
Синтез жиров
Синтез жиров представляет собой активный метаболический процесс, поскольку животные могут запасать в значительных количествах данные соединения. В среднем, у мужчины весом 70 кг на долю жиров приходится 12 кг, накопленной в них энергии хватит для поддержания основного обмена человека в течение 8 недель.
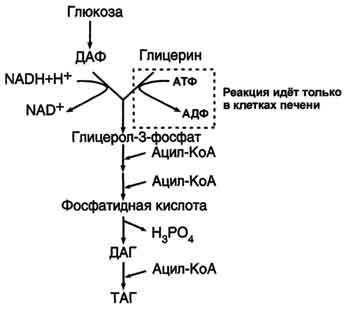
Синтез эндогенного жира из углеводов протекает в печени и в жировой ткани. Жир может синтезироваться как из продуктов распада жира, так и из углеводов. Для синтеза нейтрального жира необходим глицерин в активной форме — глицерол-3-фосфат (фосфоглицерин) и жирные кислоты в активной форме – Ацил-КоА. Глицерол-3-фосфат может быть получен путем активации глицерина с помощью глицеринкиназы, либо путем восстановления фосфодиоксиацетона, полученного при распаде глюкозы:
Вначале образуется фосфатидная кислота:
Две последующие реакции являются завершающими в синтезе триацилглицерина.
Вопросы для самоконтроля к главе 10 «Обмен жиров»
1.Какие ферменты участвуют в переваривании жиров?
2. Какова роль желчных кислот в переваривании жиров?
3. Как происходит активирование жирных кислот?
4. В чем суть — окисления жирных кислот?
5. Как происходит биосинтез жирных кислот? С чем связана распространенность в природе высших кислот с четным числом углеродных атомов?
6. Какова роль ацетил-КоА в биосинтезе жирных кислот?
Глава 11 обмен белков
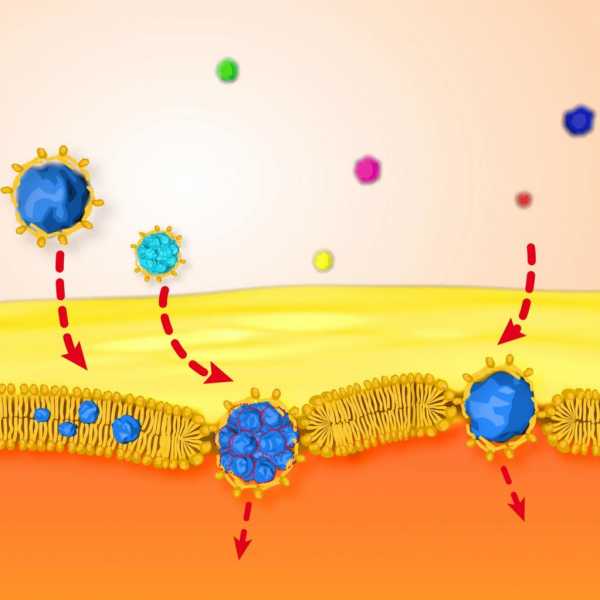
Переваривание белков происходит в желудочно-кишечном тракте под действием целого комплекса гидролаз, которые гидролитически (т.е. с участием воды) расщепляют пептидные связи, поэтому являются пептидазами (протеиназами). Гидролиз белка начинается в желудке под действием пепсина и гастриксина, которые образуются в неактивной форме, в виде проферментов – пепсиногена и гастриксиногена. Активирование данных ферментов происходит в кислой среде, создаваемой соляной кислотой желудочного сока. Пепсин легко гидролизует белки мышц, а также яичный белок и казеин. Слабее проявляется протеолитическое действие пепсина на белки соединительной ткани (коллаген, эластин). Не расщепляются пепсином протамины (белки, входящие в состав хроматина) и кератины (белки шерсти и волос).
Пепсин действует в сильно кислой среде (рН =1,0—1,5), гастриксин – в менее кислой, оптимум рН для него составляет 3,5—4,5. Данные гидролазы расщепляют не все пептидные связи, а только между определенными аминокислотами. Таким образом, в желудке образуются не отдельные аминокислоты, а фрагменты полипептидов. Дальнейший гидролиз протекает в тонком кишечнике под действием нескольких гидролаз, поступающих с пищеварительным соком поджелудочной железы и собственно кишечным соком (трипсин, химотрипсин, эластаза, аминопептидаза, карбоксипептидаза). Ферменты поджелудочной железы – трипсин, химотрипсин и карбоксипептидаза, образуются в виде проферментов – трипсиногена, химотрипсиногена и прокарбоксипептидазы, проэластазы, аминопептидаза образуется в самом тонком кишечнике сразу в активной форме. Здесь образуется также энтерокиназа, которая путем ограниченного протеолиза превращает трипсиноген в активную форму — трипсин, который также как энтерокиназа действует на трипсиноген и другие проферменты, что делает активацию лавинообразной. (Рис.29).
Энтерокиназа
Трипсиноген
Трипсин
Химотрипсиноген
Прокарбоксипептидаза
Проэластаза
Химотрипсин
Карбоксипептидаза
Эластаза
Рис.29. Активация панкреатических ферментовstudfiles.net
Синтез углеводов из жиров
Поиск ЛекцийПроцесс синтеза углеводов из жиров можно представить общей схемой:
Рисунок 7 – Общая схема синтеза углеводов из жиров
Один из основных продуктов расщепления липидов – глицерин – легко используется в синтезе углеводов через образование глицеральдегид-3-фосфата и его вступление в глюнеогенез. У растений и микроорганизмов столь же легко используется на синтез углеводов и другой важный продукт расщепления липидов– жирные кислоты (ацетил-КоА), через глиоксилатный цикл.
