
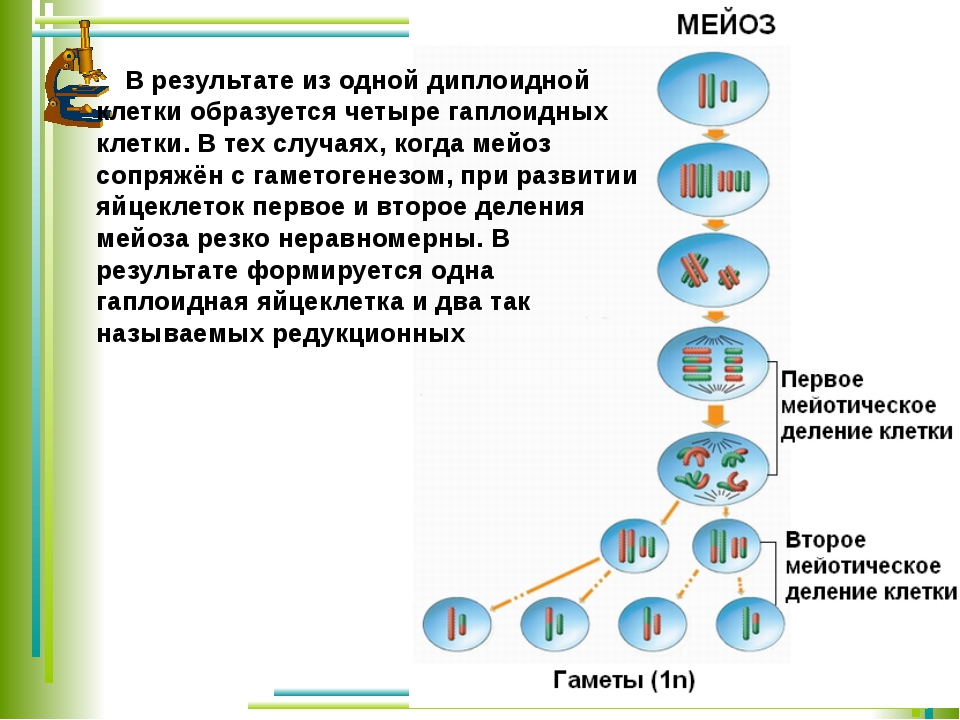
Найдите три ошибки в приведённом тексте. Укажите номера предложений, в которых сделаны ошибки, и после исправьте их.1. Спорогенезом называют процесс образования половых клеток у растений. 2. Споры образуются у растений путём мейотического деления материнских клеток спор. 3. Процесс образования микроспор или пыльцевых зёрен у растений называют микроспорогенезом, а процесс образования мегаспор — макроспорогенезом. 4. Из микроспоры в результате мейоза образуются вегетативное и генеративное ядра. 5. Генеративное ядро делится митозом и образует два спермия. 6. Макроспора в результате двойного митотического деления образует восьмиядерный зародышевый мешок. 7. Зародышевый мешок
3007. Найдите три ошибки в приведённом тексте. Укажите номера предложений, в которых сделаны ошибки, и после исправьте их.1. Спорогенезом называют процесс образования половых клеток у растений. 2. Споры образуются у растений путём мейотического деления материнских клеток спор. 3. Процесс образования микроспор или пыльцевых зёрен у растений называют микроспорогенезом, а процесс образования мегаспор — макроспорогенезом.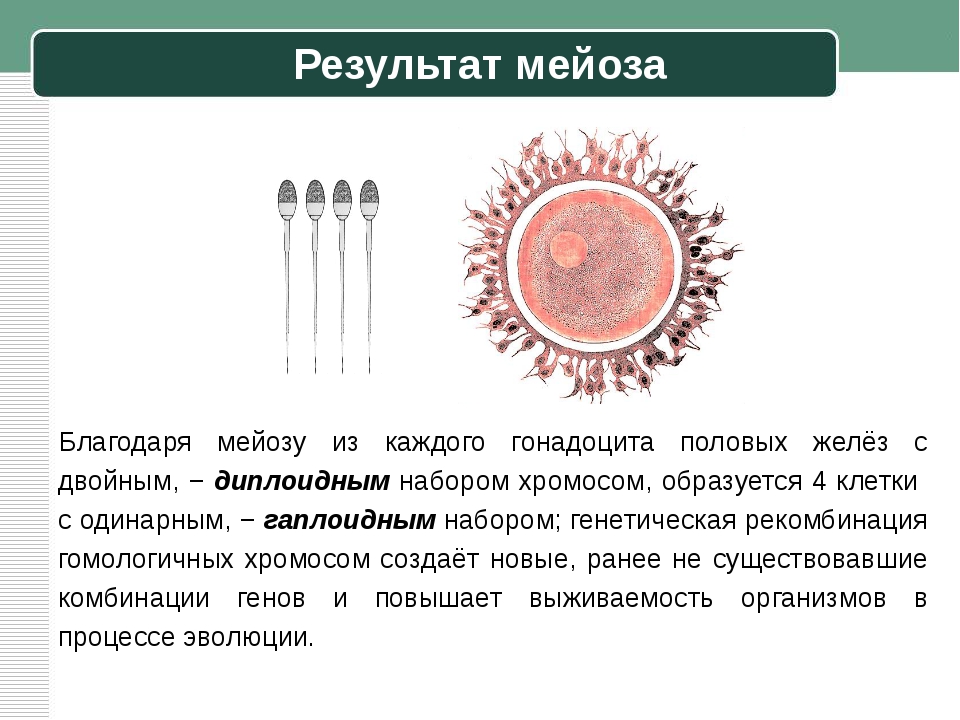 4. Из микроспоры в результате мейоза образуются вегетативное и генеративное ядра. 5. Генеративное ядро делится митозом и образует два спермия. 6. Макроспора в результате двойного митотического деления образует восьмиядерный зародышевый мешок. 7. Зародышевый мешок — это женский гаметофит цветкового
растения.
4. Из микроспоры в результате мейоза образуются вегетативное и генеративное ядра. 5. Генеративное ядро делится митозом и образует два спермия. 6. Макроспора в результате двойного митотического деления образует восьмиядерный зародышевый мешок. 7. Зародышевый мешок — это женский гаметофит цветкового
растения.
Показать подсказку
1) Спорогенезом называют процесс образования спор у растений (процесс образования половых клеток — гаметогенез)
4) Вегетативное и генеративное ядра образуются путем митотических делений
6) Зародышевый мешок образуется из макроспоры в результате трёх митотических делений
P.S. Нашли ошибку в задании? Пожалуйста, сообщите о вашей находке 😉
При обращении указывайте id этого вопроса — 3007.
Проверочная работа по теме «Мейоз»
Проверочная работа по теме «Мейоз»
Половое размножение не характерно…
А) грибам
Б) животным
В) простейшим (амеба, эвглена)
Г) растениям
2. Половые клетки у тополя узколистного
А) сперматозоиды
Б) спермии
В) споры
Г) яйцеклетки
3. Деление клеток, в результате которых образуются гаметы
А) мейоз
Б) митоз
В) сперматогенез
Г) овогенез
4. Процесс деления, в результате которого из исходной диплоидной клетки образуются четыре клетки, называют
А) оплодотворением
Б) дроблением
В) митозом
Г) мейозом
5. В процессе мейоза образуются клетки с набором хромосом
А) диплоидным
Б) равным материнскому
В) удвоенным
Г) гаплоидным
6. Первое деление мейоза называют
А) модификационным
Б) редукционным
В) мутационным
Г) регенерацией
7. Благодаря конъюгации и кроссинговеру происходит
Благодаря конъюгации и кроссинговеру происходит
А) уменьшение числа хромосом вдвое
Б) увеличение числа хромосом
В) обмен генетической информацией между гомологичными хромосомами
Г) увеличение числа гамет
8. Расхождение хромосом при делении клетки происходит в
А) анафазу
Б) метафазу
В) профазу
Г) телофазу
9. Какие клетки образуются путем мейоза
А) мышечные
Б) эпителиальные
В) нервные
Г) половые
10. Каковы причины образования большого разнообразия гамет в процессе мейоза
А) Независимое расхождение гомологичных хромосом
Б) Равномерное распределение хромосом между дочерними клетками
В) Наличие одной интерфазы и двух делений
Г) Строгая зависимость расхождения негомологичных хромосом
11. Первое деление мейоза заканчивается образованием
А) гамет
Б) ядер с гаплоидным набором хромосом
В) диплоидных клеток
Г) клеток разной плоидности
12. В соматических клетках шимпанзе 48 хромосом. Сколько хромосом содержится
В соматических клетках шимпанзе 48 хромосом. Сколько хромосом содержится
А) в сперматозоидах шимпанзе_____
Б) в яйцеклетке_______
В) в соматических клетках после митоза_______
Г) в оплодотворенной яйцеклетке________
13. Второе деление мейоза заканчивается образованием
А) образованием 4 гаплоидных клеток
Б) диплоидных клеток
В) соматических клеток
Г) клеток разной плоидности
14. Размножение, осуществляемое путем слияния гамет, называют
А) бесполым
Б) вегетативным
В) половым
Г) споровым
15. Первое деление мейоза отличается от второго деления мейоза
А) расхождением дочерних хроматид в образующиеся клетки
Б) расхождением гомологичных хромосом и образованием двух гаплоидных клеток
В) делением на две части первичной перетяжки хромосом
Г) образованием двух диплоидных клеток
16. Сколько хромосом содержит ядро исходной клетки, если при мейозе образуется ядро с 12 хромосомами?
А) 6
Б) 12
В) 18
Г) 24
17. Сперматозоиды млекопитающих отличаются от спермиев цветковых растений
Сперматозоиды млекопитающих отличаются от спермиев цветковых растений
А) гаплоидным набором хромосом
Б) крупными размерами
В) наличием запаса питательных веществ
Г) подвижностью
18. Яйцеклетка млекопитающего отличается от сперматозоида тем, что она
А) неподвижна, крупнее, округлой формы
Б) имеет гаплоидный набор хромосом
В) имеет диплоидный набор хромосом
Г) имеет плазматическую мембрану
19. На переднем конце головки сперматозоида имеется
А) центриоль
Б) рыхлый слой
В) ядро
Г) акросома
20. Ядро соматической клетки лягушки содержит 26 хромосом. Сколько молекул ДНК содержит сперматозоид лягушки?
А) 13
Б) 26
В) 39
Г) 52
21. Путем мейоза НЕ образуются
А) гаметы
Б) соматические клетки
В) яйцеклетки
Г) сперматозоиды
22. Сперматозоид, в отличие от яйцеклетки, не имеет
А) запаса питательных веществ
Б) клеточной оболочки
В) обособленного ядра
Г) митохондрий
23. Для представителей какого отдела растений характерно двойное оплодотворение?
Для представителей какого отдела растений характерно двойное оплодотворение?
А) Покрытосеменные
Б) Моховидные
В) Папоротниковидные
Г) Голосеменные
№2. Сравнительная характеристика мейоза и митоза.
Заполните таблицу на основе сравнения митоза и мейоза.
СравнениеМитоз
Мейоз
Сходство
Различие
№3. Дайте определения следующим понятиям:
Мейоз-
Митоз-
Кроссинговер-
Конъюгация хромосом-
Гомологичные хромосомы-
Ответы
1 в
2 б
3 а
4 г
5 г
6 б
7 в
8 а
9 г
10 а
11 б
12 А-24, Б -24,В-48, Г-48
13 а
14 в
15 б
16 г
17 г
18 а
19 г
20 а
21 б
22 а
23 а
Половые клетки человека, хромосомы, оплодотворение — статья МЦРМ
Половые клетки — гаметы (от греч. gametes — «супруг») можно обнаружить уже у двухнедельного эмбриона человека. Их называют первичными половыми клетками. В это время они совсем не похожи на сперматозоиды или яйцеклетки и выглядят абсолютно одинаковыми. Никаких различий, присущих зрелым гаметам, на этой стадии развития зародыша обнаружить у первичных половых клеток не удается. Это не единственная их особенность. Во-первых, первичные половые клетки появляются у зародыша гораздо раньше собственно половой железы (гонады), а во-вторых, они возникают на значительном удалении от того места, где эти железы сформируются позднее. В определенный момент происходит совершенно удивительный процесс — первичные половые клетки дружно устремляются к половой железе и заселяют, «колонизируют» ее.
gametes — «супруг») можно обнаружить уже у двухнедельного эмбриона человека. Их называют первичными половыми клетками. В это время они совсем не похожи на сперматозоиды или яйцеклетки и выглядят абсолютно одинаковыми. Никаких различий, присущих зрелым гаметам, на этой стадии развития зародыша обнаружить у первичных половых клеток не удается. Это не единственная их особенность. Во-первых, первичные половые клетки появляются у зародыша гораздо раньше собственно половой железы (гонады), а во-вторых, они возникают на значительном удалении от того места, где эти железы сформируются позднее. В определенный момент происходит совершенно удивительный процесс — первичные половые клетки дружно устремляются к половой железе и заселяют, «колонизируют» ее.
После того, как будущие гаметы попали в половые железы, они начинают интенсивно делиться, и количество их увеличивается. На этом этапе половые клетки содержат пока то же количество хромосом, что и «телесные» (соматические) клетки — 46. Однако для успешного осуществления своей миссии половые клетки должны иметь в 2 раза меньше хромосом. В противном случае после оплодотворения, то есть слияния гамет, клетки зародыша будут содержать не 46, как установлено природой, а 92 хромосомы. Нетрудно догадаться, что в следующих поколениях их число прогрессивно бы увеличивалось. Чтобы избежать такой ситуации формирующиеся половые клетки проходят специальное деление, которое в эмбриологии называется
Однако для успешного осуществления своей миссии половые клетки должны иметь в 2 раза меньше хромосом. В противном случае после оплодотворения, то есть слияния гамет, клетки зародыша будут содержать не 46, как установлено природой, а 92 хромосомы. Нетрудно догадаться, что в следующих поколениях их число прогрессивно бы увеличивалось. Чтобы избежать такой ситуации формирующиеся половые клетки проходят специальное деление, которое в эмбриологии называется 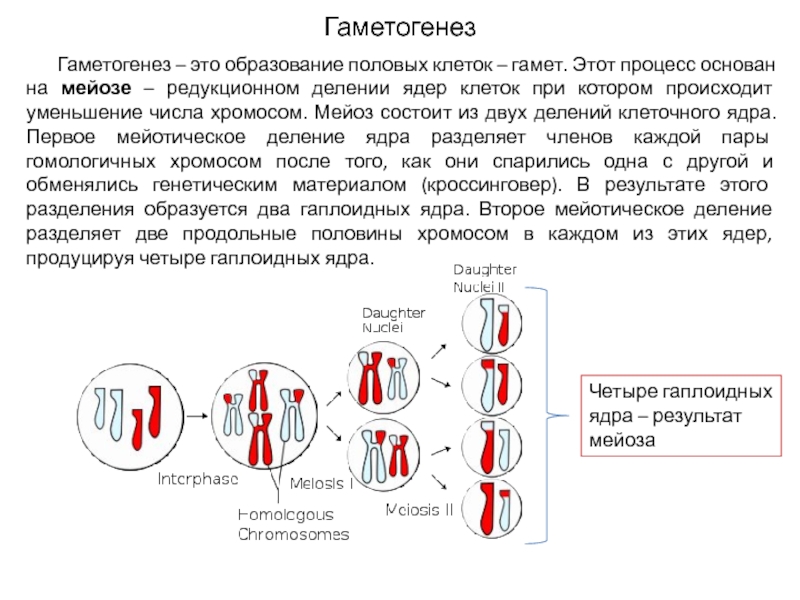 Таким образом, в результате описанных двух делений из одной первичной половой клетки образуется 4 новых.
Таким образом, в результате описанных двух делений из одной первичной половой клетки образуется 4 новых.
Причем, в сперматогенезе (греч. genesis — зарождение, развитие) в результате мейоза появляется 4 зрелых сперматозоида с гаплоидным набором хромосом, а в процессе формирования яйцеклетки — в оогенезе (от греч. oon — «яйцо») только одна. Это происходит потому, что образовавшийся в результате мейоза второй гаплоидный набор хромосом яйцеклетка не использует для формирования новой зрелой половой клетки — ооцита, а «выбрасывает» их, как «лишние», наружу в своеобразном «мусорном контейнере», который называется полярным тельцем. Первое деление хромосомного набора завершается в оогенезе выделением первого полярного тельца непосредственно перед овуляцией. Второе репликационное деление происходит только после проникновения сперматозоида внутрь яйцеклетки и сопровождается выделением второго полярного тельца. Для эмбриологов полярные тельца — очень важные диагностические показатели.
Первичные половые клетки, оказавшиеся в мужской половой железе, до поры до времени не делятся. Их деление начинается только в период полового созревания и приводит к образованию когорты так называемых стволовых диплоидных клеток, из которых и формируются сперматозоиды. Запас стволовых клеток в яичках постоянно пополняется. Здесь уместно напомнить описанную выше особенность сперматогенеза — из одной клетки образуется 4 зрелых сперматозоида. Таким образом, после полового созревания у мужчины в течение всей жизни формируются сотни миллиардов новых сперматозоидов.
Формирование яйцеклеток протекает иначе. Едва заселив половую железу, первичные половые клетки начинают интенсивно делиться. К 5 месяцу внутриутробного развития их количество достигает 6-7 миллионов, но затем происходит массовая гибель этих клеток. В яичниках новорожденной девочки их остается не более 1-2 миллионов, к 7-летнему возрасту — всего лишь около 300 тысяч, а в период полового созревания 30 —50 тысяч. Общее же число яйцеклеток, которые достигнут зрелого состояния за период половой зрелости, будет еще меньше. Хорошо известно, что в течение одного менструального цикла в яичнике обычно созревает лишь один фолликул. Нетрудно подсчитать, что в течение репродуктивного периода, продолжающегося у женщин 30 — 35 лет, образуется около 400 зрелых яйцеклеток.
Общее же число яйцеклеток, которые достигнут зрелого состояния за период половой зрелости, будет еще меньше. Хорошо известно, что в течение одного менструального цикла в яичнике обычно созревает лишь один фолликул. Нетрудно подсчитать, что в течение репродуктивного периода, продолжающегося у женщин 30 — 35 лет, образуется около 400 зрелых яйцеклеток.
Если мейоз в сперматогенезе начинается в период полового созревания и повторяется миллиарды раз в течение жизни мужчины, в оогенезе формирующиеся женские гаметы вступают в мейоз еще в периоде внутриутробного развития. Причем начинается этот процесс почти одновременно у всех будущих яйцеклеток. Начинается, но не заканчивается! Будущие яйцеклетки доходят только до середины первой фазы мейоза, а дальше процесс деления блокируется на 12 — 50 лет! Лишь с приходом половой зрелости мейоз в оогенезе продолжится, причем не всех клеток сразу, а лишь для 1- 2 яйцеклеток ежемесячно. Полностью же процесс мейотического деления яйцеклетки завершится, как уже было сказано выше, только после ее оплодотворения! Таким образом, сперматозоид проникает в яйцеклетку, еще не завершившую деление, имеющую диплоидный набор хромосом!
Сперматогенез и оогенез — очень сложные и во многом загадочные процессы. Вместе с тем очевидна подчиненность их законам взаимосвязи и обусловленности природных явлений. Для оплодотворения одной яйцеклетки in vivo (лат. в живом организме) необходимы десятки миллионов сперматозоидов. Мужской организм вырабатывает их в гигантских количествах практически всю жизнь.
Вместе с тем очевидна подчиненность их законам взаимосвязи и обусловленности природных явлений. Для оплодотворения одной яйцеклетки in vivo (лат. в живом организме) необходимы десятки миллионов сперматозоидов. Мужской организм вырабатывает их в гигантских количествах практически всю жизнь.
Вынашивание и рождение ребенка является чрезвычайно тяжелой нагрузкой на организм. Врачи говорят, что беременность — это проба на здоровье. Каким родится ребенок — напрямую зависит от состояния здоровья матери. Здоровье, как известно, не вечно. Старость и болезни, к сожалению, неотвратимы. Природа дает женщине строго ограниченное невосполнимое число половых клеток. Снижение способности к деторождению развивается медленно, но постепенно по наклонной. Наглядное доказательство того, что это действительно так, мы получаем, ежедневно оценивая результаты стимуляции яичников в программах ВРТ. Большая часть яйцеклеток обычно израсходована к 40 годам, а к 50 годам весь их запас полностью исчерпывается. Нередко так называемое истощение яичников наступает значительно раньше. Следует также сказать, что яйцеклетка подвержена «старению», с годами ее способность к оплодотворению снижается, процесс деления хромосом все чаще нарушается. Заниматься деторождением в позднем репродуктивном возрасте рискованно из-за возрастающей опасности рождения ребенка с хромосомной патологией. Типичным примером является синдром Дауна, который возникает из-за оставшейся при делении третьей лишней 21 хромосомы. Таким образом, ограничив репродуктивный период, природа охраняет женщину и заботится о здоровом потомстве.
