узнали, что я француженка по имени, – краснея, сказала m lle Bourienne.
Княжна Марья с бумагой в руке встала от окна и с бледным лицом вышла из комнаты и пошла в бывший кабинет князя Андрея.
– Дуняша, позовите ко мне Алпатыча, Дронушку, кого нибудь, – сказала княжна Марья, – и скажите Амалье Карловне, чтобы она не входила ко мне, – прибавила она, услыхав голос m lle Bourienne. – Поскорее ехать! Ехать скорее! – говорила княжна Марья, ужасаясь мысли о том, что она могла остаться во власти французов.
«Чтобы князь Андрей знал, что она во власти французов! Чтоб она, дочь князя Николая Андреича Болконского, просила господина генерала Рамо оказать ей покровительство и пользовалась его благодеяниями! – Эта мысль приводила ее в ужас, заставляла ее содрогаться, краснеть и чувствовать еще не испытанные ею припадки злобы и гордости. Все, что только было тяжелого и, главное, оскорбительного в ее положении, живо представлялось ей. «Они, французы, поселятся в этом доме; господин генерал Рамо займет кабинет князя Андрея; будет для забавы перебирать и читать его письма и бумаги. M lle Bourienne lui fera les honneurs de Богучарово. [Мадемуазель Бурьен будет принимать его с почестями в Богучарове.] Мне дадут комнатку из милости; солдаты разорят свежую могилу отца, чтобы снять с него кресты и звезды; они мне будут рассказывать о победах над русскими, будут притворно выражать сочувствие моему горю… – думала княжна Марья не своими мыслями, но чувствуя себя обязанной думать за себя мыслями своего отца и брата. Для нее лично было все равно, где бы ни оставаться и что бы с ней ни было; но она чувствовала себя вместе с тем представительницей своего покойного отца и князя Андрея. Она невольно думала их мыслями и чувствовала их чувствами. Что бы они сказали, что бы они сделали теперь, то самое она чувствовала необходимым сделать. Она пошла в кабинет князя Андрея и, стараясь проникнуться его мыслями, обдумывала свое положение.
Требования жизни, которые она считала уничтоженными со смертью отца, вдруг с новой, еще неизвестной силой возникли перед княжной Марьей и охватили ее. Взволнованная, красная, она ходила по комнате, требуя к себе то Алпатыча, то Михаила Ивановича, то Тихона, то Дрона. Дуняша, няня и все девушки ничего не могли сказать о том, в какой мере справедливо было то, что объявила m lle Bourienne. Алпатыча не было дома: он уехал к начальству. Призванный Михаил Иваныч, архитектор, явившийся к княжне Марье с заспанными глазами, ничего не мог сказать ей. Он точно с той же улыбкой согласия, с которой он привык в продолжение пятнадцати лет отвечать, не выражая своего мнения, на обращения старого князя, отвечал на вопросы княжны Марьи, так что ничего определенного нельзя было вывести из его ответов. Призванный старый камердинер Тихон, с опавшим и осунувшимся лицом, носившим на себе отпечаток неизлечимого горя, отвечал «слушаю с» на все вопросы княжны Марьи и едва удерживался от рыданий, глядя на нее.
Наконец вошел в комнату староста Дрон и, низко поклонившись княжне, остановился у притолоки.
Княжна Марья прошлась по комнате и остановилась против него.
– Дронушка, – сказала княжна Марья, видевшая в нем несомненного друга, того самого Дронушку, который из своей ежегодной поездки на ярмарку в Вязьму привозил ей всякий раз и с улыбкой подавал свой особенный пряник. – Дронушка, теперь, после нашего несчастия, – начала она и замолчала, не в силах говорить дальше.
– Все под богом ходим, – со вздохом сказал он. Они помолчали.
– Дронушка, Алпатыч куда то уехал, мне не к кому обратиться. Правду ли мне говорят, что мне и уехать нельзя?
– Отчего же тебе не ехать, ваше сиятельство, ехать можно, – сказал Дрон.
– Мне сказали, что опасно от неприятеля. Голубчик, я ничего не могу, ничего не понимаю, со мной никого нет. Я непременно хочу ехать ночью или завтра рано утром. – Дрон молчал. Он исподлобья взглянул на княжну Марью.
