
Тестирование на тему «Клеточный цикл митоз»
Клеточный цикл. Митоз.
1. Назовите фазу митоза животной клетки, во время которой происходит расхождение хроматид каждой хромосомы к разным полюсам веретена деления за счет взаимодействия микротрубочек веретена деления друг с другом и с белками первичной перетяжки хромосом.
1) анафаза 2) телофаза 3) метафаза 4) профаза 5) интерфаза
2.Клетки многих представителей одной из систематических групп животных делятся путем ми-
тоза без разрушения ядерной оболочки. Назовите эту группу животных.
простейшие 2) кишечнополостные 3) насекомые 4) кольчатые черви
З. Назовите фазу митоза животной клетки, во время которой в клетке происходит формирование веретена деления, расхождение центриолей к противоположным участкам клетки и формирование полюсов веретена деления, спирализация дНК, исчезновение ядрышка, разрушение ядерной оболочки и формирование компактных, видимых в световой микроскоп хромосом.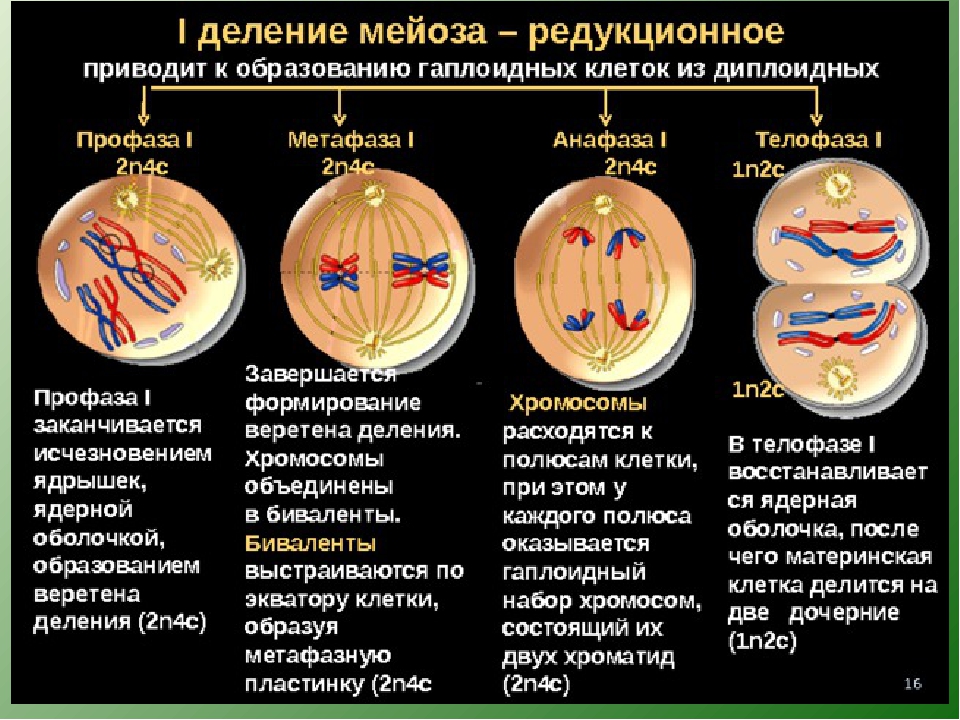
1) анафаза 2) профаза 3) телофаза 4) метафаза
4. Назовите фазу клеточного (жизненного) цикла, во время которой происходит редупликация ДНК, в результате чего каждая хромосома состоит из двух хроматид — одинаковых копий материнской хромосомы.
1) профаза 2) анафаза 3) интерфаза 4) метафаза 5) телофаза
5. Укажите клетки, которые после митоза не специализируются, не дифференцируются, а сразу делятся.
1) клетки печени, кожи 2) нервные клетки 3) бластомеры — клетки зародыша животных
6. Одна из хромосом делящейся диплоидной клетки женщины имеет определенный размер, окраску, расположение центромеры и характерный для этой хромосомы набор генов. Сколько еще похожих на нее по этим признакам хромосом находится в этой же клетке?
1)1 2)2 3)4 4) ни одной 5) 6—8 6)45
7. Сколько молекул ДНК находится в каждой хромосоме во время анафазы митоза?
1)1 2)2 3)4
8*. Назовите период интерфазы, во время которого каждая хромосома состоит из двух хроматид и
Назовите период интерфазы, во время которого каждая хромосома состоит из двух хроматид и
в клетке происходит преимущественно синтез РНК и белков, необходимых для ее деления.
1) синтетический 2) пресинтетический 3) постсинтетический
9. Сколько молекул дНК находится в каждой хроматиде во время профазы митоза?
1)1 2)2 3)4
10. В делящейся животной клетке имеются два участка, называемые «полюсами веретена деления«, в состав которых входят определенные клеточные структуры. Назовите эти структуры.
1) центриоли 2) полости аппарата Гольджи 3) вакуоли 4) ядрышки
11*. Назовите систематическую группу эукариотических организмов, при делении клеток которых наблюдается следующая особенность: цитоплазма делится путем образования перетяжки. Перетяжка формируется следующим образом: под наружной плазматической мембраной образуется кольцо из сократительных белков, взаимодействие которых друг с другом ведет к сближению и последующему соединению противоположных участков наружной плазматической мембраны, после чего дочерние клетки оказываются обособленными друг от друга.
1) животные 2) растения 3)грибы
12. Сколько молекул ДНК находится в каждой хромосоме во время телофазы митоза?
1)1 2)2 3) 4
13. В каком ответе правильно указана последовательность фаз митоза?
1) метафаза, профаза, телофаза, анафаза
2) профаза, анафаза, телофаза, метафаза
3) телофаза, метафаза, анафаза, профаза
4) профаза, метафаза, анафаза, телофаза
14. Соматическая клетка кожи человека содержит 46 хромосом. Сколько хромосом будет содержаться в каждой из ее дочерних клеток, образовавшихся в результате двух митотических делений этой соматической клетки?
1)23 2) 46 3)92 4) 138 5) 184
15. У большинства организмов негомологичные друг другу хромосомы, находящиеся в метафазе митоза, отличаются друг от друга по ряду признаков. Найдите их среди ответов и укажите признак, НЕ являющийся отличительной особенностью.
1) длина 2) толщина 3) соотношение плеч 4) положение первичной перетяжки
5) наличие или отсутствие вторичной перетяжки 6) наличие первичной перетяжки
16. Назовите структуры, из которых состоит один из важнейших компонентов митотического аппарата эукариотической клетки — веретено деления.
1) актиновые волокна (микрофиламенты) 2) миозиновые волокна
3) микротрубочки 4) миофибриллы
5) микроворсинки 6) коллагеновые волокна
17. Назовите участок хромосомы, в области которого у большинства эукариот к хромосомам прикрепляются микротрубочки веретена деления.
1) короткое плечо 2) длинное плечо 3) первичная перетяжка 4) вторичная перетяжка
18*. Назовите систематическую группу эукариотических организмов, при делении клеток которых наблюдается следующая особенность: цитоплазма делится путем образования перемычки.
1) животные 2) растения 3) грибы
19. Сколько хроматид входит в состав каждой хромосомы в конце интерфазы непосредственно перед митозом?
1) 1 2)2 3)4
20. Какая из фаз митоза является последней?
1) метафаза 2) профаза 3) телофаза 4) анафаза
21*. Назовите период интерфазы, во время которого происходит редупликация молекул ДНК.
1) синтетический 2) пресинтетический 3) постсинтетический
22. Одна из десяти хромосом делящейся гаплоидной клетки жгутикового простейшего имеет определенный размер, окраску, расположение центромеры и характерный для этой хромосомы набор генов. Сколько еще похожих на нее по этим признакам хромосом находится в этой же клетке?
1)1 2) 2 3) 4 4)6-8 5) ни одной 6) 9
23. Клетка печени обезьяны содержит 48 хромосом. Сколько хромосом будет содержаться в каждой из ее дочерних клеток, образовавшихся в результате трех митотических делений этой клетки печени?
Клетка печени обезьяны содержит 48 хромосом. Сколько хромосом будет содержаться в каждой из ее дочерних клеток, образовавшихся в результате трех митотических делений этой клетки печени?
1)6 2)12 3)24 4) 48 5) 72 6) 96 7) 144
24. Хромосома делящейся и та же хромосома неделящейся эукариотической клетки отличаются друг от друга по ряду признаков. Найдите их среди ответов и укажите признак, по которому они НЕ отличаются друг от друга.
1) длина 2) толщина 3) плотность упаковки в пространстве 4) набор генов
25. Назовите фазу митоза, в которой происходит уплотнение (конденсация) хромосом, исчезновение ядрышка и разрушение ядерной оболочки.
1) профаза 2) телофаза З) интерфаза 4)анафаза 5) метафаза
26. В одну из фаз митоза хромосомы перемещаются к полюсу веретена деления одним из своих участков вперед. Назовите этот участок хромосомы.
1) короткое плечо 2) длинное плечо З) первичная перетяжка 4) вторичная перетяжка
27. Назовите форму, которую имеет большинство хромосом эукариот в анафазу митоза.
Назовите форму, которую имеет большинство хромосом эукариот в анафазу митоза.
1) кольцо 2) шар З) трубка 4) шпилька 5) Х-образная
28. Назовите структуры, которые во время анафазы митоза подходят к одному и тому же полюсу веретена деления клетки.
1) только гомологичные друг другу хромосомы
2) только негомологичные друг другу хромосомы
З) только хроматиды негомологичных хромосом, с момента расхождения называемые соответствующими хромосомами
4) только хроматиды гомологичных хромосом, с момента расхождения называемые соответствующими хромосомами
5) хроматиды гомологичных и негомологичных хромосом, с момента расхождения называемые соответствующими хромосомами
29. Назовите фазу митоза животной клетки, во время которой происходит деспирализация хромосом, формирование ядрышка и ядерной оболочки, деление цитоплазмы (цитокинез) и в конечном итоге образование двух дочерних клеток — одинаковых копий материнской клетки.
1) профаза 2) телофаза З) анафаза 4) метафаза 5) интерфаза
30. На образование, каких структур клетки влияют некоторые яды, например колхицин, останавливающие митоз в метафазе?
1) актиновые волокна (микрофиламенты) 2) микротрубочки 3) миофибриллы
31*. Назовите одну из особенностей, которая отличает митоз растительной клетки от митоза животной клетки.
1) формируются компактные короткие и толстые хромосомы
2) образуется веретено деления из микротрубочек
З) в начале деления ядерная оболочка разрушается на множество мелких пузырьков
4) цитоплазма клетки делится путем формирования перегородки из пузырьков комплекса Гольджи
32. Назовите фазу митоза животной клетки, во время которой хромосомы выстраиваются в плоскости экватора веретена деления и заканчивается прикрепление к их центромерам микротрубочек веретена деления.
1)анафаза 2) профаза 3) метафаза 4) телофаза 5) интерфаза
33. Назовите структуры, которые, будучи ранее соединенными, отходят друг от друга в разные стороны во время анафазы митоза.
1) гомологичные друг другу хромосомы
2) негомологичные друг другу хромосомы
З) только хроматиды негомологичных хромосом, с момента расхождения называемые соответствующими хромосомами
4) только хроматиды гомологичных хромосом, с момента расхождения называемые соответствующими хромосомами
5) хроматиды гомологичных и негомологичных хромосом, с момента расхождения называемые соответствующими хромосомами
34. При большом увеличении микроскопа видна животная клетка, в центре которой в одной плоскости расположены интенсивно окрашенные структуры — хромосомы, которые имеют вид шпилек, обращенных согнутыми участками к середине клетки, а свободными — к периферии. Эта клетка находится в одной из фаз митоза. Назовите эту фазу митоза.
Эта клетка находится в одной из фаз митоза. Назовите эту фазу митоза.
1) профаза 2) телофаза З) интерфаза 4) анафаза 5) метафаза
35. Какие структурные компоненты животной клетки входят в состав полюсов веретена деления, к которым во время анафазы митоза расходятся дочерние хромосомы — хроматиды метафазных хромосом?
1) митохондрии 2) лизосомы 3) центриоли 4) аппарат Гольджи
36*. Митоз животной клетки по ряду особенностей отличается от митоза растительной клетки. Найдите эти особенности среди ответов и укажите признак, который к числу таких особенностей НЕ относится.
1) цитоплазма клетки делится путем формирования перетяжки
2) полюса веретена деления содержат центриоли, участвующие в расхождении хромосом
З) формируются компактные короткие и толстые хромосомы
37. Представьте, что вы изучаете фотографии делящихся клеток животных, относящихся к разным биологическим видам. делящиеся клетки находятся в метафазе митоза. Назовите основной признак, по которому метафазные клетки животных разных видов отличаются друг от друга.
делящиеся клетки находятся в метафазе митоза. Назовите основной признак, по которому метафазные клетки животных разных видов отличаются друг от друга.
1) число хромосом 2) расположение хромосом 3) форма хромосом
4) размер хромосом 5) форма клеток
38. При большом увеличении микроскопа среди неделящихся клеток животного обнаружена клетка с двумя маленькими клубочками, состоящими из интенсивно окрашенных нитей — хромосом. В какой фазе митоза находится эта клетка?
1) метафаза 2) интерфаза 3) телофаза 4) профаза 5) анафаза
39. Укажите клетки, которые после митоза приобретают характерные им особенности химического состава и строения, выполняют характерные дня них функции и никогда больше не делятся.
1) клетки печени 2) нервные клетки 3) бластомеры — клетки зародыша животных
40*. Назовите систематическую группу эукариотических организмов, при делении клеток которых наблюдается следующая особенность: деление ядер не сопровождается делением цитоплазмы клетки. В результате этого формируются тонкие и длинные многоядерные клетки.
В результате этого формируются тонкие и длинные многоядерные клетки.
1) животные 2) растения 3) грибы
41. В ходе редупликации ДНК из одной исходной (материнской) хромосомы и свободных нуклеотидов образуются две новые структуры, для обозначения которых используют специальный термин. Назовите этот термин.
1) гомологичные хромосомы 2) негомологичные хромосомы 3) хроматиды
42. При большом увеличении микроскопа среди неделящихся клеток животного обнаружена клетка, в которой в разных ее концах находятся две небольшие фигуры в виде звезд, состоящих из интенсивно окрашенных структур — хромосом. В какой фазе митоза находится эта клетка?
1) метафаза 2) интерфаза 3) телофаза 4) профаза 5) анафаза
43. Назовите форму, которую имеет большинство хромосом человека в метафазу митоза.
1) кольцо 2) шар 3) трубка 4) шпилька 5) Х-образная
44. Какова в среднем продолжительность митоза?
Какова в среднем продолжительность митоза?
1) 1—2 минуты 2) 1—2 часа 3) 1—2 суток
45. Каким термином называется совокупность признаков хромосомного набора (число, размер и форма хромосом), характерных для того или иного биологического вида животных, растений, грибов?
1) генотип 2) фенотип 3) кариотип 4) геном 5) кариоплазма
46. Парные, или гомологичные друг другу, хромосомы одинаковы по строению и набору генов. Каким термином называется в клетке набор хромосом, в котором каждая хромосома имеет гомологичную себе хромосому?
1) гаплоидный 2) диплоидный
Ответы по теме: «Клеточный цикл. Митоз».
Митоз — это… Что такое Митоз?
Фазы митозаМито́з (греч. μιτος — нить) — непрямое деление клетки, наиболее распространенный способ репродукции эукариотических клеток. Биологическое значение митоза состоит в строго одинаковом распределении хромосом между дочерними ядрами, что обеспечивает образование генетически идентичных дочерних клеток и сохраняет преемственность в ряду клеточных поколений. [1]
[1]
Митоз — один из фундаментальных процессов онтогенеза. Митотическое деление обеспечивает рост многоклеточных эукариот за счёт увеличения популяции тканевых клеток. В результате митотического деления клеток меристем увеличиваются тканевые популяции растительных клеток. Дробление оплодотворённого яйца и рост большинства тканей у животных также происходит путём митотических делений.[2]
На основании морфологических особенностей митоз условно подразделяется на стадии: профазу, прометафазу, метафазу, анафазу, телофазу. Первые описания митотических фаз и установление их последовательности были предприняты в 70—80-х годах XIX века. В конце 1870-х — начале 1880-х годов немецкий гистолог Вальтер Флемминг для обозначения процесса непрямого деления клетки ввёл термин «митоз».[3]
Продолжительность митоза в среднем составляет 1—2 часа.[1][4] В клетках животных митоз, как правило, длится 30—60 минут, а в растительных — 2—3 часа. [5] Клетки человека за 70 лет суммарно претерпевают порядка 1014 клеточных делений.[6]
[5] Клетки человека за 70 лет суммарно претерпевают порядка 1014 клеточных делений.[6]
История исследования
Первые неполные описания, касающиеся поведения и изменения ядер в делящихся клетках, встречаются в работах учёных начала 1870-х годов. В работе русского ботаника Э. Руссова, датируемой 1872 годом, отчётливо описаны и изображены метафазные и анафазные пластинки, состоящие из отдельных хромосом.[7] Годом позже немецкий зоолог А. Шнейдер ещё более отчётливо и последовательно, но, конечно, не совсем полно описал митотическое деление на примере дробящихся яиц прямокишечной турбеллярии Mesostomum. В его работе, в сущности, описаны и проиллюстрированы в правильной последовательности основные фазы митоза: профаза, метафаза, анафаза (ранняя и поздняя). В 1874 году московский ботаник И. Д. Чистяков также наблюдал отдельные фазы клеточного деления в спорах плаунов и хвощей. Несмотря на первые успехи ни Руссову, ни Шнейдеру, ни Чистякову не удалось дать чёткое и последовательное описание митотического деления. [8]
[8]
В 1875 году вышли работы, содержащие более детальные описания митозов. О. Бючли дал описание цитологических картин в дробящихся яйцах круглых червей и моллюсков и в сперматогенных клетках насекомых. Э. Страсбургер исследовал митотическое деление в клетках зелёной водоросли спирогиры, в материнских клетках пыльцы лука и в материнских споровых клетках плауна. Ссылаясь на работу О. Бючли и основываясь на собственных исследованиях, Э. Страсбургер обратил внимание на единство процессов клеточного деления в растительных и животных клетках.[9]
К концу 1878 — началу 1879 года появились подробные работы В. Шлейхера (о делении хрящевых клеток амфибий), В. Флемминга (о размножении клеток в разных тканях саламандры и её личинок), П. И. Перемежко (о делении клеток в эпидермисе личинок тритона). В своей работе в 1879 году Шлейхер предложил термин «кариокинез» для обозначения сложных процессов клеточного деления, подразумевая перемещения составных частей ядра.[10] Вальтер Флемминг впервые для обозначения непрямого деления клетки ввёл термин «митоз», который впоследствии стал общепринятым. [3] Также Флеммингу принадлежит окончательная формулировка определения митоза как циклического процесса, завершающегося разделением хромосом между дочерними клетками.[11]
[3] Также Флеммингу принадлежит окончательная формулировка определения митоза как циклического процесса, завершающегося разделением хромосом между дочерними клетками.[11]
-
Деление клеток по Э. Руссову (1872)
-
Деление клеток по Э. Страсбургеру (1875)
-
Деление клеток по В. Флеммингу (1882)
-
Деление клеток по Э. Б. Уилсону (1900)
В 1880 году О. В. Баранецкий установил спиральное строение хромосом. В ходе дальнейших исследований были развиты представления о спирализации и деспирализации хромосом во время митотического цикла.[11] В начале 1900-х годов хромосомы были идентифицированы в качестве носителей наследственной информации, что в дальнейшем дало объяснение биологической роли митоза, заключающейся в образовании генетически идентичных дочерних клеток.
В 1970-х годах началась расшифровка и детальное изучение регуляторов митотического деления,[12] благодаря серии экспериментов по слиянию клеток, находящихся на разных этапах клеточного цикла. В тех опытах, когда клетку в М-фазе объединяли с клеткой, находящейся в любой из стадий интерфазы (S или G2), интерфазные клетки переходили в митотическое состояние (начиналась конденсация хромосом и распадалась ядерная оболочка).[13] В итоге был сделан вывод, что в цитоплазме митотической клетки присутствует фактор (или факторы), стимулирующий митоз,[14] или, иначе, М-стимулирующий фактор (МСФ, от англ. M-phase-promoting factor, MPF).[15]
В тех опытах, когда клетку в М-фазе объединяли с клеткой, находящейся в любой из стадий интерфазы (S или G2), интерфазные клетки переходили в митотическое состояние (начиналась конденсация хромосом и распадалась ядерная оболочка).[13] В итоге был сделан вывод, что в цитоплазме митотической клетки присутствует фактор (или факторы), стимулирующий митоз,[14] или, иначе, М-стимулирующий фактор (МСФ, от англ. M-phase-promoting factor, MPF).[15]
Впервые «фактор стимуляции митоза» был открыт в зрелых неоплодотворенных яйцах шпорцевой лягушки, находящихся в М-фазе клеточного цикла. Цитоплазма такого яйца, инъецированная в ооцит, приводила к преждевременному переходу в М-фазу и к началу созревания ооцита (первоначально сокращение MPF означало Maturation Promoting Factor, что переводится как «фактор, способствующий созреванию»). В ходе дальнейших экспериментов были установлены универсальное значение и вместе с тем высокая степень консервативности «фактора стимуляции митоза»: экстракты, приготовленные из митотических клеток весьма разнообразных организмов (млекопитающих, морских ежей, моллюсков, дрожжей), при введении в ооциты шпорцевой лягушки переводили их в М-фазу. [16]
[16]
В ходе последующих исследований выяснилось, что фактор, стимулирующий митоз, представляет собой гетеродимерный комплекс, состоящий из белка циклина и зависимой от циклина протеинкиназы. Циклин является регуляторным белком и обнаруживается у всех эукариот. Его концентрация периодически возрастает в течение клеточного цикла, достигая максимума в метафазе митоза. С началом анафазы наблюдается резкое сокращение концентрации циклина, вследствие его расщепления с помощью сложных белковых протеолитических комплексов — протеосом. Зависимая от циклина протеинкиназа представляет собой фермент (фосфорилазу), модифицирующий белки за счёт переноса фосфатной группы от АТФ на аминокислоты серин и треонин. Таким образом с установления роли и структуры основного регулятора митотического деления начались исследования тонких регуляторных механизмов митоза, которые продолжаются до настоящего времени.
Типы митоза
Выработка единой типологии и классификации митозов осложняется целым спектром признаков,[~ 1] которые в различных комбинациях создают разнообразие и неоднородность картин митотического деления. При этом отдельные варианты классификации, разработанные применительно к одним таксонам, являются неприемлемыми в отношении других, поскольку не учитывают специфики их митозов. Например, отдельные варианты классификации митозов, свойственных животным или растительным организмам, оказываются неприемлемыми для водорослей.[17]
При этом отдельные варианты классификации, разработанные применительно к одним таксонам, являются неприемлемыми в отношении других, поскольку не учитывают специфики их митозов. Например, отдельные варианты классификации митозов, свойственных животным или растительным организмам, оказываются неприемлемыми для водорослей.[17]
Одним из ключевых признаков, лежащих в основе различных типологий и классификаций митотического деления, является поведение ядерной оболочки. Если образование веретена и само митотическое деление протекает внутри ядра без разрушения ядерной оболочки, то такой тип митоза называют закрытым. Митоз с распадом ядерной оболочки, соответственно, называется открытым, а митоз с распадом оболочки только на полюсах веретена, с образованием «полярных окон» — полузакрытым.[18][17]
Ещё одним характерным признаком является тип симметрии митотического веретена. При плевромитозе веретено деления билатерально симметрично либо асимметрично и состоит, как правило, из двух полуверетён, располагающихся в метафазе-анафазе под углом друг к другу. Для категории ортомитозов характерна биполярная симметрия веретена деления, а в метафазе зачастую наблюдается различимая экваториальная пластинка.[18]
Для категории ортомитозов характерна биполярная симметрия веретена деления, а в метафазе зачастую наблюдается различимая экваториальная пластинка.[18]
В рамках обозначенных признаков наиболее многочисленным является типичный открытый ортомитоз, на примере которого ниже рассматриваются принципы и стадии митотического деления. Данный тип митоза характерен для животных, высших растений и некоторых простейших.[19]
Варианты классификации митозов
|
7 типов митоза простейших[18]:
|
6 типов митоза водорослей[17]:
|
Основные типы митоза у водорослей: 1. Закрытый центрический; Закрытый центрический;2. Закрытый ацентрический; 3. Полузакрытый центрический; 4. Полузакрытый ацентрический; 5. Открытый центрический; 6. Открытый ацентрический |
Происхождение и эволюция митоза
| Информация в этом разделе устарела. Вы можете помочь проекту, обновив его и убрав после этого данный шаблон. |
Условные обозначения: 1. Закрытый эвгленоидный митоз; 2. Закрытый внутриядерный плевромитоз;
3. Закрытый внеядерный плевромитоз; 4. Полузакрытый плевромитоз; 5. Закрытый внутриядерный ортомитоз;
6. Полузакрытый ортомитоз; 7. Открытый ортомитоз;
Предполагается, что сложный митотический процесс высших организмов развивался постепенно из механизмов деления прокариот.[20] В клетках прокариот пара нуклеоидов после репликации сохраняет связь с плазматической мембраной посредством мезосомы. Расхождение реплицированных нуклеоидов происходит в результате роста мембраны между ними.