Нередко так называемое истощение яичников наступает значительно раньше. Следует также сказать, что яйцеклетка подвержена «старению», с годами ее способность к оплодотворению снижается, процесс деления хромосом все чаще нарушается. Заниматься деторождением в позднем репродуктивном возрасте рискованно из-за возрастающей опасности рождения ребенка с хромосомной патологией. Типичным примером является синдром Дауна, который возникает из-за оставшейся при делении третьей лишней 21 хромосомы. Таким образом, ограничив репродуктивный период, природа охраняет женщину и заботится о здоровом потомстве.
По каким законам происходит деление хромосом? Как передается наследственная информация? Для того чтобы разобраться с этим вопросом, можно привести простую аналогию с картами. Представим себе молодую супружескую пару. Назовем их условно — Он и Она. В каждой его соматической клетке находятся хромосомы черной масти — трефы и пики. Набор треф от шестерки до туза он получил от своей мамы. Набор пик — от своего папы. В каждой ее соматической клетке хромосомы красной масти — бубны и червы. Набор бубен от шестерки до туза она получила от своей мамы. Набор червей — от своего папы.
Набор пик — от своего папы. В каждой ее соматической клетке хромосомы красной масти — бубны и червы. Набор бубен от шестерки до туза она получила от своей мамы. Набор червей — от своего папы.
Для того чтобы получить из диплоидной соматической клетки половую клетку, число хромосом должно быть уменьшено вдвое. При этом половая клетка обязательно должна содержать полный одинарный (гаплоидный) набор хромосом. Ни одна не должна потеряться! В случае карт такой набор можно получить следующим образом. Взять наугад из каждой пары карт черной масти по одной и таким образом сформировать два одиночных набора. Каждый набор будет включать все карты черной масти от шестерки до туза, однако, какие именно это будут карты (трефы или пики) определил случай. Например, в одном таком наборе шестерка может быть пиковой, а в другом — трефовой. Нетрудно прикинуть, что в примере с картами при таком выборе одиночного набора из двойного мы можем получить 2 в девятой степени комбинаций — более 500 вариантов!
Точно также будем составлять одиночный набор из ее карт красной масти. Получим еще более 500 разных вариантов. Из его одиночного и ее одиночного набора карт составим двойной набор. Он получится мягко сказать «пестреньким»: в каждой паре карт одна будет красной масти, а другая — черной. Общее число таких возможных наборов 500×500, то есть 250 тысяч вариантов.
Получим еще более 500 разных вариантов. Из его одиночного и ее одиночного набора карт составим двойной набор. Он получится мягко сказать «пестреньким»: в каждой паре карт одна будет красной масти, а другая — черной. Общее число таких возможных наборов 500×500, то есть 250 тысяч вариантов.
Примерно также, по закону случайной выборки, поступает и природа с хромосомами в процессе мейоза. В результате из клеток с двойным, диплоидным набором хромосом получаются клетки, каждая из которых содержит одиночный, гаплоидный полный набор хромосом. Предположим, в результате мейоза в вашем теле образовалась половая клетка. Сперматозоид или яйцеклетка — в данном случае не важно. Она обязательно будет содержать гаплоидный набор хромосом — ровно 23 штуки. Что именно это за хромосомы? Рассмотрим для примера хромосому № 7. Это может быть хромосома, которую вы получили от отца. С равной вероятностью она может быть хромосомой, которую вы получили от матери. То же самое справедливо для хромосомы № 8, и для любой другой.
Поскольку у человека число хромосом гаплоидного набора равно 23, то число возможных вариантов половых гаплоидных клеток, образующихся из диплоидных соматических, равно 2 в степени 23. Получается более 8 миллионов вариантов! В процессе оплодотворения две половые клетки соединяются между собой. Следовательно, общее число таких комбинаций будет равно 8 млн. х 8 млн. = 64000 млрд. вариантов! На уровне пары гомологичных хромосом основа этого разнообразия выглядит так. Возьмем любую пару гомологичных хромосом вашего диплоидного набора. Одну из таких хромосом вы получили от матери, но это может быть хромосома либо вашей бабушки, либо вашего дедушки по материнской линии. Вторую гомологичную хромосому вы получили от отца. Однако она опять-таки может быть независимо от первой либо хромосомой вашей бабушки, либо вашего дедушки уже по отцовской линии. А таких гомологических хромосом у вас 23 пары! Получается невероятное число возможных комбинаций. Неудивительно, что при этом у одной пары родителей, рождаются дети, которые отличаются друг от друга и внешностью, и характером.
Кстати, из приведенных выше расчетов следует простой, но важный вывод. Каждый человек, ныне здравствующий, или когда-либо живший в прошлом на Земле, абсолютно уникален. Шансы появления второго такого же практически равны нулю. Поэтому не надо себя ни с кем сравнивать. Каждый из вас неповторим, и тем уже интересен!
Однако вернемся к нашим половым клеткам. Каждая диплоидная клетка человека содержит 23 пары хромосом. Хромосомы с 1 по 22 пару называются соматическим и по форме они одинаковы. Хромосомы же 23-й пары (половые хромосомы) одинаковы только у женщин. Они и обозначаются латинскими буквами ХХ. У мужчин хромосомы этой пары различны и обозначаются ХY. В гаплоидном наборе яйцеклетки половая хромосома всегда только Х, сперматозоид же может нести или Х или Y хромосому. Если яйцеклетку оплодотворит Х сперматозоид, родится девочка, если Y сперматозоид — мальчик. Все просто!
Почему мейоз у яйцеклетки так долго растянут во времени? Каким образом ежемесячно происходит выбор когорты фолликулов, которые начинают свое развитие и как из них выделяется лидирующий, доминантный, овуляторный фолликул, в котором созреет яйцеклетка? На все эти непростые вопросы у биологов нет пока однозначных ответов. Процесс формирования зрелых яйцеклеток у человека ждет новых исследователей!
Процесс формирования зрелых яйцеклеток у человека ждет новых исследователей!
Образование и созревание сперматозоидов, как уже было сказано, происходит в семенных канальцах мужской половой железы — яичках. Сформированный сперматозоид имеет длину около 50-60 микрон. Ядро сперматозоида находится в его головке. Оно содержит отцовский наследственный материал. За головкой располагается шейка, в которой имеется крупная извитая митохондрия — органоид, обеспечивающий движения хвоста. Иначе говоря, это своеобразная «энергетическая станция». На головке сперматозоида есть «шапочка». Благодаря ей форма головки — овальная. Но, дело не в форме, а в том, что содержится под «шапочкой». «Шапочка» эта на самом деле является контейнером и называется акросомой, а содержатся в ней ферменты, которые способны растворять оболочку яйцеклетки, что необходимо для проникновения сперматозоида внутрь — в цитоплазму яйцеклетки. Если у сперматозоида нет акросомы, головка у него не овальная, а круглая.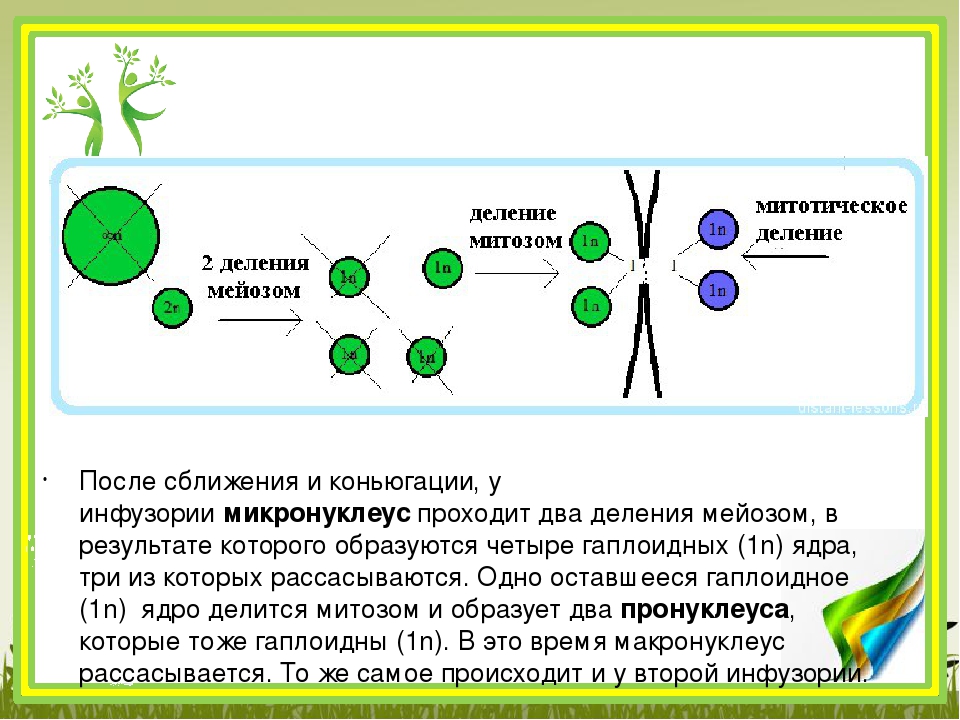 Эта патология сперматозоидов называется глобулоспермия (круглоголовые сперматозоиды). Но, беда опять не в форме, а в том, что такой сперматозоид не может оплодотворить яйцеклетку, и мужчина с таким нарушением сперматогенеза до начала 90-х прошлого столетия был обречен на бездетность. Сегодня благодаря ВРТбесплодие у этих мужчин может быть преодолено, но об этом мы расскажем позднее в главе, посвященной микроманипуляциям, в частности, ИКСИ.
Эта патология сперматозоидов называется глобулоспермия (круглоголовые сперматозоиды). Но, беда опять не в форме, а в том, что такой сперматозоид не может оплодотворить яйцеклетку, и мужчина с таким нарушением сперматогенеза до начала 90-х прошлого столетия был обречен на бездетность. Сегодня благодаря ВРТбесплодие у этих мужчин может быть преодолено, но об этом мы расскажем позднее в главе, посвященной микроманипуляциям, в частности, ИКСИ.
Перемещение сперматозоида осуществляется за счет движения его хвостика. Скорость движения сперматозоида не превышает 2-3 мм в минуту. Казалось бы, немного, однако, за 2-3 часа в женском половом тракте сперматозоиды проходят путь, в 80000 раз превышающий их собственные размеры! Будь на месте сперматозоида в этой ситуации человек, ему пришлось бы двигаться вперед со скоростью 60-70 км/час — то есть со скоростью автомобиля!
Сперматозоиды, находящиеся в яичке, неподвижны. Способность к движению они приобретают лишь, проходя по семявыводящим путям под воздействием жидкостей семявыводящих протоков и семенных пузырьков, секрета предстательной железы. В половых путях женщины сперматозоиды сохраняют подвижность в течение 3 — 4 суток, но оплодотворить яйцеклетку они должны в течение 24 часов. Весь процесс развития от стволовой клетки до зрелого сперматозоида длится примерно 72 дня. Однако, поскольку сперматогенез происходит непрерывно и в него одномоментно вступает громадное число клеток, то в яичках всегда есть большое количество спермиев, находящихся на разных этапах сперматогенеза, а запас зрелых сперматозоидов постоянно пополняется. Активность сперматогенеза индивидуальна, но с возрастом снижается.
В половых путях женщины сперматозоиды сохраняют подвижность в течение 3 — 4 суток, но оплодотворить яйцеклетку они должны в течение 24 часов. Весь процесс развития от стволовой клетки до зрелого сперматозоида длится примерно 72 дня. Однако, поскольку сперматогенез происходит непрерывно и в него одномоментно вступает громадное число клеток, то в яичках всегда есть большое количество спермиев, находящихся на разных этапах сперматогенеза, а запас зрелых сперматозоидов постоянно пополняется. Активность сперматогенеза индивидуальна, но с возрастом снижается.
Как мы уже говорили, яйцеклетки находятся в фолликулах яичника. В результате овуляции яйцеклетка попадает в брюшную полость, откуда она «вылавливается» фимбриями маточной трубы и переносится в просвет ее ампулярного отдела. Именно здесь происходит встреча яйцеклетки со сперматозоидами.
Какое же строение имеет зрелая яйцеклетка? Она довольно крупная и достигает 0,11-0,14 мм в диаметре.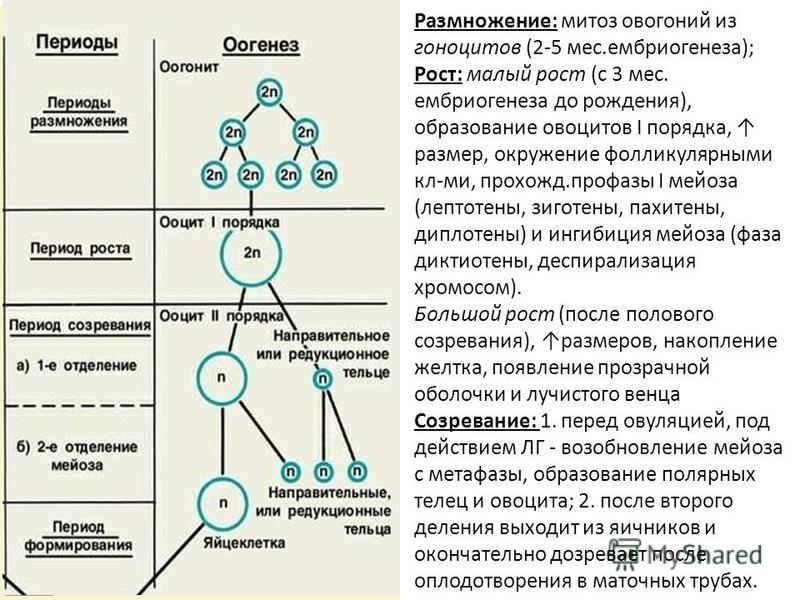 Сразу после овуляции яйцеклетка окружена скоплением мелких клеток и желатинообразной массой (так называемым лучистым венцом). Видимо, в таком виде фимбриям маточной трубы удобнее захватывать яйцеклетку. В просвете маточной трубы с помощью ферментов и механического воздействия (биения ресничек эпителия), происходит «очистка» яйцеклетки от лучистого венца. Окончательно освобождение яйцеклетки от лучистого венца происходит после встречи ее со сперматозоидами, которые буквально облепляют яйцеклетку. Каждый сперматозоид выделяет из акросомы фермент, растворяющий не только лучистый венец, но и действующий на оболочку самой яйцеклетки. Эта оболочка называется блестящей, так она выглядит под микроскопом. Выделяя фермент, все сперматозоиды стремятся оплодотворить яйцеклетку, но блестящая оболочка пропустит лишь один из них. Получается, что устремляясь к яйцеклетки, воздействуя на нее коллективно, сперматозоиды «расчищают дорогу» только для одного счастливчика. Отбором сперматозоида роль блестящей оболочки не ограничивается, на ранних стадиях развития эмбриона она поддерживает упорядоченное расположение его клеток (бластомеров).
Сразу после овуляции яйцеклетка окружена скоплением мелких клеток и желатинообразной массой (так называемым лучистым венцом). Видимо, в таком виде фимбриям маточной трубы удобнее захватывать яйцеклетку. В просвете маточной трубы с помощью ферментов и механического воздействия (биения ресничек эпителия), происходит «очистка» яйцеклетки от лучистого венца. Окончательно освобождение яйцеклетки от лучистого венца происходит после встречи ее со сперматозоидами, которые буквально облепляют яйцеклетку. Каждый сперматозоид выделяет из акросомы фермент, растворяющий не только лучистый венец, но и действующий на оболочку самой яйцеклетки. Эта оболочка называется блестящей, так она выглядит под микроскопом. Выделяя фермент, все сперматозоиды стремятся оплодотворить яйцеклетку, но блестящая оболочка пропустит лишь один из них. Получается, что устремляясь к яйцеклетки, воздействуя на нее коллективно, сперматозоиды «расчищают дорогу» только для одного счастливчика. Отбором сперматозоида роль блестящей оболочки не ограничивается, на ранних стадиях развития эмбриона она поддерживает упорядоченное расположение его клеток (бластомеров). В какой-то момент блестящая оболочка становится тесной, она разрывается и происходит хетчинг (от анг. hatching — «вылупление») — вылупление эмбриона. Эмбрион готов к имплантации в эндометрий.
В какой-то момент блестящая оболочка становится тесной, она разрывается и происходит хетчинг (от анг. hatching — «вылупление») — вылупление эмбриона. Эмбрион готов к имплантации в эндометрий.
Мейоз у грибов — Справочник химика 21
У человека и высших животных в результате мейоза образуются гаметы— яйцеклетка и сперматозоиды. При их слиянии возникает снова диплоидное ядро, из которого путем последовательных митозов развивается взрослый организм. Стадия мейоза характерна для жизненного цикла всех эукариот, однако отнюдь не всегда этот процесс протекает в период, аналогичный соответствующему моменту жизненного цикла человека. Так, клетки многих простейших и грибов обычно гаплоидны. После слияния двух гаплоидных ядер с образованием диплоидной клетки быстро наступает мейотическое деление, в результате которого вновь возникают гаплоидные особи. Чередование гаплоидных и диплоидных фаз жизненного цикла часто встречается у низших растений и примитивных животных. Например, гаметы папоротника падают на почву и [c.42]
Например, гаметы папоротника падают на почву и [c.42]
У животных мейоз происходит непосредственно перед образованием гамет. Иными словами, в клетках каждой особи данного вида на протяжении большей части жизненного цикла содержится 2N хромосом. Такие организмы называют диплоидными. Однако это отнюдь не всеобщее правило у эукариот, размножающихся половым путем. У многих протистов мейоз происходит сразу после образования зиготы, так что эти организмы на протяжении большей части жизненного цикла содержат N хромосом. Такие организмы называют гаплоидными. У многих водорослей и растений, а также у некоторых грибов и простейших происходит чередование гаплоидных и диплоидных поколений. При таком типе жизненного цикла из диплоидной зиготы возникает диплоидная особь, образующая путем мейоза гаплоидные клетки, предназначенные для бесполого размножения. Из каждой такой гаплоидной клетки возникает гаплоид- [c.
 50]
50]Основной орган спороношения аскомицетов-сумка, или аск (отсюда название этого класса грибов). В аске развиваются аскоспоры, и здесь же происходят кариогамия и мейоз. Стадия аска-конечная стадия полового размножения аск называют также совершенной или главной фор- [c.166]
Тетрадный анализ — анализ четырех клеток (гаплонтов), возникших из одной тетрады. Такой анализ у грибов и мхов позволяет получить интересные данные относительно расхождения генов и хромосом и их рекомбинации во время мейоза. [c.465]
В общем виде митоз и мейоз у грибов происходят так же, как у высших эукариот, но без разрушения ядерной мембраны и, по-видимому, без плотной компактизации хромосом, поскольку последние обычно неразличимы в световом микроскопе. [c.183]
У грибов различают вегетативное размножение, бесполое и половое. Последнее сопровождается мейозом. У некоторых гри- [c.133]
Какой бы механизм рекомбинации ни был предложен, в нем всегда должно быть учтено явление генной конверсии, или нереципрокной рекомбинации [220].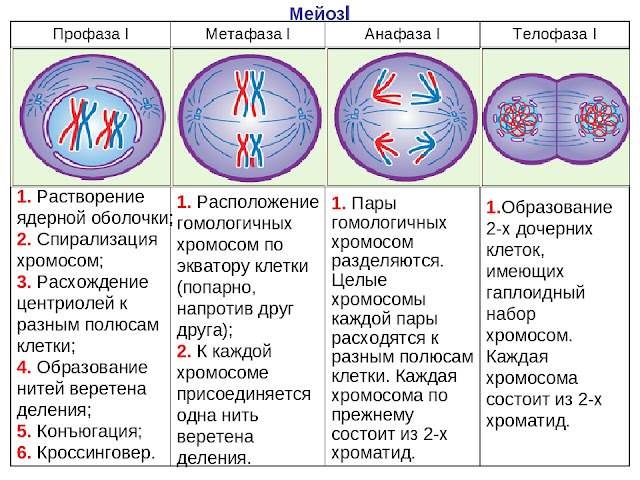 Это явление впервые было обнаружено при изучении генетики грибов, у которых можно отдельно исследовать каждый из четырех гаплоидных продуктов мейоза (тетрадный анализ, гл. 1, разд. Г, 2). Иногда вместо обычного менделевского отношения 2 2 для распределения генов в случае гетерозиготного локуса в потомстве наблюдали отношение 3 1. Это означает, что в одной из рекомбинантных. хромосом произошел возврат к родительскому типу. Механизм, лежащий в основе этого явления, может быть связан с неправильным спариванием оснований в гетеродуплексных участках. Чаще всего в точке,. [c.286]
Это явление впервые было обнаружено при изучении генетики грибов, у которых можно отдельно исследовать каждый из четырех гаплоидных продуктов мейоза (тетрадный анализ, гл. 1, разд. Г, 2). Иногда вместо обычного менделевского отношения 2 2 для распределения генов в случае гетерозиготного локуса в потомстве наблюдали отношение 3 1. Это означает, что в одной из рекомбинантных. хромосом произошел возврат к родительскому типу. Механизм, лежащий в основе этого явления, может быть связан с неправильным спариванием оснований в гетеродуплексных участках. Чаще всего в точке,. [c.286]
Половое размножение у грибов так же, как у других эукариот, включает слияние двух ядер. Такое слияние ядер у разных грибов происходит через различные промежутки времени после первого контакта между родительскими клетками. В процессе полового размножения можно различить три фазы. Первая фаза плазмогамия связана с соединением двух протопластов. Возникшая в результате этого клетка содержит два ядра. Эта пара ядер (дикарион) не обязательно сливается сразу же. Во время последующих делений клетки могут оставаться в дикариотической фазе. Оба ядра делятся при этом одновременно (сопряженное деление). Лишь позднее, часто только после образования плодового тела, происходит слияние обоих гаплоидных ядер (кариогамия) с образованием диплоидного ядра зиготы. За кариогамией следует мейоз, или редукционное деление, при котором число хромосом умень- [c.56]
Эта пара ядер (дикарион) не обязательно сливается сразу же. Во время последующих делений клетки могут оставаться в дикариотической фазе. Оба ядра делятся при этом одновременно (сопряженное деление). Лишь позднее, часто только после образования плодового тела, происходит слияние обоих гаплоидных ядер (кариогамия) с образованием диплоидного ядра зиготы. За кариогамией следует мейоз, или редукционное деление, при котором число хромосом умень- [c.56]
Основной орган спороношения аскомицетов сумка, или аск (отсюда название этого класса грибов). В аске развиваются аскоспоры, и здесь же происходят кариогамия и мейоз. Стадия аска — конечная стадия полового размножения аск называют также совершенной или главной формой плодоношения. Многие аскомицеты размножаются бесполым способом с помощью конидий. Эту форму плодоношения называют несовершенной. Существует много видов грибов, у которых известна только несовершенная стадия — образование конидий.
