
Животный мир палеозойской эры
Рождение Земли и жизни на Земле
Возраст нашей старушки Земли, которая является родным домом всему человечеству, очень велик. Его даже трудно себе представить. Земля возникла в звездном пространстве около 5 миллиардов лет назад! Попробуйте вообразить, почувствовать это время. 5 миллиардов – это значит 5 тысяч раз взять по миллиону лет! Вот как далек от нашего времени день ее рождения!
Для удобства изучения этапов образования нашей планеты ученые разделили историю ее развития и эволюцию жизни на отдельные эры, используя для этого некоторые естественные, характерные особенности. В свою очередь эры делятся на периоды. Продолжительность существования каждой эры и каждого периода – их возраст – определена в миллионах лет.
Эра звездного существования Земли – тогда раскаленной и безжизненной – удалена от нашего времени на миллиарды лет. Атмосфера была насыщена горячими газами и водяными парами. Тучи вулканического пепла сплошной пеленой окутывали планету и не пропускали солнечных лучей.
Следующая за архейской была протерозойская, или древнейшая, эра развития Земли. Ее продолжительность определена от 1,5 миллиарда до 520 миллионов лет. Для этой эры характерно дальнейшее образование морских водоемов, развитие в них массы водорослей и разнообразных беспозвоночных животных.

После протерозойской наступила палеозойская эра (от 520 до 185 миллионов лет), которую ученые делят на 6 периодов: кембрий, ордовик, силур, девон, карбон, или каменноугольный период, и пермский. Причем каждый период имеет свои характерные черты развития и для каждого определена продолжительность его существования (см. таблицу) и типичный для периода растительный и животный мир. В течение существования палеозойской эры очень активно развивались беспозвоночные животные, заселявшие водные просторы: одни группы вымирали, их сменяли другие, возникали новые формы растений и животных. В палеозое впервые появились ПОЗВОНОЧНЫЕ животные – первые рыбы: панцирные, акуловые и кистеперые, первые земноводные и к концу эры – первые пресмыкающиеся. Все эти животные быстро расселялись, захватывая все новые и новые водные пространства, а выйдя из воды на сушу, они стали распространяться на берегах, постепенно продвигаясь в глубь континентов, приспосабливаясь к самым различным условиям обитания.

Вот об этом времени развития нашей планеты – ПАЛЕОЗОЕ, о некоторых характерных для этой эры животных мы и хотим рассказать!
Представь себе, что мы с тобой решили прогуляться по берегу Кембрийского моря… Море было теплое-теплое. Мы разулись и пошли побродить среди зарослей морских водорослей. И вдруг, как только мы вошли в море, из-под ног выскользнули какие-то небольшие животные и, взмутив воду, скрылись из глаз. Мы остановились и стали присматриваться.
– Да вот же они! Смотри! Спрятались – закопались в песок. А ну-ка, давай рассмотрим их как следует, что это за животные? Да это ведь трилобиты! Одни из самых первых беспозвоночных, появившихся на Земле. Это самые массовые и очень характерные животные Кембрийского моря. В начале кембрийского периода они превосходили всех других животных и по своей численности, и по видовому разнообразию. Но позже, в Девонском море, их стало уже значительно меньше, а к концу палеозоя они и вовсе вымерли. До наших времен дошли только их ископаемые окаменелые останки.

Присмотрись к ним хорошенько: видишь, их тело как бы разделяется и поперечно и продольно на три части. За это их и назвали трилобитами. Ведь слово «лобос» по-гречески значит «лопасть». Подобно ракообразным, трилобиты покрыты известковым, достаточно прочным спинным щитом, состоящим из отдельных подвижных сегментов. У разных видов – их разное количество. Трилобиты были небольшими, их длина от 3 до 10 см. Правда, некоторые редкие виды достигали большей длины, иногда даже до 70 см. В головной части у трилобитов были небольшие «усики» – антенны.
В море встречался и птериготус – крупный хищник, ракоскорпион, достигавший длины до 2 м, обладавший мощными клешнями.
В кембрии впервые на Земле появились и головоногие моллюски – прародители современных кальмаров, каракатиц и осьминогов.
 Правда, в те далекие времена они имели внешнюю раковину трубчатой формы, украшенную разноцветным рисунком. Но современные головоногие (кроме наутилуса) давно потеряли раковины по пути эволюционного развития. Мы показали здесь только трех представителей типичных беспозвоночных кембрийского периода, хотя население Кембрийского моря было очень богатым и разнообразным. За длительный период его существования одни формы вымирали, исчезали, нарождались новые, расселялись на огромных водных пространствах, снова вымирали… Шел бурный, активный процесс видообразования как среди водорослей, которые в кембрии впервые «вышли» на сушу и образовали первые своеобразные «леса», так и среди животных. Ведь именно в кембрии возникли первые позвоночные животные. Это были древние рыбы.
Правда, в те далекие времена они имели внешнюю раковину трубчатой формы, украшенную разноцветным рисунком. Но современные головоногие (кроме наутилуса) давно потеряли раковины по пути эволюционного развития. Мы показали здесь только трех представителей типичных беспозвоночных кембрийского периода, хотя население Кембрийского моря было очень богатым и разнообразным. За длительный период его существования одни формы вымирали, исчезали, нарождались новые, расселялись на огромных водных пространствах, снова вымирали… Шел бурный, активный процесс видообразования как среди водорослей, которые в кембрии впервые «вышли» на сушу и образовали первые своеобразные «леса», так и среди животных. Ведь именно в кембрии возникли первые позвоночные животные. Это были древние рыбы.
Вслед за кембрием наступил новый период – ордовик, в течение которого (от 440 до 360 миллионов лет) продолжался активный процесс развития и изменения растительности и животного мира Земли. Его сменил силурийский период, начавшийся 360 миллионов лет назад и закончившийся 320 миллионов лет назад.
 Эти 40 миллионов лет силура прежде всего характеризуются появлением на Земле многих видов рыб. Самыми первыми были мелкие (до 10 см) панцирнокожие рыбы, обитавшие в пресноводных болотах и небольших лужах. Их существование охватывает два геологических периода – силур и девон, в конце которого они все вымерли. Второй класс панцирных рыб – пластинокожие – появился в конце силура. Эти рыбы из пресных вод постепенно перешли и приспособились к жизни в море, сделавшись типичными морскими жителями. В их числе появились и такие крупные хищники, как изображенный здесь динихтис, достигавший 10 м длины! Динихтис питался разными морскими животными, всасывая их вместе с водой, так же как это делали панцирнокожие рыбы, так как никто из них не обладал развитыми челюстями, способными хватать и удерживать свою добычу.
Эти 40 миллионов лет силура прежде всего характеризуются появлением на Земле многих видов рыб. Самыми первыми были мелкие (до 10 см) панцирнокожие рыбы, обитавшие в пресноводных болотах и небольших лужах. Их существование охватывает два геологических периода – силур и девон, в конце которого они все вымерли. Второй класс панцирных рыб – пластинокожие – появился в конце силура. Эти рыбы из пресных вод постепенно перешли и приспособились к жизни в море, сделавшись типичными морскими жителями. В их числе появились и такие крупные хищники, как изображенный здесь динихтис, достигавший 10 м длины! Динихтис питался разными морскими животными, всасывая их вместе с водой, так же как это делали панцирнокожие рыбы, так как никто из них не обладал развитыми челюстями, способными хватать и удерживать свою добычу.
 Это были относительно небольшие (около 70 см) быстроплавающие хищники, «вооруженные» мощным движителем – хвостом, с двумя крупными треугольными грудными плавниками и двумя небольшими лопастями, «посаженными» по бокам хвостового стебля. Эти лопасти были своеобразными «рулями высоты» (глубины) и обеспечивали кладоселахиям наилучшую маневренность движения при добыче быстроплавающих морских животных, которыми они питались.
Это были относительно небольшие (около 70 см) быстроплавающие хищники, «вооруженные» мощным движителем – хвостом, с двумя крупными треугольными грудными плавниками и двумя небольшими лопастями, «посаженными» по бокам хвостового стебля. Эти лопасти были своеобразными «рулями высоты» (глубины) и обеспечивали кладоселахиям наилучшую маневренность движения при добыче быстроплавающих морских животных, которыми они питались.
Наступивший после силура девонский период отличается от всех предыдущих богатейшим развитием ихтиофауны. Он по праву должен быть назван «царством рыб»!
Впервые, именно в Девонском море, возникли кистеперые рыбы (от 1,5 до 2 м), один из потомков которых дожил и до наших времен. Я имею в виду знаменитую латимерию – целаканта, живущую до сих пор в Индийском океане около Сейшельских и Коморских островов. Одна из пойманных там латимерий хранится у нас в Москве.
Кистеперыми эти рыбы были названы потому, что они обладали крепкими и сильными грудными плавниками особого строения, на которые рыбы сперва опирались при передвижении по морскому дну.

Выход кистеперых рыб на сушу стал великим событием в эволюции животных нашей планеты! На Земле появился новый класс ЗЕМНОВОДНЫХ животных, положивший начало всему богатству наземной (сухопутной) фауны позвоночных.
В конце силурийского периода палеозоя ПЕРВЫЕ ЗЕМНОВОДНЫЕ начали осваиваться и приспосабливаться к многообразным условиям жизни на суше.
ПЕРВЫЕ ЗЕМНОВОДНЫЕ
Древнейшие земноводные – ихтиостеги – обитали в верхнем девоне около 300–320 миллионов лет назад. Эти примитивные амфибии сохраняли еще очень много сходных и даже общих черт (признаков) с кистеперыми рыбами. Поэтому происхождение земноводных от кистеперых рыб не подлежит никакому сомнению. В дальнейшем история эволюции земноводных нашей планеты развивалась неравномерно. Изобилие и процветание этих животных было отмечено в карбоне, триасе и кайнозое, когда они были представлены множеством разнообразных форм. В то же время в юрском и меловом периодах произошло замедление их развития, уменьшилась численность и видовое разнообразие. Однако начиная с верхнего карбона (в палеозое) и до конца триаса (в мезозое) амфибии первенствовали в тогдашней фауне. В качестве одного из представителей крупных земноводных мы показываем вам мастодонзавра, появившегося на Земле в конце каменноугольного периода. Это был крупный хищник, питавшийся почти исключительно рыбами, населявший пресноводные водоемы (озера и болота). Он вел водный образ жизни. Его привычки и поведение были весьма сходными с образом жизни обыкновенных лягушек.
Эти примитивные амфибии сохраняли еще очень много сходных и даже общих черт (признаков) с кистеперыми рыбами. Поэтому происхождение земноводных от кистеперых рыб не подлежит никакому сомнению. В дальнейшем история эволюции земноводных нашей планеты развивалась неравномерно. Изобилие и процветание этих животных было отмечено в карбоне, триасе и кайнозое, когда они были представлены множеством разнообразных форм. В то же время в юрском и меловом периодах произошло замедление их развития, уменьшилась численность и видовое разнообразие. Однако начиная с верхнего карбона (в палеозое) и до конца триаса (в мезозое) амфибии первенствовали в тогдашней фауне. В качестве одного из представителей крупных земноводных мы показываем вам мастодонзавра, появившегося на Земле в конце каменноугольного периода. Это был крупный хищник, питавшийся почти исключительно рыбами, населявший пресноводные водоемы (озера и болота). Он вел водный образ жизни. Его привычки и поведение были весьма сходными с образом жизни обыкновенных лягушек.
Земноводные прошли лишь самые первые этапы освоения суши, но именно поэтому их биология до сих пор представляет огромный научный интерес, так как дальнейшая эволюция этих животных, их полный отрыв от водной среды, положила начало для возникновения следующей группы – высших позвоночных (пресмыкающихся).

Впервые именно пресмыкающиеся стали размножаться на суше в отрыве от воды. У них появились яйца с плотной внешней оболочкой, предохраняющей их от высыхания и механических повреждений. Благодаря этому в дальнейшем возникли новые группы высших позвоночных – птицы и млекопитающие.
Пресмыкающиеся по сравнению с земноводными являются уже типичными наземными (сухопутными) животными. Появление первых пресмыкающихся относится к концу каменноугольного периода палеозоя. Они развивались как сухопутные формы, однако были и такие, которые приспособились к жизни в воздухе (птерозавры), а также и такие, которые, сохраняя все признаки пресмыкающихся, вели водный образ жизни. Большая часть были хищниками, но черепахи и птерозавры питались и растительной пищей. Мезозавр – один из немногих древних ящеров, относящийся к отряду среднеящеров, который приспособился к жизни в воде. Он обитал в пресных водоемах Южной Африки и Бразилии в конце каменноугольного – начале пермского периода. По своему внешнему виду он был похож на современного крокодила: сжатый с боков хвост, пятипалые конечности с плавательными перепонками между пальцев, длинная морда с вытянутыми челюстями, вооруженными массой тонких и длинных зубов.
 Но по своим размерам он был в несколько раз меньше. Самые крупные мезозавры достигали всего от 60 до 100 сантиметров длины. Мезозавр был хищником – питался в основном рыбами.
Но по своим размерам он был в несколько раз меньше. Самые крупные мезозавры достигали всего от 60 до 100 сантиметров длины. Мезозавр был хищником – питался в основном рыбами.
На рисунке вы видите эдафозавра. Он является представителем отряда тероморфа – звероподобных. Эти животные, сохраняя типичный облик пресмыкающихся, в то же время приобрели уже некоторые признаки, свойственные млекопитающим животным, почему их и называли «звероподобные». В частности, эти изменения отмечаются палеонтологами в области скелета (костей черепа, зубов…). Ученые высказали предположение, что некоторые пресмыкающиеся отряда звероподобных были уже теплокровными животными.
Возникнув одновременно с мезозаврами в конце каменноугольного периода, звероподобные пресмыкающиеся достигли особого расцвета в пермском периоде, но уже к началу триаса (мезозой) они угасли и вымерли.
Звероподобные были хищниками, но среди них встречались отдельные виды, которые питались растительной пищей. Эдафозавр также был растительноядным, питался различными болотными травами.
 Длина взрослых эдафозавров достигла 2,5–3 метров. Мы очень кратко постарались рассказать вам о палеозое.
Длина взрослых эдафозавров достигла 2,5–3 метров. Мы очень кратко постарались рассказать вам о палеозое.
Палеозойская эра была одной из самых важных в развитии нашей планеты, одной из самых плодотворных, определившая коренные изменения в эволюции растительного мира и животных Земли. Впервые в палеозое растения вышли из воды и заселили сушу. Впервые возникли на Земле лесные массивы. Впервые в палеозое отмечено интенсивное развитие беспозвоночных животных. Вслед за ними именно в палеозое возникли и первые позвоночные животные. Впервые позвоночные вышли из воды на сушу и начали ее заселять. Впервые возникли высшие позвоночные, эволюция которых в дальнейшем привела к появлению на Земле теплокровных животных. Решающие изменения в развитии нашей планеты, происшедшие в течение палеозоя, подготовили почву для появления на Земле человека!
Рисунки А. Павлова.
|
Геохронологическая шкала -> Палеозой Geochronological scale -> Paleozoic Era Палеозойская эра началась около 540 миллионов лет назад и закончилась примерно 250 миллионов лет назад. На протяжении Палеозоя возникает огромное количество типов и классов живых существ. Жизнь очень сильно усложняется. Если в самом начале Палеозоя все живые организмы обитают в морях, а самыми развитыми живыми существами являются головоногие моллюски, то в конце последнего периода Палеозойской эры – пермского, на суше, покрытой лесами, уже существуют не только амфибии и рептилии, но и примитивные млекопитающие. Именно в Палеозое происходит освоение суши, сначала растениями, затем членистоногим, а затем уже и позвоночными. Морская жизнь не только «выплескивается» на сушу, но и постоянно усложняется в родной среде обитания. Безраздельно властвовавших в начале Палеозоя в толще воды головоногих моллюсков теснят рыбы. Часть головоногих вымирает, но возникают все более сложно организованные виды, появляются аммониты, расцвет которых придется на следующую эру – Мезозойскую. С середины Палеозоя жизнь начинает осваивать еще одну среду – воздушную. Но пока в воздух поднимаются только членистоногие – насекомые. Для позвоночных воздух еще закрыт – они освоят эту среду только в триасе – первом периоде Мезозоя. Конечно, на протяжении всего Палеозоя идет не только возникновение новых групп живых организмов, но и вымирание старых, не успевающих приспособится к новой, изменяющейся обстановке. К середине Палеозоя вымирают широко распространенные в кембрии членистоногие хищники – аномалокарисы и подобные им виды. Трилобиты, доминировавшие в донной фауне в начале Палеозоя, и достигавшие в ордовикском периоде длины в 90 сантиметров, в конце Палеозойской эры – в карбоне и перми, становятся редкими и мелкими – 1-2 сантиметра в длину. А завершается Палеозой грандиозным вымиранием в конце пермского периода. Это вымирание по своим масштабам превзошло все другие известные вымирания, в том числе и знаменитое вымирание динозавров в конце Мезозоя. В конце перми исчезло до 95% видов земной фауны. Причины этого катастрофического вымирания, впрочем, как и других подобных событий, точно неизвестны. Глобальность и массовость вымирания свидетельствуют о том, что у него была какая-то общая и масштабная причина. Admin 14 января 2016 года Окаменелости этого возраста: Acanthodes, Acanthodes lopatini, Acanthodiformes, Acanthodii, Acanthohoplites, Acanthopecten, Acastella, Acastella tiro, Acastidae, Acastoides, Acrocrinus, Acrodus, Actinocerida, Actinocyathus, Admoskovia, Admoskovia inflatiformis, Agnatha, Agnostida, Agnostidae, Agnostus, Agoniatitida, Alethopteris, Alethopteris bertrandi, Alethopteris lonchitica, Alveolites, Ammonites, Ammonoidea, Anathyris, Angiospirifer, Annularia, Anomalocaris, Anthozoa, Antigonambonites, Antraconauta, Aphlebia crispa, Araucaria, Araucariaxylon, Archaeocidaris, Archaeocidaris mosquensis, Archaeocidaris rossica, Archaeopteris, Archosaurus, Arthropoda, Articulata, Artisia, Asaphida, Asaphidae, Asaphiscus, Asaphiscus wheeleri, Asaphus, Asaphus cornutus, Asaphus expansus, Asaphus holmi, Asaphus intermedius, Asaphus kotlukovi, Asaphus kowalewskii, Asaphus latus, Asaphus lepidurus, Asaphus plautini, Asaphus punctatus, Ascodictyon, Asolanus, Asteroidea, Asterolepis, Asterolepis ornata, Asterophyllites, Asteropyginae, Athyridida, Athyris, Athyris concentrica, Atractopyge, Atrypa, Atrypida, Atrypidae, Aulacopteris, Aulopora, Aulopora macrostoma, Auloporida, Aulosteges, Aulosteges horrescens, Aulosteges horrescens sokensis, Aulosteges wangenheimi, Aulostegidae, Aviculopecten, Aviculopectinidae, Bactritida, Bajtugania, Baltoeurypterus, Barrandeops, Barrandeops forteyi, belemnites, Belemnitida, Bellerophon, Bellerophontidae, Billingsellida, bivalvia, Boeckops, Boeckops stelcki, Bolboporites, Boroviczia Палеозой, публикации из палеонтологического форума:
Палеозой, все палеонтологические публикации >>> |
 Она продолжалась 290 миллионов лет. Первый период Палеозойской эры — кембрийский, начался с массового распространения живых организмов с минеральным скелетом. Долгое время считалось, что тогда же возникли и многоклеточные организмы, но изучение вендской (эдиакарской) фауны показало, что мягкотелые многоклеточные, лишенные минерального скелета, возникли значительно раньше. Сейчас палеонтологи полагают, что отдельные виды, обладавшие различными скелетными элементами, могли появится и до начала Палеозоя, но они не были массовыми.
Она продолжалась 290 миллионов лет. Первый период Палеозойской эры — кембрийский, начался с массового распространения живых организмов с минеральным скелетом. Долгое время считалось, что тогда же возникли и многоклеточные организмы, но изучение вендской (эдиакарской) фауны показало, что мягкотелые многоклеточные, лишенные минерального скелета, возникли значительно раньше. Сейчас палеонтологи полагают, что отдельные виды, обладавшие различными скелетными элементами, могли появится и до начала Палеозоя, но они не были массовыми.
 Освоение новой среды обитания приводит к возникновению новых приспособлений и адаптаций, появляются совершенно новые организмы, способные жить в новых условиях. Осваивающие мелководья и полузатопленные участки побережий потомки рыб – амфибии, живут на размытой границе воды и суши, но все-таки еще в воде. Рептилии, благодаря более плотным кожным покровам и размножению защищенными от высыхания, в отличие от икры амфибий, яйцами, уже по-настоящему осваивают сушу.
Освоение новой среды обитания приводит к возникновению новых приспособлений и адаптаций, появляются совершенно новые организмы, способные жить в новых условиях. Осваивающие мелководья и полузатопленные участки побережий потомки рыб – амфибии, живут на размытой границе воды и суши, но все-таки еще в воде. Рептилии, благодаря более плотным кожным покровам и размножению защищенными от высыхания, в отличие от икры амфибий, яйцами, уже по-настоящему осваивают сушу.

 Сторонники катастроф винят в пермском кризисе мощнейшее извержение вулканов на территории современной Сибири или падение крупного астероида, след от которого пока не найден. Есть предположение, что падение астероида и последовавшая за ним вспышка вулканизма связаны друг с другом. Другие исследователи винят в катастрофе резкое глобальное потепление, перегрев Земли, который привел к уменьшению содержания кислорода в воде океанов и гибели как наземных, так и водных экосистем. Есть и другие гипотезы. Ни одна из них не может сейчас считаться более-менее подтвержденной. Ясно одно – жизнь пережила этот кризис и вышла на новый виток своего развития.
Сторонники катастроф винят в пермском кризисе мощнейшее извержение вулканов на территории современной Сибири или падение крупного астероида, след от которого пока не найден. Есть предположение, что падение астероида и последовавшая за ним вспышка вулканизма связаны друг с другом. Другие исследователи винят в катастрофе резкое глобальное потепление, перегрев Земли, который привел к уменьшению содержания кислорода в воде океанов и гибели как наземных, так и водных экосистем. Есть и другие гипотезы. Ни одна из них не может сейчас считаться более-менее подтвержденной. Ясно одно – жизнь пережила этот кризис и вышла на новый виток своего развития.  Сорокаметровые гиганты с диаметром ствола до двух метров и ветвлением, которое начинается в верхней четверти высоты ствола, если не выше. Классическое описание коры – спиралью поднимающиеся по стволу ромбические формы («чешуйчатое дерево»). Достаточно много классических примеров орнамента коры (как позитивных, так и негативных отпечатков, в том числе, частично обескоренных) можно посмотреть в моей галерее Лепидодендроны . … >>>
Сорокаметровые гиганты с диаметром ствола до двух метров и ветвлением, которое начинается в верхней четверти высоты ствола, если не выше. Классическое описание коры – спиралью поднимающиеся по стволу ромбические формы («чешуйчатое дерево»). Достаточно много классических примеров орнамента коры (как позитивных, так и негативных отпечатков, в том числе, частично обескоренных) можно посмотреть в моей галерее Лепидодендроны . … >>> .. >>>
.. >>>Развитие жизни в протерозойской и палеозойской эрах
«Общая биология. 11 класс». В.Б. Захаров и др. (гдз)
11 класс». В.Б. Захаров и др. (гдз)
Вопрос 1. Когда появились первые наземные растения?