– Лошадей нет, – сказал он, – я и Яков Алпатычу говорил.
wiki-org.ru
Эволюция птиц — это… Что такое Эволюция птиц?
Эволюция птиц началась, согласно традиционным представлениям, в Юрском периоде. Согласно этой версии, самым древним из известных видов птиц считается археоптерикс (Archaeopteryx lithographica). Птицы в этом случае рассматриваются как специализированные представители теропод, близкие к другим оперенным динозаврам из группы манирапторов, например дромеозавридам.
Вместе с тем, ряд авторов придерживаются альтернативной версии, согласно которой первой достоверной находкой птиц является довольно спорное ископаемое протоавис из позднего триаса. Согласно этой версии, и птицы, и тероподы произошли от одной из более ранних групп групп архозавров.
В настоящее время выдвинуто несколько различных гипотез происхождения и эволюции птиц, но общепринятой филогении нет.
Классические гипотезы
«с деревьев вниз» (arboreal; trees down)
- Текодонтная гипотеза
Эту гипотезу предложил в начале XX века Герхард Хейльман (Gerhard Heilmann), основываясь на детальном обзоре биологии птиц (включая археоптерикса) и рептилий. По его мнению, наиболее близкими родственниками птиц следует считать триасовых псевдозухий (из надотряда текодонтов). Текодонтная гипотеза была общепринятой в середине XX столетия и поддерживается сегодня рядом специалистов. Однако вплоть до настоящего времени среди псевдозухий не удалось обнаружить форм филогенетически близких к археоптериксу или другим ранним птицам.
- Гипотеза Уокера (архазавроморфная)
Уокер обосновывал в 70-х годах XX века родство археоптерикса и всех птиц с триасовыми архозаврами из подотряда сфенозухий, родственных современным крокодилам. Аргументы Уокера неоднократно критиковались, и сам автор впоследствии признал гипотезу несостоятельной.
До недавнего времени обе гипотезы постепенно теряли сторонников. Однако после открытия четырёхкрылого оперённого динозавра в Китае, предсказанного ещё Бибой в 1915 году[1] под именем «Тетраптерикс» (Tertrapteryx), группа гипотез «с деревьев вниз» вновь переживает расцвет. Что любопытно, подтвердить гипотезу Биба в 1926 году пытался Хейльман[2], исследовавший ноги птенцов горлиц и яканы, однако его исследования были тогда восприняты с большим скепсисом.
«с земли вверх» (cursorial; ground up)
- Тероподная гипотеза
Гипотеза первоначально была выдвинута Томасом Генри Хаксли в XIX веке. В 70-х годах XX века её возродил Джон Остром. Основываясь на сравнении пересмотренной им остеологии археоптерикса и тероподных динозавров (в особенности описанного им дейнониха), Остром заключил, что ближайшими родственниками археоптерикса являются целурозавры из семейства дромеозаврид.
Впоследствии выводы Острома были подтверждены кладистическими исследованиями, обнаружением экземпляров целурозавров с отпечатками перьев, сравнением микроструктуры яичной скорлупы и репродуктивной биологии целурозавров и птиц[3][4][5]. Сегодня тероподная гипотеза принимается большинством палеонтологов.
Современная филогения птиц и наиболее близких к ним групп тероподов выглядит следующим образом:
Однако после открытия четырёхкрылого оперённого динозавра в Китае, основная идея гипотезы — «с земли вверх», стала всё чаще подвергаться острой критике. При этом надо заметить, что остальные находки оперённых динозавров в Китае довольно хорошо вписываются в концепцию «с земли вверх».
Другие гипотезы
После открытия протоависа Санкаром Чаттерджи (en:Sankar Chatterjee) в 1984 году, появился ряд новых гипотез:
- По мнению самого С. Чаттерджи (Chatterjee S.), древнейшей птицей является протоавис, а не археоптерикс. При этом ближайшими родственниками птиц являются триасовые цератозавры — группа тероподных динозавров, обладавшая рядом существенных особенностей, характерных для птиц (близкое к гетероцильному строение шейных позвонков, сплав тазовых костей и др.). Эти особенности скелета цератозавров традиционно расцениваются как результат конвергенции. При этом полёт появился в результате планирования дромерозаврид с деревьев.