Расхождение реплицированных нуклеоидов происходит в результате роста мембраны между ними.
У многих простейших эукариот митоз также остался процессом, связанным с мембраной, однако теперь уже не плазматической, а ядерной.[21] Возможно, в связи с увеличением размера и числа хромосом, структура типа мезосомы разделилась на два элемента: ЦОМТ на ядерной оболочке и кинетохор на хромосоме. Для соединения данных структур между собой в процессе эволюции развилась промежуточная система микротрубочек. В рамках данного представления, наиболее древним и примитивным считается закрытый внутриядерный плевромитоз. Сегрегация хромосом при этом происходит путём расхождения ЦОМТ, к которым хромосомы крепятся посредством микротрубочек. В свою очередь ЦОМТ закреплены на ядерной оболочке и расходятся за счёт роста ядерной мембраны между ними.[22]
От разных вариантов закрытого внутриядерного плевромитоза, вероятно, берут своё начало несколько параллельных эволюционных линий. [22] В качестве эволюционно прогрессивных признаков при этом рассматриваются: распад ядерной оболочки во время митоза; переход ЦОМТ из ядра в цитоплазму; образование биполярного веретена; усиление спирализации хромосом; формирование экваториальной пластинки в метафазе. Таким образом, эволюция митотического деления идёт в направлении от закрытого внутриядерного плевромитоза к открытому ортомитозу.[23]
[22] В качестве эволюционно прогрессивных признаков при этом рассматриваются: распад ядерной оболочки во время митоза; переход ЦОМТ из ядра в цитоплазму; образование биполярного веретена; усиление спирализации хромосом; формирование экваториальной пластинки в метафазе. Таким образом, эволюция митотического деления идёт в направлении от закрытого внутриядерного плевромитоза к открытому ортомитозу.[23]
Регуляция митоза
Основными регулято́рными механизмами митоза являются процессы фосфорилирования и протеолиза.[24] Обратимые реакции фосфорилирования и дефосфорилирования обеспечивают протекание обратимых событий митоза, таких как сборка/распад веретена деления или распад/восстановление ядерной оболочки. Протеолиз лежит в основе необратимых событий митоза, таких как разделение сестринских хроматид в анафазе или разрушение митотических циклинов на поздних стадиях митоза.
Контрольные точки
Общая схема регуляции митоза[25]Рассматривая вопрос регуляции митоза, можно условно выделить два периода митотического деления: от начала профазы до анафазы, и далее, от анафазы до конца телофазы. [26] Каждый из двух обозначенных периодов начинается с прохождения контрольной точки клеточного цикла.
[26] Каждый из двух обозначенных периодов начинается с прохождения контрольной точки клеточного цикла.
Первой контрольной точкой является переход из фазы G2 к M-фазе. Главным условием для преодоления контрольной точки G2/M являестя завершённая репликация ДНК, так как старт митотического деления блокируется у большинства эукариот при повреждениях или неоконченной репликации ДНК. События от начала профазы и до окончания метафазы инициируются и протекают при участии белковых комплексов, состоящих из митотических циклинов и циклин-зависимых киназ (англ. M-Cdk).
Вторая контрольная точка служит разделительным барьером на границе метафазы/анафазы. На данном этапе критичным показателем является состояние веретена деления, потому что вступление в анафазу у всех эукариот блокируется в случае наличия дефектов веретена. Ключевым активатором событий анафазы является убиквитин-лигаза APCCdc20.[25]
Схема активации циклин-зависимой киназы: 1 — неактивная циклин-зависимая протеинкиназа; 2 — циклин; 3 — активный циклин-киназный комплекс Принцип положительной обратной связи в процессе активации митоза: путём фосфорилирования циклин-киназный комплекс циклин B-Cdk1 активирует собственных активаторов семейства Cdc25 и ингибируют собственных ингибиторов семейства Wee1. В качестве возможных инициаторов петли обратной связи указаны фосфатаза Cdc25B
В качестве возможных инициаторов петли обратной связи указаны фосфатаза Cdc25Bи комплекс циклин A-Cdk.[27] Активность основных регуляторов митоза на стадии профазы (на примере позвоночных). Синим цветом обозначен график активности ингибиторов семейства Wee1 (Wee1, Myt1). Зелёным цветом обозначен график активности фосфатаз семейства Cdc25 (Cdc25A, Cdc25C). График активности комплекса циклин B-Cdk1 обозначен красным цветом. Отдельно фиолетовым цветом обозначен график активности
фосфатазы Cdc25B.[28]
Основные регуляторы митоза
Циклин-киназы
Ключевыми активаторами митоза, обеспечивающими инициацию событий профазы—метафазы, являются циклин-киназные комплексы (англ. M-Cdk). Данные комплексы представляют собой гетеродимеры, состоящие из двух субъединиц: регуляторной — митотического циклина (англ. M cyclin) и каталитической — циклин-зависимой киназы (англ. Cdk — cyclin-dependent kinase).
В регуляцию митоза у всех эукариот вовлечена циклин-зависимая киназа Cdk1,[29] которая представляет собой фермент (фосфорилазу), модифицирующий белки за счёт переноса фосфатной группы от АТФ на аминокислоты серин и треонин. Концентрация Cdk1 постоянна на протяжении всего клеточного цикла,[30] поэтому активность циклин-зависимой киназы в процессе митоза зависит главным образом от её соединения с митотическим циклином. Концентрация митотических циклинов увеличивается по мере приближения к митозу и достигает максимума в метафазе. Различным таксонам свойственны различные митотические циклины. Так, у почкующихся дрожжей в регуляцию митоза вовлечены четыре циклина Clb1, 2, 3 и 4; у дрозофилы — циклины A, B, B3; у позвоночных — циклин B.[31]
Концентрация Cdk1 постоянна на протяжении всего клеточного цикла,[30] поэтому активность циклин-зависимой киназы в процессе митоза зависит главным образом от её соединения с митотическим циклином. Концентрация митотических циклинов увеличивается по мере приближения к митозу и достигает максимума в метафазе. Различным таксонам свойственны различные митотические циклины. Так, у почкующихся дрожжей в регуляцию митоза вовлечены четыре циклина Clb1, 2, 3 и 4; у дрозофилы — циклины A, B, B3; у позвоночных — циклин B.[31]
Регуляторы активности циклин-киназ
Накопление митотических циклинов начинается ещё на стадии G2. Увеличение концентрации циклинов обеспечивается за счёт транскрипции соответствующих им генов.[32] Новосинтезированные циклины сразу объединяются с неактивной киназой Cdk1. Однако образующиеся при этом циклин-киназные комплексы сохраняются в неактивном состоянии до момента активации митоза. Сдерживание ативности комплексов M-Cdk1 на протяжении фазы G2 осуществляется благодаря ингибирующему фосфорилированию молекулы Cdk1.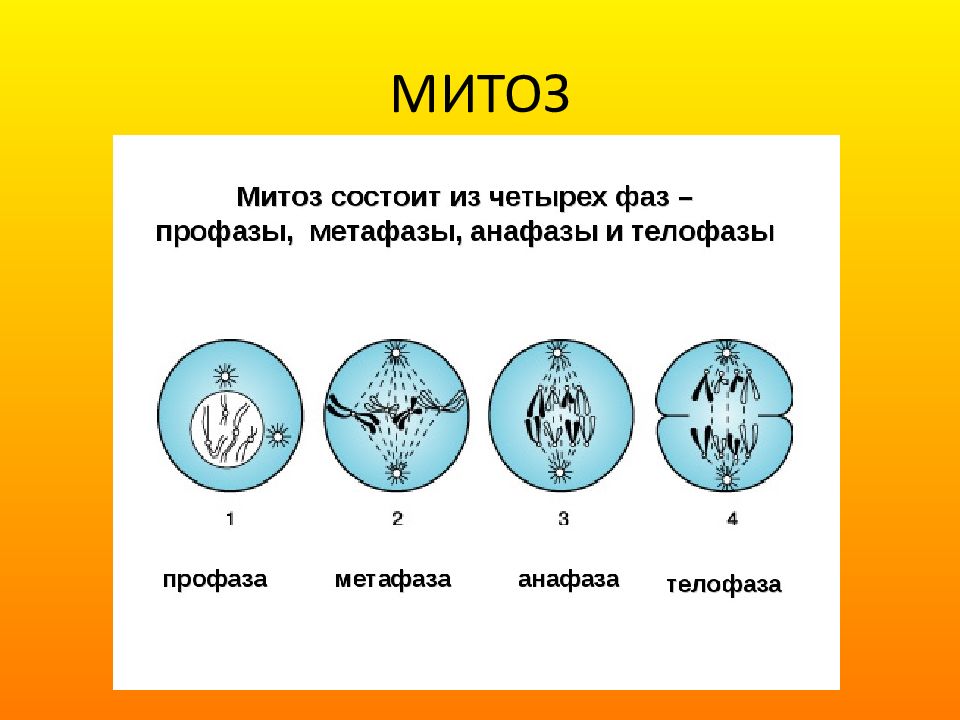 [33] За ингибирование Cdk1 ответственна группа протеинкиназ семейства Wee1.[31][32] В итоге, к началу митоза в клетке накапливается значительное количество неактивных комплексов M-Cdk1.
[33] За ингибирование Cdk1 ответственна группа протеинкиназ семейства Wee1.[31][32] В итоге, к началу митоза в клетке накапливается значительное количество неактивных комплексов M-Cdk1.
Собственно начало профазы на молекулярном уровне знаменуется резкой активацией киназных комплексов M-Cdk1. В основе скачкообразного увеличения активности M-Cdk1 лежат как минимум два взаимосвязанных события. Во-первых, к началу профазы приурочена активация фосфатаз семейства Cdc25, которые освобождают комплекс M-Cdk1 от ингибирующих фосфатных групп. Во-вторых, активированные таким образом киназы M-Cdk1 включаются в цепочку положительной обратной связи: путём фосфорилирования они активируют собственных активаторов семейства Cdc25 и ингибируют собственных ингибиторов семейства Wee1. В итоге, в начале профазы наблюдается взаимосвязанное увеличение активности фосфатаз семейства Cdc25 и циклин-киназ M-Cdk1 на фоне параллельного снижения активности ингибиторов семейства Wee1. Таким образом, в основу активации митоза заложен принцип положительной обратной связи. Но, несмотря на то, что уже известно об инициирующих механизмах митоза, остаётся до сих пор неясно, какой именно стимул изначально активирует Cdc25 или Cdk1, тем самым обеспечивая запуск цепочки положительной обратной связи.[~ 2][35][32]
Таким образом, в основу активации митоза заложен принцип положительной обратной связи. Но, несмотря на то, что уже известно об инициирующих механизмах митоза, остаётся до сих пор неясно, какой именно стимул изначально активирует Cdc25 или Cdk1, тем самым обеспечивая запуск цепочки положительной обратной связи.[~ 2][35][32]
Polo- и aurora-подобные киназы
Помимо циклин-зависимых киназ в регуляцию митотических событий вовлечены как минимум ещё два типа киназ: polo-подобные киназы и киназы семейства aurora. Polo-подобные киназы (англ. polo-like kinase, Plk) представляют собой серин-треониновые протеинкиназы, активирующиеся на начальных и инактивирующиеся на поздних стадиях митоза или в начале фазы G1. Данные киназы вовлечены в различные митотические процессы: сборку веретена деления, функционирование кинетохора, цитокинез.[36] Киназы семейства aurora также относятся к группе серин-треониновых протеинкиназ. У многоклеточных выделяются два основных представителя данного семейства: aurora A и aurora B. Киназа aurora A участвует в регуляции функционирования центросом и митотического веретена. Киназа aurora B участвует в регуляции процессов конденсации и разделения сестринских хроматид, а также обеспечивает присоединение кинетохоров к микротрубочкам веретена деления.[37]
Киназа aurora A участвует в регуляции функционирования центросом и митотического веретена. Киназа aurora B участвует в регуляции процессов конденсации и разделения сестринских хроматид, а также обеспечивает присоединение кинетохоров к микротрубочкам веретена деления.[37]
Активатор анафазы APC
Cdc20 Комплекс стимуляции анафазы[38]Комплекс, стимулирующий анафазу (англ. anaphase-promoting complex, APC), также называемый циклосома, представляет собой крупное белковое соединение, которому отводится решающая роль в активации анафазы. Функционально комплекс стимуляции анафазы представляет собой убиквитинлигазу и катализирует реакции присоединения молекул убиквитина к различным целевым белкам, которые в итоге подвергаются протеолизу.[39]
В структуре комплекса стимуляции анафазы выделяется порядка 11—13 субъединиц. Ядро комплекса составляют: субъединица куллина (Apc2) и RING-домен (Apc11), к которому присоединяется убиквитин-конъюгирующий фермент (E2). Функционирование комплекса регулируется за счёт присоединения активирующей субъединицы в нужный момент клеточного цикла.[38]
Функционирование комплекса регулируется за счёт присоединения активирующей субъединицы в нужный момент клеточного цикла.[38]
Белок Cdc20 (англ. cell division cycle protein 20 — «белок клеточного цикла 20») активирует комплекс APC при переходе делящейся клетки из метафазы в анафазу. Происходит это следующим образом. На стадии метафазы циклин-киназный комплекс M-Cdk путём фосфорилирования трансформирует ядро комплекса APC. В результате указанного конформационного изменения повышается вероятность присоединения активатора Cdc20. В итоге, активированный комплекс APCCdc20 обретает убиквитин-лигазную активность и убиквитинирует свои главные цели — секьюрин и митотические циклины.[38]
Секьюрин (одна из главных мишеней APCCdc20) представляет собой ингибирующий белок, сдерживающий в неактивном состоянии фермент сепаразу. Вследствие реакции убиквитинирования секьюрин разрушается, а высвободившаяся при этом сепараза разрушает когезин. После деградации когезина, обеспечивающего сцепление сестринских хроматид, происходит разделение и расхождение хромосом к полюсам деления клетки.[40]
После деградации когезина, обеспечивающего сцепление сестринских хроматид, происходит разделение и расхождение хромосом к полюсам деления клетки.[40]
Убиквитинирование и, как следствие, разрушение митотических циклинов (ещё одной важной мишени APCCdc20) запускает цепочку отрицательной обратной связи. Выглядит это следующим образом. Циклин-киназный комплекс M-Cdk активирует убиквитин-лигазный комплекс APCCdc20, который целенаправленно разрушает митотические циклины, что ведёт к деградации циклин-киназного комплекса M-Cdk, т. е. цепочка реакций приводит к разрушению изначального активатора этой цепочки. Но поскольку активность APCCdc20 зависит от комплекса M-Cdk, инактивация циклин-киназы M-Cdk приводит к инактивации APCCdc20. В итоге APCCdc20 деактивируется к концу митоза.[38]
Продолжительность митоза
Собственно митоз зачастую протекает сравнительно быстро. Средняя продолжительность составляет 1—2 часа,[1][4] что занимает всего около 10 % времени клеточного цикла. К примеру, у делящихся клеток меристемы корней интерфаза составляет 16—30 часов, а митоз длится всего 1—3 часа. Для эпителиальных клеток кишечника мыши интерфазный период составляет порядка 20—22 часов, а митоз продолжается в течение 1 часа.[41] В клетках животных митоз обычно протекает быстрее и длится в среднем 30—60 минут, в то время как в растительных клетках средняя продолжительность митоза составляет 2—3 часа.[5] Известны исключения с противоположными показателями. К примеру, в животных клетках продолжительность митоза может достигать 3,8 часов (эпидермис мыши). Или же встречаются растительные объекты с длительностью митоза в 5 минут (Chilomonas).[42] Наиболее интенсивно митоз протекает в эмбриональных клетках (10—40 минут в дробящихся яйцеклетках).
К примеру, у делящихся клеток меристемы корней интерфаза составляет 16—30 часов, а митоз длится всего 1—3 часа. Для эпителиальных клеток кишечника мыши интерфазный период составляет порядка 20—22 часов, а митоз продолжается в течение 1 часа.[41] В клетках животных митоз обычно протекает быстрее и длится в среднем 30—60 минут, в то время как в растительных клетках средняя продолжительность митоза составляет 2—3 часа.[5] Известны исключения с противоположными показателями. К примеру, в животных клетках продолжительность митоза может достигать 3,8 часов (эпидермис мыши). Или же встречаются растительные объекты с длительностью митоза в 5 минут (Chilomonas).[42] Наиболее интенсивно митоз протекает в эмбриональных клетках (10—40 минут в дробящихся яйцеклетках).
Длительность митоза находится в зависимости от целого ряда факторов: размеров делящейся клетки, её плоидности, числа ядер. Частота клеточных делений также зависит от степени дифференцировки клеток и специфики выполняемых функций. Так, нейроны или клетки скелетной мышцы человека не делятся совсем; клетки печени обычно делятся раз в один или два года, а некоторые эпителиальные клетки кишечника делятся чаще, чем 2 раза в сутки.[43]
Так, нейроны или клетки скелетной мышцы человека не делятся совсем; клетки печени обычно делятся раз в один или два года, а некоторые эпителиальные клетки кишечника делятся чаще, чем 2 раза в сутки.[43]
Темп клеточного деления зависит также от условий окружающей среды, в частности, от температуры. Повышение температуры окружающей среды в физиологических пределах повышает скорость митоза, что может быть объяснено обычной закономерностью кинетики химических реакций.[44]
Аппарат клеточного деления
Деление всех эукариотических клеток сопряжено с формированием специального аппарата клеточного деления. Активная роль в митотическом делении клеток зачастую отведена цитоскелетным структурам. Универсальным как для животных, так и для растительных клеток является двухполюсное митотическое веретено, состоящее из микротрубочек и связанных с ними белков.[45] Веретено деления обеспечивает строго одинаковое распределение хромосом между полюсами деления, в области которых в телофазе образуются ядра дочерних клеток.
Ещё одна не менее важная структура цитоскелета отвечает за разделение цитоплазмы (цитокинез) и, как следствие, за распределение клеточных органелл. В животных клетках за цитокинез отвечает сократимое кольцо из актиновых и миозиновых филаментов. В большинстве клеток высших растений из-за наличия жёсткой клеточной стенки цитокинез протекает с образованием клеточной пластинки в плоскости между двумя дочерними клетками. При этом область образования новой клеточной перегородки определяется заранее предпрофазным пояском из актиновых микрофиламентов, а поскольку актин участвует также в формировании клеточных септ у грибов, возможно, что он направляет цитокинез у всех эукариот.[46]
Веретено деления
Поздняя метафаза митоза в клетке лёгкого тритона (использованы иммунофлуоресцентные красители).[47] Четко просматривается веретено деления, образованное микротрубочками (зелёные), и хромосомы (синие)Формирование веретена деления начинается в профазе. В его образовании принимают участие полярные тельца (полюса) веретена и кинетохоры хромосом, и те и другие взаимодействуют с микротрубочками — биополимерами, состоящими из субъединиц тубулина. Главным центром организации микротрубочек (ЦОМТ) во многих эукариотических клетках является центросома — скопление аморфного фибриллярного материала, причём в большинстве животных клеток в состав центросом также входят пары центриолей.[48] Во время интерфазы ЦОМТ, как правило, располагающийся вблизи клеточного ядра, инициирует рост микротрубочек, расходящихся к периметру клетки и образующих цитоскелет. В S-фазе материал центросомы удваивается, а в профазе митоза начинается расхождение дочерних центросом. От них в свою очередь «отрастают» микротрубочки, которые удлиняются вплоть до соприкосновения друг с другом, после чего центросомы расходятся. Затем, в прометафазе, после разрушения ядерной мембраны, микротрубочки проникают в область клеточного ядра и взаимодействуют с хромосомами. Две дочерние центросомы теперь называют полюсами веретена.
В его образовании принимают участие полярные тельца (полюса) веретена и кинетохоры хромосом, и те и другие взаимодействуют с микротрубочками — биополимерами, состоящими из субъединиц тубулина. Главным центром организации микротрубочек (ЦОМТ) во многих эукариотических клетках является центросома — скопление аморфного фибриллярного материала, причём в большинстве животных клеток в состав центросом также входят пары центриолей.[48] Во время интерфазы ЦОМТ, как правило, располагающийся вблизи клеточного ядра, инициирует рост микротрубочек, расходящихся к периметру клетки и образующих цитоскелет. В S-фазе материал центросомы удваивается, а в профазе митоза начинается расхождение дочерних центросом. От них в свою очередь «отрастают» микротрубочки, которые удлиняются вплоть до соприкосновения друг с другом, после чего центросомы расходятся. Затем, в прометафазе, после разрушения ядерной мембраны, микротрубочки проникают в область клеточного ядра и взаимодействуют с хромосомами. Две дочерние центросомы теперь называют полюсами веретена. [49]
[49]
По морфологии различают два типа митотического веретена: астральный (или конвергентный) и анастральный (дивергентный).[~ 3][51]
Астральный тип митотической фигуры, характерный для животных клеток, отличают благодаря небольшим зонам, на полюсах веретена, в которых сходятся (конвергируют) микротрубочки. Зачастую центросомы, располагающиеся в области полюсов астрального веретена, содержат центриоли. От полюсов деления также расходятся во всех направлениях радиальные микротрубочки, не входящие в состав веретена, а образующие звездчатые зоны — цитастеры.
Анастральный тип митотической фигуры отличается широкими полярными областями веретена, так называемыми полярными шапочками, в их состав не входят центриоли. Микротрубочки при этом расходятся широким фронтом (дивергируют) от всей зоны полярных шапочек. Этот тип митотической фигуры также отличает отсутствие цитастеров. Анастральный тип митотического веретена наиболее характерен для делящихся клеток высших растений, хотя иногда наблюдается и в некоторых клетках животных.
Микротрубочки
Микротрубочки — динамичные структуры, принимающие активное участие в построении веретена деления во время митоза. Химически они представляют собой биополимеры, состоящие из субъединиц белка тубулина. Количество микротрубочек в клетках различных организмов может значительно отличаться. В метафазе веретено деления в клетках высших животных и растений может содержать до нескольких тысяч микротрубочек, тогда как у некоторых грибов их всего около 40.[49]
Митотические микротрубочки веретена деления «динамически нестабильны». Их «положительные» или «плюс-концы», расходящиеся во всех направлениях от центросом резко переходят от равномерного роста к стремительному укорочению, при котором часто деполимеризуется вся микротрубочка. Согласно этим данным образование митотического веретена объясняется селективной (выборочной) стабилизацией микротрубочек взаимодействующих в экваториальной области клетки с кинетохорами хромосом и с микротрубочками, идущими от противоположного полюса деления.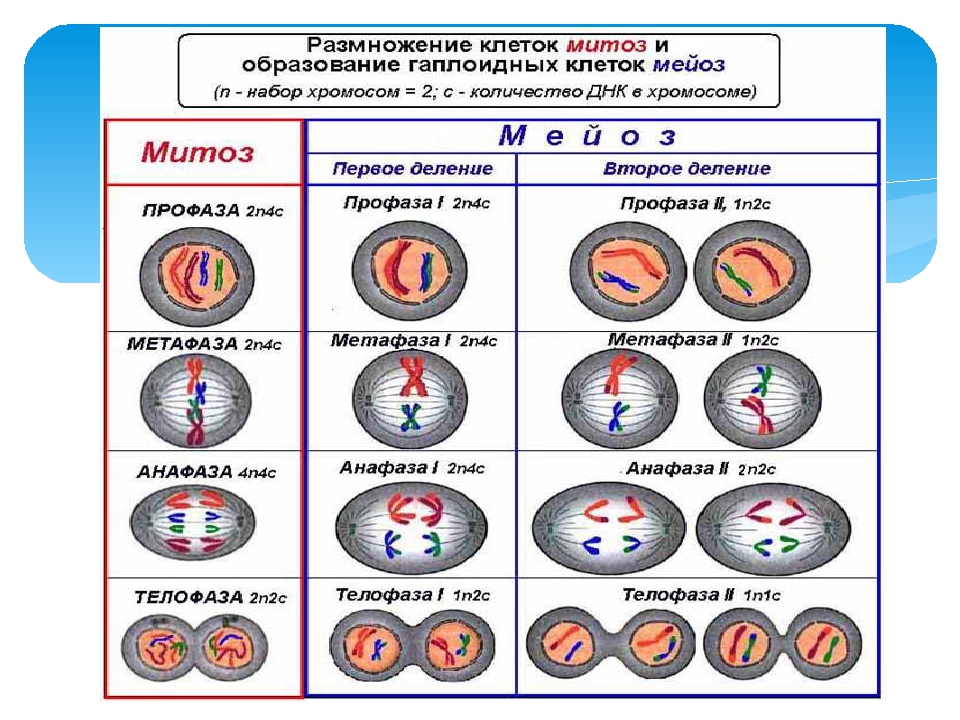 Данная модель объясняет характерную двухполюсную фигуру митотического веретена.[49]
Данная модель объясняет характерную двухполюсную фигуру митотического веретена.[49]
Центромеры и кинетохоры
Центромеры — специализированные последовательности ДНК, необходимые для связывания с микротрубочками веретена деления и для последующего расхождения хромосом. В зависимости от локализации различают несколько типов центромер. Для голоцентрических центромер характерно образование связей с микротрубочками веретена по всей длине хромосомы (некоторые насекомые, нематоды, некоторые растения). В противоположность голоцентрическим моноцентрические центромеры служат для связи с микротрубочками в единственной области хромосомы.[51]
В центромерной области обычно располагаются кинетохоры хромосом — сложные белковые комплексы, морфологически очень сходные по своей структуре для различных групп эукариот, как, например, для диатомовых водорослей, так и для человека.[52] Обычно на каждую хроматиду (хромосому) приходится по одному кинетохору. На электронных микрофотографиях кинетохор обычно выглядит как пластинчатая трехслойная структура.[53] Порядок слоев следующий: внутренний плотный слой, примыкающий к телу хромосомы; средний рыхлый слой; внешний плотный слой, от которого отходит множество фибрилл, образуя т. н. фиброзную корону кинетохора.