В начале палеозойской эры растения населяют в основном моря, но в ордовике — силуре появляются первые наземные растения — псилофиты (рис. 1).
Рис. 1. Первое наземное растение
Это были небольшие растения, занимающие промежуточное положение между водорослями и наземными сосудистыми растениями. Псилофиты имели уже проводящую (сосудистую) систему, первые слабодифференцированные ткани, могли укрепляться в почве, хотя корни еще (как и другие вегетативные органы) отсутствовали. Дальнейшая эволюция растений на суше была направлена на дифференцировку тела на вегетативные органы и ткани, совершенствование- сосудистой системы (обеспечивающей быстрое поднятие воды на большую высоту).
Вопрос 2. В каком направлении шла эволюция растений на суше?
После появления псилофитов эволюция растений на суше шла в направлении расчленения тела на вегетативные органы и ткани, совершенствования сосудистой системы (обеспечивающей быстрое перемещение воды на большую высоту). Уже в засушливом девоне широко распространяются хвощи, плауны, папоротникообразные. Еще большего развития достигает наземная растительность в каменноугольном периоде (карбоне), характеризующемся влажным и теплым климатом на протяжении всего года. Появляются голосеменные растения, произошедшие от семенных папоротников. Переход к семенному размножению дал много преимуществ: зародыш в семенах защищен от неблагоприятных условий оболочками и обеспечен пищей, имеет диплоидное число хромосом. У части голосеменных (хвойных) процесс полового размножения уже не связан с водой. Опыление у голосеменных осуществляется ветром, а семена имеют приспособления для распространения животными. Эти и другие преимущества способствовали широкому распространению семенных растений. Крупные споровые растения вымирают в пермском периоде в связи с иссушением климата.
Уже в засушливом девоне широко распространяются хвощи, плауны, папоротникообразные. Еще большего развития достигает наземная растительность в каменноугольном периоде (карбоне), характеризующемся влажным и теплым климатом на протяжении всего года. Появляются голосеменные растения, произошедшие от семенных папоротников. Переход к семенному размножению дал много преимуществ: зародыш в семенах защищен от неблагоприятных условий оболочками и обеспечен пищей, имеет диплоидное число хромосом. У части голосеменных (хвойных) процесс полового размножения уже не связан с водой. Опыление у голосеменных осуществляется ветром, а семена имеют приспособления для распространения животными. Эти и другие преимущества способствовали широкому распространению семенных растений. Крупные споровые растения вымирают в пермском периоде в связи с иссушением климата.
Вопрос 3. Охарактеризуйте эволюцию животных в палеозойскую эру.
Животный мир в палеозойской эре развивался чрезвычайно бурно и был представлен большим количеством разнообразных форм. Пышного расцвета достигает жизнь в морях. В кембрийском периоде уже существуют все основные типы животных, кроме хордовых. Губки, кораллы, иглокожие, разнообразные моллюски, громадные хищные ракоскорпионы — вот неполный перечень обитателей кембрийских морей.
Пышного расцвета достигает жизнь в морях. В кембрийском периоде уже существуют все основные типы животных, кроме хордовых. Губки, кораллы, иглокожие, разнообразные моллюски, громадные хищные ракоскорпионы — вот неполный перечень обитателей кембрийских морей.
В ордовике продолжается совершенствование и специализация основных типов. Впервые обнаруживаются остатки животных, имевших внутренний осевой скелет, — бесчелюстных позвоночных, отдаленными потомками которых являются современные миноги и миксины. Рот этих своеобразных организмов представлял собой простое отверстие, ведущее в пищеваритель¬ный тракт. Передний отдел пищеварительной трубки был пронизан жаберными щелями, между которыми располагались опорные хрящевые жаберные дуги. Бесчелюстные питались организмами, обитающими в илистом дне рек и озер, и детритом (органическими остатками), засасывая пищу ртом. У части бесчелюстных возникло расчленение жаберных дуг, что позволило изменять просвет глотки с помощью жаберной мускулатуры и, следовательно, удерживать попавшую в пищеварительную трубку подвижную добычу.
Появление хватательного ротового аппарата — крупный ароморфоз — вызвало перестройку всей организации позвоночных.
Возникновение парных плавников — конечностей — следующий крупный ароморфоз в эволюции позвоночных.
В силурийском периоде на сушу вместе с псилофитами вышли первые дышащие воздухом животные — членистоногие. В водоемах продолжалось интенсивное развитие низших позвоночных. Предполагается, что позвоночные возникли в мелких пресноводных водоемах и лишь затем переселились в моря. Девонский период ознаменован освоением суши другими членистоногими — пауками; в конце периода появляются первые наземные позвоночные — амфибии (стегоцефалы). В каменноугольном периоде возникают рептилии (котилозавры), летающие насекомые и легочные моллюски. В последнем, пермском периоде палеозойской эры наблюдается быстрое развитие и увеличение систематических групп рептилий; появляются зверозубые пресмыкающиеся — предки млекопитающих.
Вопрос 4. Какие особенности строения позвоночных животных послужили предпосылками выхода их на сушу?
В силурийском периоде на сушу вместе с псилофитами вышли первые дышащие воздухом животные — членистоногие. В водоемах продолжалось интенсивное развитие низших позвоночных. Предполагается, что позвоночные возникли в мелких пресноводных водоемах и лишь затем переселились в моря. В девоне позвоночные представлены тремя группами: двоякодышащими, лучеперыми и кистеперыми рыбами. Именно кистепёрые рыбы дали развитие наземным позвоночным животным. Кистеперые рыбы были типично водными животными, но могли дышать атмосферным воздухом с помощью примитивных легких, представлявших собой выпячивания стенки кишки. Приспособиться к жизни на суше смогли только кистеперые рыбы. Их плавники представляли собой лопасти, состоящие из отдельных костей с прикрепленными к ним мышцами (рис. 2). С помощью плавников кистеперые рыбы — крупные животные длиной от 1,5 до нескольких метров — могли ползать по дну. Таким образом, они имели две основные предпосылки для перехода в наземную среду обитания — мускулистые конечности и легкие. В конце девона кистеперые рыбы дали начало первым земноводным — стегоцефалам.
В водоемах продолжалось интенсивное развитие низших позвоночных. Предполагается, что позвоночные возникли в мелких пресноводных водоемах и лишь затем переселились в моря. В девоне позвоночные представлены тремя группами: двоякодышащими, лучеперыми и кистеперыми рыбами. Именно кистепёрые рыбы дали развитие наземным позвоночным животным. Кистеперые рыбы были типично водными животными, но могли дышать атмосферным воздухом с помощью примитивных легких, представлявших собой выпячивания стенки кишки. Приспособиться к жизни на суше смогли только кистеперые рыбы. Их плавники представляли собой лопасти, состоящие из отдельных костей с прикрепленными к ним мышцами (рис. 2). С помощью плавников кистеперые рыбы — крупные животные длиной от 1,5 до нескольких метров — могли ползать по дну. Таким образом, они имели две основные предпосылки для перехода в наземную среду обитания — мускулистые конечности и легкие. В конце девона кистеперые рыбы дали начало первым земноводным — стегоцефалам.
Рис. 2. Скелет парного плавника кистеперой рыбы и стегоцефала:
2. Скелет парного плавника кистеперой рыбы и стегоцефала:
А — плечевой пояс и плавник кистеперой рыбы;
Б — внутренний скелет плавника;
В — скелет передней конечности стегоцефала:
1 — элемент, соответствующий плечевой кости;
2 — элемент, соответствующий лучевой кости;
8 — элемент, соответствующий локтевой кости;
4, 5, 6 — кости запястья; 7 — фаланги пальцев.
ТРИЛОБИТЫ — ОБИТАТЕЛИ ПАЛЕОЗОЯ | Наука и жизнь
Трилобиты — морские членистоногие, которых на Земле уже нет. Они полностью вымерли более 200 миллионов лет назад. Временем их появления, расцвета и гибели была вся палеозойская эра. А она началась 550 миллионов лет назад и длилась около 300 миллионов лет. Временами (особенно в раннем палеозое) трилобитов было так много, что по численности и разнообразию видов они превосходили большинство обитавших тогда групп многоклеточных животных. Поэтому если мезозойская эра (примерно 70-230 миллионов лет назад) может быть названа эрой динозавров, то палеозойская — эрой трилобитов.
Членистоногие в наше время — это самый процветающий, самый многочисленный тип животных. Число известных видов приближается к трем миллионам. Их намного больше, чем всех остальных многоклеточных животных, вместе взятых. Раки, крабы, скорпионы, клещи, пауки, многоножки, насекомые — все относятся к членистоногим. А наиболее просто устроенными из всех этих летающих, ползающих, бегающих существ были трилобиты, о которых и пойдет рассказ.
Трилобиты сохранились на Земле только в виде окаменелых остатков. Понять, каким был образ их жизни, что помогло трилобитам просуществовать на Земле почти 300 миллионов лет, палеонтологам, биологам помогают наблюдения за нынешними членистоногими, которые сейчас распространены практически везде. Они живут на земле и под землей, в пресной воде и в соленой, в лужах и на дне океанов, на снегу и в горячих источниках, встречаются в Арктике и Антарктике, горах и пустынях. Членистоногие освоили, наверное, все способы питания, возможные для многоклеточных животных. Среди них есть хищники, растительноядные, паразиты, фильтраторы, собиратели отмершей органики. Членистоногие могут питаться такими трудноперевариваемыми веществами, как целлюлоза, воск и рог, могут потреблять углеводороды нефти и даже, возможно, метан. Словом, они удивительно хорошо приспособлены к жизни. Потому и заселяют Землю вот уже 500 миллионов лет. А трилобиты были, видимо, одними из самых древних среди них.
Среди них есть хищники, растительноядные, паразиты, фильтраторы, собиратели отмершей органики. Членистоногие могут питаться такими трудноперевариваемыми веществами, как целлюлоза, воск и рог, могут потреблять углеводороды нефти и даже, возможно, метан. Словом, они удивительно хорошо приспособлены к жизни. Потому и заселяют Землю вот уже 500 миллионов лет. А трилобиты были, видимо, одними из самых древних среди них.
Тело членистоногих покрыто хитиновым панцирем, жестким и очень устойчивым к химическим воздействиям. Панцирь не только защищает животное снаружи, но и служит для прикрепления внутренних органов, прежде всего развитой двигательной мускулатуры. Поэтому его можно считать своеобразным наружным скелетом этих животных. Для мелких и среднего размера членистоногих (длиной от долей миллиметра до нескольких сантиметров) прочности чисто хитинового панциря вполне достаточно. У более крупных (а трилобиты, некоторые виды которых достигали 80 сантиметров длины, могут считаться крупными членистоногими) панцирь пропитан еще минеральными солями, в основном карбонатом кальция, что придает ему особую прочность. Именно благодаря этой известковой пропитке панцири трилобитов, пролежав в земле не одну сотню миллионов лет, хорошо сохранились.
Именно благодаря этой известковой пропитке панцири трилобитов, пролежав в земле не одну сотню миллионов лет, хорошо сохранились.
Панцирь трилобитов условно может быть поделен как в продольном, так и в поперечном направлении на три части (из-за этого они и получили свое название). При делении в продольном направлении это — головной щит, туловище и хвостовой щит; в поперечном — осевая и две боковые части. Известью пропитана только спинная сторона панциря, а брюшная, на которой располагались конечности — органы движения, питания, дыхания и осязания, наоборот, была очень мягкой и нежной. В случае опасности, чтобы защитить мягкое брюшко, трилобиты могли сворачиваться. Интересно, что этому они научились не сразу. В кембрийском периоде (первый период палеозойской эры), когда они только-только появились и размножились, способностью сворачиваться обладали лишь немногие виды, а уже в следующем геологическом периоде — в ордовике — почти не было несворачивающихся видов. Возможно, что прежде в такой способности не было нужды, поскольку головоногих моллюсков (они стали главными врагами крупных морских членистоногих) тогда было еще очень мало. В ордовике головоногие сильно размножились и достигали порой гигантских размеров. Например, в море, которое в ордовикском периоде было на территории нынешней Ленинградской области, жили головоногие моллюски с раковиной пятиметровой длины.
В ордовике головоногие сильно размножились и достигали порой гигантских размеров. Например, в море, которое в ордовикском периоде было на территории нынешней Ленинградской области, жили головоногие моллюски с раковиной пятиметровой длины.
Свернутый панцирь трилобита Pliomera fischeri (вид спереди). Сворачиваясь, трилобиты защищали мягкое брюшко. Ордовикский период Ленинградской области. Фото А. Бронникова.
Окаменелые остатки трилобитов в больших количествах экспортируются из Ленинградской области. На снимке — коллекционные экземпляры, подготовленные к вывозу в Германию.
Трилобиты рода Asaphus в породе. Ордовикский период Ленинградской области.Уплощенный широкий панцирь трилобита Ptychopyge свидетельствует о том, что он, вероятно, вел малоактивный (ползающий) образ жизни. Фото А. Бронникова.
У представителей вида Asaphus kowalewskii глаза подняты на длинных стебельках. Это позволяло им глубоко погружаться в поисках пищи в донные осадки и при этом наблюдать за появлением врагов. Ордовикский период Ленинградской области. Фото А. Мазина.
Ордовикский период Ленинградской области. Фото А. Мазина.
У трилобитов рода Asaphus был толстостенный выпуклый панцирь с гладкой поверхностью. Это указывает на то, что они постоянно рылись в грунте.
По огромным пространствам океана трилобиты расселялись в виде маленьких планктонных личинок. В некоторых кембрийских местонахождениях Якутии эти личинки встречаются в больших количествах.
На то, чтобы выделить (препарировать) такой сложно устроенный панцирь (здесь показан Cybele sp. из ордовикского периода Ленинградской области), даже если он был заключен в сравнительно мягкую породу, уходят недели кропотливого ручного труда.
‹
›
У большинства современных членистоногих хорошо развиты глаза. Они бывают простые (с одной-двумя линзами) и сложные или фасеточные, состоящие из нескольких десятков, сотен или даже тысяч простых глазков. Такие же глаза были и у палеозойских трилобитов. Зрительные поверхности огромных фасеточных глаз трилобитов были ориентированы таким образом, что многие из этих животных могли видеть сразу на 360 градусов вокруг. Но только вдоль линии горизонта и на один-другой десяток градусов выше нее. Для наблюдения за «мертвым» пространством в зените в затылочной части головного щита трилобитов располагался маленький одиночный глазок.
Но только вдоль линии горизонта и на один-другой десяток градусов выше нее. Для наблюдения за «мертвым» пространством в зените в затылочной части головного щита трилобитов располагался маленький одиночный глазок.
Глаза у некоторых видов трилобитов, которые искали пищу, зарываясь в верхний слой ила, были подняты на длинных стебельках, как у современных ракообразных и брюхоногих моллюсков. Но в отличие от них трилобиты не могли стебельки глаз втягивать внутрь головы, поворачивать или складывать. Стебельки глаз трилобитов — это твердые выросты панциря, и они всегда стояли торчком и потому, конечно же, подвергались опасности быть поломанными. Но ломались они все же очень редко. Из сотен виденных мною панцирей трилобита Asaphus kowalewskii, у которого, наверное, самые высокие глаза, встретился только один экземпляр с прижизненно обломанным стебельком.
Все конечности (а их у трилобитов несколько десятков), как и остальная часть брюшка, были мягкими и потому сохраняются в ископаемом состоянии лишь в исключительных случаях. Впервые в России трилобит с сохранившимися конечностями найден в Якутии три года назад.
Впервые в России трилобит с сохранившимися конечностями найден в Якутии три года назад.
Хитин панциря членистоногих не может растягиваться. Поэтому их рост сопровождается несколькими линьками. Когда панцирь становится тесным, он лопается (обычно спереди и на спине) и животное его сбрасывает. В тот короткий период, когда старый панцирь сброшен, а новый еще не затвердел, размеры животного быстро увеличиваются. Поза, в которой происходит «вылупление» из старого панциря, очень характерная и разная у разных видов трилобитов. Для палеонтолога очень ценно найти панцирь в «линочном положении», потому что это свидетельствует о том, что порода, где он найден, не была переработана роющими животными или перемешана какими-то другими силами. Значит, здесь можно увидеть детали процесса формирования породы, можно восстановить условия среды на дне древнего водоема, в котором обитали трилобиты.
Поражает разнообразие форм панциря трилобитов: гладкие, бугристые, шипастые, с глазами огромными и редуцированными, низкими или поднятыми на длинных стебельках, с длинными ветвящимися выростами, с туловищем, состоящим из двух сегментов или из нескольких десятков, и так далее. Известно, что форма и степень расчлененности панциря у членистоногих связаны с их внутренней анатомией, говорят о преимущественном развитии тех или иных групп мышц. Все это позволяет судить об образе жизни и характере питания животных.
Известно, что форма и степень расчлененности панциря у членистоногих связаны с их внутренней анатомией, говорят о преимущественном развитии тех или иных групп мышц. Все это позволяет судить об образе жизни и характере питания животных.
Плоский широкий панцирь был у тех видов, которые медленно ползали по поверхности дна. Выпуклый с глубокими бороздами — у активно передвигавшихся по дну и зарывавшихся в грунт. Выпуклый толстостенный со сглаженной поверхностью — у тех видов, которые постоянно рылись в грунте. По панцирям некоторых трилобитов можно понять, что они вели планктонный образ жизни. У них маленькие размеры тела и огромные глаза, а при сворачивании по бокам панциря оставались большие незащищенные отверстия — места выхода длинных плавательных конечностей.
В одном и том же месте могли обитать десятки видов трилобитов с панцирями разнообразной формы. Это означает, что их питание и образ жизни сильно различались.
Раньше считалось, что трилобиты (кроме планктонных видов) могли питаться только заглатывая богатый органикой верхний слой грунта, поскольку у них были мягкие, казалось бы, не приспособленные к захвату жертвы конечности. Недавно появились новые данные, показывающие, что некоторые виды трилобитов, несомненно, были хищниками. Об этом говорит находка в Швеции. Там обнаружены следы каких-то живших в грунте животных и следы, оставленные трилобитами. При этом след трилобита накрывает след жившего в грунте животного, и тот обрывается. Следовательно, трилобиты данного вида разыскивали и поедали обитавших в грунте животных. В Якутии найдены трилобиты с сохранившимся содержимым кишечника. В нем обнаружены частицы тел донных животных — губок и брахиопод.
Недавно появились новые данные, показывающие, что некоторые виды трилобитов, несомненно, были хищниками. Об этом говорит находка в Швеции. Там обнаружены следы каких-то живших в грунте животных и следы, оставленные трилобитами. При этом след трилобита накрывает след жившего в грунте животного, и тот обрывается. Следовательно, трилобиты данного вида разыскивали и поедали обитавших в грунте животных. В Якутии найдены трилобиты с сохранившимся содержимым кишечника. В нем обнаружены частицы тел донных животных — губок и брахиопод.
Подобно многим морским членистоногим, трилобиты в своем развитии проходили стадию планктонной (то есть пассивно плавающей в толще воды) личинки. Именно благодаря своим маленьким, по виду совсем не похожим на взрослых животных личинкам трилобиты смогли расселиться на огромных пространствах палеозойских океанов.
Остатки трилобитов встречаются во многих местах России, там, где на дневную поверхность выходят палеозойские и особенно древнепалеозойские морские отложения. Наиболее известные из них — в Ленинградской области и в Восточной Сибири (в Якутии). Якутские трилобиты весьма многочисленны и разнообразны. Но их панцири почти всегда раздавлены и разделены по щиткам и сегментам. В Ленинградской области окаменелые остатки трилобитов находятся в меньших количествах. Зато среди них немало таких, которые поражают великолепной сохранностью. Многие панцири сохранили первоначальную форму и обычно имеют красивую каштаново-коричневую окраску. Ее придает им остаток не полностью разложившегося органического вещества. В местах утолщения панциря (где органического вещества было больше) видны темные пятна, а, например, зрительная поверхность глаз остается бесцветной и прозрачной. Известен случай, когда на панцире сохранилась прижизненная раскраска, именно раскраска, то есть узор, а не цвет.
Наиболее известные из них — в Ленинградской области и в Восточной Сибири (в Якутии). Якутские трилобиты весьма многочисленны и разнообразны. Но их панцири почти всегда раздавлены и разделены по щиткам и сегментам. В Ленинградской области окаменелые остатки трилобитов находятся в меньших количествах. Зато среди них немало таких, которые поражают великолепной сохранностью. Многие панцири сохранили первоначальную форму и обычно имеют красивую каштаново-коричневую окраску. Ее придает им остаток не полностью разложившегося органического вещества. В местах утолщения панциря (где органического вещества было больше) видны темные пятна, а, например, зрительная поверхность глаз остается бесцветной и прозрачной. Известен случай, когда на панцире сохранилась прижизненная раскраска, именно раскраска, то есть узор, а не цвет.
Особенности поиска и добычи остатков трилобитов зависят от типа местонахождения. Мягкие глинистые известняки Ленинградской области на открытом воздухе разрушаются легче, чем панцири трилобитов. Поэтому, как только обнажат пласт, трилобиты начинают как бы «выглядывать» из породы. Но здесь окаменелые остатки встречаются редко и на большом расстоянии одни от других. В Якутии твердые колкие известняки почти неотличимы по цвету и механическим свойствам от заключенных в них остатков трилобитов, и визуальный осмотр обнажений здесь обычно ничего не дает. Но когда окаменелости обнаружены, их обычно бывает много и в породе они распределены равномерно.
Поэтому, как только обнажат пласт, трилобиты начинают как бы «выглядывать» из породы. Но здесь окаменелые остатки встречаются редко и на большом расстоянии одни от других. В Якутии твердые колкие известняки почти неотличимы по цвету и механическим свойствам от заключенных в них остатков трилобитов, и визуальный осмотр обнажений здесь обычно ничего не дает. Но когда окаменелости обнаружены, их обычно бывает много и в породе они распределены равномерно.
Поиск и сбор трилобитов ведутся методичным последовательным дроблением больших блоков породы слой за слоем.
Чтобы найденные панцири отделить от породы, иногда используют различные пневмо- и виброинструменты, но чаще всего работают обычной стальной иглой. Работа долгая, кропотливая, требующая терпения и аккуратности.
Трилобиты Ленинградской области, благодаря своим эстетическим качествам и относительной легкости добычи, сейчас стали одним из главных объектов экспорта палеонтологических остатков из России. Интерес коллекционеров к ним очень большой, это, конечно же, хорошо, но связано и со многими бедами. Интенсивно разрабатываемые обнажения быстро нищают, а то и совсем гибнут. Отношение к уникальным окаменелостям у сборщиков обычно варварское, поскольку их интересует лишь комплектность добываемого панциря. При этом наука безвозвратно теряет возможность получить очень важную информацию о последовательности залегания видов в слоях и о сопутствующей фауне. А порой искажается и морфология трилобитов, поскольку случается, что сборщики заменяют отсутствующие фрагменты частями панцирей трилобитов других видов, а то и просто лепят из пластика.