- Гипотеза дифилетического происхождения птиц, разрабатываемая Е. Н. Курочкиным частично основывается на изучении протоависа. Согласно этой гипотезе, веерохвостые птицы произошли от некоторой группы юрских архозавров, близким родственником которых является протоавис. Ящерохвостые птицы (например, археоптерикс и энанциорнисы) являются потомками тероподов. При этом предки птицы обитали не на настоящих деревьях, а в кустарниках, на которые они запрыгивали, используя силу ног, а спрыгивали, замедляя прыжок быстрыми махами оперённых передних конечностей. Ключевым в этой гипотезе, является отсутствие стадии планирующего полёта, которая неоднократно критиковалась сторонниками гипотезы «с земли верх», согласно мнению, которых планирующий полёт самодостаточен, и не способен перейти в машущий. Тем не менее, согласно Курочкину планирующий полёт всё же имел место, но относился не к настоящим птицам, а к тупиковой ветви развития, представленной арехоптериксом. Другим ключевым моментом в гипотезе Курочкина является то, что в отличие от авторов классических гипотез «с деревьев вниз» он считает, что передние конечности при лазании играли вспомогательную роль, и не обхватывали ствол дерева, а цеплялись за мелкие соседние ветви, помогая балансировать. Основная роль при лазании в гипотезе Курочкина отводится сильным задним конечностям. Это является вторым ответом критикам гипотезы «с деревьев вниз», по мнению которых лазание по деревьям с обхватом передними конечностями ствола дерева несовместимо с машущим полётом.
- Гипотеза Диала — выдвинута в 2003 году, на основе наблюдения птенцов кекликов, взбиравшихся по наклонным поверхностям (вплоть до почти вертикальных), помогая себе взмахами крыльев.
- Гипотеза Гарнера «атакующий проавис» («prouncing proavis») — выдвинута в 1999 году, отвергает как идею «с земли вверх», так и «с деревьев вниз». Согласно этой гипотезе, полёт появился в результате прыжков на добычу из засады с возвышения, а появление перьев служило повышению точности прыжка. В подтверждение гипотезы Гарнер приводит кладистическую линию Sinosauropteryx-Protarchaeopteryx-Caudipteryx-Archaeopteryx-Iberomesornis-Enantiornithes-Ornithurae, годом ранее построенную Джи.
- Гипотеза Петерса — выдвинута в 2002 году, как развитие идеи Гарнера. В качестве одного из доказательств своей гипотезы Петерс раскритиковал как классическую гипотезу «с деревьев вниз», так и классическую «с земли вверх». Так, например, по его мнению приспособление передних конечностей к лазанию противоречит приспособлению к полёту.
- Гипотеза Лакаса-Рюза (Lacasa-Ruiz, 1993) и Лопарёва (1996) — изначально выдвинута Лакасом-Рюзм в 1993 году, а затем доработана в 1996 году Лопарёвым, который в качестве доказательства привёл сравнительный анализ строения таза птиц, а также онтогенез современных птиц. Согласно гипотезе, предки птиц обитали на заросшем мелководье, и перья на передних конечностях изначально служили для создания тени с целью привлечения рыбы (так охотятся некоторые болотные птицы). При спасении от хищников, махи передними конечностями, облегчали бег по заросшему болотистому мелководью, что постепенно перешло в использование экранного эффекта, затем в транзитный полёт (cruising flight).
- Гипотеза Савельева — выдвинута в 2006. Согласно этой гипотезе птицы изначально обитали на мелководье, а затем научились нырять, используя передние конечности подобно оляпке и пингвину, что явилось преадаптацией к полёту.
- Гипотеза Острома (Ostrom) — выдвинута в 1976 году, и согласно ей перья на передних конечностях изначально использовались в качестве ловчих сетей для насекомых. Гипотеза неоднократно критиковалась, и в 1997 году Остром от неё отказался.
Найденные ископаемые
Эволюционное происхождение перьев можно проследить начиная с Carnotaurus покрытого многочисленными пупыршками[6], похожими на фолликулы. Следующий этап виден у Sinosauropteryx и Dilong покрытых волокнистым пухом[6]. И наконец у Caudipteryx, Protarchaeopteryx, Sinornithosaurus, и Microraptorа можно наблюдать настоящие перья[6].