На электронных микрофотографиях кинетохор обычно выглядит как пластинчатая трехслойная структура.[53] Порядок слоев следующий: внутренний плотный слой, примыкающий к телу хромосомы; средний рыхлый слой; внешний плотный слой, от которого отходит множество фибрилл, образуя т. н. фиброзную корону кинетохора.
К основным функциям кинетохора относят: закрепление микротрубочек веретена деления, обеспечение движения хромосом во время митоза при участии микротрубочек, связывание между собой сестринских хроматид и регуляцию их последующего разделения в анафазе митоза.[54] Минимально достаточно одной микротрубочки (например, для дрожжей) ассоциированной с кинетохором, чтобы обеспечить движение хромосомы. Однако с одним кинетохором могут быть связаны целые пучки, состоящие из 20—40 микротрубочек (например, у высших растений или человека), чтобы обеспечить расхождение хромосом к полюсам клетки.[54][53]
Фазы митоза
Временной ход митоза и цитокинеза, типичный для клетки млекопитающего. Точные цифры для разных клеток различны. Цитокинез берёт своё начало в анафазе и завершается, как правило,
Точные цифры для разных клеток различны. Цитокинез берёт своё начало в анафазе и завершается, как правило,к окончанию телофазы
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой (от слова «митоз»). М-фазу условно подразделяют на шесть стадий, постепенно и непрерывно переходящих одна в другую.[48][41] Первые пять — профаза, прометафаза (метакинез), метафаза, анафаза и телофаза (или цитотомия) — составляют митоз,[~ 4] а берущий своё начало в анафазе процесс разделения цитоплазмы клетки, или цитокинез, протекает вплоть до завершения митотического цикла и, как правило, рассматривается в составе телофазы.
Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны стадии сопряженные с процессами внутриклеточного синтеза: профаза (2—270 минут) и телофаза (1,5—140 минут). Наиболее быстротечны фазы митоза, в ходе которых происходит движение хромосом: метафаза (0,3—175 минут) и анафаза (0,3—122 минуты).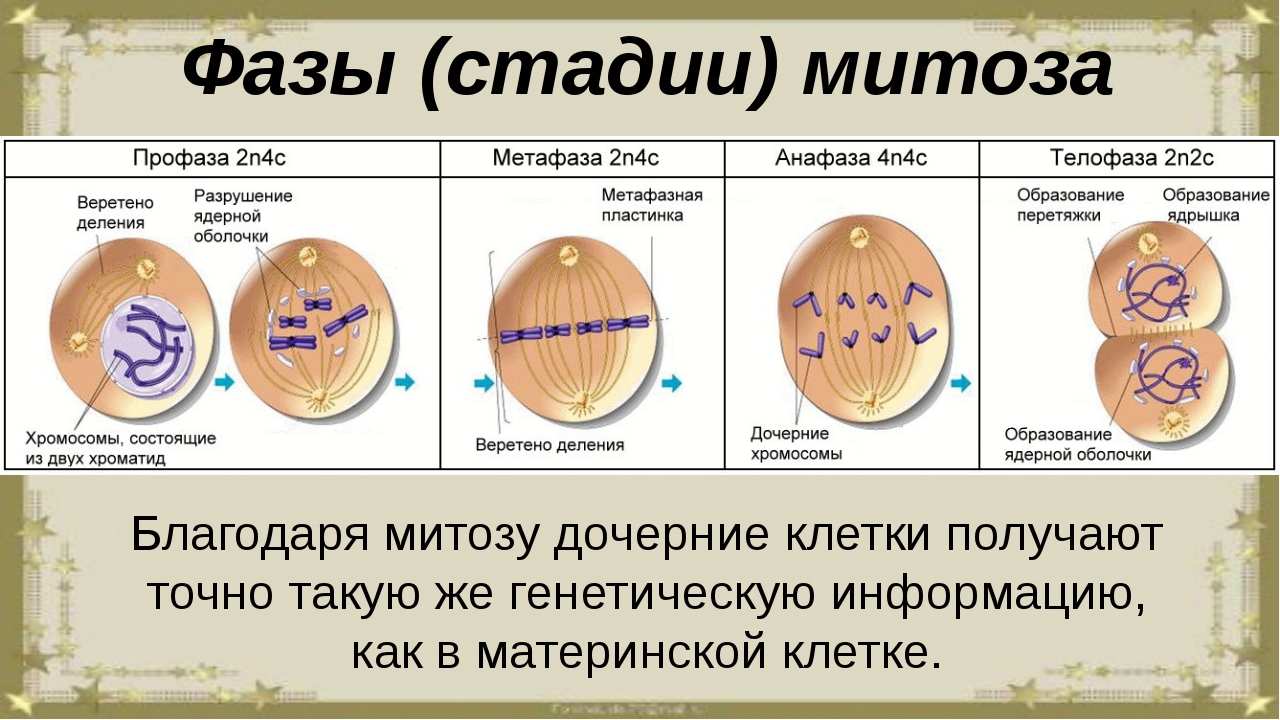 Непосредственно процесс расхождения хромосом к полюсам обычно не превышает 10 минут.[56]
Непосредственно процесс расхождения хромосом к полюсам обычно не превышает 10 минут.[56]
Профаза
ПрофазаК основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки.[57] Распад ядрышка в профазе является характерной, но не обязательной для всех клеток особенностью.[58]
Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.[59]
К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.[59]
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено — одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца (центросомы), микротрубочки и кинетохоры хромосом.[51]
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут (в интерфазе) до 15 секунд.[49][59] Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками.[59] Полимеризующиеся плюс-концы («+»-концы) являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. [49] Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.[~ 5]
[49] Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.[~ 5]
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами («-»-концами) микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления. [61]
[61]
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.[62]
Прометафаза
ПрометафазаОкончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны.[57] Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают.[63] После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин.[63] Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом.[64][63] В результате такого взаимодействия кинетохорные микротрубочки (связанные с кинетохором) стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена.[60] Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.
Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин.[63] Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом.[64][63] В результате такого взаимодействия кинетохорные микротрубочки (связанные с кинетохором) стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена.[60] Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки. [65]
[65]
В клетках млекопитающих прометафаза протекает, как правило, в течение 10—20 минут.[64] В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона — около 30 минут.[66] В дрожжевых клетках невозможно чётко разграничить стадии профазы и прометафазы по причине сохранения ядерной оболочки в процессе деления. Аналогичным образом, частичное или более позднее разрушение ядерной мембраны затрудняет разграничение стадий профазы и прометафазы в клетках Drosophila и C. elegans. В подобных случаях для описания всех ранних событий митотического деления используется обобщающий термин «профаза».[57]
Метафаза
МетафазаВ завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена (а не всей клетки[67]) примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи — к периферии клетки (фигура «материнской звезды»).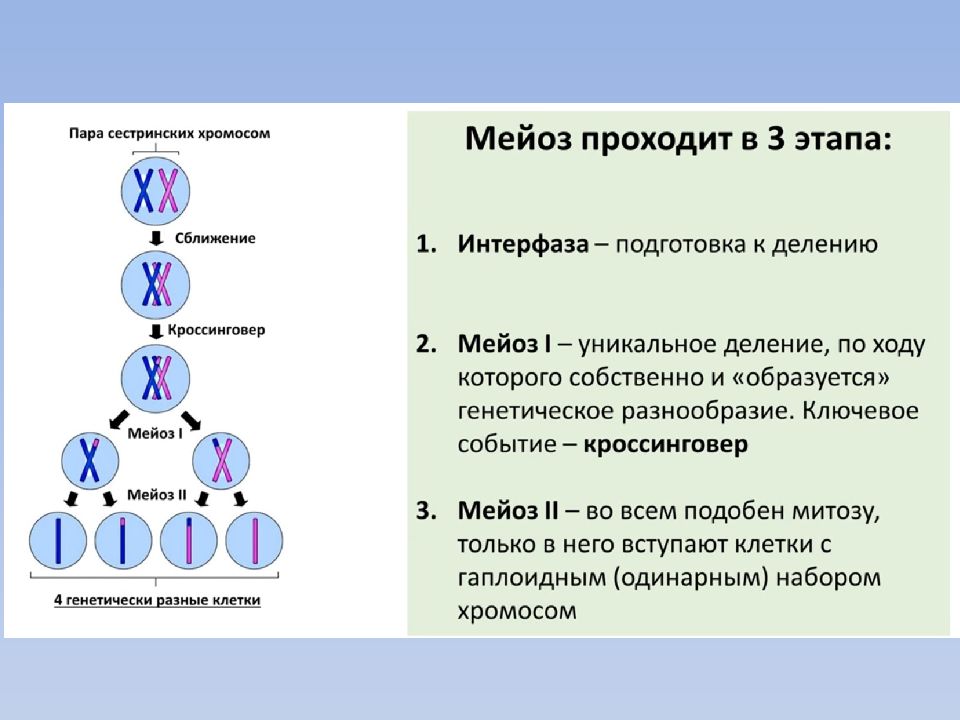 В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.[68][69] В дрожжевых клетках хромосомы тоже не выстраиваются в экваториальной плоскости, а располагаются произвольно вдоль волокон веретена деления.[57]
В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.[68][69] В дрожжевых клетках хромосомы тоже не выстраиваются в экваториальной плоскости, а располагаются произвольно вдоль волокон веретена деления.[57]
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.[70]
В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума. [68]
[68]
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.[68]
Анафаза
АнафазаАнафаза — самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид в направлении противоположных полюсов клетки.[71] Хроматиды расходятся с равномерной скоростью достигающей 0,5—2 мкм/мин.[1][72] (0,2—5 мкм/мин.[73]), при этом они часто принимают V-образную форму. Их движение обусловлено воздействием значительных сил, оценочно 10−5дин на хромосому, что в 10 000 раз превышает усилие, необходимое для простого продвижения хромосомы через цитоплазму с наблюдаемой скоростью.[74]
Как правило, расхождение хромосом в анафазе состоит из двух относительно независимых процессов называемых анафазой А и анафазой В.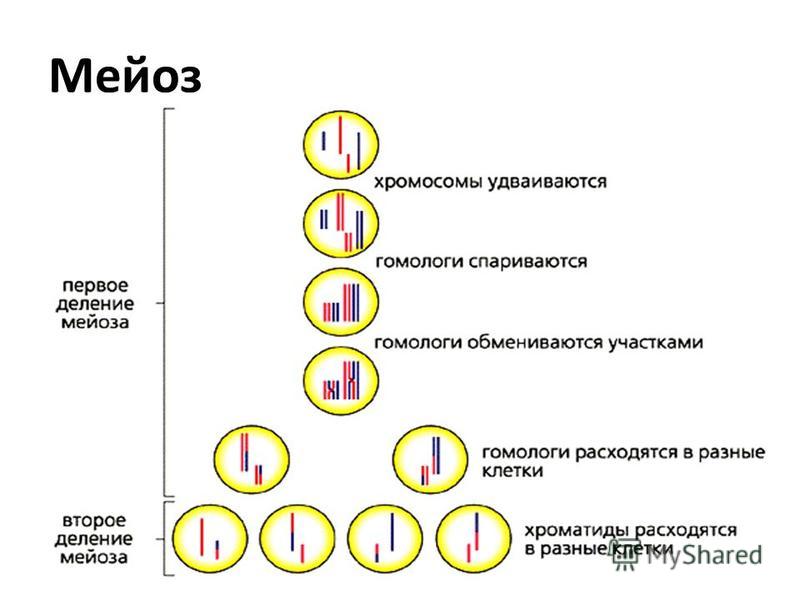
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки.[57] За их движение при этом отвечают те же силы, что ранее удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения хроматид сопровождается сокращением длины деполимеризующихся кинетохорных микротрубочек. Причем их распад наблюдается преимущественно (на 80 %[75]) в области кинетохоров, со стороны плюс-концов (ранее, с начала профазы и вплоть до начала анафазы, на плюс-концах преобладали процессы сборки субъединиц тубулина).[74] Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения сестринских хроматид, так как их движение прекращается при добавлении таксола или тяжёлой воды (D2O), оказывающих стабилизирующее воздействие на микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока остается неизвестным.[~ 6][74]
Во время анафазы В расходятся сами полюса деления клетки,[57] и, в отличие от анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей разных видов.[76] Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки.[77]
Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей разных видов.[76] Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки.[77]
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны. Так в клетках млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к противоположным полюсам и продолжается вплоть до удлинения митотического веретена в 1,5—2 раза по сравнению с метафазным. В некоторых других клетках (например, дрожжевых) анафаза В начинается только после того как хроматиды достигают полюсов деления. У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным.[71] В растительных клетках анафаза В отсутствует.[77]
У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным.[71] В растительных клетках анафаза В отсутствует.[77]
Телофаза
ТелофазаТелофаза (от греч. telos — конец) рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки.[77] В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума.[78] При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры.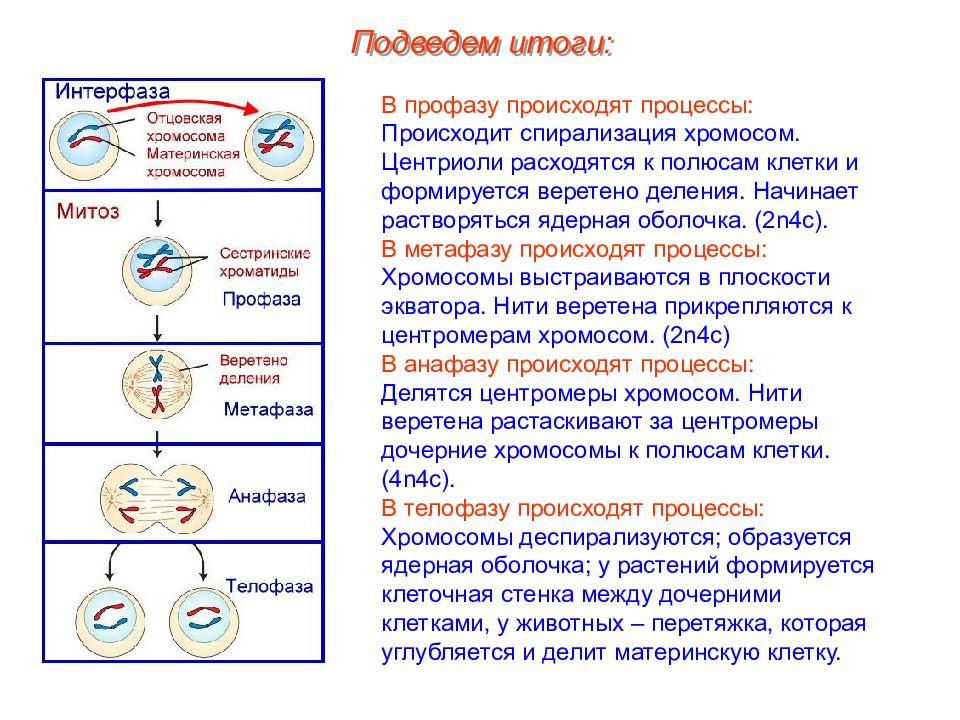 В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса.[79] Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса.[79] Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга.[80]
Окончание телофазы преимущественно совпадает с разделением тела материнской клетки — цитокинезом (цитотомией). [81][82] При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут свое начало ещё в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы.[~ 7]
[81][82] При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут свое начало ещё в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы.[~ 7]
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки (наиболее характерно для клеток животных) и деление путём образования клеточной пластинки (свойственно растениям в связи с наличием жёсткой клеточной стенки). Плоскость деления клетки детерминируется положением митотического веретена и проходит под прямым углом к длинной оси веретена.[83]
При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно ещё в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократимое кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса.[84]
В дальнейшем, вследствие активности сократимого кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса.[84]
Деление путём образования клеточной пластинки начинается с перемещения мелких ограниченных мембраной пузырьков по направлению к экваториальной плоскости клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру — раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных микротрубочек веретена деления, образующих цилиндрическую структуру, называемую фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых мембранных пузырьков продолжается рост клеточной пластинки вплоть до её окончательного слияния с мембраной материнской клетки.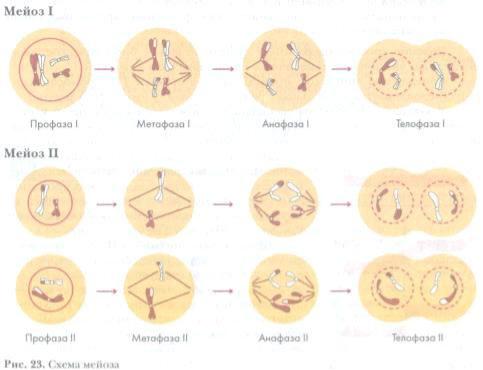 После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки.[85]
После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки.[85]
Патология митоза
Патология митоза развивается при нарушении нормального течения митотического деления и зачастую приводит к возникновению клеток с несбалансированными кариотипами, следовательно, ведёт к развитию мутаций и анеуплоидии. Также в результате развития отдельных форм патологии наблюдаются хромосомные аберрации. Незавершённые митозы, прекращающиеся по причине дезорганизации или разрушения митотического аппарата приводят к образованию полиплоидных клеток. Полиплоидия и формирование дву- и многоядерных клеток возникают в случае нарушений механизмов цитокинеза. При значительных последствиях патологии митоза возможна гибель клетки.
В нормальных тканях патология встречается в незначительных количествах. Например, в эпидермисе мышей встречается около 0,3 % патологических митозов; в эпителии гортани и матки человека — около 2 %. Патологические митозы часто наблюдаются при канцерогенезе, при различных экстремальных воздействиях, при лучевой болезни или вирусной инфекции,[~ 8] при раке и предраковых гиперплазиях.[~ 9] Частота патологических митозов также увеличивается с возрастом.[88]
Патологические митозы часто наблюдаются при канцерогенезе, при различных экстремальных воздействиях, при лучевой болезни или вирусной инфекции,[~ 8] при раке и предраковых гиперплазиях.[~ 9] Частота патологических митозов также увеличивается с возрастом.[88]
Условно различают патологию митоза функционального и органического типа. К функциональным нарушениям относят, например, гипореактивность вступающих в митоз клеток — снижение реакции на физиологические регуляторы, определяющие интенсивность пролиферации нормальных клеток. Органические нарушения возникают при повреждении структур, участвующих в митотическом делении (хромосомы, митотический аппарат, клеточная поверхность), а также при нарушении процессов, связанных с данными структурами (репликация ДНК, образование веретена деления, движение хромосом, цитокинез).[88]
Классификация и общая характеристика различных форм патологии митоза
На основании морфологических признаков и цитохимических нарушений митотического процесса выделяют три основных группы патологии митоза: патология, связанная с повреждением хромосом; патология, связанная с повреждением митотического аппарата; нарушение цитокинеза. [89]
[89]
I. Патология митоза, связанная с повреждением хромосом
1) Задержка митоза в профазе наблюдается при нарушениях репликации ДНК.
2) Нарушение спирализации и деспирализации хромосом прослеживается в результате действия на делящуюся клетку различными митотическими ядами. Например, воздействие колхицина приводит к гиперспирализации хромосом, которые приобретают укороченную и утолщенную форму.[89]
3) Раннее (преждевременное) разделение хроматид в профазе (в норме разделение хроматид происходит на рубеже перехода метафазы в анафазу). Обозначенная патология наблюдается, к примеру, при изменении осмотического давления в фибробластах кролика в культуре ткани или же при воздействии канцерогенов (бензопирена, метилхолантрена) на мышиные фибробласты.[89]
В центральной делящейся клетке, находящейся в стадии телофазы, наблюдается парный фрагмент. Микрофотография сделана во время проведения эксперимента Allium test В центре клеточного поля видна делящаяся клетка в стадии анафазы. Отчётливо заметен хроматидный мост и одиночный фрагмент хромосомы. Микрофотография сделана во время проведения эксперимента Allium test по изучению влияния активного излучения сотового телефона на клетки in vivo
Отчётливо заметен хроматидный мост и одиночный фрагмент хромосомы. Микрофотография сделана во время проведения эксперимента Allium test по изучению влияния активного излучения сотового телефона на клетки in vivo
4) Фрагментация и пульверизация хромосом возникает в опухолевых клетках, при вирусной инфекции, в результате воздействия на нормальные клетки ионизирующего излучения или мутагенов. Фрагменты могут быть одиночными, парными и множественными. Те из них, которые лишены центромерного участка, не участвуют в метакинезе, и, соответственно, не расходятся к полюсам деления в анафазе. При массовой фрагментации хромосом (пульверизация) большинство фрагментов также беспорядочно рассеиваются в цитоплазме и не участвуют в метакинезе.[90]
В итоге часть фрагментов хромосом может попасть в одно из дочерних ядер, либо резорбироватья, либо образовать обособленное микроядро. Также отдельные фрагменты обладают способностью воссоединяться своими концами, причём подобные воссоединения носят случайный характер и приводят к хромосомным аберрациям. [91]
[91]
5) Хромосомные и хроматидные мосты являются следствием фрагментации хромосом. При воссоединении фрагментов содержащих центромер образуется дицентрическая хромосома, которая в ходе анафазы растягивается между противоположными полюсами деления, образуя мост. Хромосомный (обычно двойной) мост возникает в результате воссоединения фрагментов хромосом, каждый из которых образован двумя хроматидами с центромерой. Хроматидный (обычно одиночный) мост возникает в результате воссоединения двух фрагментов отдельных хроматид с центромерой.[92]
К концу анафазы — в начале телофазы мосты обычно быстро рвутся в результате чрезмерного растягивания дицентрических фрагментов хромосом. Образование мостов приводит к генотипической разнородности дочерних клеток, а также нарушает течение завершающих стадий деления и задерживает цитокинез.[92]
6) Отставание хромосом в метакинезе и при расхождении к полюсам возникает при повреждении хромосом в области кинетохора. Поврежденные хромосомы пассивно «дрейфуют» в цитоплазме и в итоге либо разрушаются и элиминируются из клетки, либо случайным образом попадают в одно из дочерних ядер, либо образуют отдельное микроядро. Отставание хромосом наблюдалось в культурах ткани опухолевых клеток, а также в экспериментах, в ходе которых кинетохоры хромосом облучались микропучком ультрафиолетовых лучей.[93]
Поврежденные хромосомы пассивно «дрейфуют» в цитоплазме и в итоге либо разрушаются и элиминируются из клетки, либо случайным образом попадают в одно из дочерних ядер, либо образуют отдельное микроядро. Отставание хромосом наблюдалось в культурах ткани опухолевых клеток, а также в экспериментах, в ходе которых кинетохоры хромосом облучались микропучком ультрафиолетовых лучей.[93]
7) Образование микроядер происходит вследствие фрагментации или отставания отдельных хромосом, вокруг которых в телофазе формируется ядерная оболочка, параллельно образованию оболочки вокруг основных дочерних ядер. Новообразованные микроядра либо сохраняются в клетке в течение всего дальнейшего клеточного цикла вплоть до очередного деления, либо подвергаются пикнозу, разрушаются и выводятся из клетки.[93]
8) При нерасхождении хромосом сестринские хроматиды не разъединяются с началом анафазы и вместе отходят к одному из полюсов, что приводит к анеуплоидии.[94]
9) Набухание и слипание хромосом наблюдается в опухолевых клетках и при воздействии токсических доз различных митотических ядов. Вследствие набухания хромосомы теряют свои нормальные очертания и слипаются, превращаясь в комковатые массы. Расхождения хромосом не происходит и клетки в таком состоянии зачастую погибают.[94]
Вследствие набухания хромосомы теряют свои нормальные очертания и слипаются, превращаясь в комковатые массы. Расхождения хромосом не происходит и клетки в таком состоянии зачастую погибают.[94]
II. Патология митоза, связанная с повреждением митотического аппарата
1) Задержка митоза в метафазе характерна для всей группы патологий митоза, связанных с повреждением митотического аппарата.
