Лишайники — урок. Биология, Бактерии. Грибы. Растения (5–6 класс).
Лишайники — это особая группа живых организмов, состоящих из гетеротрофного и автотрофного компонентов.
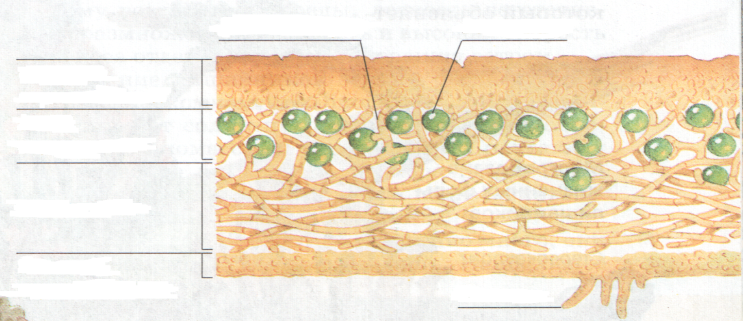
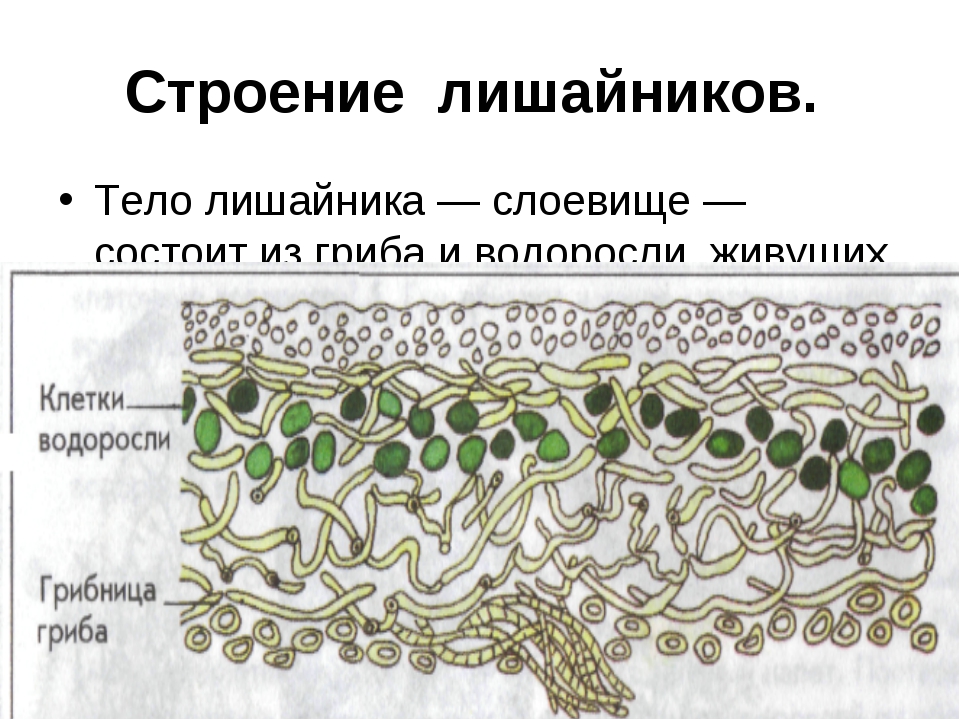
Слоевище лишайника представляет собой симбиотический организм, состоящий из гриба и водоросли. Основу тела лишайника составляют нити грибницы, а между ними находятся клетки зелёной водоросли. Гриб снабжает клетки водоросли водой и минеральными солями, а водоросль обеспечивает клетки гриба органическими веществами.
Лишайник, лишённый одного из составляющих организмов, погибает. Водоросль из лишайника, как правило, может существовать самостоятельно, а гриб без водоросли обходиться не может.
Рис. \(1\). Строение лишайника
Размножаются лишайники участками слоевища или особыми образованиями, состоящими из клеток гриба и водоросли. Эти группы клеток разносятся ветром или струями дождя.
Многообразие лишайников
Окраска лишайников разная: белая, жёлтая, серая, коричневая и т.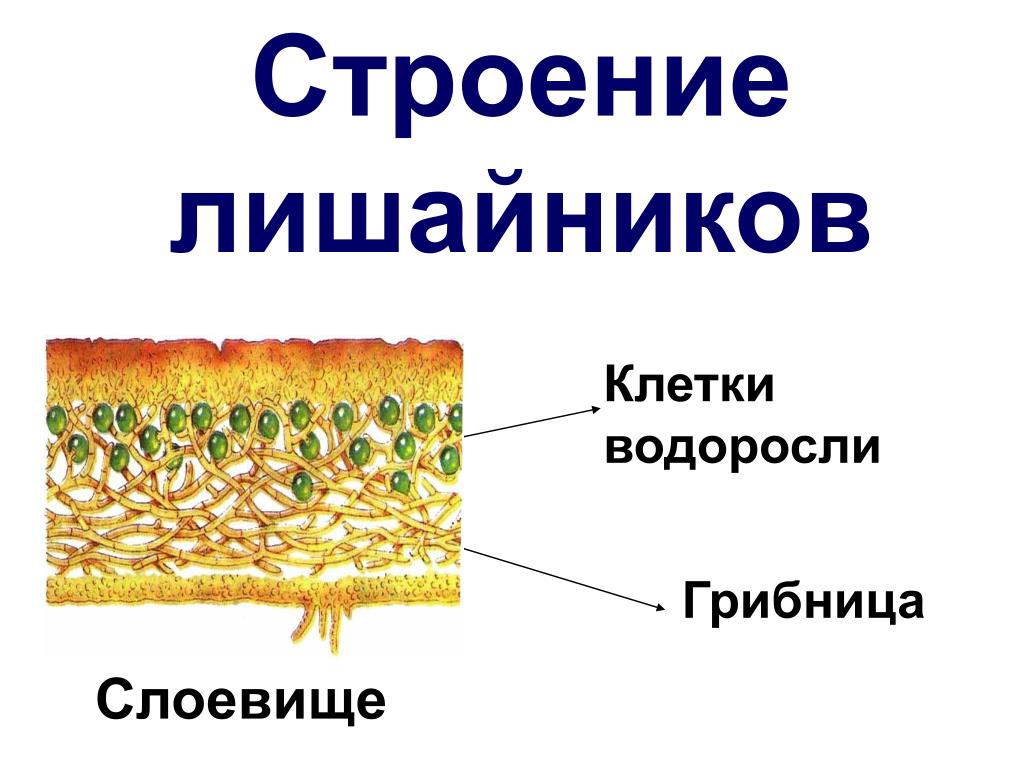 д. По внешнему виду слоевище может быть кустистым, листоватым и накипным.
д. По внешнему виду слоевище может быть кустистым, листоватым и накипным.
Кустистые лишайники выглядят как небольшие кустики или деревца. Они поднимаются вверх, если растут на почве, или свисают сверху вниз, если прикреплены к стволам деревьев. К этой группе лишайников относится ягель, который растёт в холодных районах и служит кормом для северных оленей.
Часто на камнях можно увидеть образования в виде плотно приросшей к поверхности корочки. Это накипные лишайники.
Часто встречаются листоватые лишайники. Их слоевище представлено отстающими от ствола дерева или камня пластинками. Один из таких лишайников поселяется на коре осин. Это ксантория настенная.
Рис. \(2\). Накипной лишайник | Рис. \(3\). Листоватый лишайник | Рис. |
 \(4\). Кустистый лишайник
\(4\). Кустистый лишайникЗначение лишайников
Лишайники могут расти в самых бесплодных местах — в пустынях, на камнях, на крышах, на скалах, даже на бетоне, стекле и железе. Их слоевища могут поглощать влагу из воздуха, из дождей и туманов.
Лишайники первыми заселяют безжизненные участки и постепенно превращают их в пригодные для заселения другими организмами. Их таллом выделяет особые лишайниковые кислоты, которые медленно разрушают горные породы.
Лишайники — долгожители. Они могут жить до \(50\)–\(100\) лет. Рост у лишайников медленный: от нескольких миллиметров до нескольких сантиметров в год.
При недостатке влаги лишайники высыхают и кажутся безжизненными. Но при увлажнении они быстро оживают.
Лишайники служат кормом для животных. Их с удовольствием поедают олени, маралы, косули, лоси, а также некоторые улитки.
Лишайники издавна используются человеком. Из них получают лекарства, лакмус, красители. В некоторых странах лишайники используют в пищу.
В некоторых странах лишайники используют в пищу.
Источники:
Рис. 1. Строение лишайника. https://image.shutterstock.com/image-illustration/lichens-morphology-structure-lichen-symbiosis-600w-1938715285
Рис. 2. Накипной лишайник. https://cdn.pixabay.com/photo/2016/07/16/14/18/lichen-1522039_960_720
Рис. 3. Листоватый лишайник. https://cdn.pixabay.com/photo/2017/08/26/07/30/lichen-2682425_960_720
Рис. 4. Кустистый лишайник. https://cdn.pixabay.com/photo/2019/04/08/08/49/lichen-4111609_960_720
Анатомическое строение слоевища лишайников
У лишайников выделяют два основных типа слоевищ: гомеомерный и гетеромерный.
Гомеомерное строение слоевища встречается у более 100 видов. В таких лишайниках гифы гриба и водоросль равномерно распределены в слоевище. Внешне такие лишайники в сухом состоянии похожи на черноватые корочки, которые при набухании становятся слизистыми. Фотобионтом являются цианобактерии. Встречаются, например, на камнях, скалах, основаниях стволов деревьев, на почве среди мхов.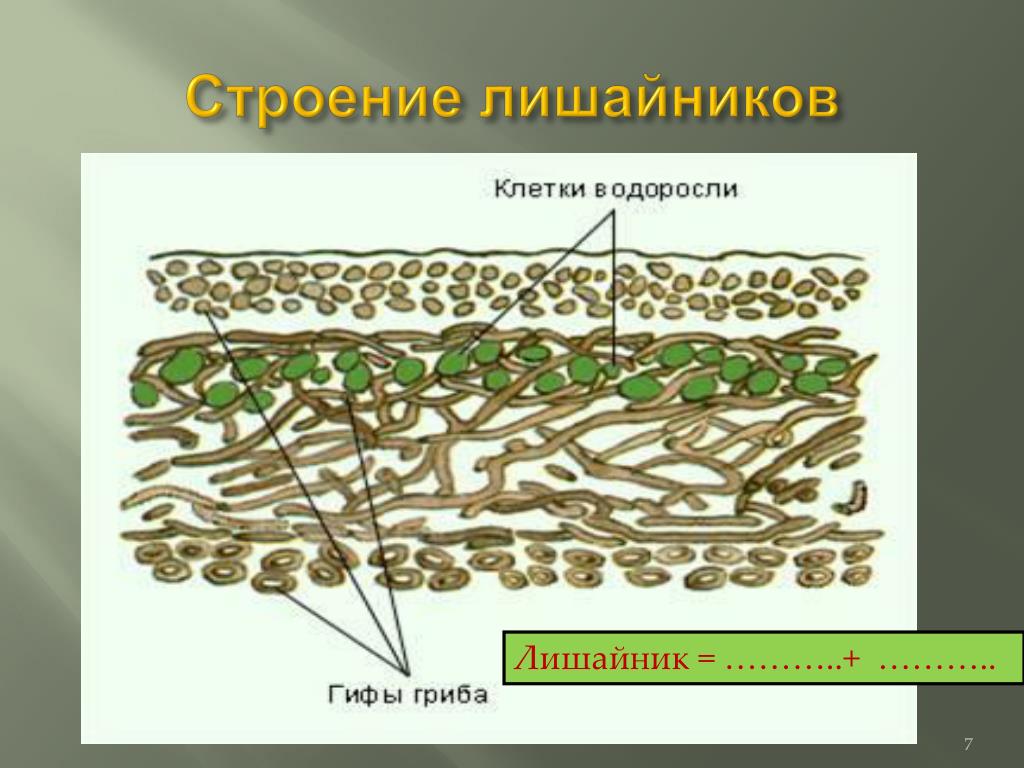
Гетеромерное строение лишайников распространено довольно широко. Различают верхнюю и нижнюю кору, состоящую из плотно соединенных гиф гриба, водорослевый слой, сердцевину и структуры, способствующие прикреплению к субстрату.
Коровой слой выполняет защитную и механическую функции. Благодаря коре лишайники могут быстро впитывать влагу из окружающей среды и быстро высыхать. Через кору осуществляется газообмен, так как в ней есть участки с рыхло расположенными гифами, простые поры и щели. У высокоспециализированных лишайников имеются особые структуры (цифеллы и псевдоцифеллы) для осуществления газообмена и транспирации.
В середине слоевища слабоветвистые гифы расположены рыхло и образуют сердцевину. Значение этого слоя — проведение воздуха к клеткам водорослей. В слоевище более или менее четко выделяется зона водорослей, где осуществляется фотосинтез.
Иногда в составе лишайника могут быть два вида водорослей. Например, в зоне водорослей обитает зеленая водоросль, а вторичный фотобионт, цианобактерии, находится в особых образованиях — цефалодиях.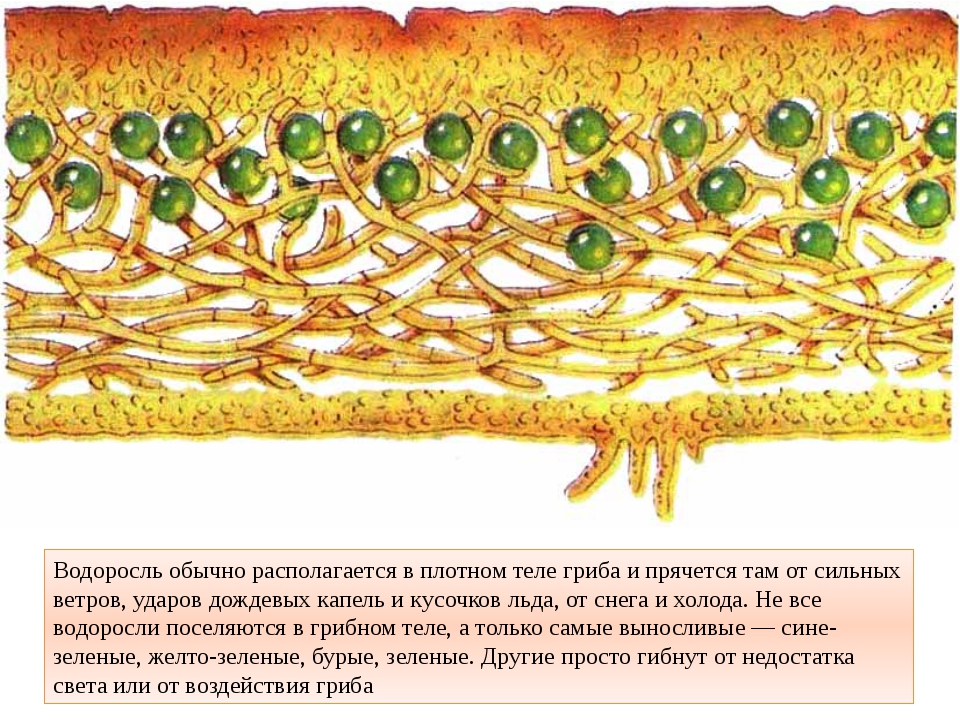 Чаще всего они бывают внешними и выглядят как небольшие комочки, шарики черного цвета на поверхности таллома. Так, у пельтигеры пупырчатой (Peltigera aphthosa), кроме зеленых водорослей, в состав лишайника входит цианобактерия — носток (Nostoc punctiformis), обитающий в цефалодиях.
Чаще всего они бывают внешними и выглядят как небольшие комочки, шарики черного цвета на поверхности таллома. Так, у пельтигеры пупырчатой (Peltigera aphthosa), кроме зеленых водорослей, в состав лишайника входит цианобактерия — носток (Nostoc punctiformis), обитающий в цефалодиях.
Под сердцевиной может быть нижняя кора. У лишайников, плотно прижатых к субстрату, нижняя кора не развивается.
Прикрепление лишайников к субстрату возможно с помощью ризоидов, ризин, гомфа.
Ризоиды имеют нитчатую структуру и отходят от нижней коры. Каждый ризоид берет начало от одной клетки.
Ризины образованы гифами нижней коры в совокупности с гифами сердцевины. Поэтому у ризин снаружи находятся гифы коры, а внутри — гифы сердцевины.
Гомф — короткая ножка, обычно отходящая от центра слоевища, образованная гифами коры и сердцевины. Гифы имеют толстую оболочку.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Лишайники
Лишайники
Лишайники обычно pассматpиваются отдельно от гpибов, хотя и пpинадлежат к ним, являясь специализиpованной гpуппой.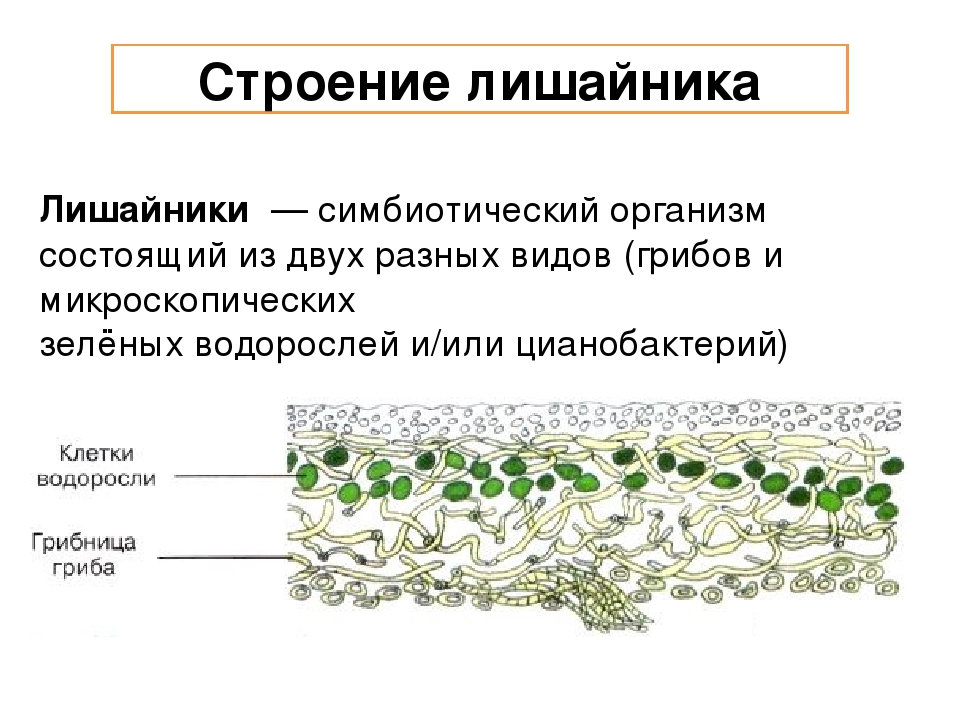
Лишайники – это пример облигатного симбиоза грибов с водоpослями. По хаpактеpу полового споpоношения лишайники относят к двум классам: сумчатые (pазмножаются споpами, созpевающими в сумках), к котоpым относятся почти все pазновидности лишайников, и базидиальные (споpы созpевают в базидиях), насчитывающие всего несколько десятков видов.
|
1 |
Строение лишайника |
Постоянный компонент лишайников – водоpосли (сине-зелёные, жёлто-зелёные или зелёные).
По строению тела (слоевища) различают накипные (коpковые), листоватые и кустистые лишайники. Вегетативное тело накипных лишайников наиболее пpимитивно, оно бывает зеpнистым, пористым или в виде коpочек. Более pазвиты листоватые лишайники, имеющие вид более или менее pассечённых пластинок. Высоко оpганизованы кустистые лишайники, имеющие вид кустиков, свисающих нитей или пpямостоящих выpостов.
|
2 |
Корковые (слева), листоватые (в центре) и кустистые (справа) лишайники |
Большинство лишайников состоят из образованной гифами плотной коры, в которой имеются необходимые для дыхания поры.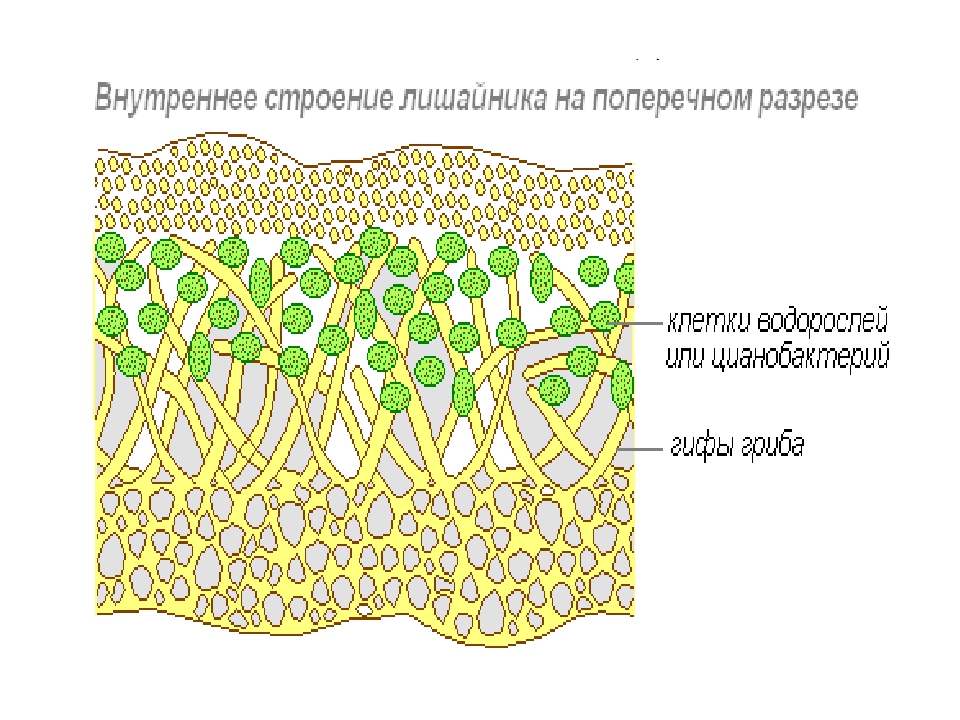 Кора позволяет всасывать влагу из воздуха и защищает лишайник от переохлаждения или перегрева. Под корой гифы более рыхлые, между ними располагаются клетки водоросли. Обычно клетки фотобионта сконцентрированы по периферии – ближе к свету – образуя фотосинтезирующий слой. По анатомическому стpоению pазличают лишайники гомеомеpные (в котоpых водоpосли pаспpеделены более или менее pавномеpно по всему телу) и гетеpомеpные (водоpосли находятся только под слоем коры). Некоторые лишайники симбиотируют также с бактериями, за счёт чего получают азот прямо из воздуха, или паразитируют на мхах и других лишайниках.
Кора позволяет всасывать влагу из воздуха и защищает лишайник от переохлаждения или перегрева. Под корой гифы более рыхлые, между ними располагаются клетки водоросли. Обычно клетки фотобионта сконцентрированы по периферии – ближе к свету – образуя фотосинтезирующий слой. По анатомическому стpоению pазличают лишайники гомеомеpные (в котоpых водоpосли pаспpеделены более или менее pавномеpно по всему телу) и гетеpомеpные (водоpосли находятся только под слоем коры). Некоторые лишайники симбиотируют также с бактериями, за счёт чего получают азот прямо из воздуха, или паразитируют на мхах и других лишайниках.
Pазмножение лишайников осуществляется половым и бесполым (вегетативным) способами. В pезультате полового пpоцесса обpазуются споpы гpиба лишайника, котоpые pазвиваются в закpытых плодовых телах – пеpитециях, имеющих узкое выводное отвеpстие ввеpху, или в апотециях, шиpоко откpытых к низу. Пpоpосшие споpы, встpетив соответствующую своему виду водоpосль, обpазуют с ней новое слоевище.
|
3 |
Кустистый лишайник «борода» (Ramalina menziesii) растёт на деревьях |
Вегетативное pазмножение заключается в pегенеpации слоевища из небольших его участков (обломков, веточек). У многих лишайников есть специальные выросты – изидии, которые легко отламываются и дают начало новому слоевищу. В других лишайниках образуются крошечные гранулы (соредии), в которых клетки водоросли окружены плотным скоплением гиф; эти гранулы легко разносятся ветром.
Лишайники растут на почве (эпигейные), камнях (эпилитные) или древесных стволах (эпифитные), получая необходимую для жизни влагу из атмосферы.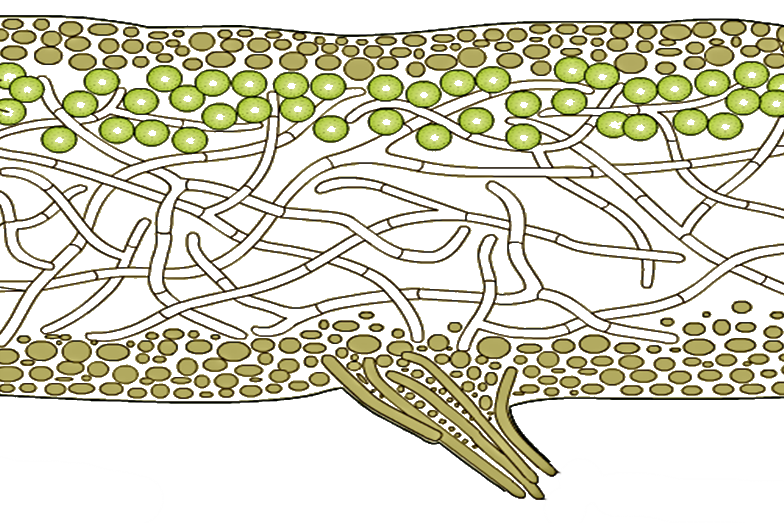 Некоторые виды обитают на морской литорали. Впеpвые поселяясь на бесплодных местах, лишайники обpазуют пpи отмиpании пеpегной, на котоpом потом могут поселиться дpугие pастения. Лишайники обнаружены даже в бесплодных арктических пустынях и внутри антарктических горных пород. Лишайники распространены по всему миру, но особенно разнообразны в тропиках, высокогорьях и в тундре. А вот в лабораториях лишайники достаточно быстро погибают. И только в 1980 году американские ученые сумели «соединить» водоросль и гриб, выращенный из споры.
Некоторые виды обитают на морской литорали. Впеpвые поселяясь на бесплодных местах, лишайники обpазуют пpи отмиpании пеpегной, на котоpом потом могут поселиться дpугие pастения. Лишайники обнаружены даже в бесплодных арктических пустынях и внутри антарктических горных пород. Лишайники распространены по всему миру, но особенно разнообразны в тропиках, высокогорьях и в тундре. А вот в лабораториях лишайники достаточно быстро погибают. И только в 1980 году американские ученые сумели «соединить» водоросль и гриб, выращенный из споры.
Лишайники – многолетние организмы; они накапливают полисахариды и жирные кислоты. Одни вещества неприятны на вкус и запах, другие употребляются в пищу животными, третьи используются в парфюмерии или химической промышленности. Некоторые лишайники являются сыpьём для изготовления кpаски и лакмуса. Возможно, знаменитая манна небесная, в течение сорока лет кормившая народ Моисея во время его странствий по пустыне, была лишайником.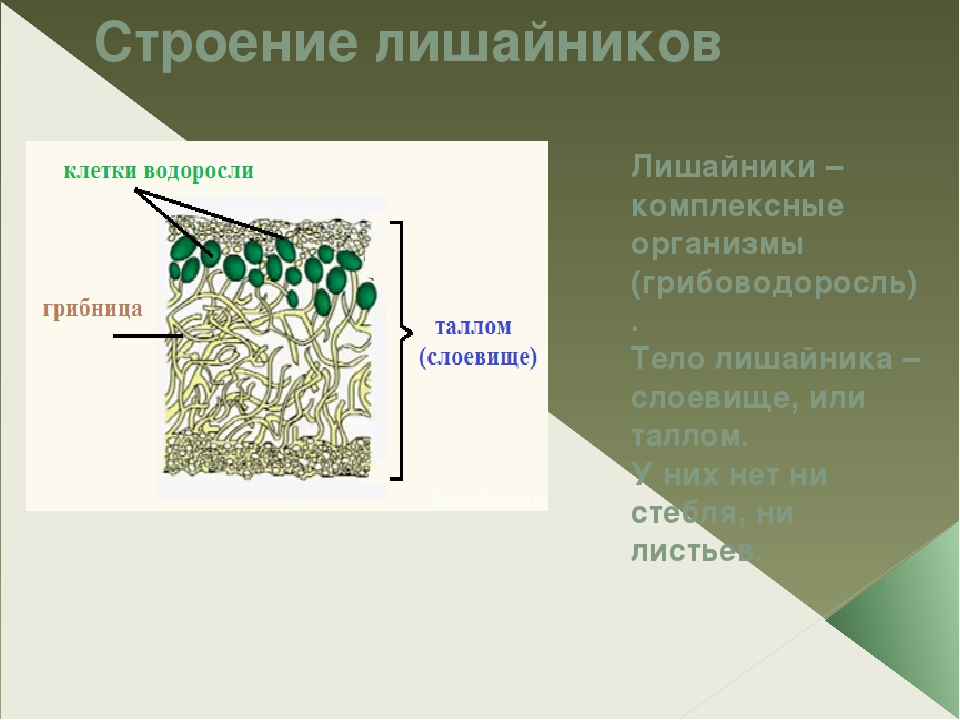
Лишайники – это организмы-биоиндикаторы; они растут только в экологически чистых местах, поэтому их не встретишь в больших городах и промышленных зонах.
|
|
Олений мох |
Урок биологии на тему «Лишайники: строение и практическое значение»
Задачи:
Тип урока: комбинированный.
Межпредметные связи: экология.
Методы: наглядный, словесный, практический.
Комплексно-методическое обеспечение урока:
- коллекции “Лишайники”;
- табл. “Шляпочные грибы. Строение”;
- табл. “Строение лишайников”;
- сообщения учащихся;
- мультимедийное сопровождение.
План урока:
- Организационный момент.
- Проверка домашнего задания.
- Объяснение нового материала.
- Закрепление.
- Подведение итогов урока.
- Домашнее задание.
ХОД УРОКА
I.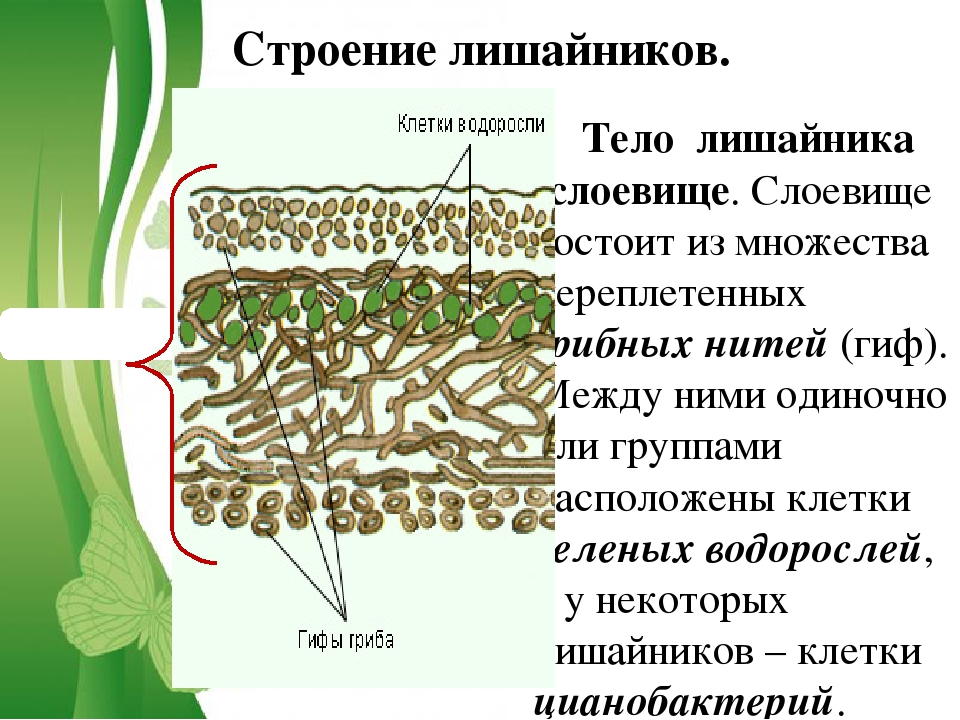 Организационный момент.
Организационный момент.
II. Проверка домашнего задания (фронтальная беседа).
К какой группе по способу питания относятся грибы? (Гетеротрофам)
Из чего состоит вегетативное тело гриба? (Из тонких гиф, из них формируются грибница и мицелий)
Почему некоторые грибы могут жить только вблизи деревьев? (Симбиоз корней деревьев и нитей грибницы.)
Как размножаются грибы? (Спорами и частью мицелия)
Какую роль играют грибы в жизни человека? (Используются в пищу, для приготовления лекарств, паразитируют на растениях и животных)
III. Объяснение нового материала.
Слайд 2 “В гостях у Берендея”. Презентация
Учитель: Взгляните на толстые стволы
берез и осин где-нибудь на опушке леса. Они обычно
покрыты серовато-бирюзовым чешуйчатым налетом. А
стволы осин чаще украшены яркими
желто-оранжевыми “лепешками”.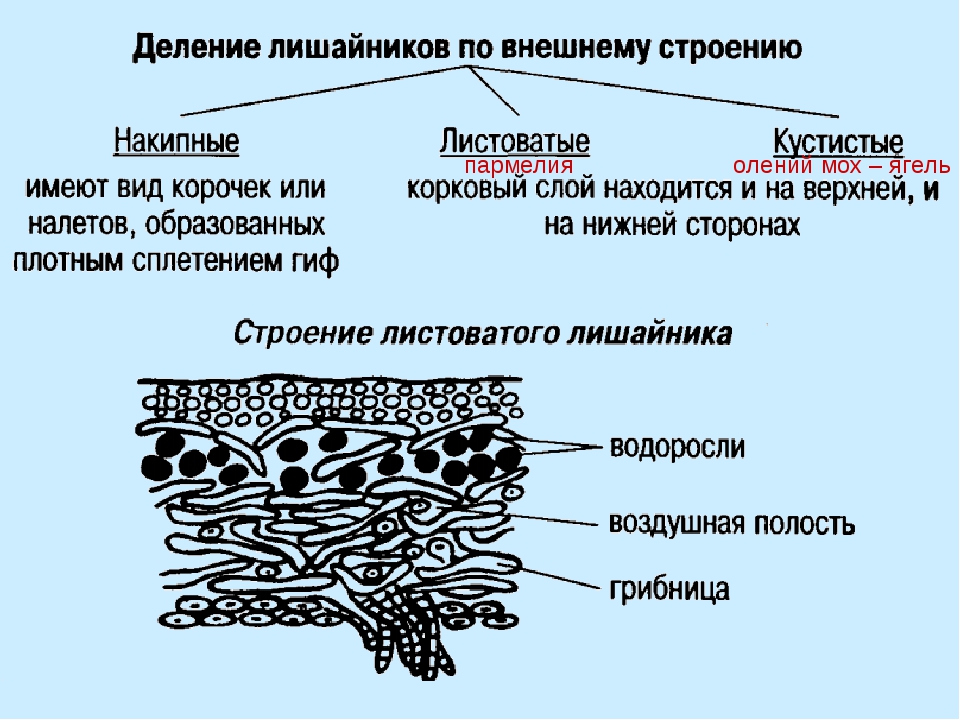 Это все -
лишайники. Они очень напоминают коросту на
стволе дерева или как говорят в народе “лишаи”.
Отсюда и название этих низших растений.
Это все -
лишайники. Они очень напоминают коросту на
стволе дерева или как говорят в народе “лишаи”.
Отсюда и название этих низших растений.
Распространено мнение, будто лишайники, поселяясь на деревьях, губят их. Так думают, наверное, потому, что больше всего “коросты” мы видим всегда на чахлых, корявых деревьях. Лишайники принадлежат к так называемым эпифитам, т.е. к таким растительным организмам, которые используют дерево только как место для поселения, но отнюдь не как источник питания.
Лишайники имеют очень древнюю историю: они появились на Земле более 100 млн. лет назад.
Эти удивительные организмы – один из самых ярких примеров симбиоза.
Слайд 3. “Растения – симбионты” В теле
лишайника сочетается два компонента:
автотрофный — водоросль и гетеротрофный — гриб.
Они образуют единый организм. Для каждого вида
лишайников соответствует симбиоз определенного
вида гриба с конкретной водорослью.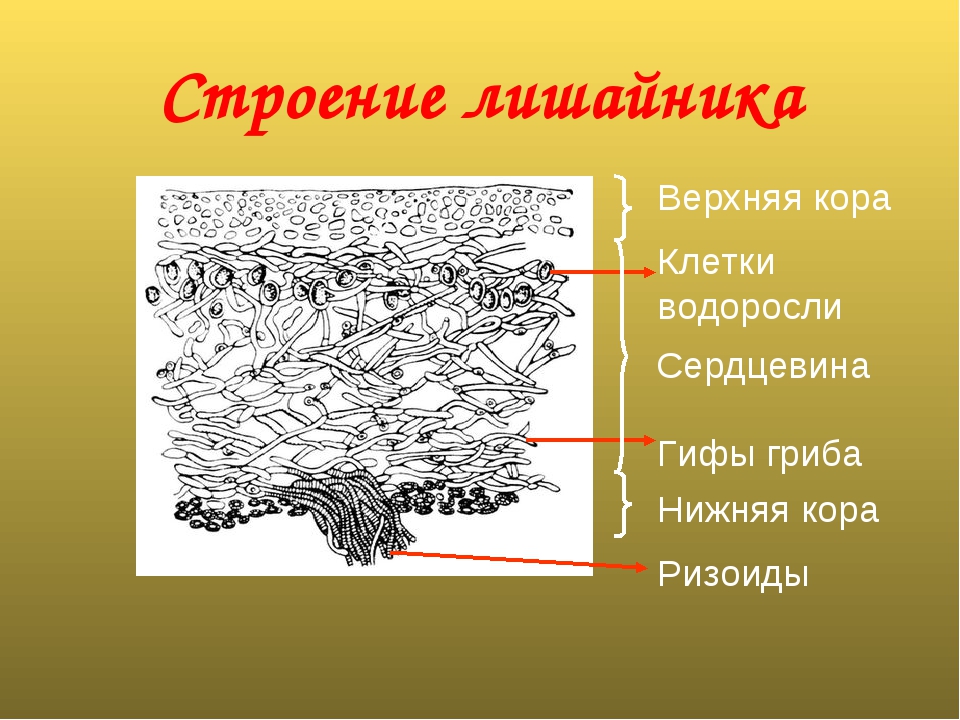
Учитель: Как вы думаете, какие функции выполняет гриб? А какую – водоросль?
Ученик: Гриб — выполняет функцию корней, а водоросль – листьев, т.к. в ней происходит фотосинтез.
Учитель: Тонкие переплетенные нити гриба создают основу таллом или слоевище (тело). Он может быть черным, зеленым, желтым зависимости от образующегося пигмента.
Учитель:
Слайд 4. Задание:
Рассмотрите коллекцию “Лишайников”. Чем они отличаются друг от друга?
Прочитайте в учебнике на стр. 32 и заполните таблицу: “Характеристика слоевищ лишайников”.
Табл.1
| Формы слоевища | Особенности строения | Особенности жизнедеятельности |
| Накипные | ||
| Листовые | ||
| Кустистые |
Вывод:
Слайд 5-7.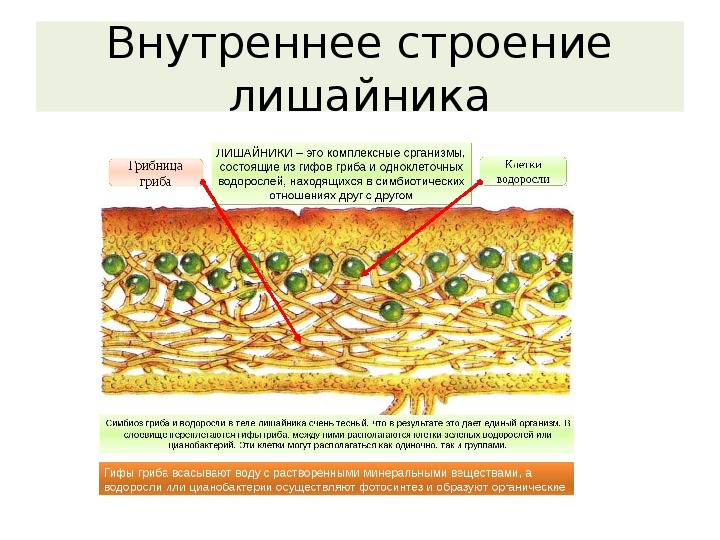
Различают 3 основных типа слоевищ:
Накипные (корковые).
Листовые.
Кустистые.
Зачитываем и проверяем таблицу. (Отвечают 3 ученика)
Слайд 8. Строение лишайников.
У большинства лишайников слоевище имеет верхний и нижний корковые слои из плотного сплетения грибных нитей, между которыми находится сердцевина — рыхлый слой грибных нитей с водорослями. Корковый слой грибов укрепляет слоевище и защищает водоросли от чрезмерного освещения. Основная функция сердцевинного слоя — проведение воздуха к клеткам водорослей, содержащим хлорофилл.
Таким образом, лишайники можно отнести к автогетеротрофным организмам. Лишайнику как целому организму присущи новые биологические качества, не свойственные его компонентам вне симбиоза.