Примечания
- ↑ Beebe W.H. «Zoologia» 1915.V.2 p.38-52
- ↑ Heilmann G. «The Origin of Birds» L.1926
- ↑ Gauthier J. «Saurischian Monophyly and the origin of birds», in K. Padian: The Origin of Birds and the Evolution of Flight. 1986. Mem. California Acad. Sci 8, 1-55
- ↑ Padian K., Chiappe L. The origin and early evolution of birds. Biol. Rev., 1998, v. 73, p. 1-42
- ↑ Chiappe L. The Closest Relatives of Birds. Ornitología Neotropical, 2004, v. 15, p. 101—116
- ↑ 1 2 3 «Dinosaurs. A Concise Natural History.» David E. Fastovsky (University of Rhode Island) and and David B. Weishampel (The Johns Hopkins University), With illustrations by John Sibbick, Cambridge University Press 2009, ISBN 978-0-511-47941-0 (e-book), ISBN 978-0-521-88996-4 (в твёрдой обложке), ISBN 978-0-521-71902-5 (в мягкой обложке), часть III: «Saurischia: meat, might, and magnitude», глава 9 «Theropoda I: nature red in tooth and claw», раздел «Theropod lives and lifestyles», подраздел «The skinny on skin», Страница 199
Ссылки
Дополнительная Литература
- Курочкин Е. Н., Богданович И. А. К проблеме происхождения птиц: компромиссный и системный подходы. Известия РАН, Серия Биологическая, 2008 № 1 с.15-17, УДК 568.2(591.174)
- Chatterjee S. The Rise of Birds. — Baltimore and London, Johns Hopkins University Press. 1997
- Chatterjee S. Protoavis and the early evolution of birds. — Palaeontographica, Abt. A, Bd. 254, Lfg., 1999, 1-3, p. 1-100
- Chatterjee, S. 1987. Skull of Protoavis and Early Evolution of Birds. Journal of Vertebrate Paleontology 7(3)(Suppl.): 14A.
- Курочкин Е. Н. Параллельная эволюция тероподных динозавров и птиц. — Зоологический журнал 2006, том 85, № 3.
- Kevin Padian and Luis M. Chiappe. The Origin of Birds and Their Flight. SCIENTIFIC AMERICAN, FEBRUARY 1998
dic.academic.ru
|
ЭВОЛЮЦИЯ ПТИЦ? Птицы обладают уникальными чертами строения, такими, как перьевой покров и особое устройство легких. Большая часть этих животных способна к полету. Эволюционисты полагают, что птицы произошли от рептилий -возможно, от динозавров. «Учение об эволюции и природа науки» даже приводит в качестве доказательства эволюции наличие якобы промежуточной формы между динозавром и птицей. В этой главе мы критически рассмотрим эту «промежуточную форму» и другие аргументы в пользу эволюции птиц. Кроме того, мы приведем подробные сведения о некоторых уникальных особенностях строения птичьего пера. АРХЕОПТЕРИКС «Учение об эволюции…» включает в себя несколько вымышленных «диалогов» между преподавателями. В одном из них, на странице 8, приводится такой обмен мнениями: Карен: У меня в университете один студент сказал, что в летописи окаменелостей есть большие пробелы. Ты что-нибудь знаешь об этом? Дуг: Ну, как же! Археоптерикс! Ископаемое, у которого перья птичьи, а скелет — как у небольшого динозавра. Это — одно из «недостающих звеньев», которое наконец-то нашлось. На той же странице помещена и фотография отпечатка археоптерикса с комментарием: Подобная иллюстрация, изображающая ископаемого археоптерикса, помещена в «Учении об эволюции…» «В 1861 году была открыта птица, которая жила 150 миллионов лет назад и имела многие черты рептилий. Этот факт стал подтверждением гипотезы об эволюции, выдвинутой Чарльзом Дарвином двумя годами раньше в «Происхождении видов»» Однако Алан Федуччиа (Alan Feduccia), орнитолог мирового уровня, работающий в университете Северной Каролины в Чейпел-Хилле, сам будучи эволюционистом, возражает против утверждений, подобных реплике «Дуга»: «Палеонтологи попытались превратить археоптерикса в наземного пернатого динозавра Но это не так. Это птица, самая настоящая птица. И никакой «палеобред» не изменит этого».059 Достоверная художественная реконструкция археоптерикса вполне соответствует его птичьим признакам060 Археоптерикс обладал полностью сформированными маховыми перьями (в том числе асимметричными бородками и особыми бороздками на вентральной стороне, усиливающими перо, как и у современных летающих птиц), классическими эллиптическими крыльями, как лесные птицы наших дней, и вилочкой, к которой прикреплялись мышцы, опускающие крыло.061 Его мозг был в точности таким же, как у летающих птиц, с развитым мозжечком и зрительной корой