2) Колхициновый митоз или к-митоз — одна из форм патологии митоза, связанная с повреждением митотического аппарата вследствие воздействия статмокинетических ядов (колхицина, колцемида, винбластина, винкристина, аценафтена, нокодазол, метанола и др.).[95] В результате воздействия статмокинетических ядов митоз задерживается на стадии метафазы в связи с дезорганизацией различных компонентов митотического веретена деления — центриолей, микротрубочек, кинетохоров. Повреждения также затрагивают клеточное ядро, плазмалемму, различные внутриклеточные органоиды (митохондрии, хлоропласты, аппарат Гольджи). Действие статмокинетических ядов усиливает спирализацию хромосом, что приводит к их укорочению и утолщению, а порой доводит до набухания и слипания хромосом. Как следствие, происходят хромосомные аберрации, образуются микроядра в результате фрагментации или отставания хромосом, развивается анеуплоидия.[96]
Действие статмокинетических ядов усиливает спирализацию хромосом, что приводит к их укорочению и утолщению, а порой доводит до набухания и слипания хромосом. Как следствие, происходят хромосомные аберрации, образуются микроядра в результате фрагментации или отставания хромосом, развивается анеуплоидия.[96]
Исход к-митоза зависит от дозы и времени воздействия статмокинетического яда на делящуюся клетку. При токсических дозах наблюдается пикноз ядра и гибель клетки. Значительные отравления приводят к полиплоидизации. Воздействие небольших доз обратимо. В течение нескольких часов может восстановиться митотический аппарат и продолжиться митотическое деление.[96]
3) Рассеивание хромосом в метафазе происходит в результате повреждения или полной дезорганизации митотического аппарата.
4) Многополюсный митоз связан с аномалией репродукции центриолей, что ведет к формированию дополнительных полюсов и веретен деления. В итоге хромосомы распределяются неравномерно между дочерними ядрами, что в свою очередь ведет к образованию анеуплоидных клеток с несбалансированным набором хромосом. [97]
[97]
5) Моноцентрический center>Анафазамитоз связан с нарушением разделения центриолей. При этом формируется лишь один полюс, от которого расходятся нити единственного полуверетена. В итоге моноцентрический митоз приводит к полиплоидизации.[98]
6) Асимметричный митоз характеризуется непропорциональным развитием противоположных полюсов деления, что приводит к неравномерному распределению хромосом между дочерними ядрами, то есть к анеуплоидии.[98] В результате асимметричный митоз приводит к образованию микроклеток и гигантских клеток с гипо- и гиперплоидными ядрами.
7) Трехгрупповая метафаза и метафаза с полярными хромосомами характеризуется наличием в метафазе помимо основной экваториальной пластинки ещё двух групп или отдельных («полярных») хромосом в области полюсов деления клетки.[98] Хромосомы сохраняются вблизи полюсов веретена из-за отставания в процессе метакинеза, а не из-за преждевременного расхождения. Причинами отставания могут служить повреждения кинетохора или дезорганизация отдельных хромосомальных нитей, участвующих в движении отстающих хромосом.[99]
Причинами отставания могут служить повреждения кинетохора или дезорганизация отдельных хромосомальных нитей, участвующих в движении отстающих хромосом.[99]
8) Полая метафаза представляет собой кольцевое скопление хромосом в экваториальной пластинке вдоль периферии клетки.[100]
III. Патология митоза, связанная с нарушением цитотомии
Различают две группы патологии митоза, связанные с нарушением цитотомии: раннюю цитотомию, берущую начало ещё в анафазе; либо наоборот, запаздывание или полное отсутствие цитотомии, в результате чего формируются двуядерные клетки, либо образуется одно полиплоидное ядро.[100]
См. также
- Амитоз (от греч. а — отрицательная частица и митоз) — прямое деление интерфазного ядра путем перетяжки. Все хромосомы во время такого деления распределяются между двумя дочерними ядрами случайным образом без предварительной конденсации. Амитоз не обязательно сопровождается делением клетки, что ведет к образованию дву- и многоядерных клеток.
 Клетки, претерпевшие амитоз, теряют возможность вступить в нормальный митотический цикл.
Клетки, претерпевшие амитоз, теряют возможность вступить в нормальный митотический цикл.
- Анеуплоидия (от греч. аn — отрицательная частица; eu — хорошо, вполне; ploos — кратный; eidos — вид) — некратное изменение числа хромосом вследствие их неравномерного распределения между дочерними клетками.
- Мейоз (от греч. meiosis — уменьшение) — деление созревания, характеризующееся редукцией (уменьшением) числа хромосомных наборов клетки; основное звено гаметогенеза.
- Полиплоидия (от греч. polyploos — многократный и eidos — вид) — кратное наследственное изменение числа наборов хромосом в клетках организма.
- Политения (от греч. poly — много и лат. taenia — повязка, лента) — образование в ядре соматических клеток многонитчатых (политенных) хромосом, превышающих по размерам обычные хромосомы в сотни раз. Размер политенных хромосом обусловлен многократной репликацией без последующего расхождения сестринских хроматид, число которых может превышать 1000.

- Эндомитоз (от греч. endon — внутри и митоз) — удвоение числа хромосом внутри ядра, протекающее без разрушения ядрышка и без образования веретена деления.
- Эуплоидия — нормальное для данного вида количество хромосом.
Примечания
- Комментарии
- ↑ К таким признакам, например, относятся: поведение ядерной оболочки со всеми переходами от интактной, в разной степени фрагментированной до полностью распадающейся; неоднозначное поведение ядрышка от сохраняющегося до частично или полностью исчезающего; разная степень спирализации (или полное отсутствие таковой у динофлагеллат) и морфологической дифференциации хромосом; особенности расположения хромосом в метафазной пластинке; наличие кинетохоров и различия в их организации; различия в морфологии, характере заложения и организации веретена, продолжительность сохранения его межзональной зоны; появление наряду с центриолями особых полярных образований различной организации и места своей локализации; разная степень развития перинуклеарной оболочки и т.
 п.[17]
п.[17] - ↑ В качестве возможных рассматриваются несколько инициирующих моделей. Например, предполагается, что комплексы M-Cdk1 неполностью блокируются семейством ингибиторов Wee1. В результате, пропорционально увеличению концентрации митотических циклинов, к началу профазы может накопиться критическая масса активных киназ M-Cdk1. Частичная активация циклин-киназ у позвоночных, возможно, обеспечивается фосфатазой Cdc25B, уровень активности которой возрастает с поздней S-фазы и достигает максимума в профазе митоза. Однако было продемонстрировано, что мышиные клетки способны делиться в отсутствии данного стимула. Ещё одним возможным активатором может быть комплекс циклин A-Cdk, сохраняющий свою активность с начала S-фазы вплоть до конца прометафазы митоза.[34]
- ↑ Сам факт разделения морфологии митотического веретена на два типа не отменяет возможности сочетания обоих в пределах одного организма. Например, в раннем эмбриогенезе млекопитающих при делении созревания ооцита и при I и II делении зиготы наблюдаются бесцентриолярные анастральные митозы.
 Но уже начиная с третьего клеточного деления и во всех последующих, клетки делятся при участии астральных веретен, в полюсах которых всегда обнаруживаются центриоли.[50]
Но уже начиная с третьего клеточного деления и во всех последующих, клетки делятся при участии астральных веретен, в полюсах которых всегда обнаруживаются центриоли.[50] - ↑ Первоначально, исходя из морфологических особенностей митоза, этот процесс был разделен лишь на четыре основные стадии: профазу, метафазу, анафазу и телофазу.[55]
- ↑ Если митотические клетки поместить в тяжелую воду (D2O) или обработать таксолом (эти воздействия подавляют разборку микротрубочек), то нити веретена будут удлиняться. Такое стабилизированное веретено не может тянуть хромосомы, и митоз останавливается. Но митоз блокируется и при прямо противоположном воздействии, если нити веретена обратимо разрушить с помощью одного из трех агентов, подавляющих сборку тубулина в микротрубочки, – колхицина, низкой температуры или высокого гидростатического давления.[60]
- ↑ Существует, как минимум, три гипотетические модели, объясняющие вероятный механизм расхождения хромосом в анафазе А.
 По одной из них перемещение хроматид объясняется наличием в кинетохоре «шагающих» белков, сходных по своей природе с динеином или кинезином; они продвигаются по микротрубочке, используя для этого энергию гидролиза АТФ. По другой гипотезе движение хромосом обусловлено распадом микротрубочек: по мере того как субъединицы тубулина диссоциируют, кинетохор, чтобы сохранить связь с микротрубочкой, должен скользить в направлении полюса. Третья возможность состоит в том, что микротрубочки не ответственны прямо за возникновение силы, движущей кинетохор к полюсам, а просто регулируют движение, вызываемое какой-то другой структурой.[74]
По одной из них перемещение хроматид объясняется наличием в кинетохоре «шагающих» белков, сходных по своей природе с динеином или кинезином; они продвигаются по микротрубочке, используя для этого энергию гидролиза АТФ. По другой гипотезе движение хромосом обусловлено распадом микротрубочек: по мере того как субъединицы тубулина диссоциируют, кинетохор, чтобы сохранить связь с микротрубочкой, должен скользить в направлении полюса. Третья возможность состоит в том, что микротрубочки не ответственны прямо за возникновение силы, движущей кинетохор к полюсам, а просто регулируют движение, вызываемое какой-то другой структурой.[74] - ↑ Существует целый ряд примеров описывающих многократные митотические деления ядер без сопутствующего разделения клеточного тела. Так, в эндосперме многих растений протекают множественные митозы без деления цитоплазмы, что приводит к образованию многоядерного симпласта. Аналогичная ситуация наблюдается при синхронных делениях многочисленных ядер миксомицетов, или на ранних этапах развития зародышей некоторых насекомых.
 [81]
[81] - ↑ После инфицирования культур диплоидных клеток легкого человека вирусом Herpes simplex число патологических митозов (к-митозы, хромосомные аберрации) увеличивалось с 3 % в контроле до 40—60 % в инфицированной культуре.[86]
- ↑ На примере эпителия гортани человека были получены данные об увеличении числа патологических митозов при раке. Если при хроническом воспалении и в папилломах «юношеского» типа количество патологических митозов всего в 2—2,5 раза превышало их число в нормальном эпителии, то при предраке количество патологических митозов составляло около 25 %, а при раке и атипическом папилломатозе с переходом в рак оно достигало 36—45 %.[87]
- Источники
- ↑ 1 2 3 4 Биологический энциклопедический словарь / Гл. редактор Гиляров М. С.. — М.: Сов. энциклопедия, 1986. — 831 с. — 100 000 экз.

- ↑ Гилберт, 1995, с. 202
- ↑ 1 2 История биологии до начала XX века, 1972, с. 489
- ↑ 1 2 Албертс и др., 1993, с. 396
- ↑ 1 2 Алов И. А. Митоз — статья из Большой советской энциклопедии
- ↑ Булдаков, Калистратова, 2003, с. 39
- ↑ История биологии до начала XX века, 1972, с. 485
- ↑ История биологии до начала XX века, 1972, с. 486
- ↑ История биологии до начала XX века, 1972, с. 487
- ↑ История биологии до начала XX века, 1972, с. 488
- ↑ 1 2 История биологии до начала XX века, 1972, с. 491
- ↑ Ченцов, 2004, с. 470
- ↑ Албертс и др., 1993, с. 400
- ↑ Ченцов, 2004, с. 471
- ↑ Албертс и др., 1993, с. 403
- ↑ Албертс и др.
 , 1993, с. 404—405
, 1993, с. 404—405 - ↑ 1 2 3 4 Седова, 1996, с. 103
- ↑ 1 2 3 Райков, 1978, с. 57
- ↑ Ченцов, 2004, с. 428
- ↑ Албертс и др., 1993, с. 465
- ↑ Райков, 1978, с. 93
- ↑ 1 2 Райков, 1978, с. 94
- ↑ Райков, 1978, с. 95
- ↑ Morgan D. O., 2007, p. 90
- ↑ 1 2 Morgan D. O., 2007, p. 91
- ↑ Alberts B. at al., 2008, p. 1071
- ↑ Morgan D. O., 2007, p. 99
- ↑ Morgan D. O., 2007, p. 97
- ↑ Morgan D. O., 2007, p. 30
- ↑ Morgan D. O., 2007, p. 28
- ↑ 1 2 Morgan D. O., 2007, p. 32
- ↑ 1 2 3 Alberts B.
 at al., 2008, p. 1074
at al., 2008, p. 1074 - ↑ Morgan D. O., 2007, p. 96
- ↑ Morgan D. O., 2007, p. 98—99
- ↑ Morgan D. O., 2007, p. 98
- ↑ Morgan D. O., 2007, p. 102
- ↑ Morgan D. O., 2007, p. 103
- ↑ 1 2 3 4 Morgan D. O., 2007, p. 48
- ↑ Morgan D. O., 2007, p. 46
- ↑ Alberts B. at al., 2008, p. 1087
- ↑ 1 2 Ченцов, 2004, с. 434
- ↑ Алов, 1972, с. 18
- ↑ Албертс и др., 1993, с. 415
- ↑ Алов, 1972, с. 21
- ↑ Албертс и др., 1993, с. 438
- ↑ Албертс и др., 1993, с. 463
- ↑ NIGMS — From Molecules to Medicines: Cell Biology and Biophysics (англ.). Архивировано из первоисточника 11 февраля 2012.
- ↑ 1 2 Албертс и др.
 , 1993, с. 439
, 1993, с. 439 - ↑ 1 2 3 4 5 Албертс и др., 1993, с. 444
- ↑ Албертс и др., 1993, с. 429
- ↑ 1 2 3 Ченцов, 2004, с. 429
- ↑ Ченцов, 2004, с. 430
- ↑ 1 2 Албертс и др., 1993, с. 446
- ↑ 1 2 Ченцов, 2004, с. 433
- ↑ Алов, 1972, с. 12
- ↑ Алов, 1972, с. 19
- ↑ 1 2 3 4 5 6 Morgan D. O., 2007, p. 89
- ↑ Алов, 1972, с. 83
- ↑ 1 2 3 Ченцов, 2004, с. 434
- ↑ 1 2 Албертс и др.
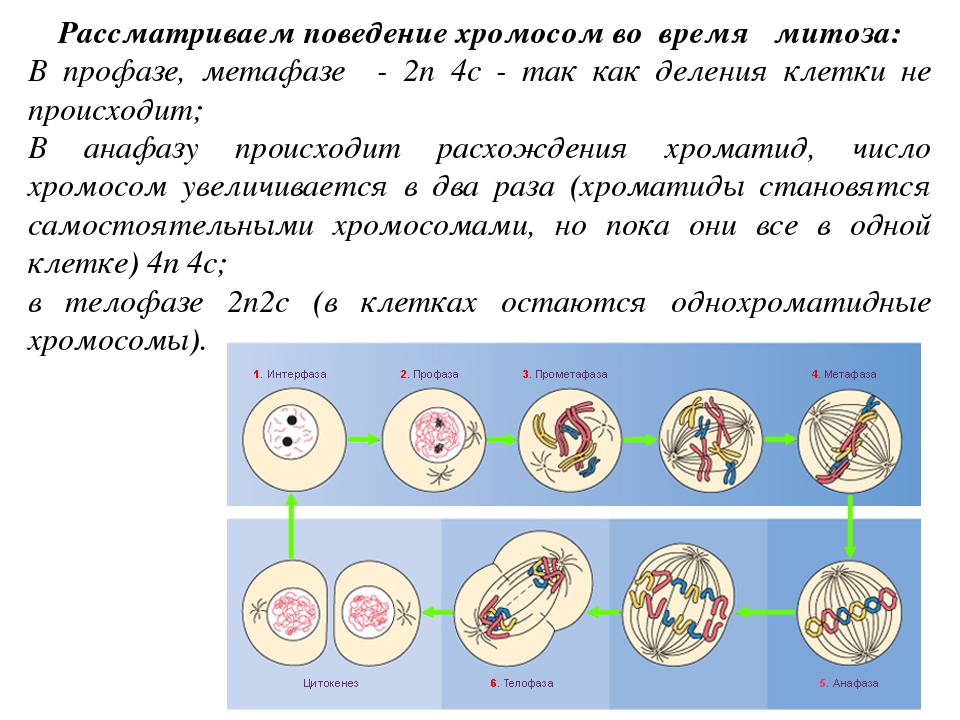 , 1993, с. 445
, 1993, с. 445 - ↑ Ченцов, 2004, с. 436
- ↑ Ченцов, 2004, с. 436—437
- ↑ 1 2 3 Ченцов, 2004, с. 436
- ↑ 1 2 Албертс и др., 1993, с. 448
- ↑ Албертс и др., 1993, с. 449
- ↑ Алов, 1972, с. 108
- ↑ Алов, 1972, с. 112
- ↑ 1 2 3 Ченцов, 2004, с. 439
- ↑ Алов, 1972, с. 113
- ↑ Албертс и др., 1993, с. 451
- ↑ 1 2 Албертс и др., 1993, с. 452
- ↑ Ченцов, 2004, с. 440
- ↑ Алов, 1972, с. 119
- ↑ 1 2 3 4 Албертс и др., 1993, с. 453
- ↑ Ченцов, 2004, с. 441
- ↑ Албертс и др.
 , 1993, с. 454
, 1993, с. 454 - ↑ 1 2 3 Ченцов, 2004, с. 442
- ↑ Алов, 1972, с. 135
- ↑ Албертс и др., 1993, с. 457
- ↑ Алов, 1972, с. 137
- ↑ 1 2 Албертс и др., 1993, с. 458
- ↑ Алов, 1972, с. 140
- ↑ Албертс и др., 1993, с. 459
- ↑ Албертс и др., 1993, с. 460
- ↑ Албертс и др., 1993, с. 461
- ↑ Алов, 1972, с. 192
- ↑ Алов, 1972, с. 193
- ↑ 1 2 Алов, 1972, с. 167
- ↑ 1 2 3 Алов, 1972, с. 169
- ↑ Алов, 1972, с. 170
- ↑ Алов, 1972, с. 171
- ↑ 1 2 Алов, 1972, с. 172
- ↑ 1 2 Алов, 1972, с.
 174
174 - ↑ 1 2 Алов, 1972, с. 176
- ↑ Алов, 1972, с. 177
- ↑ 1 2 Алов, 1972, с. 183
- ↑ Алов, 1972, с. 184
- ↑ 1 2 3 Алов, 1972, с. 185
- ↑ Алов, 1972, с. 186
- ↑ 1 2 Алов, 1972, с. 188
Литература
- Alberts B. at al. Molecular biology of the cell. — 5th edition. — Garland science, 2008. — 1601 p. — ISBN 978-0-8153-4105
- Morgan D. O. The cell cycle: principles of control. — New science press, 2007. — 297 p. — ISBN 978-0-9539181-2-6
- Алов И. А. Цитофизиология и патология митоза. — М.: «Медицина», 1972. — 264 с. — 3700 экз.
- Албертс Б. и др. Молекулярная биология клетки: В 3-х т. — 2-ое, переработанное.
 — М.: «Мир», 1993. — Т. 2. — 539 с. — ISBN 5-03-001987-1
— М.: «Мир», 1993. — Т. 2. — 539 с. — ISBN 5-03-001987-1
- Гилберт С. Биология развития: в 3-х томах. — М.: «Мир», 1995. — Т. 3. — 352 с. — 5000 экз. — ISBN 5-03-001833-6
- История биологии с древнейших времён до начала XX века / Под редакцией С. Р. Микулинского. — М.: «Наука», 1972. — 564 с. — 9600 экз.
- Райков И. Б. Ядро простейших. Морфология и эволюция. — Л.: «Наука», 1978. — 328 с. — 1600 экз.
- Седова Т. В. Кариология водорослей. — СПб.: «Наука», 1996. — 386 с. — 500 экз. — ISBN 5-02-026058-4
- Ченцов Ю. С. Введение в клеточную биологию: Учебник для вузов. — 4-ое, переработанное и дополненное. — М.: ИКЦ «Академкнига», 2004. — 495 с. — 3000 экз. — ISBN 5-94628-105-4
Ссылки
Иллюстрации
Анимация
Видео
Цитогенетический анализ клеток костного мозга (кариотип)
Цитогенетический анализ клеток костного мозга (кариотип) – исследование, целью которого является оценка количества и структуры хромосом клеток кроветворной ткани.
Синонимы русские
Стандартное кариотипирование клеток костного мозга, стандартная цитогенетика в метафазных пластинках.
Синонимы английские
Chromosome Analysis, Bone Marrow, Karyotype, Cytogenetics, Cytogenetic Analysis, Chromosome Studies, Chromosome Karyotype.
Метод исследования
Дифференциальное окрашивание хромосом.
Какой биоматериал можно использовать для исследования?
Костный мозг.
Общая информация об исследовании
Взятие костного мозга для исследования проводится посредством пункции кости. Манипуляцию выполняет врач. Чаще всего костный мозг аспирируется из грудины или задних остей подвздошных костей. После выбора места пункции и обработки кожи над ним раствором антисептика врач проводит послойную местную анестезию до надкостницы. После введения анестетика ждут наступления анестезии не менее одной минуты. Пункцию выполняют специальной иглой, которая имеет внутри стержень. Её продвигают сквозь мягкие ткани до надкостницы, затем с усилием внедряют вглубь костного вещества. После этого убирают стержень и шприцем набирают костный мозг. Во время аспирации возможно возникновение болевых ощущений из-за перепада давления в полости кости. Полученный костный мозг переливают в вакутейнер, содержащий гепарин натрия, и отправляют в лабораторию.
Её продвигают сквозь мягкие ткани до надкостницы, затем с усилием внедряют вглубь костного вещества. После этого убирают стержень и шприцем набирают костный мозг. Во время аспирации возможно возникновение болевых ощущений из-за перепада давления в полости кости. Полученный костный мозг переливают в вакутейнер, содержащий гепарин натрия, и отправляют в лабораторию.
Большинство опухолей кроветворной системы связаны с возникновением генетических изменений в столовой клетке крови, которые приводят к её злокачественной трансформации и появлению опухолевого клона. Генетические нарушения специфичны для определенного заболевания, поэтому их обнаружение считается достоверным диагностическим критерием. Кроме того, наличие некоторых генетических аномалий может влиять на прогноз заболевания, его чувствительность к тем или иным способам лечения и, соответственно, помогает выбрать наилучшую тактику терапии. Стандартное цитогенетическое исследование клеток костного мозга представляет собой изучение количества и структуры хромосом клеток и позволяет выявить геномные (нарушения в количестве хромосом) и хромосомные (изменения в их структуре) мутации.
Хромосомы – структуры в ядре клетки, в которых компактно собрана генетическая информация в виде ДНК. Когда клетка находится в состоянии покоя, хромомосы упакованы в клеточном ядре и визуально различить их друг от друга невозможно. Однако в одну из фаз деления клетки, которая называется метафаза, хромосомы выстраиваются в одну плоскость в центре клетки, формируя так называемую метафазную пластинку. Именно такие метафазные пластинки и изучаются в процессе цитогенетического исследования. Кроветворные клетки костного мозга постоянно делятся, поэтому часть из них во время взятия как раз будет находиться в метафазе своего деления. Преимущество такого метода цитогенетического исследования в том, что оно не требует дополнительных воздействий на клетки для стимуляции деления и получения метафаз. После обработки биоматериала различными химическими веществами клетки фиксируют на предметных стеклах и производят окраску. Существует несколько методов окраски хромосом, каждый из которых позволяет лучше дифференцировать те или иные особенности их строения. Хромосомы анализируется как с помощью микроскопа, так и с использованием компьютерных аналитических систем, которые существенно повышают качество и скорость исследования.