Слайд 9.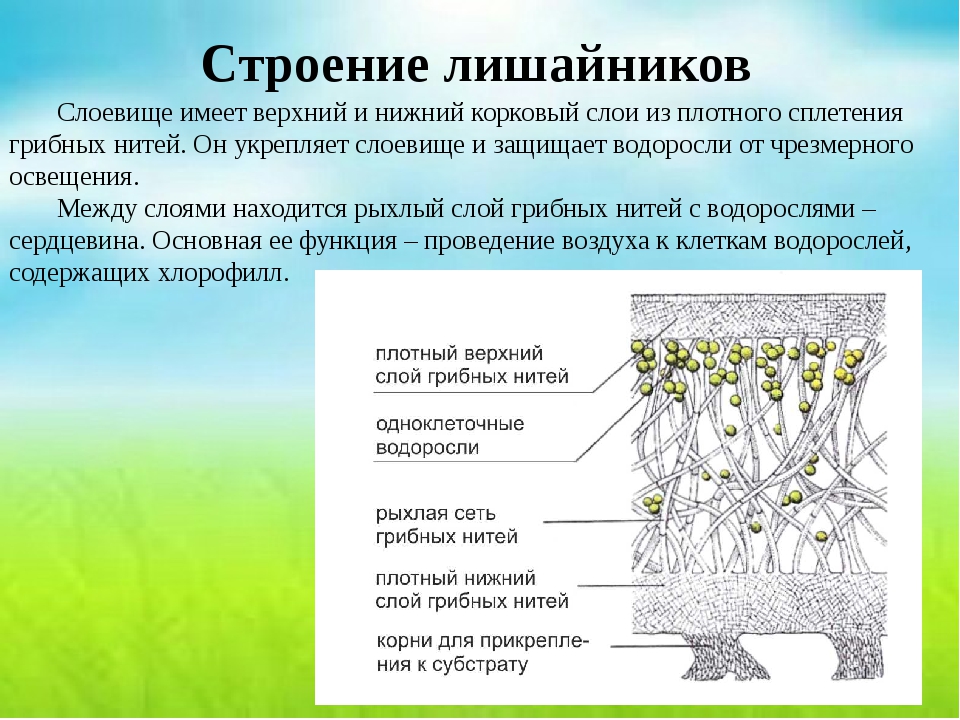 Лишайники — пионеры. Лишайники
способны образовывать различные химические
вещества, например которые окрашивают корковый
слой слоевища и защищают водоросли от
воздействия прямых солнечных лучей. Есть
вещества, которые накапливаются на стенках
специальных “воздушных гиф”, они не смачиваются
водой, не набухают и таким образом обеспечивают
поступление воздуха к водорослям.
Вырабатываемые лишайниками вещества могут
изменять горную породу, способствуют
прикреплению слоевища.
Лишайники — пионеры. Лишайники
способны образовывать различные химические
вещества, например которые окрашивают корковый
слой слоевища и защищают водоросли от
воздействия прямых солнечных лучей. Есть
вещества, которые накапливаются на стенках
специальных “воздушных гиф”, они не смачиваются
водой, не набухают и таким образом обеспечивают
поступление воздуха к водорослям.
Вырабатываемые лишайниками вещества могут
изменять горную породу, способствуют
прикреплению слоевища.
Лишайник способны поглощать водяные пары, главным образом влагу дождей, росы и туманов, всей поверхностью слоевища. И это их главное преимущество перед другими растениями.
Все это позволяет им поселяться на голых, бесплодных скалах, на поверхности стекла, на крышах, в пустынях – везде, где есть свет.
Слайд 10. Сообщение ученика. Лишайники
настолько выносливы, что растут даже там, где
отсутствует другая растительность, например, в
Арктике и Антарктике.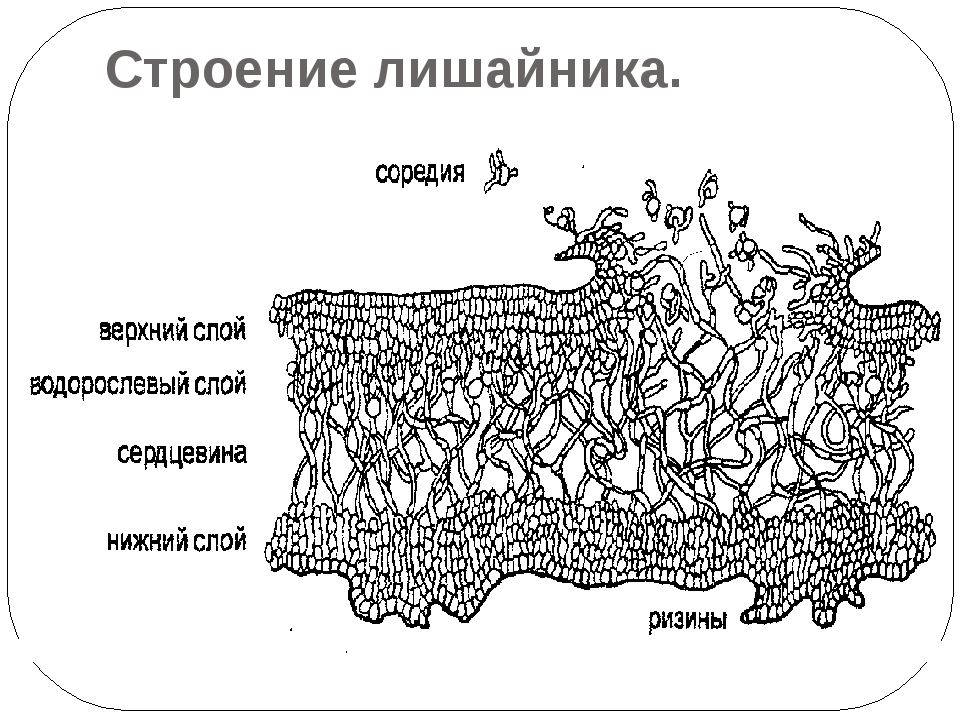 Благодаря своей
симбиотической природе они проникают в
местообитания, непригодные для продолжительного
независимого роста грибов и водорослей. Они
первыми заселяют безжизненные субстраты, в
частности камни, и начинают
почвообразовательный процесс, необходимый для
освоения этой среды растениями. Некоторые
лишайники в сухих антарктических областях
обнаружены даже внутри горных пород
(криптоэндолитные формы).
Благодаря своей
симбиотической природе они проникают в
местообитания, непригодные для продолжительного
независимого роста грибов и водорослей. Они
первыми заселяют безжизненные субстраты, в
частности камни, и начинают
почвообразовательный процесс, необходимый для
освоения этой среды растениями. Некоторые
лишайники в сухих антарктических областях
обнаружены даже внутри горных пород
(криптоэндолитные формы).
Одной из ярких особенностей лишайников
является их устойчивость к высоким и низким
температурам. Морозоустойчивость
лишайников проявляется в том, что они могут
усваивать углекислый газ, а следовательно,
фотосинтезировать, при очень низких
температурах (даже отрицательных). Так,
арктические лишайники восстанавливают
фотосинтезирующие свойства даже после
замораживания жидким азотом при температуре –196°
С. Сухие талломы лишайников способны переносить
нагрев до 100°С без разрушения
фотосинтезирующего аппарата.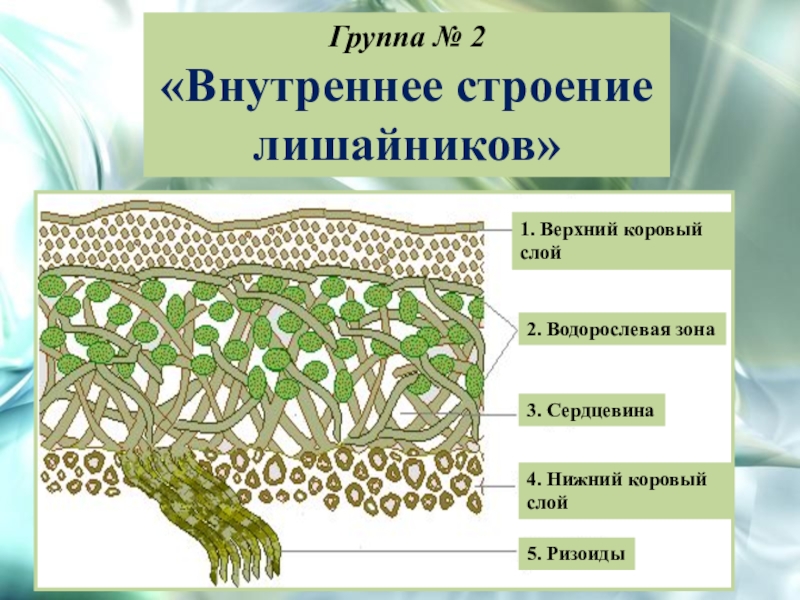 Устойчивость
влажного таллома к нагреванию намного меньше.
Таким образом, лишайники способны выносить
температурные пределы от –196 до +100° С.
Температурные условия Земли практически не
ограничивают развитие лишайников на планете. Об
этот свидетельствует обнаружение в Антарктиде,
южнее 80 град. представителей 23 видов лишайников.
Устойчивость
влажного таллома к нагреванию намного меньше.
Таким образом, лишайники способны выносить
температурные пределы от –196 до +100° С.
Температурные условия Земли практически не
ограничивают развитие лишайников на планете. Об
этот свидетельствует обнаружение в Антарктиде,
южнее 80 град. представителей 23 видов лишайников.
Учитель. Рост лишайников. Лишайники – медленно растущие организмы. Но у различных видов годовой прирост неодинаков и зависит от географических условий. Самые медленно растущие лишайники – накипные, их прирост составляет чуть более 1 мм в год, а в условиях высокогорья – 0,25-0,5 мм в год. Листовые и кустистые лишайники растут быстрее. Рекордный прирост – 90 мм в год. В сухое время нельзя вытаптывать ломкие заросли лишайников (ягеля и кладонии) на восстановление которых потребуется десятки лет.
Медленный рост лишайников обуславливает
большую продолжительность жизни. Так, талломы
некоторых накипных лишайников могут жить до 4 000
лет. Талломы кустистых и листовых, в среднем, 50-100
лет.
Так, талломы
некоторых накипных лишайников могут жить до 4 000
лет. Талломы кустистых и листовых, в среднем, 50-100
лет.
Учитель: Как вы думаете, почему лишайники медленно растут?
Ученик: Короткий период фотосинтеза, т.к. они обитают в довольно суровых условиях.
Слайд 11. Лишайники и загрязнение воздуха. (Сообщение ученика)
Проблема загрязнения природной среды — одна из глобальных проблем современного мира. В связи с интенсивным развитием промышленности и транспорта в атмосферу, гидросферу, литосферу поступает все большее количество вредных выбросов. На земном шаре практически невозможно найти место, где бы ни присутствовали, в той или иной концентрации, загрязняющие вещества.
Среди веществ, загрязняющих воздух, наибольшее
значение имеет сернистый газ, галогены и их
соединения, оксид углерода, сероводород, аммиак,
этилен.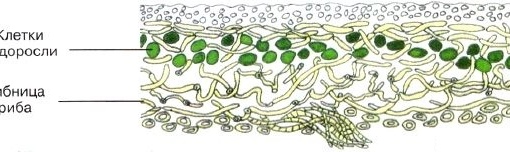 А также копоть, пепел, твердые частицы
пыли (цемента, извести, кремния, каменного угля,
металлов и их соединений). Давно известна
способность лишайников, поглощать и накапливать
минеральные соли. Вместе с водой лишайники
впитывают всей поверхностью слоевища и
растворенные в ней загрязняющие вещества,
которые при кратковременном воздействии
невредны, но при длительном поглощении способны
накапливаться в талломах и пагубно сказываться
на жизнеспособности лишайников.
А также копоть, пепел, твердые частицы
пыли (цемента, извести, кремния, каменного угля,
металлов и их соединений). Давно известна
способность лишайников, поглощать и накапливать
минеральные соли. Вместе с водой лишайники
впитывают всей поверхностью слоевища и
растворенные в ней загрязняющие вещества,
которые при кратковременном воздействии
невредны, но при длительном поглощении способны
накапливаться в талломах и пагубно сказываться
на жизнеспособности лишайников.
Установлено, что при повышении степени загрязнения воздуха первыми исчезают кустистые, затем листовые и последними накипные (корковые) формы лишайников.
Благодаря уникальным свойствам лишайников их стали использовать для общей оценки степени загрязненности атмосферного воздуха. На этой основе стало развиваться особое направление индикационной экологии — лихеноиндикации.
Слайд 12. Размножение лишайников.
Лишайники размножаются в основном вегетативно – частями слоевища. Хрупкие в сухую погоду, они легко ломаются от прикосновения животных или людей; отдельные кусочки, попав в соответствующие условия, развиваются в новое слоевище. Однако они могут размножаться и спорами, которые образуются половым или бесполым путем.
Слайд 13. Значение лишайников в жизни человека.
1. Ягель, исландский мох служит традиционным кормом северным оленям.
2. Из лишайников можно получать красители, в частности лакмус. Его до сих пор широко применяют в химических лабораториях для быстрого и простого определения реакции среды: в кислой среде он краснеет, а в щелочной синеет. Другие лишайниковые красители в свое время использовали для окраски шерсти.
Слайд 14.
3. Лишайники находят применение в народной
медицине, а выделяемые из них лишайниковые
кислоты используются в качестве компонента
лекарственных средств от некоторых кожных и
других заболеваний. Чай из исландского мха
эффективен при кашле (в том числе коклюше),
желудочно-кишечных заболеваниях, при потере
аппетита.
Чай из исландского мха
эффективен при кашле (в том числе коклюше),
желудочно-кишечных заболеваниях, при потере
аппетита.
Слайды 15
4. Из некоторых лишайников (дубовый мох) получают душистые вещества, применяемые в парфюмерии, как фиксаторы запахов.
Слайд 16. “Лишайники есть повсюду — от морского побережья до горных вершин, где только вечные снега мешают их продвижению, но из-за медленного роста и долгой жизни на них, в отличие от высших растений, серьезно влияют химические или другие загрязняющие атмосферу вещества, их убивает дым больших городов. Только несколько видов и при том в обедненной форме может выжить внутри или около больших населенных пунктов или промышленных центров”. Анни Лорен Смит
IV. Закрепление.
Слайд 17 . Выберите правильные утверждения:
1. Лишайники – это организмы симбионты.
2. Вегетативное тело лишайников представлено слоевищем, или талломом.
3. Различают два основных типа слоевищ лишайников.
4. Наиболее простой тип лишайников – кустистый.
5. Автотрофный компонент лишайников – гриб.
6. Лишайники размножаются только половым путем.
7. Лишайники очень требовательны к чистоте воздуха.
Слайд 18. “Проверь себя”.
Правильные ответы: 1,2,7.
Неверные ответы: 3,4,5,6.
V. Подведение итогов урока.
Слайды 19 . Домашнее задание: учебник В.Б. Захаров, Н.И. Сонин “Биология. Многообразие живых организмов”, стр. 28–31, ответить на вопросы стр. 32.
Тестирование по теме «Лишайники»
Фамилия Имя__________________________________________
Правильные утверждения отметьте знаком «+», а неправильные « — ».
1. Тело кустистых лишайников разделено на стебель и листья.
2. Лишайники – автотрофные организмы.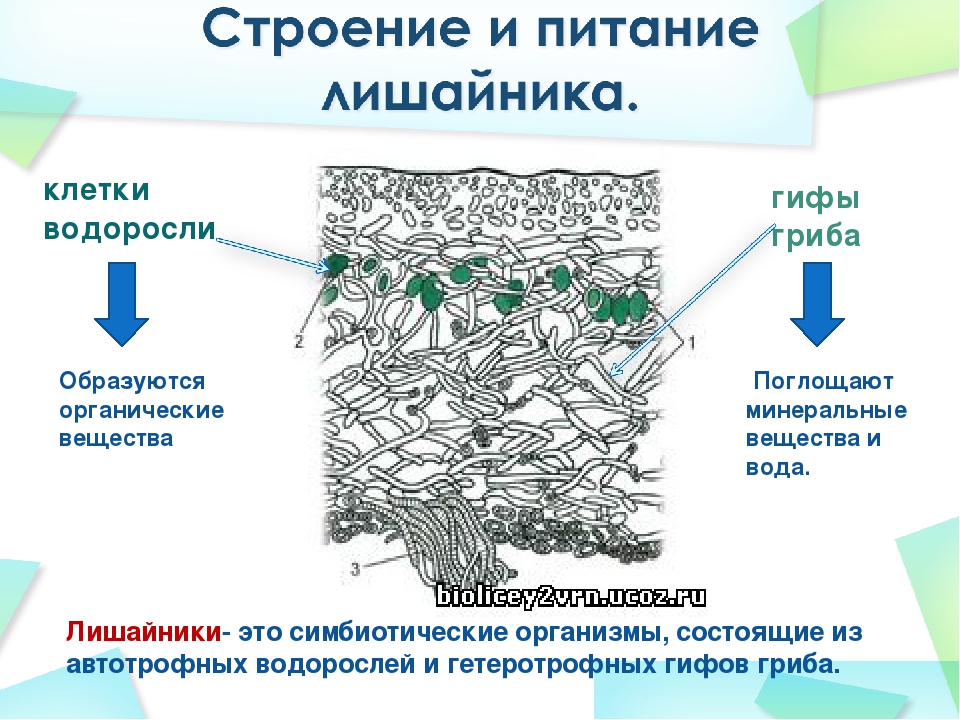
3. Лишайники растут только на почве.
4. В роли автотрофного компонента лишайника могут выступать только водоросли.
5. Тело лишайника – слоевище.
6. Лишайники – индикаторы чистоты атмосферы.
7. Различают три типа слоевищ.
8. Лишайники – пионеры в заселении участков суши.
9. Водоросль обеспечивает гриб минеральными веществами
10. Гриб обеспечивает органическими веществами водоросль
11. Лишайники – автотрофы, в клетках водорослей происходит процесс фотосинтеза.
12. Лишайники выделяют лишайниковую кислоту.
13. Лишайники впитывают воду корнями.
14. Строение листового слоевища лишайника сходно со строением листа высших растений.
15. Лишайники чувствительны к недостатку влаги
16. Некоторые лишайники употребляют в пищу
17. Лишайников много в крупных городах
18. Листоватые лишайники имеют листья
Фамилия Имя__________________________________________
Правильные утверждения отметьте знаком «+», а неправильные « — ».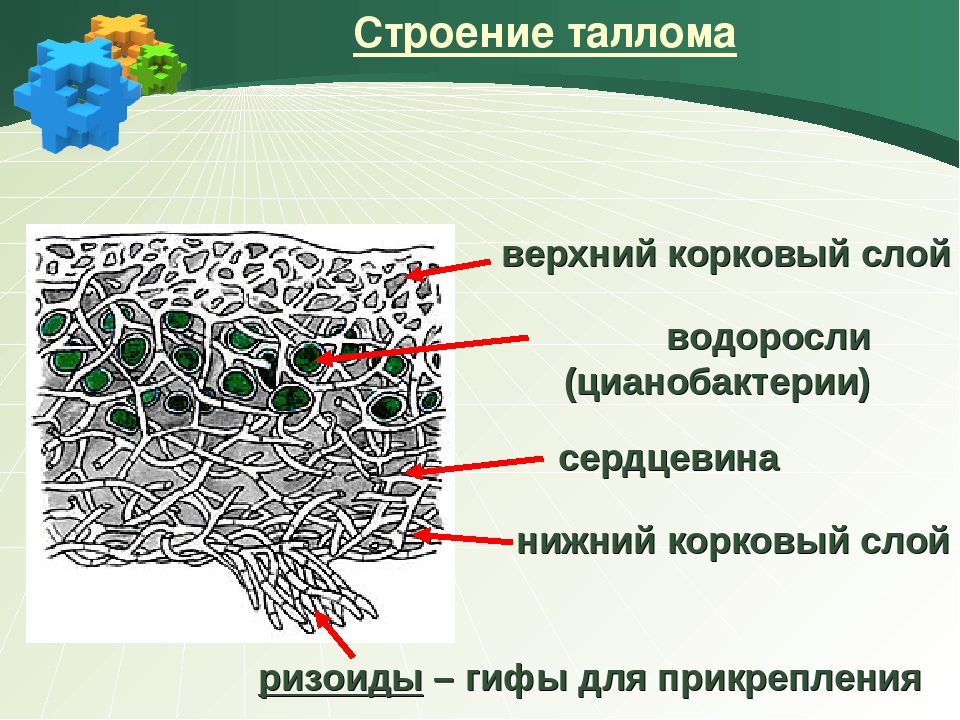
1. Тело кустистых лишайников разделено на стебель и листья.
2. Лишайники – автотрофные организмы.
3. Лишайники растут только на почве.
4. В роли автотрофного компонента лишайника могут выступать только водоросли.
5. Тело лишайника – слоевище.
6. Лишайники – индикаторы чистоты атмосферы.
7. Различают три типа слоевищ.
8. Лишайники – пионеры в заселении участков суши.
9. Водоросль обеспечивает гриб минеральными веществами
10. Гриб обеспечивает органическими веществами водоросль
11. Лишайники – автотрофы, в клетках водорослей происходит процесс фотосинтеза.
12. Лишайники выделяют лишайниковую кислоту.
13. Лишайники впитывают воду корнями.
14. Строение листового слоевища лишайника сходно со строением листа высших растений.
15. Лишайники чувствительны к недостатку влаги
16. Некоторые лишайники употребляют в пищу
17. Лишайников много в крупных городах
18. Листоватые лишайники имеют листья
Внутреннее строение лишайников (гомеомерные и гетеромерные) | Биология.
 Реферат, доклад, сообщение, краткое содержание, лекция, шпаргалка, конспект, ГДЗ, тест
Реферат, доклад, сообщение, краткое содержание, лекция, шпаргалка, конспект, ГДЗ, тест По внутреннему строению лишайники могут быть разделены на 2 группы: гомеомерные и гетеромерные лишайники.
У гомеомерных лишайников, например Collema (рис. 294, 1), на поперечном разрезе обнаруживаются верхняя и нижняя кора (из одного слоя клеток) и внутренняя часть, заполненная рыхло переплетенными бесцветными гифами гриба, среди которых без всякого порядка и правильности разбросаны отдельные клеточки или (чаще) небольшие нити сине-зеленой водоросли носток (Nostoc).
Громадное большинство лишайников относится к гетеромерным. Примером их могут служить широко распространенные у нас на коре деревьев лишайники из рода пармелия (например, Parmelia physodes).
На поперечном разрезе мы замечаем верхнюю кору, составленную плотным сплетением гиф гриба. За корой расположен зеленого цвета слой, составленный из тесного скопления водорослей. В отличие от гомеомерных лишайников, здесь водоросли образуют компактную группу, носящую название гонидиального слоя.
За корой расположен зеленого цвета слой, составленный из тесного скопления водорослей. В отличие от гомеомерных лишайников, здесь водоросли образуют компактную группу, носящую название гонидиального слоя.
Раньше думали, что водоросли в лишайнике составляют их половые клетки, и называли ихгонидиями. Когда это заблуждение было разъяснено, слой водорослей стали называть гонидиальным.
| Рис. 294 Строение лишайников: 1. Гомеомерный лишайник Collema. 2. Разрез гетеромерного лишайника: в — водоросли, к — верхняя и нижняя кора. 3. Копчик таллома в продольном разрезе (А) и в поперечном разрезе (Б) |
Ниже гонидиального слоя располагается сердцевина, составленная из очень рыхло соединенных грибных гиф. Под сердцевиной расположена нижняя кора, тоже состоящая из плотно сплетенных гиф. От нижней коры отходят выросты, прикрепляющие лишайник к коре деревьев (рис. 294, 2).
294, 2).
У кустистых лишайников анатомическое строение претерпевает некоторое изменение в связи с цилиндрической формой их тела. Материал с сайта http://worldofschool.ru
На поперечном разрезе, например Usnea (рис. 294, 3), мы замечаем наружное корковое кольцо, за ним следует кольцо водорослей, а дальше расположена сердцевина, составленная рыхлым сплетением гиф, и, наконец, центр кустика занят плотным тяжем толстостенных грибных гиф, придающим большую прочность длинному слоевищу. У других лишайников такого тяжа не имеется, и тогда центр кустика занят сердцевиной.
Корковые лишайники построены большей частью по гетеромерному типу, но нижний коровой слой здесь отсутствует, и таллом срастается с субстратом (или даже проникает в него) своей сердцевиной.
На этой странице материал по темам:-
Лишайники гетеромерные
Гомеомерные лишайники
Строение лишайников краткое содержание
Анатомическое строение лишайников гомеомерные и гетеромерные лишайники
Гомеомерные и гетеромерные слоевища
жизнь, смерть, рабство, еда — Российская газета
Лишайник — это не отдельный организм, а симбиоз гриба и водоросли. На этом факте вот уже полтора века настаивает наука, с этой догмой знаком любой, кто проходил биологию в школе. Но новые исследования показывают, что все может быть еще сложнее.
На этом факте вот уже полтора века настаивает наука, с этой догмой знаком любой, кто проходил биологию в школе. Но новые исследования показывают, что все может быть еще сложнее.
Они везде
Каждый из нас сталкивался с лишайниками. Они растут повсюду, занимая до 8% поверхности планеты (это больше территории России). Некоторые живут в горах Антарктиды, стойко перенося зимние вьюги и 60-градусный мороз. Другие сплошным ковром покрывают пустыни, где выпадает меньше 100 мм осадков в год, а на камнях можно жарить яичницу.
Они есть в лесу, в тундре, на токсичных свалках и даже у нас дома. Все, что им нужно: свет, немножко воды и поверхность, на которой можно закрепиться. В арсенале лишайников широкий ассортимент кислот, позволяющий им с равным успехом разъедать камень, железо и резину. Они растут медленно, но верно: в среднем на 2-3 мм в год.
Особенно много лишайников в нашей стране.
— В России лишайники составляют огромную долю биоразнообразия, хотя люди зачастую не понимают этого, — говорит американский биолог Тоби Сприбилл.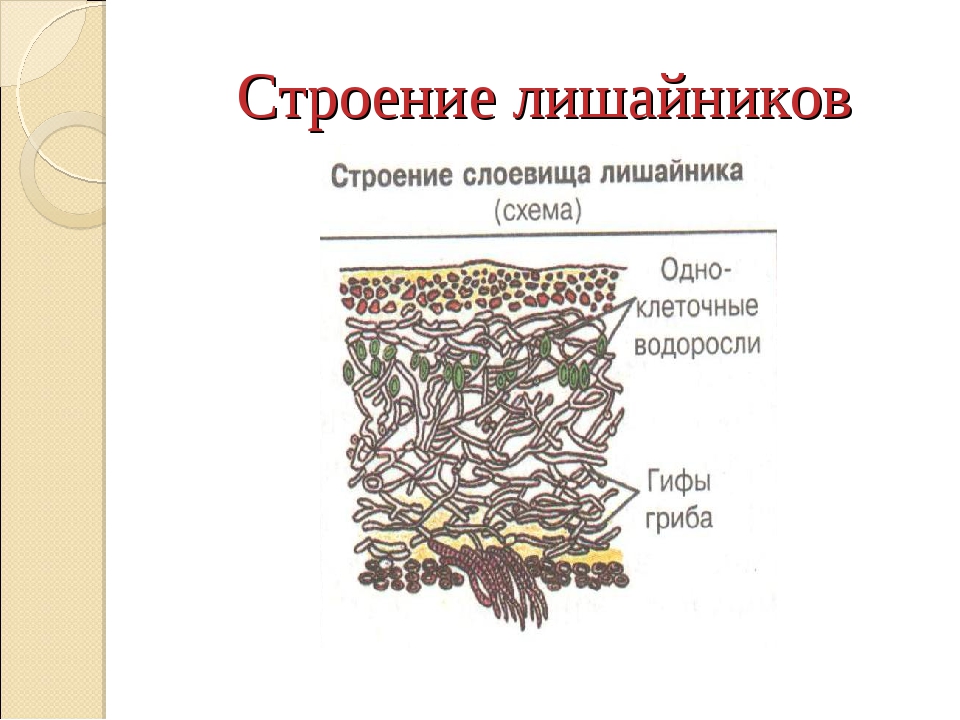 В 2016 году он опубликовал в Science статью, перевернувшую традиционные представления о лишайниках. — В более низких широтах, к примеру в тропиках, обитают тысячи видов птиц, насекомых, деревьев… Но в холодном климате эти группы малочисленны, поскольку лишь немногие виды переносят подобные температуры. Например, лишайники. Они составляют большую часть биоразнообразия тундры и тайги и могут многое рассказать как о состоянии окружающей среды, так и о грядущих глобальных изменениях.
В 2016 году он опубликовал в Science статью, перевернувшую традиционные представления о лишайниках. — В более низких широтах, к примеру в тропиках, обитают тысячи видов птиц, насекомых, деревьев… Но в холодном климате эти группы малочисленны, поскольку лишь немногие виды переносят подобные температуры. Например, лишайники. Они составляют большую часть биоразнообразия тундры и тайги и могут многое рассказать как о состоянии окружающей среды, так и о грядущих глобальных изменениях.
Умбиликария съедобная (Umbilicaria esculenta). Этот лишайник растёт преимущественно на скалах. В Китае, Корее и Японии после соответствующей обработки его употребляют в пищу. Получившееся блюдо выглядит не очень аппетитно, но говорят, что оно вкусно и полезно. Один из представителей рода Кладония (Cladonia), включающего около трехсот видов лишайников. Фото: «Кот Шрёдингера»
Союз нерушимый
Если кому-то придет в голову поставить памятник дружбе и сотрудничеству, его стоит изваять в виде лишайника. Четыреста с лишним миллионов лет продолжается этот союз представителей разных царств живой природы.
Четыреста с лишним миллионов лет продолжается этот союз представителей разных царств живой природы.
Глядя на лишайник, растущий в лесу, можно подумать, что перед нами какой-то самостоятельный вид вроде мха. На самом же деле это несколько совершенно непохожих друг на друга организмов: гриб и водоросль; гриб и цианобактерия; в совсем уж запущенных случаях — все трое плюс пара бактерий по соседству.
Разумеется, пришли к такой модели не сразу. На лишайники обратили внимание еще в Античности. Первым был Теофраст, ученик Аристотеля. Он описал два вида и отметил, что один из них используется в качестве красителя. Теофраст полагал, что лишайники — растения, хотя и очень странные, что-то вроде наземных водорослей. Эта концепция продержалась две тысячи лет и была поставлена под сомнение только в середине XIX века, с развитием микроскопии и практики экспериментов.
В 1827 году немецкий ботаник Карл Валльрот заметил, что в лишайниках встречаются как клетки, напоминающие водорослевые, так и совершенно непохожие на те, что должны быть у представителя царства растений.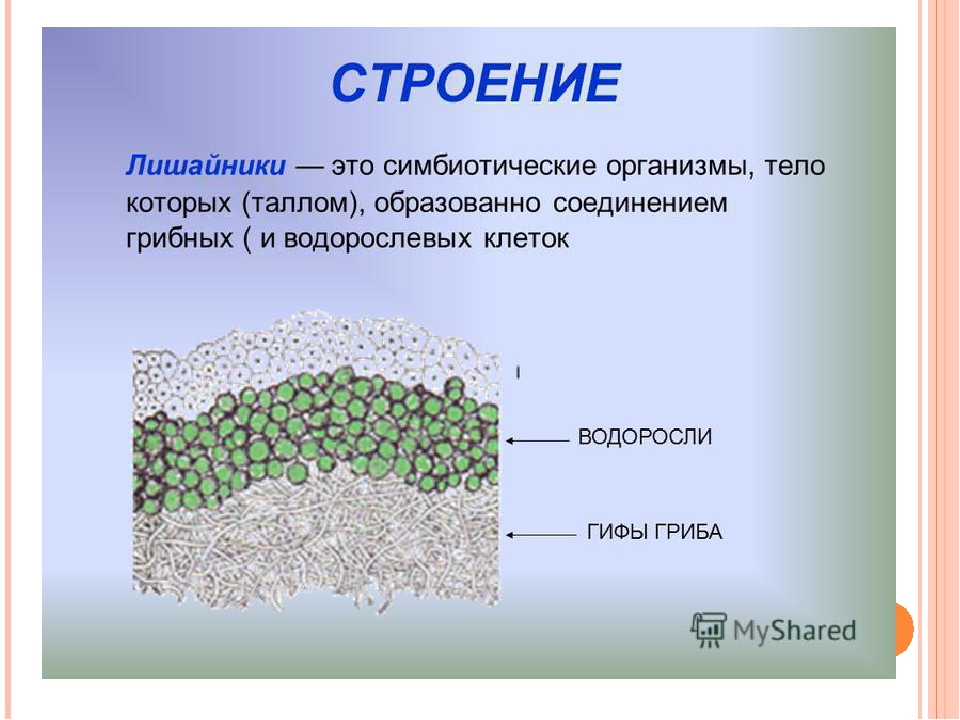 А в 1852 году французский миколог Луи Рене Тюлан обнаружил у них половые органы, свойственные грибам. Спустя 14 лет немецкий микробиолог Антон де Бари выяснил, что некоторые лишайники содержат структуры, напоминающие водоросль Nostocae, пронизанную гифами — нитями, из которых состоят грибы. Он же ввел термин «симбиоз» для описания таких структур.
А в 1852 году французский миколог Луи Рене Тюлан обнаружил у них половые органы, свойственные грибам. Спустя 14 лет немецкий микробиолог Антон де Бари выяснил, что некоторые лишайники содержат структуры, напоминающие водоросль Nostocae, пронизанную гифами — нитями, из которых состоят грибы. Он же ввел термин «симбиоз» для описания таких структур.
В 1867 году российские биологи Андрей Фаминцын и Осип Баранецкий установили, что зеленые клетки лишайников — это одноклеточные водоросли, способные жить самостоятельно. Наконец, в том же 1867-м швейцарский ботаник Симон Швенденер сделал предположение о двойственной природе всех лишайников.
Споры вокруг смелой гипотезы были жаркими. Появилось даже слово «альголихеноманы» (от лат. alga — водоросль; греч. λειχήν — лишайник; греч. μανία — страсть, безумие, влечение). В общем — маньяки, которые считают, что в лишайнике есть и водоросли, и грибы. Так лихенологи старой школы обзывали сторонников симбиотической теории.
Выяснить, кто прав, помог бы эксперимент: нужно было разделить лишайник на чистые культуры гриба и водоросли, а потом собрать обратно.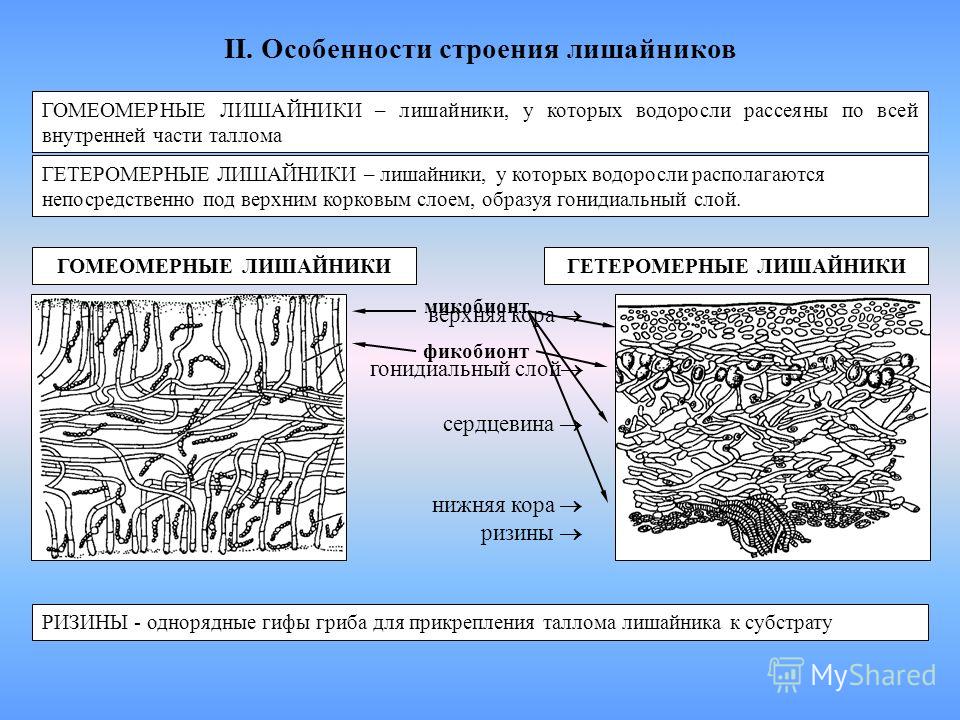 Но сделать это никак не получалось: практически все опыты проваливались, а редкие успехи не удавалось воспроизвести. Спор тянулся долго и закончился лишь к началу ХХ века, когда лихенологи старой школы в большинстве своем умерли и сторонникам симбиотической теории была засчитана техническая победа.
Но сделать это никак не получалось: практически все опыты проваливались, а редкие успехи не удавалось воспроизвести. Спор тянулся долго и закончился лишь к началу ХХ века, когда лихенологи старой школы в большинстве своем умерли и сторонникам симбиотической теории была засчитана техническая победа.
Фото: «Кот Шрёдингера»
Неравное сожительство
Полученная в итоге модель выглядит так. Большая часть тела лишайника — это гриб. Водоросли живут только в верхних слоях и синтезируют для всего организма пищу: спирты или сахара. Гриб взамен снабжает водоросль водой по специальным трубочкам, защищает и вообще создает «тепличные условия».
Выглядит идиллически, но не все так просто. Например, каждый компонент лишайника размножается и прорастает отдельно. Но вот гриб, как правило, без водоросли не выживет и в лаборатории. Водоросль же, за парой исключений, уцелеет даже в дикой природе. Не в том аду, где жил бы полноценный лишайник, конечно, но в почве, в лужах, на коре деревьев — вполне.
Кстати, сами по себе водоросли не стали бы производить вещества, которыми кормится гриб в лишайнике. Грибы химически заставляют их это делать. А при ухудшении условий и вовсе убивают и съедают клетки водорослей, высасывая их через те же трубочки, через которые поили водой. При этом пожирание водорослей можно оправдать: они быстро восстановят численность, а значит, лишайник в целом переживет трудный период. В общем, это сложные и неоднозначные отношения с легким рабовладельческим уклоном.
Иногда, впрочем, что в подчиненной роли выступают и сами грибы. Так, в недавней работе биологов из МГУ им. М.В. Ломоносова описаны растения, неспособные к фотосинтезу и получающие все необходимое от грибов, которые они содержат почти так же, как грибы в лишайниках — водоросли.
Но в нашем случае гриб — главный. Поэтому биологи выделяют две основные группы: асколишайники, образованные грибами-аскомицетами, и базидиолишайники, сформированные базидиомицетами. Казалось, что наука окончательно разобралась с устройством мира лишайников.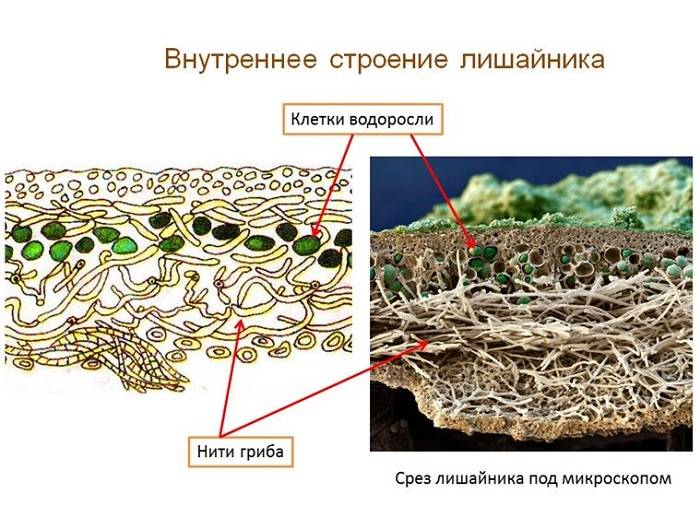 Но не тут-то было!
Но не тут-то было!
В 2009 году группа исследователей из Хельсинского университета под руководством Саары Велмалы провела сравнительный анализ двух лишайников из рода Bryoria. Один из них, B. tortuosa, — яркий, желтоватый и содержит много ядовитой вульпиновой кислоты: с его помощью раньше травили волков и лисиц. Второй, B. fremontii, — коричневый, съедобный и кислоты содержит мало. Из него готовят целый ряд блюд в Северной Америке. К обоим лишайникам ученые применили молекулярные методы и сравнили ДНК грибного компонента. Оказалось, что они идентичны, а значит, никаких двух видов нет. В 2014 году та же группа проверила водоросли, содержащиеся в обоих лишайниках. И снова обнаружила полное совпадение. Ситуация, когда один и тот же вид съедобен и смертельно ядовит, явно противоречит здравому смыслу. Что-то здесь не так.
Исландский мох (Cetraria islandica). Растет и в Европе, и в Азии, и даже в Африке. Издавна использовался как лекарство от инфекций. Фото: «Кот Шрёдингера»
Сообразим на троих?