Тот факт, что у археоптерикса были зубы, не имеет никакого отношения к его статусу «переходной формы» — у многих вымерших птиц были зубы, а у многих современных рептилий их нет. Кроме того, археоптериксу, как и другим птицам, была свойственна подвижность и верхней, и нижней челюсти. У большинства позвоночных, в том числе у рептилий подвижна только нижняя челюсть 062 ПЕРНАТЫЕ ДИНОЗАВРЫ? За последние несколько лет в средствах массовой информации не раз появлялись сообщения о так называемых «пернатых динозаврах», где делались попытки доказать, что птицы произошли от динозавров. Эти мнимые предки птиц — тероподы, группа плотоядных динозавров, в которую входит тиранозавр. Следует помнить, что СМИ обычно преподносят «доказательства» эволюции как сенсацию, а вот опровержения этих «доказательств» проходят незамеченными, даже если авторы опровержений — эволюционисты. Так, в 1996 году первые страницы газет пестрели заголовками типа: «Найдено пернатое ископаемое существо — значит, некоторые динозавры эволюционировали в птиц!»063 Речь шла об окаменелости, получившей название Sinosauropteryxprima.064Креационисты в своих публикациях призывали читателей относиться к таким сообщениям со здоровым скептицизмом.065 И их подозрения подтвердились: четверо известных палеонтологов, в том числе Джон Остром (John Ostrom) из Йельского университета, позже выяснили, что «перья» на самом деле оказались параллельными рядами волокон066 — вероятно, коллагеновых. Следующей «переходной формой от динозавров к птицам» был объявлен Mononykus, названный «нелетающей птицей».067 На обложке журнала «Time» это существо даже было изображено с перьями, хотя на нем не было обнаружено и следа перьев.068 Последующие исследования показали, что «Mononykus явно не был птицей… очевидно, он был бегающим и роющим тероподом».069 В июне 1998 года многие агентства новостей сообщили о двух ископаемых, найденных в Северном Китае и объявленных пернатыми тероподами (плотоядными динозаврами). Утверждалось, что эти ископаемые, Protarchaeopteryx robusta и Caudipteryx zoui, — «прямые предки первых птиц».070 Две последние находки были «датированы» как жившие 120-136 миллионов лет назад, в то время как «возраст» археоптерикса, настоящей птицы, — 140 — 150 миллионов лет; выходит, что «предки птиц» куда моложе своих потомков! Но Федуччиа, как и его коллегу Лэрри Мартина (Larry Martin) из университета штата Канзас, это не убеждает. Мартин пишет: «Следует иметь в виду, что для авторов подобных работ курица — это тоже пернатый динозавр!»071 Федуччиа и Мартин полагают, что Protarchaeopteryx и Caudipteryx более походят на нелетающих птиц, таких, как страус. Во-первых, зубы у них, как у древних птиц, а во-вторых, у них нет длинного хвоста, свойственного тероподам. У Caudipteryx даже имелись гастролиты — камешки в желудке, как у современных растительноядных птиц; у тероподов же таких камешков не было.072 «Теория» происхождения птиц от динозавров вообще полна проблем. Федуччиа отмечает: «С точки зрения биофизики, у таких крупных двуногих существ с укороченными передними конечностями и тяжелыми хвостами-балансирами не может развиться умение летать»; подобная анатомия максимально неудобна для полета.073 Кроме того, из строения передних конечностей динозавров неопровержимо следует, что они не могли быть предками птиц. Группа ученых под руководством Федуччиа исследовала под микроскопом эмбрионы птиц и опубликовала результаты в журнале «Science».074Вот что они доложили: Новые исследования показывают, что у птиц в эмбриональном состоянии нет большого пальца, который имелся у динозавров; следовательно, эти виды «почти невозможно» назвать близкородственными.075 ОТ ПАРЕНИЯ — К ПОЛЕТУ? Федуччиа и Мартин по ряду веских причин отвергли идею происхождения птиц от динозавров. Но, не желая отказываться от теории эволюции, они предложили другую гипотезу — о происхождении птиц от рептилий, называемых крокодиломорфы. По их предположению, эти маленькие крокодилоподобные пресмыкающиеся обитали на деревьях и «вначале перепрыгивали с ветку на ветку, а затем перешли к планированию».076 Однако планирование — не промежуточная стадия между передвижением по земле и полетом. Животные, которым свойственен