
Одомашнивание как начальный этап селекции. Методы современной селекции
Loading…
Уважаемые ученики, для изучения нового материала вам понадобиться учебник «Биология. 10 класс.» К. Беляева и Г. М. Дымшица, Швецов
- 42,43 «Одомашнивание как начальный этап селекции. Методы современной селекции.»
Выполните следующие шаги:
- Прочитайте текст параграфа.
- Изучите теоретический материал
Селекция – это наука, основанная на выведении новых сортов, видов растений или животных с целью повышения качества породы или сорта. Целью селекции является улучшение наследственных свойств растений, животных в том направлении, которое принесет максимум пользы и будет выгодно для человека. Пример: Выведение новых пород норки (с голубой, сапфировой, жемчужной окраской меха) является заслугой селекции.
В широком смысле слова селекция как процесс изменения домашних животных и культурных растений, по выражению Н. И. Вавилова, «представляет собой эволюцию, направляемую волей человека». Как род практической деятельности людей селекция возникла еще на заре человеческой культуры. Однако селекция стала наукой сравнительно недавно. Теория селекции стала успешно развиваться благодаря эволюционной теории Ч. Дарвина о творческой роли отбора, а затем на основе генетики.
Все современные сорта растений и породы животных, без которых немыслима современная цивилизация, созданы человеком благодаря селекции. В наше время перед селекцией стоят громадные задачи по созданию новых высокопродуктивных пород животных и сортов растений, приспособленных к условиям современного индустриального сельского хозяйства, а также нужных человеку штаммов микроорганизмов.
Важное место в теории селекции занимает генетика, поэтому развитие генетических основ селекции необходимо для сельскохозяйственной практики.
Основные методы селекции – отбор и гибридизация. Отбор бывает массовый и индивидуальный. При индивидуальном отборе выбирают отдельную особь с нужными признаками и получают от неё потомство. Индивидуальный отбор применяется для самоопыляющихся растений и животных. Массовый отбор применятся в селекции перекрёстноопыляемых растений некоторых животных.
Одомашнивание или иначе доместикация (от лат. domesticus — «домашний») — это процесс изменения диких животных или растений, при котором на протяжении многих поколений они содержатся человеком генетически изолированными от их дикой формы и подвергаются искусственному отбору.
Все современные домашние животные и культурные растения произошли от диких предков. Процесс превращения диких животных и растений в культурные формы называют одомашниванием. Главным фактором одомашнивания служит искусственный отбор организмов, отвечающих требованиям человека. Если самка первобытного тура — предка современного крупного рогатого скота — продуцировала лишь десятки или сотни килограммов молока, необходимые ей для кормления теленка, то отдельные коровы современных пород дают до 15 000 кг молока за лактацию, т.
Для животных, например, первым условием, а вместе с тем и показателем одомашнивания было создание (путем отбора) особей, способных к контактам с человеком, к сосуществованию с ним. Иначе говоря, человек изменял поведение животных, превращая их из диких в домашних. В условиях естественной природы культурные, т. е. одомашненные, формы существовать, как правило, не могут.
Уже на первых этапах процесс одомашнивания вызвал резкое повышение изменчивости животных и растений, что создало предпосылки для успешного осуществления искусственного отбора. В результате этого между породами современных домашних животных и сортами растений возникли большие различия. Эти различия во многих случаях превышали различия не только между видами, но и родами.
Первые попытки одомашнивания животных и растений предпринимались людьми еще за 20—30 тыс. лет до н. э. Одомашнивание животных началось, вероятно, со случайного выращивания диких животных человеком. Некоторые из этих еще диких животных могли существовать в контакте с человеком и начали размножаться в условиях, созданных для них.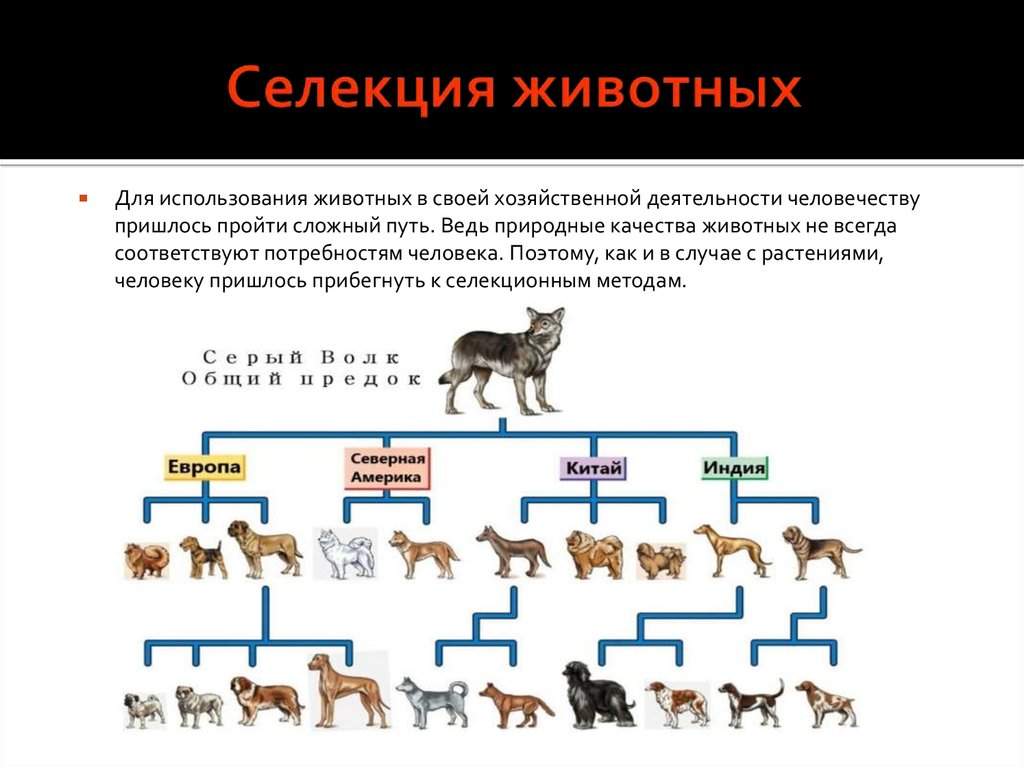 Так начался первый этап их одомашнивания.
Так начался первый этап их одомашнивания.
Широкое одомашнивание начинается с VIII—VI тыс. до н. э. Именно тогда человек вовлек в культуру подавляющее большинство животных и растений.
Некоторые виды животных и растений были одомашнены значительно позднее. Так, кроликов люди стали разводить лишь в Средневековье, сахарную свеклу стали возделывать как полевое сахарное растение только в XIX в., а мяту — в XX в.
В наше время человек для удовлетворения своих потребностей продолжает одомашнивать новые виды животных и растений. Для получения высококачественной пушнины в XX столетии создана новая отрасль животноводства — пушное звероводство. Несомненно, человек в дальнейшем будет вовлекать в одомашнивание все новые виды животных и растений.
Центры происхождения культурных растений. Выдающийся вклад в развитие представлений о центрах происхождения культурных растений внесли Н. И. Вавилов и его сотрудники.
В результате многочисленных экспедиций, организованных Н. И. Вавиловым в самые отдаленные уголки планеты, где возникли древние земледельческие цивилизации, была собрана уникальная, самая крупная в мире коллекция разнообразных культурных растений. Именно она послужила фундаментом той огромной коллекции, которая ныне находится в Институте растениеводства им. Н. И. Вавилова (Санкт-Петербург) и активно служит интересам науки и практики.
И. Вавиловым в самые отдаленные уголки планеты, где возникли древние земледельческие цивилизации, была собрана уникальная, самая крупная в мире коллекция разнообразных культурных растений. Именно она послужила фундаментом той огромной коллекции, которая ныне находится в Институте растениеводства им. Н. И. Вавилова (Санкт-Петербург) и активно служит интересам науки и практики.
В 1760-е годы английский селекционер Р. Бейкуэлл сформулировал два правила селекции крупного рогатого скота: “Скрещивай лучшее с лучшим” и “Подобное рождает подобное”. Трудами этого специалиста Англия во многом обязана своим лидирующим положением в племенном животноводстве. О каких методах селекции идёт речь в данных высказываниях?
Методы селекции растений, животных и микроорганизмов
Селекция
— это наука о методах создания новых и улучшении уже существующих пород
животных, сортов растений, штаммов микроорганизмов, которые имеют полезные для
человека свойства.
Что благодаря селекции можно разработать новые формы, которые имеют какие-либо особенности, отличающие их от исходных диких видов.
Каким же образом это происходит?
Сегодня на уроке мы рассмотрим основные методы селекции. Их достаточно много, однако все они направлены на создание новых улучшенных форм.
Основными методами селекции являются, отбор, который подразделяется на массовый отбор и индивидуальный отбор. А также (скрещивание) гибридизация, которая подразделяется на неродственное скрещивание, включающее межвидовую (отдаленную) гибридизацию и внутривидовую гибридизацию, и близкородственное скрещивание.
А также к основным методам селекции относят искусственный (индуцированный) мутагенез.
Давайте с ними познакомимся с перечисленными методами.
На прошлом уроке мы
говорили, что основным методом селекции является искусственный
отбор.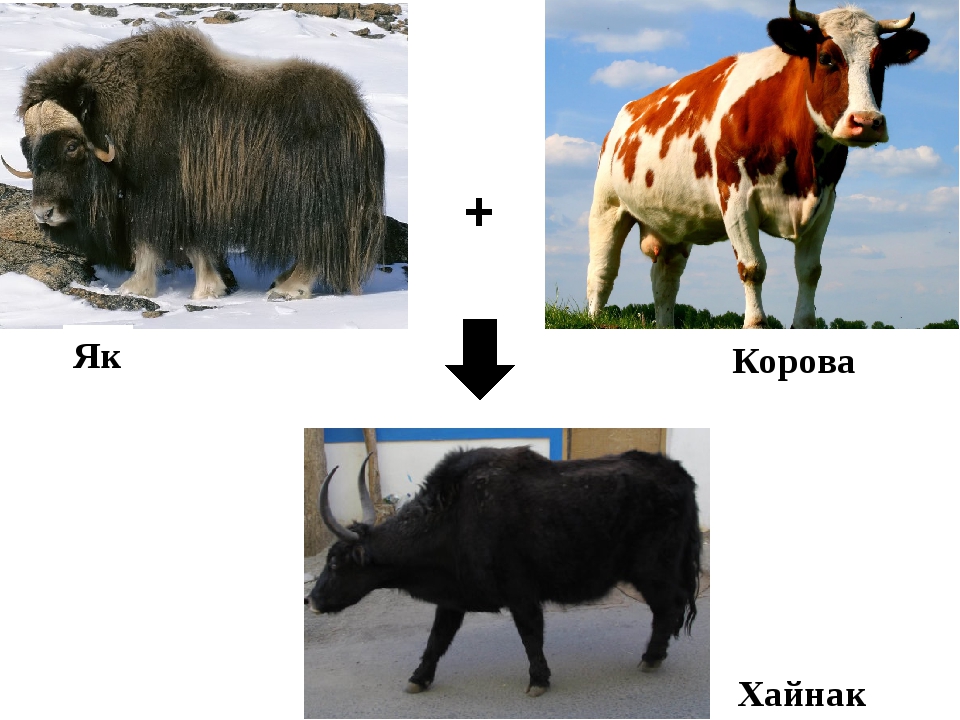
То есть отбираются формы с какими-либо измененными и лучшими признаками, либо приспособлениями, которых нет у других форм. Полезные не для животного или растения, а для самого человека.
В селекции различают два основных типа отбора: массовый и индивидуальный.
При массовом отборе, отбираются группы особей по внешним (фенотипическим) признакам без проверки их генотипа.
Скрещивают их между собой, получая гибриды второго поколения. Среди них опять производят массовый отбор особей с нужными признаками и так далее.
К примеру, возьмём целое поле высаженной люцерны на котором произрастает около одной тысячи растений.
Люцерна посевная ─ многолетнее травянистое растение из семейства бобовых, одна из самых ценных кормовых трав.
Внимательно рассмотрев
каждое растение, учтя их продуктивность по семенам и зеленой
массе при уборке. Мы выбираем 100 лучших по всем показателям. Объединив семена
лучших растений, высаживаем их на следующий год. И ожидаем получить улучшенную
продуктивность. Если все прошло удачно, и мы добились улучшения, то можно
считать, что массовый отбор был эффективен.
Мы выбираем 100 лучших по всем показателям. Объединив семена
лучших растений, высаживаем их на следующий год. И ожидаем получить улучшенную
продуктивность. Если все прошло удачно, и мы добились улучшения, то можно
считать, что массовый отбор был эффективен.
Так, например, в хозяйствах из всей популяции кур оставляют для размножения особей с большой яйцекладностью. Которые при размножении дают аналогичное потомство. Таким образом, благодаря этому методу возможно быстро улучшить сорта и породы.
Индивидуальный отбор
Он проводиться уже не по фенотипу, а по генотипу. В таком случае полученное потомство четко оценивают на наличие интересующих селекционера признаков. На последующих этапах отбора используют только тех особей, которые дали наибольшее число потомков с лучшими показателями.
Для примера вернёмся к тому же полю с люцерной. Опять отбираем из тысячи высаженных растений 100 лучших по всем показателям.
Однако, в случае
индивидуального отбора мы не станем объединять их семена, а посеем в следующем
году семена каждого растения отдельно. Затем оценим и генотипы отобранных
растений, и их фенотипические показатели.
Затем оценим и генотипы отобранных
растений, и их фенотипические показатели.
Если каждое отобранное из популяции по выдающимся показателям растение или животное сохраняет свои показатели в потомстве, то индивидуальный отбор продолжается и в последующих поколениях.
Сейчас при искусственном осеменении коров, от одного быка с интересующими свойствами можно получить до тридцати пяти тысяч телят.
Благодаря индивидуальному отбору от одного вида дикого сизого голубя выведено около ста пятидесяти пород домашних голубей;
Большинство сортов пшеницы, ячменя, овса были получены методом индивидуального отбора.
Метод индивидуального отбора наиболее применим к самоопыляющимся растениям (пшеница, ячмень, овес). Потомство одной самоопыляющейся особи называют чистой линией. Так как в размножении участвует одна особь, которая опыляет себя сама. Чистая линия, в которой генетическая информация не меняется.
В отличие от
перекрёстного опыления, где происходит обмен генетической информацией между
особями.
Гибридизация — это процесс скрещивания родительских особей и получения от них гибридов.
То есть объединяются генетические материалы разных клеток в одной клетке.
Гибрид – это организм или клетка, полученные в результате скрещивания генетически различающихся форм.
Первые программы гибридного разведения были внедрены уже в конце XIX века в Италии, когда от фитофторы погибли все цитрусовые плантации. Примерно в это же время во Флориде сильные заморозки погубили практически весь урожай лимонов и апельсинов. В надежде защититься от дальнейших напастей, селекционеры прибегли к помощи гибридизации. Именно из этих экспериментов и получились «улучшенные» виды цитрусовых.
Фрукты-гибриды клементины, танжерины, плуоты, пичерины удивительны на слух и превосходны на вкус. Кроме того, они ещё и полезны для здоровья.
Клементи́н — гибрид мандарина и апельсина-королька.
Грейпфрут получился путем естественного скрещивания апельсина с помело.
Пичерин − результат скрещивания персика и нектарина. По вкусу — нечто среднее между нектарином и персиком.
Нектакотум «состоит» из равных частей нектарина, абрикоса и сливы.
Перейдём к родственной гибридизации.
Инбридинг — это близкородственное скрещивание (внутрипородное или внутрисортовое), при котором в качестве исходных форм используются потомки одних и тех же родителей, либо потомки скрещиваются с родительскими формами.
Такое скрещивание применяется для того, чтобы перевести большинство генов породы или сорта в гомозиготное состояние и избежать расщепления по хозяйственно ценным признакам в ряду поколений.
Например, заводчики животных для поддержания породы часто пользуются таким методом гибридизации. Инбридинг позволяет закрепить уникальный, неожиданно возникший признак и передать его по наследству.
Если скрещивать
близкородственные особи, то появляется потомство с необходимыми усиленными
признаками. Однако другие признаки могут резко ухудшаться.
Однако другие признаки могут резко ухудшаться.
Такие неблагоприятные последствия близкородственного скрещивания называют депрессией. Снижение жизнеспособности и продуктивности потомства.
Например, щенки будут рождаться больными, нежизнеспособными, с генетическими отклонениями, и от них нельзя будет получить потомство.
При депрессии, родственные спаривания характеризуются генетическими изменениями.
Ещё Дарвин проанализировал данные результаты самоопыления растений и открыл закон, согласно которому, все существа, получающиеся при скрещивании особей, не состоящих в родстве, получают от этого только пользу. В то время как скрещивание родственных особей приносит только вред.
Чем объясняется такое неблагоприятное влияние? Одной из основных причин служит переход большинства генов в гомозиготное состояние.
Рассмотрим внутривидовую неродственную гибридизацию.
Внутривидовое скрещивание
— это скрещивание между особями, принадлежащими к одному виду, которое приводит
к образованию гибридного организма.
При скрещивании между собой разных сортов растений или пород животных одного вида первое гибридное поколение будет отличаться улучшенными признаками. Например, крупными размерами, повышенной устойчивостью и плодовитостью.
Гетерозис — это увеличение жизнеспособности гибридов вследствие унаследования определённого набора аллелей различных генов от своих разнородных родителей.
Сущность гетерозиса заключается в том, что первое гибридное поколение обладает повышенной урожайностью и жизнеспособностью. Однако уже начиная со второго поколения эффект гетерозиса обычно снижается.
Гетерозис наблюдается как между видами, так и внутри видов.
Межвидовая гибридизация.
Это ещё один главный метод селекции.
Межвидовая (отдалённая) гибридизация — это скрещивание особей, принадлежащих к разным видам, часто приводящее к существенному снижению жизнеспособности, частичной или полной стерильности.
Стерильность ─
отсутствие способности к оплодотворению, т. е. бесплодие.
е. бесплодие.
Межвидовые скрещивания используют для обогащения генетической основы устойчивости сортов.
Например, при скрещивании самого крупного представителя дикого барана Архар и овцы породы прекос, получается баран породы архаромеринос.
Такая порода имеет улучшенные признаки мясо-шерстного направления продуктивности.
Так межродовая гибридизация позволяет передать новому сорту более широкую экологическую пластичность, устойчивость к неблагоприятным факторам среды, и другие ценные свойства.
Приведем примеры межвидовой гибридизации
Тигон ─ гибрид тигра и львицы
Гролар ─ гибрид белого и бурого медведя
Лошак ─ гибрид жеребца и ослицы
Леопон ─ гибрид леопарда и львицы
Однако, как мы уже сказали организмы, которые появились в результате межвидовой гибридизации, частично или полностью стерильны.
Это происходит потому что
число хромосом и их форма, у особей отличаются.
Поэтому при мейозе хромосомы не сходятся гомологичными парами и не конъюгируют между собой.
Вспомним уже изученные определения.
Конъюгация — процесс точного и тесного сближения гомологичных хромосом.
Кроссинговер − процесс обмена участками гомологичных хромосом во время конъюгации в профазе I мейоза.
Таким образом, в результате межвидовой гибридизации происходят нарушения при кроссинговере и межвидовые гибриды становятся бесплодными.
При помощи методов межвидовой гибридизации был выведен гибрид кобылицы с ослом, который называют мул.
Как и все гибриды межвидовой гибридизации мулы бесплодны, однако они очень сильны, выносливы и долго живучи.
Ещё один пример межвидовой гибридизации, который широко применяется. Это гибрид пшеницы и ржи, названный «тритикале» является полиплоидом. То есть он имеет увеличенное число хромосом.
Тритикале обладает
повышенной морозостойкостью (больше чем у озимой пшеницы), устойчивостью против
грибковых и вирусных болезней, пониженной требовательностью к плодородию почвы.
Содержание белка в зерне тритикале выше, чем у пшеницы на один полтора процента и на три четыре процента, чем у ржи.
Советский ботаник Пётр Миха́йлович Жуко́вский, говорил «что человек питается преимущественно продуктами полиплоидами. Многие из полиплоидов ─ важнейший источник сырья для промышленности.»
Сахарный тростник, земляной орех, земляника, банан, ананас, груша, слива, являются естественными полиплоидами.
Наряду с высокой продуктивностью полиплоиды характеризуются повышенной концентрацией белка, витаминов, углеводов, имеют более мощное строение и оказываются гораздо устойчивее к неблагоприятным условиям.
Ещё один важный способ получения новых сортов искусственный мутагенез.
Он осуществляется путём применения ионизирующих излучений и химических мутагенов, которые значительно увеличивают число мутаций.
Таким образом, учёные пытаются получить организмы с новыми полезными свойствами.
Процесс получения
необходимых человеку веществ с помощью живых клеток называют – биотехнологией.
В начале XX века активно развивалась бродильная и микробиологическая промышленность. Пивоварение было одним из первых применений биотехнологии.
В эти же годы были предприняты первые попытки наладить производство антибиотиков, пищевых концентратов, полученных из дрожжей. В 1940 году удалось выделить и очистить первый антибиотик – пенициллин.
Так же благодаря биотехнологии, решается проблема обеспечения населения нашей планеты продуктами питания. При помощи увеличения синтеза пищевого белка микроорганизмами.
Микроорганизмы примерно в 10-100 тысяч раз быстрее синтезируют белок, чем животные. Например, 400-килограммовая корова производит в день 400 граммов белка, а 400 килограммов бактерий — 40 тысяч тонн белка.
Мутационная селекция | МАГАТЭ
Классическая селекция в самой простой ее форме предполагает избирательное выведение растений с желаемыми характеристиками и элиминирование или «выбраковку» тех, характеристики которых менее желательны. Еще одна методика, получившая название «кроссбридинг», предполагает преднамеренное скрещивание близко или отдаленно связанных, но совместимых по половому признаку родительских линий с целью выведения новых сортов сельскохозяйственных культур или линий с желаемыми свойствами. Обе методики предполагают работу с несколькими поколениями, при этом для устранения нежелательных характеристик и развития нужных требуется пять или более лет.
Еще одна методика, получившая название «кроссбридинг», предполагает преднамеренное скрещивание близко или отдаленно связанных, но совместимых по половому признаку родительских линий с целью выведения новых сортов сельскохозяйственных культур или линий с желаемыми свойствами. Обе методики предполагают работу с несколькими поколениями, при этом для устранения нежелательных характеристик и развития нужных требуется пять или более лет.
МАГАТЭ совместно с ФАО содействует государствам-членам в разработке и внедрении ядерных технологий, которые за счет использования гамма-излучения и рентгеновского излучения могут вызывать мутацию растений и тем самым значительно ускорять процесс селекции. Смежных биотехнологии также могут применяться с целью идентификации и отбора требуемых мутаций.
Наращивание темпов работы без ущерба для окружающей среды
Мутационная селекция в случае самоопыляемых культур, размножение которых происходит за счет семян, основана на самооплодотворении (или самоопылении) мутантов до тех пор, пока индуцированная желаемая характеристика не будет стабильно экспрессироваться во многих поколениях мутантов. Зачастую для сохранения желаемых характеристик требуется обратное скрещивание с исходным не мутантным генотипом (часть последовательности ДНК клетки, которая определяет специфические характеристики).
Зачастую для сохранения желаемых характеристик требуется обратное скрещивание с исходным не мутантным генотипом (часть последовательности ДНК клетки, которая определяет специфические характеристики).
Мутационная селекция основана на индуцировании мутаций и обнаружении мутаций. Она обладает множеством сравнительных преимуществ: это экономичная, быстрая, проверенная и надежная методика. Кроме того, эта технология является передаваемой, широко применяемой, не опасной и безвредной для окружающей среды. На основе более 210 видов растений из более чем 70 стран официально выпущено для коммерческого использования более 3200 мутантных сортов, включая многочисленные сельскохозяйственные культуры, декоративные растения и деревья (источник: Совместная база данных ФАО/МАГАТЭ по мутантным сортам).