Интерес коллекционеров к ним очень большой, это, конечно же, хорошо, но связано и со многими бедами. Интенсивно разрабатываемые обнажения быстро нищают, а то и совсем гибнут. Отношение к уникальным окаменелостям у сборщиков обычно варварское, поскольку их интересует лишь комплектность добываемого панциря. При этом наука безвозвратно теряет возможность получить очень важную информацию о последовательности залегания видов в слоях и о сопутствующей фауне. А порой искажается и морфология трилобитов, поскольку случается, что сборщики заменяют отсутствующие фрагменты частями панцирей трилобитов других видов, а то и просто лепят из пластика.
Специалисты считают, что трилобиты окончательно вымерли около 230 миллионов лет назад. Вероятность того, что какие-то их потомки дожили до настоящего времени, чрезвычайно мала. Для этого они должны были бы в корне изменить образ жизни (например, перейти к паразитизму, при котором происходит очень сильное упрощение строения и индивидуального развития) и видоизмениться, так что их невозможно узнать.
Однако в современной фауне есть группа членистоногих, которые по внешнему виду удивительно похожи на поздних трилобитов. Это равноногие раки, или изоподы. При взгляде на панцирь сверху некоторых из них трудно отличить от трилобитов, выдают только толстые, состоящие из крупных сегментов антенны. Изоподы, подобно трилобитам, обладают способностью сворачиваться и имеют большие фасеточные глаза. Например, обыкновенные мокрицы (наземные изоподы), если их потревожить, сворачиваются в плотный, подобный горошине шарик, способный кататься, отскакивать при ударе о твердые предметы и т.п. Сходство изопод и трилобитов обусловлено не столько родством (в общем-то довольно далеким — они принадлежат к разным классам типа членистоногих), сколько похожим принципом построения тела, а значит, одинаковым образом жизни. Очень может быть, что в экологии моря изоподы заняли освободившуюся нишу вымерших трилобитов.
Палеозойская эра. Как развивалась жизнь на Земле
Палеозойская эра
Палеозойская эра началась 600 млн. лет тому назад. Различают шесть ее периодов: кембрийский, ордовикский, силурийский, девонский, каменноугольный, пермский.
лет тому назад. Различают шесть ее периодов: кембрийский, ордовикский, силурийский, девонский, каменноугольный, пермский.
В палеозое прошли два горообразовательных процесса: каледонский (в кембрии — нижнем девоне) и герцинский (в верхнем карбоне — перми), в результате которых неоднократно изменялись очертания материков и морей. Палеозойские отложения представлены главным образом глинами, известняками, доломитом, мергелями, песчаниками, солью и каменным углем.
На протяжении палеозойской эры органический мир завоевал сушу. Среди животных появились первые позвоночные, среди растений — споровые и хвойные.
Кембрийский период
Кембрийский период получил свое название от графства Кембрий (Англия), так как здесь были впервые описаны отложения кембрийского периода.
На месте Северной Америки и Гренландии в кембрии находился материк Лавренция. Южнее Лавренции простирался Бразильский материк.
Африканский материк включал в то время Африку, Мадагаскар и Аравию. Севернее его был расположен небольшой Русский материк.
Довольно широкий морской бассейн отделял Русский материк от Сибирского, находившегося на месте современной Западной Сибири. Там, где сейчас Китай, был Китайский материк, а на юге от него — громадный Австралийский материк, охватывавший территорию современной Индии и Западной Австралии. В это время сформировались Северные Аппалачи, горы Чингизтау в Казахстане и Салаиро-Саянский хребет.
Из кембрийских отложений наиболее распространены известняки, доломиты, глинистые сланцы. Часто встречаются и отложения мелководных лагун: песчаники и глины с пластами каменной соли и гипса.
В северном полушарии можно выделить несколько зон, в которых климат был сухим и жарким. В этих местах откладывались мощные слои солей и гипса. Известняковые отложения Австралии с трещинами высыхания также свидетельствуют о жарком и сухом климате на Австралийском материке в кембрийский период.
Климат Африканского материка был, по-видимому, теплым и влажным. В Южной Австралии, Китае и Норвегии были ледники. При сравнении берегов кембрийских морей с берегами современных морей можно сделать вывод о том, что большая часть земной поверхности в кембрийский период представляла собой сушу. В мелководных морях было множество вулканических островов. Кембрийская флора и фауна распространились на земном шаре из кембрийских тропических морей.
При сравнении берегов кембрийских морей с берегами современных морей можно сделать вывод о том, что большая часть земной поверхности в кембрийский период представляла собой сушу. В мелководных морях было множество вулканических островов. Кембрийская флора и фауна распространились на земном шаре из кембрийских тропических морей.
Вся жизнь в кембрийский период была тесно связана с водной средой. На суше жизни еще не было. Из растений кембрийского периода известны жившие в морской воде известковые водоросли. После их отмирания образовывались скопления известняка, известные под названием онкоидов. Без сомнения, в кембрийских морях были и другие водоросли: сине-зеленые, красные. Но у них не имелось твердых образований, поэтому до нашего времени их остатки не сохранились.
Водоросли, выделяя свободный кислород, значительно изменили состав кембрийской атмосферы. Это создало возможность для развития других форм жизни, в частности тех групп животных, которые потребляют свободный кислород. Наши знания о животном мире кембрийского периода весьма ограничены. Кембрийские породы неоднократно подвергались метаморфизации, приведшей к исчезновению многих отпечатков и окаменелостей. Многие кембрийские отложения еще не изучены. Лучше всего изучены животные, обитавшие в мелководных морях близ побережья. Фауна же относительного глубоководья и открытого океана нам почти неизвестна.
Наши знания о животном мире кембрийского периода весьма ограничены. Кембрийские породы неоднократно подвергались метаморфизации, приведшей к исчезновению многих отпечатков и окаменелостей. Многие кембрийские отложения еще не изучены. Лучше всего изучены животные, обитавшие в мелководных морях близ побережья. Фауна же относительного глубоководья и открытого океана нам почти неизвестна.
Наряду с многочисленными представителями одноклеточных организмов в морях кембрийского периода обитали и колониальные организмы. Сохранилось множество окаменевших ходов, проложенных какими-то червеобразными существами. Из кембрийских отложений известны лишь отдельные представители распространенного в наше время типа моллюсков. Раковины двустворчатых и брюхоногих моллюсков похожи на современные пресноводные формы. Из головоногих моллюсков известны крупные роговые организмы, раковины которых делились на камеры-трубки длиной около 8 мм и шириной 1 мм. Внутри камер размещалась тонкая трубка (сифон).
Очень распространены были в то время брахиоподы — животные, раковина которых открывалась снизу вверх. На раковинах многих брахиопод имелись боковые выступы. Внутри раковины находились мясистые органы, выполнявшие дыхательные и выделительные функции. Маленькое червеобразное тело этих животных украшали две жаберные «руки». Все брахиоподы — морские животные. Некоторые из них достигали 25 см в длину. У одних были известковые раковины, у других — хитиновые, по своему составу напоминающие хитиновые скелеты и панцири современных насекомых, раков и других беспозвоночных. Образовывались подобные раковины из выделяемого кожей углеобразного азотсодержащего вещества.
На раковинах многих брахиопод имелись боковые выступы. Внутри раковины находились мясистые органы, выполнявшие дыхательные и выделительные функции. Маленькое червеобразное тело этих животных украшали две жаберные «руки». Все брахиоподы — морские животные. Некоторые из них достигали 25 см в длину. У одних были известковые раковины, у других — хитиновые, по своему составу напоминающие хитиновые скелеты и панцири современных насекомых, раков и других беспозвоночных. Образовывались подобные раковины из выделяемого кожей углеобразного азотсодержащего вещества.
Характерные брахиоподы кембрийского периода — лингула и оболюс. Они обладали хитиновыми раковинами, пропитанными соединениями кальция. Примечательно, что брахиоподы дожили до наших дней, почти не изменившись.
Из животных, которые, подобно брахиоподам, прикреплялись к морскому дну, можно указать еще морских губок. Кембрийские морские губки относятся к семействам четырех- и шестилучевых губок. Они, в отличие от современных, имели иглы, свободно расположенные в мягкой ткани.
Представителями иглокожих являлись многочисленные роды так называемых цистоидей. Тело этих животных покрывал панцирь из известковых пластинок.
Настоящие кораллы в кембрийских отложениях не найдены. Наиболее характерная группа животных для кембрийского периода — археоциаты. Они были от нескольких сантиметров до метра высотой. Вымерли археоциаты в кембрии. По своему строению животные походили на кишечнополостных очень примитивной организации. Название «археоциаты» означает «древние чаши», и, действительно, по внешнему виду они напоминали бокалы или чаши. В теле животных имелась центральная полость, окруженная двойными стенками с вертикальными и горизонтальными перегородками и многочисленными отверстиями. По мнению многих ученых, археоциаты — предки губок и кораллов, которые в дальнейшем, заселив те участки моря, где обитали археоциаты, вытеснили своих предшественников, так как обладали гораздо более современной организацией. В наше время от этих удивительных животных остались лишь отдельные известковые рифы.
Археоциаты и трилобиты.
Характерными представителями животного мира всей палеозойской эры были трилобиты — морские жабродышащие членистоногие животные. Спинная часть тела трилобита делится по длине на среднюю выступающую часть и более плоские боковые части. По ширине тело делится на три отдела — щита: головной (на нем были расположены глаза), грудной (движущиеся сегменты) и брюшной (с разным числом более или менее сросшихся сегментов). На всех грудных и брюшных сегментах имелись двуветвистые парные конечности. Хвостовой щит имел вид круглого придатка, длинных шипов либо небольшой пластинки. Брюшная сторона тела трилобитов была мягкой, кожистой и, видимо, мало содержала соединений кальция. Подрастая, животные линяли. Отсутствие челюстных зубов говорит о том, что трилобиты питались органическим илом, а яйцеобразное тело и вытянутый хвостовой щит — о том, что они жили преимущественно в стоячей воде. Одни из них были подвижными и быстро бегали по дну, другие — малоподвижными, некоторые жили в иле. Приспосабливаясь к различным условиям обитания, трилобиты постепенно изменялись. У тех из них, которые зарывались в ил, были глаза на стебельках, как у современных раков, те же, которые жили в мутной воде, совсем утратили орган зрения. В среднем кембрии в Америке появляются крупные овальные трилобиты с гладкими щитами. Их головные и хвостовые щиты были почти равной величины, мало расчлененными. В иле жили и черви. В морях кембрийского периода было необычайно много медуз.
Приспосабливаясь к различным условиям обитания, трилобиты постепенно изменялись. У тех из них, которые зарывались в ил, были глаза на стебельках, как у современных раков, те же, которые жили в мутной воде, совсем утратили орган зрения. В среднем кембрии в Америке появляются крупные овальные трилобиты с гладкими щитами. Их головные и хвостовые щиты были почти равной величины, мало расчлененными. В иле жили и черви. В морях кембрийского периода было необычайно много медуз.
Нам не известны предковые формы, от которых произошли беспозвоночные кембрийского периода. Все они появились в протерозое, но, не обладая твердыми скелетами или раковинами, не оставили после себя следов. Возможно, следы исчезли в результате различных геологических процессов. Вероятнее всего, предками моллюсков, трилобитов, брахиопод являлись червеобразные существа, обитавшие в морях раннего протерозоя.
Продолжительность кембрийского периода — 70 млн. лет.
Основные полезные ископаемые этого времени: медь, серный колчедан, платина, золото, мышьяк, полиметаллы, гипсы, газы, каменная соль.
Ордовикский период
Отложения ордовика были выделены в Англии и описаны английским геологом Р. Мурчисоном. По решению XXI сессии Международного геологического конгресса ордовик выделен в самостоятельную систему.
Материк Лавренция в ордовикский период распался на четыре больших и ряд более мелких островов. На месте Русского материка образовались два больших острова, разделенных узким проливом. Почти половина территории Сибирского и Китайского материков была залита мелким морем.
В южном полушарии образовался огромный материк — Гондвана, включавший современную Южную Америку, южную часть Атлантического океана, Африку, Индийский океан, Австралию, Северную Азию. Начинают формироваться Северный Тянь-Шань, Алтай, Австралийские Кордильеры, Западно-Сибирские хребты.
В существовавших на территории Урала, Чукотки и Кордильер морских бассейнах действовали тысячи вулканов, давших мощные отложения вулканических пород.
Среди пород ордовика преобладают морские отложения — песчаники, известняки, сланцы. По сравнению с отложениями кембрия среди ордовикских отложений меньше лагунных образований — гипсов, солей, известняков, доломитов. Климат в ордовикский период становится более теплым и мягким, о чем свидетельствует большое распространение известняков: строматопоровых, коралловых, криноидных, трилобитовых и цефалоподовых. Значительно возросла площадь моря. Экваториальное первичное море затопило обширные площади кембрийских материков.
По сравнению с отложениями кембрия среди ордовикских отложений меньше лагунных образований — гипсов, солей, известняков, доломитов. Климат в ордовикский период становится более теплым и мягким, о чем свидетельствует большое распространение известняков: строматопоровых, коралловых, криноидных, трилобитовых и цефалоподовых. Значительно возросла площадь моря. Экваториальное первичное море затопило обширные площади кембрийских материков.
Полностью исчезает южная сухая зона. Сокращается площадь северных пустынь. В результате этих изменений изменяется также животный и растительный мир. Горные материки, вклинившиеся между морскими бассейнами, препятствовали расселению животных и растений по земному шару. Вот почему фауна и флора европейского ордовика отличаются от индийской и восточно-азиатской.
В конце кембрийского периода вулканические извержения заполняют туфами и лавами морские бассейны. Одновременно морское дно значительно опускается. Все это обусловило накопление мощных толщ осадочных пород, в частности черного ила, состоящего из вулканического пепла, песка, обломочных пород.
Почти не претерпели изменений в этот период водоросли. Морская фауна характеризовалась таким богатством форм, что ордовикский период представляется нам важнейшей эпохой всей истории Земли. Именно в ордовике сформировались основные типы морских организмов. По сравнению с кембрием значительно возрастает количество трилобитов. В ордовике много крупных трилобитов (до 50–70 см) появляется и в Европе. Это свидетельствует о том, что они себя хорошо чувствовали в новых условиях.
Благодаря миграции фауны с запада на восток и приспособлению к новым условиям в ордовикских морях появляется 77 новых родов трилобитов. Внешнее строение тела свидетельствует о том, что трилобиты вели разный образ жизни. Их глаза имели от 10 до 1200 фасеток. Встречались и слепые трилобиты. Число туловищных члеников (сегментов) колебалось у разных видов от 2 до 29. Тело было покрыто шипами для защиты от врагов или совершенно гладким, хорошо приспособленным к ползанию в иле. Иногда тело покрывали длинные острые шипы, увеличивавшие его поверхность, что позволяло животному свободно держаться в воде.
В ордовикских отложениях найдены все важнейшие группы животных, обитавших в морях в более позднее время. В рыхлых зеленых песчаниках вблизи Ленинграда встречается много ядер фораминифер. В черных сланцах находят радиолярий. Довольно многочисленны в ордовикских отложениях губки, в скелете которых имелись кремнеземные иголочки: циатофикас, высотой до 12 см, и брахиоспондия, высотой до 30 см с 12 корневыми отростками.
Морские губки были четырех- и шестилучевые. Особенно красивыми иглами отличались четырехлучевая эутаксиксадима шестилучевая рецептакулитида. Тело первой, величиной с вишню, обладало волокнистой структурой. Каждое из волоконец представляло собой шестигранную трубку, состоявшую из маленьких четырехлучевых иголочек, сплетавшихся так тесно, что отделить хотя бы одну из них очень трудно. Шестилучевые губки появились впервые в раннем ордовике. Круглое, плоское, грушевидное или блюдцевидное тело этого существа было покрыто щитом из ромбических пластинок. Под каждой пластинкой находился пустой заостренный столбик. Столбики соединялись с внутренними пластинками. Все это составляло внутреннюю раковину.
Столбики соединялись с внутренними пластинками. Все это составляло внутреннюю раковину.
Появились первые кораллы, однако особого значения в природе они пока не имели. Из моллюсков самыми распространенными были наутилоидеи и гастроподы. Раковины наутилоидей были прямыми. Сам моллюск помещался в жилой камере, остальные камеры заполнял газ. Заполняя эти камеры водой, моллюск мог погружаться на значительные глубины, а вытесняя воду газом, всплывать на поверхность. Появились граптолиты, имевшие вид веточек, спиралей, петелек. Они жили колониями, прикреплялись к водорослям или свободно плавали с помощью пузыря.
В ордовикский период впервые появляются мшанки и табуляты, особенно распространившиеся в силурийский период.
Бурно развиваются брахиоподы. Если в кембрии их было 18 родов, то в ордовике существовал уже 41 род этих животных.
Иглокожих в ордовике представляли многие виды цистоидей, тело которых покрывал известковый панцирь. Круглое ротовое отверстие защищала пластинка. Значительное распространение форм цистоидей дает основание считать их предками морских лилий, морских ежей и морских звезд, поскольку разные цистоидеи имели много общего в строении с этими большими группами животных.
Значительное распространение форм цистоидей дает основание считать их предками морских лилий, морских ежей и морских звезд, поскольку разные цистоидеи имели много общего в строении с этими большими группами животных.
Ордовикский период продолжался 60 млн. лет. В его отложениях залегают полиметаллические и железные руды, фосфориты, горючие сланцы, строительное материалы, нефть.
Силурийский период
Силурийский период назван в честь древнего кельтского племени силуров. Он подразделяется на два отдела: нижний и верхний силур. В силуре в северном полушарии вновь образовался материк Лавренция. Наступавшее с юга на территорию Гондваны море образовало большой мелкий залив, почти разделивший Гондвану на две части. Другие материки и острова мало изменили свои очертания, приобретенные в кембрии.
Характернейшая особенность силурийского периода — постепенное опускание суши под воду. Море размывало многие сформировавшиеся прежде горные массивы и залило огромные площади. Медленное погружение суши и опускание дна океана привели к накоплению осадочных пород — мергелей, песчаников, доломитов, граптолитовых сланцев, брахноподовых и корраловых известняков.
В конце силура происходят горообразовательные процессы, благодаря которым образовались Скандинавские, Кембрийские горы, а также горы Южной Шотландии и Восточной Гренландии. На месте Сибири образовался большой материк Ангарида, частично сформировались Кордильеры. Климат на протяжении всего силурийского периода, вероятно, был теплым, влажным и лишь в конце силура на севере он стал сухим и жарким.
В силурийском периоде жизнь проникает на сушу. Первые наземные растения, остатки которых найдены в силурийских отложениях, получили название псилофитов, что значит безлистые, голые растения. Высотой они были не более полуметра. По внешнему виду растения напоминали современные сфагновые мхи, однако имели более простую организацию. По своему строению псилофиты похожи на бурые водоросли, от которых они, по-видимому, и произошли. Росли псилофиты в увлажненных местах или в мелких водоемах.
Ветвление у псилофитов было дихотомическим, то есть каждая ветка разделялась на две. Их тело еще четко не расчленялось на корневую и стебельную части. Вместо корней у них были отростки — ризоиды, которыми они прикреплялись к почве. Роль листьев выполняла чешуя. На концах веток псилофитов располагались органы размножения — спорангии, в которых развивались споры.
Их тело еще четко не расчленялось на корневую и стебельную части. Вместо корней у них были отростки — ризоиды, которыми они прикреплялись к почве. Роль листьев выполняла чешуя. На концах веток псилофитов располагались органы размножения — спорангии, в которых развивались споры.
Среди растений силурийских водных бассейнов преобладали водоросли: зеленые, сине-зеленые, красные, сифоновые, бурые, почти не отличавшиеся по своему строению от современных водорослей. Такое сходство натолкнуло некоторых исследователей на мысль о том, что в отдельных частях современных океанов температура, соленость и другие особенности воды сохранились такими же, какими они были в то отдаленное время.
Животный мир силурийского периода представлен главным образом теми же типами беспозвоночных, которые жили и в ордовике. Довольно распространенными были трилобиты (свыше 80 видов), моллюски (свыше 760 видов), брахиоподы (свыше 290 видов) и морские лилии, чашечки которых имели характерные для цистоидей ромбические поры. В позднем силуре появляются многочисленные представители морских звезд и морских ежей.
В позднем силуре появляются многочисленные представители морских звезд и морских ежей.
Среди силурийских двустворчатых моллюсков большое значение приобретают таксодонты, гетеродонты, десмодонты. Характерной особенностью некоторых из этих животных было то, что их створки загибались в противоположные стороны.
В солоноватоводных бухтах обитало множество тонкораковинных форм.
Брюхоногие моллюски силура выделялись весьма любопытными особенностями. У подавляющего большинства раковина была завернута вправо. Кроме того, некоторые из них имели шаровидную раковину с разрезом посредине, постепенно зараставшим либо превращавшимся в ряд дырочек.
Значительно распространились в морях силурийского периода головоногие моллюски. Представители небольшого рода — вольбортелла — с роговой раковиной, жившие в кембрийский и ордовикский периоды, дали многочисленных потомков (крупных и малых) с округлыми и гладкими известняковыми раковинами. Это свидетельствует об их большой подвижности.
В отличие от брюхоногих моллюсков, тело которых почти полностью заполняло раковину, головоногие моллюски жили в камере, отделенной от других камер перегородкой. В перегородках между нежилыми камерами имелись круглые отверстия, через которые проходила ткань в виде тяжа, так называемый сифон.
К наступлению половой зрелости тело моллюска полностью заполняло жилую камеру. Отложив яйца, моллюск сжимался, и жилая камера становилась слишком большой для моллюска; тогда и возникала перегородка, уменьшавшая объем жилой камеры. Моллюск снова рос, увеличивалась жилая камера, и со временем возникала новая перегородка.
Самые известные представители головоногих — ортоцерасы. Их мягкое тело напоминало современных осьминогов, но в отличии от осьминогов ортоцерасы имели длинную прямую раковину, сильно напоминавшую прямой рог. Отсюда и их название «ортоцерасы», что в переводе значит «прямой рог». Их длина достигала 1 метра. Плавали ортоцерасы раковиной вперед, а в спокойном состоянии повисали с помощью воздушных камер и хватательных щупалец, распуская их как парашюты. Ортоцерасы — предки всех головоногих, имевших перегородки. Их потомок — наутилус — живет и в настоящее время.
Ортоцерасы — предки всех головоногих, имевших перегородки. Их потомок — наутилус — живет и в настоящее время.
В силурийский период наряду с трилобитами появляется своеобразная группа животных, тело которых было покрыто плотным панцирем с многочисленными шипами и состояло из сегментов (5 головных, 7 грудных и 6 брюшных) и овального хвостового плавника или концевой иглы. Этих животных называют ракоскорпионами. Подвижные, хорошо вооруженные, они являлись подлинными властителями силурийских морей.
У наиболее характерного представителя ракоскорпионов — эвриптеруса — на ногах были иглы. У птериготуса первая пара ног преобразовалась в длинные клешни. На конце тела у него имелись шипы, которыми он убивал свою добычу.
Ракоскорпион птериготус.
В позднем силуре появляются первые животные, дышавшие легкими. Близкие родственники современных скорпионов, они, однако, имели много общего с ракоскорпионами, то есть были переходной группой от ракоскорпионов к современным скорпионам.
