
Тест по биологии на тему «Критерии вида» (11 класс)
КРИТЕРИИ ВИДА вариант 1
1. Критерий, в основе которого лежит сходство процессов жизнедеятельности у особей вида, называют
1) физиологическим
2) генетическим
3) морфологическим
4) биохимическим
2. Для определения вида недостаточно использовать только морфологический критерий, так как
1) существуют виды-двойники
2) виды разделены на популяции
3) виды между собой скрещиваются
4) близкие виды могут занимать разные ареалы
3. Генетическому критерию вида соответствует утверждение
1) оперение самки и самца утки кряквы различно
2) нерест популяций форели происходит в разное время
3) лютик едкий и лютик ползучий имеют разные ареалы
4) виды-двойники малярийного комара имеют разные кариотипы
4. Совокупность факторов внешней среды, в которой обитает вид, — это критерий вида
1) экологический
2) генетический
4) морфологический
5. Морфологический критерий вида характеризуется
Морфологический критерий вида характеризуется
1) положением вида в биогеоценозе
2) соотношением числа популяций внутри вида
3) совокупностью внешних и внутренних признаков особей
4) положением вида в системе органического мира
6. По какому морфологическому признаку можно отличить млекопитающих от других позвоночных животных
1) Пятипалая конечность
2) Волосяной покров
3) Наличие хвоста
4) Глаза, прикрытые веками
7. Вид — совокупность особей, обладающих
1) сходным набором хромосом
2) способностью образовывать пищевые связи между его особями
3) обитающих в разных экосистемах
4) созданных человеком на основе отбора
8. Прочитайте текст. Выберите три предложения, в которых даны описания генетического критерия вида волка европейского. Запишите цифры, под которыми они указаны.
(1) Волк европейский распространён в Прибалтике, России, Белоруссии и других странах. (2) Кариотип волка представлен 78 хромосомами. (3) Европейский волк – хищник, достигающий 1,6 м в длину и 90 см в высоту. (4) Ведёт стайный образ жизни. (5) Гибриды волков и немецких овчарок плодовиты. (6) При скрещивании волка, например, с американским волком (койотом) рождается бесплодное потомство.
(3) Европейский волк – хищник, достигающий 1,6 м в длину и 90 см в высоту. (4) Ведёт стайный образ жизни. (5) Гибриды волков и немецких овчарок плодовиты. (6) При скрещивании волка, например, с американским волком (койотом) рождается бесплодное потомство. 9. Установите соответствие между характеристикой вида Дикобраз азиатский и критерием вида, к которому её относят: 1) морфологический, 2) физиологический, 3) экологический. Запишите цифры 1, 2 и 3 в правильном порядке.
А) Лапы снабжены длинными когтями.
Б) Животные питаются растительной пищей.
В) Беременность самок длится 110-115 дней.
Г) Самые длинные и более редкие иглы растут на пояснице животных.
Е) Животные ведут ночной образ жизни
КРИТЕРИИ ВИДА вариант 2
1. Виды-двойники сходны по строению, поэтому их нельзя определить только по критерию
1) генетическому
2) географическому
3) морфологическому
4) физиологическому
2. В чем проявляется относительный характер морфологического критерия вида
В чем проявляется относительный характер морфологического критерия вида
1) ареалы разных видов совпадают
2) наборы хромосом у разных видов одинаковые
3) самцы и самки одного вида различаются внешне
4) разные виды обитают в сходных условиях
3. Видом называют группу особей
1) скрещивающихся и дающих плодовитое потомство
2) обитающих в одном природном сообществе
3) обитающих на общей территории
4) принадлежащих к одному сорту или породе
4. Совокупность факторов внешней среды, в которой обитают особи вида, — критерий
1) экологический
2) географический
3) физиологический
4) морфологический
5. Бурого медведя и очкового медведя ученые считают разными видами потому, что
1) они отличаются внешним видом
2) они живут на разных территориях
3) между ними существует репродуктивная изоляция
4) они питаются разной пищей
6. В каком случае двух зайцев можно отнести к разным видам?
1) если у них разное число хромосом в клетках
2) если они живут на разных территориях
3) если одна группа менее плодовита, чем другая
4) если по численности одна группа больше другой
7. По морфологическому критерию птицы отличаются от других хордовых
По морфологическому критерию птицы отличаются от других хордовых
1) хромосомным набором
2) перьевым покровом
3) способностью к полету
4) интенсивным обменом веществ
Установите соответствие между признаками свиного цепня и критериями вида: 1) морфологический, 2) экологический, 3) физиологический. Запишите цифры 1, 2, 3 в порядке, соответствующем буквам.
А) размер тела до 3 м
Б) на голове кроме присосок имеются крючья
В) взрослый червь обитает в тонком кишечнике человека
Г) размножается партеногенетически
Д) личинки развиваются в теле домашних и диких свиней
Е) свиные цепни высоко плодовиты
6.
 1 Вид, критерий вида — Биология Егэ
1 Вид, критерий вида — Биология ЕгэРаздел ЕГЭ: 6.1. Вид, его критерии. Популяция — структурная единица вида и элементарная единица эволюции. Микроэволюция. Образование новых видов. Способы видообразования. Сохранение многообразия видов как основа устойчивости биосферы
Вид — совокупность особей, имеющих наследственное сходство морфологических, физиологических и биохимических особенностей, способных свободно скрещиваться и давать плодовитое потомство, занимающих определённый ареал и одинаково изменяющихся под влиянием факторов внешней среды.
Чтобы отнести особей к одному и тому же или к разным видам, их сравнивают по критериям — эволюционно устойчивым диагностическим признакам, характерным для одного вида, но отсутствующим у других.
Подвид — географически или экологически обособленная часть вида, особи которой под влиянием факторов среды приобрели устойчивые особенности, отличающие ее от других особей того же вида; особи разных подвидов могут свободно скрещиваться в природе.
Экологическая ниша — пространственное и трофическое место вида в биогеоценозе (экосистеме), комплекс его связей с другими видами и требований к физической среде обитания.
Популяция — совокупность свободно скрещивающихся особей одного вида, длительно существующих на определённой части ареала и относительно обособленных от других совокупностей. Термин «популяция» ввёл датский биолог В. Иогансен в 1903 г.
Каждая популяция характеризуется плотностью (количество особей на единицу площади), численностью (общее количество особей популяции), возрастным составом (соотношение молодых, зрелых и старых особей), половой структурой (соотношение самок и самцов), пространственной структурой (характер размещения особей).
Типы пространственного распределения особей:
Популяции (по характеру использования территории):
Численность популяции, ее плотность могут периодически и не периодически изменяться под влиянием разнообразных факторов. Такие колебания численности популяции называются популяционными волнами, или «волнами жизни». Это понятие ввел русский биолог С. С. Четвериков.
Это понятие ввел русский биолог С. С. Четвериков.
«Волны жизни» могут резко изменить генофонд популяции за счет дрейфа генов.
Тест: Критерии вида.
Вариант № 1.
1. Морфологический критерий вида основан на:
1) сходстве внешнего и внутреннего строения особей
2) сходстве всех процессов жизнедеятельности у особей
3) специфичном для каждого вида наборе хромосом
4) однообразии факторов среды обитания вида
2. Область распространения северного оленя – пример критерия:
1) экологического 2) генетического 3) морфологического 4) географического
3. Критерий, в основе которого лежит сходство процессов жизнедеятельности у особей вида,
называют:
1) физиологическим 2) генетическим 3) морфологическим 4) биохимическим
4. Для определения вида недостаточно использовать только морфологический критерий,
так как:
1) существуют виды-двойники 2) виды разделены на популяции
3) виды между собой скрещиваются 4) близкие виды могут занимать разные ареалы
5. Найдите название критерия вида в указанном перечне
Найдите название критерия вида в указанном перечне
1) цитологический 2) гибридологический 3) генетический 4) популяционный
6. Укажите положение, отражающее генетический критерий вида:
1) вид дифференцирован в пространстве
2) особи вида имеют одинаковый набор хромосом
3) особи вида характеризуются сходными внешними признаками
4) особи разных видов скрещиваются между собой
7. Генетическому критерию вида соответствует утверждение:
1) оперение самки и самца утки кряквы различно
2) нерест популяций форели происходит в разное время
3) лютик едкий и лютик ползучий имеют разные ареалы
4) виды-двойники малярийного комара имеют разные кариотипы
8. Совокупность факторов внешней среды, в которой обитает вид, — это критерий вида:
1) экологический 2) генетический 3) физиологический 4) морфологический
9. Морфологический критерий вида характеризуется:
Морфологический критерий вида характеризуется:
1) положением вида в биогеоценозе
2) соотношением числа популяций внутри вида
3) совокупностью внешних и внутренних признаков особей
4) положением вида в системе органического мира
10. Совокупность внешних признаков особей относят к критерию вида:
1) географическому 2) генетическому 3) морфологическому 4) экологическому
11. По морфологическому критерию птицы отличаются от других хордовых:
1) хромосомным набором 2) перьевым покровом
3) способностью к полету 4) интенсивным обменом веществ
12. При распознавании видов-двойников учитывается главным образом критерий:
1) генетический 2) географический 3) морфологический 4) физиологический
13. Общую территорию, которую занимает вид, называют:
1) экологической нишей 2) биоценозом 3) ареалом 4) кормовой территорией
14. По какому морфологическому признаку можно отличить млекопитающих от других
По какому морфологическому признаку можно отличить млекопитающих от других
позвоночных животных:
1) пятипалая конечность 2) волосяной покров 3) наличие хвоста 4) глаза, прикрытые веками
15. Вид — совокупность особей, обладающих:
1) сходным набором хромосом
2) способностью образовывать пищевые связи между его особями
3) обитающих в разных экосистемах
4) созданных человеком на основе отбора
16. Почему нельзя определить вид только по генетическому критерию:
1) ареалы разных видов совпадают
2) набор хромосом у разных видов может совпадать
3) разные виды обитают в сходных условиях
4) особи разных видов сходны по строению
17. Физиологический критерий вида проявляется у всех его особей в сходстве:
1) процессов жизнедеятельности
2) строения и формы хромосом
3) химического состава
4) внешнего и внутреннего строения
Тест: Критерии вида.
Вариант № 2.
1. Ареал обитания вида является критерием:
1) физиологическим 2) генетическим 3) географическим 4) морфологическим
2. Виды-двойники сходны по строению, поэтому их нельзя определить только по критерию:
1) генетическому 2) географическому 3) морфологическому 4) физиологическому
3. Определенный набор хромосом у особей одного вида считают критерием:
1) биологическим 2) морфологическим 3) генетическим 4) цитологическим
4. В чем проявляется относительный характер морфологического критерия вида:
1) ареалы разных видов совпадают
2) наборы хромосом у разных видов одинаковые
3) самцы и самки одного вида различаются внешне
4) разные виды обитают в сходных условиях
5. Большая синица живет в кронах деревьев, питается крупными насекомыми и их
личинками — это описание критерия вида:
1) географического 2) экологического 3) морфологического 4) генетического
6. Морфологический критерий вида – это:
Морфологический критерий вида – это:
1) его область распространения
2) особенности процессов жизнедеятельности
3) особенности внешнего и внутреннего строения
4) определенный набор хромосом и генов
7. Видом называют группу особей:
1) скрещивающихся и дающих плодовитое потомство
2) обитающих в одном природном сообществе
3) обитающих на общей территории
4) принадлежащих к одному сорту или породе
8. Генетический критерий вида характеризуется:
1) сходством биохимического состава 2) внешними признаками
3) набором хромосом 4) сходством процессов жизнедеятельности
9. Пространственное размещение вида в природе — это критерий:
1) физиологический 2) генетический 3) географический 4) морфологический
10. Относительность географического критерия вида состоит в том, что:
1) ареалы разных видов совпадают
2) наборы хромосом у разных видов одинаковые
3) самцы и самки одного вида различаются внешне
4) разные виды обитают в сходных условиях
11. Совокупность факторов внешней среды, в которой обитают особи вида, — критерий:
Совокупность факторов внешней среды, в которой обитают особи вида, — критерий:
1) экологический
2) географический
3) физиологический
4) морфологический
12. Бурого медведя и очкового медведя ученые считают разными видами потому, что:
1) они отличаются внешним видом
2) они живут на разных территориях
3) между ними существует репродуктивная изоляция
4) они питаются разной пищей
13. Прочитайте описание песца: «В отличие от лисицы, тело у песца приземистое, морда укороченная; уши закруглены, слабо выступают из зимней шерсти (это предохраняет их от обморожения)». В этом описании используется критерий вида:
1) генетический 2) географический 3) морфологический 4) физиологический
14 . Наличие у человека 46 хромосом, характеризует критерий:
1) физиологический 2) генетический 3) географический 4) биохимический
15. Двух зайцев можно отнести к разным видам, в случае:
Двух зайцев можно отнести к разным видам, в случае:
1) если у них разное число хромосом в клетках
2) если они живут на разных территориях
3) если одна группа менее плодовита, чем другая
4) если по численности одна группа больше другой
16. К экологическму критерию вида обыкновенного ежа можно отнести:
1) распространение в Европе
2) число хромосом в соматических клетках
3) наземный образ жизни
4) наличие игл на теле
17. Заяц-беляк и заяц-русак принадлежат:
1) к одному виду
2) к одной популяции
3) к разным отрядам
4) к разным видам
Критерии вида ЕГЭ.docx
Тест по теме «Критерии вида»1 вариант 2 вариант
1.Совокупность особей, сходных по строению, имеющих общее происхождение, свободно скрещивающихся между собой и дающих плодовитое потомство, называются…
А. Популяцией
Б. Видом
В. Классом
Г. Верного ответа нет 1.
 Сходство внешнего и внутреннего строения особей одного вида.
Сходство внешнего и внутреннего строения особей одного вида.А) генетический критерий
Б) морфологический критерий
В) экологический критерий
Г) географический критерий
2. Для видов обитающих в Байкале, ареал ограничивается этим озером, — это пример … критерия
А. Экологического
Б. Морфологического В. Географического
Г. Физиологического 2. Сходство всех процессов жизнедеятельности у особей одного вида.
А) биохимический критерий
Б) генетический критерий
В) морфологический критерий
Г) физиологический
3. Критерий вида, включающий в себя совокупность факторов внешней среды, составляющих непосредственную среду обитания вида, — это … критерий А. Экологический
В. Морфологический Г. Верного ответа нет
3. Сходство химического состава особей одного вида.
А) морфологический критерий
Б) биохимический критерий
В) генетический критерий
Г) экологический критерий
4. Часто скрещиваются между собой виды тополей и ив, — это пример не абсолютности … критерия
А.
 Генетического
Генетического Б. Биохимического В. Физиологического
Г. Морфологического
4. Определённый набор и форма хромосом у особей одного вида.
А) биохимический критерий
Б) морфологический критерий
В) генетический критерий
Г) физиологический
5. Основополагающим для вида критерием является… А. МорфологическийБ. Генетический В. Физиологический
Г. Биохимический
5. Обитание особей одного вида в сходных условиях.
А) морфологический критерий
Б) биохимический критерий
В) географический критерий
Г) экологический критерий
6. Для определения видовой принадлежности необходимо использовать: А. Морфологический и генетический критерии
Б. Биохимический и физиологический критерии
В. Географический и экологический критерий
Г. Все ответы верны
6. Общий ареал, занимаемый видом в природе.
А) географический критерий
Б) экологический критерий
В) морфологический критерий
Г) генетический критерий
7. Болотная камышовка и тростниковая камышовка внешне не отличаются, но не скрещиваются и имеют совершенно разные брачные песни, — это пример не абсолютности
А.
 Морфологического критерия
Морфологического критерия Б. Экологического критерия В. Географического критерия
Г. Биохимического критерия
7. Пеночки — теньковки и пеночки — веснички, обитающие в одном лесу, составляют:
в) две популяции одного вида;
б) две популяции двух видов;
г) одну популяцию разных видов.
8. Критерий, характеризующий определённый ареал, занимаемый видом в природе, — это…
А. Экологический критерий
Б. Морфологический критерий В. Географический критерий
Г. Физиологический критерий
8. Исходная единица систематики организмов:
а) вид;
б) род;
в) популяция;
г) отдельная особь
9. В природных условиях популяции не смешиваются друг с другом. Этому препятствуют…
А. Географические преграды
Б. Морфологические отличия В. Разные сроки размножения
Г. Все ответы верны
9. Видом называется группа особей,
А) обитающих на общей территории
Б) появившаяся в результате эволюции
В) скрещивающихся и дающих плодовитое потомство
Г) созданных человеком на основе отбора
10.
 Виды-двойники
Виды-двойникиА) способны скрещиваться между собой и давать плодовитое потомство
Б) отличаются морфологически
В) обладают разным кариотипом
Г) способны скрещиваться между собой, но не дают плодовитое потомство
10. Ночной образ жизни сурка обыкновенного относится к критерию вида
А) Генетическому
Б)Морфологическому
В) Географическому
Г) Экологическому
11. Какому критерию вида соответствует следующее описание: большая синица живет в кронах деревьев, питается крупными насекомыми и их личинками?
А) географическому
Б) экологическому
В) морфологическому
Г) генетическому 11. . Пространственное размещение вида в природе — это критерий
А) физиологический
Б) генетический
В) географический
Г) морфологический
12. Совокупность внешних признаков особей относят к критерию вида
А) географическому
Б) генетическому
В) морфологическому
 Определите количество видов и критерии, которыми надо руководствоваться:
Определите количество видов и критерии, которыми надо руководствоваться:а) один вид, морфологическим критерием;
б) два вида, физиологическим критерием;
в) один вид, генетическим критерием;
г) два вида, морфологическим критерием.
В1.Установите соответствие между признаками большого прудовика и критериями вида, для которых они характерны.
Признаки большого прудовика:
Критерии вида:
1.Органы чувств – одна пара щупалец
2. Коричневый цвет раковины.
3. Населяет пресные водоёмы.
4. Питается мягкими тканями растений.
5. Раковина спирально закрученная.
А. Морфологический.
Б. Экологический.
Вид — это совокупность особей, сходных по критериям вида до такой степени, что они могут в естественных условиях скрещиваться и давать плодовитое потомство.
Плодовитое потомство — которое само может размножаться. Пример неплодовитого потомства — мул, гибрид осла и лошади.
Критерии вида
— это признаки, по которым сравнивают 2 организма, чтобы определить, относятся они к одному виду или к разным.
1) Морфологический — внутреннее и внешнее строение
2) Физиолого-биохимический — как работают органы и клетки.
3) Поведенческий — поведение, особенно в момент размножения
4) Экологический — совокупность факторов внешней среды, необходимых для жизни вида
5) Географический — ареал, т.е. территория, на которой живет данный вид.
6) Генетико-репродуктивный — сходство по количеству и строению хромосом, позволяющее организмам давать плодовитое потомство.
По одному критерию нельзя судить о виде. Например:
1) Существуют виды-двойники, которые морфологически друг от друга не отличаются, но имеют разное количество хромосом и генетико-репродуктивный критерий не дает им размножаться.
2) Все критерии, кроме генетико-репродуктивного, позволяют ослу и лошади размножаться.
Тесты
1. Морфологический критерий вида основан на
А) сходстве внешнего и внутреннего строения особей
Б) сходстве всех процессов жизнедеятельности у особей
В) специфичном для каждого вида наборе хромосом
Г) однообразии факторов среды обитания вида
2. К какому критерию вида относят область распространения северного оленя
К какому критерию вида относят область распространения северного оленя
А) экологическому
Б) генетическому
В) морфологическому
Г) географическому
3. Критерий, в основе которого лежит сходство процессов жизнедеятельности у особей вида, называют
А) физиологическим
Б) генетическим
В) морфологическим
Г) биохимическим
4. Для определения вида недостаточно использовать только морфологический критерий, так как
А) существуют виды-двойники
Б) виды разделены на популяции
В) виды между собой скрещиваются
Г) близкие виды могут занимать разные ареалы
5. Найдите название критерия вида в указанном перечне
А) цитологический
Б) гибридологический
В) генетический
Г) популяционный
6. Укажите положение, отражающее генетический критерий вида
А) вид дифференцирован в пространстве
Б) особи вида имеют одинаковый набор хромосом
В) особи вида характеризуются сходными внешними признаками
Г) особи разных видов скрещиваются между собой
7. Генетическому критерию вида соответствует утверждение
Генетическому критерию вида соответствует утверждение
А) оперение самки и самца утки кряквы различно
Б) нерест популяций форели происходит в разное время
В) лютик едкий и лютик ползучий имеют разные ареалы
Г) виды-двойники малярийного комара имеют разные кариотипы
8. Совокупность факторов внешней среды, в которой обитает вид, — это критерий вида
А) экологический
Б) генетический
В) физиологический
Г) морфологический
9. Морфологический критерий вида характеризуется
А) положением вида в биогеоценозе
Б) соотношением числа популяций внутри вида
В) совокупностью внешних и внутренних признаков особей
Г) положением вида в системе органического мира
10. Совокупность внешних признаков особей относят к критерию вида
А) географическому
Б) генетическому
В) морфологическому
Г) экологическому
11) По морфологическому критерию птицы отличаются от других хордовых
А) хромосомным набором
Б) перьевым покровом
В) способностью к полету
Г) интенсивным обменом веществ
12. При распознавании видов-двойников учитывается главным образом критерий
При распознавании видов-двойников учитывается главным образом критерий
А) генетический
Б) географический
В) морфологический
Г) физиологический
13. Реальность существования видов в природе подтверждается
А) группой критериев, отличающих один вид от другого
Б) наличием популяций разных видов, обитающих в одной среде
В) совпадением ареалов организмов разных видов
Г) морфологическим различием мужских и женских особей одного вида
14. Определенный набор хромосом у особей одного вида считают критерием
А) биологическим
Б) морфологическим
В) генетическим
Г) цитологическим
15. Общую территорию, которую занимает вид, называют
А) экологической нишей
Б) биоценозом
В) ареалом
Г) кормовой территорией
16. Способность растений скрещиваться и давать плодовитое потомство — это основной признак
А) рода
Б) отдела
В) класса
Г) вида
17. Почему нельзя определить вид только по генетическому критерию
Б) набор хромосом у разных видов может совпадать
В) разные виды обитают в сходных условиях
Г) особи разных видов сходны по строению
18. Физиологический критерий вида проявляется у всех его особей в сходстве
Физиологический критерий вида проявляется у всех его особей в сходстве
А) процессов жизнедеятельности
Б) строения и формы хромосом
В) химического состава
Г) внешнего и внутреннего строения
19. Ареал обитания вида является критерием
А) физиологическим
Б) генетическим
В) географическим
Г) морфологическим
20. Виды-двойники сходны по строению, поэтому их нельзя определить только по критерию
А) генетическому
Б) географическому
В) морфологическому
Г) физиологическому
21. Определенный набор хромосом у особей одного вида считают критерием
А) биологическим
Б) морфологическим
В) генетическим
Г) цитологическим
22. В чем проявляется относительный характер морфологического критерия вида
А) ареалы разных видов совпадают
Б) наборы хромосом у разных видов одинаковые
В) самцы и самки одного вида различаются внешне
Г) разные виды обитают в сходных условиях
23. Большая синица живет в кронах деревьев, питается крупными насекомыми и их личинками — это описание критерия вида
Большая синица живет в кронах деревьев, питается крупными насекомыми и их личинками — это описание критерия вида
А) географического
Б) экологического
В) морфологического
Г) генетического
24. Морфологический критерий вида — это
А) его область распространения
Б) особенности процессов жизнедеятельности
В) особенности внешнего и внутреннего строения
Г) определенный набор хромосом и генов
25. Видом называют группу особей
А) скрещивающихся и дающих плодовитое потомство
Б) обитающих в одном природном сообществе
В) обитающих на общей территории
Г) принадлежащих к одному сорту или породе
26. Генетический критерий вида характеризуется
А) сходством биохимического состава
Б) внешними признаками
В) набором хромосом
Г) сходством процессов жизнедеятельности
27. Число хромосом в клетке
А) индивидуально у каждого организма в популяции
Б) одинаково, как в животных, так и в растительных клетках
В) одинаково у всех представителей данного вида
Г) одинаково у всех представителей семейства
28. Применять к описанию вида растения географический критерий — это значит охарактеризовать
Применять к описанию вида растения географический критерий — это значит охарактеризовать
А) среду его обитания
Б) область его распространения
В) особенности строения цветков
Г) особенности строения вегетативных органов
29. Критерии вида — это
А) популяционная структура вида
Б) способность особей к эволюции
В) совокупность признаков, свойственных данному виду
Г) способность организмов к саморегуляции
30. Способность различных видов люпина синтезировать и накапливать определенные алкалоиды — это показатель критерия
А) экологического
Б) морфологического
В) биохимического
Г) географического
31. Какую характеристику вида Росянка круглолистная следует отнести к физиологическому критерию?
А) цветки правильные, белые, собраны в соцветие кисть
Б) использует в пищу белки насекомых
В) распространен по торфяным болотам
Г) листья образуют прикорневую розетку
32. Применить к описанию вида растения морфологический критерий — это значит охарактеризовать его
А) область распространения
Б) среду обитания
В) время цветения
Г) особенности строения
33. Применить к описанию вида животного экологический критерий — это значит охарактеризовать
Применить к описанию вида животного экологический критерий — это значит охарактеризовать
А) изменчивость признаков в пределах нормы реакции
Б) совокупность внешних признаков
В) размер его ареала
Г) набор предполагаемых кормов
34) Какой из перечисленных признаков можно отнести к экологическому критерию вида обыкновенного ежа?
А) распространение в Европе
Б) число хромосом в соматических клетках
В) наземный образ жизни
Г) наличие игл на теле
35. К какому критерию вида следует отнести совокупность факторов среды обитания лютика едкого?
А) географическому
Б) генетическому
В) морфологическому
Г) экологическому
36. Виды — двойники малярийного комара отличаются друг от друга, главным образом, по критерию
А) морфологическому
Б) географическому
В) генетическому
Г) экологическому
«Вид. Критерии вида» ЕГЭ
1. 835 Процветанию вида способствует: 1) гомозиготность особей 2) медленная смена поколений
3) межвидовая гибридизация 4) генетическая неоднородность особей
2. 420 Скрещиванию разных видов синиц, обитающих в пределах одного лесного массива, препятствует:
420 Скрещиванию разных видов синиц, обитающих в пределах одного лесного массива, препятствует:
1) разный хромосомный набор 2) различие потребляемых кормов
3) нарушение светового режима 4) отсутствие мест для гнездования
3. 125. Верны ли следующие суждения о критериях вида.
А. Экологический критерий вида характеризует процессы жизнедеятельности особей.
Б. Географический критерий вида описывает место обитания особей.
1) верно только А 2) верно только Б 3) верны оба суждения 4) оба суждения неверны
4. 26.Совокупность свойств организма, проявляющихся при индивидуальном развитии в определенных условиях
обитания, это: 1) фенотип 2) генотип 3) ген 4) аллель
5. 220. В соответствии с физиологическим критерием особи одного вида:
1) имеют сходную окраску тела 2) особи обитают в одном биогеоценозе
3) образуют плодовитое потомство 4) имеют одинаковый набор генов
6. 1020. Образование нового вида организмов называют:
1) борьбой за существование 2) макроэволюцией 3) микроэволюцией 4) искусственным отбором
7. 320. Окраска и размеры тела особей бурого медведя характеризуют критерий вида:
320. Окраска и размеры тела особей бурого медведя характеризуют критерий вида:
1) физиологический 2) морфологический 3) генетический 4) биохимический
8. 419. Какой критерий вида иллюстрирует одинаковое число и размер хромосом в ядрах клеток особей Клена
остролистного? 1) физиологический 2) морфологический 3) генетический 4) биологический
9. 520. Сроки цветения и плодоношения растений Ночной красавцы характеризуют критерий вида:
1) физиологический 2) экологический 3) генетический 4) морфологический
10. 620. Ареал обитания особей Лягушки остромордой характеризует критерий:
1) физиологический 2) географический 3) генетический 4) экологический
11. 719. Согласно современным научным представлениям вид это:
1) совокупность особей, обитающих в одном биоценозе
2) группа особей, свободно скрещивающихся между собой и дающих плодовитое потомство
3) естественный биоценоз, включающий особей из разных царств живой природы
4) сообщество особей одного пола или возраста
12. 720. Ночной образ жизни и питание насекомыми Ежа европейского характеризуют критерий вида:
720. Ночной образ жизни и питание насекомыми Ежа европейского характеризуют критерий вида:
1) генетический 2) физиологический 3) географический 4) экологический
13. Генетический критерий вида описывает: 1) внешнее строение организмов
2) процессы жизнедеятельности 3) состав основных химических соединений 4) число и строение хромосом
14. 819. Вид представляет собой совокупность: 1) отрядов 2) родов 3) популяций 4) семейств
15. 820. Какой критерий вида характеризует особенность состава веществ клеток живого организма?
1) генетическим 2) морфологическим 3) биохимическим 4) физиологическим
16. 124. Число хромосом постоянно для представителей одного: 1) царства 2) вида 3) рода 4) семейства
17. 1920. Сходство внешнего и внутреннего строения характерно для особей одного:
1) вида 2) биоценоза 3) класса 4) царства
18. Д44. Изменение частоты встречаемости генов в популяции и ограничение обмена генами вызывает:
1) межвидовая борьба 2) мутационная изменчивость 3) модификационная изменчивость 4) изоляция
19. 2120. Какой критерий вида характеризует местообитание его особей в природе?
2120. Какой критерий вида характеризует местообитание его особей в природе?
20. 2225. Верны ли следующие суждения о способах видообразования.
А. Географическая изоляция популяции приводит к возникновению нового вида.
Б. Примером географической изоляции внутри вида служит невозможность скрещивания растений, обитающих
на одной территории, изза разных сроков цветения.
1) верно только А 2) верно только Б 3) оба суждения верны 4) неверны оба суждения
21. 2320. Морфологический критерий вида – это: 1) процессы жизнедеятельности, протекающие в организме
2) место его обитания 3) набор хромосом в клетках организмов 4) внешнее и внутреннее строение особей
22. 2520. Сроки цветения и плодоношения сирени обыкновенной характеризуют критерий вида:
1) морфологический 2) генетический 3) физиологический 4) биохимический
23. 4720. В чем проявляется недостаточность географического критерия вида?
1) существуют видыдвойники 2) структура хромосом меняется по причине мутаций
3) процессы жизнедеятельности в разных условиях протекают сходно
4) некоторые виды, существующие повсеместно
24. 2720. Какой критерий вида характеризует различие в форме кроны и высоты берёзы пушистой, выросшей
2720. Какой критерий вида характеризует различие в форме кроны и высоты берёзы пушистой, выросшей
в лесу и на опушке? 1) морфологический 2) генетический 3) географический 4) экологический
25. 3120. Какой критерий вида характеризует пищевые связи особей в природе?
1) генетический 2) экологический 3) морфологический 4) географический
26. 3025. Выберите верное суждение о критериях вида.
А. Морфологические признаки не обеспечивают определение вида, так как существуют виды двойники.
Б. Согласно биохимическому критерию вида существует видовая специфичность белков и нуклеиновых кислот.
27. 2820. Какой критерий характеризует особенности процессов жизнедеятельности, отличающие между собой
близкородственные виды? 1) географический 2) экологический
3) физиологический 4) популяционно – статистический
28. 3219. Согласно морфологическому критерию виды синиц отличаются друг от друга:
1) хромосомным набором 2) способностью к полету
3) окраской перьевого покрова 4) интенсивностью обмена веществ
29. 4636. Выберите верное суждение о критериях вида.
4636. Выберите верное суждение о критериях вида.
А. В соответствии с экологической характеристикой особи одного вида имеют одинаковый ареал распространения.
Б. Каждый вид имеет свой кариотип, набор хромосом.
1) верно только А 2) верно только Б 3) оба суждения верны 4) оба суждения неверны
30. 4620. В чем состоит недостаточность биохимического критерия вида?
1) процессы жизнедеятельности в разных условиях протекают сходно
2) в пределах вида возможно изменение состава нуклеиновых кислот
3) существуют виды двойники 4) разные виды занимают в природе сходные ареалы
31. 4536. Выберите верное суждение о критериях вида.
А. Морфологические признаки являются главными в определении вида.
Б. Согласно биохимическому критерию вида все его особи отличаются кариотипом.
1) верно только А 2) верно только Б 3) оба суждения верны 4) оба суждения неверны
32. 4020. Виды сохраняются в природе за счет:
1) дрейфа генов 2) наследственной изменчивости 3) борьбы за существование 4) действия факторов эволюции
33. 4120. Главным признаком формирования нового вида считают: 1) дрейф генов
4120. Главным признаком формирования нового вида считают: 1) дрейф генов
2) наличие новых мутаций 3) интенсивное размножение 4) репродуктивную изоляцию
34. 4220. В чем проявляется недостаточность генетического критерия вида?
1) разнополые особи имеют внешнее различие 2) особи имеют сходные процессы жизнедеятельности
3) организмы обладают наследственной изменчивостью 4) один вид организмов занимает определенный ареал
35. 4420. Смешиванию популяций внутри вида мешает:
1) географическая преграда 2) половой состав 3) возраст особей 4) саморегуляция
36. Совокупность внешних признаков особей относят к критерию вида:
1) географическому 2) генетическому 3) морфологическому 4) экологическому
37. По морфологическому критерию птицы отличаются от других хордовых: 1) хромосомным набором
2) перьевым покровом 3) способностью к полету 4) интенсивным обменом веществ
38. При распознавании видовдвойников учитывается главным образом критерий:
1) географический 2) генетический 3) морфологический 4) физиологический
39. Общую территорию, которую занимает вид, называют:
Общую территорию, которую занимает вид, называют:
1) экологической нишей 2) биоценозом 3) ареалом 4) кормовой территорией
40. По какому морфологическому признаку можно отличить млекопитающих от других позвоночных животных:
1) пятипалая конечность 2) волосяной покров 3) наличие хвоста 4) глаза, прикрытые веками
41. Вид совокупность особей, обладающих: 1) способностью образовывать пищевые связи между его особями
2) сходным набором хромосом 3) обитающих в разных экосистемах 4) созданных человеком на основе отбора
42. Почему нельзя определить вид только по генетическому критерию:
1) ареалы разных видов совпадают 2) набор хромосом у разных видов может совпадать
3) разные виды обитают в сходных условиях 4) особи разных видов сходны по строению
43. В чем проявляется относительный характер морфологического критерия вида:
1) ареалы разных видов совпадают 2) наборы хромосом у разных видов одинаковые
3) самцы и самки одного вида различаются внешне 4) разные виды обитают в сходных условиях
44. 120.Какой из перечисленных признаков можно отнести к экологическому критерию вида обыкновенного ежа?
120.Какой из перечисленных признаков можно отнести к экологическому критерию вида обыкновенного ежа?
45. В каком из приведённых примеров дана характеристика морфологического критерия вида птицы большой синицы?
1) питается мелкими насекомыми 2) гнездится на деревьях
3) длина хвоста синицы не превышает длины её тела 4) предпочитает мелколиственные леса
46. Распространение большого пёстрого дятла в пределах ареала соответствует критерию вида:
1) морфологическому 2) физиологическому 3) экологическому 4) географическому
47. 220. К чему приводит появление новых аллелей в популяции?
1) возникновению преград для свободного скрещивания 2) усилению гомозиготности популяции
3) образованию нового вида 4) генетической разновидности популяции
48. 320. Какая из указанных структур является элементарной единицей эволюции?
1) вид 2) популяция 3) сорт 4) биоценоз
49. Признаком вида служит: 1) обитание его особей в одной экосистеме 2) получение плодовитого потомства
3) расселение особей в разные среды обитания 4) размножение его особей бесполым путем
50. Какая характеристика иллюстрирует физиологический критерий вида пастушьей сумки?
Какая характеристика иллюстрирует физиологический критерий вида пастушьей сумки?
1) распространение по полям, дорогам, сорным местам 2) обитание на Европейской части России
3) особенности обмена веществ в растении 4) поочередное расположение листьев на стебле
51. Найдите название критерия вида в указанном перечне:
1) цитологический 2) гибридологический 3) генетический 4) популяционный
52. 620. Систематическая категория, которая реально существует в природе:
1) класс 2) семейство 3) род 4) вид
53. 720. На образование новых видов в природе не влияет: 1) мутационная изменчивость
2) борьба за существование 3) естественный отбор 4) модификационная изменчивость
54. 920. Пространственное размещение вида в природе – это критерий:
1) физиологический 2) генетический 3) географический 4) морфологический
55. 1120.Ареал, занимаемый видом в природе, это критерий:
1) морфологический 2) физиологический 3) экологический 4) географический
56. 2220. Целостность вида обусловлена:
Целостность вида обусловлена:
1) сходными пищевыми потребностями 2) генетическим единством его особей
3) колебаниями численности его особей 4) связями между популяциями разных видов
57. 2320. Генетическое единство популяции животных поддерживается:
1) скрещиванием особей 2) широким расселением особей 3) саморегуляцией 4) пищевыми ценностями
58. Для определения вида недостаточно использовать только морфологический критерий, так как:
1) существуют видыдвойники 2) виды разделены на популяции
3) виды между собой скрещиваются 4) близкие виды могут занимать разные ареалы
59. Применять к описанию вида растения географический критерий – это значит охарактеризовать:
1) среду его обитания 2) область его распространения
3) особенности строения цветков 4) особенности строения вегетативных органов
60. Способность растений скрещиваться и давать плодовитое потомство это основной признак:
1) рода 2) отдела 3) класса 4) вида
61. Большая синица живет в кронах деревьев, питается крупными насекомыми и их личинками это описание критерия
вида: 1) географического 2) экологического 3) морфологического 4) генетического
62. Число хромосом в клетке: 1) индивидуально у каждого организма в популяции
Число хромосом в клетке: 1) индивидуально у каждого организма в популяции
2) одинаково, как в животных, так и в растительных клетках
3) одинаково у всех представителей данного вида 4) одинаково у всех представителей семейства
63. Критерии вида – это: 1) популяционная структура вида 2) способность особей к эволюции
3) совокупность признаков, свойственных данному виду 4) способность организмов к саморегуляции
64. Способность различных видов люпина синтезировать и накапливать определенные алкалоиды – это показатель
критерия: 1) экологического 2) морфологического 3) биохимического 4) географического
65. Какую характеристику вида Росянка круглолистная следует отнести к физиологическому критерию?
1) цветки правильные, белые, собраны в соцветие кисть 2) использует в пищу белки насекомых
3) распространен по торфяным болотам 4) листья образуют прикорневую розетку
66. Применить к описанию вида животного экологический критерий – это значит охарактеризовать:
1) изменчивость признаков в пределах нормы реакции 2) совокупность внешних признаков
3) размер его ареала 4) набор предполагаемых кормов
67. Какой из перечисленных признаков можно отнести к экологическому критерию вида обыкновенного ежа?
Какой из перечисленных признаков можно отнести к экологическому критерию вида обыкновенного ежа?
1) распространение в Европе 2) число хромосом в соматических клетках
3) наземный образ жизни 4) наличие игл на теле
68. Виды – двойники малярийного комара отличаются друг от друга, главным образом, по критерию:
1) морфологическому 2) географическому 3) генетическому 4) экологическому
69. Древесный образ жизни обыкновенной квакши соответствует критерию вида:
1) экологическому 2) географическому 3) морфологическому 4) генетическому
70. Распространение большой синицы в Евразии и СевероЗападной Африки относят к критерию вида:
1) географического 2) экологического 3) морфологического 4) генетического
71. Характерные для вида признаки и свойства называют:
1) альтернативными признаками 2) модификациями 3) критериями 4) аллелями
72. *** 3528. К морфологическому критерию вида относят сходство: 1) поведения особей в период размножения
2) требований к условиям среды обитания 3) строения внутренних органов
4) внешнего строения 5) покровов тела 6) процессов жизнедеятельности
73. 232. Установите соответствие между признаком медузы корнерота и критерием вида, для которого он характерен.
232. Установите соответствие между признаком медузы корнерота и критерием вида, для которого он характерен.
Признаки медузы корнерота
А) обитает в морях и океанах
Б) тело состоит из двух слоев: эктодермы и энтодермы
В) внутри тела имеется кишечная полость
Г) является хищником
Д) на щупальцах расположены стрекательные клетки
1) морфологический
74. Установите соответствие между признаком печеночного сосальщика и критерием вида, для которого он характерен:
ПРИЗНАК ПЕЧЕНОЧНОГО СОСАЛЬЩИКА
А)
живет
Б)
В)
Г)
Д) пищеварительная система имеет ротовое отверстие
жизни
две
тело
образу
имеет
—
по
в
экологический
75. *** 3628. Согласно генетическому критерию вида особи имеют сходство: 1) размера и формы хромосом
2) кариотипа 3) процессов гаметогенеза 4) набора хромосом в соматических клетках
5) процессов пластического обмена 6) строения внутренних органов
76. Установите соответствие между характеристикой вида Дикая свинья (кабан) и критерием вида, к которому эту
характеристику относят:
Свиньи
Животные
ХАРАКТЕРИСТИКА ВИДА ДИКАЯ СВИНЬЯ
А) Число поросят в выводке зависит от упитанности самки и её возраста.
днем.
Б)
жизни.
В)
Г) Окраска особей от светлобурой или серой до чёрной, поросята
полосатые.