Но общая схема не отражает всех биохимических процессов, происходящих в результате образования углеводов из жиров.
Поэтому рассмотрим все этапы данного процесса.
Схема синтеза углеводов и жиров более полно представлена на рисунке 8 и происходит в ряд этапов.
1 этап. Гидролитическое расщепление жира под действием фермента липазы на глицерин и высшие жирные кислоты (см. п.1.2). Продукты гидролиза должны, пройдя ряд превращений, превратиться в глюкозу.
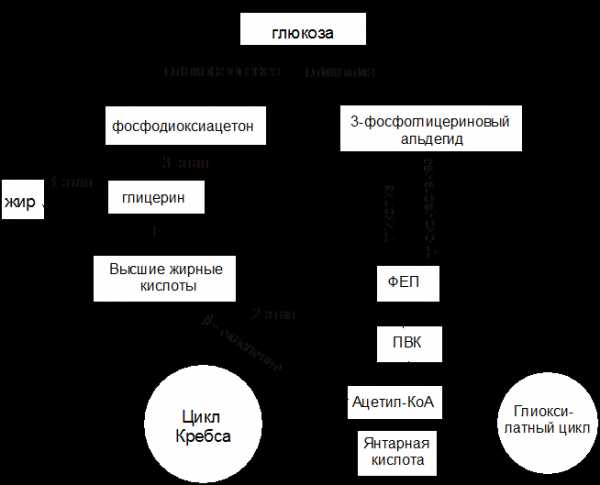
Рисунок 8 – Схема биосинтеза углеводов из жиров
2 этап. Превращение высших жирных кислот в глюкозу. Высшие жирные кислоты, которые образовались в результате гидролиза жира, разрушаются преимущественно путем b-окисления (этот процесс был рассмотрен ранее в разделе 1.2 пункт 1.2.2). Окончательным продуктом этого процесса является ацетил-КоА.
Глиоксилатный цикл
Растения, некоторые бактерии и грибы могут использовать ацетил-КоА не только в цикле Кребса, но и в цикле, получившим название глиоксилатного. Этот цикл играет важную роль в качестве связующего звена в метаболизме жиров и углеводов.
Особенно интенсивно глиоксилатный цикл функционирует в особых клеточных органеллах–глиоксисомах – при прорастании семян масличных растений. При этом происходит превращение жира в углеводы, необходимые для развития проростка семени. Этот процесс функционирует до тех пор, пока у проростка не разовьется способность к фотосинтезу. Когда в конце прорастания запасной жир истощается, глиоксисомы в клетке исчезают.
Глиоксилатный путь специфичен только для растений и бактерий, у животных организмов он отсутствует. Возможность функционирования глиоксилатного цикла связана с тем, что растения и бактерии способны синтезировать такие ферменты, как изоцитратлиаза и малатсинтаза, которые вместе с некоторыми ферментами цикла Кребса участвуют в глиоксилатном цикле.
Схема окисления ацетил-КоА по глиоксилатному пути показана на рисунке 9.
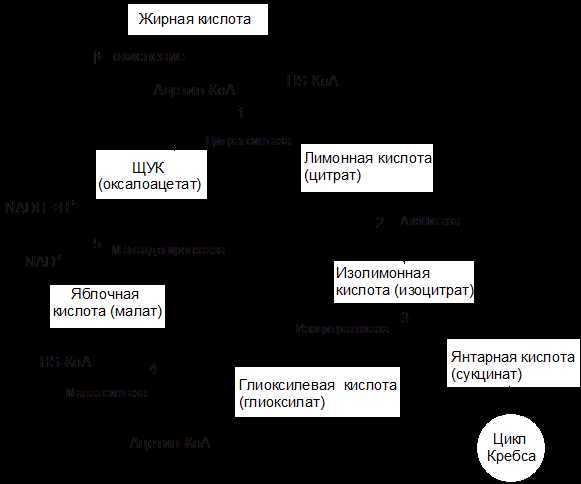
Рисунок 9 – Схема глиоксилатного цикла
Две начальные реакции (1 и 2) глиоксилатного цикла идентичны таковым цикла трикарбоновых кислот. В первой реакции (1) ацетил-КоА конденсируется с оксалоацетатом под действием цитратсинтазы, с образованием цитрата. Во второй реакции цитрат изомеризуется в изоцитрат при участии аконитатгидратазы. Следующие реакции, специфичные для глиоксилатного цикла, катализируются специальными ферментами. В третьей реакции изоцитрат под действием изоцитратлиазы расщепляется на глиоксилевую кислоту и янтарную кислоту:
В ходе четвертой реакции, катализируемой малатсинтазой, глиоксилат конденсируется с ацетил-КоА (второй молекулой ацетил-КоА, вступающей в глиоксилатный цикл) с образованием яблочной кислоты (малат):
Затем в пятой реакции малат окисляется до оксалоацетата. Эта реакция идентична конечной реакции цикла трикарбоновых кислот; она же является конечной реакцией глиоксилатного цикла, т.к. образовавшийся оксалоацетат вновь конденсируется с новой молекулой ацетил-КоА, начиная тем самым новый оборот цикла.
Образовавшаяся в третьей реакции глиоксилатного цикла янтарная кислота не используется этим циклом, а подвергается дальнейшим превращениям.