Из представителей кораллов самыми распространенными являлись табуляты — червеобразные животные с известняковыми трубочками. Они жили колониями. Трубочки разделялись на камеры перегородками. Иногда, помимо перегородок, у них были еще длинные ряды коротких шипов или продольных ребер.
В среднем силуре появляются первые представители настоящих кораллов. Они жили отдельными особями. Их чашечка высотой до 20 см имела прочную наружную стенку. У одних кораллов было четкое четырехлучевое строение, у других — двусторонне-симметричное строение, лежащее в основе строения всех кораллов и наблюдающееся даже в зародышевых формах современных кораллов. От четырех отрядов ордовикских плавающих остракод произошли 23 рода силурийских, величиной 22–80 мм. Среди иглокожих в силуре появляются настоящие бластоидеи, офиуры, морские звезды, настоящие морские ежи.
Силурийские рыбы еще не имели внутреннего костного скелета. Их тело и ротовая полость были сплошь покрыты маленькими кожными зубами. Среди рыб встречались костно-щитковые, бесщитковые и разнощитковые. В позднем силуре появились настоящие челюстные рыбы с парными плавниками и сложным скелетом.
Среди рыб встречались костно-щитковые, бесщитковые и разнощитковые. В позднем силуре появились настоящие челюстные рыбы с парными плавниками и сложным скелетом.
В среднем силуре от Европы до Сибири, от Канады до Аргентины распространились конусовидные, прямые или свернутые в спираль граптолиты. В конце силура они почти полностью вымерли. Близкие родственники иглокожих — граптолиты большими группами прикреплялись ко дну, скалам, водорослям. У отдельных граптолитов были нежные парашюты, благодаря которым они свободно плавали в морской воде. Их наружные скелеты состояли из хитинообразного вещества. Животные обитали главным образом вблизи низких берегов, в лагунах, на небольших глубинах, где откладывались глинистые осадки, богатые органическими веществами.
Когда в конце силура в результате тектонических движений берега поднялись, вблизи их началось отложение грубого обломочного материала. Усилился прибой. Сложившиеся условия оказывали отрицательное влияние на граптолитов, поэтому их жизненный ареал значительно сократился. Вблизи берегов стали появляться кораллы, брахиоподы, мшанки, для которых новые условия были чрезвычайно благоприятными. Новые роды рыб наутилоидей питались граптолитами, что также привело к значительному уменьшению их численности. Граптолитами питались, вероятно, и ракоскорпионы. Когда же в начале девона появилось множество хороших пловцов — позвоночных и аммоноидей, граптолиты совершенно исчезли.
Вблизи берегов стали появляться кораллы, брахиоподы, мшанки, для которых новые условия были чрезвычайно благоприятными. Новые роды рыб наутилоидей питались граптолитами, что также привело к значительному уменьшению их численности. Граптолитами питались, вероятно, и ракоскорпионы. Когда же в начале девона появилось множество хороших пловцов — позвоночных и аммоноидей, граптолиты совершенно исчезли.
Основные полезные ископаемые силурийского периода: железные руды, золото, медь, горючие сланцы, фосфориты и барит.
Силурийский период длился 35 млн. лет.
Девонский период
Девонские отложения впервые были описаны в английском графстве Девоншир. Девонский период подразделяется на три отдела: нижний, средний и верхний. В девоне северные материки образовали единый большой материк Атлантию, восточнее которого находилась Азия. Продолжает существовать Гондвана. Огромные материки были перегорожены горными хребтами, которые, разрушаясь, заполняли обломками впадины между горами. Климат стал сухим и жарким. Пересыхали озера и лагуны, а входившие в состав их вод соли и гипсы выпадали в осадок, образуя соленосные и гипсоносные толщи. Усиливается вулканическая деятельность.
Климат стал сухим и жарким. Пересыхали озера и лагуны, а входившие в состав их вод соли и гипсы выпадали в осадок, образуя соленосные и гипсоносные толщи. Усиливается вулканическая деятельность.
В среднем девоне море вновь наступает на сушу. Возникают многочисленные впадины. Их постепенно заливает море. Климат становится теплым и влажным. В верхнем девоне моря снова обмелевают, возникают небольшие горы, которые в дальнейшем были почти полностью разрушены. Наиболее характерные отложения девонского периода — континентальные красные песчаники, сланцы, гипсы, соль, известняки.
Существенным образом изменились физико-географические условия, приведшие к изменению флоры и фауны.
В водах девонских морей и океанов жили многочисленные водоросли: сифоновые, сине-зеленые, красные, в лагунах — харовые.
Появившиеся в силурийский период псилофиты в раннем девоне уже имели более сложную организацию. Их тело довольно четко разделялось на корень, стебель и ветки. От них в среднем девоне произошли примитивные папоротники. У псилофитов уже был деревянистый стебель. Ветки этих растений начинают выполнять различные функции, а их концевые части постепенно превращаются в рассеченные листья, с помощью которых осуществляется фотосинтез. Произрастают и другие потомки псилофитов: плаунообразные и членистостебельные, с более сложной организацией, чем у псилофитов. Они постепенно вытесняют своих предков, захватывая их места и расселяясь во влажных местностях, в мелких лагунах и болотах. В верхнем девоне псилофиты исчезают. Появляются первые семенные папоротники, кордаитовые и настоящие папоротники.
У псилофитов уже был деревянистый стебель. Ветки этих растений начинают выполнять различные функции, а их концевые части постепенно превращаются в рассеченные листья, с помощью которых осуществляется фотосинтез. Произрастают и другие потомки псилофитов: плаунообразные и членистостебельные, с более сложной организацией, чем у псилофитов. Они постепенно вытесняют своих предков, захватывая их места и расселяясь во влажных местностях, в мелких лагунах и болотах. В верхнем девоне псилофиты исчезают. Появляются первые семенные папоротники, кордаитовые и настоящие папоротники.
Споровые псилофиты, примитивные папоротники, плауновые, членистостебельные произрастали в увлажненных и болотистых местах, образуя густые заросли. Они достигали 30 м высоты и одного метра толщины. Растения размножались спорами, прораставшими в росток лишь во влажной среде.
У первых семенных растений на верхушках специализированных листьев были семенные зачатки, лежавшие открыто на листьях. Поэтому растения и получили название голосеменных. Они представляли собой уже настоящие деревья с настоящими листьями и органами размножения в виде шишек. Голосеменные могли размножаться непосредственно на суше, поскольку для прорастания семян водная среда не нужна. Кроме того, семена являются многоклеточным органом со значительным количеством запасных питательных веществ, которые обеспечивают зародыш в начале его жизни всем необходимым, а семенной покров хорошо предохраняет его от воздействия неблагоприятных условий. Все это дало возможность голосеменным широко распространиться на суше. И хотя споровые растения продолжали существовать, господствующее положение среди растений постепенно занимают голосеменные.
Они представляли собой уже настоящие деревья с настоящими листьями и органами размножения в виде шишек. Голосеменные могли размножаться непосредственно на суше, поскольку для прорастания семян водная среда не нужна. Кроме того, семена являются многоклеточным органом со значительным количеством запасных питательных веществ, которые обеспечивают зародыш в начале его жизни всем необходимым, а семенной покров хорошо предохраняет его от воздействия неблагоприятных условий. Все это дало возможность голосеменным широко распространиться на суше. И хотя споровые растения продолжали существовать, господствующее положение среди растений постепенно занимают голосеменные.
Сухой жаркий климат на материках привел к высыханию многих рек, озер, болот, лагун, мелких внутриконтинентальных морей. Из водных животных выживали лишь те, которые, помимо жабер, позволявших им жить в воде, обладали еще и легкими. При высыхании водоемов они могли дышать атмосферным воздухом. К ним относятся прежде всего двоякодышащие рыбы, имевшие роговые зубы и острые ребра. В 1870 г. в двух небольших реках Австралии были выявлены живые экземпляры двоякодышащих рыб, строение которых сильно напоминало их ископаемых предков. Впоследствии в Африке и Южной Америке также были найдены живые двоякодышащие рыбы.
В 1870 г. в двух небольших реках Австралии были выявлены живые экземпляры двоякодышащих рыб, строение которых сильно напоминало их ископаемых предков. Впоследствии в Африке и Южной Америке также были найдены живые двоякодышащие рыбы.
Кроме них, в пересыхающих водоемах девонского периода водились кистеперые рыбы. С помощью плавников, напоминавших кисть, кистеперые рыбы способны были ползать. Плавательный пузырь у них был обогащен кровеносными сосудами и играл роль легких. Таким образом, кистеперые рыбы могли дышать воздухом и переползать из лагуны в лагуну в поисках пищи и воды. Скелет кистеперых почти полностью окостенел. Череп состоял из костей, присутствующих в черепах высших позвоночных животных. Следовательно, кистеперые рыбы являлись предками всех наземных позвоночных, в том числе и амфибий, появившихся в верхнем девоне. Это были уже настоящие наземные животные. Они обитали на суше, хотя и имели еще много общего с рыбами — форму черепа, чешую, жаберные крышки.
В 1938 г. в водах Индийского океана у юго-восточных берегов Африки были найдены живые ископаемые — кистеперые рыбы. Их называют целакантусами, или латимериями. Живут целакантусы на значительных глубинах. Они — хищники. Особый интерес вызывают найденные в Пенсильвании окаменевшие следы лап. На трех пальцах из пяти имелись когти. Хорошо виден след хвоста, который тянулся за туловищем животного. Вероятно, этот след принадлежит кистеперой рыбе, передвигавшейся в поисках водоемов по девонской суше.
в водах Индийского океана у юго-восточных берегов Африки были найдены живые ископаемые — кистеперые рыбы. Их называют целакантусами, или латимериями. Живут целакантусы на значительных глубинах. Они — хищники. Особый интерес вызывают найденные в Пенсильвании окаменевшие следы лап. На трех пальцах из пяти имелись когти. Хорошо виден след хвоста, который тянулся за туловищем животного. Вероятно, этот след принадлежит кистеперой рыбе, передвигавшейся в поисках водоемов по девонской суше.
Иногда девонский период называют периодом рыб. И действительно, в морях и озерах девонского периода обитало огромное множество различных рыб. Помимо кистеперых и двоякодышащих, до конца палеозоя там жили ганоидные рыбы. Они имели толстую чешую ромбической формы, располагавшуюся косыми рядами. Позднее ганоидных рыб вытеснили более совершенные формы. В морях размножились рыбы с твердым кожным скелетом, с панцирными щитками, кожными зубами. В то отдаленное время, когда достигшие своего первого расцвета, позвоночные дали многочисленные ответвления, существовали формы и с примитивной внутренней организацией. Несколько десятков лет назад особое внимание привлекли небольшие ископаемые рыбы — палеоспондулюсы. Их нашли в сером тонкослоистом песчанике среди множества остатков других рыб и растений. Венчик нитевидных ротовых присосок напоминал присоску бесплавниковых рыб, находившихся на самой низкой ступени развития. Поэтому их отнесли к бесплавниковым. При помощи специального метода исследования удалось установить анатомические особенности палеоспондулюсов. Они являлись паразитами, их рот был приспособлен к сосанию. Широкие носовые полости свидетельствуют о высоком развитии чутья. Четыре пары хрящевых дуг, соединяющихся у глотки, а также пятая пара несколько более крупных по размерам пластинок образовывали жабры. Подобные образования наблюдаются лишь в черепе акулы и у молодых амфибий. Ребра и плавники у палеоспондулюсов отсутствовали. У них имелась довольно широкая хорда. По мнению некоторых исследователей, строение тела палеоспондулюсов сходно со строением тела древнейших предков всех рыб.
Несколько десятков лет назад особое внимание привлекли небольшие ископаемые рыбы — палеоспондулюсы. Их нашли в сером тонкослоистом песчанике среди множества остатков других рыб и растений. Венчик нитевидных ротовых присосок напоминал присоску бесплавниковых рыб, находившихся на самой низкой ступени развития. Поэтому их отнесли к бесплавниковым. При помощи специального метода исследования удалось установить анатомические особенности палеоспондулюсов. Они являлись паразитами, их рот был приспособлен к сосанию. Широкие носовые полости свидетельствуют о высоком развитии чутья. Четыре пары хрящевых дуг, соединяющихся у глотки, а также пятая пара несколько более крупных по размерам пластинок образовывали жабры. Подобные образования наблюдаются лишь в черепе акулы и у молодых амфибий. Ребра и плавники у палеоспондулюсов отсутствовали. У них имелась довольно широкая хорда. По мнению некоторых исследователей, строение тела палеоспондулюсов сходно со строением тела древнейших предков всех рыб.
В девонских отложениях встречаются остатки форм быстроплавающих рыб из числа акулообразных — маленьких гибких рыбок, предков акул более позднего времени. В отложениях больших внутренних морей девонского периода находят представителей довольно крупных панцирных рыб, по форме сильно напоминающих трилобитов. Передняя часть их тела была покрыта прочным панцирем, задняя — почти не защищена. Головной щит широкий, полукруглой формы, украшен длинными шипами. Глаза сближены между собой. Рот беззубый или с режущим челюстным краем. Эти рыбы обитали в воде, иногда ходили по дну с помощью острых плавниковых шипов. Отдельные панцирные рыбы были очень крупными. Так, голова динихтиса («страшной рыбы») достигала в длину одного метра.
В отложениях больших внутренних морей девонского периода находят представителей довольно крупных панцирных рыб, по форме сильно напоминающих трилобитов. Передняя часть их тела была покрыта прочным панцирем, задняя — почти не защищена. Головной щит широкий, полукруглой формы, украшен длинными шипами. Глаза сближены между собой. Рот беззубый или с режущим челюстным краем. Эти рыбы обитали в воде, иногда ходили по дну с помощью острых плавниковых шипов. Отдельные панцирные рыбы были очень крупными. Так, голова динихтиса («страшной рыбы») достигала в длину одного метра.
Динихтис.
Панцирь некоторых рыб покрывал не только переднюю часть тела, но и плавники. Он спасал рыб от нападения хищников, но в то же время стеснял их движения. В морях, лагунах и болотах панцирных рыб было так много, что они погибали от недостатка пищи и кислорода. В дальнейшем эти рыбы полностью вымерли. В морях стали господствовать беспанцирные рыбы с прочными плавниками и гибким хвостом. На смену плоскотелым малоподвижным придонным рыбам, не имевшим настоящих челюстей, пришли высокотелые, подвижные, гибкие рыбы с мощными режущими или перетирающими челюстями, в частности кистеперые, лучеперые и костистые рыбы. Их легкие плавники состояли из гибких роговых лучей, имевших опору в костях. С помощью мощного хвостового плавника рыбы развивали значительную скорость. Чешуя рыб была очень тонкой и легкой. У костно-хрящевых, в незначительном количестве доживших до нашего времени, скелет слабо окостеневший, их тело покрывает ганоидная чешуя, хвост имеет длинную чешуйную ось.
Их легкие плавники состояли из гибких роговых лучей, имевших опору в костях. С помощью мощного хвостового плавника рыбы развивали значительную скорость. Чешуя рыб была очень тонкой и легкой. У костно-хрящевых, в незначительном количестве доживших до нашего времени, скелет слабо окостеневший, их тело покрывает ганоидная чешуя, хвост имеет длинную чешуйную ось.
В районах действующих вулканов иногда образовывались озера, в иле которых жили червеобразные членистые животные. На берегах этих озер произрастали похожие на камыш растения с длинными узкими листьями.
Из числа беспозвоночных в морях девонского периода были распространены: 15 родов табулят, 24 рода кораллов, 11 родов трилобитов, 40 родов плеченогих, 56 родов двустворчатых, 28 родов головоногих моллюсков. Кроме того, водились многочисленные мшанки, брюхоногие моллюски, иглокожие. Продолжал существовать род граптолитов. Встречались и пресноводные моллюски.
Среди растительных остатков были найдены раковины наземного брюхоногого моллюска строфитес.
Важное место среди беспозвоночных девонских морей и океанов занимали исполинские ракоскорпионы. Часть их приспособилась к условиям жизни в солоноватых и опресненных водоемах. Главной добычей ракоскорпионов являлись трилобиты и рыбы. Происхождение этих загадочных животных пока неизвестно. Однако форма и строение некоторых из них напоминает трилобита с длинной заостренной хвостовой иглой. Это дает основание полагать, что ракоскорпионы произошли от пресноводных предков морских трилобитов.
В наше время ближайшими родичами ракоскорпионов являются мечехвосты, обитающие в мелководных зонах Атлантического, Индийского и Тихого океанов. Питаются мечехвосты главным образом моллюсками. Их личинки напоминают некоторых трилобитов. Развитие современных мечехвостов свидетельствует о том, что они являются промежуточными формами между трилобитами и паукообразными.
У девонских насекомых были перепончатые крылья. Большую часть своей жизни они проводили в воде.
В девонских морях впервые появились хищные головоногие моллюски — аммоноидеи. В их спирально завернутых раковинах имелись перегородки.
В их спирально завернутых раковинах имелись перегородки.
Среди иглокожих наиболее распространенными в данный период были морские лилии, морские звезды и морские ежи.
В девонский период началось вымирание многих силурийских форм: трилобитов, ракоскорпионов, древних иглокожих.
Основные изменения фауны и флоры происходили на суше. В конце девона на Земле росли леса из папоротников, хвощей, плаунов. В этих лесах уже жили насекомые и древние пауки. Появились первые земноводные животные — стегоцефалы.
С девонскими отложениями связано большое количество полезных ископаемых: нефть, каменная соль, горючие сланцы, бокситы, железная руда, медь, золото, марганцевые руды, фосфориты, гипсы, известняки.
Девонский период длился 55 млн. лет.
Каменноугольный период
В отложениях этого периода находят огромные залежи каменного угля. Отсюда и произошло название периода. Есть и другое его наименование — карбон.
Каменноугольный период подразделяется на три отдела: нижний, средний и верхний. В этот период физико-географические условия Земли претерпели значительные изменения. Неоднократно менялись очертания материков и морей, возникали новые горные хребты, моря, острова. В начале карбона происходит значительное опускание суши. Морем были затоплены обширные площади Атлантии, Азии, Гондваны. Уменьшилась площадь больших островов. Исчезли под водой пустыни северного континента. Климат стал очень теплым и влажным.
В этот период физико-географические условия Земли претерпели значительные изменения. Неоднократно менялись очертания материков и морей, возникали новые горные хребты, моря, острова. В начале карбона происходит значительное опускание суши. Морем были затоплены обширные площади Атлантии, Азии, Гондваны. Уменьшилась площадь больших островов. Исчезли под водой пустыни северного континента. Климат стал очень теплым и влажным.
В нижнем карбоне начинается интенсивный горообразовательный процесс: формируются Арденны, Гарц, Рудные горы, Судеты, Атласские горы, Австралийские Кордильеры, Западно-Сибирские горы. Море отступает.
В среднем карбоне суша вновь опускается, но гораздо меньше, чем в нижнем. В межгорных котловинах накопляются мощные толщи континентальных отложений. Формируется Восточный Урал, Пеннинские горы.
В верхнем карбоне море вновь отступает. Значительно уменьшаются внутриконтинентальные моря. На территории Гондваны возникают большие ледники, в Африке и Австралии — несколько меньшие.
В конце карбона в Европе и Северной Америке климат претерпевает изменения, становится частично умеренным, а частично жарким и сухим. В это время происходит формирование Центрального Урала.
Морские осадочные отложения каменноугольного периода представлены главным образом глинами, песчаниками, известняками, сланцами и вулканогенными породами. Континентальные — преимущественно углем, глинами, песками и другими породами.
Усиленная вулканическая деятельность в карбоне привела к насыщению атмосферы углекислым газом. Вулканический пепел, являющийся чудесным удобрением, сделал плодородными карбоновые почвы.
Теплый и влажный климат господствовал на материках длительное время. Все это создало чрезвычайно благоприятные условия для развития наземной флоры, в том числе высших растений каменноугольного периода — кустов, деревьев и травянистых растений, жизнь которых была тесно связана с водой. Они росли главным образом среди громадных болот и озер, близ солоноватоводных лагун, на побережье морей, на влажной илистой почве. По образу жизни они походили на современных мангровых, которые растут на низинных берегах тропических морей, в устьях больших рек, в болотистых лагунах, поднимаясь над водой на высоких корнях-ходулях.
По образу жизни они походили на современных мангровых, которые растут на низинных берегах тропических морей, в устьях больших рек, в болотистых лагунах, поднимаясь над водой на высоких корнях-ходулях.
Значительное развитие в каменноугольный период получили плауновидные, членистостебельные и папоротниковые, давшие большое количество древовидных форм.
Древовидные плаунообразные достигали 2 м в диаметре и 40 м в высоту. У них еще не было годичных колец. Пустой ствол с мощной разветвленной кроной надежно удерживался в рыхлой почве большим корневищем, разветлявшимся на четыре главные ветви. Эти ветви в свою очередь дихотомически разделялись на корневые отростки. Их листья, до метра в длину, густыми султанообразными пучками украшали концы ветвей. На концах листьев располагались почки, в которых развивались споры. Стволы плаунообразных были покрыты чешуей — рубцами. К ним прикреплялись листья. В этот период были распространены гигантские плаунообразные — лепидодендроны с ромбическими рубцами на стволах и сигиллярии с шестигранными рубцами. В отличие от большинства плаунообразных у сигиллярий был почти неразветвленный ствол, на котором вырастали спорангии. Среди плаунообразных были и травянистые растения, полностью вымершие в пермский период.
В отличие от большинства плаунообразных у сигиллярий был почти неразветвленный ствол, на котором вырастали спорангии. Среди плаунообразных были и травянистые растения, полностью вымершие в пермский период.
Членистостебельные растения разделяются на две группы: клинолистые и каламиты. Клинолистые являлись водными растениями. У них был длинный, членистый, слегка ребристый стебель, к узлам которого кольцами прикреплялись листья. Почковидные образования содержали споры. На воде клинолистые держались с помощью длинных разветвленных стеблей, похожих на современный водяной лютик. Клинолистые появились в среднем девоне и вымерли в пермский период.
Каламиты были древовидными растениями до 30 м высотой. Они образовывали болотные леса. Некоторые виды каламитов проникли далеко на материк. Их древние формы имели дихотомические листья. Впоследствии преобладали формы с простыми листьями и годичными кольцами. Эти растения имели сильно разветвленное корневище. Нередко из ствола вырастали дополнительные корни и ветви, покрытые листьями.
В конце карбона появляются первые представители хвощевых — небольшие травянистые растения. Среди карбоновой флоры видную роль играли папоротники, в частности травянистые, по своему строению напоминавшие псилофитов, и настоящие папоротники — большие древовидные растения, корневищем закреплявшиеся в мягком грунте. У них был шершавый ствол с многочисленными ветвями, на которых росли широкие папоротниковидные листья.
Голосеменные карбоновых лесов относятся к подклассам семенных папоротников и стахиоспермид. Их плоды развивались на листьях, что является признаком примитивной организации. В то же время у линейных или ланцетных листьев голосеменных было довольно сложное жилкование. Наиболее совершенные растения карбона — кордаиты. Их цилиндрические безлистые стволы до 40 м высотой разветвлялись. Ветви имели на концах широкие линейные или ланцетные листья с сетчатым жилкованием, Мужские спорангии (микроспорангии) имели вид почек. Из женских спорангиев развивались ореховидные плоды. Результаты микроскопического исследования плодов показывают, что эти растения, похожие на цикадовых, являлись переходными формами к хвойным растениям.