Д)
Е) Свиньи предпочитают дубовые и буковые леса.
77. 78. 832. Установите соответствие между признаком моллюска беззубки и критерием вида, который его характеризует.
1) морфологический
2) физиологический
3)
КРИТЕРИЙ ВИДА
активны
экологический
образ
Способ
добывания
стадный
земли.
рытьё
ведут
пищи
А) тело покрыто мантией
Б) раковина имеет две створки
В) обитает в пресных водоемах
Г) кровеносная система незамкнутая
Д) питается водными организмами
1) морфологический
2) экологический
78. Установите соответствие между признаком медоносной пчелы и критерием вида, к которому он относится:
ПРИЗНАК МЕДОНОСНОЙ ПЧЕЛЫ
А)
общественный
в
самцов
размерах
различие
Б)
развитие
В)
наличие
Г)
Д)
Е) глаза фасеточные
79. 1832. Установите соответствие между признаком африканского слона и критерием вида.
1832. Установите соответствие между признаком африканского слона и критерием вида.
Критерий вида
личинок
волосков
и
в
на
пыльцой
нектаром
питание
2)
и
жизни
самок
сотах
теле
цветков
КРИТЕРИЙ ВИДА
1) морфологический
экологический
Признак африканского слона
А) ведут дневной образ жизни
Б) ареал – многие районы Африканского континента
В) встречаются в саваннах и нагорьях
Г) питаются растительными кормами
Д) образуют группы стада
1) экологический
2) географический
80. Установите соответствие между характеристикой вида Дельфин обыкновенный (дельфинбелобочка) и критерием вида,
к которому эту характеристику относят:
ПРИЗНАК ДЕЛЬФИНА ОБЫКНОВЕННОГО
А)
КРИТЕРИЙ ВИДА
Хищники
разными
питаются
на
610
освоили
Самцы
Животные
видами
рыб.
крупнее
самок.
среду обитания.
сантиметров.
Беременность самок продолжается 1011 месяцев.
Б)
В)
Г)
Д)
Е) Животные ведут стадный образ жизни.
81. 3132. Установите соответствие между характеристикой особей ежа обыкновенного и критерием вида,
который она иллюстрирует.
2) физиологический
3)
см
водную
1) морфологический
экологический
Величина
160260
тела
Характеристика
Критерий
А) имеется вытянутая мордочка Б) ведут ночной образ жизни
В) тело покрыто иглами
Г) питаются дождевыми червями и насекомыми
Д) передние конечности копательные
1) морфологический
2) экологический
82. 3532. Установите соответствие между характеристикой и критерием вида, которую она иллюстрирует.
Характеристика
А) последовательность триплетов в ДНК
Б) особенности первичной структуры белка
В) специфичность пищи
Г) своеобразие поведения в брачный период
Д) сходство в составе выдыхаемого воздуха
Критерий
1) биохимический
2) экологический
83. 3732. Установите соответствие между признаком зеленой жабы и критерием вида, для которого он характерен.
Признак
А) ведут сумеречный образ жизни
Б) имеют толстую кожу В) питаются слизнями
Г) задние конечности длиннее передних
Д) встречаются в пустынных областях
Критерий
1) экологический
2) морфологический
84. 4143. Установите соответствие между признаком речного рака и критерием вида, для которого он характерен.
Признак
А) нервная система узлового типа
Б) обитает в проточных водах
В) питается падалью
Г) имеет хитиновый панцирь
Д) наличие 2х пар усиков
Критерий
1) экологический
2) морфологический
*** 85. Выберите три предложения, в которых даны описания морфологического критерия вида сосны обыкновенной.
1) Сосна обыкновенная – светолюбивое растение.
2) Она имеет высокий стройный ствол, крона формируется только вблизи верхушки.
3) Сосна растёт на песчаных почвах, меловых горах.
4) У неё хорошо развиты главный и боковые корни, листья игловидные, по две хвоинки в узле на побеге.
5) На молодых побегах развиваются зеленоватожёлтые мужские шишки и красноватые женские шишки.
6) Пыльца переносится ветром и попадает на женские шишки, где происходит оплодотворение.
*** 86. Выберите из текста три предложения, в которых даны описания экологического критерия вида.
1) Комнатная муха – двукрылое насекомое служит пищей для насекомоядных птиц.
2) Её ротовой аппарат лижущего типа. 3) Взрослые особи мух и их личинки питаются полужидкой пищей.
4) Самки мух откладывают яйца на гниющие органические остатки.
5) Личинки белого цвета, не имеют ног, быстро растут и превращаются в краснобурых куколок.
6) Из куколки развивается взрослая муха.
87. Установите соответствие между признаком крота и критерием вида, к которому этот признак относят:
КРИТЕРИЙ ВИДА
ПРИЗНАК КРОТА
А) тело покрыто короткой шерстью
Б)
роет
очень
ход
глаза
Г) передние лапы широкие
Д)
Е) размножается в гнездовой камере
88. Установите соответствие между признаком большого прудовика и критерием вида, для которого он характерен:
маленькие
почве
копательные
насекомыми
1) морфологический
экологический
питается
2)
в
ПРИЗНАК БОЛЬШОГО ПРУДОВИКА
А)
коричневый
населяет
органы чувств – одна пара щупалец
раковины
водоемы
питается мягкими тканями растений
Б)
В)
Г)
Д) раковина спирально закрученная
89. Установите соответствие между критерием вида и характеристикой голого слизня:
Установите соответствие между критерием вида и характеристикой голого слизня:
1) морфологический
цвет
пресные
2)
КРИТЕРИЙ ВИДА
экологический
ПРИЗНАК ГОЛОГО СЛИЗНЯ
А)
в
и
садах
раковина
огородах
обитает
отсутствует
Б)
В)
мускулистое
Г) питается мягкими тканями наземных растений
Д)
органы чувств 2 пары щупалец
Е) ведет наемный образ жизни
мягкое
тело
КРИТЕРИЙ ВИДА
1) морфологический
2)
экологический
90. Установите соответствие между признаком прыткой ящерицы и критерием вида, который он иллюстрирует:
ПРИЗНАК ПРЫТКОЙ ЯЩЕРИЦЫ
А) зимнее оцепенение Б) длина тела 2528 см
В)
формы
Г) различия в окраске самцов и самок
Д) обитание на опушках лесов, в оврагах и садах
Е) питание насекомыми
91. Установите соответствие между признаком, характеризующим прыткую ящерицу, и критерием вида:
1) морфологический
КРИТЕРИЙ ВИДА
веретеновидной
тело
2)
экологический
ПРИЗНАК ПРЫТКОЙ ЯЩЕРИЦЫ
А) тело имеет бурую окраску Б) питается
КРИТЕРИЙ ВИДА
насекомыми
при низкой температуре неактивна
В)
легкие
Г)
Д) размножается на суше Е) кожа не
имеет желез
92. Установите соответствие между характеристикой вида Дикобраз азиатский и критерием вида, к которому её относят:
Установите соответствие между характеристикой вида Дикобраз азиатский и критерием вида, к которому её относят:
1) морфологический
2) экологический
дыхания
органы
–
ПРИЗНАК ДИКОБРАЗА АЗИАТСКОГО
А)
снабжены
длинными
растительной
питаются
самок
Лапы
Животные
Беременность
когтями.
пищей.
Б)
В)
дней.
Г) Самые длинные и более редкие иглы растут на пояснице
животных.
Д) У самки выделяется молоко после рождения детёнышей.
Е) Животные ведут ночной образ жизни.
длится
110115
КРИТЕРИЙ ВИДА
1) морфологический
2) физиологический
3)
экологический
93.1532. Установите соответствие между признаком серой крысы и критерием вида, для которого он характерен.
КРИТЕРИЙ ВИДА
ПРИЗНАК СЕРОЙ КРЫСЫ
А) живет обычно в постройках человека
Б)
характерно
раннее
пищей
половое
питается
2) экологический
1) физиологический
созревание
В) в южных районах может селиться по берегам рек и других
водоемов
Г)
разнообразной
Тип | Задание | ||
A1 | |||
A2 | |||
A3 | |||
A4 | |||
A5 | |||
A6 | |||
A7 | |||
A8 | |||
A9 | |||
A10 | |||
A11 | |||
A12 | |||
A13 | |||
A14 | |||
A15 | |||
A16 | |||
A17 | |||
A18 | |||
A19 | |||
A20 | |||
A21 | |||
A22 | |||
A23 | |||
A24 | |||
A25 | |||
A26 | |||
A27 | |||
A28 | |||
A29 | |||
A30 | |||
A31 | |||
A32 | |||
A33 | |||
A34 | |||
A35 | |||
A36 | |||
B1 | |||
B2 | |||
B3 | |||
B4 | |||
B5 | |||
B6 | |||
B7 | |||
B8 | |||
C1 |
| ||
C2 |
| ||
C3 |
| ||
C4 |
| ||
C5 |
| ||
C6 |
|
 Изменится ли размер ушей у зайцев в процессе эволюции при действии этой формы естественного отбора? Ответ обоснуйте
Изменится ли размер ушей у зайцев в процессе эволюции при действии этой формы естественного отбора? Ответ обоснуйте Объясните результат в каждом случае
Объясните результат в каждом случаеТема №5166 Эволюция живой природы (Часть 2)
24. Задание 19 Способность к размножению на суше в процессе эволюции животных впервые появилась
Задание 19 Способность к размножению на суше в процессе эволюции животных впервые появилась
1) у земноводных
2) у пресмыкающихся
3) у птиц
4) у млекопитающих
Ответ: 2
25. Задание 19 В палеозойскую эру на суше и в воздухе господствовали
1) птицы
2) млекопитающие
3) пресмыкающиеся
4) членистоногие
Ответ: 4
26. Задание 19 В Мезозойскую доминирующей группой наземных животных были
1) членистоногие
2) птицы
3) стегоцефалы
4) рептилии
Ответ: 4
27. Задание 19 . Современный человек живет в периоде, который называется
1) неоген
2) палеоген
3) антропоген
4) карбон
Ответ: 3
28. Задание 19 . Какие органы являются гомологичными?
1) клешни рака и ногочелюсти паука
2) крыло бабочки и крыло птицы
3) спинной плавник окуня и ласт кита
4) шерсть собаки и шерстинки шмеля
Ответ: 1
29. Задание 19 . Метанефридии кольчатых червей гомологичны
Задание 19 . Метанефридии кольчатых червей гомологичны
1) почке
2) мочевому пузырю
3) нефрону
4) мочеиспускательному каналу
Ответ: 3
30. Задание 19 . К биологическому регрессу ведёт
1) избыточная численность популяции
2) значительное снижение численности популяции
3) увеличение плодовитости
4) расширение ареала
Ответ: 2
31. Задание 19 . К ароморфозам птиц относится появление
1) двух кругов кровообращения
2) теплокровности
3) легких
4) мозжечка
Ответ: 2
32. Задание 19 . Сколько в соответствии с правилом экологической пирамиды необходимо планктона, чтобы в море вырос дельфин весом в 500 кг, при условии, что он будет питаться хищной рыбой?
1) 5 тонн
2) 50 тонн
3) 500 кг
4) 500 тонн
Ответ: 4
33. Задание 19 . Роль мутационного процесса в эволюции состоит в
Задание 19 . Роль мутационного процесса в эволюции состоит в
1) уничтожении наименее приспособленных особей
2) формировании приспособленности организмов к среде обитания
3) усилении борьбы за существование
4) повышении генетической неоднородности особей в популяции
Ответ: 4
34. Задание 19 . На каждый последующий трофический уровень переходит только часть энергии, которая составляет около
1) 1%
2) 10%
3) 50%
4) 90%
Ответ: 2
35. Задание 19 . Творческая роль естественного отбора проявляется в
1) усилении внутривидовой борьбы
2) освоении организмами новых сред обитания
3) возникновении новых мутаций
4) возникновении новых видов
Ответ: 4
36. Задание 19 . Что служит материалом для естественного отбора?
Что служит материалом для естественного отбора?
1) адаптации организмов
2) фенотипическая изменчивость
3) пищевые связи между популяциями
4) комбинации генов
Ответ: 4
37. Задание 19 . Палеонтологическими доказательствами антропогенеза служат
1) признаки сходства человека и позвоночных животных
2) признаки сходства эмбрионов человека и животных
3) атавизмы и рудименты у современных людей
4) ископаемые останки предков человека
Ответ: 4
38. Задание 19 . Какой признак первых наземных позвоночных указывает на их происхождение от древних кистепёрых рыб?
1) особенности строения покровов
2) дыхание с помощью жабр
3) наличие замкнутой кровеносной системы
4) строение парных конечностей
Ответ: 4
39. Задание 19 . К ароморфным изменениям у членистоногих относят появление
Задание 19 . К ароморфным изменениям у членистоногих относят появление
1) нервной системы в виде цепочки
2) конечностей, состоящих из отделов
3) замкнутой кровеносной системы
4) органов зрения и осязания
Ответ: 2
40. Задание 19 . Организмы в экосистеме изменяют среду обитания, создавая тем самым условия для
1) естественной смены сообщества
2) действия массового отбора
3) возникновения мутаций
4) сезонных изменений
Ответ: 1
41. Задание 19 . Эффективность естественного отбора снижается при
1) возникновении рецессивных мутаций
2) увеличении гомозиготных особей в популяции
3) изменении нормы реакции признака
4) увеличении числа видов в экосистеме
Ответ: 2
42. Задание 19 . Какая особенность отличает лицевую часть черепа человека и человекообразной обезьяны?
Задание 19 . Какая особенность отличает лицевую часть черепа человека и человекообразной обезьяны?
1) небольшие носовые кости
2) направленные вперёд глазницы
3) развитые скуловые кости
4) наличие подбородочного выступа
Ответ: 4
43. Задание 19 . Быстрее будут эволюционировать популяции
1) гаплоидных трутней
2) гетерозиготных по многим признакам окуней
3) самцов домашних тараканов
4) мартышек в зоопарке
Ответ: 2
44. Задание 19 . Случайная смена частот аллелей в популяции называется
1) мутация
2) дрейф генов
3) модификация
4) естественный отбор
Ответ: 2
45. Задание 19 . Человек в отличие от человекообразных обезьян
1) обладает рассудочной деятельностью
2) создает сложные орудия труда
3) имеет объемное зрение
4) имеет четырехкамерное сердце
Ответ: 2
46. Задание 19 . Идея биологической эволюции органического мира согласуется с представлениями о
Задание 19 . Идея биологической эволюции органического мира согласуется с представлениями о
1) мутационном процессе
2) самозарождении организмов
3) стремлении организмов к прогрессу
4) наследовании приобретённых признаков
Ответ: 4
47. Задание 19 . Новые приспособления к условиям среды формируются у особей в зависимости от
1) стремления организмов к прогрессу
2) благоприятных условий окружающей среды
3) нормы реакции организмов
4) направления и формы естественного отбора
Ответ: 4
48. Задание 19 . Основным признаком экосистемы является
1) отсутствие конкуренции между видами
2) разнообразие рельефа
3) однообразный видовой состав
4) наличие пищевых связей
Ответ: 4
49. Задание 19 . Какая пара водных позвоночных животных иллюстрирует конвергентное сходство, возникшее в процессе эволюции?
Задание 19 . Какая пара водных позвоночных животных иллюстрирует конвергентное сходство, возникшее в процессе эволюции?
1) европейский осётр и белуга
2) синий кит и кашалот
3) морской котик и морской лев
4) голубая акула и дельфин афалина
Ответ: 4
50. Задание 19 . Малые популяции вымирают из-за того, что в них
1) меньше рецессивных мутаций, чем в больших популяциях
2) меньше вероятность перевода мутаций в гомозиготное состояние
3) больше вероятность близкородственных скрещиваний и наследственных заболеваний
4) выше степень гетерозиготности особей
Ответ: 3
51. Задание 19 . В результате микроэволюционных процессов образуются
1) виды
2) классы
3) типы
4) отряды
Ответ: 1
1. Задание 20 Морфологический критерий вида основан на
Задание 20 Морфологический критерий вида основан на
1) сходстве внешнего и внутреннего строения особей
2) сходстве всех процессов жизнедеятельности у особей
3) специфичном для каждого вида наборе хромосом
4) однообразии факторов среды обитания вида
Ответ: 1
2. Задание 20 К какому критерию вида относят область распространения северного оленя
1) экологическому
2) генетическому
3) морфологическому
4) географическому
Ответ: 4
3. Задание 20 Критерий, в основе которого лежит сходство процессов жизнедеятельности у особей вида, называют
1) физиологическим
2) генетическим
3) морфологическим
4) биохимическим
Ответ: 1
4. Задание 20 Для определения вида недостаточно использовать только морфологический критерий, так как
1) существуют виды-двойники
2) виды разделены на популяции
3) виды между собой скрещиваются
4) близкие виды могут занимать разные ареалы
Ответ: 1
5. Задание 20 Найдите название критерия вида в указанном перечне
Задание 20 Найдите название критерия вида в указанном перечне
1) цитологический
2) гибридологический
3) генетический
4) популяционный
Ответ: 3
6. Задание 20 Укажите положение, отражающее генетический критерий вида
1) вид дифференцирован в пространстве
2) особи вида имеют одинаковый набор хромосом
3) особи вида характеризуются сходными внешними признаками
4) особи разных видов скрещиваются между собой
Ответ: 2
7. Задание 20 Генетическому критерию вида соответствует утверждение
1) оперение самки и самца утки кряквы различно
2) нерест популяций форели происходит в разное время
3) лютик едкий и лютик ползучий имеют разные ареалы
4) виды-двойники малярийного комара имеют разные кариотипы
Ответ: 4
8. Задание 20 Совокупность факторов внешней среды, в которой обитает вид, — это критерий вида
Задание 20 Совокупность факторов внешней среды, в которой обитает вид, — это критерий вида
1) экологический
2) генетический
3) физиологический
4) морфологический
Ответ: 1
9. Задание 20 Морфологический критерий вида характеризуется
1) положением вида в биогеоценозе
2) соотношением числа популяций внутри вида
3) совокупностью внешних и внутренних признаков особей
4) положением вида в системе органического мира
Ответ: 3
10. Задание 20 Совокупность внешних признаков особей относят к критерию вида
1) географическому
2) генетическому
3) морфологическому
4) экологическому
Ответ: 3
11. Задание 20 По морфологическому критерию птицы отличаются от других хордовых
1) хромосомным набором
2) перьевым покровом
3) способностью к полету
4) интенсивным обменом веществ
Ответ: 2
12. Задание 20 При распознавании видов-двойников учитывается главным образом критерий
Задание 20 При распознавании видов-двойников учитывается главным образом критерий
1) генетический
2) географический
3) морфологический
4) физиологический
Ответ: 1
13. Задание 20 Общую территорию, которую занимает вид, называют
1) экологической нишей
2) биоценозом
3) ареалом
4) кормовой территорией
Ответ: 3
14. Задание 20 По какому морфологическому признаку можно отличить млекопитающих от других позвоночных животных
1) Пятипалая конечность
2) Волосяной покров
3) Наличие хвоста
4) Глаза, прикрытые веками
Ответ: 2
15. Задание 20 Вид — совокупность особей, обладающих
1) сходным набором хромосом
2) способностью образовывать пищевые связи между его особями
3) обитающих в разных экосистемах
4) созданных человеком на основе отбора
Ответ: 1
16. Задание 20 Почему нельзя определить вид только по генетическому критерию
Задание 20 Почему нельзя определить вид только по генетическому критерию
1) ареалы разных видов совпадают
2) набор хромосом у разных видов может совпадать
3) разные виды обитают в сходных условиях
4) особи разных видов сходны по строению
Ответ: 2
17. Задание 20 Физиологический критерий вида проявляется у всех его особей в сходстве
1) процессов жизнедеятельности
2) строения и формы хромосом
3) химического состава
4) внешнего и внутреннего строения
Ответ: 1
18. Задание 20 Ареал обитания вида является критерием
1) физиологическим
2) генетическим
3) географическим
4) морфологическим
Ответ: 3
19. Задание 20 Виды-двойники сходны по строению, поэтому их нельзя определить только по критерию
1) генетическому
2) географическому
3) морфологическому
4) физиологическому
Ответ: 3
20. Задание 20 Определенный набор хромосом у особей одного вида считают критерием
Задание 20 Определенный набор хромосом у особей одного вида считают критерием
1) биологическим
2) морфологическим
3) генетическим
4) цитологическим
Ответ: 3
21. Задание 20 В чем проявляется относительный характер морфологического критерия вида
1) ареалы разных видов совпадают
2) наборы хромосом у разных видов одинаковые
3) самцы и самки одного вида различаются внешне
4) разные виды обитают в сходных условиях
Ответ: 3
22. Задание 20 Большая синица живет в кронах деревьев, питается крупными насекомыми и их личинками — это описание критерия вида
1) географического
2) экологического
3) морфологического
4) генетического
Ответ: 2
23. Задание 20 Морфологический критерий вида — это
1) его область распространения
2) особенности процессов жизнедеятельности
3) особенности внешнего и внутреннего строения
4) определенный набор хромосом и генов
Ответ: 3
24. Задание 20 Видом называют группу особей
Задание 20 Видом называют группу особей
1) скрещивающихся и дающих плодовитое потомство
2) обитающих в одном природном сообществе
3) обитающих на общей территории
4) принадлежащих к одному сорту или породе
Ответ: 1
25. Задание 20 Генетический критерий вида характеризуется
1) сходством биохимического состава
2) внешними признаками
3) набором хромосом
4) сходством процессов жизнедеятельности
Ответ: 3
26. Задание 20 Пространственное размещение вида в природе — это критерий
1) физиологический
2) генетический —
3) географический
4) морфологический
Ответ: 3
27. Задание 20 Относительность географического критерия вида состоит в том, что
1) ареалы разных видов совпадают
2) наборы хромосом у разных видов одинаковые
3) самцы и самки одного вида различаются внешне
4) разные виды обитают в сходных условиях
Ответ: 1
28. Задание 20 Относительность экологического критерия вида состоит в том, что
Задание 20 Относительность экологического критерия вида состоит в том, что
1) ареалы разных видов совпадают
2) наборы хромосом у разных видов одинаковые
3) самцы и самки одного вида различаются внешне
4) разные виды обитают в сходных условиях
Ответ: 4
29. Задание 20 . Совокупность факторов внешней среды, в которой обитают особи вида, — критерий
1) экологический
2) географический
3) физиологический
4) морфологический
Ответ: 1
30. Задание 20 . Бурого медведя и очкового медведя ученые считают разными видами потому, что
1) они отличаются внешним видом
2) они живут на разных территориях
3) между ними существует репродуктивная изоляция
4) они питаются разной пищей
Ответ: 3
31. Задание 20 . К одному виду относятся
Задание 20 . К одному виду относятся
1) овчарка и сенбернар
2) стрептококк и плеврококк
3) хламидомонада и хлорелла
4) головня и ржавчина
Ответ: 1
32. Задание 20 . К разным видам относятся
1) овчарка и сенбернар
2) африканский и индийский слон
3) европеец и китаец
4) сиамский кот и сфинкс
Ответ: 2
33. Задание 20 . Прочитайте описание песца: «В отличие от лисицы, тело у песца приземистое, морда укороченная; уши закруглены, слабо выступают из зимней шерсти (это предохраняет их от обморожения)». Какой критерий вида используется в этом описании?
1) генетический
2) географический
3) морфологический
4) физиологический
Ответ: 3
34. Задание 20 . К какому критерию вида относится наличие у человека 46 хромосом?
1) физиологическому
2) генетическому
3) географическому
4) биохимическому
Ответ: 2
35. Задание 20 . В каком случае двух зайцев можно отнести к разным видам?
Задание 20 . В каком случае двух зайцев можно отнести к разным видам?
1) если у них разное число хромосом в клетках
2) если они живут на разных территориях
3) если одна группа менее плодовита, чем другая
4) если по численности одна группа больше другой
Ответ: 1
36. Задание 20 . Выберите признак, характеризующий виды-двойники.
1) могут скрещиваться между собой
2) не обладают морфологическим сходством
3) формируют на общей территории одну популяцию
4) репродуктивно изолированы друг от друга
Ответ: 4
37. Задание 20 . Какая характеристика иллюстрирует физиологический критерий вида пастушьей сумки?
1) распространение по полям, дорогам, сорным местам
2) обитание на Европейской части России
3) особенности обмена веществ в растении
4) поочерёдное расположение листьев на стебле
Ответ: 3
38. Задание 20 . Признаком вида служит
Задание 20 . Признаком вида служит
1) обитание его особей в одной экосистеме
2) получение плодовитого потомства
3) расселение особей в разные среды обитания
4) размножение его особей бесполым путём
Ответ: 2
39. Задание 20 . Виды — двойники малярийного комара отличаются друг от друга, главным образом, по критерию
1) экологическому
2) генетическому
3) географическому
4) морфологическому
Ответ: 2
40. Задание 20 . Какой из перечисленных признаков можно отнести к экологическму критерию вида обыкновенного ежа?
1) распространение в Европе
2) число хромосом в соматических клетках
3) наземный образ жизни
4) наличие игл на теле
Ответ: 3
41. Задание 20 . К какому критерию вида следует отнести совокупность факторов среды обитания лютика едкого?
1) географическому
2) генетическому
3) морфологическому
4) экологическому
Ответ: 4
42. Задание 20 . Какое из высказываний иллюстрирует морфологический критерий вида?
Задание 20 . Какое из высказываний иллюстрирует морфологический критерий вида?
1) эти два животных способны свободно скрещиваться между сообой
2) оба животных питаются одними и теми же растениями
3) оба животных проживают на одной территории
4) оба животных внешне похожи и имеют сходное строение
Ответ: 4
43. Задание 20 . Какое из высказываний иллюстрирует экологический критерий вида?
1) у обоих животных отсутствует один и тот же фермент
2) у этих животных одинаковый набор тканей и органов
3) оба животных питаются одними и теми же растениями
4) оба животных проживают на одной территории
Ответ: 3
44. Задание 20 . Среда обитания животных — это критерий вида
1) генетический
2) морфологический
3) экологический
4) биохимический
Ответ: 3
45. Задание 20 . Заяц-беляк и заяц-русак принадлежат
Задание 20 . Заяц-беляк и заяц-русак принадлежат
1) к одному виду
2) к одной популяции
3) к разным отрядам
4) к разным видам
Ответ: 4
46. Задание 20 . В каком случае двух зайцев можно отнести к разным видам?
1) если у них разное число хромосом в клетках
2) если они живут на разных территориях
3) если одна группа менее плодовита, чем другая
4) если по численности одна группа больше другой
Ответ: 1
47. Задание 20 . В каком из приведённых примеров дана характеристика морфологического критерия вида птицы большой синицы?
1) питается мелкими насекомыми
2) гнездится на деревьях
3) длина хвоста синицы не превышает длины её тела
4) предпочитает мелколиственные леса
Ответ: 3
48. Задание 20 . Древесный образ жизни обыкновенной квакши соответствует критерию вида
Древесный образ жизни обыкновенной квакши соответствует критерию вида
1) экологическому
2) географическому
3) морфологическому
4) генетическому
Ответ: 1
49. Задание 20 . Распространение большого пёстрого дятла в пределах ареала соответствует критерию вида
1) морфологическому
2) физиологическому
3) экологическому
4) географическому
Ответ: 4
50. Задание 20 . Способность белены чёрной синтезировать и накапливать алкалоиды — показатель критерия вида
1) морфологического
2) генетического
3) биохимического
4) географического
Ответ: 3
51. Задание 20 . Характерные для вида признаки и свойства называют
1) альтернативными признаками
2) модификациями
3) критериями
4) аллелями
Ответ: 3
52. Задание 20 . Ареал, занимаемый видом в природе, — это критерий
Задание 20 . Ареал, занимаемый видом в природе, — это критерий
1) экологический
2) географический
3) физиологический
4) морфологический
Ответ: 2
53. Задание 20 . О возможной принадлежности двух внешне похожих насекомоядных птиц к разным видам надежнее можно судить по
1) разным срокам гнездования
2) количеству яиц в кладке
3) возрасту
4) характеру пищи
Ответ: 1
1. Задание 20 Особи, относящиеся к одному виду,
1) имеют большие различия между собой, чем особи одного рода
2) обладают меньшей степенью родства, чем особи одного класса
3) занимают различные ярусы в природном сообществе
4) обладают наибольшей степенью родства
Ответ: 4
2. Задание 20 Половая структура популяции определяется количеством
1) женских и мужских особей
2) новорожденных особей
3) различных возрастных групп
4) старых особей
Ответ: 1
3. Задание 20 Возрастная структура популяции характеризуется
Задание 20 Возрастная структура популяции характеризуется
1) соотношением женских и мужских особей
2) численностью особей
3) соотношением молодых и половозрелых особей
4) её плотностью
Ответ: 3
4. Задание 20 Популяция клеста-еловика более устойчива, если
1) в ней преобладают мужские особи
2) в лесу нарушена ярусность
3) особи в ней генотипически различны
4) особи в ней генотипически однородны
Ответ: 3
5. Задание 20 Расширению ареала вида способствует
1) наличие в нём большого числа популяций
2) генетическое родство особей
3) отсутствие генетического родства особей
4) наличие в нем небольшого числа популяций
Ответ: 1
6. Задание 20 Элементарная структура, на уровне которой проявляется в природе действие естественного отбора
1) организм
2) биоценоз
3) вид
4) популяция
Ответ: 4
7. Задание 20 Сокращение в природе ареала вида способствует
Задание 20 Сокращение в природе ареала вида способствует
1) близкородственному скрещиванию
2) упрощению строения
3) усложнению строения
4) возникновению гибридов
Ответ: 1
8. Задание 20 Саморегуляция численности популяций обеспечивается
1) возникновением изоляции
2) модификационной изменчивостью
3) наследственной изменчивостью
4) действием ограничивающих факторов
Ответ: 4
9. Задание 20 Увеличению численности популяции жертв способствует
1) сокращение численности хищников
2) увеличение численности паразитов
3) увеличение численности конкурентов
4) уменьшение численности симбионтов
Ответ: 1
10. Задание 20 Популяция является структурной единицей
1) отряда
2) семейства
3) рода
4) вида
Ответ: 4
11. Задание 20 Группа наиболее сходных особей вида, относительно обособленных от других групп этого вида, длительно проживающая на определенной территории, представляет собой
Задание 20 Группа наиболее сходных особей вида, относительно обособленных от других групп этого вида, длительно проживающая на определенной территории, представляет собой
1) стадо
2) популяцию
3) подвид
4) род
Ответ: 2
12. Задание 20 Популяцию считают элементарной единицей эволюции, так как
1) она обладает целостным генофондом, способным изменяться
2) особи популяций имеют сходный обмен веществ
3) особи популяции отличаются размерами
4) она не способна изменяться во времени
Ответ: 1
13. Задание 20 Особи объединяются в одну популяцию на основе
1) их роли в биогеоценозе
2) равного соотношения полов
3) общности питания
4) свободного скрещивания
Ответ: 4
14. Задание 20 При длительном сохранении относительно постоянных условий среды в популяциях вида
Задание 20 При длительном сохранении относительно постоянных условий среды в популяциях вида
1) возрастает число спонтанных мутаций
2) проявляется стабилизирующий отбор
3) проявляется движущий отбор
4) усиливаются процессы дивергенции
Ответ: 2
15. Задание 20 Неограниченный рост численности популяции сдерживается
1) расширением ареала
2) подготовкой особей к зиме
3) действием факторов внешней среды
4) связями между особями разных поколений
Ответ: 3
16. Задание 20 Приспособленность вида к жизни в разных условиях в пределах своего ареала обеспечивает его существование в форме
1) популяций
2) отдельных особей
3) колоний
4) сообществ
Ответ: 1
17. Задание 20 Генофонд популяции — это совокупность всех составляющих популяцию
Задание 20 Генофонд популяции — это совокупность всех составляющих популяцию
1) особей
2) модификаций
3) генов
4) фенотипов
Ответ: 3
18. Задание 20 . В двух не сообщающихся между собой лесах Подмосковья живут кабаны, лоси, обыкновенные лисицы, зайцы-русаки и волки. Сколько популяций названных животных живёт в двух лесах?
1) 4
2) 5
3) 8
4) 10
Ответ: 4
19. Задание 20 . Пространственная структура вида в природе представлена
1) особями разного пола
2) сезонными группами
3) популяциями
4) сообществами
Ответ: 3
20. Задание 20 . Элементарной единицей эволюции считают
1) организм
2) семейство
3) популяцию
4) вид
Ответ: 3
21. Задание 20 . Основной причиной для выделения группы особей в популяцию является
Задание 20 . Основной причиной для выделения группы особей в популяцию является
1) нескрещиваемость членов группы между собой
2) внешнее отличие особей группы друг от друга
3) внутренние отличия особей групп друг от друга
4) ареал обитания особей группы
Ответ: 4
22. Задание 20 . Основным критерием возникновения нового вида является
1) появление внешних различий у особей популяции
2) экологическая изоляция популяций
3) географическая изоляция популяций
4) возникновение репродуктивной изоляции между популяциями
Ответ: 4
23. Задание 20 . Разделение популяций одного вида по срокам размножения может привести к
1) популяционным волнам
2) конвергенции признаков
3) усилению межвидовой борьбы
4) экологическому видообразованию
Ответ: 4
24. Задание 20 . Группа особей, способных скрещиваться и давать плодовитое потомство, − это
Задание 20 . Группа особей, способных скрещиваться и давать плодовитое потомство, − это
1) вид
2) пищевая цепь
3) экосистема
4) экологическая ниша
Ответ: 1
25. Задание 20 . Группа особей, способных свободно скрещиваться и проживающих на одной территории, − это
1) вид
2) популяция
3) экосистема
4) экологическая ниша
Ответ: 2
26. Задание 20 . Закон Харди – Вайнберга описывает
1) эволюционирующую популяцию
2) идеальную популяцию
3) малочисленную популяцию
4) популяцию неограниченной численности
Ответ: 2
27. Задание 20 . Закон Мюллера – Геккеля утверждает
1) единство органического мира
2) сходство ранних стадий эмбрионального развития хордовых
3) сходство мутаций, возникающих у близких родов растений
4) общность строения всех организмов
Ответ: 2
28. Задание 20 . Материал для действия естественного отбора поставляет
Задание 20 . Материал для действия естественного отбора поставляет
1) наследственная изменчивость
2) борьба за существование
3) ненаследственная изменчивость
4) изоляция организмов
Ответ: 1
29. Задание 20 . Стабилизирующий отбор способствует
1) расширению ареала обитания вида
2) колебанию численности особей в популяции
3) сохранению адаптивных признаков в популяции
4) свободному скрещиванию особей вида
Ответ: 3
30. Задание 20 . К результатам эволюции относят
1) генную мутацию
2) многообразие видов
3) изоляцию
4) искусственный отбор
Ответ: 2
31. Задание 20 . Какой из перечисленных факторов эволюции усиливает действие остальных?
1) мутационная изменчивость
2) комбинативная изменчивость
3) популяционные волны
4) изоляция
Ответ: 4
32. Задание 20 . Что является единицей эволюции в соответствии с синтетической теорией эволюции?
Задание 20 . Что является единицей эволюции в соответствии с синтетической теорией эволюции?
1) вид
2) особь
3) популяция
4) род
Ответ: 3
33. Задание 20 . Основным критерием возникновения нового вида является
1) появление морфологических различий между особями
2) разрыв ареала
3) репродуктивная изоляция популяций
4) экологическая изоляция
Ответ: 3
34. Задание 20 . Биологическими предпосылками микроэволюции в популяции являе(ю)тся
1) мутационный процесс и естественный отбор
2) модификационная изменчивость
3) избыток или недостаток корма
4) поведенческие различия
Ответ: 1
35. Задание 20 . Разрыв ареала вида приводит к
1) экологическому видообразованию
2) дрейфу генов
3) географической изоляции популяций
4) повышенной мутационной изменчивости
Ответ: 3
36. Задание 20 . Дивергенция в популяциях ведёт к
Задание 20 . Дивергенция в популяциях ведёт к
1) появлению мутаций
2) появлению разновидностей и видов
3) гибели популяций
4) модификационным изменениям
Ответ: 2
1. Задание 21 Сокращение численности и ареала уссурийского тигра в современную эпоху — пример
1) биологического прогресса
2) биологического регресса
3) идиоадаптации
4) ароморфоза
Ответ: 2
2. Задание 21 Возрастание численности серой вороны в населённых пунктах — пример
1) ароморфоза
2) дегенерации
3) биологического регресса
4) биологического прогресса
Ответ: 4
3. Задание 21 Увеличение численности насекомых-вредителей сельскохозяйственных растений, — пример
1) ароморфоза
2) дегенерации
3) биологического регресса
4) биологического прогресса
Ответ: 4
4. Задание 21 Показатель биологического прогресса вида
Задание 21 Показатель биологического прогресса вида
1) симбиоз
2) взаимопомощь
3) конкуренция
4) высокая численность
Ответ: 4
5. Задание 21 Главная причина биологического регресса многих видов в настоящее время
1) изменение климата
2) хозяйственная деятельность человека
3) изменение рельефа
4) увеличение численности хищников
Ответ: 2
Питание какой критерий вида. Какими бывают критерии вида? Морфологический критерий вида
Вид является одной из основных форм организации жизни на Земле (наряду с клеткой, организмом и экосистемой) и основной единицей классификации биологического разнообразия. Но в то же время термин «вид» до сих пор остается одним из наиболее сложных и неоднозначных биологических понятий.
Проблемы, связанные с понятием биологического вида легче понять, рассматривая их в историческом аспекте.
История вопроса
Термин «вид» употреблялся для обозначения имен биологических объектов с древнейших времен. Изначально он не был чисто биологическим: виды уток (кряква, шилохвость, чирок) не имели принципиальных отличий от видов кухонной утвари (сковородка, кастрюля и т.д.).
Биологический смысл термину «вид» придал шведский натуралист Карл Линней. Он употреблял это понятие для обозначения важного свойства биологического разнообразия – его дискретности (прерывистости; от латинского discretio – разделять). К. Линней рассматривал виды как объективно существующие группы живых организмов, достаточно легко отличимые друг от друга. Он считал их неизменными, раз и навсегда созданными богом.
Выделение видов в то время происходило на основе различий между особями по ограниченному числу внешних признаков. Этот метод получил название типологического подхода. Отнесение особи к тому или иному виду осуществлялось на основе сличения ее признаков с описаниями уже известных видов. Если ее признаки не удавалось соотнести ни с одним из существующих видовых диагнозов, то по данному экземпляру (он получал название типового) описывался новый вид. Иногда это приводило к казусным ситуациям: самцы и самки одшого вида описывались как разные виды.
Если ее признаки не удавалось соотнести ни с одним из существующих видовых диагнозов, то по данному экземпляру (он получал название типового) описывался новый вид. Иногда это приводило к казусным ситуациям: самцы и самки одшого вида описывались как разные виды.
С развитием эволюционных идей в биологии возникла дилемма: либо виды без эволюции, либо эволюция без видов. Авторы эволюционных теорий – Жан-Батист Ламарк и Чарльз Дарвин отрицали реальность видов. Ч.Дарвин, автор «Происхождения видов путем естественного отбора…», считал их «искусственными понятиями, придуманными ради удобства».
К концу XIX века, когда разнообразие птиц и млекопитающих было достаточно полно изучено на значительной территории Земли, стали очевидны недостатки типологического подхода: выяснилось, что животные из разных мест порой хоть и незначительно, но достаточно надежно отличаются друг от друга. В соответствии с установленными правилами им надо было присваивать статус самостоятельных видов. Число новых видов росло лавинообразно. Наряду с этим крепло сомнение: стоит ли различным популяциям близкородственных животных присваивать видовой статус только на том основании, что они слегка друг от друга отличаются?
Наряду с этим крепло сомнение: стоит ли различным популяциям близкородственных животных присваивать видовой статус только на том основании, что они слегка друг от друга отличаются?
В XX веке с развитием генетики и синтетической теории вид стали рассматривать как группу популяций с общим уникальным генофондом, обладающую собственной «системой защиты» целостности своего генофонда. Таким образом, типологический подход к выделению видов сменился эволюционным: виды определяются не различием, а обособленностью. Популяциям вида, морфологически отличным друг от друга, но способных свободно скрещиваться друг с другом, придается статус подвидов. Эта система взглядов легла в основу биологической концепции вида, получившей мировое признание благодаря заслуге Эрнста Майра. Смена концепций вида «примирила» представления о морфологической обособленности и эволюционной изменяемости видов и позволила с большей объективностью подойти к задаче описания биологического разнообразия.
Вид и его реальность. Ч. Дарвин в своей книге «Происхождение видов» и в других работах исходил из факта изменяемости видов, превращения одного вида в другой. Отсюда его трактовка вида как устойчивого и одновременно изменяющегося с течением времени, приводящего сначала к появлению разновидностей, которые он назвал «зарождающимися видами».
Ч. Дарвин в своей книге «Происхождение видов» и в других работах исходил из факта изменяемости видов, превращения одного вида в другой. Отсюда его трактовка вида как устойчивого и одновременно изменяющегося с течением времени, приводящего сначала к появлению разновидностей, которые он назвал «зарождающимися видами».
Вид – совокупность географически и экологически близких популяций, способных в природных условиях скрещиваться между собой, обладающих общими морфофизиологи-ческими признаками, биологически изолированных от популяций других видов.
