
1. Состав нуклеиновых кислот
ПЛАН
Состав нуклеиновых кислот
Состав ДНК
Состав РНК
Макромолекулярная структура ДНК
Выделение дезоксирибонуклеиновых кислот
Выделение рибонуклеиновых кислот
Природа межнуклеотидных связей
Межнуклеотидная связь в ДНК
Межнуклеотидная связь в РНК
Значение нуклеиновых кислот
Список литературы
Нуклеиновые кислоты — это биополимеры, макромолекулы которых состоят из многократно повторяющихся звеньев — нуклеотидов. Поэтому их называют также полинуклеотидами. Важнейшей характеристикой нуклеиновых кислот является их нуклеотидный состав. В состав нуклеотида — структурного звена нуклеиновых кислот — входят три составные части:
азотистое основание — пиримидиновое или пуриновое. В нуклеиновых кислотах содержатся основания 4-х разных видов: два из них относятся к классу пуринов и два – к классу пиримидинов. Азот, содержащийся в кольцах, придает молекулам основные свойства.
моносахарид — рибоза или 2-дезоксирибоза. Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т.е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два вида нуклеиновых кислот – рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дизоксирибозу.
остаток фосфорной кислоты. Нуклеиновые кислоты являются кислотами потому, что в их молекулах содержится фосфорная кислота.
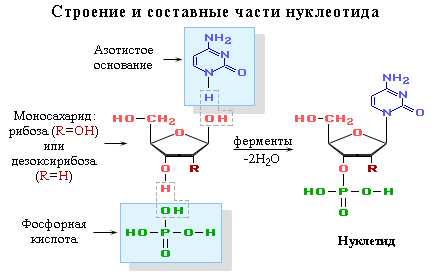
Нуклеотид — фосфорный эфир нуклеозида. В состав нуклеозида входят два компонента: моносахарид (рибоза или дезоксирибоза) и азотистое основание.
В конце 40-х — начале 50-х годов, когда появились такие методы исследования, как хроматография на бумаге и УФ-спектроскопия, были проведены многочисленные исследования нуклеотидного состава НК (Чаргафф, А. Н. Белозерский). Полученные данные позволили решительно отбросить старые представления о нуклеиновых кислотах, как о полимерах, содержащих повторяющиеся тетрануклеотидные последовательности (так называемая тстрануклеотидная теория строения ПК. господствовавшая в 30—40-е годы), и подготовили почву для создания современных представлений не только о первичной структуре ДНК и РНК, но и об их макромолекулярной структуре и функциях.
Поскольку в НК число нуклеотидов каждого вида равно числу соответствующих оснований, для установления нуклеотидного состава данной НК достаточно определить количественное соотношение оснований. Для этой цели из гидролизатов с помощью хроматографии на бумаге или электрофореза (когда в результате гидролиза получают нуклеотиды) выделяют индивидуальные соединения. Каждое основание независимо от того, связано оно с углеводным фрагментом или нет, обладает характерным максимумом поглощения в УФ, интенсивность которого зависит от концентрации. По этой причине, исходя из УФ-спектров выделенных соединений, можно определить количественное соотношение оснований, а следовательно, и нуклеотидный состав исходной НК.
При количественном определении минорных нуклеотидов, особенно таких неустойчивых, как дигидроуридиловая кислота, пользуются ферментативными методами гидролиза (ФДЭ змеиного яда и селезенки).
Использование описанных выше аналитических приемов показало, что ПК различного происхождения состоят за редким исключением из четырех основных нуклеотидов и что содержание минорных нуклеотидов может меняться в значительных пределах.
Как будет показано далее, при изучении нуклеотидного состава ДНК были получены данные, которые помогли установить ее пространственную структуру.
studfiles.net
Состав нуклеиновых кислот
Нуклеиновые кислоты — это биополимеры, макромолекулы которых состоят из многократно повторяющихся звеньев — нуклеотидов. Поэтому их называют также полинуклеотидами. Важнейшей характеристикой нуклеиновых кислот является их нуклеотидный состав. В состав нуклеотида — структурного звена нуклеиновых кислот — входят три составные части:
· азотистое основание — пиримидиновое или пуриновое. В нуклеиновых кислотах содержатся основания 4-х разных видов: два из них относятся к классу пуринов и два – к классу пиримидинов. Азот, содержащийся в кольцах, придает молекулам основные свойства.
· моносахарид — рибоза или 2-дезоксирибоза. Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т.е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два вида нуклеиновых кислот – рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дизоксирибозу.
· остаток фосфорной кислоты. Нуклеиновые кислоты являются кислотами потому, что в их молекулах содержится фосфорная кислота.
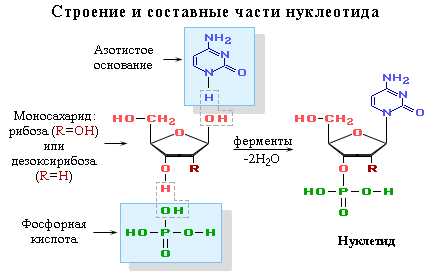 |
Нуклеотид — фосфорный эфир нуклеозида. В состав нуклеозида входят два компонента: моносахарид (рибоза или дезоксирибоза) и азотистое основание.
В конце 40-х — начале 50-х годов, когда появились такие методы исследования, как хроматография на бумаге и УФ-спектроскопия, были проведены многочисленные исследования нуклеотидного состава НК (Чаргафф, А. Н. Белозерский). Полученные данные позволили решительно отбросить старые представления о нуклеиновых кислотах, как о полимерах, содержащих повторяющиеся тетрануклеотидные последовательности (так называемая тстрануклеотидная теория строения ПК. господствовавшая в 30—40-е годы), и подготовили почву для создания современных представлений не только о первичной структуре ДНК и РНК, но и об их макромолекулярной структуре и функциях.
Метод определения состава ПК основан на анализе гндролизатов, образующихся при их ферментативном или химическом расщеплении. Обычно используются три способа химического расщепления НК. Кислотный гидролиз в жестких условиях (70%-ная хлорная кислота, 100°С, 1 ч или 100%-ная муравьиная кислота, 175 °C, 2 ч), применяемый для анализа как ДНК, так и РНК, приводит к разрыву всех N-гликозидных связей и образованию смеси пуриновых и пиримидиновых оснований. При исследовании РНК могут использоваться как мягкий кислотный гидролиз (1 н. соляная кислота, lOO°C, 1 ч), в результате которого образуются пуриновые основания и пирамидиповые нуклеозид-2′(3′)-фосфаты, так и щелочной гидролиз (0,3 н. едкий кали, 37 °С, 20 ч), дающий смесь нуклеозид -2′ (3′) -фосфатов.