Растительные биотехнологии играют важную роль в мутационной селекции. Методы культивирования тканей растений являются мощными инструментами, позволяющими сократить время, необходимое для создания разводимых линий мутантов (т. е. таких, которые стабильно передают конкретные признаки своему потомству). Это ограничивает использование индуцированных мутаций культур, которые являются рецессивными (в генетике – когда одна характеристика гена не экспрессируется, поскольку имеется экспрессия другой, доминантной).
е. таких, которые стабильно передают конкретные признаки своему потомству). Это ограничивает использование индуцированных мутаций культур, которые являются рецессивными (в генетике – когда одна характеристика гена не экспрессируется, поскольку имеется экспрессия другой, доминантной).
Одним из таких средств селекции растений является методика двойного гаплоида, которая предполагает удвоение хромосомного набора гаплоида – организма или клетки, которая имеет только один элемент каждой хромосомной пары.
Другим методом является определение молекулярных маркеров, тесно связанных с определенными искомыми признаками, которые затем могут быть использованы для быстрого подтверждения этих признаков. Разработка и распространение таких молекулярных маркеров имеет потенциал в отношении дальнейшего укрепления программ мутационной селекции растений, в частности, для основных пищевых культур, таких как рис.
Ветеринары рассказали об опасности модных пород собак и кошек
Селекция, в результате которой собаки и кошки приобретают новую форму головы, ушей или носа, меняют окрас или размер, приносит животным постоянные боли и грозит преждевременной гибелью. Такими выводами представители палаты ветеринаров Берлина поделились с информационным агентством МИА «Россия сегодня».
Такими выводами представители палаты ветеринаров Берлина поделились с информационным агентством МИА «Россия сегодня».
В качестве примера немецкие ветеринары приводят моду на собак и кошек с напоминающей человеческую округленной головой. Такая селекция приводит к проблемам дыхания, вплоть до приступов удушья. Из-за уменьшения размеров челюсти не остается места для языка, зубов и животные не могут нормально есть, а слишком выступающие вперед глаза подвергаются дополнительным повреждениям.
К проблемным признакам берлинские эксперты также относят отсутствующий или слишком короткий хвост, неполное количество пальцев на лапах, округленную «человекоподобную» голову, изменение густоты и цвета меха, а также карликовость. По данным немецкого сайта partner-hund.de, больше других от нездоровой селекции страдают австралийские овчарки, французские бульдоги, чихуахуа, мопсы, таксы, немецкие овчарка, шарпеи и китайские хохлатые собачки.
Фото pixabay.com
С мнением коллег из Германии согласны и российские ветеринары. Как рассказала изданию «Ветеринария и жизнь» ветеринарный врач-кардиолог кандидат ветеринарных наук Наталья Анисимова, чтобы закрепить желаемые свойства, селекционеры не всегда внимательно относятся к состоянию здоровья особей и нередко используют инбридинг – близкородственное скрещивание. Чистопородные животные часто обладают генетическими дефектами, которые влияют на строение и функции организма, в том числе на сердечно-сосудистую систему, вызывают структурные изменения тканей, приводящие к развитию болезней сердца, считает ветеринар.
Как рассказала изданию «Ветеринария и жизнь» ветеринарный врач-кардиолог кандидат ветеринарных наук Наталья Анисимова, чтобы закрепить желаемые свойства, селекционеры не всегда внимательно относятся к состоянию здоровья особей и нередко используют инбридинг – близкородственное скрещивание. Чистопородные животные часто обладают генетическими дефектами, которые влияют на строение и функции организма, в том числе на сердечно-сосудистую систему, вызывают структурные изменения тканей, приводящие к развитию болезней сердца, считает ветеринар.
Среди искусственно выведенных пород собак и кошек, продолжает Наталья Анисимова, чаще встречаются врожденные пороки и нарушения ритма, дегенеративные заболевания клапанного аппарата и миокарда. По ее словам, часто единственным признаком врожденного порока становится внезапная гибель щенка или котенка.
Фото rony michaud/pixabay.com
Тем, кто собирается завести дома собаку или кошку, Наталья Анисимова советует перед приобретением ознакомиться с особенностями породы и предрасположенностью ее представителей к болезням в специальной литературе или у ветеринарного врача, который проведет обследование животного на предмет врожденных патологий.
Селекция, подготовка к ЕГЭ по биологии
Селекция (лат. selectio — выбирать) — наука и отрасль практической деятельности, направленная на создание новых сортов растений, пород животных и штаммов микроорганизмов, обладающих полезными для человека свойствами.
Этими полезными свойствами могут быть размер и форма плодов, урожайность, удойность у коров, устойчивость к факторам внешней среды (к засушливому климату, к морозу).
Основы селекции
В основе селекции лежит способность генотипа живых организмов к изменениям, что происходит главным образом за счет комбинативной и мутационной изменчивости. В процессе селекции происходит искусственный отбор организмов с полезными для человека свойствами и их размножение.
В результате множества последовательных скрещиваний, в конце концов, селекционерам удается достичь желаемой цели: вывести гибридов
с нужными признаками.
Мутационная изменчивость существует благодаря мутациям — случайным ненаправленным изменениям генотипа. Благодаря мутациям, к примеру, возник безалкалоидный сорт люпина. И.В. Мичуриным на яблоне сорта Антоновка Могилевская были обнаружены необычайно крупные плоды, ветвь с которым послужила для появления нового сорта — Антоновки шестистограммовой. Эти плоды — результат произошедшей в естественных условиях мутации соматических клеток.
«Сколько ждать этой естественной мутации?» — спросите вы. Может один день, а может и 100, и 10000 лет — всем властвует случайность. На наш век может не выпасть удача, а мы такого допустить не можем! 🙂
Именно по этой причине в селекции растений часто используются искусственно вызванные мутации — авто- и аллополиплоидию.
Автополиплоидия
Автополиплоидия — кратное (4n,6n,8n) увеличение исходного набора хромосом, который характерен для
особей вида.
Автополиплоидия возникает в результате обработки почек колхицином, который нарушает образование нитей веретена деления, и, соответственно, нарушает расхождение хромосом в мейозе, в результате чего набор хромосом в половых клетках (гаметах) оказывается удвоенным. Таким способом получают полиплоиды — сорта растений, обладающие повышенной урожайностью.
Существуют различные тетраплоидные сорта свеклы, мака, кукурузы и других сельскохозяйственных культур, которые отличаются большими размерами плодов.
Аллополиплоидия
Аллополиплоидия (греч. állos — другой и polýploos — многократный) — соединение в клетках организма хромосомного набора от разных видов или родов, в результате которого образуется гибридная зигота.
Благодаря аллополиплоидии получают новые сорта растений. Наиболее известным примером является гибрид ржи и пшеницы — тритикале. Некоторые межвидовые гибриды табака обладают повышенной устойчивостью к возбудителям заболеваний мучнистой росы, табачной мозаики.
Некоторые межвидовые гибриды табака обладают повышенной устойчивостью к возбудителям заболеваний мучнистой росы, табачной мозаики.
В рамках биотехнологии разработаны методы, с помощью которых стало возможным создание бактерий, синтезирующих полезные для человека белки, многие из которых используются как лекарства: аминокислоты, антибиотики, инсулин.
Скрещивание особей в селекции
Каждое скрещивание как сдача новых карт: может повезет, а может и нет. Вполне возможно, что особь унаследует полезные признаки от родителей и сможет передать их своим потомкам, всегда есть и шанс того, что появятся новые полезные для человека признаки, равно как и шанс, что ничего полезного из проводимого скрещивания не выйдет.
Возможны несколько вариантов скрещивания:
- Близкородственное скрещивание (инбридинг — от англ.
 in — внутри + breeding — разведение)
in — внутри + breeding — разведение) - Неродственное скрещивание (аутбридинг — от англ.
 out — вне + breeding — разведение)
out — вне + breeding — разведение) - Отдаленная гибридизация
Близкородственное скрещивание в течение нескольких поколений приводит к переходу генов в гомозиготное состояние, вследствие чего потомство ослабевает и становится более подвержено наследственным заболеваниям.
Замечу, что под инбридингом подразумевают близкородственное скрещивание животных. Для самоопыления у растений существует иной термин — инцухт.
В селекции инбридинг применяют для выведения чистых линий (гомозиготных особей — aa, AA, bb, BB), которые используются, например, для анализирующего скрещивания. Инбридинг использовался при выведении абсолютно всех пород животных, и в настоящее время активно используется в питомниках для выведения нужных пород животных (кошек, собак и т.д.)
Аутбридинг заключается в скрещивании неродственных особей, которые могут принадлежать к одному сорту, породе, виду или роду. Аутбридинг ведет к явлению гетерозиса — получения гетерозисных форм, которые превосходят родительских особей по ряду признаков.
Гетерозис — явление увеличения жизнеспособности особей у гибридов, которые получены при скрещивании двух чистых линий. Такой эффект связан с переходом генов в гетерозиготное состояние, что повышает выживаемость организмов, плодовитость, и множество других полезных свойств.
Применение отдаленной гибридизации заключается в скрещивании особей, принадлежащих к разным родам и видам. Такие особи обладают крайне полезными для человека свойствами, но часто бесплодны (стерильны).
Известным примером отдаленной гибридизации является мул — гибрид осла (самца) и лошади (самки). Отличаются большой выносливостью и
работоспособностью, живут до 40 лет, обладают хорошим иммунитетом к заболеваниям, не требовательны в корме и уходе.
Отличаются большой выносливостью и
работоспособностью, живут до 40 лет, обладают хорошим иммунитетом к заболеваниям, не требовательны в корме и уходе.
Обратный пример: гибрид ослицы (самки) и жеребца (самца) — лошак. Встречаются гораздо реже по сравнению с мулом, так как обладают меньшей выносливостью и работоспособностью. В большинстве случаев бесплодны.
Отбор в селекции
Отбор в селекции осуществляет человек с единственной целью: размножить особей с нужными и полезными признаками, свойствами. Очевидно, что такой отбор называется искусственным, в противовес естественному отбору, главный критерий которого — приспособленность.
Отбор может осуществляться двумя способами:
- Массовый отбор
- Индивидуальный отбор
Отбор организмов исключительно на основе внешних данных (фенотипа). Основным критерием для человека служит проявление признака:
размер плодов, цвет лепестков, цвет листьев и т.д. Этот вид отбора характеризуется массовостью и быстротой.
Основным критерием для человека служит проявление признака:
размер плодов, цвет лепестков, цвет листьев и т.д. Этот вид отбора характеризуется массовостью и быстротой.
В результате массового отбора формируется группа особей, которые обладают нужными и полезными для человека признаками. В дальнейшем они подвергаются размножению.
Выборочный отбор и сохранение особей с ценными для человека признаками. В ходе индивидуального отбора оценивается не только фенотип, но и генотип, вследствие чего данный вид отбора занимает большее время, но оказывается более эффективен.
Индивидуальный отбор требует оценки потомства от выбранной особи в ряду поколений. Иногда подобный отбор применяют у самоопыляемых
растений: пшеницы, ячменя — с целью получения чистых линий. Как было сказано ранее, чистые линии характеризуются гомозиготностью и являются исходным материалом для селекции.
Как было сказано ранее, чистые линии характеризуются гомозиготностью и являются исходным материалом для селекции.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Селекция: основные методы и достижения
Вспомните!
Что такое селекция?
Селекция (от лат. selectio — отбор) — наука о создании новых и улучшении существующих сортов растений, пород животных и штаммов микроорганизмов. Одновременно под селекцией понимают и сам процесс создания сортов, пород и штаммов. Теоретической основой селекции является генетика.
Одновременно под селекцией понимают и сам процесс создания сортов, пород и штаммов. Теоретической основой селекции является генетика.
Приведите примеры известных вам пород животных и сортов растений.
Сорта яблок Антоновка, груша Северянка, породы собак: ротвейлер, карликовый пудель, колли.
Вопросы для повторения и задания
1. Что такое селекция?
Селекция (от лат. selectio — отбор) — наука о создании новых и улучшении существующих сортов растений, пород животных и штаммов микроорганизмов. Одновременно под селекцией понимают и сам процесс создания сортов, пород и штаммов. Теоретической основой селекции является генетика.
2. Что называют породой, сортом, штаммом?
Порода, сорт или штамм — это совокупность особей одного вида, искусственно созданная человеком и характеризующаяся определёнными наследственными свойствами.
3. Какие основные методы селекции вы знаете?
4. Выберите критерии и сравните массовый и индивидуальный отбор.
5. Какие сложности возникают при постановке межвидовых скрещиваний?
Отдалённая гибридизация заключается в скрещивании разных видов. В растениеводстве с помощью отдалённой гибридизации создана новая зерновая культура — тритикале, гибрид ржи с пшеницей. Эта культура сочетает многие свойства пшеницы (высокие хлебопекарные качества) и ржи (способность расти на бедных песчаных почвах). Классическим примером межвидовых гибридов в животноводстве является мул, полученный при скрещивании осла с кобылицей, который значительно превосходит родителей по выносливости и работоспособности. В Казахстане при скрещивании диких горных баранов-архаров с тонкорунными овцами была создана знаменитая архаромериносная порода овец. Однако применение межвидовых скрещиваний имеет определённые сложности, потому что получаемые гибриды часто оказываются бесплодными (стерильными) или низкоплодовитыми. Стерильность гибридов связана с отсутствием у них парных гомологичных хромосом. Это делает невозможным процесс конъюгации. Следовательно, мейоз не может завершиться, и половые клетки не образуются.
Стерильность гибридов связана с отсутствием у них парных гомологичных хромосом. Это делает невозможным процесс конъюгации. Следовательно, мейоз не может завершиться, и половые клетки не образуются.
6. Получают ли и используют ли в вашем регионе межвидовые гибриды? Используя дополнительные источники информации, выясните, гибридами каких видов являются такие организмы, как бестер, хонорик, лошак, рафанобрассика. Какой интерес представляют они для сельского хозяйства?
Подумайте! Вспомните!
1. Что схожего и чем отличаются методы селекции растений и животных?
2. Почему для каждого региона нужны свои сорта растений и породы животных? Какие сорта и породы характерны для вашего региона? В чём их особенности и преимущества?
Так как условия среды в различных регионах разные, и сорта и породы должны быть приспособлены к конкретным условиям. Особенности растениеводства Южного Урала
Особенности растениеводства Южного Урала
Основные культуры района
3. Из большого разнообразия видов животных, обитающих на Земле, человек отобрал для одомашнивания сравнительно немного видов. Как вы считаете, чем это объясняется?
Процесс одомашнивания диких животных начинается с искусственной селекции отдельных индивидов для получения потомства с определенными признаками, необходимыми человеку. Индивиды, как правило, выбираются в соответствии с определёнными желаемыми характеристиками, включая снижение агрессивности по отношению к человеку и представителям собственного вида. В этом отношении принято говорить об укрощении дикого вида. Целью одомашнивания является использование животного в сельском хозяйстве в качестве сельскохозяйственного животного или в качестве домашнего питомца. Если эта цель достигнута, можно говорить об одомашненном животном. Одомашнивание животного коренным образом изменяет условия для дальнейшего развития вида. Естественное эволюционное развитие заменяется искусственной селекцией по критериям разведения. Таким образом, в рамках одомашнивания меняются генетические свойства вида.
Естественное эволюционное развитие заменяется искусственной селекцией по критериям разведения. Таким образом, в рамках одомашнивания меняются генетические свойства вида.
4. Гетерозис в последующих поколениях обычно не сохраняется, затухает. Почему это происходит?
При скрещивании разных пород животных или сортов растений, а также при межвидовых скрещиваниях в первом поколении у гибридов повышается жизнеспособность и наблюдается мощное развитие. Явление превосходства гибридов по своим свойствам родительских форм получило название гетерозиса, или гибридной силы. Проявляется в первом поколении, а во втором затухает.
5. Как вы думаете, почему лигры рождаются только в зоопарках и не встречаются в дикой природе? Объясните свою точку зрения.
Лигры — межвидовые гибриды между львом и тигрицей — выглядят как огромные львы с размытыми полосами. Следовательно, его родители относятся к одному и тому же биологическому роду пантер, но разным видам. Внешне он заметно отличается от своего противоположного гибрида, тигрольва. Является крупнейшим представителем семейства кошачьих, существующих в настоящее время. Выглядит как гигантский лев с размытыми полосами. Лигры не встречаются в природе главным образом потому, что в естественной среде львы и тигры почти не имеют шансов встретиться: современный ареал льва включает в основном центральную и южную Африку (хотя в Индии существует последняя уцелевшая популяция азиатских львов), в то время как тигр — исключительно азиатский вид. Поэтому скрещивание видов происходит, когда животные долгое время живут в одном вольере или клетке (например, в зоопарке или цирке), но потомство дают лишь 1—2 % пар, из-за чего в мире сегодня числится не более двух десятков лигров.
Внешне он заметно отличается от своего противоположного гибрида, тигрольва. Является крупнейшим представителем семейства кошачьих, существующих в настоящее время. Выглядит как гигантский лев с размытыми полосами. Лигры не встречаются в природе главным образом потому, что в естественной среде львы и тигры почти не имеют шансов встретиться: современный ареал льва включает в основном центральную и южную Африку (хотя в Индии существует последняя уцелевшая популяция азиатских львов), в то время как тигр — исключительно азиатский вид. Поэтому скрещивание видов происходит, когда животные долгое время живут в одном вольере или клетке (например, в зоопарке или цирке), но потомство дают лишь 1—2 % пар, из-за чего в мире сегодня числится не более двух десятков лигров.
6. Как вы считаете, может ли применяться массовый отбор при разведении животных? Докажите свое мнение.
Не применяют. массовый отбор — это отбор по фенотипу. Индивидуальный — по генотипу. Производители у животных — особи с хорошо прописанной родословной т.е. генотип по нужным признакам достаточно хорошо известен. Да и особенности животных — необходимо время для достижения половой зрелости, небольшое число потомства (по сравнению с растениями — сейчас можно считать решенной проблемой — искусственное осеменение, суррогатные самки) и невозможность бесполого размножения.
Производители у животных — особи с хорошо прописанной родословной т.е. генотип по нужным признакам достаточно хорошо известен. Да и особенности животных — необходимо время для достижения половой зрелости, небольшое число потомства (по сравнению с растениями — сейчас можно считать решенной проблемой — искусственное осеменение, суррогатные самки) и невозможность бесполого размножения.
7. Используя дополнительную литературу и ресурсы Интернета, подготовьте сообщение или презентацию об истории селекции с древних времён до настоящего времени.
Селекция как способ выведения пород домашних животных и сортов культурных растений существует издавна. Около 8000-9000 лет назад с появлением сельского хозяйства на Ближнем Востоке, а позже в Европе и Азии началось развитие растениеводства и животноводства. Уже с тех времен люди стали заниматься искусственным отбором с целью выведения пород животных и сортов растений с хозяйственно-ценными качествами. О первых селекционных мероприятиях, известных еще почти 6000 лет назад в Эламе (Двуречье), можно судить по изображе¬нию родословной лошадей, обнаруженной на печатке. Существуют также сведения, что арабы задолго до новой эры применяли искусственное опыление финиковых пальм. В Римской империи сохранились документы с подробным описанием приемов, используемых при разведении животных. В трудах ученых Древнего Китая и Древнего Рима имеются указания на значение отбора колосьев у злаков и даются рекомендации по проведению такого отбора.
О первых селекционных мероприятиях, известных еще почти 6000 лет назад в Эламе (Двуречье), можно судить по изображе¬нию родословной лошадей, обнаруженной на печатке. Существуют также сведения, что арабы задолго до новой эры применяли искусственное опыление финиковых пальм. В Римской империи сохранились документы с подробным описанием приемов, используемых при разведении животных. В трудах ученых Древнего Китая и Древнего Рима имеются указания на значение отбора колосьев у злаков и даются рекомендации по проведению такого отбора.
На первых порах селекционные мероприятия ограничивались отбором. Он носил бессознательный характер, велся длительное время (10—15 лет). Селекционеры, не имея теоретической базы, руководствовались опытом и интуицией. Они учитывали полезные свойства родительских особей, но целенаправленно проводить селекцию не могли. Результаты скрещивания часто оказывались неожиданными, и в потомстве не обнаруживалось ожидаемого признака. Тем не менее, безвестные селекционеры оставили в наследство немало ценных сортов культурных растений и пород домашних животных. Например, ряд лучших сортов хлопчатника, возделываемых ныне в России и США, позаимствован у крестьян старых мексиканских деревень. Методом бессознательного отбора выведены сорта льна-долгунца в некоторых районах Пско¬ва: низкорослые растения шли на хозяйственные нужды, а семена высоких использовались на посев. Известны сорта озимой (например, Крымка, Полтавка, Сандомирка) и яровой (Улька, Гирка, Сыр-Бидай и др.) пшеницы с ценными хозяйственными качествами, выведенные в давние времена.
Например, ряд лучших сортов хлопчатника, возделываемых ныне в России и США, позаимствован у крестьян старых мексиканских деревень. Методом бессознательного отбора выведены сорта льна-долгунца в некоторых районах Пско¬ва: низкорослые растения шли на хозяйственные нужды, а семена высоких использовались на посев. Известны сорта озимой (например, Крымка, Полтавка, Сандомирка) и яровой (Улька, Гирка, Сыр-Бидай и др.) пшеницы с ценными хозяйственными качествами, выведенные в давние времена.
Однако отбор по хозяйственно-полезным признакам и свойствам без учета механизмов их наследуемости и изменчивости нередко давал нежелательные результаты. К примеру, отбор по экстерьеру тонкорунных овец на комолость приводил к появлению крипторхизма; избавление от пегости на шее у романовских овец ослабляло их жизнеспособность; повышение оброслости шерстью у овец сопровождалось снижением их веса. Не удавалось вывести и чистую линию виандоттов (порода кур) с розовидным гребнем; несмотря на выбраковывание цыплят с листовидным гребнем, они появлялись в потомстве. Очевидно, порода состояла из генерозигот по этому гену, так как гомозиготы обладали сниженной плодовитостью.
Очевидно, порода состояла из генерозигот по этому гену, так как гомозиготы обладали сниженной плодовитостью.
Все это свидетельствовало о том, что желаемый ре¬зультат нельзя получить без теоретических знаний. С конца XVIII — начала XIX в. работы селекционеров носили уже научный характер. Главной задачей селекции стало изучение генетики таких признаков, как продуктивность животных и урожайность растений. Разрешение задач селекции невозможно без знаний, касающихся генетического анализа, т. е. без знаний типа наследования признаков (доминантный или рецессивный), типа доминирования, характера наследования (аутосомное или сцепленное с полом, независимое или сцепленное), типа и характера взаимодействия генов в онтогенезе. Главное внимание селекционеры должны уделять проблемам взаимоотношения генотипа и среды, ибо от факторов последней во многом зависит экспрес¬сивность и пенетрантность изучаемых признаков.
8. Существуют ли в вашем регионе селекционные станции или центры? Какие исследования они проводят? Каковы их достижения? Вместе с учителем организуйте экскурсию на такую станцию.
Ю-У НИИ плодоовощеводства и картофелеводства, г. Челябинск
Эх, яблочко, ранеточка…
В 1931 году по инициативе И. В. Мичурина было создано первое на Южном Урале научно-исследовательское учреждение по садоводству — Уральская зональная плодово-ягодная опытная станция. Организатором этой станции был Валерий Павлович Ярушин.