Критерии вида – совокупность определенных признаков, свойственных только одному какому-либо виду (Т.А. Козлова, В.С. Кучменко. Биология в таблицах. М.,2000)
|
Критерии вида |
Показатели каждого критерия |
|
Морфологический |
Сходство внешнего и внутреннего строения особей одного вида; характеристика особенностей строения представителей одного вида |
|
Физиологический |
Сходство всех процессов жизнедеятельности, и прежде всего размножения. |
|
Биохимический |
Видовая специфичность белков и нуклеиновых кислот |
|
Генетический |
Каждый вид характеризуется определенным, присущим только ему набором хромосом, их структурой и дифференцированной окраской |
|
Эколого-географический |
Ареал обитания и непосредственная среда обитания – экологическая ниша. Каждый вид имеет свою собственную нишу обитания и ареал распространения |
 Представители разных видов, как правило, не скрещиваются или их потомство бесплодно
Представители разных видов, как правило, не скрещиваются или их потомство бесплодноСущественно и то, что вид представляет собой универсальную дискретную (дробимую) единицу организации жизни. Вид – это качественная ступень живой природы, он существует как результат внутривидовых взаимоотношений, которые обеспечивают его жизнь, воспроизведение и эволюцию.
Главная черта вида- относительная стабильность его генофонда, поддерживаемая репродуктивной изоляцией особей от других таких же видов. Единство вида поддерживается свободным скрещиванием между особями, в результате которого осуществляется постоянный поток генов во внутривидовом сообществе. Поэтому каждый вид на протяжении многих поколений устойчиво существует на той или иной местности, в этом проявляется его реальность. В то же время генетическая структура вида постоянно перестраивается под влиянием эволюционных факторов (мутации, рекомбинации, отбор), и поэтому вид оказывается неоднородным. Он распадается на популяции, расы, подвиды.
Единство вида поддерживается свободным скрещиванием между особями, в результате которого осуществляется постоянный поток генов во внутривидовом сообществе. Поэтому каждый вид на протяжении многих поколений устойчиво существует на той или иной местности, в этом проявляется его реальность. В то же время генетическая структура вида постоянно перестраивается под влиянием эволюционных факторов (мутации, рекомбинации, отбор), и поэтому вид оказывается неоднородным. Он распадается на популяции, расы, подвиды.
Генетическая обособленность видов достигается географической (родственные группы разделены морем, пустыней, горным хребтом) и экологической изоляцией (несовпадение сроков и мест размножения, обитание животных в различных ярусах биоценоза). В тех случаях, когда межвидовое скрещивание все же происходит, гибриды либо ослаблены, либо стерильны (например, гибрид осла и лошади – мул), что указывает на качественную обособленность вида и его реальность. По определению К. А. Тимирязева, «вида как категории строго определенной, всегда себе равной и неизменной, в природе не существует. Но вместе с этим мы должны признать, что виды, в наблюдаемый нами момент, имеют реальное существование».
Но вместе с этим мы должны признать, что виды, в наблюдаемый нами момент, имеют реальное существование».
Популяция. В пределах ареала какого-либо вида его особи распространены неравномерно, так как в природе нет идентичных условий для существования и размножения. Например, колонии кротов встречаются лишь в отдельных луговинах, заросли крапивы – по оврагам и канавам, лягушки одного озера бывают отделены от другого соседнего озера и т. д. Население вида распадается на естественные группировки – популяции. Однако эти разграничения не устраняют возможности скрещивания между особями, занимающими пограничные участки. Плотность населения популяции подвержена значительным колебаниям в разные годы и различные сезоны года. Популяция – это форма существования вида в конкретных условиях среды и единица его эволюции.
Популяция представляет собой совокупность свободно скрещивающихся особей одного вида, длительно существующих на определенной части ареала внутри вида и относительно обособленных от других популяций. Особи одной популяции обладают наибольшим, сходством по всем признакам, присущим виду, вследствие того, что возможность скрещивания внутри популяции выше, чем между особями соседних популяций и они испытывают одинаковое давление отбора. Несмотря на это популяции генетически неоднородны вследствие непрерывно возникающей наследственной изменчивости.
Особи одной популяции обладают наибольшим, сходством по всем признакам, присущим виду, вследствие того, что возможность скрещивания внутри популяции выше, чем между особями соседних популяций и они испытывают одинаковое давление отбора. Несмотря на это популяции генетически неоднородны вследствие непрерывно возникающей наследственной изменчивости.
Дарвиновская дивергенция (расхождение признаков и свойств потомков по отношению к исходным формам) может происходить только путем дивергенции популяций. Впервые это положение в 1926 г. обосновал С. С. Четвериков, показав, что за кажущейся внешней однородностью любой вид обладает огромным скрытым запасом генетической изменчивости в форме множества разнообразных рецессивных генов. Этот генетический резерв неодинаков в разных популяциях. Именно поэтому популяция является элементарной единицей вида и элементарной эволюционной единицей.
Типы видов
Выделение видов происходит на основании двух принципов (критериев). Это морфологический критерий (выявляющий различия между видами) и критерий репродуктивной изоляции (оценивающий степень их генетической обособленности). Процедура описания новых видов зачастую сопряжена с определенными сложностями, связанных как с неоднозначным соответствием критериев вида друг другу, так и с постепенностью и незавершенностью процесса видообразования. В зависимости от того, какого рода сложности возникали при выделении видов и каким образом они были решены, выделяют так называемые «типы видов».
Процедура описания новых видов зачастую сопряжена с определенными сложностями, связанных как с неоднозначным соответствием критериев вида друг другу, так и с постепенностью и незавершенностью процесса видообразования. В зависимости от того, какого рода сложности возникали при выделении видов и каким образом они были решены, выделяют так называемые «типы видов».
Монотипический вид. Часто никаких сложностей при описании новых видов не возникает. Такие виды обычно обладают обширным неразорванным ареалом, на протяжении которого географическая изменчивость выражена слабо.
Политипический вид.
Нередко с помощью морфологического критерия выделяется целая группа близкородственных форм, обитающих, как правило, в сильно расчлененной местности (в горах, либо на островах). Каждая из этих форм имеет свой, обычно довольно ограниченный ареал. Если между сравниваемыми формами существует географический контакт, то возможно применение критерия репродуктивной изоляции: если гибриды не возникают, либо относительно редки, этим формам придается статус самостоятельных видов; в противном случае описывают разные подвиды одного вида. Вид, включающий в себя несколько подвидов называется политипическим. Когда же анализируемые формы географически изолированы, оценка их статуса довольно субъективна и происходит только на основании морфологического критерия: если различия между ними «существенны», то перед нами – разные виды, если нет – подвиды. Не всегда в группе близкородственных форм удается однозначно определить статус каждой формы. Иногда группа популяций замыкается в кольцо, охватывающее горный массив или земной шар. В таком случае может оказаться, что «хорошие» (обитающие совместно и не гибридизирующие) виды связаны друг с другом цепью подвидов.
Вид, включающий в себя несколько подвидов называется политипическим. Когда же анализируемые формы географически изолированы, оценка их статуса довольно субъективна и происходит только на основании морфологического критерия: если различия между ними «существенны», то перед нами – разные виды, если нет – подвиды. Не всегда в группе близкородственных форм удается однозначно определить статус каждой формы. Иногда группа популяций замыкается в кольцо, охватывающее горный массив или земной шар. В таком случае может оказаться, что «хорошие» (обитающие совместно и не гибридизирующие) виды связаны друг с другом цепью подвидов.
Полиморфный вид.
Иногда в пределах единой популяции вида существуют две или несколько морф – групп особей, резко отличных по окраске, но способных свободно скрещиваться друг с другом. Как правило, генетическая основа полиморфизма проста: различия между морфами обуславливается действием разных аллелей одного гена. Пути возникновения этого явления могут быть весьма различны.
|
Адаптивный полиморфизм богомола |
Гибридогенный полиморфизм испанской каменки |
|
У богомола существует зеленая и бурая морфы. Первая плохо заметна на зеленых частях растений, вторая – на ветках деревьев и сухой траве. В экспериментах по пересаживанию богомолов на фон не соответствующий их окраске удалось показать, что полиморфизм в данном случае мог возникнуть и поддерживается благодаря естественному отбору: зеленая и бурая окраска богомолов является защитой от хищников и позволяет этим насекомым меньше конкурировать друг с другом. |
У самцов испанской каменки существуют белогорлая и черногорлая морфы. Характер соотношения этих морф в разных частях ареала позволяет предположить, что черногорлая морфа образовалась в результате гибридизации с близкородственным видом – каменкой-плешанкой. |
Виды-двойники
– виды, обитающие совместно и не скрещивающиеся друг с другом, но очень слабо отличающиеся морфологически. Сложность различения таких видов связана со сложностью выделения или неудобстве использования их диагностических признаков – ведь сами виды-двойники прекрасно разбираются в собственной «таксономии». Чаще виды-двойники встречаются среди групп животных, использующих для поиска полового партнера запах (насекомые, грызуны) и реже – у тех, которые пользуются зрительной и акустической сигнализацией (птицы).
Сложность различения таких видов связана со сложностью выделения или неудобстве использования их диагностических признаков – ведь сами виды-двойники прекрасно разбираются в собственной «таксономии». Чаще виды-двойники встречаются среди групп животных, использующих для поиска полового партнера запах (насекомые, грызуны) и реже – у тех, которые пользуются зрительной и акустической сигнализацией (птицы).
Клесты еловик (Loxia curvirostra) и сосновик (Loxia pytyopsittacus). Эти два вида клестов являются одним из немногочисленных примеров видов-двойников среди птиц. Обитая совместно на значительной территории, охватывающую Северную Европу и Скандинавский полуостров, эти виды не скрещиваются друг с другом. Морфологические различия между ними, незначительные и весьма ненадежные, выражаются в размерах клюва: у сосновика он несколько толще, чем у еловика.
«Полувиды».
Видообразование – длительный процесс, и поэтому можно столкнуться с такими формами, статус которых нельзя оценить объективно. Они еще не являются самостоятельными видами, поскольку гибридизируют в природе, но это уже и не подвиды, так как морфологические различия между ними весьма существенны. Такие формы называются «пограничными случаями», «проблемными видами» или «полувидами». Формально им приписываются бинарные латинские названия, как у «нормальных» видов, и в таксономических списках они помещаются рядом друг с другом. «Полувиды» встречаются не так уж и редко, и мы сами зачастую не подозреваем, что окружающие нас виды являются типичными примерами «пограничных случаев». В Средней Азии домовый воробей обитает совместно с другим близкородственным видом – черногрудым воробьем, от которого хорошо отличается по окраске. Гибридизация между ними в этом районе отсутствует. Их систематический статус как самостоятельных видов не вызывал бы сомнений, если бы не существовала вторая зона контакта в Европе. Италию населяет особая форма воробьев, возникшая в результате гибридизации домового и испанского. При этом в Испании, где домовый и испанский воробьи также обитают совместно, гибриды встречаются редко.
Они еще не являются самостоятельными видами, поскольку гибридизируют в природе, но это уже и не подвиды, так как морфологические различия между ними весьма существенны. Такие формы называются «пограничными случаями», «проблемными видами» или «полувидами». Формально им приписываются бинарные латинские названия, как у «нормальных» видов, и в таксономических списках они помещаются рядом друг с другом. «Полувиды» встречаются не так уж и редко, и мы сами зачастую не подозреваем, что окружающие нас виды являются типичными примерами «пограничных случаев». В Средней Азии домовый воробей обитает совместно с другим близкородственным видом – черногрудым воробьем, от которого хорошо отличается по окраске. Гибридизация между ними в этом районе отсутствует. Их систематический статус как самостоятельных видов не вызывал бы сомнений, если бы не существовала вторая зона контакта в Европе. Италию населяет особая форма воробьев, возникшая в результате гибридизации домового и испанского. При этом в Испании, где домовый и испанский воробьи также обитают совместно, гибриды встречаются редко.
Цели: сформировать понятия «вид» и «критерии вида»; показать механизмы репродуктивной изоляции в природе; продолжить формирование умений давать морфологическое описание растений, работать с текстами, составлять таблицы, анализировать, формулировать выводы.
Оборудование: картины по зоологии: «Заяц- беляк», «Заяц-русак», «Медведь бурый», «Медведь белый»; разда- точный материал «Заяц-беляк и заяц-русак», «Ворон и ворона».
Ход урока
I . Организационный момент
Учитель объявляет тему и задачи урока.
11. Актуализация знаний
1. Выполнение заданий (устно).
Задание № 1
Назовите знакомые вам виды растений и животных, обитающих рядом с вашим домом или школой.
Задание № 2
Учитель демонстрирует картину «Бурый медведь» и задает вопросы:
Название этого вида животного — Медведь бурый. Какое из этих двух слов относится к родовому названию, какое — к видовому?
Назовите другой вид животного из этого же рода. (Это Медведь
белый).
(Это Медведь
белый).
Учитель вывешивает картину, иллюстрирующую белого медведя, рядом с картиной «Медведь бурый».
Сравните два вида одного рода. Покажите черты сходства и отличия.
Задание № 3 ,
В указанном списке животных подсчитайте количество особей, видов и родов.
1. Еж обыкновенный.
2. Лисица обыкновенная.
3. Медведь гималайский или белогрудый.
4. Хомячок джунгарский.
5. Заяц-беляк.
6. Медведь бурый.
7. Хомячок сирийский или золотистый.
8. Заяц-русак.
9. Еж ушастый.
10. Лисица обыкновенная.
(Ответ: количество особей — 10; видов — 9; родов — 5 (Еж, Лиси- ца, Медведь, Хомячок, Заяц).)
При выполнении последнего задания у ряда учащихся возникает проблема: отнести зайца-беляка и зайца-русака к одному виду или к двум разным видам. Правдиво или ошибочно мнение, что заяц- беляк — это заяц-русак зимой?
Доклад «Заяц русак и заяц беляк».
— Какой вывод можно сделать по результатам выполнения всех заданий по актуализации знаний?
Вывод:
1. Для обозначения вида используется двойная (бинарная) но менклатура, согласно которой сначала указывают род, к которому относится вид (существительное), а затем видовое название (прила гательное).
2. Особи разных видов отличаются друг от друга по местам оби тания, внешним признакам и др.
3. Сходные виды объединяются в один род.
4. Вид — это основная категория биологической классификации.
III . Изучение нового материала
1. Рассказ учителя.
— Что же такое вид и каковы его критерии?
Вопрос о виде и критериях вида занимает центральное место в теории эволюции и служит предметом многочисленных иссле дований в области систематики, зоологии, ботаники и других наук. И это вполне объяснимо: четкое понимание сущности вида необходимо для выяснения механизмов эволюционного процесса.
Строгое общепринятое определение вида до сих пор не разра
ботано. В биологическом энциклопедическом словаре мы на
ходим следующее определение вида:
В биологическом энциклопедическом словаре мы на
ходим следующее определение вида:
«Вид — совокупность популяций особей, способных к скрещива нию с образованием плодовитого потомства, населяющих определ енный ареал, обладающих рядом общих морфофизиологических признакови отдаленных от других таких же групп особей практи чески полным отсутствием гибридных форм».
Сравните это определение с тем, что дается в вашем учебнике (учебник А.А. Каменского, § 4.1, с. 134 ).
Поясним понятия, которые встречаются. в определении вида:
Ареал — область распространения данного вида или популяции в природе.
Популяция (от лат. «рор uius » — народ, население) — совокуп ность особей одного вида, обладающих общим генофондом и зани мающих определенную территорию — ареал.
Генофонд — совокупность генов, которые имеются у особей данной популяции.
Рассмотрим историю развития взглядов на вид в биологии.
Понятие вида впервые ввел в науку английский ботаник Джон
Рей в
XVII
веке
. Основополагающая работа по проблеме вида
была написана шведским натуралистом и естествоиспытателем
Карлом Линнеем в
XVIII
веке
, в которой он предложил первое
научное определение вида, уточнил его критерии.
Основополагающая работа по проблеме вида
была написана шведским натуралистом и естествоиспытателем
Карлом Линнеем в
XVIII
веке
, в которой он предложил первое
научное определение вида, уточнил его критерии.
Комментарии учителя. К. Линней считал, что вид — универ сальная, реально существующая единица живой материи, морфо логически однородная и неизменная . Все особи вида, по мнению ученого, имеют типичный морфологический облик, а вариации представляют собой случайные отклонения , результат несовершенного воплощения идеи вида (своего рода уродства ). Ученый считал, что виды неизменны, неизменна природа. Идея неизмен ной природы покоилась на концепции креационизма, согласно которой все сущее было сотворено Богом. Применительно к биологии эту концепцию Линней выразил в своей знаменитой фор муле «Видов столько, сколько разных форм вначале произвело Бесконечное существо».
Другая концепция принадлежит Тому Батисту Ламарку
— вели
кому французскому натуралисту. По его концепции виды реально не
существуют
, это чисто умозрительное понятие, изобретенное для
того, чтобы легче было рассматривать совокупно большее количество
особей, поскольку по словам Ламарка, «в природе нет ни
чего кроме особей». Индивидуальная изменчивость непрерывна,
поэтому границу между видами можно провести и здесь, и там —
где удобнее.
По его концепции виды реально не
существуют
, это чисто умозрительное понятие, изобретенное для
того, чтобы легче было рассматривать совокупно большее количество
особей, поскольку по словам Ламарка, «в природе нет ни
чего кроме особей». Индивидуальная изменчивость непрерывна,
поэтому границу между видами можно провести и здесь, и там —
где удобнее.
Третья концепция подготавливалась в первой четверти XIX века. Она была обоснована Чарльзом Дарвином и последующими биолога ми. По этой концепции виды имеют независимую реальность. Вид неоднороден, представляет собой систему соподчиненных единиц. С реди них основная элементарная единица — популяция. Виды, по Дарвину, изменяются, они относительно постоянны и являются рез ультатом эволюционного развития .
Таким образом, понятие «вид» имеет длительную историю становления в биологической науке.
Порой самые опытные биологи становятся в тупик, определяя,
принадлежат ли данные особи к одному виду или нет
. Почему так
происходит, существуют ли точные и строгие критерии, которые по
могли бы разрешить все сомнения?
Почему так
происходит, существуют ли точные и строгие критерии, которые по
могли бы разрешить все сомнения?
Критерии вида — это признаки, по которым один вид отлича ется от другого. Они же являются изолирующими механизмами, препятствующими скрещиванию, независимости, самостоятельно сти видов.
Нам известно, что одной из главных особенностей биологической материи на нашей планете является дискретность. Это выражается в том, что она представлена отдельными видами, не скрещивающимися друг с другом, изолированными один от дру гого.
Существование вида обеспечивается его генетическим единством (особи вида способны скрещиваться и давать жизнеспособное плодовитое потомство) и его генетической самостоятельностью (невоз можность скрещивания с особями другого вида, нежизнеспособно сть или стерильность гибридов).
Генетическая самостоятельность вида обусловливается совокуп
ностью его характерных признаков: морфологических, физиологических, биохимических, генетических, особенностей образа жизни, поведения, географического распространения и т. д. Это и есть крит
ериивида.
д. Это и есть крит
ериивида.
Познакомимся с ними.
2. Работа в группах
Каждая группа получает текст, иллюстрирующий один из критериев вида. Через 5 минут необходимо рассказать о сути данного критерияи о том,в чем недостаток данного критерия. По мере выступления групп классом заполняется таблица «Критерии вида».
Таблица № 1
Критерии вида.
|
Название критерия |
Признаки особей по критерию |
Исключение |
|
1. Морфологический |
Сходство внешнего и внутреннего строения организмов. |
Виды-двойники, половой диморфизм, полиморфизм. |
|
2. Физиологический |
Сходство всех процессов жизнедеятельности и возможность получения плодовитого потомства при скрещивании. |
У разных видов сходство процессов жизнедеятельности. |
|
3. Экологический |
Сходство по способам питания, местам обитания, наборам факторов внешней среды, необходимых для существования. |
Экологические ниши разных видов перекрываются. |
|
4. Географический |
Занимают определённый ареал. |
Космополиты. Совпадение ареалов разных видов. |
|
5. Биохимический |
Сходство по биохимическим параметрам – состав и структура белков, нуклеиновых кислот. |
Есть очень близкие по биохимическому составу виды. |
|
6. Этологический |
Сходство в поведении. Особенно в брачный период (ритуалы ухаживания, брачные песни и т. д.). |
Существуют виды с близким поведением. |
|
7. Цито-генетический а)Цитологический |
Особи одного вида скрещиваются между собой и дают плодовитое потомство (основан на сходстве числа хромосом, их формы и строения). |
Хромосомный полиморфизм в пределах вида; у многих разных видов число хромосом одинаково. |
|
б) Генетический |
Генетическая изоляция видов. Наличие постпопуляционных механизмов изоляции. Важнейшие из них — это гибель мужских гамет (генетическая несовместимость), гибель зигот, нежизнеспособность гибридов, их стерильность, наконец, невозможность найти полового партнера и дать жизнеспособное плодовитое потомство |
Собака и волк, тополь и ива, канарейка и зяблик дают плодовитое потомство. (Наличие межвидовых гибридов) |
|
8. Исторический |
Общность предков, единая история возникновения и развития вида. |
 Наличие межвидовых гибридов.
Наличие межвидовых гибридов.

Итак, видовые критерии, по которым мы отличаем один вид другого, в совокупности обусловливают генетическую изоляцию ви дов, обеспечивая самостоятельность каждого вида и разнообразие в природе. По сути, в выработке этих изолирующих видовых призна ков и заключается процесс формирования видов. Именно поэтому изучение видовых критериев имеет определяющее значение для по нимания механизмов процесса эволюции, происходящего на нашей планете.
3. Формулирование выводов.
После заполнения таблицы формулируются выводы:
1) Видовые критерии, по которым отличается один вид от друг го, в совокупности обусловливают генетическую изоляцию видов, обеспечивая самостоятельность каждого вида и разнообразие их в природе.
2) Нет ни одного видового критерия, который можно было признать абсолютным и универсальным.
3) К способам изоляции, препятствующим скрещиванию разных видов относятся:
а) различия ареалов, мест обитания => невозможность встречи;
б) разные сроки размножения;
в) различия в строении половых органов;
г) нежизнеспособность или стерильность гибридов;
д)
разные ритуалы «ухаживания» в период размножения.
4) Вид представляет собой генетически относительно изолиро ванную систему, что доказывает реальность существования видов в природе.
Вспомните, о чем говорилось в тексте «Заяц-беляк и заяц- русак». Какие критерии вида использованы при описании зай цев?
Ответить на вопрос:
— Какие критерии вида использованы при описанииживотных?
1). Лебедь-шипун часто изгибает шею в виде буквы S , а клюв и голову держит наклонно к воде. При раз дражении издает характерный шипящий звук, по которому и получил свое название. Распространен лебедь-шипун на изолированных участках в средней и южной полосе Европы и Азии от южной Швеции, Дании и Польши на западе до Монголии, Приморского края и Китая на Востоке. Всюду на этой территории редок, часто пара от пары гнездится на гро мадном расстоянии, а во многих районах вовсе отсутствует. Населяет заросшие водной растительностью лиманы, озера, иногда даже болота, предпочитая глухие, мало посещаемые человеком.
Малый, или тундровый лебедь распространен по тундре
Азии от Кольского полуострова на западе до дельты Колымы на Востоке,
заходя в область лесотундры и западные острова Северного Ледовитого. Для гнездования
выбирает заболоченные и низкие травянистые участки
разбросанными по ним озерами, а также речные долины, изобилующие
старицами и протоками.
Для гнездования
выбирает заболоченные и низкие травянистые участки
разбросанными по ним озерами, а также речные долины, изобилующие
старицами и протоками.
Брачные игры своеобразны и проходят на суше. При этомсамец ходит перед самкой, вытягивает шею, временами приподнимает крылья, издавая ими особый хлопающий звук и звонко кричит.
2). Городская ласточка. Верх го ловы, спина, крылья и хвост сине-черные, надхвостье и весь низ тела белые. Хвост с резкой треугольной вырезкой на конце. Обитатель горных и культурного ландшафтов. Гнездится на стенах скал и зданий. П ерелетная птица. Держится стаями в воздухе или сидя на проводах , чаще других ласточек садится на з емлю. Гнездится колониями . Гнездо лепит из комочков глины в форме полушара с боковым входом . Кладка из 4-6 белых яиц в мае – июне. Гол ос – звонкое «тиррч-тиррч»
Береговая ласточка. Верх головы, шеи, спина, крылья, хвост и полоса поперёк груди серовато-бурые, горло, грудь и брюшко белые. Хвост с неглубокой вырезкой.
Населяет долины рек, где гнездится по обрывистым глиняным или песчаным берегам. Обычная или многочисленная перелетная птица. Держится стаями, гнездится колониями. Гнезда устраивает в норах по обрывистым берегам рек. Кладка из 4-6 белых яиц в мае-июле. Голос — негромкое «чирр- чирр
Домашнее задание
По учебнику А.А. Каменского, § 4.1, вопросы после параграфа, термины.
Индивидуально:
1) Сообщение «Правда ли, что ворон — это муж вороны?»
2 ) Используя литературные источники, приведите конкретные пр имеры географического, экологического и этологического критериев.
Дополнительный материал для работы в группах.
Критерии вида
Морфологический критерий
Это был первый и долгое время единственный критерий, используемый для описания видов.
Морфологический критерий самый удобный и заметный, поэтому и сейчас широко используется в систематике растений и животных.
Мы можем легко отличить по размерам и окраске оперения большого
пестрого дятла от зеленого дятла, малого пестрого дятла и желны
(черного дятла), большую синицу от хохлатой, длиннохвостой, голубой
и гаички, клевер луговой от ползучего и люпиновидного и т. д.
д.
Несмотря на удобство, этот критерий не всегда «работает». Им не воспользуешься для разграничения видов-двойников, практически не отличающихся морфологически. Таких видов много среди малярийных комаров, дрозофил, сиговых рыб. Даже у птиц 5% видов-двойников, а одном ряду североамериканских сверчков их 17.
Использование одного только морфологического критерия может
привести к ошибочным выводам. Так, К. Линней по особенности
внешнего строения отнес самца и самку утки кряквы к разным видам. Сибирские охотники по окраске меха лисиц выделили пять вариаций: сиводушки, огневки, крестовки, черно-бурые и черные. В Англии у 70 видов бабочек, наряду с особями со светлой окраской, имеются и тем
ные морфы, численность которых в популяциях стала увеличиваться в
связи с загрязнением лесов. Полиморфизм — широко распространенное
явление. Он встречается у всех видов. Затрагивает он и те признаки, по которым различаются виды. У жуков дровосеков, например у усача цве
точного, встречающегося поздней весной на купальнице, помимо ти
пичной формы в популяциях встречается до 100 аберраций окраски. Во времена Линнея морфологический критерий был главным, так как счи
тали, что существует одна типичная для вида форма.
Во времена Линнея морфологический критерий был главным, так как счи
тали, что существует одна типичная для вида форма.
Сейчас, когда установлено, что у вида может быть много форм, типо логическая концепция вида отброшена и морфологический критерий не всегда удовлетворяет ученых. Все же следует признать, что этот критерий очень удобен для систематизации видов и в большинстве определителей животных и растений он играет главную роль.
Физиологический критерий
Физиологические особенности различных видов растений и живот
ных часто являются фактором, обеспечивающим их генетическую само
стоятельность. Например, у многих дрозофил сперма особей чужого ви
да вызывает иммунологическую реакцию в половых путях самки, что приводит к гибели сперматозоидов. Гибридизация различных видов и
подвидов козлов часто приводит к нарушению периодичности плодо
ношения — приплод появляется зимой, что ведет к его гибели. Скрещи
вание разных подвидов косуль, например сибирской и европейской,
подчас приводит к гибели самок и потомства из-за крупных размеров
плода.
Биохимический критерий
Интерес к этому критерию появился в последние десятилетия в связи с
развитием биохимических исследований. Он не находит широкого применения
, так как не существует каких-либо специфических веществ, характерных
только для одного вида и, кроме того, он весьма трудоемкий и далеко не универсальный. Однако им можно воспользоваться в тех случаях,
когда другие критерии «не работают». Например, для двух видов-двойников
бабочек из рода амата (А. р
h
е
g
еа и А. г
ugazzii
) диагностическим
и признаками являются два фермента — фосфоглюкомутаза и эстераза-5, позволяющие даже определять гибридов этих двух видов. В последнее время
широкое применение получило сравнительное изучение состава ДН
К в практической таксономии микробов. Изучение состава ДНК позволило
произвести ревизию филогенетической системы различных групп микроорганизмов. Разработанные методы дают возможность сравнивать состав
ДНК у законсервированных в толщах земли бактерий и ныне живущих
форм.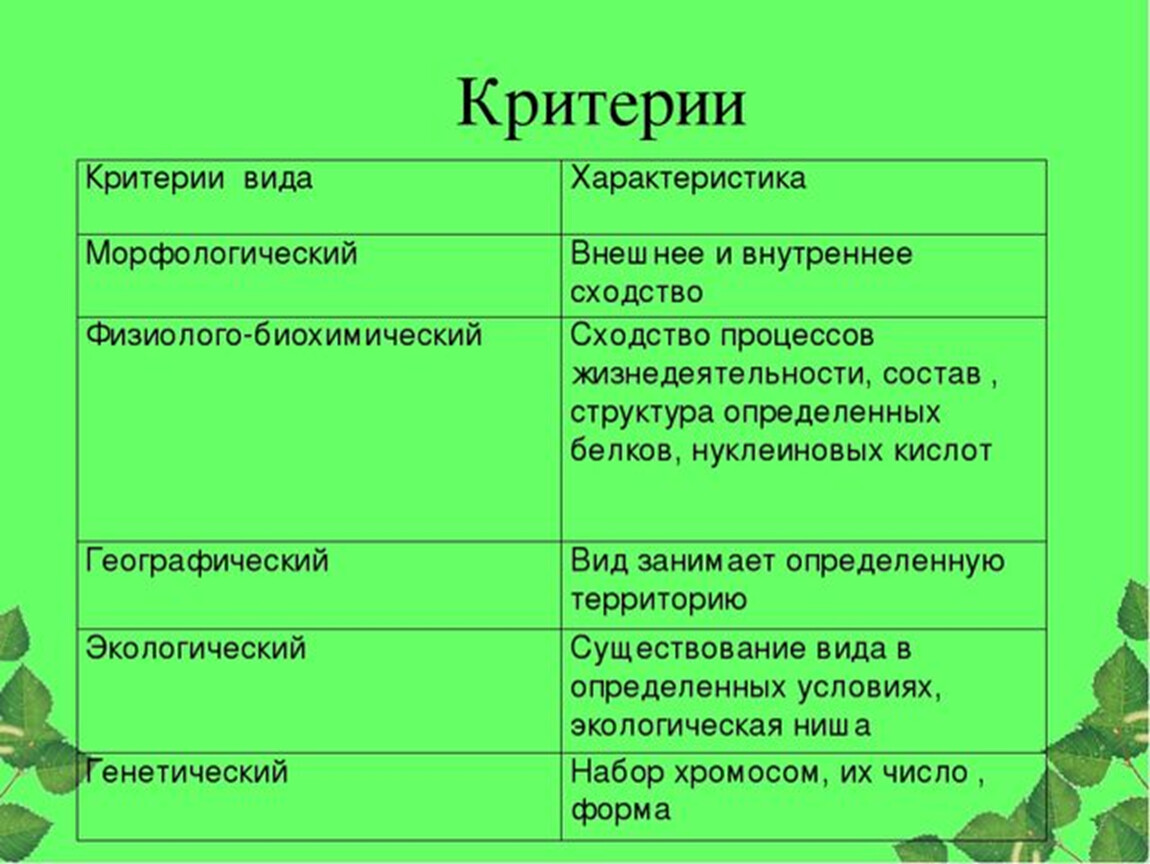 Было проведено, например, сравнение состава ДНК у пролежавшей
около 200 млн. лет в толще солей палеозойской бактерии псевдо
монады солелюбивой и у ныне живущих псевдомонад. Состав их ДНК оказалдся
идентичным, а биохимические свойства — сходными.
Было проведено, например, сравнение состава ДНК у пролежавшей
около 200 млн. лет в толще солей палеозойской бактерии псевдо
монады солелюбивой и у ныне живущих псевдомонад. Состав их ДНК оказалдся
идентичным, а биохимические свойства — сходными.
Цитологический критерий
Развитие цитологических методов позволило ученым исследовать фо рму и число хромосом у многих видов животных и растений. Появилось новое направление — кариосистематика, которое внесло некоторые поправки и уточнения в построенную на основе морфологических критериев филогенетическую систему. В ряде случаев число хромосом служит характерной особенностью вида. Кариологический анализ позволил , например, упорядочить систематику диких горных баранов, у кото рых разными исследователями выделялось от 1 до 17 видов. Анализ показал наличие трех кариотипов: 54-хромосомный — у муфлонов, 56-х ромосомный — у архаров и аргали и 58-хромосомный — у обитателей гор Средней Азии — уриалов.
Однако и этот критерий не является универсальным. Во-первых, у
многих разных видов число хромосом одинаково и форма их сходна. Во-вторых, в пределах одного и того же вида могут встречаться особи с разным числом хромосом. Это так называемые хромосомный и геномный
полиморфизм. Например, козья ива имеет диплоидное — 38 и тетраплоидное число хромосом — 76. У серебряного карася встречаются популяции с набо
ром хромосом 100, 150, 200, тогда как нормальное число их равно 50. У
радужной форели число хромосом варьирует от 58 до 64, у беломорской сель
ди встречаются особи с 52 и 54 хромосомами. В Таджикистане на участке
протяженностью всего 150 км, зоологами была обнаружена популяция слепушонки обыкновенной с набором хромосом от 31 до 54. У песчанок из разных мест обитания число хромосом различное: 40 — у песчанок алжир
ских популяций, 52 — у израильских и 66 — у египетских. К настоящему времени внутривидовой хромосомный полиморфизм обнаружен у 5 % ц
ито-генетически изученных видов млекопитающих.
Во-первых, у
многих разных видов число хромосом одинаково и форма их сходна. Во-вторых, в пределах одного и того же вида могут встречаться особи с разным числом хромосом. Это так называемые хромосомный и геномный
полиморфизм. Например, козья ива имеет диплоидное — 38 и тетраплоидное число хромосом — 76. У серебряного карася встречаются популяции с набо
ром хромосом 100, 150, 200, тогда как нормальное число их равно 50. У
радужной форели число хромосом варьирует от 58 до 64, у беломорской сель
ди встречаются особи с 52 и 54 хромосомами. В Таджикистане на участке
протяженностью всего 150 км, зоологами была обнаружена популяция слепушонки обыкновенной с набором хромосом от 31 до 54. У песчанок из разных мест обитания число хромосом различное: 40 — у песчанок алжир
ских популяций, 52 — у израильских и 66 — у египетских. К настоящему времени внутривидовой хромосомный полиморфизм обнаружен у 5 % ц
ито-генетически изученных видов млекопитающих.
Иногда этот критерий неверно трактуется как генетический. Безусловно
, число и форма хромосом важный признак, препятствующий скрещи
ванию особей разных видов. Однако это скорее цитоморфологичес
кий критерий, так как речь идет о внутриклеточной морфологии: числе
и форме хромосом, а не о наборе и структуре генов.
Безусловно
, число и форма хромосом важный признак, препятствующий скрещи
ванию особей разных видов. Однако это скорее цитоморфологичес
кий критерий, так как речь идет о внутриклеточной морфологии: числе
и форме хромосом, а не о наборе и структуре генов.
Э тологический критерий
Для некоторых видов животных механизмом, препятствующим с крещиванию и нивелированию различий между ними, являются осо бенности их поведения, особенно в брачный период. Узнавание партнера своего вида и отвергание попыток ухаживания самцов другого вида основаны на специфических раздражителях — зрительных, звуковых химических, тактильных, механических и др.
В широко распространенном роде пеночки разные виды очень похо
жи друг на друга морфологически, в природе их не различить ни по окраске, ни по размерам. Зато все они очень хорошо различаются по песне и по повадкам. Песня пеночки веснички сложная, похожа на песнь зяблика
, только без его завершающего колена, а песня теньковки — про
стенькие монотонные посвисты. Многочисленные виды-двойники аме
риканских светлячков из рода Р
hotinus
были впервые опознаны лишь по
различиям в их световых сигналах. Самцы светлячков в полёте производят вспышки света, частота, продолжительность и чередуемость которых
специфичны для каждого вида
. Хорошо известно, что ряд видов прямокрылых и равнокрылых, обитающих в пределах,
одного биотопа и размножающихся синхронно, различаются лишь
характеру их призывных сигналов. Такие виды-двойники с акустической
репродуктивной изоляцией обнаружены, например, у сверчков, кобылок-коньков, цикад и других насекомых. Два близких вида американских
жабне скрещиваются также из-за различий в зове самцов.
Многочисленные виды-двойники аме
риканских светлячков из рода Р
hotinus
были впервые опознаны лишь по
различиям в их световых сигналах. Самцы светлячков в полёте производят вспышки света, частота, продолжительность и чередуемость которых
специфичны для каждого вида
. Хорошо известно, что ряд видов прямокрылых и равнокрылых, обитающих в пределах,
одного биотопа и размножающихся синхронно, различаются лишь
характеру их призывных сигналов. Такие виды-двойники с акустической
репродуктивной изоляцией обнаружены, например, у сверчков, кобылок-коньков, цикад и других насекомых. Два близких вида американских
жабне скрещиваются также из-за различий в зове самцов.
Решающую роль в репродуктивной изоляции часто играют различия в демонстративном поведении. Например, близкие виды мух дрозофил от
личаются спецификой ритуала ухаживания (по характерувибрации
крыльев, дрожания лапок, кружения, тактильных контактов). Два близких
вида — серебристая чайка и клуша имеют различия в степени выраженно
сти демонстративных поз, а семь видов ящериц рода
S
се1ороги
s
различаются степенью поднятия головы при ухаживании половых партнеров.
Экологический критерий
Особенности поведения подчас тесно связаны с экологической спецификой вида, например, с особенностями устройства гнезда. Три вида наших обычных синиц гнездятся в дуплах лиственных деревьев, преимущественно берез. Большая синица на Урале выбирает обычно глубокое дупло в нижней части ствола березы или ольхи, образовавшееся в ре зультате выгнивания сучка и смежной с ним древесины. Это дупло недоступно ни дятлам, ни воронам, ни хищным млекопитающим. Синица московка заселяет морозобойные трещины в стволах березы и ольхи. Га ичка же предпочитает строить дупло сама, выщипывая полости в трухля вых или старых стволах березы и ольхи, и без этой трудоемкой процедуры она не отложит яиц.
Особенности образа жизни, присущие каждому виду, определяют
его положение, его роль в биогеоценозе, то есть его экологическую
нишу. Даже самые близкие виды, как правило, занимают разные экониши, то есть различаются хотя бы по одному-двум экологическим
признакам.
Так, экониши всех наших видов дятлов различаются по характеру питания. Большой пестрый дятел зимой питается семенами лиственницы и сосны, раздалбливая шишки в своих «кузницах». Черный дятел желна добывает личинок усачей и златок из-под коры и из древесины елей, а малый пестрый дятел долбит мягкую древесину ольхи либо добывает насекомых из стеблей травянистых растений.
Каждый из 14 видов дарвиновых вьюрков (названных так в честь Ч. Дарвина, впервые обратившего на них внимание), обитающих на Галапагосских островах, имеет свою специфическую эконишу, отличающуюся от других прежде всего характером корма и путями его добывания.
Ни экологический, ни рассмотренный выше этологический крите
рии не являются универсальными. Очень часто особи одного вида, но раз
ных популяций отличаются целым рядом особенностей образа жизни
и поведения. И напротив, разные виды, даже очень далекие, в системат
ическом отношении, могут иметь сходные этологические признаки
или играть одинаковую роль в сообществе (например, роли травоядных млекопитающих и насекомых, скажем, таких, как саранча, вполне сопоставимы).
Географический критерий
Этот критерий наряду с экологическим в большинстве определителей занимает второе (после морфологического) место. При определении многих видов растений, насекомых, птиц, млекопитающих и других групп организмов, распространение которых хорошо изучено, зна ние ареала играет существенную роль. У подвидов ареалы, как правило не совпадают, что обеспечивает их репродуктивную изоляцию и, по сути , их существование в качестве самостоятельных подвидов. Многие виды занимают разные ареалы (такие виды называют аллопатрическими). Но огромное число видов имеют совпадающие или перекрываю щиеся ареалы (симпатрические виды). Кроме того, существуют виды, не имеющие четких границ распространения, а также виды-кос мополиты, обитающие на огромных пространствах суши или океана. В силу этих обстоятельств географический критерий не может быть у ниверсальным.