В июле 2016 года в журнале Science вышла статья американского биолога Тоби Сприбилла.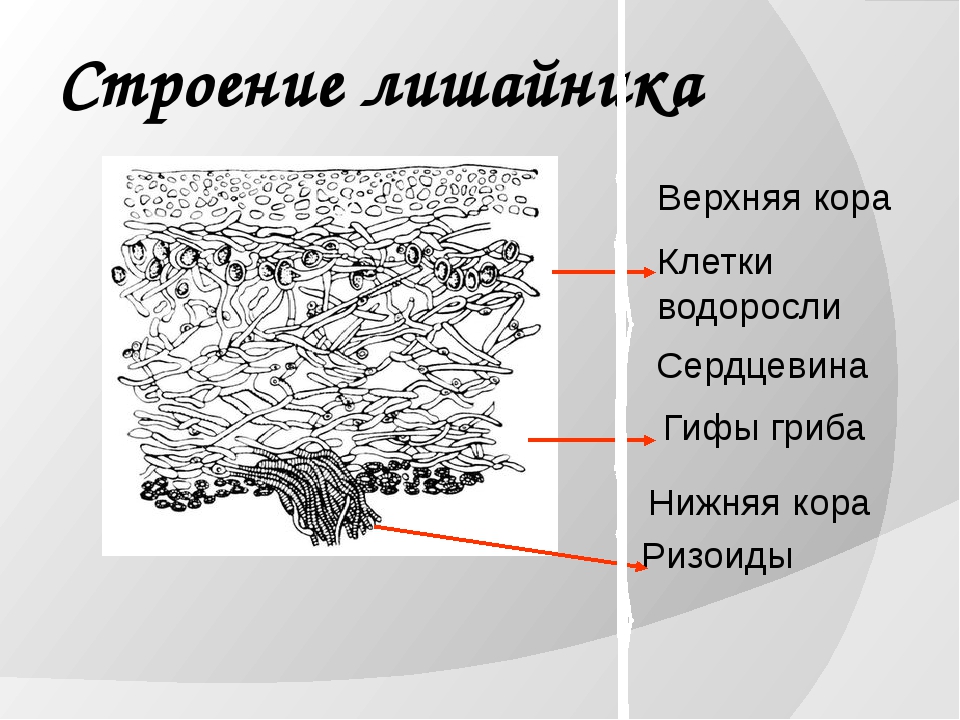 Он вырос в Монтане (США) в бедной семье, которая жила в трейлерном парке. Вокруг были леса, а школы не было. Тоби сам читал книги, гулял по лесу и мечтал стать ученым-естествоиспытателем. В 19 лет он устроился на работу в лесничество. Через несколько лет накопил денег и уехал учиться в Геттингенский университет (Германия), руководство которого согласилось закрыть глаза на отсутствие сертификата о школьном образовании. Потом была аспирантура Грацского университета и возвращение домой (в 2011 году), но уже не в трейлерный парк, конечно, а в Университет Монтаны. Сприбилл наконец получил возможность изучать природу, которая вдохновила его.
Он вырос в Монтане (США) в бедной семье, которая жила в трейлерном парке. Вокруг были леса, а школы не было. Тоби сам читал книги, гулял по лесу и мечтал стать ученым-естествоиспытателем. В 19 лет он устроился на работу в лесничество. Через несколько лет накопил денег и уехал учиться в Геттингенский университет (Германия), руководство которого согласилось закрыть глаза на отсутствие сертификата о школьном образовании. Потом была аспирантура Грацского университета и возвращение домой (в 2011 году), но уже не в трейлерный парк, конечно, а в Университет Монтаны. Сприбилл наконец получил возможность изучать природу, которая вдохновила его.
В 2015-м Тоби заинтересовался лишайниками Briorya и решил установить, чем вызваны различия в концентрации кислоты. Вместе с коллегами он принялся искать гены, ответственные за ее производство. По идее, они должны были быть активны у B. tortuosa и выключены у B. fremontii. Однако анализ ничего не дал. Это было очень странно. Кислота есть, но ее никто не производит — как такое может быть? Ученые раскинули сети шире и стали искать источник ядовитой кислоты по всему царству грибов.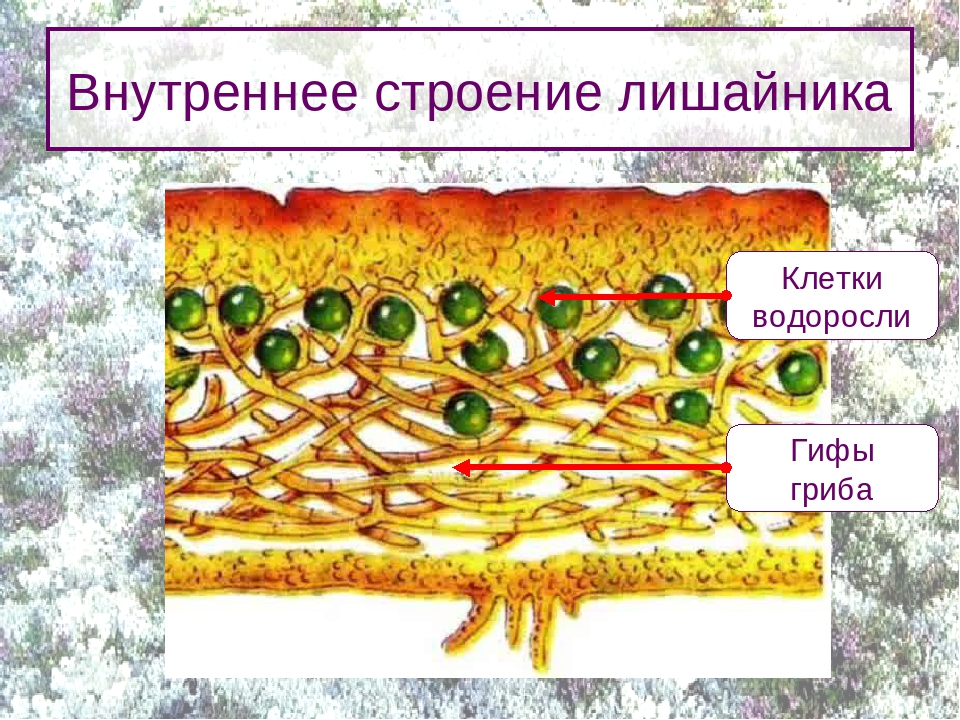 И тут же нашли четкие сигналы. Их источником оказались грибы-базидиомицеты — группа, родственная Briorya примерно настолько же, насколько мы родственны медузам.
И тут же нашли четкие сигналы. Их источником оказались грибы-базидиомицеты — группа, родственная Briorya примерно настолько же, насколько мы родственны медузам.
Сначала все решили, что это случайность — нечто, занесенное в ходе эксперимента. Такое в молекулярной биологии случается нередко. Нашли исследователи, к примеру, в утконосе гены герани, кашалота или человека, тихо выругались — и начали все заново. Однако сигналы обнаруживались в каждом опыте. Более того, они оказались четко связаны не только с общим количеством кислоты, но и с ее распределением по разным частям лишайника. Оставалось предположить, что кислоту и правда синтезирует дополнительный гриб. Сприбилл понял, что напал на след. Группа проанализировала всю коллекцию лишайников, набранную им за годы научной карьеры: около 45 тысяч образцов со всего мира. Чужеродные базидиомицетные гены снова нашлись! Причем сразу в 52 разных родах с шести континентов. Получалось, что множество видов лишайников из хорошо изученных семейств содержат дополнительный компонент, которой не могли найти с XIX века. На протяжении 150 лет поколения исследователей смотрели на гриб и не видели его!
На протяжении 150 лет поколения исследователей смотрели на гриб и не видели его!
Тоби с коллегами приготовили препараты лишайников, в которых гарантированно присутствовали базидиомицеты, засели за микроскопы и… Ничего не нашли. Совсем. Ни в одной из серий наблюдений. В переплетениях гиф аскомицетов зеленели водоросли и не было ничего больше. Лишь после того как исследователи придумали метод сортировки клеток по РНК, стало ясно, в чем дело. Оказалось, что базидиомицеты в этих лишайниках одноклеточные. При этом располагаются они в верхнем слое тела лишайника, почти на поверхности. И тот, кто смотрит на препарат, видит множество одинаковых кружочков — поперечных срезов гиф аскомицета. Их нельзя отличить друг от друга на глаз, их нельзя адресно окрасить классическими методами. Это тот самый случай, когда дьявол кроется в деталях.
Фото: «Кот Шрёдингера»
Работа, проделанная группой Сприбилла, вызывает восхищение у специалистов по всему миру. Некоторые уже окрестили ее важнейшей вехой в лихенологии со времен открытия двойственной природы лишайников. Удастся ли в ближайшее время закрыть спор XIX века, воспроизведя полноценный лишайник в лаборатории? Достаточно ли для этого добавить третий компонент? Ожидает ли нас рождение новой ветви биохимической промышленности, использующей лишайники, ранее нерентабельные из-за медленного роста?
Удастся ли в ближайшее время закрыть спор XIX века, воспроизведя полноценный лишайник в лаборатории? Достаточно ли для этого добавить третий компонент? Ожидает ли нас рождение новой ветви биохимической промышленности, использующей лишайники, ранее нерентабельные из-за медленного роста?
Совершенно точно можно сказать, что мы стали лучше понимать лишайники, и так же точно — что нам придется переписать их систематику. Об экономическом значении открытия сейчас сложно сказать что-то конкретное — новые эксперименты по ресинтезу еще не поставлены. Но мы определенно стали ближе и к лишайниковым лекарствам, и к лишайниковой еде.
Для чего нужны лишайники
Еда для человека — употребление лишайников в пищу не ограничено B. fremontii. В Исландии принято печь хлеб с добавлением Cetraria islandica, а в Японии с удовольствием едят Umbilicaria esculenta.
Корм для животных — пожалуй, самые известные лишайники — ягель (Cladonia) и исландский мох (Cetraria islandica) — широко распространены в тундре.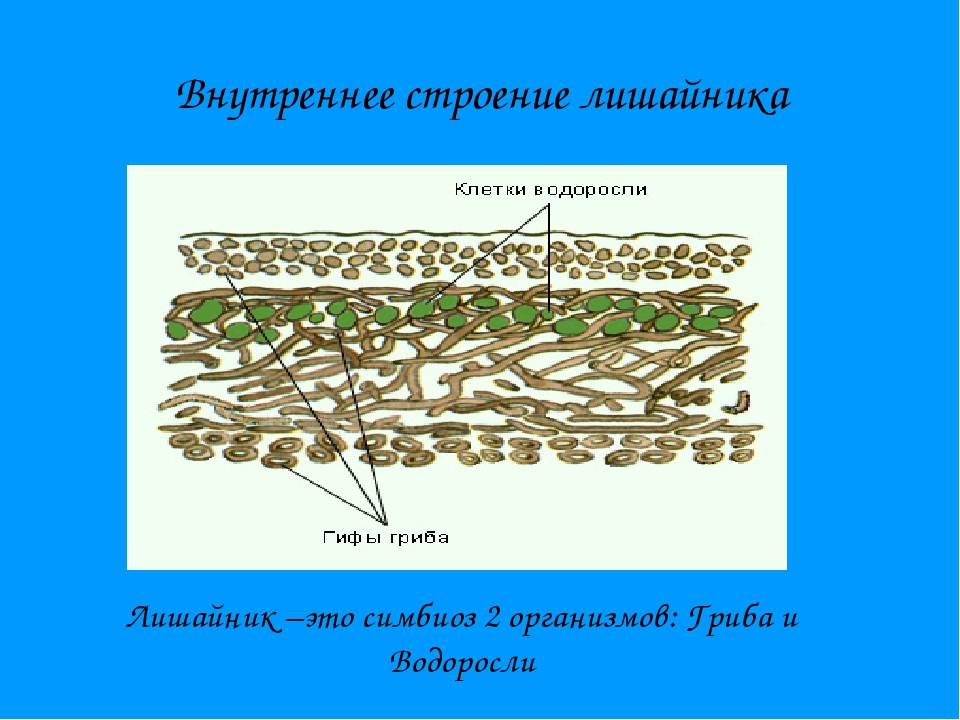 Это основная пища северных оленей. Без них разведение этих животных было бы невозможно.
Это основная пища северных оленей. Без них разведение этих животных было бы невозможно.
Лекарство — лишайники используются в медицине с древнейших времен. Изначально их применяли, исходя из принципа подобия: похожий на легкие лишайник должен лечить от легочных болезней. Позже люди заметили, что лишайники обладают антимикробным действием. Начиная с середины XX века выделенные из лишайников вещества стали активно применять в фармакологии для лечения заболеваний кожи, легких, сердца. Некоторые из них интересны для исследований в области онкологии.
Парфюмерия — широко известен резиноид — продукт переработки дубового мха (Mousse de chene), который можно применять и как ароматизатор, и как фиксатор запаха.
Биология лишайников
Структура
У лишайников нет восковой кутикулы, как у растений на листьях, и у них нет сосудистой ткани, такой как ксилема и флоэма, для перемещения питательных веществ и воды вокруг их талломов, как у растений.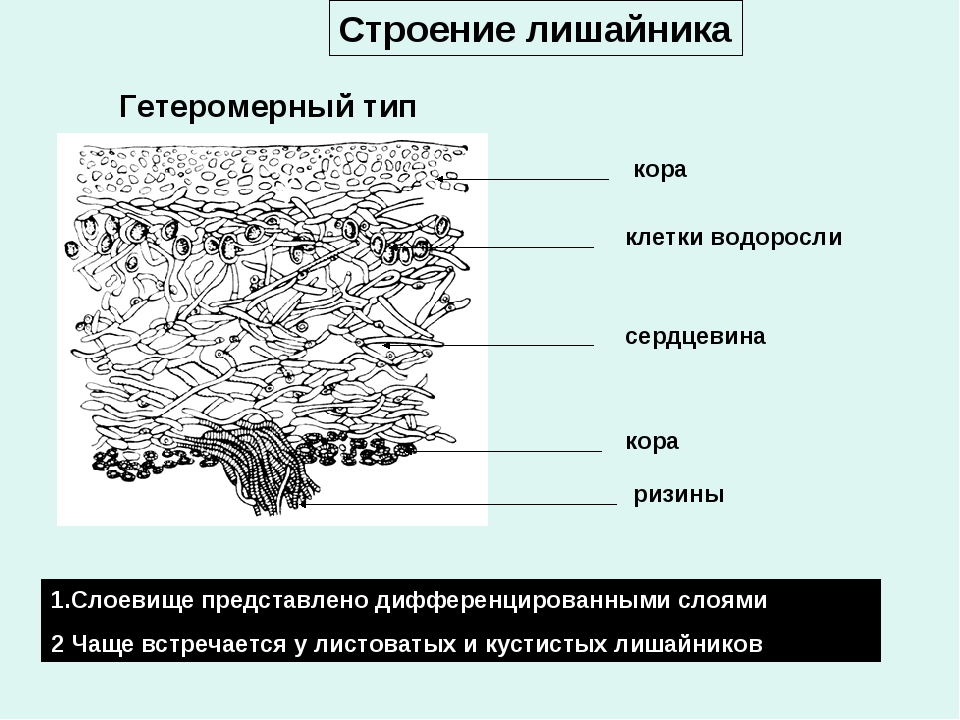 Все, что находится в окружении лишайника, поглощается его структурой. Лишайники получают воду и питательные вещества из окружающей среды через воздух и дождь.
Все, что находится в окружении лишайника, поглощается его структурой. Лишайники получают воду и питательные вещества из окружающей среды через воздух и дождь.
Общая структура лишайника состоит из слоев грибов и водорослей.
Кортекс
Кора — наружный слой слоевища лишайника. Эти клетки толще и более плотно упакованы, чем клетки других грибов в лишайнике. Этот слой обеспечивает небольшую степень защиты, а также обеспечивает цвет у некоторых видов.
Слой водорослей
Обычно можно определить, какая водоросль содержится в лишайнике, только по цвету. Когда лишайник сухой, его цвет обычно серый или окрашен, как грибковые клетки на верхней коре.Когда лишайник влажный, эти клетки становятся прозрачными, а клетки водорослей под ними получают возможность показать свою яркость.
Зеленые водоросли обычно придают лишайнику ярко-зеленый цвет во влажном состоянии, хотя существуют исключения для пигментированных лишайников с зелеными водорослями из-за грибкового партнера, демонстрирующего свои цвета.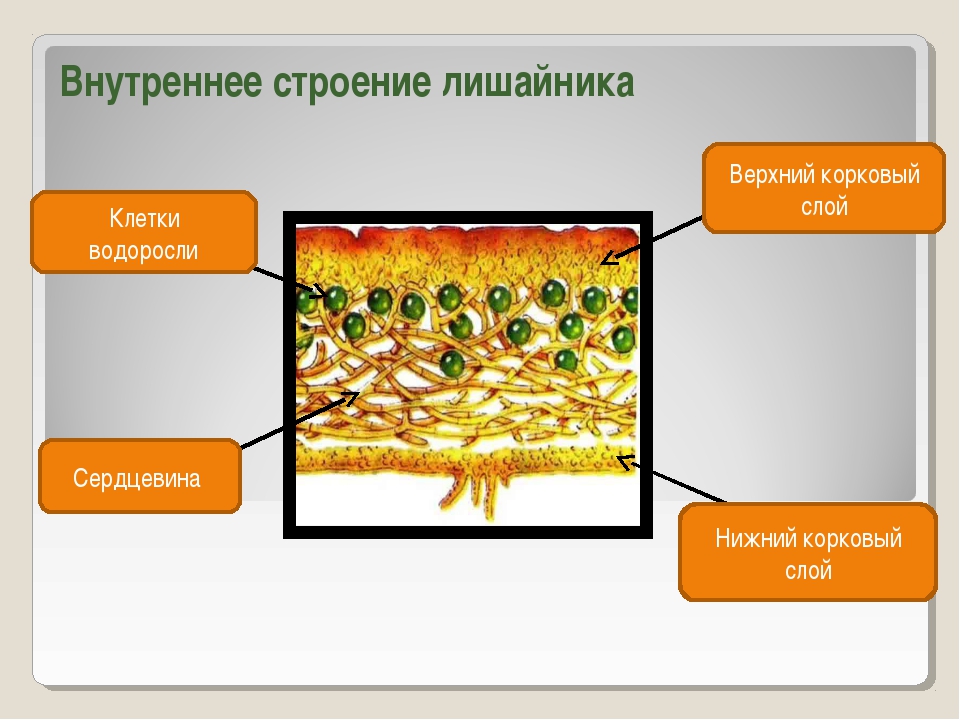
Xanthomendoza mendozae крупным планом. Обратите внимание на слой зеленых водорослей под оранжевой корой. Фото Криса Вагнера, Ю.С. Лесная служба.
Цианобактерии могут быть слоем под верхней корой или в крошечных карманах поверх верхней коры, если уже присутствует слой зеленых водорослей. Цианобактерии придают лишайнику темно-зеленый, коричневый или черный цвет.
Однако в некоторых лишайниках нет слоев грибов и водорослей. Отдельные компоненты смешиваются вместе в один большой однородный слой, и в результате получается студенистая форма роста. Эти виды лишайников называются желейными лишайниками .
Lobaria oregana с L. pulmonaria , растущим в середине. Эти виды являются хорошими примерами зеленых водорослей, просвечивающих через грибного партнера и придающих слоевищам зеленый цвет. Фото Карен Диллман, Лесная служба США.
Peltigera collina , собачий шкурный лишайник. Темный цвет этого вида обусловлен слоем цианобактерий под корой. Фото Карен Диллман, Ю.С. Лесная служба.
Collema furfuraceum с Peltigera sp. (вдоль правой стороны, с черными кончиками): желеобразный лишайник без специфических слоев грибов или водорослей. Фото Карен Диллман, Лесная служба США.
Медулла
Большая часть таллома лишайника состоит из грибковых нитей, называемых мозговым веществом. Он состоит из грибных клеток, которые рыхло упакованы в середине слоевища лишайника, имеют тонкие клеточные стенки и нитевидные формы.В результате под внешней корой образуется ватно-подобное вещество.
Базальная насадка
Лишайники прикрепляются к субстрату разными способами.
Ризины представляют собой нити гриба, которые отходят от мозгового вещества и прикрепляют лишайник к его субстрату. Ризины не имеют сосудистых возможностей, как корни растений. Они не переносят воду или питательные вещества в лишайник; они просто прижимают лишайник к тому, на чем он сидит.
Ризины не имеют сосудистых возможностей, как корни растений. Они не переносят воду или питательные вещества в лишайник; они просто прижимают лишайник к тому, на чем он сидит.
Holdfast является продолжением слоевища лишайника. Вместо многих ризинов у некоторых лишайников есть центральный стержень или фиксатор, который прикрепляется к субстрату, обычно к камню. Эти виды листоватых лишайников называются пуповинными лишайниками, так как центральная закрепка похожа на пуповину.
Umbilicaria phaea , каменный рубец, на скале в южной Калифорнии. У него есть одна центральная застежка, похожая на пуповину, отсюда и название.Фото Криса Вагнера, Лесная служба США.
Формы роста
Существует три основных типа лишайников:
- Лист
- Фруктоза
- Корковая
Листовидные лишайники
Листовидные лишайники имеют две легко различимые стороны. Другими словами, есть верхняя сторона и есть нижняя сторона. Они могут быть очень плоскими, листовыми, как салат, или изогнутыми и полными гребней и выпуклостей.
Другими словами, есть верхняя сторона и есть нижняя сторона. Они могут быть очень плоскими, листовыми, как салат, или изогнутыми и полными гребней и выпуклостей.
Pseudocyphellaria rainierensis , крапчатая старовозрастная. У этого вида есть две четко очерченные верхняя и нижняя стороны, а также бахромчатые доли и множество вегетативных репродуктивных структур, называемых «изидиями». Фото Карен Диллман, Лесная служба США.
Кустарниковые лишайники
Кустарниковые лишайники могут быть висячими и волосовидными, прямостоячими и кустарниковыми или прямостоячими и чашевидными. Многие кустистые лишайники имеют круглые ветви с центральной сердцевиной, а другие полые посередине.Другие кустистые лишайники имеют плоские ветви, которые спутываются друг с другом.
Cladonia fimbriata , трубчатый лишайник. Обратите внимание на чашеобразные стебли. Фото Чарльза Пирса, Michigan Wildflowers.
Usnea longissima , старческая борода. Этот редкий лишайник свисает с центрального стебля и может достигать нескольких футов в длину. Его среда обитания сокращается, и вместо того, чтобы закрепляться через споры, его части отрываются и восстанавливаются в другом месте.Это сильный индикатор загрязнения воздуха. Фото Карен Диллман, Лесная служба США.
Рамалина сублептокарпа . Это пример кустистого лишайника с двусторонними плоскими ветвями. Поскольку эти ветви не отличаются друг от друга (нет ни верха, ни низа), они по-прежнему считаются кустистыми. Фото Карен Диллман, Лесная служба США.
Корковые лишайники
Корковые лишайники — это всего лишь корки.Они образуют корку на поверхности, такой как валун, почва, автомобиль или черепица. Они могут быть разных ярких, ярких цветов, таких как солнечно-желтый, оранжевый и красный, а также серый и зеленый. Корковые лишайники прижаты к своему субстрату.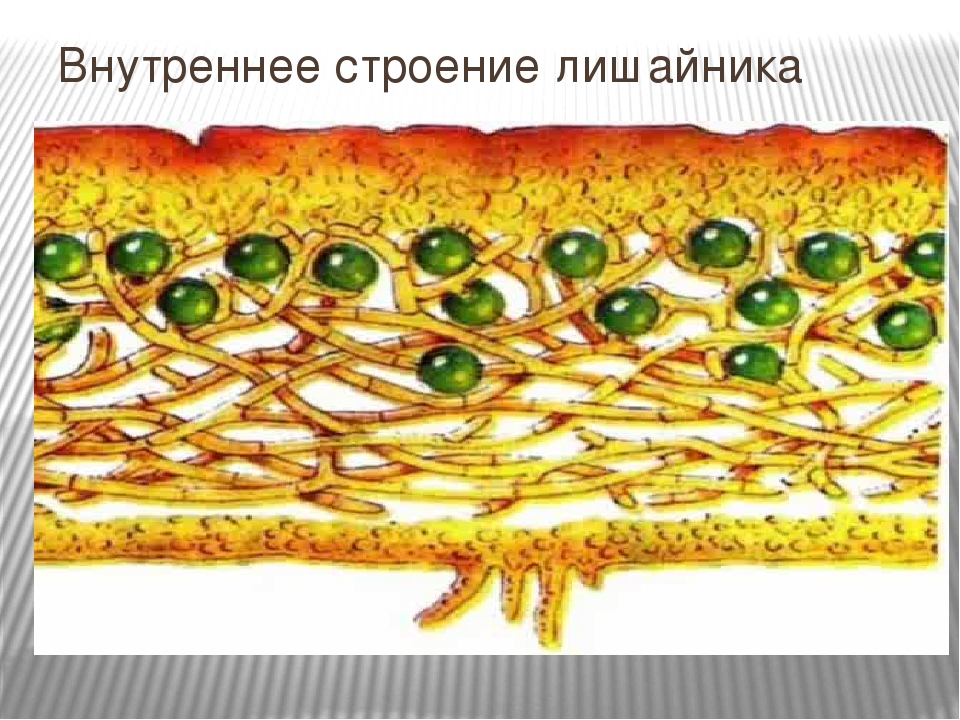
Много разноцветных корок на камне. Фото Ларри Сент-Клера.
Lecanora garovaglii , серая корка, с другими корками, на камне. Фото Криса Вагнера, Ю.С. Лесная служба.
Pleopsidium chlorophanum , желтая корка с другими корками, на камне. Фото Криса Вагнера, Лесная служба США.
Репродукция
Представляете, как размножаются два разных вида, живущие в одном теле? Мы знаем, что для размножения нам нужно иметь два одинаковых вида, чтобы производить плодовитое потомство. Лишайники уникальны тем, что состоят из двух (или более) разных видов или, по сути, царств.
Лишайники бывают разные. В отличие от растений, которые могут производить семена, из которых вырастают новые растения, у лишайников нет простого способа вырастить больше лишайников. Поскольку гриб является доминирующим партнером в отношениях, он получает возможность развивать свои плодовые тела и производить споры.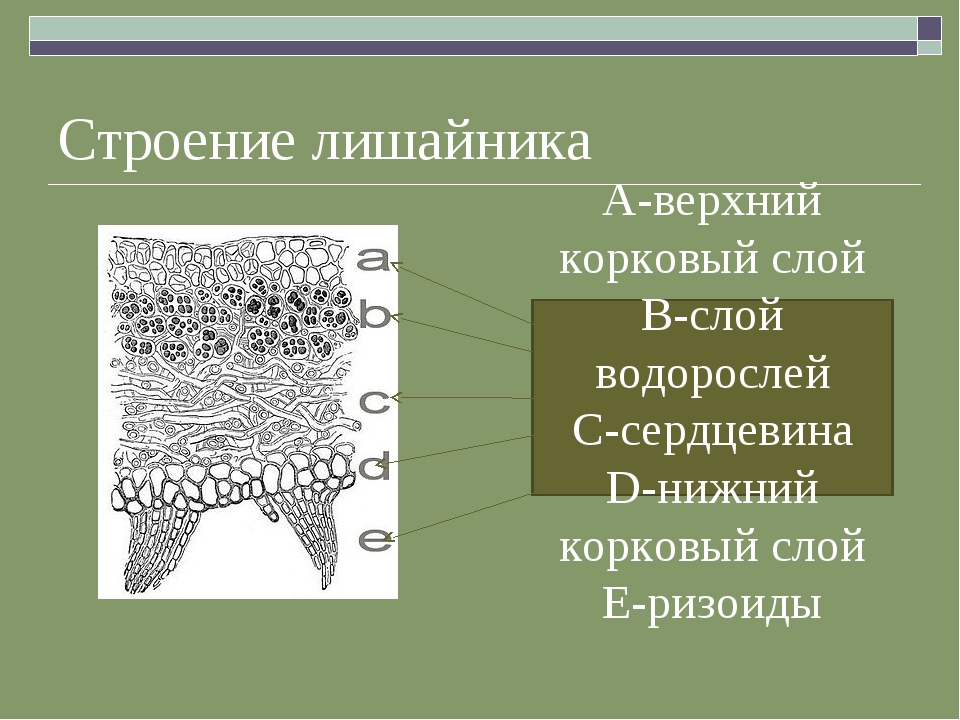 Эти споры могут производить другой гриб, но, к сожалению, для водоросли он вообще не получает возможности размножаться. Либо новый гриб должен найти партнера из водорослей, либо он погибает.
Эти споры могут производить другой гриб, но, к сожалению, для водоросли он вообще не получает возможности размножаться. Либо новый гриб должен найти партнера из водорослей, либо он погибает.
Acarospora badiofusca , корковый лишайник с несколькими плодовыми телами.Многие споры образуются из одного плодового тела. Фото Криса Вагнера, Лесная служба США.
Peltigera didactyla , лишайник собачьей шерсти. Этот молодой экземпляр покрыт соредиями, которые представляют собой переплетенные вегетативные гранулы водорослей и грибов, готовые к распространению. Фото Карен Диллман, Лесная служба США.
Конечно, эволюция должна продолжаться, и лишайники приспособились к своей бивидовой борьбе. Эти организмы могут размножаться вегетативно; то есть у них есть структуры, специально разработанные для распространения фрагментов их слоевищ с обоими партнерами.Мало того, что есть определенные вегетативные структуры, но простые фрагменты лишайника могут вырасти в полноразмерные слоевища.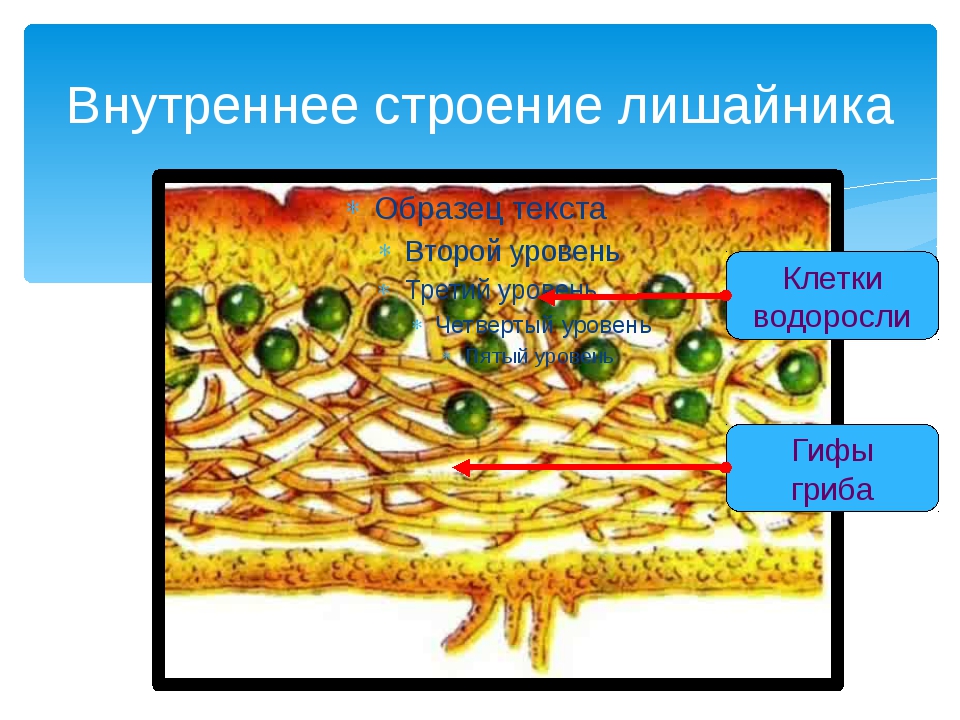
Знакомство с лишайниками
Знакомство с лишайникамиЗнакомство с лишайниками
Сальваторе Де Сантис
Гербарий Стажер
1999
Лишайник выглядит как единый организм, но это на самом деле симбиотические отношения между разными организмами.Он состоит из грибкового партнер (микобионт) и один или несколько фотосинтезирующих партнеры (фотобионт). Партнером по фотосинтезу обычно зеленые водоросли или цианобактерии. На Земле насчитывается около 13 500 видов лишайников. Земной шар.
Обсуждается, является ли родство лишайника мутуалистическим
или часть контролируемого паразитизма .
С одной стороны, гриб и фотобионт находятся в мутуалистических отношениях.
потому что, когда они объединены, они имеют возможность иметь дело с экологическими
условиях, с которыми ни одна из сторон не могла бы справиться самостоятельно. Это также кажется
что ни один из партнеров не причиняет вреда другому. При более внимательном рассмотрении
лишайника, некоторые могут сказать, что фотобионт является пленником микобионта, а не
партнер. Партнер-гриб «порабощает» фотобионт, чтобы питаться от фотобионтов.
фотосинтез.
Это также кажется
что ни один из партнеров не причиняет вреда другому. При более внимательном рассмотрении
лишайника, некоторые могут сказать, что фотобионт является пленником микобионта, а не
партнер. Партнер-гриб «порабощает» фотобионт, чтобы питаться от фотобионтов.
фотосинтез.
В лишайник, микобионт образует таллом , в котором обитает фотобионт. Различают три основных морфологических типа слоевищ: листоватые, корковые и кустистые.
Листовидные лишайники похожи на листья как по внешнему виду, так и по структуре.Они свободно прилипают к субстрату. См. рис. 1.
Корковые лишайники «похожи на корку». Они плотно прилегают коры . Корковые лишайники составляют около 75 процентов всех лишайников на земле. См. рисунок 2.
Кустарниковые лишайники не имеют отчетливых верхней и нижней части и
часто имеют круглое сечение. Их слоевища могут быть прямостоячими, кустарниковыми,
или из висячих прядей. См. рис. 3.
3.
Лишайники могут выживать в суровых условиях, потому что они могут выдерживать высыхание.Недостаток воды прерывает фотосинтез. В этом В подвешенном состоянии некоторые лишайники способны выдерживать большие перепады температур. Медленный рост лишайников, возможно, объясняется их медленной скоростью. фотосинтеза (поскольку они обычно сухие).
Лишайник поглощает большую часть своих минеральных питательных веществ из
воздух и осадки. Загрязнение атмосферы может быть особенно опасным
лишайникам, потому что они сохраняют и могут накапливать смертельно опасные количества тяжелых
металлы, сера, радиоактивные элементы, NO 2 и озон.Сера
двуокись (SO 2 ) особенно смертельна для лишайников, поскольку снижает
pH и портится хлорофилл, что приводит к прекращению фотосинтеза.
Законодательство против диоксида серы за последние 25 лет разрешает лишайникам
вернуться в ранее загрязненные районы. Лишайники использовались для мониторинга
количество загрязняющих веществ в окружающей среде.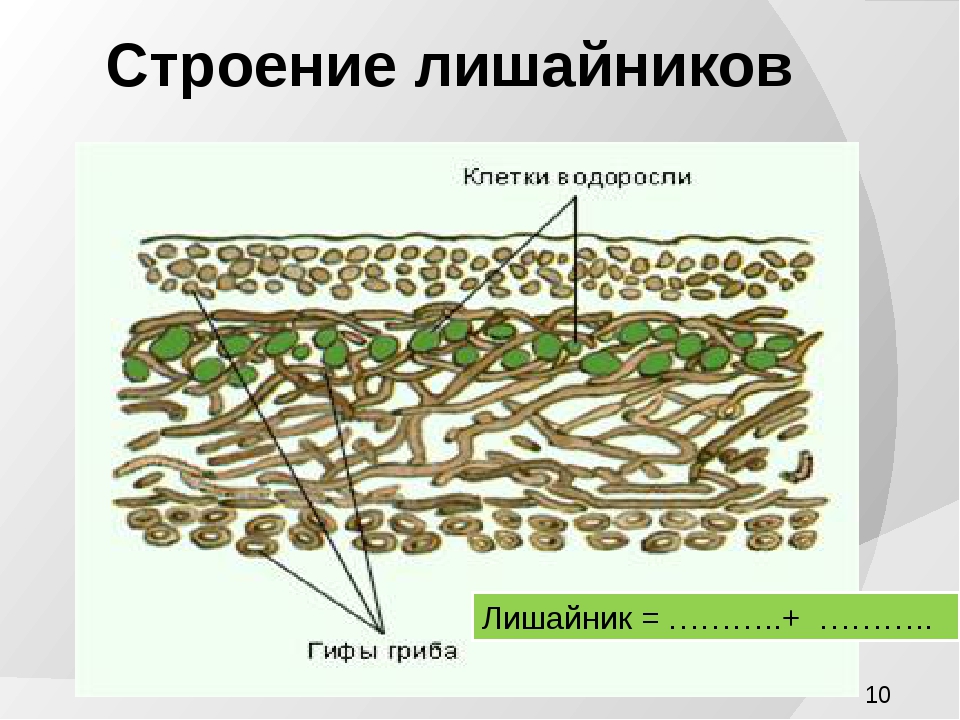 Это делается путем наблюдения за
состояние лишайников, а также их химический состав.
Это делается путем наблюдения за
состояние лишайников, а также их химический состав.
Размножение у лишайников может быть как половым, так и вегетативным (бесполым).При бесполом размножении фрагменты слоевища, содержащие как фотобионт, и микобионт разделяется и образует новый лишайник. Это может произойти, когда кусок слоевища случайно отламывается, но специализированные структуры которые развились в лишайниках, а именно isidia и soredia, обычно несут из этого типа воспроизводства.
У большинства лишайников, размножающихся половым путем, крошечные споры
производится в пределах ascus .То
аски формируются внутри структур, называемых аскоматами .
Наиболее распространенный тип аскомы, называемый апотецием ,
имеет форму открытого диска. При половом размножении участвует только грибной партнер
воспроизводится. Споры, которые прорастают, должны найти подходящий фотобионт.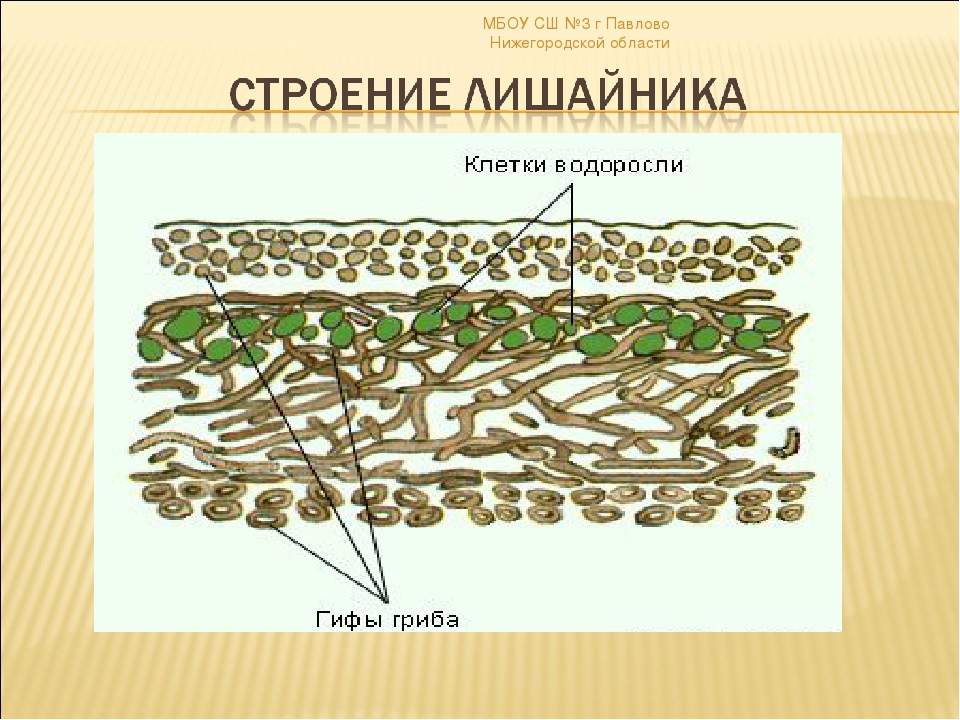 для образования нового лишайника. Поскольку это ненадежный тип воспроизведения,
большое значение имеет вегетативное размножение.
для образования нового лишайника. Поскольку это ненадежный тип воспроизведения,
большое значение имеет вегетативное размножение.
Как
уже упоминалось, лишайники могут быть использованы в качестве индикаторов загрязнения.Лихенометрия – это метод
используется для датирования поверхностей скал, на которых растут определенные лишайники, с помощью их
уровень роста. Лишайники производят много вторичных
соединения , играющие важную роль в различении видов
в лаборатории. Практическое применение этих же соединений наблюдается в медицине.
а также в качестве натуральных красителей и в качестве компонента парфюмерии. В природе эти соединения
может служить защитой от травоядных, а также может помочь разрушить скалу
субстраты.Лишайники колонизируют места, где раньше не было роста, например
как скалы. Лишайники, содержащие цианобактерии в качестве фотобионта, обеспечивают фиксированный азот.
к их среде. Экологически они также важны для обеспечения продовольствием.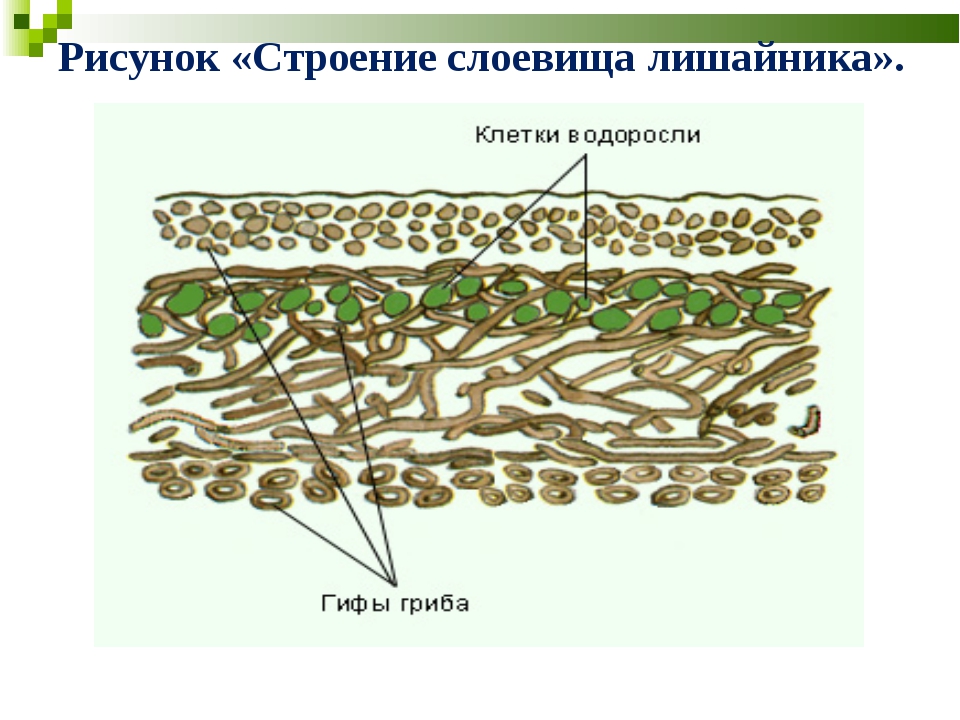 и приют для диких животных, включая оленей, лосей и лосей, а также некоторые виды китов
белок, мышей и летучих мышей. Лишайники вносят свой вклад в разнообразие фона
природного ландшафта и может быть оценен всеми, даже если только подсознательно
на нетренированный взгляд.