Результаты микроскопического исследования плодов показывают, что эти растения, похожие на цикадовых, являлись переходными формами к хвойным растениям.
В каменноугольных лесах появляются первые грибы, моховидные растения (наземные и пресноводные), образовывавшие иногда колонии, и лишайники.
В морских и пресноводных бассейнах продолжают существовать водоросли: зеленые, красные и харовые.
При рассмотрении каменноугольной флоры в целом поражает разнообразие форм листьев древовидных растений. Рубцы на стволах растений на протяжении всей жизни держали длинные, ланцетные листья. Концы ветвей были украшены огромными лиственными кронами. Иногда листья росли по всей длине ветвей.
Другой характерный признак каменноугольной флоры — развитие подземной корневой системы. Сильно разветвленные корни разрастались в илистой почве и из них произрастали новые побеги. Порой значительные площади были изрезаны подземными корнями.
В местах быстрого накопления илистых осадков корни удерживали стволы многочисленными побегами. Важнейшая особенность каменноугольной флоры заключается в том, что растения не отличались ритмическим ростом в толщину.
Важнейшая особенность каменноугольной флоры заключается в том, что растения не отличались ритмическим ростом в толщину.
Распространение одних и тех же каменноугольных растений от Северной Америки до Шпицбергена свидетельствует о том, что от тропиков до полюсов господствовал относительно равномерный теплый климат, на смену которому в верхнем карбоне пришел довольно прохладный. В прохладном климате росли голосеменные папоротники и кордаиты.
Рост каменноугольных растений почти не зависел от времен года. Он напоминал рост пресноводных водорослей. Времена года, вероятно, мало отличались друг от друга.
При изучении каменноугольной флоры можно проследить эволюцию растений. Схематически она выглядит следующим образом: бурые водоросли — папоротники — псилофиты — птеридоспермиды (семенные папоротники) — хвойные.
Отмирая, растения каменноугольного периода падали в воду, их заносило илом, и, пролежав миллионы лет, они постепенно превратились в уголь. Каменный уголь образовывался изо всех частей растения: древесины, коры, ветвей, листьев, плодов. В уголь превращались и остатки животных. Об этом свидетельствует тот факт, что остатки пресноводных и наземных животных в карбоновых отложениях встречаются относительно редко.
В уголь превращались и остатки животных. Об этом свидетельствует тот факт, что остатки пресноводных и наземных животных в карбоновых отложениях встречаются относительно редко.
Морской животный мир карбона характеризовался разнообразием видов. Чрезвычайно распространенными были фораминиферы, в частности фузулиниды с веретеновидными раковинами величиной с зерно.
В среднем карбоне появляются швагерины. Их шаровидная раковина была величиной с небольшую горошину. Из раковин фораминифер позднего карбона в отдельных местах образовались залежи известняков.
Среди кораллов еще встречались немногочисленные роды табулят, но начинали преобладать хететиды. Одиночные кораллы нередко имели толстые известковые стенки. Колониальные кораллы образовывали рифы.
В это время интенсивно развиваются иглокожие, в частности морские лилии и морские ежи. Многочисленные колонии мшанок иногда образовывали мощные известняковые отложения.
Чрезвычайно развились плеченогие моллюски, в частности продуктусы, по приспособляемости и географическому распространению намного превосходившие всех встречающихся на Земле плеченогих. Величина их раковин достигала 30 см в поперечнике. Одна створка раковины была выпуклой, а другая — в виде плоской крышечки. Прямой удлиненный замочный край часто имел полые шипы. У некоторых форм продуктусов шипы в четыре раза превышали диаметр раковины. С помощью шипов продуктусы держались на листьях водных растений, переносивших их по течению. Иногда своими шипами они прикреплялись к морским лилиям или водорослям и жили возле них в висячем положении. У рихтофений одна створка раковины преобразовалась в рог длиной до 8 см.
Величина их раковин достигала 30 см в поперечнике. Одна створка раковины была выпуклой, а другая — в виде плоской крышечки. Прямой удлиненный замочный край часто имел полые шипы. У некоторых форм продуктусов шипы в четыре раза превышали диаметр раковины. С помощью шипов продуктусы держались на листьях водных растений, переносивших их по течению. Иногда своими шипами они прикреплялись к морским лилиям или водорослям и жили возле них в висячем положении. У рихтофений одна створка раковины преобразовалась в рог длиной до 8 см.
В каменноугольный период почти полностью вымирают наутилоидеи, за исключением наутилусов. Данный род, распавшийся на 5 групп (которые были представлены 84 видами), дожил до нашего времени. Продолжают существовать ортоцерасы, раковины которых имели резко выраженное внешнее строение. Роговидно согнутые раковины циртоцерасов почти не отличались от раковин их девонских предков. Аммониты были представлены двумя отрядами — гониатитами и агониатитами, как и в девонском периоде; двустворчатые моллюски — одномышечными формами. Среди них — множество пресноводных форм, населявших карбоновые озера и болота.
Среди них — множество пресноводных форм, населявших карбоновые озера и болота.
Появляются первые наземные гастроподы — животные, дышавшие легкими.
Трилобиты достигли значительного расцвета в ордовикский и силурийский периоды. В каменноугольный период сохранились лишь немногочисленные их роды и виды.
К концу каменноугольного периода трилобиты почти полностью вымерли. Этому способствовало то, что головоногие моллюски и рыбы питались трилобитами и потребляли ту же пищу, что и трилобиты. Строение тела трилобитов было несовершенным: панцирь не защищал брюха, конечности были небольшими и слабыми. Трилобиты не имели органов нападения. Некоторое время они могли защищаться от хищников, свертываясь наподобие современных ежей. Но в конце карбона появились рыбы с мощными челюстями, разгрызавшими их панцирь. Поэтому от многочисленного типа в перми сохранился только один род.
В озерах каменноугольного периода появляются ракообразные, скорпионы, насекомые.
Карбоновые насекомые обладали признаками многих родов современных насекомых, поэтому отнести их к какому-либо одному известному нам теперь роду невозможно. Несомненно, предками насекомых каменноугольного периода были ордовикские трилобиты. У девонских и силурийских насекомых было много общего с некоторыми их предками. Они играли уже значительную роль в животном мире.
Несомненно, предками насекомых каменноугольного периода были ордовикские трилобиты. У девонских и силурийских насекомых было много общего с некоторыми их предками. Они играли уже значительную роль в животном мире.
Однако подлинного расцвета насекомые достигли в каменноугольном периоде. Представители самых малых из известных видов насекомых были 3 см в длину; размах крыльев у самых крупных (например, у стенодиктии) достигал 70 см, у древней стрекозы меганевры — одного метра. Тело меганевры имело 21 сегмент. Из них 6 составляли голову, 3 — грудь с четырьмя крыльями, 11 — брюшко, конечный сегмент походил на шиловидное продолжение хвостового щита трилобитов. Многочисленные пары конечностей были расчленены. С их помощью животное и ходило, и плавало. Молодые меганевры обитали в воде, превращаясь во взрослых насекомых в результате линьки. У меганевры были крепкие челюсти и сложные глаза.
В верхнем каменноугольном периоде древние насекомые вымерли, их потомки были более приспособленными к новым условиям жизни. Прямокрылые в ходе эволюции дали термитов и стрекоз, эвриптерусы — муравьев. Большинство древних форм насекомых переходили к наземному образу жизни лишь в зрелом возрасте. Размножались они исключительно в воде. Таким образом, перемена влажного климата на более сухой для многих древних насекомых была катастрофой.
Прямокрылые в ходе эволюции дали термитов и стрекоз, эвриптерусы — муравьев. Большинство древних форм насекомых переходили к наземному образу жизни лишь в зрелом возрасте. Размножались они исключительно в воде. Таким образом, перемена влажного климата на более сухой для многих древних насекомых была катастрофой.
Палеозойская эра
Вряд ли можно мысленно охватить отрезок времени длиной в 370 млн. лет.
Именно столько продолжался следующий этап истории Земли — палеозойская эра. Геологи подразделяют ее на шесть периодов: кембрийский — самый древний из них, — ордовикский, силурийский, девонский, карбоновый и пермский.
Палеозой начался колоссальным разливом морей, последовавшим за появлением обширных кусков суши в конце протерозоя. Многие геологи полагают, что в те времена существовал единый огромный континентальный блок, называемый Пангея (в переводе с греческого — “вся земля”), который был со всех сторон окружен мировым океаном. Со временем этот единый континент распался на части, ставшие ядрами современных континентов. В ходе дальнейшей истории Земли эти ядра могли увеличиваться за счет процессов горообразования или же вновь распадаться на части, которые продолжали удаляться друг от друга, пока не заняли положение современных континентов.
Со временем этот единый континент распался на части, ставшие ядрами современных континентов. В ходе дальнейшей истории Земли эти ядра могли увеличиваться за счет процессов горообразования или же вновь распадаться на части, которые продолжали удаляться друг от друга, пока не заняли положение современных континентов.
Впервые гипотезу о разрыве и взаимном расхождении континентов (“континентальный дрейф”) высказал в 1912 г. немецкий геолог Альфред Вегенер. По его представлениям Пангея первоначально разделилась на два сверхконтинента:
Лауразию в северном полушарии и Гондвану на юге. Впадина между ними была затоплена морем, носящим название Тетис. Позднее, в силурийском периоде вследствие каледонского и герцинского горообразовательных процессов на севере поднялся обширный континент. Его сильно пересеченный рельеф в ходе девонского периода был занесен продуктами выветривания мощных горных массивов; в .сухом и горячем климате их частицы обволакивались окисью железа, что придавало им красноватую окраску. Подобное явление можно наблюдать и в некоторых современных пустынях. Вот почему этот девонский континент часто называется Древним красным континентом. На нем в девоне пышно развивались многочисленные новые группы наземных растений, а в некоторых его частях были обнаружены остатки первых наземных позвоночных — рыбообразных амфибий.
Подобное явление можно наблюдать и в некоторых современных пустынях. Вот почему этот девонский континент часто называется Древним красным континентом. На нем в девоне пышно развивались многочисленные новые группы наземных растений, а в некоторых его частях были обнаружены остатки первых наземных позвоночных — рыбообразных амфибий.
В это время Гондвана, включавшая в себя всю современную Южную Америку, почти всю Африку, Мадагаскар, Индию и Антарктиду, оставалась еще единым сверхконтинентом.
К концу палеозоя море отступило, и герцинское горообразование стало понемногу слабеть, сменившись варисцийской складчатостью Центральной Европы.
В конце палеозоя вымирают многие наиболее примитивные растения и животные.
Растения завоевывают сушу.
В течение палеозоя одни группы растений постепенно сменялись другими.
В начале эры, от кембрия до силура, доминировали морские водоросли, но уже в силуре появляются высшие сосудистые растения, произрастающие на суше. До конца каменноугольного периода преобладали споровые растения, но в пермском периоде, особенно, в его второй половине, значительную часть наземной растительности составляют семянные растения из группы голосеменных (Gymnospermae). До начала палеозоя, за исключением нескольких сомнительных находок спор, признаков развития наземных растений нет. Однако, вполне вероятно, что некоторые растения (лишайники, грибы) начали проникать во внутренние районы суши еще в протерозое, так как нередко отложения этого времени содержат значительные количества необходимых растениям питательных веществ.
До начала палеозоя, за исключением нескольких сомнительных находок спор, признаков развития наземных растений нет. Однако, вполне вероятно, что некоторые растения (лишайники, грибы) начали проникать во внутренние районы суши еще в протерозое, так как нередко отложения этого времени содержат значительные количества необходимых растениям питательных веществ.
Для того, чтобы приспособиться к новым условиям жизни на суше, многим растениям пришлось коренным образом изменить свое анатомическое строение.
Так, например, растениям нужно было приобрести наружный эпидермальный покров для защиты от быстрой потери влаги и высыхания; их нижние части должны были одеревянеть и превратиться в подобие опорного каркаса, чтобы противостоять силе тяжести, столь чувствительной после выхода из воды. Корня ми они уходили в почву, откуда черпали воду и питательные вещества. Поэтому растениям нужно было выработать сеть каналов для доставки этих веществ к верхним частям своего тела. Кроме того, они нуждались в плодородной почве, а условием этого была жизнедеятельность множества почвенных микроорганизмов, бактерий, синезеленых водорослей, грибов, лишайников и почвенных животных. Продукты жизнедеятельности и мертвые тела этих организмов постепенно превращали кристаллические горные породы в плодородную почву, способную прокормить прогрессирующие растения. Попытки освоения суши становились все более удачными. Уже в отложениях силурийских морей Центральной Чехии встречаются хорошо сохранившиеся остатки древнейших сосудистых растений — псилофитов (в переводе с греческого — “лишенных листьев”). Эти первичные высшие растения, стебель которых нес пучок сосудов, проводящих жидкости, имели наиболее сложную и комплексную организацию из всех автотрофныхрастенийтоговремени,исключая,возможно,существовавшие
Продукты жизнедеятельности и мертвые тела этих организмов постепенно превращали кристаллические горные породы в плодородную почву, способную прокормить прогрессирующие растения. Попытки освоения суши становились все более удачными. Уже в отложениях силурийских морей Центральной Чехии встречаются хорошо сохранившиеся остатки древнейших сосудистых растений — псилофитов (в переводе с греческого — “лишенных листьев”). Эти первичные высшие растения, стебель которых нес пучок сосудов, проводящих жидкости, имели наиболее сложную и комплексную организацию из всех автотрофныхрастенийтоговремени,исключая,возможно,существовавшие
уже в то время мхи, наличие которых в силуре, однако, еще не доказано. Псилофитовые флоры, появившиеся к концу силурийского периода, процветали вплоть до конца девона. Таким образом, силурийский период положил конец многовековому господству водорослей в растительном мире планеты.
Хвощи, плауны и папоротники.
В нижних слоях девона, в отложениях Древнего красного континента, в изобилии встречаются остатки новых групп растений с развитой сосудо-проводящей системой, размножающихся спорами, как и псилофиты. Среди них преобладают плауны, хвощи и — с середины девонского периода — папоротники. Множество находок остатков этих растений в девонских породах, позволяет заключить, что после протерозоя растения прочно обосновались на суше.
Среди них преобладают плауны, хвощи и — с середины девонского периода — папоротники. Множество находок остатков этих растений в девонских породах, позволяет заключить, что после протерозоя растения прочно обосновались на суше.
Уже в среднем девоне папоротники начинают вытеснять псилофитовую флору, а в верхнедевонских слоях появляются уже древовидные папоротники. Параллельно идет развитие различных хвощей и плаунов. Иногда эти растения достигали крупных размеров, и в результате накопления их остатков в некоторых местах в конце девона образовались первые значительные залежи торфа, который постепенно превращался в каменный уголь. Таким образом, в девоне Древний красный континент мог предоставить растениям все необходимые условия для миграции из прибрежных вод на сушу, для чего потребовались миллионы лет.
Следующий, каменноугольный период палеозойской эры принес с собой мощные горообразовательные процессы, в результате которых на поверхность вышли части морского дна. В бесчисленных лагунах, дельтах рек, топях в зоне литорали воцарилась буйная тепло- и влаголюбивая флора. В местах ее массового развития скоплялись колоссальные количества торфообразного растительного вещества, и, со временем, под действием химических процессов, они преобразовывались в обширные залежи каменного угля В пластах угля часто встречаются прекрасно сохранившиеся остатки растений, свидетельствующие о том, что в ходе каменноугольного периода на Земле появилось много новых групп флоры. Большое распространение получили в это время птеридоспермиды, или семенные папоротники, которые, в отличие от папоротников обыкновенных, размножаются не спорами, а Семенами. Они представляют собой промежуточный этап эволюции между папоротниками и цикадовыми — растениями, похожими на современные пальмы, — с которыми птеридоспермиды находятся в тесном родстве. Новые группы растений появлялись в течение всего каменноугольного периода, в том числе такие прогрессивные формы, как кордаитовые и хвойные. Вымершие кордаитовые были, как правило, крупными деревьями с листьями длиной до 1 м. Представители этой группы активно участвовали в образовании местонахождений каменного угля.
В местах ее массового развития скоплялись колоссальные количества торфообразного растительного вещества, и, со временем, под действием химических процессов, они преобразовывались в обширные залежи каменного угля В пластах угля часто встречаются прекрасно сохранившиеся остатки растений, свидетельствующие о том, что в ходе каменноугольного периода на Земле появилось много новых групп флоры. Большое распространение получили в это время птеридоспермиды, или семенные папоротники, которые, в отличие от папоротников обыкновенных, размножаются не спорами, а Семенами. Они представляют собой промежуточный этап эволюции между папоротниками и цикадовыми — растениями, похожими на современные пальмы, — с которыми птеридоспермиды находятся в тесном родстве. Новые группы растений появлялись в течение всего каменноугольного периода, в том числе такие прогрессивные формы, как кордаитовые и хвойные. Вымершие кордаитовые были, как правило, крупными деревьями с листьями длиной до 1 м. Представители этой группы активно участвовали в образовании местонахождений каменного угля.
Одними из наиболее распространенных растений карбона были гигантские древовидные плауны и хвощи. Из числа первых наиболее известны лепидодендроны — гиганты высотой в 30 м, и сигиллярии, имевшие немногим более 25 м. Стволы этих плаунов разделялись у вершины на ветви, каждая из которых заканчивалась кроной из узких и длинных листьев. Среди гигантских плауновидных были также каламитовые — высокие древовидные растения, листья которых были разделены на нитевидные сегменты; они произрастали на болотах и в других влажных местах, будучи, как и другие плауны, привязанными к воде.
Но самыми замечательными и причудливыми растениями карбоновых лесов были, вне всякого сомнения, папоротники. Остатки их листьев и стволов можно найти в любой крупной палеонтологической коллекции. Особенно поразительный облик имели древовидные папоротники, достигавшие от 10 до 15м в высоту, их тонкий стебель венчала крона из сложно расчлененных листьев ярко-зеленого цвета.
В начале пермского периода еще доминировали спороносные растения, но уже к концу этого последнего этапа палеозойской эры их сильно потеснили голосеменные. Среди этих последних мы находим типы, достигшие своего расцвета лишь в мезозое. Разница между растительностью начала и конца пермского времени огромна. В середине перми совершается переход от начальных фаз эволюции наземных растений к его среднему этапу — мезофиту, для которого характерно господство голосеменных.
В нижнепермских отложениях постепенно исчезают гигантские плауны, как и большинство спороносных папоротников и некоторых хвощей. Зато появляются новые виды папоротникообразных растений (Callipteris conferma, Taeniepteris и др.), которые быстро расселяются по территории тогдашней Европы. Среди пермских находок особенно часты окремнелые стволы папоротников, известные под названием Psaronius. Все реже попадаются в нижней перми кордаитовые, зато расширяется состав гинктовых (GinKgoales) и цикадовых. В сухом климате того времени прекрасно чувствовали себя хвойные. В ранней перми широко распространены были роды Lebachia и Ernestiodendron, а в поздней — Ullmannia и Voltzia. В Южном полушарии процветала так называемая гондванская, или глоссоптерисовая флора. Характерный представитель этой флоры — Glossopteris — принадлежит уже к семенным папоротникам. Леса каменноугольного, а во многих районах Земли также и раннепермского времени приобрели теперь огромное экономическое значение, поскольку за их счет образовались основные промышленные местонахождения каменного угля.
В ранней перми широко распространены были роды Lebachia и Ernestiodendron, а в поздней — Ullmannia и Voltzia. В Южном полушарии процветала так называемая гондванская, или глоссоптерисовая флора. Характерный представитель этой флоры — Glossopteris — принадлежит уже к семенным папоротникам. Леса каменноугольного, а во многих районах Земли также и раннепермского времени приобрели теперь огромное экономическое значение, поскольку за их счет образовались основные промышленные местонахождения каменного угля.
Животный мир палеозоя.
В протерозое тела животных были построены весьма примитивно и обычно не имели скелета. Однако типичные ископаемые палеозойских отложений уже обладали прочным наружным скелетом, или раковиной, защищавшим уязвимые части тела. Под этим покровом животные меньше опасались естественных врагов, что создало предпосылки для быстрого увеличения размеров тела и усложнения организации животных. Появление скелетных животных произошло в самом начале палеозоя — в раннем кембрии, после чего началось их стремительное развитие. Хорошо сохранившиеся окаменелые остатки скелетных животных встречаются повсеместно во множестве, что резко контрастирует с крайней редкостью протерозойских находок.
Хорошо сохранившиеся окаменелые остатки скелетных животных встречаются повсеместно во множестве, что резко контрастирует с крайней редкостью протерозойских находок.
Некоторые ученые рассматривают эту взрывообразную эволюцию как доказательство того, что концентрация атмосферного кислорода достигла к началу кембрия уровня, необходимого для развития высших организмов. В верхней части земной атмосферы образовался озоновый экран, поглощающий губительное ультрафиолетовое излучение, что стимулировало развитие жизни в океане.
Повышение содержания кислорода в атмосфере неизбежно вело к росту интенсивности жизненных процессов. Доктор Э. О. Кангеров полагает, что раковины и внутренний скелет у животных могли появиться только тогда, когда организмы получили в свое распоряжение источник энергии, перекрывающий минимум, необходимый для поддержания внутреннего метаболизма. Таким источником энергии оказалась повышенная концентрация кислорода в атмосфере. Животные, быстро приспосабливаясь к измененной среде, приобретали различные типы раковин, панцирей и внутреннего скелета. При всем своем разнообразии все эти животные пока еще жили в морях, и лишь позднее в ходе эволюции некоторые из них приобрели способность дышать атмосферным кислородом.
При всем своем разнообразии все эти животные пока еще жили в морях, и лишь позднее в ходе эволюции некоторые из них приобрели способность дышать атмосферным кислородом.
Фауна раннего палеозоя была уже столь разнообразной, что в ней были представлены практически все основные разделы беспозвоночных. Такому высокому уровню дифференциации животных, начиная с кембрийского периода, неизбежно должна была предшествовать длительная эволюция, хотя скудные материалы докембрия и не позволяют нам восстановить в деталях картину такого развития.
Трилобиты и другие членистоногие.
Самыми типичными представителями палеозойской фауны являются, вне всякого сомнения, относящиеся к членистоногим животные, известные под названием трилобиты, что в переводе значит “трехдольные”. Их сегментированное тело было покрыто прочным панцирем, подразделенным на три отдела: голову, туловище и хвост. Известно, что 60 % всех видов животного царства раннего палеозоя принадлежало именно к этой группе. До настоящего времени только в одном-единственном случае удалось найти докембрийские остатки членистоногих — в 1964 г. в Австралии. Но уже с самого начала кембрия трилобиты начинают свое триумфальное развитие, разделяясь на сотни родов и видов, многие из которых исчезли с лица планеты столь же быстро, как и появились. Трилобиты во множестве обитали в ордовикских морях, продолжая, хотя и не так интенсивно, свою эволюцию, о чем можно судить по отложениям этого времени, богатым новыми родами трилобитообразных. Трилобиты пошли на убыль в силурийском периоде и стали еще более редкими в девоне. В карбоне и перми существовало одно-единственное семейство трилобитов (Proetidae), последние представители которого вымерли к концу перми. Трилобиты имели повсеместное распространение в палеозое, поэтому они играют важную роль при определении возраста и сравнении отложений разных континентов.