Генетический критерий
Генетическое единство вида и, соответственно, генетическая изоляция
его от других видов — главный критерий вида, основной видовой
признак, обусловленный комплексом особенностей строения и жизне
деятельности организмов данного вида. Генетическая совмести
мость, сходство морфологических, физиологических, цитологических
и других признаков, одинаковое поведение, совместное обитание — все эт
о создает необходимые условия для успешного размножения и вос
производства вида. В то же время все эти признаки обеспечивают генетическую
изоляцию вида от других сходных с ним видов. Например, раз
личия в песне дроздов, пеночек, славок, зябликов и вьюрка, глухой
и обыкновенной кукушек препятствуют образованию смешанных пар,
несмотря на сходство их окраски и экологии (у птиц, обладающих специфической песней, почти не встречаются гибриды). Даже в тех случа
ях,
когда, несмотря на изоляционные барьеры, произошло скрещива
ние особей разных видов, гибридная популяция, как правило, не возникает, так как начинает действовать целый ряд постпопуляционных
механизмов изоляции. Важнейшие из них — это гибель мужских гамет (генет
ическая несовместимость), гибель зигот, нежизнеспособность гиб
ридов, их стерильность, наконец, невозможность найти полового
партнера и дать жизнеспособное плодовитое потомство.
Генетическая совмести
мость, сходство морфологических, физиологических, цитологических
и других признаков, одинаковое поведение, совместное обитание — все эт
о создает необходимые условия для успешного размножения и вос
производства вида. В то же время все эти признаки обеспечивают генетическую
изоляцию вида от других сходных с ним видов. Например, раз
личия в песне дроздов, пеночек, славок, зябликов и вьюрка, глухой
и обыкновенной кукушек препятствуют образованию смешанных пар,
несмотря на сходство их окраски и экологии (у птиц, обладающих специфической песней, почти не встречаются гибриды). Даже в тех случа
ях,
когда, несмотря на изоляционные барьеры, произошло скрещива
ние особей разных видов, гибридная популяция, как правило, не возникает, так как начинает действовать целый ряд постпопуляционных
механизмов изоляции. Важнейшие из них — это гибель мужских гамет (генет
ическая несовместимость), гибель зигот, нежизнеспособность гиб
ридов, их стерильность, наконец, невозможность найти полового
партнера и дать жизнеспособное плодовитое потомство.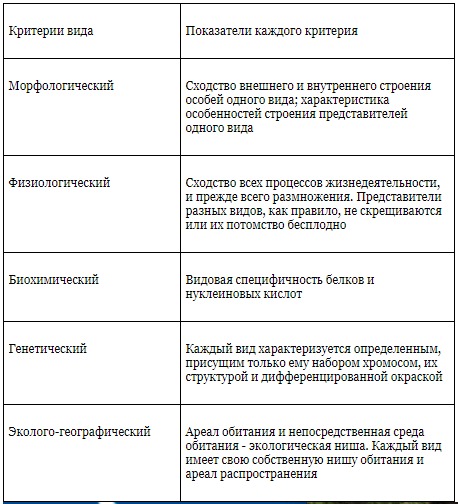 Мы знаем, что
для каждого вида характерен свой набор специфических признаков. Межвидовой гибрид будет иметь признаки, промежуточные между
признаками двух исходных родительских форм. Его песня, к примеру
не будет понята ни зябликом, ни вьюрком, если это гибрид этих видов, и он не найдет полового партнера. У такого гибрида при обра
зовании гамет содержащиеся в его клетках хромосомы зяблика «не уз
нают» хромосомы вьюрка и, не найдя гомологичного партнера, не
конъюгируют. В результате образуются гаметы с нарушенным набором
хромосом, которые, как правило, нежизнеспособны. И вследствие
го этого гибрид окажется стерильным.
Мы знаем, что
для каждого вида характерен свой набор специфических признаков. Межвидовой гибрид будет иметь признаки, промежуточные между
признаками двух исходных родительских форм. Его песня, к примеру
не будет понята ни зябликом, ни вьюрком, если это гибрид этих видов, и он не найдет полового партнера. У такого гибрида при обра
зовании гамет содержащиеся в его клетках хромосомы зяблика «не уз
нают» хромосомы вьюрка и, не найдя гомологичного партнера, не
конъюгируют. В результате образуются гаметы с нарушенным набором
хромосом, которые, как правило, нежизнеспособны. И вследствие
го этого гибрид окажется стерильным.
Ворон и ворона
Скажу сразу: Ворон — это не «муж» вороны, а самостоятельный вид.
Ворон — один из наиболее крупных представителей семейства вороновые , весит от 0,8 до 1,5 кг. Окраска оперения, клюва и ног у него однотонного черного цвета.
Распространен ворон почти по всему северному полушарию: встречается
почти по всей Европе, Азии, исключая Юго-Восточную, в Северной
Африке и Северной Америке. Повсеместно он ведет оседлый образ жизни. Населяет леса, пустыни и горы. В безлесных местностях держится у
скал, береговых обрывов речных долин. Спаривание и брачные игры на
юге страны отмечаются в первой половине февраля, на севере — в
марте. Пары постоянны. Гнезда обычно помещаются на вершинах высоких деревьев. В кладке от 3 до 7, чаще 4-6, яиц голубовато-зеленой окрас
ки с темными отметинами.
Повсеместно он ведет оседлый образ жизни. Населяет леса, пустыни и горы. В безлесных местностях держится у
скал, береговых обрывов речных долин. Спаривание и брачные игры на
юге страны отмечаются в первой половине февраля, на севере — в
марте. Пары постоянны. Гнезда обычно помещаются на вершинах высоких деревьев. В кладке от 3 до 7, чаще 4-6, яиц голубовато-зеленой окрас
ки с темными отметинами.
Ворон — всеядная птица. Основной его корм — падаль, которую он чаще всего находит на свалках и у скотобоен. Поедая падаль, он выступает как санитарная птица. Кормится он также грызунами, яйцами, и птенцами, рыбой, различными беспозвоночными животными, а места ми и зёрнами хлебных злаков.
Ворона по общему телосложению напоминает ворона, но значительно мельче его: весит от 460 до 690 г.
Описываемый вид интересен тем, что по окраске оперения распадается
на две группы: серую и черную. Серая ворона имеет хорошо извест
ную двуцветную окраску: голова, горло, крылья, хвост, клюв и ноги черные, остальное оперение серое. Черная ворона вся черная, с металлическим синим и пурпуровым блеском.
Черная ворона вся черная, с металлическим синим и пурпуровым блеском.
Каждая из этих групп имеет локальное распространение. Серая ворона широко распространена в Европе, Западной Азии, черная – в Центральной и Западной Европе, с одной стороны, в Средней, Восточной Азии и Северной Америке – с другой.
Населяет ворона опушки и окраины лесов, сады, рощи, заросли речных долин, реже скалы и склоны береговых обрывов. Это частично осёдлая, частично перелётная птица.
В начале марта в южных частях страны и в апреле-мае в северных и восточных начинается откладывание яиц. В кладке обычно 4-5 яиц бледно-зелёной, голубовато-зелёной или частично зелёной окраски с тёмными пятнышками и крапинками. Ворона – всеядная птица. Из животных она поедает различных беспозвоночных – жуков, муравьев, моллюсков, а также грызунов, ящериц, лягушек и рыб. Из растений клюёт зёрна культурных злаков, семена ели, вьюнка полевого, птичьей гречишки и др. Зимой питается в основном отбросами.
Заяц-беляк и заяц-русак
Род собственно зайцев, к которому относятся русак и беляк, а также еще 28 видов, довольно многочислен. Наиболее известные в России зайцы — беляк и русак
. Беляка можно встретить на территории от побережья Северного Ледовитого океана до южной границы лесной зоны, в Сибири — до границ с Казахста
ном, Китаем и Монголией, а на Дальнем Востоке — от Чукотки до
и Северной Кореи. Распространен беляк и в лесах Европы, а также на востоке Северной Америки. Русак обитает на территории Европейской России от Карелии
юга Архангельской области до южных границ страны, на Украине и в Закав
казье. А вот в Сибири этот заяц обитает только на юге и к западу от Байкала.
Наиболее известные в России зайцы — беляк и русак
. Беляка можно встретить на территории от побережья Северного Ледовитого океана до южной границы лесной зоны, в Сибири — до границ с Казахста
ном, Китаем и Монголией, а на Дальнем Востоке — от Чукотки до
и Северной Кореи. Распространен беляк и в лесах Европы, а также на востоке Северной Америки. Русак обитает на территории Европейской России от Карелии
юга Архангельской области до южных границ страны, на Украине и в Закав
казье. А вот в Сибири этот заяц обитает только на юге и к западу от Байкала.
Беляк получил свое название благодаря белоснежному зимнему меху. Только кончики ушей остаются у него черными весь год. Русак же в некоторых северных местностях тоже сильно светлеет к зиме, но снежно-белым он не бывает никогда. А на юге он вовсе не меняет окраски.
Русак больше приспособлен к жизни в открытых ландшафтах, поскольку он крупнее беляка, да и бегает лучше. На коротких дистанциях этот заяц может развивать
скорость до 50 км/час. У беляка лапы широкие, с густым опушением чтобы меньше проваливаться в рыхлые лесные сугробы. А у русака лапы уже,
ведь на открытых местах снег, как правило, твердый, слежавшийся, «утоптанный ветром».
У беляка лапы широкие, с густым опушением чтобы меньше проваливаться в рыхлые лесные сугробы. А у русака лапы уже,
ведь на открытых местах снег, как правило, твердый, слежавшийся, «утоптанный ветром».
Длина тела беляка — 45-75 см, масса — 2,5-5,5 кг. Уши короче, чем у русака. Длина тела русака — 50-70 см, масса до 5 (иногда 7) кг.
Размножаются зайцы обычно два, а на юге три или даже четыре раза в год. У зай цев-беляков в выводе может быть по два, три пять, семь зайчат, а у русаков — обычно всего один или два зайчонка. Русаки начинают пробовать траву через две недели после рождения, а беляки и того быстрее — через неделю.
Вид (лат. species) — таксономическая, систематическая единица, группа особей с общими морфофизиологическими, биохимическими и поведенческими признаками, способная к взаимному скрещиванию, дающему в ряду поколений плодовитое потомство, закономерно распространённая в пределах определённого ареала и сходно изменяющаяся под влиянием факторов внешней среды. Вид — реально существующая генетически неделимая единица живого мира, основная структурная единица в системе организмов, качественный этап эволюции жизни.
Вид — реально существующая генетически неделимая единица живого мира, основная структурная единица в системе организмов, качественный этап эволюции жизни.
Долгое время считалось, что любой вид — это закрытая генетическая система, то есть между генофондами двух видов нет обмена генами. Это утверждение верно для большинства видов, однако из него есть исключения. Так, к примеру, львы и тигры могут иметь общее потомство (лигры и тиграны), самки которого плодовиты — могут рожать как от тигров, так и львов. В неволе скрещиваются и многие другие виды, которые в природных условиях не скрещиваются из-за географической или репродуктивной изоляции. Скрещивание (гибридизация) между разными видами может происходить и в природных условиях, особенно при антропогенных нарушениях среды обитания, нарушающих экологические механизмы изоляции. Особенно часто гибридизуются в природе растения. Заметный процент видов высших растений имеет гибридогенное происхождение — они образовались при гибридизации в результате частичного или полного слияния родительских видов.
Основные критерии вида
1. Морфологический критерий вида. Основан на существовании морфологических признаков, характерных для одного вида, но отсутствующих у других видов.
Например: у гадюки обыкновенной ноздря находится в центре носового щитка, а у всех других гадюк (носатая, малоазиатская, степная, кавказская, гюрза) ноздря смещена к краю носового щитка.
В то же время в пределах видов существуют значительные индивидуальные морфологические различия. Например, гадюка обыкновенная представлена множеством цветовых форм (черные, серые, голубоватые, зеленоватые, красноватые и другие оттенки). Эти признаки не могут использоваться для разграничения видов.
2. Географический критерий. Основан на том, что каждый вид занимает определенную территорию (или акваторию) – географический ареал. Например, в Европе одни виды малярийного комара (род Anopheles) населяют Средиземноморье, другие – горы Европы, Северную Европу, Южную Европу.
Однако географический критерий не всегда применим.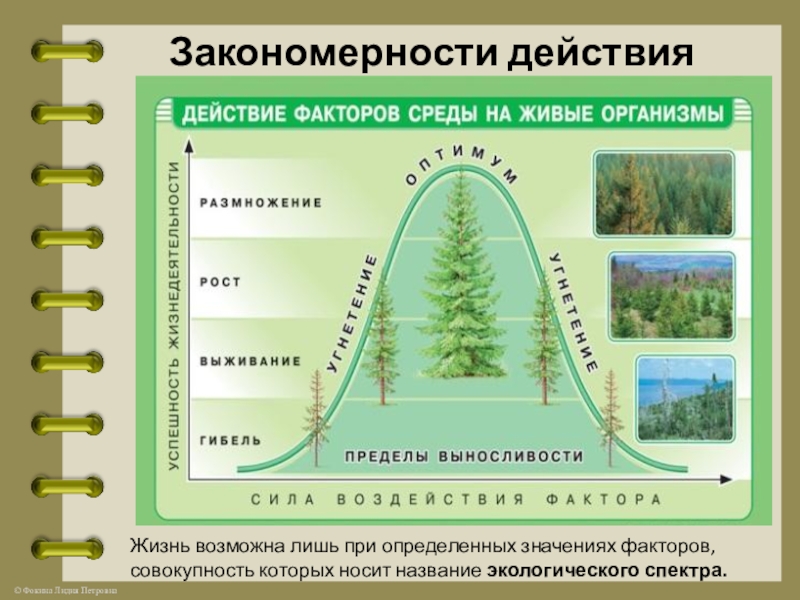 Ареалы разных видов могут перекрываться, и тогда один вид плавно переходит в другой. В этом случае образуется цепь викарирующих видов (надвид, или серия), границы между которыми часто можно установить только путем специальных исследований (например, чайка серебристая, клуша, западная, калифорнийская).
Ареалы разных видов могут перекрываться, и тогда один вид плавно переходит в другой. В этом случае образуется цепь викарирующих видов (надвид, или серия), границы между которыми часто можно установить только путем специальных исследований (например, чайка серебристая, клуша, западная, калифорнийская).
3. Экологический критерий. Основан на том, что два вида не могут занимать одну экологическую нишу. Следовательно, каждый вид характеризуется своими собственными отношениями со средой обитания.
Однако в пределах одного вида разные особи могут занимать разные экологические ниши. Группы таких особей называются экотипами. Например, один экотип сосны обыкновенной населяет болота (сосна болотная), другой – песчаные дюны, третий – выровненные участки боровых террас.
Совокупность экотипов, образующих единую генетическую систему (например, способных скрещиваться между собой с образованием полноценного потомства) часто называется эковидом.
4.
Молекулярно-генетический критерий.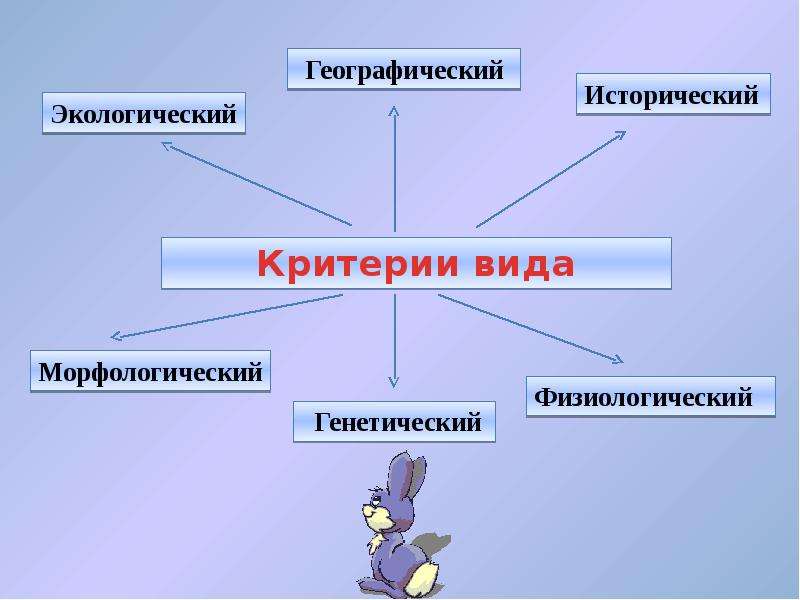 Основан на степени сходства и различия последовательностей нуклеотидов в нуклеиновых кислотах. Как правило, для оценки степени сходства или различия используются «некодирующие» последовательности ДНК (молекулярно-генетические маркеры). Однако в пределах одного и того же вида существует полиморфизм ДНК, а разные виды могут характеризоваться сходными последовательностями.
Основан на степени сходства и различия последовательностей нуклеотидов в нуклеиновых кислотах. Как правило, для оценки степени сходства или различия используются «некодирующие» последовательности ДНК (молекулярно-генетические маркеры). Однако в пределах одного и того же вида существует полиморфизм ДНК, а разные виды могут характеризоваться сходными последовательностями.
5. Физиолого-биохимический критерий. Основан на том, что разные виды могут различаться по аминокислотному составу белков. В то же время в пределах вида существует белковый полиморфизм (например, внутривидовая изменчивость многих ферментов), а разные виды могут иметь сходные белки.
6.
Цитогенетический (кариотипический) критерий. Основан на том, что каждый вид характеризуется определенным кариотипом – числом и формой метафазных хромосом. Например, у всех твердых пшениц в диплоидном наборе 28 хромосом, а всех мягких – 42 хромосомы. Однако у разных видов могут быть очень сходные кариотипы: например, у большинства видов семейства кошачьих 2n=38 . В то же время, в пределах одного вида может наблюдаться хромосомный полиморфизм. Например, у лосей евразийских подвидов 2n=68, а у лосей североамериканских видов 2n=70 (в кариотипе североамериканских лосей на 2 метацентрика меньше и на 4 акроцентрика больше). У некоторых видов существуют хромосомные расы, например, у черной крысы – 42-хромосомная (Азия, Маврикий), 40-хромосомная (Цейлон) и 38-хромосомная (Океания).
В то же время, в пределах одного вида может наблюдаться хромосомный полиморфизм. Например, у лосей евразийских подвидов 2n=68, а у лосей североамериканских видов 2n=70 (в кариотипе североамериканских лосей на 2 метацентрика меньше и на 4 акроцентрика больше). У некоторых видов существуют хромосомные расы, например, у черной крысы – 42-хромосомная (Азия, Маврикий), 40-хромосомная (Цейлон) и 38-хромосомная (Океания).
7. Репродуктивный критерий. Основан на том, что особи одного вида могут скрещиваться между собой с образованием плодовитого потомства, похожего на родителей, а особи разных видов, обитающих совместно, не скрещиваются между собой, или их потомство бесплодно.
Однако известно, что в природе часто распространена межвидовая гибридизация: у многих растений (например, ивы), ряда видов рыб, земноводных, птиц и млекопитающих (например, волк и собака). В то же время в пределах одного вида могут существовать группировки, репродуктивно изолированные друг от друга.
8. Этологический критерий. Связан с межвидовыми различиями в поведении у животных. У птиц для распознавания видов широко используется анализ песен. По характеру издаваемых звуков различаются разные виды насекомых. Разные виды североамериканских светляков различаются по частоте и цвету световых вспышек.
Этологический критерий. Связан с межвидовыми различиями в поведении у животных. У птиц для распознавания видов широко используется анализ песен. По характеру издаваемых звуков различаются разные виды насекомых. Разные виды североамериканских светляков различаются по частоте и цвету световых вспышек.
9. Исторический (эволюционный) критерий. Основан на изучении истории группы близких видов.
ГЕОГРАФИЧЕСКИМ КРИТЕРИЕМ ВИДА ЯВЛЯЕТСЯ ЧТО
Этот критерий носит комплексный характер, поскольку включает сравнительный анализ современных ареалов видов (географический критерий), сравнительный анализ геномов (молекулярно-генетический критерий), сравнительный анализ цитогеномов (цитогенетический критерий) и другие.
Ни один из рассмотренных критериев вида не является главным или наиболее важным. Для четкого разделения видов необходимо их тщательное изучение по всем критериям.
В связи с неодинаковыми условиями среды особи одного вида в пределах ареала распадаются на более мелкие единицы — популяции. Реально вид существует именно в виде популяций.
Реально вид существует именно в виде популяций.
Виды бывают монотипическими — со слабо дифференцированной внутренней структурой, они характерны для эндемиков. Политипические виды отличаются сложной внутривидовой структурой.
Внутри видов могут быть выделены подвиды — географически или экологически обособленные части вида, особи которых под влиянием факторов среды в процессе эволюции приобрели устойчивые морфофизиологические особенности, отличающие их от других частей этого вида. В природе особи разных подвидов одного вида могут свободно скрещиваться и давать плодовитое потомство.
Название вида
Научное название вида биномиально, то есть состоит из двух слов: названия рода, к которому принадлежит данный вид, и второго слова, называемого в ботанике видовым эпитетом, а в зоологии — видовым названием.
Первое слово — существительное в единственном числе; второе — либо прилагательное в именительном падеже, согласованное в роде (мужском, женском или среднем) с родовым названием, либо существительное в родительном падеже. Первое слово пишется с заглавной буквы, второе -со строчной.
Первое слово пишется с заглавной буквы, второе -со строчной.
- Petasites fragrans — научное название вида цветковых растений из рода Белокопытник (Petasites) (русское название вида — Белокопытник душистый). В качестве видового эпитета использовано прилагательное fragrans («душистый»).
- Petasites fominii — научное название ещё одного вида из этого же рода (русское название — Белокопытник Фомина). В качестве видового эпитета использована латинизированная фамилия (в родительном падеже) ботаника Александра Васильевича Фомина (1869-1935), исследователя флоры Кавказа.
Иногда используются также записи для обозначения неопределённых таксонов в ранге вида:
- Petasites sp. — запись обозначает, что имеется в виду таксон в ранге вида, относящийся к роду Petasites.
- Petasites spp. — запись обозначает, что имеются в виду все таксоны в ранге вида, входящие в род Petasites (либо все остальные таксоны в ранге вида, входящие в родPetasites, но не включённые в некий данный список таких таксонов).

Понятие вида. Вид как таксономическая категория
Чтобы изучать многообразие жизни, человеку необходимо было разработать систему классификации организмов для разделения их на группы. Как вы уже знаете, наименьшей структурной единицей в систематике живых организмов является вид.
Вид — исторически сложившаяся совокупность особей, которые сходны по морфологическим, физиологическим и биохимическим признакам, свободно скрещиваются и дают плодовитое потомство, приспособлены к определенным условиям среды и занимают в природе общую территорию — ареал.
Для того чтобы отнести особей к одному и тому же или к разным видам, их сравнивают между собой по ряду определенных характерных признаков — критериев.
Критерии вида
Совокупность характерных однотипных признаков, по которым особи одного вида схожи, а особи разных видов различаются между собой, называют критерием вида. В современной биологии выделяют следующие основные критерии вида: морфологический, физиологический, биохимический, генетический, экологический, географический.
Морфологический критерий отражает совокупность характерных признаков внешнего строения. Например, виды клевера различаются по окраске соцветий, форме и окраске листьев. Этот критерий относительный. В пределах вида особи могут заметно различаться по строению. Эти различия зависят от пола (половой диморфизм), стадии развития, стадии в цикле размножения, условий среды обитания, принадлежности к сортам или породам.
Например, у кряквы самец ярко окрашен, а самка темно-бурая, у благородного оленя самцы имеют рога, а у самок их нет. У бабочки капустной белянки гусеница отличается от взрослой особи внешними признаками. У папоротника щитовника мужского спорофит имеет листья и корни, а гаметофит представлен зеленой пластинкой с ризоидами. В то же время некоторые виды настолько схожи по морфологическим признакам, что их называют видами-двойниками. Например, некоторые виды малярийных комаров, дрозофил, североамериканских сверчков внешне не различаются, но не скрещиваются между собой.
Таким образом, на основании одного морфологического критерия нельзя судить о принадлежности особи к тому или иному виду.
Физиологический критерий
— совокупность характерных особенностей процессов жизнедеятельности (размножение, пищеварение, выделение и др.). Одним из важных признаков является способность особей скрещиваться. Особи разных видов не могут скрещиваться из-за несовместимости половых клеток, несоответствия половых органов. Этот критерий относительный, поскольку и особи одного вида иногда не могут скрещиваться. У мух дрозофил невозможность спаривания может быть обусловлена различием в строении полового аппарата. Это приводит к нарушению процессов размножения. И наоборот, известны такие виды, представители которых могут скрещиваться между собой. Например, лошадь и осел, представители некоторых видов ив, тополей, зайцев, канареек. Из этого следует, что для определения видовой принадлежности особей недостаточно сравнивать их только по физиологическому критерию.
Биохимический критерий отражает характерный химический состав тела и обмен веществ. Это самый ненадежный критерий. Нет веществ или биохимических реакций, характерных только для определенного вида. Особи одного вида могут значительно различаться по этим показателям. Тогда как у особей разных видов синтез белков и нуклеиновых кислот происходит одинаково. Ряд биологически активных веществ играют аналогичную роль в обмене веществ у разных видов. Например, хлорофилл у всех зеленых растений участвует в фотосинтезе. Значит, определение видовой принадлежности особей на основании одного биохимического критерия тоже невозможно.
Генетический критерий
характеризуется определенным набором хромосом, сходных по размерам, форме и составу. Это самый надежный критерий, так как он является фактором репродуктивной изоляции, поддерживающей генетическую целостность вида. Однако и этот критерий не является абсолютным. У особей одного вида число, размеры, форма и состав хромосом могут различаться в результате геномных, хромосомных и генных мутаций. В то же время при скрещивании некоторых видов иногда появляются жизнеспособные плодовитые межвидовые гибриды. Например, собака и волк, тополь и ива, канарейка и зяблик при скрещивании дают плодовитое потомство. Таким образом, сходства по данному критерию также недостаточно, чтобы отнести особей к одному виду.
В то же время при скрещивании некоторых видов иногда появляются жизнеспособные плодовитые межвидовые гибриды. Например, собака и волк, тополь и ива, канарейка и зяблик при скрещивании дают плодовитое потомство. Таким образом, сходства по данному критерию также недостаточно, чтобы отнести особей к одному виду.
Экологический критерий — это совокупность характерных факторов среды, необходимых для существования вида. Каждый вид может обитать в той среде, где климатические условия, особенности почвы, характер рельефа и источники пищи соответствуют его пределам толерантности. Но в этих же условиях среды могут обитать и организмы других видов. Выведение человеком новых пород животных и сортов растений показало, что особи одного вида (дикие и окультуренные) могут жить в сильно различающихся условиях среды.
ГЕОГРАФИЧЕСКИМ КРИТЕРИЕМ ВИДА ЯВЛЯЕТСЯ…
Это доказывает относительный характер экологического критерия. Следовательно, есть необходимость использования других критериев при определении принадлежности особей к определенному виду.
Географический критерий характеризует способность особей одного вида населять в природе определенную часть земной поверхности (ареал).
Например, лиственница сибирская распространена в Сибири (Зауралье), а лиственница даурская — в Приморском крае (Дальний Восток), морошка — в тундре, а черника — в умеренной зоне.
Этот критерий указывает на приуроченность вида к определенному местообитанию. Но есть виды, не имеющие четких границ расселения, а обитающие практически повсеместно (лишайники, бактерии). У некоторых видов ареал совпадает с ареалом человека. Такие виды называются синантропными (комнатная муха, постельный клоп, домовая мышь, серая крыса). У разных видов могут быть совпадающие местообитания. Значит, и этот критерий имеет относительный характер. Он не может использоваться в качестве единственного для определения видовой принадлежности особей.
Таким образом, ни один из описанных критериев не является абсолютным и универсальным. Поэтому при определении принадлежности особи к определенному виду следует учитывать все его критерии.
Ареал вида. Понятие об эндемиках и космополитах
Согласно географическому критерию, каждый вид в природе занимает определенную территорию — ареал.
Ареал (от лат. area — площадь, пространство) — часть земной поверхности, в пределах которой распространены и проходят полный цикл своего развития особи данного вида.
Ареал может быть сплошным или прерывистым, обширным или ограниченным. Виды, имеющие обширный ареал в пределах разных континентов, называются видами-космополитами (некоторые виды протистов, бактерий, грибов, лишайников). Когда ареал распространения очень узкий и находится в пределах небольшого региона, то населяющий его вид называется эндемиком (от греч. endemos — местный).
Например, кенгуру, ехидна и утконос обитают только в Австралии. Гинкго в естественных условиях произрастает только в Китае, рододендрон остроконечный и лилия даурская — только на Дальнем Востоке.
Вид — совокупность особей, которые сходны по морфологическим, физиологическим и биохимическим признакам, свободно скрещиваются и дают плодовитое потомство, приспособлены к определенным условиям среды и занимают в природе общую территорию — ареал. Для каждого вида характерны следующие критерии: морфологический, физиологический, биохимический, генетический, экологический, географический. Все они носят относительный характер, поэтому при определении видовой принадлежности особей используются все возможные критерии.
Для каждого вида характерны следующие критерии: морфологический, физиологический, биохимический, генетический, экологический, географический. Все они носят относительный характер, поэтому при определении видовой принадлежности особей используются все возможные критерии.
Согласно упрощенной морфологической концепции вида, природные популяции, морфологически отличные друг от друга, признаются видами .
Точнее и более правильно определять виды как природные популяции, в пределах которых изменчивость морфологических (обычно количественных) признаков носит непрерывный характер, отделенные от других популяций разрывом. Если различия невелики, но непрерывность распределения нарушена, то такие формы должны приниматься за разные виды. В афористичной форме это выражается так: критерием вида служит дискретность границ распределения признаков .
При определении видов часто возникают затруднения, вызванные двумя обстоятельствами. Во-первых, причиной затруднений может быть сильная внутривидовая изменчивость, а во-вторых, наличие так называемых видов-двойников. Рассмотрим эти случаи.
Рассмотрим эти случаи.
Внутривидовая вариабельность может достигать большого размаха. Прежде всего это различия между самцами и самками одного и того же вида. Такие различия четко проявляются у многих птиц, дневных бабочек, ос-немок, некоторых рыб и других организмов. Подобные факты были использованы Дарвином в работах о половом отборе. У ряда животных резкие различия наблюдаются между взрослыми и неполовозрелыми особями. Подобные факты широко известны зоологам. Поэтому очень полезны выборки из популяций видов на разных стадиях их жизненного цикла. Теоретическая основа внутривидовой изменчивости (индивидуальной или групповой) изложена в ряде руководств. Здесь будут рассмотрены только признаки, которые чаще всего используются при установлении видового статуса особей из выборки.
Морфологические признаки
— это общая наружная морфология и, если необходимо, структура генитального аппарата. Наиболее важные морфологические признаки имеются у животных с наружным скелетом, таких как членистоногие или моллюски, но их можно найти и у многих других животных, без панцирей или раковин. Это всевозможные отличия в шерстном покрове зверей, оперении птиц, рисунке крыльев бабочек и т. д.
Это всевозможные отличия в шерстном покрове зверей, оперении птиц, рисунке крыльев бабочек и т. д.
Во многих случаях критерием, позволяющим различить близкие виды, является структура гениталий. Это особенно подчеркивается сторонниками биологической концепции вида, поскольку различия в форме хитинизированных или склеротизированных частей генитального аппарата препятствуют скрещиванию между самцами одного вида и самками другого. В энтомологии известно правило Дюфура, по которому у видов с хитинизированными частями гениталий самцов и совокупительными органами самок наблюдается такое соотношение, как у ключа и замка. Иногда его так и называют — правило «ключа и замка». Следует, однако, помнить о том, что признаки гениталий, как и другие морфологические признаки, у некоторых видов также варьируют (например, у жуков-листоедов рода Altica), что было неоднократно показано. Тем не менее в тех группах, где систематическое значение строения гениталий доказано, оно служит очень ценным признаком, так как при дивергенции видов их структура должна меняться одной из первых.
Анатомические признаки, такие как детали строения черепа или форма зубов, обычно используются в надвидовой систематике позвоночных животных.
Экологические признаки . Известно, что каждый вид животных характеризуется определенными экологическими предпочтениями, зная которые, нередко можно если не совсем точно решить, с каким видом мы имеем дело, то хотя бы значительно облегчить идентификацию. Согласно правилу конкурентного исключения (правило Гаузе), два вида не могут существовать в одном и том же месте, если их экологические требования одинаковы.
При изучении галлообразующих или минирующих насекомых-фитофагов (мушки-галлицы, осы-орехотворки, минирующие личинки бабочек, жуков и других насекомых) зачастую главными признаками оказываются формы мин, для которых даже разработана классификация, или галлов. Так, на шиповниках иди на дубах развивается несколько видов орехотворок, вызывающих образование галлов на листьях или побегах растений. И во всех случаях галлы каждого вида имеют свою характерную форму.
Пищевые предпочтения животных достигли большого размаха — от строгой монофагии через олигофагию до полифагии. Известно, что гусеницы тутового шелкопряда питаются исключительно листьями тутовника, или шелковицы. Гусеницы бабочек-белянок (капустницы, репницы и др.) грызут листья крестоцветных растений, не переходя на растения других семейств. А медведь или кабан, будучи полифагами, питаются и животной, и растительной пищей.
В группах животных, где установлен строгий выбор пищи, по характеру погрыза определенного вида растений можно установить их видовую принадлежность. Так поступают энтомологи в полевых условиях. Лучше, конечно, собрать для дальнейшего изучения и самих растительноядных насекомых. Опытный натуралист, хорошо знающий природные условия определенной местности, заранее может предсказать, с каким набором видов животных можно встретиться, посещая те или иные биотопы — лес, луг, песчаные дюны или берег реки. Поэтому на этикетках, сопровождающих коллекционные сборы, обязательно нужно указывать условия, в которых были собраны те или иные виды. Это сильно облегчает дальнейшую обработку коллекции и идентификацию видов.
Это сильно облегчает дальнейшую обработку коллекции и идентификацию видов.
Этологические признаки . На таксономическую ценность этологических признаков указывает ряд авторов. Известный этолог Хайнд считает поведение таксономическим признаком, который может быть использован для уточнения систематического положения видов. К этому следует добавить, что наиболее полезными оказываются стереотипные действия. Они столь же характерны для каждого вида, как и любые морфологические признаки. Это следует иметь в виду при изучении близкородственных видов или видов-двойников. Если даже элементы поведения могут быть сходными, то выражение этих элементов специфично для каждого вида.
Вопрос: ГЕОГРАФИЧЕСКИМ КРИТЕРИЕМ ВИДА ЯВЛЯЕТСЯ ЧТО
Дело в том, что особенности поведения являются у животных важными изолирующими механизмами, препятствующими скрещиванию между разными видами. Примерами этологической изоляции являются случаи, когда потенциальные брачные партнеры встречаются, но не спариваются.
Как показывают многочисленные наблюдения в природе и эксперименты в лабораторных условиях, отологические признаки вида в первую очередь проявляются в особенностях брачного поведения. К ним относятся характерные позы самцов в присутствии самки, а также голосовые сигналы. Изобретение звукозаписывающих аппаратов, особенно сонографов, позволяющих представить звук в графической форме, окончательно убедило исследователей в видоспецифичности песен не только птиц, но и сверчков, кузнечиков, цикадок, а также голосов лягушек и жаб.
Но не только позы или голоса животных являются этологическими видовыми признаками. К ним относятся и особенности строительства гнезд у птиц и насекомых из отряда перепончатокрылых (пчел и ос), типы и характер яйцекладок у насекомых, форма паутинных ловчих сетей у пауков и многое другое. Видоспецифичны оотеки богомолов и кубышки саранчовых, световые вспышки жуков-светляков.
Иногда различия имеют количественный характер, но и этого достаточно, чтобы распознать видовую принадлежность объекта исследования.
Географические признаки . Часто географические признаки являются удобным средством различения популяций, точнее, решения вопроса о том, являются ли две изучаемые популяции одним или разными видами. Если ряд форм географически замещают друг друга, образуя цепь или кольцо форм, каждая из которых отличается от своих соседей, то они называются аллопатрическими формами . Считается, что аллопатрические формы представляют собой политипический вид, состоящий из нескольких подвидов.
Противоположную картину представляют случаи, когда ареалы форм частично или полностью совпадают. Если между этими формами нет переходов, то их называют симпатрическими формами . Такой характер распространения указывает на полную видовую самостоятельность данных форм в связи с тем, что симпатрическое (совместное) существование, не сопровождающееся скрещиванием, служит одним из основных критериев вида.
В практике систематика часто возникают затруднения в отнесении конкретной аллопатрической формы к виду или подвиду. Если аллопатрические популяции соприкасаются, но не скрещиваются в зоне контакта, то такие популяции следует считать видами. В противоположность этому, в случае если аллопатрические популяции соприкасаются и в узкой зоне контакта свободно скрещиваются или в широкой зоне контакта связаны переходами, то их почти всегда следует считать подвидами.
Если аллопатрические популяции соприкасаются, но не скрещиваются в зоне контакта, то такие популяции следует считать видами. В противоположность этому, в случае если аллопатрические популяции соприкасаются и в узкой зоне контакта свободно скрещиваются или в широкой зоне контакта связаны переходами, то их почти всегда следует считать подвидами.
Сложнее обстоит дело, когда между ареалами аллопатрических популяций есть разрыв, из-за которого контакт невозможен. В этом случае мы можем иметь дело либо с видами, либо с подвидами. Классическим примером подобного рода является географическая разобщенность популяций голубой сороки. Один подвид (С. с. cooki) населяет Пиренейский полуостров, а другой (С. с. cyanus) — юг Дальнего Востока (Приморье и смежные части Китая). Считается, что это результат разрыва прежнего сплошного ареала, возникшего в ледниковом периоде. Многие систематики придерживаются мнения, что сомнительные аллопатрические популяции целесообразнее считать подвидами.
Другие признаки
. Во многих случаях близкородственные виды легче различать по морфологии хромосом, чем по другим признакам, что и было продемонстрировано на видах рода Drosophila и на клопах семейства Lygaeidae. Все больший размах приобретает использование физиологических признаков, по которым можно различать близкие таксоны. Было показано, что у близкородственных видов комаров значительно различаются скорость роста и длительность стадии яйца. Все большее признание получает вывод о том, что основная часть белков специфична для каждого вида. На этом феномене базируются выводы в области серосистематики. Полезным оказалось также изучение специфических выделений, образующих на теле определенный рисунок или восковые сооружения в виде колпачков, как у щитовок или мучнистых червецов из класса насекомых. Они также видоспецифичны. Нередко приходится использовать всю совокупность признаков различного характера для решения сложных таксономических проблем. В современных работах по зоологической систематике, как показывает ознакомление с новейшими публикациями, авторы не ограничиваются одними только морфологическими признаками.
Во многих случаях близкородственные виды легче различать по морфологии хромосом, чем по другим признакам, что и было продемонстрировано на видах рода Drosophila и на клопах семейства Lygaeidae. Все больший размах приобретает использование физиологических признаков, по которым можно различать близкие таксоны. Было показано, что у близкородственных видов комаров значительно различаются скорость роста и длительность стадии яйца. Все большее признание получает вывод о том, что основная часть белков специфична для каждого вида. На этом феномене базируются выводы в области серосистематики. Полезным оказалось также изучение специфических выделений, образующих на теле определенный рисунок или восковые сооружения в виде колпачков, как у щитовок или мучнистых червецов из класса насекомых. Они также видоспецифичны. Нередко приходится использовать всю совокупность признаков различного характера для решения сложных таксономических проблем. В современных работах по зоологической систематике, как показывает ознакомление с новейшими публикациями, авторы не ограничиваются одними только морфологическими признаками. Чаще всего присутствуют указания на хромосомный аппарат.
Чаще всего присутствуют указания на хромосомный аппарат.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Вконтакте
Одноклассники
Вид, критерии вида. Популяции.
Вид – совокупность особей, обладающих наследственным сходством морфологических, физиологических и биологических особенностей, свободно скрещивающихся и дающих плодовитое потомство, приспособившихся к определенным условиям жизни и занимающих в природе определенный ареал.
Виды представляют собой устойчивые генетические системы, так как в природе отделены друг от друга целым рядом барьеров.
Вид представляет собой одну из основных форм организации живого. Однако определить, принадлежат ли данные особи к одному виду или нет, иногда бывает трудно. Поэтому для решения вопроса о принадлежности особей к данному виду используется целый ряд критериев:
Морфологический критерий
– главный критерий, основанный на внешних различиях между видами животных или растений. Этот критерий служит для разделения организмов, которые четко отличаются по внешним или внутренним морфологическим признакам. Но следует отметить, что очень часто между видами существуют очень тонкие отличия, которые можно выявить лишь при длительном изучении данных организмов.
Этот критерий служит для разделения организмов, которые четко отличаются по внешним или внутренним морфологическим признакам. Но следует отметить, что очень часто между видами существуют очень тонкие отличия, которые можно выявить лишь при длительном изучении данных организмов.
Географический критерий – основан на том, что каждый вид обитает в пределах определенного пространства (ареала). Ареал – это географические границы распространения вида, размеры, форма и расположение в биосфере которого отлично от ареалов других видов. Однако этот критерий также недостаточно универсален по трем причинам. Во-первых, ареалы многих видов географически совпадают, во-вторых, существуют виды космополиты, для которых ареалом является практически вся планета (кит-касатка). В-третьих, у некоторых быстрорасселяющихся видов (домового воробья, домовой мухи и др.) ареал настолько быстро изменяет свои границы, что не может быть определен.
Экологический критерий
– предполагает, что каждый вид характеризуется определенным типом питания, местом обитания, сроками размножения, т. е.
е.
занимает определенную экологическую нишу.
Этологический критерий – заключается в том, что поведение животных одних видов отличается от поведения других.