Рекомендуемые страницы:
Поиск по сайту
poisk-ru.ru
общие сведения, причины и значение
Жиры содержатся как в организмах животных, так и в растениях. Представляют собой сложные эфиры трехатомного спирта (глицерина) и кислот (олеиновой,стеариновой, линолевой, линоленовой и пальмитиновой). Это доказывается их расщеплением на кислоты и глицерин, а также синтезом жиров из описанных соединений.
Образование жиров в человеческом организме
Жиры являются сложными эфирами глицерина. При пищеварительном процессе они эмульгируются солями желчных кислот и входят в контакт с ферментами, при помощи которых гидролизуются. Таким образом, высвободившиеся жирные кислоты всасываются в слизистую пищеварительного тракта, что является окончанием процесса синтеза жиров. После этого жир проходит всю портальную систему организма в качестве микрочастиц, которые связываются с белками в крови. Метаболизм происходит в печени.
Синтез жиров возможен благодаря излишкам углеводов, которые не участвуют в образовании гликогена. Помимо этого, липиды получаются из некоторых аминокислот.
В сравнении с гликогеном, жиры являются компактным хранилищем энергии. При этом оно никак не ограничивается, так как имеет вид нейтральных липидов в жировых клетках. Липогенез происходит за счет синтеза жирных кислот, так как они содержатся почти во всех липидных группах.
Стадии липидного обмена
Жиры и жироподобные соединения проходят в теле человека следующий цикл:
- поступление в организм с пищей;
- распад на более простые соединения, процесс переваривания, всасывание;
- перенос из пищеварительной системы при помощи хилопротеинов;
- обмен сложного белка, представленного нейтральными жирами, жирными кислотами, холестеридами или фосфолипидами;
- обмен сложных липидов, сложных эфиров многоатомных спиртов и высших жирных кислот;
- обмен полициклического липофильного спирта;
- взаимопереход жирных кислот и кетоновых тел;
- процесс превращения ацетил-КоА в жирные кислоты;
- расщепление жиров на составляющие под действием липазы;
- деградация продуктов распада жирных кислот.
Значение жирных кислот для организма человека
Для нормального синтеза жиров в организме человека важны фосфолипиды. При их недостатке затормаживаются обменные процессы в печени.
Фосфолипиды распадаются на глицерин, жирные кислоты, фосфорную кислоту и азотистые основания. Первые два вещества могут либо превращаться в воду и углекислый газ, либо участвовать в синтезе жиров.
Холин (азотистое основание) важен для образования метионина и креатина. Метионин необходим для нормальной работы печени, снижения уровня холестерина в крови, а также антидепрессивного эффекта. Креатин отвечает за энергетический обмен в мышечных и нервных клетках. Ацетилхолин (продукт холина) нормализует передачу нервного возбуждения.
Именно жиры дают энергию адезинтрифосфат-молекулам, которые отвечают за все биохимические процессы в организме.
Таким образом, синтез жиров в клеточных мембранах важен для протекания многочисленных химических реакций. Без них человеческий организм не сможет нормально функционировать.
Причины нарушений переваривания жиров
Сбои усвоения жиров могут быть вызваны следующими причинами:
- Закупорка желчевыводящих протоков, что ведет к проблемам с секрецией. Вызвано подобное состояние может быть наличием камней или опухолей. Уменьшенная выработка желчной секреции ведет к трудностям смешения жиров и, таким образом, невозможности гидролизовать жирные соединения.
- Проблемы с выработкой сока в поджелудочной железе. Это также влияет на гидролиз жиров.
Каждая из описанных выше проблем ведет к увеличению количества жира в твердых продуктах жизнедеятельности человека. Возникает так называемый «жирный стул». Это состояние чревато тем, что перестают усваиваться жирорастворимые витамины A, E, D и К, а также крайне важные для организма жирные кислоты. Длительный «жирный стул» ведет к недостаточности этих веществ и развитию соответствующих клинических симптомов.

Также сбой переваривания жиров влечет трудности в усвоении нелипидных веществ, так как жир имеет свойство обволакивать пищу, чем препятствует воздействию на нее ферментов.
Заболевания, вызванные сбоем синтеза жиров
Нарушение липидного обмена может вести за собой следующие состояния:
- Ожирение. Возникает как при нарушении пищевых привычек, сопряженных с малоподвижным образом жизни, так и при наличии гормонального дисбаланса.
- Абеталипопротеинемия. Редкое наследственное заболевание, при котором в крови отсутствуют определенные липопротеиды. Жиры скапливаются в слизистой. Развивается деформация эритроцитов.
- Кахексия. Малое употребление калорий приводит к уменьшению жировой ткани в организме. Такое состояние может возникать при наличии опухолей, при хронических заболеваниях инфекционного характера, плохом питании или сбоях в метаболизме.
- Атеросклероз. Хроническое заболевание артерий, вызванное нарушением обмена липидов, сопряженное с отложением холестериновых бляшек на сосудистых стенках. В дальнейшем это чревато появлением склероза (разрастанием соединительной ткани), что ведет к деформации сосудов вплоть до их полной закупорки. Атеросклероз провоцирует ишемическую болезнь сердца.
- Артериосклероз Менкеберга. Данное заболевание похоже на атеросклероз. Однако его принципиальное отличие состоит в том, что сосуды деформируются и закупориваются не под влиянием соединительных тканей, а из-за кальциноза — скопления отложения солей. При таком поражении не формируются бляшки. Кроме того, болезнь провоцирует иные осложнения, главным из которых является аневризм.