А уже в следующем году начались научные исследования по селекции и подбору сортов, пригодных для выращивания в суровых условиях Челябинской и Курганской областей, в том числе и входивших в них тогда Камышловском и Каменск-Уральском районах ныне Свердловской области. Ученые приступили к обследованию и сбору лучших форм плодово-ягодных культур восточнее Уральского хребта. В 1934 году сотрудники станции зарегистрировали крупный массив дикорастущей вишни — 2270 га — в Карагайской лесной даче (Анненский бор). В том же году научная экспедиция, возглавляемая доктором сельскохозяйственных наук Э. П. Сюбаровой (БелНИИ плодоводства) и челябинским ученым М. Н. Саламатовым, обследовала заросли дикой степной вишни в Верхнеуфалейском и Полтавском районах Челябинской области. Одновременно были завезены обширные коллекции сливы с Дальнего Востока, из Канады, Северной Америки, средней полосы России, Поволжья, карзинские сливы из Сибири. Из отобранного материала в 1937 году ученые выделили сорта и предложили первый уральский сортимент ягодных культур.
П. Сюбаровой (БелНИИ плодоводства) и челябинским ученым М. Н. Саламатовым, обследовала заросли дикой степной вишни в Верхнеуфалейском и Полтавском районах Челябинской области. Одновременно были завезены обширные коллекции сливы с Дальнего Востока, из Канады, Северной Америки, средней полосы России, Поволжья, карзинские сливы из Сибири. Из отобранного материала в 1937 году ученые выделили сорта и предложили первый уральский сортимент ягодных культур.
В те годы, как в народе, так и агрономической науке, считалось, что из-за сурового климата с морозными и продолжительными зимами, садоводство на Урале невозможно. Южноуральским ученым потребовалось всего двадцать лет — по селекционным меркам весьма небольшой срок, чтобы опровергнуть это широко распространенное мнение. Благодаря селекции и изучению новых сортов плодово-ягодных культур на челябинской опытной станции, у нас стало стремительно развиваться садоводство.
Первые коллективные сады в области появились вскоре после войны. В 1948 году образовались «Тракторосад», «Дружба» в Металлургическом районе, «Локомотив» в Советском районе, сады Магнитогорского металлургического комбината. Их появлению предшествовала длительная и вязкая борьба в коридорах тогдашней власти с противниками создания таких садов. Тем не менее, коллективное садоводство развивалось и развивается до сих пор. В настоящее время садоводы производят основную часть плодоовощной продукции.
В 1948 году образовались «Тракторосад», «Дружба» в Металлургическом районе, «Локомотив» в Советском районе, сады Магнитогорского металлургического комбината. Их появлению предшествовала длительная и вязкая борьба в коридорах тогдашней власти с противниками создания таких садов. Тем не менее, коллективное садоводство развивалось и развивается до сих пор. В настоящее время садоводы производят основную часть плодоовощной продукции.
К началу 50-х годов нашими учеными были собраны и изучались 442 сортообразца яблони, в том числе 210 сортов урало-сибирской селекции. Первые официально оформленные авторскими свидетельствами сорта плодово-ягодной опытной станции были именно яблони. Основой довоенного сортимента стали местные ранетки. Они отличаются приспособленностью к уральскому и сибирскому климату, высокой урожайностью и малыми размерами плодов весом от 15 до 50 г. Старшее поколение еще помнит эти яркие красивые яблочки-ранетки — Любимец, Анисик омский, Пониклое. Они казались тогда пределом мечтаний, особенно для детворы. Но к 60-м годам в активе института было уже 25 районированных сортов:14 яблонь, 4 груши, 4 сливы и 3 ягодных культуры.
Но к 60-м годам в активе института было уже 25 районированных сортов:14 яблонь, 4 груши, 4 сливы и 3 ягодных культуры.
Расцвет
В 1964 году Уральская зональная плодово-ягодная опытная станция была переименована в Челябинскую плодоовощную селекционную станцию им. И. В. Мичурина. К тому времени в области уже успешно работал специализированный трест «Плодопром», который возглавлял Всеволод Иванович Назаров. Тесное сотрудничество науки и производства, увлеченность и энтузиазм их сотрудников подняли южноуральское садоводство на невиданные высоты. При поддержке руководителей области были созданы плодовые питомники: «Смолинский» в пригороде Челябинска, «Мичуринский» в Карталах, «Радужный» в Магнитогорске, «Тюбелясский» в горнозаводской зоне. При Смолинском и Мичуринском плодопитомниках были организованы государственные сортоиспытательные участки.
Именно тогда, в 50-60-е годы в колхозах и совхозах, находящихся при МТС, закладывались небольшие сады от 20 до 100 га. Некоторые из них сохранились до сих пор. На пике развития садоводства области общественные сады занимали у нас площадь 8 тысяч гектаров, а сама отрасль была в целом прибыльной.
Некоторые из них сохранились до сих пор. На пике развития садоводства области общественные сады занимали у нас площадь 8 тысяч гектаров, а сама отрасль была в целом прибыльной.
В пятидесятые годы в нашей области началась также работа по научному обеспечению картофелеводства. К началу 80-х годов Челябинская плодоовощная селекционная станция им. И. В. Мичурина имела в своем активе 29 собственных сортов плодово-ягодных культур и картофеля, занесенных в Государственный реестр селекционных достижений, допущенных к использованию.
Ученые станции начали использовать в селекции растений мировую коллекцию форм плодово-ягодных культур и картофеля из Всесоюзного института растениеводства и других научно-исследовательских учреждений, в том числе зарубежных. В селекционный процесс вовлекался генетический фонд, внедрялись новые селекционные технологии, позволяющие сокращать селекционный процесс, возросла подготовка кадров высшей квалификации. Научные разработки южноуральских ученых стали внедряться в хозяйствах Челябинской, Курганской, Кустанайской, Оренбургской и других областях, в Республике Башкортостан.
Новое имя — новые цели
В ноябре 1991-го по решению правительства РФ Челябинская плодоовощная опытная станция им. И. В. Мичурина была преобразована в Южно-Уральский НИИ плодоовощеводства и картофелеводства. Изменилось не только название. Перед институтом были поставлены более серьезные цели и направления научно-исследовательской деятельности. Помимо селекции плодово-ягодных культур и картофеля, жизнь потребовала создания новых ресурсосберегающих экологически чистых технологий селекции и возделывания этих культур, научных исследований по гибридизации, а также производства элитных саженцев новых перспективных сортов садовых культур и семян картофеля на оздоровленной, безвирусной основе с применением биотехнологии.
За весь период деятельности института, начиная с Уральской зональной плодово-ягодной станции, челябинскими селекционерами создано более 200 сортов плодово-ягодных культур, 18 сортов картофеля, разработаны технологии промышленного и любительского садоводства, технологии производства картофеля с урожайностью до 70 т/га.
В государственный реестр селекционных достижений, допущенных к использованию, в разные годы было внесено 110 сортов. Об уровне научно-исследовательской работы говорит тот факт, что на сегодняшний день институт имеет более 100 авторских свидетельств и патентов на сорта и изобретения.
Впервые на Южном Урале созданы модели интенсивного сорта плодовых, ягодных культур, картофеля до 2020 года, разработаны схемы селекции на продуктивность, зимостойкость, в т.ч. устойчивость цветков к весенним заморозкам, качество продукции, иммунитет. Институт располагает богатым генетическим фондом садовых культур, который насчитывает 64 тысячи гибридных растений, в т.ч. 39 — плодовых, 25 тысяч сеянцев ягодных культур. Объемы гибридных скрещиваний составляют 45 тысяч цветков.
Научные исследования по селекции и агротехнике садовых культур и картофеля ведутся в творческом содружестве с ведущими научно-исследовательскими институтами и опытными станциями России, ближнего и дальнего зарубежья.
На сегодняшний день институт имеет селекционный сад площадью более 100 гектаров. Именно здесь сосредоточена основная научная деятельность южноуральских селекционеров.
Яблоня. Впервые в мировой практике садоводства селекционеры института по оригинально разработанной методике вывели сорта естественных карликов с высотой деревьев в 1,5-2,5 м. При размножении их на клоновых вегетативно размножаемых карликовых подвоях, они становятся естественными стланцами (0,8-1,5 м).
Груша — удивительная культура на Урале. Ежегодно плодоносит. Плоды вкусные, сладкие, пригодные для переработки. Привлечение в селекцию отборных форм уссурийской груши позволило создать сорта, отличающиеся высокой зимостойкостью и продуктивностью, высокими вкусовыми качествами плодов, моногенной устойчивостью к парше и полевой устойчивостью к грушевому галловому клещу.
Абрикос и слива. Выведены местные сорта и выделены отборные формы, отличающиеся повышенной зимостойкостью плодовых почек и высоким качеством плодов.
Вишня. Накоплены экспериментальные данные для создания генотипов, устойчивых к коккомикозу. Продолжается выделение доноров с геном моноустойчивости к коккомикозу и выявление форм степной вишни с полевой устойчивостью к коккомикозу. С этой целью и для пополнения коллекции проведены экспедиции по обследованию дикоросов вишни на территории Башкирии, Челябинской и Курганской областей. Отобраны формы степной и лесной вишни, устойчивые к коккомикозу, крупноплодные, хорошим вкусом плодов.
Ягодные культуры. Ведутся исследования по селекции новых сортов, устойчивых к неблагоприятным факторам среды, высокозимостойких, с повышенной устойчивостью цветков к весенним заморозкам, высокопродуктивных, с отличным качеством плодов. Составлены модели оптимального сорта ягодных культур с учетом технологических запросов селекции 2020-2025 годов. Разработаны технологии возделывания шиповника, крыжовника, жимолости, смородины, технология размножения смородины в пленочных теплицах.
Вернуть былую славу
Реформы, проводимые в аграрном секторе России с начала 90-х годов, оказались разрушительными для садоводства. Как отрасль народного хозяйства оно фактически перестало существовать. Только в Челябинской области доля промышленного садоводства с 65% снизилась до 1%. Почти не осталось плодоносящих насаждений, опытно-производственные плодопитомники практически прекратили свою деятельность, а их продукция удовлетворяет спрос рынка всего на 12-15%.
В области отсутствует целевая программа развития промышленного садоводства, основанная на достоверных материалах инвентаризации насаждений и новых разработках научных учреждений последних лет.
В этом нет вины науки. Ученые Южно-Уральского научно-исследовательского института плодоовощеводства и картофелеводства не раз предлагали сформировать научные основы возрождения отрасли. Однако состояние макроэкономики и чиновничий консерватизм не дают возможности продвинуться вперед. Расчет на то, что коллективное любительское садоводство (т.е. частный сектор) восполнит потери промышленного садоводства, не оправдывается. Да и не может оправдаться. Не смотря на то, что ЮУНИИПОК ежегодно производит более 60 тысяч саженцев плодовых и ягодных культур для населения, частный сектор страдает от недостатка посадочного материала, особенно новых сортов, потребность в котором удовлетворяется на 47-50%. Эту нишу пытаются заполнить мелкие производители саженцев — частники. Но качество их посадочного материала часто не выдерживает никакой критики по всем параметрам.
Расчет на то, что коллективное любительское садоводство (т.е. частный сектор) восполнит потери промышленного садоводства, не оправдывается. Да и не может оправдаться. Не смотря на то, что ЮУНИИПОК ежегодно производит более 60 тысяч саженцев плодовых и ягодных культур для населения, частный сектор страдает от недостатка посадочного материала, особенно новых сортов, потребность в котором удовлетворяется на 47-50%. Эту нишу пытаются заполнить мелкие производители саженцев — частники. Но качество их посадочного материала часто не выдерживает никакой критики по всем параметрам.
Реально ли возродить общественное садоводство в новых экономических условиях?
Вполне, считают южноуральские ученые и селекционеры. Но для этого на государственном и областном уровне, по их мнению, необходимо решить самые насущные вопросы:
С площадей под садами от начала подготовки территорий и до вступления в плодоношение насаждений отменить налог на землю
На возвратной основе выделять капитальные вложения для посадки многолетних насаждений
На промышленных предприятиях области организовать производство специальной техники для садоводства
Необходимы законы по защите отечественного производителя на собственном рынке
Стимулировать малый бизнес для организации переработки продукции садоводства
В системе профтехобразования необходимо организовать подготовку кадров для садоводства
Весьма актуально и объединение научного потенциала уральских ученых-аграрников. Для этих целей необходимо создание Уральского научно-методического центра с размещением его в Челябинске. Польза от такого объединения огромная. Доказательством тому координационный совет по картофелю, созданный в апреле 2000 года по инициативе ГНУ ЮУНИИПОК на общественных началах. За 8 лет совместной работы институты картофелеводства провели мощную мобилизацию генофонда, пополнили коллекции, договорились о комбинациях скрещивания, регулярно обмениваются информацией, исходным и селекционным материалом. Практика координационного совета показала, что эта форма работы чрезвычайно эффективна, жизнеспособна и поэтому заслуживает развития и совершенствования.
Для этих целей необходимо создание Уральского научно-методического центра с размещением его в Челябинске. Польза от такого объединения огромная. Доказательством тому координационный совет по картофелю, созданный в апреле 2000 года по инициативе ГНУ ЮУНИИПОК на общественных началах. За 8 лет совместной работы институты картофелеводства провели мощную мобилизацию генофонда, пополнили коллекции, договорились о комбинациях скрещивания, регулярно обмениваются информацией, исходным и селекционным материалом. Практика координационного совета показала, что эта форма работы чрезвычайно эффективна, жизнеспособна и поэтому заслуживает развития и совершенствования.
Начата работа и по объединению садоводческих учреждений. Подписано многостороннее соглашение между шестью научными учреждениями Уральского и прилегающих к нему регионов — ГНУ ЮУНИИПОК, БашНИИСХ, Удмуртский НИИСХ, Костанайский НИИСХ, Казахский НИИКОХ, Карабалыкская опытная станция.
Аграрная наука сегодня переживает трудные времена, но она жива и готова внести свой вклад в возрождение сельскохозяйственного производства, в том числе и южноуральского промышленного садоводства.
«Сколково» ищет прорывные проекты в агрогенетике
В последние годы сельское хозяйство переживает серьезную трансформацию. Все чаще на полях можно видеть снабженные десятками датчиков комбайны-беспилотники. Они управляются при помощи данных со спутника и передают информацию о состоянии поля и посевов на компьютер фермера
Летающие над полями дроны, ПО для оценки количества подземных вод, сенсоры, которые устанавливаются на сельхозтехнике и в режиме реального времени передают информацию о качестве собираемого урожая. Это лишь некоторые примеры цифровизации сельского хозяйства, которая бурно происходит в развитых странах и в значительно более ограниченных масштабах — в России. Технологическая революция заметна и за пределами полей: автоматика дозирует корм для коров на основе анализа физиологических параметров животных; на птицефабриках инфракрасные камеры отслеживают заболевших птиц.
Это внешние проявления того, как меняется сектор агро. Впечатляют и темпы его внутренней трансформации, связанной с успехами сельскохозяйственных наук, в частности, генетики и селекции. Например, ученые выводят супермускулистых животных с блокированным миостатином, белком, который подавляет рост мышечной ткани. В растениеводстве таких примеров еще больше, рассказывает в беседе с Sk.ru руководитель сельскохозяйственного направления Фонда «Сколково» Роман Куликов. «Мировая статистика такова: на один селекционный проект с животными приходится 10 проектов с растениями. Это естественно – растения служат более существенной кормовой базой для людей и животных, а также платформой для получения многих терапевтических соединений. Если продуктивных сельскохозяйственных животных — около полутора десятков, то культурных растений — сотни, и опытов с ними проводят очень много. Так, в последнее время мировые концерны ведут активную работу по созданию устойчивых к засухе сортов с особым метаболизмом и водным обменом». Адаптация сельскохозяйственных культур к изменению климата крайне актуальна и для России. Те районы, где еще 20 лет назад успешно выращивали зерновые (Волгоградская область, регионы на левом берегу Волги), сегодня страдают от засухи.
Например, ученые выводят супермускулистых животных с блокированным миостатином, белком, который подавляет рост мышечной ткани. В растениеводстве таких примеров еще больше, рассказывает в беседе с Sk.ru руководитель сельскохозяйственного направления Фонда «Сколково» Роман Куликов. «Мировая статистика такова: на один селекционный проект с животными приходится 10 проектов с растениями. Это естественно – растения служат более существенной кормовой базой для людей и животных, а также платформой для получения многих терапевтических соединений. Если продуктивных сельскохозяйственных животных — около полутора десятков, то культурных растений — сотни, и опытов с ними проводят очень много. Так, в последнее время мировые концерны ведут активную работу по созданию устойчивых к засухе сортов с особым метаболизмом и водным обменом». Адаптация сельскохозяйственных культур к изменению климата крайне актуальна и для России. Те районы, где еще 20 лет назад успешно выращивали зерновые (Волгоградская область, регионы на левом берегу Волги), сегодня страдают от засухи. Это большая проблема и для производителей, и для потребителей, и для страховых компаний.
Это большая проблема и для производителей, и для потребителей, и для страховых компаний.
Сельское хозяйство — это отрасль, где люди решают сугубо практические задачи производства определенной продукции. Чтобы производить эффективнее (больше и дешевле), им необходимы новые технологии, которые разрабатываются сторонними компаниями, продолжает Роман. «Тем, кто выращивает пшеницу, нужно взаимодействовать с производителями семян, пестицидов, техники. Специфика российского рынка в том, что крупные агрохолдинги (их в стране около сотни) за время своего активного развития применяли, в основном, западные технологии. Потому что отечественная сельскохозяйственная наука и технологии в начале 1990-х годов — по очевидным причинам — затормозили. Но в новой реальности агрохолдинги заинтересованы в поиске российских аналогов западных технологий, поскольку последние резко подорожали или вовсе стали недоступны. Поэтому сейчас в России повсеместно возникают небольшие компании, готовые продавать крупным игрокам рынка необходимые технологии, причем они дешевле и адаптированы к нашим климатическим условиям».
В СССР приоритетной считали максимальную универсальность сорта, в жертву которой приносили все остальные характеристики, в том числе продуктивность
Проблема в том, что те, кто предлагает и потребляет решения, зачастую не знают о существовании друг друга. Местом встречи может стать «Фонд Сколково», который несколько недель запустил конкурс инновационных проектов в области генетики и селекции сельскохозяйственных культур, животноводства и аквакультуры «Агрогенетика 2016».
«У нас весьма насыщенные и разноплановые форсайты. На их основе мы планируем компоновать в этом году три больших конкурса, чтобы не упустить ни одну из сфер. Первый конкурс, «Агрогенетика-2016», как представляется, станет самой непростой историей, ведь генетика и селекция — это максимально технологичная и наиболее пострадавшая в переходный период в РФ область», — говорит Роман Куликов.
На конкурс принимают проекты по четырем направлениям. Во-первых, это селекция и семеноводство сельскохозяйственных культур, генетические ресурсы и современные методы создания новых сортов и гибридов, технологии семеноводства. Второе направление -селекция в животноводстве, аквакультуре и агропромышленной микробиологии. Третье — биоинформационные технологии в генетике, селекции и диагностике. Четвертое — технологии, способствующие реализации генетического потенциала и продуктивности сельскохозяйственных растений, животных и аквакультуры: защита и питание растений, методы подготовки и технологии улучшения почв, кормление, ветеринария, зоотехния.
Второе направление -селекция в животноводстве, аквакультуре и агропромышленной микробиологии. Третье — биоинформационные технологии в генетике, селекции и диагностике. Четвертое — технологии, способствующие реализации генетического потенциала и продуктивности сельскохозяйственных растений, животных и аквакультуры: защита и питание растений, методы подготовки и технологии улучшения почв, кормление, ветеринария, зоотехния.
«Сколково» проводит конкурс совместно с МФТИ. Физтех очень активен в том, что касается life science, рассказывает Роман Куликов. Так, на базе МФТИ действует биофармкластер «Северный»; институт поддерживает масштабные биологические проекты. «Исследования в МФТИ охватывают широкий круг областей, в том числе, в биомедицине. Некоторые исследователи, прежде специализировавшиеся на фарме, все больше концентрируются на разработке стимуляторов роста и пестицидов для сельского хозяйства. Темы смежные, общие биологические принципы никто не отменял. Кроме того, в МФТИ исторически очень сильная физико-математическая школа, и это важно с учетом того, насколько современная селекция и генетика завязаны на биоинформатике», — объясняет выбор партнера по конкурсу руководитель аграрного направления «Сколково». Все чаще рабочее место селекционеров оказывается не в поле, а в лаборатории, где исследователи высчитывают и моделируют последовательность скрещиваний, необходимую для получения растения или животного с заданными свойствами. Это ускоряет и упрощает процесс выведения новых сортов и пород. Олег Корзинов, руководитель биофармкластера «Северный» на базе МФТИ, так отозвался о решении участвовать в конкурсе «Агрогенетика-2016»: «Современные прорывные технологии в области генетики уже нашли свое применение в диагностике заболеваний. Следующий большой скачок развития человечества будет связан уже с редактированием генома, и первые результаты будут получены в области агротехнологий. Поэтому поиск и поддержка в России конкурентоспособных проектов в области агрогенетики является очень важной стратегической задачей».
Все чаще рабочее место селекционеров оказывается не в поле, а в лаборатории, где исследователи высчитывают и моделируют последовательность скрещиваний, необходимую для получения растения или животного с заданными свойствами. Это ускоряет и упрощает процесс выведения новых сортов и пород. Олег Корзинов, руководитель биофармкластера «Северный» на базе МФТИ, так отозвался о решении участвовать в конкурсе «Агрогенетика-2016»: «Современные прорывные технологии в области генетики уже нашли свое применение в диагностике заболеваний. Следующий большой скачок развития человечества будет связан уже с редактированием генома, и первые результаты будут получены в области агротехнологий. Поэтому поиск и поддержка в России конкурентоспособных проектов в области агрогенетики является очень важной стратегической задачей».
Роман Куликов. Фото: Sk.ru
Инструментарий геномной селекции позволяет значительно ускорить традиционные методы без фактического редактирования генома, что в итоге усилит конкурентные преимущества отечественного сельского хозяйства на мировых рынках. Роман Куликов в беседе с Sk.ru был категоричен: селекция и генетика не несут человечеству зла, напротив, они ускоряют природные, естественные процессы. «Эта технология позволяет быстрее отбирать и адаптировать признаки, чтобы успеть за изменениями климата и диетических предпочтений потребителей, а также появлением новых вредителей и пестицидов. Большинство животных и растений, которые мы употребляем в пищу, и так являются результатами селекции. У яблок, которые мы едим, уже много сотен лет «новый» генотип, отличающий их от дичков. Дикий арбуз внешне больше похож на тыкву. С помощью достижений сельскохозяйственной науки мы ускоряем селекционный процесс так, что вместо десятка лет он занимает 3-5 лет».
Роман Куликов в беседе с Sk.ru был категоричен: селекция и генетика не несут человечеству зла, напротив, они ускоряют природные, естественные процессы. «Эта технология позволяет быстрее отбирать и адаптировать признаки, чтобы успеть за изменениями климата и диетических предпочтений потребителей, а также появлением новых вредителей и пестицидов. Большинство животных и растений, которые мы употребляем в пищу, и так являются результатами селекции. У яблок, которые мы едим, уже много сотен лет «новый» генотип, отличающий их от дичков. Дикий арбуз внешне больше похож на тыкву. С помощью достижений сельскохозяйственной науки мы ускоряем селекционный процесс так, что вместо десятка лет он занимает 3-5 лет».
Все чаще рабочее место селекционеров оказывается не в поле, а в лаборатории, где исследователи высчитывают и моделируют последовательность скрещиваний, необходимую для получения растения или животного с заданными свойствами
В животноводстве на первый план тоже выходят биоинформатика и молекулярные технологии. Страны, добившиеся высоких показателей по молочному стаду (Израиль, США, Канада, некоторые европейские государства) реализуют национальные программы по мониторингу поголовья. Животные при рождении чипируются, информация об их болезнях, надоях и т.д. накапливается в базе данных и соотносится с генетикой. Компиляция этих данных дает селекционерам мощнейший инструмент, позволяющий просчитать и смоделировать, скрещивание каких животных приведет к появлению потомства с требуемыми характеристиками. В России, рассказывает Роман Куликов, подобных проектов мало.