Генетический критерий – заключает в себе главное свойство вида – его генетическую изоляцию от других. Животные и растения разных видов почти никогда не скрещиваются между собой. Конечно, вид не может быть полностью изолирован от потока генов со стороны близкородственных видов, но при этом он сохраняет постоянство генетического состава на протяжении эволюционно длительного времени. Самые четкие границы между видами проходят именно с генетической точки зрения.
Физиолого-биохимический критерий
– этот критерий не может служить надежным способом разграничения видов, так как основные биохимические процессы протекают у сходных групп организмов одинаково. А в пределах каждого вида существует большое число приспособлений к конкретным условиям обитания путем изменения протекания физиолого-биохимических процессов.
По одному из критериев нельзя точно различать виды между собой. Определить принадлежность особи к конкретному виду можно только на основании совокупности всех или большинства критериев. Особи, занимающие определенную территорию и свободно скрещивающиеся между собой, называются популяцией.
Популяция
– совокупность особей одного вида, занимающих определенную территорию и обменивающихся генетическим материалом. Совокупность генов всех особей в популяции называется генофондом популяции. В каждом поколении отдельные особи вносят больший или меньший вклад в общий генофонд в зависимости от их приспособительной ценности. Неоднородность организмов, входящих в популяцию, создает условия для действия естественного отбора, поэтому популяция считается наименьшей эволюционной единицей, с которой начинается эволюционные преобразования вида — видообразования. Популяция, таким образом, представляет собой надорганизменную формулу организации жизни. Популяция не является полностью изолированной группой. Иногда происходит скрещивание между особями различных популяций. Если какая-то популяция окажется полностью географически или экологически изолированной от других, то она может дать начало новому подвиду, а впоследствии и виду.
Иногда происходит скрещивание между особями различных популяций. Если какая-то популяция окажется полностью географически или экологически изолированной от других, то она может дать начало новому подвиду, а впоследствии и виду.
Каждая популяция животных или растений состоит из особей разного пола и различного возраста. Соотношение численности этих особей может быть различно в зависимости от времени года, природных условий. Численность популяции определяется соотношением рождаемости и смертности, составляющих ее организмов. Если на протяжении достаточно длительного времени эти показатели равны, то численность популяции не изменяется. Факторы среды, взаимодействие с другими популяциями может изменять численность популяции.
Качественным этапом процесса эволюции является вид. Buд
— это совокупность особей|особей, которые сходны по морфофизиологическим признакам, способны скрещиваться между собой, давать плодовитое потомство и формируют систему популяций, образующих общий ареал.
Каждый вид живых организмов можно описать исходя из совокупности характерных|характерных чёрт|черт, свойств, которые называются признаками. Признаки вида, с помощью которых один вид отличают от другого, называются критериями вида. Наиболее часто используют шесть общих критериев вида: морфологический, физиологический, географический, экологический, генетический и биохимический.
Морфологический критерий предполагает описание внешних (морфологических) признаков особей|особей, входящих в состав определённого вида. По внешнему виду, размерам и окраске оперения можно, например, легко отличить большого пёстрого дятла от зелёного, малого пёстрого дятла от желны, большую|большую синицу от хохлатой, длиннохвостой, голубой и от гаички. По внешнему виду побегов и соцветий, размерам и расположению листьев легко различают виды клевера|клевера: луговой, ползучий, люпиновый, горный.
Морфологический критерий самый удобный и поэтому широко используется в систематике. Однако этот критерий недостаточен для различения видов, которые имеют значительное морфологическое сходство. К настоящему времени накоплены факты, свидетельствующие о существовании видов-двойников, не имеющих заметных морфологических различий, но в природе не скрещивающихся из-за наличия разных хромосомных наборов. Так, под названием «крыса чёрная» различают два вида-двойника: крыс, имеющих в кариотипе 38 хромосом и живущих на всей территории Европы, Африки, Америки, Австралии, Новой Зеландии, Азии к западу|западу от Индии, и крыс, имеющих 42 хромосомы, распространение которых связано с монголоидными оседлыми цивилизациями, населяющими Азию к востоку от Бирмы. Установлено также, что под названием «малярийный комар» существует 15 внешне не различимых видов.
Однако этот критерий недостаточен для различения видов, которые имеют значительное морфологическое сходство. К настоящему времени накоплены факты, свидетельствующие о существовании видов-двойников, не имеющих заметных морфологических различий, но в природе не скрещивающихся из-за наличия разных хромосомных наборов. Так, под названием «крыса чёрная» различают два вида-двойника: крыс, имеющих в кариотипе 38 хромосом и живущих на всей территории Европы, Африки, Америки, Австралии, Новой Зеландии, Азии к западу|западу от Индии, и крыс, имеющих 42 хромосомы, распространение которых связано с монголоидными оседлыми цивилизациями, населяющими Азию к востоку от Бирмы. Установлено также, что под названием «малярийный комар» существует 15 внешне не различимых видов.
Физиологический критерий
заключается в сходстве жизненных процессов, в первую очередь в возможности скрещивания между особями одного вида с образованием плодовитого потомства. Между разными видами существует физиологическая изоляция. Например, у многих видов дрозофилы сперма особей|особей чужого вида вызывает иммунологическую реакцию в половых путях самки, что приводит к гибели сперматозоидов. В то же время между некоторыми видами живых организмов скрещивание возможно; при этом могут образовываться плодовитые гибриды (зяблики, канарейки, вороны|вороны, зайцы, тополя|тополя, ивы и др.)
Например, у многих видов дрозофилы сперма особей|особей чужого вида вызывает иммунологическую реакцию в половых путях самки, что приводит к гибели сперматозоидов. В то же время между некоторыми видами живых организмов скрещивание возможно; при этом могут образовываться плодовитые гибриды (зяблики, канарейки, вороны|вороны, зайцы, тополя|тополя, ивы и др.)
Географический критерий (географическая определённость вида)
основан на том, что каждый вид занимает определённую территорию или акваторию. Иными словами, каждый вид характеризуется определённым географическим ареалом.
Многие виды занимают разные ареалы. Но огромное число видов имеет совпадающие (накладывающиеся) или перекрывающиеся ареалы. Кроме того, существуют виды, не имеющие чётких границ распространения, а также виды-космополиты, обитающие на огромных пространствах суши|суши или океана. Космополитами являются некоторые обитатели внутренних водоёмов — рек и пресноводных озёр (виды рдестов, ряски, тростник). Обширный набор космополитов имеется среди сорных и мусорных растений, синантропных животных (виды, обитающие рядом с человеком или его
жилищем) — постельный клоп, рыжий таракан, комнатная муха, а также одуванчик лекарственный, ярутка полевая, пастушья сумка и др.
Обширный набор космополитов имеется среди сорных и мусорных растений, синантропных животных (виды, обитающие рядом с человеком или его
жилищем) — постельный клоп, рыжий таракан, комнатная муха, а также одуванчик лекарственный, ярутка полевая, пастушья сумка и др.
Существуют также виды, которые имеют разорванный ареал. Так, например, липа растёт в Европе, встречается в Кузнецком Алатау и Красноярском крае. Голубая сорока|сорока имеет две части ареала — западноевропейскую и восточносибирскую. В силу этих обстоятельств географический критерий, как и другие, не является абсолютным.
Экологический критерий основан на том, что каждый вид может существовать только в определённых условиях, выполняя соответствующую функцию в определённом биогеоценозе. Иными словами, каждый вид занимает определённую экологическую нишу. Например, лютик едкий произрастает на пойменных лугах|лугах, лютик ползучий — по берегам рек и канав, лютик жгучий — на заболоченных местах. Существуют, однако, виды, которые не имеют строгой экологичекой приуроченности. Во-первых, это синантропные виды. Во-вторых, это виды, которые находятся под опекой человека: комнатные и культурные растения, домашние животные.
Генетический (цитоморфологический) критерий основан на различии видов по кариотипам, т. е. по числу, форме и размерам хромосом. Для подавляющего большинства видов характерен|характерен строго определённый кариотип. Однако и этот критерий не является универсальным. Во-первых, у многих разных видов число хромосом одинаково и форма их сходна. Так, многие виды из семейства бобовых имеют 22 хромосомы (2n = 22). Во-вторых, в пределах одного и того же вида могут встречаться особи с разным числом хромосом, что является результатом геномных мутаций. Например, ива козья имеет диплоидное (38) и тетраплоидное (76) число хромосом. У серебристого карася встречаются популяции с набором хромосом 100, 150,200, тогда как нормальное число их равно 50. Таким образом, в случае возникновения полиплоидных или анеушюидных (отсутствие одной хромосомы или появление лишней в геноме) форм на основе генетического критерия нельзя достоверно определить принадлежность особей|особей к конкретному виду.
Биохимический критерий позволяет различить виды по биохимическим параметрам (состав и структура определённых белков, нуклеиновых кислот и других веществ). Известно, что синтез определённых высокомолекулярных веществ присущ лишь отдельным группам видов. Например, по способности образовывать и накапливать алкалоиды различаются виды растений в пределах семейств паслёновых, сложноцветных, лилейных, орхидных. Или, к примеру, для двух видов бабочек из рода|рода амата диагностическим признаком является наличие двух ферментов — фосфоглюкомутазы и эстеразы-5. Однако этот критерий не находит широкого применения — он трудоёмкий и далеко не универсальный. Существует значительная внутривидовая изменчивость практически всех биохимических показателей вплоть до последовательности аминокислот в молекулах белков и нуклеотидов в отдельных участках ДНК.
Таким образом, ни один из критериев в отдельности не может служить для определения вида. Охарактеризовать вид можно только по их совокупности.
Определение вида
Во все времена у разных исследователей были свои взгляды на вид, как таковой. Жан-Батист Ламарк признавал лишь группы особей, живущих в определенных условиях, то есть – популяции. Карл Линей, признавая реальность видов, отрицал факт из эволюции.
Согласно утверждениям Чарльза Дарвина и его последователей, виды реально существуют в виде совокупности особей. Каждый вид более-менее четко отделен от других, имеет определенные свойства и ареал. В результате эволюции виды изменяются. Каждому виду предшествует целая цепочка предковых форм, образуя филогенетический ряд.
Современная биологическая концепция дает такое определение вида:
Определение 1
Вид – это совокупность популяций особей, подобных между собой по строению, функциям, местом в биогеоценозе (экологическая ниша), которые населяют определенную часть биосферы (ареал), свободно скрещиваются, дают плодовитое потомство и не гибридизируются с другими видами.
Но в последнее время тезис о невозможности гибридизации с другими видами пересмотрен. Ученые перестали рассматривать виды как генетически закрытые системы. Некоторые виды могут скрещиваться между собой и давать плодовитое гибридное потомство. Но это лишь исключение из общей закономерности.
Критерии вида
Для того, чтобы четко отличить один вид от другого, учеными-систематиками (классификаторами) был разработан четкий перечень правил и признаков. Эти признаки называются критериями вида. Ознакомимся с ними детальнее.
- В основу морфологического критерия положен анализ наличия или отсутствия сходства по внешним или внутренним признакам особей одного вида (от строения хромосом, до особенностей строения отдельных органов и их частей). Морфологические признаки, уникальные для определенного вида, называют диагностическими.
- Генетический критерий – это характерное для каждого вида количество хромосом, их размер, форма. Этот критерий дает возможность судить о главных, ключевых признваках вида.
- Физиологический критерий базируется на анализе сходства и различиях в процессах жизнедеятельности организмов. К ним принадлежит и способность к парованию и рождения плодовитого потомства.
- Биохимический критерий – это особенности строения и состава макромолекул (в первую очередь – белков) и хода химических реакций, характерных для особей данного вида.
- Географический критерий заключается в том, что особи одного вида занимают какую-то определенную часть биосферы, которая отличается от ареалов близких видов. Но этот критерий не может быть решающим, так как существуют виды. Распространены повсеместно (часто с помощью человека) – например некоторые грызуны или насекомые-вредители.
- Экологический критерий подразумевает совокупность факторов внешней среды, в которой обитает вид. Условия обитания в каждом отдельном месте являются уникальными и неповторимыми. Они влияют на организмы, там проживающие, вызывают в них адаптивные реакции. Каждый вид занимает свою экологическую нишу в биогеоценозе.
Замечание 1
Для четкого и достоверного определения видовой принадлежности особи недостаточно использовать какой-либо один критерий. Только их совокупность, взаимное подтверждение могут дать верную характеристику вида.
Целостность вида
Как мы уже знаем, вид существует в форме популяций. Каждая из этих популяций под воздействием движущих сил эволюции приспособилась к жизни в определенных условиях (адаптировалась). Благодаря этому обстоятельству вид, состоящий из многочисленных популяций. Занимает обширный ареал, несмотря на разнообразие природных условий в его пределах.
Замечание 2
Но, несмотря на это, любой вид, как состоящий из одной популяции, так и слагающийся из их большого количества, составляет единое целое. Эта целостность достигается обособленностью одного вида от остальных.
Целостность вида обусловлена, также, связями между его отдельными особями (в стае, стаде, семье). Вся эта система разнообразных связей обеспечивает существование вида как целостной системы.
У особей каждого вида в процессе эволюции могут вырабатываться взаимные приспособления (забота о потомстве, система коммуникации, защита от врагов). Иногда видовые приспособления могут привести к гибели отдельных особей, но они оказываются полезными виду в целом.
Анатомический критерий вида. Урок Вид. Критерии вида. Географическим критерием вида является…
Определение вида
Во все времена у разных исследователей были свои взгляды на вид, как таковой. Жан-Батист Ламарк признавал лишь группы особей, живущих в определенных условиях, то есть – популяции. Карл Линей, признавая реальность видов, отрицал факт из эволюции.
Согласно утверждениям Чарльза Дарвина и его последователей, виды реально существуют в виде совокупности особей. Каждый вид более-менее четко отделен от других, имеет определенные свойства и ареал. В результате эволюции виды изменяются. Каждому виду предшествует целая цепочка предковых форм, образуя филогенетический ряд.
Современная биологическая концепция дает такое определение вида:
Определение 1
Вид – это совокупность популяций особей, подобных между собой по строению, функциям, местом в биогеоценозе (экологическая ниша), которые населяют определенную часть биосферы (ареал), свободно скрещиваются, дают плодовитое потомство и не гибридизируются с другими видами.
Но в последнее время тезис о невозможности гибридизации с другими видами пересмотрен. Ученые перестали рассматривать виды как генетически закрытые системы. Некоторые виды могут скрещиваться между собой и давать плодовитое гибридное потомство. Но это лишь исключение из общей закономерности.
Критерии вида
Для того, чтобы четко отличить один вид от другого, учеными-систематиками (классификаторами) был разработан четкий перечень правил и признаков. Эти признаки называются критериями вида. Ознакомимся с ними детальнее.
- В основу морфологического критерия положен анализ наличия или отсутствия сходства по внешним или внутренним признакам особей одного вида (от строения хромосом, до особенностей строения отдельных органов и их частей). Морфологические признаки, уникальные для определенного вида, называют диагностическими.
- Генетический критерий – это характерное для каждого вида количество хромосом, их размер, форма. Этот критерий дает возможность судить о главных, ключевых признваках вида.
- Физиологический критерий базируется на анализе сходства и различиях в процессах жизнедеятельности организмов. К ним принадлежит и способность к парованию и рождения плодовитого потомства.
- Биохимический критерий – это особенности строения и состава макромолекул (в первую очередь – белков) и хода химических реакций, характерных для особей данного вида.
- Географический критерий заключается в том, что особи одного вида занимают какую-то определенную часть биосферы, которая отличается от ареалов близких видов. Но этот критерий не может быть решающим, так как существуют виды. Распространены повсеместно (часто с помощью человека) – например некоторые грызуны или насекомые-вредители.
- Экологический критерий подразумевает совокупность факторов внешней среды, в которой обитает вид. Условия обитания в каждом отдельном месте являются уникальными и неповторимыми. Они влияют на организмы, там проживающие, вызывают в них адаптивные реакции. Каждый вид занимает свою экологическую нишу в биогеоценозе.
Замечание 1
Для четкого и достоверного определения видовой принадлежности особи недостаточно использовать какой-либо один критерий. Только их совокупность, взаимное подтверждение могут дать верную характеристику вида.
Целостность вида
Как мы уже знаем, вид существует в форме популяций. Каждая из этих популяций под воздействием движущих сил эволюции приспособилась к жизни в определенных условиях (адаптировалась). Благодаря этому обстоятельству вид, состоящий из многочисленных популяций. Занимает обширный ареал, несмотря на разнообразие природных условий в его пределах.
Замечание 2
Но, несмотря на это, любой вид, как состоящий из одной популяции, так и слагающийся из их большого количества, составляет единое целое. Эта целостность достигается обособленностью одного вида от остальных.
Целостность вида обусловлена, также, связями между его отдельными особями (в стае, стаде, семье). Вся эта система разнообразных связей обеспечивает существование вида как целостной системы.
У особей каждого вида в процессе эволюции могут вырабатываться взаимные приспособления (забота о потомстве, система коммуникации, защита от врагов). Иногда видовые приспособления могут привести к гибели отдельных особей, но они оказываются полезными виду в целом.
В биологии видом называют определенную совокупность особей, которые обладают наследственным сходством физиологических, биологических и морфологических особенностей, способны свободно скрещиваться и производить жизнеспособное потомство. Виды являются устойчивыми генетическими системами, поскольку в природе они отделены некоторым рядом барьеров друг от друга. Ученые различают их между собой по ряду основных признаков. Обычно выделяют следующие критерии вида: морфологический, географический, экологический, генетический, физико-биохимический.
Морфологический критерий
Такие признаки являются главными в этой системе. Морфологические критерии вида основаны на внешних различиях между отдельными группами животных или растений. Данное условие подразделяет на виды организмы, четко отличающиеся между собой по внутренним или внешним морфологическим признакам.
Географические критерии вида
Основаны на том, что представители каждой устойчивой генетической системы обитают в пределах ограниченных пространств. Такие зоны называют ареалами. Вместе с тем географический критерий имеет некоторые недочеты. Он недостаточно универсален по следующим причинам. Во-первых, существуют некоторые космополитические виды, которые распространены по всей планете (например, кит-касатка). Во-вторых, у многих биологических совокупностей ареалы географически совпадают. В-третьих, в случае некоторых слишком быстро расселяющихся популяций ареалы очень изменчивы (например, воробей или домовая муха).
Экологический критерий вида
Предполагается, что каждому виду свойственны определенные характеристики вроде типа питания, сроков размножения, места обитания и всего того, что определяет занимаемую им экологическую нишу. В основе данного критерия лежит предположение о том, что поведение одних животных отличается от поведения других.
Генетический критерий вида
Здесь учитывается главное свойство любого вида — его генетическая изоляция от других. Растения и животные различных устойчивых генетических систем практически никогда между собой не скрещиваются. Разумеется, вид совершенно полностью не может быть изолирован от притока генов от родственных видов. Однако же при этом он в целом сохраняет постоянство своего генетического состава в течение действительно длительного периода времени. Именно в генетической составляющей лежат самые четкие различия между представителями разных биологических популяций.
Физико-биохимические критерии вида
Подобные критерии также не могут служить абсолютно надежным способом для разграничения видов, поскольку фундаментальные биохимические процессы
происходят у подобных групп одинаково. А в среде каждого отдельного вида существует какое-то количество приспособлений к определенным условиям обитания, что сказывается на изменении биохимических процессов.
Выводы
Таким образом, по какому-то отдельному критерию весьма затруднительно различать между собой виды. Принадлежность особи к какому-либо конкретному виду должна определяться лишь через комплексное сравнение по целому ряду критериев — всех или хотя бы большинства. Особи, которые занимают некоторую территорию и способны свободно скрещиваться между собой, являются видовой популяцией.
Вид. Критерии вида
Вертьянов С. Ю.
Различить надвидовые таксоны, как правило, довольно легко, но четкое разграничение самих видов встречает определенные трудности. Часть видов занимает географически разделенные области обитания (ареалы) и потому не скрещивается, а в искусственных условиях дает плодовитое потомство. Линнеевское краткое определение вида как группы особей, свободно скрещивающихся между собой и дающих плодовитое потомство, неприменимо к организмам, размножающимся партеногенетически или бесполым путем (бактерии и одноклеточные животные, многие высшие растения), а также к вымершим формам.
Совокупность отличительных признаков вида называют его критерием.
Морфологический критерий основан на сходстве особей одного вида по комплексу признаков внешнего и внутреннего строения. Морфологический критерий — один из основных, но в ряде случаев морфологического сходства оказывается недостаточно. Малярийным комаром ранее называли шесть не скрещивающихся похожих видов, из которых только один разносит малярию. Существуют так называемые виды-двойники. Два вида черных крыс, внешне практически неразличимых, живут раздельно и не скрещиваются. Самцы многих существ, например птиц (снегирей, фазанов), внешне мало похожи на самок. Взрослые самец и самка нитехвостого угря так непохожи, что ученые полвека помещали их в разные роды, а иногда даже в разные семейства и подпорядки.
Физиолого-биохимический критерий
В его основе лежит сходство процессов жизнедеятельности особей одного вида. Часть видов грызунов обладает способностью впадать в спячку, у других она отсутствует. Многие близкие виды растений различаются по способности синтезировать и накапливать определенные вещества. Биохимический анализ позволяет различить виды одноклеточных организмов, не размножающихся половым путем. Бациллы сибирской язвы, например, вырабатывают белки, которые не встречаются у других видов бактерий.
Возможности физиолого-биохимического критерия имеют ограничения. Часть белков обладает не только видовой, но и индивидуальной специфичностью. Существуют биохимические признаки, одинаковые у представителей не только разных видов, но даже отрядов и типов. Сходным образом могут протекать у разных видов и физиологические процессы. Так, интенсивность обмена веществ у некоторых арктических рыб такая же, как у других видов рыб южных морей.
Генетический критерий
Все особи одного вида обладают сходным кариотипом. Особи разных видов имеют разные хромосомные наборы, не могут скрещиваться и живут в естественных условиях отдельно друг от друга. У двух видов-двойников черных крыс разное количество хромосом — 38 и 42. Кариотипы шимпанзе, горилл и орангутанов различаются расположением генов в гомологичных хромосомах. Аналогичны отличия кариотипов зубра и бизона, имеющих в диплоидном наборе по 60 хромосом. Различия в генетическом аппарате некоторых видов могут быть еще более тонкими и состоять, например, в разном характере включения и выключения отдельных генов. Применение только генетического критерия иногда оказывается недостаточным. Один вид долгоносика объединяет диплоидные, триплоидные и тетраплоидные формы, домовая мышь также имеет различные наборы хромосом, а ген ядерного белка-гистона Н1 человека отличается от гомологичного ему гена гороха всего одним нуклеотидом. В геноме растений, животных и человека обнаружены такие изменчивые последовательности ДНК, что по ним у людей можно различать братьев и сестер.
Репродуктивный критерий
(Лат. reproducere воспроизводить) основан на способности особей одного вида давать плодовитое потомство. Важную роль при скрещивании играет поведение особей — брачный ритуал, видоспецифические звуки (пение птиц, стрекотание кузнечиков). По характеру поведения особи узнают брачного партнера своего вида. Особи сходных видов могут не скрещиваться по причине несоответствия поведения при спаривании или несовпадения мест размножения. Так, самки одного вида лягушек мечут икру по берегам рек и озер, а другого — в лужах. Похожие виды могут не скрещиваться из-за различия брачных периодов или сроков спаривания при обитании в разных климатических условиях. Разные сроки цветения у растений препятствуют перекрестному опылению и служат критерием принадлежности к разным видам.
Репродуктивный критерий тесно связан с генетическим и физиологическим критериями. Жизнеспособность гамет зависит от осуществимости конъюгации хромосом в мейозе, а значит, от сходства или различия кариотипов скрещивающихся особей. Резко понижает возможность скрещивания различие в суточной физиологической активности (дневной или ночной образ жизни).
Применение только репродуктивного критерия не всегда позволяет четко разграничить виды. Существуют виды, хорошо различимые по морфологическому критерию, но дающие при скрещивании плодовитое потомство. Из птиц это некоторые виды канареек, зябликов, из растений — разновидности ив и тополей. Представитель отряда парнокопытных бизон обитает в степях и лесостепях Северной Америки и никогда в естественных условиях не встречается с зубром, обитающим в лесах Европы. В условиях зоопарка эти виды дают плодовитое потомство. Так была восстановлена популяция европейских зубров, практически истребленная во время мировых войн. Скрещиваются и дают плодовитое потомство яки и крупный рогатый скот, белые и бурые медведи, волки и собаки, соболи и куницы. В царстве растений межвидовые гибриды встречаются еще чаще, среди растений существуют даже межродовые гибриды.
Эколого-географический критерий
Большинство видов занимает определенную территорию (ареал) и экологическую нишу. Лютик едкий растет на лугах и полях, в более сырых местах распространен другой вид — лютик ползучий, по берегам рек и озер — лютик жгучий. Сходные виды, обитающие в одном ареале, могут различаться экологическими нишами — например, если питаются различной пищей.
Применение эколого-географического критерия ограничено рядом причин. Ареал вида может быть прерывист. Видовой ареал зайца-беляка — острова Исландия и Ирландия, север Великобритании, Альпы и северо-запад Европы. Некоторые виды имеют одинаковый ареал, например два вида черных крыс. Есть организмы, распространенные почти повсеместно, — многие сорные растения, ряд насекомых-вредителей и грызунов.
Проблема определения вида иногда вырастает в сложную научную задачу и решается с привлечением комплекса критериев. Таким образом, вид — совокупность особей, занимающих определенный ареал и обладающих единым генофондом, обеспечивающим наследственное сходство морфологических, физиолого-биохимических и генетических признаков, в природных условиях скрещивающихся и дающих плодовитое потомство.
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://www.portal-slovo.ru
Генетический (цитогенетический) критерий вида наряду с другими применяется для разграничения элементарных систематических групп, анализа состояния вида. В данной статье рассмотрим характеристику критерия, а также сложности, с которыми может столкнуться исследователь, применяющий его.
В разных отраслях биологической науки вид определяется по-своему. В эволюционном ракурсе можно сказать, что вид — это совокупность особей, имеющих сходство внешнего строения и внутренней организации, физиологических и биохимических процессов, способных к неограниченному скрещиванию, оставлению плодовитого потомства и генетически обособленная от сходных групп.
Вид может быть представлен одной или несколькими популяциями и, соответственно, иметь цельный или рассеченный ареал (территорию/акваторию обитания)
Номенклатура вида
Каждый вид имеет свое название. В соответствии с правилами бинарной номенклатуры, оно состоит из двух слов: существительного и прилагательного. Имя существительное является родовым названием, а прилагательное — видовым. Например, в названии «Одуванчик лекарственный», вид «лекарственный» является одним из представителей растений рода «Одуванчик».
Особи родственных видов внутри рода имеют некоторые различия внешнего вида, физиологии, экологических предпочтений. Но если они слишком похожи, то их видовую принадлежность определяет генетический критерий вида на основе анализа кариотипов.
Для чего виду нужны критерии
Карл Линней, первым давший современные названия и описавший множество видов живых организмов, считал их неизменными и невариабельными. То есть все особи соответствуют единому видовому образу, а любые отклонения от него являются ошибкой воплощения видовой идеи.
С первой половины XIX века Чарльз Дарвин и его последователи обосновывают совершенно другую концепцию вида. В соответствии с ней, вид изменчив, неоднороден и включает переходные формы. Постоянство вида относительно, оно зависит от изменчивости условий окружающей среды. Элементарная единица существования вида — популяция. Она репродуктивно обособлена и соответствует генетическому критерию вида.
Учитывая неоднородность особей одного вида, ученым бывает сложно определить видовую принадлежность организмов или распределить их между систематическими группами.
Морфологический и генетический критерии вида, биохимический, физиологический, географический, экологический, поведенческий (этологический) — все это комплексы различий между видами. Они определяют изолированность систематических групп, их репродуктивную дискретность. И по ним можно отличить один вид от другого, установить степень их родства и положение в биологической системе.
Характеристика генетического критерия вида
Сущность данного признака в том, что все особи одного вида обладают одинаковым кариотипом.
Кариотип — это своеобразный хромосомный «паспорт» организма, он определяется количеством хромосом, присутствующих в зрелых соматических клетках организма, их размерами и особенностями строения:
- соотношением длины плеч хромосом;
- положением в них центромер;
- наличием вторичных перетяжек и спутников.
Принадлежащие к разным видам особи не смогут скрещиваться. Даже если получение потомства возможно, как у осла и лошади, тигра и льва, то межвидовые гибриды не будут плодовитыми. Это объясняется тем, что половинки генотипа неодинаковы и конъюгация между хромосомами не может произойти, поэтому гаметы не образуются.
На фото: мул — бесплодный гибрид осла и кобылы.
Объект исследования — кариотип
Кариотип человека представлен 46 хромосомами. У большинства исследованных видов количество отдельных молекул ДНК в ядре, образующих хромосомы, укладывается в диапазон 12 — 50. Но есть исключения. Плодовая мушка дрозофила имеет по 8 хромосом в ядрах клеток, а у мелкого представителя семейства Чешуекрылые Lysandra диплоидный хромосомный набор равен 380.
Электронная микрофотография кондесированных хромосом, позволяющая оценить их форму и размер, отражает кариотип. Анализ кариотипа в рамках исследования генетического критерия, а также сравнение кариотипов между собой помогает определить видовую принадлежность организмов.
Когда два вида как один
Общий признак критериев вида в том, что они не являются абсолютными. Это значит, что применение только одного из них может быть недостаточным для точного определения. Организмы, внешне неотличимые друг от друга, могут оказаться представителями разных видов. Здесь морфологическому приходит на помощь генетический критерий. Примеры двойников:
- На сегодняшний день известно два вида черных крыс, которые раньше определялись как один благодаря внешней идентичности.
- Существует не менее 15 видов малярийных комаров, которые различимы лишь благодаря цитогенетическому анализу.
- В Северной Америке найдено 17 видов сверчков, имеющих генетические различия, но фенотипически относимых к единому виду.
- Считается, что среди всех видов птиц есть 5% двойников, для идентификации которых нужно применять генетический критерий.
- Путаница в систематике горных полорогих была устранена благодаря кариологическому анализу. Выделено 3 разновидности кариотипов (у муфлонов 2n=54, у архаров и аргали — 56, уриалы имеют по 58 хромосом).
Один из видов черных крыс имеет 42 хромосомы, кариотип другого представлен 38 молекулами ДНК.
Когда один вид как два
Для видовых групп с большой площадью ареала и численностью особей, когда внутри них действует географическая изоляция или особи имеют широкую экологическую валентность, характерно наличие особей с различными кариотипами. Такое явление — еще один вариант исключений в генетическом критерии вида.
Примеры хромосомного и геномного полиморфизма часты у рыб:
- у форели радужной количество хромосом меняется от 58 до 64;
- две кариоморфы, с 52 и 54 хромосомами, обнаружены у сельди беломорской;
- при диплоидном наборе в 50 хромосом, представители разных популяций карася серебряного имеют по 100 (тетраплоиды), 150 (гексаплоиды), 200 (октаплоиды) хромосом.
Полиплоидные формы находятся и у растений (козья ива), и у насекомых (долгоносики). Домовые мыши и песчанки могут иметь разное количество хромосом, не кратное диплоидному набору.
Двойники по кариотипу
У представителей разных классов и типов могут встречаться кариотипы с одним числом хромосом. Гораздо больше таких совпадений среди представителей одних семейств и родов:
- У горилл, орангутанов и шимпанзе кариотип, состоящий из 48 хромосом. На вид различия не определяются, тут нужно сравнивать порядок нуклеотидов.
- Незначительны отличия в кариотипах североамериканского бизона и европейского зубра. У обоих по 60 хромосом в диплоидном наборе. Они будут отнесены к одному виду, если проводить анализ только по генетическому критерию.
- Примеры генетических двойников находятся и среди растений, особенно внутри семейств. Среди ив возможно даже получение межвидовых гибридов.
Для выявления тонких различий генетического материала у таких видов нужно определять последовательности генов и порядок их включения.
Влияние мутаций на анализ критерия
Количество хромосом кариотипа может быть изменено в результате геномных мутаций — анеуплоидии или эуплоидии.
При анеуплоидии в кариотипе появляется одна или несколько добавочных хромосом, а также может быть число хромосом меньшее, чем у полноценной особи. Причина такого нарушения в нерасхождении хромосом на стадии образования гамет.
На рисунке пример анеуплоидии у человека (синдром Дауна).
Зиготы с уменьшенным числом хромосом, как правило, не приступают к дроблению. А полисомные организмы (с «лишними» хромосомами») вполне могут оказаться жизнеспособными. В случае трисомии (2n+1) или пентасомии (2n+3) нечетное число хромосом укажет на аномалию. Тетрасомия же (2n+2) может привести к фактической ошибке определения вида по генетическому критерию.
Умножение кариотипа — полиплоидия — также может ввести в заблуждение исследователя, когда кариотип мутанта представляет сумму нескольких диплоидных наборов хромосом.
Сложность критерия: неуловимые ДНК
Диаметр нити ДНК в раскрученном состоянии составляет 2 нм. Генетический критерий определяет кариотип в период, предшествующий делению клетки, когда тонкие молекулы ДНК многократно спирализуются (конденсируются) и представляют собой плотные палочковидные структуры — хромосомы. Толщина хромосомы в среднем 700 нм.
Школьные и университетские лаборатории обычно оснащены микроскопами с небольшим увеличением (от 8 до 100), рассмотреть в них детали кариотипа не представляется возможным. Разрешающая способность светового микроскопа, кроме того, позволяет на любом, даже самом большом увеличении увидеть объекты не меньше половины длины самой короткой световой волны. Наименьшая длина — у волн фиолетового цвета (400 нм). Это значит, что мельчайший объект, различимый в световой микроскоп будет от 200 нм.
Получается, что окрашенный деконденсированный хроматин будет выглядеть как замутненные области, а хромосомы будут видны без деталей. Четко увидеть и сравнить между собой разные кариотипы позволяет электронный микроскоп с разрешающей способностью от 0,5 нм. Учитывая толщину нитевидной ДНК (2 нм), она будет хорошо различима под таким прибором.
Цитогенетический критерий в школе
По причинам, описанным выше, использование микропрепаратов на лабораторных работах по генетическому критерию вида нецелесообразно. В заданиях можно применять фотографии хромосом, полученные под электронным микроскопом. Для удобства работы на фото отдельные хромосомы объединяют в гомологичные пары и располагают по порядку. Такая схема называется кариограммой.
Пример задания для лабораторной работы
Задание. Рассмотрите приведенные фотографии кариотипов, сравните их и сделайте вывод о принадлежности особей к одному или двум видам.
Фотографии кариотипов для сравнения на лабораторной работе.
Работа над заданием. Подсчитать общее количество хромосом на каждом фото кариотипа. В случае совпадения сравнить их по внешнему виду. Если представлена не кариограмма, среди хромосом средней длины найти самые короткие и самые длинные на обоих изображениях, сопоставить их по размеру и расположению центромер. Сделать вывод о различии/сходстве кариотипов.
Ответы на задание:
- Если количество, размер и форма хромосом совпадают, то две особи, чей генетический материал представлен для изучения, относятся к одному виду.
- Если количество хромосом отличается в два раза, и на обеих фотографиях встречаются одинаковые по размеру и форме хромосомы, то, скорее всего, особи являются представителями одного вида. Это будут кариотипы диплоидной и тетраплоидной форм.
- Если число хромосом неодинаково (отличается на одну — две), но в целом форма и размеры хромосом обоих кариотипов совпадают, речь идет о нормальной и мутантной формах одного вида (явление анеуплоидии).
- При разном количестве хромосом, а также несовпадении характеристик размера и формы, критерий отнесет представленных особей к двум разным видам.
В выводе требуется указать, можно ли на основании генетического критерия (и только его) определять видовую принадлежность особей.
Ответ: нельзя, поскольку любой видовой критерий, в том числе и генетический, имеет исключения и может дать ошибочный результат определения. Точность может гарантировать лишь применение комплекса критериев вида.
Цели: сформировать понятия «вид» и «критерии вида»; показать механизмы репродуктивной изоляции в природе; продолжить формирование умений давать морфологическое описание растений, работать с текстами, составлять таблицы, анализировать, формулировать выводы.
Оборудование: картины по зоологии: «Заяц- беляк», «Заяц-русак», «Медведь бурый», «Медведь белый»; разда- точный материал «Заяц-беляк и заяц-русак», «Ворон и ворона».
Ход урока
I . Организационный момент
Учитель объявляет тему и задачи урока.
11. Актуализация знаний
1. Выполнение заданий (устно).
Задание № 1
Назовите знакомые вам виды растений и животных, обитающих рядом с вашим домом или школой.
Задание № 2
Учитель демонстрирует картину «Бурый медведь» и задает вопросы:
Название этого вида животного — Медведь бурый. Какое из этих двух слов относится к родовому названию, какое — к видовому?
Назовите другой вид животного из этого же рода. (Это Медведь белый).
Учитель вывешивает картину, иллюстрирующую белого медведя, рядом с картиной «Медведь бурый».
Сравните два вида одного рода. Покажите черты сходства и отличия.
Задание № 3 ,
В указанном списке животных подсчитайте количество особей, видов и родов.
1. Еж обыкновенный.
2. Лисица обыкновенная.
3. Медведь гималайский или белогрудый.
4. Хомячок джунгарский.
5. Заяц-беляк.
6. Медведь бурый.
7. Хомячок сирийский или золотистый.
8. Заяц-русак.
9. Еж ушастый.
10. Лисица обыкновенная.
(Ответ: количество особей — 10; видов — 9; родов — 5 (Еж, Лиси- ца, Медведь, Хомячок, Заяц).)
При выполнении последнего задания у ряда учащихся возникает проблема: отнести зайца-беляка и зайца-русака к одному виду или к двум разным видам. Правдиво или ошибочно мнение, что заяц- беляк — это заяц-русак зимой?
Доклад «Заяц русак и заяц беляк».
— Какой вывод можно сделать по результатам выполнения всех заданий по актуализации знаний?
Вывод:
1. Для обозначения вида используется двойная (бинарная) но менклатура, согласно которой сначала указывают род, к которому относится вид (существительное), а затем видовое название (прила гательное).
2. Особи разных видов отличаются друг от друга по местам оби тания, внешним признакам и др.
3. Сходные виды объединяются в один род.
4. Вид — это основная категория биологической классификации.
III . Изучение нового материала
1. Рассказ учителя.
— Что же такое вид и каковы его критерии?
Вопрос о виде и критериях вида занимает центральное место в теории эволюции и служит предметом многочисленных иссле дований в области систематики, зоологии, ботаники и других наук. И это вполне объяснимо: четкое понимание сущности вида необходимо для выяснения механизмов эволюционного процесса.
Строгое общепринятое определение вида до сих пор не разра ботано. В биологическом энциклопедическом словаре мы на ходим следующее определение вида:
«Вид — совокупность популяций особей, способных к скрещива нию с образованием плодовитого потомства, населяющих определ енный ареал, обладающих рядом общих морфофизиологических признакови отдаленных от других таких же групп особей практи чески полным отсутствием гибридных форм».
Сравните это определение с тем, что дается в вашем учебнике (учебник А.А. Каменского, § 4.1, с. 134 ).
Поясним понятия, которые встречаются. в определении вида:
Ареал — область распространения данного вида или популяции в природе.
Популяция (от лат. «рор uius » — народ, население) — совокуп ность особей одного вида, обладающих общим генофондом и зани мающих определенную территорию — ареал.
Генофонд — совокупность генов, которые имеются у особей данной популяции.
Рассмотрим историю развития взглядов на вид в биологии.
Понятие вида впервые ввел в науку английский ботаник Джон Рей в XVII веке . Основополагающая работа по проблеме вида была написана шведским натуралистом и естествоиспытателем Карлом Линнеем в XVIII веке , в которой он предложил первое научное определение вида, уточнил его критерии.
Комментарии учителя. К. Линней считал, что вид — универ сальная, реально существующая единица живой материи, морфо логически однородная и неизменная . Все особи вида, по мнению ученого, имеют типичный морфологический облик, а вариации представляют собой случайные отклонения , результат несовершенного воплощения идеи вида (своего рода уродства ). Ученый считал, что виды неизменны, неизменна природа. Идея неизмен ной природы покоилась на концепции креационизма, согласно которой все сущее было сотворено Богом. Применительно к биологии эту концепцию Линней выразил в своей знаменитой фор муле «Видов столько, сколько разных форм вначале произвело Бесконечное существо».
Другая концепция принадлежит Тому Батисту Ламарку — вели кому французскому натуралисту. По его концепции виды реально не существуют , это чисто умозрительное понятие, изобретенное для того, чтобы легче было рассматривать совокупно большее количество особей, поскольку по словам Ламарка, «в природе нет ни чего кроме особей». Индивидуальная изменчивость непрерывна, поэтому границу между видами можно провести и здесь, и там — где удобнее.
Третья концепция подготавливалась в первой четверти XIX века. Она была обоснована Чарльзом Дарвином и последующими биолога ми. По этой концепции виды имеют независимую реальность. Вид неоднороден, представляет собой систему соподчиненных единиц. С реди них основная элементарная единица — популяция. Виды, по Дарвину, изменяются, они относительно постоянны и являются рез ультатом эволюционного развития .
Таким образом, понятие «вид» имеет длительную историю становления в биологической науке.