 Их называют несовершенными грибами, или дейтеромицетами. [c.65]
Их называют несовершенными грибами, или дейтеромицетами. [c.65]Базидиомицеты считаются наиболее высокоразвитой группой грибов. Характерный для них орган спороношения — базидия — по своему расположению (это клетка на конце гифы) соответствует аску. От бази-дии отшнуровываются, как правило, четыре одноядерные гаплоидные базидиоспоры. Подобно аскоспорам, они образуются в результате плазмогамии, кариогамии и мейоза. Два последних процесса протекают соответственно в аске или в базидии. [c.72]
Нельзя, однако, говорить о полном отсутствии полового процесса у несовершенных грибов. У них, так же, как у базидиомицетов или у других аскомицетов, доказано наличие парасексуального процесса. Плазмогамия, кариогамия и мейоз имеют место и у них, но не в определенных участках вегетативного тела и не на определенных стадиях развития. В норме первичный мицелий несовершенных грибов бывает гомокариотическим, т. е. содержит ядра только одного типа. В результате объединения протопластов, содержащих ядра разного типа, возникают гетерокарионы. Введенное в мицелий чужое ядро размножается, и образовавшиеся дочерние ядра распространяются по мицелию. Время от времени происходят кариогамия и мейоз. Таким образом, парасексуаль-ный цикл обеспечивает примерно столь же эффективную рекомбинацию ядерного материала, как и истинный половой процесс. [c.75]
Введенное в мицелий чужое ядро размножается, и образовавшиеся дочерние ядра распространяются по мицелию. Время от времени происходят кариогамия и мейоз. Таким образом, парасексуаль-ный цикл обеспечивает примерно столь же эффективную рекомбинацию ядерного материала, как и истинный половой процесс. [c.75]
У некоторых высших грибов половые различия имеют более сложную генотипическую основу по сравнению с описанными выше случаями. Так, у гриба Aleurodis us двудомность зависит от взаимодействия двух разных пар генов. Плодовые тела всегда представляют собой двойные гетерозиготы АаВЬ и мейоз приводит к образованию четырех типов спор и мицелиев АВ, АЬ, аВ и аЬ. [c.140]
Аскоспора—продукт мейоза у некоторых грибов (Азсо-тусе1ез). В мейозе образуются 4 гаплоидных ядра, которые после митотического деления дают 8 аскоспор. Эти споры заключены в асках. [c.451]
До сих пор мы рассматривали относительно простой случай, когда слияние гамет и мейоз непосредственно следуют друг за другом, т. е. когда первое деление диплоидной зиготы представляет собой редукционное деление. Поскольку при этом образуется 4 гаплоидных гона, то развившиеся из них организмы называют гаплобионтами почти всю свою жизнь они проводят с одинарным, гаплоидным, набором хромосом. Таковы, например, многие водоросли и грибы. [c.136]
е. когда первое деление диплоидной зиготы представляет собой редукционное деление. Поскольку при этом образуется 4 гаплоидных гона, то развившиеся из них организмы называют гаплобионтами почти всю свою жизнь они проводят с одинарным, гаплоидным, набором хромосом. Таковы, например, многие водоросли и грибы. [c.136]
Иначе обстоит дело у некоторых плесневых грибов, вроде аспергил-лов (А spergillus) и пенициллов Peni illium). У них очень редко (примерно один раз на миллион) происходит истинное слияние ядер. Здесь возможен и возврат к гаплоидному состоянию, на сей раз уже с рекомбинацией (с частотой примерно 1 i 1(Х)0), однако при этом мейоз отсутствует просто скопления хромосом случайно делятся на две части. [c.140]
Удобным объектом для изучения последовательных стадий рекомбинаций служат грибы-аскомицеты, так как у них продукты одного мейотического деления удерживаются вместе в одном аске. Более того, четыре гаплоидных ядра, образуемые в результате мейоза, распо- [c. 452]
452]
Конвдии. Вегетативные споры сумчатых и базидиальных грибов. Если конидии гаплоидные, то их слияние приводит к появлению диплоидных клеток, которые, пройдя мейоз, дают начало аскам. [c.309]
Одни из фундаментальных законов генетики гласит, что оба родителя вносят равный вклад в генетическую конституцию потомства, поскольку одни полный набор генов потомок получает от матери, а другой — от отца. Таким образом, когда из одной диплоидной клетки путем мейоза образуются четыре гаплоидные (разд. 15.2.1), в каждой из этих клеток ровно половину всех геиов должны составлять материнские гены, а другую половину — отцовские. Проверить справедливость этого утверждеиия для сложного организма в частности организма человека, разумеется, невозможно. К счастью, существуют и такие организмы, например грибы, у которых можно выделить и подвергнуть анализу все четыре дочерние клетки, образовавшиеся в результате мейоза из одной-единственной клетки. Подобный анализ показал, что из строгих генетических правил есть исключеиия. Иногда мейоз дает три копии материнского варианта (аллеля) данного гена и лишь одну копию отцовского аллеля, что свидетельствует о превращении одной из двух копий отцовского аллеля в копию материнского аллеля. Этот феномен получил название конверсии генов. Часто конверсия генов бывает связана с общей генетической рекомбинацией, и возможно, это явление играет немаловажную роль в эволюции некоторых генов (см. разд. 10.5.2). Полагают, что конверсия генов представляет собой прямое следствие действия двух механизмов -общей генетической рекомбинации и репарации ДНК. [c.309]
Иногда мейоз дает три копии материнского варианта (аллеля) данного гена и лишь одну копию отцовского аллеля, что свидетельствует о превращении одной из двух копий отцовского аллеля в копию материнского аллеля. Этот феномен получил название конверсии генов. Часто конверсия генов бывает связана с общей генетической рекомбинацией, и возможно, это явление играет немаловажную роль в эволюции некоторых генов (см. разд. 10.5.2). Полагают, что конверсия генов представляет собой прямое следствие действия двух механизмов -общей генетической рекомбинации и репарации ДНК. [c.309]
Принадлежность насекомых к империи ядерных соответствует признанию, что клетки их тела имеют оформленные ядра с хромосомами и другие органеллы, облеченные двухслойными биологическими мембранами. Наряду с другими свойствами, отличающими всех представителей империи (растения, грибы, животные), они противопоставляются доядерным (Prokaryota) специфичными формами клеточного деления (митоз, мейоз) и жизненными циклами, предполагающими развитие полового процесса и полового размножения. В соответствии с этим они демонстрируют признанные в генетике и биохимии стандарты, поскольку сами послужили главными объектами исследований такого рода. [c.55]
В соответствии с этим они демонстрируют признанные в генетике и биохимии стандарты, поскольку сами послужили главными объектами исследований такого рода. [c.55]
Образование гибридов у дрожжей, грибов и водорослей происходит в результате слияния клеток (копуляции). Если исходные клетки были гаплоидными (т. е. содержали только один набор хромосом), то в результате последующего слияния ядер (кариогамии) появится диплоидная клетка (зигота), несущая два набора хромосом в одном ядре (рис. 9). У некоторых микроорганизмов, например у Neurospora rassa, диплоидное ядро сразу же подвергается мейозу. Вегетативные диплоиды у этого организма неизвестны. В ходе мейоза каждая из хромосом продольно расщепляется и какое-то время состоит из двух сестринских хроматид. Гомологичные хромосомы образуют пары и обмениваются частями своих хроматид в результате кроссинго-вера (рассмотрение механизмов кроссинговера не входит в задачи этой книги).
 Затем формируются гаплоидные половые споры, каждая из которых может содержать новый набор генов, которыми различались родительские клетки, в результате рекомбинации генов одной и той же хромосомы, а также разных хромосом при перераспределении хромосомных пар (рис. 9). [c.84]
Затем формируются гаплоидные половые споры, каждая из которых может содержать новый набор генов, которыми различались родительские клетки, в результате рекомбинации генов одной и той же хромосомы, а также разных хромосом при перераспределении хромосомных пар (рис. 9). [c.84] Разнообразие жизненных циклов и типов несовместимости у грибов накладывает отпечаток и на приемы, используемые при их гибридологическом анализе. У одних грибов половой процесс осуществляется на основе гетерогамии, как у нейроспоры, что позволяет ставить реципрокные скрещивания. У других — на основе изогамии, как у дрожжей сахаромицетов. Наряду с половым размножением существует полный или неполный парасексуальный цикл в зависимости от вида грибов. Парасексуальный цикл — это процесс объединения и последующей рекомбинации генов на основе событий, происходящих в митозе, а не в мейозе, без участия оплодотворения половым путем. Остановимся только на двух подходах, внесших существенный вклад в разработку проблем общей генетики тетрадном анализе и генетическом анализе на основе парасексуального процесса. [c.185]
[c.185]
В цикле развития высших сумчатых грибов закономерно сменяют друг друга три ядерные фазы длительная гаплоидная, представленная долгоживущим мицелием, на котором развивается конидиальное спороношение, кратковременная дикарионтическая — от плазмогамии до кариогамии (существует в виде аскогенных гиф) и самая кратковременная диплоидная (диплоидное ядро материнской клетки сумки) — от кариогамии до мейоза. [c.177]
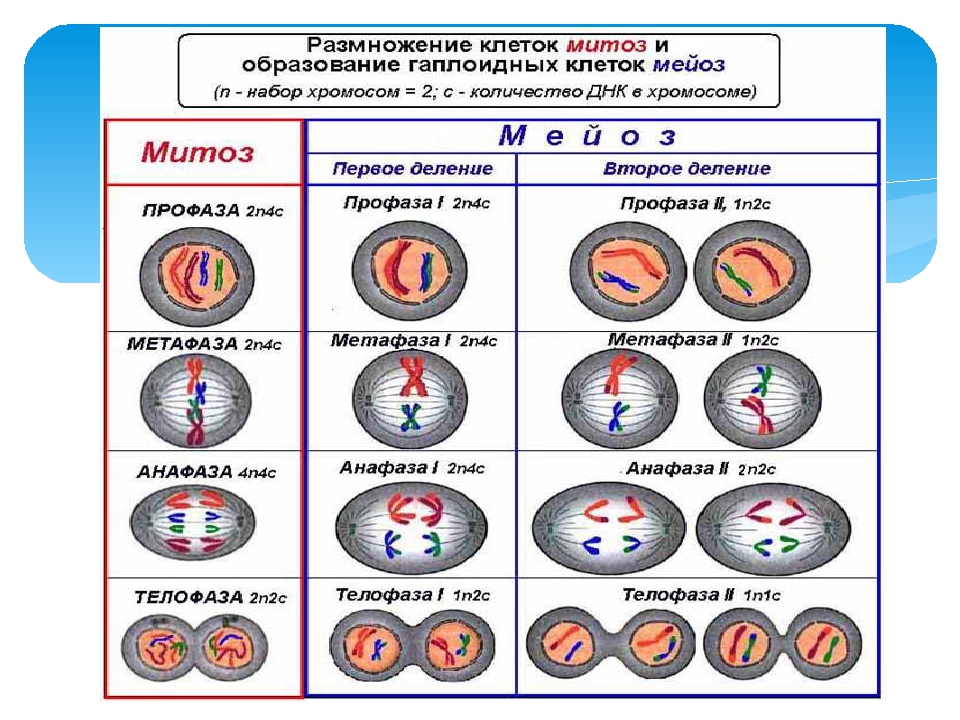
Сравнение мейоза и митоза — Биология муниципального колледжа Маунт-Худ 102
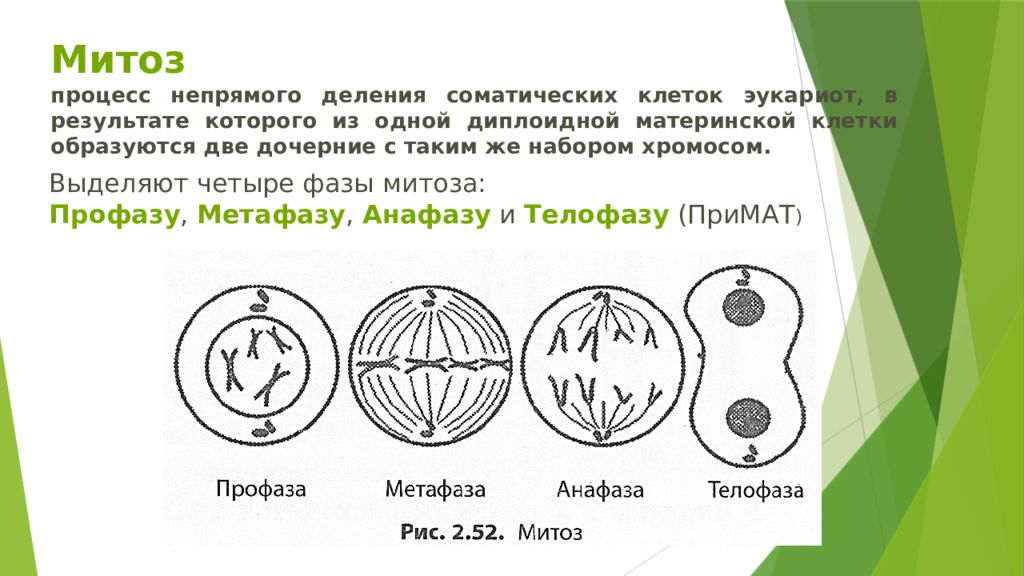
Митоз и мейоз, которые являются формами деления ядра в эукариотических клетках, имеют некоторое сходство, но также обнаруживают явные различия, которые приводят к их очень разным результатам. Митоз — это единичное ядерное деление, в результате которого образуются два ядра, обычно разделяемые на две новые клетки. Ядра, полученные в результате митотического деления, генетически идентичны исходным. У них одинаковое количество наборов хромосом: один в случае гаплоидных клеток и два в случае диплоидных клеток. С другой стороны, мейоз — это два деления ядра, в результате которых образуются четыре ядра, обычно разделенных на четыре новых клетки. Ядра, полученные в результате мейоза, никогда не бывают генетически идентичными, и они содержат только один набор хромосом — это половина количества исходной клетки, которая была диплоидной ( Рисунок 6 ).
С другой стороны, мейоз — это два деления ядра, в результате которых образуются четыре ядра, обычно разделенных на четыре новых клетки. Ядра, полученные в результате мейоза, никогда не бывают генетически идентичными, и они содержат только один набор хромосом — это половина количества исходной клетки, которая была диплоидной ( Рисунок 6 ).
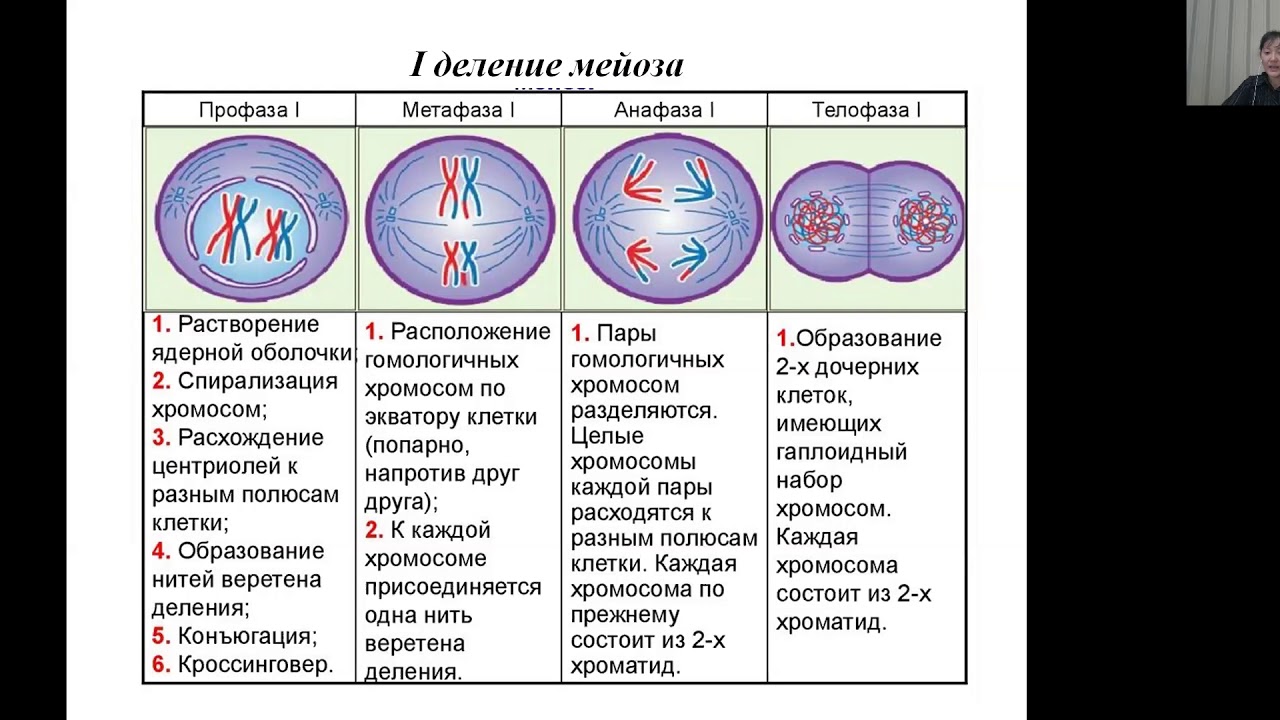
Различия в результатах мейоза и митоза происходят из-за различий в поведении хромосом во время каждого процесса. Большинство этих различий в процессах происходит в мейозе I, который представляет собой совершенно иное деление ядра, чем митоз.В мейозе I пары гомологичных хромосом становятся ассоциированными друг с другом, связаны друг с другом, испытывают хиазмы и кроссовер между сестринскими хроматидами и выстраиваются вдоль метафазной пластинки в тетрады с волокнами веретена с противоположных полюсов веретена, прикрепленными к каждой кинетохоре гомолога в гене. тетрада. Все эти события происходят только в мейозе I, а не в митозе.
Гомологичные хромосомы перемещаются к противоположным полюсам во время мейоза I, поэтому количество наборов хромосом в каждом будущем ядре сокращается с двух до одного.По этой причине мейоз I называют делением редукции . Такого снижения уровня плоидности при митозе нет.
Мейоз II намного больше аналогичен митотическому делению. В этом случае дублированные хромосомы (только один их набор) выстраиваются в центре клетки с разделенными кинетохорами, прикрепленными к волокнам веретена с противоположных полюсов. Во время анафазы II, как и в митотической анафазе, кинетохоры делятся, и одна сестринская хроматида притягивается к одному полюсу, а другая сестринская хроматида притягивается к другому полюсу.Если бы не тот факт, что были кроссоверы, два продукта каждого деления мейоза II были бы идентичны, как в митозе; напротив, они разные, потому что всегда имел место по крайней мере один кроссовер на хромосому. Мейоз II не является редукционным делением, потому что, хотя в полученных клетках меньше копий генома, остается один набор хромосом, как это было в конце мейоза I.