планирующий полет, либо имеют крылья, причем намного длиннее, чем у настоящих летунов (сравним размах крыла планера и аэроплана, или вспомним размах крыла альбатроса, который большую часть времени парит), либо обладают широкой перепонкой, сильно отличающейся от крыла (вспомним дельтаплан или белку-летягу). Для машущего полета необходимы сложные мышечные движения, что, в свою очередь, требует развитого мозга, где была бы заложена программа для таких движений. В итоге, для всего этого необходима новая генетическая информация, которой нет у нелетающих животных. Еще одна проблема заключается в следующем: «До сих пор не было найдено ни их гипотетического предка, ни промежуточных форм, связывающих их с ископаемыми птицами. И хотя верно говорят, что кладистический анализ [сравнение комплекса признаков] действенен ровно настолько, насколько верны данные, лежащие в его основе, ни одно кладистическое исследование еще не выявило предка, не принадлежащего к тероподам». Короче говоря, Федуччиа и Мартин не оставили камня на камне от той идеи, что птицы эволюционировали от бегающих динозавров, «оторвавшись от земли». Но защитники гипотезы происхождения птиц от динозавров приводят столь же убедительные контраргументы против теории Федуччиа и Мартина о том, что птицы «спустились с деревьев». Факты свидетельствуют, что и те, и другие правы, — птицы не эволюционировали ни от бегавших по земле динозавров, ни от крокодильчиков, обитавших на деревьях. Птицы вообще ни от кого не эволюционировали! И это полностью согласуется с библейским повествованием о сотворении различных родов птиц в День Пятый (Быт. 1:20 — 23). РАЗЛИЧИЯ МЕЖДУ РЕПТИЛИЯМИ И ПТИЦАМИ Все эволюционисты считают, что птицы произошли от той или иной группы рептилий, хотя и не могут прийти к согласию относительно того, от какой именно. Но рептилии и птицы сильно отличаются друг от друга по многим показателям. У летающих птиц обтекаемая форма тела, и в полете центр тяжести соответствует центру тела. Кости у них полые, заполненные воздухом — это снижает их массу, а воздух участвует в дыхании. У птиц сильные летательные мышцы с особыми длинными сухожилиями, которые проходят через отверстия в плечевых костях — таким образом, создается подобие блока. Кроме того, птицам свойственно очень острое зрение. Но особенно удивительно у птиц устроены перья и легкие. ПЕРЬЯ Федуччиа называет перья «почти идеальными приспособлениями для полета». И в самом деле, перья — легкие, но мощные образования, их форма соответствует требованиям аэродинамики, они имеют сложную структуру, образуемую бородками и крючочками. Эта структура придает перьям водонепроницаемость, а с помощью клюва птица без труда укладывает их в аэродинамически выгодную форму.077 Рассмотрим удивительное устройство бородочек пера (на фото слева), снабженных крохотными крючочками и бороздками (увеличение — в 20 000 раз).078 Эволюционист и атеист Ричард Докинс в книге, рекомендуемой в «Учении об эволюции…», с легкостью заявляет: «Перья — видоизмененные чешуи рептилий».079И это мнение широко распространено среди эволюционистов. Но чешуи — просто складчатые производные кожи; перья же — сложные структуры, включающие в себя бородки, бородочки и крючочки. Кроме того, перья и развиваются совершенно иначе — наподобие волос, из фолликул, расположенных под кожей. Обратите внимание на различие между микроструктурами пера (вверху) и чешуи (внизу). Увеличение в 80 раз. Фото любезно предоставлено доктором Дэвидом Ментоном (David Menton). В главе 2 мы показали, что каждый орган и каждая структура на генетическом уровне должны быть представлены информацией, записанной химическими «буквами» на длинной молекуле ДНК. Очевидно, что информация, необходимая для кодирования устройства пера, существенно отличается от информации, необходимой для кодирования устройства чешуи. Для эволюции чешуи в перья необходимо, чтобы в ДНК птиц появилось значительное количество генетической информации, отсутствующей в ДНК рептилий — предполагаемого предка птиц. Как обычно, естественный отбор не благоприятствует гипотетическим переходным формам. Многие эволюционисты утверждают, что перья развились у динозавров для теплоизоляции, а затем эволюционировали таким