Порой самые опытные биологи становятся в тупик, определяя, принадлежат ли данные особи к одному виду или нет . Почему так происходит, существуют ли точные и строгие критерии, которые по могли бы разрешить все сомнения?
Критерии вида — это признаки, по которым один вид отлича ется от другого. Они же являются изолирующими механизмами, препятствующими скрещиванию, независимости, самостоятельно сти видов.
Нам известно, что одной из главных особенностей биологической материи на нашей планете является дискретность. Это выражается в том, что она представлена отдельными видами, не скрещивающимися друг с другом, изолированными один от дру гого.
Существование вида обеспечивается его генетическим единством (особи вида способны скрещиваться и давать жизнеспособное плодовитое потомство) и его генетической самостоятельностью (невоз можность скрещивания с особями другого вида, нежизнеспособно сть или стерильность гибридов).
Генетическая самостоятельность вида обусловливается совокуп ностью его характерных признаков: морфологических, физиологических, биохимических, генетических, особенностей образа жизни, поведения, географического распространения и т.д. Это и есть крит ериивида.
Познакомимся с ними.
2. Работа в группах
Каждая группа получает текст, иллюстрирующий один из критериев вида. Через 5 минут необходимо рассказать о сути данного критерияи о том,в чем недостаток данного критерия. По мере выступления групп классом заполняется таблица «Критерии вида».
Таблица № 1
Критерии вида.
|
Название критерия |
Признаки особей по критерию |
Исключение |
|
1. Морфологический |
Сходство внешнего и внутреннего строения организмов. |
Виды-двойники, половой диморфизм, полиморфизм. |
|
2. Физиологический |
Сходство всех процессов жизнедеятельности и возможность получения плодовитого потомства при скрещивании. |
У разных видов сходство процессов жизнедеятельности. Наличие межвидовых гибридов. |
|
3. Экологический |
Сходство по способам питания, местам обитания, наборам факторов внешней среды, необходимых для существования. |
Экологические ниши разных видов перекрываются. |
|
4. Географический |
Занимают определённый ареал. |
Космополиты. Совпадение ареалов разных видов. |
|
5. Биохимический |
Сходство по биохимическим параметрам – состав и структура белков, нуклеиновых кислот. |
Есть очень близкие по биохимическому составу виды. |
|
6. Этологический |
Сходство в поведении. Особенно в брачный период (ритуалы ухаживания, брачные песни и т. д.). |
Существуют виды с близким поведением. |
|
7. Цито-генетический а)Цитологический |
Особи одного вида скрещиваются между собой и дают плодовитое потомство (основан на сходстве числа хромосом, их формы и строения). |
Хромосомный полиморфизм в пределах вида; у многих разных видов число хромосом одинаково. |
|
б) Генетический |
Генетическая изоляция видов. Наличие постпопуляционных механизмов изоляции. Важнейшие из них — это гибель мужских гамет (генетическая несовместимость), гибель зигот, нежизнеспособность гибридов, их стерильность, наконец, невозможность найти полового партнера и дать жизнеспособное плодовитое потомство |
Собака и волк, тополь и ива, канарейка и зяблик дают плодовитое потомство. (Наличие межвидовых гибридов) |
|
8. Исторический |
Общность предков, единая история возникновения и развития вида. |
Итак, видовые критерии, по которым мы отличаем один вид другого, в совокупности обусловливают генетическую изоляцию ви дов, обеспечивая самостоятельность каждого вида и разнообразие в природе. По сути, в выработке этих изолирующих видовых призна ков и заключается процесс формирования видов. Именно поэтому изучение видовых критериев имеет определяющее значение для по нимания механизмов процесса эволюции, происходящего на нашей планете.
3. Формулирование выводов.
После заполнения таблицы формулируются выводы:
1) Видовые критерии, по которым отличается один вид от друг го, в совокупности обусловливают генетическую изоляцию видов, обеспечивая самостоятельность каждого вида и разнообразие их в природе.
2) Нет ни одного видового критерия, который можно было признать абсолютным и универсальным.
3) К способам изоляции, препятствующим скрещиванию разных видов относятся:
а) различия ареалов, мест обитания => невозможность встречи;
б) разные сроки размножения;
в) различия в строении половых органов;
г) нежизнеспособность или стерильность гибридов;
д) разные ритуалы «ухаживания» в период размножения.
4) Вид представляет собой генетически относительно изолиро ванную систему, что доказывает реальность существования видов в природе.
Вспомните, о чем говорилось в тексте «Заяц-беляк и заяц- русак». Какие критерии вида использованы при описании зай цев?
Ответить на вопрос:
— Какие критерии вида использованы при описанииживотных?
1). Лебедь-шипун часто изгибает шею в виде буквы S , а клюв и голову держит наклонно к воде. При раз дражении издает характерный шипящий звук, по которому и получил свое название. Распространен лебедь-шипун на изолированных участках в средней и южной полосе Европы и Азии от южной Швеции, Дании и Польши на западе до Монголии, Приморского края и Китая на Востоке. Всюду на этой территории редок, часто пара от пары гнездится на гро мадном расстоянии, а во многих районах вовсе отсутствует. Населяет заросшие водной растительностью лиманы, озера, иногда даже болота, предпочитая глухие, мало посещаемые человеком.
Малый, или тундровый лебедь распространен по тундре Азии от Кольского полуострова на западе до дельты Колымы на Востоке, заходя в область лесотундры и западные острова Северного Ледовитого. Для гнездования выбирает заболоченные и низкие травянистые участки разбросанными по ним озерами, а также речные долины, изобилующие старицами и протоками.
Брачные игры своеобразны и проходят на суше. При этомсамец ходит перед самкой, вытягивает шею, временами приподнимает крылья, издавая ими особый хлопающий звук и звонко кричит.
2). Городская ласточка. Верх го ловы, спина, крылья и хвост сине-черные, надхвостье и весь низ тела белые. Хвост с резкой треугольной вырезкой на конце. Обитатель горных и культурного ландшафтов. Гнездится на стенах скал и зданий. П ерелетная птица. Держится стаями в воздухе или сидя на проводах , чаще других ласточек садится на з емлю. Гнездится колониями . Гнездо лепит из комочков глины в форме полушара с боковым входом . Кладка из 4-6 белых яиц в мае – июне. Гол ос – звонкое «тиррч-тиррч»
Береговая ласточка. Верх головы, шеи, спина, крылья, хвост и полоса поперёк груди серовато-бурые, горло, грудь и брюшко белые. Хвост с неглубокой вырезкой.
Населяет долины рек, где гнездится по обрывистым глиняным или песчаным берегам. Обычная или многочисленная перелетная птица. Держится стаями, гнездится колониями. Гнезда устраивает в норах по обрывистым берегам рек. Кладка из 4-6 белых яиц в мае-июле. Голос — негромкое «чирр- чирр
Домашнее задание
По учебнику А.А. Каменского, § 4.1, вопросы после параграфа, термины.
Индивидуально:
1) Сообщение «Правда ли, что ворон — это муж вороны?»
2 ) Используя литературные источники, приведите конкретные пр имеры географического, экологического и этологического критериев.
Дополнительный материал для работы в группах.
Критерии вида
Морфологический критерий
Это был первый и долгое время единственный критерий, используемый для описания видов.
Морфологический критерий самый удобный и заметный, поэтому и сейчас широко используется в систематике растений и животных.
Мы можем легко отличить по размерам и окраске оперения большого пестрого дятла от зеленого дятла, малого пестрого дятла и желны (черного дятла), большую синицу от хохлатой, длиннохвостой, голубой и гаички, клевер луговой от ползучего и люпиновидного и т. д.
Несмотря на удобство, этот критерий не всегда «работает». Им не воспользуешься для разграничения видов-двойников, практически не отличающихся морфологически. Таких видов много среди малярийных комаров, дрозофил, сиговых рыб. Даже у птиц 5% видов-двойников, а одном ряду североамериканских сверчков их 17.
Использование одного только морфологического критерия может привести к ошибочным выводам. Так, К. Линней по особенности внешнего строения отнес самца и самку утки кряквы к разным видам. Сибирские охотники по окраске меха лисиц выделили пять вариаций: сиводушки, огневки, крестовки, черно-бурые и черные. В Англии у 70 видов бабочек, наряду с особями со светлой окраской, имеются и тем ные морфы, численность которых в популяциях стала увеличиваться в связи с загрязнением лесов. Полиморфизм — широко распространенное явление. Он встречается у всех видов. Затрагивает он и те признаки, по которым различаются виды. У жуков дровосеков, например у усача цве точного, встречающегося поздней весной на купальнице, помимо ти пичной формы в популяциях встречается до 100 аберраций окраски. Во времена Линнея морфологический критерий был главным, так как счи тали, что существует одна типичная для вида форма.
Сейчас, когда установлено, что у вида может быть много форм, типо логическая концепция вида отброшена и морфологический критерий не всегда удовлетворяет ученых. Все же следует признать, что этот критерий очень удобен для систематизации видов и в большинстве определителей животных и растений он играет главную роль.
Физиологический критерий
Физиологические особенности различных видов растений и живот ных часто являются фактором, обеспечивающим их генетическую само стоятельность. Например, у многих дрозофил сперма особей чужого ви да вызывает иммунологическую реакцию в половых путях самки, что приводит к гибели сперматозоидов. Гибридизация различных видов и подвидов козлов часто приводит к нарушению периодичности плодо ношения — приплод появляется зимой, что ведет к его гибели. Скрещи вание разных подвидов косуль, например сибирской и европейской, подчас приводит к гибели самок и потомства из-за крупных размеров плода.
Биохимический критерий
Интерес к этому критерию появился в последние десятилетия в связи с развитием биохимических исследований. Он не находит широкого применения , так как не существует каких-либо специфических веществ, характерных только для одного вида и, кроме того, он весьма трудоемкий и далеко не универсальный. Однако им можно воспользоваться в тех случаях, когда другие критерии «не работают». Например, для двух видов-двойников бабочек из рода амата (А. р h е g еа и А. г ugazzii ) диагностическим и признаками являются два фермента — фосфоглюкомутаза и эстераза-5, позволяющие даже определять гибридов этих двух видов. В последнее время широкое применение получило сравнительное изучение состава ДН К в практической таксономии микробов. Изучение состава ДНК позволило произвести ревизию филогенетической системы различных групп микроорганизмов. Разработанные методы дают возможность сравнивать состав ДНК у законсервированных в толщах земли бактерий и ныне живущих форм. Было проведено, например, сравнение состава ДНК у пролежавшей около 200 млн. лет в толще солей палеозойской бактерии псевдо монады солелюбивой и у ныне живущих псевдомонад. Состав их ДНК оказалдся идентичным, а биохимические свойства — сходными.
Цитологический критерий
Развитие цитологических методов позволило ученым исследовать фо рму и число хромосом у многих видов животных и растений. Появилось новое направление — кариосистематика, которое внесло некоторые поправки и уточнения в построенную на основе морфологических критериев филогенетическую систему. В ряде случаев число хромосом служит характерной особенностью вида. Кариологический анализ позволил , например, упорядочить систематику диких горных баранов, у кото рых разными исследователями выделялось от 1 до 17 видов. Анализ показал наличие трех кариотипов: 54-хромосомный — у муфлонов, 56-х ромосомный — у архаров и аргали и 58-хромосомный — у обитателей гор Средней Азии — уриалов.
Однако и этот критерий не является универсальным. Во-первых, у многих разных видов число хромосом одинаково и форма их сходна. Во-вторых, в пределах одного и того же вида могут встречаться особи с разным числом хромосом. Это так называемые хромосомный и геномный полиморфизм. Например, козья ива имеет диплоидное — 38 и тетраплоидное число хромосом — 76. У серебряного карася встречаются популяции с набо ром хромосом 100, 150, 200, тогда как нормальное число их равно 50. У радужной форели число хромосом варьирует от 58 до 64, у беломорской сель ди встречаются особи с 52 и 54 хромосомами. В Таджикистане на участке протяженностью всего 150 км, зоологами была обнаружена популяция слепушонки обыкновенной с набором хромосом от 31 до 54. У песчанок из разных мест обитания число хромосом различное: 40 — у песчанок алжир ских популяций, 52 — у израильских и 66 — у египетских. К настоящему времени внутривидовой хромосомный полиморфизм обнаружен у 5 % ц ито-генетически изученных видов млекопитающих.
Иногда этот критерий неверно трактуется как генетический. Безусловно , число и форма хромосом важный признак, препятствующий скрещи ванию особей разных видов. Однако это скорее цитоморфологичес кий критерий, так как речь идет о внутриклеточной морфологии: числе и форме хромосом, а не о наборе и структуре генов.
Э тологический критерий
Для некоторых видов животных механизмом, препятствующим с крещиванию и нивелированию различий между ними, являются осо бенности их поведения, особенно в брачный период. Узнавание партнера своего вида и отвергание попыток ухаживания самцов другого вида основаны на специфических раздражителях — зрительных, звуковых химических, тактильных, механических и др.
В широко распространенном роде пеночки разные виды очень похо жи друг на друга морфологически, в природе их не различить ни по окраске, ни по размерам. Зато все они очень хорошо различаются по песне и по повадкам. Песня пеночки веснички сложная, похожа на песнь зяблика , только без его завершающего колена, а песня теньковки — про стенькие монотонные посвисты. Многочисленные виды-двойники аме риканских светлячков из рода Р hotinus были впервые опознаны лишь по различиям в их световых сигналах. Самцы светлячков в полёте производят вспышки света, частота, продолжительность и чередуемость которых специфичны для каждого вида . Хорошо известно, что ряд видов прямокрылых и равнокрылых, обитающих в пределах, одного биотопа и размножающихся синхронно, различаются лишь характеру их призывных сигналов. Такие виды-двойники с акустической репродуктивной изоляцией обнаружены, например, у сверчков, кобылок-коньков, цикад и других насекомых. Два близких вида американских жабне скрещиваются также из-за различий в зове самцов.
Решающую роль в репродуктивной изоляции часто играют различия в демонстративном поведении. Например, близкие виды мух дрозофил от личаются спецификой ритуала ухаживания (по характерувибрации крыльев, дрожания лапок, кружения, тактильных контактов). Два близких вида — серебристая чайка и клуша имеют различия в степени выраженно сти демонстративных поз, а семь видов ящериц рода S се1ороги s различаются степенью поднятия головы при ухаживании половых партнеров.
Экологический критерий
Особенности поведения подчас тесно связаны с экологической спецификой вида, например, с особенностями устройства гнезда. Три вида наших обычных синиц гнездятся в дуплах лиственных деревьев, преимущественно берез. Большая синица на Урале выбирает обычно глубокое дупло в нижней части ствола березы или ольхи, образовавшееся в ре зультате выгнивания сучка и смежной с ним древесины. Это дупло недоступно ни дятлам, ни воронам, ни хищным млекопитающим. Синица московка заселяет морозобойные трещины в стволах березы и ольхи. Га ичка же предпочитает строить дупло сама, выщипывая полости в трухля вых или старых стволах березы и ольхи, и без этой трудоемкой процедуры она не отложит яиц.
Особенности образа жизни, присущие каждому виду, определяют его положение, его роль в биогеоценозе, то есть его экологическую нишу. Даже самые близкие виды, как правило, занимают разные экониши, то есть различаются хотя бы по одному-двум экологическим признакам.
Так, экониши всех наших видов дятлов различаются по характеру питания. Большой пестрый дятел зимой питается семенами лиственницы и сосны, раздалбливая шишки в своих «кузницах». Черный дятел желна добывает личинок усачей и златок из-под коры и из древесины елей, а малый пестрый дятел долбит мягкую древесину ольхи либо добывает насекомых из стеблей травянистых растений.
Каждый из 14 видов дарвиновых вьюрков (названных так в честь Ч. Дарвина, впервые обратившего на них внимание), обитающих на Галапагосских островах, имеет свою специфическую эконишу, отличающуюся от других прежде всего характером корма и путями его добывания.
Ни экологический, ни рассмотренный выше этологический крите рии не являются универсальными. Очень часто особи одного вида, но раз ных популяций отличаются целым рядом особенностей образа жизни и поведения. И напротив, разные виды, даже очень далекие, в системат ическом отношении, могут иметь сходные этологические признаки или играть одинаковую роль в сообществе (например, роли травоядных млекопитающих и насекомых, скажем, таких, как саранча, вполне сопоставимы).
Географический критерий
Этот критерий наряду с экологическим в большинстве определителей занимает второе (после морфологического) место. При определении многих видов растений, насекомых, птиц, млекопитающих и других групп организмов, распространение которых хорошо изучено, зна ние ареала играет существенную роль. У подвидов ареалы, как правило не совпадают, что обеспечивает их репродуктивную изоляцию и, по сути , их существование в качестве самостоятельных подвидов. Многие виды занимают разные ареалы (такие виды называют аллопатрическими). Но огромное число видов имеют совпадающие или перекрываю щиеся ареалы (симпатрические виды). Кроме того, существуют виды, не имеющие четких границ распространения, а также виды-кос мополиты, обитающие на огромных пространствах суши или океана. В силу этих обстоятельств географический критерий не может быть у ниверсальным.
Генетический критерий
Генетическое единство вида и, соответственно, генетическая изоляция его от других видов — главный критерий вида, основной видовой признак, обусловленный комплексом особенностей строения и жизне деятельности организмов данного вида.Генетическая совмести мость, сходство морфологических, физиологических, цитологических и других признаков, одинаковое поведение, совместное обитание — все эт о создает необходимые условия для успешного размножения и вос производства вида. В то же время все эти признаки обеспечивают генетическую изоляцию вида от других сходных с ним видов. Например, раз личия в песне дроздов, пеночек, славок, зябликов и вьюрка, глухой и обыкновенной кукушек препятствуют образованию смешанных пар, несмотря на сходство их окраски и экологии (у птиц, обладающих специфической песней, почти не встречаются гибриды). Даже в тех случа ях, когда, несмотря на изоляционные барьеры, произошло скрещива ние особей разных видов, гибридная популяция, как правило, не возникает, так как начинает действовать целый ряд постпопуляционных механизмов изоляции. Важнейшие из них — это гибель мужских гамет (генет ическая несовместимость), гибель зигот, нежизнеспособность гиб ридов, их стерильность, наконец, невозможность найти полового партнера и дать жизнеспособное плодовитое потомство. Мы знаем, что для каждого вида характерен свой набор специфических признаков. Межвидовой гибрид будет иметь признаки, промежуточные между признаками двух исходных родительских форм. Его песня, к примеру не будет понята ни зябликом, ни вьюрком, если это гибрид этих видов, и он не найдет полового партнера. У такого гибрида при обра зовании гамет содержащиеся в его клетках хромосомы зяблика «не уз нают» хромосомы вьюрка и, не найдя гомологичного партнера, не конъюгируют. В результате образуются гаметы с нарушенным набором хромосом, которые, как правило, нежизнеспособны. И вследствие го этого гибрид окажется стерильным.
Ворон и ворона
Скажу сразу: Ворон — это не «муж» вороны, а самостоятельный вид.
Ворон — один из наиболее крупных представителей семейства вороновые , весит от 0,8 до 1,5 кг. Окраска оперения, клюва и ног у него однотонного черного цвета.
Распространен ворон почти по всему северному полушарию: встречается почти по всей Европе, Азии, исключая Юго-Восточную, в Северной Африке и Северной Америке. Повсеместно он ведет оседлый образ жизни. Населяет леса, пустыни и горы. В безлесных местностях держится у скал, береговых обрывов речных долин. Спаривание и брачные игры на юге страны отмечаются в первой половине февраля, на севере — в марте. Пары постоянны. Гнезда обычно помещаются на вершинах высоких деревьев. В кладке от 3 до 7, чаще 4-6, яиц голубовато-зеленой окрас ки с темными отметинами.
Ворон — всеядная птица. Основной его корм — падаль, которую он чаще всего находит на свалках и у скотобоен. Поедая падаль, он выступает как санитарная птица. Кормится он также грызунами, яйцами, и птенцами, рыбой, различными беспозвоночными животными, а места ми и зёрнами хлебных злаков.
Ворона по общему телосложению напоминает ворона, но значительно мельче его: весит от 460 до 690 г.
Описываемый вид интересен тем, что по окраске оперения распадается на две группы: серую и черную. Серая ворона имеет хорошо извест ную двуцветную окраску: голова, горло, крылья, хвост, клюв и ноги черные, остальное оперение серое. Черная ворона вся черная, с металлическим синим и пурпуровым блеском.
Каждая из этих групп имеет локальное распространение. Серая ворона широко распространена в Европе, Западной Азии, черная – в Центральной и Западной Европе, с одной стороны, в Средней, Восточной Азии и Северной Америке – с другой.
Населяет ворона опушки и окраины лесов, сады, рощи, заросли речных долин, реже скалы и склоны береговых обрывов. Это частично осёдлая, частично перелётная птица.
В начале марта в южных частях страны и в апреле-мае в северных и восточных начинается откладывание яиц. В кладке обычно 4-5 яиц бледно-зелёной, голубовато-зелёной или частично зелёной окраски с тёмными пятнышками и крапинками. Ворона – всеядная птица. Из животных она поедает различных беспозвоночных – жуков, муравьев, моллюсков, а также грызунов, ящериц, лягушек и рыб. Из растений клюёт зёрна культурных злаков, семена ели, вьюнка полевого, птичьей гречишки и др. Зимой питается в основном отбросами.
Заяц-беляк и заяц-русак
Род собственно зайцев, к которому относятся русак и беляк, а также еще 28 видов, довольно многочислен. Наиболее известные в России зайцы — беляк и русак . Беляка можно встретить на территории от побережья Северного Ледовитого океана до южной границы лесной зоны, в Сибири — до границ с Казахста ном, Китаем и Монголией, а на Дальнем Востоке — от Чукотки до и Северной Кореи. Распространен беляк и в лесах Европы, а также на востоке Северной Америки. Русак обитает на территории Европейской России от Карелии юга Архангельской области до южных границ страны, на Украине и в Закав казье. А вот в Сибири этот заяц обитает только на юге и к западу от Байкала.
Беляк получил свое название благодаря белоснежному зимнему меху. Только кончики ушей остаются у него черными весь год. Русак же в некоторых северных местностях тоже сильно светлеет к зиме, но снежно-белым он не бывает никогда. А на юге он вовсе не меняет окраски.
Русак больше приспособлен к жизни в открытых ландшафтах, поскольку он крупнее беляка, да и бегает лучше. На коротких дистанциях этот заяц может развивать скорость до 50 км/час. У беляка лапы широкие, с густым опушением чтобы меньше проваливаться в рыхлые лесные сугробы. А у русака лапы уже, ведь на открытых местах снег, как правило, твердый, слежавшийся, «утоптанный ветром».
Длина тела беляка — 45-75 см, масса — 2,5-5,5 кг. Уши короче, чем у русака. Длина тела русака — 50-70 см, масса до 5 (иногда 7) кг.
Размножаются зайцы обычно два, а на юге три или даже четыре раза в год. У зай цев-беляков в выводе может быть по два, три пять, семь зайчат, а у русаков — обычно всего один или два зайчонка. Русаки начинают пробовать траву через две недели после рождения, а беляки и того быстрее — через неделю.
Экологическая ниша – обзор
Водные экологические ниши
Экологические ниши встречаются во всех типах экосистем. На уровне понятия или определения нет различия между водными и наземными нишами. Тем не менее, водная среда отличается тем, что некоторые (но не все) оси ниш, которые, скорее всего, будут важными, отличаются от осей наземной среды. Важные параметры экологической ниши для водных организмов включают температуру, растворенный кислород, структуру среды обитания, хищничество и питательные вещества для растений.
Диапазон температур для водных экосистем намного уже, чем диапазон для наземных экосистем, поскольку жидкая вода имеет минимальную температуру 0 °C. Термические пороги слабы для фитопланктона и зоопланктона, о чем свидетельствует распределение видов в широком диапазоне широт и высот. Термические пороги более важны для крупных беспозвоночных и особенно для рыб. Например, семейство Salmonidae (лосось и форель) содержит много видов, не переносящих воду с температурой выше 15–20 °C.Точно так же вечно прохладные воды, такие как горные или субарктические озера, не могут поддерживать популяции многих видов тепловодных рыб, таких как большинство видов семейства солнечных рыб (Centrarchidae).
Вода удерживает примерно 10 мг/л кислорода при низких температурах и 6–7 мг/л при высоких температурах. Таким образом, дыхание может сделать воду бескислородной, если оно не компенсируется кислородом, образующимся в результате фотосинтеза, или контактом воды с атмосферой. Например, высокая скорость дыхания в воде, находящейся в контакте с отложениями, может удалить весь кислород из воды за несколько дней.
Некоторые водные среды гораздо больше подвержены истощению кислорода, чем другие. Горные потоки с большим уклоном и низким уровнем дыхания находятся на одном конце спектра, поскольку они имеют очень небольшой потенциал для истощения кислорода, тогда как плодородные озера или водно-болотные угодья, где много дыхания и менее эффективный газообмен с атмосферой, демонстрируют гораздо более высокая вероятность истощения кислорода.
Распространение организмов частично отражает их способность переносить кислородное голодание.Рыбы, например, демонстрируют широкий диапазон толерантности к истощению кислорода. На одном полюсе находятся рыбы, которые могут получать кислород из атмосферы (например, тропические лабиринтные рыбы, такие как бойцовая петушка, Betta ), и рыбы с маленьким перевернутым ртом, способным втягивать кислород из верхних 1–2 мм воды. при контакте с атмосферой (карповые рыбы, включая рыбу-комара, Gambusia ). Промежуточными по толерантности являются рыбы, не имеющие специальных способов получения кислорода, но обладающие высокой физиологической устойчивостью к кислородному голоданию (некоторые сомы, например бычок, италурид, Ameiurus , и сазан, карповый, Cyprinus ). .Напротив, другие рыбы умеренно или сильно чувствительны к истощению кислорода: форель и лосось, а также большеротый окунь ( Micropterus ), синежаберный ( Lepomis ) и другие солнечные рыбы, и многие другие. Таким образом, ихтиофауна конкретного водоема отражает вероятность кислородного голодания. Беспозвоночные следуют аналогичным схемам. Водные личинки видов мошек, например, различаются по толерантности к кислороду, что объясняет их контрастное распределение в водной среде.
Структура среды обитания (покрытие) является постоянным компонентом большинства земных сред; это в значительной степени обеспечивается растительностью, которая появляется на всех поверхностях, кроме самых сухих или на ледяных щитах. Напротив, пелагическая (открытая вода) среда озер не имеет структуры. Таким образом, организмы, живущие в пелагических зонах, приспособлены для жизни в среде, лишенной пространственной стабильности и не предлагающей убежища от хищников. Различия в покрытии, начиная от открытых вод озер и ручьев с песчаным дном и заканчивая богатой растительностью прибрежной зоной, заболоченными местами с растительностью и каменистыми ручьями, предлагают очень широкий спектр возможностей для осей ниш, связанных со средой обитания.
Лимнологи и водные экологи неоднократно показывали, что хищничество оказывает сильное влияние на видовой состав внутренних вод. Высшие хищники особенно важны тем, что они часто удаляют или сильно подавляют численность своей добычи. В частности, визуальные хищники озер могут гораздо эффективнее уничтожать свою добычу, чем в большинстве других сред. Фактически, некоторые крупные беспозвоночные не могут обитать в водных экосистемах, содержащих рыб.Небольшие безрыбные озера, например, содержат большое количество крупных беспозвоночных, которых нет в аналогичных озерах, зарыбленных рыбой. Таким образом, размер ниши, связанный с хищничеством, имеет решающее значение для распространения многих беспозвоночных.
Водная среда обладает исключительным потенциалом для производства автотрофной биомассы. Воды с очень низким содержанием азота и фосфора обладают производственным потенциалом, который намного ниже, чем у любой почвенной среды, где присутствует влага.Напротив, водные экосистемы с высокой концентрацией фосфора и азота имеют продуктивный потенциал для автотрофов, который может быть в 1000 раз выше, чем наименее продуктивные воды. Конкуренция за питательные вещества имеет первостепенное значение в непродуктивных водах, содержащих виды с очень высоким сродством к фосфору и азоту. Наиболее продуктивные водные среды также поддерживают специализированные таксоны, но с различными видами адаптации. Баланс питательных веществ также может быть важным фактором, определяющим нишу.Например, азотфиксирующие сине-зеленые водоросли (гетероцистные цианобактерии), в отличие от других водорослей, способны превращать газообразный N2 в аммиак, который можно использовать для получения аминокислот. Когда неорганического азота не хватает, эти организмы способны продолжать рост, в то время как другие автотрофы не могут.
3 Стрессоры: факторы окружающей среды и их влияние на экосистему залива и дельты | Устойчивое управление водными ресурсами и окружающей средой в дельте Калифорнийского залива
Линдли, С.Т., Р. Шик, А. Агравал, М. Гослин, Т. Пирсон, Э. Мора, Дж. Дж. Андерсон, Б. Мэй, С. Грин, К. Хэнсон, А. Лоу, Д. Макьюэн, Р. Б. Макфарлейн, К. , Суонсон и Дж. Г. Уильямс. 2006. Историческая структура населения стальной головы Центральной долины и ее изменение плотинами. Наука об устье и водоразделе Сан-Франциско 4 (1): статья 3, 1–19. Доступно по адресу http://repositories.cdlib.org/jmie/sfews/vol4/iss1/art3. По состоянию на 17 июля 2012 г.
Линдли, С. Т., Р. С. Шик, Э. Мора, П. Б. Адамс, Дж. Дж.Андерсон, С. Грин, К. Хэнсон, Б. П. Мэй, Д. Макьюэн, Р. Б. Макфарлейн, К. Суонсон и Дж. Г. Уильямс. 2007. Схема оценки жизнеспособности находящихся под угрозой исчезновения чавычи и стальноголового лосося в бассейне Сакраменто-Сан-Хоакин. Устье Сан-Франциско и наука о водоразделах 5 (1). Доступно по адресу http://escholarship.org/uc/item/3653x9xc#page-2. По состоянию на 25 июля 2012 г.
Линдли, С. Т., М. Л. Мозер, Д. Л. Эриксон, М. Белчик, Д. В. Уэлч, Э. Л. Речиски, Дж. Т. Келли, Дж. Хьюблейн и А.П. Климли. 2008. Морская миграция североамериканского зеленого осетра. Труды Американского общества рыболовства 137: 182-194.
Линдли, С.Т., С.Б. Граймс, М.С. Мор, В. Петерсон, Дж. Стейн, Дж.Р. Андерсон, Л.В. Ботсфорд, Д.Л. Боттон, К.А. Басак, Т.К. Коллиер, Дж. Фергюсон, Дж.К. Гарза, А.М. Гровер, Д.Г. П. В. Лоусон, А. Лоу, Р. Б. Макфарлейн, К. Мур, М. Пламер-Цвален, Ф. Б. Швинг, Дж. Смит, К. Трейси, Р. Уэбб, Б. К. Уэллс и Т. Х. Уильямс. 2009.Что вызвало обвал акций чавычи из-за падения реки Сакраменто? Национальная служба морского рыболовства, Юго-западный научный центр рыболовства. NOAA_TM_SWFSC-447. Доступно на http://swr.nmfs.noaa.gov/media/salmondeclinereport.pdf. По состоянию на 17 июля 2012 г.
Lluch-Coat, S.E., E.N. Curchitser, and S.Ito. 2010 г. Сендайский семинар по совмещению моделей «климат-рыба-рыболовство», 2010 г. Информационный бюллетень Северо-Тихоокеанской организации морских наук 18 (2): 20-21.
Лодж, Д. М., С. Уильямс, Х.Дж. МакИсаак, К. Р. Хейс, Б. Леунг, С. Райхард, Р. Н. Мак, П. Б. Мойл, М. Смит, Д. А. Андоу, Дж. Т. Карлтон и А. МакМайкл. 2006. Биологические вторжения: рекомендации для политики и управления США. Экологические приложения 16:2035-2054.
Лукас, Л. 2009. Влияние эстуарного переноса на качество воды. Стр. 273–307 в « Современные проблемы физики эстуариев» под редакцией А. Велле-Левинсона. Кембридж: Издательство Кембриджского университета.
Лукас, Л. В., Дж. Э. Клерн, Дж.Р. Косефф, С. Г. Монисмит и Дж. К. Томпсон. 1998. Объясняет ли модель критической глубины Свердрупа динамику цветения в эстуариях? Журнал морских исследований 56 (2): 375-415.
Лукас, Л. В., Дж. Э. Клерн, Дж. К. Томпсон и Н. Э. Монсен. 2002. Функциональная изменчивость мест обитания в дельте Сакраменто-Сан-Хоакин: последствия восстановления. Экологические приложения. 12: 1528-1547.
Лукас, Л., Дж. Р. Косефф, С. Г. Монисмит и Дж. К. Томпсон. 2009. Процессы на мелководье определяют динамику цветения фитопланктона: исследование моделирования.Дж. Мар Системы 75:70-86.
Луома, С. Н. и Дж. Э. Клерн. 1982. Воздействие сброса сточных вод на биологические сообщества в заливе Сан-Франциско. Стр. 137–160 в заливе Сан-Франциско: использование и защита, под редакцией В. Дж. Коклемана. Тихоокеанское отделение AAAS. Доступно по адресу http://sfbay.wr.usgs.gov/publications/pdf/luoma_1982_discharge.pdf. По состоянию на 25 июля 2012 г.
Луома, С. Н. и Д. Дж. Х. Филлипс. 1988. Распространение, изменчивость и воздействие микроэлементов в заливе Сан-Франциско.Бюллетень о загрязнении морской среды 19:413-425.
Луома, С. Н. и П. С. Радуга. 2008. Загрязнение металлами в водной среде: наука и боковое управление. Кембридж: Издательство Кембриджского университета.
Мак Налли, Р., Дж. Р. Томсон, У. Дж. Киммерер, Ф. Фейрер, К. Б. Ньюман, А. Сих, В. А. Беннетт, Л. Браун, Э. Флейшман, С. Д. Калберсон и Г. Кастильо. 2010. Анализ сокращения пелагических видов в верхнем эстуарии Сан-Франциско с использованием многомерного авторегрессионного моделирования (MAR).Экологические приложения 20:1417-1430.
Стол NAAQS | Агентство по охране окружающей среды США
Закон о чистом воздухе, в который в последний раз вносились поправки в 1990 году, требует, чтобы Управление по охране окружающей среды установило национальные стандарты качества атмосферного воздуха (40 CFR часть 50) для шести основных загрязнителей («критериальные» загрязнители воздуха), которые могут быть вредными для здоровья населения и окружающей среды. Закон о чистом воздухе определяет два типа национальных стандартов качества окружающего воздуха. Первичные стандарты обеспечивают защиту здоровья населения, включая защиту здоровья «чувствительных» групп населения, таких как астматики, дети и пожилые люди. Второстепенные стандарты обеспечивают защиту общественного благосостояния, включая защиту от ухудшения видимости и повреждения животных, сельскохозяйственных культур, растительности и зданий.
Периодически стандарты пересматриваются и иногда могут пересматриваться, устанавливая новые стандарты. Ниже перечислены самые последние установленные стандарты. В некоторых регионах США также могут сохраняться определенные нормативные требования для внедрения ранее установленных стандартов.
Единицами измерения для стандартов являются объемные части на миллион (ppm), объемные части на миллиард (ppb) и микрограммы на кубический метр воздуха (мкг/м 3 ).
| Загрязнитель [ссылки на исторические таблицы обзоров NAAQS] |
Первичный/ Вторичный |
Время усреднения | Уровень | Форма | |
|---|---|---|---|---|---|
| Окись углерода (СО) | первичный | 8 часов | 9 частей на миллион | Не должен превышаться более одного раза в год | |
| 1 час | 35 частей на миллион | ||||
| Свинец (Pb) | первичный и вторичный |
Скользящая средняя за 3 месяца | 0.15 мкг/м 3 (1) | Не должен превышать | |
| Двуокись азота (NO 2 ) | первичный | 1 час | 100 частей на миллиард | 98-й процентиль 1-часовых максимальных суточных концентраций, усредненных за 3 года | |
| первичный и вторичный |
1 год | 53 частей на миллиард (2) | Среднее годовое значение | ||
| Озон (O 3 ) | первичный и вторичный |
8 часов | 0.070 частей на миллион (3) | Годовая четвертая по величине суточная максимальная 8-часовая концентрация, усредненная за 3 года | |
| Загрязнение твердыми частицами (PM) | ПМ 2,5 | первичный | 1 год | 12,0 мкг/м 3 | среднегодовое, усредненное за 3 года |
| вторичный | 1 год | 15,0 мкг/м 3 | среднегодовое, усредненное за 3 года | ||
| первичный и вторичный |
24 часа | 35 мкг/м 3 | 98-й процентиль, усредненный за 3 года | ||
| ПМ 10 | первичный и вторичный |
24 часа | 150 мкг/м 3 | Не более одного раза в год в среднем за 3 года | |
| Двуокись серы (SO 2 ) | первичный | 1 час | 75 частей на миллиард (4) | 99-й процентиль 1-часовых максимальных суточных концентраций, усредненных за 3 года | |
| вторичный | 3 часа | 0.5 частей на миллион | Не должен превышаться более одного раза в год | ||
(1) В областях, в которых до обнародования текущих (2008 г.) стандартов не были достигнуты стандарты Pb, и для которых не были представлены и утверждены планы реализации для достижения или сохранения текущих (2008 г.) стандартов, прежние нормы (1,5 мкг/м3 в среднем за календарный квартал) также остаются в силе.
(2) Уровень годового стандарта № 2 равен 0.053 промилле. Он показан здесь в единицах ppb для более четкого сравнения со стандартным уровнем за 1 час.
(3) Окончательное правило подписано 1 октября 2015 г. и вступает в силу 28 декабря 2015 г. Предыдущие (2008 г.) стандарты O 3 не отменяются и остаются в силе для определенных областей. Кроме того, в некоторых областях могут действовать определенные постоянные обязательства по внедрению в соответствии с ранее отмененными стандартами 1 часа (1979 г.) и 8 часов (1997 г.) O 3 .
(4) Предыдущие стандарты SO 2 (0.14 частей на миллион в сутки и 0,03 частей на миллион в год) дополнительно останутся в силе в определенных областях: (1) любая область, для которой еще не прошел 1 год с даты вступления в силу действующих стандартов (2010 г.), и (2) любая область, для которой план реализации, предусматривающий достижение текущего (2010 г.) стандарта, не был представлен и утвержден и который обозначен как несоответствующий в соответствии с предыдущими стандартами SO 2 или не соответствует требованиям вызова SIP в соответствии с предыдущим SO 2 стандартов (40 CFR 50.4(3)). SIP-вызов — это действие EPA, требующее от штата повторно представить весь или часть своего Плана реализации штата, чтобы продемонстрировать достижение требуемого NAAQS.
Меню мер контроля для реализации NAAQS
Меню мер контроля (MCM) предоставляет государственным, местным и местным авиационным агентствам информацию о существующих мерах по сокращению выбросов, а также соответствующую информацию об эффективности и рентабельности мер. Государственные, местные и племенные агентства смогут использовать эту информацию при разработке стратегий, планов и программ по сокращению выбросов, чтобы обеспечить соблюдение и соблюдение национальных стандартов качества атмосферного воздуха (NAAQS).MCM — это «живой» документ, который может обновляться новыми или более актуальными данными по мере их появления.
исчезающих видов | Что мы делаем | Список и критическая среда обитания
Самец очковой гаги, находящийся под угрозой исчезновения ( Somateria fischeri ), Центр морской жизни Аляски, Сьюард, Аляска.
Фото: Лаура Л. Уайтхаус/USFWS
Прежде чем вид сможет получить защиту, предусмотренную Законом об исчезающих видах (ESA), он должен быть сначала добавлен в федеральные списки исчезающих и находящихся под угрозой исчезновения диких животных и растений.Список исчезающих и находящихся под угрозой исчезновения диких животных (50 CFR 17.11) и Список находящихся под угрозой исчезновения и находящихся под угрозой исчезновения растений (50 CFR 17.12) содержат названия всех видов, которые были определены Службой рыболовства и дикой природы США (Службой) или Национальным морским рыболовством. Служба (для большинства морских обитателей) больше всего нуждается в федеральной защите.
Вид добавляется в список, когда определяется, что он находится под угрозой исчезновения из-за любого из следующих факторов:
- существующее или угрожающее уничтожение, изменение или сокращение его среды обитания или ареала;
- чрезмерное использование в коммерческих, развлекательных, научных или образовательных целях;
- болезнь или хищничество;
- неадекватность существующих механизмов регулирования; или
- других природных или техногенных факторов, влияющих на его выживаемость.
Существует два способа включения вида в список ESA: через процесс подачи петиции или через процесс оценки кандидата. ESA предусматривает, что любое заинтересованное лицо может обратиться к министру внутренних дел или министру торговли большинства морских обитателей с просьбой добавить вид в списки исчезающих и находящихся под угрозой исчезновения видов или удалить их из них. В процессе оценки кандидатов биологи Службы определяют виды в качестве кандидатов для включения в список.
Для получения подробной информации загрузите информационный бюллетень [328 КБ].
Узнайте больше о нашем национальном рабочем плане по рассмотрению вопросов, связанных с внесением в список ЕКА, и критически важными решениями по среде обитания.