Хромосомы анализируется как с помощью микроскопа, так и с использованием компьютерных аналитических систем, которые существенно повышают качество и скорость исследования.
Кариотипирование проводится как минимум на двадцати хорошо окрашенных, полных метафазных пластинках с хорошим разбросом хромосом. Исследователь подсчитывает их количество и проводит оценку структуры каждой путем сопоставления двух аналогичных хромосом и сравнения их с цитогенетическими картами.
Для чего используется исследование?
- Для выявления хромосомных аномалий в клетках костного мозга при диагностике и последующем мониторинге эффективности лечения при заболеваниях системы кроветворения.
Когда назначается исследование?
- Для верификации диагноза и определения прогностических факторов при злокачественных новообразованиях гемопоэтической ткани (острые и хронические лейкозы, лимфомы, множественная миелома и другие моноклональные гаммапатии, миелодиспластические синдромы).

- В целях мониторинга эффективности лечения.
- В целях мониторинга сохранения ремиссии заболевания.
Что означают результаты?
Нормальный кариотип человека выглядит следующим образом:
-
46,ХХ – нормальный кариотип женщины,
-
46,XY – нормальный кариотип мужчины,
— где первая цифра — это общее количество хромосом, а после запятой перечислены половые хромосомы.
Количество проанализированных метафаз, а также метафаз, в которых выявлены мутации, записывается в квадратных скобках. Для записи генетических аномалий существуют специальные правила, соответственно которым будет написан результат исследования при выявлении какой-либо мутации.
Количественные аномалии хромосом записываются как их общее число, перечисление половых хромосом и номер отсутствующей или лишней хромосомы с соответствующим знаком минус или плюс.
Для обозначения структурных аномалий используются специальные буквенные символы:
-
t – транслокация – участок одной хромосомы переносится на другую,
-
del – делеция – потеря участка хромосомы,
-
inv – инверсия – поворот участка хромосомы на 180 градусов,
— после которых указывается номер хромосомы, на которой они обнаружены, а также ее участок.
В заключении цитогенетического исследования содержится информация о виде мутации и доле метафазных пластинок, в которой она выявлена.
Что может влиять на результат?
- Соблюдение правил взятия и преаналитической подготовки биоматериала, опыт и квалификация врача-цитогенетика.
- При аспирации костного мозга для нескольких анализов последние его порции могут быть значительно разведены периферической кровью. Поэтому материал для цитогенетического исследования необходимо набирать сразу после пункции. Разведение кровью приведет к невозможности получить необходимое количество метафазных пластинок, так как клетки периферической крови не находятся в состоянии деления.
- Недостаточное количество костного мозга также может быть причиной невозможности правильного выполнения анализа.
Дидактический материал по биологии (ЕГЭ линия 27)
ПРИЛОЖЕНИЕ 8
ЗАДАНИЕ №27 ЕГЭ
РЕШЕНИЕ ЗАДАЧ ПО ДЕЛЕНИЮ КЛЕТКИ (МИТОЗ, МЕЙОЗ) НА ПРИМЕНЕНИЕ ЗНАНИЙ
НЕОБХОДИМЫЕ ПОЯСНЕНИЯ:
В задачах на определение числа хромосом или ДНК в
клетках или организме частично правильный элемент ответа не может оцениваться в
1 балл.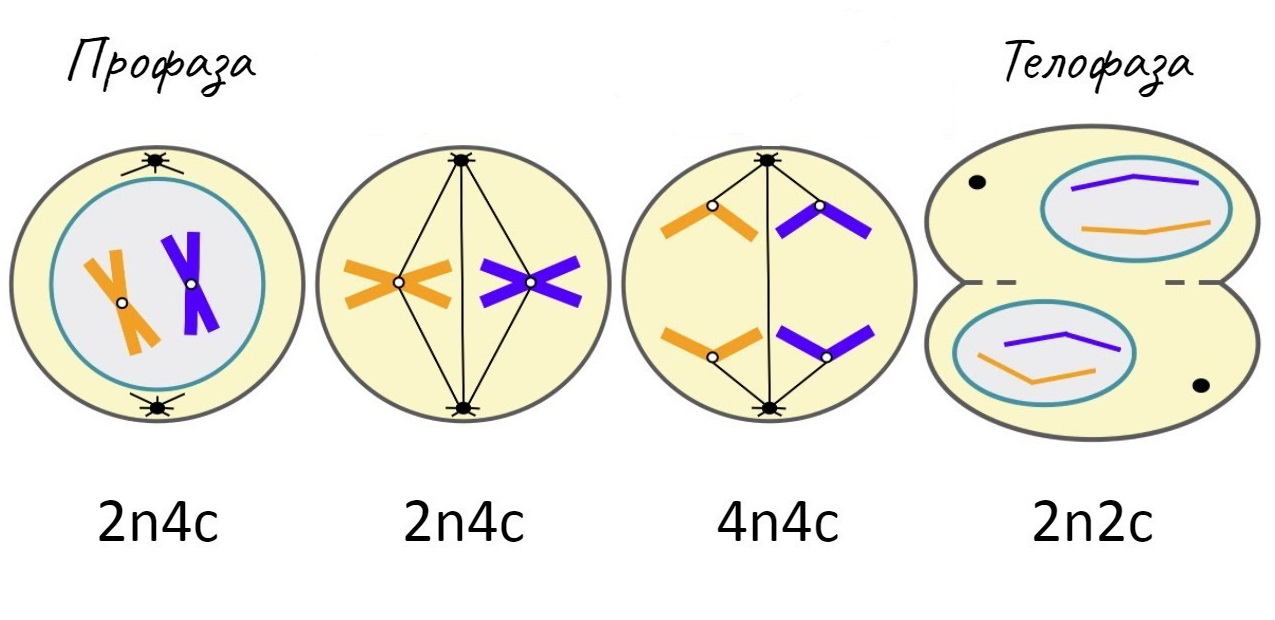 Ответ участника должен соответствовать требованиям эталона.
Ответ участника должен соответствовать требованиям эталона.
По материалам курса февраль 2018 наборы числа хромосом и ДНК в митозе и мейозе:
Таблица 1
|
ФАЗА МИТОЗА |
ЧИСЛО ХРОМОСОМ И ДНК |
ФАЗА МЕЙОЗ |
ЧИСЛО ХРОМОСОМ И ЧИСЛО ДНК |
|
интерфаза |
|
интерфаза |
|
|
профаза |
|
профазаI |
|
|
метофаза |
|
метофазаI |
|
|
анафаза |
|
анафазаI |
|
|
телофаза |
|
телофазаI |
|
|
|
|
профазаII |
|
|
|
|
метофазаII |
|
|
|
|
анафазаII |
|
|
|
|
телофазаII |
|
Пример 1: Какой хромосомный набор характерен для
макроспоры, из которой в дальнейшем формируется восьмиядерный зародышевый
мешок и яйцеклетка цветкового растения? Объясните, из каких исходных клеток и в
результате какого деления образуются микроспора и яйцеклетка.
|
Содержание верного ответа и указания по оцениванию (правильный ответ должен содержать следующие позиции) |
Баллы |
|
Схема решения задачи включает: 1) набор хромосом макроспоры – n, яйцеклетки – n; 2) микроспора образуется из клетки макроспорангия семязачатка путём мейоза; 3) яйцеклетка образуется из макроспоры путём митоза |
|
|
Ответ включает в себя все названные выше элементы и не содержит биологических ошибок |
3 |
|
Ответ включает в себя два из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает в себя три названных выше элемента, но содержит биологические ошибки |
2 |
|
Ответ включает в себя один из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает в себя два из названных выше элементов, но содержит биологические ошибки |
1 |
|
Ответ неправильный |
0 |
|
Максимальный балл |
3 |
Пример 2:
Хромосомный набор соматических клеток пшеницы равен 28. Определите хромосомный
набор и число молекул ДНК в одной из клеток семязачатка перед началом мейоза, в
анафазе мейоза 1 и в анафазе мейоза 2. Объясните, какие процессы происходят в
эти периоды и как они влияют на изменение числа ДНК и хромосом.
Определите хромосомный
набор и число молекул ДНК в одной из клеток семязачатка перед началом мейоза, в
анафазе мейоза 1 и в анафазе мейоза 2. Объясните, какие процессы происходят в
эти периоды и как они влияют на изменение числа ДНК и хромосом.
|
Содержание верного ответа и указания по оцениванию (правильный ответ должен содержать следующие позиции) |
Баллы |
|
Элементы ответа: 1) перед началом мейоза число молекул ДНК – 56, происходит репликация и число ДНК удваивается, число хромосом не изменяется – 28, но каждая хромосома состоит из двух хроматид; 2) в анафазе I мейоза число молекул ДНК – 56, число хромосом – 28, к полюсам клетки расходятся гомологичные хромосомы, но все хромосомы находятся в одной клетке; 3) в анафазе II мейоза число молекул ДНК – 28, хромосом – 28, после первого мейотического деления число ДНК и хромосом уменьшилось в 2 раза, к полюсам клетки расходятся сестринские хромосомы (дочерние однохроматидные хромосомы) |
|
|
Ответ включает в себя все названные выше элементы и не содержит биологических ошибок |
3 |
|
Ответ включает в себя два из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает в себя три названных выше элемента, но содержит биологические ошибки |
2 |
|
Ответ включает в себя один из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает в себя два из названных выше элементов, но содержит биологические ошибки |
1 |
|
Ответ неправильный |
0 |
|
Максимальный балл |
3 |
Пример 3:
В кариотипе одного из видов рыб 56 хромосом. Определите число хромосом и
молекул ДНК в клетках при овогенезе в зоне роста в конце интерфазы и в конце
зоны созревания гамет. Объясните полученные результаты.
Определите число хромосом и
молекул ДНК в клетках при овогенезе в зоне роста в конце интерфазы и в конце
зоны созревания гамет. Объясните полученные результаты.
|
Содержание верного ответа и указания к оцениванию (правильный ответ должен содержать следующие позиции) |
Баллы |
|
Элементы ответа: 1) в зоне роста в период интерфазы в клетках число хромосом 56; число молекул ДНК равно 112; 2) в зоне окончательного созревания гамет в клетках 28 хромосом; число молекул ДНК – 28; 3) в зоне роста в период интерфазы число хромосом не изменяется; число молекул ДНК удваивается за счёт репликации; 4) в конце
зоны созревания гамет происходит мейоз, число хромосом уменьшается в 2 раза,
образуются гаплоидные клетки – гаметы, каждая хромосома содержит одну
молекулу ДНК. |
|
|
Ответ правильный и полный, включает все названные выше элементы |
3 |
|
Ответ включает два-три из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает четыре названных выше элемента, но содержит биологические ошибки |
2 |
|
Ответ включает один из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает два-три из названных выше элементов, но содержит биологические ошибки |
1 |
|
Ответ неправильный |
0 |
|
Максимальный балл |
3 |

Пример 4: Какой хромосомный набор характерен для клеток
спороносных побегов и заростка плауна? Объясните, из каких исходных клеток и в
результате какого деления они образуются.
|
Содержание верного ответа и указания к оцениванию (правильный ответ должен содержать следующие позиции) |
Баллы |
|
Элементы ответа: 1) в клетках спороносных побегов диплоидный набор хромосом – 2n; 2) в клетках заростка гаплоидный набор хромосом – n; 3) спороносные побеги развиваются на взрослом растении в результате митоза клеток спорофита; 4) заросток развивается из споры в результате митоза |
|
|
Ответ правильный и полный, включает все названные выше элементы |
3 |
|
Ответ включает два-три из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает четыре названных выше элемента, но содержит биологические ошибки |
2 |
|
Ответ включает один из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает два-три из названных выше элементов, но содержит биологические ошибки |
1 |
|
Ответ неправильный |
0 |
|
Максимальный балл |
3 |
Пример 5: Какой
хромосомный набор характерен для клеток восьмиядерного зародышевого мешка и
зародышевой почечки семени пшеницы.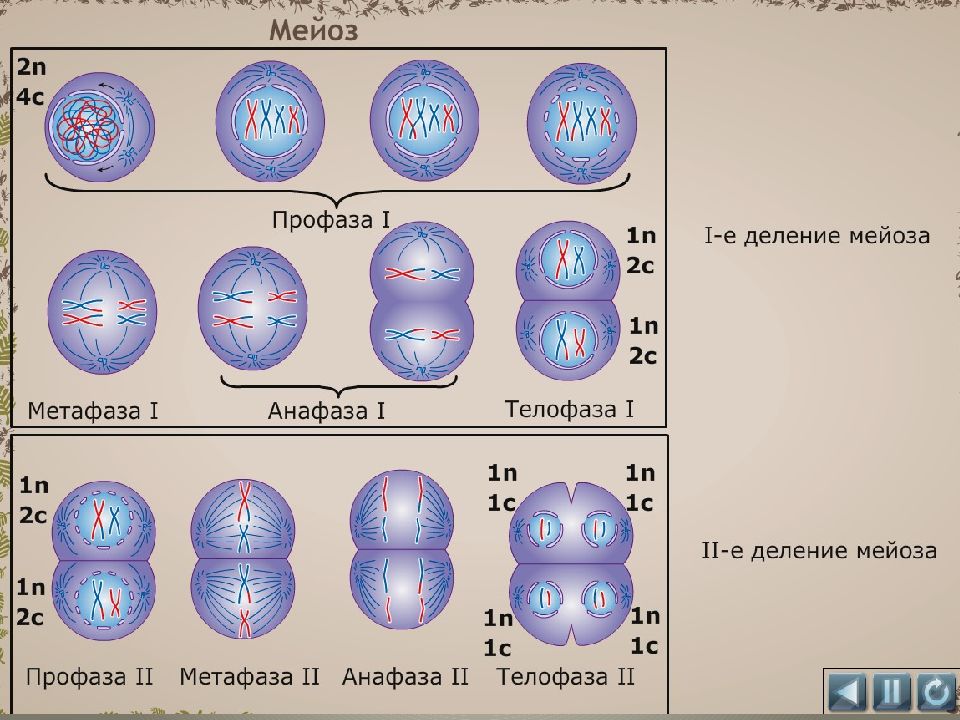 Объясните, из каких исходных клеток и в
результате какого деления они образуются.
Объясните, из каких исходных клеток и в
результате какого деления они образуются.
|
Содержание верного ответа и указания по оцениванию (правильный ответ должен содержать следующие позиции) |
Баллы |
|
Элементы ответа: 1) клетки восьмиядерного зародышевого мешка гаплоидные – n; 2) в клетках зародышевой почечки диплоидный набор хромосом – 2n; 3) клетки зародышевой почечки развиваются из зиготы в результате митоза; 4) клетки восьмиядерного зародышевого мешка развиваются из женской споры митозом |
|
|
Ответ правильный и полный, включает все названные выше элементы |
3 |
|
Ответ включает два-три из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает четыре названных выше элемента, |
2 |
|
Ответ включает один из названных выше элементов и не содержит биологических ошибок, ИЛИ ответ включает два-три из названных выше элементов,
|
1 |
|
Ответ неправильный |
0 |
|
Максимальный балл |
3 |
ЗАДАЧИ ДЛЯ САМОСТОЯТЕЛЬНОЙ РАБОТЫ
1. Общая масса всех молекул ДНК в 46 соматических хромосомах одной
соматической клетки человека составляет 6х10-9 мг. Определите, чему равна масса
всех молекул ДНК в сперматозоиде и в соматической клетке перед началом деления
и после его окончания. Ответ поясните.
Общая масса всех молекул ДНК в 46 соматических хромосомах одной
соматической клетки человека составляет 6х10-9 мг. Определите, чему равна масса
всех молекул ДНК в сперматозоиде и в соматической клетке перед началом деления
и после его окончания. Ответ поясните.
2. Какое деление мейоза сходно с митозом? Объясните, в чем оно выражается. К какому набору хромосом в клетке приводит мейоз.
3. Объясните, в чем заключается сходство и различие мутационной и комбинативной изменчивости.
4. Какой хромосомный набор характерен для клеток зародыша и эндосперма семени, листьев цветкового растения. Объясните результат в каждом случае.
5. Хромосомный набор соматических клеток пшеницы равен 28. Определите хромосомный набор и число молекул ДНК в одной из клеток семязачатка перед началом мейоза, в анафазе мейоза 1 и в анафазе мейоза 2. Объясните, какие процессы происходят в эти периоды и как они влияют на изменение числа ДНК и хромосом.
6. Какой хромосомный набор характерен для ядер клеток эпидермиса листа
и восьмиядерного зародышевого мешка семязачатка цветкового растения? Объясните,
из каких исходных клеток и в результате какого деления образуются эти клетки.
Какой хромосомный набор характерен для ядер клеток эпидермиса листа
и восьмиядерного зародышевого мешка семязачатка цветкового растения? Объясните,
из каких исходных клеток и в результате какого деления образуются эти клетки.
7. Синдром Дауна у человека проявляется при трисомии по 21 паре хромосом. Объясните причины появления такого хромосомного набора у человека.
8. Укажите число хромосом и количество молекул ДНК в профазе первого и второго мейотического деления клетки. Какое событие происходит с хромосомами в профазе первого деления?
9. Для соматической клетки животного характерен диплоидный набор хромосом. Определите хромосомный набор (n) и число молекул ДНК (с) в клетке в профазе мейоза I и метафазе мейоза II. Объясните результаты в каждом случае.
10. Какой хромосомный набор характерен для гамет и спор растения мха кукушкина льна? Объясните, из каких клеток и в результате какого деления они образуются.
11.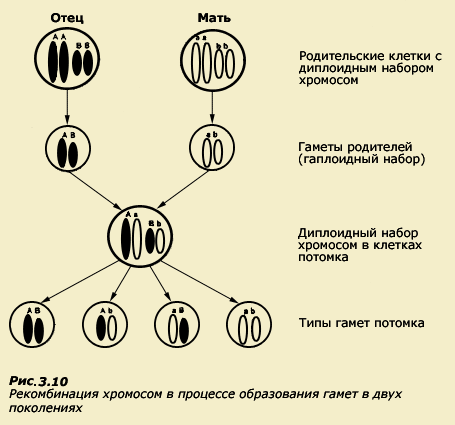 Соматические клетки дрозофилы содержат 8 хромосом. Как изменится
число хромосом и молекул ДНК в ядре при гаметогенезе перед началом деления и в
конце телофазы мейоза I? Объясните результаты в каждом случае.
Соматические клетки дрозофилы содержат 8 хромосом. Как изменится
число хромосом и молекул ДНК в ядре при гаметогенезе перед началом деления и в
конце телофазы мейоза I? Объясните результаты в каждом случае.
12. Для соматической клетки животного характерен диплоидный набор хромосом. Определите хромосомный набор (n) и число молекул ДНК(с) в клетке в конце телофазы мейоза I и анафазе мейоза II. Объясните результаты в каждом случае.
13. Каковы особенности мейоза, которые обеспечивают разнообразие гамет?
Укажите не менее трёх особенностей.
14. Для соматической клетки животного характерен диплоидный набор хромосом. Определите хромосомный набор (n) и число молекул ДНК (с) в клетке перед мейозом I и профазе мейоза II. Объясните результаты в каждом случае.
15. Какой хромосомный набор характерен для гаметофита и гамет мха сфагнума? Объясните из каких исходных клеток и в результате какого деления образуются эти клетки?
16. Укажите число хромосом и количество молекул ДНК в профазе
первого и второго мейотического деления клетки. Какое событие происходит
с хромосомами в профазе первого деления?
Укажите число хромосом и количество молекул ДНК в профазе
первого и второго мейотического деления клетки. Какое событие происходит
с хромосомами в профазе первого деления?
17. У крупного рогатого скота в соматических клетках 60 хромосом. Определите число хромосом и молекул ДНК в клетках яичников при овогенезе в интерфазе перед началом деления и после деления мейоза I. Объясните полученные результаты на каждом этапе.
18. В соматических клетках дрозофилы содержится 8 хромосом. Какое число хромосом и молекул ДНК содержится в ядре при гаметогенезе перед началом мейоза I и мейоза II? Объясните, как образуется такое число хромосом и молекул ДНК.
21. Какой хромосомный набор характерен для вегетативной, генеративной
клеток и спермиев пыльцевого зерна цветкового растения? Объясните, из
каких исходных клеток и в результате какого деления образуются
эти клетки.
22. Соматические клетки кролика содержат 44 хромосомы. Как изменится число хромосом и молекул ДНК в ядре при гаметогенезе перед началом деления и в конце телофазы мейоза I? Объясните результаты в каждом случае.
23. В соматических клетках мухи дрозофилы содержится 8 хромосом. Определите число хромосом и молекул ДНК в клетках при сперматогенезе в зоне размножения и в конце зоны созревания гамет. Ответ обоснуйте. Какие процессы происходят в этих зонах?
24. Какой хромосомный набор характерен для споры, гаметофита и спорофита мха кукушкин лён? Из каких исходных клеток и в результате какого деления образуются эти стадии развития мха?
25. Какой хромосомный набор характерен для клеток мякоти иголок и спермиев сосны? Объясните, из каких исходных клеток и в результате какого деления образуются эти клетки
26. Общая масса всех молекул ДНК в 46 хромосомах одной соматической
клетки человека составляет около 6·10−9 мг. Определите
массу всех молекул ДНК в ядре клетки при сперматогенезе перед началом
мейоза и после окончания мейоза. Объясните полученные результаты.
Определите
массу всех молекул ДНК в ядре клетки при сперматогенезе перед началом
мейоза и после окончания мейоза. Объясните полученные результаты.
27. Какой хромосомный набор характерен для вегетативной, генеративной клеток и спермиев пыльцевого зерна цветкового растения? Объясните, из каких исходных клеток и в результате какого деления образуются эти клетки.
28. Общая масса всех молекул ДНК в 46 хромосомах одной соматической клетки человека составляет около 6 · 10−9 мг. Определите, чему равна масса всех молекул ДНК в ядре при сперматогенезе перед началом мейоза, после мейоза I и мейоза II. Объясните полученные результаты.
29. Кариотип собаки включает 78 хромосом. Определите число хромосом
и число молекул ДНК в клетках при овогенезе в зоне размножения и в
конце зоны созревания гамет. Какие процессы происходят в этих зонах?
Ответ обоснуйте (в ответе должно содержаться четыре критерия).
30. У полевой мыши 40 хромосом. Сколько хромосом у самца мыши в сперматогониях, с которых начинается формирование сперматозоидов, в зрелых сперматозоидах и в клетках зародыша? Какое деление приводит к образованию этих клеток? Из каких клеток они образуются?
31. В кариотипе яблони 34 хромосомы. Сколько хромосом и ДНК будет содержаться в яйцеклетке яблони, клетках эндосперма её семени и клетках листа? Из каких клеток образуются указанные клетки?
32. Хромосомный набор соматических клеток картофеля равен 48. Определите хромосомный набор и число молекул ДНК в клетках при мейозе в профазе мейоза I и метафазе мейоза II. Объясните все полученные результаты.
33. Масса молекул ДНК в соматических клетках равна 6*10-9. Чему равна масса молекул ДНК перед началом мейоза и в анафазе мейоза 1, ответ поясните.
34. В соматических клетках овса 42 хромосомы. Определите хромосомный
набор и количество молекул ДНК перед началом мейоза I и в метафазе мейоза II.
Ответ поясните.
Определите хромосомный
набор и количество молекул ДНК перед началом мейоза I и в метафазе мейоза II.
Ответ поясните.
35. У хламидомонады преобладающим поколением является гаметофит. Определите хромосомный набор взрослого организма и его гамет. Объясните из каких исходных клеток образуются взрослые особи и их гаметы, в результате какого деления формируются половые клетки.
36. Количество хромосом в соматической клетке дрозофилы — 8. Определите число хромосом и молекул ДНК у дрозофилы в период овогенеза в анафазе мейоза 1 и профазе мейоза 2. Объясните результаты.
37. Какой хромосомный набор характерен для клеток восьмиядерного зародышевого мешка и зародышевой почечки семени пшеницы. Объясните, из каких исходных клеток и в результате какого деления они образуются.
38. Какой хромосомный набор характерен для вегетативной, генеративной
клеток и спермиев пыльцевого зерна вишни? Объясните, из каких исходных клеток и
в результате какого деления образуются эти клетки.
39. У шимпанзе в соматических клетках 48 хромосом. Определите хромосомный набор и число молекул ДНК в клетках перед началом мейоза, в анафазе мейоза I и в профазе мейоза II. Объясните ответ в каждом случае.
40. Укажите:1) способы деления клеток при образовании микроспор из спорогенной ткани;
2) способы деления при образовании вегетативной и генеративной клеток;3) число хромосом и молекул ДНК в микроспоре, вегетативной и генеративной клетках покрытосеменного растения (выразить формулой).