Страны, добившиеся высоких показателей по молочному стаду (Израиль, США, Канада, некоторые европейские государства) реализуют национальные программы по мониторингу поголовья. Животные при рождении чипируются, информация об их болезнях, надоях и т.д. накапливается в базе данных и соотносится с генетикой. Компиляция этих данных дает селекционерам мощнейший инструмент, позволяющий просчитать и смоделировать, скрещивание каких животных приведет к появлению потомства с требуемыми характеристиками. В России, рассказывает Роман Куликов, подобных проектов мало.
Найти их – цель конкурса «Агрогенетика». Его организаторы надеются, что разрозненные научные команды, которые занимаются селекцией, получат шанс наладить взаимодействие как друг с другом, так и с бизнесом. «В институтах реализуют проекты молекулярных маркеров на свиньях, крупном рогатом скоте, некоторых растениях. Есть программа, нацеленная на создание отечественного племенного и посевного фонда. По сути это реанимация советской, а позже и российской генетики и селекции. Большинство этих разработок принадлежат научно-исследовательским институтам, — говорит Роман Куликов. — Наша задача в том, чтобы они узнали друг о друге и о Фонде «Сколково», структуре, которая может подставить им плечо, сопровождать и развивать их наработки».
Большинство этих разработок принадлежат научно-исследовательским институтам, — говорит Роман Куликов. — Наша задача в том, чтобы они узнали друг о друге и о Фонде «Сколково», структуре, которая может подставить им плечо, сопровождать и развивать их наработки».
В этом году под эгидой ФАНО стартовал прорывной проект, цель которого – ревизия отечественных сортов и проверка их соответствия современным вызовам рынка. Если у этих сортов есть перспектива, то для их коммерциализации в новых экономических реалиях потребуется большая работа: менторская помощь, поиск индустриальных партнеров, анализ рынка. По словам Р. Куликова, «чтобы привлечь в проект инвестиции, превратить его в коммерчески привлекательный продукт, который захотят продавать семеноводческие компании, его нужно как следует упаковать. Это огромная работа, и «Сколково» — тот институт, который может помочь на всех этапах».
«У нас по многим культурам дефицит конкурентоспособных гибридов — на рынке присутствуют устаревшие сорта, которым 10-15 лет»
При этом, уверят Надежда Долматова, проектный менеджер направления «Биотехнологии в сельском хозяйстве», существует очевидный запрос на качественные селекционные разработки со стороны крупных сельскохозяйственных холдингов. Опытами в селекции и генетике заняты на крупных мясоперерабатывающих предприятиях.
Опытами в селекции и генетике заняты на крупных мясоперерабатывающих предприятиях.
Плоды усилий российских селекционеров не полностью устраивают бизнес по причине их не слишком высокого соответствия требованиям рынка. «В институтах вроде бы есть понимание, что нужны засухоустойчивые сорта. Но там не задаются вопросом о том, как соблюсти правильный экономический баланс, выводя растение с требуемыми признаками. В результате у нас по многим культурам дефицит конкурентоспособных гибридов, на рынке присутствуют устаревшие и далеко не оптимальные сорта, которым 10-15 лет. А ведь селекционер должен очень чутко прислушиваться к запросам рынка. Залог успеха именно в этом», — обращает внимание Надежда Долматова.
О той же проблеме на проходящей в Сколково Летней школе ОтУС, посвященной агротехнологиям, говорила и Екатерина Журавлева, помощник руководителя Федерального агентства научных организаций. Хотя селекция в России имеет долгую историю, современные методы в ней используются мало, а генотипы используемых сортов в большинстве случаев не расшифрованы. Генетические исследования в области растениеводства помогут улучшить процесс отбора нужных культур и продемонстрировать конкурентоспособность российских сортов. Например, сейчас из 10 главных возделываемых сортов картофеля всего 3 отечественных, причем не на лидирующих позициях.
Генетические исследования в области растениеводства помогут улучшить процесс отбора нужных культур и продемонстрировать конкурентоспособность российских сортов. Например, сейчас из 10 главных возделываемых сортов картофеля всего 3 отечественных, причем не на лидирующих позициях.
Отчасти это следствие советской модели развития селекции и генетики. В СССР приоритетной считали максимальную универсальность сорта, в жертву которой приносили все остальные характеристики, в том числе продуктивность. На задний план отошли те свойства, которые отвечают технологичность, содержание питательных веществ, сроки хранения и так далее, рассказывает Роман Куликов. «Западная селекция уже тогда начала фокусироваться именно на технологичности. Разработчики исходили из того, что сорт должен быть максимально интегрирован в технологию, и создавали коробочное технологическое решение, например, в случае с сельскохозяйственными культурами – какой техникой и каких диапазонах работать, на какую глубину засевать, какие удобрения использовать».
животноводство | Britannica
Селекция и вариация
Английский агроном Роберт Бейкуэлл был очень успешным животноводом в 18 веке. Его работа была основана на традиционном методе визуальной оценки выбранных им животных. Хотя он не писал о своих методах, известно, что он много путешествовал верхом и собирал овец и крупный рогатый скот, которые считал полезными. Считается, что он проводил широкомасштабные ауткроссы различных пород, а затем практиковал инбридинг с целью закрепления желаемых характеристик у скрещенных животных.Он также был первым, кто систематически отдавал своих животных на вязку. По этим причинам он общепризнан как первый научный селекционер.
В животноводстве популяция представляет собой группу скрещивающихся особей, т. е. породу или породу внутри породы, которая в некоторых аспектах отличается от других пород или линий. Как правило, некоторые животные внутри породы считаются чистокровными. Существенное различие между чистокровными и нечистокровными животными заключается в том, что генеалогия чистокровных животных тщательно записывается, обычно в племенной книге или племенной книге, которую ведет какая-либо санкционирующая ассоциация. Чистокровные ассоциации предоставляют другие услуги, которые полезны их членам для развития их бизнеса.
Чистокровные ассоциации предоставляют другие услуги, которые полезны их членам для развития их бизнеса.
Селекционное разведение использует естественные различия в признаках, которые существуют среди членов любой популяции. Прогресс в разведении требует понимания двух источников изменчивости: генетики и окружающей среды. Для некоторых признаков существует взаимодействие генетики и окружающей среды. Различия в окружающей среде животных, такие как количество корма, уход и даже погода, могут влиять на их рост, размножение и продуктивность.Такие изменения производительности из-за окружающей среды не передаются следующему поколению. Для большинства признаков, измеренных у домашних животных, окружающая среда оказывает большее влияние на изменчивость, чем генетические различия. Например, только около 30% различий в молочной продуктивности молочного скота можно объяснить генетическими эффектами; остальная часть вариации связана с воздействием окружающей среды. Таким образом, факторы окружающей среды должны учитываться и контролироваться при выборе племенного поголовья.
Генетическая изменчивость необходима для того, чтобы добиться прогресса в разведении последующих поколений. Каждый ген, который является основной единицей наследственности, занимает определенное место или локус на хромосоме. Два или более генов могут быть связаны с конкретным локусом и, следовательно, с определенным признаком. (Признаки, которые можно непосредственно наблюдать, такие как размер, цвет, форма и т. д., составляют фенотип организма.) Эти гены известны как аллели. Если парные аллели одинаковы, организм называется гомозиготным по этому признаку; если они разные, организм гетерозиготен.Как правило, один из аллелей будет экспрессироваться с исключением другого аллеля, и в этом случае два аллеля называются доминантным и рецессивным соответственно. Однако иногда ни один из них не доминирует, и в этом случае два аллеля называются кодоминантными.
Мышь внизу гетерозиготна с мутантным геном, придающим ей пятнистый хвост. Две мыши над ним парамутированы; у них также есть пятнистые хвосты, хотя они не несут гена этой черты.
Хотя полных знаний о генетическом составе любой породы домашнего скота пока не существует, генетические вариации можно использовать для улучшения поголовья.Исследователи разделяют общую генетическую изменчивость на аддитивные, доминантные и эпистатические типы действия генов, которые определены в следующих параграфах. Аддитивную изменчивость легче всего использовать в селекции, потому что она распространена, и эффект каждого аллеля в локусе просто добавляется к эффекту других аллелей в том же локусе. Генетические улучшения, полученные с помощью аддитивных генетических эффектов, являются постоянными и накапливаются от одного поколения к другому.
Хотя изменчивость доминирования в теории не является более сложной, ее сложнее контролировать на практике из-за того, что один аллель маскирует эффект другого.Например, пусть и обозначают локус, где и 1 и и 2 представляют два возможных аллеля в этом месте. Тогда A 1 A 1 , A 1 A 2 (который идентичен A 2 A 1 ), а также A 2 a 2 — три возможных генотипа. Если A 1 доминирует на A 2 , генотипы 2 1 A 2 и 9002 2 и A 1 A 1 нельзя отличить внешность.Таким образом, неспособность наблюдать различия между , , , 1, , , , , 2, и , , , 1, , , , , 1, представляет большую трудность в использовании доминирования.
Тогда A 1 A 1 , A 1 A 2 (который идентичен A 2 A 1 ), а также A 2 a 2 — три возможных генотипа. Если A 1 доминирует на A 2 , генотипы 2 1 A 2 и 9002 2 и A 1 A 1 нельзя отличить внешность.Таким образом, неспособность наблюдать различия между , , , 1, , , , , 2, и , , , 1, , , , , 1, представляет большую трудность в использовании доминирования.
Аддитивные и доминантные вариации вызываются генами в одном локусе. Эпистатическая изменчивость вызывается совместным действием генов в двух или более локусах. Преднамеренного использования этого типа генетической изменчивости в разведении было мало из-за сложной природы идентификации и контроля соответствующих генов.
животноводство | Britannica
Селекция и вариация
Английский агроном Роберт Бейкуэлл был очень успешным животноводом в 18 веке. Его работа была основана на традиционном методе визуальной оценки выбранных им животных. Хотя он не писал о своих методах, известно, что он много путешествовал верхом и собирал овец и крупный рогатый скот, которые считал полезными. Считается, что он проводил широкомасштабные ауткроссы различных пород, а затем практиковал инбридинг с целью закрепления желаемых характеристик у скрещенных животных.Он также был первым, кто систематически отдавал своих животных на вязку. По этим причинам он общепризнан как первый научный селекционер.
В животноводстве популяция представляет собой группу скрещивающихся особей, т. е. породу или породу внутри породы, которая в некоторых аспектах отличается от других пород или линий. Как правило, некоторые животные внутри породы считаются чистокровными. Существенное различие между чистокровными и нечистокровными животными заключается в том, что генеалогия чистокровных животных тщательно записывается, обычно в племенной книге или племенной книге, которую ведет какая-либо санкционирующая ассоциация. Чистокровные ассоциации предоставляют другие услуги, которые полезны их членам для развития их бизнеса.
Чистокровные ассоциации предоставляют другие услуги, которые полезны их членам для развития их бизнеса.
Селекционное разведение использует естественные различия в признаках, которые существуют среди членов любой популяции. Прогресс в разведении требует понимания двух источников изменчивости: генетики и окружающей среды. Для некоторых признаков существует взаимодействие генетики и окружающей среды. Различия в окружающей среде животных, такие как количество корма, уход и даже погода, могут влиять на их рост, размножение и продуктивность.Такие изменения производительности из-за окружающей среды не передаются следующему поколению. Для большинства признаков, измеренных у домашних животных, окружающая среда оказывает большее влияние на изменчивость, чем генетические различия. Например, только около 30% различий в молочной продуктивности молочного скота можно объяснить генетическими эффектами; остальная часть вариации связана с воздействием окружающей среды. Таким образом, факторы окружающей среды должны учитываться и контролироваться при выборе племенного поголовья.
Генетическая изменчивость необходима для того, чтобы добиться прогресса в разведении последующих поколений. Каждый ген, который является основной единицей наследственности, занимает определенное место или локус на хромосоме. Два или более генов могут быть связаны с конкретным локусом и, следовательно, с определенным признаком. (Признаки, которые можно непосредственно наблюдать, такие как размер, цвет, форма и т. д., составляют фенотип организма.) Эти гены известны как аллели. Если парные аллели одинаковы, организм называется гомозиготным по этому признаку; если они разные, организм гетерозиготен.Как правило, один из аллелей будет экспрессироваться с исключением другого аллеля, и в этом случае два аллеля называются доминантным и рецессивным соответственно. Однако иногда ни один из них не доминирует, и в этом случае два аллеля называются кодоминантными.
Мышь внизу гетерозиготна с мутантным геном, придающим ей пятнистый хвост. Две мыши над ним парамутированы; у них также есть пятнистые хвосты, хотя они не несут гена этой черты.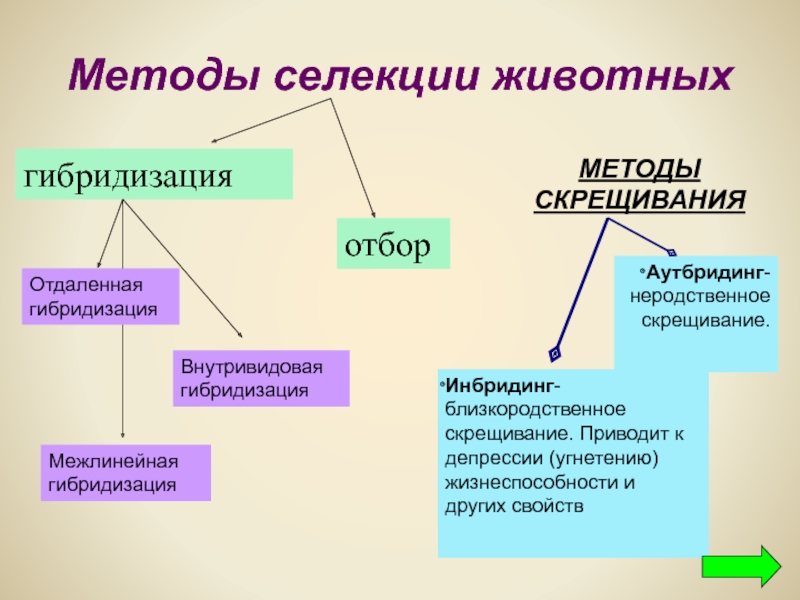
Хотя полных знаний о генетическом составе любой породы домашнего скота пока не существует, генетические вариации можно использовать для улучшения поголовья.Исследователи разделяют общую генетическую изменчивость на аддитивные, доминантные и эпистатические типы действия генов, которые определены в следующих параграфах. Аддитивную изменчивость легче всего использовать в селекции, потому что она распространена, и эффект каждого аллеля в локусе просто добавляется к эффекту других аллелей в том же локусе. Генетические улучшения, полученные с помощью аддитивных генетических эффектов, являются постоянными и накапливаются от одного поколения к другому.
Хотя изменчивость доминирования в теории не является более сложной, ее сложнее контролировать на практике из-за того, что один аллель маскирует эффект другого.Например, пусть и обозначают локус, где и 1 и и 2 представляют два возможных аллеля в этом месте. Тогда A 1 A 1 , A 1 A 2 (который идентичен A 2 A 1 ), а также A 2 a 2 — три возможных генотипа. Если A 1 доминирует на A 2 , генотипы 2 1 A 2 и 9002 2 и A 1 A 1 нельзя отличить внешность.Таким образом, неспособность наблюдать различия между , , , 1, , , , , 2, и , , , 1, , , , , 1, представляет большую трудность в использовании доминирования.
Тогда A 1 A 1 , A 1 A 2 (который идентичен A 2 A 1 ), а также A 2 a 2 — три возможных генотипа. Если A 1 доминирует на A 2 , генотипы 2 1 A 2 и 9002 2 и A 1 A 1 нельзя отличить внешность.Таким образом, неспособность наблюдать различия между , , , 1, , , , , 2, и , , , 1, , , , , 1, представляет большую трудность в использовании доминирования.
Аддитивные и доминантные вариации вызываются генами в одном локусе. Эпистатическая изменчивость вызывается совместным действием генов в двух или более локусах. Преднамеренного использования этого типа генетической изменчивости в разведении было мало из-за сложной природы идентификации и контроля соответствующих генов.
Селективное разведение | Биониндзя
Понимание:
• Селекционное разведение домашних животных показывает, что искусственный отбор может вызвать эволюцию
Селективное разведение является формой искусственного отбора , при котором человек вмешивается в разведение видов для получения желаемых признаков у потомства
- Путем размножения представителей вида с желаемым признаком частота этого признака становится более распространенной в последующих поколениях
- Селекционное разведение свидетельствует об эволюции, поскольку целевые породы могут демонстрировать значительные вариации за (относительно) короткий период
Селекционное разведение растительных культур позволило получить новые виды пищевых продуктов из одного и того же исходного растительного источника
- Растения рода Brassica были выведены для производства различных пищевых продуктов путем модификации частей растения путем искусственного отбора
- Сюда входят брокколи (модифицированные цветочные бутоны), белокочанная капуста (модифицированные листовые бутоны) и капуста (модифицированные листья)
Селекционное разведение одомашненных животных также привело к появлению d разные породы потомства
- Примеры селекционного разведения домашних животных можно увидеть у лошадей, коров и собак
Пример 1: Коневодство
Лошадей селекционно разводили на протяжении многих поколений для получения вариаций в соответствии со многими поколениями целевая функция
- Скаковые лошади были выведены для скорости и, следовательно, обычно стройнее, легче, выше и быстрее
- Тяжелые лошади были выведены для силы и выносливости и, следовательно, они более крепкие и коренастые
Пример 2: Разведение коров
Коров селекционно разводили на протяжении многих поколений, чтобы получить потомство с улучшенной молочной продуктивностью
Фермеры также нацелились на разведение коров с мутацией, приводящей к увеличению мышечной массы ) имеют чрезмерную массу и производят более съедобное нежирное мясо
Пример 3: Собаководство
Породы собак демонстрируют огромное разнообразие благодаря целенаправленному отбору определенных признаков человеком
- Охотничьи собаки (например,г.
 биглей) обычно разводили, чтобы они были меньше ростом, чтобы проникать в лисьи норы
биглей) обычно разводили, чтобы они были меньше ростом, чтобы проникать в лисьи норы - Пастушьи собаки (например, овчарки) были выведены для повышенного интеллекта, чтобы следовать пастушьим командам
- Гоночные собаки (например, борзые) были специально выведены, чтобы быть гладкими и быстрые
- Игрушечные собаки (например, чихуахуа) были селекционно выведены из-за их миниатюрных размеров
Вариации пород одомашненных собак
Животноводы — обзор
Естественный отбор
Дарвин был впечатлен успехом селекционеров растений и животных в получении с помощью искусственного отбора новых сортов с признаками, которые считались желательными.Это включает в себя отбор особей, демонстрирующих требуемые черты в большей степени, чем другие особи, и размножение от них, причем этот процесс повторяется на протяжении многих поколений. Таким путем получают устойчивые к болезням культурные растения путем селекции наиболее устойчивых растений.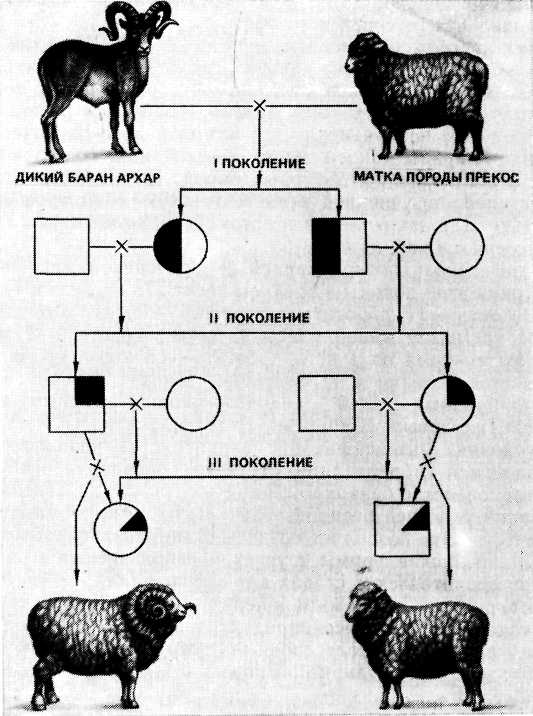 Дарвин также понял, что в естественных популяциях многие особи не доживают до размножения. Он предположил, что любые признаки, способствующие выживанию, будут естественным отбором и имеют тенденцию распространяться в популяции, что в конечном итоге приводит к эволюционным изменениям.Естественный отбор благоприятных вариаций в настоящее время общепризнан как главная и, вероятно, единственная основа для приобретения признаков, в результате которых популяция становится лучше приспособленной к окружающей среде, в процессе адаптивной эволюции .
Дарвин также понял, что в естественных популяциях многие особи не доживают до размножения. Он предположил, что любые признаки, способствующие выживанию, будут естественным отбором и имеют тенденцию распространяться в популяции, что в конечном итоге приводит к эволюционным изменениям.Естественный отбор благоприятных вариаций в настоящее время общепризнан как главная и, вероятно, единственная основа для приобретения признаков, в результате которых популяция становится лучше приспособленной к окружающей среде, в процессе адаптивной эволюции .
Естественный отбор в значительной степени ответственен за адаптивные генетические изменения в популяции, включая стабилизирующие, направленные (при которых изменчивость уменьшается) и разрушительные изменения (при которых изменчивость увеличивается) (рис. 4.10). Чаще всего это консервативная сила, устраняющая девиантов и поддерживающая статус-кво. Это связано с тем, что популяция обычно хорошо адаптирована к окружающей среде, в которой она встречается, и генетические изменения, будь то в результате мутации, рекомбинации или потока генов, с большей вероятностью приведут к снижению, а не к увеличению приспособленности. Естественный отбор, действующий на устранение вариантов, заметно отличающихся от среднего, называется стабилизирующим отбором . Например, гетерокариотические изоляты базидиомицета Schizophyllum commune , полученные из плодовых тел и, таким образом, представляющие мицелий, переживший естественный отбор, имеют ограниченный диапазон скоростей радиального удлинения.Гетерокарионы, полученные в лаборатории путем спаривания гомокарионов, имеют гораздо более широкий диапазон скоростей удлинения, но примерно с таким же средним значением. Таким образом, в естественной популяции особи с темпами расширения намного ниже или намного выше среднего были элиминированы стабилизирующим отбором. Гибридизация и искусственный отбор могут давать варианты, сильно отличающиеся от форм, встречающихся в природе, что указывает на широкое действие стабилизирующего отбора.
Естественный отбор, действующий на устранение вариантов, заметно отличающихся от среднего, называется стабилизирующим отбором . Например, гетерокариотические изоляты базидиомицета Schizophyllum commune , полученные из плодовых тел и, таким образом, представляющие мицелий, переживший естественный отбор, имеют ограниченный диапазон скоростей радиального удлинения.Гетерокарионы, полученные в лаборатории путем спаривания гомокарионов, имеют гораздо более широкий диапазон скоростей удлинения, но примерно с таким же средним значением. Таким образом, в естественной популяции особи с темпами расширения намного ниже или намного выше среднего были элиминированы стабилизирующим отбором. Гибридизация и искусственный отбор могут давать варианты, сильно отличающиеся от форм, встречающихся в природе, что указывает на широкое действие стабилизирующего отбора.
Рисунок 4.10. Естественный отбор.Отбор может привести к (а, б) стабилизации частот аллелей в популяциях или к (в) направленным или (г) разрушительным изменениям. Верхний ряд указывает начальное распределение аллелей, а заштрихованные области представляют предпочтительные аллели. Нижний ряд показывает распределение частот аллелей после одного или нескольких поколений отбора. (e) Направленная селекция по потере чувствительности популяций Rhynchosporium secalis на ячмене в Великобритании к фунгициду пропиконазолу. Средняя минимальная ингибирующая концентрация увеличилась примерно в 10 раз за 3 года.