Поскольку в НК число нуклеотидов каждого вида равно числу соответствующих оснований, для установления нуклеотидного состава данной НК достаточно определить количественное соотношение оснований. Для этой цели из гидролизатов с помощью хроматографии на бумаге или электрофореза (когда в результате гидролиза получают нуклеотиды) выделяют индивидуальные соединения. Каждое основание независимо от того, связано оно с углеводным фрагментом или нет, обладает характерным максимумом поглощения в УФ, интенсивность которого зависит от концентрации. По этой причине, исходя из УФ-спектров выделенных соединений, можно определить количественное соотношение оснований, а следовательно, и нуклеотидный состав исходной НК.
При количественном определении минорных нуклеотидов, особенно таких неустойчивых, как дигидроуридиловая кислота, пользуются ферментативными методами гидролиза (ФДЭ змеиного яда и селезенки).
Использование описанных выше аналитических приемов показало, что ПК различного происхождения состоят за редким исключением из четырех основных нуклеотидов и что содержание минорных нуклеотидов может меняться в значительных пределах.
Как будет показано далее, при изучении нуклеотидного состава ДНК были получены данные, которые помогли установить ее пространственную структуру.
Состав ДНК
Исследуя нуклеотидный состав нативных ДНК различного происхождения, Чаргафф обнаружил следующие закономерности.
1. Все ДНК независимо от их происхождения содержат одинаковое число пуриновых и пиримидиновых оснований. Следовательно, в любой ДНК на каждый пуриновый нуклеотид приходится один пиримидиновый.
2. Любая ДНК всегда содержит в равных количествах попарно аденин и тимин, гуанин и цитозин, что обычно обозначают как А=Т и G=C. Из этих закономерностей вытекает третья.
3. Количество оснований, содержащих аминогруппы в положении 4 пиримидинового ядра и 6 пуринового (цитозин и аденин), равно количеству оснований, содержащих оксо-группу в тех же положениях (гуанин и тимин), т. е. A+C=G+T. Эти закономерности получили название правил Чаргаффа. Наряду с этим было установлено, что для каждого типа ДНК суммарное содержание гуанина и цитозина не равно суммарному содержанию аденина и тимина, т. е. что (G+C)/(A+T), как правило, отличается от единицы (может быть как больше, так и меньше ее). По этому признаку различают два основных типа ДНК: А Т-тип с преимущественным содержанием аденина и тимина и G C-тип с преимущественным содержанием гуанина и цитозина.
Величину отношения содержания суммы гуанина и цитозина к сумме содержания аденина и тимина, характеризующую нуклеотидный состав данного вида ДНК, принято называть коэффициентом специфичности. Каждая ДНК имеет характерный коэффициент специфичности, который может изменяться в пределах от 0,3 до 2,8. При подсчете коэффициента специфичности учитывается содержание минорных Оснований, а также замены основных оснований их производными. Например, при подсчете коэффициента специфичности для ЭДНК зародышей пшеницы, в которой содержится 6% 5-метилцитозина, Последний входит в сумму содержания гуанина (22,7%) и цитозина (16,8%). Смысл правил Чаргаффа для ДНК стал понятным после установления ее пространственной структуры.
Состав РНК
Первые сведения о нуклеотидном составе РНК относились к препаратам, представляющим собой смеси клеточных РНК (рибосомных, информационных и транспортных) и называемым обычно суммарной фракцией РНК. Правила Чаргаффа в этом случае не соблюдаются, хотя определенное соответствие между содержанием гуанина и цитозина, а также аденина и урацила все же имеет, место.
Данные, полученные в последние годы при анализе индивидуальных РНК, показывают, что и на них правила Чаргаффа не распространяются. Однако различия в содержании аденина и урацила, а также гуанина и цитозина для большинства РНК невелики и что, следовательно, тенденция к выполнению указанных правил все же наблюдается. Этот факт объясняется особенностями макроструктуры РНК.
Характерными структурными элементами некоторых РНК являются минорные основания. Соответствующие им нуклеотидные остатки обычно входят в состав транспортных и некоторых других РНК в очень небольших количествах, поэтому определение полного нуклеотидного состава таких РНК представляет собой иногда весьма сложную задачу.
biohimist.ru
Строение нуклеиновых кислот — Науколандия
По общему строению все нуклеиновые кислоты являются молекулами-полимерами, т. е. биополимерами, их мономерами являются нуклеотиды. Строение конкретного типа нуклеиновой кислоты имеет свои особенности как на уровне нуклеотидов, так и на уровне вторичной и третичной структуры молекул.
Существует два основных типа нуклеиновых кислот — ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая кислота).
Общее строение каждого нуклеотида-мономера, входящего в состав молекулы любой нуклеиновой кислоты, таково:
- азотистое основание,
- пентоза,
- остаток фосфорной кислоты.
То есть каждый нуклеотид состоит из трех компонентов.
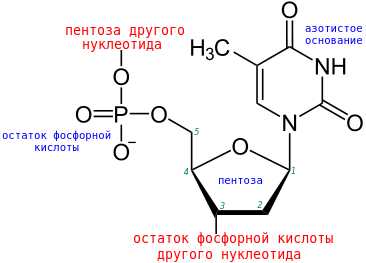 Строение и связи одного нуклеотида, входящего в состав нуклеиновой кислоты.
Строение и связи одного нуклеотида, входящего в состав нуклеиновой кислоты.
Числами обозначены номера атомов углерода в пентозе.
Пентоза — это углевод, включающий пять атомов углерода. В состав нуклеиновых кислот входят кольцевые (а не линейные) формы рибозы и дезоксирибозы. Первая входит в состав РНК, вторая — в ДНК. Дезоксирибоза отличается от рибозы тем, что лишена одной гидроксильной (-OH) группы. Пятичленное кольцо формируют четыре атома углерода и один атом кислорода. Один из атомов углерода (пятый) остается вне кольца и образует связь с остатком фосфорной кислоты.