41. Как изменяется число хромосом и ДНК в клетке мужчины в процессе сперматогенеза на стадиях: интерфаза I , телофаза I, анафаза II, телофаза II.
42. В соматической клетке кукурузы 20 хромосом. Определить набор хромосом в клетке верхушки растения и в ядре пыльцевого зерна.
43. Какой хромосомный набор характерен для мегаспоры и клеток
эндосперма сосны? Объясните, из каких исходных клеток и в результате, какого
деления образуются эти клетки.
44. Какой хромосомный набор характерен для клеток чешуй женских шишек и женской споры ели? Объясните, из каких исходных клеток и в результате какого деления образуются клетки шишки и мегаспора ели.
45. Какой хромосомный набор характерен для мегаспоры и клеток эндосперма лиственницы? Объясните, из каких исходных клеток и в результате какого деления образуются эти клетки.
46. Какой хромосомный набор характерен для заростка и зародыша плауна? Объясните, из каких исходных клеток и в результате какого деления образуются клетки заростка и зародыша плауна?
47. Определите хромосомный набор макроспоры, из которой формируется восьмиядерный зародышевый мешок, и яйцеклетки. Объясните, из каких клеток и каким делением образованы макроспора и яйцеклетка.
48. Для соматической клетки животного характерен диплоидный набор
хромосом. Определите хромосомный набор (n) и число молекул ДНК (с) в ядре
клетки при гаметогенезе в метафазе I мейоза и анафазе II мейоза. Объясните
результаты в каждом случае.
Объясните
результаты в каждом случае.
Задачи по теме «Размножение: мейоз, двойное оплодотворение»
1.Хромосомный набор соматических клеток пшеницы равен 28. Определите хромосомный набор и число молекул ДНК в одной из клеток семязачатка перед началом мейоза, в анафазе мейоза 1 и в анафазе мейоза 2. Объясните, какие процессы происходят в эти периоды и как они влияют на изменение числа ДНК и хромосом ?
Элементы ответа:
1) Клетки семязачатка содержат диплоидный набор хромосом – 28 (2n2c). Перед началом мейоза в S-периоде интерфазы — удвоение ДНК: 28 хромосом, 56 ДНК (2n4c).
2) В анафазе мейоза 1 – к полюсам клетки расходятся хромосомы, состоящие из двух хроматид. Генетический материал клетки будет (2n4c = n2c+n2c) — 28 хромосом, 56 ДНК .
3) В мейоз 2 вступают 2 дочерние клетки с гаплоидным набором хромосом (n2c) — 14 хромосом,28ДНК . В анафазе мейоза 2– к полюсам клетки расходятся хроматиды. После расхождения хроматид число хромосом увеличивается в 2 раза (хроматиды становятся самостоятельными хромосомами, но пока они все в одной клетке) – (2n2с= nc+nc) – 28 хромосом,28ДНК
2. В кариотипе клевера 14 хромосомы. Сколько хромосом и ДНК будет содержаться в яйцеклетке клевера, клетках эндосперма его семени и клетках листа?
В кариотипе клевера 14 хромосомы. Сколько хромосом и ДНК будет содержаться в яйцеклетке клевера, клетках эндосперма его семени и клетках листа?
Элементы ответа:
1) 7 хромосом и 7 ДНК будет содержаться в яйцеклетке (nc)
2) В клетках эндосперма 21 хромосома и 21 ДНК (3n3c)
3) В клетках листа 14 хромосом 14ДНК (2n2c) О
3.Количество хромосом в соматической клетке дрозофилы а 8. Определите число хромосом и молекул ДНК у дрозофилы в период овогенеза в анафазе мейоза 1 и профазе мейоза 2. Объясните результаты.
Элементы ответа:
1) В соматических клетках диплоидный набор хромосом — 8 хромосом, 8 молекул ДНК.
2) При овогенезе в анафазе мейоза I — 8 хромосом 16 хроматид. В интерфазе — репликация ДНК (число молекул ДНК становится вдвое больше). Далее I деление мейоза — редукционное, число хромосом и молекул ДНК на каждом полюсе клетки в анафазе I будет меньше вдвое (гомологичные хромосомы расходятся к полюсам клетки).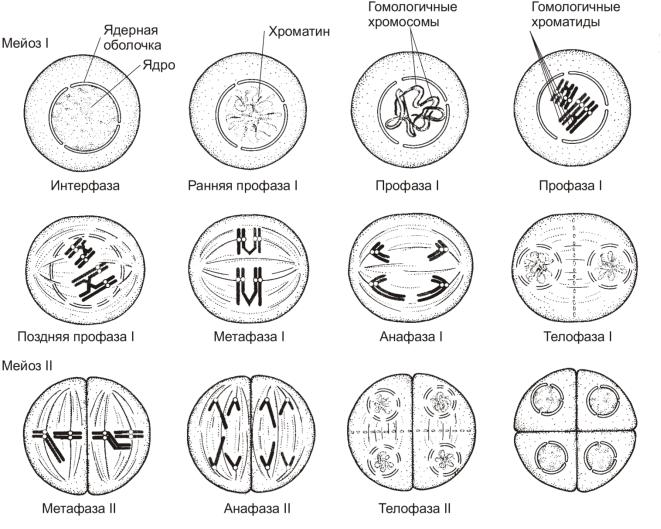
3) При овогенезе в профазе II — 4 хромосомы, 8 молекул ДНК. В телофазе I образуются две гаплоидные клетки, хромосомы двухроматидные (1n2с). После редукционного деления репликации ДНК не происходит, число хромосом и молекул ДНК остаётся таким же (1n2c), как в телофазе I. То есть в профазе II набор 1n2c. Ответ включает два названных выше элемента и не содержит
4.Для соматической клетки животного характерен диплоидный набор хромосом. Определите хромосомный набор (n) и число молекул ДНК (с) в клетке перед мейозом I и профазе мейоза II. Объясните результаты в каждом случае.
Элементы ответа:
1) Диплоидный набор хромосом 2n2c . Перед началом мейоза в S-периоде интерфазы — удвоение ДНК: Профаза мейоза I – 2n4с
2) Первое деление редукционное. В мейоз 2 вступают 2 дочерние клетки с гаплоидным набором хромосом (n2c)
3) Метафаза мейоза II — хромосомы выстраиваются на экваторе n2с
5.Найдите ошибки в приведенном тексте. Укажите номера предложений, в которых сделаны ошибки, исправьте их.
1. Оплодотворение у цветковых растений имеет свои особенности. 2. В завязи цветка образуются гаплоидные пыльцевые зерна. 3. Гаплоидное ядро пыльцевого зерна делится на два ядра – генеративное и вегетативное. 4. Генеративное ядро делится на два спермия. 5. Спермии направляются к пыльнику. 6. Один из них оплодотворяет находящуюся там яйцеклетку, а другой центральную клетку. 7. В результате двойного оплодотворения из зиготы развивается диплоидный зародыш семени, а из центральной клетки триплоидный эндосперм
Элементы ответа:
Ошибки допущены в предложениях 2, 5, 6.
1) 2 – пыльцевые зерна образуются в пыльниках тычинок.
2) 5 – спермии направляются к завязи цветка.
3) 6 – яйцеклетки находятся в завязи цветка, а не в пыльниках
6.Общая масса всех молекул ДНК в 46 соматических хромосомах одной соматической клетки человека составляет 6х10-9 мг. Определите, чему равна масса всех молекул ДНК в сперматозоиде и в соматической клетке перед началом деления и после его окончания. Ответ поясните
Ответ поясните
1) В половых клетках 23 хромосомы, т.е. в два раза меньше, чем в соматических, поэтому масса ДНК в сперматозоиде в два раза меньше и составляет 6х 10-9 : 2 = 3х 10-9мг.
2) Перед началом деления (в интерфазе) количество ДНК удваивается и масса ДНК равна 6х 10-9 х2 = 12 х 10-9мг.
3) После митотического деления в соматической клетке число хромосом не меняется и масса ДНК равна 6х 10-9 мг
Подразделение сотовой связи — AP Biology
Если вы считаете, что контент, доступный через Веб-сайт (как определено в наших Условиях обслуживания), нарушает одно
или больше ваших авторских прав, сообщите нам, отправив письменное уведомление («Уведомление о нарушении»), содержащее
в
информацию, описанную ниже, назначенному ниже агенту. Если репетиторы университета предпримут действия в ответ на
ан
Уведомление о нарушении, оно предпримет добросовестную попытку связаться со стороной, которая предоставила такой контент
средствами самого последнего адреса электронной почты, если таковой имеется, предоставленного такой стороной Varsity Tutors.
Ваше Уведомление о нарушении прав может быть отправлено стороне, предоставившей доступ к контенту, или третьим лицам, таким как в виде ChillingEffects.org.
Обратите внимание, что вы будете нести ответственность за ущерб (включая расходы и гонорары адвокатам), если вы существенно искажать информацию о том, что продукт или действие нарушает ваши авторские права. Таким образом, если вы не уверены, что контент находится на Веб-сайте или по ссылке с него нарушает ваши авторские права, вам следует сначала обратиться к юристу.
Чтобы отправить уведомление, выполните следующие действия:
Вы должны включить следующее:
Физическая или электронная подпись правообладателя или лица, уполномоченного действовать от их имени;
Идентификация авторских прав, которые, как утверждается, были нарушены;
Описание характера и точного местонахождения контента, который, по вашему мнению, нарушает ваши авторские права, в \
достаточно подробностей, чтобы позволить репетиторам Varsity найти и точно идентифицировать этот контент; например нам требуется
а
ссылка на конкретный вопрос (а не только на название вопроса), который содержит содержание и описание
к какой конкретной части вопроса — изображению, ссылке, тексту и т. д. — относится ваша жалоба;
Ваше имя, адрес, номер телефона и адрес электронной почты; и
Ваше заявление: (а) вы добросовестно считаете, что использование контента, который, по вашему утверждению, нарушает
ваши авторские права не разрешены законом, владельцем авторских прав или его агентом; (б) что все
информация, содержащаяся в вашем Уведомлении о нарушении, является точной, и (c) под страхом наказания за лжесвидетельство вы
либо владелец авторских прав, либо лицо, уполномоченное действовать от их имени.
д. — относится ваша жалоба;
Ваше имя, адрес, номер телефона и адрес электронной почты; и
Ваше заявление: (а) вы добросовестно считаете, что использование контента, который, по вашему утверждению, нарушает
ваши авторские права не разрешены законом, владельцем авторских прав или его агентом; (б) что все
информация, содержащаяся в вашем Уведомлении о нарушении, является точной, и (c) под страхом наказания за лжесвидетельство вы
либо владелец авторских прав, либо лицо, уполномоченное действовать от их имени.
Отправьте жалобу нашему уполномоченному агенту по адресу:
Чарльз Кон
Varsity Tutors LLC
101 S. Hanley Rd, Suite 300
St. Louis, MO 63105
Или заполните форму ниже:
1.3: Митоз — Биология LibreTexts
Цели обучения
- Перечислите и опишите ключевые события, которые происходят во время клеточного цикла и митоза.

- Опишите движение хромосом и хроматид во время митоза и роль микротрубочек.
- Определите содержание хромосом в клетках на различных этапах с учетом информации о n или 2n номерах клетки.
Копирование и деление хромосом — вызов клетки
Рисунок \ (\ PageIndex {1} \): ДНК, выделенная из хромосомы человека. Он показывает немного (не более 3%) одной молекулы ДНК, высвобожденной из одной хромосомы человека. (Хромосома была обработана для удаления гистонов). Помня, что это 3% ДНК только одной из 46 хромосом в диплоидной клетке человека, вы можете оценить проблему, с которой сталкивается клетка: как безошибочно разделить эти огромные отрезки ДНК, не создавая ужасных путаниц.(любезно предоставлено Дж. Р. Полсоном и У. К. Лэммли)
Решение этой проблемы:
- Дублируйте каждую хромосому во время фазы S клеточного цикла.
- Это дает диады, каждая из которых состоит из 2 идентичных сестринских хроматид.
 Они удерживаются вместе белковым кольцом, называемым когезином.
Они удерживаются вместе белковым кольцом, называемым когезином. - Уплотните хромосомы в компактную форму. Для этого требуются АТФ и белковые комплексы, называемые конденсинами.
- Разделите сестринские хроматиды и
- Распределите их поровну между двумя дочерними ячейками
Цикл клетки
Клеточный цикл — это упорядоченная серия событий, включающих рост и деление клеток, в результате которых образуются две новые дочерние клетки.Клетки на пути к клеточному делению проходят через серию точно рассчитанных и тщательно регулируемых стадий роста, репликации ДНК и деления, в результате которых образуются две идентичные (клонированные) клетки. Клеточный цикл состоит из двух основных фаз: интерфазы и митотической фазы. Во время интерфазы клетка растет, и ДНК реплицируется. Во время митотической фазы реплицированная ДНК и цитоплазматическое содержимое разделяются, и клетка делится.
Рисунок \ (\ PageIndex {2} \): клеточный цикл состоит из интерфазы и митотической фазы. Во время интерфазы клетка растет, и ядерная ДНК дублируется. За интерфазой следует митотическая фаза. Во время митотической фазы дублированные хромосомы разделяются и распределяются по дочерним ядрам. Цитоплазма также обычно делится, в результате чего образуются две дочерние клетки. (OpenStaxCNX)
Во время интерфазы клетка растет, и ядерная ДНК дублируется. За интерфазой следует митотическая фаза. Во время митотической фазы дублированные хромосомы разделяются и распределяются по дочерним ядрам. Цитоплазма также обычно делится, в результате чего образуются две дочерние клетки. (OpenStaxCNX) Клеточный цикл — это упорядоченная серия событий, включающих рост и деление клеток, в результате которых образуются две новые дочерние клетки. Клетки на пути к клеточному делению проходят через серию точно рассчитанных и тщательно регулируемых стадий роста, репликации ДНК и деления, в результате которых образуются две идентичные (клонированные) клетки.Клеточный цикл состоит из двух основных фаз: интерфазы и митотической фазы. Во время интерфазы клетка растет, и ДНК реплицируется. Во время митотической фазы реплицированная ДНК и цитоплазматическое содержимое разделяются, и клетка делится.
Межфазный
Во время интерфазы клетка претерпевает нормальные процессы роста, а также готовится к клеточному делению. Чтобы клетка перешла из интерфазы в митотическую фазу, должны быть выполнены многие внутренние и внешние условия.Три этапа межфазного взаимодействия называются G 1 , S и G 2 .
Чтобы клетка перешла из интерфазы в митотическую фазу, должны быть выполнены многие внутренние и внешние условия.Три этапа межфазного взаимодействия называются G 1 , S и G 2 .
Видео \ (\ PageIndex {1} \): Interphase | Клетки | MCAT | Хан Академия
G
1 Фаза (первый разрыв)Первая стадия межфазной границы называется фазой G 1 (первый промежуток), потому что с микроскопической точки зрения заметны небольшие изменения. Однако на стадии G 1 клетка достаточно активна на биохимическом уровне. Клетка накапливает строительные блоки хромосомной ДНК и связанных белков, а также накапливает достаточные запасы энергии для выполнения задачи репликации каждой хромосомы в ядре.
S-фаза (синтез ДНК)
На протяжении всей интерфазы ядерная ДНК остается в полуконденсированной конфигурации хроматина. В S-фазе репликация ДНК может происходить с помощью механизмов, которые приводят к образованию идентичных пар молекул ДНК — сестринских хроматид, которые прочно прикрепляются к центромерной области через белки, называемые когезинами.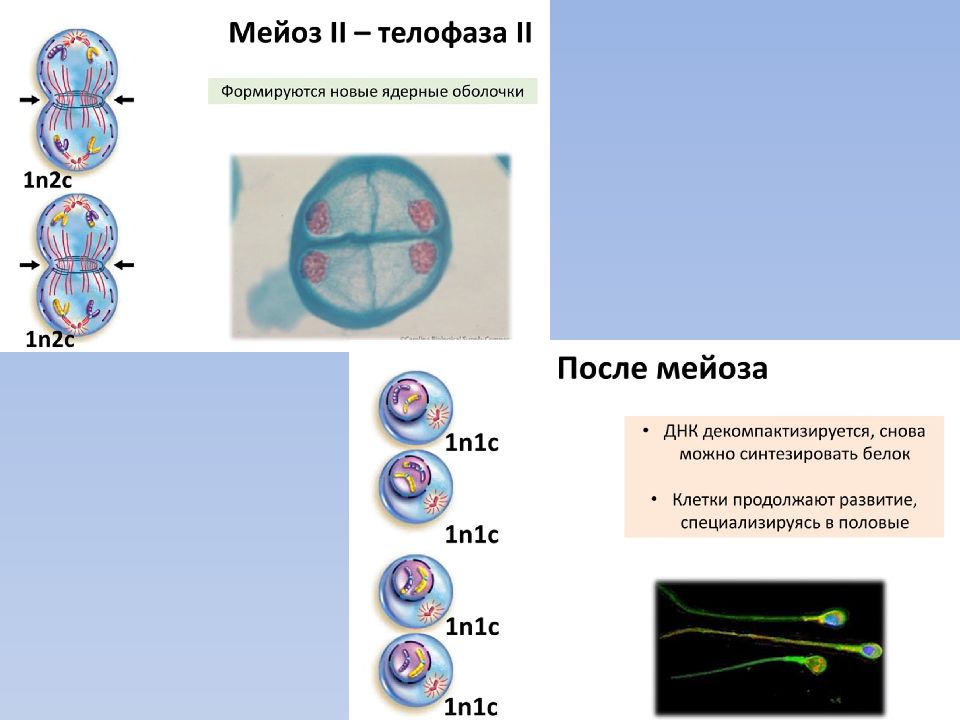
Подсчет хромосом и хроматид
Обратите внимание, что две сестринские хроматиды, которые удерживаются вместе с помощью белков когезина, по-прежнему считаются одной хромосомой, пока они не разделятся во время митоза.
Центросома также дублируется во время S фазы. Две центросомы дадут начало митотическому веретену, аппарату, который управляет движением хромосом во время митоза. В центре каждой животной клетки центросомы животных клеток связаны с парой стержневидных объектов, центриолей, которые расположены под прямым углом друг к другу. Центриоли помогают организовать деление клеток. Центриоли отсутствуют в центросомах других видов эукариот, таких как растения и большинство грибов.
G
2 Фаза (второй разрыв) В фазе G 2 клетка пополняет запасы энергии и синтезирует белки, необходимые для манипуляций с хромосомами. Некоторые клеточные органеллы дублируются, а цитоскелет разбирается, чтобы обеспечить ресурсы для митотической фазы. Во время G 2 может происходить дополнительный рост клеток. Последние приготовления к митотической фазе должны быть завершены до того, как клетка сможет вступить в первую стадию митоза.
Во время G 2 может происходить дополнительный рост клеток. Последние приготовления к митотической фазе должны быть завершены до того, как клетка сможет вступить в первую стадию митоза.
Что в пробелах?
Какие конкретные типы белков необходимы клетке во время G 1 ? Во время G 2 ?
Митоз
Митотическая фаза — это многоступенчатый процесс, во время которого дублированные хромосомы выравниваются, разделяются и переходят в две новые идентичные дочерние клетки. Первая часть митотической фазы называется кариокинезом или делением ядра. Вторая часть митотической фазы, называемая цитокинезом, — это физическое разделение цитоплазматических компонентов на две дочерние клетки.
Митоз делится на ряд фаз — профаза, прометафаза, метафаза, анафаза и телофаза, которые приводят к делению ядра клетки. Кариокинез еще называют митозом.
Рисунок \ (\ PageIndex {3} \): Кариокинез (или митоз) делится на пять стадий — профаза, прометафаза, метафаза, анафаза и телофаза. Изображения внизу были сделаны с помощью флуоресцентной микроскопии (отсюда черный фон) клеток, искусственно окрашенных флуоресцентными красителями: синяя флуоресценция указывает на ДНК (хромосомы), а зеленая флуоресценция указывает на микротрубочки (веретенообразный аппарат).(«Рисунки митоза»: модификация работы Марианы Руис Вильярреал; кредитные «микрофотографии»: модификация работы Роя ван Хисбина; кредит «цитокинезная микрофотография»: Центр Уодсворта / Департамент здравоохранения штата Нью-Йорк; данные шкалы от Мэтта Рассел)
Изображения внизу были сделаны с помощью флуоресцентной микроскопии (отсюда черный фон) клеток, искусственно окрашенных флуоресцентными красителями: синяя флуоресценция указывает на ДНК (хромосомы), а зеленая флуоресценция указывает на микротрубочки (веретенообразный аппарат).(«Рисунки митоза»: модификация работы Марианы Руис Вильярреал; кредитные «микрофотографии»: модификация работы Роя ван Хисбина; кредит «цитокинезная микрофотография»: Центр Уодсворта / Департамент здравоохранения штата Нью-Йорк; данные шкалы от Мэтта Рассел)
Видео \ (\ PageIndex {2} \): Настоящий микроскопический митоз (Сообщество медицинских исследований через www.youtube.com/watch?v=L61Gp_d7evo)
Профаза
Во время профазы ядерная оболочка начинает диссоциировать на маленькие пузырьки, а мембранные органеллы (такие как комплекс Гольджи или аппарат Гольджи и эндоплазматический ретикулум) фрагментируются и рассеиваются к периферии клетки.Ядрышко исчезает (разойдется).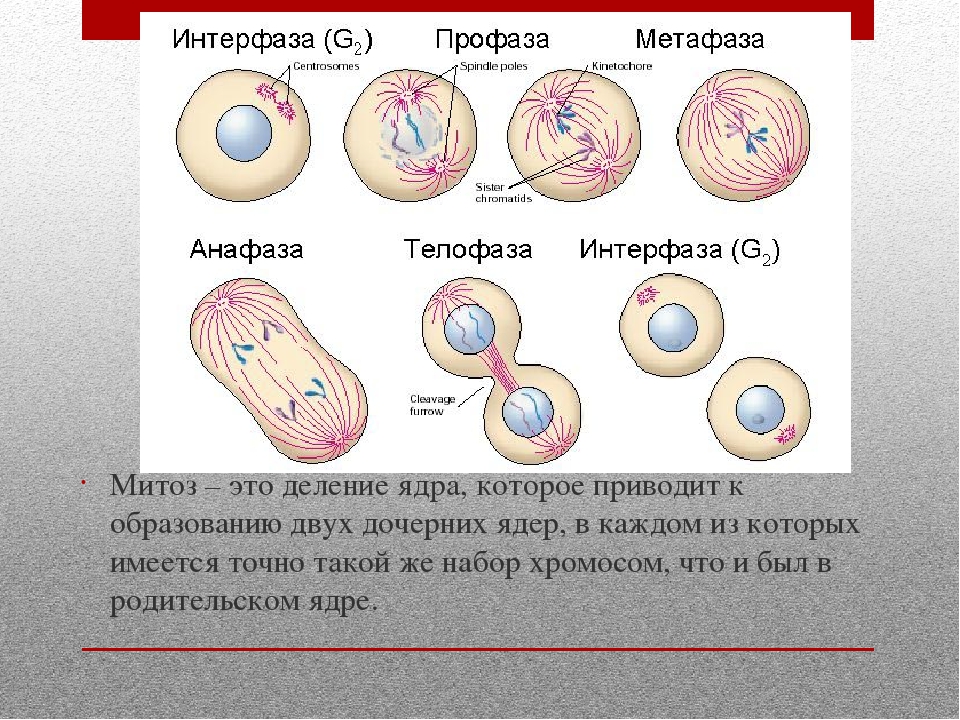 Центросомы начинают двигаться к противоположным полюсам клетки. Микротрубочки, которые образуют митотическое веретено, проходят между центросомами, раздвигая их дальше друг от друга по мере удлинения волокон микротрубочек. Сестринские хроматиды начинают более плотно сплетаться с помощью белков конденсина и становятся видимыми под световым микроскопом.
Центросомы начинают двигаться к противоположным полюсам клетки. Микротрубочки, которые образуют митотическое веретено, проходят между центросомами, раздвигая их дальше друг от друга по мере удлинения волокон микротрубочек. Сестринские хроматиды начинают более плотно сплетаться с помощью белков конденсина и становятся видимыми под световым микроскопом.