Верхний ряд указывает начальное распределение аллелей, а заштрихованные области представляют предпочтительные аллели. Нижний ряд показывает распределение частот аллелей после одного или нескольких поколений отбора. (e) Направленная селекция по потере чувствительности популяций Rhynchosporium secalis на ячмене в Великобритании к фунгициду пропиконазолу. Средняя минимальная ингибирующая концентрация увеличилась примерно в 10 раз за 3 года.
Естественный отбор также может вызывать изменения, называемые направленным отбором , как в отношении признаков, демонстрирующих непрерывную изменчивость, так и в отношении признаков, определяемых отдельными генами. Направленный отбор, действующий на непрерывно изменяющийся признак, будет благоприятствовать особям с одной крайности (рис. 4.10). Воздействуя на признаки, определяемые отдельными генами, он будет либо уменьшать, либо увеличивать частоту аллеля, в крайних случаях приводя к исчезновению аллеля или вызывая его «фиксацию», полностью заменяя альтернативные аллели. Появление в течение нескольких десятилетий нескольких патотипов в австралийской популяции Puccinia graminis f. сп. tritici клонального происхождения является примером происходящей направленной селекции. Давление мутаций не могло не повлиять на распространение мутантов патотипа за столь короткое время. При мутуализме высших термитов/ Termitomyces термиты питаются богатыми питательными веществами грибковыми конкрециями – незрелыми плодовыми телами, содержащими бесполые споры, некоторые из которых выживают при прохождении через кишечник и действуют как инокуляты, когда в гнездо вносится новая пища (стр.332). Поскольку только несколько спор успешно образуют мицелий, это приводит к уменьшению изменчивости внутри гнезда. Поскольку генотипы, образующие много конкреций, с наибольшей вероятностью будут инокулированы на свежее органическое вещество, термиты эффективно отбираются для образования большого количества конкреций. Если возникает мутант, который производит мало клубеньков, он будет недопредставлен в инокуляции свежего органического вещества и, следовательно, будет «автоматически» отобран против него.
Появление в течение нескольких десятилетий нескольких патотипов в австралийской популяции Puccinia graminis f. сп. tritici клонального происхождения является примером происходящей направленной селекции. Давление мутаций не могло не повлиять на распространение мутантов патотипа за столь короткое время. При мутуализме высших термитов/ Termitomyces термиты питаются богатыми питательными веществами грибковыми конкрециями – незрелыми плодовыми телами, содержащими бесполые споры, некоторые из которых выживают при прохождении через кишечник и действуют как инокуляты, когда в гнездо вносится новая пища (стр.332). Поскольку только несколько спор успешно образуют мицелий, это приводит к уменьшению изменчивости внутри гнезда. Поскольку генотипы, образующие много конкреций, с наибольшей вероятностью будут инокулированы на свежее органическое вещество, термиты эффективно отбираются для образования большого количества конкреций. Если возникает мутант, который производит мало клубеньков, он будет недопредставлен в инокуляции свежего органического вещества и, следовательно, будет «автоматически» отобран против него.
Подрывной (или диверсифицирующий ) отбор благоприятствует особям на двух крайних точках спектра изменчивости и действует против промежуточных форм.Это происходит, когда есть две возможные среды для эксплуатации, и успех достигается за счет высокой эффективности в одной, а не умеренной эффективности в обеих (рис. 4.10). Он объясняет происхождение внутри патогенного вида специализированных для разных хозяев форм, например Puccinia graminis f. сп. tritici , поражающий пшеницу, и Puccinia graminis f. сп. avenae , поражающий овес. Эти две формы все еще могут гибридизоваться и сохранять ограниченную способность заражать хозяина другой формы, что предполагает относительно недавнее происхождение в результате разрушительного отбора от менее специализированного общего предка.
Хотя естественный отбор может привести аллель к фиксации или исчезновению, он может поддерживать баланс между двумя или более аллелями, встречающимися с довольно высокой частотой. Действующий таким образом естественный отбор называется уравновешивающим отбором . Это может произойти, когда в диплоидной или дикариотической фазе гетерозигота более приспособлена, чем любая гомозигота (преимущество гетерозиготы – стр. 110). Балансирующий отбор может также принимать форму частотно-зависимого отбора , при котором отбор увеличивает частоту аллеля, если он встречается редко, и снижает его частоту, если он распространен.Например, аллель редкого типа спаривания будет увеличиваться по частоте, поскольку люди, обладающие им, смогут спариваться с большинством других людей. Однако, если аллель становится обильной, то особи, обладающие им, с большой вероятностью столкнутся с особями идентичного типа спаривания, что приведет к неудаче спаривания. Отбор, зависящий от частоты, также будет иметь место при отношении ген к гену, которое определяет патогенность (стр. 256). Редкий аллель вирулентности будет увеличиваться по частоте, так как будет мало отбора на устойчивость хозяина.
Действующий таким образом естественный отбор называется уравновешивающим отбором . Это может произойти, когда в диплоидной или дикариотической фазе гетерозигота более приспособлена, чем любая гомозигота (преимущество гетерозиготы – стр. 110). Балансирующий отбор может также принимать форму частотно-зависимого отбора , при котором отбор увеличивает частоту аллеля, если он встречается редко, и снижает его частоту, если он распространен.Например, аллель редкого типа спаривания будет увеличиваться по частоте, поскольку люди, обладающие им, смогут спариваться с большинством других людей. Однако, если аллель становится обильной, то особи, обладающие им, с большой вероятностью столкнутся с особями идентичного типа спаривания, что приведет к неудаче спаривания. Отбор, зависящий от частоты, также будет иметь место при отношении ген к гену, которое определяет патогенность (стр. 256). Редкий аллель вирулентности будет увеличиваться по частоте, так как будет мало отбора на устойчивость хозяина. Если, однако, он станет массовым, то соответствующий ген устойчивости хозяина также станет массовым, и отбор будет действовать против особей, несущих теперь уже почти бесполезный аллель вирулентности.
Если, однако, он станет массовым, то соответствующий ген устойчивости хозяина также станет массовым, и отбор будет действовать против особей, несущих теперь уже почти бесполезный аллель вирулентности.
Естественный отбор может действовать по-разному у гаплоидных и диплоидных/гетерокариотичных грибов. У гаплоидных грибов ген будет экспрессироваться либо на протяжении всего жизненного цикла, либо на какой-то его стадии, и естественный отбор может влиять на частоту его аллелей. У диплоидных грибов рецессивный аллель не будет влиять на фенотип и, таким образом, будет частично защищен от естественного отбора; если он вреден, его частота будет уменьшаться медленнее, чем если бы он частично или полностью доминировал или присутствовал в гаплоидном грибе.Его исчезновение, вероятно, будет отсрочено надолго, поскольку при достижении низких частот он редко будет присутствовать в гомозиготном состоянии, в котором он подвергался бы отбору. У гетерокариотических базидиомицетов аргументы в пользу гаплоидных грибов относятся к фазе гомокариона, а аргументы в пользу диплоидных грибов относятся к фазе гетерокариона. Следовательно, грибы, которые являются диплоидными или гетерокариотическими на протяжении большей части своего жизненного цикла, вероятно, несут много вредных в настоящее время рецессивных аллелей, которые в другом геноме или среде могут оказаться полезными.
Следовательно, грибы, которые являются диплоидными или гетерокариотическими на протяжении большей части своего жизненного цикла, вероятно, несут много вредных в настоящее время рецессивных аллелей, которые в другом геноме или среде могут оказаться полезными.
Границы | Возможности повышения устойчивости в программах разведения животных
Введение
Современное животноводство характеризуется интенсификацией, т. е. увеличением поголовья на ферму. Для успешной интенсификации животноводческого производства без негативных последствий для животных, фермеров и ферм необходимо выполнение определенных требований. Одним из таких требований интенсификации животноводческого производства является способность фермера заботиться о большем количестве животных.Для этого требуются здоровые и простые в уходе животные, которым требуется мало/меньше времени на внимание (Elgersma et al., 2018). Устойчивые животные — это животные, которым требуется мало/меньше времени на внимание: поэтому желательно повысить устойчивость. Повышение устойчивости может быть реализовано с помощью различных стратегий. Одна из стратегий заключается в повышении устойчивости путем генетического отбора в программах разведения. Преимущество генетической селекции, по сравнению с улучшением управления, заключается в том, что она носит кумулятивный характер и влияет на все последующие поколения скота.
Повышение устойчивости может быть реализовано с помощью различных стратегий. Одна из стратегий заключается в повышении устойчивости путем генетического отбора в программах разведения. Преимущество генетической селекции, по сравнению с улучшением управления, заключается в том, что она носит кумулятивный характер и влияет на все последующие поколения скота.
Мы определяем устойчивость как «способность животного подвергаться минимальному влиянию беспокойства или быстро возвращаться в состояние, в котором оно было до воздействия беспокойства» (скорректировано из Colditz and Hine, 2016). В литературе обсуждалось несколько определений устойчивости (например, Colditz and Hine, 2016) и концепций, связанных с устойчивостью: надежность, толерантность, устойчивость, взаимодействие GxE, генетическая гетерогенность изменчивости окружающей среды, пластичность, чувствительность к окружающей среде, канализация (развитие). стабильность и остаточная внутрииндивидуальная фенотипическая изменчивость (т. г., Холлинг, 1973; Дебат и Дэвид, 2001 г .; Де Йонг и Биджма, 2002 г.; Флэтт, 2005; Кнап, 2005; Малдер и др., 2007, 2013; Бишоп, 2012 г.; Вестнит и др., 2015; Кольдиц и Хайн, 2016 г.; Марьянович и др., 2018). Мы обобщили эти определения и концепции во вставке 1, но обсуждение различий и сходств между ними выходит за рамки данной статьи.
г., Холлинг, 1973; Дебат и Дэвид, 2001 г .; Де Йонг и Биджма, 2002 г.; Флэтт, 2005; Кнап, 2005; Малдер и др., 2007, 2013; Бишоп, 2012 г.; Вестнит и др., 2015; Кольдиц и Хайн, 2016 г.; Марьянович и др., 2018). Мы обобщили эти определения и концепции во вставке 1, но обсуждение различий и сходств между ними выходит за рамки данной статьи.
Вставка 1. Определения устойчивости и связанных с устойчивостью понятий, обсуждаемых в литературе (цитируется). Обратите внимание, что эта вставка служит не критическим отражением литературы, а обзором интерпретации различных концепций .
Устойчивость : способность животного подвергаться минимальному влиянию беспокойства или быстро возвращаться в состояние, в котором оно находилось до воздействия беспокойства (с поправкой на Colditz and Hine, 2016; этот документ).
Устойчивость : животные, которые сочетают высокий производственный потенциал с устойчивостью к внешним стрессорам, что позволяет беспроблемно проявлять высокий производственный потенциал в самых разных условиях окружающей среды (скорректировано из Knap, 2005).
Допуск : чистое влияние на производительность заданного уровня помех (с поправкой на Bishop, 2012).
Устойчивость : способность животного-хозяина контролировать нарушение (с поправкой на Bishop, 2012).
Взаимодействие GxE : лучший генотип в одной среде может не быть лучшим генотипом в другой среде, и эти генотипы различаются по своей реакции на факторы окружающей среды (Mulder et al., 2013).
Чувствительность к окружающей среде : потеря гибкости при работе в интенсивных или ограничивающих условиях из-за несбалансированного распределения ресурсов (Knap, 2005).
Наследуемая изменчивость в изменчивости окружающей среды или наследственная изменчивость : изменчивость значений признаков генотипа, измеренная либо повторно на одном и том же индивидууме, либо на нескольких индивидуумах, принадлежащих к одной семье (Marjanovic et al., 2018).
(фенотипическая) пластичность : способность организма изменять свою физиологию, морфологию или развитие в ответ на изменения в окружающей среде (Debat and David, 2001).
Макросредовая чувствительность: свойство организмов одного генотипа систематически развивать разные фенотипы в разных средах = норма реакции (Debat and David, 2001).
(Генетическая) канализация : сходство проявления фенотипического признака при разных условиях развития; процесс, при котором фенотипическая изменчивость снижается механизмами развития или фенотип генотипа остается относительно инвариантным, когда особи одного и того же одно- или многолокусного генотипа различаются по своему генетическому фону; особый вид эпистаза (Debat and David, 2001; Flatt, 2005).
(Развитие) стабильность : способность системы возвращаться в равновесное состояние после временного нарушения (Holling, 1973).
Остаточная внутрииндивидуальная фенотипическая вариация : величина внутрииндивидуальной дисперсии, не объясняемая конкретной статистической моделью (т. е. средний квадрат отклонений наблюдений от нормы индивидуальной реакции), усредненный по выборке индивидуумов (Westneat et al. , 2015).
, 2015).
Нарушения могут быть разной природы: физическими (например, болезнь, температурный стресс) или психологическими (например, новая среда, социальный стрессор, человеческое взаимодействие) (см. обзор Colditz and Hine, 2016).Таким образом, «общая» устойчивость является составным признаком, состоящим из различных типов устойчивости в зависимости от характера нарушения (Colditz and Hine, 2016; Elgersma et al., 2018). Возмущения можно классифицировать как «факторы макросреды» или «факторы микросреды» (Falconer and Mackay, 1996; Mulder et al., 2013). Факторы макроокружающей среды являются характеристиками окружающей среды и, таким образом, влияют на (большинство) всего населения в этой макросреде (например, распространенность болезней, температура окружающей среды).Генетическая изменчивость реакции на эти факторы макросреды может быть выражена как генетическая дисперсия наклона нормы реакции в разных средах или при разных количествах возмущения (Mulder et al., 2013). Факторы микросреды возникают в макросреде и, таким образом, влияют только на меньшинство всего населения в этой макросреде (например, болезни, социальные взаимодействия). Генетические различия в реакции на факторы микроокружения могут быть выражены как генетическая дисперсия в размере изменчивости окружающей среды (Mulder et al., 2013). Несмотря на то, что макро- и микроэкологическая чувствительность относятся к разным понятиям и лучше всего исследуются разными методами (Mulder et al., 2015a), расчетная генетическая корреляция между ними оказалась высокой (0,76 при SE = 0,10; Mulder et al. , 2013). Это указывает на то, что, несмотря на то, что нарушения имеют разный масштаб, устойчивость к этим нарушениям имеет общий генетический фон и будет реагировать на повышение устойчивости в сходном направлении.Тем не менее, с практической точки зрения, устойчивость к случайным макроэкологическим нарушениям, таким как вспышки болезней и периоды сильной жары, встречается реже и, следовательно, имеет меньшее значение, по крайней мере, на большинстве ферм в умеренном климате. Таким образом, простой в управлении домашний скот — это домашний скот с повышенной устойчивостью к повседневным нарушениям микросреды в макросреде (например, на ферме), но также и с повышенной устойчивостью к нарушениям макросреды.
Генетические различия в реакции на факторы микроокружения могут быть выражены как генетическая дисперсия в размере изменчивости окружающей среды (Mulder et al., 2013). Несмотря на то, что макро- и микроэкологическая чувствительность относятся к разным понятиям и лучше всего исследуются разными методами (Mulder et al., 2015a), расчетная генетическая корреляция между ними оказалась высокой (0,76 при SE = 0,10; Mulder et al. , 2013). Это указывает на то, что, несмотря на то, что нарушения имеют разный масштаб, устойчивость к этим нарушениям имеет общий генетический фон и будет реагировать на повышение устойчивости в сходном направлении.Тем не менее, с практической точки зрения, устойчивость к случайным макроэкологическим нарушениям, таким как вспышки болезней и периоды сильной жары, встречается реже и, следовательно, имеет меньшее значение, по крайней мере, на большинстве ферм в умеренном климате. Таким образом, простой в управлении домашний скот — это домашний скот с повышенной устойчивостью к повседневным нарушениям микросреды в макросреде (например, на ферме), но также и с повышенной устойчивостью к нарушениям макросреды.
Показатели устойчивости, насколько нам известно, еще не включены в индексы селекции, несмотря на их очевидную значимость для здорового и легкого в управлении скота (а также однородность производства и эффективность производства).Это, вероятно, связано с незнанием того, как определять, измерять и взвешивать показатели устойчивости и их экономическую ценность, а также из-за убеждения, что текущие связанные со здоровьем характеристики в индексах отбора (например, продолжительность жизни, смертность, рост) охватывают устойчивость, по крайней мере, в основном. (например, Кнап, 2005). Несмотря на предложения по измерению экологической чувствительности (Knap, 2009b) (т.е. устойчивости) и определению ее экономической ценности (Knap, 2005), надлежащие инструменты сбора и обработки данных считались серьезными проблемами для реализации (Knap, 2009b). .Однако последние технологические разработки позволяют собирать большие/продольные данные и выводить из них новые фенотипы (Mulder, 2017). Эти разработки будут только расширяться и станут более важными в будущем. Требуются новые идеи для изучения и использования больших данных и новых фенотипов. Кроме того, признаки, связанные со здоровьем, не обязательно отражают общую устойчивость, а в основном отражают устойчивость к болезням (например, устойчивость к болезням; Mulder and Rashidi (2017). Кроме того, оценка племенной ценности признаков, связанных с болезнями, ограничивается лишь несколькими заболеваниями. , если есть.Таким образом, представляется, что включение общих показателей устойчивости в индексы отбора дает дополнительные преимущества.
Эти разработки будут только расширяться и станут более важными в будущем. Требуются новые идеи для изучения и использования больших данных и новых фенотипов. Кроме того, признаки, связанные со здоровьем, не обязательно отражают общую устойчивость, а в основном отражают устойчивость к болезням (например, устойчивость к болезням; Mulder and Rashidi (2017). Кроме того, оценка племенной ценности признаков, связанных с болезнями, ограничивается лишь несколькими заболеваниями. , если есть.Таким образом, представляется, что включение общих показателей устойчивости в индексы отбора дает дополнительные преимущества.
Таким образом, целью данной статьи было:
1. определить черты индикатора устойчивости на основе больших/лонгитюдных данных,
2. определить экономические значения показателей устойчивости на основе затрат на оплату труда и лечение, и
3. показать потенциал повышения устойчивости домашнего скота за счет включения устойчивости в цели селекции.
Статья посвящена описанным задачам последовательно и заканчивается общими выводами, поэтому статья отличается по своей структуре от обычных исследовательских работ.
Определение индикаторов устойчивости
Устойчивость была популярной концепцией в широком диапазоне научных дисциплин в последние два десятилетия (например, Scheffer et al., 2015; Ge et al., 2016). Однако было доказано, что определить индикаторы устойчивости сложно, и были предложены различные варианты, что говорит о том, что «правильный» индикатор устойчивости еще предстоит определить. Недавние технологические разработки открывают новые возможности для изучения альтернативных показателей устойчивости на основе продольных данных (Mulder, 2017).Здесь мы предлагаем индикаторы устойчивости, которые используют наличие множества повторных наблюдений за человеком или за (родителем) семьей (т. е. большие данные). Мы также опишем некоторые условия, которым эти индикаторы устойчивости должны соответствовать, чтобы быть информативными, и их потенциальные недостатки.
Индикаторы устойчивости
Измерить общую устойчивость сложно. Многие исследования были сосредоточены на одном конкретном типе устойчивости (особенно устойчивости к болезням) и использовали экспериментальные установки для выявления основных физиологических механизмов. Однако эти механизмы сильно зависят от характера нарушения, часто выбираются исходя из интересов исследования и могут характеризовать фенотип, который (слишком) зависит от исследуемого нарушения (Colditz and Hine, 2016). Несмотря на то, что эти исследования могут предоставить полезную информацию и понимание физиологии, результаты могут не отражать общую устойчивость. Кроме того, как правило, эти сложные среды отличаются от коммерческих сред и поэтому могут быть менее репрезентативными из-за взаимодействий GxE.Вместо этого было предложено измерять « сводных характеристики переменных отклика » (цитата из Colditz and Hine, 2016) в качестве общих показателей устойчивости (Knap, 2009b; Doeschl-Wilson et al., 2012; Colditz and Hine, 2016; Elgersma). и др., 2018).
Однако эти механизмы сильно зависят от характера нарушения, часто выбираются исходя из интересов исследования и могут характеризовать фенотип, который (слишком) зависит от исследуемого нарушения (Colditz and Hine, 2016). Несмотря на то, что эти исследования могут предоставить полезную информацию и понимание физиологии, результаты могут не отражать общую устойчивость. Кроме того, как правило, эти сложные среды отличаются от коммерческих сред и поэтому могут быть менее репрезентативными из-за взаимодействий GxE.Вместо этого было предложено измерять « сводных характеристики переменных отклика » (цитата из Colditz and Hine, 2016) в качестве общих показателей устойчивости (Knap, 2009b; Doeschl-Wilson et al., 2012; Colditz and Hine, 2016; Elgersma). и др., 2018).
Кольдиц и Хайн (2016) предложили разнообразный набор « сводных характеристик переменных реакции » для измерения устойчивости к возмущению, включая типичные производственные характеристики, такие как потребление корма, скорость роста и другие производственные переменные. Люди, испытывающие нарушения, едят и производят меньше (т. е. отклоняются от) своего потенциала без нарушений (например, Van der Waaij et al., 2000). Однако отклонение между наблюдаемым средним значением продуктивности и предполагаемым потенциалом (как это было предложено Кольдитцем и Хайном, 2016 г.) не обязательно полностью соответствует определению устойчивости: у устойчивых животных может наблюдаться резкое падение продуктивности, но они также могут иметь возможность быстро восстановиться. к состоянию до воздействия беспокойства по сравнению с менее устойчивыми животными.Отклонения в течение определенного периода времени (по крайней мере, в течение продолжительности нарушения), вероятно, лучше отражают устойчивость. Таким образом, устойчивость может быть измерена на основе отклонений ожидаемого производства и наблюдаемого производства (т. е. остатков) за определенный период времени.
Люди, испытывающие нарушения, едят и производят меньше (т. е. отклоняются от) своего потенциала без нарушений (например, Van der Waaij et al., 2000). Однако отклонение между наблюдаемым средним значением продуктивности и предполагаемым потенциалом (как это было предложено Кольдитцем и Хайном, 2016 г.) не обязательно полностью соответствует определению устойчивости: у устойчивых животных может наблюдаться резкое падение продуктивности, но они также могут иметь возможность быстро восстановиться. к состоянию до воздействия беспокойства по сравнению с менее устойчивыми животными.Отклонения в течение определенного периода времени (по крайней мере, в течение продолжительности нарушения), вероятно, лучше отражают устойчивость. Таким образом, устойчивость может быть измерена на основе отклонений ожидаемого производства и наблюдаемого производства (т. е. остатков) за определенный период времени.
Было описано множество признаков потенциального индикатора устойчивости, предназначенных для прогнозирования изменений в состоянии экосистем (подробный обзор см. в Таблице 1 Шеффера и др., 2015 г.). Однако многие из этих индикаторов устойчивости трудно применить к видам домашнего скота или генетическому улучшению скота, потому что индикаторы устойчивости не подходят для данных о животноводстве (например,г., пространственная корреляция, спектральное покраснение). Кроме того, большое количество исследованных показателей устойчивости схожи, поскольку они используют данные сходным образом, т. е. количество уникальных показателей ограничено. Тем не менее, мы видим возможности для некоторых из них. В этой статье мы подробно остановились на четырех индикаторах устойчивости, для которых мы предлагаем, чтобы они отражали различные перспективы устойчивости [см. также Scheffer et al. (2018)]. Подходящими индикаторами устойчивости, основанными на одном признаке (например, продуктивность, потребление корма) в животноводстве, могут быть дисперсия отклонений, автокорреляция отклонений, асимметрия отклонений продуктивных признаков за период времени и наклон нормы реакции (см.