и приют для диких животных, включая оленей, лосей и лосей, а также некоторые виды китов
белок, мышей и летучих мышей. Лишайники вносят свой вклад в разнообразие фона
природного ландшафта и может быть оценен всеми, даже если только подсознательно
на нетренированный взгляд.
Форма и строение — Сайт лишайников
| |
Эта страница будет посвящена объяснению общих структур лишайников, которые можно увидеть невооруженным глазом, а также предоставлению некоторой базовой информации о внутреннем строении лишайников. Чтобы помочь проиллюстрировать некоторые пункты, будут ссылки на рисунки, взятые из книги Альберта Шнайдера Учебник общей лихенологии , опубликованной в 1897 году, и есть больше об этих рисунках (и некоторые предупреждения) на странице SCHNEIDER’S BOOK.
Когда вы смотрите на лишайник, такой как листовой Xanthoria filsonii , вы видите желтоватый базальный лист, на котором есть многочисленные более темные желтые диски.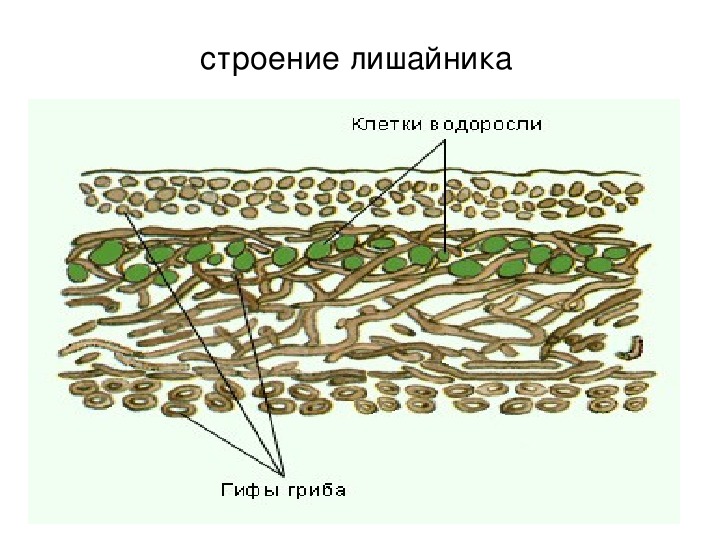 Базальный лист — это таллом (состоит из ткани грибов и фотобионтов), а диски — это апотеции (в которых грибной партнер производит половые споры). У некоторых родов апотеции узкие и удлиненные (иногда даже ветвящиеся) и называются тогда lirellae . Другой тип плодового тела гриба — это перитеций , который невооруженным глазом выглядит как маленький полушаровидный прыщик, обычно черного цвета.Внутри перитеция находятся аски, в которых образуются половые споры. На поверхности слоевища могут быть видны мельчайшие порошкообразные гранулы (называемые soredia ), каждый из которых состоит из нескольких фотобионтных клеток, окруженных грибковыми нитями. Кроме того, слоевище может производить крошечные, простые или разветвленные колючие отростки (называемые isidia ), опять же смесь грибковых и фотобионтных клеток. Изидии легко ломаются, и они, и соредии легко рассеиваются и содержат все необходимое для образования новых слоевищ.
Базальный лист — это таллом (состоит из ткани грибов и фотобионтов), а диски — это апотеции (в которых грибной партнер производит половые споры). У некоторых родов апотеции узкие и удлиненные (иногда даже ветвящиеся) и называются тогда lirellae . Другой тип плодового тела гриба — это перитеций , который невооруженным глазом выглядит как маленький полушаровидный прыщик, обычно черного цвета.Внутри перитеция находятся аски, в которых образуются половые споры. На поверхности слоевища могут быть видны мельчайшие порошкообразные гранулы (называемые soredia ), каждый из которых состоит из нескольких фотобионтных клеток, окруженных грибковыми нитями. Кроме того, слоевище может производить крошечные, простые или разветвленные колючие отростки (называемые isidia ), опять же смесь грибковых и фотобионтных клеток. Изидии легко ломаются, и они, и соредии легко рассеиваются и содержат все необходимое для образования новых слоевищ. Соредии и изидии – средства вегетативного размножения. Существует отдельная страница РЕПРОДУКТИВНЫЕ СТРУКТУРЫ, а остальная часть текущей страницы будет сосредоточена на нерепродуктивных особенностях таллома. Кроме того, почти у всех лишайников микобионтом является аскомицет, и эта страница будет посвящена формам и структурам асколишайников. О БАЗИДИОЛИХЕНАХ есть отдельная страница, которая содержит подробности как вегетативной, так и репродуктивной структуры таких лишайников.
Соредии и изидии – средства вегетативного размножения. Существует отдельная страница РЕПРОДУКТИВНЫЕ СТРУКТУРЫ, а остальная часть текущей страницы будет сосредоточена на нерепродуктивных особенностях таллома. Кроме того, почти у всех лишайников микобионтом является аскомицет, и эта страница будет посвящена формам и структурам асколишайников. О БАЗИДИОЛИХЕНАХ есть отдельная страница, которая содержит подробности как вегетативной, так и репродуктивной структуры таких лишайников.
На странице ЧТО ТАКОЕ ЛИШАЙНИК представлены три основные формы роста слоевища: корковое, листоватое и кустистое.В первых двух слоевище имеет верхнюю (или дорсальную ) поверхность и нижнюю (или вентральную ) поверхность, тогда как в третьем термины верхний и нижний часто не имеют смысла, когда речь идет о слоевище. Обычно у кустистого слоевища есть только внешняя поверхность. У ряда родов таллом имеет компоненты двух различных форм и такой таллом называется диморфным талломом . Cladonia является примером очень распространенного рода, в котором виды имеют диморфные слоевища, и чуть позже на этой странице будет еще несколько о Cladonia .
Cladonia является примером очень распространенного рода, в котором виды имеют диморфные слоевища, и чуть позже на этой странице будет еще несколько о Cladonia .
Слоевище
На следующих очень упрощенных диаграммах поперечного сечения показано общее внутреннее строение многих корковых, листоватых и кустистых лишайников. Крайний слева — корковый вид. Черным цветом обозначена кора , полоса уплотненных гиф грибов, служащая защитной оболочкой. Под корой, зеленым цветом, находится слой фотобионтных клеток. Если осторожно соскоблить верхнюю кору, то можно увидеть слой фотобионтов — зеленый, если это водоросли, и сине-зеленый, если это цианобактерии.На этой фотографии показаны несколько талломов Endocarpon pusillum , часть верхней коры одного из которых соскоблена. Под слоем фотобионта находится мозговое вещество , рыхлое переплетение грибковых гиф, показанное здесь серым цветом. Вы также можете увидеть некоторые серые гифы, растущие из мозгового вещества, и эти гифы проникают в субстрат и, таким образом, очень плотно закрепляют слоевище.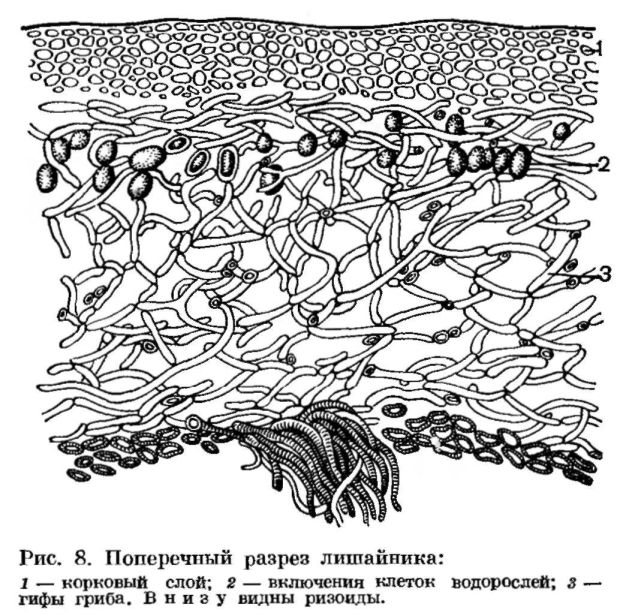
Прежде чем продолжить, я еще раз подчеркну, что эти диаграммы очень упрощены и служат только для передачи взаимного положения коры, мозгового вещества, фотобионтов и ризин в слоевищах многих лишайников.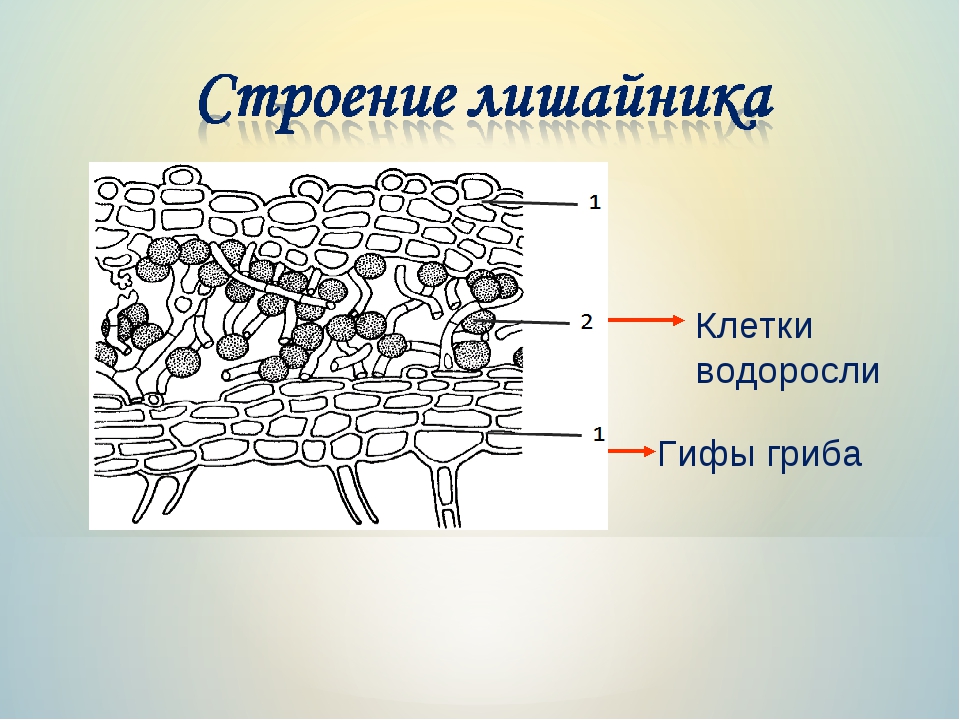 Стоит отметить, что у ряда видов в коре могут быть кристаллические отложения, полученные из вторичных метаболитов, продуцируемых лишайниками. Вскоре я остановлюсь немного подробнее, что позволит вам увидеть некоторые различия в структуре слоевища, а также исключения из некоторых приведенных выше утверждений.Прежде чем сделать это, я приведу более реалистичную схему части слоевища лишайника в конце этого абзаца. У этого вида кора состоит из двух слоев: тонкого верхнего слоя крошечных желтых гранул и более толстого бесцветного слоя под ним. Ниже коры находятся сферические клетки фотобионта (показаны здесь зеленым цветом), а ниже — мозговое вещество. В мозговом веществе хорошо видны отдельные нитевидные гифы, но в нижнем слое коры гифы уплотнены.
Стоит отметить, что у ряда видов в коре могут быть кристаллические отложения, полученные из вторичных метаболитов, продуцируемых лишайниками. Вскоре я остановлюсь немного подробнее, что позволит вам увидеть некоторые различия в структуре слоевища, а также исключения из некоторых приведенных выше утверждений.Прежде чем сделать это, я приведу более реалистичную схему части слоевища лишайника в конце этого абзаца. У этого вида кора состоит из двух слоев: тонкого верхнего слоя крошечных желтых гранул и более толстого бесцветного слоя под ним. Ниже коры находятся сферические клетки фотобионта (показаны здесь зеленым цветом), а ниже — мозговое вещество. В мозговом веществе хорошо видны отдельные нитевидные гифы, но в нижнем слое коры гифы уплотнены.
Несмотря на то, что корковое слоевище имеет отчетливые верхнюю и нижнюю поверхности, из-за плотного прилегания к субстрату нижняя поверхность не видна.Ризины на листовидном талломе могут быть густыми или редкими, встречаются листоватые виды без ризинов. Лишайник с ризином называется ризинат , а ризинат без ризина. Когда ризины присутствуют (и независимо от того, густые они или редкие), у одних видов они могут быть обнаружены где угодно под слоевищем, а у других видов ризины приурочены к определенным участкам. Эта вариация в плотности и размещении ризины означает, что некоторые листоватые талломы довольно слабо прикреплены к своему субстрату, в то время как другие очень прочно прикреплены.У некоторых листоватых видов слоевище прикрепляется только по центру, оставляя остальную часть слоевища свободной от субстрата и, таким образом, позволяя легко увидеть нижнюю сторону. У других листоватых видов слоевища настолько хорошо прикреплены к большей части их нижней поверхности, что их можно было бы сначала принять за корковые виды. Существует также разнообразие формы ризины между видами. Ризины могут быть простыми линейными пучками, как показано на диаграмме выше, однако они также могут быть любыми, от редко до сильно разветвленных.
Лишайник с ризином называется ризинат , а ризинат без ризина. Когда ризины присутствуют (и независимо от того, густые они или редкие), у одних видов они могут быть обнаружены где угодно под слоевищем, а у других видов ризины приурочены к определенным участкам. Эта вариация в плотности и размещении ризины означает, что некоторые листоватые талломы довольно слабо прикреплены к своему субстрату, в то время как другие очень прочно прикреплены.У некоторых листоватых видов слоевище прикрепляется только по центру, оставляя остальную часть слоевища свободной от субстрата и, таким образом, позволяя легко увидеть нижнюю сторону. У других листоватых видов слоевища настолько хорошо прикреплены к большей части их нижней поверхности, что их можно было бы сначала принять за корковые виды. Существует также разнообразие формы ризины между видами. Ризины могут быть простыми линейными пучками, как показано на диаграмме выше, однако они также могут быть любыми, от редко до сильно разветвленных.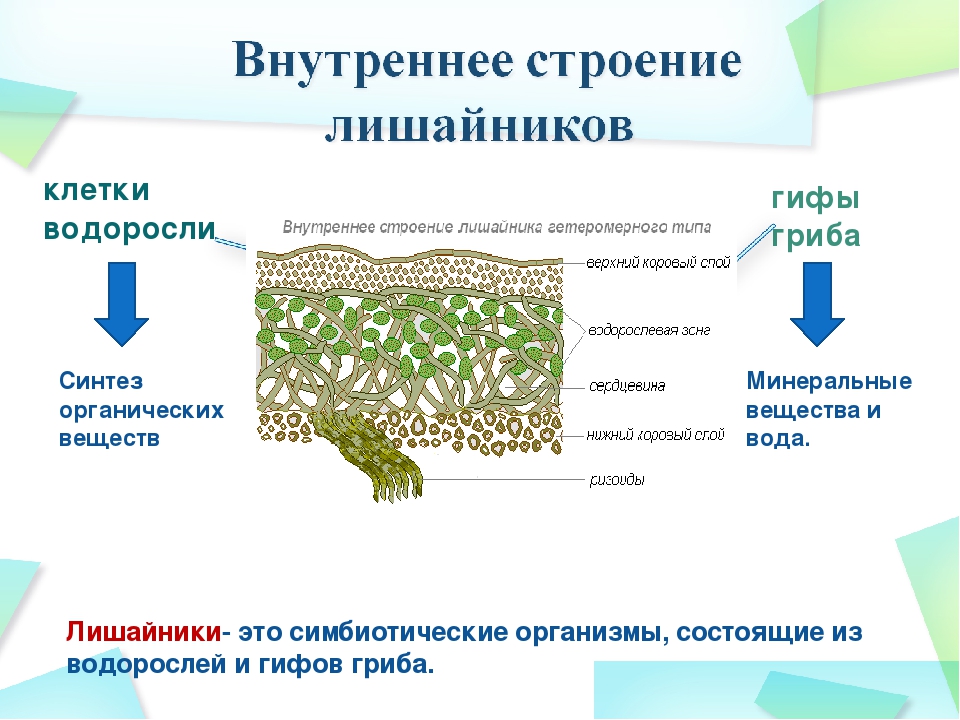 У некоторых видов ризины разветвляются и могут делать это неоднократно. У других каждая ризина имеет главную ось, но с короткими боковыми ответвлениями, отходящими от главной оси, а третьи напоминают потертые кисти для рисования, у которых во все стороны расходятся щетинки. Как уже упоминалось, некоторые лиственные виды не имеют ризины, и ярким примером является Xanthoparmelia semiviridis . Талломы свободно сидят на почве, скрученные в сухом состоянии, но плоские и более темные во влажном состоянии.
У некоторых видов ризины разветвляются и могут делать это неоднократно. У других каждая ризина имеет главную ось, но с короткими боковыми ответвлениями, отходящими от главной оси, а третьи напоминают потертые кисти для рисования, у которых во все стороны расходятся щетинки. Как уже упоминалось, некоторые лиственные виды не имеют ризины, и ярким примером является Xanthoparmelia semiviridis . Талломы свободно сидят на почве, скрученные в сухом состоянии, но плоские и более темные во влажном состоянии.
У некоторых видов гифы могут вырастать из коры, придавая коре волосистый вид.Среди волосатых видов есть такие, у которых волосы редкие, и, с другой стороны, те, у которых волосы густые. Существуют также различия в структуре и расположении таких волос, и для всех этих вариантов существуют технические описательные термины, но нет смысла вдаваться в такие подробности на этом веб-сайте.
Кора значительно различается по своему строению, и существуют лишайники без коры, такие как различные лишайники. Биссоидный таллом имеет несколько тонкий вид, как вата, в некоторой степени выдранная, и состоит из слабо переплетенных гиф и фотобионтных клеток.Если кора присутствует, она варьируется от тонкой до относительно толстой в зависимости от вида. Шнайдер предоставил рисунки ТОНКИХ слоевищ, а также РАЗЛИЧНЫХ более крепких слоевищ. Первая диаграмма листовидного слоевища на этой странице показала, что оно имеет две коры, и это обычное явление, но есть листоватые лишайники, у которых отсутствует нижняя кора. Австралийским примером является Heterodea muelleri , вид, который широко распространен и довольно обычен на юге Австралии. Хотя у него нет нижней коры, в слоевище есть ризины.
Биссоидный таллом имеет несколько тонкий вид, как вата, в некоторой степени выдранная, и состоит из слабо переплетенных гиф и фотобионтных клеток.Если кора присутствует, она варьируется от тонкой до относительно толстой в зависимости от вида. Шнайдер предоставил рисунки ТОНКИХ слоевищ, а также РАЗЛИЧНЫХ более крепких слоевищ. Первая диаграмма листовидного слоевища на этой странице показала, что оно имеет две коры, и это обычное явление, но есть листоватые лишайники, у которых отсутствует нижняя кора. Австралийским примером является Heterodea muelleri , вид, который широко распространен и довольно обычен на юге Австралии. Хотя у него нет нижней коры, в слоевище есть ризины.
На каждой из приведенных выше простых диаграмм клетки фотобионта показаны ограниченными четко определенной полосой непосредственно под корой и отличной от мозгового вещества. Так обстоит дело с очень многими лишайниками (и такие лишайники описываются как гетеромерных ), но есть несколько родов, в которых фотобионтные клетки распределены по слоевищу случайным образом, и Шнайдер включил рисунки такого вида, описанного как гомомерных.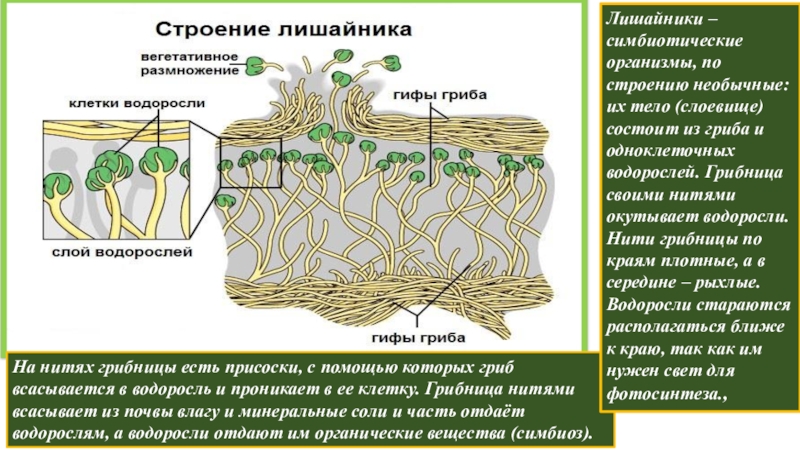 . В любом случае гифы грибов «собирают» большую часть питательных веществ, производимых фотобионтом, и следующие иллюстрации (опять же взятые из книги Шнайдера) показывают, что гифы грибов разветвляются и окружают ряд шаровидных клеток фотобионта.Существуют некоторые различия в том, как гифы прикрепляются к клеткам фотобионта или проникают в них, чтобы собрать продукты фотосинтеза.
. В любом случае гифы грибов «собирают» большую часть питательных веществ, производимых фотобионтом, и следующие иллюстрации (опять же взятые из книги Шнайдера) показывают, что гифы грибов разветвляются и окружают ряд шаровидных клеток фотобионта.Существуют некоторые различия в том, как гифы прикрепляются к клеткам фотобионта или проникают в них, чтобы собрать продукты фотосинтеза.
Подеция и псевдоподеция
Cladonia — очень широко распространенный род лишайников, многие виды которого первоначально развиваются в виде чешуйчатых слоевищ. Чешуя представляет собой небольшое чешуйчатое слоевище, а колония чешуйчатого лишайника имеет вид россыпи мелких чешуек или чешуек на субстрате.Однако вид Cladonia также образует прямостоячие кустистые структуры, называемые podetia , иногда с появлением довольно простых стеблей, иногда расширяющихся на вершине и, таким образом, имеющих несколько трубчатую форму.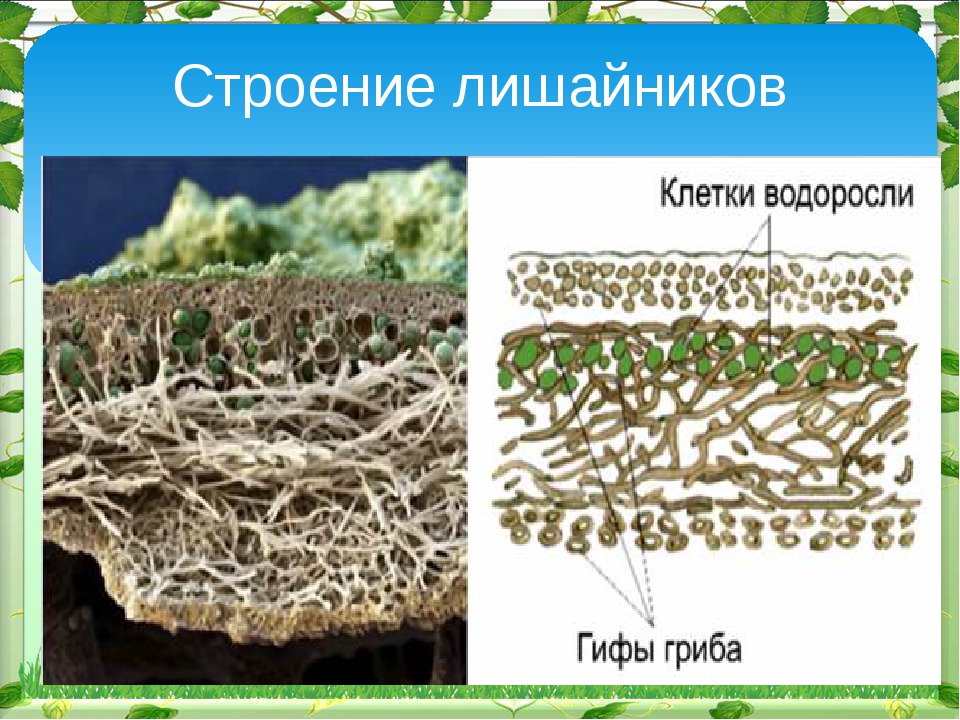 На этом фото вы видите растущую на почве колонию Cladonia . На почве много чешуек, но можно увидеть и ряд прямостоячих подеций с более широкими вершинами. Более того, по краям некоторых более широких вершин вы можете увидеть развитие дополнительных подеций, а также чешуйчатые наросты на подециях, как показано на этом увеличенном фрагменте предыдущей фотографии.Справа значительное увеличение другой части первой фотографии. Справа значительное увеличение части предыдущей фотографии. Желтые стрелки указывают на доли чешуйчатого слоевища, из которого растет подеций. Этот молодой подиум имеет высоту несколько миллиметров. Белая стрелка указывает на то, что выглядит как еще один таллом, который находится в контакте с талломом, указанным желтыми стрелками, а не частью слоевища, отмеченного желтой стрелкой. Чешуйки, такие как те, что на почве вокруг оснований подеций, часто называют базальными чешуйками .У ряда из видов Cladonia базальные чешуйки исчезают и их не видно после того, как подеции разовьются.
На этом фото вы видите растущую на почве колонию Cladonia . На почве много чешуек, но можно увидеть и ряд прямостоячих подеций с более широкими вершинами. Более того, по краям некоторых более широких вершин вы можете увидеть развитие дополнительных подеций, а также чешуйчатые наросты на подециях, как показано на этом увеличенном фрагменте предыдущей фотографии.Справа значительное увеличение другой части первой фотографии. Справа значительное увеличение части предыдущей фотографии. Желтые стрелки указывают на доли чешуйчатого слоевища, из которого растет подеций. Этот молодой подиум имеет высоту несколько миллиметров. Белая стрелка указывает на то, что выглядит как еще один таллом, который находится в контакте с талломом, указанным желтыми стрелками, а не частью слоевища, отмеченного желтой стрелкой. Чешуйки, такие как те, что на почве вокруг оснований подеций, часто называют базальными чешуйками .У ряда из видов Cladonia базальные чешуйки исчезают и их не видно после того, как подеции разовьются. Здесь и здесь другие примеры Cladonia podetia. Цветные области на вершине подеций — это апотеции. Гифы, помогающие формировать подеции, образуются из генеративной ткани . У лишайников эта ткань окружает половые органы и дает начало грибковым спорообразующим структурам (например, апотециям). Таким образом, с точки зрения развития весь подеций можно рассматривать как часть плодового тела.Существует ряд родов лишайников, у которых апотеции держатся вверху на стеблях, но стебель состоит из вегетативной ткани слоевища, а генеративная ткань находится только на верхушке стебля. В таком случае стебель представляет собой просто удлинение слоевища и, хотя внешне он может быть похож на подеций, в развитии совсем другой и называется псевдоподеций . Например, коричневые апотеции Stereocaulon ramulosum образуются на псевдоподециях.Многие виды Stereocaulon имеют диморфные талломы с непостоянным первичным талломом (часто корковым или чешуйчатым), из которого развиваются кустистые и часто сильно разветвленные псевдоподеции.
Здесь и здесь другие примеры Cladonia podetia. Цветные области на вершине подеций — это апотеции. Гифы, помогающие формировать подеции, образуются из генеративной ткани . У лишайников эта ткань окружает половые органы и дает начало грибковым спорообразующим структурам (например, апотециям). Таким образом, с точки зрения развития весь подеций можно рассматривать как часть плодового тела.Существует ряд родов лишайников, у которых апотеции держатся вверху на стеблях, но стебель состоит из вегетативной ткани слоевища, а генеративная ткань находится только на верхушке стебля. В таком случае стебель представляет собой просто удлинение слоевища и, хотя внешне он может быть похож на подеций, в развитии совсем другой и называется псевдоподеций . Например, коричневые апотеции Stereocaulon ramulosum образуются на псевдоподециях.Многие виды Stereocaulon имеют диморфные талломы с непостоянным первичным талломом (часто корковым или чешуйчатым), из которого развиваются кустистые и часто сильно разветвленные псевдоподеции.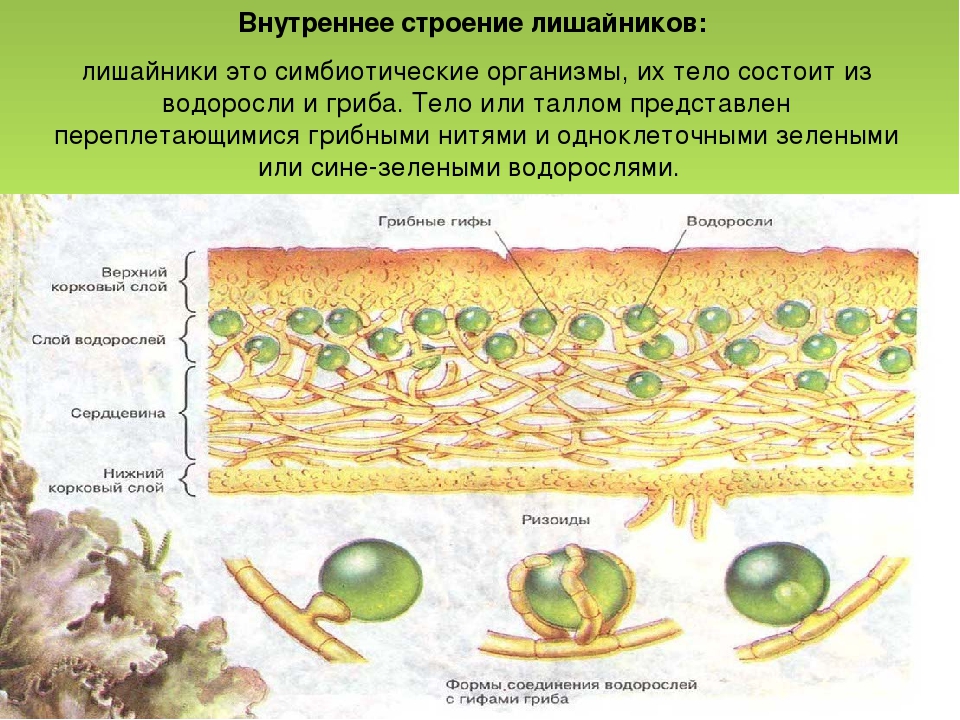 У рода Cladia талломы состоят из перфорированных сетчатых псевдоподеций, поэтому мозговой слой подвергается контакту с наружным воздухом.
У рода Cladia талломы состоят из перфорированных сетчатых псевдоподеций, поэтому мозговой слой подвергается контакту с наружным воздухом.
Цифеллы, псевдоцифеллы и пятна
Цифеллы и псевдоцифеллы представляют собой разрывы в коре слоевища, но имеют две разные формы.Невооруженным глазом cyphella выглядит как гладкое углубление в слоевище. Поверхность углубления выстлана грибковыми клетками, отличными по форме от гиф мозгового вещества. Хотя псевдоцифелла также представляет собой разрыв в коре, это тот случай, когда, в отличие от цифеллы, гифы мозгового вещества подвергаются воздействию воздуха, а не имеют дифференцированный слой гиф, отделяющий мозговое вещество от внешней среды. Вы можете найти диаграммы Шнайдера структур цифеллы и псевдоцифеллы ЗДЕСЬ.Cyphellae встречаются только у листовидного рода Sticta и почти всегда приурочены к нижней стороне слоевища. Псевдоцифеллы встречаются у небольшого числа родов листоватых, кустистых и даже накипных лишайников. При наличии в листоватых лишайниках псевдоцифеллы могут быть обнаружены на нижней или верхней поверхности в зависимости от вида. Цифеллы и псевдоцифеллы являются местами газообмена.
При наличии в листоватых лишайниках псевдоцифеллы могут быть обнаружены на нижней или верхней поверхности в зависимости от вида. Цифеллы и псевдоцифеллы являются местами газообмена.
У лишайников со слоем фотобионтов под корой слой фотобионтов не обязательно должен быть непрерывным.Поскольку слой фотобионта вносит свой вклад в общий цвет слоевища, область, свободная от фотобионта, будет отображаться в виде более светлого пятна или линии на слоевище. Такие пятна или линии, обычно неправильной формы, называются макул (единственное число: макула).
Смешанные фотобионты
У ряда лишайников водоросль является основным фотобионтом, но цианобактерии присутствуют в качестве второстепенного фотобионта в дискретных карманах, и такой карман называется cephalodium .Цефалодии могут быть обнаружены внутри слоевища или в виде наростов на слоевище. В ряде лишайников свободноживущие клетки цианобактерий могут попадать в волосистую кору, а затем вокруг таких клеток вырастают дополнительные гифы, которые в конечном итоге образуют внешние цефалодии. Внутренние цефалодии распространены у представителей родов Lobaria и Sticta , где они развиваются в виде комков в мозговом веществе. Эти цефалодии могут стать настолько большими, что вызывают видимые вздутия поверхности слоевища. Внешние цефалодии различаются по форме у разных видов и могут быть дисковидными, округлыми или даже разветвленными.У некоторых видов разграниченные области менее разграничены, чем у других, что приводит к незначительным разногласиям во мнениях. Например, вид Solorina crocea из северного полушария имеет как водорослевые, так и цианобактериальные фотобионты, причем последние обнаружены в пятнистом слое под слоем водорослей. Я видел, как в одной книге этот вид описывается как имеющий двойной слой фотобионтов, тогда как в другой книге пятна описываются как цефалодии. У вида Compsocladium archboldianum (известного из Ириан-Джая и Папуа-Новой Гвинеи) клетки водорослей образуют фотобионтный слой под корой, а отдельные цианобактериальные нити обнаруживаются в мозговом веществе.
Внутренние цефалодии распространены у представителей родов Lobaria и Sticta , где они развиваются в виде комков в мозговом веществе. Эти цефалодии могут стать настолько большими, что вызывают видимые вздутия поверхности слоевища. Внешние цефалодии различаются по форме у разных видов и могут быть дисковидными, округлыми или даже разветвленными.У некоторых видов разграниченные области менее разграничены, чем у других, что приводит к незначительным разногласиям во мнениях. Например, вид Solorina crocea из северного полушария имеет как водорослевые, так и цианобактериальные фотобионты, причем последние обнаружены в пятнистом слое под слоем водорослей. Я видел, как в одной книге этот вид описывается как имеющий двойной слой фотобионтов, тогда как в другой книге пятна описываются как цефалодии. У вида Compsocladium archboldianum (известного из Ириан-Джая и Папуа-Новой Гвинеи) клетки водорослей образуют фотобионтный слой под корой, а отдельные цианобактериальные нити обнаруживаются в мозговом веществе.
Обратите внимание, что в некоторых ссылках цефалодия определяется как ограниченная область одного типа фотобионта в лишайнике, где основной фотобионт принадлежит к другому типу, что позволяет учитывать водорослевые карманы в лишайнике с доминирующим фотобионтом — цианобактерией. Однако такой пример, по-видимому, не известен, но даже если такие примеры существуют, наиболее распространенным расположением цефалодия по-прежнему будет цианобактериальный фотобионт в лишайнике, где водоросль является доминирующим фотобионтом.
Что доминирует — гриб или фотобионт?
У большинства лишайников грибная ткань составляет основную часть слоевища, как показано на простых диаграммах в верхней части этой страницы.Существует небольшое число родов, в которых преобладает фотобионтная ткань. В эту категорию попадает ряд биссоидных родов с ватноподобными слоевищами, а также некоторые более сильные роды, такие как Collema и Leptogium . В последних двух фотобионт является цианобактерией рода Nostoc , а талломы Collema и Leptogium можно было первоначально принять за Nostoc , особенно если у лишайников отсутствовали апотеции. На следующих фотографиях показаны Nostoc (слева) и плодородный Collema (справа).Без белого апотеция таллом Collema безликий и студенистый по текстуре, точно так же, как Nostoc .
На следующих фотографиях показаны Nostoc (слева) и плодородный Collema (справа).Без белого апотеция таллом Collema безликий и студенистый по текстуре, точно так же, как Nostoc .
Учитывая, что грибная ткань обычно доминирует, можно было бы сделать вывод, что в таких случаях именно гриб определяет, будет ли слоевище корковым, листоватым или кустистым. Однако случай STICTA AND ‘DENDRISCOCAULON’ поучителен. На нем показан один гриб, связанный с разными фотобионтами (один с водорослями, другой с цианобактериями), но без цианобактерий, приуроченных к цефалодиям.Вместо этого существует диморфный таллом, при этом одна форма таллома содержит водоросль, а другая — цианобактерии, так что на форму таллома влияет фотобионт. Обратите внимание, что у Cladonia и Stereocaulon разные компоненты диморфного таллома имеют один и тот же фотобионт, хотя вы также найдете цефалодии у многих видов Stereocaulon .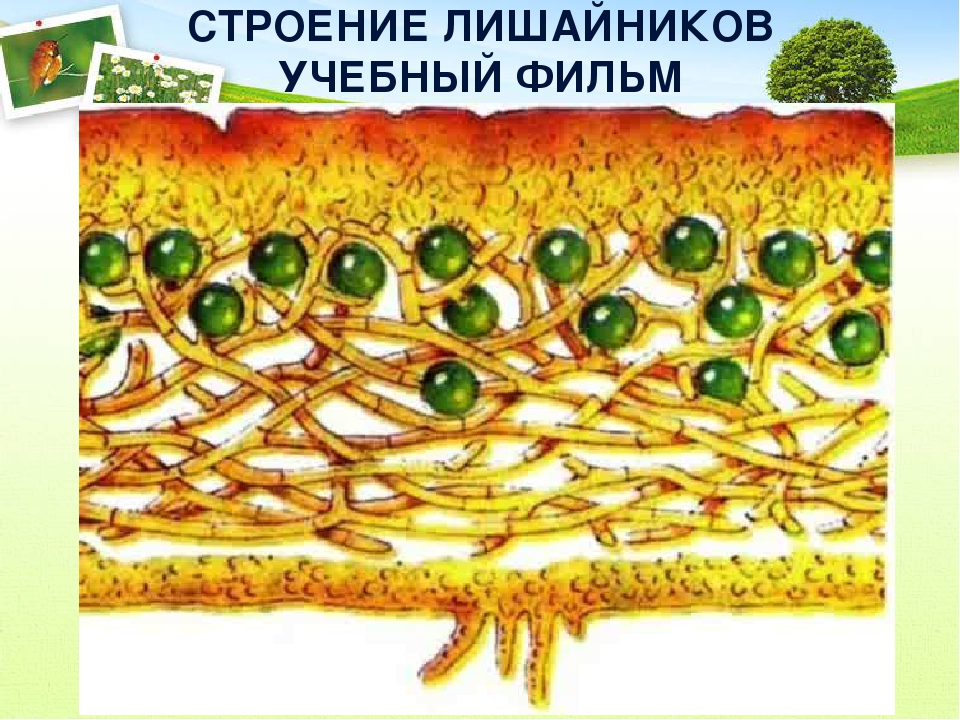
Определение, структура и воспроизведение (с диаграммой)
В этой статье мы обсудим:- 1. Определение лишайников 2. Строение лишайников 3. Взаимоотношения 4. Размножение.
Определение лишайников:
Лишайники представляют собой двойные организмы или объекты, содержащие постоянную ассоциацию гриба или микобионта и водоросли или фикобионта. Грибным партнером обычно является Ascomycota, а иногда и базидиомицет. Партнером из водорослей в основном является зеленая водоросль или цианобактерия (сине-зеленая водоросль).
Термин лишайник был введен Теофрастом (370-285 гг.С.). Насчитывается около 400 родов и 15 000 видов лишайников.
Лишайники часто растут в самых негостеприимных и необитаемых местах, таких как бесплодные скалы, остывшая вулканическая лава, ледяная тундра или альпийские горы, песчаные дюны, крыши, стены, оконные стекла, кора деревьев, листья и т.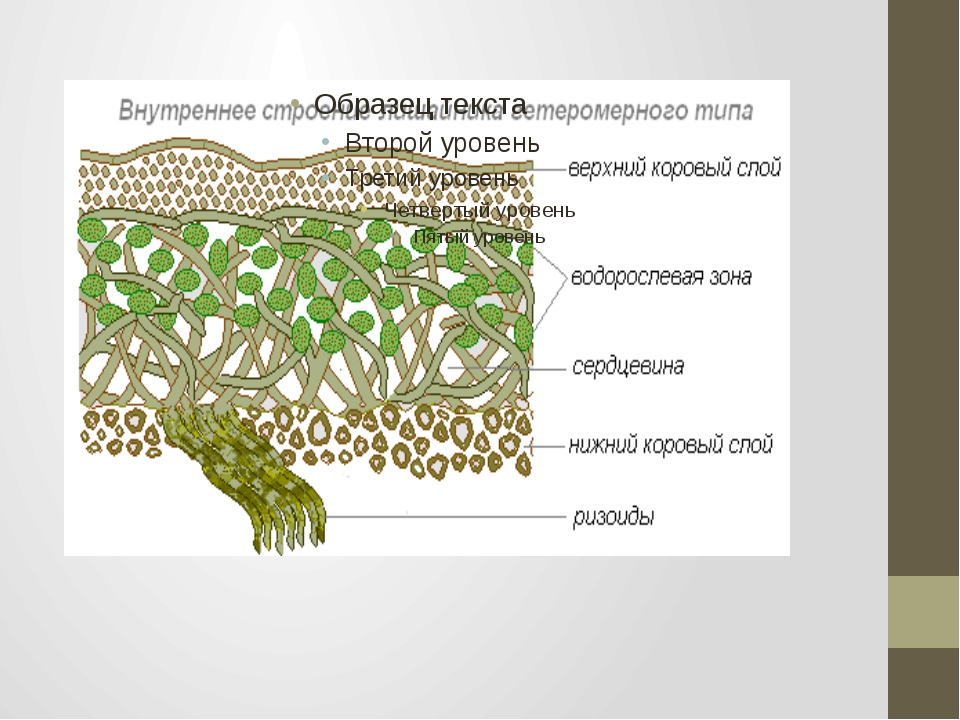 д. Они обычно живут во влажных и открытых условиях, но могут переносят сильную засуху.