Азотистые основания, входящие в состав нуклеиновых кислот, делятся на две группы — пуриновые и пиримидиновые. Пуриновые основания состоят из двух колец, одно из которых имеет 5 атомов, а другое — 6, при этом кольца соединены так, что у них два атома общие. Пиримидиновые основания включают одно шестичленное кольцо. К пуринам относятся гуанин (Г) и аденин (А). К пиримидинам относятся тимин (Т), цитозин (Ц) и урацил (У).
В ДНК входят Г, А, Т, Ц. В РНК — Г, А, У, Ц. То есть в РНК тимин заменен на урацил.
Между собой нуклеотиды соединены через остатки фосфорной кислоты, чем и определяют строение полимера — нуклеиновой кислоты. Остаток фосфорной кислоты через один атом кислорода соединен с 5-м атомом углерода пентозы одного нуклеотида, а через другой атом кислорода – с 3-м атомом углерода пентозы другого нуклеотида. Таким образом между нуклеотидами образуется фосфодиэфирная связь. В итоге получается одна полинуклеотидная цепочка.
Сборка нуклеиновой кислоты из нуклеотидов катализирует фермент полимераза. Он присоединяет фосфатную группу очередного нуклеотида к гидроксильной группе, находящейся при 3-м атоме углерода пентозы нуклеотида, который уже является частью нуклеиновой кислоты. Таким образом, синтез полинуклеотидной цепи всегда осуществляется с 3′-конца.
5′-конец первого нуклеотида цепи всегда несет фосфатную группу с одной эфирной связью. Этот конец считается началом цепи.
По строению рибонуклеиновые кислоты — одноцепочечные молекулы. Молекулы дезоксирибонуклеиновой кислоты имеют в своем строении две полимерные цепочки, закрученные относительно друг друга в спираль. Этим определяется вторичная структура ДНК. Цепочки соединены между собой водородными связями, образующимися между азотистыми основаниями.
Азотистые основания в молекуле ДНК соединены между собой по принципу комплиментарности: аденин с тимином (образуют две водородные связи), а гуанином с цитозином (три связи).
Многие молекулы ДНК — самые длинные из существующих полимеров.
Существует несколько типов РНК. Наиболее известные — информационная (она же матричная), транспортная и рибосомальная. Транспортная РНК имеет вторичную структуру, по форме напоминающую клеверный лист. Ее третичная структура похожа на букву Г.
Третичную структуру ДНК определяют различные уровни ее компактизации.
scienceland.info
Химический состав и структура нуклеиновых кислот.
План: СТРОЕНИЕ И ТИПЫ РНК.
ГЕНЕТИЧЕСКИЙ КОД. СИНТЕЗ БЕЛКА В КЛЕТКЕ
Нуклеиновые кислоты впервые открыл И. Ф. Мишер в 1868 г. Он выделил из ядер клеток особое вещество кислотной природы и назвал его нуклеином. Впоследствии ему дали название «нуклеиновая кислота». Было обнаружено два типа нуклеиновых кислот. Их назвали в зависимости от углеводного компонента, входящего в состав. Нуклеиновую кислоту, в состав которой входит углевод дезоксирибоза, назвали дезоксирибонуклеиновой кислотой (ДНК), а в состав которой входит углевод рибоза, — рибонуклеиновой кислотой (РНК)- В период с 1900 по 1932 г. был определен химический состав нуклеиновых кислот. Они включают следующие компоненты:
РНК
Аденин, гуанин Цитозин, урацил Рибоза
ДНК
Аденин, гуанин Цитозин, тимин Дезоксирибоза
Пуриновые основания Пиримидиновые основания Углеводный компонент
Обе нуклеиновые кислоты включают остатки фосфорной кислоты. Различие заключается в том, что в состав РНК входит азотистое основание урацил вместо тимина и рибоза вместо дезоксирибозы.
В 1936 г. на кафедре биохимии растений Московского университета А. Н. Белозерский с И. И. Дубровской впервые выделили ДНК в чистом виде из растительного материала. К середине 40-х годов было выяснено, что ДНК и РНК одновременно присутствуют в каждом живом организме.
В конце 40-х — начале 50-х годов при изучении нуклеиновых кислот стали использовать новые физические и химические методы исследования. В 1950 г. Э. Чаргафф установил правила нукле-отидных отношений, лежащие в основе строения всех ДНК.
Правила Чаргаффа заключаются в том, что в ДНК содержание аденина равно содержанию тимина (А = Т), а содержание гуанина равно содержанию цитозина (Г = Ц), отсюда А + Г/Т + Ц = 1; сумма пуриновых нуклеотидов равна сумме пиримидиновых нуклео-тидов. В соответствии с этим правилом нуклеотидный состав разных организмов может варьировать только по величине А + Т/Г+ Ц.
К 1952 г. Р. Франклин и М. Уилкинс добились получения высококачественных рентгенограмм ДНК, показавших, что она имеет форму спирали и двойственную структуру.
В 1953 г. Дж. Уотсон и Ф. Крик, опираясь на данные рентге-ноструктурного анализа и правила Чаргаффа, установили структуру ДНК. Согласно их модели молекула ДНК имеет двойную спираль, состоящую из двух полинуклеотидных цепей с общей осью (рис. 17). Диаметр двойной спирали ДНК равен 2 нм, а расстояние между витками 3,4 нм. На каждый виток спирали приходится 10 пар нуклеотидов, отсюда расстояние между азотистыми основаниями равно 0,34 нм.
Структурными единицами полинуклеотидных цепей являются нуклеотиды. В состав нуклеотида входят: одно из азотистых оснований — пуриновое (аденин или гуанин) или пиримидиновое (тимин или цитозин), дезоксирибоза, фосфатный остаток. Эти компоненты соединены друг с другом в следующем порядке: азотистое основание — дезоксирибоза — фосфатный остаток. Соединение одного из оснований с дезоксирибозой приводит к образованию нуклеозида. В случае присоединения фосфатной группы к углеводной части нуклеозида образуется нуклеотид.
Дезоксирибоза в нуклеотидах соединяется с основаниями гли-козидной связью, а с фосфорной кислотой — эфирными связями. Следовательно, по химическому составу любой нуклеотид — это фосфорный эфир нуклеозидов. В соответствии с этим нукле-отиды называются дезоксиадениловой, двзоксигуаниловой, дезокси-цитидиловой и тимидиловой кислотами.