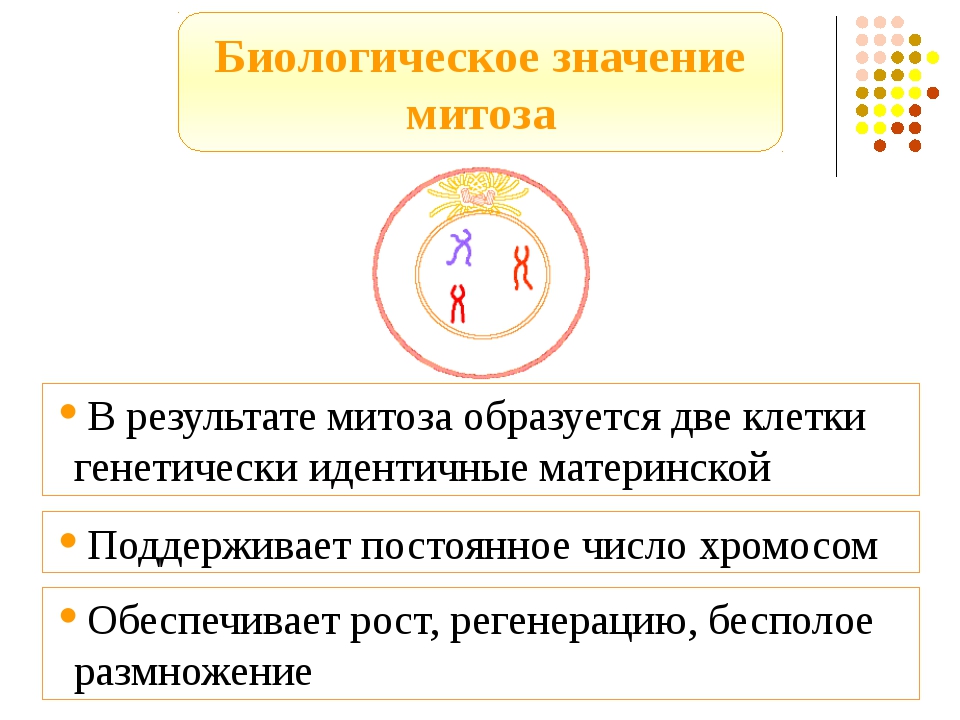
Клетки, продуцируемые митозом, будут функционировать в разных частях тела, обеспечивая рост или замену мертвых или поврежденных клеток.У некоторых организмов они могут даже участвовать в бесполом размножении. Клетки, продуцируемые мейозом в диплоидно-доминантном организме, таком как животное, будут участвовать только в половом размножении.
Рисунок 6: Мейозу и митозу предшествует один раунд репликации ДНК; однако мейоз включает два ядерных подразделения. Четыре дочерние клетки, полученные в результате мейоза, гаплоидны и генетически различны. Дочерние клетки, полученные в результате митоза, диплоидны и идентичны родительской клетке.Если не указано иное, изображения на этой странице лицензированы OpenStax по лицензии CC-BY 4.0.
OpenStax, Биология. OpenStax CNX. 27 мая 2016 г. http://cnx.org/contents/[email protected]: 1Q8z96mT @ 4 / Meiosis
11.1D: Сравнение мейоза и митоза
-
- Последнее обновление
- Сохранить как PDF
- Ключевые моменты
- Ключевые термины
- Сравнение мейоза и митоза
Митоз и мейоз имеют некоторое сходство, но также и некоторые различия, большинство из которых наблюдается во время мейоза I.
Задачи обучения
- Сравнить и сопоставить митоз и мейоз
Ключевые моменты
- По большей части при митозе диплоидные клетки делятся на две новые диплоидные клетки, тогда как в мейозе диплоидные клетки делятся на четыре новых гаплоидных клетки.
- В митозе дочерние клетки имеют такое же количество хромосом, что и родительская клетка, в то время как в мейозе дочерние клетки имеют половину количества хромосом по сравнению с родительскими.
- Дочерние клетки, продуцируемые митозом, идентичны, тогда как дочерние клетки, продуцируемые мейозом, отличаются, потому что произошел кроссинговер.
- События, которые происходят в мейозе, но не в митозе, включают спаривание гомологичных хромосом, кроссинговер и выстраивание вдоль метафазной пластинки в тетрадах.
- Мейоз II и митоз не являются делением редукции, как мейоз I, потому что количество хромосом остается тем же; поэтому мейоз II называют экваториальным делением.
- Когда гомологичные хромосомы разделяются и перемещаются к противоположным полюсам во время мейоза I, уровень плоидности снижается с двух до одного, что называется редукционным делением.
Ключевые термины
- редукционное деление : первое из двух делений мейоза, тип деления клетки
- плоидность : количество гомологичных наборов хромосом в клетке
- экваториальное деление : процесс ядерного деления, при котором каждая хромосома делится поровну, так что количество хромосом остается одинаковым от родительских клеток к дочерним
Сравнение мейоза и митоза
Митоз и мейоз являются формами деления ядра в эукариотических клетках.У них есть некоторые общие черты, но также есть явные различия, которые приводят к очень разным результатам. Целью митоза является регенерация, рост и бесполое размножение клеток, в то время как цель мейоза — производство гамет для полового размножения.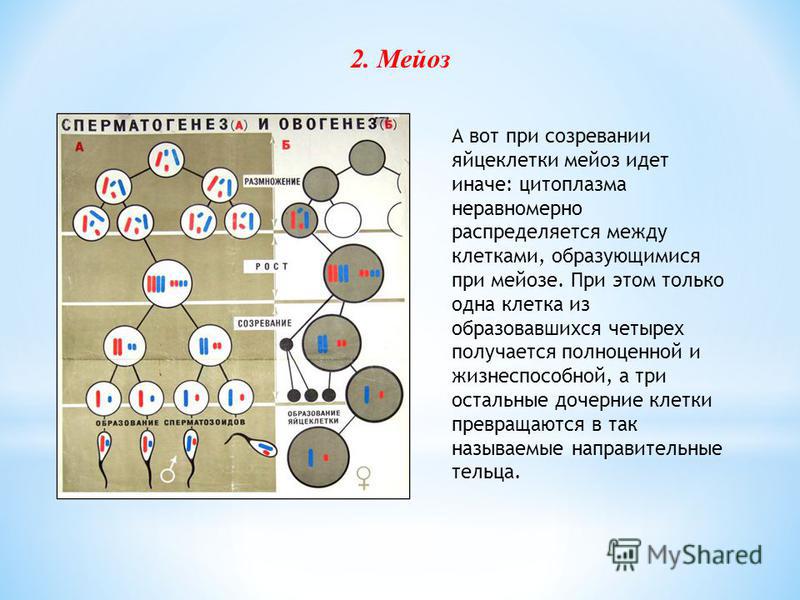 Митоз — это деление одного ядра, в результате которого образуются два ядра, которые обычно делятся на две новые дочерние клетки. Ядра, полученные в результате митотического деления, генетически идентичны исходному ядру. У них одинаковое количество наборов хромосом, один набор в случае гаплоидных клеток и два набора в случае диплоидных клеток.У большинства растений и всех видов животных, как правило, именно диплоидные клетки подвергаются митозу с образованием новых диплоидных клеток. Напротив, мейоз состоит из двух ядерных делений, в результате чего образуются четыре ядра, которые обычно делятся на четыре новых гаплоидных дочерних клетки. Ядра, полученные в результате мейоза, не являются генетически идентичными и содержат только один набор хромосом. Это половина количества хромосомных наборов в исходной клетке, которая является диплоидной.
Митоз — это деление одного ядра, в результате которого образуются два ядра, которые обычно делятся на две новые дочерние клетки. Ядра, полученные в результате митотического деления, генетически идентичны исходному ядру. У них одинаковое количество наборов хромосом, один набор в случае гаплоидных клеток и два набора в случае диплоидных клеток.У большинства растений и всех видов животных, как правило, именно диплоидные клетки подвергаются митозу с образованием новых диплоидных клеток. Напротив, мейоз состоит из двух ядерных делений, в результате чего образуются четыре ядра, которые обычно делятся на четыре новых гаплоидных дочерних клетки. Ядра, полученные в результате мейоза, не являются генетически идентичными и содержат только один набор хромосом. Это половина количества хромосомных наборов в исходной клетке, которая является диплоидной.
 Четыре дочерние клетки, полученные в результате мейоза, гаплоидны и генетически различны. Дочерние клетки, полученные в результате митоза, диплоидны и идентичны родительской клетке.
Четыре дочерние клетки, полученные в результате мейоза, гаплоидны и генетически различны. Дочерние клетки, полученные в результате митоза, диплоидны и идентичны родительской клетке. Основные различия между митозом и мейозом происходят в мейозе I. В мейозе I гомологичные пары хромосом становятся ассоциированными друг с другом и связываются вместе синаптонемным комплексом. Развиваются хиазмы и происходит кроссовер между гомологичными хромосомами, которые затем выстраиваются вдоль метафазной пластинки в тетрадах с кинетохорными волокнами от противоположных полюсов веретена, прикрепленными к каждой кинетохоре гомолога в тетраде.Все эти события происходят только в мейозе I.
Когда тетрада распадается и гомологичные хромосомы перемещаются к противоположным полюсам, уровень плоидности снижается с двух до одного. По этой причине мейоз I называют редукционным делением. Такого снижения уровня плоидности во время митоза нет.
Meiosis II намного больше похож на митотическое деление. В этом случае дублированные хромосомы (только один набор, поскольку гомологичные пары теперь разделены на две разные клетки) выстраиваются на метафазной пластине с разделенными кинетохорами, прикрепленными к кинетохорным волокнам с противоположных полюсов.Во время анафазы II и митотической анафазы кинетохоры делятся, и сестринские хроматиды, теперь называемые хромосомами, притягиваются к противоположным полюсам. Однако две дочерние клетки митоза идентичны, в отличие от дочерних клеток, продуцируемых мейозом. Они разные, потому что на каждой хромосоме был хотя бы один кроссовер. Мейоз II не является редукционным делением, потому что, хотя в полученных клетках меньше копий генома, остается один набор хромосом, как это было в конце мейоза I.Поэтому Мейоз II называют экваториальным делением.
В этом случае дублированные хромосомы (только один набор, поскольку гомологичные пары теперь разделены на две разные клетки) выстраиваются на метафазной пластине с разделенными кинетохорами, прикрепленными к кинетохорным волокнам с противоположных полюсов.Во время анафазы II и митотической анафазы кинетохоры делятся, и сестринские хроматиды, теперь называемые хромосомами, притягиваются к противоположным полюсам. Однако две дочерние клетки митоза идентичны, в отличие от дочерних клеток, продуцируемых мейозом. Они разные, потому что на каждой хромосоме был хотя бы один кроссовер. Мейоз II не является редукционным делением, потому что, хотя в полученных клетках меньше копий генома, остается один набор хромосом, как это было в конце мейоза I.Поэтому Мейоз II называют экваториальным делением.
ЛИЦЕНЗИИ И АТРИБУЦИИ
CC ЛИЦЕНЗИОННЫЙ КОНТЕНТ, ПРЕДЫДУЩИЙ РАЗДЕЛ
- Курирование и проверка. Источник : Boundless.com. Лицензия : CC BY-SA: Attribution-ShareAlike
CC ЛИЦЕНЗИОННОЕ СОДЕРЖАНИЕ, СПЕЦИАЛЬНАЯ АТРИБУЦИЯ
- Колледж OpenStax, Биология.
 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44468/latest…ol11448/latest . Лицензия : CC BY: Attribution
16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44468/latest…ol11448/latest . Лицензия : CC BY: Attribution - Колледж OpenStax, Процесс мейоза. 28 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest/ . Лицензия : CC BY: Attribution
- гаплоид. Источник : Викисловарь. Адрес: : http://en.wiktionary.org/wiki/haploid . Лицензия : CC BY-SA: Attribution-ShareAlike
- диплоид. Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/diploid .
 Лицензия : CC BY-SA: Attribution-ShareAlike
Лицензия : CC BY-SA: Attribution-ShareAlike - гамет. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/gamete . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44468/latest…7_00_02abc.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…ol11448/latest . Лицензия : CC BY: Attribution
- переход.
 Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/crossing%20over . Лицензия : CC BY-SA: Attribution-ShareAlike
Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/crossing%20over . Лицензия : CC BY-SA: Attribution-ShareAlike - тетрад. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/tetrad . Лицензия : CC BY-SA: Attribution-ShareAlike
- хроматида. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/chromatid . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44468/latest…7_00_02abc.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Процесс мейоза.
 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_02.jpg . Лицензия : CC BY: Attribution
16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_02.jpg . Лицензия : CC BY: Attribution - Колледж OpenStax, Процесс мейоза.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_01.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Процесс мейоза. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_03.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.16 октября 2013 г. Предоставлено : OpenStax CNX.
 Расположен по адресу : http://cnx.org/content/m44469/latest…ol11448/latest . Лицензия : CC BY: Attribution
Расположен по адресу : http://cnx.org/content/m44469/latest…ol11448/latest . Лицензия : CC BY: Attribution - мейоз II. Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/meiosis%20II . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44468/latest…7_00_02abc.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Процесс мейоза. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest.
 ..e_11_01_02.jpg . Лицензия : CC BY: Attribution
..e_11_01_02.jpg . Лицензия : CC BY: Attribution - Колледж OpenStax, Процесс мейоза.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_01.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Процесс мейоза. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_03.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Процесс мейоза.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_05.jpg .
 Лицензия : CC BY: Attribution
Лицензия : CC BY: Attribution - Колледж OpenStax, Процесс мейоза. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_04.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Безграничный. Источник : Безграничное обучение. Расположен по адресу : www.boundless.com//biology/de…orial-division . Лицензия : CC BY-SA: Attribution-ShareAlike
- плоидность.
 Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/ploidy . Лицензия : CC BY-SA: Attribution-ShareAlike
Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/ploidy . Лицензия : CC BY-SA: Attribution-ShareAlike - деление редуктора. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/reduction_division . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44468/latest…7_00_02abc.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Процесс мейоза. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.
 org/content/m44469/latest…e_11_01_02.jpg . Лицензия : CC BY: Attribution
org/content/m44469/latest…e_11_01_02.jpg . Лицензия : CC BY: Attribution - Колледж OpenStax, Процесс мейоза.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_01.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Процесс мейоза. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_03.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Процесс мейоза.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest.
 ..e_11_01_05.jpg . Лицензия : CC BY: Attribution
..e_11_01_05.jpg . Лицензия : CC BY: Attribution - Колледж OpenStax, Процесс мейоза. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_04.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Процесс мейоза.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44469/latest…e_11_01_06.jpg . Лицензия : CC BY: Attribution
Ядро во время митоза — Клетка
Уникальной особенностью ядра является то, что оно разбирается и переформируется каждый раз, когда большинство клеток делится. В начале митоза хромосомы конденсируются, ядрышко исчезает, а ядерная оболочка разрушается, что приводит к выбросу большей части содержимого ядра в цитоплазму. В конце митоза процесс меняется на противоположный: хромосомы деконденсируются, и ядерные оболочки заново формируются вокруг разделенных наборов дочерних хромосом. Глава 14 представляет всестороннее обсуждение митоза; в этом разделе мы рассмотрим механизмы, участвующие в разборке и воссоздании ядра. Этот процесс в значительной степени контролируется обратимым фосфорилированием и дефосфорилированием ядерных белков в результате действия протеинкиназы Cdc2, которая является критическим регулятором митоза во всех эукариотических клетках.
В конце митоза процесс меняется на противоположный: хромосомы деконденсируются, и ядерные оболочки заново формируются вокруг разделенных наборов дочерних хромосом. Глава 14 представляет всестороннее обсуждение митоза; в этом разделе мы рассмотрим механизмы, участвующие в разборке и воссоздании ядра. Этот процесс в значительной степени контролируется обратимым фосфорилированием и дефосфорилированием ядерных белков в результате действия протеинкиназы Cdc2, которая является критическим регулятором митоза во всех эукариотических клетках.
Растворение ядерной оболочки
В большинстве клеток разборка ядерной оболочки отмечает конец профазы митоза (). Однако такая разборка ядра не является универсальным признаком митоза и происходит не во всех клетках. Некоторые одноклеточные эукариоты (например, дрожжи) подвергаются так называемому закрытому митозу, при котором ядерная оболочка остается неповрежденной (). При закрытом митозе дочерние хромосомы мигрируют к противоположным полюсам ядра, которое затем делится на две части. Однако клетки высших эукариот обычно подвергаются открытому митозу, который характеризуется разрушением ядерной оболочки. Затем дочерние хромосомы мигрируют к противоположным полюсам митотического веретена, и новые ядра снова собираются вокруг них.
Однако клетки высших эукариот обычно подвергаются открытому митозу, который характеризуется разрушением ядерной оболочки. Затем дочерние хромосомы мигрируют к противоположным полюсам митотического веретена, и новые ядра снова собираются вокруг них.
Рисунок 8.29
Ядро во время митоза. Микрофотографии, иллюстрирующие прогрессивные стадии митоза в растительной клетке. Во время профазы хромосомы конденсируются, ядрышко исчезает, а ядерная оболочка разрушается.В метафазе конденсированные хромосомы (подробнее …)
Рисунок 8.30
Закрытый и открытый митоз. При закрытом митозе ядерная оболочка остается неповрежденной, а хромосомы мигрируют к противоположным полюсам веретена внутри ядра. При открытом митозе ядерная оболочка разрушается, а затем повторно формируется вокруг двух наборов разделенных (подробнее …)
Разборка ядерной оболочки, которая параллельна аналогичному разрушению эндоплазматического ретикулума, включает изменения во всех трех из них. его компоненты: ядерные мембраны фрагментируются на везикулы, комплексы ядерных пор диссоциируют, а ядерная пластинка деполимеризуется.Наиболее понятным из этих событий является деполимеризация ядерной пластинки — сети нитей, лежащих в основе ядерной мембраны. Ядерная пластинка состоит из волокнистых белков, ламинов, которые соединяются друг с другом, образуя нити. Разборка ядерной пластинки происходит в результате фосфорилирования ламинов, в результате чего филаменты распадаются на отдельные димеры ламинов (). Фосфорилирование ламинов катализируется протеинкиназой Cdc2, которая была представлена в главе 7 (см.) И будет подробно обсуждаться в главе 14 как центральный регулятор митоза.Cdc2 (а также другие протеинкиназы, активируемые в митотических клетках) фосфорилируют все различные типы ламинов, и было показано, что обработки изолированных ядер Cdc2 достаточно для индукции деполимеризации ядерной пластинки. Более того, потребность в фосфорилировании ламина при разрушении ядерной пластинки была продемонстрирована непосредственно путем создания мутантных ламинов, которые больше не могут фосфорилироваться.
его компоненты: ядерные мембраны фрагментируются на везикулы, комплексы ядерных пор диссоциируют, а ядерная пластинка деполимеризуется.Наиболее понятным из этих событий является деполимеризация ядерной пластинки — сети нитей, лежащих в основе ядерной мембраны. Ядерная пластинка состоит из волокнистых белков, ламинов, которые соединяются друг с другом, образуя нити. Разборка ядерной пластинки происходит в результате фосфорилирования ламинов, в результате чего филаменты распадаются на отдельные димеры ламинов (). Фосфорилирование ламинов катализируется протеинкиназой Cdc2, которая была представлена в главе 7 (см.) И будет подробно обсуждаться в главе 14 как центральный регулятор митоза.Cdc2 (а также другие протеинкиназы, активируемые в митотических клетках) фосфорилируют все различные типы ламинов, и было показано, что обработки изолированных ядер Cdc2 достаточно для индукции деполимеризации ядерной пластинки. Более того, потребность в фосфорилировании ламина при разрушении ядерной пластинки была продемонстрирована непосредственно путем создания мутантных ламинов, которые больше не могут фосфорилироваться. Когда гены, кодирующие эти мутантные ламины, были введены в клетки, их экспрессия, как было обнаружено, блокировала нормальное разрушение ядерной пластинки, когда клетки вступали в митоз.
Когда гены, кодирующие эти мутантные ламины, были введены в клетки, их экспрессия, как было обнаружено, блокировала нормальное разрушение ядерной пластинки, когда клетки вступали в митоз.
Рисунок 8.31
Растворение ядерной пластинки. Ядерная пластинка состоит из сети ламинатных нитей. Во время митоза Cdc2 и другие протеинкиназы фосфорилируют ламины, вызывая диссоциацию нитей на димеры свободных ламинов.
Вместе с растворением ядерной пластинки ядерная мембрана фрагментируется на пузырьки (). Ламины B-типа остаются связанными с этими пузырьками, но ламины A и C диссоциируют от ядерной мембраны и высвобождаются в виде свободных димеров в цитозоле.Это различие возникает из-за того, что ламины B-типа постоянно модифицируются путем добавления липидов (пренильных групп), тогда как C-концевые пренильные группы ламинов A- и C-типа удаляются протеолизом после их включения в пластинку. Комплексы ядерных пор также диссоциируют на субъединицы в результате фосфорилирования нескольких белков ядерных пор. Интегральные белки ядерной мембраны также фосфорилируются при митозе, и фосфорилирование этих белков может быть важным для образования пузырьков, а также для диссоциации ядерной мембраны как от хромосом, так и от ядерной пластинки.
Интегральные белки ядерной мембраны также фосфорилируются при митозе, и фосфорилирование этих белков может быть важным для образования пузырьков, а также для диссоциации ядерной мембраны как от хромосом, так и от ядерной пластинки.
Рисунок 8.32
Разрушение ядерной мембраны. Когда ядерная пластинка диссоциирует, ядерная мембрана фрагментируется на везикулы. Ламины B-типа остаются связанными с этими пузырьками, в то время как ламины A и C высвобождаются в виде свободных димеров.