Синтез жиров в клетках растений
Обменные процессы в растительных тканях претерпевают изменения по завершении периода цветения. Когда ослабевает синтез белков, жиры начинают образовываться из углеводов. Этот процесс протекает вплоть до полного созревания семян. Синтез жиров из углеводов и синтез белков из аминокислот важны для периода размножения.
Наибольшим содержанием жиров характеризуются масличные культуры. Это необходимо учитывать тем, что желает скорректировать собственный вес.
Липидный обмен в науке
Сегодня синтез жиров, подходящих для питания, возможен путем этерификации глицерином жирных кислот, которые, в свою очередь, создаются за счет окисления парафинов. Так как и жирные кислоты, и глицерин получают из каменного угля, существует реальный способ проведения полного синтеза пищевых жиров. Эти открытия стали возможны благодаря работам Ф. Велера, А. В. Г. Кольбе, М. Бертло и А. М. Бутлерова. Именно они доказали связь органических и неорганических веществ, а также возможность их взаимопревращения.
Полученные знания успешно применяются в пищевой, фармацевтической и химической промышленности. Однако сегодня целесообразнее получать жиры из естественных источников (растительных и животных), так как синтез не является выгодной экономической процедурой.
fb.ru
4. Синтез жиров
Синтезируются жиры из глицерина и жирных кислот.
Глицерин в организме возникает при распаде жира (пищевого и собственного), а также легко образуются из углеводов.
Жирные кислоты синтезируются из ацетилкофермента А. Ацетилкофермент А – универсальный метаболит. Для его синтеза необходимы водород и энергия АТФ. Водород же получается из НАДФ.Н2. В организме синтезируются только насыщенные и мононасыщенные (имеющие одну двойную связь) жирные кислоты. Жирные кислоты, имеющие две и более двойных связей в молекуле, называемые полинасыщенные, в организме не синтезируются и должны поступать с пищей. Для синтеза жира могут быть использованы жирные кислоты – продукты гидролиза пищевого и собственного жиров.
Все участники синтеза жира должны быть в активном виде: глицерин в форме глицерофосфата, а жирные кислоты в формеацетилкофермента А. Синтез жира осуществляется в цитоплазме клеток (преимущественно жировой ткани, печени, тонкой кишки). Пути синтеза жиров представлены в схеме.
Следует отметить, что глицерин и жирные кислоты могут быть получены из углеводов. Поэтому при избыточном потреблении их на фоне малоподвижного образа жизни развивается ожирение.
ДАФ –дигидроацетонфосфат,
ДАГ – диацилглицерин.
ТАГ – триацилглицерол.
Общая характеристика липопротеинов. Липиды в водной среде (а значит, и в крови) нерастворимы, поэтому для транспорта липидов кровью в организме образуются комплексы липидов с белками – липопротеины.
Все типы липопротеинов имеют сходное строение – гидрофобное ядро и гидрофильный слой на поверхности. Гидрофильный слой образован белками, которые называют апопротеинами,и амфифильными молекулами липидов – фосфолипидами и холестеролом. Гидрофильные группы этих молекул обращены к водной фазе, а гидрофобные части – к гидрофобному ядру липопротеина, в котором находятся транспортируемые липиды.
Апопротеины выполняют несколько функций:
• формируют структуру липопротеинов;
• взаимодействуют с рецепторами на поверхности клеток и таким образом определяют, какими тканями будет захватываться данный тип липопротеинов;
• служат ферментами или активаторами ферментов, действующих на липопротеины.
Липопротеины. В организме синтезируются следующие типы липопротеинов: хиломикроны (ХМ), липопротеины очень низкой плотности (ЛПОНП), липопротеины промежуточной плотности (ЛППП), липопротеины низкой плотности (ЛПНП) и липопротеины высокой плотности (ЛПВП).Каждый из типов ЛП образуется в разных тканях и транспортирует определѐнные липиды. Например, ХМ транспортируют экзогенные (пищевые жиры) из кишечника в ткани, поэтому триацилглицеролы составляют до 85% массы этих частиц.
Свойства липопротеинов. ЛП хорошо растворимы в крови, неопалесцируют, так как имеют небольшой размер иотрицательный зарядна
поверхности. Некоторые ЛП легко проходят через стенки капилляров кровеносных сосудов и доставляют липиды к клеткам. Большой размер ХМ не позволяет им проникать через стенки капилляров, поэтому из клеток кишечника они сначала попадают в лимфатическую систему и потом через главный грудной проток вливаются в кровь вместе с лимфой. Судьба жирных кислот, глицерола и остаточных хиломикронов. В результате действия ЛП-липазы на жиры ХМ образуются жирные кислоты и глицерол. Основная масса жирных кислот проникает в ткани. В жировой ткани в абсорбтивный период жирные кислоты депонируются в виде триацилглицеролов, в сердечной мышце и работающих скелетных мышцах используются как источник энергии. Другой продукт гидролиза жиров, глицерол, растворим в крови, транспортируется в печень, где в абсорбтивный период может быть использован для синтеза жиров.
Гиперхиломикронемия, гипертриглицеронемия.После приѐма пищи, содержащей жиры, развивается физиологическая гипертриглицеронемия и, соответственно, гиперхиломикронемия, которая может продолжаться до нескольких часов.Скорость удаления ХМ из кровотока зависит от:
• активности ЛП-липазы;
• присутствия ЛПВП, поставляющих апопротеины С-II и Е для ХМ;
• активности переноса апоС-II и апоЕ на ХМ.
Генетические дефекты любого из белков, участвующих в метаболизме ХМ, приводят к развитию семейной гиперхиломикронемии – гиперлипопротеинемии типа I.