Узнайте больше о методологии, которую мы используем для определения приоритетов нашей работы по листингу.
Что значит быть в списке?
Виды, соответствующие определениям ESA исчезающих видов или видов, находящихся под угрозой исчезновения, добавляются в списки находящихся под угрозой исчезновения диких животных или растений и имеют право на определение критической среды обитания.Среди природоохранных преимуществ, разрешенных для находящихся под угрозой исчезновения растений и животных, которые перечислены в соответствии с ESA, являются: защита от опасности в результате федеральной деятельности; защита критически важной среды обитания, которая разрушается или подвергается неблагоприятным изменениям; ограничения на получение и торговлю; требование о том, чтобы Служба разрабатывала и реализовывала планы восстановления перечисленных видов под юрисдикцией США; разрешение на покупку земли или обмен на важную среду обитания; и федеральная помощь департаментам охраны природы штата и Содружества в рамках совместных соглашений об исчезающих видах.Внесение в список также способствует большему признанию ненадежного статуса вида, поощряя усилия по сохранению со стороны других агентств (иностранных, федеральных, государственных и местных), независимых организаций и заинтересованных лиц.
Политика Конгресса, изложенная в разделе 2 ESA, заключается в том, что все федеральные агентства должны стремиться сохранять исчезающие и находящиеся под угрозой исчезновения виды и должны использовать свои полномочия для достижения целей ESA. Точно так же раздел 7 ESA предписывает федеральным агентствам использовать свои законные полномочия для выполнения программ сохранения перечисленных видов.Он также требует, чтобы эти агентства гарантировали, что любые действия, которые они финансируют, санкционируют или осуществляют, не могут поставить под угрозу выживание любого исчезающего или исчезающего вида или уничтожить или неблагоприятно изменить его обозначенную критическую среду обитания (если таковая имеется). Когда агентство обнаруживает, что одно из его действий может затронуть внесенные в список виды, оно должно проконсультироваться со Службой, чтобы избежать опасности для вида или разрушения или неблагоприятного изменения обозначенной критической среды обитания. При необходимости предлагаются «разумные и разумные альтернативы», такие как изменение проекта или изменение графика, чтобы позволить завершить предлагаемую деятельность, сохраняя при этом вид или критически важную среду обитания.В тех случаях, когда федеральное действие может поставить под угрозу выживание вида, предложенного для включения в список, но еще не доработанного и не добавленного в список, федеральное агентство обязано «посоветоваться» со Службой (хотя рекомендации, вытекающие из такой конференции, не имеют юридической силы).
Дополнительная защита разрешена разделом 9 ESA, который объявляет незаконным вывоз, импорт, экспорт или участие в межгосударственной или международной торговле внесенными в список животными, за исключением случаев, когда имеется разрешение для определенных целей сохранения.ESA также объявляет незаконным владение, продажу или транспортировку любых перечисленных видов, изъятых в нарушение закона. Для растений торговые ограничения такие же, но правила «приобретения» другие. Сбор или злонамеренное повреждение любого исчезающего растения на землях, находящихся под федеральной юрисдикцией, является незаконным. Удаление или повреждение внесенных в список растений на государственных и частных землях при сознательном нарушении законодательства штата или в ходе нарушения закона штата об уголовном посягательстве также является незаконным в соответствии с ESA. Кроме того, в некоторых штатах действуют более строгие законы, прямо запрещающие вывоз растений и животных, внесенных в список штата или федерального уровня.
Просмотрите или загрузите последние объявления о листинге или объявления о действиях в Федеральном реестре .
Узнайте больше о сохранении кандидатов.
Используйте наши инструменты поиска видов, чтобы узнать, сколько и какие виды перечислены, где они перечислены, получить дополнительную информацию об отдельных видах, информацию о предлагаемых видах и видах-кандидатах или другую информацию о видах, находящихся под угрозой исчезновения.
Прочтите о критической среде обитания, необходимой для сохранения вида, и о том, как она связана с процессом внесения в список.
Узнайте о правилах защиты, изданных в соответствии с разделом 4 (d) ESA для видов, находящихся под угрозой исчезновения.
Доступ к полной библиотеке документов, связанных с листингом.
Ознакомьтесь с политикой службы в отношении листинга.
границ | Распределение видов растений по градиентам окружающей среды: имеют ли значение подземные взаимодействия с грибами?
Влияние микоризных грибов на распространение видов растений: обзор данных
Виды растений не распределены в ландшафте случайным образом по градиентам окружающей среды (Pottier et al., 2013), но проявляют видовую толерантность, формируя неслучайные сообщества (Киквидзе и др., 2005). Среди абиотических факторов признано, что свойства климата и почвы сильно ограничивают распространение видов (например, Dubuis et al., 2013). Виды не являются независимыми субъектами в экосистеме, но взаимодействуют экологически позитивно (например, содействие, мутуализм) и негативно (например, конкурентное исключение; Pellissier et al., 2012). Таким образом, поиск принципов, объясняющих локальное объединение сообществ, должен интегрировать биотические процессы (Guisan, Rahbek, 2011; Weiher et al., 2011). Точно так же, как абиотические переменные определяют распространение видов, биотические взаимодействия ограничивают ареалы отдельных видов и, таким образом, пространственную изменчивость видовых ассоциаций (Wisz et al., 2013). Грибы проявляют мутуалистические и антагонистические взаимодействия с растениями и влияют на рост растений (Smith and Read, 2008). Поскольку распределение грибов и состав сообщества также формируются в соответствии с абиотическими градиентами (Opik et al., 2010; Tedersoo et al., 2012), мы ожидаем, что в дополнение к климатическим и почвенным абиотическим факторам сдвиги в структуре грибных сообществ вдоль экологических градиентов должны способствовать дальнейшему развитию. ограничивают распространение видов растений.
Большинство растений вступают в микоризные симбиозы с грибами. Некоторые из этих симбиотических грибов, арбускулярные микоризные грибы (AMF), являются облигатными биотрофами, которым требуется растение-хозяин для завершения своего жизненного цикла (Smith and Read, 2008). Другие, такие как эктомикоризные грибы (ECM), в меньшей степени зависят от растения-хозяина. В этих ассоциациях гриб снабжает растение неорганическими питательными веществами, такими как нитраты и фосфаты, а растение обеспечивает грибных партнеров фотосинтатами (Li et al., 2006). Симбиозы растений и грибов влияют на различные ключевые аспекты жизни растений, такие как клональное размножение (Streitwolf-Engel et al., 2001), рост и репродуктивный успех (Sanders, 2010). Кроме того, симбиозы с AMF значительно повышают устойчивость растения-хозяина к абиотическим стрессам, таким как засуха и засоление, а также к биотическим стрессам, включая надземные и подземные патогены и паразиты (Whipps, 2004; Jung et al., 2012). Кроме того, было показано, что АМФ положительно влияет на ассоциацию растения с азотфиксирующими или фосфатсолюбилизирующими бактериями (Jung et al., 2012). Присутствие видов грибов ECM может влиять на приспособленность растений, регулируя содержание нитратов в почве. Действительно, эти виды грибов обладают способностью иммобилизовать нитраты в своей цитоплазме и доставлять их партнерам по растению таким образом, который не зависит от доступных уровней нитратов в почве. Учитывая, как микориза влияет на отдельные растения, ожидается, что структура сообщества микоризных грибов будет влиять как на конкурентоспособность растения, так и на устойчивость к абиотическим условиям, и, следовательно, на состав растительного сообщества (van der Heijden et al., 1998; ван дер Хейден и Сандерс, 2002).
Влияние микоризы на растительные сообщества варьируется от изменения относительной численности видов до изменения состава и разнообразия растительного сообщества (Grime et al., 1987; Hartnett and Wilson, 1999; van der Heijden, 2003). Микоризный симбиоз также может изменять структуру растительных сообществ, сдвигая конкурентный баланс между видами растений внутри сообществ и предоставляя относительные преимущества видам растений, которые в противном случае могли бы быть низшими конкурентами (Moora and Zobel, 2010).В частности, влияние арбускулярной микоризы на растения может быть особенно выражено на стадии укоренения всходов (Francis, Read, 1995), что имеет решающее значение для формирования локального видового состава и разнообразия растительных сообществ (Dickson, Foster, 2008; Koorem et al. ., 2012). Например, Курем и др. (2012) предполагают, что микоризация Oxalis acetosella и Prunella vulgaris увеличивает их способность приживаться в условиях низкого плодородия почвы. Другие попытки оценить взаимодействие между сообществами грибов и растений были предприняты с использованием обработки фунгицидами в экспериментах и документально подтвержденных изменений в составе сообщества после обработки (Hartnett and Wilson, 1999; Liu et al., 2012). Хотя потенциальная роль микоризы в структурировании растительных сообществ обсуждается на протяжении десятилетий (Zobel et al., 1997), доказательств того, что эта роль выходит за рамки сообщества, остается мало (Klironomos et al., 2011).
Ожидается, что почвенные грибы, и особенно микоризные грибы, будут играть ключевую роль в определении крупномасштабного распространения видов растений (Fitter et al., 2005; Rosendahl, 2008; van der Heijden et al., 2008). Неспособность многих эктомикоризных растений прижиться в отсутствие внеклеточного матрикса позволяет предположить, что влияние внеклеточного матрикса на распространение и сообщества растений, вероятно, распространяется на большие пространственные масштабы (Klironomos et al., 2011). Недавние исследования показывают, что богатство грибов ECM связано с температурой и осадками в глобальном масштабе, достигая пика в биомах умеренных и бореальных лесов (Tedersoo et al., 2012). Это может ограничивать глобальное распространение растений, связанных с грибами ECM. Тем не менее, если распределение видов растений по градиентам среды относительно хорошо описано, то биогеография микроорганизмов, как и почвенных грибов, находится в зачаточном состоянии. Хотя были достигнуты некоторые успехи (т.г., Мура и др., 2004, 2011; Фиттер и др., 2005 г.; Треседер и Кросс, 2006 г.; Чаудхари и др., 2008 г.; Кивлин и др., 2011; Тедерсоо и др., 2012 г.; Туррини и Джованнетти, 2012 г.; Ян и др., 2012; Opik et al., 2013), из-за скрытного образа жизни почвенных грибов знания о глобальных закономерностях распространения видов остаются скудными. Наше понимание совместной изменчивости растений и связанных с ними грибковых сообществ в ландшафтном масштабе все еще ограничено.
Инвазивные виды растений могут служить хорошим примером для понимания взаимоотношений между растениями и моделей распространения их грибковых симбионтов.Хорошо известный пример важности микоризных грибов относится к роду Pinus после интродукции в южное полушарие. Хотя этот род в настоящее время считается одной из наиболее инвазивных групп деревьев (Richardson and Rejmánek, 2004), первоначальные насаждения Pinus sp. в южном полушарии не удалось из-за отсутствия правых видов грибов ECM (Pringle et al., 2009; Dickie et al., 2010). Глобальное распространение коммерческого грибкового инокулята ECM в значительной степени преодолело это ограничение (Vellinga et al., 2009), предполагая, что отсутствие совместно эволюционировавших мутуалистов было важным ограничением для укоренения сосны до введения грибов ECM. Арбускулярные микоризные грибы ассоциируются с большинством видов растений, распространены по всему миру и, как правило, обладают низкой специфичностью к хозяину (Smith and Read, 2008). Как таковые, они могут показаться маловероятными кандидатами на роль основных вторжений растений (Richardson et al., 2000). Тем не менее, глобальное распространение таксонов AMF ставится под сомнение тем фактом, что на самом деле большинство таксонов, выделенных молекулярным анализом, демонстрируют ограниченное распространение (Opik et al., 2010). Следовательно, крупномасштабное распространение видов растений может быть ограничено пространственно ограниченным распространением таксонов AMF, как показано для грибов ECM (Dickie et al., 2010).
Одним из основных препятствий в нашей способности понять важность грибкового симбиоза в формировании моделей распределения растений является наше ограниченное понимание микоризного статуса большинства видов растений (Reinhart et al., 2012). В нескольких публикациях (Trappe, 1962; Harley, Harley, 1987; Wang, Qiu, 2006) сообщается о наличии микоризных грибов в корнях большого числа видов растений.Однако затрагивается удивительно небольшое число потенциальных растений-хозяев. Хемпель и др. (2013) показывают, что для одного из наиболее изученных регионов мира, Центральной Европы, доступная информация о микоризном статусе включает менее одной трети видов региона (Wang and Qiu, 2006). Хотя простое документирование микоризного статуса вида растений может не полностью раскрыть механизм взаимодействия растения со своим симбиотическим партнером (Reinhart et al., 2012), оно обеспечивает основу для понимания закономерностей совместного распространения в ландшафтном масштабе (Wang and Qiu, 2006), кроме того, он может выявить экологические отношения между растениями, их симбиотическими грибами и окружающей средой (Peat and Fitter, 1993; Hempel et al., 2013). Хемпель и др. (2013) изучили взаимосвязь между микоризным статусом, характеристиками среды обитания, особенностями жизненного цикла и моделями распространения растений для 1752 представителей немецкой флоры. Они показали, что облигатно микоризные (ОМ) и немикоризные (НМ) виды растений, как правило, заметно отличаются от факультативно микоризных (ФМ) видов практически по всем анализируемым критериям. Самое интересное, что виды FM демонстрируют самый широкий географический и экологический ареал, за ними следуют виды OM, а затем виды NM. Это открытие предполагает, что факультативные взаимодействия, включая способность выживать без грибкового мутуалиста, когда он локально отсутствует, дают преимущество и обеспечивают более широкое распространение.Однако для понимания роли грибов в формировании распространения растений потребуются будущие исследования совместного существования растений и грибов в естественной среде.
Влияние почвенных грибов на распространение видов растений: новые взгляды из Альп
Как и растения, грибы также предъявляют особые требования к среде обитания (Fitter et al., 2005). Следовательно, в меньших масштабах предположительно важную роль играют местные факторы, такие как характеристики почвы. Например, микоризные грибы менее распространены во влажных, кислых и холодных средах обитания (Peat and Fitter, 1993; Gavito et al., 2005) и более многочисленны при умеренном стрессе питательных веществ (Smith and Read, 2008; Brundrett, 2009). Поскольку сообщества грибов, потенциально взаимодействующих с растениями, различаются в зависимости от условий окружающей среды, можно ожидать, что сдвиг в их распределении повлияет как на распределение видов растений, так и на состав сообщества. Однако эту гипотезу и детали того, как совокупность грибов связана с распространением видов растений, еще предстоит проверить на многих видах грибов и растений и в различных типах среды обитания.В частности, известно, что AMF Glomeromycetes взаимодействуют с большинством видов травянистых растений умеренного пояса (Harley and Harley, 1987; Wang and Qiu, 2006), и ожидается, что совокупность AMF будет влиять на толерантность растений к факторам окружающей среды, а также на их конкурентоспособность. Агарикомицеты, однако, могут функционировать как патогены растений и как грибы ECM, и, таким образом, воздействовать на растения непосредственно положительным или отрицательным образом. Грибы ECM взаимодействуют в основном с деревьями, но также было показано, что они образуют мутуализмы с недревесными видами, такими как альпийские растения (Read and Haselwandter, 1981).Агарикомицеты также могут косвенно воздействовать на растительные сообщества как разлагатели, влияя на разложение растительной подстилки и доступность питательных веществ (Zak et al., 2011).
Одна из трудностей в обнаружении эффекта биотических взаимодействий заключается в том, что они обычно коррелируют с абиотическими переменными (Meier et al., 2010; Pellissier et al., 2010a,b). Сообщества грибов изменяются по тем же абиотическим градиентам, что и растения. Что касается растений (Körner, 2003), колебаний температуры (Bahram et al., 2012), осадков (Hawkes et al., 2011) и снежного покрова (Zinger et al., 2009) могут быть связаны с изменением состава грибного сообщества. Аналогичным образом структура почвы (Hartmann et al., 2008), кислотность (Rousk et al., 2010), содержание азота (Egerton-Warburton and Allen, 2000) и органическое вещество (Zinger et al., 2011) влияют на грибковое сообщество. состав так же, как и в растительных сообществах (Dubuis et al., 2013). Следовательно, необходимо также учитывать абиотические переменные при попытке обнаружить взаимосвязь между распространением грибов и видов растений.
Методы
Набор данных о 213 сообществах растительных и почвенных грибов был собран в Западных Швейцарских Альпах (рис. 1) на высоте от 400 до 3210 м над уровнем моря, чтобы улучшить наше понимание пространственной изменчивости состава грибных сообществ (см. и др., 2013). Исчерпывающая инвентаризация всех видов сосудистых растений была проведена в каждом месте отбора проб в пределах 4 м 2 площадок (Dubuis et al., 2013). С каждого участка растительности (четыре угла и центр каждого участка) было взято пять образцов почвы для выделения ДНК и химического анализа почвы (см. Приложение).Состав сообществ почвенных грибов определяли путем пиросеквенирования ампликонов внутреннего транскрибируемого спейсера 1 (ITS1) грибов, полученных с помощью универсальных пар грибных праймеров ITS1F—ITS2 (Gardes, Bruns, 1993) из каждого образца почвенной ДНК. После анализа и кластеризации в оперативные таксономические единицы (OTU) с использованием алгоритма иерархической кластеризации DBC454 (подробно описанного в Pagni et al., 2013) было определено количество OTU на участок.
Рис. 1. Район исследования в Западных Швейцарских Альпах (700 км 2 ). Градиент от коричневого до синего соответствует высоте (400–3210 м). Зеленая поверхность представляет собой лесные массивы. Размер точки соответствует количеству OTU на участке (A) Glomeromycetes и (B) Agaricomycetes на инвентаризированных участках.
На отобранных участках были обнаружены OTU из 14 таксономических классов, в том числе двух классов, потенциально взаимодействующих с растениями, Glomeromycetes и Agaricomycetes. Чтобы иметь более полное представление о структуре сборки AMF в почвах, частичная область малой субъединицы гена рибосомной ДНК (SSU), созданная с помощью специфичных для Glomeromycetes (AMF) пар праймеров NS31-AM1 (Helgason et al., 1999), также был секвенирован. Состав этих сообществ будет представлен в другом месте. Здесь мы оставляем только информацию о количестве ОТЕ агарикомицетов и гломеромицетов. Уровень Agaricomycetes оценивали по количеству OTU ITS1 на участок, суммированному по подучастку, отнесенному к этому классу грибов, тогда как показатель Glomeromycetes оценивали по количеству OTU SSU на участок.
Используя модели распределения видов (SDM), мы проверили, включает ли предиктор, представляющий количество OTU грибов, потенциально взаимодействующих с растениями (т.т. е., мутуалисты или патогены) позволили подобрать статистические модели, лучше описывающие распространение видов растений. SDM обычно связывают присутствие и отсутствие данного вида с набором параметров окружающей среды (Guisan and Zimmermann, 2000). Мы запустили обобщенные линейные модели (GLM) с биномиальным распределением и функцией логистической связи для 124 видов растений (с более чем 10 экземплярами на участках выборки) с учетом семи абиотических факторов [градусо-дни, влажность, солнечная радиация, почва pH, фосфат почвы, уклон и топография, как это сделано в Dubuis et al.(2012, 2013)]. Мы провели GLM, включая количество OTU Glomeromycetes и Agaricomycetes, полученных на участок. Мы проверили производительность моделей с помощью 10-кратной перекрестной проверки, измеренной по площади под кривой рабочей характеристики приемника (ROC) (площадь под кривой, AUC). Мы сравнили оценку подгонки модели со скорректированным объясненным отклонением ( R 2 ). Мы рассчитали R 2 глобального соответствия, а также AUC с предиктором биотики грибов и без него.
Результаты
Когда количество OTU было включено в модели распространения растений, мы обнаружили значительное увеличение объясняемого отклонения ( R 2 , парный критерий Уилкоксона, Glomeromycetes V = 1306, p < 0,0001, Agaricomycetes V = 1326, p < 0,0001) и прогностической способности (AUC, парный критерий Вилкоксона, Glomeromycetes V = 6713, p < 0,0001, Agaricomycetes V < , p , , 0001, рис. 2). Даже после включения в модели абиотических факторов, которые с наибольшей вероятностью будут определять распространение видов растений (например, топография, климат и почва), информация о потенциальных мутуалистических грибах в почве последовательно повышала прогностическую точность SDM растений (рис. 2). Таким образом, количество OTU грибов Agaricomycetes и Glomeromycetes в почве дает важную дополнительную информацию для объяснения распределения видов растений. Поскольку агарикомицеты также включают другие функциональные группы, помимо ЭМП, такие как гнилостные и патогенные микроорганизмы, эти последние функции также могут играть роль в наблюдаемом сигнале в наших анализах.Для каждого вида растений мы также рассчитали среднюю высоту места, где этот вид был обнаружен. Параллельно мы рассчитали объясненное отклонение ( R 2 ) GL-моделей, когда в модели распределения видов растений было включено только количество OTU. Мы обнаружили, что те виды растений с наиболее сильной связью ( R 2 ) с числом OTU грибов встречались на больших высотах ( t -test, Glomeromycetes: t = 4.01, р = 0,0001, агарикомицеты: т = 2,97, р = 0,03; Рисунок 3).
Рисунок 2. Блок-диаграмма прогностической способности моделей распределения видов, измеренная с помощью площади под кривой (AUC) при рассмотрении только абиотических предикторов или дополнительном включении количества OTU потенциально мутуалистических грибов. Показаны результаты для Glomeromycetes (A) и Agaricomycetes (B) .
Рис. 3.Соотношение между средней высотой, на которой встречается вид растений, и объясняемым отклонением моделей, когда в модели распределения видов включено только количество OTU. Показаны результаты для Glomeromycetes (A) и Agaricomycetes (B) , когда они рассматривались отдельно как предиктор в моделях распределения видов. Пунктирные линии представляют 10-й и 90-й процентили доверительного интервала из квантильной регрессии.
Высокогорная почва особенно изменчива по количеству OTU (рис. 1), и грибковые различия между сообществами могут влиять на состав растений.В частности, было показано, что даже если микоризные ассоциации все еще достаточно часты в альпийской среде, уровень колонизации сильно различается между местообитаниями (Read and Haselwandter, 1981). Таким образом, дифференциальный состав мутуалистических грибов в альпийских местообитаниях может играть роль в модулировании состава растительных сообществ. Преимущества, которые растения получают от ассоциации с микоризными грибами, могут быть особенно важны для высокогорных видов растений, произрастающих в суровых условиях.Это может быть связано с тем, что высокогорная среда характеризуется более бедными почвами и более суровыми условиями окружающей среды (Körner, 2003), что делает укоренение и рост видов растений более опасным и делает симбиоз с микроорганизмами, который обеспечивает питательные вещества, отсутствующие в почве, более полезным. Подтверждая наши результаты, Wagg et al. (2011) исследовали рост сеянцев двух видов Pinus на двух высотах. Низинная почва была наиболее плодородной со сложным почвенным микробным сообществом, но последнее в целом отрицательно влияло на рост проростков.Напротив, высокогорная почва была наименее плодородной, но имела микробное сообщество, которое усиливало рост проростков. Это говорит о том, что в суровых условиях с более низкой продуктивностью, например, на больших высотах, зависимость растений от почвенных микробов может быть выше (Johnson et al., 1997; Thrall et al., 2007). Однако, поскольку разнообразие грибов может различаться в высокогорных местообитаниях, это может повлиять на формирование растительных сообществ.
Несколько исследований показали уменьшение разнообразия микоризных грибов, связанных с корнями растений, с увеличением высоты (Bahram et al., 2012; Гай и др., 2012; Луго и др., 2012). Хотя эти исследования зафиксировали изменения в разнообразии микоризных грибов с высотой, ни одно экспериментальное исследование еще не исследовало важность микоризных грибов для укоренения растений вдоль градиента высоты. В известняковом альпийском районе, похожем на изучаемый регион, Блашке (1991) показал, что микоризные грибы обычно обнаруживаются на корнях альпийской флоры, даже если интенсивность и характер колонизации микоризными грибами различаются между видами.Следовательно, даже если общее разнообразие микоризных грибов уменьшается с высотой, это не обязательно означает меньшую важность микоризных грибов для укоренения растений, но может означать более устойчивые отношения.
Даже если микоризные ассоциации по-прежнему довольно часто встречаются в альпийской среде, уровень колонизации сильно различается между видами растений и местообитаниями (Read and Haselwandter, 1981). Это объясняет, почему распределение некоторых видов показало связь с количеством OTU грибов, а другие — нет (рис. 3).Кроме того, Блашке (1991) предположил, что микоризная колонизация была наибольшей там, где формировались пространственные кластеры подходящих растений-хозяев (т. е. гильдии растений-хозяев), что предполагает облегчение колонизации из-за близости корней с установленными микоризными ассоциациями. Микоризная колонизация, как правило, положительно коррелирует с плотностью растений-хозяев, и уменьшение растительного покрова по мере увеличения высоты может также влиять на микоризную колонизацию корней растений (Hartnett et al., 1993; Genney et al., 2001; Руотсалайнен и Вэре, 2004). Следовательно, грибы могут влиять на укоренение альпийских растений, но структура растительных сообществ может также усиливать колонизацию корневых систем растений грибами посредством механизмов обратной связи.
Ограничения корреляционного подхода при изучении взаимодействия растений и грибов
Первое ограничение возникает из-за того, что, поскольку предложенный здесь подход является коррелятивным, мы не можем исключить возможность того, что включение в модели важных дескрипторов грибов AMF и ECM косвенно отражает отсутствующий абиотический компонент (Wisz et al., 2013). Многие абиотические факторы влияют на установление и рост видов растений в сообществе. Это означает, что выявление и измерение всего набора абиотических условий, способствующих сборке растительных сообществ, является особенно сложной задачей. Относительно легко упустить некоторые важные абиотические предикторы распространения растений (Austin and Van Niel, 2011). В частности, мы измеряли влажность почвы как отношение между осадками и потенциальной эвапотранспирацией (Zimmermann and Kienast, 1999), поскольку прямые измерения влажности почвы по-прежнему трудно провести стандартизированным способом на больших территориях (Austin and Van Niel, 2011). ).Тем не менее влажность почвы считается сильным фактором распространения видов растений даже в холодных условиях (Le Roux et al., 2013). Поскольку ожидается, что грибы также сильно зависят от влажности почвы, разнообразие микоризных грибов может косвенно влиять на условия влажности на участке, но это еще предстоит проверить с помощью более качественных измерений влажности почвы в полевых условиях.
Поскольку процент микоризной колонизации обычно положительно коррелирует с плотностью растений-хозяев (Genney et al., 2001), разнообразие микоризных грибов может также влиять на конкурентное давление между видами растений за укоренение и устойчивость. Межвидовая конкуренция растений может быть важным фактором, определяющим распространение видов даже в холодных условиях (Pellissier et al., 2010a). Совокупность грибов в почве может коррелировать с плотностью растений (Gilbert et al., 2002). Поскольку в условиях высокой плотности ожидается усиление конкуренции между видами растений, что приведет к их исключению из сообществ, влияние грибов на растения можно спутать с конкурентными эффектами растений.Чтобы проанализировать потенциальное смешанное влияние плотности растений, мы проверили взаимосвязь между суммой растительного покрова и количеством OTU. Мы не обнаружили связи между количеством грибных OTU и суммой растительного покрова на данном участке (Коэффициент детерминации, Glomeromycetes R 2 = 0,06, Agaricomycetes R 2 = 0,04). Таким образом, взаимосвязь между распространением грибов и растений не зависит от потенциально искажающего эффекта плотности растений.Тем не менее, мы не учитывали конкуренцию растений под землей, поскольку такую меру трудно измерить в полевых условиях. Как следствие, полностью исключить этот смешанный эффект нельзя.
Еще одно ограничение анализа связано с низким разрешением назначения OTU. OTU, идентифицированные из прочтений ITS, были отнесены на грубом таксономическом уровне, что не позволяет нам узнать, к какой функциональной группе они принадлежат. Таким образом, в то время как мы ожидаем последовательной микоризной функции для гломеромицетов, агарикомицеты перегруппировывают виды из различных функциональных групп, включая гнилостные, патогены растений и мутуалистов.Как следствие, богатство OTU агарикомицетов в почве не может быть напрямую связано с функцией и должно интерпретироваться с осторожностью. Поэтому, даже если бы мы обнаружили в наших анализах биологический сигнал, мы не можем сделать вывод о функциональной связи, связывающей разнообразие грибов с распространением растений. В этом отношении корреляционные подходы предоставляют интересные проверяемые гипотезы, но только экспериментальные проекты могут в конечном итоге раскрыть причинно-следственную связь между растениями и грибами.
Заключение и перспективы
Наш обзор литературы выявил отсутствие у нас знаний о пространственных вариациях взаимодействия между растениями и микоризными грибами в различных экологических условиях.Большинство исследований до сих пор ограничивали свой объем несколькими сообществами в стандартных условиях окружающей среды и подчеркивали роль микоризных грибов в воздействии на состав растительных сообществ. Однако некоторые данные свидетельствуют о том, что роль микоризных грибов может выходить за рамки отдельных сообществ и может меняться в зависимости от широких экологических градиентов. Наши результаты подтверждают роль сообщества почвенных грибов в распределении видов растений в ландшафтном масштабе, особенно на больших высотах, но также требуют более контролируемого подхода к проблеме.Потребуются манипулятивные полевые эксперименты, в которых рассматривается манипулирование как растениями, так и микоризными грибами, чтобы узнать больше об их взаимодействии в градиентах окружающей среды. Насколько нам известно, ни одно исследование еще не разработало явные в пространстве эксперименты, которые могли бы формально продемонстрировать роль грибов в управлении пространственным распределением видов растений. Для этого потребуется воздействовать на грибковые сообщества в полевых условиях либо с помощью дополнительных обработок против грибков (Mendes et al., 2010), либо с помощью фунгицидов для удаления грибков (Helgason et al., 2007), или их сочетание. Тем не менее, полевые манипуляции с микроорганизмами сложны, и дополнительные корреляционные подходы, такие как использованные здесь, вероятно, останутся ценными для сбора информации о взаимодействии растений и грибов в более широких пространственных масштабах.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Благодарим С.Джованеттина, С. Года, Ж.-Н. Pradervand, C. Purro, V. Rion и P. Vittoz за их помощь в инвентаризации растительности и сборе образцов, а также Y. Ben Saida за помощь в выделении ДНК, A. von Ungern-Sternberg за помощь в анализе данных и Jut Wynne за английские исправления. Часть расчетов выполнена в Центре высокопроизводительных вычислений Vital-IT (http://www.vital-it.ch) SIB Швейцарского института биоинформатики (SIB) (http://www.vital-it .ч). SIB получает финансовую поддержку от федерального правительства Швейцарии через Государственный секретариат по вопросам образования и исследований (SER).Это исследование было поддержано Швейцарским национальным научным фондом (грант № 31003A-125145 Антуану Гизану). Лоик Пеллиссье получил поддержку гранта Датского совета по независимым исследованиям № 12-126430, Мари Моора — грантов Эстонского научного фонда (9050, 9157) и Европейского фонда регионального развития (Центр передового опыта FIBIR), Элен Никулита-Хирзель — гранта Эстонского научного фонда (9050, 9157). Грант Швейцарского национального научного фонда PMPDP3_129027.
Ссылки
Остин, М. П., и Ван Нил, К.П. (2011). Улучшение моделей распределения видов для изучения изменения климата: выбор переменных и масштаб. Ж. Биогеогр . 38, 1–8. doi: 10.1111/j.1365-2699.2010.02416.x
Полнотекстовая перекрестная ссылка
Бахрам, М., Пылме, С., Кыльялг, У., Зарре, С., и Тедерсоо, Л. (2012). Региональные и локальные закономерности разнообразия эктомикоризных грибов и структуры сообщества вдоль высотного градиента в гирканских лесах северного Ирана. Новый Фитол . 193, 465–473.doi: 10.1111/j.1469-8137.2011.03927.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Блашке, Х. (1991). Распространение, микоризная инфекция и структура корней цветочных элементов calcicole на границе деревьев, Баварские Альпы, Германия. Арт. Альп. Рез . 23, 444–450. дои: 10.2307/1551686
Полнотекстовая перекрестная ссылка
Брундретт, MC (2009). Микоризные ассоциации и другие средства питания сосудистых растений: понимание глобального разнообразия растений-хозяев путем разрешения противоречивой информации и разработки надежных средств диагностики. Почва для растений 320, 37–77. doi: 10.1007/s11104-008-9877-9
Полнотекстовая перекрестная ссылка
Buée, M., Reich, M., Murat, C., Morin, E., Nilsson, R.H., Uroz, S., et al. (2009). 454 Пиросеквенирование лесных почв выявило неожиданно высокое разнообразие грибов. Новый Фитол . 184, 449–456. doi: 10.1111/j.1469-8137.2009.03003.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Чаудхари В., Капур Р. и Бхатнагар А.К.(2008). Влияние двух арбускулярных микоризных грибов на концентрации эфирного масла и артемизинина в трех образцах Artemisia annua L. Appl. Грунт Экол . 40, 174–181. doi: 10.1016/j.apsoil.2008.04.003
Полнотекстовая перекрестная ссылка
Диксон Т.Л. и Фостер Б.Л. (2008). Относительная важность видового пула, продуктивности и нарушений в регулировании видового богатства пастбищных растений: полевой эксперимент. Дж. Экол . 96, 937–946.doi: 10.1111/j.1365-2745.2008.01420.x
Полнотекстовая перекрестная ссылка
Дюбюи, А., Джованеттина, С., Пеллиссье, Л., Потье, Дж., Виттоз, П., и Гисан, А. (2012). Улучшение предсказания распределения видов растений и состава сообщества путем добавления эдафических к топоклиматическим переменным. Дж. Вег. Наука . 24, 593–606. doi: 10.1111/jvs.12002
Полнотекстовая перекрестная ссылка
Дюбюи, А., Россье, Л., Потье, Ж., Пеллиссье, Л., Виттоз, П. и Гисан, А.(2013). Прогнозирование текущих и будущих моделей пространственного сообщества функциональных признаков растений. Экография 36, 1158–1168. doi: 10.1111/j.1600-0587.2013.00237.x
Полнотекстовая перекрестная ссылка
Эгертон-Уорбертон, Л. М., и Аллен, Э. Б. (2000). Сдвиги арбускулярных микоризных сообществ по градиенту антропогенного осаждения азота. Экол. Заявка . 10, 484–496. doi: 10.1890/1051-0761(2000)010[0484:SIAMCA]2.0.CO;2
Полнотекстовая перекрестная ссылка
Слесарь, А., Гиллиган С., Холлингворт К., Клечковски А., Твайман Р. и Питчфорд Дж. (2005). Биоразнообразие и экосистемная функция в почве. Функц. Экол . 19, 369–377. doi: 10.1111/j.0269-8463.2005.00969.x
Полнотекстовая перекрестная ссылка
Фрэнсис Р. и Рид Д. Дж. (1995). Мутуализм и антагонизм в микоризном симбиозе с особым акцентом на влияние на структуру растительного сообщества. Кан. Дж. Бот . 73, С1301–С1309. дои: 10.1139/b95-391
Полнотекстовая перекрестная ссылка
Гай, Дж.П., Тиан Х., Ян Ф.Ю., Кристи П., Ли С.Л. и Клирономос Дж.Н. (2012). Разнообразие арбускулярных микоризных грибов вдоль тибетского градиента высот. Педобиология 55, 145–151. doi: 10.1016/j.pedobi.2011.12.004
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Гардес М. и Брунс Т. Д. (1993). Праймеры ITS с повышенной специфичностью к базидиомицетам — применение для идентификации микоризы и ржавчины. Мол. Экол . 2, 113–118.doi: 10.1111/j.1365-294X.1993.tb00005.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Гавито, М.Е., Олссон, П.А., Рухье, Х., Медина-Пенафиэль, А., Якобсен, И., Баго, А., и соавт. (2005). Температурные ограничения роста и функционирования культур корневых органов с арбускулярными микоризными грибами. Новый Фитол . 168, 179–188. doi: 10.1111/j.1469-8137.2005.01481.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Генни, Д.Р., Хартли С.Х. и Александр И.Дж. (2001). Арбускулярная микоризная колонизация увеличивается с увеличением плотности хозяев в сообществе вересковых пустошей. Новый Фитол . 152, 355–363. doi: 10.1046/j.0028-646X.2001.00248.x
Полнотекстовая перекрестная ссылка
Гилберт Г.С., Феррер А. и Карранса Дж. (2002). Разнообразие трутовиков и плотность хозяев во влажном тропическом лесу. Биодайверы. Консервировать . 11, 947–957. дои: 10.1023/A:1015896204113
Полнотекстовая перекрестная ссылка
Грайм, Дж.П., Макки, Дж.М.Л., Хиллер, С.Х., и Рид, Д.Дж. (1987). Флористическое разнообразие в модельной системе с использованием экспериментальных микрокосмов. Природа 328, 420–422. дои: 10.1038/328420a0
Полнотекстовая перекрестная ссылка
Гисан, А., и Рахбек, К. (2011). SESAM — новая структура, объединяющая макроэкологические модели и модели распределения видов для прогнозирования пространственно-временных закономерностей группировок видов. Ж. Биогеогр . 38, 1433–1444. doi: 10.1111/j.1365-2699.2011.02550.x
Полнотекстовая перекрестная ссылка
Гизан, А.и Циммерманн, Н. Е. (2000). Прогнозные модели распределения местообитаний в экологии. Экол. Модель . 135, 147–186. doi: 10.1016/S0304-3800(00)00354-9
Полнотекстовая перекрестная ссылка
Харли, Дж. Л., и Харли, Э. Л. (1987). Контрольный список микоризы в британской флоре. Новый Фитол . 105, 741–749. doi: 10.1111/j.1469-8137.1987.tb00674.x
Полнотекстовая перекрестная ссылка
Хартманн А., Шмид М., ван Туйнен Д. и Берг Г. (2008). Растительный отбор микробов. Почва для растений 321, 235–257. doi: 10.1007/s11104-008-9814-y
Полнотекстовая перекрестная ссылка
Hartnett, C.D., and Wilson, G.W.T. (1999). Микоризы влияют на структуру и разнообразие растительных сообществ в высокотравных прериях. Экология 80, 1187–1195. doi: 10.1890/0012-9658(1999)080[1187:MIPCSA]2.0.CO;2
Полнотекстовая перекрестная ссылка
Хартнетт Д., Хетрик Б., Уилсон Г. и Гибсон Д. (1993). Влияние микоризы на внутри- и межвидовые взаимодействия соседей среди сосуществующих степных трав. Дж. Экол . 81, 787–795. дои: 10.2307/2261676
Полнотекстовая перекрестная ссылка
Хоукс, К.В., Кивлин, С.Н., Рокка, Дж.Д., Хьюге, В., Томсен, М.А., и Саттл, К.Б. (2011). Реакция грибного сообщества на осадки. Глоб. Сменить Биол . 17, 1637–1645. doi: 10.1111/j.1365-2486.2010.02327.x
Полнотекстовая перекрестная ссылка
Хелгасон, Т., Фиттер, А. Х., и Янг, Дж. П. В. (1999). Молекулярное разнообразие арбускулярных микоризных грибов, колонизирующих Hyacinthoides non-scripta (колокольчик) в полуестественном лесу. Мол. Экол . 8, 659–666. doi: 10.1046/j.1365-294x.1999.00604.x
Полнотекстовая перекрестная ссылка
Хельгасон, Т., Мерривезер, Дж. В., Янг, Дж. П. В., и Фиттер, А. Х. (2007). Специфичность и устойчивость арбускулярных микоризных грибов естественного лесного сообщества. Дж. Экол . 95, 623–630. doi: 10.1111/j.1365-2745.2007.01239.x
Полнотекстовая перекрестная ссылка
Хемпель С., Гётценбергер Л., Кюн И., Михальски Г. С., Риллиг С. М., Зобель М., и другие. (2013). Микоризы во флоре Центральной Европы — связь с особенностями истории жизни растений и экологией. Экология 94, 1389–1399. дои: 10.1890/12-1700.1
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Джонсон, Северная Каролина, Грэм, Дж. Х., и Смит, Ф. А. (1997). Функционирование микоризных ассоциаций в континууме мутуализм-паразитизм. Новый Фитол . 135, 575–585. doi: 10.1046/j.1469-8137.1997.00729.x
Полнотекстовая перекрестная ссылка
Юнг, С.К., Мартинес-Медина, А., Лопес-Раес, Дж. А., и Позо, М. Дж. (2012). Устойчивость к микоризе и активация защитных сил растений. J. Chem. Экол . 38, 651–664. doi: 10.1007/s10886-012-0134-6
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Киквидзе З., Пугнер Ф.И., Брукер Р.В., Чолер П., Лорти С.Дж., Мишале Р. и др. (2005). Связывание закономерностей и процессов в альпийских растительных сообществах: глобальное исследование. Экология 86, 1395–1400.дои: 10.1890/04-1926
Полнотекстовая перекрестная ссылка
Кивлин, С. Н., Хоукс, К. В., и Треседер, К. К. (2011). Мировое разнообразие и распространение арбускулярных микоризных грибов. Почвенный биол. Биохим . 43, 2294–2303. doi: 10.1016/j.soilbio.2011.07.012
Полнотекстовая перекрестная ссылка
Клирономос, Дж., Зобель, М., и Тиббет, М. (2011). Силы, структурирующие растительные сообщества: количественная оценка важности микоризного симбиоза. Новый Фитол .189, 366–370. doi: 10.1111/j.1469-8137.2010.03550.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Коорем, К., Сакс, Ю., Собер, В., и Уйбопуу, А. (2012). Влияние арбускулярной микоризы на состав сообщества и пополнение сеянцев в подлеске леса умеренного пояса. Базовое приложение Экол . 13, 663–672. doi: 10.1016/j.baae.2012.09.008
Полнотекстовая перекрестная ссылка
Кёрнер, К. (2003). Жизнь альпийских растений, 2-е издание . Гейдельберг: Спрингер.дои: 10.1007/978-3-642-18970-8
Полнотекстовая перекрестная ссылка
Le Roux, X., Schmid, B., Poly, F., Barnard, R.L., Niklaus, P.A., Guillaomaud, N., et al. (2013). Условия окружающей среды в почве и накопление микробов опосредуют влияние разнообразия растений на активность нитрифицирующих и денитрифицирующих ферментов почвы на пастбищах умеренного пояса. PLoS ONE 8:e61069. doi: 10.1371/journal.pone.0061069
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Ли, Х., Smith, S.E., Holloway, R.E., Zhu, Y., and Smith, F.A. (2006). Арбускулярные микоризные грибы способствуют поглощению фосфора пшеницей, выращенной на фосфорфиксирующей почве, даже при отсутствии положительной реакции роста. Новый Фитол . 172, 536–543. doi: 10.1111/j.1469-8137.2006.01846.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Лю, Ю., Ю, С., Се, З. П., и Штехелин, К. (2012). Анализ отрицательной обратной связи между растением и почвой в субтропическом муссонном лесу. Дж. Экол . 100, 1019–1028. doi: 10.1111/j.1365-2745.2012.01953.x
Полнотекстовая перекрестная ссылка
Луго, Массачусетс, Негритто, Массачусетс, Жофре, М., Антон, А., и Галетто, Л. (2012). Колонизация местных трав Анд арбускулярными микоризными грибами в Пуне: вопрос высоты, пути фотосинтеза хозяина и жизненных циклов хозяина. FEMS микробиол. Экол . 81, 455–466. doi: 10.1111/j.1574-6941.2012.01373.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Мейер, Э.S., Kienast, F., Pearman, P.B., Svenning, J.-C., Thuiller, W., Araújo, M.B., et al. (2010). Биотические и абиотические переменные демонстрируют небольшую избыточность в объяснении распределения видов деревьев. Экография 33, 1038–1048. doi: 10.1111/j.1600-0587.2010.06229.x
Полнотекстовая перекрестная ссылка
Мендес П.Ф., Васконселлос Р.Л.Ф., де Паула А.М. и Кардосо Э.Дж.Б.Н. (2010). Оценка потенциала лесных пород при «микробиологическом управлении» для восстановления деградировавших районов добычи полезных ископаемых. Вода. Воздуха. Загрязнение почвы . 208, 79–89. doi: 10.1007/s11270-009-0150-5
Полнотекстовая перекрестная ссылка
Мура, М., и Зобель, М. (2010). «Арбускулярная микориза и взаимодействие растений между собой», в Положительные взаимодействия растений и динамика сообщества , изд. FI Pugnaire (Лондон: CRC Press), 79–98.