Наряду с главными азотистыми основаниями ДНК содержит также метилированные основания, такие, как 5-метилцитозин, 5-оксиметилцитозин и др. У животных количество 5-метилцито-зина в ДНК обычно не превышает 1,5—2 %.
В каждой из цепей ДНК нуклеотиды последовательно соединены друг с другом с помощью остатка фосфорной кислоты и молекулы дезоксирибозы. Дезоксирибоза связывается с одной молекулой фосфорной кислоты через углерод в положении 3′, а с другой — через углерод 5′, образуя углеводно-фосфатный остов (рис. 18).
Обе цепи в молекуле ДНК имеют противоположную полярность. Это означает, что межнуклеотидная связь в одной цепи имеет направление 5′-*3′, а в другой 3′-»5′.
Азотистые основания нуклеотидов обеих цепей ДНК заключены внутри между витками спирали и соединены водородными связями. В соответствии с правилами Чаргаффа аденин одной цепи связан только с тимином другой цепи, а гуанин — только с цитозином. Пара аденин — тимин соединена двумя водородными связями, а пара гуанин — цитозин — тремя. Такой порядок соответствия азотистых оснований (А **Г и Г **Ц) называется
комплементарностью, и, следовательно, цепи в ДНК комплементарны, они взаимно дополняют друг друга.
Углеводно-фосфатный остов по всей длине во всех молекулах ДНК имеет однотипную структуру и не может нести генетической информации. В противоположность этому рас-
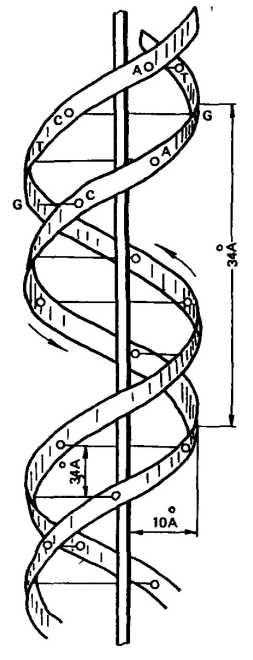
Рис. 17. Схема строения молекулы ДНК из
двух спирально закрученных ценей (по Д. Уотсо-
ну в Ф, Кряку) (Цифры указывают на расстоя-
ним в А между разными точками молекулы)
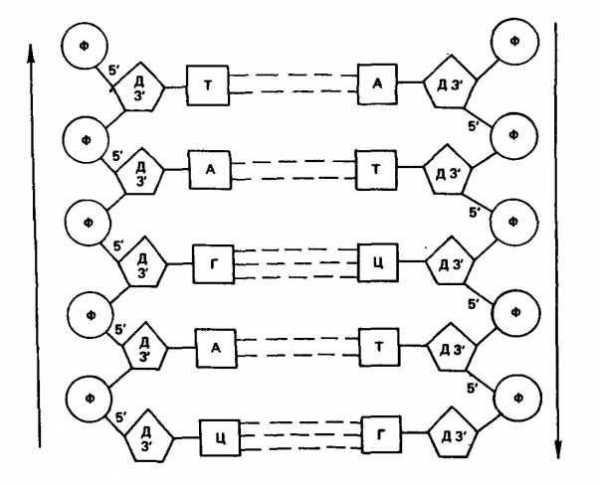
Рис. 18. Схема отрезка двухцепочной молекулы ДНК (по С. М. Гершензону)
положение пуриновых и пиримидиновых оснований нуклеотидов вдоль цепи ДНК очень изменчиво и характерно для каждого данного типа молекул ДНК. Значит, наследственная информация зашифрована различной последовательностью оснований.
Нуклеотидный состав ДНК значительно варьирует в зависимости от принадлежности организма к той или иной систематической группе (табл. 7). Специфичность ДНК выражается соотношением А + Т/Г + Ц, получившим название коэффициента видовой специфичности.
7. Нуклеотидный состав ДНК из различных источников (по А. Ленинджеру, 1976)
|
Нуклеотидный |
состав, к |
ЛОЛ. % |
Коэффициент | ||
|
Объект |
специфичности | ||||
|
A |
Г |
Ц |
Т |
А + Т/Г + Ц | |
|
Человек |
30,9 |
19,9 |
19,8 |
29,4 |
1,52 |
|
Животные: | |||||
|
овиа |
29,3 |
21,4 |
21,0 |
28,3 |
1,36 |
|
курица |
28,8 |
20,5 |
21,5 |
29,2 |
1,38 |
|
Растения, грибы, зерна пшеницы |
27,3 |
22,7 |
22,8 |
27,1 |
1,19 |
|
Дрожжи |
31,3 |
18,7 |
17,1 |
32,9 |
1,79 |
|
Aspergillus niger |
25,0 |
25,1 |
25,0 |
24,9 |
1,00 |
|
Бактерии: | |||||
|
Е. coli |
24,7 |
26,0 |
25,7 |
23,6 |
0,93 |
|
Staphylococcus ayreus |
30,8 |
21,0 |
19,0 |
29,2 |
1,50 |
|
Clostridium perfnngens |
36,9 |
14,0 |
12,8 |
36,3 |
2,70 |
|
Brucella abortus |
21,0 |
29,0 |
28,9 |
21,1 |
0,72 |
|
Sarcina lutea |
13,4 |
37,1 |
37,1 |
12,4 |
0,35 |
В ДНК животных наблюдается избыток А + Т по отношению к Г + Ц. У грибов и бактерий встречаются формы как богатые А + Т, так и с преобладанием Г + Ц, в то же время есть близкие по коэффициенту специфичности к животным. Это говорит о том, что изменчивость в расположении оснований уже достаточна для того, чтобы обеспечить различия между генами этих организмов.
Молекулы ДНК состоят примерно из 2-103—1-Ю8 и более
нуклеотидов и имеют большую относительную молекулярную массу.