Содержание жиров в пищевых продуктах.Жиры как запасные вещества находятся в соединительной ткани животных и рыб, в жировой ткани, подкожной клетчатке, печени и костях, а также в семенах растений и иногда в мякоти плодов.
В растениях одного и того же вида состав и свойства жира могут колебаться в зависимости от климатических условий произрастания. Содержание и качество жиров в животном сырье также зависит от породы, возраста, степени упитанности, пола, сезона года и т.д.
Жиры широко используют, при производстве многих пищевых продуктов, они обладают высокой калорийностью и пищевой ценностью, вызывают длительное чувство насыщения. Жиры являются важными вкусовыми и структурными компонентами в процессе приготовления пищевых продуктов, оказывают значительное влияние на внешний вид пищи. При жарке жир играет роль среды, передающей тепло.
Данные о содержании жиров в некоторых продуктах
|
Название продукта |
Примерное содержа-ние жиров в пищевых продуктах, % на сырую массу |
Название продукта |
Примерное содержа-ние жиров в пищевых продуктах, % на сырую массу |
|
Семена: |
Хлеб ржаной |
1,20 | |
|
Подсолнечника |
35-55 |
Овощи свежие |
0,1-0,5 |
|
Конопли |
31-38 |
Плоды свежие |
0,2-0,4 |
|
Мака |
40 |
Говядина |
3,8-25,0 |
|
Какао-бобы |
55 |
Свинина |
6,3-41,3 |
|
Орехи арахиса |
40-55 |
Баранина |
5,8-33,6 |
|
Орехи грецкие (ядра) |
58-74 |
Рыба |
0,4-20 |
|
Хлебные злаки: |
Молоко коровье |
3,2-4,5 | |
|
Пшеница |
2,3 |
Масло сливочное |
61,5-82,5 |
|
Рожь |
2,0 |
Маргарин |
82,5 |
|
Овес |
6,2 |
Яйца |
12,1 |
В жирах, полученных из растительных и животных тканей, кроме глицеридов, могут находиться свободные жирные кислоты, фосфатиды, стеролы, пигменты, витамины, вкусовые и ароматические вещества, ферменты, белки и др., которые влияют на качество и свойства жиров. На вкус и запах жиров также оказывают влияние вещества, образующиеся в жирах при хранении (альдегиды, кетоны, перекисные и другие соединения).
Жиры в организм человека должны постоянно поступать с пищей. Потребность в жирах зависит от возраста, характера работы, климатических условий и других факторов, но в среднем в сутки взрослому человеку необходимо от 80 до 100 г жиров. В суточном рационе должно быть примерно 70 % животных и 30 % растительных жиров.
studfiles.net
Липогенез.
АТФ необходим для синтеза активных форм субстратов, используемых в процессе синтеза жира.
Для синтеза нейтрального жира необходим глицерин в активной форме — глицерол-3-фосфат (фосфоглицерин). Глицерол-3-фосфат может быть получен двумя способами:
Путем активации глицерина с помощью глицеринкиназы.
2. Путем восстановления фосфодиоксиацетона, полученного при распаде глюкозы.
Кроме глицерина, для синтеза нейтрального жира необходимы жирные кислоты в активной форме. Активная форма любой жирной кислоты – Ацил-КоА. Образуется при участии фермента ацил-КоА-синтазы.
Здесь наблюдается глубокий распад АТФ до АМФ. АМФ не может вступить в окислительное фосфорилирование. Поэтому существует реакция: АТФ + АМФ 2АДФ. Поэтому затраты на активацию молекулы жирной кислоты эквивалентны затрате двух АТФ. Следующим этапом на пути синтеза жира является реакция образования фосфатидной кислоты:
Реакция катализируется ключевым ферментом липогенеза – глицерол-3-фосфатацилтрансферазой. Для этого фермента нет аллостерических эффекторов, но обнаружен адипсин (ацилстимулирующий белок), который облегчает взаимодействие Ацил-КоА с ферментом. Адипсин является продуктом протеолиза одного из компонентов системы комплемента. Относится к гормонам местного действия, так как вырабатывается в жировой ткани и действует там же.
Две последующие реакции являются завершающими в синтезе триацилглицерина.
Реакции синтеза не зависят от того, каково происхождение веществ – участников реакций.
Жир может синтезироваться как из продуктов распада жира, так и из углеводов. Синтез эндогенного жира из углеводов протекает в печени и в жировой ткани. Ацил-КоА синтезируется из Ацетил-КоА. ГБФ-путь распада углеводов обеспечивает синтез энергией. Образование Ацетил-КоА происходит в матриксе митохондрий. Синтез жирных кислот идет в цитоплазме на мембранах эндоплазматического ретикулума путем постепенного удлинения ацетил-КоА на 2 углеродных атома за каждый цикл. Удлинение высших жирных кислот, содержащих более 16 углеродных атомов, идёт путём реакций, обратных -окислению (О-окислении смотрите ниже, раздел «Катаболизм жирных кислот»). Однако реакции синтеза жирных кислот до 16 углеродных атомов принципиально отличаются от реакций, обратных-окислению. Они протекают обходным обратным путём.
Отличия реакций синтеза высших жирных кислот от обратных бета-окислению:
1. -окисление протекает в митохондриях, а синтез жирных кислот протекает в цитоплазме на мембранах эндоплазматического ретикулума. Но образуется Ацетил-КоА в митохондриях, а через мембраны сам проходить не может. Поэтому существуют механизмы транспорта Ацетил-КоА из митохондрий в цитоплазму.