Мура М., Опик М., Сен Р. и Зобель М. (2004). Местные сообщества арбускулярных микоризных грибов по-разному влияют на продуктивность проростков редких и обычных видов Pulsatilla . Функц. Экол . 18, 554–562. doi: 10.1111/j.0269-8463.2004.00876.x
Полнотекстовая перекрестная ссылка
Мура, М., Бергер, С., Дэвисон, Дж., Эпик, М., Боммарко, Р., Брюльхайде, Х., и другие. (2011). Чужеродные растения связаны с широко распространенными таксонами арбускулярных микоризных грибов: данные исследования континентального масштаба с использованием массового параллельного секвенирования 454. Ж. Биогеогр . 38, 1305–1317. doi: 10.1111/j.1365-2699.2011.02478.x
Полнотекстовая перекрестная ссылка
Опик, М., Метсис, М., Даниэлл, Т.Дж., Зобель, М., и Мура, М. (2009). Крупномасштабное параллельное секвенирование 454 выявляет специфичность экологической группы хозяев арбускулярных микоризных грибов в боронеморальном лесу. Новый Фитол . 184, 424–437. doi: 10.1111/j.1469-8137.2009.02920.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Opik, M., Vanatoa, A., Vanatoa, E., Moora, M., Davison, J., Kalwij, J.M., et al. (2010). Онлайн-база данных MaarjAM раскрывает глобальные и экосистемные модели распространения арбускулярных микоризных грибов (Glomeromycota). Новый Фитол . 188, 223–241. doi: 10.1111/j.1469-8137.2010.03334.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Opik, M., Zobel, M., Cantero, J.J., Davison, J., Facelli, J.M., Hiiesalu, I., et al. (2013). Глобальный отбор корней растений расширяет описанное молекулярное разнообразие арбускулярных микоризных грибов. Микориза 23, 411–430. doi: 10.1007/s00572-013-0482-2
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Паньи, М., Никулита-Хирзель, Х., Пеллиссье, Л., Дюбюи, А., Ксенариос, И., Гизан, А., и соавт. (2013). Иерархическая группировка пиропоследовательностей на основе плотности в больших масштабах — случай ITS1 грибов. Биоинформатика 15, 1268–1274. doi: 10.1093/биоинформатика/btt149
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Торф Х. и Фиттер А. (1993). Распространение арбускулярной микоризы во флоре Великобритании. Новый Фитол . 125, 845–854. дои: 10.1111/j.1469-8137.1993.tb03933.x
Полнотекстовая перекрестная ссылка
Pellissier, L., Anne Bråthen, K., Pottier, J., Randin, C.F., Vittoz, P., Dubuis, A., et al. (2010а). Модели распределения видов показывают очевидное конкурентное и стимулирующее влияние доминирующих видов на распространение растений тундры. Экография 33, 1004–1014. doi: 10.1111/j.1600-0587.2010.06386.x
Полнотекстовая перекрестная ссылка
Пеллиссье Л., Потье Ж., Виттоз П., Дюбюи А.и Гисан, А. (2010b). Пространственная структура цветочной морфологии: возможное понимание влияния опылителей на распространение растений. Ойкос 119, 1805–1813. doi: 10.1111/j.1600-0706.2010.18560.x
Полнотекстовая перекрестная ссылка
Pellissier, L., Fiedler, K., Ndribe, C., Dubuis, A., Pradervand, J.-N., Guisan, A., et al. (2012). Сдвиги в видовом богатстве, специализации травоядных и устойчивости растений по градиенту высоты. Экол. Эвол . 2, 1818–1825 гг. дои: 10.1002/экс3.296
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Pottier, J., Dubuis, A., Pellissier, L., Maiorano, L., Rossier, L., Randin, C.F., et al. (2013). Точность предсказания растительного сообщества на основе моделей распределения видов варьируется в зависимости от градиента окружающей среды. Глоб. Экол. Биогеогр . 22, 52–63. doi: 10.1111/j.1466-8238.2012.00790.x
Полнотекстовая перекрестная ссылка
Прингл, А., Бевер, Дж. Д., Гардес, М., Паррент, Дж. Л., Риллиг, М.C. и Klironomos, JN (2009). Микоризные симбиозы и инвазии растений. год. Преподобный Экол. Эвол. Сист . 40, 699–715. doi: 10.1146/annurev.ecolsys.39.110707.173454
Полнотекстовая перекрестная ссылка
Рид, Д. Дж., и Хазелвандтер, К. (1981). Наблюдения за микоризным статусом некоторых альпийских растительных сообществ. Новый Фитол . 88, 341–352. doi: 10.1111/j.1469-8137.1981.tb01729.x
Полнотекстовая перекрестная ссылка
Рейнхарт, К.О., Уилсон, Г.В.Т. и Ринелла, М. Дж. (2012). Прогнозирование реакции растений на микоризу: интеграция истории эволюции и свойств растений. Экол. Письмо . 15, 689–695. doi: 10.1111/j.1461-0248.2012.01786.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Ричардсон Д.М. и Рейманек М. (2004). Хвойные деревья как инвазивные пришельцы: глобальный обзор и прогностическая основа. Разнообразие и распространение , 10, 321–331. doi: 10.1111/j.1366-9516.2004.00096.x
Полнотекстовая перекрестная ссылка
Ричардсон, Д.М., Пишек П., Рейманек М., Барбур М. Г., Панетта Ф. Д. и Уэст С. Дж. (2000). Натурализация и инвазия чужеродных растений: понятия и определения. Разнообразие и распространение , 6, 93–107. doi: 10.1046/j.1472-4642.2000.00083.x
Полнотекстовая перекрестная ссылка
Rousk, J., Båath, E., Brookes, P.C., Lauber, C.L., Lozupone, C., Caporaso, J.G., et al. (2010). Почвенные бактериальные и грибковые сообщества поперек градиента рН в пахотной почве. ИСМЭ J . 4, 1340–1351.doi: 10.1038/ismej.2010.58
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Руотсалайнен, А., и Вэре, Х. (2004). Колонизация корневых грибов вдоль высотного градиента в Северной Норвегии. Арт. Антаркт. Альп. Рез . 36, 239–243. doi: 10.1657/1523-0430(2004)036[0239:RFCAAA]2.0.CO;2
Полнотекстовая перекрестная ссылка
Смит, С.Э., и Рид, Д.Дж. (2008). Микоризный симбиоз . Лондон: Академическая пресса.
Штрайтвольф-Энгель, Р., ван дер Хейден, М.Г.А., и Сандерс, И.Р. (2001). Экологическое значение действия арбускулярных микоризных грибов на клональное размножение растений. Экология 82, 2846–2859. doi: 10.1890/0012-9658(2001)082[2846:TESOAM]2.0.CO;2
Полнотекстовая перекрестная ссылка
Tedersoo, L., Bahram, M., Toots, M., Diedhiou, A.G., Henkel, T.W., Kjöller, R., et al. (2012). К глобальным закономерностям разнообразия и структуры сообщества эктомикоризных грибов. Мол. Экол .21, 4160–4170. doi: 10.1111/j.1365-294X.2012.05602.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Тралл, П. Х., Слэттери, Дж. Ф., Бродхерст, Л. М., и Бикфорд, С. (2007). Географические закономерности численности и адаптации симбионтов во взаимодействиях коренных австралийских акаций и ризобий. Дж. Экол . 95, 1110–1122. doi: 10.1111/j.1365-2745.2007.01278.x
Полнотекстовая перекрестная ссылка
Траппе, Дж. М. (1962). Грибные ассоциаты эктотрофных микориз. Бот. Версия . 28, 538–606. дои: 10.1007/BF02868758
Полнотекстовая перекрестная ссылка
Треседер, К.К., и Кросс, А. (2006). Глобальное распространение арбускулярных микоризных грибов. Экосистемы 9, 305–316. doi: 10.1007/s10021-005-0110-x
Полнотекстовая перекрестная ссылка
Туррини, А., и Джованнетти, М. (2012). Арбускулярные микоризные грибы в национальных парках, заповедниках и охраняемых территориях по всему миру: стратегическая перспектива их сохранения in situ. Микориза 22, 81–97. doi: 10.1007/s00572-011-0419-6
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
van der Heijden, M., Klironomos, J., Ursic, M., Moutoglis, P., Streitwolf-Engel, R., Boller, T., et al. (1998). Разнообразие микоризных грибов определяет биоразнообразие растений, изменчивость экосистем и продуктивность. Природа 396, 69–72. дои: 10.1038/23932
Полнотекстовая перекрестная ссылка
ван дер Хейден, MG (2003). Различные арбускулярные микоризные грибы изменяют сосуществование и распределение ресурсов между сосуществующими растениями. Новый Фитол . 157, 569–578. doi: 10.1046/j.1469-8137.2003.00688.x
Полнотекстовая перекрестная ссылка
van der Heijden., M.G.A., Bardgett, R., and van Straalen, N. (2008). Невидимое большинство: почвенные микробы как движущие силы разнообразия и продуктивности растений в наземных экосистемах. Экол. Письмо . 11, 296–310. doi: 10.1111/j.1461-0248.2007.01139.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
ван дер Хейден, М. Г. А., и Сандерс, И.Р. (ред.). (2002). «Экология микоризы», в Ecological Studies Vol. 157 (Гейдельберг: Springer Verlag), 243–265. дои: 10.1007/978-3-540-38364-2_10
Полнотекстовая перекрестная ссылка
Вагг, К., Антунес, П.М., и Петерсон, Р.Л. (2011). Взаимодействия арбускулярных микоризных грибов, связанные с филогенезом, с нехозяином. Симбиоз 53, 41–46. doi: 10.1007/s13199-011-0107-5
Полнотекстовая перекрестная ссылка
Вейхер Э., Фройнд Д., Бантон Т., Стефански А., Ли, Т., и Бентивенга, С. (2011). Достижения, проблемы и развивающийся синтез теории сборки экологического сообщества. Филос. Транс. Р. Соц. Лонд. Б биол. Наука . 366, 2403–2413. doi: 10.1098/rstb.2011.0056
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Уиппс, Дж. (2004). Перспективы и ограничения использования микоризы в биоконтроле корневых патогенов. Кан. Дж. Бот . 82, 1198–1227. дои: 10.1139/b04-082
Полнотекстовая перекрестная ссылка
Виш, М.S., Pottier, J., Kisslig, W.D., Pellissier, L., Lenoir, J., Damgaard, C.F., et al. (2013). Роль биотических взаимодействий в формировании распределения и реализованных совокупностей видов: значение для моделирования распределения видов. Биол. Версия . 88, 15–30. doi: 10.1111/j.1469-185X.2012.00235.x
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Ян, Х., Занг, Ю. и Юань, Ю. (2012). Селективность растений-хозяев влияет на распространение арбускулярных микоризных грибов: данные метаданных о последовательности рДНК ITS. BMC Evol. Биол . 12:50. дои: 10.1186/1471-2148-12-50
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Зак, Д. Р., Прегитцер, К. С., Бертон, А. Дж., Эдвардс, И. П., и Келлнер, Х. (2011). Реакция микробов на изменяющуюся среду: последствия для будущего функционирования наземных экосистем. Грибной Экол . 4, 386–395. doi: 10.1016/j.funeco.2011.04.001
Полнотекстовая перекрестная ссылка
Циммерманн Н. и Кинаст Ф.(1999). Прогнозное картирование альпийских пастбищ в Швейцарии: подход «виды против сообщества». Дж. Вег. Наука . 10, 469–482. дои: 10.2307/3237182
Полнотекстовая перекрестная ссылка
Зингер Л., Куасак Э., Холер П. и Геремия Р. А. (2009). Оценка микробных сообществ методом расчленения графа при изучении почвенных грибов на двух альпийских лугах. Заяв. Окружающая среда. Микробиол . 75, 5863–5870. doi: 10.1128/AEM.00748-09
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Зингер, Л., Lejon, D.P., Baptist, F., Bouasria, A., Aubert, S., Geremia, R.A., et al. (2011). Контрастные модели разнообразия кренархейных, бактериальных и грибковых почвенных сообществ в альпийском ландшафте. PLoS ONE 6:e19950. doi: 10.1371/journal.pone.0019950
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Зобель М., Мура М. и Хаукиоя Э. (1997). Сосуществование растений в интерактивной среде: нельзя забывать об арбускулярной микоризе. Ойкос 78, 202–208.
Приложение
Отбор проб был проведен в Западных швейцарских Альпах (рис. 1). Градиент высот области охватывает высоты от 400 до 3210 м. Следуя сбалансированной схеме случайной стратификации выборки, основанной на высоте, уклоне и экспозиции, мы отобрали 213 участков с разнообразной нелесной растительностью. Исчерпывающая инвентаризация всех видов сосудистых растений была проведена в каждом месте отбора проб в пределах 4 м 2 площадок. Для каждого участка растительности было взято пять образцов почвы (из четырех углов и по одному из середины каждого участка) для выделения ДНК и химического анализа почвы.Для химического анализа почвы пять образцов были смешаны, чтобы выровнять возможную вариацию внутри делянки. После высушивания образцов мы измерили pH, текстуру почвы, общий азот (N), общий фосфор (P) и органический углерод (C) (подробности о конкретных методах см. Dubuis et al., 2013).
Мы рассчитали климатические предикторы по территории на основе данных о среднемесячных значениях средней температуры (°C) и суммы осадков (мм), зарегистрированных за период 1961–2010 гг. швейцарской сетью метеорологических станций, интерполированных с использованием цифровой модели рельефа. с разрешением 25 м.Мы рассчитывали градусо-дни как сумму дней, умноженных на температуру выше 0°C, а индекс влажности как разницу между осадками и потенциальной эвапотранспирацией для вегетационного периода июня, июля и августа (Zimmermann, Kienast, 1999). Топографические переменные были получены из цифровой модели рельефа.
Условия ПЦР и пиросеквенирование
Образцы почвы были независимо гомогенизированы и просеяны с помощью сита с размером ячеек 2 мм. 3 г почвы каждого образца высушивали на воздухе при 30°C и из 250 мг сухой почвы выделяли геномную ДНК в соответствии с рекомендациями дистрибьютора PowerSoil ® -htp 96 Well Soil DNA Isolation Kit (MoBio Laboratory, Carlsbad, США).Универсальные грибковые праймеры ITS1F и ITS2 были выбраны для амплификации всего грибкового сообщества, присутствующего в образцах, также были использованы специфичные для Glomeroycota праймеры NS31 и AM1 для амплификации частичной области малой субъединицы гена рибосомной ДНК (SSU). ITS1F и NS31 были связаны с праймером для пиросеквенирования A Genome Sequencer FLX Titanium, также ITS2 и AM1 были связаны с праймером для пиросеквенирования B. Ключ штрих-кода длиной 10 п.н. был вставлен между праймером A и последовательностью праймера ITS1F или NS31.Таким образом, составной прямой праймер представлял собой: 5′- CGTATCGCCTCCCTCGCGCCATCAG -(X)10- CTTGGTCATTTAGAGGAAGTAA -3’/5′- CGTATCGCCTCCCTCGCGCCATCAG -(X)10- TTGGAGTG9GCAAGTC соответственно. Обратный праймер: 5′- CTATGCGCCTTGCCAGCCCGCTCAG — GCTGCGTTCTTCATCGATGC -3’/5′- CTATGCGCCTTGCCAGCCCGCTCAG — GTTTCCCGTAA GGCGCCGAA -3′, где A и B праймеры подчеркнуты. 10, а специфичные для грибов праймеры показаны курсивом.Использовался набор из 162 различных ключей штрих-кода по 10 п.н. каждый, разработанных в соответствии с указаниями компании «Рош». Основным критерием этих ключей штрих-кода является то, что между всеми используемыми штрих-кодами должно различаться не менее 3 нуклеотидов из 10. Реакции ПЦР ITS1 проводили, как описано у Buée et al. (2009), также проводили ПЦР-реакции гена SSU, как описано Opik et al. (2009). Количество ДНК в продуктах ПЦР измеряли с помощью NanoDrop 1000 (Thermo Scientific, Уилмингтон, Делавэр, США).Ампликоны с разными ключами штрих-кода объединяли в эквимолярных пропорциях. Таким образом, для каждого маркера (ITS1 или SSU) было создано 6 библиотек ампликонов по 167 образцов в каждой. Затем библиотеки ампликонов разделяли электрофорезом через 1,5%-ный агарозный гель в 0,5×ТВЕ и очищали ПЦР-ампликоны от геля с помощью набора Qiagen QIAquick Gel Extraction (Qiagen, Швейцария). Количество ДНК в очищенных библиотеках ампликонов измеряли с помощью NanoDrop 1000 (Thermo Scientific, Уилмингтон, Делавэр, США).Пиросеквенирование шести библиотек ампликонов ITS1 (из праймера ITS1F) и 6 библиотек ампликонов SSU (из праймера NS31) на системе Genome Sequencer FLX Titanium 454 (454 Life Sciences/Roche Applied Biosystems, Натли, Нью-Джерси, США) проводили в Microsynth (Балгач, Швейцария).
Из общего числа полученных прочтений 70% последовательностей соответствовали полной длине. Все детали биоинформатического анализа и сравнительного анализа приведены в Pagni et al. (2013). Поскольку каждый кластер, сгенерированный DBC454, содержит чтения, идентифицированные в настоящем проекте, и чтения, описанные в Genbank, процедура позволяет отнести чтения, содержащиеся в каждом кластере, к таксономической группе.Кроме того, для дальнейшего статистического анализа можно извлечь количество OTU для каждой таксономической группы на участке, включая Glomeromycetes (полученные с помощью маркера SSU) и Agaricomycetes (полученные с помощью маркера ITS1).
Глава 6: Оценка воздействия: Оценка путей воздействия | Руководство PHA
Ссылки
Александр М. 2000. Старение, биодоступность и переоценка риска от загрязнителей окружающей среды. Environ Sci Technol 34 (20).
АЦДР.1992. Оценка населения на объектах опасных отходов. Атланта: Министерство здравоохранения и социальных служб США. 15 октября 1992 г.
АЦДР. 2000. Указания по заполнению файла экспозиции и демографической структуры (файл EDS). Атланта: Министерство здравоохранения и социальных служб США. Июль 2000 г. (пересмотр).
АЦДР. 2001а. Сводный отчет о семинаре ATSDR по почвенному пайку. Июнь 2000 г., Атланта, Джорджия. Атланта: Министерство здравоохранения и социальных служб США. 20 марта 2001 г. Доступно по адресу: http://www.atsdr.cdc.gov/child/soilpica.html.
АЦДР. 2001б. Краткий отчет для совещания группы экспертов ATSDR о воздействии на племена загрязнителей окружающей среды в растениях. Атланта: Министерство здравоохранения и социальных служб США. 23 марта 2001 г.
АЦДР. 2002. Тематические исследования в области медицины окружающей среды. Экологические триггеры астмы. Атланта: Министерство здравоохранения и социальных служб США. Апрель 2002 г. Доступно по адресу: http://www.atsdr.cdc.gov/csem/asthma/
.Lyman WJ, Reehl WF, Rosenblatt DH, редакторы.1982. Справочник по методам оценки химических свойств. Нью-Йорк: McGraw-Hill Book Co.
Nwosu JU, Harding, AK, Linder G. 1995. Поглощение кадмия и свинца пищевыми культурами, выращенными на илистых суглинках. Bull Environ Contam Toxicol 54:570-8.
Нили В.Б., Брэнсон Д.Р., Блау Г.Э. 1974. Коэффициент распределения для измерения потенциала биоконцентрации органических химических веществ в рыбе. Environ Sci Technol 8:1113-5.
Папа Калифорния 3-й, Бейтс Д.В., Райзенн М.Э. 1995. Влияние загрязнения воздуха твердыми частицами на здоровье: время для переоценки? Environment Health Perspect 103(5):472-80
Самет Дж.М., Доминики Ф., Курьеро ФК, Курсак И., Зегер С.Л.Загрязнение воздуха мелкими частицами и смертность в 20 городах США, 1987-1994 гг. N Engl J Med 343(24):1742-9
Бюро переписи населения США. 2000. Сводный файл 1. Профиль общих демографических характеристик (DP-1) и владения, размера домохозяйства и возраста домохозяина (QT-h3). Доступно по адресу: http://factfinder.census.gov/External
.Van der Zee S, Hoek G, Boezen HM, Schouten JP, van Wijnen JH, Brunekreef B. 1999. Острое воздействие загрязнения воздуха в городах на респираторное здоровье детей с хроническими респираторными симптомами и без них.Оккупируйте Environ Med 56 (12): 802-12.
Что это такое и как это работает?
Тимоти Ф. Слейпер, доктор философии.Директор по экономическому анализу, Центр бизнес-исследований Индианы, Школа бизнеса Келли Университета Индианы
Таня Дж. ХоллАналитик экономических исследований, Центр бизнес-исследований Индианы, Школа бизнеса Келли Университета Индианы
Устойчивое развитие было часто упоминаемой целью предприятий, некоммерческих организаций и правительств в последнее десятилетие, однако измерение степени, в которой организация является устойчивой или стремится к устойчивому росту, может быть затруднена.
Джон Элкингтон стремился измерить устойчивость в середине 1990-х годов, внедрив новую структуру для измерения эффективности в корпоративной Америке. 1 Эта система учета, называемая тройной итоговой строкой (TBL), выходит за рамки традиционных показателей прибыли, рентабельности инвестиций и акционерной стоимости и включает экологические и социальные аспекты. Сосредоточив внимание на комплексных результатах инвестиций, то есть на показателях по взаимосвязанным аспектам прибыли, людей и планеты, тройная итоговая отчетность может стать важным инструментом для поддержки целей устойчивого развития.
Интерес к тройному бухгалтерскому учету растет в коммерческом, некоммерческом и государственном секторах. Многие предприятия и некоммерческие организации приняли концепцию устойчивого развития TBL для оценки своей деятельности, и аналогичный подход получил распространение в правительствах на федеральном, государственном и местном уровнях.
В этой статье рассматривается концепция TBL, объясняется, чем она может быть полезна для бизнеса, политиков и специалистов по экономическому развитию, а также освещаются некоторые текущие примеры применения TBL на практике.
Определение тройного результата
TBL — это система учета, которая включает три аспекта деятельности: социальный, экологический и финансовый. Это отличается от традиционных рамок отчетности, поскольку включает экологические (или природоохранные) и социальные показатели, которым может быть трудно назначить соответствующие средства измерения. Измерения TBL также обычно называют тремя P: люди, планета и прибыль. Мы будем называть их 3P.
Задолго до того, как Элкингтон представил концепцию устойчивости как «тройной результат», защитники окружающей среды боролись с мерами и рамками устойчивости. Академические дисциплины, организованные вокруг устойчивости, увеличились за последние 30 лет. Люди в академических кругах и за их пределами, которые изучали и практиковали устойчивость, согласятся с общим определением Эндрю Савица для TBL. TBL «улавливает суть устойчивости, измеряя влияние деятельности организации на мир… включая его прибыльность и ценность для акционеров, а также его социальный, человеческий и экологический капитал. 2
Хитрость заключается не в определении TBL. Хитрость заключается в измерении.
Расчет TBL
3P не имеют общей единицы измерения. Прибыль измеряется в долларах. В чем измеряется социальный капитал? Как насчет окружающей среды или экологического здоровья? Найти единую единицу измерения — одна из задач.
Некоторые выступают за монетизацию всех аспектов TBL, включая социальное обеспечение или ущерб окружающей среде.Хотя было бы полезно иметь общую единицу — доллары — многие возражают против определения долларовой стоимости водно-болотных угодий или исчезающих видов по чисто философским причинам. Другие ставят под сомнение метод определения правильной цены за утраченные водно-болотные угодья или исчезающие виды.
Другим решением может быть вычисление TBL в терминах индекса. Таким образом, устраняется проблема несовместимости единиц и, при наличии общепринятого метода учета, можно проводить сравнения между объектами, например.g., сравнивая производительность между компаниями, городами, девелоперскими проектами или каким-либо другим эталоном.
Примером индекса, который сравнивает показатели округа и страны по различным компонентам, является индекс инноваций Центра бизнес-исследований Индианы. Однако некоторая субъективность остается даже при использовании индекса. Например, как взвешиваются компоненты индекса? Будет ли каждый «P» иметь равный вес? А как насчет подкомпонентов внутри каждого «P»? Имеют ли они равные веса? Является ли категория людей более важной, чем планета? Кто решает?
В другом варианте можно отказаться от измерения устойчивости с использованием долларов или индекса.Если бы у пользователей TBL хватило смелости, каждая мера устойчивости была бы самостоятельной. Например, мерой будет «акров водно-болотных угодий», а прогресс будет оцениваться на основе создания, разрушения водно-болотных угодий или статус-кво с течением времени. Обратной стороной этого подхода является увеличение количества показателей, которые могут иметь отношение к измерению устойчивости. Пользователь TBL может получить метрическую усталость.
Обсудив трудности с расчетом TBL, мы обращаем внимание на возможные показатели для включения в расчет TBL.После этого мы обсудим, как предприятия и другие организации применяют структуру TBL.
Какие показатели входят в индекс?
Универсального стандартного метода расчета TBL не существует. Также не существует общепринятого стандарта для мер, включающих каждую из трех категорий TBL. Это можно рассматривать как преимущество, поскольку оно позволяет пользователю адаптировать общую структуру к потребностям различных организаций (предприятий или некоммерческих организаций), различных проектов или политик (инвестиции в инфраструктуру или образовательные программы) или различных географических границ (города, региона). или страны).
Как бизнес, так и местное государственное учреждение могут оценивать экологическую устойчивость в одних и тех же условиях, например, уменьшая количество твердых отходов, попадающих на свалки, но местный общественный транспорт может измерять успех с точки зрения пассажиро-миль, в то время как коммерческая автобусная компания будет измерять успех с точки зрения прибыли на акцию. TBL может учесть эти различия.
Кроме того, TBL может быть специфичным для конкретного случая (или проекта) или допускать широкий охват — измерение воздействия через большие географические границы — или узкий географический охват, например небольшой город.TBL для конкретного случая (или проекта) будет измерять влияние конкретного проекта в конкретном месте, например, сообщество, строящее парк. TBL также может применяться к инфраструктурным проектам на уровне штата или энергетической политике на национальном уровне.
Уровень организации, тип проекта и географический охват будут определять многие решения о том, какие меры следует включить. При этом набор мер в конечном итоге будет определяться заинтересованными сторонами и профильными экспертами, а также возможностью сбора необходимых данных.Несмотря на то, что имеется обширная литература о соответствующих мерах, которые следует использовать для обеспечения устойчивости на уровне штата или на национальном уровне, в конечном итоге наличие данных будет определять расчеты TBL. Ниже представлены многие из традиционных мер устойчивости, проверенных академическими рассуждениями.
Экономические меры
Экономические переменные должны быть переменными, связанными с прибылью и потоком денег. Он может рассматривать доходы или расходы, налоги, факторы делового климата, занятость и факторы разнообразия бизнеса.Конкретные примеры включают:
- Личный доход
- Стоимость неполной занятости
- Текучесть предприятия
- Установочные размеры
- Рост занятости
- Распределение занятости по секторам
- Доля фирм в каждом секторе
- Доходы по секторам, вносящие вклад в валовой государственный продукт
Экологические меры
Переменные окружающей среды должны представлять измерения природных ресурсов и отражать потенциальное влияние на их жизнеспособность.Он может включать качество воздуха и воды, потребление энергии, природные ресурсы, твердые и токсичные отходы и землепользование/почвенный покров. В идеале наличие долгосрочных трендов для каждой из переменных окружающей среды помогло бы организациям определить влияние проекта или политики на регион. Конкретные примеры включают:
- Концентрация диоксида серы
- Концентрация оксидов азота
- Отдельные приоритетные загрязнители
- Чрезмерное количество питательных веществ
- Потребление электроэнергии
- Потребление ископаемого топлива
- Управление твердыми отходами
- Управление опасными отходами
- Изменение землепользования/почвенного покрова
Социальные меры
Социальные переменные относятся к социальным параметрам сообщества или региона и могут включать показатели образования, справедливости и доступа к социальным ресурсам, здоровья и благополучия, качества жизни и социального капитала.Примеры, перечисленные ниже, представляют собой небольшой фрагмент потенциальных переменных:
- Уровень безработицы
- Уровень участия женщин в рабочей силе
- Средний доход домохозяйства
- Относительная бедность
- Процент населения с высшим образованием или сертификатом
- Среднее время в пути
- Насильственные преступления на душу населения
- Ожидаемая продолжительность жизни с поправкой на здоровье
Данные по многим из этих мер собираются на уровне штата и на национальном уровне, но также доступны на местном уровне или на уровне сообщества.Многие из них подходят для использования сообществом при создании TBL. Однако по мере сужения географического охвата и характера проекта набор соответствующих мер может меняться. Для местных или общественных проектов показатели успеха TBL лучше всего определять на местном уровне.
Существует несколько аналогичных подходов к обеспечению участия заинтересованных сторон и их вклада в разработку структуры TBL: разработка матрицы решений для включения общественных предпочтений в планирование проекта и принятие решений, 3 использование «формата повествования» для привлечения акционеров к участию и комплексному проекту. оценка, 4 , а заинтересованные стороны ранжируют и взвешивают компоненты системы устойчивого развития в соответствии с приоритетами сообщества. 5 Например, сообщество может считать важной мерой успеха программы развития предпринимательства количество компаний, принадлежащих женщинам, созданных за пятилетний период. В конечном счете, организация будет нести ответственность за разработку окончательного набора мер, применимых к поставленной задаче.
Варианты измерения тройного результата
Применение TBL предприятиями, некоммерческими организациями и правительствами мотивировано принципами экономической, экологической и социальной устойчивости, но различается в отношении того, как они измеряют три категории результатов.Сторонники, разработавшие и применившие системы оценки устойчивости, такие как TBL, столкнулись со многими проблемами, главная из которых заключалась в том, как составить всеобъемлющий и содержательный индекс и как определить подходящие данные для переменных, составляющих индекс.
Индикатор подлинного прогресса (GPI), например, состоит из 25 переменных, охватывающих экономические, социальные и экологические факторы. Эти переменные конвертируются в денежные единицы и суммируются в единую меру, выраженную в долларах. 6 Миннесота разработала свой собственный индикатор прогресса, состоящий из 42 переменных, которые сосредоточены на целях здоровой экономики и оценивают прогресс в достижении этих целей. 7
Существует большое количество литературы по комплексной оценке 8 и мерам устойчивости, которые выросли из дисциплин, измеряющих воздействие на окружающую среду. Они не ограничены строгой экономической теорией измерения изменений в общественном благосостоянии. 9 Исследователи экологической политики утверждают, что три категории — экономическая, социальная и экологическая — должны быть объединены, чтобы увидеть полную картину последствий, которые могут иметь регулирование, политика или проект экономического развития, и оценить варианты политики и компромиссы.
Кто использует тройной результат?
Предприятия, некоммерческие и государственные организации могут использовать TBL.
Предприятия
TBL и его основная ценность устойчивости стали привлекательными в деловом мире из-за накопления неподтвержденных данных о большей долгосрочной прибыльности. Например, сокращение отходов упаковки также может снизить затраты. Примерами таких подходов являются фирмы General Electric, Unilever, Proctor and Gamble, 3M и Cascade Engineering. 10 Хотя у этих компаний нет TBL на основе индекса, можно увидеть, как они измеряют устойчивость, используя концепцию TBL. Cascade Engineering, например, частная фирма, которой не нужно подавать подробные финансовые документы публичных компаний, определила следующие переменные для своей системы показателей TBL:
- Экономический
- Социальный
- Среднее количество часов обучения на одного сотрудника
- От социального обеспечения к сохранению карьеры
- Благотворительные взносы
- Защита окружающей среды/безопасность
- Частота инцидентов, связанных с безопасностью
- Коэффициент потерянных/ограниченных рабочих дней
- Доллары продаж за киловатт-час
- Выбросы парниковых газов
- Использование бывших в употреблении и промышленных переработанных материалов
- Расход воды
- Количество отходов на свалку
Некоммерческие организации
Многие некоммерческие организации приняли TBL, а некоторые сотрудничают с частными фирмами для решения широких вопросов устойчивости, которые затрагивают взаимных заинтересованных сторон.Компании признают, что сотрудничество с некоммерческими организациями имеет смысл для бизнеса, особенно с теми некоммерческими организациями, которые преследуют цели экономического процветания, социального благополучия и защиты окружающей среды. 11
Фонд Форда финансировал исследования, в которых использовались варианты TBL для измерения результатов программ повышения благосостояния в десятках сельских регионов США. 12 Другим примером является RSF Social Finance, 13 , некоммерческая организация, которая уделяет особое внимание тому, как их инвестиции улучшают все три категории TBL.Хотя RSF использует оригинальный подход к концепции TBL, можно увидеть, как TBL можно адаптировать практически к любой организации. Их подход включает следующее:
- Продовольствие и сельское хозяйство (экономический): Изучите новые экономические модели, которые поддерживают устойчивое производство продуктов питания и сельского хозяйства, повышая при этом осведомленность общественности о ценности органического и биодинамического земледелия.
- Экологическое управление (окружающая среда): Предоставление финансирования организациям и проектам, посвященным поддержанию, восстановлению и сохранению экосистем Земли, особенно комплексным, системным и культурно значимым подходам.
- Образование и искусство (социальный): финансирование образовательных и художественных проектов, которые являются целостными и терапевтическими.
Правительство
Государственные, региональные и местные органы власти все чаще используют TBL и аналогичные системы оценки устойчивости в качестве инструментов для принятия решений и мониторинга эффективности. Мэриленд, Миннесота, Вермонт, Юта, район залива Сан-Франциско и район северо-восточного Огайо провели анализ с использованием TBL или аналогичной системы устойчивого развития.
Разработчики политики используют эти рамки оценки устойчивости, чтобы решить, какие действия они должны или не должны предпринимать, чтобы сделать общество более устойчивым. Разработчики политики хотят знать причинно-следственную связь между действиями — проектами или политикой — и движут ли результаты общество к устойчивости или от нее. Штат Мэриленд, например, использует смешанную структуру GPI-TBL для сравнения инициатив — например, инвестирования в чистую энергию — с базовым планом «ничего не делать» или с другими вариантами политики. 14
На международном уровне Европейский союз использует комплексную оценку для выявления «вероятных положительных и отрицательных последствий предлагаемых политических действий, что позволяет выносить обоснованные политические суждения о предложении и определять компромиссы в достижении конкурирующих целей». 15 Руководство ЕС само по себе подвергалось критике и претерпело несколько этапов усовершенствования. 16 Процесс уточнения руководящих принципов демонстрирует как прозрачность процесса, так и приверженность ЕС комплексной оценке.
Инициативы регионального экономического развития
Концепция тройного результата может использоваться сообществами на региональном уровне для поощрения экономического роста на устойчивой основе. Это требует повышенного уровня сотрудничества между бизнесом, некоммерческими организациями, правительствами и гражданами региона. Следующие примеры в Соединенных Штатах демонстрируют различные способы использования концепции TBL для устойчивого роста экономической базы региона.
Кливленд, Огайо
В 2009 году мэр Кливленда созвал саммит «Устойчивый Кливленд 2019» (SC2019), чтобы собрать вместе сотни людей, заинтересованных в применении принципов устойчивого развития к моделированию местной экономики. 17 SC2019 — это 10-летняя инициатива по созданию устойчивой экономики в Кливленде с упором на концепцию, подобную TBL. Город использует четыре ключевые области для измерения устойчивости: личная и социальная среда, природная среда, застроенная среда (т.г., инфраструктура и модели роста городов) и деловая среда. Каждая ключевая область имеет шесть целей. На данный момент конкретные показатели измерения еще не полностью разработаны; тем не менее, городские власти стремятся создать информационную панель, которую можно было бы объединить для создания индекса общего успеха проекта. Эта информационная панель позволит быстро оценивать ход выполнения SC2019 из года в год.
Гранд-Рапидс, Мичиган и его окрестности
В 2005 году регион Гранд-Рапидс создал первое в стране «Партнерство по устойчивому развитию сообщества» для разработки дорожной карты, которая приведет Гранд-Рапидс к устойчивому развитию.В регионе используются 14 основных показателей, связанных с качеством жизни в регионе и факторами окружающей среды, для определения прогресса, достигнутого в направлении устойчивого развития. Вместо создания индекса для каждого индикатора были установлены целевые показатели. Более подробную информацию о метриках, используемых для каждого индикатора, можно найти в их отчете TBL. 18 Ниже приведены краткие пояснения переменных, используемых для измерения их TBL.
- Качество окружающей среды
- Отходы: тенденции переработки, мусор и дворовые отходы
- Энергетика: потребление энергии, потребление природного газа и использование альтернативных видов топлива
- Вода: потребление воды
- Качество воздуха: инвентаризация выбросов токсичных веществ и количество дней действия озонового загрязнения воздуха
- Застроенная среда: количество зарегистрированных и сертифицированных проектов LEED
- Землепользование и естественная среда обитания: инвентаризация землепользования и лесного полога
- Транспорт: общественный транспорт
- Экономическое процветание
- Личный доход: личный доход на душу населения
- Безработица: уровень безработицы
- Редевелопмент, реинвестирование и рабочие места: результаты инвестиций в редевелопмент старых месторождений и создание рабочих мест
- Конкурентоспособность знаний: рейтинг сторонних отчетов U.С. регионов
- Социальный капитал и акционерный капитал
- Охрана и безопасность: криминальная статистика
- Уровень образования: уровни получения степени
- Здоровье и благополучие: тенденции младенческой смертности и уровня свинца в крови
- Качество жизни: домовладение, бедность, тенденции снижения цен и бесплатных обедов
- Столица сообщества: 211 обращений за помощью, участие избирателей, население и этническая принадлежность
Резюме
Концепция Triple Bottom Line, разработанная Джоном Элкингтоном, изменила способ, которым предприятия, некоммерческие организации и правительства оценивают устойчивость и эффективность проектов или политики.Помимо измерения устойчивости по трем направлениям — люди, планета и прибыль — гибкость TBL позволяет организациям применять эту концепцию в соответствии с их конкретными потребностями.
Есть проблемы с применением TBL на практике.