Репликация (удвоение) ДНК. ДНК находится в хромосомах, и репликация ее происходит перед каждым удвоением хромосом и делением клетки. Дж. Уотсон и Ф. Крик предложили схему удвоения ДНК, согласно которой спиралевидная двухцепочная ДНК сначала раскручивается (расплетается) вдоль оси. При этом водородные связи между азотистыми основаниями рвутся и цепи расходятся. Одновременно к нуклеотидам каждой цепи пристраиваются комплементарные азотистые основания нуклеотидов второй цепи, где против аденина встает тимин, против тимина — аденин, против гуанина — цитозин и т. д., которые с помощью ферментов ДНК-полимераз связываются в новые полинуклеотидные цепи. В результате из одной образуются две новые дочерние молекулы ДНК. Каждая дочерняя молекула, наследуя структуру одной цепи материнской молекулы, строго сохраняет специфичность заключенной в ней информации. Поскольку матрицей для репликации служит одна из двух цепей молекулы, такой тип синтеза ДНК носит название полуконсервативной ауторепродукции.
Дальнейшие исследования показали, что репликация бактериальных и других молекул ДНК начинается в определенной точке старта. В хромосомах эукариот обнаружено по нескольку таких начальных точек. Цепи ДНК в точке инициации репликации разъединяются под влиянием особого белка геликазы (рис. 19). Возникают одноцепочные участки ДНК, которые становятся матрицами для репликации-притяжения комплементарных нуклеотидов. Эти одноцепочные участки связываются с особыми белками, которые их стабилизируют (препятствуют их комплементарному взаимодействию). Особый фермент топоизомераза (у прокариот назьшается ДНК-гиразой) способствует расщеплению спирали ДНК в области репликационной вилки.
Репликация на материнской цепи, идущей от точки старта в направлении 5′->3′, идет в виде сплошной линии. Эта цепь получила название лидирующей. Синтез на второй цепи 3′->5′ идет отдельными фрагментами в противоположном направлении (тоже 5′->3′)- Эта цепь получила название запаздывающей. Фрагментами являются небольшие участки ДНК (у кишечной палочки около 2000 нуклеотидов, у эукариот около 200). Они называются по
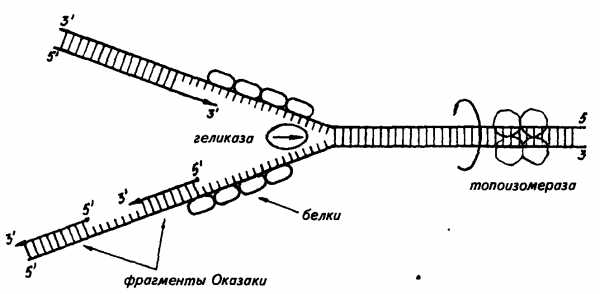
Рис. 19. Синтез лидирующей (вверху) и запаздывающей (внизу) цеией ДНК в вилке репликации (по В. Alberts, R. Sternglanz)
имени открывшего их японского ученого Р. Оказаки. После завершения синтеза фрагменты Оказаки соединяются при помощи фермента ли газы в общую полинуклеотидную цепочку. У эукариот репликация ДНК и соединение различных ее репликационных участков происходят в фазе S-периода интерфазы. После завершения этой фазы в каждой хромосоме имеется две молекулы ДНК, которые становятся двумя идентичными хроматидами.
Структура, способная к репликации (хромосома, плазмида, вирусный геном), называется репликоном.
Самоудвоение молекул ДНК — основа устойчивости генетической информации данного вида и обеспечивает материальную непрерывность наследственного вещества клетки.
СТРОЕНИЕ И ТИПЫ РНК
Многочисленными исследованиями было установлено, что синтез белка в клетке происходит не в ядре, где находится ДНК, а в цитоплазме. Следовательно, сама ДНК не может служить матрицей для синтеза белка. Вставал вопрос о молекулярных механизмах переноса информации, закодированной в ДНК (генах), из ядра в цитоплазму к месту синтеза белка. Сравнительно недавно выяснилось, что молекулами, ответственными за считывание и перенос информации, а также за преобразование этой информации в последовательность аминокислот в структуре белковой молекулы, являются рибонуклеиновые кислоты (РНК). Молекулы рибонуклеиновой кислоты имеют одну полинуклеотидную цепь, Нуклеотиды молекулы РНК называются адениловой гуаниловой, уридиловой и цитцдиловой кислотами. На долю РНК приходится около 5—10 % общей массы клетки.
Существует три основных вида РНК: информационная (иРНК), или матричная (мРНК), рибосомная (рРНК) и транспортная (тРНК). Они различаются по величине молекул и функциям. Все типы РНК синтезируются на ДНК при участии ферментов — РНК-полимераз. Информационная, или матричная, РНК составляет 2—3 % всей клеточной РНК, рибосомная — 80—85, транспортная — около 15 %.
Информационная РНК (иРНК) впервые была обнаружена в 1957 г. Роль ее в том, что она считывает наследственную информацию с участка ДНК (гена) и в форме скопированной последовательности азотистых оснований переносит ее в рибосомы, где происходит синтез определенного белка. Каждая из молекул иРНК по порядку расположения нуклеотидов и по размеру соответствует гену в ДНК, с которого она была транскрибирована. В среднем иРНК содержит 1500 нуклеотидов (75— 3000). Каждый триплет (три нуклеотида) на иРНК называется кодоноц. От кодона зависит, какая аминокислота встанет в данном ме*сте при синтезе белка.’ Информационная РНК может обладать относительной молекулярной массой от 250 до 1000 тыс. Д (дальтон).
Существует большое разнообразие иРНК как в отношении состава, так и величины молекулы. Это связано с тем, что в клетке находится большое количество разнообразных белков, а строение каждого белка обусловлено своим геном, с которого иРНК считала информацию.
Транспортная РНК (тРНК) обладает относительно невысокой молекулярной массой порядка 24—29 тыс. Д и содержит в молекуле от 75 до 90 нуклеотидов. До 10 % всех нуклеотидов тРНК приходится на долю минорных оснований, что, по-видимому, защищает ее от действия гидролитических ферментов.