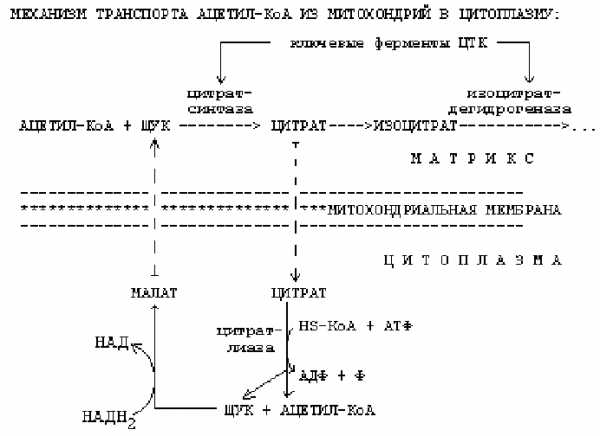
Ключевыми ферментами ЦТК являются цитратсинтаза и изоцитратдегидрогеназа. Основные аллостерические регуляторы этих ферментов — это АТФ и АДФ. Если в клетке много АТФ, то АТФ выступает как ингибитор этих ключевых ферментов. Однако изоцитратдегидрогеназа угнетается АТФ сильнее, чем цитратсинтетаза. Это приводит к накоплению цитрата и изоцитрата в матриксе митохондрии. При накоплении цитрат выходит из митохондрии в цитоплазму. В цитоплазме есть фермент цитратлиаза. Этот фермент расщепляет цитрат на ЩУК и Ацетил-КоА.
Условием для выхода Ацетил-КоА из митохондрии в цитоплазму является хорошее обеспечение клетки АТФ. Если АТФ в клетке мало, то Ацетил-КоА расщепляется до СО2 и Н2О.
2. В ходе -окисления промежуточные продукты связаны с HS-КоА, а при синтезе жирных кислот промежуточные продукты связаны с особым ацил-переносящим белком (АПБ). Это сложный белок. Его небелковая часть похожа по строению на КоА. Небелковая часть АПБ состоит из тиоэтиламина, пантотеновой кислоты (витамин В3) и фосфата.
3. При -окислении в качестве окислителя используются НАД и ФАД. При синтезе ЖК нужен восстановитель — используется НАДФ*Н2.
В клетке существует 2 основных источника НАДФ*Н2 для синтеза жирных кислот:
а) ГМФ-путь распада углеводов
б) В реакции окисления малата (смотрите рисунок).
Эта реакция протекает в цитоплазме и катализируется ферментом МАЛАТДЕГИДРОГЕНАЗОЙ ДЕКАРБОКСИЛИРУЮЩЕЙ, тривиальное название которого – ЯБЛОЧНЫЙ ФЕРМЕНТ.
4. В ходе синтеза жирных кислот в каждом цикле удлинения используется не сам Ацетил-КоА, а его производное — малонил-КоА (при -окислении каждый цикл укорочения приводит к образованию Ацетил-КоА).
Эту реакцию катализирует фермент АЦЕТИЛ-КоА-КАРБОКСИЛАЗА. Это ключевой фермент в мультиферментной системе синтеза ЖК. Этот фермент регулируется по типу отрицательной обратной связи. Ингибитором является продукт синтеза: ацил-КоА с длинной цепью (n=16) — пальмитоил-КоА. Активатором является цитрат. В состав небелковой части этого фермента входит витамин H (биотин).
Далее происходит поэтапное удлинение молекулы Ац-КоА на 2 углеродных атома за каждый этап за счет малонил-КоА. В процессе удлинения малонил-КоА теряет СО2. После образования малонил-КоА основные реакции синтеза жирных кислот катализируются одним ферментом — синтетазой жирных кислот (фиксирован на мембранах эндоплазматического ретикулума). Синтетаза жирных кислот содержит 7 активных центров. Участок, связывающий малонил-КоА, содержит небелковый компонент – витамин B3 (пантотеновую кислоту).
РЕАКЦИИ СИНТЕЗА ЖИРНЫХ КИСЛОТ.
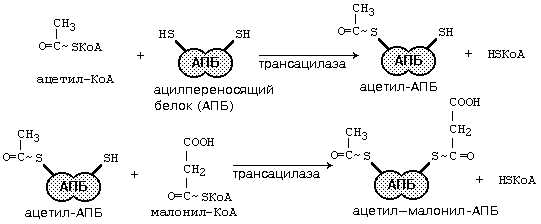
После этого ацил-АПБ вступает в новый цикл синтеза. К свободной SH-группе ацилпереносящего белка присоединяется новая молекула малонил-КоА. Затем происходит отщепление ацильного остатка, и он переносится на малонильный остаток с одновременным декарбоксилированием, и цикл реакций повторяется.
Таким образом, углеводородная цепочка будущей жирной кислоты постепенно растет (за каждый цикл – на два углеродных атома). Это происходит до момента, пока она не удлинится до 16 углеродных атомов (в случае синтеза пальмитиновой кислоты) или более (синтез других жирных кислот). Вслед за этим происходит тиолиз, и образуется в готовом виде активная форма жирной кислоты – ацил-КоА.
УСЛОВИЯ ДЛЯ ПРОТЕКАНИЯ СИНТЕЗА ВЫСШИХ ЖИРНЫХ КИСЛОТ:
1) Поступление углеводов, при окислении которых образуются необходимые субстраты и НАДФН2.
2) Высокий энергетический заряд клетки – высокое содержание АТФ, которое обеспечивает выход цитрата из митохондрий в цитоплазму.