Конденсация хромосом
Другим важным изменением ядерной структуры во время митоза является конденсация хромосом. Интерфазный хроматин, который уже упакован в нуклеосомы, конденсируется примерно в тысячу раз дальше, образуя компактные хромосомы, наблюдаемые в митотических клетках (2).Эта конденсация необходима для того, чтобы хромосомы могли перемещаться по митотическому веретену, не запутываясь и не ломаясь во время распределения по дочерним клеткам. ДНК в этом сильно конденсированном состоянии больше не может быть транскрибирована, поэтому весь синтез РНК останавливается во время митоза. Когда хромосомы конденсируются и транскрипция прекращается, ядрышко также исчезает.
Когда хромосомы конденсируются и транскрипция прекращается, ядрышко также исчезает.
Рисунок 8.33
Хромосомная конденсация. Электронная микрофотография, показывающая конденсацию отдельных хромосом во время профазы митоза.(К. Г. Мурти / Visuals Unlimited.)
Конденсированная ДНК в метафазных хромосомах, по-видимому, организована в большие петли, каждая из которых включает около ста килобаз ДНК, которые прикреплены к белковой структуре (см.). Несмотря на его фундаментальное значение, механизм конденсации хромосом во время митоза не изучен. Основной единицей структуры хроматина является нуклеосома, которая состоит из 146 пар оснований ДНК, обернутых вокруг гистонового ядра, содержащего по две молекулы гистонов h3A, h3B, h4 и h5 каждая (см.).Одна молекула гистона h2 связана с ДНК, когда она входит в каждую ядерную частицу нуклеосомы, и взаимодействия между этими молекулами h2 участвуют в сворачивании хроматина в более компактные структуры более высокого порядка. Гистон h2 является субстратом протеинкиназы Cdc2 и фосфорилируется во время митоза большинства клеток, что согласуется с его фосфорилированием, играющим роль в конденсации митотических хромосом. Однако недавние эксперименты показали, что фосфорилирование гистона h2 не требуется для конденсации хромосом, поэтому потенциальная роль фосфорилирования h2 неясна.Напротив, было обнаружено, что фосфорилирование гистона h4 необходимо для конденсации митотических хромосом, хотя механизм, с помощью которого фосфорилирование h4 влияет на конденсацию хромосом, еще предстоит выяснить.
Однако недавние эксперименты показали, что фосфорилирование гистона h2 не требуется для конденсации хромосом, поэтому потенциальная роль фосфорилирования h2 неясна.Напротив, было обнаружено, что фосфорилирование гистона h4 необходимо для конденсации митотических хромосом, хотя механизм, с помощью которого фосфорилирование h4 влияет на конденсацию хромосом, еще предстоит выяснить.
Недавние исследования также выявили белковые комплексы, называемые конденсинами, которые играют важную роль в конденсации хромосом. Конденсины необходимы для конденсации хромосом в экстрактах митотических клеток и, по-видимому, функционируют, оборачивая ДНК вокруг себя, тем самым уплотняя хромосомы в конденсированную митотическую структуру.Конденсины фосфорилируются и активируются протеинкиназой Cdc2, обеспечивая прямую связь между активацией Cdc2 и конденсацией митотических хромосом.
Реформирование интерфазного ядра
Во время завершения митоза (телофаза) вокруг разделенных наборов дочерних хромосом образуются два новых ядра (см. ). О деконденсации хромосом и повторной сборке ядерной оболочки, по-видимому, свидетельствует инактивация Cdc2, который был ответственен за инициирование митоза путем фосфорилирования клеточных белков-мишеней, включая ламины, гистон h4 и конденсины.Переход от метафазы к анафазе включает активацию убиквитин-опосредованной системы протеолиза, которая инактивирует Cdc2 за счет деградации его регуляторной субъединицы, циклина B (см.). Инактивация Cdc2 приводит к дефосфорилированию белков, которые были фосфорилированы при инициации митоза, что приводит к выходу из митоза и повторному образованию интерфазных ядер.
). О деконденсации хромосом и повторной сборке ядерной оболочки, по-видимому, свидетельствует инактивация Cdc2, который был ответственен за инициирование митоза путем фосфорилирования клеточных белков-мишеней, включая ламины, гистон h4 и конденсины.Переход от метафазы к анафазе включает активацию убиквитин-опосредованной системы протеолиза, которая инактивирует Cdc2 за счет деградации его регуляторной субъединицы, циклина B (см.). Инактивация Cdc2 приводит к дефосфорилированию белков, которые были фосфорилированы при инициации митоза, что приводит к выходу из митоза и повторному образованию интерфазных ядер.
Первым шагом в реформировании ядерной оболочки является связывание пузырьков, образовавшихся во время разрушения ядерной мембраны, с поверхностью хромосом ().Это взаимодействие мембранных везикул с хромосомами может быть опосредовано как ламинами, так и интегральными мембранными белками внутренней ядерной мембраны. Затем везикулы сливаются, образуя двойную мембрану вокруг хромосом. За этим следует повторная сборка комплексов ядерных пор, реформирование ядерной пластинки и деконденсация хромосом. Везикулы сначала сливаются, образуя мембраны вокруг отдельных хромосом, которые затем сливаются друг с другом, образуя единое целое ядро.
За этим следует повторная сборка комплексов ядерных пор, реформирование ядерной пластинки и деконденсация хромосом. Везикулы сначала сливаются, образуя мембраны вокруг отдельных хромосом, которые затем сливаются друг с другом, образуя единое целое ядро.
Рисунок 8.34
Восстановление ядерной оболочки. Первым шагом в повторной сборке ядерной оболочки является связывание мембранных везикул с хромосомами, которое может опосредоваться как интегральными мембранными белками, так и ламинами B-типа. Затем везикулы сливаются, ядерное (подробнее …)
Первоначальное восстановление ядерной оболочки вокруг конденсированных хромосом исключает цитоплазматические молекулы из вновь собранного ядра. Новое ядро затем способно расширяться за счет избирательного импорта ядерных белков из цитоплазмы.Поскольку сигналы ядерной локализации не отщепляются от белков, которые импортируются в ядро, те же ядерные белки, которые были выпущены в цитоплазму после разборки ядерной оболочки в начале митоза, могут быть повторно импортированы в новые ядра, сформированные после митоза. Ядрышко также переформируется, когда хромосомы деконденсируются и начинается транскрипция генов рРНК, завершая возврат из митоза в интерфазное ядро.
Ядрышко также переформируется, когда хромосомы деконденсируются и начинается транскрипция генов рРНК, завершая возврат из митоза в интерфазное ядро.
Клетка: Руководство по гистологии
Мейоз
На диаграмме слева показано деление мейотических клеток.Справа показано деление митотических (нормальных) клеток. Мейоз происходит от греческого слова «уменьшение».
- Диплоидное ядро содержит две пары хромосом каждого типа (аутосомы) вместе с половыми хромосомами (X и X или X и Y).
- Одна из этих хромосом происходит от мужского родителя (родительская хромосома), а одна — от женского (материнская хромосома).
- Хромосомы в этой паре называются гомологами — есть один отцовский и один материнский гомолог.
Мейоз |
Митоз |
|
| Гомологичные хромосомы дублируются и остаются вместе на веретене экватора как «сестринские хроматиды» или «биваленты» Плечи сестринских хроматид склеены белком, называемым когезином. |
Гомологичные хромосомы дублируются, но остаются отдельными. | |
| В длинной профазе мейотического деления 1 генетический материал может «переходить» между материнскими и отцовскими парами хромосом. В среднем у человека происходит 2-3 перехода на хромосому. Этот обмен генетическим материалом между материнскими и отцовскими хромосомами известен как «генетическая рекомбинация» Каждый кроссовер между двумя несестринскими хроматидами называется хиазмой (множественное число «хиазмы»)
Эти случайные перекрестные события перемешивают генетический материал каждой из хромосом в гаметах, помогая произвести разнообразное потомство. Хиазмы также важны для удержания вместе материнских и отцовских гомологов до тех пор, пока они не разделятся в анафазе 1. |
Кроссоверы между несестринскими хроматидами не происходят, так как они раздельны. |
|
| В анафазе мейотического деления 1 плечи сестринских хроматид становятся «отсоединенными», и сестринские хроматиды разделяются, так что одна клетка-двойник наследует одного в основном отцовского гомолога, а другая — в основном материнского гомолога. |
В анафазе одна из каждой пары хромосом переходит к каждой дочерней клетке, так что каждая дочерняя клетка наследует одну копию отцовской хромосомы и одну копию материнской хромосомы — как на диаграмме справа. | |
| Полученные клетки после мейотического деления 1 являются диплоидными, но в отличие от митотических делений: |
||
| Затем клетки вступают в следующий раунд клеточного деления без дупликации ДНК — мейотическое деление 2, чтобы сформировать гамету. |
Полученные клетки диплоид . Произведено 2 диплоидных клетки. |

У людей, которые имеют 23 пары хромосом, в результате случайного смешения 2 23 = 8.4 x 10 6 генетически различных гамет. Поскольку происходит некоторый обмен (или рекомбинация) генетического материала между парами гомологичных хромосом, фактическое количество генетически различных гамет может быть намного выше.
Иногда гомологичные хромосомы не разделяются нормально в мейозе 1 (нерасхождение). В результате могут образовываться эмбрионы, которые либо имеют дополнительные копии, либо не имеют копий одной или нескольких хромосом. Многие из этих эмбрионов умирают, но при состоянии «синдрома Дауна» появляется дополнительная копия хромосомы 21 (Х-хромосома).Дополнительная копия аутосомной хромосомы известна как «трисомия».
При образовании женских гамет (ооцитов) в результате мейоза образуется только одна яйцеклетка. После первого мейотического деления одна из дочерних клеток больше и способна образовывать яйцеклетку (вторичный ооцит), а вторая клетка мала (первое полярное тельце) и не может производить яйцеклетку. Точно так же после второго мейотического деления одна из дочерних клеток становится большой (яйцеклетка), а другая — маленькой (второе полярное тельце) и не может производить яйцеклетку.
Перейдите по этим ссылкам, чтобы узнать больше:
Ооциты
Сперма
Оплодотворение
Раннее развитие эмбриона мыши
hillis2e_ch07
Концепция 7.
 4: Мейоз сокращает вдвое содержание ядерной хромосомы и создает разнообразие
4: Мейоз сокращает вдвое содержание ядерной хромосомы и создает разнообразие
В Концепции 7.l мы описали роль и важность мейоза в половом размножении. Теперь посмотрим, как происходит упорядоченное и точное создание гаплоидных клеток.
Мейоз состоит из двух ядерных делений , которые уменьшают количество хромосом до гаплоидного числа.Хотя во время мейоза ядро делится дважды, ДНК реплицируется только один раз. В отличие от продуктов митоза гаплоидные клетки, продуцируемые мейозом, генетически отличаются друг от друга и от родительской клетки. РИСУНОК 7.11 сравнивает два процесса.
Рисунок 7.11: Митоз и мейоз: сравнение Мейоз включает два деления клеток, первое из которых сильно отличается от единичного деления митоза. Мейоз II подобен митозу в том, что центромеры разделяются во время анафазы, позволяя хроматидам двух гомологичных пар разделиться на четыре дочерние хромосомы, которые генетически отличаются от родительских хромосом.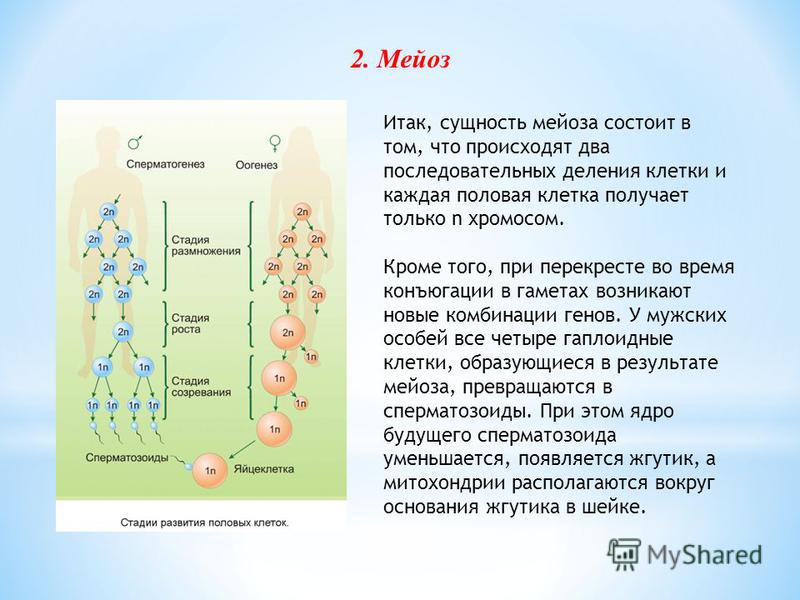
Чтобы понять процесс мейоза и его конкретные детали, полезно иметь в виду общие функции, которым мейоз призван служить:
- Для уменьшения числа хромосом с диплоидной до гаплоидной
- Чтобы гарантировать, что каждый из гаплоидных продуктов имеет полный набор хромосом
- Для создания генетического разнообразия продуктов (гамет)
События мейоза проиллюстрированы на РИСУНОК 7.12 . В оставшейся части этого раздела мы обсудим некоторые ключевые особенности, которые отличают мейоз от митоза.
Рисунок 7.12: Мейоз : создание гаплоидных клеток В мейозе два набора хромосом делятся между четырьмя дочерними клетками, каждая из которых имеет вдвое меньше хромосом, чем исходная клетка. Четыре гаплоидных клетки являются результатом двух последовательных делений ядра. На микрофотографиях показан мейоз в мужском репродуктивном органе лилии; диаграммы показывают соответствующие фазы в животной клетке. (В учебных целях хромосомы одного родителя исходного организма окрашены в синий цвет, а хромосомы другого родителя — в красный.)
(В учебных целях хромосомы одного родителя исходного организма окрашены в синий цвет, а хромосомы другого родителя — в красный.)
Перейти к ДЕЙСТВИЮ 7.4 Изображения Meiosis
PoL2e.com/ac7.4
Перейти к УПРАЖНЕНИЮ 7.2 Мейоз
PoL2e.com/at7.2
Деление мейоза уменьшает число хромосом
Как отмечалось выше, мейоз состоит из двух ядерных делений, мейоза I и мейоза II.Мейоз I :
характеризуют две уникальные особенности.- Гомологичные хромосомы объединяются и выстраиваются в линию по всей своей длине. В митозе такого спаривания не происходит.
- Гомологичные пары хромосом разделяют , но отдельные хромосомы, каждая из которых состоит из двух сестринских хроматид, остаются нетронутыми.
 (Хроматиды разделятся во время мейоза II.)
(Хроматиды разделятся во время мейоза II.)
Подобно митозу, мейозу I предшествует интерфаза с S-фазой, во время которой реплицируется каждая хромосома.В результате каждая хромосома состоит из двух сестринских хроматид. В конце мейоза I образуются два ядра, каждое с половиной исходных хромосом (по одному члену каждой гомологичной пары). Поскольку центромеры не разделились, эти хромосомы все еще двойные — состоят из двух сестринских хроматид. Сестринские хроматиды разделяются во время мейоза II , что составляет , а не , которому предшествует репликация ДНК. В результате продуктами мейоза I и II являются четыре клетки, каждая из которых содержит гаплоидное число хромосом.Но эти четыре клетки генетически не идентичны .
Пересечение и независимый ассортимент создают разнообразие
Диплоидный организм имеет два набора хромосом (2 n ): один набор происходит от его родителя-мужчины, а другой — от его родителя-женщины. По мере роста и развития организма его клетки подвергаются митотическим делениям. В митозе каждая хромосома ведет себя независимо от своего гомолога, и две ее хроматиды направляются к противоположным полюсам во время анафазы.Каждое дочернее ядро заканчивается идентичным набором из 2 n хромосом. В мейозе все обстоит иначе (см. Рис. 7.11).
По мере роста и развития организма его клетки подвергаются митотическим делениям. В митозе каждая хромосома ведет себя независимо от своего гомолога, и две ее хроматиды направляются к противоположным полюсам во время анафазы.Каждое дочернее ядро заканчивается идентичным набором из 2 n хромосом. В мейозе все обстоит иначе (см. Рис. 7.11).
Важным следствием мейоза является то, что четыре полученные клетки отличаются друг от друга генетически. Перетасовка генетического материала происходит за счет двух процессов: кроссинговера и независимого отбора.
Мейоз I начинается с длинной профазы I (первые три панели на рис. 7.12), во время которой хромосомы заметно меняются.Гомологичные хромосомы образуют пару, сцепляясь по своей длине в процессе, называемом синапсисом. (Этого не происходит при митозе.) Этот процесс спаривания длится от профазы I до конца метафазы I. Четыре хроматиды каждой пары гомологичных хромосом образуют тетраду или бивалент. Например, в клетке человека в конце профазы I имеется 23 тетрады, каждая из которых состоит из четырех хроматид. Четыре хроматиды происходят от двух партнеров в каждой гомологичной паре хромосом.
Четыре хроматиды происходят от двух партнеров в каждой гомологичной паре хромосом.
На протяжении профазы I и метафазы I хроматин продолжает скручиваться и уплотняться, а хромосомы становятся более конденсированными.В определенный момент пары гомологичных хромосом, кажется, отталкиваются друг от друга, особенно возле центромер, но они остаются прикрепленными. Х-образные точки крепления называются хиазмы (сингулярная хиазма, «крест»):
. Хиазма — это точка, в которой происходит обмен генетическим материалом между несестринскими хроматидами на гомологичных хромосомах — процесс, называемый кроссинговером ( РИСУНОК 7.13 ). Любая из четырех хроматид в тетраде может участвовать в этом обмене, и одна хроматида может обмениваться материалом более чем в одной точке по своей длине.Кроссинговер происходит вскоре после начала синапсиса, но хиазмы становятся видимыми только позже, когда гомологи отталкиваются друг от друга. Кроссинговер приводит к получению рекомбинантных хроматид и увеличивает генетическую изменчивость среди продуктов мейоза за счет перетасовки генетической информации между гомологичными парами хромосом. В Концепции 8.3 мы подробнее рассмотрим генетические последствия кроссинговера.
В Концепции 8.3 мы подробнее рассмотрим генетические последствия кроссинговера.
Рисунок 7.13: Кроссинговер формирует генетически разнообразные хромосомы Обмен генетическим материалом посредством кроссинговера приводит к новым комбинациям генетической информации о рекомбинантных хромосомах.Два разных цвета различают хромосомы, предоставленные родителями мужского и женского пола организма, чья клетка претерпевает мейоз.
Митоз редко длится более часа или двух, но мейоз может длиться намного дольше, чем . У мужчин-мужчин клеткам семенников, которые претерпевают мейоз, требуется около недели для профазы I и около месяца для всего мейотического цикла. У женщин профаза I начинается задолго до рождения женщины, во время ее раннего развития плода. Мейоз продолжается несколько десятилетий спустя, во время месячного цикла яичников, и завершается только после оплодотворения.
Помимо кроссинговера, мейоз является вторым источником генетического разнообразия.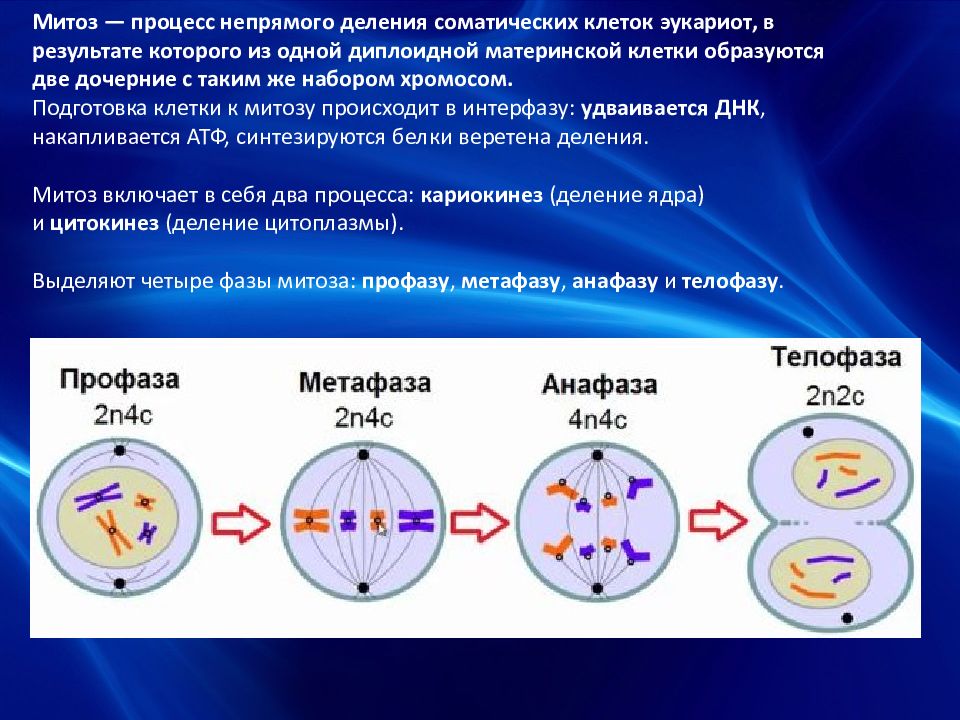 Это вопрос случайности, какой член гомологичной пары переходит к какой дочерней клетке в анафазе I. Например, рассмотрим диплоидный организм с двумя парами гомологичных хромосом (пары 1 и 2). Один член каждой пары происходил от мужчины-родителя организма (отцовские 1 и 2), а другой — от женского родителя (материнские 1 и 2). Когда клетки в этом организме подвергаются мейозу, конкретное дочернее ядро может получить отцовскую 1 и материнскую 2, отцовскую 2 и материнскую 1, обе материнские или обе отцовские хромосомы.Все зависит от того, как гомологичные пары выстраиваются в метафазу I. Это явление называется независимым ассортиментом.