Роль тРНК заключается в том, что они переносят аминокислоты к рибосомам и участвуют в процессе синтеза белка. Каждая аминокислота присоединяется к определенной тРНК. Ряд аминокислот обладает более одной тРНК. К настоящему времени обнаружено более 60 тРНК, которые отличаются между собой первичной структурой (последовательностью оснований). Вторичная структура у всех тРНК представлена в виде клеверного листа с двухцепочным стеблем и тремя одноцепочными петлями (рис. 20). На конце одной из цепей находится акцепторный участок — триплет ЦЦА, к аденину которого присоединяется специфическая аминокислота. Аминокислота присоединяется к тРНК под действием фермента аминоацил-тРНК-синтетазы, который «узнает» одновременно и аминокислоту, и тРНК. В головке средней петли тРНК находится антикодон — триплет, состоящий из трех нуклеотидов. Антикодон комплементарен определенному кодону мРНК. При помощи антикодона тРНК «узнает» соответствующий кодон в иРНК, т. е. определяет место, куда
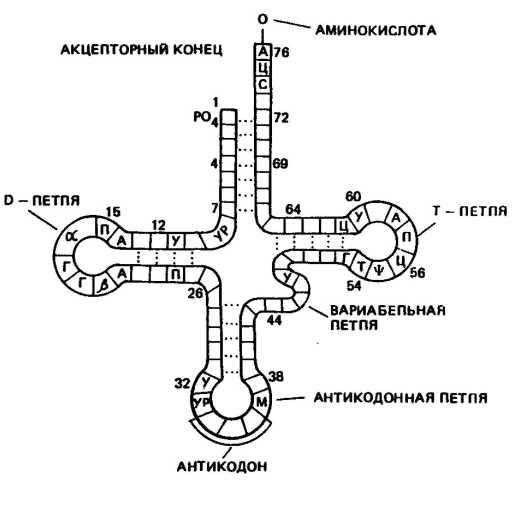
Рис, 20. Вторичная структура тРНК (по Ричу п Киму)
должна быть поставлена данная аминокислота в синтезируемой молекуле белка.
Предполагается, что петли тРНК, не вовлеченные в связывание и выполнение декодирующей функции аминокислоты, используются для связывания тРНК с рибосомой и со специфической аминоацил-тРНК-синтетазой.
Рибосомная РНК (рРНК). Размер рибосомных РНК эукариот составляет 5—28S (S — единица Сведберга, характеризующая скорость осаждения, седиментации частиц при ультрацентрифугировании), молекулярная масса 3,5-104— 1,510^ Д. Они содержат 120—3100 нуклеотидов. Рибосомная РНК накапливается в ядре, в ядрышках. В ядрышки из цитоплазмы транспортируются рибосомные белки, и там происходит спонтанное образование субчастиц рибосом путем объединения белков с соответствующими рРНК. Субчастицы рибосомы вместе или врозь транспортируются через поры ядерной мембраны в цитоплазму.
Рибосомы представляют собой органеллы величиной 20— 30 нм. Они построены из двух субчастиц разного размера и формы. На определенных стадиях белкового синтеза в клетке происходит разделение рибосом на субчастицы. Рибосомная РНК служит как бы каркасом рибосом и способствует первоначальному связыванию иРНК с рибосомой в процессе биосинтеза белка. Субчастицы обозначают у эукариот как 60 и 40S. Целые рибосомы
осаждаются при 80S. 40£-субчастица содержит 18S РНК и примерно 30 белков; бОБ-субчастица содержит 28S РНК, 5S РНК и 5,8 S РНК. В состав этой частицы входит примерно 50 различных белков. У прокариот функциональная рибосома имеет константу седиментации 70S. 708-рибосомы состоят из малой (30S) и большой (50S) субчастиц. 808-рибосомы содержат примерно равное количество рРНК и белка, у 70S-pn6ocoM соотношение РНК и белка составляет 2:1. Число рибосом в клетке прокариот равно примерно 104, у эукариот — около 105. В период синтеза белка рибосомы могут объединяться в полисомы, образуя более высокоорганизованные комплексы.
studfiles.net
Нуклеиновые кислоты.
Нуклеиновые кислоты были открыты в 1868г. швейцарским врачом Ф. Мишером. Биологическая функция этого вещества оставалась неизвестной еще в течение почти столетия, и только в 40-х годах прошлого века Эвери, Маклеод и Маккарти установили, что нуклеиновые кислоты, отвечают за хранение, репликацию (воспроизведение), транскрипцию (передачу) и трансляцию (воспроизведение на белок) генетической (наследственной) информации. Короче, именно нуклеиновые кислоты определяют вид, форму, химический состав и функции живой клетки и всего организма в целом.
В 1953 г. Уотсон и Крик сообщили о расшифровке молекулярной структуры ДНК. В каждом живом организме присутствуют два типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). В то же время вирусы содержат только один какой-нибудь тип нуклеиновых кислот: либо РНК, либо ДНК.
Нуклеиновые кислоты – это высокомолекулярные соединения, размер которых сильно варьирует. Молярная масса транспортной РНК составляет 25000, тогда как отдельные молекулы ДНК обладают массой от 1 000 000 до 1 000 000 000.
Количественное содержание ДНК в клетках одного и того же организма постоянно и исчисляется несколькими пикограммами, однако в клетках разных видов живых организмов имеются существенные количественные различия в содержании ДНК. ДНК преимущественно сосредоточено в ядре, митохондриях и хлоропластах. РНК большей частью содержится в цитоплазме клеток. Содержание РНК, как правило, в 5-10 раз больше, чем ДНК. Соотношение РНК/ДНК в клетках тем выше, чем интенсивнее в них синтез белка.
Нуклеиновые кислоты обладают сильно выраженными кислотными свойствами и при физиологических значениях рН несут высокий отрицательный заряд. В связи с этим в клетках организмов они легко взаимодействуют с различными катионами и прежде всего с основными белками, образуя нуклеопротеины.
-
Состав нуклеиновых кислот.
Нуклеиновые кислоты при полном их гидролизе распадаются на три типа веществ – азотистые основания (пуриновые и пиримидиновые основания), сахара (пентозы) и фосфорную кислоту.
Пентозы нуклеиновых кислот представлены D-рибозой или 2-D-дезоксирибозой. Оба эти сахара содержатся в составе нуклеиновых кислот в фуранозной форме и имеют -конфигурацию:

Нуклеиновая кислота называется рибонуклеиновой (РНК), если в ее состав входит рибоза, или дезоксирибонуклеиновой (ДНК), если в ее состав входит дезоксирибоза. Недавно установлено, что рибоза и дезоксирибоза не являются единственными углеводами, входящими в состав нуклеиновых кислот: в ряде фаговых ДНК и РНК некоторых видов раковых клеток найдена глюкоза.