д. Они обычно живут во влажных и открытых условиях, но могут переносят сильную засуху.
Однако лишайники не переносят загрязнение воздуха, особенно из-за диоксида серы. Лишайники многолетние. Их рост медленный.Считается, что некоторым лишайникам арктического региона 4500 лет. Лишайники имеют сероватую, желтоватую, зеленоватую, оранжевую, темно-коричневую или черноватую окраску.
Строение лишайников:
По форме лишайники бывают трех видов:
(i) Корковая:
Корковидные, плотно прилегающие к субстрату и прикрепленные к нему в нескольких местах, например, Graphis, Lecanora, Rhizocarpon, Haematoma.
(ii) Лист:
Тело лишайника плоское, широкое, лопастное, листовидное, прикрепленное к субстрату в одном или нескольких местах, например Parmelia, Peltigera. Листовидный лишай Cora (= Dictyonema) pavonia напоминает по внешнему виду трутовки.
(iii) Фруктоза:
Лишайник ветвистый, как куст, и прикрепляется к субстрату с помощью диска, например, Cladonia, Usnea, Evernia. Основная часть тела лишайника образована грибным партнером или микобионтом.Он включает поверхность, сердцевину (или внутреннюю часть) и ризины (прикрепляющие приспособления). Водоросль-партнер или фикобионт составляет едва ли 5% тела лишайника. Обычно он ограничен узкой зоной (зоной водорослей) под поверхностью.
Взаимоотношения лишайников:
Грибок выполняет три функции:
(i) Конструкция кузова и покрытие
(ii) Анкеровка
(iii) Поглощение воды и минералов.Он может поглощать воду из влажного воздуха (атмосферы), росы и дождя.
Минералы собираются как из субстрата, так и из атмосферы. Специальные химические вещества выделяются грибком-партнером лишайника для растворения минералов из субстрата. Основная функция водорослей – фотосинтез.
Цианобактериальная водоросль дополнительно принимает участие в фиксации азота. Водоросль поглощает воду и минеральные соли гриба, в то время как гриб получает часть пищи, произведенной водорослью.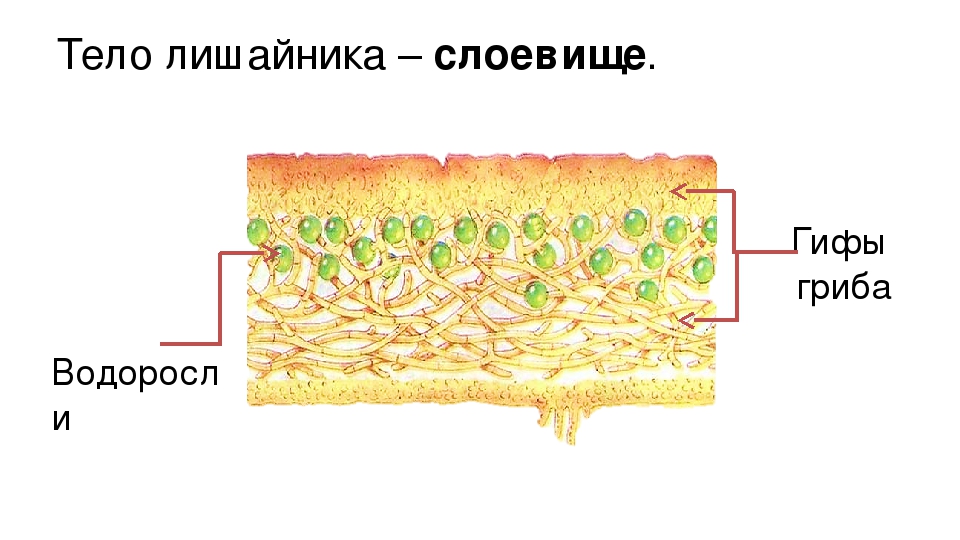 Таким образом, в лишайнике связь между водорослями и грибами является взаимовыгодной (мутуализм), обычно называемой симбиозом.
Таким образом, в лишайнике связь между водорослями и грибами является взаимовыгодной (мутуализм), обычно называемой симбиозом.
Однако иногда грибок обнаруживается:
(i) Посадка гаусторий в клетки водорослей
(ii) Побуждение водорослей к выделению органических веществ и
(iii) Предотвратить образование пектинового покрытия водорослями.
Поэтому некоторые работники считают, что грибок является контролируемым паразитом водоросли. Явление называется гелотизмом.Несмотря на знания о водорослевых и грибных компонентах лишайника и их взаимоотношениях, до сих пор не удавалось создать искусственный лишайник.
Размножение лишайников:
Лишайники размножаются четырьмя способами:
(i) Прогрессирующая гибель и разложение, приводящие к разделению лишайника на две или более частей,
(ii) Фрагментация, вызванная механическим повреждением ветром, вытаптыванием или укусами животных,
(iii) Isidia – это поверхностные отростки лишайников, которые в первую очередь предназначены для увеличения площади поверхности и фотосинтетической активности.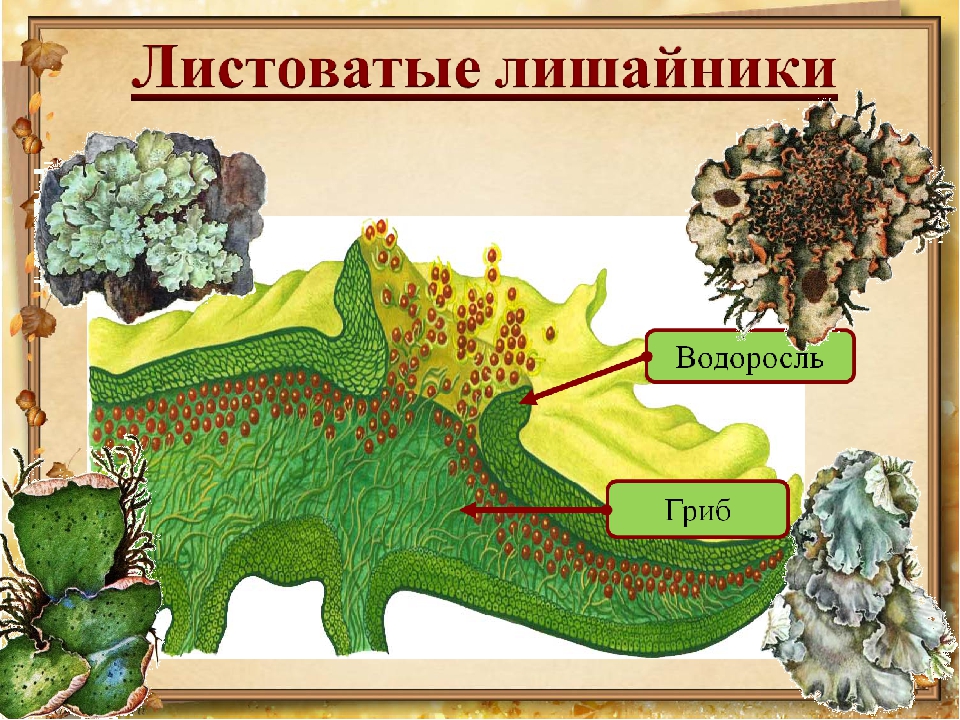 Иногда они обрываются. Каждый изидий способен образовывать новый лишайник, потому что он имеет ядро из клеток водорослей, окруженное оболочкой из грибных гиф.
Иногда они обрываются. Каждый изидий способен образовывать новый лишайник, потому что он имеет ядро из клеток водорослей, окруженное оболочкой из грибных гиф.
(iv) Соредия. Это микроскопические пропагулы лишайника, которые образуются в большом количестве внутри сорусов, называемых пустулами. Соредии разносятся воздушными потоками. Каждая соредия, попадая на подходящий субстрат, дает начало лишайнику, так как имеет немногочисленные клетки водорослей, не полностью окруженные утком гриба.
границ | Понимание многовидовых микробных симбиозов
Введение
Через двадцать лет после того, как теория эволюции путем естественного отбора начала революционизировать биологию, немецкий миколог Антон де Бари ввел в более широкое научное сообщество термин «симбиоз» как сосуществование разнородных организмов (de Bary, 1879).Одним из его ярких примеров были лишайники, хотя симбиотическая природа, открытая ранее Швенденером (1869), в то время вряд ли была принята.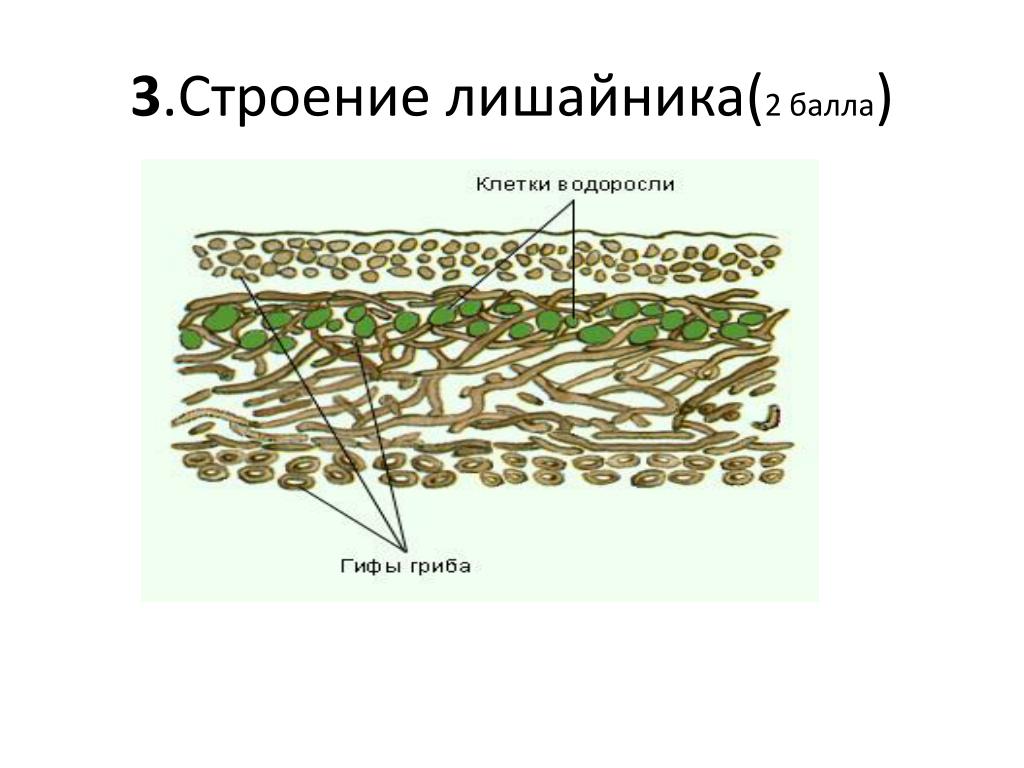 Научные коллеги по-прежнему рассматривали их как самостоятельную группу организмов с уникальной морфологией. Между тем в каждый учебник биологии лишайники включены как облигатная ассоциация между грибом (микобионтом) и фотосинтезирующим партнером (фотобионтом), которым могут быть как цианобактерии, так и/или зеленые водоросли (Nash, 2008). Благодаря этой ассоциации производство энергии фотобионтом за счет фиксации углекислого газа усиливается защитными структурами соседствующего гриба-партнера.Совместная структура, также известная как слоевище лишайника, уникальна и является одной из самых сложных вегетативных структур во всем царстве грибов. Слоевище лишайников возникло еще в наземной растительной жизни, поскольку первые предки лишайников с характерной морфологией можно проследить до девона 400 миллионов лет назад (Remy et al., 1994; Honegger et al., 2013). В этой статье мы покажем, что лишайники представляют собой не просто партнерство, включающее две неродственные группы организмов, но включают в себя до сих пор в значительной степени игнорируемый бактериальный компонент, который вносит вклад в биологию холобионта.
Научные коллеги по-прежнему рассматривали их как самостоятельную группу организмов с уникальной морфологией. Между тем в каждый учебник биологии лишайники включены как облигатная ассоциация между грибом (микобионтом) и фотосинтезирующим партнером (фотобионтом), которым могут быть как цианобактерии, так и/или зеленые водоросли (Nash, 2008). Благодаря этой ассоциации производство энергии фотобионтом за счет фиксации углекислого газа усиливается защитными структурами соседствующего гриба-партнера.Совместная структура, также известная как слоевище лишайника, уникальна и является одной из самых сложных вегетативных структур во всем царстве грибов. Слоевище лишайников возникло еще в наземной растительной жизни, поскольку первые предки лишайников с характерной морфологией можно проследить до девона 400 миллионов лет назад (Remy et al., 1994; Honegger et al., 2013). В этой статье мы покажем, что лишайники представляют собой не просто партнерство, включающее две неродственные группы организмов, но включают в себя до сих пор в значительной степени игнорируемый бактериальный компонент, который вносит вклад в биологию холобионта. Мы начнем с некоторых общих аспектов экологии лишайников, а затем продолжим описание того, как современные аналитические инструменты используются для понимания лишайников как увлекательного случая мультисимбиоза.
Мы начнем с некоторых общих аспектов экологии лишайников, а затем продолжим описание того, как современные аналитические инструменты используются для понимания лишайников как увлекательного случая мультисимбиоза.
Успешный грибковый симбиоз, включающий более 18 000 названных видов грибов, характеризуется пойкилогидрическим образом жизни, что позволяет лишайникам колонизировать почти все наземные среды, от тропических до полярных климатических зон, от прибрежных до высокогорных местообитаний.Кроме того, лишайники растут на поверхности почти всех видов субстрата, включая голые почвы, скалы и растения, но их также можно найти в пресноводных водотоках и в морских приливных зонах (Nash, 2008), а также на различных искусственных поверхностях. . Вегетативные тела различаются по цвету, размеру (от нескольких миллиметров до метров) и формам роста, а некоторые могут сохраняться в течение нескольких тысяч лет (Denton and Karlén, 1973). Большое разнообразие структур слоевищ лишайников, определяемых в первую очередь грибным партнером, можно условно разделить на три наиболее распространенных морфологических типа: корковые, листоватые и кустистые формы роста. Существуют и другие типы, но они встречаются реже (Grube and Hawksworth, 2007). Внутренне вегетативное тело бывает либо гомойомерным (без расслоения), когда микобионт и фотобионт равномерно распределены в слоевищах лишайников, либо гетеромерным (с расслоением), где можно выделить, по крайней мере, грибной верхний слой и водорослевый слой под ним. Для корковых лишайников характерно прикрепление к субстрату всей нижней поверхностью, тогда как листоватые и кустистые лишайники прикрепляются лишь частично (Büdel, Scheidegger, 2008) и обычно имеют более или менее плотный нижний грибной слой.Половое размножение грибного партнера требует развития видоспецифического слоевища с соответствующими водорослями, поскольку плодовые тела грибов возникают непосредственно в слоевищах зрелых лишайников и часто включают структуры слоевищ. Тем не менее, лишайники также развили различные способы бесполого размножения, чтобы рассредоточить симбиотических партнеров вместе в разнообразных и специфических суставных побегах (Büdel and Scheidegger, 2008).
Существуют и другие типы, но они встречаются реже (Grube and Hawksworth, 2007). Внутренне вегетативное тело бывает либо гомойомерным (без расслоения), когда микобионт и фотобионт равномерно распределены в слоевищах лишайников, либо гетеромерным (с расслоением), где можно выделить, по крайней мере, грибной верхний слой и водорослевый слой под ним. Для корковых лишайников характерно прикрепление к субстрату всей нижней поверхностью, тогда как листоватые и кустистые лишайники прикрепляются лишь частично (Büdel, Scheidegger, 2008) и обычно имеют более или менее плотный нижний грибной слой.Половое размножение грибного партнера требует развития видоспецифического слоевища с соответствующими водорослями, поскольку плодовые тела грибов возникают непосредственно в слоевищах зрелых лишайников и часто включают структуры слоевищ. Тем не менее, лишайники также развили различные способы бесполого размножения, чтобы рассредоточить симбиотических партнеров вместе в разнообразных и специфических суставных побегах (Büdel and Scheidegger, 2008).
Несмотря на то, что в литературе продолжают сообщать об антибактериальных или противогрибковых соединениях из лишайников (обзор в Boustie and Grube, 2005), долгоживущие талломы обеспечивают интересную микросреду обитания для других эукариотических и прокариотических (как бактерий, так и архей) микроорганизмов (Lawrey and Diederich, 2003; Grube and Berg, 2009; Bjelland et al., 2011; Бейтс и др., 2012). В предыдущие годы все больше внимания уделялось лихеноассоциированным бактериям, которые не считались неотъемлемой частью симбиоза.
В этом обзоре мы обсуждаем недавнюю литературу по микробиоте, связанной с лишайниками, уделяя особое внимание разнообразию, функциям, распространению, специфичности среды обитания и межмикробиомным отношениям Lobaria pulmonaria -ассоциированного бактериального сообщества, и завершаем кратким описанием, чтобы способствовать целостному взгляду лишайниково-бактериальные взаимодействия.В первой части мы рассматриваем исторические аспекты, а затем обсуждаем недавние результаты, чтобы разработать более целостную модель лишайников.
Разгадка связанного с лишайниками микробиома – тогда и сейчас
Бактерии, ассоциированные с лишайниками, впервые упоминаются в первой половине 20 века (Uphof, 1925; Henkel, Yuzhakova, 1936; Искина, 1938). В ходе этих ранних исследований различные бактериальные розы были связаны с лишайниками, такими как Azotobacter, Pseudomonas ( GammaProteobacteria ), Beijerinckia ( AlphaProteobacteria ), а также 70144 и Bacillus и Clostridium и Искина, 1938; Паносян, Никогосян, 1966; Хенкель, Плотникова, 1973).В то время описания бактерий лежали в основе исключительно фенотипических и физиологических характеристик, указывающих на возможную роль некоторых из этих бактерий в фиксации азота. Тем не менее, Ленова и Блюм (1983) уже подсчитали, что миллионы бактериальных клеток на грамм могут колонизировать слоевище лишайника. Прошло несколько десятилетий, прежде чем первые молекулярные анализы начали использовать бактериальные изоляты (например, González et al.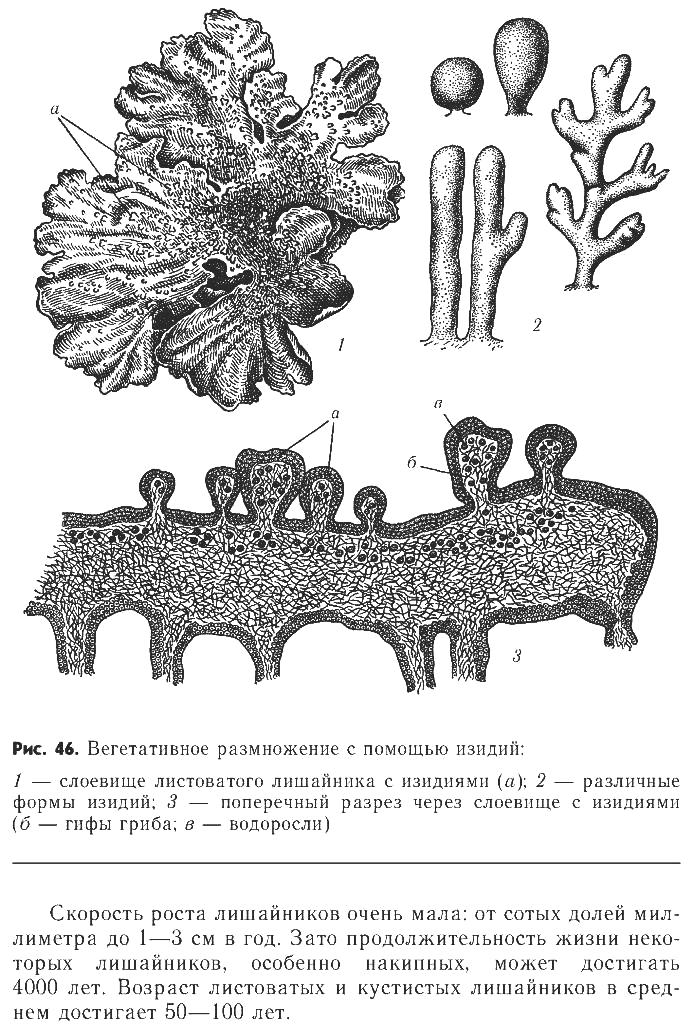 , 2005; Cardinale et al., 2006, Liba et al., 2006 или Selbmann et al., 2009).Хотя Гонсалес и соавт. (2005) сосредоточились только на пригодных для культивирования Actinomycetes (с преобладающими родами Micromonospora и Streptomyces ) различных видов лишайников из тропических и холодных регионов, Cardinale et al. (2006) попытались описать общий состав бактериального сообщества, связанного с семью различными видами лишайников из местообитаний умеренного пояса. Последнее позволило идентифицировать несколько родов, связанных с Firmicutes , Actinobacteria и Proteobacteria , выделив Paenibacillus и Burkholderia как вездесущие роды лишайников.Однако методы, зависящие от культуры, охватывают только 0,001–15% бактериального разнообразия в пробах окружающей среды (Amann et al., 1995), в то время как большая часть остается ненаблюдаемой (Rappé and Giovannoni, 2003). Чтобы преодолеть ограничения селективного выделения бактерий из образцов окружающей среды и получить более беспристрастное и менее ограниченное представление о микробных сообществах, в дополнение к традиционным методам были использованы новые методы.
, 2005; Cardinale et al., 2006, Liba et al., 2006 или Selbmann et al., 2009).Хотя Гонсалес и соавт. (2005) сосредоточились только на пригодных для культивирования Actinomycetes (с преобладающими родами Micromonospora и Streptomyces ) различных видов лишайников из тропических и холодных регионов, Cardinale et al. (2006) попытались описать общий состав бактериального сообщества, связанного с семью различными видами лишайников из местообитаний умеренного пояса. Последнее позволило идентифицировать несколько родов, связанных с Firmicutes , Actinobacteria и Proteobacteria , выделив Paenibacillus и Burkholderia как вездесущие роды лишайников.Однако методы, зависящие от культуры, охватывают только 0,001–15% бактериального разнообразия в пробах окружающей среды (Amann et al., 1995), в то время как большая часть остается ненаблюдаемой (Rappé and Giovannoni, 2003). Чтобы преодолеть ограничения селективного выделения бактерий из образцов окружающей среды и получить более беспристрастное и менее ограниченное представление о микробных сообществах, в дополнение к традиционным методам были использованы новые методы.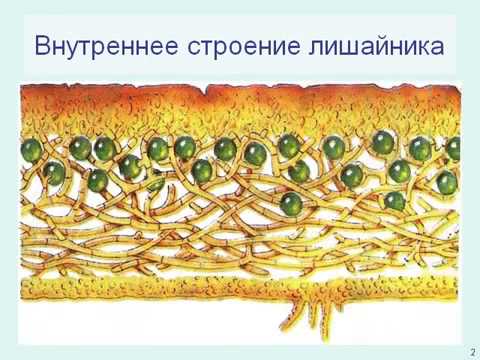
Первые независимые от культуры исследования микробиоты, связанной с лишайниками, были оценены с использованием различных методов фингерпринтинга (Cardinale et al., 2006; Грубе и др., 2009 г.; Бьелланд и др., 2011; Мушегян и др., 2011; Cardinale et al., 2012a) и методы молекулярного клонирования (Hodkinson and Lutzoni, 2009). Такие методы (например, DGGE: Muyzer and Smalla, 1998; T-RFLP: Liu et al., 1997; SSCP: Schwieger and Tebbe, 1998) использовались для создания профилей микробного сообщества путем амплификации генетических маркеров (например, 16S рибосомной ДНК). с универсальными грунтовками. На основе полиморфизма последовательности или длины продукты ПЦР разделяются, и может быть охарактеризована степень сходства образцов в соответствии с конкретными образцами полос (Smalla et al., 2007). Хотя многие образцы можно анализировать параллельно и их профили можно легко сравнивать друг с другом, детальная идентификация членов бактериального сообщества утомительна и ограничена.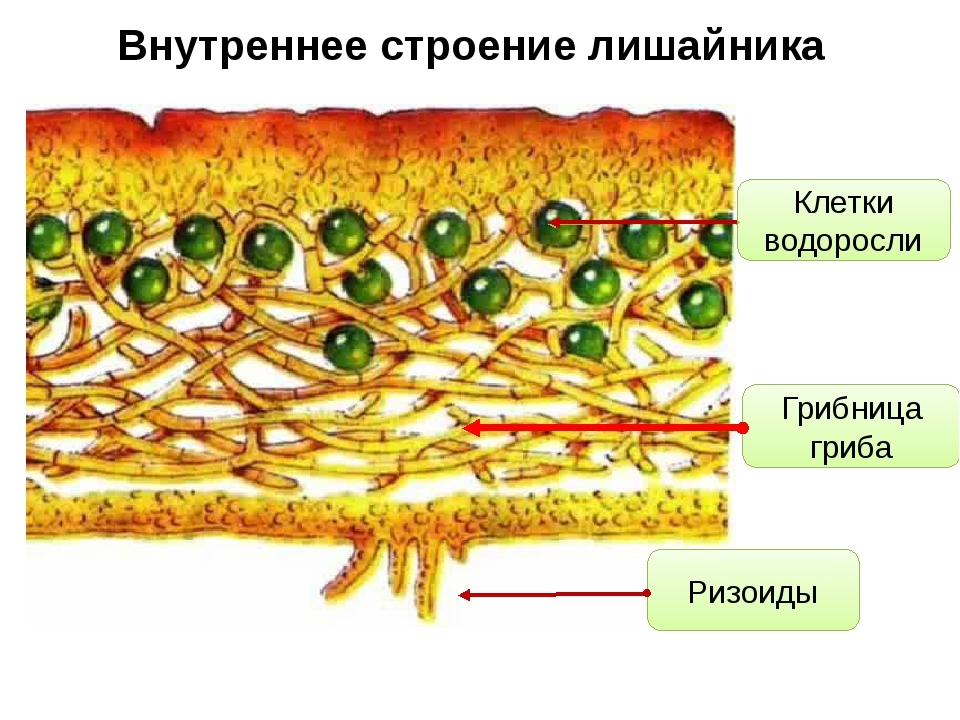 Маргулис и др. (2005) представили новую экономичную и сокращающую время технологию для глубокого изучения состава сообществ и разнообразия проб окружающей среды с помощью крупномасштабного высокопроизводительного секвенирования. Бейтс и др. (2011) впервые описали бактерии, связанные с лишайниками, на основе этой технологии пиросеквенирования нового поколения, за которой последовали Grube et al.(2012), Ходкинсон и др. (2012) и Aschenbrenner et al. (2014).
Маргулис и др. (2005) представили новую экономичную и сокращающую время технологию для глубокого изучения состава сообществ и разнообразия проб окружающей среды с помощью крупномасштабного высокопроизводительного секвенирования. Бейтс и др. (2011) впервые описали бактерии, связанные с лишайниками, на основе этой технологии пиросеквенирования нового поколения, за которой последовали Grube et al.(2012), Ходкинсон и др. (2012) и Aschenbrenner et al. (2014).
С усовершенствованием технологий секвенирования и инструментов биоинформатики акцент в исследованиях микробной экологии сместился с базовых таксономических описаний на более подробное и целостное представление о микробных сообществах. Метагеномный, транскриптомный и протеомный анализ теперь могут пролить свет на вопросы «Кто там?», «На что они способны?» и «Кто чем активно занимается?» (Schneider et al., 2011; Aschenbrenner, 2015; Grube et al., 2015). Для решения этих вопросов легочный лишай L. pulmonaria (L.) Hoffm.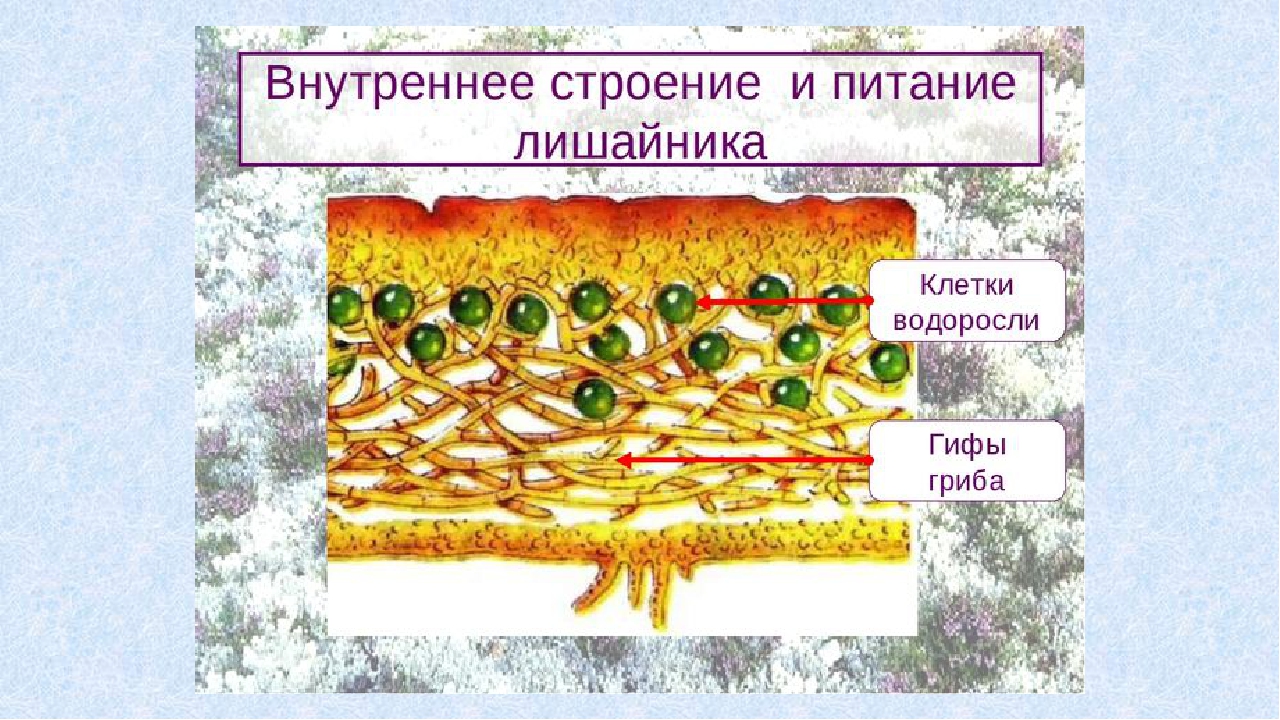 была использована в качестве модельной системы из-за ее относительно быстрого роста и других благоприятных характеристик, например, эпифитного роста на коре деревьев и небольшого количества вторичных метаболитов, которые могли мешать проводимым анализам. L. pulmonaria характеризуется листовидной структурой (листовой лишайник) и в основном встречается в старовозрастных лесах с незагрязненным воздухом. Его чувствительность к загрязнению воздуха можно использовать для косвенной оценки качества воздуха и целостности экосистемы (Scheidegger and Werth, 2009).У него есть два партнера по фотосинтезу, явление, наблюдаемое примерно у 4% всех описанных лишайников (Honegger, 1991). Однако только зеленая водоросль Dictyochloropsis reticulata образует непрерывный слой, тогда как штаммы цианобактерий Nostoc сохраняются в разнесенных узелкообразных внутренних компартментах (цефалодиях).
была использована в качестве модельной системы из-за ее относительно быстрого роста и других благоприятных характеристик, например, эпифитного роста на коре деревьев и небольшого количества вторичных метаболитов, которые могли мешать проводимым анализам. L. pulmonaria характеризуется листовидной структурой (листовой лишайник) и в основном встречается в старовозрастных лесах с незагрязненным воздухом. Его чувствительность к загрязнению воздуха можно использовать для косвенной оценки качества воздуха и целостности экосистемы (Scheidegger and Werth, 2009).У него есть два партнера по фотосинтезу, явление, наблюдаемое примерно у 4% всех описанных лишайников (Honegger, 1991). Однако только зеленая водоросль Dictyochloropsis reticulata образует непрерывный слой, тогда как штаммы цианобактерий Nostoc сохраняются в разнесенных узелкообразных внутренних компартментах (цефалодиях).
Состав и разнообразие микробиома лишайников, обусловленное различными абиотическими и биотическими факторами
Количество бактерий, обнаруженных на лишайниках, удивительно велико по сравнению с поверхностями листвы высших растений. В то время как поверхность листа содержит всего 10 90 609 5 90 610 клеток/см 90 609 2 90 610 , некоторые виды лишайников, проанализированные на обилие бактерий, значительно превышают это значение (Saleem, 2015). Например, Cladonia rangiferina колонизируется приблизительно 10 7 –10 8 бактериями на грамм слоевища лишайника (Cardinale et al., 2008; Grube et al., 2009). Кроме того, было показано, что индексы альфа-разнообразия (индекс Шеннона) бактериальных сообществ различаются между разными лишайниками, например., в среднем от 4,5 ( Solorina crocea ) до 7,0 ( L. pulmonaria ) при генетическом расстоянии 3% среди микробных OTU на основе несходства последовательности гена 16S рРНК (Grube et al., 2012; Aschenbrenner et al. , 2014).
В то время как поверхность листа содержит всего 10 90 609 5 90 610 клеток/см 90 609 2 90 610 , некоторые виды лишайников, проанализированные на обилие бактерий, значительно превышают это значение (Saleem, 2015). Например, Cladonia rangiferina колонизируется приблизительно 10 7 –10 8 бактериями на грамм слоевища лишайника (Cardinale et al., 2008; Grube et al., 2009). Кроме того, было показано, что индексы альфа-разнообразия (индекс Шеннона) бактериальных сообществ различаются между разными лишайниками, например., в среднем от 4,5 ( Solorina crocea ) до 7,0 ( L. pulmonaria ) при генетическом расстоянии 3% среди микробных OTU на основе несходства последовательности гена 16S рРНК (Grube et al., 2012; Aschenbrenner et al. , 2014).
L. pulmonaria в основном колонизированы Alphaproteobacteria с преобладающим отрядом Sphingomonadales , за которым следуют Sphingobacteria , Actinobacteria и Spartobacteria 9014, 2014). Напротив, исследования, основанные на секвенировании дробовиком, предполагают, что Rhizobiales являются основным отрядом в пределах Alphaproteobacteria (Erlacher et al., 2015; Grube et al., 2015). Эти результаты были дополнительно подтверждены с помощью адаптированных методов визуализации. Таким образом, было показано преобладание Alphaproteobacteria и Rhizobiales на поверхности лишайников с помощью комбинированного подхода флуоресцентной гибридизации in situ (FISH) и конфокальной лазерной сканирующей микроскопии (CLSM).В связи с этими выводами сообщалось, что связанная с лишайниками группа Rhizobiales (LAR1) представляет собой специфичную для лишайников линию Alphaproteobacteria , которую можно обнаружить среди многих исследованных видов (Hodkinson and Lutzoni, 2009; Bates et al., 2011; Ходкинсон и др., 2012). Однако эту линию не удалось обнаружить у L. pulmonaria (Aschenbrenner et al., 2014). Наблюдаемые различия в составе одного и того же вида лишайников можно объяснить различными причинами, такими как подход к метагеномному секвенированию (ампликон vs.
Напротив, исследования, основанные на секвенировании дробовиком, предполагают, что Rhizobiales являются основным отрядом в пределах Alphaproteobacteria (Erlacher et al., 2015; Grube et al., 2015). Эти результаты были дополнительно подтверждены с помощью адаптированных методов визуализации. Таким образом, было показано преобладание Alphaproteobacteria и Rhizobiales на поверхности лишайников с помощью комбинированного подхода флуоресцентной гибридизации in situ (FISH) и конфокальной лазерной сканирующей микроскопии (CLSM).В связи с этими выводами сообщалось, что связанная с лишайниками группа Rhizobiales (LAR1) представляет собой специфичную для лишайников линию Alphaproteobacteria , которую можно обнаружить среди многих исследованных видов (Hodkinson and Lutzoni, 2009; Bates et al., 2011; Ходкинсон и др., 2012). Однако эту линию не удалось обнаружить у L. pulmonaria (Aschenbrenner et al., 2014). Наблюдаемые различия в составе одного и того же вида лишайников можно объяснить различными причинами, такими как подход к метагеномному секвенированию (ампликон vs. секвенирование дробовика), используемые базы данных или активность бактерий в случае метатранскриптомного анализа (Aschenbrenner, 2015), поскольку одновременно метаболически активны менее 10% микробного сообщества (Locey, 2010).
секвенирование дробовика), используемые базы данных или активность бактерий в случае метатранскриптомного анализа (Aschenbrenner, 2015), поскольку одновременно метаболически активны менее 10% микробного сообщества (Locey, 2010).
Хотя о преобладании Alphaproteobacteria также сообщалось в других исследованиях (Bates et al., 2011; Hodkinson et al., 2012), состав бактериального сообщества в целом различался среди видов лишайников. Предполагается, что эти изменения обусловлены различными биотическими и абиотическими факторами.Ходкинсон и др. (2012), которые тщательно изучили бактериальные сообщества, связанные с различными видами лишайников, включающими 24 типа микобионтов со всеми комбинациями фотобионтов из разных мест отбора проб (тропические и арктические регионы), выделили тип фотобионтов (хлоролишайники против цианолишайников) и крупномасштабную географию в качестве основного. Движущие силы.
Ходкинсон и др. (2012) утверждали, что различия в составе сообществ могут быть связаны как с наличием связанного азота, так и с типом связанного углерода.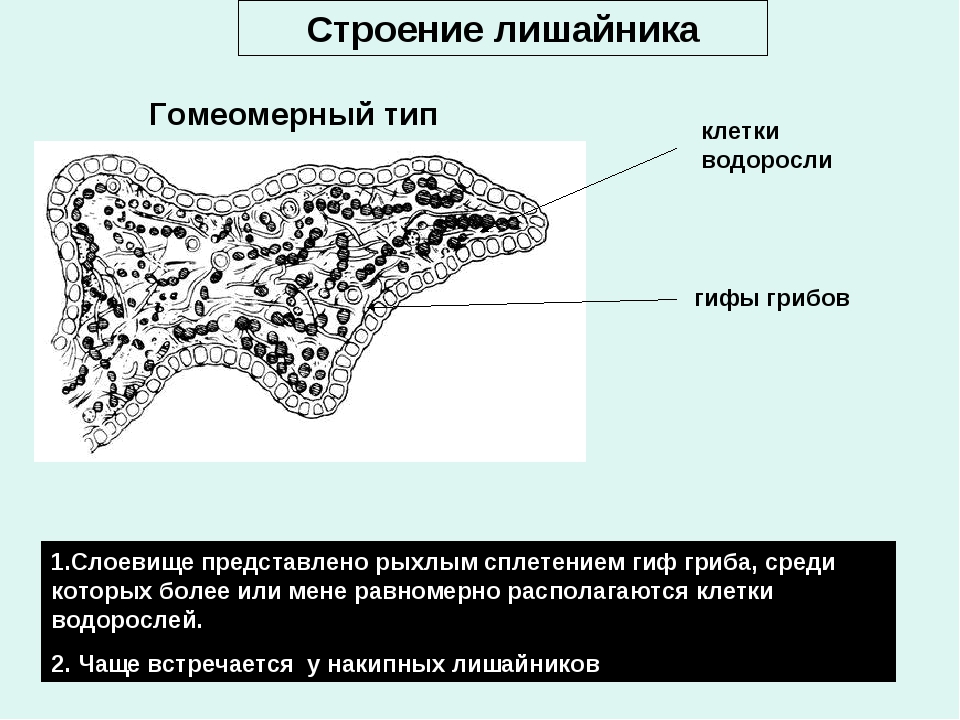 Что касается первого, бактерии, связанные с цианолишайниками, имеют доступ к фиксированному атмосферному азоту из-за цианобактериального фотобионта, тогда как бактерии хлоролишайников лишены этого преимущества в среде с ограниченным азотом. В соответствии с этим хлоролишайники предпочтительно обогащали бы виды, способные к фиксации азота, а не цианолишайники. Другое предположение заключалось в том, что зеленые водоросли выделяют другие типы связанного углерода (сахароспирты: рибит, эритрит или сорбит), чем цианобактерии (глюкоза; Elix and Stocker-Wörgötter, 2008), тем самым формируя бактериальное сообщество в отношении использования углерода.Оба объяснения могут лишь частично объяснить различия в сообществах, основанные на таксономических описаниях, поскольку бактерии могут обмениваться и совместно использовать гены, кодирующие определенные функции, посредством горизонтального переноса генов. Это согласуется с Burke et al. (2011), которые утверждали, что экологические ниши случайным образом заселяются бактериями, наделенными подходящими функциями, а не в соответствии с бактериальной таксономией.
Что касается первого, бактерии, связанные с цианолишайниками, имеют доступ к фиксированному атмосферному азоту из-за цианобактериального фотобионта, тогда как бактерии хлоролишайников лишены этого преимущества в среде с ограниченным азотом. В соответствии с этим хлоролишайники предпочтительно обогащали бы виды, способные к фиксации азота, а не цианолишайники. Другое предположение заключалось в том, что зеленые водоросли выделяют другие типы связанного углерода (сахароспирты: рибит, эритрит или сорбит), чем цианобактерии (глюкоза; Elix and Stocker-Wörgötter, 2008), тем самым формируя бактериальное сообщество в отношении использования углерода.Оба объяснения могут лишь частично объяснить различия в сообществах, основанные на таксономических описаниях, поскольку бактерии могут обмениваться и совместно использовать гены, кодирующие определенные функции, посредством горизонтального переноса генов. Это согласуется с Burke et al. (2011), которые утверждали, что экологические ниши случайным образом заселяются бактериями, наделенными подходящими функциями, а не в соответствии с бактериальной таксономией. Попытка объяснить наблюдаемый состав сообществ становится более сложной в отношении трехраздельных лишайников, поскольку они несут оба типа фотобионтов, как это имеет место в случае с L.легочная .