Прометафаза и метафаза
Во время прометафазы многие процессы, которые были начаты в профазе, продолжают развиваться.Остатки фрагмента ядерной оболочки. Митотическое веретено продолжает развиваться, поскольку все больше микротрубочек собираются и растягиваются по длине бывшей ядерной области. Хромосомы становятся более конденсированными и дискретными. Каждая сестринская хроматида в центромерной области развивает белковую структуру, называемую кинетохорой. Белки кинетохоры привлекают и связывают микротрубочки митотического веретена. Поскольку микротрубочки веретена отходят от центросом, некоторые из этих микротрубочек входят в контакт и прочно связываются с кинетохорами. Как только митотическое волокно прикрепляется к хромосоме, хромосома будет ориентирована до тех пор, пока кинетохоры сестринских хроматид не окажутся напротив противоположных полюсов. В конце концов, все сестринские хроматиды будут прикреплены своими кинетохорами к микротрубочкам с противоположных полюсов. Микротрубочки веретена, которые не взаимодействуют с хромосомами, называются полярными микротрубочками. Эти микротрубочки перекрывают друг друга на полпути между двумя полюсами и способствуют удлинению клеток. Астральные микротрубочки расположены рядом с полюсами, помогают в ориентации веретена и необходимы для регуляции митоза.
Как только митотическое волокно прикрепляется к хромосоме, хромосома будет ориентирована до тех пор, пока кинетохоры сестринских хроматид не окажутся напротив противоположных полюсов. В конце концов, все сестринские хроматиды будут прикреплены своими кинетохорами к микротрубочкам с противоположных полюсов. Микротрубочки веретена, которые не взаимодействуют с хромосомами, называются полярными микротрубочками. Эти микротрубочки перекрывают друг друга на полпути между двумя полюсами и способствуют удлинению клеток. Астральные микротрубочки расположены рядом с полюсами, помогают в ориентации веретена и необходимы для регуляции митоза.
Во время метафазы все хромосомы выровнены в плоскости, называемой метафазной пластиной, или экваториальной плоскостью, посередине между двумя полюсами клетки. Сестринские хроматиды все еще плотно связаны друг с другом с помощью белков когезина.В это время хромосомы максимально уплотнены.
Сестринские хроматиды все еще плотно связаны друг с другом с помощью белков когезина.В это время хромосомы максимально уплотнены.
Анафаза
Во время анафазы белки когезина разрушаются, и сестринские хроматиды разделяются на центромере. Каждая хроматида, теперь называемая хромосомой, быстро тянется к центросоме, к которой прикреплена ее микротрубочка. Клетка становится заметно удлиненной (овальной формы), когда полярные микротрубочки скользят друг относительно друга в метафазной пластинке, где они перекрываются.
Телофаза
Во время телофазы хромосомы достигают противоположных полюсов и начинают деконденсироваться (распадаться), расслабляясь в конфигурацию хроматина.Митотические веретена деполимеризуются в мономеры тубулина, которые будут использоваться для сборки компонентов цитоскелета для каждой дочерней клетки. Ядерные оболочки образуются вокруг хромосом, а нуклеосомы появляются в ядерной области.
Упражнение \ (\ PageIndex {1} \)
Что из следующего является правильным порядком событий в митозе?
- Сестринские хроматиды выстраиваются на метафазной пластинке.
 Кинетохора прикрепляется к митотическому веретену.Ядро реформируется, и клетка делится. Белки когезина распадаются, и сестринские хроматиды отделяются.
Кинетохора прикрепляется к митотическому веретену.Ядро реформируется, и клетка делится. Белки когезина распадаются, и сестринские хроматиды отделяются. - Кинетохора прикрепляется к митотическому веретену. Белки когезина распадаются, и сестринские хроматиды отделяются. Сестринские хроматиды выстраиваются у метафазной пластинки. Ядро реформируется, и клетка делится.
- Кинетохора прикрепляется к белкам когезина. Сестринские хроматиды выстраиваются у метафазной пластинки. Кинетохора разрушается, и сестринские хроматиды отделяются.Ядро реформируется, и клетка делится.
- Кинетохора прикрепляется к митотическому веретену. Сестринские хроматиды выстраиваются у метафазной пластинки. Белки когезина распадаются, и сестринские хроматиды отделяются. Ядро реформируется, и клетка делится.
- Ответ
-
4
Упражнение \ (\ PageIndex {2} \)
Химиотерапевтические препараты, такие как винкристин и колхицин, нарушают митоз, связываясь с белком тубулином (субъединицей микротрубочек) и препятствуя сборке и разборке микротрубочек. На какую именно митотическую структуру воздействуют эти препараты и какое влияние это окажет на деление клеток?
На какую именно митотическую структуру воздействуют эти препараты и какое влияние это окажет на деление клеток?
- Ответ
-
Митотическое веретено образовано микротрубочками. Микротрубочки — это полимеры протеина тубулина; следовательно, эти препараты разрушают митотическое веретено. Без функционального митотического веретена хромосомы не будут отсортированы или разделены во время митоза. Клетка остановится в митозе и умрет.
Цитокинез
Цитокинез, или «движение клетки», — это вторая основная стадия митотической фазы, во время которой завершается деление клетки за счет физического разделения цитоплазматических компонентов на две дочерние клетки.Деление не завершается до тех пор, пока компоненты клетки не будут распределены и полностью разделены на две дочерние клетки. Хотя стадии митоза у большинства эукариот схожи, процесс цитокинеза у эукариот, имеющих клеточные стенки, таких как клетки растений, совершенно иной.
В таких клетках, как клетки животных, у которых отсутствуют клеточные стенки, цитокинез следует за наступлением анафазы. Сократительное кольцо, состоящее из актиновых филаментов, образуется внутри плазматической мембраны на бывшей метафазной пластинке.Нити актина притягивают экватор клетки внутрь, образуя щель. Эта трещина или «трещина» называется бороздой дробления. По мере того как актиновое кольцо сжимается, борозда углубляется, и в конечном итоге мембрана расщепляется, чтобы разделить две новые клетки.
В клетках растений между дочерними клетками должна образоваться новая клеточная стенка. Во время интерфазы аппарат Гольджи накапливает ферменты, структурные белки и молекулы глюкозы, прежде чем они разбиваются на везикулы и распределяются по делящейся клетке.Во время телофазы эти везикулы Гольджи транспортируются по микротрубочкам с образованием фрагмопласта (везикулярной структуры) на метафазной пластинке. Там пузырьки сливаются и сливаются от центра к стенкам клетки; эта структура называется клеточной пластиной. По мере слияния большего количества пузырьков клеточная пластинка увеличивается, пока не сливается с клеточными стенками на периферии клетки. Ферменты используют глюкозу, скопившуюся между слоями мембраны, для создания новой клеточной стенки. Мембраны Гольджи становятся частями плазматической мембраны по обе стороны от новой клеточной стенки.
По мере слияния большего количества пузырьков клеточная пластинка увеличивается, пока не сливается с клеточными стенками на периферии клетки. Ферменты используют глюкозу, скопившуюся между слоями мембраны, для создания новой клеточной стенки. Мембраны Гольджи становятся частями плазматической мембраны по обе стороны от новой клеточной стенки.
Упражнение \ (\ PageIndex {3} \)
Рассмотрим события интерфазы и митоза. Какие события клеточного цикла будут затронуты в клетке, продуцирующей мутировавшие (нефункциональные) белки конденсина?
- Ответ
-
Если конденсин не функционирует, хромосомы не упаковываются после репликации ДНК в S-фазе интерфазы.
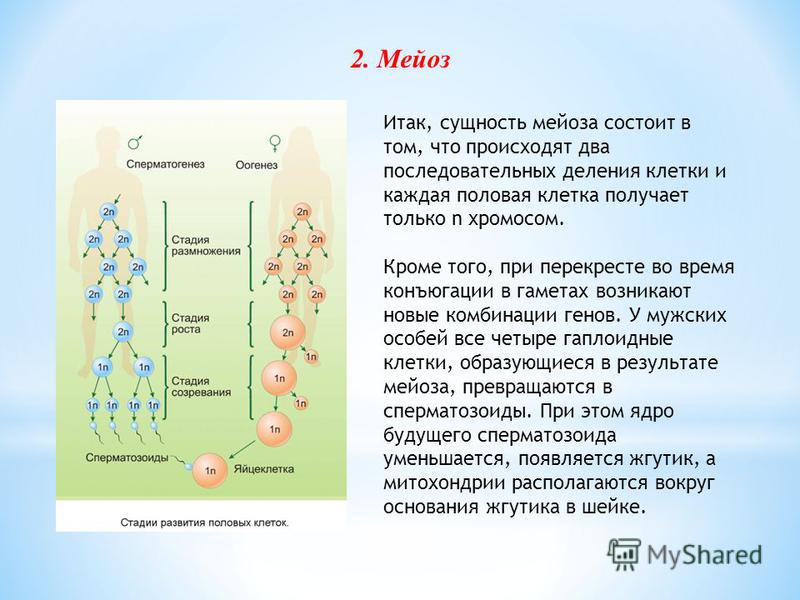 Вероятно, что белки центромерной области, такой как кинетохора, не будут формироваться. Даже если бы волокна митотического веретена могли прикрепляться к хроматидам без упаковки, хромосомы не были бы отсортированы или разделены во время митоза.
Вероятно, что белки центромерной области, такой как кинетохора, не будут формироваться. Даже если бы волокна митотического веретена могли прикрепляться к хроматидам без упаковки, хромосомы не были бы отсортированы или разделены во время митоза.
Авторы и авторство
-
Конни Рай (Общественный колледж Восточного Миссисипи), Роберт Уайз (Университет Висконсина, Ошкош), Владимир Юруковский (Общественный колледж округа Саффолк), Жан ДеСе (Университет Северной Каролины в Чапел-Хилл), Джунг Чой (Институт Джорджии Technology), Яэль Ависсар (Колледж Род-Айленда) среди других авторов. Исходный контент от OpenStax (CC BY 4.0; Скачать бесплатно по адресу http: // cnx.org / contents / 185cbf87-c72 … [email protected]).
Как «посчитать» хромосомы
После того, как репликация ДНК копирует хромосомы во время S фазы, сестринские хроматиды остаются скрепленными белками когезина. Эти сестры по-прежнему считаются одной хромосомой, пока они не отделяются во время анафазы митоза, что гарантирует, что каждая из двух новых клеток унаследует одну копию (сестринскую хроматиду) каждой хромосомы.
Видео \ (\ PageIndex {3} \): отслеживание движений хромосом в митозе
Пример \ (\ PageIndex {1} \)
Если в клетке 10 хромосом (всего), сколько хромосом будет присутствовать в метафазе митоза? В анафазе? В двух дочерних клетках?
Решение
10 хромосом (каждая состоит из двух сестринских хроматид, потому что репликация ДНК произошла в S-фазе перед митозом) присутствуют во время метафазы.
20 хромосом присутствуют во время анафазы, потому что сестринские хроматиды были разделены.
После телофазы и цитокинеза каждая новая дочерняя клетка будет иметь по 10 хромосом, что идентично родительской клетке.
Упражнение \ (\ PageIndex {4} \)
Бурые медведи ( Ursus arctos) имеют 74 (2n = 74) хромосомы на диплоидную клетку. Сколько хромосом и сестринских хроматид присутствует на каждой из следующих стадий: G 1 , G 2 , анафаза митоза и после цитокинеза?
- Ответ
-
# Хромосомы # Хроматиды G 1 74 74 G 2 74 148 Анафаза митоза 148 148 Цитокинез 74 74
Упражнение \ (\ PageIndex {5} \)
Клетки человека имеют 23 пары хромосом.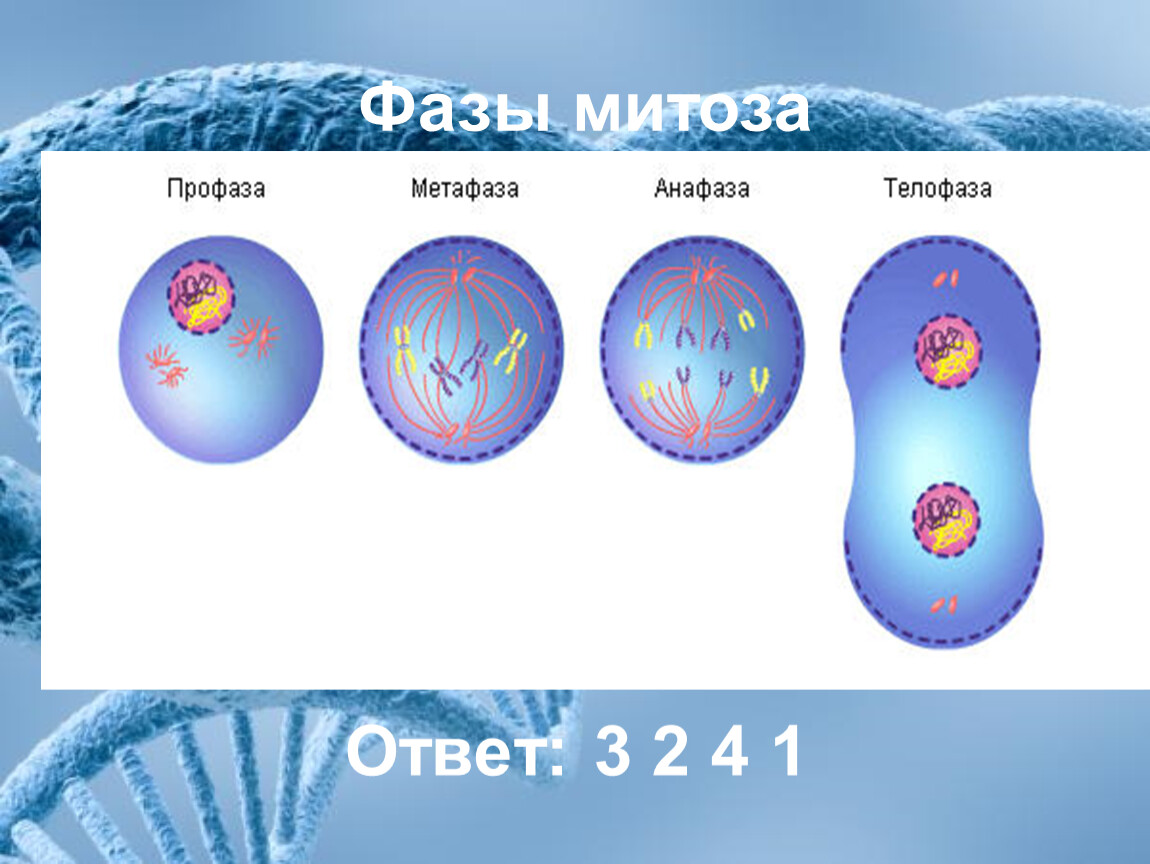 Сколько хромосом присутствует в метафазе? В анафазе? В клетках, завершивших митоз?
Сколько хромосом присутствует в метафазе? В анафазе? В клетках, завершивших митоз?
- Ответ
-
Метафаза: 46 хромосом
Анафаза: 92 хромосомы
После митоза: Каждая клетка имеет 46 хромосом.
Мейоз
Большинство клеток растений и животных диплоидны. Термин диплоид происходит от греческого слова diplos, означает «двойной» или «два»; термин подразумевает, что клетки растений и животных имеют пары хромосом.Например, в клетках человека 46 хромосом организованы в 23 пары. Следовательно, человеческие клетки диплоидны в том смысле, что они имеют пару из 23 отдельных хромосом.
Во время полового размножения половые клетки родительских организмов соединяются друг с другом и образуют оплодотворенную яйцеклетку (зиготу). В этой ситуации каждая половая клетка представляет собой гамету . Гаметы человеческих клеток — гаплоидов, от греческого гаплос, означает «одиночный». Этот термин означает, что каждая гамета содержит половину из 46 хромосом — 23 хромосомы у человека.Когда человеческие гаметы соединяются друг с другом, восстанавливается первоначальное диплоидное состояние 46 хромосом. Затем митоз вызывает развитие диплоидной клетки в многоклеточный организм.
Процесс, посредством которого число хромосом уменьшается вдвое во время образования гамет, составляет мейоз. При мейозе клетка, содержащая диплоидное число хромосом, превращается в четыре клетки, каждая из которых имеет гаплоидное число хромосом. Например, в клетках человека, подвергающихся мейозу, клетка, содержащая 46 хромосом, дает четыре клетки, каждая с 23 хромосомами.
Мейоз происходит в несколько этапов, которые напоминают этапы митоза. Происходят две основные фазы мейоза: мейоз I и мейоз II. Во время мейоза I одна клетка делится на две. Во время мейоза II каждая из этих двух клеток снова делится. Те же самые разграничивающие фазы митоза имеют место в мейозе I и мейозе II — профаза, метафаза, анафаза и телофаза, но с некоторыми содержащимися в них вариациями.
Во время мейоза II каждая из этих двух клеток снова делится. Те же самые разграничивающие фазы митоза имеют место в мейозе I и мейозе II — профаза, метафаза, анафаза и телофаза, но с некоторыми содержащимися в них вариациями.
Как показано на рис. 8-1, сначала хромосомы клетки делятся на две клетки.Затем хромосомы двух клеток разделяются и переходят в четыре дочерние клетки. Родительская клетка диплоидна, а каждая из дочерних клеток имеет один набор хромосом и является гаплоидной. Синапсис и кроссинговер происходят на стадии I профазы.
Рисунок 8-1 Процесс мейоза, в котором образуются четыре гаплоидных клетки.
Члены каждой пары хромосом в клетке называются гомологичными хромосомами. Гомологичные хромосомы похожи, но не идентичны.Они могут нести разные версии одной и той же генетической информации. Например, одна гомологичная хромосома может нести информацию о светлых волосах, а другая гомологичная хромосома может нести информацию о черных волосах.
Когда клетка готовится к вступлению в мейоз, каждая из ее хромосом дублируется на стадии синтеза (S) клеточного цикла, как в митозе. Таким образом, каждая хромосома состоит из двух сестринских хроматид.
Мейоз I
В начале мейоза I клетка человека содержит 46 хромосом или 92 хроматиды (столько же, сколько во время митоза).Мейоз I проходит следующие фазы:
■ Профаза I: Профаза I в некоторых отношениях похожа на профазу в митозе. Хроматиды укорачиваются, утолщаются и становятся видимыми под микроскопом. Однако важное отличие состоит в том, что происходит процесс, называемый синапсис . Синапсис — это когда гомологичные хромосомы мигрируют друг к другу и соединяются, образуя тетрад (комбинация четырех хроматид, по две от каждой гомологичной хромосомы).Второй процесс, названный пересечением , также происходит во время профазы I. В этом процессе сегменты ДНК от одной хроматиды в тетраде переходят на другую хроматиду в тетраде.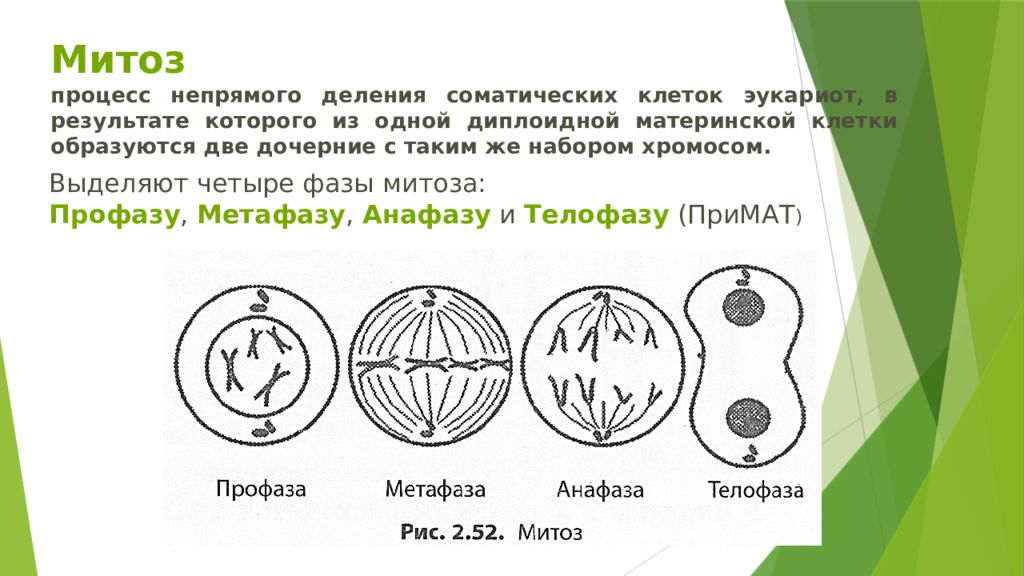 Эти обмены хромосомными сегментами происходят сложным и малоизученным образом. В результате получается генетически новая хроматида. Кроссовер — важная движущая сила эволюции. После того, как произошел кроссинговер, гомологичная пара хромосом отличается генетически.
Эти обмены хромосомными сегментами происходят сложным и малоизученным образом. В результате получается генетически новая хроматида. Кроссовер — важная движущая сила эволюции. После того, как произошел кроссинговер, гомологичная пара хромосом отличается генетически.
■ Метафаза I: В метафазе I мейоза тетрады выстраиваются на экваториальной пластине (как в митозе).Центромеры прикрепляются к волокнам веретена, которые отходят от полюсов клетки. К каждому волокну веретена прикрепляется одна центромера.
■ Анафаза I: В анафазе I гомологичные хромосомы или тетрады разделяются. Одна гомологичная хромосома (состоящая из двух хроматид) перемещается на одну сторону клетки, а другая гомологичная хромосома (состоящая из двух хроматид) перемещается на другую сторону клетки. В результате 23 хромосомы (каждая из которых состоит из двух хроматид) перемещаются на один полюс, а 23 хромосомы (каждая из которых состоит из двух хроматид) перемещается на другой полюс.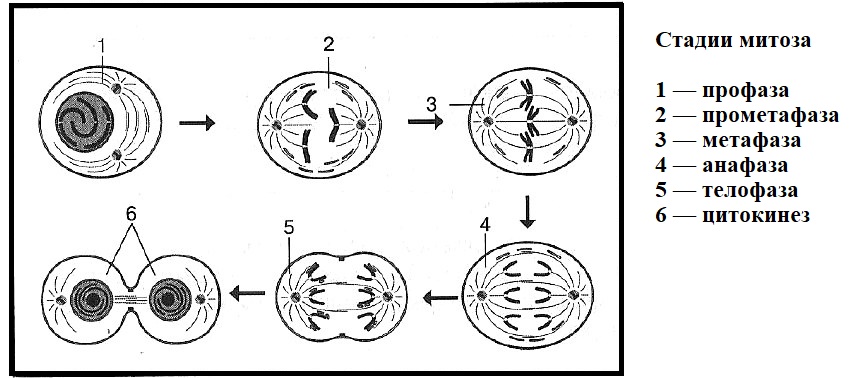 По сути, число хромосом клетки уменьшается вдвое после завершения мейоза I. По этой причине процесс является редукционно-делительным.
По сути, число хромосом клетки уменьшается вдвое после завершения мейоза I. По этой причине процесс является редукционно-делительным.
■ Телофаза I: В телофазе I мейоза ядро реорганизуется, хромосомы превращаются в хроматин, и клеточная мембрана начинает защемляться внутрь. Цитокинез происходит сразу после телофазы I. Этот процесс происходит по-разному в клетках растений и животных, как и в митозе.
Мейоз II
Мейоз II является вторым по значимости подразделением мейоза.Это происходит практически так же, как митоз. В мейозе II клетка содержит единственный набор хромосом. Однако к каждой хромосоме по-прежнему прикреплена двойная сестринская хроматида. Мейоз II разделяет сестринские хроматиды на отдельные клетки. Мейоз II проходит следующие фазы:
■ Профаза II: Профаза II подобна профазе митоза. Материал хроматина конденсируется, и каждая хромосома содержит две хроматиды, прикрепленные центромерой.Затем 23 пары хроматид, всего 46 хроматид, перемещаются к экваториальной пластине.
■ Метафаза II: В метафазе II мейоза 23 пары хроматид собираются в центре клетки до разделения. Этот процесс идентичен метафазе в митозе, за исключением того, что он происходит в гаплоидной, а не в диплоидной клетке.
■ Анафаза II: Во время анафазы II мейоза центромеры делятся, и сестринские хроматиды разделяются, и в это время их называют нереплицируемыми хромосомами.Волокна веретена перемещают хромосомы к каждому полюсу. Всего на каждый полюс движутся 23 хромосомы. Силы и связи, которые действуют в митозе, также действуют в анафазе II.
■ Телофаза II: Во время телофазы II хромосомы собираются на полюсах клеток и становятся нечеткими. Опять же, они образуют массу хроматина. Ядерная оболочка развивается, ядрышки появляются снова, и клетки подвергаются цитокинезу.
Во время мейоза II каждая клетка, содержащая 46 хроматид, дает две клетки, каждая с 23 хромосомами.Первоначально было две клетки, подвергшиеся мейозу II; Следовательно, результатом мейоза II являются четыре клетки, каждая с 23 хромосомами. Каждая из четырех клеток гаплоидна; то есть каждая клетка содержит единственный набор хромосом.