Обмен углеводов и обмен жиров очень тесно связаны. Углеводы легко могут превращаться в жиры, а вот превращение жиров в углеводы невозможно. Жиры не могут превращаться в углеводы, так как Ацетил-КоА не может превращаться в пируват. Обмен жиров и углеводов объединяется как энергетический обмен, который находится под контролем гормонов.
studfiles.net
Липогенез.
АТФ необходим для синтеза активных форм субстратов, используемых в процессе синтеза жира.
Для синтеза нейтрального жира необходим глицерин в активной форме — глицерол-3-фосфат (фосфоглицерин). Глицерол-3-фосфат может быть получен двумя способами:
Путем активации глицерина с помощью глицеринкиназы.
2. Путем восстановления фосфодиоксиацетона, полученного при распаде глюкозы.
Кроме глицерина, для синтеза нейтрального жира необходимы жирные кислоты в активной форме. Активная форма любой жирной кислоты – Ацил-КоА. Образуется при участии фермента ацил-КоА-синтазы.
Здесь наблюдается глубокий распад АТФ до АМФ. АМФ не может вступить в окислительное фосфорилирование. Поэтому существует реакция: АТФ + АМФ 2АДФ. Поэтому затраты на активацию молекулы жирной кислоты эквивалентны затрате двух АТФ. Следующим этапом на пути синтеза жира является реакция образования фосфатидной кислоты:
Реакция катализируется ключевым ферментом липогенеза – глицерол-3-фосфатацилтрансферазой. Для этого фермента нет аллостерических эффекторов, но обнаружен адипсин (ацилстимулирующий белок), который облегчает взаимодействие Ацил-КоА с ферментом. Адипсин является продуктом протеолиза одного из компонентов системы комплемента. Относится к гормонам местного действия, так как вырабатывается в жировой ткани и действует там же.
Две последующие реакции являются завершающими в синтезе триацилглицерина.
Реакции синтеза не зависят от того, каково происхождение веществ – участников реакций.
Жир может синтезироваться как из продуктов распада жира, так и из углеводов. Синтез эндогенного жира из углеводов протекает в печени и в жировой ткани. Ацил-КоА синтезируется из Ацетил-КоА. ГБФ-путь распада углеводов обеспечивает синтез энергией. Образование Ацетил-КоА происходит в матриксе митохондрий. Синтез жирных кислот идет в цитоплазме на мембранах эндоплазматического ретикулума путем постепенного удлинения ацетил-КоА на 2 углеродных атома за каждый цикл. Удлинение высших жирных кислот, содержащих более 16 углеродных атомов, идёт путём реакций, обратных -окислению (О -окислении смотрите ниже, раздел «Катаболизм жирных кислот»). Однако реакции синтеза жирных кислот до 16 углеродных атомов принципиально отличаются от реакций, обратных -окислению. Они протекают обходным обратным путём.
Отличия реакций синтеза высших жирных кислот от обратных бета-окислению:
1. -окисление протекает в митохондриях, а синтез жирных кислот протекает в цитоплазме на мембранах эндоплазматического ретикулума. Но образуется Ацетил-КоА в митохондриях, а через мембраны сам проходить не может. Поэтому существуют механизмы транспорта Ацетил-КоА из митохондрий в цитоплазму.
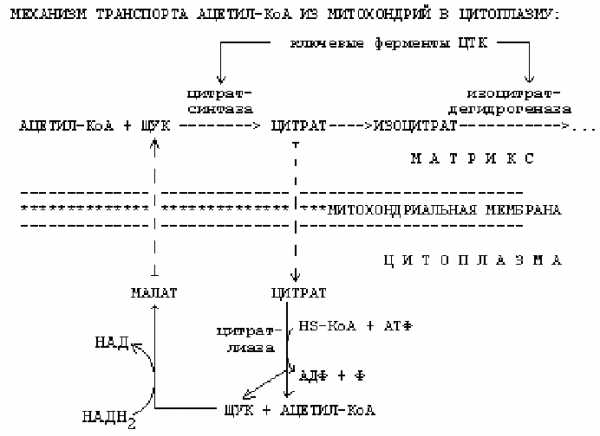
Ключевыми ферментами ЦТК являются цитратсинтаза и изоцитратдегидрогеназа. Основные аллостерические регуляторы этих ферментов — это АТФ и АДФ. Если в клетке много АТФ, то АТФ выступает как ингибитор этих ключевых ферментов. Однако изоцитратдегидрогеназа угнетается АТФ сильнее, чем цитратсинтетаза. Это приводит к накоплению цитрата и изоцитрата в матриксе митохондрии. При накоплении цитрат выходит из митохондрии в цитоплазму. В цитоплазме есть фермент цитратлиаза. Этот фермент расщепляет цитрат на ЩУК и Ацетил-КоА.
Условием для выхода Ацетил-КоА из митохондрии в цитоплазму является хорошее обеспечение клетки АТФ. Если АТФ в клетке мало, то Ацетил-КоА расщепляется до СО2 и Н2О.
2. В ходе -окисления промежуточные продукты связаны с HS-КоА, а при синтезе жирных кислот промежуточные продукты связаны с особым ацил-переносящим белком (АПБ). Это сложный белок. Его небелковая часть похожа по строению на КоА. Небелковая часть АПБ состоит из тиоэтиламина, пантотеновой кислоты (витамин В3) и фосфата.
3. При -окислении в качестве окислителя используются НАД и ФАД. При синтезе ЖК нужен восстановитель — используется НАДФ*Н2.