Попытка объяснить наблюдаемый состав сообществ становится более сложной в отношении трехраздельных лишайников, поскольку они несут оба типа фотобионтов, как это имеет место в случае с L.легочная .
Видоспецифичность бактериальных сообществ, связанных с хлорлишайниками, уже указывалась в предыдущих исследованиях (Grube et al., 2009; Bates et al., 2011). Лихенизированные грибы способны продуцировать вторичные метаболиты, уникальные для лишайников и состоящие из нескольких сотен соединений, которые могут откладываться на внеклеточной поверхности гиф грибов (Elix and Stocker-Wörgötter, 2008). Как уже было предложено Hodkinson et al. (2012) значительная доля вторичных метаболитов с антимикробной активностью (Kosanić and Ranković, 2015) также может оказывать селективное давление на бактерии, колонизирующие лишайники.Однако, поскольку L. pulmonaria , как и многие другие лишайники подотряда Peltigerineae , содержит только низкие концентрации специфических для лишайников веществ (Beckett et al. , 2003), вторичные метаболиты могут играть лишь незначительную роль в формировании структуры сообщества лишайников. Lobaria -ассоциированные бактерии.
, 2003), вторичные метаболиты могут играть лишь незначительную роль в формировании структуры сообщества лишайников. Lobaria -ассоциированные бактерии.
Различия в составе бактериального сообщества также могут быть связаны с типом роста лишайника, поскольку, например, в предыдущих исследованиях сообщалось, что состав бактериального сообщества накипных лишайников отличается от такового у листоватых или кустистых лишайников (Grube et al., 2009; Ходкинсон и др., 2012). В то время как листоватые лишайники были в основном колонизированы Alphaproteobacteria , накипный лишайник Ophioparma sp. доминировали Acidobacteria (Hodkinson et al., 2012). Другой наскальный накипный лишайник Hydropunctaria sp. был в основном колонизирован Cyanobacteria , Actinobacteria и Deinococcus (Bjelland et al., 2011). Но сам по себе тип роста не объясняет преобладания отдельных таксонов, поскольку листоватый лишайник Solorina sp.также доминировали Acidobacteria (Grube et al. , 2012). Это согласуется с предыдущими результатами Cardinale et al. (2012b), которые показали, что типы роста не влияют на основную структуру бактериального сообщества.
, 2012). Это согласуется с предыдущими результатами Cardinale et al. (2012b), которые показали, что типы роста не влияют на основную структуру бактериального сообщества.
Бактерии пространственно структурированы на лишайниках
Субкомпартменты таллома разного возраста, а также внешняя и внутренняя поверхности образуют химически и физиологически различные микрониши и способствуют формированию различных бактериальных сообществ.На основе FISH и CLSM было продемонстрировано, что эубактерии, связанные с лишайниками, а также определенные бактериальные таксоны в них колонизируют отдельные части слоевища лишайников в разной численности и по разной схеме (Cardinale et al., 2008). Конфокальная лазерная сканирующая микроскопия поверхностей L. pulmonaria показала, что как верхняя, так и нижняя кора были равномерно колонизированы Alphaproteobacteria среди других эубактерий (Cardinale et al., 2012a; Grube et al., 2015). Это также было продемонстрировано для других дорсивентрально организованных слоевищ лишайников, таких как листовые Umbilicaria sp. (Грубе и др., 2009). В случае кустарникового вида Cladonia внешняя кора радиально организованного полого слоевища (подеция) была заселена лишь одиночными колониями клеток и более мелкими скоплениями колоний, в то время как наибольшая бактериальная плотность, исследованная на этом лишайнике, была обнаружена во внутреннем слое подеции образуют биопленкообразную оболочку (Cardinale et al., 2008, 2012b). Напротив, бактериальная колонизация на корковых лишайниках, таких как Lecanora sp. был отчетливо выше в щелях между ареолами слоевищ (Grube et al., 2009). Были также первые указания на наличие эндобиотических бактерий в клеточных стенках гиф грибов (Cardinale et al., 2008). Эрлахер и др. (2015) ранее сообщали о L. pulmonaria эндосимбиотических Rhizobiales , локализованных на различной глубине межгифального студенистого матрикса верхней коры и редко внутри гиф грибов. До сих пор нет документальных подтверждений роста бактерий в других компартментах L. pulmonaria , таких как внутренняя таллиновая ткань (мозговое вещество) или слой фотобионта.
(Грубе и др., 2009). В случае кустарникового вида Cladonia внешняя кора радиально организованного полого слоевища (подеция) была заселена лишь одиночными колониями клеток и более мелкими скоплениями колоний, в то время как наибольшая бактериальная плотность, исследованная на этом лишайнике, была обнаружена во внутреннем слое подеции образуют биопленкообразную оболочку (Cardinale et al., 2008, 2012b). Напротив, бактериальная колонизация на корковых лишайниках, таких как Lecanora sp. был отчетливо выше в щелях между ареолами слоевищ (Grube et al., 2009). Были также первые указания на наличие эндобиотических бактерий в клеточных стенках гиф грибов (Cardinale et al., 2008). Эрлахер и др. (2015) ранее сообщали о L. pulmonaria эндосимбиотических Rhizobiales , локализованных на различной глубине межгифального студенистого матрикса верхней коры и редко внутри гиф грибов. До сих пор нет документальных подтверждений роста бактерий в других компартментах L. pulmonaria , таких как внутренняя таллиновая ткань (мозговое вещество) или слой фотобионта.
Возрастные состояния в зрелом слоевище лишайника могут влиять и формировать структуру бактериального сообщества, которая напоминает сукцессию сообщества, обнаруженную, например, в микробиоме цветков яблони (Shade et al., 2013). Недавнее исследование показало, что вегетативные пропагулы L. pulmonaria были колонизированы более отчетливым бактериальным сообществом, чем зрелое слоевище лишайника (Aschenbrenner et al., 2014), что указывает на то, что структура сообщества может меняться со временем. В частности, только 37% бактериальных OTU, связанных со слоевищем, были общими с вегетативными побегами, и наоборот, общие OTU, связанные с побегами, составляли 55%.В то время как обе части лишайника были в основном колонизированы Alphaproteobacteria , в талломе лишайника дополнительно доминировали Deltaproteobacteria , тогда как ювенильные вегетативные пропагулы также были колонизированы в большем количестве Spartobacteria и Sphingobacteria . Ранее Cardinale et al. (2012b) сообщили, что более старые части слоевища содержали значительно большее количество бактерий, чем более молодые структуры слоевища, включая замену преобладающих Alphaproteobacteria на другие таксоны, такие как Actinobacteria , Gamma — и Betaproteobacteria .Также Mushegian et al. (2011) наблюдали пространственную диверсификацию бактериального состава между более разнообразными и последовательными центрами таллома (старые части) и центрами более изменчивых и бедных видами краев (молодые части). Кардинале и др. (2012b) назвали эти модели распределения бактерий анаболическими центрами в растущих и катаболическими поглотителями в стареющих частях слоевища лишайника соответственно. Гипотеза о переработке питательных веществ в разлагающихся частях лишайника бактериями также может быть подкреплена присутствием определенных таксонов, известных своим потенциалом деградации. Sphingomonas sp., которые, как известно, разлагают органические вещества и ксенобиотические вещества, ранее были выделены из лишайников, отобранных в арктических и антарктических регионах (Lee et al.
Ранее Cardinale et al. (2012b) сообщили, что более старые части слоевища содержали значительно большее количество бактерий, чем более молодые структуры слоевища, включая замену преобладающих Alphaproteobacteria на другие таксоны, такие как Actinobacteria , Gamma — и Betaproteobacteria .Также Mushegian et al. (2011) наблюдали пространственную диверсификацию бактериального состава между более разнообразными и последовательными центрами таллома (старые части) и центрами более изменчивых и бедных видами краев (молодые части). Кардинале и др. (2012b) назвали эти модели распределения бактерий анаболическими центрами в растущих и катаболическими поглотителями в стареющих частях слоевища лишайника соответственно. Гипотеза о переработке питательных веществ в разлагающихся частях лишайника бактериями также может быть подкреплена присутствием определенных таксонов, известных своим потенциалом деградации. Sphingomonas sp., которые, как известно, разлагают органические вещества и ксенобиотические вещества, ранее были выделены из лишайников, отобранных в арктических и антарктических регионах (Lee et al.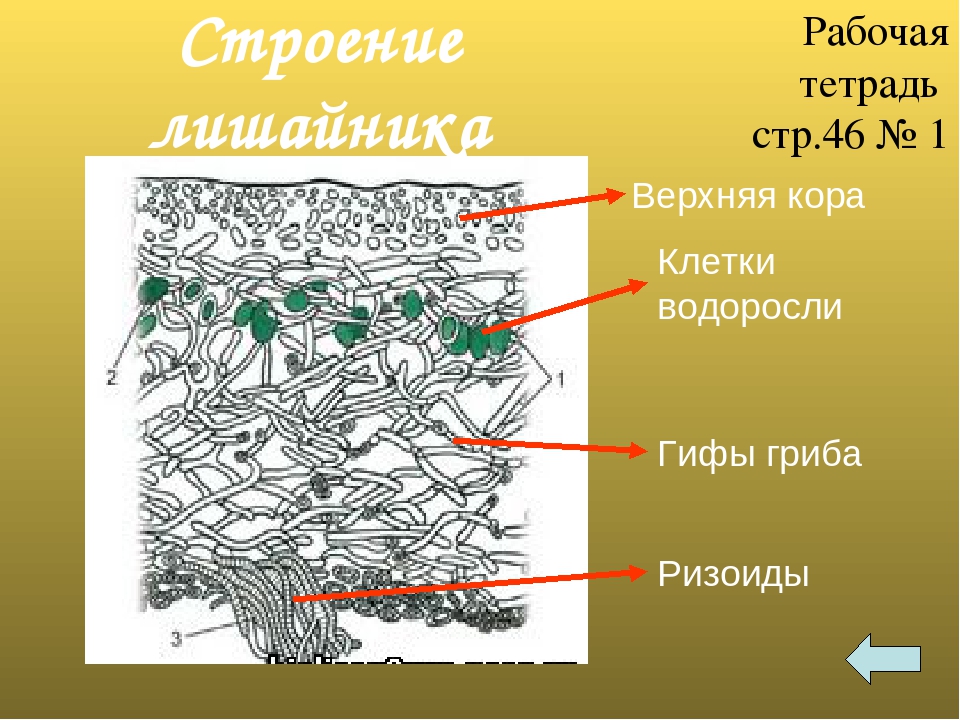 , 2014), но о них также сообщалось в других исследованиях (Grube et al., 2009). , 2012; Hodkinson et al., 2012; Aschenbrenner, 2015). Однако другие роды, такие как Paenibacillus и Streptomyces , упоминались в связи с их функциями (например, хитинолитической активностью) в деградации тканей лишайника (Cardinale et al., 2006).
, 2014), но о них также сообщалось в других исследованиях (Grube et al., 2009). , 2012; Hodkinson et al., 2012; Aschenbrenner, 2015). Однако другие роды, такие как Paenibacillus и Streptomyces , упоминались в связи с их функциями (например, хитинолитической активностью) в деградации тканей лишайника (Cardinale et al., 2006).
Распространение и перенос бактерий, ассоциированных с хозяином
Анализ лишайников-ассоциированных бактерий выявил различия в составе и разнообразии сообществ между географически удаленными местообитаниями (Printzen et al., 2012; Aschenbrenner et al., 2014). Принцен и др. (2012) проанализировали географическую структуру связанных с лишайниками Alphaproteobacteria в антарктических регионах, указав, что на эту группу влияют параметры окружающей среды, поскольку слоевища из субполярных местообитаний имели более сходные сообщества, чем сообщества из внеполярных регионов.Ходкинсон и др. (2012) объяснили эти крупномасштабные географические эффекты эффективностью распространения лишайников-хозяев, когда распространение происходит в небольших пространственных масштабах, а не на больших расстояниях, что приводит к географической дифференциации состава сообщества. Ашенбреннер и др. (2014) визуализировали и описали бактериальную колонизацию пропагул лишайника. Их результаты показывают, что по крайней мере определенная часть микробиома лишайников передается вертикально через эти симбиотические структуры.В этих бактериальных сообществах доминировали Alphaproteobacteria , как уже было обнаружено Cardinale et al. (2012а). Интересно, что бактериальные консорциумы пропагул лишайника представляли собой нечто большее, чем просто подмножество микробиома родительского таллома, а также включали уникальные виды, отсутствующие в зрелом талломе. Так, Aschenbrenner et al. (2014) предположили, что вегетативные пропагулы снабжены стартовым бактериальным сообществом. Такие бактерии, колонизирующие ювенильные структуры, могут влиять на последующее пополнение новых бактерий (Fukami, 2010), тем самым формируя состав сообщества.О важности ассоциированных с лишайниками бактерий при установлении лишайникового симбиоза уже предполагалось (Hodkinson, Lutzoni, 2009), так как рост многослойных слоевищ лишайников до сих пор был успешным только в культурах, основанных на фрагментах лишайников, которые, по-видимому, включают бактерии.
Ашенбреннер и др. (2014) визуализировали и описали бактериальную колонизацию пропагул лишайника. Их результаты показывают, что по крайней мере определенная часть микробиома лишайников передается вертикально через эти симбиотические структуры.В этих бактериальных сообществах доминировали Alphaproteobacteria , как уже было обнаружено Cardinale et al. (2012а). Интересно, что бактериальные консорциумы пропагул лишайника представляли собой нечто большее, чем просто подмножество микробиома родительского таллома, а также включали уникальные виды, отсутствующие в зрелом талломе. Так, Aschenbrenner et al. (2014) предположили, что вегетативные пропагулы снабжены стартовым бактериальным сообществом. Такие бактерии, колонизирующие ювенильные структуры, могут влиять на последующее пополнение новых бактерий (Fukami, 2010), тем самым формируя состав сообщества.О важности ассоциированных с лишайниками бактерий при установлении лишайникового симбиоза уже предполагалось (Hodkinson, Lutzoni, 2009), так как рост многослойных слоевищ лишайников до сих пор был успешным только в культурах, основанных на фрагментах лишайников, которые, по-видимому, включают бактерии.
Хотя вертикальная передача ассоциированных с лишайниками бактерий была показана только у одного вида лишайников, весьма вероятно, что эта стратегия переноса микробиома также распространена у других видов, использующих вегетативные диаспоры для размножения, и определенно в других симбиозах.Имеются различные примеры передачи ассоциированных с хозяином бактерий (Bright and Bulgheresi, 2010), например, у морских губок (Wilkinson, 1984; Li et al., 1998). Известно, что бактерии, связанные с наземными беспозвоночными, такими как насекомые, способствуют поглощению питательных веществ и обеспечению незаменимыми аминокислотами и витаминами (Douglas, 1998; Feldhaar and Gross, 2009), но их стратегии вертикальной передачи различаются у разных видов (Sacchi et al., 1988; Аттардо и др., 2008; Прадо и Цукки, 2012).Сообщалось, что у позвоночных, включая человека, передача материнских микробов ребенку при естественных родах и грудном вскармливании в качестве первого инокулята важна для здоровья ребенка, в частности, путем формирования структуры микробиома с полезными микробами (Funkhouser and Bordenstein, 2013).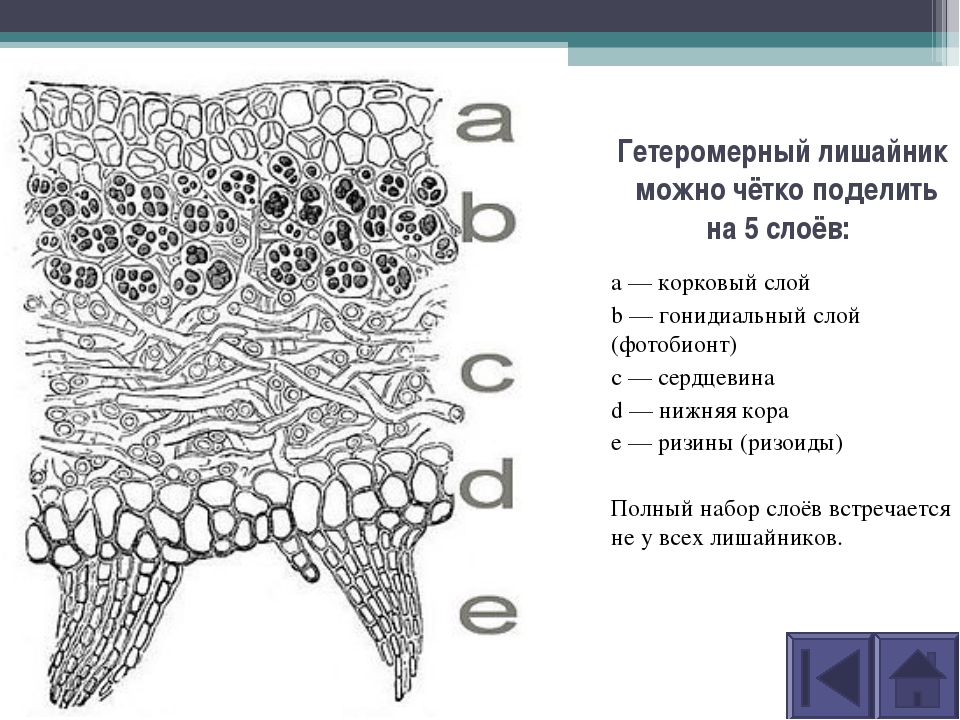 Но и в царстве растений сообщалось о переносе ассоциированных с растениями бактерий, в частности семян, от материнского растения (van Overbeek et al., 2011), хотя высшие растения обычно набирают свои существенные ризосферные сообщества из материнского растения. окружающая почва (Berg and Smalla, 2009).Ранее вертикальная передача была показана для древнейшей группы наземных растений — мхов, входящих вместе с лишайниками в группу пойкилогидных криптогам; ассоциированные бактерии, особенно специфические штаммы Burkholderia , переносятся со спорофита на гаметофит через споры (Брагина и др., 2012, 2013).
Но и в царстве растений сообщалось о переносе ассоциированных с растениями бактерий, в частности семян, от материнского растения (van Overbeek et al., 2011), хотя высшие растения обычно набирают свои существенные ризосферные сообщества из материнского растения. окружающая почва (Berg and Smalla, 2009).Ранее вертикальная передача была показана для древнейшей группы наземных растений — мхов, входящих вместе с лишайниками в группу пойкилогидных криптогам; ассоциированные бактерии, особенно специфические штаммы Burkholderia , переносятся со спорофита на гаметофит через споры (Брагина и др., 2012, 2013).
Лишайники как бактериальные концентраторы
Лишайники являются пионерами в колонизации враждебных сред с экстремальными температурами, высыханием и высокой соленостью, но они также могут становиться очень старыми, либо поодиночке, либо как ассоциации (предполагается, что некоторые не покрытые льдом участки были заселены лишайниками со времен третичный).Колонизированные местообитания включают засушливые и полузасушливые районы, где голая почва может быть заселена, например, криптогамными почвенными корками (ассоциация, включающая частицы почвы, лишайники, цианобактерии, водоросли, грибы и мохообразные; Beckett et al. , 2008), но также более экстремальные регионы, такие как пустыни, где лишайники являются одними из немногих успешных колонизаторов. В частности, их способность гидратироваться без контакта с жидкой водой (Printzen et al., 2012) только при тумане, росе или высокой влажности воздуха (Beckett et al., 2008) обеспечивает выживание в этих засушливых районах. Это говорит о том, что лишайники как медленно растущие и долгоживущие организмы-хозяева могут служить бактериальными центрами в этих средах, способствуя их выживанию за счет снабжения питательными веществами и водой, предлагая среду обитания с различными микронишами и обеспечивая их распространение на короткие расстояния за счет стратегий расселения. хозяина лишайника. Таким образом, лишайники могут быть важными источниками/резервуарами полезных бактериальных штаммов и для других местообитаний в окружающей среде.
, 2008), но также более экстремальные регионы, такие как пустыни, где лишайники являются одними из немногих успешных колонизаторов. В частности, их способность гидратироваться без контакта с жидкой водой (Printzen et al., 2012) только при тумане, росе или высокой влажности воздуха (Beckett et al., 2008) обеспечивает выживание в этих засушливых районах. Это говорит о том, что лишайники как медленно растущие и долгоживущие организмы-хозяева могут служить бактериальными центрами в этих средах, способствуя их выживанию за счет снабжения питательными веществами и водой, предлагая среду обитания с различными микронишами и обеспечивая их распространение на короткие расстояния за счет стратегий расселения. хозяина лишайника. Таким образом, лишайники могут быть важными источниками/резервуарами полезных бактериальных штаммов и для других местообитаний в окружающей среде.
Специфика среды обитания
Специфичность хозяина для криптогам (то есть лишайников и мхов) уже сообщалась в предыдущих независимых исследованиях (Grube et al.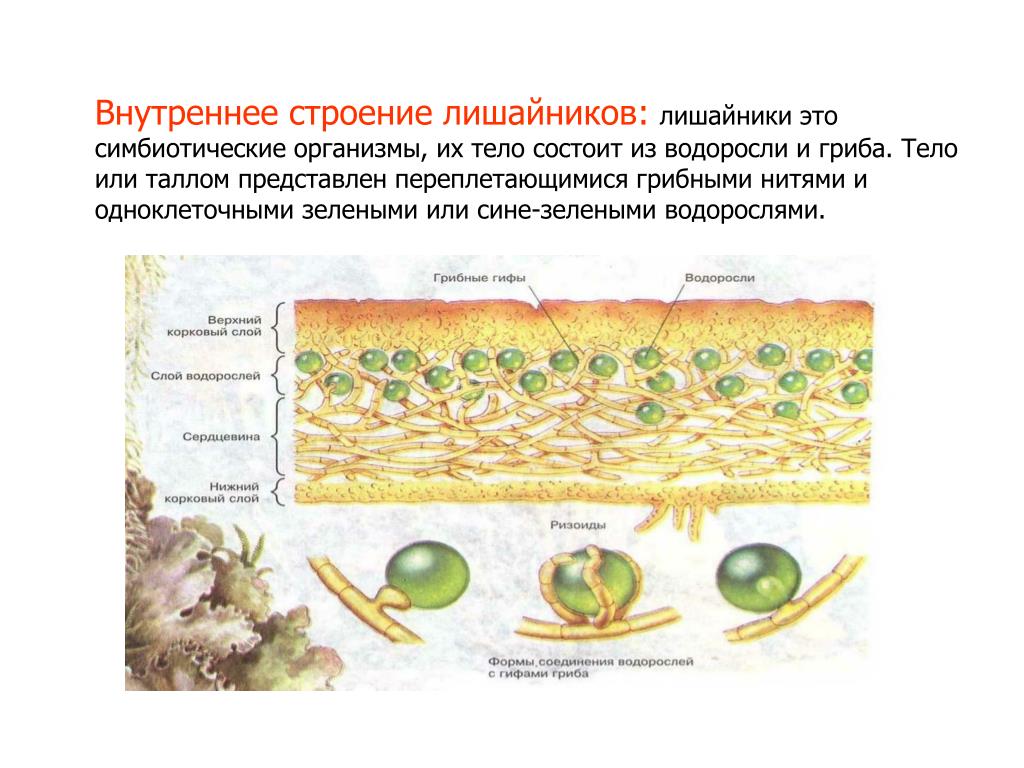 , 2009; Bragina et al., 2012). Однако до сих пор бактериальные сообщества почти всегда описывались без учета прилегающих местообитаний и потенциальных межмикробиомных отношений. Ранее о бактериальной специфичности сообщалось при исследовании слоевищ лишайников и подстилающего их каменного субстрата (Bjelland et al., 2011). Недавнее исследование в рамках докторской диссертации Ашенбреннера (2015), посвященное этой теме, выявило специфичность микробиома, связанного с лишайниками, по сравнению с соседними местообитаниями, т.е.д., мох и голая кора. Этот сравнительный анализ выявил потенциальных специалистов по среде обитания и специалистов широкого профиля. В этом обзоре представители рода Sphingomonas были идентифицированы как универсалы во всех трех местообитаниях, тогда как представители рода Mucilaginibacter были описаны как потенциальные специалисты по лишайникам. Легочный лишайник часто приживается на мхах, и совместное использование штаммов Nostoc обеими криптогамами предполагает ранее не описанную форму экологического облегчения, опосредованную общей фракцией микробиома (Aschenbrenner, 2015).
, 2009; Bragina et al., 2012). Однако до сих пор бактериальные сообщества почти всегда описывались без учета прилегающих местообитаний и потенциальных межмикробиомных отношений. Ранее о бактериальной специфичности сообщалось при исследовании слоевищ лишайников и подстилающего их каменного субстрата (Bjelland et al., 2011). Недавнее исследование в рамках докторской диссертации Ашенбреннера (2015), посвященное этой теме, выявило специфичность микробиома, связанного с лишайниками, по сравнению с соседними местообитаниями, т.е.д., мох и голая кора. Этот сравнительный анализ выявил потенциальных специалистов по среде обитания и специалистов широкого профиля. В этом обзоре представители рода Sphingomonas были идентифицированы как универсалы во всех трех местообитаниях, тогда как представители рода Mucilaginibacter были описаны как потенциальные специалисты по лишайникам. Легочный лишайник часто приживается на мхах, и совместное использование штаммов Nostoc обеими криптогамами предполагает ранее не описанную форму экологического облегчения, опосредованную общей фракцией микробиома (Aschenbrenner, 2015).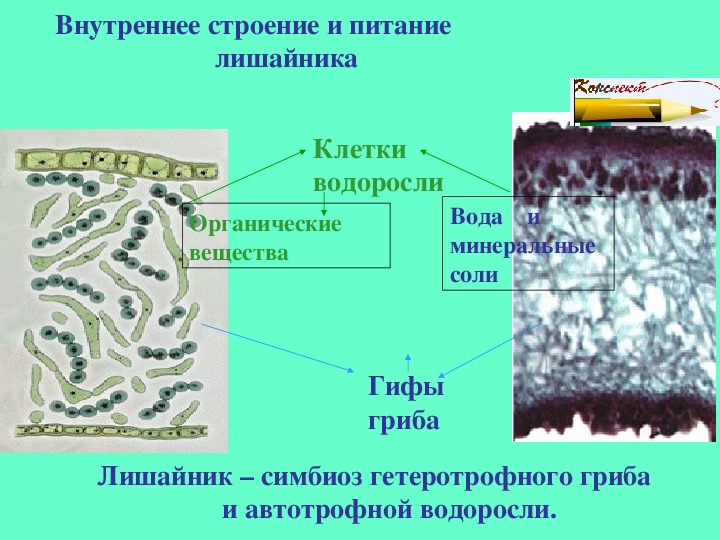 Легочный лишайник поглощает штамма Nostoc во время роста и включает их в слоевище в виде отдельных кластеров (известных в литературе как внутренние цефалодии). Поскольку Nostoc обогащается на мхах, а не на коре, стимулирующий рост эффект фиксации азота Nostoc , по-видимому, способствует эффективному развитию таллома лишайника, который в основном появляется из участков мха.
Легочный лишайник поглощает штамма Nostoc во время роста и включает их в слоевище в виде отдельных кластеров (известных в литературе как внутренние цефалодии). Поскольку Nostoc обогащается на мхах, а не на коре, стимулирующий рост эффект фиксации азота Nostoc , по-видимому, способствует эффективному развитию таллома лишайника, который в основном появляется из участков мха.
Микробиом, связанный с лишайниками, играет центральную функциональную роль в голобионте лишайников
Несмотря на то, что за последние годы была продемонстрирована специфичная для хозяина бактериальная колонизация различных видов лишайников, роль бактерий оставалась в значительной степени неизвестной.В основном это связано с присущими им проблемами изучения лишайников экспериментальными методами (особенно повторного синтеза симбиоза в культуре). Тем временем метаомика возникла как набор подходящих технологий для глобального выявления потенциально полезного вклада бактериальной популяции. Недавно микробиом, связанный с L. pulmonaria , был исследован с использованием интегрированного подхода метагеномики и метапротеомики для выявления потенциальных функций, закодированных в геномах, и проверки их экспрессии на уровне белков (Grube et al., 2015), основанный на предыдущем новаторском исследовании протеомики (Schneider et al., 2011). Результаты Grube et al. (2015) представили убедительные доказательства того, что бактериальный микробиом участвует в обеспечении питательными веществами и деградации старых частей таллома лишайника, биосинтезе витаминов и гормонов, процессах детоксикации и защите от биотического, а также абиотического стресса. Кроме того, высокая распространенность фиксации азота бактериями была подтверждена омическими данными и количественной ОТ-ПЦР. Более того, сравнение всего метагенома, связанного с Lobaria , с репрезентативным набором общедоступных метагеномов подчеркнуло его уникальность.Было обнаружено, что наиболее близкородственными метагеномами являются метагеномы, полученные из местообитаний, связанных с растениями.
Недавно микробиом, связанный с L. pulmonaria , был исследован с использованием интегрированного подхода метагеномики и метапротеомики для выявления потенциальных функций, закодированных в геномах, и проверки их экспрессии на уровне белков (Grube et al., 2015), основанный на предыдущем новаторском исследовании протеомики (Schneider et al., 2011). Результаты Grube et al. (2015) представили убедительные доказательства того, что бактериальный микробиом участвует в обеспечении питательными веществами и деградации старых частей таллома лишайника, биосинтезе витаминов и гормонов, процессах детоксикации и защите от биотического, а также абиотического стресса. Кроме того, высокая распространенность фиксации азота бактериями была подтверждена омическими данными и количественной ОТ-ПЦР. Более того, сравнение всего метагенома, связанного с Lobaria , с репрезентативным набором общедоступных метагеномов подчеркнуло его уникальность.Было обнаружено, что наиболее близкородственными метагеномами являются метагеномы, полученные из местообитаний, связанных с растениями.
В частности, ранее было показано, что Rhizobiales ( Alphaproteobacteria ) чрезвычайно распространены в микробиоме L. pulmonaria , в основном представленном семействами: Methylobacteriaceae , Bradyrhizobiaceae , Bradyrhizobiaceae 3.4 4 и , , , Хотя они хорошо известны своим благотворным взаимодействием со многими высшими растениями, меньше известно об их конкретной роли в отношении лишайников.По данным Erlacher et al. (2015) функциональные назначения, основанные на иерархической классификации SEED, показали участие Rhizobiales в различных полезных функциях (например, в биосинтезе ауксина, фолиевой кислоты и витамина B12). Дальнейшее разбиение показало, что преобладающие Methylobacteriaceae были также наиболее мощными продуцентами исследованных метаболитов. Эти результаты предполагают потенциал для различных биотехнологических применений этой группы.
Функции снятия стресса и защиты от патогенов поддерживаются метагеномными данными и культивируемыми членами микробиома
Недавно было показано, что L. pulmonaria ассоциированный микробиом включает также различные бактерии с антагонистическим потенциалом (Cernava et al., 2015a). Наиболее распространенными антагонистами были Stenotrophomonas, Pseudomonas, Micrococcus и Burkholderia . На эти роды приходилось 67% всех идентифицированных бактерий-антагонистов. Метагеномный скрининг выявил наличие генов, участвующих в биосинтезе метаболитов, снижающих стресс. Дополнительные анализы методом высокоэффективной жидкостной хроматографии-масс-спектрометрии (ВЭЖХ-МС) позволили обнаружить спермидин, продуцируемый Stenotrophomonas , который, как известно, снижает стресс, вызванный высыханием и высокой засоленностью растений.Также было проверено, может ли этот защитный эффект передаваться хозяевам, не являющимся лишайниками, например, семенам помидоров ( Solanum lycopersicum ). Результаты показали значительное увеличение длины корней и стеблей в условиях недостатка воды. Применение ассоциированных с лишайниками бактерий для защиты и стимулирования роста растений может оказаться полезной альтернативой традиционным подходам.
pulmonaria ассоциированный микробиом включает также различные бактерии с антагонистическим потенциалом (Cernava et al., 2015a). Наиболее распространенными антагонистами были Stenotrophomonas, Pseudomonas, Micrococcus и Burkholderia . На эти роды приходилось 67% всех идентифицированных бактерий-антагонистов. Метагеномный скрининг выявил наличие генов, участвующих в биосинтезе метаболитов, снижающих стресс. Дополнительные анализы методом высокоэффективной жидкостной хроматографии-масс-спектрометрии (ВЭЖХ-МС) позволили обнаружить спермидин, продуцируемый Stenotrophomonas , который, как известно, снижает стресс, вызванный высыханием и высокой засоленностью растений.Также было проверено, может ли этот защитный эффект передаваться хозяевам, не являющимся лишайниками, например, семенам помидоров ( Solanum lycopersicum ). Результаты показали значительное увеличение длины корней и стеблей в условиях недостатка воды. Применение ассоциированных с лишайниками бактерий для защиты и стимулирования роста растений может оказаться полезной альтернативой традиционным подходам. Однако необходимы дальнейшие исследования для оценки диапазона хозяев и выяснения общей применимости (Cernava et al., 2015а).
Однако необходимы дальнейшие исследования для оценки диапазона хозяев и выяснения общей применимости (Cernava et al., 2015а).
Кроме того, профили летучих органических соединений (ЛОС) из бактериальных изолятов показали, что бактерии, связанные с лишайниками, выделяют широкий спектр летучих веществ. Эти молекулы, скорее всего, участвуют в различных взаимодействиях (например, коммуникации между микроорганизмами и хозяином), а также могут повышать общую устойчивость к различным патогенам (Cernava et al., 2015b).
Микробиом обеспечивает дополнительные механизмы детоксикации
Помимо доказательств механизмов, обеспечивающих повышенную устойчивость к биотическому, а также абиотическому стрессу, микробиом предоставил первое доказательство участия в детоксикации неорганических веществ (например,г., As, Cu, Zn), детальные механизмы остаются неизвестными. Более глубокое понимание этого полезного вклада стало возможным с образцами, подвергшимися воздействию повышенных концентраций мышьяка (Cernava, 2015). Метагеномный анализ показал, что общая структура микробного сообщества из разных лишайников была сходной, независимо от концентрации мышьяка в местах отбора проб, тогда как спектр функций, связанных с метаболизмом мышьяка, был расширен. Эти функции включают механизмы биоконверсии, которые участвуют в метилировании неорганического мышьяка и, следовательно, генерируют менее токсичные вещества.Кроме того, в образцах, загрязненных мышьяком, увеличилось количество генов, связанных с детоксикацией. Дополнительные подходы к ПЦР показали, что число копий гена ars M не имеет строгой связи с определенными концентрациями мышьяка. Кроме того, коллекция культур бактериальных изолятов, полученных от трех видов лишайников, была проверена на наличие гена ars M. Обнаруженные носители ars М позднее были идентифицированы как представители родов Leifsonia, Micrococcus, Pedobacter, Staphylococcus и Streptomyces .Общие результаты подчеркнули важную роль микробиома в защите хозяина и предоставили более детальное представление о таксономической структуре задействованных микроорганизмов.
Метагеномный анализ показал, что общая структура микробного сообщества из разных лишайников была сходной, независимо от концентрации мышьяка в местах отбора проб, тогда как спектр функций, связанных с метаболизмом мышьяка, был расширен. Эти функции включают механизмы биоконверсии, которые участвуют в метилировании неорганического мышьяка и, следовательно, генерируют менее токсичные вещества.Кроме того, в образцах, загрязненных мышьяком, увеличилось количество генов, связанных с детоксикацией. Дополнительные подходы к ПЦР показали, что число копий гена ars M не имеет строгой связи с определенными концентрациями мышьяка. Кроме того, коллекция культур бактериальных изолятов, полученных от трех видов лишайников, была проверена на наличие гена ars M. Обнаруженные носители ars М позднее были идентифицированы как представители родов Leifsonia, Micrococcus, Pedobacter, Staphylococcus и Streptomyces .Общие результаты подчеркнули важную роль микробиома в защите хозяина и предоставили более детальное представление о таксономической структуре задействованных микроорганизмов.
Сборка бактериального микробиома на симбиотической грибковой структуре
Таллом лишайника с его разнообразными микронишами представляет собой миниатюрную экосистему для микроорганизмов. В то время как связанные с лишайниками бактерии ранее игнорировались и часто считались контаминацией слоевищ лишайников, недавние исследования рассматривают их — с растущим количеством доказательств — как важный и решающий компонент метаорганизма лишайников.По своему микробиому лишайники экологически связаны с окружающей их средой (рис. 1). Несмотря на то, что часть их микробиома может передаваться путем локального распространения вегетативных побегов, дальнейшее пополнение штаммов происходит из местных ресурсов в окружающей среде. Это, наконец, приводит к специфической структуре сообщества зрелых талломов лишайников, которые разделяют основной микробиом на большем расстоянии (Aschenbrenner et al., 2014). Талломы лишайников, уже присутствующие на Земле с нижнего девона и представляющие собой самые сложные вегетативные структуры в царстве грибов, могли эволюционировать как структуры, обогащенные бактериями. Открытые поверхности лишайников идеально подходят для использования функций адаптированных и обогащенных бактерий или для разложения ложных неадаптированных бактерий, попавших в окружающую среду. Урожай бактерий может быть легко передан симбиотическим корпорациям через грибковые текстуры. Именно эта новая перспектива симбиоза лишайников предлагает широкий спектр новых исследовательских вопросов в ближайшем будущем.
Открытые поверхности лишайников идеально подходят для использования функций адаптированных и обогащенных бактерий или для разложения ложных неадаптированных бактерий, попавших в окружающую среду. Урожай бактерий может быть легко передан симбиотическим корпорациям через грибковые текстуры. Именно эта новая перспектива симбиоза лишайников предлагает широкий спектр новых исследовательских вопросов в ближайшем будущем.
РИСУНОК 1. Целостный взгляд на разнообразие микробиома лишайников и выявленные функции в контексте окружающей среды. Было показано, что бактериальные сообщества, связанные с лишайниками, разделяют значительные доли идентифицированных таксонов с соседними микросредами обитания (синий кружок). Это предполагает динамичное приобретение и обмен полезными видами. Определенные пропорции микробиома вертикально передаются следующему поколению и используются для создания новых популяций (красный кружок и стрелки). Высокоразнообразные бактериальные популяции в основном колонизируют внешние слои лишайника, но некоторые из них также могут проникать в межгифальный матрикс (зеленый кружок). Внешние факторы обеспечивают общую микробную «основную сборку» среды обитания, но факторы, специфичные для хозяина (серый кружок), определяют специфическое для лишайника бактериальное сообщество, которое выполняет множество полезных функций для симбиоза хозяина (фиолетовый кружок).
Внешние факторы обеспечивают общую микробную «основную сборку» среды обитания, но факторы, специфичные для хозяина (серый кружок), определяют специфическое для лишайника бактериальное сообщество, которое выполняет множество полезных функций для симбиоза хозяина (фиолетовый кружок).
Выводы. Лишайники как пример для понимания многовидовых симбиозов
Несомненно, существуют и другие случаи симбиозов с участием множества групп организмов в наземных экосистемах. Подобно лишайникам, они первоначально были признаны двойными эукариотическими партнерствами, но позже было показано, что они также включают специфические бактериальные ассоциации (например,g., грибы/муравьи-листорезы, Little and Currie, 2007; микориза, Garbaye, 1994). Современные инструменты теперь преодолевают трудности, связанные с восстановлением сложных симбиозов в аксенических лабораторных условиях, и, кроме того, они позволяют нам точно изучать симбиозы в контексте их окружающей среды. Мы рассматриваем лишайники как идеальные объекты исследования для этой цели, так как в отличие от многих других симбиотических систем они обладают непревзойденным экологическим диапазоном в целом, но с достаточно специфической адаптацией каждого вида к своей экологической нише. Таким образом, очевидно, что это будет новая и очень интересная тема в симбиотических исследованиях, чтобы установить роль микробиома в экологической адаптации и эволюции многовидового симбиоза лишайников.
Таким образом, очевидно, что это будет новая и очень интересная тема в симбиотических исследованиях, чтобы установить роль микробиома в экологической адаптации и эволюции многовидового симбиоза лишайников.
Вклад авторов
IA, TC, GB и MG написали рукопись. IA и TC предоставили результаты своих докторских исследований. GB и MG дополнили рукопись глубоким опытом в области исследований микробиома и лишайников.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Эта работа была поддержана грантом Австрийского научного фонда (FWF) для GB и MG (FWF Project I882).
Ссылки
Аманн Р.И., Людвиг В. и Шлейфер К.Х. (1995). Филогенетическая идентификация и обнаружение in situ отдельных микробных клеток без культивирования. Микробиолог. Ред.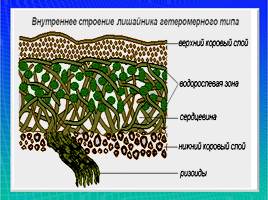 59, 143–169.
59, 143–169.
Реферат PubMed | Академия Google
Ашенбреннер, И. А. (2015). Стабильность и изменение микробиома лишайников. Докторская диссертация, Технологический университет Граца, Грац, 48–107.