Это вопрос случайности, какой член гомологичной пары переходит к какой дочерней клетке в анафазе I. Например, рассмотрим диплоидный организм с двумя парами гомологичных хромосом (пары 1 и 2). Один член каждой пары происходил от мужчины-родителя организма (отцовские 1 и 2), а другой — от женского родителя (материнские 1 и 2). Когда клетки в этом организме подвергаются мейозу, конкретное дочернее ядро может получить отцовскую 1 и материнскую 2, отцовскую 2 и материнскую 1, обе материнские или обе отцовские хромосомы.Все зависит от того, как гомологичные пары выстраиваются в метафазу I. Это явление называется независимым ассортиментом.
Обратите внимание, что из четырех возможных исходов на рисунке выше только два дочерних ядра получают либо все материнские, либо все отцовские хромосомы (за исключением материала, обмененного посредством кроссинговера). Чем больше количество хромосом, тем ниже вероятность восстановления исходных родительских комбинаций и, следовательно, тем выше потенциал генетического разнообразия.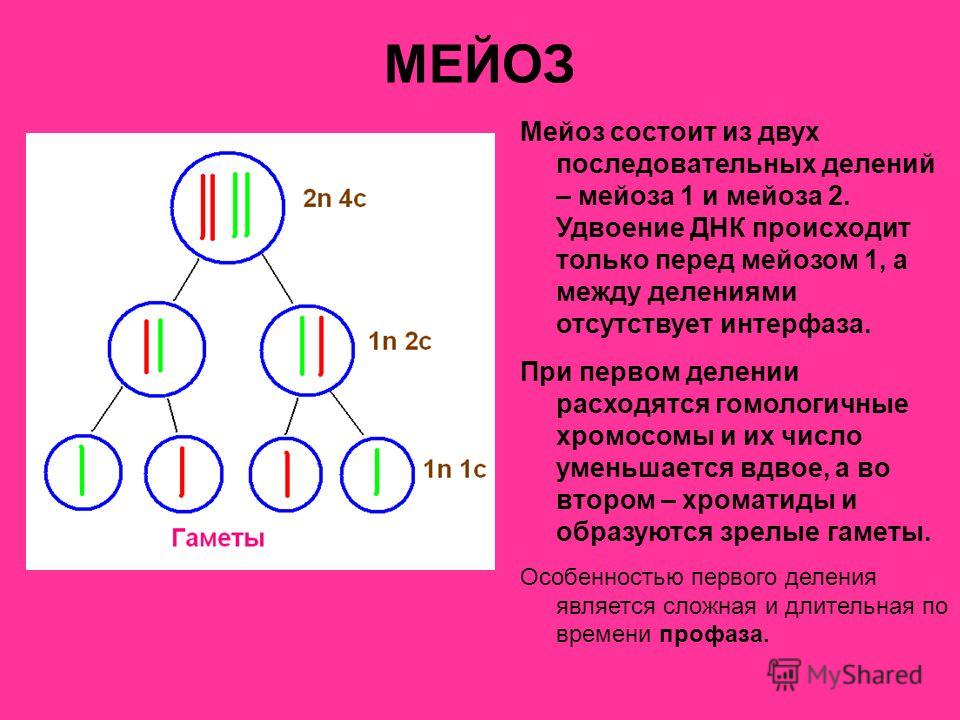 Большинство видов диплоидных организмов имеют более двух пар хромосом. У человека с 23 парами хромосом 2 23 (8 388 608) различных комбинаций материнских и отцовских хромосом могут быть произведены только с помощью механизма независимого набора! Принимая во внимание дополнительную генетическую перетасовку, обеспечиваемую кроссинговером, количество возможных комбинаций практически бесконечно. Скрещивание и независимый ассортимент, а также процессы, приводящие к мутациям, обеспечивают генетическое разнообразие, необходимое для эволюции путем естественного отбора.
Большинство видов диплоидных организмов имеют более двух пар хромосом. У человека с 23 парами хромосом 2 23 (8 388 608) различных комбинаций материнских и отцовских хромосом могут быть произведены только с помощью механизма независимого набора! Принимая во внимание дополнительную генетическую перетасовку, обеспечиваемую кроссинговером, количество возможных комбинаций практически бесконечно. Скрещивание и независимый ассортимент, а также процессы, приводящие к мутациям, обеспечивают генетическое разнообразие, необходимое для эволюции путем естественного отбора.
Мы видели, насколько мейоз I принципиально отличается от митоза. Однако мейоз II похож на митоз в том, что он включает разделение хроматид на дочерние ядра (см. Шаги 7–11 на рис. 7.12). Конечными продуктами мейоза I и мейоза II являются четыре гаплоидные дочерние клетки, каждая с одним набором ( n ) хромосом.
Мейотические ошибки приводят к аномальным структурам и номерам хромосом
Мейоз — сложный процесс, и иногда что-то идет не так. Например, хромосомы могут сломаться, гомологи могут не разделиться в анафазе I или хроматиды могут не разделиться в анафазе II. Гаметы, образованные в результате мейотических ошибок, несут аномальные хромосомы, и когда аномальные хромосомы принимают участие в оплодотворении, последствия для потомства могут быть значительными.
Например, хромосомы могут сломаться, гомологи могут не разделиться в анафазе I или хроматиды могут не разделиться в анафазе II. Гаметы, образованные в результате мейотических ошибок, несут аномальные хромосомы, и когда аномальные хромосомы принимают участие в оплодотворении, последствия для потомства могут быть значительными.
Иногда гомологичная пара хромосом не может разделиться (не может «разъединиться») в анафазе I или пара хроматид не может разделиться в анафазе II. Этот отказ от разделения называется нерасхождением.Если пара хромосом не может разделиться в анафазе I, каждое из четырех дочерних ядер окажется с обоими членами этой гомологичной пары, а два других не будут иметь ни одного члена пары. Если нерасхождение происходит в анафазе II, только два из четырех дочерних ядер будут затронуты: у одного будет дополнительная хромосома, а у другого — на один меньше, чем полный набор хромосом.
На примере человека, если во время анафазы I два гомолога хромосомы 10 не разделятся, половина гамет будет иметь две копии хромосомы 10, всего 24 хромосомы вместо 23. Если одна из этих гамет сливается с нормальной гаметой во время оплодотворения, у зиготы будет 47 (23 + 24) хромосом с тремя копиями хромосомы 10. Состояние аномального числа хромосом называется анеуплоидией ; с одной дополнительной хромосомой называется трисомией, а отсутствие одной хромосомы называется моносомией ( РИСУНОК 7.14 ).
Если одна из этих гамет сливается с нормальной гаметой во время оплодотворения, у зиготы будет 47 (23 + 24) хромосом с тремя копиями хромосомы 10. Состояние аномального числа хромосом называется анеуплоидией ; с одной дополнительной хромосомой называется трисомией, а отсутствие одной хромосомы называется моносомией ( РИСУНОК 7.14 ).
Рисунок 7.14: Нерасхождение ведет к анеуплоидии Нерасхождение, показанное здесь, происходит в мейозе I, приводит к анеуплоидии: одна или несколько хромосом либо отсутствуют, либо присутствуют в избытке.Как правило, анеуплоидия смертельна для развивающегося эмбриона.
По неясным причинам анеуплоидия является распространенным и опасным заболеванием у людей. Около 10–30 процентов всех зачатий демонстрируют анеуплоидию, но большинство эмбрионов, которые развиваются из таких зигот, не доживают до рождения, а те, которые действительно умирают, часто умирают в возрасте до 1 года. По крайней мере, пятая часть всех признанных беременностей у людей прерывается самопроизвольно (выкидышами) в течение первых 2 месяцев, в основном из-за трисомии и моносомии. Фактическая доля самопроизвольно прерванных беременностей, безусловно, выше, потому что самые ранние из них часто остаются нераспознанными. Наиболее распространенной формой анеуплоидии у людей является трисомия 16 (три копии хромосомы 16), но почти ни один из этих эмбрионов не доживает до рождения. Среди немногих анеуплоидий, способствующих выживанию, — синдром Дауна — трисомия 21. Такие люди обычно имеют умственную отсталость, но могут вести долгую и продуктивную жизнь.
Фактическая доля самопроизвольно прерванных беременностей, безусловно, выше, потому что самые ранние из них часто остаются нераспознанными. Наиболее распространенной формой анеуплоидии у людей является трисомия 16 (три копии хромосомы 16), но почти ни один из этих эмбрионов не доживает до рождения. Среди немногих анеуплоидий, способствующих выживанию, — синдром Дауна — трисомия 21. Такие люди обычно имеют умственную отсталость, но могут вести долгую и продуктивную жизнь.
Большинство организмов либо диплоидны (например, большинство животных), либо гаплоидны (например, большинство грибов).При некоторых обстоятельствах могут образовываться триплоидные (3 n ), тетраплоидные (4 n ) или полиплоидные ядра более высокого порядка. Это может происходить по-разному. Например, может быть дополнительный раунд репликации ДНК, предшествующий мейозу, или может не быть веретена, сформированного в мейозе II. Полиплоидия встречается в природе у некоторых видов животных и многих растений.
ССЫЛКА
Полиплоидия может привести к репродуктивной изоляции (неспособность двух особей произвести плодовитое потомство) и, вероятно, привела к видообразованию — эволюции новых видов — как описано в Концепции 17 .3
Диплоидное ядро может подвергаться нормальному мейозу, потому что существует два набора хромосом, образующих гомологичные пары, которые разделяются во время анафазы I. Точно так же тетраплоидное ядро имеет четное количество хромосом каждого типа, поэтому каждая хромосома может спариваться со своим гомологом. . Однако триплоидное ядро не может подвергнуться нормальному мейозу, потому что одна треть хромосом не имеет партнеров. Полиплоидия имеет значение для сельского хозяйства, особенно в производстве гибридных растений.Например, при селекции пшеницы необходимо учитывать плоидность (количество хромосом в ядре), поскольку существуют диплоидные, тетраплоидные и гексаплоидные сорта пшеницы.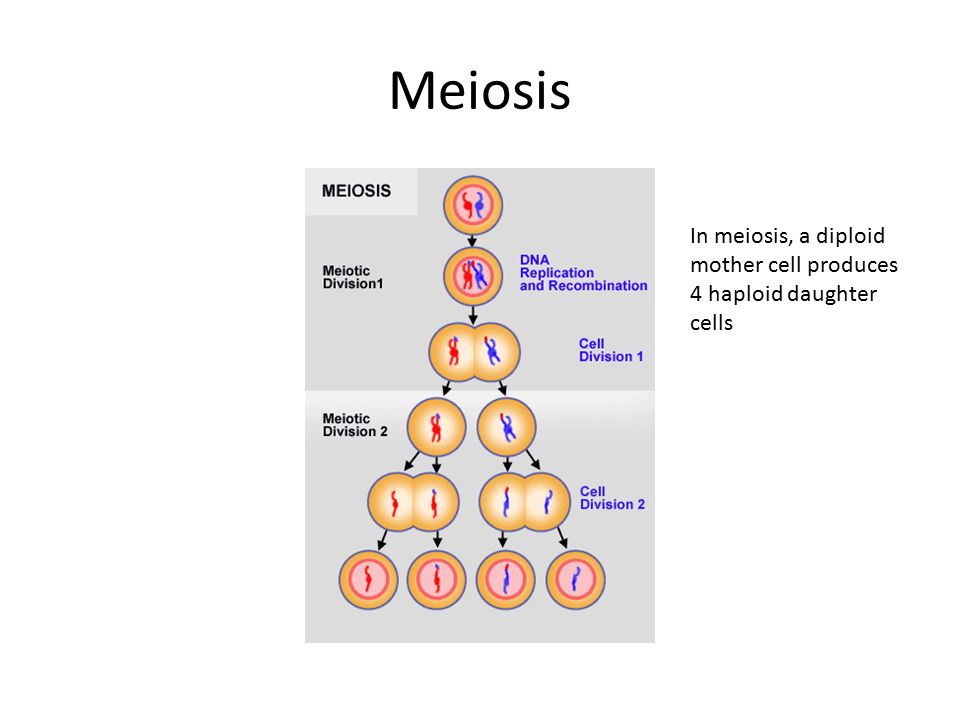 Полиплоидия может быть желательным признаком сельскохозяйственных культур и декоративных растений, поскольку она часто приводит к появлению более крепких растений с более крупными цветами, плодами и семенами. Кроме того, желательны триплоидные сорта плодов, потому что они бесплодны и, следовательно, не имеют косточек.
Полиплоидия может быть желательным признаком сельскохозяйственных культур и декоративных растений, поскольку она часто приводит к появлению более крепких растений с более крупными цветами, плодами и семенами. Кроме того, желательны триплоидные сорта плодов, потому что они бесплодны и, следовательно, не имеют косточек.
ПРИМЕНЯЙТЕ КОНЦЕПЦИЮ: Мейоз сокращает вдвое содержание ядерных хромосом и создает разнообразие
Клетки диплоидного организма (2 n = 4) показаны на диаграммах в процессе деления.Для каждой диаграммы укажите тип деления клетки (митоз или мейоз), фазу деления и любые изображенные особые состояния.
Во время кроссинговера в мейозе I хроматиды из гомологичных пар хромосом разрываются и воссоединяются. Иногда это может происходить между негомологичными хромосомами. Результатом является транслокация, которая довольно часто встречается даже в митотических клетках.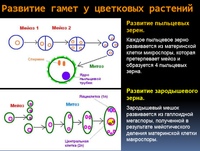 Как мы укажем в нашем обсуждении экспрессии генов и ее регуляции в главах 10 и 11, расположение генов по отношению к другим последовательностям ДНК важно, и транслокации могут иметь огромное влияние на экспрессию генов.
Как мы укажем в нашем обсуждении экспрессии генов и ее регуляции в главах 10 и 11, расположение генов по отношению к другим последовательностям ДНК важно, и транслокации могут иметь огромное влияние на экспрессию генов.
Пример транслокации, которая, как известно, происходит у людей, — это обмен материала между хромосомами 9 и 22:
В этом случае часть последовательности гена BCR на хромосоме 22 оказывается рядом с частью последовательности гена ABL , которая была перемещена с хромосомы 9. Если транслокация происходит в митотической клетке, образующей белые кровяные тельца, Результатом такого сочетания является форма лейкемии, рака белых кровяных телец.
Транслокация, происходящая во время мейоза, может переноситься образовавшимися гаметами и передаваться потомству при оплодотворении.
КОНЦЕПЦИЯ ПРОВЕРКИ 7.4
- Как кроссинговер и независимый ассортимент во время мейоза приводят к генетически различающимся дочерним ядрам?
- В чем разница между мейозом и митозом?
- У позвоночного животного диплоидное число 6.
 Сколько хромосом присутствует в следующих клетках: Гамета? Гамета с моносомией хромосомы 2? Клетка кожи? Сперматозоид в мейотической анафазе II?
Сколько хромосом присутствует в следующих клетках: Гамета? Гамета с моносомией хромосомы 2? Клетка кожи? Сперматозоид в мейотической анафазе II?
Существенная роль клеточного деления в сложных эукариотах состоит в замене погибающих клеток.Что заставляет клетки умирать?
Telophase — обзор | Темы ScienceDirect
D ТЕЛОФАЗА
Последняя стадия митоза, телофаза, во многих отношениях противоположна профазе. Когда два набора половинных хромосом достигают своего назначения, веретено исчезает, и вокруг каждого нового ядра образуется ядерная мембрана. Хромосомы удлиняются за счет деспирализации или набухания (Mazia, 1961) и исчезают из поля зрения. Два интерфазных ядра напоминают ядра родительской клетки, из которой они были сформированы, за исключением того, что они меньше.Они растут за счет синтеза ядерного материала во время межфазной границы.
Форма отдельных хромосом — важный аспект описательной цитологии. Но поскольку характерной чертой деления клеток является то, что хромосомы подвергаются циклам конденсации с последующей деконденсацией, очевидно, что при описании хромосом необходимо указать стадию деления клетки, на которой они происходят; и, кроме того, поскольку фиксация и другие виды обработки будут влиять на морфологию хромосом, метод, которым был изготовлен препарат, также имеет значение.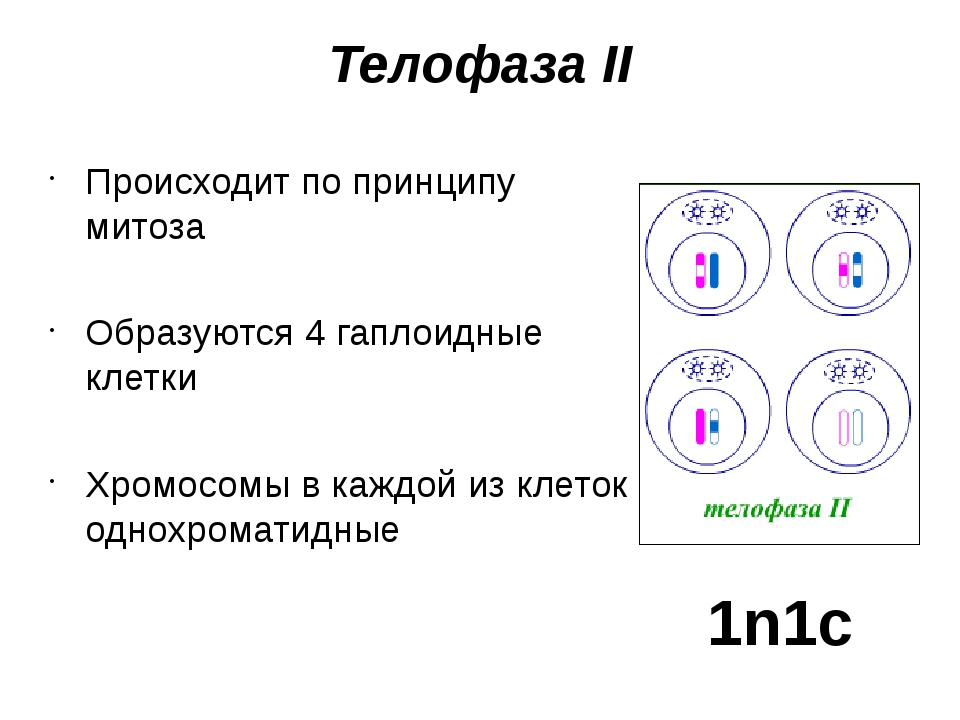
Для многих целей обычно выбирают метафазные хромосомы, поскольку на этой стадии хромосомы достигли максимальной степени конденсации и поэтому лучше всего подходят для индивидуального изучения.
Чтобы облегчить рассредоточение хромосом внутри клетки и минимизировать перекрытие, в настоящее время принято, особенно при изучении хромосом млекопитающих, погружать нефиксированные клетки в гипотонический солевой раствор, который вызывает их набухание за счет поглощения вода (Сюй, 1952).Также распространенной практикой является воздействие на клетки лекарственного препарата колхицина, который ингибирует формирование митотического веретена, в результате чего хромосомы, неспособные перейти в анафазу, накапливаются в метафазе. Присутствие колхицина также усиливает сокращение хромосом и раздвигает две хроматиды хромосомы друг от друга (см. Clarke, 1962).
Еще одним предварительным условием для получения чистых препаратов хромосом является обеспечение того, чтобы все хромосомы лежали в одной плоскости. В современной практике для этого обычно используется один из двух методов. Один из способов — раздавить ячейку между предметным стеклом и покровным стеклом, приложив давление. Этот метод был использован Бовери в 1890 году, но в начале двадцатого века его заменила практика резки тонких срезов. Техника сквоша была вновь представлена Хайцем (1935b). Второй метод заключается в прижатии клеток к предметному стеклу путем их сушки в токе воздуха. Этот метод был только недавно внедрен для изучения хромосом (Tjio and Puck, 1958), но ранее использовался для приготовления препаратов клеток крови.
В современной практике для этого обычно используется один из двух методов. Один из способов — раздавить ячейку между предметным стеклом и покровным стеклом, приложив давление. Этот метод был использован Бовери в 1890 году, но в начале двадцатого века его заменила практика резки тонких срезов. Техника сквоша была вновь представлена Хайцем (1935b). Второй метод заключается в прижатии клеток к предметному стеклу путем их сушки в токе воздуха. Этот метод был только недавно внедрен для изучения хромосом (Tjio and Puck, 1958), но ранее использовался для приготовления препаратов клеток крови.