В клетке существует 2 основных источника НАДФ*Н2 для синтеза жирных кислот:
а) ГМФ-путь распада углеводов
б) В реакции окисления малата (смотрите рисунок).
Эта реакция протекает в цитоплазме и катализируется ферментом МАЛАТДЕГИДРОГЕНАЗОЙ ДЕКАРБОКСИЛИРУЮЩЕЙ, тривиальное название которого – ЯБЛОЧНЫЙ ФЕРМЕНТ.
4. В ходе синтеза жирных кислот в каждом цикле удлинения используется не сам Ацетил-КоА, а его производное — малонил-КоА (при -окислении каждый цикл укорочения приводит к образованию Ацетил-КоА).
Эту реакцию катализирует фермент АЦЕТИЛ-КоА-КАРБОКСИЛАЗА. Это ключевой фермент в мультиферментной системе синтеза ЖК. Этот фермент регулируется по типу отрицательной обратной связи. Ингибитором является продукт синтеза: ацил-КоА с длинной цепью (n=16) — пальмитоил-КоА. Активатором является цитрат. В состав небелковой части этого фермента входит витамин H (биотин).
Далее происходит поэтапное удлинение молекулы Ац-КоА на 2 углеродных атома за каждый этап за счет малонил-КоА. В процессе удлинения малонил-КоА теряет СО2. После образования малонил-КоА основные реакции синтеза жирных кислот катализируются одним ферментом — синтетазой жирных кислот (фиксирован на мембранах эндоплазматического ретикулума). Синтетаза жирных кислот содержит 7 активных центров. Участок, связывающий малонил-КоА, содержит небелковый компонент – витамин B3 (пантотеновую кислоту).
РЕАКЦИИ СИНТЕЗА ЖИРНЫХ КИСЛОТ.
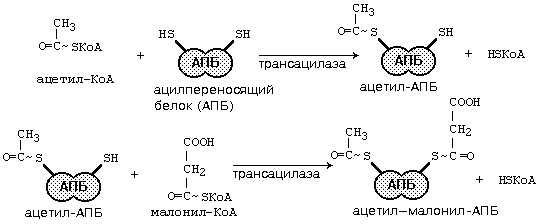
После этого ацил-АПБ вступает в новый цикл синтеза. К свободной SH-группе ацилпереносящего белка присоединяется новая молекула малонил-КоА. Затем происходит отщепление ацильного остатка, и он переносится на малонильный остаток с одновременным декарбоксилированием, и цикл реакций повторяется.
Таким образом, углеводородная цепочка будущей жирной кислоты постепенно растет (за каждый цикл – на два углеродных атома). Это происходит до момента, пока она не удлинится до 16 углеродных атомов (в случае синтеза пальмитиновой кислоты) или более (синтез других жирных кислот). Вслед за этим происходит тиолиз, и образуется в готовом виде активная форма жирной кислоты – ацил-КоА.
УСЛОВИЯ ДЛЯ ПРОТЕКАНИЯ СИНТЕЗА ВЫСШИХ ЖИРНЫХ КИСЛОТ:
1) Поступление углеводов, при окислении которых образуются необходимые субстраты и НАДФН2.
2) Высокий энергетический заряд клетки – высокое содержание АТФ, которое обеспечивает выход цитрата из митохондрий в цитоплазму.
Обмен углеводов и обмен жиров очень тесно связаны. Углеводы легко могут превращаться в жиры, а вот превращение жиров в углеводы невозможно. Жиры не могут превращаться в углеводы, так как Ацетил-КоА не может превращаться в пируват. Обмен жиров и углеводов объединяется как энергетический обмен, который находится под контролем гормонов.
studfiles.net
Биосинтез триацилглицеролов. Синтез жиров из углеводов. Регуляция процессов.
Опубликовано в Шпоры по биохимии
Кроме пищевых жиров источником жирных кислот в организме служит их синтез из глюкозы. Непосредственным предшественником жирных кислот при их синтезе в организме является ацетил-КоА, т. е. то же вещество, которое образуется при р-окислении жирных кислот. Несмотря на то, что все реакции р-окисления обратимы, они не используются для синтеза жирных кислот.Ацетил-КоА для синтеза жирных кислот образуется путем окислительного де- карбоксилирования пирувата. Кроме того, окисление и синтез жирных кислот разделены в пространстве: окисление происходит в митохондриях, а синтез — в цитозоле.
Большинство жирных кислот, необходимых человеку, может синтезироваться в организме из углеводов. Часть углеводов, поступающих с пищей, превращается в организме в жиры, особенно если количество углеводов превышает необходимое для возобновления запасов гликогена в печени и мышцах. Схема этого превращения представлена на рис. 10.15.
Глюкоза служит источником ацетил-КоА, из которого синтезируются жирные кислоты. Необходимый для восстановительных реакций НАДФН поставляется за счет окисления глюкозы в пентозофосфатном пути, а также за счет дегидрирования яблочной кислоты НАДФ-зависимой малатдегидрогеназой. Глице- рол-3-фосфат получается путем восстановления диоксиацетонфосфата — промежуточного продукта гликолиза Таким образом, из глюкозы образуется все, что необходимо для синтеза жиров.Синтез жиров из углеводов наиболее активно происходит в печени, жировой ткани и лактирующих молочных железах. Синтез триацилглицеринов из глицерол-3-фосфата и ацил-КоА представлен на схеме:
Лизофосфатидная кислота
Фосфатидная кислота
+Вода — Н3РО4
Диацилглицерин
Триацилглицерин
biohimist.ru