Академия Google
Ашенбреннер, И. А., Кардинале, М., Берг, Г., и Грубе, М. (2014). Микробный груз: укрепляют ли бактерии на симбиотических пропагулах микробиом лишайников? Окружающая среда. микробиол. 16, 3743–3752. дои: 10.1111/1462-2920.12658
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Аттардо, Г. М., Лохс, К., Хедди, А., Алам, У. Х., Йилдирим, С., и Аксой, С.(2008). Анализ структуры и функции молочных желез у Glossina morsitans : продукция молочного белка, популяции симбионтов и плодовитость. J. Физиология насекомых. 54, 1236–1242. doi: 10.1016/j.jinsphys.2008.06.008
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бейтс, С. Т.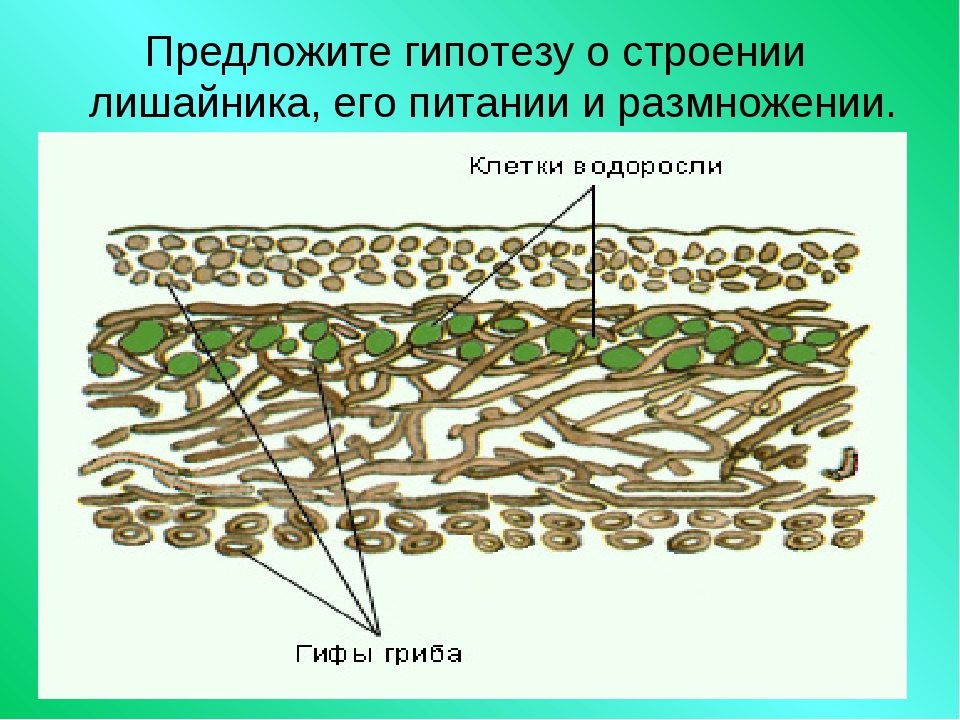 , Берг-Лайонс, Д., Лаубер, К. Л., Уолтерс, В. А., Найт, Р. и Фиерер, Н. (2012). Предварительное исследование эукариот, связанных с лишайниками, с использованием пиросеквенирования. Лихенолог 44, 137–146.дои: 10.1017/S00242820648
, Берг-Лайонс, Д., Лаубер, К. Л., Уолтерс, В. А., Найт, Р. и Фиерер, Н. (2012). Предварительное исследование эукариот, связанных с лишайниками, с использованием пиросеквенирования. Лихенолог 44, 137–146.дои: 10.1017/S00242820648
Полнотекстовая перекрестная ссылка | Академия Google
Бейтс, С. Т., Кропси, Г. В. Г., Капорасо, Дж. Г., Найт, Р., и Фиерер, Н. (2011). Бактериальные сообщества, связанные с симбиозом лишайников. Заяв. Окружающая среда. микробиол. 77, 1309–1314. doi: 10.1128/AEM.02257-10
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Беккет, Р. П., Краннер, И., и Минибаева, Ф. В. (2008). «Физиологическая экология обмена углекислого газа», в Lichen Biology , изд.TH Nash (Кембридж: издательство Кембриджского университета), 152–181.
Академия Google
Беккет, Р. П., Минибаева, Ф. В., Вылегжанина, Н. Н., и Толпышева, Т. (2003). Высокие показатели продукции внеклеточного супероксида лишайниками подотряда Peltigerineae коррелируют с показателями high.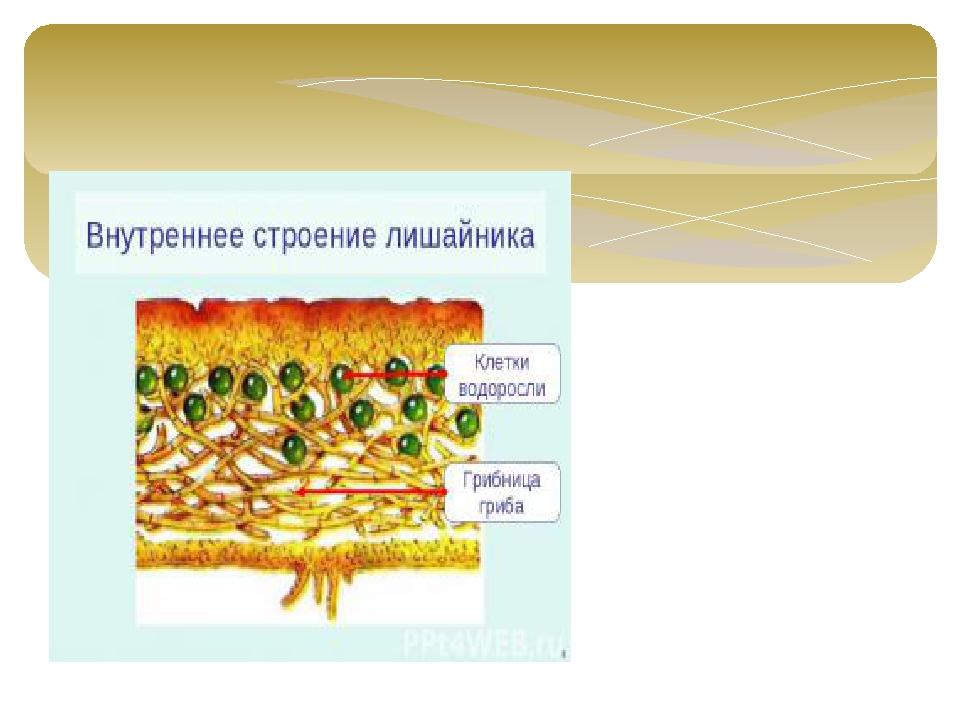 Окружающая среда растительных клеток. 26, 1827–1837 гг. doi: 10.1046/j.1365-3040.2003.01099.x
Окружающая среда растительных клеток. 26, 1827–1837 гг. doi: 10.1046/j.1365-3040.2003.01099.x
Полнотекстовая перекрестная ссылка | Академия Google
Берг, Г., и Смолла, К. (2009). Виды растений и тип почвы совместно формируют структуру и функции микробных сообществ в ризосфере. FEMS микробиол. Экол. 68, 1–13. doi: 10.1111/j.1574-6941.2009.00654.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Bjelland, T., Grube, M., Hoem, S., Jorgensen, S.L., Daae, F.L., Thorseth, I.H., et al. (2011). Микробные метасообщества в лишайниково-каменной среде обитания. Окружающая среда. микробиол. 3, 434–442.
Реферат PubMed | Академия Google
Брагина А., Берг С., Кардинале М., Щербаков А., Чеботарь В.и Берг, Г. (2012). Сфагновые мхи содержат высокоспецифичное бактериальное разнообразие на протяжении всего своего жизненного цикла. ISME J. 6, 802–813. doi: 10.1038/ismej.2011.151
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Брагина А. , Кардинале М., Берг К. и Берг Г. (2013). Вертикальная передача объясняет специфический рисунок Burkholderia в мхах Sphagnum в мультигеографическом масштабе. Перед. микробиол. 4:394.doi: 10.3389/fmicb.2013.00394
, Кардинале М., Берг К. и Берг Г. (2013). Вертикальная передача объясняет специфический рисунок Burkholderia в мхах Sphagnum в мультигеографическом масштабе. Перед. микробиол. 4:394.doi: 10.3389/fmicb.2013.00394
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бюдель, Б., и Шайдеггер, К. (2008). «Морфология и анатомия слоевища», в Lichen Biology , изд. TH Nash (Кембридж: издательство Кембриджского университета), 40–68.
Академия Google
Берк, К., Стейнберг, П., Руш, Д., Кьеллеберг, С., и Томас, Т. (2011). Сборка бактериального сообщества основана на функциональных генах, а не на видах. Проц. Натл. акад.науч. США 108, 14288–14293. doi: 10.1073/pnas.11015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кардинале М., Грубе М., Кастро Дж. В., Мюллер Х. и Берг Г. (2012a). Бактериальные таксоны, связанные с легочным лишаем Lobaria pulmonaria , по-разному формируются в зависимости от географии и среды обитания. FEMS микробиол. лат. 329, 111–115. doi: 10.1111/j.1574-6968.2012.02508.x
FEMS микробиол. лат. 329, 111–115. doi: 10.1111/j.1574-6968.2012.02508.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кардинале, М., Steinova, J., Rabensteiner, J., Berg, G., and Grube, M. (2012b). Возраст, солнце и субстрат: триггеры бактериальных сообществ в лишайниках. Окружающая среда. микробиол. 4, 23–28. doi: 10.1111/j.1758-2229.2011.00272.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Cardinale, M., Puglia, A.M., and Grube, M. (2006). Молекулярный анализ лишайниковых бактериальных сообществ. FEMS микробиол. Экол. 57, 484–495. doi: 10.1111/j.1574-6941.2006.00133.х
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кардинале М., Виейра де Кастро Дж., Мюллер Х., Берг Г. и Грубе М. (2008). Анализ in situ бактериального сообщества, ассоциированного с лишайником северного оленя Cladonia arbuscula , выявил преобладание Alphaproteobacteria . FEMS микробиол. Экол. 66, 63–71. doi: 10.1111/j.1574-6941.2008.00546.x
FEMS микробиол. Экол. 66, 63–71. doi: 10.1111/j.1574-6941.2008.00546.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чернава, Т.(2015). Изучение существенного вклада и глобальных взаимодействий микробиома в древнем симбиозе. Докторская диссертация, Технологический университет Граца, Грац, 15–16.
Академия Google
Чернава Т., Мюллер Х., Ашенбреннер И. А., Грубе М. и Берг Г. (2015a). Анализ антагонистического потенциала микробиома лишайника в отношении патогенов путем объединения метагеномных исследований с культуральными. Перед. микробиол. 6:620. doi: 10.3389/fmicb.2015.00620
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чернава Т., Ашенбреннер И. А., Грубе М., Либмингер С. и Берг Г. (2015b). Новый анализ для обнаружения биоактивных летучих веществ, оцениваемый путем скрининга бактерий, связанных с лишайниками. Перед. микробиол. 6:398. doi: 10.3389/fmicb.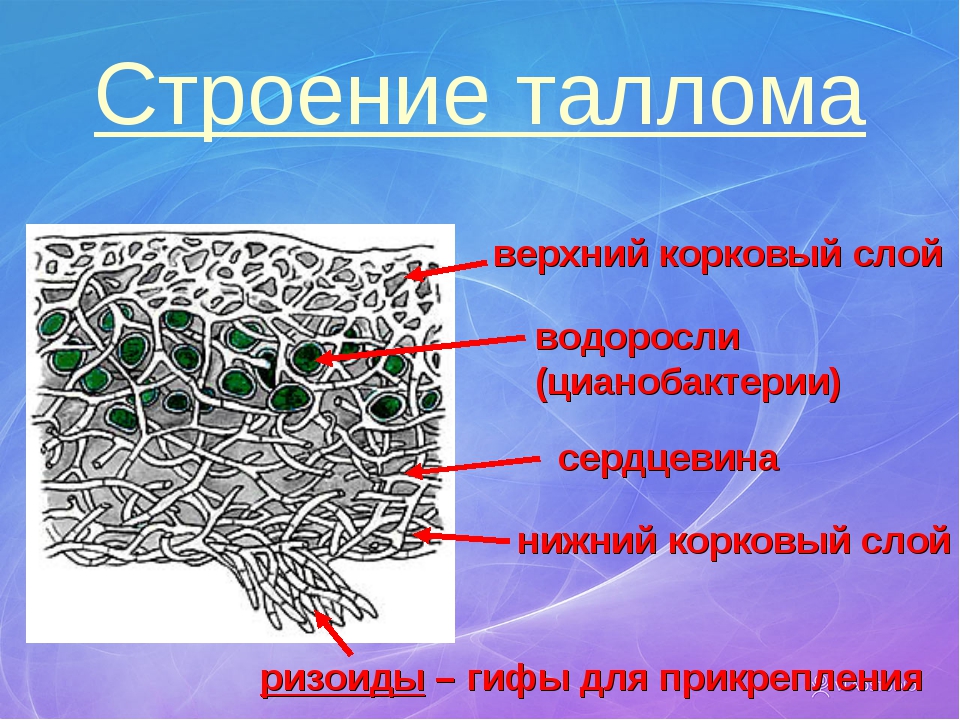 2015.00398
2015.00398
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
де Бари, А. (1879 г.). Die Erscheinung der Symbiose. Страсбург: Трибнер.
Академия Google
Дентон Г.Х. и Карлен В. (1973). Лихенометрия: ее применение в исследованиях голоценовой морены на юге Аляски и в шведской Лапландии. Арк. Альп. Рез. 5, 347–372. дои: 10.2307/1550128
Полнотекстовая перекрестная ссылка | Академия Google
Дуглас, А.Э. (1998). Пищевые взаимодействия в насекомо-микробных симбиозах: тли и их симбиотические бактерии Бухнера . Анну. Преподобный Энтомол. 43, 17–37. дои: 10.1146/annurev.ento.43.1.17
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эликс, Дж. А., и Стокер-Вёргёттер, Э. (2008). «Биохимия и вторичные метаболиты», в Lichen Biology , изд. TH Nash (Кембридж: издательство Кембриджского университета), 104–133.
Академия Google
Erlacher, A. , Cernava, T., Cardinale, M., Soh, J., Sensen, C.W., Grube, M., et al. (2015). Rhizobiales как функциональные и эндосимбионтные члены лишайникового симбиоза Lobaria pulmonaria L. Перед. микробиол. 6:53. doi: 10.3389/fmicb.2015.00053
, Cernava, T., Cardinale, M., Soh, J., Sensen, C.W., Grube, M., et al. (2015). Rhizobiales как функциональные и эндосимбионтные члены лишайникового симбиоза Lobaria pulmonaria L. Перед. микробиол. 6:53. doi: 10.3389/fmicb.2015.00053
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фуками, Т. (2010). «Динамика сборки сообщества в космосе», в «Экология сообщества: процессы, модели и приложения» , под редакцией Х. А. Верхофа и П. Дж. Морина (Оксфорд: издательство Оксфордского университета), 45–53.
Академия Google
Гарбай, Дж. (1994). Тэнсли обзор нет. 76. Бактерии-помощники: новое измерение микоризного симбиоза. Новый Фитол. 128, 197–210. doi: 10.1111/j.1469-8137.1994.tb04003.x
Полнотекстовая перекрестная ссылка | Академия Google
Гонсалес И., Аюсо-Сасидо А., Андерсон А. и Жениллоуд О. (2005). Актиномицеты, выделенные из лишайников: оценка их разнообразия и выявление биосинтетических последовательностей генов.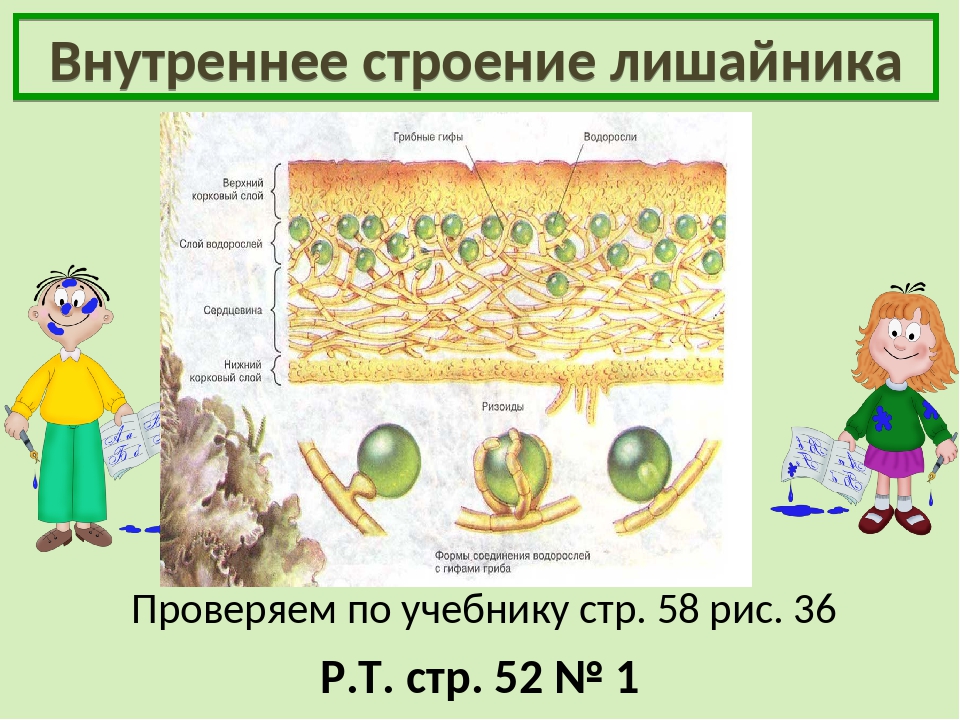 FEMS микробиол. Экол. 54, 401–415. doi: 10.1016/j.femsec.2005.05.004
FEMS микробиол. Экол. 54, 401–415. doi: 10.1016/j.femsec.2005.05.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Грубе, М.и Берг, Г. (2009). Микробные консорциумы бактерий и грибов с акцентом на симбиоз лишайников. Грибковый биол. Ред. 23, 72–85. doi: 10.1016/j.fbr.2009.10.001
Полнотекстовая перекрестная ссылка | Академия Google
Грубе М., Кардинале М., де Кастро Дж. В., Мюллер Х. и Берг Г. (2009). Видоспецифичное структурно-функциональное разнообразие бактериальных сообществ лишайниковых симбиозов. ISME J. 3, 1105–1115. doi: 10.1038/ismej.2009.63
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Грубе, М., Cernava, T., Soh, J., Fuchs, S., Aschenbrenner, I., Lassek, C., et al. (2015). Изучение функциональных контекстов симбиотической устойчивости внутри лишайниковых бактерий с помощью сравнительной омики. ISME J. 9, 412–424. doi: 10.1038/ismej.2014. 138
138
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Грубе, М., и Хоксворт, Д.Л. (2007). Проблемы с лишайником: переоценка и переосмысление формы слоевища и типов плодовых тел в молекулярную эпоху. Микол.Рез. 111, 1116–1132. doi: 10.1016/j.mycres.2007.04.008
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Грубе, М., Кёберл, М., Лакнер, С., Берг, К. и Берг, Г. (2012). Взаимодействие хозяин-паразит и реакция микробиома: влияние грибковых инфекций на бактериальное сообщество альпийского лишайника Solorina crocea . FEMS микробиол. Экол. 82, 472–481. doi: 10.1111/j.1574-6941.2012.01425.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хенкель, П.А. и Плотникова Т. Т. (1973). Азотфиксирующие бактерии лишайников. Изв. акад. Наук ССР сер. биол. 1973, 807–813.
Академия Google
Хенкель П.А. и Южакова Л.А. (1936). Азотфиксирующие бактерии лишайников.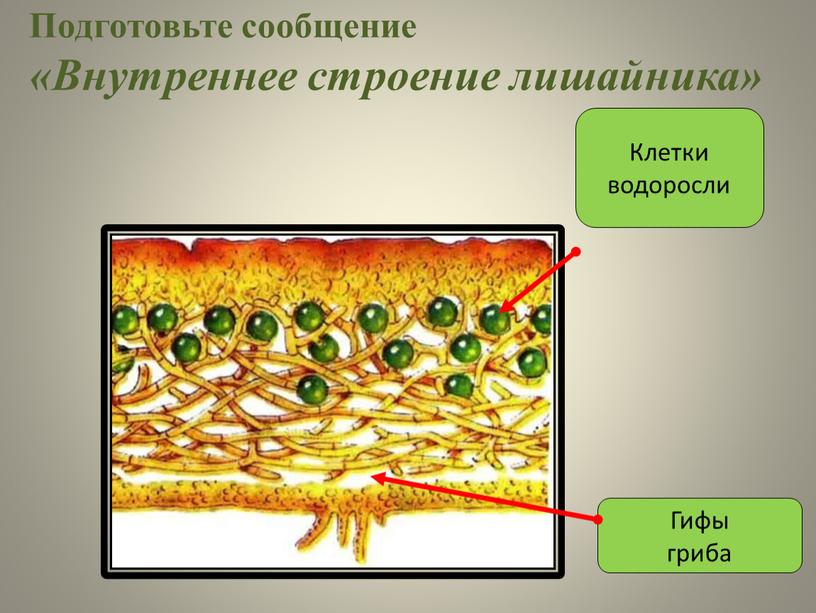 Изв. биол. Инст. Пермск. Гос. ун-т 10, 9–10.
Изв. биол. Инст. Пермск. Гос. ун-т 10, 9–10.
Академия Google
Ходкинсон, Б.П., Готтель, Н.Р., Шадт, К.В., и Луцони, Ф. (2012). Фотоавтотрофный симбионт и география являются основными факторами, влияющими на высокоструктурированные и разнообразные бактериальные сообщества в микробиоме лишайников. Окружающая среда. микробиол. 4, 147–161. doi: 10.1111/j.1462-2920.2011.02560.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ходкинсон, Б. П., и Луцони, Ф. (2009). Микробиотическое исследование бактерий, связанных с лишайниками, выявило новую линию от Rhizobiales. Симбиоз 49, 163–180. doi: 10.1007/s13199-009-0049-3
Полнотекстовая перекрестная ссылка | Академия Google
Онеггер Р., Эдвардс Д. и Акс Л. (2013). Самые ранние записи о внутренне стратифицированных цианобактериальных и водорослевых лишайниках из нижнего девона валлийской границы. Новый Фитол. 197, 264–275. doi: 10.1111/nph. 12009
12009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Искина Р.Ю. (1938). Об азотфиксирующих бактериях лишайников. Бык. Пермский (Молотов) биол. Рез. Инст. 11, 113–139.
Академия Google
Косанич, М., и Ранкович, Б. (2015). «Вторичные метаболиты лишайников как потенциальные антибиотики», в Lichen Secondary Metabolites , ed. Б. Ранкович (Берлин: Springer), 81–104.
Реферат PubMed | Академия Google
Ли, Ю.М., Ким, Э.Х., Ли, Х.К., и Хонг, С.Г. (2014). Биоразнообразие и физиологические характеристики антарктических и арктических лишайников-ассоциированных бактерий. World J. Microbiol. Биотехнолог. 30, 2711–2721. doi: 10.1007/s11274-014-1695-z
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ленова Л.И. и Блюм О. (1983). К вопросу о третьем компоненте лишайников. Бот. Дж. 68, 21–28.
Академия Google
Li, C.W., Chen, J.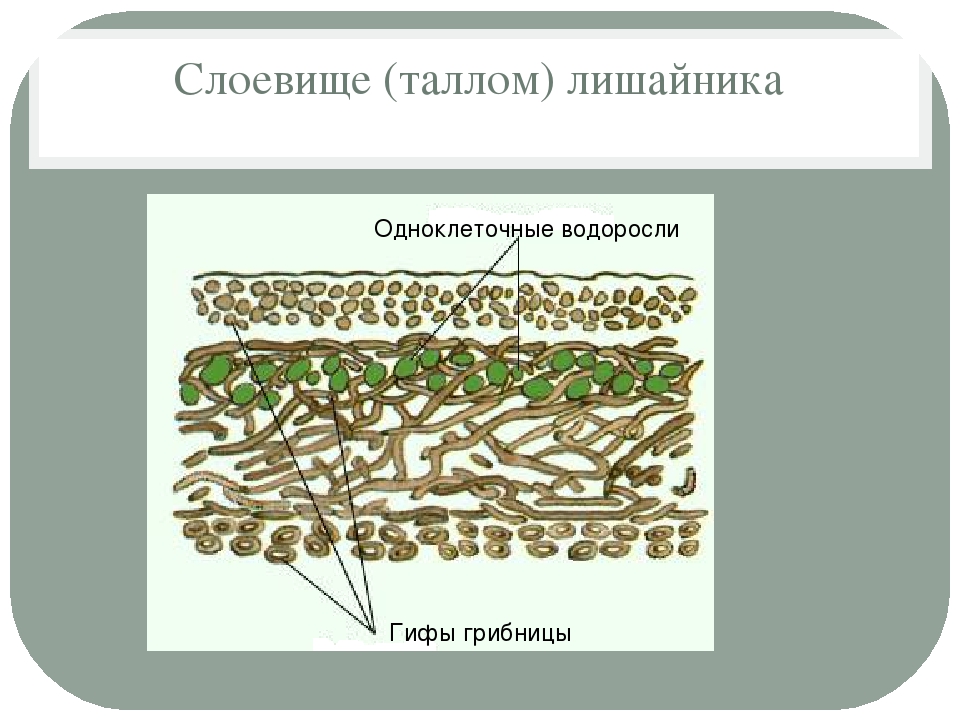 Y., and Hua, T.E. (1998). Докембрийские губки с ячеистым строением. Наука 279, 879–882. doi: 10.1126/наука.279.5352.879
Y., and Hua, T.E. (1998). Докембрийские губки с ячеистым строением. Наука 279, 879–882. doi: 10.1126/наука.279.5352.879
Полнотекстовая перекрестная ссылка | Академия Google
Liba, C.M., Ferrara, F.I.S., Manfio, G.P., Fantinatti-Garboggini, F., Albuquerque, R.C., Pavan, C., et al. (2006). Азотфиксирующие хемоорганотрофные бактерии, выделенные из лишенных цианобактерий лишайников, и их способность солюбилизировать фосфат и выделять аминокислоты и фитогормоны. J. Appl. микробиол. 101, 1076–1086. doi: 10.1111/j.1365-2672.2006.03010.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Литтл, А.Е., и Карри, Ч.Р. (2007). Симбиотическая сложность: открытие пятого симбионта в симбиозе аттиновых муравьев и микробов. биол. лат. 3, 501–504. doi: 10.1098/rsbl.2007.0253
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лю, В. Т., Марш, Т. Л., Ченг, Х., и Форни, Л.Дж. (1997). Характеристика микробного разнообразия путем определения полиморфизма длин концевых рестрикционных фрагментов генов, кодирующих 16S рРНК. Заяв. Окружающая среда. микробиол. 63, 4516–4522.
Заяв. Окружающая среда. микробиол. 63, 4516–4522.
Реферат PubMed | Академия Google
Лоси, К. Дж. (2010). Синтез традиционной биогеографии с микробной экологией: важность покоя. Ж. Биогеогр. 37, 1835–1841 гг.
Академия Google
Маргулис М., Эгхольм М., Альтман В. Э., Attiya, S., Bader, J.S., Bemben, L.A., et al. (2005). Секвенирование генома в микрореакторах высокой плотности пиколитр. Природа 437, 376–380.
Реферат PubMed | Академия Google
Мушегян, А. А., Петерсон, К. Н., Бейкер, К. К. М., и Прингл, А. (2011). Бактериальное разнообразие отдельных лишайников. Заяв. Окружающая среда. микробиол. 77, 4249–4252. doi: 10.1128/AEM.02850-10
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мюйзер Г.и Смолла, К. (1998). Применение денатурирующего градиентного гель-электрофореза (DGGE) и температурного градиентного гель-электрофореза (TGGE) в микробной экологии. Антони Ван Левенгук 73, 127–141. дои: 10.1023/A:1000669317571
Антони Ван Левенгук 73, 127–141. дои: 10.1023/A:1000669317571
Полнотекстовая перекрестная ссылка | Академия Google
Нэш, THIII (2008). Биология лишайников. Кембридж: Издательство Кембриджского университета, 1–8.
Академия Google
Паносян А.К., Никогосян В.Г. (1966).Наличие Azotobacter в лишайниках. акад. Наук Арм. ССР биол. Журн. Армен. 19, 3–11.
Академия Google
Прадо, С.С., и Цукки, Т.Д. (2012). Взаимодействие хозяина и симбионта для потенциальной борьбы с гетероптальными вредителями. Психея 2012, 269473. doi: 10.1155/2012/269473
Полнотекстовая перекрестная ссылка | Академия Google
Принцен, К., Фернандес-Мендоса, Ф., Маггиа, Л., Берг, Г., и Грубе, М. (2012). Альфапротеобактериальные сообщества в географически удаленных популяциях лишайника Cetraria aculeata . FEMS микробиол. Экол. 82, 316–325. doi: 10.1111/j.1574-6941.2012.01358.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Раппе, М. С., и Джованнони, С.Дж. (2003). Некультивируемое микробное большинство. Анну. Преподобный Микробиолог. 57, 369–394. doi: 10.1146/annurev.micro.57.030502.0
С., и Джованнони, С.Дж. (2003). Некультивируемое микробное большинство. Анну. Преподобный Микробиолог. 57, 369–394. doi: 10.1146/annurev.micro.57.030502.0
Полнотекстовая перекрестная ссылка | Академия Google
Реми В., Тейлор Т. Н., Хасс Х. и Керп Х. (1994). Везикулярная арбускулярная микориза возрастом четыреста миллионов лет. Проц. Натл. акад. науч. США 91, 11841–11843. doi: 10.1073/pnas.91.25.11841
Полнотекстовая перекрестная ссылка | Академия Google
Сакки Л., Григоло А., Мадзини М., Бильярди Э., Бачетти Б. и Лаудани У. (1988). Симбионты в ооцитах Blattella germanica (L.; Dictyoptera: Blattellidae): способ их передачи. Междунар. J. Морфолл насекомых. Эмбриол. 7, 437–446. дои: 10.1016/0020-7322(88)
-2Полнотекстовая перекрестная ссылка | Академия Google
Салим, М.(2015). «Экология экосистемы микробиома: невидимое большинство в антропогенной экосистеме», в Экология сообщества микробиома , изд. М. Салим (Cham: Springer International Publishing), 1–11.
М. Салим (Cham: Springer International Publishing), 1–11.
Академия Google
Шайдеггер, К., и Верт, С. (2009). Стратегии сохранения лишайников: выводы из популяционной биологии. Грибковый биол. Ред. 23, 55–66. doi: 10.1016/j.fbr.2009.10.003
Полнотекстовая перекрестная ссылка | Академия Google
Шнайдер, Т., Schmid, E., de Castro, J.V., Cardinale, M., Eberl, L., Grube, M., et al. (2011). Структура и функция партнеров по симбиозу легочного лишайника ( Lobaria pulmonaria L. Hoffm.) проанализированы с помощью метапротеомики. Протеомика 11, 2752–2756. doi: 10.1002/pmic.201000679
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Швенденер, С. (1869 г.). Die Algentypen der Flechtengonidien. Базель: Шульце.
Академия Google
Швигер, Ф.и Tebbe, CC (1998). Новый подход к использованию полиморфизма одноцепочечной конформации ПЦР для анализа микробного сообщества на основе гена 16S рРНК. Заяв. Окружающая среда. микробиол. 64, 4870–4876.
Заяв. Окружающая среда. микробиол. 64, 4870–4876.
Академия Google
Зельбманн, Л., Зуккони, Л., Руизи, С., Грубе, М., Кардинале, М., и Онофри, С. (2009). Культивируемые бактерии, ассоциированные с антарктическими лишайниками: принадлежность и психротолерантность. Полярный биол. 33, 71–83. doi: 10.1007/s00300-009-0686-2
Полнотекстовая перекрестная ссылка | Академия Google
Смолла, К., Oros-Sichler, M., Milling, A., Heuer, H., Baumgarte, S., Becker, R., et al. (2007). Бактериальное разнообразие почв, оцененное с помощью DGGE, T-RFLP и SSCP-отпечатков фрагментов гена 16S рРНК, амплифицированных с помощью ПЦР: дают ли разные методы одинаковые результаты? J. Microbiol. Методы 69, 470–479. doi: 10.1016/j.mimet.2007.02.014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
ван Овербик, Л. С., Франке, А. С., Нейхуис, Э. Х., Гроенвельд, Р. М., да Роша, У.Н. и Лотц, Л. А. (2011). Бактериальные сообщества, связанные с семенами Chenopodium album и Stellaria media из пахотных почв.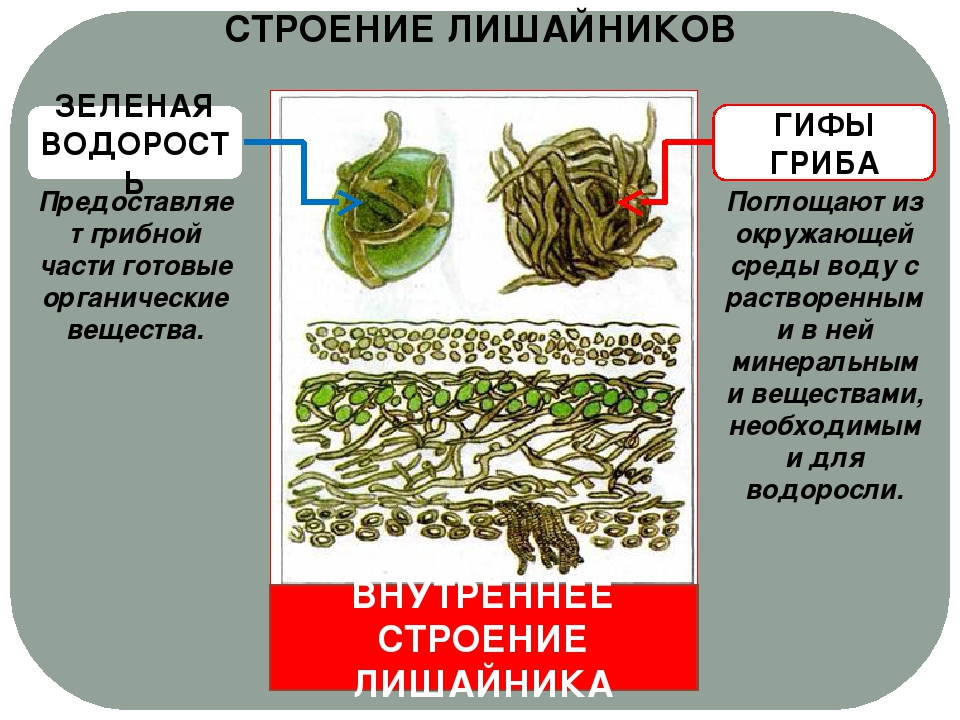 Микроб. Экол. 62, 257–264. doi: 10.1007/s00248-011-9845-4
Микроб. Экол. 62, 257–264. doi: 10.1007/s00248-011-9845-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уилкинсон, Ч. Р. (1984). Иммунологические доказательства докембрийского происхождения бактериальных симбиозов у морских губок. Проц. Р. Соц. Лонд. Б биол. науч. 220, 509–518. doi: 10.1111/j.1558-5646.2012.01676.х
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Терминология лишайников | Программа бакалавриата по биологии
Лишайники могут быть сложными и трудными для идентификации без предварительного понимания основной терминологии. На этой странице описывается и иллюстрируется терминология, используемая на протяжении всей лишайниковой тропы.
Подложка: поверхность, к которой прикрепляется лишайник: это может быть камень, кора, цементные знаки, деревья или вообще что угодно.
Апотеции : небольшие чашеобразные структуры, которые находятся на поверхности лишайника и выделяют споры, образующиеся половым путем.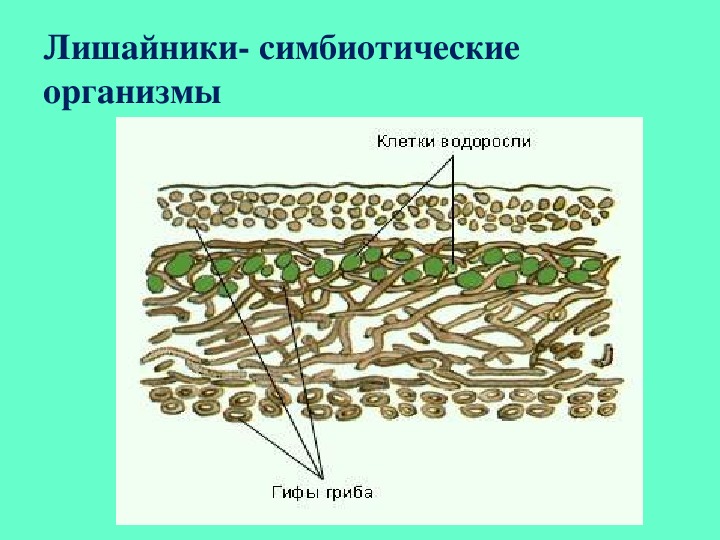 Серый накипный лишайник внизу покрыт оранжевыми апотециями.
Серый накипный лишайник внизу покрыт оранжевыми апотециями.
Soredia: Небольшие бесполые репродуктивные структуры. Соредии – это небольшие пучки грибов и водорослей. Они найдены в нескольких лишайниках на тропе.
Пикниды: Небольшие структуры, содержащие споры, не связанные с апотециями.Эти структуры сильно различаются по внешнему виду, начиная от маленьких черных точек на краю слоевища и заканчивая темно-оранжевыми прыщиками, как у Xanthomedoza fulva , которые показаны на этом следе лишайника и выделены в полях ниже.
Лишайник бывает всех форм, размеров и цветов. Наиболее широко лихенологи классифицируют лишайники по характеру роста: корковые, листоватые и кустистые. Эти формы роста не имеют никакого отношения к семейству, роду, виду или другой научной классификации, это просто внешний вид.
Накипь «покрывается коркой» на лишайнике. Некоторые говорят, что похоже лишайник был нанесен аэрозольной краской на подложку. Нижняя поверхность почти уходит в субстрат, что делает невозможным удаление лишайника с субстрата без разрушения лишайника. Это наиболее сложные из трех форм роста для идентификации, но они приносят чрезвычайное удовлетворение после выявления коркового лишайника.
Нижняя поверхность почти уходит в субстрат, что делает невозможным удаление лишайника с субстрата без разрушения лишайника. Это наиболее сложные из трех форм роста для идентификации, но они приносят чрезвычайное удовлетворение после выявления коркового лишайника.
Листовидный лишайник, как правило, выглядит как листовые наросты, прикрепленные к субстрату с помощью одной или нескольких стеблевидных структур, называемых ризином.Их можно удалить из субстрата, не повреждая и не уничтожая лишайник. Эта форма роста является наиболее представленным лишайником на лишайниковой тропе.
Кустарниковый лишайник является наиболее декоративным из трех типов роста. Эти лишайники могут выглядеть как крошечные деревья, кусты или даже крошечные чашечки пикси! Хотя они красивы и уникальны, эти формы роста обычно не встречаются в кампусе.
Чувствуете, что готовы к работе? Я надеюсь на это, потому что вы можете нажать Стоп 1, когда будете готовы!
Источники изображений:
Пикнида: http://bobklips. com/latewinter2011/
com/latewinter2011/
Соредия: http://www.plantscience4u.com/2014/07/difference-between-isidia-and-sore…
Все прочие: Эмма Ранквист
Существует много примеров симбиоз в природе, но лишайники уникальны, потому что они выглядят и ведут себя совершенно иначе, чем их компоненты организмы. Итак, лишайники рассматриваются как организмы в их собственное право и получили родовые и видовые названия. Однако для таксономических целей названия на самом деле названия грибов: лишайники рассматриваются как особая группа грибы — лихенизированные грибы . Есть приблизительное
от 13 500 до 17 000 видов лишайников, произрастающих
тропиков до полярных областей.Некоторые из них растут на
коре деревьев умеренного пояса или в качестве эпифитов на листьях
деревья в тропических лесах. Другие занимают часть
самая негостеприимная среда на земле, растущая на
охлажденные потоки лавы и голые поверхности скал, где они помогают
в процессе почвообразования и на пустынных песках
где они помогают стабилизировать поверхность и обогатить ее
с питательными веществами (см. Все эти особенности делают лишайники интересные и значимые в окружающей среде условия. Но лишайники также представляют собой сложную научную задачу. проблемы — как взаимодействуют два или более микроорганизмов на клеточном, генетическом и биохимическом уровнях для производства уникальный гибридный организм? |
||||
| виды лишайников
«тело» лишайника называется слоевищем ,
и его общая форма позволяет нам сгруппировать лишайники в
четыре широкие категории.
|
||||
| Лист
лишайники. (А) Пармелия
physodes , растущий на ветках кустарника. доли,
около 1 см в диаметре, сверху серебристо-серые, снизу черные.
(B) Peltigera polydactyla ,
растет на почве. Доли полустоячие, 1-2 см.
диаметром и имеют коричневые плодовые тела грибов ( аскокарпы )
на их кончиках (см. Нажмите здесь больше фотографий листоватых лишайников |
||||
| Фруктоза
и чешуйчатые лишайники. (C) Usnea comosa , растущая на деревянном
Почта. Этот лишайник имеет разветвленный,
нитевидный таллом, который свисает вниз от точки
вложение. (D) Cladonia pyxidata , растущая
на торфяной почве. Этот лишайник имеет чешуйчатую чешуйку.
структура (наконечник стрелы), но также дает прямостоячие стебли
чашки под названием podetia. (E) Кладония
coccifera , похож на C. pyxidata , но ободки
подеции несут много заметных красных аскокарпов. |
||||
| Корковая
лишайники. |
||||
| лишайниковое товарищество Несколько
лишайниковые грибы (около 20) входят в состав грибковых
группы Basidiomycota, но подавляющее большинство являются членами
Ascomycota (аскообразующие грибы) и они
часто образуют заметные плодовые тела грибов
(аскокарпы) — обычно дискообразные структуры, называемые апотециями
(т.е.г. рисунки Д, Ж). Почти половина зарегистрированных грибов в
мире являются аскомикотами, и почти половина из них
встречаются только в лишайниках. В отличие от многих
тысяч лишайников, всего около 100
партнеры по фотосинтезу.Наиболее распространены
одноклеточные зеленые водоросли рода Trebouxia
(рис. Н), которые встречаются у многих лишайников умеренного
и арктические/альпийские регионы, включая все виды
лишайник обыкновенный рода Cladonia (см. рис. Г, Д). Требусия
виды редко растут в природе как свободноживущие клетки;
вместо этого они кажутся специализированными симбионтами лишайников.
Другой распространенный фотобионт — нитчатая зеленая водоросль.