в Таблице 1 Шеффера и др., 2015 г.). Однако многие из этих индикаторов устойчивости трудно применить к видам домашнего скота или генетическому улучшению скота, потому что индикаторы устойчивости не подходят для данных о животноводстве (например,г., пространственная корреляция, спектральное покраснение). Кроме того, большое количество исследованных показателей устойчивости схожи, поскольку они используют данные сходным образом, т. е. количество уникальных показателей ограничено. Тем не менее, мы видим возможности для некоторых из них. В этой статье мы подробно остановились на четырех индикаторах устойчивости, для которых мы предлагаем, чтобы они отражали различные перспективы устойчивости [см. также Scheffer et al. (2018)]. Подходящими индикаторами устойчивости, основанными на одном признаке (например, продуктивность, потребление корма) в животноводстве, могут быть дисперсия отклонений, автокорреляция отклонений, асимметрия отклонений продуктивных признаков за период времени и наклон нормы реакции (см. табл. 1).Мы предполагаем, что в целом отрицательные отклонения в основном наблюдаются у менее устойчивых животных, но в зависимости от признака, например, температуры тела, это может быть и противоположным.
табл. 1).Мы предполагаем, что в целом отрицательные отклонения в основном наблюдаются у менее устойчивых животных, но в зависимости от признака, например, температуры тела, это может быть и противоположным.
• Дисперсия отклонений (также известная как однородность, унаследованная изменчивость, остаточная дисперсия или генетическая неоднородность изменчивости окружающей среды (см. обзоры Hill and Mulder, 2010; Elgersma et al., 2018) дает представление о влиянии нарушений с: низкой (более высокой) дисперсией для животных без нарушений или не подверженных воздействию нарушений и высокой (более высокой) дисперсией для животных, подвергающихся воздействию нарушений.Мы предполагаем, что дисперсия отклонений отражает тяжесть и продолжительность (т. е. скорость восстановления) возмущений окружающей среды, с которыми сталкивается человек. Вклад серьезности и продолжительности не может быть выделен в рамках этого показателя устойчивости.
• (Lag-one) автокорреляция отклонений дает представление о продолжительности воздействия нарушений с: автокорреляцией около 0 (т. е. последующие отклонения не связаны) для животных без нарушений, без воздействия нарушений или с быстрое восстановление после нарушений; автокорреляция в сторону +1 (т.т. е. последующие отклонения более сходны) у животных, находящихся под влиянием нарушений и с более медленным (более длительным) восстановлением после нарушений; и автокорреляция в сторону -1 (т. е. последующие отклонения противоположны) для животных, находящихся под влиянием нарушений, и быстрая и сверхкомпенсирующая реакция на нарушения, например, компенсаторный рост. Мы предполагаем, что автокорреляция отклонений отражает продолжительность (то есть скорость восстановления) возмущений окружающей среды, с которыми сталкивается человек.
е. последующие отклонения не связаны) для животных без нарушений, без воздействия нарушений или с быстрое восстановление после нарушений; автокорреляция в сторону +1 (т.т. е. последующие отклонения более сходны) у животных, находящихся под влиянием нарушений и с более медленным (более длительным) восстановлением после нарушений; и автокорреляция в сторону -1 (т. е. последующие отклонения противоположны) для животных, находящихся под влиянием нарушений, и быстрая и сверхкомпенсирующая реакция на нарушения, например, компенсаторный рост. Мы предполагаем, что автокорреляция отклонений отражает продолжительность (то есть скорость восстановления) возмущений окружающей среды, с которыми сталкивается человек.
• Асимметрия отклонений указывает направление отклонений с помощью: асимметрии около 0 для животных без нарушений или не подвергающихся воздействию нарушений; положительная асимметрия для животных, имеющих (в основном) положительные отклонения за счет положительной реакции на улучшение среды; и отрицательная асимметрия для животных, имеющих (в основном) отрицательные отклонения из-за нарушений. Мы предполагаем, что асимметрия отклонений отражает серьезность возмущений окружающей среды, которые испытывает человек.
Мы предполагаем, что асимметрия отклонений отражает серьезность возмущений окружающей среды, которые испытывает человек.
• Наклон нормы реакции указывает на устойчивость к макроэкологическим нарушениям (Finlay and Wilkinson, 1963), например, к заболеваемости (Herrero-Medrano et al., 2015) или аномальной жаре (Ravagnolo and Misztal). , 2000). Наклон нормы реакции оценивается на основе продукции особи с учетом уровня возмущения с: наклоном 0 для животных, не подверженных влиянию возмущения, и наклоном ниже 0 для животных, подвергшихся воздействию возмущения с более крутым, отрицательным склоны для животных, которые больше подвержены влиянию.Мы предполагаем, что наклон нормы реакции отражает тяжесть макросредовых возмущений, которые испытывает человек.
Таблица 1 . Обзор возможных индикаторов устойчивости на основе больших/продольных данных по животным с описанием, интерпретацией и примечаниями.
Таким образом, ожидается, что менее устойчивые животные будут иметь большую дисперсию, положительную автокорреляцию, отрицательную асимметрию и более крутой наклон, чем в среднем по популяции, если предположить, что нарушение снижает значение признака. Ожидается, что устойчивые животные будут иметь меньшую дисперсию (т. е. ближе к 0), автокорреляцию и асимметрию около 0, а наклон ближе к нулю, чем в среднем по популяции.
Ожидается, что устойчивые животные будут иметь меньшую дисперсию (т. е. ближе к 0), автокорреляцию и асимметрию около 0, а наклон ближе к нулю, чем в среднем по популяции.
Необходимо сделать несколько замечаний относительно предлагаемых индикаторов устойчивости. Следует сделать общее замечание: масштабирование может привести к тому, что животные с более высоким средним значением будут иметь более высокую дисперсию отклонений, но они также могут быть генетически более чувствительными и менее устойчивыми по сравнению с животными с низкой продуктивностью (Falconer and Mackay, 1996).Следует сделать следующие конкретные замечания: основное допущение как для дисперсии, так и для автокорреляции как индикаторов устойчивости состоит в том, что отклонения в основном (или исключительно) отрицательны, поскольку возмущения уменьшат производительность. Однако дисперсия и автокорреляция не различают отрицательные и положительные отклонения. Следовательно, животные, демонстрирующие продуктивность выше ожидаемой, например, компенсаторную продуктивность (возможно, желательную черту, часть устойчивости), будут характеризоваться как не/менее устойчивые. С другой стороны, такие животные, вероятно, показали серьезное снижение продуктивности из-за беспокойства, явно демонстрируя отсутствие устойчивости к беспокойству. Напротив, асимметрия действительно различает положительные и отрицательные отклонения, но, вероятно, более чувствительна к выбросам из-за кубических членов. Автокорреляция также может быть отрицательной, что означает, что животное демонстрирует быструю и гиперкомпенсирующую реакцию на нарушения, например, компенсаторный рост. Хотя отрицательная автокорреляция предпочтительнее (сильной) положительной автокорреляции, животное с автокорреляцией около 0 вообще не подвержено влиянию возмущения и, следовательно, предпочтительнее отрицательной автокорреляции.Наконец, для оценки наклона нормы реакции требуется количественная оценка на уровне фермы либо по известной коварианте окружающей среды, например по температуре (Ravagnolo and Misztal, 2000; Bohmanova et al., 2007; Carabaño et al., 2017), либо по общее падение производства на ферме в результате многих отдельных падений производства на этой ферме (Rashidi et al.
С другой стороны, такие животные, вероятно, показали серьезное снижение продуктивности из-за беспокойства, явно демонстрируя отсутствие устойчивости к беспокойству. Напротив, асимметрия действительно различает положительные и отрицательные отклонения, но, вероятно, более чувствительна к выбросам из-за кубических членов. Автокорреляция также может быть отрицательной, что означает, что животное демонстрирует быструю и гиперкомпенсирующую реакцию на нарушения, например, компенсаторный рост. Хотя отрицательная автокорреляция предпочтительнее (сильной) положительной автокорреляции, животное с автокорреляцией около 0 вообще не подвержено влиянию возмущения и, следовательно, предпочтительнее отрицательной автокорреляции.Наконец, для оценки наклона нормы реакции требуется количественная оценка на уровне фермы либо по известной коварианте окружающей среды, например по температуре (Ravagnolo and Misztal, 2000; Bohmanova et al., 2007; Carabaño et al., 2017), либо по общее падение производства на ферме в результате многих отдельных падений производства на этой ферме (Rashidi et al. , 2014; Herrero-Medrano et al., 2015). Кроме того, каждое возмущение имеет свой наклон, а также практически невозможно оценить несколько наклонов для нескольких возмущений, происходящих одновременно.Однако наклон нормы реакции также можно интерпретировать как (форму) общей устойчивости, хотя наклон, вероятно, не отражает реакцию на нарушения микросреды, которые возникают на уровне отдельных животных, например, эндемические заболевания. Таким образом, наклон предоставляет информацию только о нарушениях, которые вызывают снижение производительности фермы, таких как тепловой стресс или вспышка болезни (например, в случае теплового стресса). Таким образом, наклон дает информацию только о нарушениях макросреды.Это отличается от трех других предлагаемых индикаторов устойчивости, основанных на отклонениях в таблице 1, которые можно использовать для эпидемических (на уровне ферм) и эндемических событий (на индивидуальном уровне). Таким образом, эти индикаторы предоставляют информацию для оценки племенной ценности при всех типах нарушений и, таким образом, общей устойчивости.
, 2014; Herrero-Medrano et al., 2015). Кроме того, каждое возмущение имеет свой наклон, а также практически невозможно оценить несколько наклонов для нескольких возмущений, происходящих одновременно.Однако наклон нормы реакции также можно интерпретировать как (форму) общей устойчивости, хотя наклон, вероятно, не отражает реакцию на нарушения микросреды, которые возникают на уровне отдельных животных, например, эндемические заболевания. Таким образом, наклон предоставляет информацию только о нарушениях, которые вызывают снижение производительности фермы, таких как тепловой стресс или вспышка болезни (например, в случае теплового стресса). Таким образом, наклон дает информацию только о нарушениях макросреды.Это отличается от трех других предлагаемых индикаторов устойчивости, основанных на отклонениях в таблице 1, которые можно использовать для эпидемических (на уровне ферм) и эндемических событий (на индивидуальном уровне). Таким образом, эти индикаторы предоставляют информацию для оценки племенной ценности при всех типах нарушений и, таким образом, общей устойчивости. Но, возможно, «лучший» показатель устойчивости еще предстоит определить на основе комбинации различных показателей устойчивости: многомерный подход к устойчивости, например, основанный на взаимных корреляциях или собственных значениях, которые отражают отношения между различными показателями устойчивости. на основе разных фенотипов (т.г., Ладе и Гросс, 2012; Шеффер и др., 2015; Gijzel et al., 2017), или индекс ряда показателей (т. е. подход индекса отбора, Hazel, 1943).
Но, возможно, «лучший» показатель устойчивости еще предстоит определить на основе комбинации различных показателей устойчивости: многомерный подход к устойчивости, например, основанный на взаимных корреляциях или собственных значениях, которые отражают отношения между различными показателями устойчивости. на основе разных фенотипов (т.г., Ладе и Гросс, 2012; Шеффер и др., 2015; Gijzel et al., 2017), или индекс ряда показателей (т. е. подход индекса отбора, Hazel, 1943).
Мало что известно о полезности предложенных показателей устойчивости, основанных на отклонениях, для генетики домашнего скота: ни в одном исследовании не изучались автокорреляции отклонений и асимметрия отклонений, и лишь в нескольких исследованиях изучалась дисперсия отклонений. Элгерсма и др. (2018) исследовали сырую фенотипическую дисперсию суточного удоя в течение всего периода дойки и ее связь с показателями здоровья и долголетия молочных коров.Необработанная дисперсия надоев была наследственной (0,10; Elgersma et al. , 2018). Более того, на генетическом уровне у коров с низкой дисперсией отклонений по удою было значительно меньше производственных заболеваний [т.е. здоровье вымени, генетическая корреляция (r g ) = -0,36; кетоз, r г = -0,52] и более высокая продолжительность жизни (r г = -0,30), что свидетельствует о более высокой устойчивости (Elgersma et al., 2018). Путц и др. (2018) продемонстрировали аналогичные результаты для свиней, содержащихся в «естественной среде заражения»: отклонения в суточном потреблении корма и отклонения в продолжительности дня в кормушке на этапе откорма положительно генетически коррелировали со смертностью и количеством обработок (Putz et al., 2018), что указывает на то, что свиньи с более низкой дисперсией имеют более низкую смертность и нуждаются в меньшем количестве лечения. Это означает, что как потребление корма, так и продолжительность кормления указывают на состояние здоровья на генетическом уровне и, таким образом, показывают, что изменение часто измеряемых признаков может указывать на устойчивость.
, 2018). Более того, на генетическом уровне у коров с низкой дисперсией отклонений по удою было значительно меньше производственных заболеваний [т.е. здоровье вымени, генетическая корреляция (r g ) = -0,36; кетоз, r г = -0,52] и более высокая продолжительность жизни (r г = -0,30), что свидетельствует о более высокой устойчивости (Elgersma et al., 2018). Путц и др. (2018) продемонстрировали аналогичные результаты для свиней, содержащихся в «естественной среде заражения»: отклонения в суточном потреблении корма и отклонения в продолжительности дня в кормушке на этапе откорма положительно генетически коррелировали со смертностью и количеством обработок (Putz et al., 2018), что указывает на то, что свиньи с более низкой дисперсией имеют более низкую смертность и нуждаются в меньшем количестве лечения. Это означает, что как потребление корма, так и продолжительность кормления указывают на состояние здоровья на генетическом уровне и, таким образом, показывают, что изменение часто измеряемых признаков может указывать на устойчивость.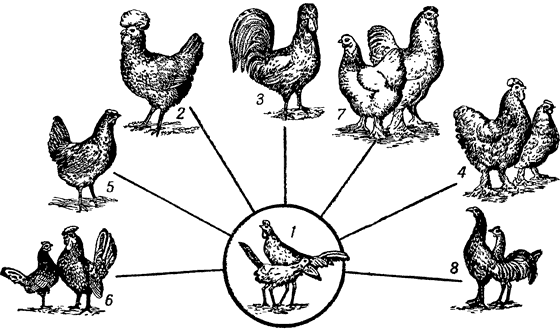 Кроме того, во многих других исследованиях изучалась наследуемость однородности производственных признаков, которая аналогична дисперсии отклонений: сообщаемая наследуемость колебалась от 0,00 до 0,15 почти у всех видов домашнего скота (см. Hill and Mulder, 2010, обзор до 2010 г.; Невес и др., 2011; Янхунен и др., 2012; Сае-Лим и др., 2015, 2017; Малдер и др., 2016; см. дополнительную информацию в Elgersma et al., 2018). Кроме того, генетические коэффициенты изменчивости, как правило, были от умеренных до высоких, что предполагает относительно большой потенциал генетического улучшения однородности. Эти исследования иллюстрируют хороший потенциал для использования производственных отклонений в качестве показателей устойчивости.
Кроме того, во многих других исследованиях изучалась наследуемость однородности производственных признаков, которая аналогична дисперсии отклонений: сообщаемая наследуемость колебалась от 0,00 до 0,15 почти у всех видов домашнего скота (см. Hill and Mulder, 2010, обзор до 2010 г.; Невес и др., 2011; Янхунен и др., 2012; Сае-Лим и др., 2015, 2017; Малдер и др., 2016; см. дополнительную информацию в Elgersma et al., 2018). Кроме того, генетические коэффициенты изменчивости, как правило, были от умеренных до высоких, что предполагает относительно большой потенциал генетического улучшения однородности. Эти исследования иллюстрируют хороший потенциал для использования производственных отклонений в качестве показателей устойчивости.
Условия измерения показателей устойчивости
Для успешного использования индикаторов устойчивости в программах разведения применяются определенные условия: сбор наблюдений должен производиться на многих животных, наблюдения должны проводиться часто и в течение более длительного периода времени, окружающая среда должна быть сложной и разнообразной для оценки общей устойчивости. , и, что наиболее важно, показатели устойчивости должны быть информативными для устойчивости.
, и, что наиболее важно, показатели устойчивости должны быть информативными для устойчивости.
Производственные признаки, подходящие для исследования показателей устойчивости на основе отклонений, могут быть измерены повторно на индивидуальном уровне или на уровне семьи (см., например, Рисунок 1). Повторные наблюдения на индивидуальном уровне позволяют более точно оценить показатели устойчивости и их генетическую изменчивость, поэтому они предпочтительны. Подходящими признаками являются, например, надои, масса яиц, живая масса/прирост и потребление корма. Важный момент, на который необходимо обратить внимание, заключается в том, что эти признаки могут различаться по размеру отклонений в зависимости от стадии (лактации) или возраста (для роста молодых животных): может потребоваться стандартизация отклонений по стадиям или возрастам или лечение разных стадий/возрастов. как разные черты.В случае, если на особь собирается слишком мало повторных наблюдений за продуктивными признаками, альтернативой являются отклонения продуктивных признаков, собранные на (отцовском) семейном уровне. Такой подход к исследованию индикаторов устойчивости особенно полезен для скота, содержащегося в большом количестве и имеющего относительно низкую экономическую ценность (например, рыба и птица), или скота, содержащегося в экстенсивных системах земледелия (например, мясной крупный рогатый скот и овцы). Подходящими признаками являются, например, признаки убоя/туши (например, Ibáñez-Escriche et al., 2008) или признаки, зарегистрированные только в течение ограниченного периода жизни (например, Neves et al., 2011; Mulder et al., 2016; Iung et al., 2017). Следует ожидать, что технические разработки в (ближайшем) будущем позволят исследовать отклонения на основе повторных наблюдений за одной и той же особью для всех видов домашнего скота.
Такой подход к исследованию индикаторов устойчивости особенно полезен для скота, содержащегося в большом количестве и имеющего относительно низкую экономическую ценность (например, рыба и птица), или скота, содержащегося в экстенсивных системах земледелия (например, мясной крупный рогатый скот и овцы). Подходящими признаками являются, например, признаки убоя/туши (например, Ibáñez-Escriche et al., 2008) или признаки, зарегистрированные только в течение ограниченного периода жизни (например, Neves et al., 2011; Mulder et al., 2016; Iung et al., 2017). Следует ожидать, что технические разработки в (ближайшем) будущем позволят исследовать отклонения на основе повторных наблюдений за одной и той же особью для всех видов домашнего скота.
Рисунок 1 . Два примера производственных признаков, подходящих для исследования показателей устойчивости на основе отклонений. (A) Показаны повторные измерения удоя на индивидуальном уровне двух дойных коров с одной и той же базовой кривой Вильминка (красным), но с разной устойчивостью: серым цветом показана менее устойчивая молочная корова с более высокими колебаниями удоя, а черная более выносливая молочная корова с меньшими колебаниями надоев. (B) Показывает измерения веса туш на уровне семейства двух семейств, отличающихся устойчивостью: серым цветом показано менее устойчивое семейство с более высокими колебаниями веса туш, а черным цветом — более устойчивое семейство с более низкими колебаниями веса туш.
(B) Показывает измерения веса туш на уровне семейства двух семейств, отличающихся устойчивостью: серым цветом показано менее устойчивое семейство с более высокими колебаниями веса туш, а черным цветом — более устойчивое семейство с более низкими колебаниями веса туш.
Необходимо соблюдать осторожность при правильной оценке отклонений: менее устойчивые животные будут иметь больше колебаний в продуктивности, но, как следствие, ожидаемая продуктивность (на основании собственных наблюдений) будет ниже. Это означает, что отклонения будут меньше, если использовать просто среднюю ожидаемую продуктивность, а жизнестойкость животного завысить.Например, Элгерсма и др. (2018) пришли к выводу, что необработанная фенотипическая дисперсия суточного удоя является составной мерой остаточной дисперсии и формы кривой лактации (Elgersma et al., 2018). Они предложили смоделировать индивидуальную кривую лактации и использовать дисперсию отклонений от кривой лактации в качестве лучшего индикатора устойчивости (Elgersma et al. , 2018), как это сделали Codrea et al. (2011). Однако у человека, постоянно страдающего от нарушений (или нарушений), кривая лактации будет ниже, чем его истинный потенциал без (каких-либо) нарушений.Как следствие, отклонения, основанные на индивидуальной кривой лактации, будут ниже, чем по сравнению с индивидуальной потенциальной кривой лактации. Моделирование кривых лактации на основе наблюдаемых данных может поглотить часть дисперсии отклонений. Вместо этого отклонения могут быть основаны на разнице между наблюдаемой выработкой и потенциальной выработкой индивидуума, например, на основе его (G)EBV. Определение ожидаемой продуктивности животного является одной из основных задач объективной оценки предлагаемых показателей устойчивости и требует дальнейшего изучения в ближайшем будущем.
, 2018), как это сделали Codrea et al. (2011). Однако у человека, постоянно страдающего от нарушений (или нарушений), кривая лактации будет ниже, чем его истинный потенциал без (каких-либо) нарушений.Как следствие, отклонения, основанные на индивидуальной кривой лактации, будут ниже, чем по сравнению с индивидуальной потенциальной кривой лактации. Моделирование кривых лактации на основе наблюдаемых данных может поглотить часть дисперсии отклонений. Вместо этого отклонения могут быть основаны на разнице между наблюдаемой выработкой и потенциальной выработкой индивидуума, например, на основе его (G)EBV. Определение ожидаемой продуктивности животного является одной из основных задач объективной оценки предлагаемых показателей устойчивости и требует дальнейшего изучения в ближайшем будущем.
Для правильной оценки общей жизнестойкости особей необходимы два фактора: продолжительность периода наблюдения и частота наблюдений. Во-первых, продолжительность периода наблюдения должна быть достаточно большой, чтобы допустить возникновение различных типов возмущений, как было предложено Mulder et al. (2013): в стрессовой среде можно наблюдать больше генетических вариаций чувствительности к микроокружающей среде по сравнению с менее стрессовой средой (Mulder et al., 2013).Потенциальный риск заключается в том, что результаты не воспроизводимы между средами или даже внутри них: например, устойчивость, основанная на нарушениях, вызванных одним конкретным типом (например, болезнями), не отражает устойчивости к другим типам нарушений (например, жаре), и, следовательно, не отражает общей устойчивости. Во-вторых, частота наблюдений должна быть достаточно высокой, чтобы фиксировать отклонения, вызванные возмущениями. Элгерсма и др. (2018) предположили, что более ранние исследования могли недооценивать потенциал показателей устойчивости из-за слишком длительных интервалов между днями тестирования, например.г., ежемесячные учеты удоев. Кроме того, в случае использования семейных отклонений существует значительный риск того, что это не отразит никаких нарушений (в случае отсутствия или наличия небольших отклонений), не говоря уже о разнообразном наборе типов нарушений для оценки общей устойчивости.
(2013): в стрессовой среде можно наблюдать больше генетических вариаций чувствительности к микроокружающей среде по сравнению с менее стрессовой средой (Mulder et al., 2013).Потенциальный риск заключается в том, что результаты не воспроизводимы между средами или даже внутри них: например, устойчивость, основанная на нарушениях, вызванных одним конкретным типом (например, болезнями), не отражает устойчивости к другим типам нарушений (например, жаре), и, следовательно, не отражает общей устойчивости. Во-вторых, частота наблюдений должна быть достаточно высокой, чтобы фиксировать отклонения, вызванные возмущениями. Элгерсма и др. (2018) предположили, что более ранние исследования могли недооценивать потенциал показателей устойчивости из-за слишком длительных интервалов между днями тестирования, например.г., ежемесячные учеты удоев. Кроме того, в случае использования семейных отклонений существует значительный риск того, что это не отразит никаких нарушений (в случае отсутствия или наличия небольших отклонений), не говоря уже о разнообразном наборе типов нарушений для оценки общей устойчивости. Напротив, слишком малые периоды времени могут вызвать больший шум, и поэтому может стать труднее найти соответствующую информацию для обеспечения устойчивости. Период времени зависит от измеряемого признака. Будущие исследования должны быть сосредоточены на поиске оптимального периода времени для определения отклонений путем изучения точности EBV и генетических корреляций между индикатором (индикаторами) устойчивости и существующими признаками здоровья.Независимо от этого, устойчивость идеально оценивается в среде со всеми видами возмущений в течение всего производственного периода с частыми измерениями.