Азотистые основания, которые обычно встречаются в нуклеиновых кислотах – это производные пурина аденин ( А) и гуанин (G)-и производные пиримидина – цитозин (С), тимин (Т) и урацил (U). Сами пурин и пиримидин в состав нуклеиновых кислот не входят.
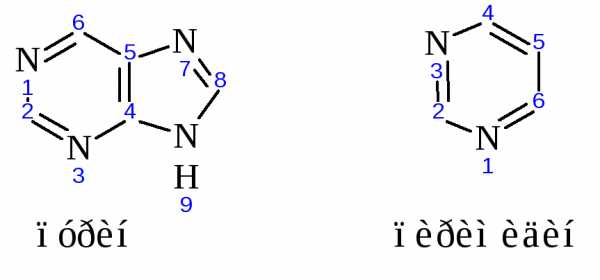
Строение основных азотистых оснований-компонентов нуклеиновых кислот:
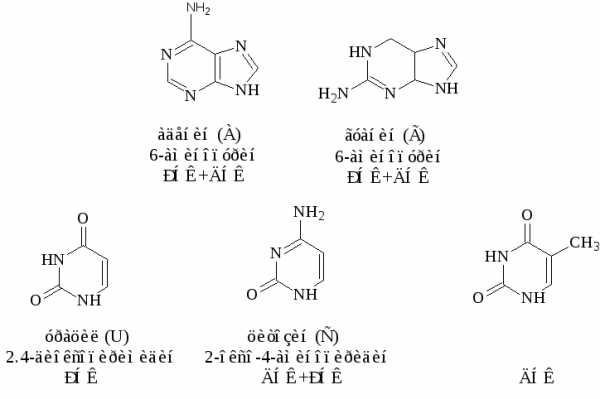
Цитозин, аденин, гуанин содержатся в нуклеиновых кислотах обоих типов, урацил входит только в состав РНК, а тимин в ДНК.
Для гуанина, цитозина, тимина и урацила известна кето-енольная таутомерия, однако кетоструктуры гораздо более стабильны и доминируют при физиологических условиях.
studfiles.net
Нуклеиновые кислоты
НУКЛЕОПРОТЕИДЫ
Нуклеопротеиды входят в состав всех клеток организма и выполняют основные функции — служат носителями генетической информации и участвуют в биосинтезе белка. По химической природе эти макромолекулы представляют собой комплекс белков, имеющих, как правило, положительный заряд (гистонов) и нуклеиновых кислот, которые при физиологических значениях рН имеют высокий отрицательный заряд. Таким образом, между белковой частью и нуклеиновой кислотой в нуклеопротеидах возникают электростатические взаимодействия.
Нуклеопротеиды
ДНП РНП
(Дезоксирибонуклеопротеид) (Рибонуклеопротеид)
Белки
(гистоны, белки
протамины и др)
ДНК РНК
Главные мононуклеотиды Главные мононуклеотиды
(дезоксирибонуклеотиды) (рибонуклеотиды)
д — АМФ АМФ
д — ГМФ ГМФ
д — ЦМФ ЦМФ
д — ТМФУМФ
Дезоксирибонуклеозиды Н3РО4Н3РО4Рибонуклеозиды
д — Аденозин Аденозин
д — Гуанозин Гуанозин
д — Цитидин Цитидин
д — ТимидинУридин
N- ОснованияДезоксирибоза РибозаN- Основания
Аденин Аденин
Гуанин Гуанин
Цитозин Цитозин
ТиминУрацил
Нуклеиновыми кислотами или полинуклеотидами называются высокомолекулярные вещества, состоящие из мононуклеотидов, связанных между собой 3,5-фосфодиэфирными связями. Нуклеотиды представляют собой соединения, в которых остаток фосфорной кислоты присоединен к нуклеозиду чаще всего 5-фосфомоноэфирной связью. К нуклеозидам относят вещества, в которых азотистые основания соединены с пентозой N-гликозидной связью. В зависимости от типа пентозы различают два вида нуклеозидов – дезоксирибонуклеозиды, содержащие 2-дезоксирибозу, и рибонуклеозиды, содержащие рибозу.
Выделяют два вида нуклеиновых кислот: дезоксирибонуклеиновую (ДНК), представляющую собой полидезоксирибонуклеотид, и рибонуклеиновую (РНК) или полирибонуклеотид. В свою очередь, нуклеопротеиды также можно подразделить на две группы: дезоксирибонуклеопротеиды (ДНП), в состав которых входит ДНК, и рибонуклеопротеиды (РНП), содержащие РНК.
Нуклеиновые кислоты также как все полимеры имеют вторичную и третичную структуры. Вторичная структура ДНК имеет вид двутяжной антипараллельной спирали. Схематично она напоминает винтовую лестницу, перила которой образованы дезоксирибозами соединенными фосфорно-эфирными связями по типу 3,5- связей, а ступени парами азотистых оснований, одно из которых представлено производными пурина, другое — пиримидина. Азотистые основания соединяются по принципу комплементарности. Так, аденин всегда соединяется с тимином (А-Т), а гуанин — с цитозином (Г — Ц) с помощью водородных связей. Третичная структура ДНК образуется в результате дополнительного скручивания в пространстве двуспиральной молекулы. Она имеет вид суперспирали или изогнутой двойной спирали. Вторичная структура РНК — это частично спирализованная одинарная полинуклеотидная цепь, участки спирализации которой удерживаются за счет водородных связей, образованных между комплементарными азотистыми основаниями — А — У и Г — Ц. Так например, вторичная структура тРНК имеет вид «клеверного листа». Она образуется вследствие внутрицепочечного спаривания комплементарных нуклеотидов отдельных участков тРНК. Те участки молекулы, которые не вовлекаются в образование водородных связей между нуклеотидами, образуют петли. Вторичная структура рРНК представляет собой спиральные участки, соединенные изогнутой одиночной цепью.
Общее содержание ДНК и РНК в клетках зависит от их функционального состояния. В сперматозоидах количество ДНК достигает 60% сухой массы клетки, в большинстве клеток 1-10%. Содержание РНК, как правило, в 5-10 раз превышает содержание ДНК. Основные различия в локализации и функционировании основных типов нуклеиновых кислот приведены в таблице 1.