образом, что стали пригодны для полета. Но, как и прочие домыслы, эта гипотеза не объясняет, как возникла новая генетическая информация, впоследствии подвергшаяся естественному отбору. Еще одна проблема заключается в том, что отбор для теплоизоляции весьма отличен от отбора для полета. У птиц, утративших способность летать, перья тоже утратили значительную часть свойств и уподобились шерсти. У нелетающих птиц мутации, ухудшающие аэродинамическую структуру пера, не приводят к таким недостаткам, к каким привели бы у летающих. Следовательно, естественный отбор не станет уничтожать эти ухудшения, а, напротив, может даже отобрать их. Утрата способности летать и ряда свойств пера — потеря информации, а значит, не имеет никакого отношения к эволюции, которая предполагает увеличение информации. Все это говорит о том, что для теплоизоляции вполне пригодны и структуры, подобные волосу млекопитающих.080 В то же время, если бы перья выполняли исключительно эту функцию, то естественный отбор препятствовал бы появлению пера, предназначенного для полета. Об этом свидетельствуют и волосоподобные перья. Пуховые перья тоже прекрасно служат теплоизоляции и свойственны нелетающим птицам. Они являются пуховыми потому, что у них, в отличие от перьев, предназначенных для полета, отсутствуют крючочки. Опять же, естественный отбор должен был воспрепятствовать эволюции теплоизолирующих перьев в перья с аэродинамическими свойствами. И наконец, белки пера (Ф-кератины) по биохимическому строению отличаются от белков кожи и чешуи (А-кератинов). Вот какой вывод сделал один исследователь: На морфологическом уровне перья традиционно считают гомологичными чешуям рептилий. Однако перья отличаются от них по морфогенезу [развитию формы], генетической структуре, форме и последовательности белков и структуре волокон.081 ЛЕГКИЕ ПТИЦ Для превращения легкого рептилии в легкое птицы потребовались бы кардинальные изменения. В легких рептилий воздух втягивается в крошечные ячейки (альвеолы), в которых кровь обогащается кислородом, высвобождая углекислый газ. Затем отработанный воздух выдыхается. У птиц же имеется сложная система воздушных мешков, включающая даже полые кости. Эта система поддерживает движение воздуха в одном направлении по особым трубочкам (парабронхам) в легкие, а по кровеносным сосудам легких кровь движется в противоположном направлении, чтобы обогатиться кислородом,082 — прекрасное техническое решение!083 Как могли легкие рептилий, действующие по принципу мехов, постепенно превратиться в легкие птиц? Гипотетические «промежуточные стадии» означают, что легкие какое-то время не могли функционировать нормально — выходит, бедное животное не могло дышать! Следовательно, естественный отбор должен был отобрать и закрепить уже существующее положение вещей, отбраковав негодные «промежуточные формы». Даже предположив теоретически, что ряд промежуточных форм все-таки существовал, как мы сможем объяснить направление изменений, производимых естественным отбором? Например, летучие мыши прекрасно себя чувствуют с легкими, устроенными по принципу мехов; некоторые из них даже охотятся на высоте более трех километров. Птичьи легкие, при всей их высокой эффективности, получают особенное преимущество на чрезвычайно больших высотах, где концентрация кислорода низка. Таким образом, эволюция легкого рептилии в легкое птицы не дала бы никаких избирательных преимуществ. Вероятно, нам не стоит удивляться тому обстоятельству, что Алан Федуччиа в своей главной работе об эволюции птиц даже не коснулся этой проблемы.084 Недавние исследования структуры легкого Sinosauropteryx показали, что «его подобные мехам легкие никак не могли эволюционировать в высокоэффективные легкие современных птиц».085 Любопытно, что некоторые сторонники эволюции птиц от динозавров пытаются умалить значимость свидетельств против своей теории, утверждая: «Защитники этого аргумента не в состоянии предъявить нам животное, от легких которого могли бы произойти легкие птиц — сложные по строению и не имеющие аналогов в животном мире».086 Разумеется, с их точки зрения птичьи легкие непременно должны были произойти от легких какого-нибудь другого животного — ведь этого требует вера в эволюцию.