в Австралии. Но уже с самого начала кембрия трилобиты начинают свое триумфальное развитие, разделяясь на сотни родов и видов, многие из которых исчезли с лица планеты столь же быстро, как и появились. Трилобиты во множестве обитали в ордовикских морях, продолжая, хотя и не так интенсивно, свою эволюцию, о чем можно судить по отложениям этого времени, богатым новыми родами трилобитообразных. Трилобиты пошли на убыль в силурийском периоде и стали еще более редкими в девоне. В карбоне и перми существовало одно-единственное семейство трилобитов (Proetidae), последние представители которого вымерли к концу перми. Трилобиты имели повсеместное распространение в палеозое, поэтому они играют важную роль при определении возраста и сравнении отложений разных континентов.
Гигантом среди палеозойских беспозвоночных был, несомненно, морской ракоскорпион Eurypterus, относящийся к группе Merostomata, до известной степени промежуточной между трилобитами и скорпионами и появившейся еще в кембрии.
Расцвета меростомовые достигли в среднем палеозое, когда произошло вселение части их из морей в пресные воды. Размеры палеозойских меростомовых в силуре и девоне достигали 3 м. До наших дней сохранились лишь представители одного семейства мечехвостов (Limulidae).
Размеры палеозойских меростомовых в силуре и девоне достигали 3 м. До наших дней сохранились лишь представители одного семейства мечехвостов (Limulidae).
В девоне и, особенно, карбоне начинают развиваться наземные членистоногие и среди них многочисленные типы наземных форм: многоножки (с силура), скорпионы (с силура), пауки и другие. Из карбона известны примитивные стрекозы рода Meganeura, размах крыльев которых достигал 57 см, и многоножки Arthropleura (класс Diplopoda), выраставшие до полутора метров в длину.
Археоциаты
В кембрийских морях в изобилии водились животные с кубковидным скелетом, известные под именем археоциаты (Archaeocyathi), которые в начале палеозойской эры играли ту же роль, что и кораллы в позднейшие времена. Они вели прикрепленный образ жизни в теплых и мелких водах. Со временем их известковые скелеты образовывали в определенных местах значительные скопления извести, говорящие о том, что ранее эти участки были дном мелкого и теплого моря.
Брахиоподы
В начале палеозоя появились также брахиоподы (Brachiopoda) — морские животные с двустворчатой раковиной, похожие на моллюсков. Они составляли 30 % видов известной кембрийской фауны. Прочные раковины большинства кембрийских видов брахиопод состояли из хитинового вещества, пропитанного фосфатом кальция, тогда как раковины более поздних форм состояли преимущественно из карбоната кальция. Скапливаясь в благоприятных местах в неисчислимых количествах, брахиоподы давали значительную часть материала при образовании подводных рифов и барьеров. В морской фауне палеозоя брахиоподы своей численностью превышают все другие типы животных. Они присутствуют практически во всех морских отложениях этого времени.
Они составляли 30 % видов известной кембрийской фауны. Прочные раковины большинства кембрийских видов брахиопод состояли из хитинового вещества, пропитанного фосфатом кальция, тогда как раковины более поздних форм состояли преимущественно из карбоната кальция. Скапливаясь в благоприятных местах в неисчислимых количествах, брахиоподы давали значительную часть материала при образовании подводных рифов и барьеров. В морской фауне палеозоя брахиоподы своей численностью превышают все другие типы животных. Они присутствуют практически во всех морских отложениях этого времени.
Иглокожие
Важным элементом палеозойской фауны были иглокожие (Echinodermata), к которым относятся и общественные морские звезды и морские ежи. Их кембрийские представители принадлежат в большинстве своем к даво вымершим группам, отличающимся, в частности, простым асимметричным строением.
Только позже в палеозое приобрели иглокожие радиальную симметрию. В слоях нижнего кембрия встречаются остатки представителей класса Eocrinoidea, настоящие морские лилии (Crinoidea) появляются только в начале ордовика. Некоторые примитивные формы иглокожих, такие, как цистоидеи (Cystoidea), имели шаровидное тело, по которому были бессистемно разбросаны кроющие пластинки (“таблички”), У прикрепленных форм развивался стебель, служащий для прикрепления к субстрату. Впоследствии стебельчатость стала общей для большинства форм. Морские лилии, вершина расцвета которых приходится на карбон, пережили все геологические эпохи с кембрия; известны и морские ежи, тогда как морские звезды и офидры известны с ордовика.
Некоторые примитивные формы иглокожих, такие, как цистоидеи (Cystoidea), имели шаровидное тело, по которому были бессистемно разбросаны кроющие пластинки (“таблички”), У прикрепленных форм развивался стебель, служащий для прикрепления к субстрату. Впоследствии стебельчатость стала общей для большинства форм. Морские лилии, вершина расцвета которых приходится на карбон, пережили все геологические эпохи с кембрия; известны и морские ежи, тогда как морские звезды и офидры известны с ордовика.
Моллюски
В начале палеозойской эры моллюсков (Mollusco) было крайне мало. (Кстати говоря, некоторые специалисты относят к мягкотелым, или моллюскам, вышеупомянутых брахиоподов.) Те, что были, относились к брюхоногим, хотя уже с кембрия известны их основные классы — и брюхоногие (Gastropoda), панцирные, или хитоны (Amphineura), раковина которых состояла из нескольких щитков, и двустворчатые (Bivalvia), и головоногие (Cephalopoda). К середине палеозоя моллюски заметно размножились. Остатки брюхоногих содержатся почти во всех изученных сериях. Быстрымитемпамишло иразвитиеголовоногих.Пресноводные двустворчатые появляются в большом числе в девоне, известны они также из карбона и перми. Большое распространение получили в палеозое и брюхоногие, первые пресноводные формы которых появились в конце каменноугольного периода.
Быстрымитемпамишло иразвитиеголовоногих.Пресноводные двустворчатые появляются в большом числе в девоне, известны они также из карбона и перми. Большое распространение получили в палеозое и брюхоногие, первые пресноводные формы которых появились в конце каменноугольного периода.
Из головоногих были наиболее широко представлены наутилоидеи (Nautiloidea), достигшие расцвета в силуре; один род — Nautilus, или “кораблик” — дожил до наших дней. К концу палеозоя наутилоидеи были вытеснены аммонитами (Ammonoidea) — головоногими со спирально закрученной раковиной, нередко с богато скульптированной поверхностью. Своим видом раковина сильно напоминает бараний рог. Свое имя аммониты ведут от “рога Аммона”; Аммон, божество древних египтян, изображался с головой барана. Среди аммонитов особое место принадлежит гониатитам (Goniatites), появившимся в девоне и занимавшим господствующее положение в морях карбона. Их остатки являются хорошим указателем геологического возраста морских пород.
Граптолиты и кишечнополостные Особого внимания заслуживают и две другие группы — граптолиты (Graptolithina) и кишечнополостные (Coelenterata). Окаменелые граптолиты часто выглядят как грифельный след на палеозойских породах; это были морские колониальные организмы, имевшие широкое распространение, что позволяет использовать их для дробного расчленения морских отложений. Граптолиты имеют отдаленное родство с хордовыми предками позвоночных.
Окаменелые граптолиты часто выглядят как грифельный след на палеозойских породах; это были морские колониальные организмы, имевшие широкое распространение, что позволяет использовать их для дробного расчленения морских отложений. Граптолиты имеют отдаленное родство с хордовыми предками позвоночных.
Из числа кишечнополостных наиболее замечательны кораллы (Anthogoo).
В раннем палеозое широко были распространены две группы кораллов: четырехлучевые, или ругозы (Rugosa) и табуляты (Tabulata). В теле первых выделялись четыре главные вертикальные перегородки, вторые представлены группами поперечных образований. Силурийские кораллы нередко образовывали массивные напластования известняков органического происхождения. Другой группой кишечнополостных, имевших чрезвычайно широкое распространение в середине палеозоя, были строматопоры (Stromatoporoidea). Эти организмы, происхождение которых по сей день является предметом дискуссий, наращивали прочный известковый скелет, часто уплощенной формы. Некоторые из них достигали 2 и более метров в поперечнике. Строматопоры активно участвовали в образовании силурийских и девонских известковых рифов. Обычно их причисляют к гидроидным полипам (Hydrozoa). Многочисленными были в палеозое и конулярии (Conulata), которых также обычно относят к кишечнополостным.
Некоторые из них достигали 2 и более метров в поперечнике. Строматопоры активно участвовали в образовании силурийских и девонских известковых рифов. Обычно их причисляют к гидроидным полипам (Hydrozoa). Многочисленными были в палеозое и конулярии (Conulata), которых также обычно относят к кишечнополостным.
Они появляются в ордовике, достигают высшей точки развития в девоне и вымирают в самом начале мезозойской эры. Конулярии представлены конусовидными “раковинами” из органического вещества, не исключено их родство с медузами.
Фораминиферы
Конец палеозоя был отмечен массовым развитием фораминифер (Foraminifera). Эти одноклеточные организмы, чье имя в буквальном переводе означает “несущие отверстия”, были заключены в раковины, снабженные особыми отверстиями. В карбоне и особенно в перми члены этой группы порой достигали внушительных размеров. Размножаясь в большом числе, они поставляли значительную часть материала, из которого строились рифы морского дна.
Беспозвоночные в конце палеозоя
К концу палеозоя (пермь) фораминиферы продолжают эволюционировать, но целый ряд других групп животных приходит в упадок: сокращается число трилобитов, вымирают ругозы, падает значение брахиоподов. Относительно распространенными остались двустворчатые, среди которых появляются формы близкие мезозойским типам. Головоногие, среди которых появились в это время первые настоящие аммониты, претерпевают к концу перми определенный кризис.
Относительно распространенными остались двустворчатые, среди которых появляются формы близкие мезозойским типам. Головоногие, среди которых появились в это время первые настоящие аммониты, претерпевают к концу перми определенный кризис.
Первые позвоночные животные
Первые позвоночные появляются в ордовикских отложениях. Остатки костного панциря примитивных позвоночных из группы бесчелюстных рыбообразных — остракодерм (Ostracodermi) были обнаружены в нижнеордовикских породах Эстонии и в среднеордовикских отложениях США. В современной фауне бесчелюстные рыбообразные представлены немногочисленными формами, полностью лишенными костного скелета и чешуйчатого покрова, к числу их относятся миноги. Очень большой вклад в наши знания об ископаемых бесчелюстных внес шведский палеонтолог профессор Э. Стеньше.
Более высоко организованные позвоночные, относимые к настоящим рыбам, обладающим хорошо развитыми челюстями и парными плавниками) появляются в силуре. Наиболее характерную группу древнейших рыб образуют панцирные пластинокожие (Placodermi), процветавшие в девоне. К их числу относятся своеобразные антиархи. В начале девона плакодермы оставались относительно некрупными формами, близкими по размерам к бесчелюстным. Но быстро увеличиваясь в размерах, они уже вскоре стали настоящими гигантами, как например,
К их числу относятся своеобразные антиархи. В начале девона плакодермы оставались относительно некрупными формами, близкими по размерам к бесчелюстным. Но быстро увеличиваясь в размерах, они уже вскоре стали настоящими гигантами, как например,
Dinichthys, длина которого достигала 11 м. Это хищное чудовище, должно быть, наводило ужас на обитателей девонских морей. Наряду с пластинокожими в середине палеозоя появляются предки настоящих акул; в верхнем палеозое некоторых из них можно встретить даже в отложениях пресноводных бассейнов.
Параллельно идет эволюция различных групп высших, или костных рыб (Osteichthyes), появившихся к началу девона, к концу девона давших начало первым земноводным — ихтиостегидам (Ichthyostegalia). Что же касается других групп рыб и рыбообразных, то почти все группы начала девона начинают исчезать к концу этого периода. Исключение составляли акантоды (Acanthodii), своеобразные рыбы с зубчатыми шипами в основании парных плавников.
Быстро распространяясь, костные рыбы уже к концу девона оказались доминирующей группой позвоночных пресноводных бассейнов. Практически с самого начала своей эволюции они распались на три основные ветви. Виды первой из них процветают и ныне, охватывая 90 % всех существующих рыб. Плавники этих рыб поддерживались длинными костными лучами, откуда и пошло название всего подкласса — лучеперые (Actinopterygii). Вторая группа костных рыб в настоящее время представлена лишь тремя родами двоякодышащих рыб (Dipnoi), распространенных на южных материках. Свое название они получили за то, что помимо жабер, эти рыбы обладают и легкими, использующимися для дыхания воздухом. Третью группу костных рыб образуют кистеперые (Crossopterygii’), получившие свое название за кистеобразное ветвление внутреннего скелета парных плавников. Кистеперые рыбы имеют огромное эволюционное значение: именно они дали начало всем наземным позвоночным, включая сюда и человека. Вместе с двоякодышащими кистеперых иногда объединяют в одну группу.
Практически с самого начала своей эволюции они распались на три основные ветви. Виды первой из них процветают и ныне, охватывая 90 % всех существующих рыб. Плавники этих рыб поддерживались длинными костными лучами, откуда и пошло название всего подкласса — лучеперые (Actinopterygii). Вторая группа костных рыб в настоящее время представлена лишь тремя родами двоякодышащих рыб (Dipnoi), распространенных на южных материках. Свое название они получили за то, что помимо жабер, эти рыбы обладают и легкими, использующимися для дыхания воздухом. Третью группу костных рыб образуют кистеперые (Crossopterygii’), получившие свое название за кистеобразное ветвление внутреннего скелета парных плавников. Кистеперые рыбы имеют огромное эволюционное значение: именно они дали начало всем наземным позвоночным, включая сюда и человека. Вместе с двоякодышащими кистеперых иногда объединяют в одну группу.
Кистеперые, замечательные своими плавниками с широким мясистым основанием, обитали не только в морях, но и в пресноводных бассейнах, и достигли апогея развития в конце девона. В последующие геологические эпохи кистеперых становилось все меньше, и в наше время они представлены единственным реликтовым родом — латимерией (Latimeria), которая встречается в глубоких водах вблизи Мадагаскара. Ближайшая к латимерии форма вымерла еще в меловом периоде.
В последующие геологические эпохи кистеперых становилось все меньше, и в наше время они представлены единственным реликтовым родом — латимерией (Latimeria), которая встречается в глубоких водах вблизи Мадагаскара. Ближайшая к латимерии форма вымерла еще в меловом периоде.
Первые наземные позвоночные
Древнейшие наземные позвоночные, остатки которых обнаружила в 30-х годах датская экспедиция на острове Имер, к востоку от Гренландии, возникли в конце девона от кистеперых рыб.
Выход животных из воды на сушу стал переломным моментом в эволюции жизни на Земле. Естественно, потребовалось продолжительное время для коренной перестройки функций организма, связанной с наземным образом жизни.
Предковые для наземных позвоночных кистеперые рыбы сначала оставляли воду лишь на непродолжительные сроки. Передвигались на суше они плохо, используя для этой цели змеевидные изгибы тела. Такой способ передвижения практически представляет собой своего рода плавание на суше. Лишь постепенно в передвижении на суше все большую роль начинали играть парные конечности, превращавшиеся по мере этого из рыбьих плавников в конечности наземных животных. Лишь когда предки наземных позвоночных приспособились к поискам пищи на суше, можно стало говорить о появлении настоящих наземных позвоночных. Первые амфибии — ихтиостеги — имели в своем строении еще много рыбьих признаков, о чем говорит и их название.
Лишь когда предки наземных позвоночных приспособились к поискам пищи на суше, можно стало говорить о появлении настоящих наземных позвоночных. Первые амфибии — ихтиостеги — имели в своем строении еще много рыбьих признаков, о чем говорит и их название.
В каменноугольном и, отчасти, пермском периодах продолжалась прогрессивная эволюция амфибий. Многообразие их форм возрастало, но все древние амфибии обитали либо в заболоченных местах, либо даже в пресных водоемах.
Основную группу древних амфибий составляют так называемые лабиринтодонты, получившие свое название за строение зубов, на поперечных срезах которых дентин и эмаль образуют глубоко вдающиеся внутрь разветвленные складки, разделенные узкими промежутками. Аналогичное строение зубов имели икистеперые рыбы. Длина тела у лабиринтодонтов колебалась от нескольких сантиметров до четырех — пяти метров; часто по размерам и форме тела они напоминали некрупных аллигаторов. В своем развитии амфибии зависят еще от водной среды, так как они размножаются, откладывая икру в воду. В воде же живут и растут их личинки.
В воде же живут и растут их личинки.
Первые пресмыкающиеся
В конце карбона происходят общие изменения климата. Если раньше климат в Северном полушарии был теплым и влажным, то теперь он становится все более засушливым и континентальным; засухи становятся все более продолжительными. Это стимулировало эволюцию новой группы животных — пресмыкающихся, или рептилий (Reptilia), ведущих свое происхождение от лабиринтодонтов. В отличие от амфибий, пресмыкающиеся потеряли связь с водной средой; они приобрели способность к внутреннему оплодотворению, их яйца содержат большое количество питательного вещества — желтка, они покрыты твердой пористой скорлупой и откладываются на суше. Личинок рептили и не имеют, и из яиц у них вылупляется вполне сформированное молодое животное. Хотя появление пресмыкающихся отмечено уже в середине карбона, их бурное развитие начинается только в перми. С этого времени прослеживается несколько основных линий эволюции, теряющихся в мезозойской эре.
Рептилии конца каменноугольного периода отличались еще крайней примитивностью. В это время наибольшее распространение среди них получили пеликозавры (Pelycosauria), выраставшие до значительных размеров. Они относятся к звероподобным пресмыкающимися, от которых в мезозое произошли млекопитающие.
В это время наибольшее распространение среди них получили пеликозавры (Pelycosauria), выраставшие до значительных размеров. Они относятся к звероподобным пресмыкающимися, от которых в мезозое произошли млекопитающие.
Пермь — последний период палеозойской эры, продолжавшийся приблизительно 345 млн. лет. За это время жизнь на Земле изменилась до неузнаваемости: животные вышли из воды и постепенно продвинулись вглубь континентов, так что к концу эры встречались формы, приспособленные к существованию даже в самых засушливых местах. Некоторые из этих форм начали линию развития, ведущую непосредственно к млекопитающим и в конечном счете к человеку.
Мультимедийная разработка урока в 9-м классе «Жизнь в палеозойскую эру»
Цели урока:
- углубить и расширить понятия о развитии жизни на Земле;
- изучить особенности изменения климатических условий, географии, а также развития растительного и животного мира в палеозойскую эру;
- продолжить формирование
понятия «главные направления эволюции» на примере развития растительного и
животного мира палеозоя.

Оборудование: процессор, видеопроектор, мультимедийная презентация (см. Приложение).
Этапы урока:
- Проверка знаний об особенностях развития жизни на Земле в архейскую и протерозойскую эру проводится в письменной форме (Слайд 2).
- Изучение нового материала.
- Вводное слово учителя о целях урока.
- Задание учащимся о заполнении в тетради таблицы в ходе изучения нового материала.
- Работа с мультимедийной презентацией.
- Обобщение знаний по данной теме.
- Домашнее задание.
Сегодня на уроке нам предстоит продолжить изучение вопроса о развитии жизни на Земле, и мы увидим, какие географические изменения лика Земли происходили в различные периоды палеозойской эры, как менялся климат, а также растительный и животный мир.
В ходе изучения нового материала вам предстоит заполнить таблицу:
Развитие жизни в палеозойскую эру
| Период | Растительный мир | Животный мир |
(Слайд 3)
Давайте познакомимся с содержанием нашего урока.
(Слайд 4)
Палеозойская эра – крупный период в истории развития земли, следовавшей за архейской или азойской эрой, и предшествовавший мезозойской эре. Отложения палеозойской эры составляют палеозойскую группу слоёв, совокупность которых достигает в некоторых местностях 30000м. мощности, почти в 10 раз превышает мощность мезозойских отложений, что указывает, конечно на весьма значительную её продолжительность. Её началом считают появление организмов, снабженных скелетами, раковинами, панцирями: причем защитные приспособления возникают сразу во многих группах организмов.
Палеозой — эра древней жизни, началась 570 миллионов лет назад и продолжалась около 320 миллионов лет.
(Слайд 5)
Палеозой включает 6 геологических систем: кембрийскую, ордовикскую,
силурийскую, девонскую, каменноугольную и пермскую. Палеозойская эра
характеризуется 2 главными эпохами складчатости: калидонской (Великобритания,
Скандинавский полуостров, Шпицберген, Казахстан и др. ) и герцинский (Центральная
Европа, Урал, Аппалачи).
) и герцинский (Центральная
Европа, Урал, Аппалачи).
Кембрий (Слайд 6)
Кембрийский период начался 570±20 млн. лет назад, длительность 80 млн. В кембрийский период впервые в геологической истории появились скелетные организмы.
География и климат. (Слайд 7)
Начало кембрийскому периоду положил поразительной силы эволюционный взрыв, в ходе которого на Земле впервые появились представители большинства основных групп животных, известных современной науке. Поперёк экватора распростёрся сверхматерик Гондвана. Наряду с ним было ещё 4 материка меньших размеров, соответствовавших нынешним Европе, Сибири, Китаю и Северной Америке. В мелких тропических водах формируются обширные строматолитовые рифы. Ближе к концу периода началось оледенение, приведшее к понижению уровня моря.
Животный мир. (Слайд 8)
В ходе грандиозного эволюционного взрыва возникло большинство современных
типов животных, включая микроскопических фораминифер, губок, морских звезд,
морских ежей, морских лилий и различных червей. В тропиках археоциаты. возводили
громадные рифовые сооружения. Появились первые твердопокровные животные; в морях
господствовали трилобиты и брахиоподы. Возникли первые хордовые. Позднее
появились головоногие моллюски и примитивные рыбы.
В тропиках археоциаты. возводили
громадные рифовые сооружения. Появились первые твердопокровные животные; в морях
господствовали трилобиты и брахиоподы. Возникли первые хордовые. Позднее
появились головоногие моллюски и примитивные рыбы.
Представители. (Слайд 9)
Ареоциаты, Биллингселла, Галлюциногения, Медузы, Аномалокарис, Виваксия, Опабиния, Бургессия, Трилобиты.
Растительный мир. (Слайд 10)
Растительный мир представлен синезелёными, и красными водораслями, примитивными высшими растениями.
Ордовик (Слайд 11)
Ордовикский период – второй период палеозойской эры. Начало ордовикского периода 490±15 млн. лет назад, продолжительность 65 млн. лет.
В раннем и начале среднего ордовика происходило максимальное расширение морских пространств.
География и климат. (Слайд 12)
В начале ордовикского периода большая часть южного полушария была по-прежнему
занята великим материком Гондваной, в то время как прочие крупные массивы суши
сосредоточились ближе к экватору.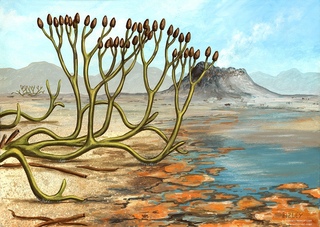 Европа и Северная Америка (Лаврентия)
постепенно отодвигались друг от друга, а океан Япентус расширялся. Сперва этот
океан достиг ширины приметро 2000км. затем вновь начал сужаться по мере того,
как массивы суши, образующие Европу, Северную Америку и Гренландию, стали
постепенно сближаться, пока, наконец, не слились в единое целое. На протяжении
периода массивы суши смещались все дальше и дальше к югу. Старые ледниковые
покровы кембрия растаяли, и уровень моря повысился. Большая часть суши была
сосредоточена в теплых широтах. В конце периода началось новое оледенение.