Напротив, слишком малые периоды времени могут вызвать больший шум, и поэтому может стать труднее найти соответствующую информацию для обеспечения устойчивости. Период времени зависит от измеряемого признака. Будущие исследования должны быть сосредоточены на поиске оптимального периода времени для определения отклонений путем изучения точности EBV и генетических корреляций между индикатором (индикаторами) устойчивости и существующими признаками здоровья.Независимо от этого, устойчивость идеально оценивается в среде со всеми видами возмущений в течение всего производственного периода с частыми измерениями.
Недавние технологические разработки позволяют значительно увеличить количество наблюдений и количество наблюдаемых фенотипов: большие/продольные данные и новые фенотипы приведут к получению большего количества новых данных об индивидуумах для более точной оценки отклонений и, следовательно, генетических параметров. В настоящее время многие (селекционные) организации используют рутинный сбор данных. Автоматические доильные системы (AMS) и автоматические системы кормления (AFS) для крупного рогатого скота и свиней являются наиболее известными и хорошо разработанными примерами. Другими разработками, которые в настоящее время находятся в стадии изучения, являются, например, автоматические системы взвешивания и автоматический учет производства яиц при групповом содержании. Ожидается, что сбор большего количества данных и новых фенотипов увеличится в ближайшем будущем, что поможет в (лучшей) оценке генетической изменчивости отклонений: более высокая наследуемость обычно обнаруживается при большем количестве наблюдений (т.е., индивидуальные измерения) на человека или семью (Hill and Mulder, 2010; Elgersma et al., 2018), что свидетельствует о том, что для точной оценки генетической изменчивости требуются большие наборы данных. Кроме того, хотя оценки наследуемости дисперсии отклонений, основанные на отдельных записях, часто очень низки (около 0,01; см., например, Hill and Mulder, 2010; Elgersma et al.
Автоматические доильные системы (AMS) и автоматические системы кормления (AFS) для крупного рогатого скота и свиней являются наиболее известными и хорошо разработанными примерами. Другими разработками, которые в настоящее время находятся в стадии изучения, являются, например, автоматические системы взвешивания и автоматический учет производства яиц при групповом содержании. Ожидается, что сбор большего количества данных и новых фенотипов увеличится в ближайшем будущем, что поможет в (лучшей) оценке генетической изменчивости отклонений: более высокая наследуемость обычно обнаруживается при большем количестве наблюдений (т.е., индивидуальные измерения) на человека или семью (Hill and Mulder, 2010; Elgersma et al., 2018), что свидетельствует о том, что для точной оценки генетической изменчивости требуются большие наборы данных. Кроме того, хотя оценки наследуемости дисперсии отклонений, основанные на отдельных записях, часто очень низки (около 0,01; см., например, Hill and Mulder, 2010; Elgersma et al. , 2018), наследуемость остаточной дисперсии многих из этих наблюдений одного животного на самом деле является умеренным (Damgaard et al., 2003; Капелл и др., 2011; Селл-Кубиак и др., 2015; Малдер и др., 2016; Элгерсма и др., 2018). На рис. 2 показана взаимосвязь между наследуемостью остаточной дисперсии, основанной на нескольких отдельных записях, и наследуемостью остаточной дисперсии, основанной на одной отдельной записи. Таким образом, последние технологические разработки, такие как AMS и AFS, позволяют или позволят собирать большие наборы данных с большим количеством наблюдений на животное (т. е. большие данные). Это значительно облегчит использование показателей устойчивости в программах разведения всех видов скота.
, 2018), наследуемость остаточной дисперсии многих из этих наблюдений одного животного на самом деле является умеренным (Damgaard et al., 2003; Капелл и др., 2011; Селл-Кубиак и др., 2015; Малдер и др., 2016; Элгерсма и др., 2018). На рис. 2 показана взаимосвязь между наследуемостью остаточной дисперсии, основанной на нескольких отдельных записях, и наследуемостью остаточной дисперсии, основанной на одной отдельной записи. Таким образом, последние технологические разработки, такие как AMS и AFS, позволяют или позволят собирать большие наборы данных с большим количеством наблюдений на животное (т. е. большие данные). Это значительно облегчит использование показателей устойчивости в программах разведения всех видов скота.
Рисунок 2 . Наследуемость остаточной дисперсии (hостаточная дисперсия2=nиндивидуальная запись21+(n-1)rзаписей) как функция количества наблюдений на одно животное ( n ) для двух наследуемости остаточной дисперсии на основе одной отдельной записи (индивидуальная запись2) и две повторяемости записей остаточной дисперсии ( r индивидуальная запись ). Используемая наследуемость остаточной дисперсии на основе одной отдельной записи (индивидуальная запись2) аналогична описанным в литературе наследуемым вариациям остаточной дисперсии на основе одной отдельной записи (т.э., одно единственное фенотипическое наблюдение), см. Hill and Mulder (2010) и Elgersma et al. (2018) для примеров.
Используемая наследуемость остаточной дисперсии на основе одной отдельной записи (индивидуальная запись2) аналогична описанным в литературе наследуемым вариациям остаточной дисперсии на основе одной отдельной записи (т.э., одно единственное фенотипическое наблюдение), см. Hill and Mulder (2010) и Elgersma et al. (2018) для примеров.
Важность устойчивости для целей разведения
Определение цели разведения является одним из важнейших элементов разведения животных. Цель селекции и соответствующий индекс селекции определяют направление генетического улучшения. У всех видов селекционные цели перешли от преимущественно продуктивных целей к сбалансированным селекционным целям, направленным на одновременное улучшение продуктивности, продуктивности, здоровья и функциональных признаков (Olesen et al., 2003; Кнап, 2009а; Нитесон-Ван Ньювенховен и др., 2013 г.; Мильор и др., 2017). Генетическое улучшение устойчивости вписывается в философию сбалансированного разведения, но, насколько нам известно, устойчивость (пока) не включена в цели разведения скота. Если устойчивость влияет на прибыль фермы, она должна быть целью разведения. Другими словами, если экономическая ценность устойчивости отлична от нуля, она должна быть целью разведения. Вопрос, однако, в том, как мы можем определить экономическую ценность устойчивости.
Если устойчивость влияет на прибыль фермы, она должна быть целью разведения. Другими словами, если экономическая ценность устойчивости отлична от нуля, она должна быть целью разведения. Вопрос, однако, в том, как мы можем определить экономическую ценность устойчивости.
Чтобы определить экономическую ценность устойчивости, мы можем рассмотреть затраты на отсутствие устойчивости, например, более высокие производственные потери, (трудовые) затраты на лечение, ветеринарные расходы и трудозатраты фермера на наблюдение за животными, у которых проявляются признаки отсутствие сопротивляемости. При определении экономической ценности признаков важно избегать двойного учета. Если устойчивость определяется как колебания производства, необходимо соблюдать осторожность, чтобы избежать двойного учета. Например, производственные потери (т.т. е., отклонения из-за отсутствия жизнестойкости) уже могут быть отражены в продуктивных признаках, особенно у молочных коров. Кроме того, затраты на лечение могут быть уже учтены в цели разведения, например, в случае мастита у молочного скота. Затраты на лечение мастита или затраты на выбрасываемое молоко не должны учитываться при устойчивости. С другой стороны, производственные потери и издержки, связанные с болезнями, не всегда включаются в цели разведения, например, свиней и домашней птицы: в этих целях разведения отсутствуют признаки здоровья, а наблюдаемые производственные потери в коммерческих или гибридных условиях обусловлены нарушения могут не наблюдаться в условиях отбора с высоким уровнем здоровья.Следовательно, экономическая ценность устойчивости свиней и птицы может включать производственные потери в коммерческой среде и затраты на здоровье. В любом случае, экономическая ценность может быть основана, например, на затратах труда на наблюдение за животными, у которых проявляются признаки болезни или другие проблемы, например, сигналы тревоги или визуальные признаки. Эти расходы часто упускают из виду, потому что они считаются частью повседневного управления. Однако, если количество животных на одного работника фермы увеличивается, а рабочее время ограничено, это явно связано с потребностью фермера в здоровых и простых в содержании животных.
Затраты на лечение мастита или затраты на выбрасываемое молоко не должны учитываться при устойчивости. С другой стороны, производственные потери и издержки, связанные с болезнями, не всегда включаются в цели разведения, например, свиней и домашней птицы: в этих целях разведения отсутствуют признаки здоровья, а наблюдаемые производственные потери в коммерческих или гибридных условиях обусловлены нарушения могут не наблюдаться в условиях отбора с высоким уровнем здоровья.Следовательно, экономическая ценность устойчивости свиней и птицы может включать производственные потери в коммерческой среде и затраты на здоровье. В любом случае, экономическая ценность может быть основана, например, на затратах труда на наблюдение за животными, у которых проявляются признаки болезни или другие проблемы, например, сигналы тревоги или визуальные признаки. Эти расходы часто упускают из виду, потому что они считаются частью повседневного управления. Однако, если количество животных на одного работника фермы увеличивается, а рабочее время ограничено, это явно связано с потребностью фермера в здоровых и простых в содержании животных. Генетическое улучшение устойчивости снизит потребность в рабочей силе и позволит фермеру содержать больше животных. Поэтому устойчивость должна быть включена в цели селекции.
Генетическое улучшение устойчивости снизит потребность в рабочей силе и позволит фермеру содержать больше животных. Поэтому устойчивость должна быть включена в цели селекции.
Для устойчивого животноводства необходимо учитывать экологические и социальные проблемы, помимо экономических (Olesen et al., 2000; Nielsen and Amer, 2007). Другими словами, в дополнение к экономической ценности устойчивости может присутствовать неэкономическая ценность. Для определения неэкономических ценностей может потребоваться обширная работа (например,г., Нильсен и др., 2011; Grimsrud et al., 2013), и может, например, основываться на улучшении здоровья и благополучия животных, а также на удовлетворенности фермеров работой. Однако, чтобы просто проиллюстрировать устойчивость как концепцию, мы не будем принимать во внимание неэкономические ценности в этой статье.
Далее мы будем использовать пример затрат на рабочую силу, чтобы показать потенциал включения устойчивости в цели разведения. Дополнительные трудозатраты на (более низкую) устойчивость связаны с вероятностью того, что животное подаст сигнал тревоги. Оповещение — это предупреждение, которое может указывать на то, что животное находится под влиянием беспокойства. Предупреждение может быть сгенерировано либо визуальным осмотром, либо датчиками, AMS или AFS. Мы предполагаем, что оповещения генерируются, когда признак (с нормальным распределением и индивидуальной дисперсией) превышает фиксированное пороговое значение, основанное на дисперсии популяции (например, пороговое значение, которое принадлежит 1% вероятности для всей популяции). Этим признаком может быть, например, надои или живая масса, но, вероятно, не ранее предложенные показатели устойчивости.Ожидается, что предлагаемые индикаторы устойчивости будут обнаруживать проблемы слишком поздно, и мы предлагаем использовать их для разведения более устойчивых животных, а не использовать их в качестве предикторов предупреждений. Разведение более устойчивых животных приведет к получению потомства с меньшей дисперсией их сенсорных значений, чем у текущего поколения, и, следовательно, с меньшей вероятностью генерирования предупреждения (рис.
Оповещение — это предупреждение, которое может указывать на то, что животное находится под влиянием беспокойства. Предупреждение может быть сгенерировано либо визуальным осмотром, либо датчиками, AMS или AFS. Мы предполагаем, что оповещения генерируются, когда признак (с нормальным распределением и индивидуальной дисперсией) превышает фиксированное пороговое значение, основанное на дисперсии популяции (например, пороговое значение, которое принадлежит 1% вероятности для всей популяции). Этим признаком может быть, например, надои или живая масса, но, вероятно, не ранее предложенные показатели устойчивости.Ожидается, что предлагаемые индикаторы устойчивости будут обнаруживать проблемы слишком поздно, и мы предлагаем использовать их для разведения более устойчивых животных, а не использовать их в качестве предикторов предупреждений. Разведение более устойчивых животных приведет к получению потомства с меньшей дисперсией их сенсорных значений, чем у текущего поколения, и, следовательно, с меньшей вероятностью генерирования предупреждения (рис. 3).
3).
Рисунок 3 . Нормальное распределение вероятности того, что животное подаст сигнал тревоги с индивидуальным средним значением и дисперсией на основе признака, достигающего определенного порога.На этом рисунке индивидуально-специфическая дисперсия «более устойчивого животного» меньше индивидуально-специфической дисперсии «среднего животного». Следовательно, «более устойчивое животное» имеет меньшую вероятность генерировать предупреждение, чем «среднее животное».
Мы проиллюстрируем пример использования затрат на рабочую силу для получения экономической стоимости в двух ситуациях: в случае, когда рабочее время не ограничено, и, что более реалистично, в случае, когда рабочее время ограничено на ферму (т. е. максимальное количество животных при данных условиях фермы).Кроме того, мы покажем, как совокупная экономическая ценность устойчивости может быть определена на основе нескольких компонентов, в данном случае упрощенных до затрат на оплату труда и (медицинских) затрат на лечение. Для простоты мы выбираем дисперсию в качестве индикатора устойчивости для (дальнейшей) иллюстрации. Мы также предполагаем, что разведение для более низкой дисперсии уменьшит вероятность генерирования предупреждений, и что генетическая корреляция между дисперсией и устойчивостью в цели разведения равна 1. Мы предполагаем, что разведение более устойчивых животных приведет к потомству с более близкой автокорреляцией. к нулю, асимметрия ближе к нулю и наклон ближе к нулю.Во всех случаях ожидается, что потомство будет иметь более низкую вероятность генерирования предупреждения, потому что основная концепция та же, хотя она менее очевидна, чем с меньшей дисперсией.
Для простоты мы выбираем дисперсию в качестве индикатора устойчивости для (дальнейшей) иллюстрации. Мы также предполагаем, что разведение для более низкой дисперсии уменьшит вероятность генерирования предупреждений, и что генетическая корреляция между дисперсией и устойчивостью в цели разведения равна 1. Мы предполагаем, что разведение более устойчивых животных приведет к потомству с более близкой автокорреляцией. к нулю, асимметрия ближе к нулю и наклон ближе к нулю.Во всех случаях ожидается, что потомство будет иметь более низкую вероятность генерирования предупреждения, потому что основная концепция та же, хотя она менее очевидна, чем с меньшей дисперсией.
Экономическая ценность устойчивости на основе одного компонента
Если рабочее время не ограничено, экономическая ценность устойчивости представляет собой изменение ожидаемого количества оповещений, умноженное на время на оповещение, и затраты труда на единицу времени в течение всего производственного цикла индивидуума; я. д., используя уравнение 8b из Mulder et al. (2008), экономическая ценность устойчивости ( v устойчивость, неограниченное рабочее время ) составляет:
д., используя уравнение 8b из Mulder et al. (2008), экономическая ценность устойчивости ( v устойчивость, неограниченное рабочее время ) составляет:
, где z — ордината стандартного нормального распределения при стандартизованном пороге x предупреждения (например, порог, принадлежащий 1% вероятности), l a — необходимое рабочее время для работы с предупреждением, c l — стоимость труда в единицу времени, а d — количество дней периода финишера.
Если рабочее время ограничено на ферму (т. е. количество животных на ферме максимально при определенном управлении фермой), мы предполагаем, что общее доступное время ( L ) является постоянным, а среднее время, доступное на одно животное для нормального управление ( l n ) изменить нельзя. Мы также предполагаем, что среднее время на животное, необходимое для обхода с оповещениями ( L 3 r = = L × P A , с P A — вероятность получения оповещения) можно изменить путем выбора, т. е.т. е. отбор на устойчивость снижает количество предупреждений на одно животное в течение всего производственного цикла. Общая прибыль фермы:
е.т. е. отбор на устойчивость снижает количество предупреждений на одно животное в течение всего производственного цикла. Общая прибыль фермы:
где n количество животных на ферму, равное:
Переписав уравнение (2) с использованием уравнения (3), вы получите:
Прибыль=Lln+lr×(Доходы-Затраты) (4)Чтобы получить экономическую стоимость, требуется производная уравнения 4 относительно l r , равная:
dProfitdlr=-L(ln+lr)2×(Доходы-Затраты) (5)Экономическая стоимость должна быть выражена в расчете на одно животное, поэтому уравнение 5 делится на уравнение 3 для получения увеличения прибыли, когда l r изменяется за 1 единицу времени:
dProfitdlr/animal=-(Доходы-Затраты)ln+lr (6) Другими словами, уравнение (6) представляет собой изменение прибыли при изменении l r в 1 единицу времени и может быть интерпретировано как себестоимость труда, затрачиваемого на обработку предупреждений за весь период животное содержится, т. е.е., произведение c l × d в уравнении (1). Фактически, экономическая ценность показывает увеличение или уменьшение прибыли фермы из-за большей или меньшей устойчивости, потому что на ферме можно держать больше или меньше животных, если труд ограничен.
е.е., произведение c l × d в уравнении (1). Фактически, экономическая ценность показывает увеличение или уменьшение прибыли фермы из-за большей или меньшей устойчивости, потому что на ферме можно держать больше или меньше животных, если труд ограничен.
Чтобы получить экономическую ценность на основе разницы в устойчивости, уравнение 1 (неограниченное рабочее время) корректируется на основе другой себестоимости ограниченного рабочего времени в уравнении 6. В результате получается:
vустойчивость,ограниченное рабочее время= 0.5×z×x×la×-(Доходы-Затраты)ln+lr (7) Чтобы показать влияние неограниченного рабочего времени и ограниченного рабочего времени на ферму, мы рассчитали экономические значения устойчивости для фермы со свиньями на откорме. Мы предполагаем, что 8 рабочих часов в день, 15 денежных единиц в час затрат на рабочую силу, 10 денежных единиц прибыли на животное (т. е. экономическая ценность прироста), вероятность предупреждения 1% (т. е. 90 443 x 90 444 = -2,33), 5 мин. время внимания на одно предупреждение ( l a ) и 125 дней ( d ), чтобы вырасти с 25 кг до 125 кг (т.д., среднесуточный прирост 800 г/сут.). Рисунок 4 показывает, что экономическая ценность устойчивости постоянна, когда рабочее время не ограничено. Однако экономическая ценность устойчивости возрастает с увеличением размера фермы, когда рабочее время ограничено на ферму (рис. 4). Эти две ситуации приводят к равной экономической ценности, когда размер фермы составляет 1500 свиней на откорме, потому что в этом случае доход фермера составит 15 денежных единиц в час, что равно цене неограниченного труда. Экономическая ценность устойчивости, основанная на ограниченном рабочем времени, достигает более 60% экономической ценности роста (в нашем примере) при размере фермы в 2000 свиней и будет продолжать расти с увеличением размера фермы.Таким образом, повышение устойчивости животных позволит содержать больше животных на ферме (т.
е. 90 443 x 90 444 = -2,33), 5 мин. время внимания на одно предупреждение ( l a ) и 125 дней ( d ), чтобы вырасти с 25 кг до 125 кг (т.д., среднесуточный прирост 800 г/сут.). Рисунок 4 показывает, что экономическая ценность устойчивости постоянна, когда рабочее время не ограничено. Однако экономическая ценность устойчивости возрастает с увеличением размера фермы, когда рабочее время ограничено на ферму (рис. 4). Эти две ситуации приводят к равной экономической ценности, когда размер фермы составляет 1500 свиней на откорме, потому что в этом случае доход фермера составит 15 денежных единиц в час, что равно цене неограниченного труда. Экономическая ценность устойчивости, основанная на ограниченном рабочем времени, достигает более 60% экономической ценности роста (в нашем примере) при размере фермы в 2000 свиней и будет продолжать расти с увеличением размера фермы.Таким образом, повышение устойчивости животных позволит содержать больше животных на ферме (т. е. интенсификация). На самом деле, при ограниченном рабочем времени время нормального содержания ( 1 n ) на одно животное уменьшается с увеличением количества животных и постоянным количеством доступного рабочего времени. Следовательно, доля времени для предупреждений ( l r ) увеличивается, а доля времени для обычного времени ( l n ) уменьшается с размером фермы.Как следствие, увеличение или уменьшение устойчивости оказывает большее влияние на прибыль фермы, когда размер фермы увеличивается. Тем не менее, обе ситуации показывают, что экономическая ценность устойчивости будет отрицательной, а это означает, что уменьшение отклонений будет иметь положительный эффект (т.е. увеличение) на прибыль фермы, и, таким образом, устойчивость должна быть включена в цель селекции.
е. интенсификация). На самом деле, при ограниченном рабочем времени время нормального содержания ( 1 n ) на одно животное уменьшается с увеличением количества животных и постоянным количеством доступного рабочего времени. Следовательно, доля времени для предупреждений ( l r ) увеличивается, а доля времени для обычного времени ( l n ) уменьшается с размером фермы.Как следствие, увеличение или уменьшение устойчивости оказывает большее влияние на прибыль фермы, когда размер фермы увеличивается. Тем не менее, обе ситуации показывают, что экономическая ценность устойчивости будет отрицательной, а это означает, что уменьшение отклонений будет иметь положительный эффект (т.е. увеличение) на прибыль фермы, и, таким образом, устойчивость должна быть включена в цель селекции.
Рисунок 4 . Экономическая ценность устойчивости, основанная на доступном рабочем времени, неограниченном или ограниченном, для повседневного ухода за животными.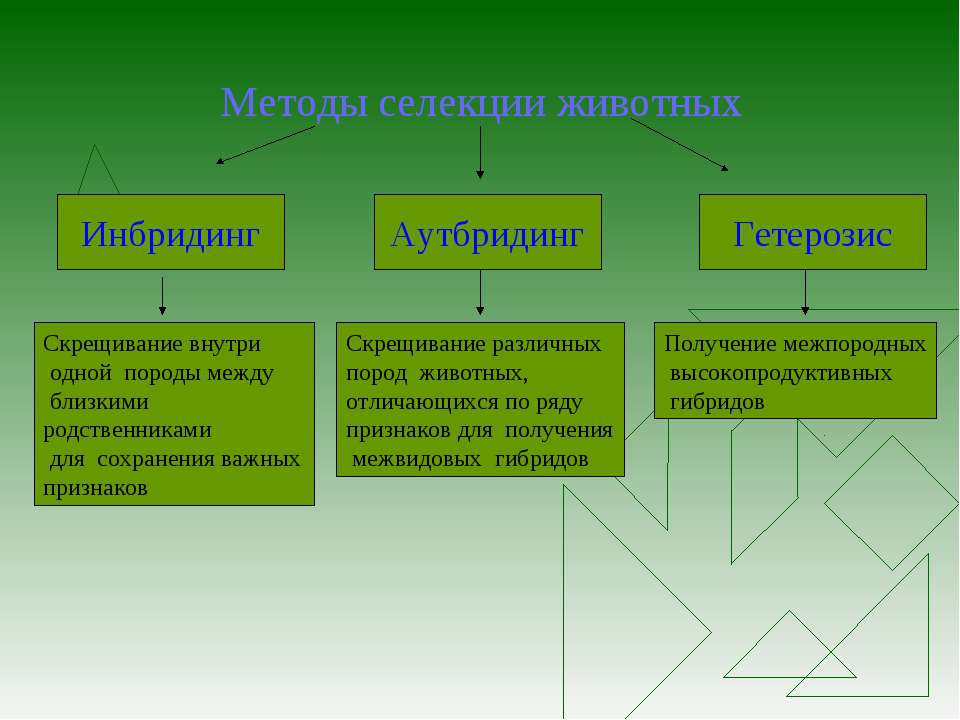 Сделаны следующие допущения: 8 рабочих часов в день, 15 денежных единиц/час затрат на оплату труда, 10 денежных единиц прибыли на животное (т. е. экономическая ценность прироста), вероятность предупреждения 1% (т. е. 90 443 x 90 444 = -2,33), 5 минут внимания на одно предупреждение ( l a ) и 125 дней ( d ) для роста с 25 кг до 125 кг (т. е. среднесуточный прирост составляет 800 г/день).