Каждая из четырех клеток гаплоидна; то есть каждая клетка содержит единственный набор хромосом.
23 хромосомы в четырех клетках мейоза не идентичны, потому что кроссинговер произошел в профазе I. Кроссинговер дает генетические вариации, так что каждая из четырех клеток, полученных в результате мейоза, отличается от трех других. Таким образом, мейоз обеспечивает механизм для создания вариаций хромосом.Кроме того, он объясняет образование четырех гаплоидных клеток из одной диплоидной клетки.
Митоз | Изучайте науку в Scitable
Когда прометафаза заканчивается и начинается метафаза , хромосомы выравниваются вдоль экватора клетки. Каждая хромосома имеет по крайней мере две микротрубочки, отходящие от ее кинетохоры — по крайней мере, одна микротрубочка, соединенная с каждым полюсом. В этот момент напряжение внутри клетки уравновешивается, и хромосомы больше не двигаются вперед и назад.Кроме того, теперь веретено завершено, и видны три группы микротрубочек веретена. Микротрубочки кинетохор прикрепляют хромосомы к полюсу веретена; Межполюсные микротрубочки проходят от полюса шпинделя через экватор, почти до противоположного полюса шпинделя; и астральных микротрубочек, простираются от полюса веретена к клеточной мембране.
Микротрубочки кинетохор прикрепляют хромосомы к полюсу веретена; Межполюсные микротрубочки проходят от полюса шпинделя через экватор, почти до противоположного полюса шпинделя; и астральных микротрубочек, простираются от полюса веретена к клеточной мембране.
Метафаза приводит к анафазе , во время которой сестринские хроматиды каждой хромосомы отделяются и перемещаются к противоположным полюсам клетки.Ферментативный распад когезина, который связывает сестринские хроматиды вместе во время профазы, вызывает это разделение. После разделения каждая хроматида становится независимой хромосомой. Между тем, изменения длины микротрубочек обеспечивают механизм движения хромосом. Более конкретно, в первой части анафазы — иногда называемой анафазой A — микротрубочки кинетохор укорачиваются и притягивают хромосомы к полюсам веретена. Затем, во второй части анафазы — иногда называемой анафазой B — астральные микротрубочки, прикрепленные к клеточной мембране, раздвигают полюса дальше друг от друга, и межполюсные микротрубочки скользят друг мимо друга, оказывая дополнительное притяжение на хромосомы (Рисунок 2). .
.
Рисунок 2: Типы микротрубочек, участвующих в митозе
Во время митоза несколько типов микротрубочек являются активными. Моторные белки, связанные с межполярными микротрубочками, управляют сборкой веретена. Обратите внимание на другие типы микротрубочек, участвующих в закреплении полюса веретена и разъединении сестринских хроматид.
Life Sciences Cyberbridge
Life Sciences CyberbridgeMEIOSIS I
Мейоз — это процесс, при котором реплицированные хромосомы подвергаются двум ядерным делениям с образованием четырех гаплоидных клеток, также называемых мейоцитами (сперматозоиды и яйца).Диплоидные (2 n ) организмы полагаются на мейоз для производства мейоцитов, которые имеют половину плоидности родительских для полового размножения. Уменьшение наполовину плоидности мейоцитов необходимо для восстановления генетического содержания зиготы до родительского. Мейоз использует механизмы, аналогичные тем, которые используются во время митоза, для разделения и перераспределения хромосом. Однако некоторые особенности, а именно спаривание и генетическая рекомбинация между гомологичными хромосомами, являются уникальными для мейоза.
Однако некоторые особенности, а именно спаривание и генетическая рекомбинация между гомологичными хромосомами, являются уникальными для мейоза.Шаги, ведущие к мейозу, аналогичны этапам митоза — центриоли и хромосомы реплицируются. Количество ДНК в клетке увеличилось вдвое, а плоидность клетки осталась такой же, как и раньше, 2 n . В мейозе I фазы аналогичны митозу: профаза I, метафаза I, анафаза I и телофаза I (рисунок ниже). Мейоз I переходит непосредственно в мейоз II, не проходя через интерфазу.
Мейоз I уникален тем, что генетическое разнообразие генерируется посредством кроссинговера и случайного расположения гомологичных хромосом (двухвалентных хромосом).Кроме того, в мейозе I количество хромосом уменьшается с диплоидного (2 n ) до гаплоидного ( n ) во время этого процесса. (См. Рисунок ниже, где мейоз I начинается с диплоидной (2 n = 4) клетки и заканчивается двумя гаплоидными ( n = 2) клетками). У людей (2 n = 46), у которых 23 пары хромосом, количество хромосом уменьшается вдвое в конце мейоза I ( n = 23).
Профаза I
Во время профазы I конденсация хромосом позволяет рассматривать хромосомы под микроскопом.В поздней профазе I гомологичные хромосомы (также называемые двухвалентными хромосомами или бивалентами) спариваются латерально или бок о бок. В это время говорят, что они находятся в синапсисе. Во время синапсов кроссоверы — перекрестные связи, которые образуются в результате разрыва и воссоединения сестринских хроматид — могут происходить между парными бивалентами, что приводит к генетической рекомбинации (обмену генетическим материалом) между вовлеченными цепями. Точка, где происходит кроссовер, называется хиазмой (множественное число хиазм) (см. Рисунок ниже).На рисунке ниже после кроссинговера синяя и красная хромосомы, которые первоначально несли AA и aa аллелей, соответственно, теперь несут аллелей Aa в обеих хромосомах в конце профазы I. Обратите внимание, что эти биваленты имеют два хромосомы и четыре хроматиды, по одной хромосоме от каждого родителя.
Метафаза I
В метафазе I каждая пара бивалентов (две хромосомы, всего четыре хроматиды) выравнивается на метафазной пластине.Это отличается от метафазы в митозе, когда все хромосомы выстраиваются в один файл на метафазной пластине. Положение каждой хромосомы в бивалентах случайное — любой родительский гомолог может появляться с каждой стороны. Это означает, что у дочерних клеток есть 50-50 шансов получить гомолог матери или отца для каждой хромосомы (см. Рисунок ниже). Как показано на рисунке ниже, во время метафазы I биваленты любого родителя могут выравниваться по обе стороны от ячейки. В организме с двумя наборами хромосом существует четыре способа организации хромосом, что приводит к различиям в хромосомном распределении в дочерних клетках после мейоза I.(Диплоидный организм с 2 n хромосом будет иметь 2 n возможных комбинаций или способов организации своих хромосом во время метафазы I.)
В диплоидной клетке с 2 парами хромосом существует 4 способа расположения хромосом в метафазе I.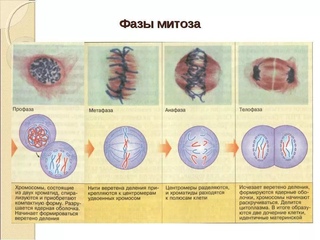
Анафаза I
В анафазе I гомологичные хромосомы разделяются. Гомологичные хромосомы, каждая из которых содержит две хроматиды, перемещаются на разные полюса. В отличие от митоза центромеры не расщепляются, и сестринские хроматиды остаются спаренными в анафазе I.
Телофаза I и цитокинез
В телофазе I гомологи каждого бивалента достигают противоположных полюсов клетки, и вокруг каждого набора хромосом формируется новая ядерная мембрана. Затем цитокинез делит клетку на две дочерние клетки. Каждая из двух дочерних клеток теперь гаплоидная ( n ), с половиной количества хромосом на ядро, как в мейозе I. У некоторых видов ядерная мембрана на короткое время образуется вокруг хромосом, а у других — нет.Теперь клетка переходит в мейоз II, при этом хромосомы остаются конденсированными.
Life Sciences Cyberbridge
Life Sciences CyberbridgeМейоз II
Хромосомная репликация не происходит между мейозом I и мейозом II; мейоз I переходит непосредственно в мейоз II, не проходя через интерфазу. Вторая часть мейоза, мейоз II, больше напоминает митоз, чем мейоз I. Хромосомные числа, которые уже были уменьшены до гаплоидных ( n ) к концу мейоза I, остаются неизменными после этого деления.В мейозе II фазы, опять же, аналогичны митозу: профаза II, метафаза II, анафаза II и телофаза II (см. Рисунок ниже). Как показано на рисунке ниже, мейоз II начинается с двух гаплоидных ( n = 2) клеток и заканчивается четырьмя гаплоидными ( n = 2) клетками. Обратите внимание, что эти четыре мейоцита генетически отличаются друг от друга. У человека (2 n = 46), имеющего 23 пары хромосом, количество хромосом остается неизменным от начала до конца мейоза II ( n = 23).
Вторая часть мейоза, мейоз II, больше напоминает митоз, чем мейоз I. Хромосомные числа, которые уже были уменьшены до гаплоидных ( n ) к концу мейоза I, остаются неизменными после этого деления.В мейозе II фазы, опять же, аналогичны митозу: профаза II, метафаза II, анафаза II и телофаза II (см. Рисунок ниже). Как показано на рисунке ниже, мейоз II начинается с двух гаплоидных ( n = 2) клеток и заканчивается четырьмя гаплоидными ( n = 2) клетками. Обратите внимание, что эти четыре мейоцита генетически отличаются друг от друга. У человека (2 n = 46), имеющего 23 пары хромосом, количество хромосом остается неизменным от начала до конца мейоза II ( n = 23).
Профаза II
Волокна веретена реформируются и прикрепляются к центромерам в профазе II.
Метафаза II
Хромосомы выравниваются на метафазной пластинке во время метафазы II, готовясь к делению центромер в следующей фазе.
Анафаза II
В анафазе II хромосомы делятся в центромерах (как в митозе), и полученные хромосомы, каждая с одной хроматидой, перемещаются к противоположным полюсам клетки.
Телофаза II и цитокинез
Четыре гаплоидных ядра (содержащие хромосомы с одиночными хроматидами) образуются в телофазе II. Деление цитоплазмы во время цитокинеза приводит к образованию четырех гаплоидных клеток. Обратите внимание, что эти четыре клетки не идентичны, поскольку случайное расположение бивалентов и кроссинговер в мейозе I приводит к разному генетическому составу этих клеток.
У человека мейоз производит генетически различные гаплоидные дочерние клетки, каждая из которых имеет 23 хромосомы, состоящие из одной хроматиды.Эти гаплоидные клетки становятся неоплодотворенными яйцеклетками у женщин и сперматозоидами у мужчин. Генетические различия гарантируют, что братья и сестры одних и тех же родителей никогда не будут полностью генетически идентичными.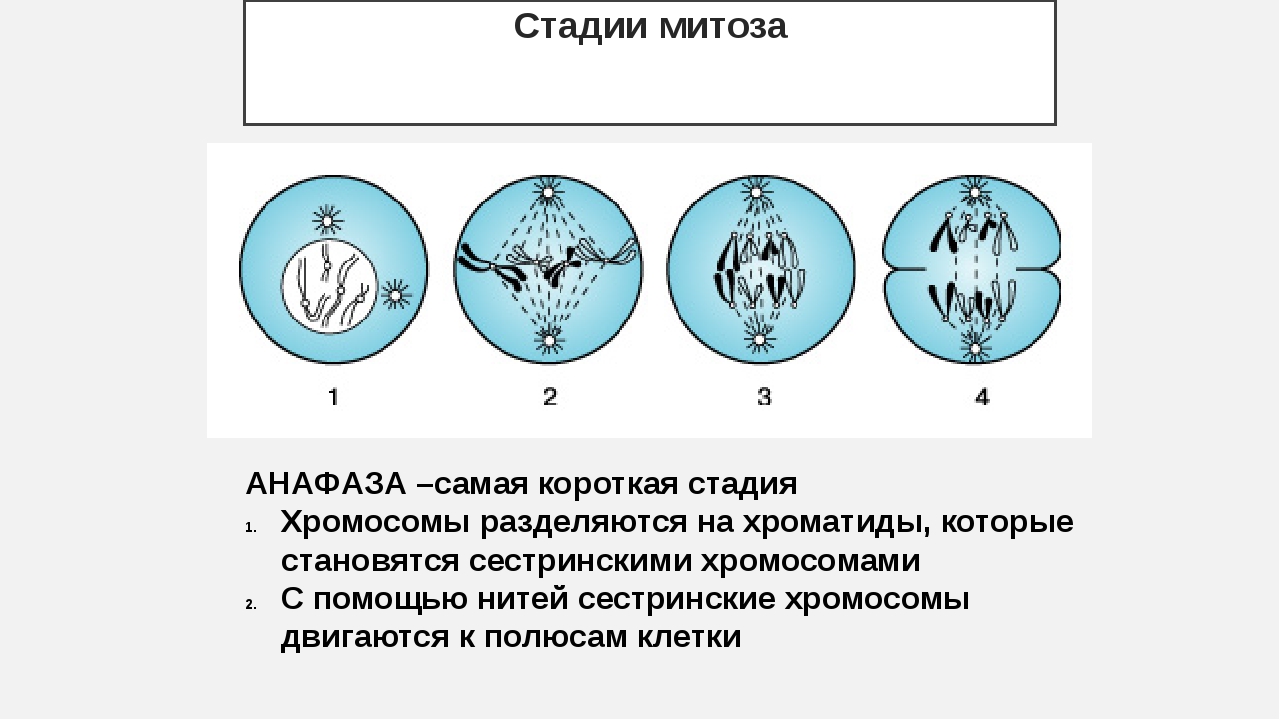
Генетика, нерасхождение — StatPearls — Книжная полка NCBI
Введение
Человеческое тело состоит из триллионов соматических клеток, способных делиться на идентичные дочерние клетки, способствуя росту, восстановлению и реакции организма на изменяющуюся среду.Этот процесс называется «митозом». При производстве гамет происходит другая форма деления клеток, называемая «мейозом». Результатом мейоза является создание четырех дочерних клеток, либо сперматозоидов, либо яйцеклеток, путем редукционного деления, в результате чего образуется гаплоидный набор хромосом в каждой гамете. При оплодотворении гаплоидное ядро сперматозоидов сливается с гаплоидным ядром яйцеклетки, что восстанавливает диплоидный хромосомный набор и подтверждает образование зиготы. Во время анафазы клеточного цикла хромосомы отделяются от противоположных концов клетки, образуя две дочерние клетки.Нерасхождение — это неспособность хромосом разделиться, в результате чего образуются дочерние клетки с ненормальным числом хромосом. [1] [2] [3]
[1] [2] [3]
Клеточный
Геном кодируется химической последовательностью нуклеотидов ДНК в наших клетках. В периоды роста клеток гистоновые белки вокруг ДНК ацетилируются, что снижает взаимодействие между ДНК и гистоновым белком. Эта открытая ДНК называется эухроматином и обеспечивает доступ транскрипционных ферментов к ДНК. Перед периодами клеточного деления гистоновые белки деацетилируются, что позволяет образовывать конденсированную форму ДНК, называемую гетерохроматином.Соматические клетки человека содержат 23 парные хромосомы или 46 полных хромосом. 46 считается «диплоидным» числом (2n), а 23 считается «гаплоидным» числом (1n) или половиной диплоидного числа. «Анеуплоидия» относится к наличию аномального количества хромосом. Моносомия (n-1) — это форма анеуплоидии, характеризующаяся отсутствием одной хромосомы, в результате чего всего 45 хромосом. Трисомия (n + 1) — еще одна форма анеуплоидии, при которой имеется дополнительная хромосома, в результате чего получается 47 хромосом. Каждый тип анеуплоидии можно объяснить нерасхождением во время митоза или мейоза. [4] [5] [6]
Каждый тип анеуплоидии можно объяснить нерасхождением во время митоза или мейоза. [4] [5] [6]
Механизм
В клеточном цикле есть 2 части: интерфаза и митоз / мейоз. Интерфазу можно подразделить на рост 1 (G1), синтез (S) и рост 2 (G2). Во время фазы G клетка растет, производя различные белки, а во время фазы S реплицируется ДНК, так что каждая хромосома включает 2 идентичные сестринские хроматиды.
Митоз состоит из 4 фаз: профаза, метафаза, анафаза и телофаза.В профазе ядерная оболочка разрушается, и хроматин конденсируется. В метафазе хромосомы выстраиваются вдоль метафазной пластинки, а микротрубочки прикрепляются к кинетохорам каждой хромосомы. В анафазе хроматиды разделяются и тянутся микротрубочками к противоположным концам клетки. Наконец, в телофазе снова появляются ядерные оболочки, хромосомы раскручиваются в хроматин, и клетка подвергается цитокинезу, который разбивает клетку на 2 идентичные дочерние клетки.
Мейоз дважды проходит все 4 фазы митоза с измененными механизмами, которые в конечном итоге создают гаплоидные клетки вместо диплоидных. Одна модификация находится в мейозе I. Гомологичные хромосомы разделяются вместо сестринских хроматид, создавая гаплоидные клетки. Именно во время этого процесса мы видим кроссинговер и независимый ассортимент, ведущие к увеличению генетического разнообразия потомства. Мейоз II развивается так же, как митоз, но с гаплоидным числом хромосом, в конечном итоге создавая 4 дочерние клетки, все генетически отличные от исходной клетки.
Одна модификация находится в мейозе I. Гомологичные хромосомы разделяются вместо сестринских хроматид, создавая гаплоидные клетки. Именно во время этого процесса мы видим кроссинговер и независимый ассортимент, ведущие к увеличению генетического разнообразия потомства. Мейоз II развивается так же, как митоз, но с гаплоидным числом хромосом, в конечном итоге создавая 4 дочерние клетки, все генетически отличные от исходной клетки.
Нерасхождение может происходить во время анафазы митоза, мейоза I или мейоза II.Во время анафазы сестринские хроматиды (или гомологичные хромосомы для мейоза I) отделяются и перемещаются к противоположным полюсам клетки, притягиваемые микротрубочками. При нерасхождении не происходит разделения, в результате чего обе сестринские хроматиды или гомологичные хромосомы притягиваются к одному полюсу клетки.
Митотическое нерасхождение может происходить из-за инактивации топоизомеразы II, конденсина или сепаразы. Это приведет к образованию 2 дочерних анеуплоидных клеток, одна с 47 хромосомами (2n + 1), а другая с 45 хромосомами (2n-1).
Нерасхождение в мейозе I происходит, когда тетрады не могут разделиться во время анафазы I. В конце мейоза I будет 2 гаплоидных дочерних клетки, одна с n + 1, а другая с n-1. Обе эти дочерние клетки затем продолжат делиться еще раз в мейозе II, производя 4 дочерние клетки, 2 с n + 1 и 2 с n-1.
Нерасхождение в мейозе II является результатом неспособности сестринских хроматид разделиться во время анафазы II. Поскольку мейоз прошел без ошибок, 2 из 4 дочерних клеток будут иметь нормальный набор из 23 хромосом.Две другие дочерние клетки будут анеуплоидными, одна с n + 1, а другая с n-1.
Тестирование
Внутриутробная диагностика хромосомной анеуплоидии плода может быть сделана путем цитогенетического анализа фетальных клеток, обычно получаемых с помощью амниоцентеза или биопсии ворсин хориона. Хромосомный набор плода анализируется путем выполнения теста кариотипа, подсчета хромосом и анализа под световой микроскопией, при этом ищутся отклонения в числе или структуре хромосом. Существует множество пренатальных скрининговых тестов, которые помогают определить возрастной риск хромосомной анеуплоидии плода посредством анализа различных маркеров или внеклеточной ДНК плода в сыворотке крови матери. [7] [8]
Существует множество пренатальных скрининговых тестов, которые помогают определить возрастной риск хромосомной анеуплоидии плода посредством анализа различных маркеров или внеклеточной ДНК плода в сыворотке крови матери. [7] [8]
При экстракорпоральном оплодотворении (ЭКО) перед имплантацией также можно провести тестирование с помощью преимплантационной генетической диагностики (PGD), диагностики полярных тел (PBD) или биопсии бластомера. ПГД — это метод, используемый для идентификации нормальных эмбрионов, которые будут имплантированы матери, хотя технологически сложный и требует дополнительных затрат по сравнению с пренатальной диагностикой.PBD может обнаруживать анеуплоидии материнского происхождения и относительно быстро выполняется по сравнению с PGD. Наконец, биопсия бластомера может быть получена до имплантации для генетического анализа. Однако биопсия бластомера подвергает развивающийся эмбрион большему риску и поэтому в настоящее время не является рекомендуемым стандартом практики.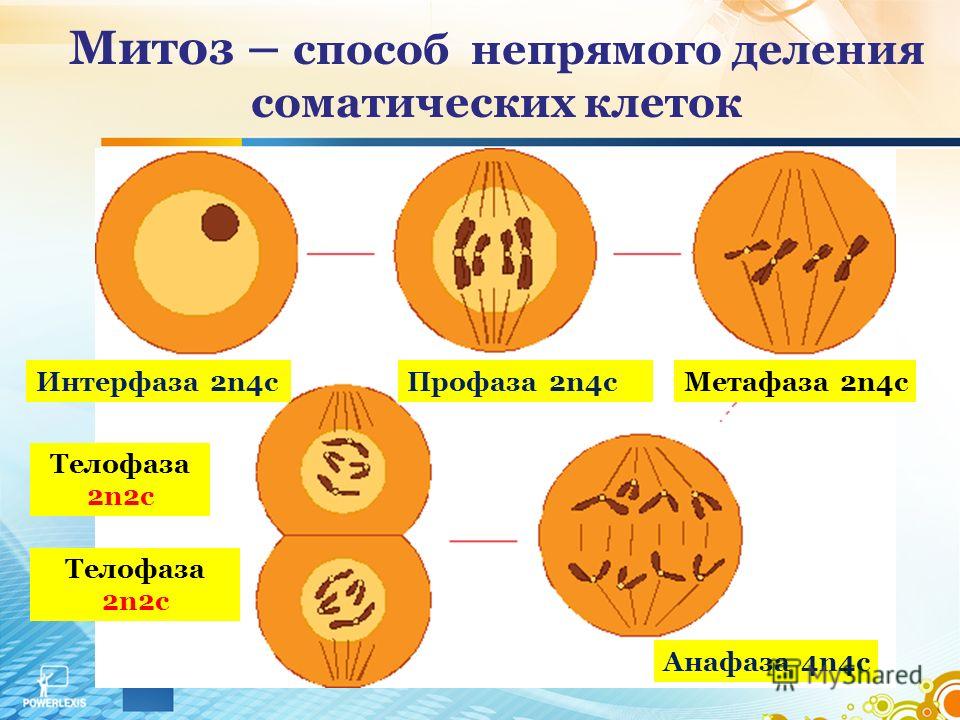
Клиническая значимость
Митотическое нерасхождение может вызывать соматический мозаицизм, при этом хромосомный дисбаланс отражается только в прямом потомстве исходной клетки, где это нерасхождение произошло.Это может вызвать некоторые формы рака, в том числе ретинобластому.
Мейотическое нерасхождение имеет большее клиническое значение, поскольку большинство анеуплоидий несовместимы с жизнью. Тем не менее, некоторые из них дают жизнеспособное потомство со спектром нарушений развития.
Аутосомные трисомии
Синдром Патау: трисомия 13 хромосомы
-
Клинические признаки: качающиеся ступни, микрофтальмия (аномально маленькие глаза), микроцефалия (аномально маленькая голова), полидактилия, голопрозэнцефалия, расщелина губы и неба, врожденный порок сердца и тяжелая форма. Интеллектуальная недееспособность.Средняя продолжительность жизни редко превышает один год.
Синдром Эдвардса: трисомия 18 хромосомы
-
Клинические признаки: качающиеся ступни, низко посаженные уши, микрогнатия (аномально маленькая челюсть), сжатые руки с перекрытием пальцев, врожденный порок сердца и тяжелая умственная отсталость. Ожидаемая продолжительность жизни обычно составляет менее одного года.
Синдром Дауна: трисомия 21 хромосомы
-
Наиболее частая жизнеспособная анеуплоидия.
-
Клинические признаки: Единственная ладонная складка, плоское лицо, выступающие эпикантальные складки, атрезия двенадцатиперстной кишки, врожденный порок сердца, болезнь Гиршпрунга, умственная отсталость. Заметно повышенный риск развития болезни Альцгеймера или лейкемии. Продолжительность жизни около 60 лет.
Трисомии половых хромосом
Синдром Клайнфельтера: лишняя Х-хромосома у мужчины (47, XXY)
-
Клинические признаки: Высокий рост, длинные конечности, гинекомастия, оволосение у женщин, атрофия яичек, задержка развития.
Синдром тройной Х-хромосомы: дополнительная Х-хромосома у женщины (47, XXX)
-
Клинические признаки: Фенотипически нормальные, некоторые имеют необычно высокий рост.
-
Х-хромосомы инактивированы как тельца Барра.