Иллюстрация человеческих хромосом в митотической метафазе, полученная с помощью современных технологий, показана на рис. 8.1. Сравнение с рис. 2.1c, на котором показана аналогичная клетка, которой просто позволили расти на покровном стекле, проясняет, что, хотя колхицин, гипотонический солевой раствор и раздавливание разрушают структуру клетки, это значительно облегчает изучение отдельных хромосом.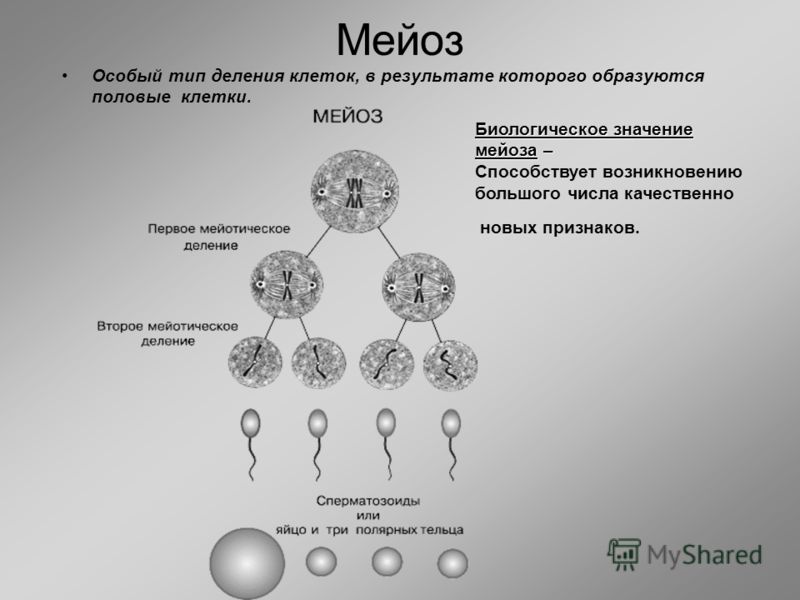 Такая заготовка — артефакт в хорошем смысле слова, настоящее произведение искусства.В результате можно подсчитать хромосомы и сравнить их индивидуальные формы. Метод анализа хромосом человека был описан Турпином и Леженом (1965).
Такая заготовка — артефакт в хорошем смысле слова, настоящее произведение искусства.В результате можно подсчитать хромосомы и сравнить их индивидуальные формы. Метод анализа хромосом человека был описан Турпином и Леженом (1965).
Наиболее очевидное различие между отдельными хромосомами заключается в размере, хотя следует иметь в виду, что размер хромосом — понятие относительное, сравнивая длины хромосом в одной и той же клетке. Как упоминалось ранее, фактический размер хромосом варьируется не только на разных стадиях клеточного деления, но также может отличаться на одной и той же стадии в разных клетках.Но относительный размер хромосом — это постоянная характеристика, которая имеет первостепенное значение в задаче идентификации отдельных хромосом. Форма хромосом определяется положением центромеры.
Было много дискуссий относительно положения центромеры и используемой номенклатуры (см. Levan et al ., 1964). Отчасти это связано с вопросом, может ли центромера располагаться на крайнем конце хромосомы или всегда есть короткое плечо, которое может быть или не быть видимым.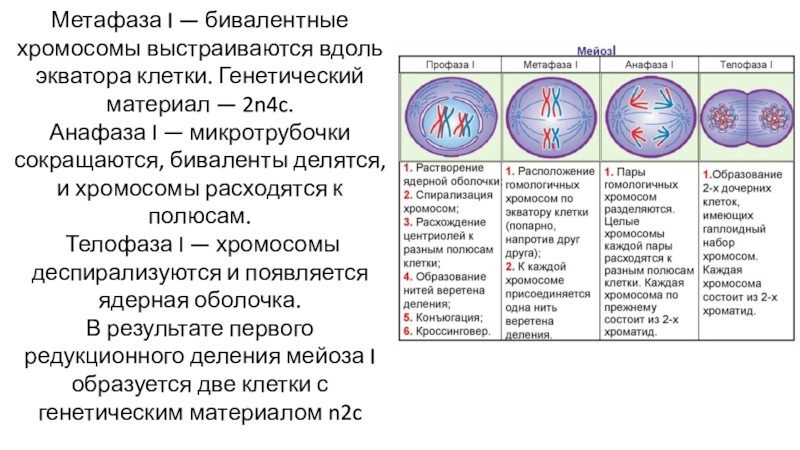 Однако с практической точки зрения наиболее важным различием является то, находится ли центромера где-то около середины хромосомы, разделяя ее на два плеча, или центромера находится так близко к одному концу, что второе плечо, если оно есть, , можно практически пренебречь. Уайт (1945) назвал последний тип хромосомы «акроцентрическим» и использовал термин «метацентрический» для хромосом, имеющих два плеча. Эти термины будут использоваться в этой книге. Многие авторы ограничивают термин «метацентрические» теми хромосомами, в которых центромера находится точно посередине, и используют термин «субметацентрические» для тех двух плеч, в которых центромера находится ближе к одному концу.Этот термин не будет использоваться, потому что с увеличением точности измерений будет меньше хромосом, в которых центромера окажется точно посередине, и, таким образом, подавляющему большинству хромосом придется дать более громоздкий термин «субметацентрические». . » Чтобы различать различные типы метацентрических хромосом, необходимо дать количественные измерения относительных длин плеч, хотя, конечно, следует иметь в виду погрешность, присущую таким измерениям.
Однако с практической точки зрения наиболее важным различием является то, находится ли центромера где-то около середины хромосомы, разделяя ее на два плеча, или центромера находится так близко к одному концу, что второе плечо, если оно есть, , можно практически пренебречь. Уайт (1945) назвал последний тип хромосомы «акроцентрическим» и использовал термин «метацентрический» для хромосом, имеющих два плеча. Эти термины будут использоваться в этой книге. Многие авторы ограничивают термин «метацентрические» теми хромосомами, в которых центромера находится точно посередине, и используют термин «субметацентрические» для тех двух плеч, в которых центромера находится ближе к одному концу.Этот термин не будет использоваться, потому что с увеличением точности измерений будет меньше хромосом, в которых центромера окажется точно посередине, и, таким образом, подавляющему большинству хромосом придется дать более громоздкий термин «субметацентрические». . » Чтобы различать различные типы метацентрических хромосом, необходимо дать количественные измерения относительных длин плеч, хотя, конечно, следует иметь в виду погрешность, присущую таким измерениям.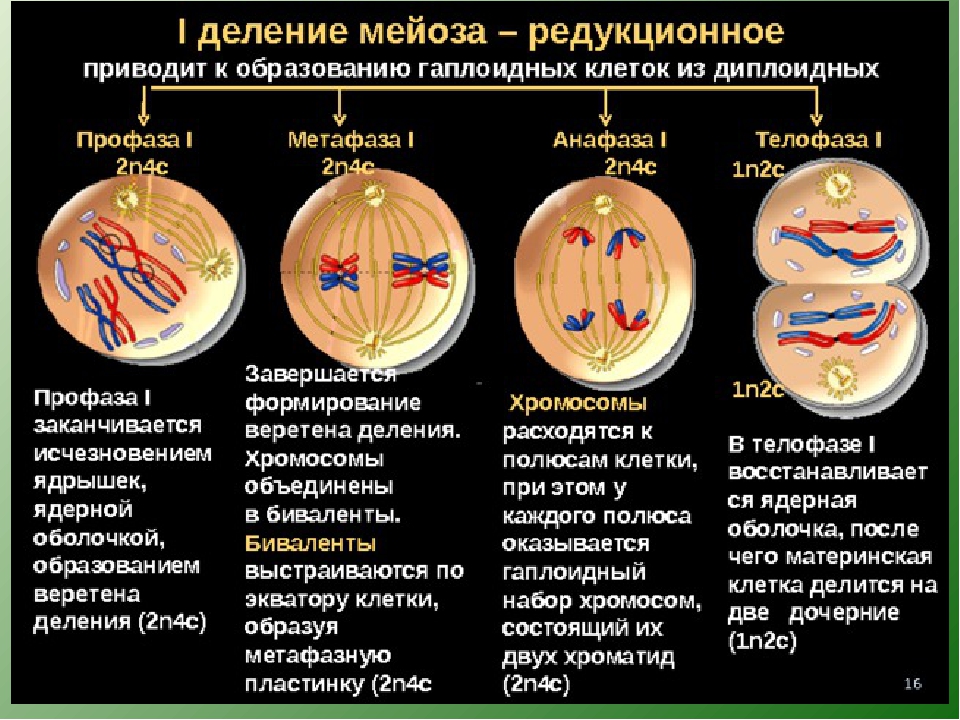 Леван и др. . (1964) используют более сложную терминологию, основанную на таких измерениях, но здесь предпочтительнее простое разделение на «акроцентрические» и «метацентрические». Ясно, что какая бы терминология ни использовалась, должны быть пограничные случаи, когда необходимо принять решение о том, какая категория применима. В данном случае иногда может возникнуть вопрос, может ли короткое плечо считаться незначительным или оно должно считаться коротким само по себе.
Леван и др. . (1964) используют более сложную терминологию, основанную на таких измерениях, но здесь предпочтительнее простое разделение на «акроцентрические» и «метацентрические». Ясно, что какая бы терминология ни использовалась, должны быть пограничные случаи, когда необходимо принять решение о том, какая категория применима. В данном случае иногда может возникнуть вопрос, может ли короткое плечо считаться незначительным или оно должно считаться коротким само по себе.
В более старом типе препарата две хроматиды хромосомы, как правило, нельзя было различить, и, следовательно, акроцентрические хромосомы в метафазе оказались палочковидными, в то время как метацентрические хромосомы имели V- или J-образную форму.Однако с помощью современных методов две хроматиды обычно четко разделены, и хромосома удерживается вместе только в центромере. Следовательно, акроцентрические хромосомы имеют V-образную форму, а метацентрические хромосомы имеют форму креста.
На основе этих двух характеристик, длины хромосом и положения центромеры, хромосомы можно расположить по порядку, как показано на рис. 8.1. Когда это делается, становится очевидным, что есть, как правило, две хромосомы, похожие по размеру и положению центромеры.Такие хромосомы образуют гомологичную пару. Половые хромосомы гетерогаметного пола составляют исключение из этого правила.
8.1. Когда это делается, становится очевидным, что есть, как правило, две хромосомы, похожие по размеру и положению центромеры.Такие хромосомы образуют гомологичную пару. Половые хромосомы гетерогаметного пола составляют исключение из этого правила.
Деление клетки
Деление клетки состоит из двух фаз — деления ядра , за которым следует цитокинез. Ядерное деление делит генетический материал в ядре, в то время как цитокинез делит цитоплазму. Есть два типа деления ядра — митоз и мейоз. Митоз делит ядро, так что обе дочерние клетки генетически идентичны.Напротив, мейоз — это деление редукции, производящее дочерние клетки, которые содержат половину генетической информации родительской клетки.
Первый шаг в митозе или мейозе начинается с конденсации генетического материала, хроматина, в плотно свернутые тела, хромосомы . Каждая хромосома состоит из двух идентичных половин, называемых сестринскими хроматидами , , которые соединены в центромере . Каждая хроматида состоит из одной плотно свернутой молекулы ДНК.Соматические клетки (все клетки тела, кроме яйцеклеток и сперматозоидов) являются диплоидными клетками, потому что каждая клетка содержит две копии каждой хромосомы. Пара таких хромосом называется гомологичной парой. В гомологичной паре хромосом один гомолог происходит от материнского родителя, другой — от отцовского родителя. У человека 46 хромосом (23 гомологичные пары). У мужчин всего 22 гомологичных пары (аутосомы) и одна негомологичная пара — половые хромосомы X и Y.
Каждая хроматида состоит из одной плотно свернутой молекулы ДНК.Соматические клетки (все клетки тела, кроме яйцеклеток и сперматозоидов) являются диплоидными клетками, потому что каждая клетка содержит две копии каждой хромосомы. Пара таких хромосом называется гомологичной парой. В гомологичной паре хромосом один гомолог происходит от материнского родителя, другой — от отцовского родителя. У человека 46 хромосом (23 гомологичные пары). У мужчин всего 22 гомологичных пары (аутосомы) и одна негомологичная пара — половые хромосомы X и Y.
Когда клетка не делится, хроматин заключен в четко выраженную ядерную оболочку, одно или несколько ядрышек видны внутри ядра, а две центросомы (каждая из которых содержит две центриоли) лежат рядом друг с другом за пределами ядерной оболочки.Эти особенности характерны для интерфазы , — неделящегося, но метаболически активного периода клеточного цикла (рис. 1). Когда начинается деление клеток, эти особенности меняются, как описано в следующих разделах.
Рисунок 1. Этапы клеточного цикла.
Митоз
В митозе (прилагательное, митотическое) есть четыре фазы: профаза, метафаза, анафаза и телофаза (рисунок 2):
- Во время профазы , ядрышки исчезают, хроматин конденсируется в хромосомы, ядерная оболочка разрушается и митотическое веретено собирается.Развитие митотического веретена начинается, когда центросомы расходятся к противоположным концам (полюсам) ядра. По мере того, как они расходятся, из каждой центросомы развиваются микротрубочки, длина которых увеличивается за счет добавления тубулиновых единиц. Микротрубочки каждой центросомы соединяются со специализированными участками центромеры, называемыми кинетохорами . Микротрубочки тянут кинетохоры, перемещая хромосомы вперед и назад к одному полюсу, затем к другому. Внутри веретена также есть микротрубочки, которые перекрываются в центре веретена и не прикрепляются к хромосомам.
- Метафаза начинается, когда хромосомы распределяются по метафазной пластинке, плоскости, лежащей между двумя полюсами веретена.
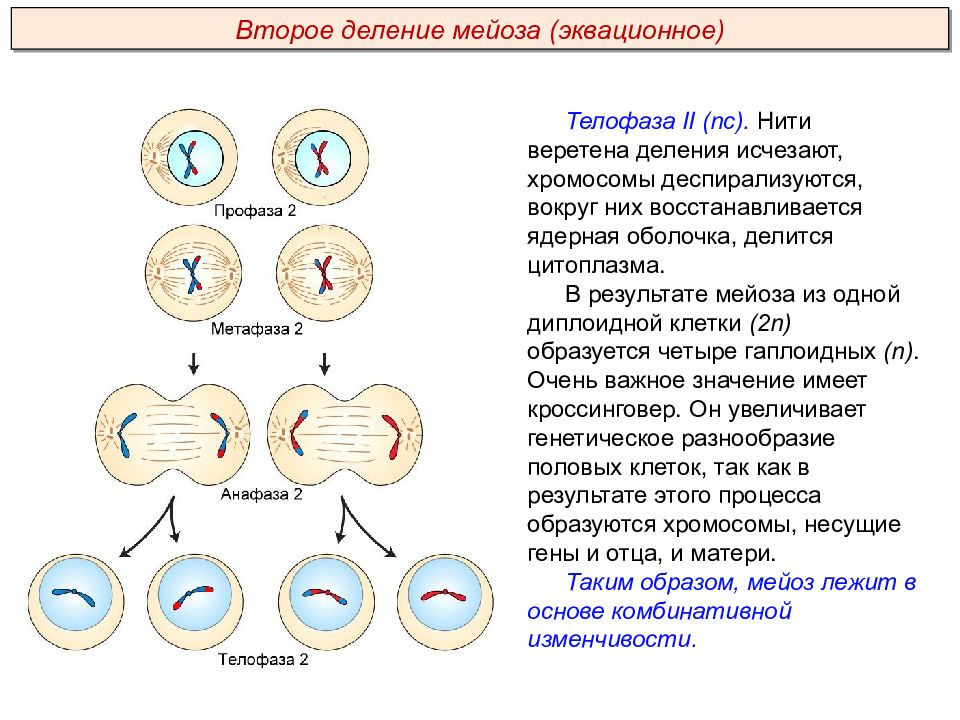 Метафаза заканчивается, когда микротрубочки, все еще прикрепленные к кинетохорам, разделяют каждую хромосому на две хроматиды. Каждая хроматида снабжена центромерой и кинетохорами. После отделения от сестринской хроматиды каждая хроматида называется хромосомой. (Чтобы подсчитать количество хромосом одновременно, подсчитайте количество центромер.)
Метафаза заканчивается, когда микротрубочки, все еще прикрепленные к кинетохорам, разделяют каждую хромосому на две хроматиды. Каждая хроматида снабжена центромерой и кинетохорами. После отделения от сестринской хроматиды каждая хроматида называется хромосомой. (Чтобы подсчитать количество хромосом одновременно, подсчитайте количество центромер.)
- Анафаза начинается после разделения хромосом на отдельные хроматиды. Во время анафазы микротрубочки, связанные с хроматидами (теперь хромосомы), укорачиваются, эффективно притягивая хромосомы к противоположным полюсам. Перекрывающиеся микротрубочки, происходящие из противоположных центросом, но не прикрепленные к хромосомам, взаимодействуют, чтобы раздвинуть полюса дальше друг от друга. В конце анафазы каждый полюс имеет полный набор хромосом, то же количество хромосом, что и исходная клетка.(Поскольку она состоит только из одной хроматиды, каждая хромосома содержит только одну копию молекулы ДНК.)
- Telophase завершает ядерное подразделение.
 Во время этой фазы вокруг каждого полюса образуется ядерная оболочка, образующая два ядра. Хромосомы в каждом из этих ядер распадаются на хроматин, и ядра появляются снова. Одновременно происходит цитокинез, разделяющий цитоплазму на две клетки. Микрофиламенты образуют кольцо внутри плазматической мембраны между двумя новообразованными ядрами.По мере того как микрофиламенты укорачиваются, они действуют как кошелек, притягивая плазматическую мембрану к центру, разделяя клетку на две дочерние клетки. Канавка, которая образуется при затягивании кошельков, называется бороздой спайности .
Во время этой фазы вокруг каждого полюса образуется ядерная оболочка, образующая два ядра. Хромосомы в каждом из этих ядер распадаются на хроматин, и ядра появляются снова. Одновременно происходит цитокинез, разделяющий цитоплазму на две клетки. Микрофиламенты образуют кольцо внутри плазматической мембраны между двумя новообразованными ядрами.По мере того как микрофиламенты укорачиваются, они действуют как кошелек, притягивая плазматическую мембрану к центру, разделяя клетку на две дочерние клетки. Канавка, которая образуется при затягивании кошельков, называется бороздой спайности .
Рисунок 2. Размножение клеток и четыре стадии митоза.
После завершения митоза и начала интерфазы клетка начинает период роста. Рост начинается во время первой фазы, называемой G 1 (разрыв), и продолжается через фазы S (синтез) и G 2 .Также во время фазы S синтезируется вторая молекула ДНК для каждой хромосомы. В результате этой репликации ДНК каждая хромосома получает вторую хроматиду. Во время периода роста G 2 готовятся материалы для следующего митотического деления. Промежуток времени от деления одной клетки до G 1 , S и G 2 называется клеточным циклом (рис. 1).
Во время периода роста G 2 готовятся материалы для следующего митотического деления. Промежуток времени от деления одной клетки до G 1 , S и G 2 называется клеточным циклом (рис. 1).
Клетка, которая начинает митоз в диплоидном состоянии, то есть с двумя копиями каждой хромосомы, завершит митоз с двумя копиями каждой хромосомы.Однако каждая из этих хромосом будет состоять только из одной хроматиды или одной молекулы ДНК. Во время интерфазы вторая молекула ДНК реплицируется из первой, так что, когда начинается следующее митотическое деление, каждая хромосома снова будет состоять из двух хроматид.
Мейоз
Мейоз (прилагательное, мейотический) очень похож на митоз. Основное различие состоит в том, что мейоз состоит из двух групп подразделений, мейоза I и мейоза II (рис. 3). В мейозе I гомологичные хромосомы спариваются на метафазной пластинке, а затем мигрируют к противоположным полюсам.В мейозе II хромосомы распространяются по метафазной пластинке, а сестринские хроматиды разделяются и мигрируют к противоположным полюсам. Таким образом, мейоз II аналогичен митозу. Ниже приводится краткое описание каждой стадии мейоза:
Таким образом, мейоз II аналогичен митозу. Ниже приводится краткое описание каждой стадии мейоза:
- Профаза I начинается как профаза митоза. Ядрышко исчезает, хроматин конденсируется в хромосомы, ядерная оболочка разрушается, и развивается веретенообразный аппарат. Однако, когда хромосомы конденсируются, их поведение отличается от митоза. Во время профазы I происходит спаривание гомологичных хромосом — процесс, называемый синапсисом . Эти пары гомологичных хромосом называются тетрадами (группа из четырех хроматид) или бивалентами. Во время синапсов соответствующие области образуют тесные ассоциации, называемые хиазмами, (единичные, хиазмы) вдоль несестринговых хроматид. Хиазмы — это участки, в которых происходит обмен генетическим материалом между несестринскими гомологичными хроматидами; этот процесс называется кроссинговером . Результат способствует смешиванию генетического материала от обоих родителей, процесс, называемый генетической рекомбинацией.

- В метафазе I гомологичные пары хромосом распределены по метафазной пластинке. Микротрубочки, отходящие от одного полюса, прикреплены к кинетохорам одного члена каждой гомологичной пары. Микротрубочки другого полюса соединены со вторым членом каждой гомологичной пары.