Таблица 1
Краткая характеристика нуклеиновых кислот клеток высших организмов
|
Тип нуклеиновой кислоты |
Молекулярная масса |
Локализация в клетке |
Функция |
|
ДНК |
1011 |
Ядро Митохондрии |
Хранение генетической информации и участие в передаче ее дочерней ДНК при делении клетки или РНК в биосинтезе белка |
|
мРНК |
4104 – 1,2106 |
Ядро Цитоплазма |
Является копией участка ДНК, содержащего информацию о структуре молекулы белка. Переносит информацию от ДНК к рибосомам – месту синтеза белка |
|
тРНК |
2,5104 |
Гиалоплазма, рибосомы, митохондрии |
Участвует в активировании аминокислот, их транспорте к рибосомам и сборке из аминокислот белков |
|
рРНК |
4104-1,75106 |
Рибосомы |
Образует скелет рибосом, который окутывается белками. Играет вспомогательную роль при сборке белка на рибосомах |
Гидролиз нуклеопротеидов дрожжей. Качественные реакции на продукты гидролиза нуклеопротеидов
Для изучения химического состава нуклеопротеидов проводят кислотный гидролиз дрожжей, богатых нуклеопротеидами, и выявляют продукты гидролиза — полипептиды, пуриновые основания, углевод и фосфорную кислоту специфическими для каждого вещества реакциями.
studfiles.net
Нуклеиновые кислоты.
Нуклеиновые кислоты были открыты в 1868г. швейцарским врачом Ф. Мишером. Биологическая функция этого вещества оставалась неизвестной еще в течение почти столетия, и только в 40-х годах прошлого века Эвери, Маклеод и Маккарти установили, что нуклеиновые кислоты, отвечают за хранение, репликацию (воспроизведение), транскрипцию (передачу) и трансляцию (воспроизведение на белок) генетической (наследственной) информации. Короче, именно нуклеиновые кислоты определяют вид, форму, химический состав и функции живой клетки и всего организма в целом.
В 1953 г. Уотсон и Крик сообщили о расшифровке молекулярной структуры ДНК. В каждом живом организме присутствуют два типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). В то же время вирусы содержат только один какой-нибудь тип нуклеиновых кислот: либо РНК, либо ДНК.
Нуклеиновые кислоты – это высокомолекулярные соединения, размер которых сильно варьирует. Молярная масса транспортной РНК составляет 25000, тогда как отдельные молекулы ДНК обладают массой от 1 000 000 до 1 000 000 000.
Количественное содержание ДНК в клетках одного и того же организма постоянно и исчисляется несколькими пикограммами, однако в клетках разных видов живых организмов имеются существенные количественные различия в содержании ДНК. ДНК преимущественно сосредоточено в ядре, митохондриях и хлоропластах. РНК большей частью содержится в цитоплазме клеток. Содержание РНК, как правило, в 5-10 раз больше, чем ДНК. Соотношение РНК/ДНК в клетках тем выше, чем интенсивнее в них синтез белка.
Нуклеиновые кислоты обладают сильно выраженными кислотными свойствами и при физиологических значениях рН несут высокий отрицательный заряд. В связи с этим в клетках организмов они легко взаимодействуют с различными катионами и прежде всего с основными белками, образуя нуклеопротеины.
-
Состав нуклеиновых кислот.
Нуклеиновые кислоты при полном их гидролизе распадаются на три типа веществ – азотистые основания (пуриновые и пиримидиновые основания), сахара (пентозы) и фосфорную кислоту.
Пентозы нуклеиновых кислот представлены D-рибозой или 2-D-дезоксирибозой. Оба эти сахара содержатся в составе нуклеиновых кислот в фуранозной форме и имеют -конфигурацию:

Нуклеиновая кислота называется рибонуклеиновой (РНК), если в ее состав входит рибоза, или дезоксирибонуклеиновой (ДНК), если в ее состав входит дезоксирибоза. Недавно установлено, что рибоза и дезоксирибоза не являются единственными углеводами, входящими в состав нуклеиновых кислот: в ряде фаговых ДНК и РНК некоторых видов раковых клеток найдена глюкоза.
Азотистые основания, которые обычно встречаются в нуклеиновых кислотах – это производные пурина аденин ( А) и гуанин (G)-и производные пиримидина – цитозин (С), тимин (Т) и урацил (U). Сами пурин и пиримидин в состав нуклеиновых кислот не входят.
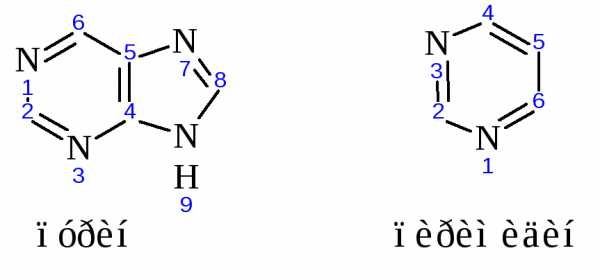
Строение основных азотистых оснований-компонентов нуклеиновых кислот:
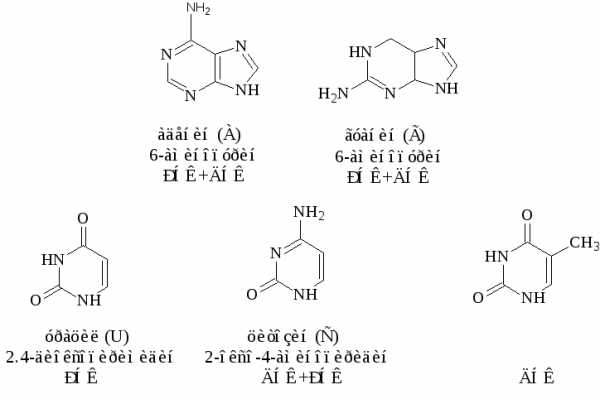
Цитозин, аденин, гуанин содержатся в нуклеиновых кислотах обоих типов, урацил входит только в состав РНК, а тимин в ДНК.
Для гуанина, цитозина, тимина и урацила известна кето-енольная таутомерия, однако кетоструктуры гораздо более стабильны и доминируют при физиологических условиях.
studfiles.net