Европа и Северная Америка (Лаврентия)
постепенно отодвигались друг от друга, а океан Япентус расширялся. Сперва этот
океан достиг ширины приметро 2000км. затем вновь начал сужаться по мере того,
как массивы суши, образующие Европу, Северную Америку и Гренландию, стали
постепенно сближаться, пока, наконец, не слились в единое целое. На протяжении
периода массивы суши смещались все дальше и дальше к югу. Старые ледниковые
покровы кембрия растаяли, и уровень моря повысился. Большая часть суши была
сосредоточена в теплых широтах. В конце периода началось новое оледенение.
Животный мир. (Слайд 13)
Резкое увеличение численности животных-фильтраторов, в том числе мшанок
(морских циновок), морских лилий, плеченогих, двустворчатых моллюсков и
граптолитов, чей расцвет пришелся как раз на ордовик. Археоциаты уже вымерли, но
эстафету рифостроительства подхватили у них строматопороидеи и первые кораллы.
Увеличилось число наутилоидей и бесчелюстных панцирных рыб. В водоёмах
ордивикского периода существовали представители почти всех типов беспозвоночных
(радиолярии, форамониферы, трилобиты и др.), появились первые беспозвоночные –
бесчелюстные рыбообразные
В водоёмах
ордивикского периода существовали представители почти всех типов беспозвоночных
(радиолярии, форамониферы, трилобиты и др.), появились первые беспозвоночные –
бесчелюстные рыбообразные
Представители. (Слайд 14)
Археокринус, Гомилозои, Брюхоногие моллюски, Астраспис, Гониоцерас, Морские бутоны, Платилихас, Платистофия, Эндоцераз, Эхиносферит.
Растительный мир. (Слайд 15)
Существовали различные виды водорослей. В позднем ордовике появились первые настоящие наземные растения.
Силур (Слайд 16)
Третий период палеозойской эры геологической истории. Начался 435 млн. лет назад, длительность 30 млн. лет. Подразделяется на 2 отдела.
Самый крупный массив суши в Силурийской системе — материк Гондвана.
Начало Силурийского периода характеризовалось глобальной морской трансгрессией, конец — завершением каледонской складчатости.
География и климат. (Слайд 17)
(Слайд 17)
Гондвана надвинулась на Южный полюс. Океан Япетус уменьшался в размерах, а массивы суши, образующие Северную Америку и Гренландию сближались. В конечном итоге они столкнулись, образовав Лавразию. Это был период бурной вулканической активности и интенсивного горообразования. Начался он с эпохи оледенения. Когда льды растаяли, уровень моря повысился и климат стал мягче.
Животный мир. (Слайд 18)
Ругозы ведут очень активное рифостроительство. Численность граптолитов снижается. В морях процветают наутилоидеи, брахиоподы, трилобиты и иглокожие. В несильносолёной воде обитают ракоскорпионы (эвриптериды). Изобилие рыб как в пресной так и в солёной воде. Появились первые челюстные рыбы-акантоды. Скорпионы, многоножки и, возможно, эвриптериды начали выбираться на сушу. Формирование основных классов беспозвоночных организмов, появились первые примитивные позвоночные (бесчелюстные и рыбы).
Представители. (Слайд 19)
(Слайд 19)
Акантоды, Арктинурус, Биркения, Дейфон, Морская лилия, Ортоцератида, Палеофонус, Птераспис, Птериготус, Спирифериды, Стилонурус, Четырёхлучевые кораллы.
Растительный мир. (Слайд 20)
Растения заселяли берега водоёмов. Первая наземная флора – псилофиты.
Девон (Слайд 21)
Четвертый период палеозойской эры геологической истории. Начался 400 млн. лет назад, длился около 55 млн. лет. Подразделяется на 3 отдела и 7 ярусов. Начало периода характеризовалось отступанием моря и накоплением толщ мощных континентальных (красноцветных)отложений. Основные полезные ископаемые — нефть и газ, каменные и калийные соли, медистые песчаники.
География и климат. (Слайд 22)
Девонский период был временем величайших катаклизмов на нашей планете.
Европа, Северная Америка и Гренландия Столкнулись между собой, образовав
огромный сверхматерик Лавразию. С океанского дна были вытолкнуты огромные
массивы осадочныхпород, сформировавшие громадные горные системы на востоке
Северной Америки и на западе Европы. Реки выносили в моря горы осадков.
Образовались обширные болотистые дельты, что создавало идеальные условия для
животных, дерзнувших сделать первые столь важные шаги из воды на сушу. К концу
периода уровень моря понизился. Климат со временем потеплел и стал более резким,
с чередованием ливневых дождей и жестокой засухи. Обширные районы материков
стали безводными.
С океанского дна были вытолкнуты огромные
массивы осадочныхпород, сформировавшие громадные горные системы на востоке
Северной Америки и на западе Европы. Реки выносили в моря горы осадков.
Образовались обширные болотистые дельты, что создавало идеальные условия для
животных, дерзнувших сделать первые столь важные шаги из воды на сушу. К концу
периода уровень моря понизился. Климат со временем потеплел и стал более резким,
с чередованием ливневых дождей и жестокой засухи. Обширные районы материков
стали безводными.
Животный мир. (Слайд 23)
Быстрая эволюция рыб, включая акул и скатов, кистеперых и лучеперых рыб.
Увеличилось число аммонитов. В морях охотились гигантские эвриптериды длиной до
2 м. В позднем девоне многие группы древних рыб, а также кораллов, плеченогих и
аммонитов вымерли. Суша подверглась нашествию множества членистоногих, в том
числе клещей, пауков и примитивных бескрылых насекомых. Появились в позднем
девоне и первые земноводные.
Представители. (Слайд 24)
Аргириаспис, Астеролепис, Диптер, Дунклеостерус, Ихтиостега, Кладоселахия, Коккостерус, Целакант.
Растительный мир. (Слайд 25)
Растения сумели отодвинуться от кромки воды и вскоре обширные районы суши поросли густыми первобытными лесами. Возросло число разнообразных сосудистых растений. Появились споровые ликофиты (плауны) и хвощи, некоторые из них развились в настоящие деревья высотой 38м.
Каменноугольный период (Слайд 26)
Каменноугольный период – пятый период палеозойской эры геологической истории. Начался каменноугольный период 345 млн. лет назад; длительность 65 млн. лет. Подразделяется на 3 или 2 отдела. В каменноугольный период происходили интенсивные тектонические движения – герцинская складчатость. На приморских равнинах формировались залежи торфа и каменного угля.
География и климат. (Слайд 27)
Два огромных сверхматерика: Лавразия на севере и Гондвана на юге — сближались
друг с другом. В раннем карбоне на обширных пространствах раскинулись мелкие
прибрежные моря и болота, и установился почти тропический климат. Громадные леса
с пышной растительностью существенно повысили содержание кислорода в атмосфере.
В дальнейшем похолодало и на Земле произошно по меньшей мере, два крупных
оледенения.
В раннем карбоне на обширных пространствах раскинулись мелкие
прибрежные моря и болота, и установился почти тропический климат. Громадные леса
с пышной растительностью существенно повысили содержание кислорода в атмосфере.
В дальнейшем похолодало и на Земле произошно по меньшей мере, два крупных
оледенения.
Животный мир. (Слайд 28)
В морях появились аммониты, возросла численность брахиоподов. Ругозы, граптолиты, трилобиты, а также некоторые мшанки, морские лилии и моллюски вымерли. Это был век земноводных, а также насекомых — кузнечиков, тараканов, чешуйниц, термитов, жуков и гигантских стрекоз. Появились первые рептилии.
Представители. (Слайд 29)
Бабочки, Вестлотиана, Кузнечит, Многоножка, Пауки, Птероплакс, Стрекоза, Таракан, Урокордил.
Растительный мир. (Слайд 30)
Дельты рек и берега обширных болот поросли густыми лесами из гигантских
плаунов, хвощей, древовидных папоротников и семенных растений высотой до 45 м. Неразложившиеся останки этой растительности со временем превратились в каменный
уголь.
Неразложившиеся останки этой растительности со временем превратились в каменный
уголь.
Пермий (Слайд 31)
Пермский период – последний период палеозойской эры. Начался 280 млн. лет назад, продолжительность 45 млн. лет. Пермский период выделен в 1841 английским геологом Р. Мурчисоном на Урале и Русской равнине (на территории Пермской губернии, отсюда название). Подразделяется на нижний и верхний отделы. Общепринятой схемы расчленения на ярусы не существует. Характеризуется интенсивными тектоническими движениями, связанными с последними фазами герцинской складчатости и обширными регрессиями моря. В отложениях пермской системы – угли, нефть и газ, каменные и калийные соли, медистые песчаники, фосфориты.
География и климат. (Слайд 32)
Весь пермский период сверхматерики Гондвана и Лавразия постепенно сближались
друг к другу. Азия столкнулась с Европой, взметнув ввысь Уральский горный
хребет. Индия «наехала» на Азию — и возникли Гималаи. В Северной Америке выросли
Аппалачи. Сформировался гигантский сверхматерик Пангея. Пермский период начался
с оледенения, вызвавшего понижение уровня моря. По мере движения Гондваны к
северу земля прогревалась, и льды постепенно растаяли. В Лавразии сделалось
очень жаркои сухо, по ней распространились обширные пустыни.
Индия «наехала» на Азию — и возникли Гималаи. В Северной Америке выросли
Аппалачи. Сформировался гигантский сверхматерик Пангея. Пермский период начался
с оледенения, вызвавшего понижение уровня моря. По мере движения Гондваны к
северу земля прогревалась, и льды постепенно растаяли. В Лавразии сделалось
очень жаркои сухо, по ней распространились обширные пустыни.
Животный мир. (Слайд 33)
Бурно эволюционировали двустворчатые моллюски. В морях в изобилии водились
аммониты. На место главных рифостроителей стали выдвигаться современные кораллы.
В пресных водоемах господствовали земноводные. Появились и водные рептилии, в
том числе мезозавры. В ходе великого вымирания конца периода исчезло свыше 50%
животных семейств. На суше рептилии взяли верх над земноводными. Также
господствовали примитивные пресмыкающиеся, насекомые. В морях обитали
фораминиферы, кораллы(ругозы), двустворчатые, брюхоногие и головоногие моллюски,
мшанки, брахиоподы, морские членистоногие, морские лилии, из позвоночных –
хрящевые акулообразные рыбы.
Представители. (Слайд 34)
Двиния, двинозавр, Диметродон, Ивантозавр, Иностранцевия, Каккопс, Лантанозух, Мезенозавр, Скутозавр, Целурозавравус, Эокапторинус, Эриопс, Эстемменозух.
Растительный мир. (Слайд 35)
На южных массивах суши распространились леса крупных семенных папоротников-глоссоптерисов. Появились первые хвойные, быстро заселившие внутриматериковые области и высокогорья.
Заключение. (Слайд 36)
- Палеозойская эра (греч. «палайос»- древний, «зое» — жизнь) – эра древней жизни.
- Её возраст 570 млн лет.
- Подразделяется на 6 периодов (кембрий, ордовик, силур, девон, каменноугольный, пермь).
- Растительный мир развивался от водорослей до первых семенных растений (семенных пааоротников).
- Животный мир развивался от примитивных морских бесчерепных хордовых до наземных пресмыкающихся.
- В силурийском периоде появились первые обитатели суши – растения-псилофиты и
беспозвоночные животные паукообразные.
 Это были первые животные, дышащие
атмосферным кислородом.
Это были первые животные, дышащие
атмосферным кислородом.
Домашнее задание. (Слайд 37)
Используя дополнительную литературу заполните представленную вам таблицу.
СПАСИБО ЗА ВНИМАНИЕ
Используемая литература:
- Большая энциклопедия Кирилла и Мефодия (2006г.)
- Н.Н.Иорданский «Эволюция Жизни» (2001г.)
Палеозойская эра – обзор
Фаза 4: Заполнение Глубокой впадины (триас)
По мере того, как палеозойская эра подходила к концу, обнаженные склоны Свердрупской впадины постепенно трансгрессировались. Значительное повышение базового уровня положило начало Фазе 4, и море достигло суши далеко за пределы прежних окраин бассейна. Между окраинами бассейна и осью бассейна существовала большая батиметрическая разница, вероятно, более 2 км. Этот глубокий бассейн начал формироваться в раннем каменноугольном периоде по мере развития рифтогенеза, а затем термического опускания и продолжал расширяться в сторону суши. Он углублялся на протяжении всего позднего палеозоя, поскольку скорость опускания центральной части Свердрупской впадины превышала многолетнюю скорость поступления наносов.
Он углублялся на протяжении всего позднего палеозоя, поскольку скорость опускания центральной части Свердрупской впадины превышала многолетнюю скорость поступления наносов.
Новый режим осадконакопления установился на заре мезозойской эры. Климат был теплым и сухим, в осадконакоплении теперь преобладали терригенные породы, а морская жизнь была скудной и не имела ничего общего с ее пермским аналогом. Фаза 4 продолжалась до позднего триаса (основной ретий), когда крупный тектонический эпизод установил очень разные режимы осадконакопления и тектонические процессы и положил начало новой фазе развития бассейна.Литостратиграфия и стратиграфия секвенций фазы 4 показаны на рис. 15, а на рис. 16 представлен стратиграфический разрез этих слоев от восточной окраины бассейна до участка в центре бассейна. Четыре последовательности второго порядка составляют последовательность первого порядка Фазы 4, и каждая из них характеризуется особым режимом осадконакопления и тектоническим режимом (рис. 16 и 17).
16 и 17).
Рис. 15. Стратиграфическая схема триаса-юры. Последовательности первого порядка соответствуют тектоническим фазам бассейна.
Рис. 16. Стратиграфический разрез пластов фазы 4 восточной части Свердрупского бассейна. За это время была заполнена глубокая центральная впадина.
Рис. 17. Обнажение на Грили-Фьорд, остров Элсмир, иллюстрирующее четыре последовательности второго порядка, которые составляют пласты фазы 4. B , Барроу Фм; Bj , Bjorne Fm; GP , Gore Point Fm; H , Хейберг Фм; HB , Хойл Бэй Фм; MH , Мюррей Харбор Фм; PB , Пэт Бэй Фм.
Раннетриасовые дельтовые и морские толщи имеют мощность до 2000 м и составляют первую толщу второго порядка (рис. 15). Основные дельты начали проградировать в бассейн с юга и востока (форма Бьорне) (рис. 18А), и это огромное количество терригенных отложений было получено главным образом из девонских терригенных отложений, которые окружали бассейн и простирались над кратоном. Внезапное увеличение запасов терригенных пород предполагает, что региональное поднятие кратонных областей было связано с тектоническим эпизодом, положившим начало этой новой фазе.Отсутствие растительного покрова из-за предшествующего вымирания P-T также, вероятно, способствовало этому увеличению предложения (Midwinter et al., 2017). На протяжении всего раннего триаса песчаные дельты продвигались в бассейн (Embry, 1991a, 2011) и отодвигали границу шельфа/склона на значительное расстояние к северу и западу. Крупные подводные конусы выноса отлагались в глубоком бассейне (форма Блайнд-Фьорд) на протяжении всего раннего триаса. Значительные отложения также поступали из Крокерленда (Embry, 1993a) (рис. 18A), и данные об обломочном цирконе из этих отложений указывают на то, что девонские обломочные клиновидные пласты были основным источником, а пермские магматические породы были локальным источником на северо-востоке (Omma et al. ., 2011; Анфинсон и др., 2016; Алонсо-Торрес и др., 2018).
Внезапное увеличение запасов терригенных пород предполагает, что региональное поднятие кратонных областей было связано с тектоническим эпизодом, положившим начало этой новой фазе.Отсутствие растительного покрова из-за предшествующего вымирания P-T также, вероятно, способствовало этому увеличению предложения (Midwinter et al., 2017). На протяжении всего раннего триаса песчаные дельты продвигались в бассейн (Embry, 1991a, 2011) и отодвигали границу шельфа/склона на значительное расстояние к северу и западу. Крупные подводные конусы выноса отлагались в глубоком бассейне (форма Блайнд-Фьорд) на протяжении всего раннего триаса. Значительные отложения также поступали из Крокерленда (Embry, 1993a) (рис. 18A), и данные об обломочном цирконе из этих отложений указывают на то, что девонские обломочные клиновидные пласты были основным источником, а пермские магматические породы были локальным источником на северо-востоке (Omma et al. ., 2011; Анфинсон и др., 2016; Алонсо-Торрес и др., 2018).
Рис. 18. (А) Палеогеография позднего раннего триаса (спатия) с крупными дельтами вдоль восточной и южной окраин бассейна. В глубоком бассейне отложились толстые подводные конусы выноса. Неглубокие шельфовые пески на северной окраине были получены из Крокерленда. (B) Палеогеография раннего позднего триаса (карния) с Крокерландом, обеспечивающим значительные объемы отложений. В западной части бассейна шельфовые пески простирались на юг по бассейну.На востоке располагалась глубоководная котловина.
18. (А) Палеогеография позднего раннего триаса (спатия) с крупными дельтами вдоль восточной и южной окраин бассейна. В глубоком бассейне отложились толстые подводные конусы выноса. Неглубокие шельфовые пески на северной окраине были получены из Крокерленда. (B) Палеогеография раннего позднего триаса (карния) с Крокерландом, обеспечивающим значительные объемы отложений. В западной части бассейна шельфовые пески простирались на юг по бассейну.На востоке располагалась глубоководная котловина.
Краевое поднятие, за которым последовала крупная трансгрессия, произошло на стыке раннего и среднего триаса, и скорость опускания и поступления наносов значительно уменьшилась. Темный битуминозный ил и ил (форма Мюррей-Харбор) были широко отложены, а с восточной и южной окраин произошло незначительное вырастание шельфовых песчаников (форма Рош-Пойнт) (рис. 16 и 17). Толщины среднего триаса, составляющие толщу второго порядка, не превышают мощности 300 м.Вдоль северо-западной окраины бассейна эти пласты состоят в основном из сланцев и алевролитов, хотя очень мелкие песчаниковые образования среднего шельфа, отобранные в скважине на северной окраине, вероятно, происходят из Крокерленда (Kondla et al., 2015).
После очередного тектонического эпизода поднятия и трансгрессии в самом раннем позднем триасе скорость опускания и поступления наносов снова значительно возросла, и смешанный карбонатно-кремнисто-обломочный шельф/склон (формации мыса Сент-Эндрю, Гор-Пойнт) значительно продвинулся в бассейн (рис.16). Мощность этих пластов достигает 1400 м. Это продвижение было остановлено еще одной трансгрессией в раннем карнии, но кремнисто-обломочные породы снова быстро преодолели опускание и песчаные отложения внутреннего шельфа (форма Пэт-бэй), образовавшиеся в сторону моря по склону с преобладанием ила и ила и отложениям внешнего шельфа (залив Хойл) (рис. 16 и 17). Примечательно, что в среднем и позднем карнии Крокерланд был преобладающим источником отложений для бассейна Свердруп, а песчаные шельфовые отложения (форма Пат Бей) простирались на юг через весь бассейн на западе (рис.18B) (Эмбри, 1993а; Анфинсон и др., 2016).
Данные по детритовому циркону из этих отложений, полученных из Крокерленда, показывают, что в юго-западной части северной окраины, в основном месте поступления отложений во время карнийского периода, девонские кремнисто-обломочные породы Крокерленда были первичными материнскими породами. Однако в северо-восточной части северной окраины каменноугольно-триасовые цирконы были обнаружены в карнийских песчаниках (Omma et al., 2011), что указывает на то, что Уральский/Новоземельский ороген также поставлял отложения в эту часть Свердрупской впадины ( Анфинсон и др., 2016). Как обсуждалось в Anfinson et al. (2016), похоже, что заключительная, позднетриасовая фаза Уральского/Нового орогена (Zhang et al., в печати) уничтожила морской путь между орогеном и Крокерлендом, позволив дренажу с верховьями орогена простираться до северо-восток Свердрупской впадины (см. рис. 7 в Anfinson et al., 2016).
Трансгрессия в середине позднего триаса (последний карний) снова отодвинула береговую линию к краям бассейна. Вслед за этим значительно увеличилось поступление терригенных пород из Крокерленда, и в норийской последовательности второго порядка не было обнаружено отложений, происходящих из кратонных областей к востоку и югу от бассейна (Embry, 2011).В течение норийского периода мелководные шельфовые пески (Romulus Mbr, Heiberg Fm) проградировали через ил и ил преддельты (Barrow Fm) (1000 м), и бассейн продолжал заполняться с северо-востока на юго-запад (рис. 19). Более 100 000 кубических километров отложений северного происхождения были отложены в бассейне Свердруп в течение норийского периода. К концу норийского века глубоководная центральная часть Свердрупской впадины, существовавшая более 100 млн лет, была заполнена, и на большей части впадины имелся мелководный морской путь (рис.19А).
Рис. 19. (A) Позднетриасовая (норийская) палеогеография с Крокерландом, являющимся единственным источником отложений. Самый высокий вход был на северо-востоке, а центральный бассейн заполнялся проградацией на юго-запад. (B) Стратиграфический разрез пластов норийской последовательности второго порядка в восточной части бассейна Свердруп, иллюстрирующий проградацию ЮЗ.
Четыре образца норийского песчаника, расположенные вдоль трансекты с севера на юг в восточной части бассейна, содержат большое количество цирконов от каменноугольного до позднетриасового возраста, что указывает на то, что Уральский/Новый ороген был важным исходным регионом для юго-западного простирания норийского обломочные породы (Анфинсон и др., 2016). Неопубликованные данные об обломочном цирконе из пробы из юго-западной части северной окраины (Л. Химан, личное сообщение, 2013 г.) указывают на то, что девонские обломочные породы на Крокерленде снабжали эту часть бассейна, хотя поступление наносов было очень низким по сравнению с таковым из северо-восточный район. Во время фазы 4 соляные купола и стены, образовавшиеся из свиты Отто-Фьорд, росли вверх в центральной части бассейна и иногда образовывали эмерджентные острова (Balkwill, 1978; Embry, 1991a; Harrison and Jackson, 2014).
Палеозойская эра
Палеозойская эраОнлайн экспонаты: Геологическая шкала времениПалеозойская эра
Палеозой занял более половины — примерно 300 миллионов лет (от 542 до 251 млн лет назад)* — фанерозоя. В палеозое было шесть основных континентальных массивов суши; каждый из них состоял из разных частей современных континентов.Например, в начале палеозоя современное западное побережье Северной Америки проходило с востока на запад вдоль экватора, а Африка находилась на Южном полюсе. Эти палеозойские континенты испытали огромное горообразование на своих окраинах, а также многочисленные вторжения и отступления мелководных морей во внутренних районах. Большие обнажения известняка, подобные изображенному здесь, являются свидетельством этих периодических вторжений континентальных морей. Палеозойская эра заключена в скобки времен глобальных суперконтинентов.Эпоха началась с распадом мирового континента Паннотии и завершилась с образованием Пангеи, когда континенты Земли снова соединились.