- Анафаза I начинается, когда гомологи внутри тетрад разъединяются, поскольку они притягиваются к противоположным полюсам.
- В телофазе I хромосомы достигли своих полюсов, и вокруг них развивается ядерная мембрана.Обратите внимание, что каждый полюс образует новое ядро, которое будет иметь половину числа хромосом, но каждая хромосома будет содержать две хроматиды. Поскольку в дочерних ядрах будет половина хромосом, клетки, которые они в конечном итоге образуют, будут гаплоидными.
- Происходит цитокинез с образованием двух дочерних клеток. Может последовать короткая интерфаза, но репликации хромосом не происходит. Вместо этого в обоих дочерних ядрах начинается вторая часть мейоза.

- В профазе II ядерная оболочка исчезает, и развивается веретено.Нет хиазм и кроссинговера генетического материала, как в профазе I.
- В метафазе II хромосомы выстраиваются поодиночке на метафазной пластинке (а не в тетрадах, как в метафазе I). Единичное выравнивание хромосом — это именно то, что происходит при митозе, за исключением того, что сейчас количество хромосом составляет только половину.
- Анафаза II начинается, когда каждая хромосома разделяется на две хроматиды микротрубочками аппарата веретена. Хроматиды (теперь хромосомы) мигрируют к своим полюсам.Опять же, это именно то, что происходит при митозе — за исключением того, что теперь количество хромосом составляет только половину.
- В телофазе II ядерная оболочка снова появляется на каждом полюсе и происходит цитокинез. Конечный результат мейоза — четыре гаплоидных клетки. Каждая клетка содержит половину числа хромосом, и каждая хромосома состоит только из одной хроматиды.

Рисунок 3. Этапы мейоза.
Мейоз заканчивается четырьмя гаплоидными дочерними клетками, каждая из которых имеет половину числа хромосом (по одной хромосоме от каждой гомологичной пары).Это гамет — то есть яйцеклетки и сперматозоиды. Слияние яйцеклетки и сперматозоидов, оплодотворение (сингамия , ), дает начало диплоидной клетке, зиготе . Затем одноклеточная зигота делится путем митоза с образованием многоклеточного эмбриона плода, а через девять месяцев — новорожденного ребенка. Обратите внимание, что одна копия каждой пары хромосом в зиготе происходит от одного родителя, а вторая копия — от другого родителя. Таким образом, пара гомологичных хромосом в диплоидной зиготе представляет собой как материнское, так и отцовское наследие.
Репликация ДНК
Во время S-фазы интерфазы собирается вторая хроматида. Вторая хроматида содержит точно такую же ДНК, что и первая хроматида. Процесс копирования, называемый репликацией ДНК , включает разделение («распаковку») молекулы ДНК на две цепи, каждая из которых служит шаблоном для сборки новой комплементарной цепи. В результате получаются две идентичные двухцепочечные молекулы ДНК, состоящие из одной цепи старой ДНК (матричная цепь) и одной цепи новой реплицированной ДНК (комплементарной цепи).
В результате получаются две идентичные двухцепочечные молекулы ДНК, состоящие из одной цепи старой ДНК (матричная цепь) и одной цепи новой реплицированной ДНК (комплементарной цепи).
Ниже приведены этапы дублирования ДНК. Изучая шаги, обращайтесь к Рисунку 4:
- Каждая цепь ДНК помечена как 3 # x2032 и 5 # x2032. Область 3 # x2032 оканчивается гидроксильной группой, а область 5 # x2032 оканчивается фосфатной группой.
- Фермент геликаза «расстегивает» (раскручивает) спираль ДНК, создавая Y-образную вилку репликации. Примечание. ДНК, показанная на рисунке 4, не имеет спиральной формы; он нарисован в параллельной форме для простоты понимания.
- Праймеры РНК «привносят» соответствующие пары оснований в каждую из исходных цепей. ДНК-полимераза — это фермент, который связывает пары оснований вместе, но он может работать только в направлении от 5 # x2032 до 3 # x2032.
- Другая исходная нить также должна быть «соединена» 5 # x2032 в 3 # x2032, чтобы она была собрана в обратном порядке.

- Чтобы связать эти пары оснований с исходной цепью, необходим другой фермент, называемый ДНК-лигазой .Это называется «запаздывающая нить», поскольку на сборку требуется больше времени.
Рисунок 4. Репликация ДНК.
Мутации
Процесс репликации ДНК чрезвычайно точен; однако ошибки могут возникать, когда нуклеотидные основания между нитями ДНК иногда неправильно спарены. Кроме того, ошибки в молекулах ДНК могут возникать в результате воздействия радиации (например, ультрафиолета или рентгеновского излучения) или различных химически активных веществ. При возникновении ошибок доступны механизмы исправления для внесения исправлений.
Если ошибку ДНК не исправить, она становится мутацией. Мутация — это любая последовательность нуклеотидов в молекуле ДНК, которая не совсем соответствует исходной молекуле ДНК, с которой она была скопирована. Мутации включают неправильный нуклеотид (замена), недостающий нуклеотид (делеция) или дополнительный нуклеотид, отсутствующий в исходной молекуле ДНК (вставка). Когда происходит инсерционная мутация, это вызывает смещение всех последующих нуклеотидов на одну позицию, вызывая мутацию сдвига рамки считывания.Радиация или химические вещества, вызывающие мутации, называются мутагенами . Мутагены, активирующие неконтролируемый рост клеток (рак), называются канцерогенами.
Когда происходит инсерционная мутация, это вызывает смещение всех последующих нуклеотидов на одну позицию, вызывая мутацию сдвига рамки считывания.Радиация или химические вещества, вызывающие мутации, называются мутагенами . Мутагены, активирующие неконтролируемый рост клеток (рак), называются канцерогенами.
Синтез белка
ДНК в хромосомах содержит генетические инструкции, которые регулируют развитие, рост и метаболическую активность клеток. Инструкции ДНК определяют, будет ли клетка клеткой гороха, человека или какого-либо другого организма, а также устанавливают конкретные характеристики клетки в этом организме.Например, ДНК в клетке может установить, что это человеческая клетка. Если во время развития он становится клеткой радужной оболочки любого глаза, ДНК будет направлять другую информацию, соответствующую ее местоположению в организме, например, образование коричневой, синей или другой пигментации. ДНК контролирует клетку таким образом, потому что она содержит коды для полипептидов. Многие полипептиды представляют собой ферменты, регулирующие химические реакции и влияющие на результирующие характеристики клетки. Таким образом, с молекулярной точки зрения признаки являются конечными продуктами метаболических процессов, регулируемых ферментами.Ген определяется как сегмент ДНК, который кодирует определенный фермент или другой полипептид (гипотеза один ген — один полипептид).
Многие полипептиды представляют собой ферменты, регулирующие химические реакции и влияющие на результирующие характеристики клетки. Таким образом, с молекулярной точки зрения признаки являются конечными продуктами метаболических процессов, регулируемых ферментами.Ген определяется как сегмент ДНК, который кодирует определенный фермент или другой полипептид (гипотеза один ген — один полипептид).
Процесс, описывающий, как ферменты и другие белки производятся из ДНК, называется синтезом белка . Синтез белка состоит из трех этапов: транскрипции, обработки РНК и трансляции. При транскрипции молекулы ДНК используются в качестве матрицы для создания РНК. После транскрипции процессинг РНК модифицирует молекулу РНК с делециями и добавлениями.При трансляции обработанные молекулы РНК используются для сборки аминокислот в полипептид.
Во время транскрипции образуются три типа молекул РНК:
- Информационная РНК (мРНК) представляет собой одну цепь РНК, которая обеспечивает матрицу, используемую для секвенирования аминокислот в полипептид.
 Триплетная группа из трех соседних нуклеотидов на мРНК, называемая кодоном , кодирует для одной конкретной аминокислоты. Существует 64 возможных способа расположения четырех нуклеотидов в триплетные комбинации (4 × 4 × 4 = 64 возможных кодона).Генетический код — это таблица информации, которая обеспечивает «декодирование» для каждого кодона, то есть идентифицирует аминокислоту, указанную каждой из возможных 64 комбинаций кодонов. Например, кодон, состоящий из трех нуклеотидов цитозин-гуанин-аденин (CGA), кодирует аминокислоту аргинин. См. Рисунок 5.
Триплетная группа из трех соседних нуклеотидов на мРНК, называемая кодоном , кодирует для одной конкретной аминокислоты. Существует 64 возможных способа расположения четырех нуклеотидов в триплетные комбинации (4 × 4 × 4 = 64 возможных кодона).Генетический код — это таблица информации, которая обеспечивает «декодирование» для каждого кодона, то есть идентифицирует аминокислоту, указанную каждой из возможных 64 комбинаций кодонов. Например, кодон, состоящий из трех нуклеотидов цитозин-гуанин-аденин (CGA), кодирует аминокислоту аргинин. См. Рисунок 5.
Как пользоваться графиком:
1. Код продукции лейцина аминокислоты —
.CUA, CUG, CUC, CUU, UUA или UUG.
2 Код продукции лизина аминокислоты — AAA или
AAG.
3. Код аминокислоты цистеина — UGU или UGC.
- Трансферная РНК (тРНК) — это короткая молекула РНК (состоящая примерно из 80 нуклеотидов), которая используется для транспортировки аминокислот к их нужным местам на матрице мРНК.
 Взаимодействия между различными частями молекулы тРНК приводят к спариванию оснований между нуклеотидами, сворачивая тРНК таким образом, что она образует трехмерную молекулу.(В двух измерениях тРНК напоминает три части листа клевера.) Один конец тРНК присоединяется к аминокислоте. Другая часть тРНК, определяемая комбинацией триплетов нуклеотидов, является антикодоном. Во время трансляции антикодон тРНК спаривается с кодоном мРНК.
Взаимодействия между различными частями молекулы тРНК приводят к спариванию оснований между нуклеотидами, сворачивая тРНК таким образом, что она образует трехмерную молекулу.(В двух измерениях тРНК напоминает три части листа клевера.) Один конец тРНК присоединяется к аминокислоте. Другая часть тРНК, определяемая комбинацией триплетов нуклеотидов, является антикодоном. Во время трансляции антикодон тРНК спаривается с кодоном мРНК.
- Молекулы рибосомной РНК (рРНК) являются строительными блоками рибосом. Ядрышко представляет собой совокупность ДНК, активно транскрибируемую в рРНК. Внутри ядрышка различные белки, импортируемые из цитозоля, собираются с рРНК с образованием больших и малых субъединиц рибосом.Вместе две субъединицы образуют рибосому, которая координирует действия мРНК и тРНК во время трансляции. Рибосомы имеют три сайта связывания — один для мРНК, один для тРНК, несущей растущую полипептидную цепь, и один для второй тРНК, которая доставляет следующую аминокислоту, которая будет вставлена в растущую полипептидную цепь.
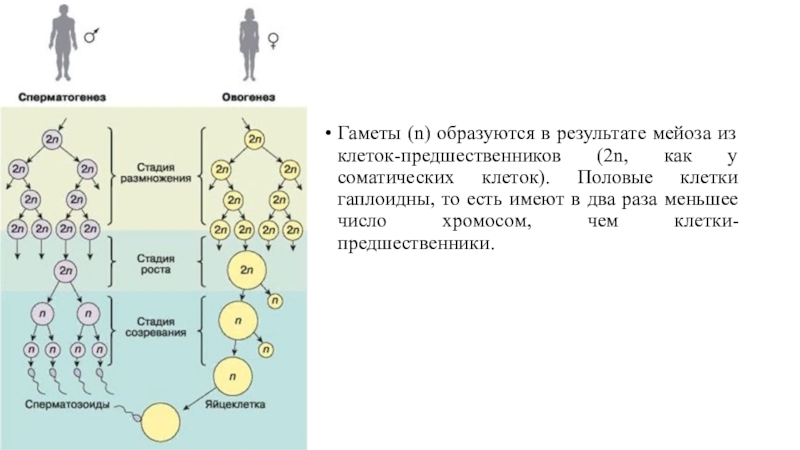
Рис. 5. Комбинации производства аминокислот.
Вот подробности транскрипции, процессинга РНК и синтеза белка (см. Также рисунки 6 и 7):
- Во время транскрипции РНК-полимераза прикрепляется к промоторным участкам ДНК и начинает распаковывать ДНК на две цепи.(См. Шаг 1 на рисунке 6.)
- По мере того, как РНК-полимераза распаковывает ДНК, она собирает новые нуклеотиды, используя одну нить ДНК в качестве матрицы. В отличие от процесса репликации ДНК, новые нуклеотиды являются нуклеотидами РНК, и транскрибируется только одна цепь ДНК. (См. Шаг 2 на рисунке 6.)
- Транскрипция продолжается до тех пор, пока РНК-полимераза не достигнет особой последовательности нуклеотидов, которая служит точкой терминации. Высвобождаются РНК-полимераза и вновь созданная молекула РНК.Эта вновь созданная молекула РНК может быть мРНК, тРНК или рРНК, в зависимости от того, какой сегмент ДНК транскрибируется. (См. Шаг 3 на рисунке 6.)
- Во время обработки РНК вновь созданные молекулы мРНК претерпевают два вида изменений.
 В первой модификации некодирующие промежуточные последовательности, называемые интронами , удаляются, оставляя только экзонов, последовательностей, которые выражают код полипептида. Вторая модификация добавляет две специальные последовательности — 5-дюймовый колпачок к одному концу мРНК и поли-A-хвост к другому концу.(См. Шаги 4A, 4B и 4C на рисунке 6.)
В первой модификации некодирующие промежуточные последовательности, называемые интронами , удаляются, оставляя только экзонов, последовательностей, которые выражают код полипептида. Вторая модификация добавляет две специальные последовательности — 5-дюймовый колпачок к одному концу мРНК и поли-A-хвост к другому концу.(См. Шаги 4A, 4B и 4C на рисунке 6.)
- МРНК, тРНК и субъединицы рибосом транспортируются через ядерную оболочку в цитоплазму. В цитоплазме аминокислоты прикрепляются к одному концу тРНК. (См. Шаги 5A, 5B и 5C на рисунке 7.)
- Трансляция начинается, когда малая и большая рибосомные субъединицы прикрепляются к одному концу мРНК. Кроме того, тРНК (с антикодоном UAC), несущая аминокислоту метонин, прикрепляется к мРНК (в «стартовом» кодоне AUG) внутри рибосомы.(См. Шаг 6 на рисунке 7.)
- Вторая тРНК, также несущая аминокислоту, прибывает и заполняет вторую позицию тРНК. Кодон мРНК определяет, какая тРНК (и, следовательно, какая аминокислота) занимает вторую позицию.
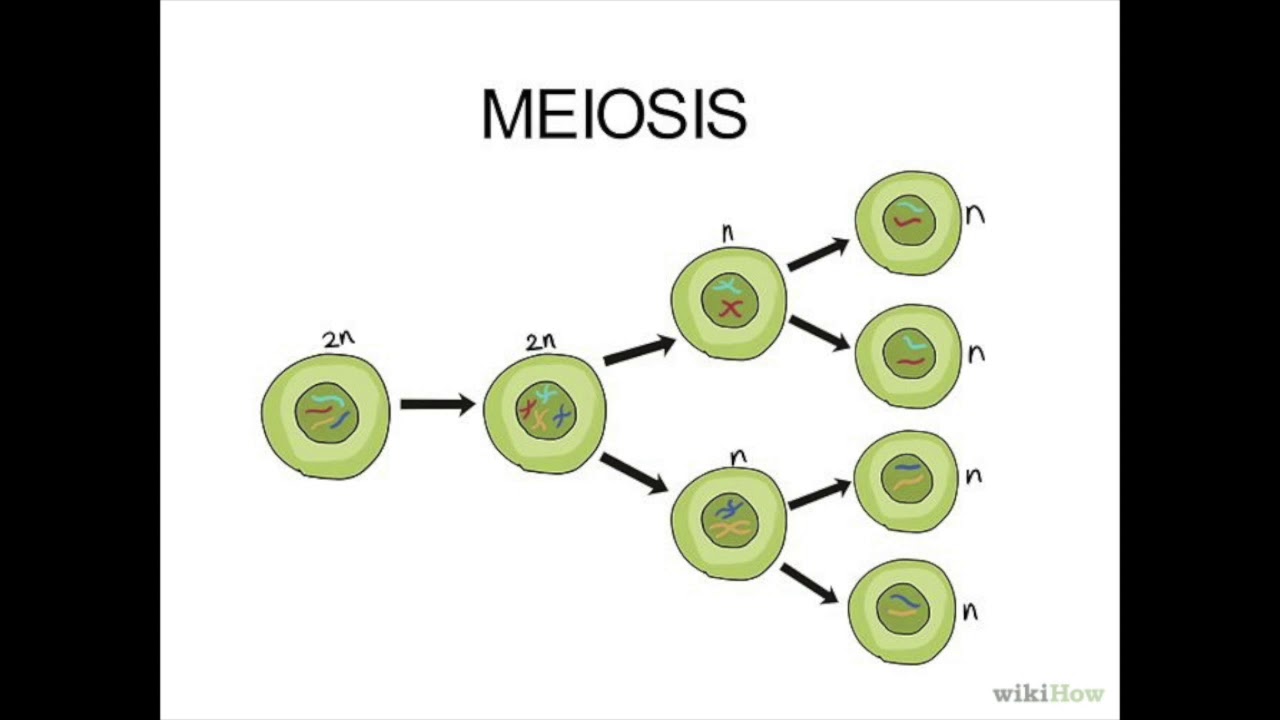 (Шаг 7 на рисунке 7 показывает входящую тРНК, приближающуюся к еще не освобожденной позиции.)
(Шаг 7 на рисунке 7 показывает входящую тРНК, приближающуюся к еще не освобожденной позиции.)
- Аминокислота первой тРНК присоединяется к аминокислоте второй тРНК, образуя пару аминокислот. Затем высвобождается первая тРНК. Рибосома перемещается над одной позицией кодона, тем самым помещая вторую тРНК в первое положение и освобождая второе положение.(Шаг 8 на рисунке 7 показывает этот процесс после того, как несколько тРНК доставили аминокислоты.)
- Новая тРНК (с ее аминокислотой) заполняет вакантное место. Теперь две аминокислоты, удерживаемые тРНК в первом положении, переносятся на аминокислоту вновь прибывшей тРНК, образуя полипептидную цепь из трех аминокислот. Опять же, тРНК в первом положении высвобождается, рибосома перемещается над одним положением кодона, а второе положение тРНК остается вакантным.
- Процесс продолжается, поскольку новые тРНК приносят больше аминокислот.По мере поступления каждой новой тРНК полипептидная цепь удлиняется на одну новую аминокислоту, увеличиваясь в последовательности и длине в соответствии с кодонами на мРНК.
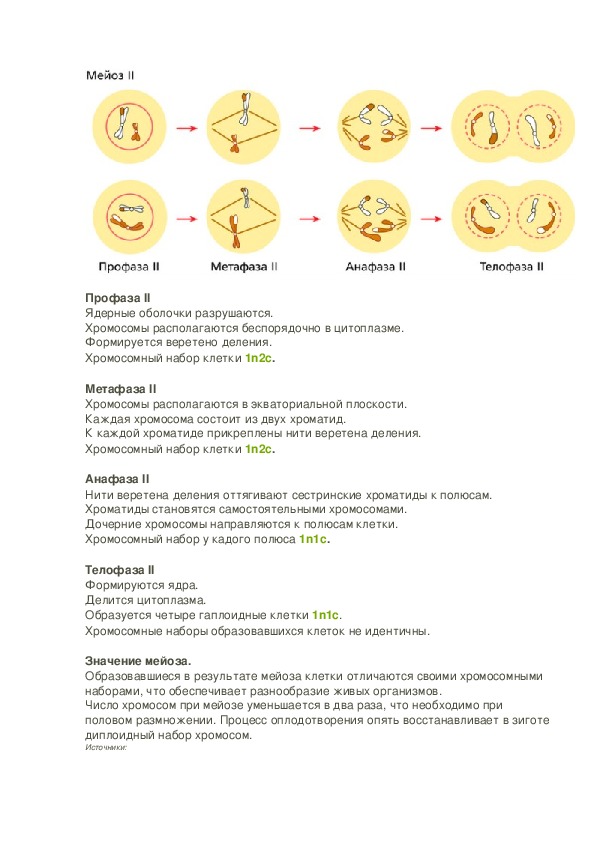 (См. Этап 9 на рисунке 7.) В конце концов, встречается «стоп-кодон», такой как UAG, и субъединицы рибосомы и полипептид высвобождаются.
(См. Этап 9 на рисунке 7.) В конце концов, встречается «стоп-кодон», такой как UAG, и субъединицы рибосомы и полипептид высвобождаются.
Рисунок 6. Транскрипция и обработка РНК.
Рис. 7. Этапы синтеза белка.
После высвобождения полипептида взаимодействия между аминокислотами придают белку особую трехмерную форму.Последующая обработка эндоплазматическим ретикулумом или телом Гольджи может произвести окончательные модификации до того, как белок начнет функционировать как структурный элемент или фермент.
.