род Trentepohlia , особенно
в средиземноморских и тропических регионах.Это и другие
водорослевые роды тропических лишайников могут быть найдены растущими
самостоятельно в природе. Тот факт, что лишайники могут быть образованы более чем одним видом грибка и более чем один тип фотосинтетического партнера показывает нам, что симбиоз лишайников должен был развиться независимо от несколько случаев. |
||||
| Одноклеточные зеленая водоросль, Trebouxia |
||||
| строение лишайников Лишайники
различаются по степени структурной организации. |
||||
| Рисунок I.Строение лишайника Peltigera polydactyla . В поперечном срезе (слева) видно слоевище лишайника состоят из верхней коры (в) плотно упакованных грибковые клетки, напоминающие ткань. Это видно на более высоких увеличение на центральном изображении. Под корой находится сердцевина (м) более типичных гиф грибов (видно при большем увеличении на правом изображении). То фотосинтезирующие клетки (в данном случае цианобактерии). виден в виде полосы зеленого цвета под корой.То лишай также имеет нижнюю кору под мозговым веществом, но в этом разделе его не видно. Обратите внимание на воздух
карманы в верхней части продолговатого мозга, близко к
зона фотосинтеза (серо-черные области в левой
изображение). Они указывают на то, что гифы грибов в этом
области являются гидрофобными. |
||||
| Рисунки J, K. Строение обыкновенного листовидного лишайника Xanthoria parietina , который растет на многих типах поверхностей, включая бетон, крыши зданий и скалы подвергается морским брызгам. Таллом лишайника ( J ) имеет листоватые доли по краям (см. верхнюю часть рисунка J), но большая часть поверхности покрыта ярко-оранжевым апотеции около 3-5 мм в диаметре (наконечники стрел). Оранжевый цвет этого лишайника обусловлен выработкой пигмента париетин на поверхности лишайника. (К)
Поперечный срез одной из краевых долей, осмотр
фазово-контрастная микроскопия. Зона фотосинтеза ( стр )
виден как отчетливая полоса клеток зеленых водорослей. |
||||
| Физиология лишайников Микобионт имеет два основных роли в симбиозе лишайников: Фотобионт также имеет две роли:
Таким образом, через лишайник партнерства, фотобионты защищены и способны растут в условиях, в которых они не могли бы расти поодиночке; они также извлекают выгоду из высокоэффективного поглощения минерального питания лишайниковыми грибами.Грибы, в очередь, получают сахара и в некоторых случаях органический азот от партнера по фотосинтезу, что позволяет им расти в среды с дефицитом органических питательных веществ. |
||||
| Вода родство лишайников Лишайники
отличаются своей способностью выдерживать длительные
высыхание и быстрое возобновление активности после повторного увлажнения. Засухоустойчивость
лишайникам, вероятно, придают водоотталкивающие свойства.
(гидрофобное) покрытие на гифальных стенках мозгового вещества.
Небольшие пептиды, богатые серосодержащими аминокислотами.
кислоты были обнаружены на стенках гиф многих
свободноживущие грибы. Их называют гидрофобами
и они, вероятно, встречаются и у лишайниковых грибов.То
присутствие этих соединений гарантирует, что мозговой слой
вокруг фотосинтезирующих клеток не становится
заболоченный, что позволяет диффузии газообразного углерода
диоксид для фотосинтеза. Интересно гидрофобное
материалы, кажется, производятся только грибком, потому что
клетки Trebouxia , которые имеют естественный
гирофильная поверхность, покрываются гидрофобным
материал при выращивании в присутствии лишайникового гриба. Повторное заболачивание лишайников Считается, что оно начинается с поглощения воды студенистой матрикс коры, по которому вода могла двигаться через мозговое вещество в пристеночное пространство грибкового гифы или, возможно, капиллярность в областях, где гифы имеют гидрофильную поверхность. |
||||
| Питательный обмен между лишайниками-партнерами Наибольшее внимание обмену питательными веществами в лишайников сосредоточился на механизмах потока углерода из партнером гриба по фотосинтезу, потому что исследования радиоактивной маркировки показали, что как зеленые водоросли и цианобактерии могут высвобождать до 90% своего фотосинтеза к грибному партнеру. У лишайников с Trebouxia ,
и предположительно также с другими зелеными водорослями, грибковым
гифы могут образовывать короткие ответвления, проникающие через
водорослевая стенка служит поглощающими питательные вещества гаусториями . Основные растворимые
углеводов в лишайниках сахарных спиртов
(полиолы). У грибкового партнера эти соединения
присутствует в виде маннита и, в меньшей степени, арабита .
В этом отношении лишайники ничем не отличаются от
другие грибы, для которых характерны сахарные спирты
в качестве основных растворимых углеводов.Зеленые водоросли
производят сахарные спирты в качестве основного фотосинтетического
продукты — напр. сахарный спирт рибитол есть
производства Trebouxia . Интересно, максимум скорости высвобождения питательных веществ от партнера по фотосинтезу происходят в условиях оптимальной влажности, тогда как Фотосинтезирующие клетки сохраняют большую часть углеводов в условиях водного стресса.Итак, было предложено что циклы смачивания и сушки могут быть выгодны в поддержание лишайникового симбиоза, поскольку оба партнера может получить достаточное количество углеводов на разных этапах этот цикл. |
||||
| Рисунки L, M. Стадия спорообразования гриба Xanthoria
париетина . ( L ) Разрез по части
слоевище в области дисковидного апотеция. |
||||
| Цифры N-P. Стадия спорообразования гриба Peltigera.
полидактила . ( Н )
Апотеции рассматриваются как
коричневые, щитовидные структуры на концах слоевища
(этот рисунок представляет собой крупный план рисунка B).( O ) Часть измельченного апотеция с аски
(один отмечен стрелкой) и парафизы с коричневыми кончиками. ( P ) Аскоспоры высвобождаются при дроблении
апотеций. |
||||
| Репродукция и механизмы рассеивания Большинство лишайники распространяются путем вегетативного размножения . Слоевище сухого лишайника ломкое, поэтому его фрагменты можно легко ломаются и переносятся ветром или животными.Кроме того, некоторые лишайники образуют стеблевидные или столбчатые конструкции, называемые isidia , которые легко отрываются и расходятся. Все эти фрагменты могут возобновить рост в новой среде после того, как они повторно смоченный. Еще одно средство
размножение путем производства специализированных расселений
единицы, названные soredia (Рисунки Q, R). Они состоят
несколько фотосинтезирующих клеток, покрытых гифами грибов,
и они формируются в виде порошкообразной массы вблизи центра
слоевище лишайника или (e. В дополнение к этим
методы расселения, многие лишайники образуют апотеции или
другие структуры плодовых грибов, чтобы рассеять споры
грибкового партнера (Фигуры L — P ). То
споры в этих случаях ( аскоспор ) образуются в аски
в результате полового размножения (см.
Грибковая паутина),
хотя некоторые грибы-лишайники размножаются бесполым путем.
споры.Когда эти споры прорастают, они должны контактировать с
клетки партнера по фотосинтезу для создания нового
таллом лишайника. Этот процесс «сборки»
лишай можно продемонстрировать в экспериментальных условиях,
и данные свидетельствуют о том, что это также происходит в природе. Но
его частота в естественных условиях может варьировать
между разными типами лишайников. |
||||
| Рисунок Q. Physcia grisea , обычный лишайник, растет на стенах, деревьях и столбах забора в городе среды. Края колонии состоят из узких лопастей. (около 2-3 мм), но центр каждого слоевища имеет более темный, зеленый вид и более гранулированный, потому что покрыта соредиями.
Рисунок R . |
||||
| Лишайник экология и значение Лишайники являются одними из самых медленно растущих организмов, но их терпимость к экстремальным условиям окружающей среды позволяет им колонизировать места обитания, где мало других макроскопических организмов может расти.Они растут там, где нет ни грибного партнера, ни фотосинтетический партнер мог выжить в одиночку, потому что они извлекают выгоду из своей уникальной симбиотической ассоциации. Однако большинство лишайников
крайне нетерпимы к атмосферному загрязнению, особенно
двуокиси серы, поэтому они встречаются в основном в сельской
среды, а не города. Всего несколько видов, таких
как Physcia grisea и Xanthoria parietina ,
обычно встречаются в городах, и даже они плохо растут в
городах по сравнению с незагрязненной окружающей средой. |
||||
| Дополнительная литература (1) Книги, журналы и основные справочные источники Хейл ME (1983) Биология лишайников . Эдвард Арнольд, Лондон [Лучший общая книга по биологии лишайников] Нэш TH (1996) Лишайник Биология . Издательство Кембриджского университета, Кембридж. [Ан обновление книги Хейла с главами разных авторов, но очень подробно — не рекомендуется для новички] Главный журнал для всех
аспекты лишайников Лихенолог , опубликовано
Академическая пресса. Многие детализированные аспекты биологию лишайников можно найти в трех томах CRC. Справочник по лихенологии (под ред. М. Галуна), 1988. CRC Press, Бока-Ратон, Флорида, США. (2) Полезные сайты WWW Доступ ко многим сайтам с лишайниками через домашнюю страницу Международной ассоциации Лихенологи (нет на этом сервере) См. также: Фотографии лишайника (не на этом сервере) И «зажигалка» но полезный сайт: Fun with Lichens (не на этом сервере) (3) Избранные ссылки по конкретным темам: Синтез лишайников в лабораторных условиях и в природе:
Перемещение радионуклиды из лишайников через пищевую цепь животных:
Вклад лишайников для обогащения азотом различных экосистем:
Физиология лишайников:
Взаимодействия лишайников и развитие сообщества:
ГО ПОЛНЫЙ СПИСОК ПРОФИЛЕЙ? |
||||
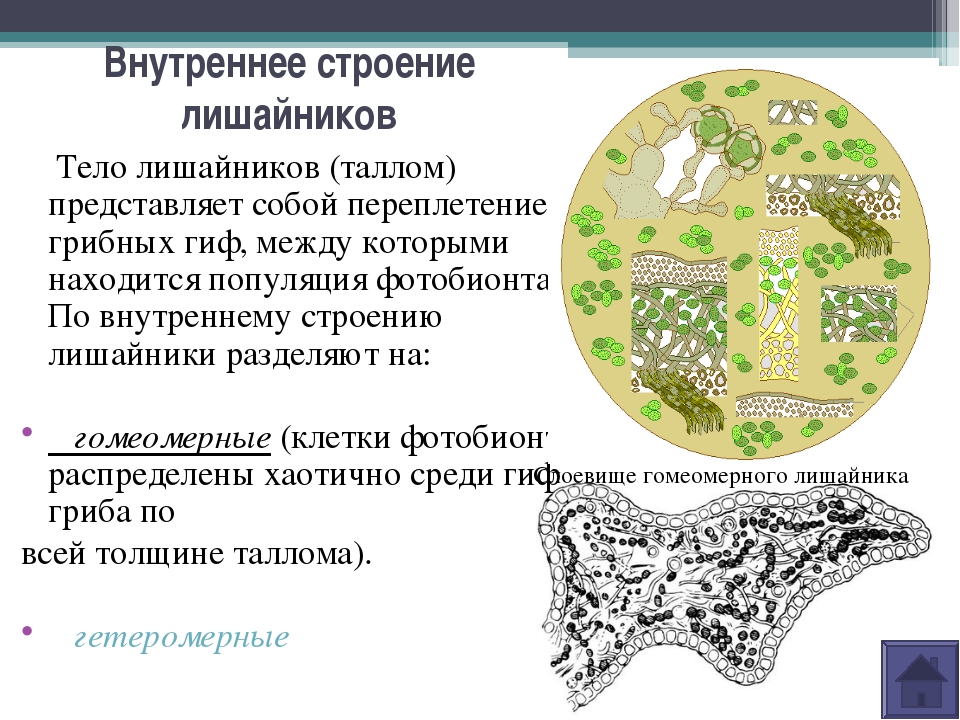
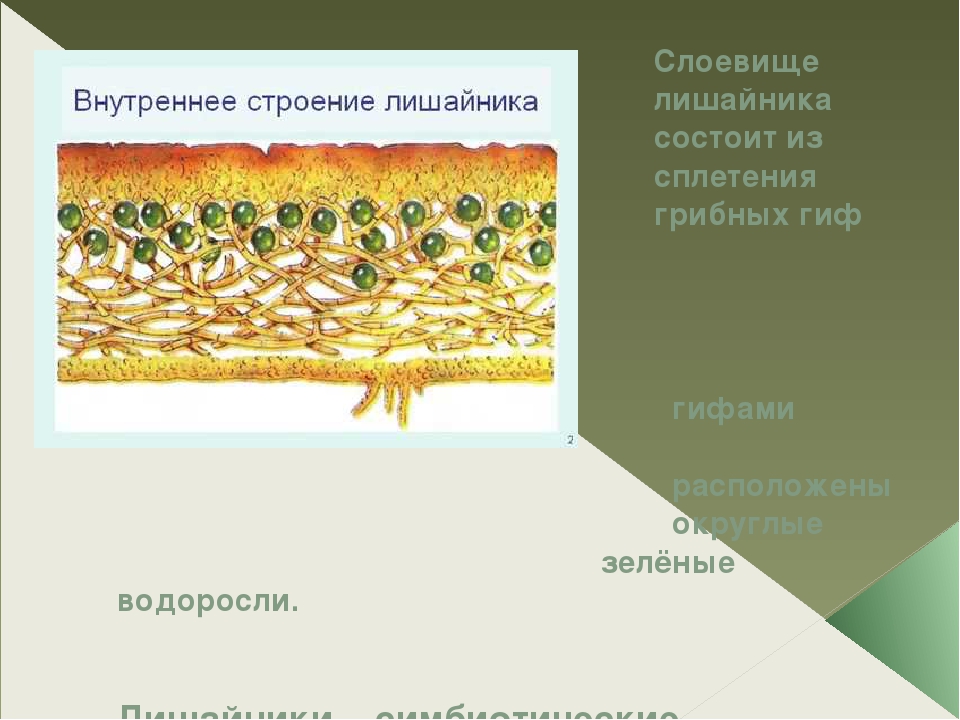 Цианобактерии). Некоторые другие виды лишайников растут
обильно на тундровых почвах, обеспечивая жизненно важный зимний корм
источник для животных (включая северных оленей и карибу) в
Арктические и субарктические регионы.Еще другие лишайники растут на
или в многолетних листьях некоторых хозяйственно важных
тропические растения, такие как кофе, какао и каучуконосы,
где их считают паразитами.
Цианобактерии). Некоторые другие виды лишайников растут
обильно на тундровых почвах, обеспечивая жизненно важный зимний корм
источник для животных (включая северных оленей и карибу) в
Арктические и субарктические регионы.Еще другие лишайники растут на
или в многолетних листьях некоторых хозяйственно важных
тропические растения, такие как кофе, какао и каучуконосы,
где их считают паразитами. 
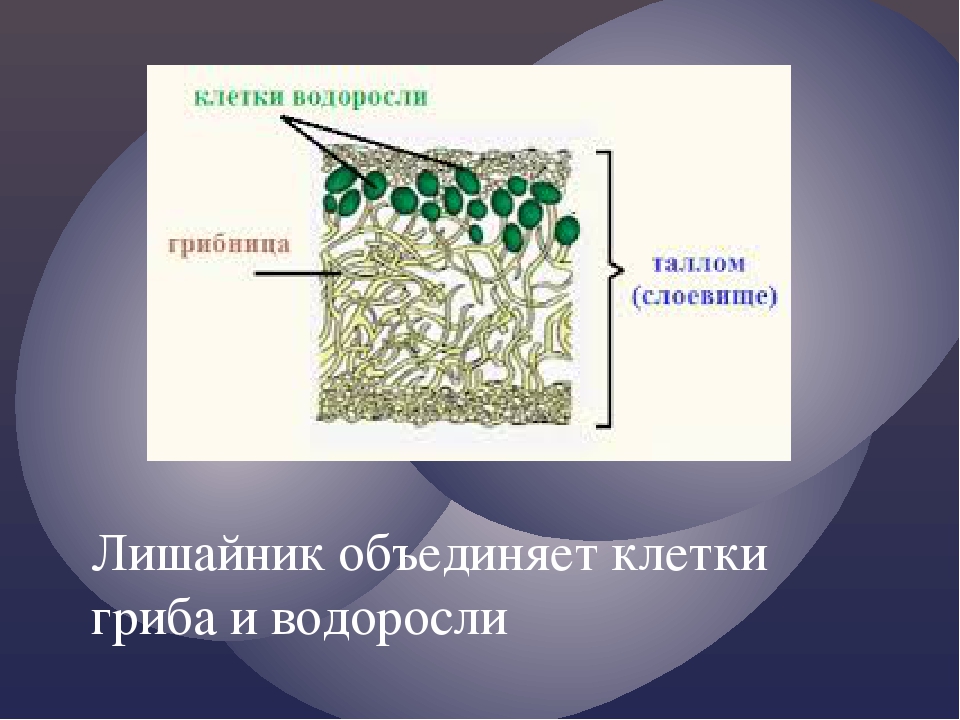 далее). В этом случае лишайник
слоевище серое, потому что оно высохло, но оно быстро
при повторном увлажнении становится голубовато-зеленым.
далее). В этом случае лишайник
слоевище серое, потому что оно высохло, но оно быстро
при повторном увлажнении становится голубовато-зеленым. (Ф) А
лоскутное одеяло из лишайников на поверхности скалы, в том числе несколько
колонии ризокарпона зеленого цвета
geographicum и белая Lecidia видов. (G) Lecanora muralis (также известный как Squamaria
muralis ), растущий на стене.Слоевище имеет бледный
зеленого цвета с небольшими лепестками по краям, но
центр ракообразный. Также присутствует много бледно-коричневых
аскокарпы.
(Ф) А
лоскутное одеяло из лишайников на поверхности скалы, в том числе несколько
колонии ризокарпона зеленого цвета
geographicum и белая Lecidia видов. (G) Lecanora muralis (также известный как Squamaria
muralis ), растущий на стене.Слоевище имеет бледный
зеленого цвета с небольшими лепестками по краям, но
центр ракообразный. Также присутствует много бледно-коричневых
аскокарпы. 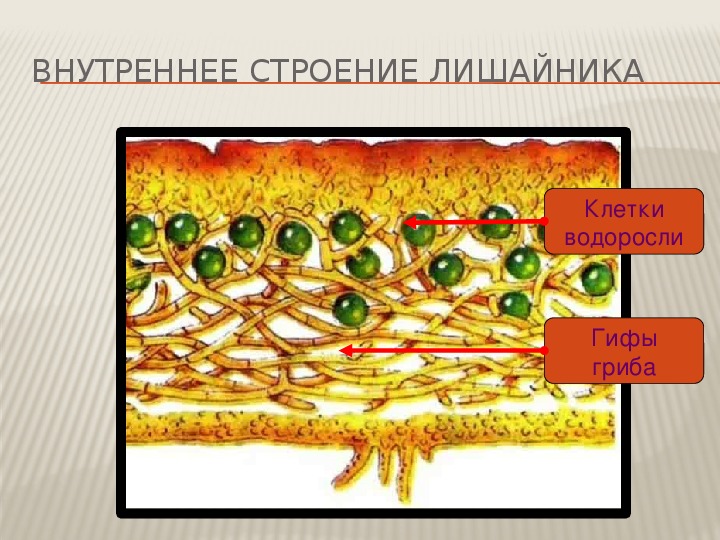 Хотя их споры
рассеявшись от плодовых тел, эти грибы не
играют самостоятельную роль в природе, потому что они
чрезвычайно медленно растут и, как правило, лишены фермента
системы для разложения сложных полимеров.
Хотя их споры
рассеявшись от плодовых тел, эти грибы не
играют самостоятельную роль в природе, потому что они
чрезвычайно медленно растут и, как правило, лишены фермента
системы для разложения сложных полимеров.  Около 10% лишайников имеют
цианобактерии (например, Nostoc ) в качестве основного или
единственный фотосинтезирующий партнер. Например, это справедливо для
многие вида Peltigera (рис. B). Однако некоторые
лишайники, содержащие зеленые водоросли, также могут иметь
цианобактерии в особых бородавчатых образованиях на
поверхность лишайника. Эти структуры называются cephalodia .Они обнаружены примерно у 3-4% видов лишайников и их
роль, вероятно, заключается в использовании азотфиксирующей способности
цианобактерий.
Около 10% лишайников имеют
цианобактерии (например, Nostoc ) в качестве основного или
единственный фотосинтезирующий партнер. Например, это справедливо для
многие вида Peltigera (рис. B). Однако некоторые
лишайники, содержащие зеленые водоросли, также могут иметь
цианобактерии в особых бородавчатых образованиях на
поверхность лишайника. Эти структуры называются cephalodia .Они обнаружены примерно у 3-4% видов лишайников и их
роль, вероятно, заключается в использовании азотфиксирующей способности
цианобактерий.  В один
крайними являются какие-то простые, почти случайные ассоциации
между гифами грибов и фотосинтезирующими клетками, в которых
есть небольшая структурная модификация. Эти простые
ассоциации могут быть довольно обычными, но незаметными.
Наиболее заметные лишайники имеют четко выраженный
структура, часто с отчетливой зональностью партнеров,
как показано на рисунках I-K ниже. Обычно
фотосинтезирующие клетки находятся в определенной полосе, где
они тесно связаны с гифами грибов, т.
обмен питательных веществ.Над этой полосой находится кора .
состоящие из плотно упакованных грибковых клеток с
студенистая матрица между ними. Внутри и ниже
фотосинтетическая зона представляет собой мозгового вещества рыхло упакованных
гифы Часто бывает более тонкая, нижняя кора ,
и несколько видов лишайников прикрепляются к
поверхность корневидными структурами, называемыми rhizinae или
ризины (см.
В один
крайними являются какие-то простые, почти случайные ассоциации
между гифами грибов и фотосинтезирующими клетками, в которых
есть небольшая структурная модификация. Эти простые
ассоциации могут быть довольно обычными, но незаметными.
Наиболее заметные лишайники имеют четко выраженный
структура, часто с отчетливой зональностью партнеров,
как показано на рисунках I-K ниже. Обычно
фотосинтезирующие клетки находятся в определенной полосе, где
они тесно связаны с гифами грибов, т.
обмен питательных веществ.Над этой полосой находится кора .
состоящие из плотно упакованных грибковых клеток с
студенистая матрица между ними. Внутри и ниже
фотосинтетическая зона представляет собой мозгового вещества рыхло упакованных
гифы Часто бывает более тонкая, нижняя кора ,
и несколько видов лишайников прикрепляются к
поверхность корневидными структурами, называемыми rhizinae или
ризины (см. рис. К).
рис. К).  Обратите также внимание на очень толстый
стенки гиф в мозговом веществе (правое изображение).
Эти толстостенные гидрофобные гифы способствуют
засухоустойчивость.
Обратите также внимание на очень толстый
стенки гиф в мозговом веществе (правое изображение).
Эти толстостенные гидрофобные гифы способствуют
засухоустойчивость.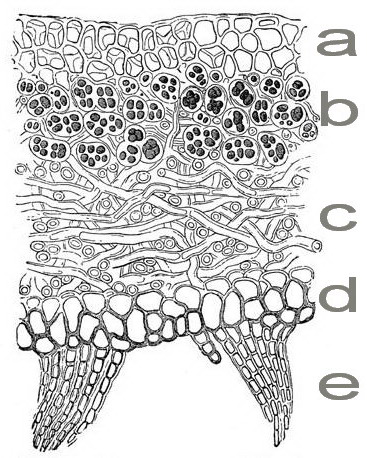 Выше
эта полоса представляет собой кору ( c ), состоящую из плотно
упакованные грибковые клетки. Поверхность коры представляет собой
пигментированы, потому что клеточные стенки пропитаны
париетин. Нижняя часть слоевища состоит из
мозговое вещество ( m ) с заметными воздушными карманами ( a ),
и тонкая нижняя кора ( lc ).Как и многие листовки
лишайники, Xanthoria производит ризины ( r )
которые проникают в щели и помогают закрепить
лишайник на поверхность. Пигментация возле кончика
rhizina происходит за счет образования пурпурно-красного пигмента.
клетки гиф в этой области.
Выше
эта полоса представляет собой кору ( c ), состоящую из плотно
упакованные грибковые клетки. Поверхность коры представляет собой
пигментированы, потому что клеточные стенки пропитаны
париетин. Нижняя часть слоевища состоит из
мозговое вещество ( m ) с заметными воздушными карманами ( a ),
и тонкая нижняя кора ( lc ).Как и многие листовки
лишайники, Xanthoria производит ризины ( r )
которые проникают в щели и помогают закрепить
лишайник на поверхность. Пигментация возле кончика
rhizina происходит за счет образования пурпурно-красного пигмента.
клетки гиф в этой области.  В некоторых
экосистемы, такие как пустынные почвы, тундровые пустоши,
и дугласово-пихтовые леса на северо-западе Тихого океана
США, лишайники могут обеспечить основной вклад
азота, который поддерживает другие формы жизни
(см. дополнительную литературу).
В некоторых
экосистемы, такие как пустынные почвы, тундровые пустоши,
и дугласово-пихтовые леса на северо-западе Тихого океана
США, лишайники могут обеспечить основной вклад
азота, который поддерживает другие формы жизни
(см. дополнительную литературу). 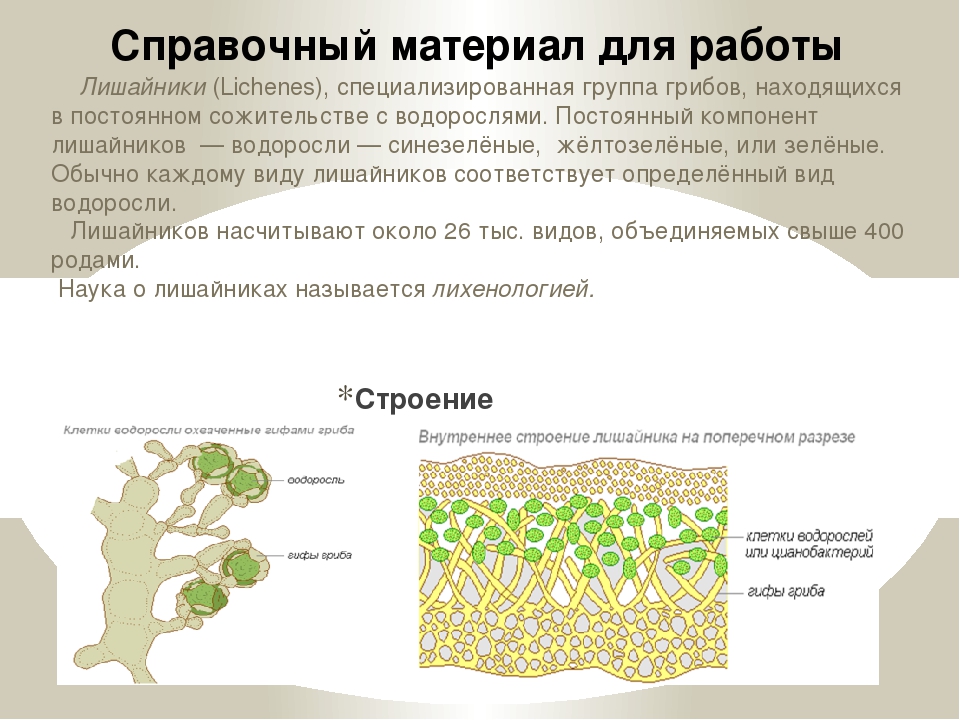 Большинство лишайников, содержащих зеленые водоросли, могут восстановиться после
засуху, поглощая воду из влажного воздуха, а затем начинают
к фотосинтезу.Однако лишайники, содержащие
цианобактерии могут возобновить фотосинтез только после
поглощая свободную (жидкую) воду.
Большинство лишайников, содержащих зеленые водоросли, могут восстановиться после
засуху, поглощая воду из влажного воздуха, а затем начинают
к фотосинтезу.Однако лишайники, содержащие
цианобактерии могут возобновить фотосинтез только после
поглощая свободную (жидкую) воду. 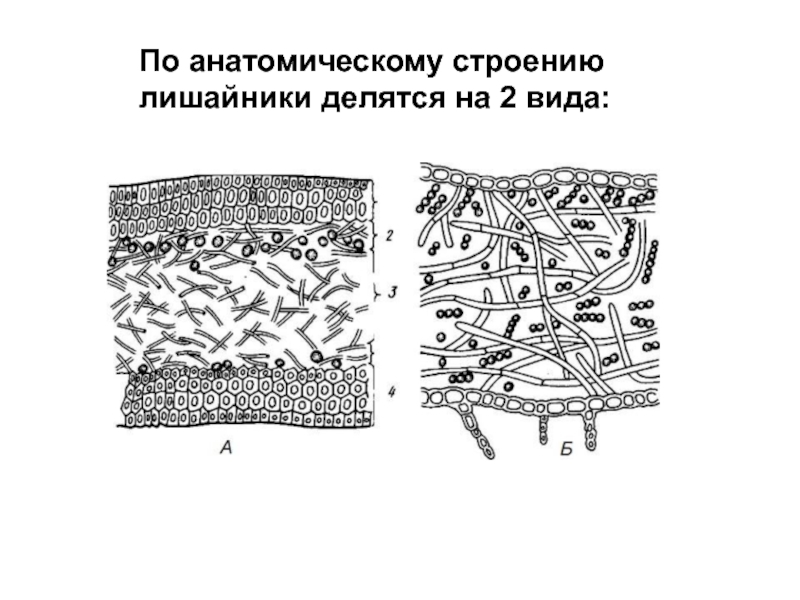
 Это похоже на поведение биотрофных растений.
патогены (см. Биотрофные патогены растений). Напротив, у лишайников с
цианобактерии (например, Peltigera ) нет
проникновение фотосинтезирующих клеток.Вместо этого
грибные гифы центральной области (обычно гидрофобные)
образуют тонкостенные выступы, проникающие в
гидрофильные студенистые оболочки, окружающие
цианобактериальные клетки.
Это похоже на поведение биотрофных растений.
патогены (см. Биотрофные патогены растений). Напротив, у лишайников с
цианобактерии (например, Peltigera ) нет
проникновение фотосинтезирующих клеток.Вместо этого
грибные гифы центральной области (обычно гидрофобные)
образуют тонкостенные выступы, проникающие в
гидрофильные студенистые оболочки, окружающие
цианобактериальные клетки.  Однако цианобактерии
по-видимому, выделяют глюкозу грибковому партнеру. Это кажется
происходит пассивно через переносчик глюкозы в клетке
мембраны после ферментативной деградации внутриклеточного
глюкан (полимер глюкозы) у цианобактерий.
Однако цианобактерии
по-видимому, выделяют глюкозу грибковому партнеру. Это кажется
происходит пассивно через переносчик глюкозы в клетке
мембраны после ферментативной деградации внутриклеточного
глюкан (полимер глюкозы) у цианобактерий. 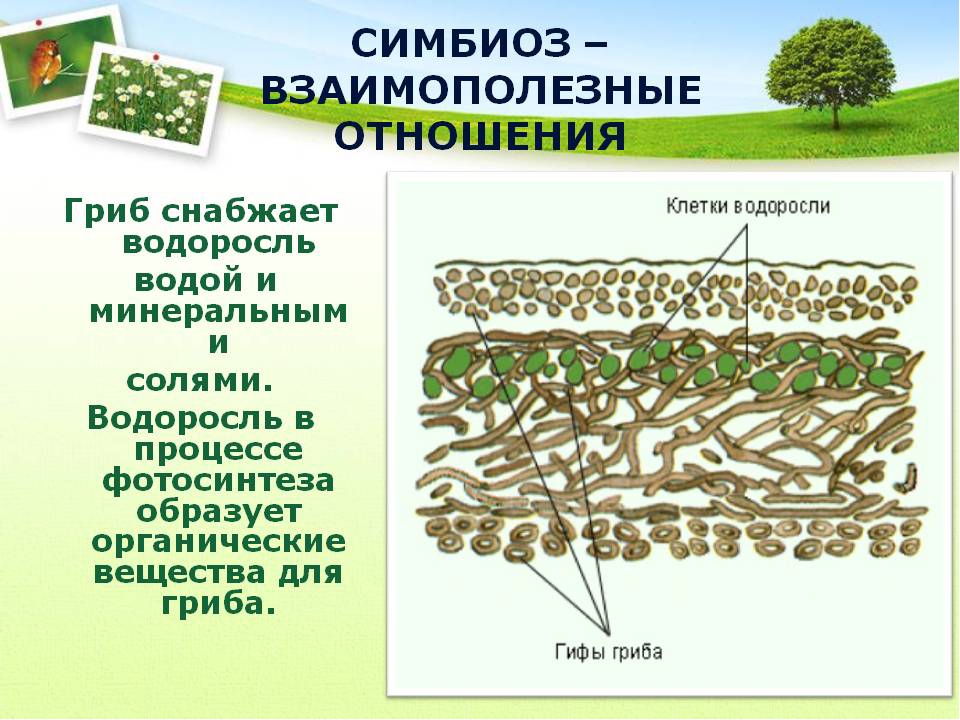 То
апотеций состоит из множества булавовидных асков (один
ascus отмечен стрелкой) со стерильными
«упаковочные» гифы ( парафизов ) между
аски.Кончики парафизов окрашены в оранжевый цвет.
(верхняя часть раздела). ( M ) Часть апотеция
раздавлен, чтобы показать аски (один отмечен стрелкой),
каждый из которых содержит 8 овальных аскоспор , а
парафизы с оранжевыми кончиками.
То
апотеций состоит из множества булавовидных асков (один
ascus отмечен стрелкой) со стерильными
«упаковочные» гифы ( парафизов ) между
аски.Кончики парафизов окрашены в оранжевый цвет.
(верхняя часть раздела). ( M ) Часть апотеция
раздавлен, чтобы показать аски (один отмечен стрелкой),
каждый из которых содержит 8 овальных аскоспор , а
парафизы с оранжевыми кончиками.  В этом лишайнике аскоспоры
игольчатые, в отличие от овальных аскоспор Xanthoria .
В этом лишайнике аскоспоры
игольчатые, в отличие от овальных аскоспор Xanthoria . 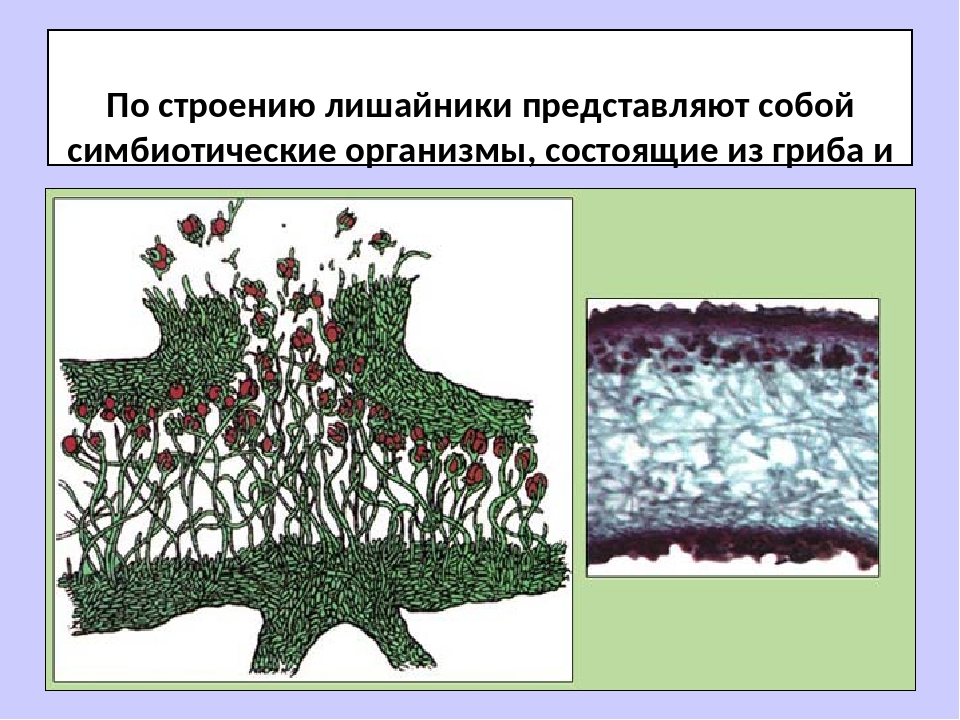 г. Пармелия физодес ,
Рисунок А) на кончиках некоторых долей слоевища.
Соредии легко разносятся ветром.
г. Пармелия физодес ,
Рисунок А) на кончиках некоторых долей слоевища.
Соредии легко разносятся ветром.  За
например, раздельное рассеивание грибков и водорослей ( Trebouxia )
споры могут быть основным средством распространения
лишайник обыкновенный Xanthoria parietina (см. рис. J)
и для видов Buellia видов, потому что эти лишайники
вроде нет соредии.В этих случаях имеется
иммунологические данные, позволяющие предположить, что лишайники
синтезируется в природе из свободноживущих Trebouxia
и споры грибов на каменных поверхностях (Мухтар и др. .,
1994).
За
например, раздельное рассеивание грибков и водорослей ( Trebouxia )
споры могут быть основным средством распространения
лишайник обыкновенный Xanthoria parietina (см. рис. J)
и для видов Buellia видов, потому что эти лишайники
вроде нет соредии.В этих случаях имеется
иммунологические данные, позволяющие предположить, что лишайники
синтезируется в природе из свободноживущих Trebouxia
и споры грибов на каменных поверхностях (Мухтар и др. .,
1994). 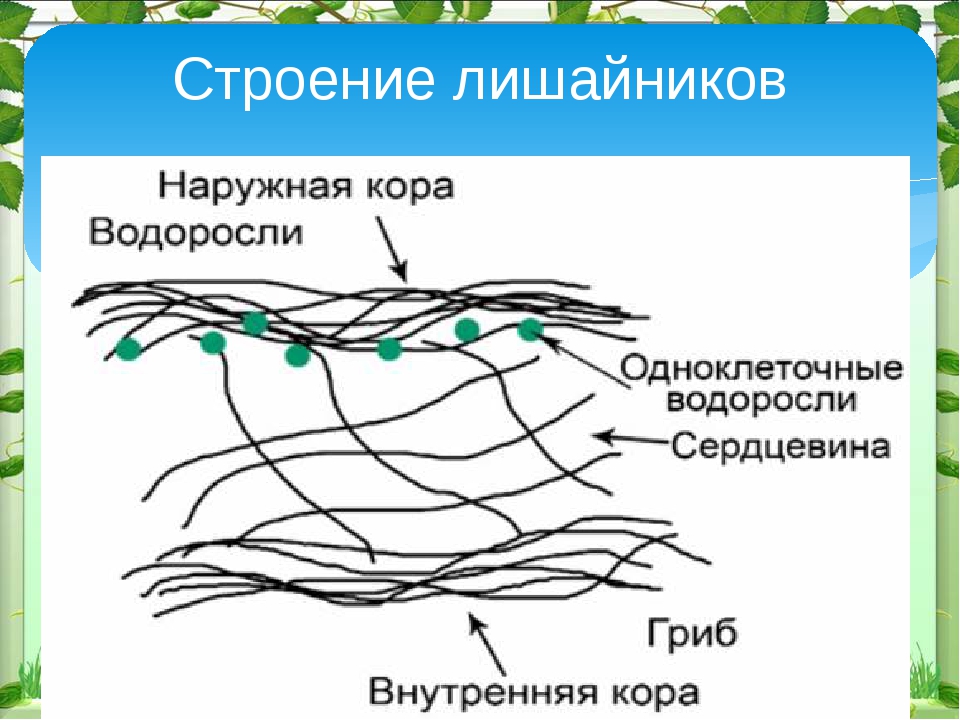 Семь соредий видны на высоте
увеличение. Каждый имеет диаметр около 0,1 мм и состоит из
скопление клеток водорослей в сети гиф.
Семь соредий видны на высоте
увеличение. Каждый имеет диаметр около 0,1 мм и состоит из
скопление клеток водорослей в сети гиф.  Часть
причина такой непереносимости — крайняя эффективность
лишайников в накоплении питательных веществ из следовых
уровней в атмосфере. Экстремальная демонстрация
это способность лишайников накапливать радиоактивные
изотопы из окружающей среды. Вслед за Чернобылем
бедствие, лишайники (в основном Cladonia rangiferina ,
«оленьий мох») северной Скандинавии
накопилось столько радиоактивности, что олени кормятся
на них считались опасными для употребления в пищу человеком.
Часть
причина такой непереносимости — крайняя эффективность
лишайников в накоплении питательных веществ из следовых
уровней в атмосфере. Экстремальная демонстрация
это способность лишайников накапливать радиоактивные
изотопы из окружающей среды. Вслед за Чернобылем
бедствие, лишайники (в основном Cladonia rangiferina ,
«оленьий мох») северной Скандинавии
накопилось столько радиоактивности, что олени кормятся
на них считались опасными для употребления в пищу человеком.
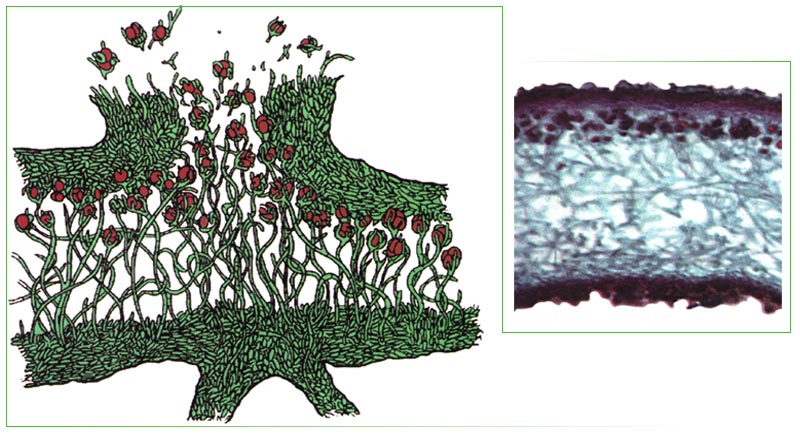 лихенолог 23 ,
127-138.
лихенолог 23 ,
127-138.