059 Cited in V Morell, »Archaeopteryx Early bird catches a can of worms,» Science 259(5096) 764 — 65, February 5, 1993 060 Рисунок Стива Кардно, 1994, любезно предоставлен художником 061 A Feduccia, «Evidence from claw geometry indicating arboreal habits of ‘Archaeopteryx,» Science 259(5096) 764 — 65, February 5, 1993 062 D Menton and С Wieland, «Bird evolution flies out the window,» Creation 16(4) 16-19, September-November 1994 063 The Examiner, Launceston, Tasmania, October 19,1996. 064 Ann Gibbons, «New feathered fossil brings dinosaurs and birds closer,» Science 274:720 -721, 1996. 065 J. D. Sarfati, «Kentucky Fried Dinosaur?» Creation 19(2):6, March — May 1997 066 New Sc;entist, 154(2077): 13, April 12, 1997; Creation 19(3):6, June-August 1997. 067 A. Perle et al., «Flightless bird from the Cretaceous of Mongolia,» Nature 362:623 — 626, 1993; отметим исправление названия Mononykus в то время как по выбору Perle et al. был уже принят вариант Mononvchus, Nature 363:188, 1993 068 Time (Australia), April 26, 1993 069 D. P. Prothero and R. M. Schroch, editors, Major features of vertebrate evolution, «On the Origin of Birds and of Avian Flight,» by J. H. Ostrom (Knoxville, TN: University of Tennessee Press, 1994), pp. 160-177. 070 Р. Qiang, P. J. Currie, M. A. Norell, and Ji Shu-An, «Two feathered dinosaurs from Northeastern China,» Nature 393(6687):753 — 761, June 25, 1998. Perspective by K. Padian, same issue, pp. 729 — 730. 071 CNN website . cited June 24, 1998. 072 Washington Post, June 25, 1998. 073 Ann Gibbons «New feathered fossil brings dinosaurs and birds closer,» Science 274:720 -721, 1996. 074 А. С. Burke and A. Feduccm, «Developmental patterns and the identification of homologies in the avian hand,» Science 278(5338):666 — 8, October 24,1997, with a perspective by R. Hinchliffe, «The forward march of the bird-dinosaurs halted?» pp. 596 — 597, J. D. Sartati, «Dmo-Bird evolution falls flat,» Creation 20(2):41, March 1998. 075 The Cincinnati Enquirer, October 25, 1997 076 P. Shipman, «Birds do it… did dinosaurs?» New Scientist 153(2067):26 -31, February 1, 1997, p. 28 077 A. Feduccia, The Origin and Evolution of Birds (New Haven, CT. Yale University Press, 1996), p 130 078 Фото любезно предоставлено доктором Дэвидом Ментоном. 079 R. Dawkms, Climbing Mount Improbable (Harmondsworth, Middlesex, England: Penguin Books, 1996), p. 113. 080 A. Feduccia, The Origin and Evolution of Birds (New Heven,CT: Yale University Press, 1996), p. 130 081 А Н. Brush, «On the origin of feathers,» Journal oj Evolutionary Biology 9:131 — 142, 1996. 082 M. Denton, Evolution, A Theory in Crisis (Bethesda, MD: Adler & Adler, 1985), pp. 199 — 213; K. Schmidt-Nielsen, «How Birds Breathe,» Scientific American, December 1971, pp. 72-79. 083 Инженеры успешно используют этот принцип, называемый принципом противотока и широко распространенный в живой природе. См. P. F. Scholander, «The wonderful net,» Scientific American, April 1957, pp. 96 — 107. 084 «Blown away by design: Michael Denton and birds’ lungs,» Creation 21(4):14 — 15, September -November 1999. 085 A. Feduccia, ссылка 20. Тем не менее, эта книга показывает, как много прорех в теории «от динозавров — к птицам». 086 К. Padian and L. M. Chiappe, «The origin of birds and their flight,» Scientific American, 278(2), 38-47, February 1998, p. 43.
|
svobd.ru