Многие палеозойские породы имеют экономическое значение. Например, большая часть известняка, добываемого для строительных и промышленных целей, а также угольные месторождения Западной Европы и востока США образовались в палеозое.
Палеозой делится на шесть периодов: кембрий, ордовик, силур, девон, карбон (в У.S., он делится на миссисипский и пенсильванский периоды) и пермский. Большинство этих названий происходит от мест, где впервые были изучены породы этого возраста. Камбрия — латинское название Уэльса, а ордовики и силуры — два валлийских кельтских племени. Девон назван в честь Девоншира, Англия. Миссисипи назван в честь верхней долины реки Миссисипи, , а не штата Миссисипи, в котором очень мало горных пород этого возраста; однако пенсильванский — это , названный в честь штата Пенсильвания.Пермь была описана по породам в районе Перми, города на Урале в России. Исключением из этого соглашения об именах является каменноугольный период; его название означает «угольоносный», и это время, когда по всему миру формировались обширные угольные пласты.
Жизнь
Две великие фауны животных господствовали в морях палеозоя. «Кембрийская фауна» типична для кембрийских океанов; хотя в кембрии присутствовали представители большинства типов, в морях преобладали трилобиты, нечленораздельные брахиоподы, моноплакофорные моллюски, гиолитиды, «мелкие раковинные окаменелости» неопределенного систематического положения и археоциатиды.Хотя все они, кроме археоциатид, выжили после кембрия, их разнообразие уменьшилось после ордовика. В позднепалеозойских морях преобладали криноидные и бластоидные иглокожие, членистые брахиоподы, граптолиты, таблитчатые и морщинистые кораллы.
К концу ордовика жизнь уже не ограничивалась морями. Растения начали колонизировать землю, за ними в силурийском периоде последовали беспозвоночные, а в верхнем девоне — позвоночные. Ранние четвероногие этого времени были похожими на амфибий животными, которые в конечном итоге дали начало рептилиям и синапсидам к концу палеозоя.Одна из самых ранних наземных фаун четвероногих, известных в мире, происходит из Джоггинса, Новая Шотландия.
Наземные растения быстро заняли освободившиеся ниши на суше. К концу девона в ландшафте преобладали леса из прогимноспермов, таких как Archaeopteris . К концу палеозоя по ландшафту распространились саговники, глоссоптериды, примитивные хвойные и папоротники.
Пермское вымирание 251,4 млн лет назад опустошило морскую биоту: вымерли таблитчатые и морщинистые кораллы, бластоидные иглокожие, граптолиты, трилобиты и большинство морских лилий.Одна линия морских лилий выжила, но они никогда больше не будут доминировать в морской среде.
Местонахождения палеозойских ископаемых
-
Река Алдан, Сибирь: Нижнекембрийская фауна с этого участка в Якутии, Сибирь, прослеживает раннюю эволюцию животных со скелетами.
-
Сланец Берджесс: Одной из величайших когда-либо сделанных ископаемых находок является фауна Берджесса в Британской Колумбии. Тысячи окаменелостей мягкотелых животных рисуют нам картину кембрийской морской жизни.
-
Бассейн Каннинг, Австралия: Большое разнообразие ископаемых брюхоногих моллюсков было обнаружено в бассейне Каннинг.
-
Гласс-Маунтинс, Техас: Пермские окаменелости из Гласс-Маунтинс относятся к мелководным тепловодным морским обитателям.
-
Диапазон Хаус, Юта: В сланцах Уилер и формации Марджум было обнаружено множество кембрийских тварей, оба из которых обнажены в районе Хаус.
-
Джоггинс, Новая Шотландия: Угольные болота каменноугольного периода были домом для первых четвероногих, таких как Dendrerpeton .
-
Озеро Виннипег, Манитоба, Канада: Известняки этого региона сохранили множество впечатляющих окаменелостей ордовикских макроводорослей.
-
Мраморные горы, Калифорния: Оленеллидных трилобитов много в сланцах Латам, здесь, в пустыне Мохаве.
-
Mazon Creek, Illinois: Это место стало известно своими железными конкрециями, в которых сохранились как растения, так и морские беспозвоночные.
-
Райни Черт, Шотландия: Это был один из самых важных источников окаменелостей ранних наземных растений и наземных членистоногих. Анатомия образцов сохранена в трехмерных деталях.
-
Горы Уайт-Инио, Калифорния: Вы можете посетить кембрийские рифы в горах восточной Калифорнии.
Ресурсы
Глубокое время | Периссодактиль
Perissodactyls впервые появились около 55,8 млн лет назад (миллион лет назад). Это кажется очень долгим. Анатомически современные люди ( Homo sapiens ) существуют всего около 200 000 лет. Таким образом, непарнокопытные существовали примерно в 275 раз дольше, чем наш вид, и более чем в 9000 раз дольше, чем зарегистрированная история человечества!
Однако по сравнению с возрастом Земли и продолжительностью жизни на этой планете непарнокопытные — пришельцы.Геологи оценивают возраст Земли в 4,54 миллиарда лет. Исходя из этого, сложная животная жизнь существовала только около 10% земной истории, а непарнокопытные — только последний 1%. Как видите, земля очень стара!
Эти последние 10%, называемые фанерозойским эоном, охватывают 545 миллионов лет. Он подразделяется на три периода, известные как «эры»: палеозой («старая жизнь»), мезозой («средняя жизнь») и кайнозой («новая жизнь»). На этом веб-сайте мы в основном будем рассматривать события, произошедшие в кайнозойскую эру.
Млекопитающие впервые появились в мезозое, около 200 млн лет назад, но на протяжении большей части эпохи доминирующими наземными животными были динозавры. Мезозойские млекопитающие, как правило, меньше и гораздо реже, чем современные; самое крупное из известных мезозойских млекопитающих, Repenomanus из мелового периода Китая, достигало примерно метра в длину.
После крупного вымирания в конце мезозоя 65 млн лет назад (наиболее вероятной причиной которого было падение огромного метеорита) единственной выжившей группой динозавров были предки современных птиц.Напротив, было много видов млекопитающих, которые пережили массовое вымирание. Среди них были предки современной фауны млекопитающих. Кайнозой был и остается веком млекопитающих.
Летопись окаменелостей кайнозоя очень обширна, и палеонтологи смогли собрать воедино очень подробную историю жизни этого временного периода. Он разделен на семь эпох. Многое изменилось за последние 65 миллионов лет, и каждая из этих эпох очень отличается с точки зрения существовавших видов (непарнокопытных и других видов), конфигурации континентов и глобального климата.Мы рассмотрим эти факторы более подробно в разделах «Эволюция» и «Окружающая среда» на этом веб-сайте
.Нажмите на имена на боковой панели, чтобы просмотреть сводку основных особенностей каждой эпохи.
Палеозойская эра (от 541 млн лет до 252 млн лет назад)
В палеозойскую эру, длившуюся 289 миллионов лет, растения и рептилии начали перемещаться с моря на сушу. Эпоха делится на шесть периодов: пермский, каменноугольный, девонский, силурийский, ордовикский и кембрийский.Несколько раз в течение этой эпохи в Канзасе появлялись и исчезали моря. Породы двух последних периодов эпохи — каменноугольного и пермского — можно найти на поверхности в некоторых частях центрального и восточного Канзаса.
Пермский период (от 299 миллионов лет назад до 252 миллионов лет назад)
Циклы мелководья, приливных отмелей и суши привели к образованию отложений известняка, сланца, песчаника, доломита, гипса и кремня. Пермские породы находятся на поверхности Флинт-Хиллз и южно-центральной части Канзас-Рэд-Хиллз.Нефть и газ добываются из пермских пород, а соль, оставшаяся после высыхания морей, добывается под землей в центральном Канзасе.
Окаменелости пермских морских животных включают моллюсков, плеченогих, мшанок, морских лилий, кораллы, акульи зубы и одноклеточные фузилиниды. Были обнаружены наземные окаменелости листьев и насекомых. Массовое вымирание произошло в конце этого периода.
Каменноугольный период (от 359 миллионов лет назад до 299 миллионов лет назад)
Каменноугольный период состоит из двух подпериодов: пенсильванского и миссисипского.
Пенсильванский подпериод (от 323 миллионов лет назад до 299 миллионов лет назад): Циклы мелководья, болот и речных русел привели к образованию отложений известняка, песчаника и сланца, обнаруженных на поверхности в восточной части Канзаса. На юго-востоке Канзаса реки и ручьи впадали в море во время пенсильванского периода и откладывали песок и другие отложения в эстуариях и дельтах. Образовавшиеся толстые песчаники образуют холмы Чатокуа. Точно так же сланцы и песчаники, отложившиеся в ранний пенсильванский подпериод, образуют низменность чероки на юго-востоке Канзаса и являются вторыми по возрасту породами, обнаруженными на поверхности в штате.Складчатость и разломы слоев горных пород в этот период создали Центральное Канзасское поднятие, горный хребет, ныне глубоко погребенный. Окаменелости морских животных Пенсильвании включают плеченогих, мшанок, кораллов, морских лилий, моллюсков и одноклеточных фузилинидов. Наземные окаменелости включают растения, земноводных и ранних рептилий. Нефть, газ и уголь добываются из подземных слоев горных пород Пенсильвании.
Миссисипский субпериод (от 359 миллионов лет назад до 323 миллионов лет назад): Циклы мелководья и суши привели к образованию отложений известняка, песчаника, сланца и кремня.Найденные на поверхности далеко на юго-востоке Канзаса, они являются самыми старыми поверхностными породами. Морские окаменелости Миссисипи включают морских лилий, брахиопод, мшанок и моллюсков. Нефть и газ добываются из подземных пород Миссисипи.
Девонский период (от 419 миллионов лет назад до 359 миллионов лет назад) 90 004 морей покрывали Канзас в течение части этого периода. В недрах встречаются девонские известняки, сланцы, доломиты, песчаники и кремни. Микроскопические окаменелости и некоторые мелкие морские окаменелости были обнаружены в кернах горных пород, поднятых из-под земли во время бурения на нефть и газ.
Силурийский период (от 443 миллионов лет назад до 419 миллионов лет назад) 90 004 морей покрывали Канзас в течение части этого периода. В недрах встречаются силурийские известняки, сланцы, доломиты, песчаники и кремни. Микроскопические окаменелости и некоторые мелкие морские окаменелости были обнаружены в кернах горных пород, поднятых из-под земли во время бурения на нефть и газ.
Ордовикский период (от 485 миллионов лет назад до 443 миллионов лет назад) 90 004 морей покрывали Канзас в течение части этого периода.Доломит, известняк, песчаник, сланцы и кремни этого периода находятся в недрах, а нефть и газ добываются из ордовикских пород. Микроскопические окаменелости и некоторые мелкие морские окаменелости были обнаружены в кернах горных пород, поднятых из-под земли во время бурения на нефть и газ.
Кембрийский период (от 541 млн лет до 485 млн лет назад)
Кембрийский период в Канзасе в начале периода был отмечен эрозией, за которой позже последовало мелководье.В недрах встречаются кембрийские доломиты, песчаники, известняки и сланцы. Микроскопические окаменелости и некоторые мелкие морские окаменелости были обнаружены в кернах горных пород, поднятых из-под земли во время бурения на нефть и газ.
|
от от 540 до 248 миллионов лет назад Введение: Если бы вы могли видеть спутниковый снимок Земли такой, какой она была 540 миллионов лет назад вы бы не признали его своим домом. Большая часть суши была объединена в один континент с несколькими небольшими разбросанными материки рядом. Большой континент, Гондвана, был близок к южный полюс, поместив то, что сегодня является Сибирью и Китаем, рядом с экватором! Если если бы вы приземлились на этой странно выглядящей Земле, вы были бы еще более смущает чужой пейзаж. Не было деревьев, травы или растения, покрывающие землю – только голые скалы и рыхлые скалистые почва.Тогда не было ни птиц, ни насекомых, ни лягушек. единственным звуком были бы океанские волны, бьющиеся о берега скалистых пляжей. и ветер дует в уши. В безветренный день далеко вдали от берега стояла тишина, какой никогда не знал ни один человек на открытом воздухе. И не было ничего, абсолютно ничего, есть на земле. В допалеозойское (докембрийское) время эра) жизнь распространилась через океан (заняв около 3 миллиардов лет сделать это), поэтому в океане вы могли найти что-нибудь поесть: водоросли, губки, другие мягкотелые беспозвоночные, кораллы, членистоногие (например, трилобиты), многощетинковые черви (6 семейств которых дожили до наших дней!), моллюсков и некоторых довольно причудливых иглокожих (морские звезды и ежи) — но ничего, что я хотел бы проглотить. Величайшее изменение появление Земли произошло в палеозойскую эру, охватывающую всего 292 миллиона лет. От конец палеозоя бесплодный, безжизненные континенты Земли превратились в густые леса и болота. В наконец воздух наполнился жужжанием насекомых и звуками земноводных и пресмыкающихся. В океане жизнь пошла от примитивные рифы и простые растительноядные беспозвоночные докембрия время появления плавающих беспозвоночных хищников и первых рыб. Движение Континентов: Больше течении палеозойской эры, Гондвана и частично разорвались вверх, и континенты двигались вокруг океана в медленном танце. Гондвана составляли Африка, Южная Америка, Австралия, Антарктида, Азия, маленький клочок земли, ставший позже Флоридой, и большую часть восточного побережья США.Гондвана путешествовал так далеко на юг за это время, что южный полюс оказалась на территории современного Марокко на северо-западе Африки, на юге Бразилии, а затем в Южной Африке к концу палеозой. Северная Америка, Гренландия и Шотландия объединились в одно целое и столкнулись с Британскими островами и континент, состоящий из Скандинавии и Европы. От к концу палеозойской эры все континенты пришли снова вместе, чтобы создать суперконтинент, который мы называем Пангеей.Это в этот момент Южная Америка присоединилась к Северной Америке, в то время как северная Африка врезалась в часть Северной Америки который станет восточным побережьем США, в результате чего Аппалачи Горы подняться. Когда это произошло, Нью-Йорк находился примерно в 20 градусах к югу от экватора и Торонто и Квебек был на экваторе! Срок службы: В В палеозойскую эру жизнь на суше быстро и фантастически разнообразилась. и в океане.За это время жизнь в море резко изменилась. время. Появились трилобиты, моллюски, ежи и морские звезды в начале палеозоя в рамках «кембрийского взрыва». Позже появились бесчелюстные рыбы, затем рыбы с челюстями и даже акулы, наполненные моря к концу палеозоя. Видовое разнообразие позвоночных и беспозвоночные сошли с ума, и некоторые довольно причудливые существа появился: включая бронированную рыбу длиной 30 футов ( Dunkleosteus ) и множество беспозвоночных кошмарной формы. Внешний вид растений на суше подготовили почву для возможной колонизации континенты животными. Нет только растения в конечном итоге станут источником пищи для этих животных, но растения изменили свойства самой земли. До растений, не было корней, чтобы стабилизировать почву от эрозии. И это также повлияло на течение рек; более устойчивая почва вызвала реки больше извивались, и поэтому реки откладывали наносы по-разному, создание разнообразных мест обитания для будущих животных.Четное гибель растений затронула землю. Мертвые растения смешанные в почву, образующую гумус, темное органическое богатство насыщенных питательными веществами почва. Большие деревья, погибшие в болотах, со временем стали крупные залежи угля. Во всех этих и многих других отношениях растения подготовили землю к нашествию животных. После распространение наземных растений, животным не потребовалось много времени в море, чтобы эволюционировать для жизни на берегу.К середине палеозоя, скорпионы и нелетающие насекомые были одними из первых существа для жизни на суше. Это Потребовалось еще 90 миллионов лет, чтобы позвоночные появились на берегу. От конце палеозоя существовали гигантские растения хвоща высотой как деревья, ранние хвойные леса, летающие насекомые, земноводные (некоторые из них довольно большие и причудливые), рептилии и терапсиды (называемые млекопитающие рептилии).Но полное отсутствие травы, цветы и птицы наверняка покажутся современным людям странными и жуткий — Земля по-прежнему отображал довольно чужеродную сцену. Смерть: Хотя жизнь сделала большой скачок в палеозойскую эру, несколько массовых вымираний жизни состоялось. С каждым угасанием до 60% или больше видов, живших в то время, погибло; но после каждого события новые виды появлялись очень быстро и диверсифицировались в еще более разновидность.Потом, в конце палеозоя произошло самое массовое вымирание со времен жизни произошло («пермское вымирание»). На суши, большинство видов терапсидов было уничтожено, но это было гораздо серьезнее в океане. Более 90% морских виды внезапно исчезли. Хотя климатические изменения были предложено в качестве причины, в окаменелостях нет прямых доказательств записывать.После После этого ужасного события мир вступил в новое время перемен, называемое мезозой Эра, также известная как Эпоха динозавров . |
Палеозойская эра: определение, хронология и события — видео и стенограмма урока
Кембрийский взрыв
542 миллиона лет назад простые формы жизни процветали в океанской среде, но на континентах все еще не было жизни.Палеозойская эра станет свидетелем взрыва разнообразия разнообразных форм жизни в океане и, в конечном итоге, совершит прыжок на сушу. Этот быстрый рост разнообразия животных известен как Кембрийский взрыв . К силурийскому периоду на сушу вышли как растения, так и беспозвоночные животные. К девонскому периоду позвоночные рыбы вышли из моря и стали обитателями суши. Оказавшись на суше, жизнь стала такой же разнообразной, как море. В каменноугольный период у крылатых насекомых появился полет.
Трилобиты
Вероятно, самым успешным животным палеозойской эры был трилобит . Небольшие членистоногие, трилобиты, средний размер которых составляет от 1 до 4 дюймов. Окаменелости трилобитов были обнаружены по всему миру в морских породах, что позволяет предположить, что они никогда не переходили на сушу. Хотя сейчас они вымерли, на пике их развития насчитывалось около 17 000 видов трилобитов. Их изобилие и разнообразие в летописи окаменелостей сделали трилобитов важными и полезными в изучении истории Земли.
Тектоника плит и пермское вымирание
На протяжении большей части палеозоя на земном шаре существовало несколько континентов. К концу эры континенты объединились, чтобы сформировать самый последний суперконтинент, Пангею.
В то время как начало палеозоя было отмечено взрывом жизни, конец был отмечен взрывом, названным Пермским вымиранием . Когда 251 миллион лет назад произошло крупнейшее массовое вымирание в истории Земли, вымерло 96% всей морской жизни и 70% наземной жизни.Существует несколько теорий относительно причины вымирания, в том числе:
- Удар астероида
- Массовое извержение вулкана
- Истощение кислорода в океане
- Значительное глобальное потепление
Жизни потребуется еще 10 миллионов лет, чтобы оправиться от этого массового вымирания. Следующая эра, триасовая, положила начало появлению крупных рептилий.
Краткий обзор урока
Палеозойская эра началась 542 миллиона лет назад с появлением сложных форм жизни и закончилась 251 миллион лет назад крупнейшим массовым вымиранием, которое когда-либо переживал мир.Это самая древняя и продолжительная эра фанерозойского эона. Эпоху обычно еще больше разбивают на шесть основных периодов.
- Кембрийский период: с 542 до 488,3 млн лет назад
- Ордовик: 488,3–443,7 млн лет назад
- Силур: от 443,7 до 416 миллионов лет назад
- Девон: 416–359,2 млн лет назад
- Каменноугольный период: 359,2–299 миллионов лет назад
- Пермь: 299–251 миллион лет назад
Палеозойская эра станет свидетелем взрыва разнообразия разнообразных форм жизни в океане и, в конечном итоге, совершит прыжок на сушу.Этот быстрый рост разнообразия животных известен как Кембрийский взрыв . Вероятно, самым успешным животным палеозойской эры был трилобит .
В то время как начало палеозоя было отмечено взрывом жизни, конец был отмечен взрывом, названным Пермским вымиранием . Когда 251 миллион лет назад произошло крупнейшее массовое вымирание в истории Земли, вымерло 96% всей морской жизни и 70% наземной жизни.К концу эры континенты воссоединились, чтобы сформировать самый последний суперконтинент, Пангею.
Шкалы времени – Ископаемые лошади
Стратиграфия – это научное исследование геологической истории. Он возник около 200 лет назад благодаря работе английского инженера по имени Уильям Смит, который видел одни и те же слои горных пород, содержащих ископаемые, при обследовании различных частей Англии.
Пример стратиграфии. Это наблюдение привело Смита к составлению карт этих слоев, по которым он мог точно предсказать расположение неоткрытых угольных пластов, потому что он знал, какие породы перекрывают известные угольные пласты в других местах Англии.
Он вывел два важных принципа стратиграфии:
- Каждый пласт горных пород имеет характерный набор окаменелостей, что позволяет связать сходные пласты в разных местах
- Более молодые пласты накладываются на более старые пласты.
Эти принципы позволяют геологам датировать слои горных пород и содержащиеся в них окаменелости относительно других слоев. Эта относительная датировка послужила геологам основой для научного разделения истории жизни на Земле.Позже открытие радиоактивности в 20 веке дало ученым инструменты для абсолютной датировки горных пород и уточнения этих подразделений.
В этом разделе мы собираемся определить основные подразделения геологического времени, начиная с самых общих подразделений и продвигаясь к более конкретным подразделениям.
Геологическое время
Жизнь на Земле зародилась более 3,5 миллиардов лет назад, но только около 570 миллионов лет назад (млн лет назад) жизнь стала в изобилии.Время, когда жизнь стала изобильной, называется фанерозой , что означает «много животных».
Фанерозойский эон делится на три эры. Самая старая эра называется Палеозойская , что означает «древняя жизнь», а самая поздняя эра — Кайнозойская , что означает «недавняя жизнь». Между ними зажата мезозойская — между прочим, «мезо» в переводе с греческого означает середина.
В течение палеозойской эры произошел драматический взрыв живых существ.Раньше было множество морских беспозвоночных, таких как черви, трилобиты, брюхоногие моллюски, кораллы и головоногие моллюски. Первые позвоночные появились около 500 миллионов лет назад, из них произошли рыбы, которые стали доминирующими позвоночными на планете. Около 410 миллионов лет назад от костистых рыб произошли амфибии. Земноводные были среди форм жизни, которые первыми колонизировали землю. Рептилии произошли от этого вида амфибий около 360 миллионов лет назад и постепенно начали вытеснять земноводных.
Иллюстрация геологического времени, созданная Геологической службой США.Рептилии были доминирующими позвоночными в течение мезозойской эры , включая господство динозавров. Млекопитающие тоже появились в эту эпоху, около 220 миллионов лет назад, но они были очень мелкими — не крупнее современных грызунов — и ни многочисленными, ни разнообразными. Около 200 миллионов лет назад от динозавров произошли первые птицы.
Кайнозойская эра началась около 65 миллионов лет назад и продолжается до сих пор! Его иногда называют «эпохой млекопитающих», потому что в эту эпоху млекопитающие стали доминирующими наземными животными.Эту эпоху мы рассмотрим наиболее подробно.
Кайнозойская эра первоначально называлась третьим и четвертым периодами. Эти два периода до сих пор называют третичным и четвертичным .
Четвертичный период состоит из нашей современной эпохи (голоцен) и плейстоцена.