Сделаны следующие допущения: 8 рабочих часов в день, 15 денежных единиц/час затрат на оплату труда, 10 денежных единиц прибыли на животное (т. е. экономическая ценность прироста), вероятность предупреждения 1% (т. е. 90 443 x 90 444 = -2,33), 5 минут внимания на одно предупреждение ( l a ) и 125 дней ( d ) для роста с 25 кг до 125 кг (т. е. среднесуточный прирост составляет 800 г/день).
Экономическая ценность устойчивости на основе нескольких компонентов
В приведенной выше ситуации учитывались только затраты на оплату труда.Затем мы расширим экономическую ценность устойчивости, включив, например, затраты на лечение болезни, если эти затраты еще не включены в цель разведения. Экономическое значение устойчивости:
vresilience=vlabor+vtreatment (8), где v труд представляет собой экономическую стоимость затрат на оплату труда (на основе неограниченного или ограниченного рабочего времени), а v лечение представляет собой экономическую стоимость затрат на лечение, которые можно определить как:
vлечение=0.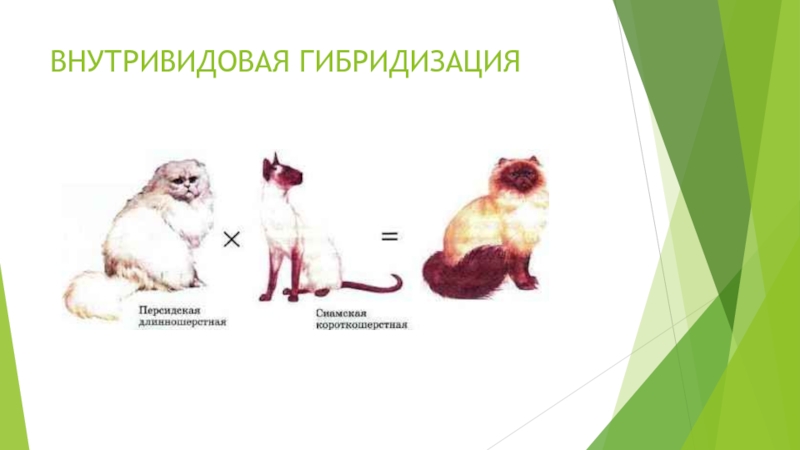 5×z×x×обработка×стоимость обработки (9)
5×z×x×обработка×стоимость обработки (9)
, где z — ордината стандартного нормального распределения при стандартизированном пороге x предупреждения (например, порог, принадлежащий 1% вероятности), p лечение — вероятность лечение при условии, что животное получило предупреждение, а Стоимость лечение — это стоимость лечения. Обратите внимание, что в этом случае затраты на лечение не должны быть частью других компонентов индекса отбора, чтобы избежать двойного учета этих затрат.Этот небольшой расширенный пример показывает, что различные компоненты, основанные на затратах, могут быть относительно легко включены в экономическую ценность устойчивости. Как показано на рис. 4, экономическая ценность устойчивости может легко достигать 60% экономической ценности роста, но эти дополнительные компоненты могут сделать экономическую ценность даже больше, чем экономическая ценность роста.
Дополнительная ценность устойчивости в программах разведения
Теперь мы продемонстрируем добавленную стоимость оценочной племенной ценности для устойчивости к разведению здоровых и простых в содержании животных для двух видов домашнего скота: (1) сценарий для свиней и (2) сценарий для молочного скота. В обоих случаях индексы отбора будут упрощены для того, чтобы сделать общие выводы. Селекция основана на усечении отбора по индексу (I) для максимизации отклика на цель разведения (H). Моделируется упрощенная схема геномики. Расчет ответов на отбор проводили в программе SelAction v2.1 (Rutten et al., 2002) с использованием принципа Dekkers (2007) для включения геномной информации.
В обоих случаях индексы отбора будут упрощены для того, чтобы сделать общие выводы. Селекция основана на усечении отбора по индексу (I) для максимизации отклика на цель разведения (H). Моделируется упрощенная схема геномики. Расчет ответов на отбор проводили в программе SelAction v2.1 (Rutten et al., 2002) с использованием принципа Dekkers (2007) для включения геномной информации.
Сценарий свинья
В этом примере мы описываем упрощенный сценарий свиней, в котором особи отбираются только по скорости роста, а цель разведения расширяется с учетом устойчивости.Мы исследуем эффект добавления индикатора устойчивости (например, дисперсии отклонений) к индексу селекции и предполагаем генетическую корреляцию 1 между индикатором устойчивости и устойчивостью в цели селекции. Предположения сделаны:
• Устойчивость имеет наследуемость 0,15 (немного более консервативно, чем Putz et al., 2018). Генетическая корреляция устойчивости и скорости роста неизвестна и будет установлена на уровне 0,25 или -0,25. Положительная генетическая корреляция указывает на то, что более низкий показатель устойчивости коррелирует с более низкой скоростью роста и , наоборот, для отрицательной корреляции.Другими словами, положительная корреляция неблагоприятна, а отрицательная корреляция благоприятна для одновременного улучшения обеих характеристик.
Положительная генетическая корреляция указывает на то, что более низкий показатель устойчивости коррелирует с более низкой скоростью роста и , наоборот, для отрицательной корреляции.Другими словами, положительная корреляция неблагоприятна, а отрицательная корреляция благоприятна для одновременного улучшения обеих характеристик.
• Отбор осуществляется при половом созревании. Отбор основан на всей доступной на данный момент информации: собственной продуктивности, BLUP, полных и полусибсах, а также геномной племенной ценности.
• Геномная племенная ценность имеет точность ~0,77, исходя из контрольной популяции в 10 000 животных и числа независимых хромосомных сегментов, равного 1 000 (Daetwyler et al., 2008).
• Экономическое значение скорости роста установлено на 1 и −1, а экономическое значение устойчивости будет варьироваться от −1000 до 1000, чтобы получить эллипс ответов выбора. Обратите внимание, что интересная часть эллипса находится там, где реакция скорости роста положительна, а реакция дисперсии отрицательна (т. е. улучшенная устойчивость).
е. улучшенная устойчивость).
Дополнительную информацию о вводе можно найти в дополнительных материалах.
На рис. 5 показана реакция отбора на скорость роста и устойчивость.Желаемое направление селекционной программы, направленное на одновременное улучшение скорости роста и устойчивости, т. е. уменьшение дисперсии, указано в правом нижнем углу каждого графика. Таким образом, в случае неблагоприятной положительной генетической корреляции между скоростью роста и резильентностью, а показатели резильентности не включаются в селекционный индекс, невозможно добиться прогресса в получении более резильентных животных (рис. 5А). Однако включение показателя устойчивости в селекционный индекс свиней может привести к более высокой реакции отбора на цель разведения (H) и более устойчивым животным, в зависимости от выбранных экономических показателей (рис. 5B).Например, при экономическом значении -0,6 для устойчивости (и экономическом значении 1 для скорости роста) реакция выбора в H улучшается на 14,6%, когда показатель устойчивости включен в индекс выбора. Хотя в этом случае наблюдается снижение селективной реакции скорости роста (-15,4%), селективная реакция устойчивости улучшается на 185,3% (см. красные кресты на рисунках 5A, B). Отсутствие индикатора устойчивости в индексе выбора увеличивает вероятность создания предупреждения до 1.16 % (начало: 1 %), а включение индикатора устойчивости в индекс выбора снижает вероятность создания предупреждения до 0,88 %. Это соответствует сокращению количества предупреждений на 24,7%. В случае благоприятной генетической корреляции между скоростью роста и устойчивостью прирост реакции отбора в H даже выше при сравнении индекса селекции без (рис. 5C) и с показателем устойчивости (рис. 5D): +42,8% для экономического значения -1,6 для устойчивости. Этот упрощенный пример показывает, что включение индикаторов устойчивости в индекс выбора может иметь большое влияние на реакцию выбора и количество предупреждений.
Хотя в этом случае наблюдается снижение селективной реакции скорости роста (-15,4%), селективная реакция устойчивости улучшается на 185,3% (см. красные кресты на рисунках 5A, B). Отсутствие индикатора устойчивости в индексе выбора увеличивает вероятность создания предупреждения до 1.16 % (начало: 1 %), а включение индикатора устойчивости в индекс выбора снижает вероятность создания предупреждения до 0,88 %. Это соответствует сокращению количества предупреждений на 24,7%. В случае благоприятной генетической корреляции между скоростью роста и устойчивостью прирост реакции отбора в H даже выше при сравнении индекса селекции без (рис. 5C) и с показателем устойчивости (рис. 5D): +42,8% для экономического значения -1,6 для устойчивости. Этот упрощенный пример показывает, что включение индикаторов устойчивости в индекс выбора может иметь большое влияние на реакцию выбора и количество предупреждений.
Рисунок 5 . Отбор ответов на скорость роста и устойчивость с экономической ценностью скорости роста, установленной на 1 и -1, и экономической ценностью устойчивости, варьирующейся от -1000 до 1000 в программе разведения свиней. На рисунке показаны эллипсы отклика скорости роста и устойчивости для индекса селекции без устойчивости (A,C) или с устойчивостью (B,D) с генетической корреляцией 0,25 (A,B) или генетической корреляция −0.25 (С,Г) . Красные кресты в A,B обсуждаются в тексте.
На рисунке показаны эллипсы отклика скорости роста и устойчивости для индекса селекции без устойчивости (A,C) или с устойчивостью (B,D) с генетической корреляцией 0,25 (A,B) или генетической корреляция −0.25 (С,Г) . Красные кресты в A,B обсуждаются в тексте.
Сценарий молочного скота
В этом примере мы описываем упрощенный сценарий молочного скота, хотя и с более сложной программой разведения, чем сценарий свиней: особи отбираются по надоям и признакам, связанным со здоровьем, которыми в данном случае являются долголетие и здоровье вымени. Мы исследуем эффект добавления индикатора устойчивости (например, дисперсии отклонений) к индексу выбора, т.е.д., выбирая по показателю устойчивости. Предположения сделаны:
• Устойчивость имеет наследуемость 0,10 (Elgersma et al., 2018). Генетическая корреляция между устойчивостью и надоем составляет 0,61, между устойчивостью и долголетием -0,30, а между устойчивостью и здоровьем вымени -0,36. Другими словами, более высокая устойчивость (т. е. меньшая дисперсия отклонений) генетически коррелирует с более низкими удоями, более высокой продолжительностью жизни и лучшим здоровьем вымени. Эти оценки были получены от Elgersma et al.(2018 г.) и CRV (2015 г.).
Другими словами, более высокая устойчивость (т. е. меньшая дисперсия отклонений) генетически коррелирует с более низкими удоями, более высокой продолжительностью жизни и лучшим здоровьем вымени. Эти оценки были получены от Elgersma et al.(2018 г.) и CRV (2015 г.).
• Отбор основан только на геномной племенной ценности. Точность геномной племенной ценности составляет ~0,79, исходя из контрольной популяции в 20 000 животных и числа независимых хромосомных сегментов, равного 1 200 (Daetwyler et al., 2008).
• Экономические значения установлены на 0,3 для надоя, 0,3 для долголетия и 0,2 для здоровья вымени. В ситуации по умолчанию экономическое значение устойчивости установлено на -0,2. Это означает, что 30 % внимания уделяется надою, 30 % — долголетию, 20 % — здоровью вымени и 20 % — устойчивости, при игнорировании корреляций между признаками (например,г., Миглиор и др., 2005). Экономическая ценность устойчивости будет варьироваться от -0,5 до -0,001 (т. е. стремление к уменьшению вариации отклонений) в качестве анализа чувствительности.
Дополнительную информацию о вводе можно найти в дополнительных материалах.
Аналогично сценарию со свиньями, включение устойчивости в цель разведения молочного скота может привести к более высокой реакции отбора в H по сравнению с отсутствием включения устойчивости в цель разведения, в зависимости от выбранной экономической ценности (Рисунок 6), даже если здоровье- родственные черты (т.е., долголетие и здоровье вымени) уже включены в цель селекции и селекционный индекс. Если устойчивость, являющаяся в данном случае дисперсией, имеет экономическое значение -0,2, реакция селекции в H увеличивается на 3,0% (см. черный крест на рис. 6): включение показателя устойчивости в индекс селекции компенсирует потерю надоев. (6,3%) за счет увеличения продолжительности жизни (1,4%), здоровья вымени (1,0%) и устойчивости (-102,6%) (таблица 2). Отсутствие индикатора устойчивости в индексе выбора снижает вероятность создания предупреждения до 0.92 % (начало: 1 %), а включение индикатора устойчивости в индекс выбора снижает вероятность создания предупреждения до 0,84 %. Это соответствует уменьшению количества предупреждений на 8,4% (красный крестик на рис. 6). Этот упрощенный пример показывает, что показатели устойчивости могут оказывать благотворное влияние. Однако эффект меньше в сценарии молочного скота по сравнению со сценарием свиней из-за наличия признаков, связанных со здоровьем, в индексе селекции.
Это соответствует уменьшению количества предупреждений на 8,4% (красный крестик на рис. 6). Этот упрощенный пример показывает, что показатели устойчивости могут оказывать благотворное влияние. Однако эффект меньше в сценарии молочного скота по сравнению со сценарием свиней из-за наличия признаков, связанных со здоровьем, в индексе селекции.
Рисунок 6 .Изменение (в %) реакции селекции в цели разведения (H) и вероятность того, что животное вызовет тревогу в индексе селекции по умолчанию без и с включением индикатора устойчивости для различных экономических значений в программе разведения молочного скота. Индекс выбора по умолчанию включает надои, продолжительность жизни и здоровье вымени. Кресты обсуждаются в тексте и подробно показаны в таблице 2.
Таблица 2 . Отклики селекции (в единицах признаков) и вероятность оповещения (%) и их относительные изменения (в %) в индексе селекции без и с включением индикатора устойчивости (экономическая ценность = -0. 2) в программе молочного скотоводства.
2) в программе молочного скотоводства.
Перспективы и другие виды домашнего скота
Потенциал устойчивости в целях разведения был четко проиллюстрирован в двух сценариях. Очевидно, что эти сценарии переоценивают влияние показателей устойчивости из-за их упрощения. Тем не менее, основная идея верна, потому что сокращение времени, затрачиваемого на животное с предупреждением (по любой причине), снизит затраты и, следовательно, увеличит прибыль фермы. Сценарий свиньи был основан только на двух признаках, но все же мог быть довольно близок к сценарию линии быка-свиньи.Программы разведения по линиям быков в первую очередь направлены на улучшение производственных признаков, в отличие от программ разведения по линиям самок, которые в большей степени ориентированы на улучшение воспроизводства и материнских качеств. Тем не менее, устойчивость маточных линий также может быть включена в программы разведения: фермам-множителям требуются выносливые свиноматки и выносливые поросята. Например, однородность размера помета оказывает благотворное влияние на устойчивость поросят в зависимости от выживаемости (Damgaard et al., 2003; Mulder et al., 2015b). Были обнаружены благоприятные корреляции между остаточной дисперсией потребления корма и продолжительностью кормления со смертностью и количеством лечебных процедур у свиней в неблагоприятных условиях (Putz et al., 2018). Это показывает, что остаточная дисперсия потребления корма и продолжительности кормления может быть использована для повышения устойчивости. Программы разведения молочного скота содержат признаки, связанные со здоровьем, которые, как ожидается, частично покрывают показатели устойчивости. Действительно, было обнаружено несколько благоприятных r g между признаками здоровья и дисперсией отклонений удоев, но ни один из них не равен 1, что указывает на то, что необработанная дисперсия удоев содержит новую информацию об устойчивости и здоровье (Elgersma et al., 2018). Это также было продемонстрировано в сценарии с молочным скотом, который показал значительное улучшение в ответ на селекционный индекс с устойчивостью. Мы предлагаем, чтобы включение устойчивости в индексы селекции программ разведения скота (аналогично свиноводству и молочному скоту) значительно увеличило бы реакцию на устойчивость скота и, вероятно, также увеличило бы реакцию селекции на H.
Например, однородность размера помета оказывает благотворное влияние на устойчивость поросят в зависимости от выживаемости (Damgaard et al., 2003; Mulder et al., 2015b). Были обнаружены благоприятные корреляции между остаточной дисперсией потребления корма и продолжительностью кормления со смертностью и количеством лечебных процедур у свиней в неблагоприятных условиях (Putz et al., 2018). Это показывает, что остаточная дисперсия потребления корма и продолжительности кормления может быть использована для повышения устойчивости. Программы разведения молочного скота содержат признаки, связанные со здоровьем, которые, как ожидается, частично покрывают показатели устойчивости. Действительно, было обнаружено несколько благоприятных r g между признаками здоровья и дисперсией отклонений удоев, но ни один из них не равен 1, что указывает на то, что необработанная дисперсия удоев содержит новую информацию об устойчивости и здоровье (Elgersma et al., 2018). Это также было продемонстрировано в сценарии с молочным скотом, который показал значительное улучшение в ответ на селекционный индекс с устойчивостью. Мы предлагаем, чтобы включение устойчивости в индексы селекции программ разведения скота (аналогично свиноводству и молочному скоту) значительно увеличило бы реакцию на устойчивость скота и, вероятно, также увеличило бы реакцию селекции на H.
Мы не исследовали устойчивость в программах разведения другого домашнего скота, такого как домашняя птица, экстенсивно содержащиеся виды скота (например,например, мясной крупный рогатый скот и овцы), или виды аквакультуры, потому что устойчивость в настоящее время труднее оценить и применить на практике. Во-первых, потому что повторные измерения сложно собрать. В основном это связано с невозможностью индивидуального измерения животных, из-за группового содержания или трудностей отлова особей. Во-вторых, трудозатраты на оповещения менее актуальны. Оповещения создаются для некоторых видов скота на основе групповых измерений (например, потребления воды), но не на индивидуальном уровне.Кроме того, предупреждения такого типа представляют собой эпидемические нарушения, а не ежедневные эндемические нарушения. Однако показатели устойчивости, основанные на относительно ограниченном наборе производственных данных (как о частоте повторных наблюдений, так и о количестве животных), уже могут предоставить ценную информацию о здоровье животных (например, 4-недельные отклонения массы тела; Berghof et al., в процессе подготовки). ), который в настоящее время не включен в селекционные индексы. Что еще более важно, разработка новых методов в ближайшем будущем позволит собирать ежедневные наблюдения на индивидуальном уровне, такие как отдельные гнезда несушек, индивидуальные измерения рыбы (без отлова), автоматический сбор данных, а также индивидуальное отслеживание и измерения с помощью камеры. или дроны.Это может привести к сбору больших данных и определению новых фенотипов и, в конечном счете, к использованию индикаторов устойчивости в программах селекции для всех видов домашнего скота.
Заключение
В этом документе показано, что включение устойчивости в программы разведения имеет большой потенциал для получения здорового и легкого в управлении скота. Показатели устойчивости могут быть основаны на отклонениях между наблюдаемым производством и ожидаемым производством. Особый интерес представляют дисперсия отклонений, автокорреляция отклонений и асимметрия отклонений.Также наклон нормы реакции может содержать информацию, хотя и ограниченную макросредовыми возмущениями. Экономическое значение показателей устойчивости в селекционном индексе может быть определено на основе приведенных затрат на рабочую силу и затраты на здоровье при условии, что эти затраты не учитываются в других признаках в селекционных индексах. Для большинства ферм, где рабочее время ограничено, экономическая ценность устойчивости возрастает с увеличением количества животных на ферме. В этой статье также показано дополнительное преимущество включения устойчивости в цель разведения: как в сценариях свиней, так и в сценариях молочного скота мы показываем улучшение реакции селекции в целях разведения и, в частности, повышение устойчивости за счет включения устойчивости в цель разведения.Ожидается, что быстрое технологическое развитие массового сбора данных (т. е. больших данных) в ближайшем будущем только усилится, в результате чего будет доступно больше данных. Сопутствующие возможности использования этих данных для определения показателей устойчивости значительно облегчат селекцию для повышения устойчивости всех видов домашнего скота.
Вклад авторов
ТБ и HM задумали проект. TB, MP и HM разработали идеи измерения устойчивости. HM рассчитал экономическую ценность устойчивости и разработал сценарии размножения.ТБ выполнил моделирование в SelAction. ТБ написал (первый черновик) статьи. MP и HM внесли свой вклад в написание статьи.
Финансирование
Это исследование было проведено при финансовой поддержке Нидерландской организации научных исследований Земли и наук о жизни (NWO-ALW; проект ALWSA.2016.4), а также Министерства экономики Нидерландов (проект TKI Agri & Food 16022) и партнеров Breed4Food Cobb Europe, CRV, Hendrix Genetics и Topigs Norsvin.
Заявление о конфликте интересов
Это исследование было частично профинансировано партнерами Breed4Food Cobb Europe, CRV, Hendrix Genetics и Topigs Norsvin.За исключением финансового вклада, между партнерами Breed4Food и авторами не существует никаких других общих интересов (например, трудоустройство, консультации, патенты, продукты). Этот документ не обсуждался и не рецензировался ни одним из партнеров Breed4Food.
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fgene.2018.00692/full#supplementary-material
Ссылки
Бохманова, Дж., Мишталь, И., и Коул, Дж. Б. (2007). Температурно-влажностные показатели как индикаторы потерь молочной продуктивности при тепловом стрессе. J. Dairy Sci . 90, 1947–1956. doi: 10.3168/jds.2006-513
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Карабаньо М. Дж., Рамон М., Диас С., Молина А., Перес-Гусман М. Д. и Серрадилья Дж.М. (2017). СИМПОЗИУМ ПО СЕЛЕКЦИИ И ГЕНЕТИКЕ: Разведение на устойчивость к воздействию теплового стресса у молочных жвачных животных. Всесторонний обзор . Дж. Аним. Наука . 95, 1813–1826 гг. дои: 10.2527/jas2016.1114
Полнотекстовая перекрестная ссылка
Кодреа, М.К., Хойсгаард, С., и Фриггенс, Северная Каролина (2011). Дифференциальное сглаживание измерений временных рядов для выявления нарушений продуктивности и количественной оценки характеристик реакции животных: пример использования профилей надоев у молочных коров. Дж.Аним. Наука . 89, 3089–3098. doi: 10.2527/jas.2010-3753
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кольдиц, И. Г., и Хайн, Британская Колумбия (2016). Устойчивость сельскохозяйственных животных: биология, управление, разведение и последствия для благополучия животных. Аним. Произв. Наука . 56, 1961–1983. дои: 10.1071/AN15297
Полнотекстовая перекрестная ссылка | Академия Google
Daetwyler, H.D., Villanueva, B., and Woolliams, J.A. (2008). Точность прогнозирования генетического риска заболевания с использованием полногеномного подхода. PLoS ONE 3:e3395. doi: 10.1371/journal.pone.0003395
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Damgaard, L.H., Rydhmer, L., Løvendahl, P., and Grandinson, K. (2003). Генетические параметры внутрипометной изменчивости веса поросят при рождении и изменения внутрипометной изменчивости во время подсоса. Дж. Аним. Наука . 81, 604–610. дои: 10.2527/2003.813604x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Де Йонг, Г.и Биджма, П. (2002). Отбор и фенотипическая пластичность в эволюционной биологии и животноводстве. Прямой эфир. Наука . 78, 195–214. doi: 10.1016/S0301-6226(02)00096-9
Полнотекстовая перекрестная ссылка | Академия Google
Дебат, В., и Дэвид, П. (2001). Картирование фенотипов: канализация, пластичность и стабильность развития. Тренды Экол. Эвол . 16, 555–561. doi: 10.1016/S0169-5347(01)02266-2
Полнотекстовая перекрестная ссылка | Академия Google
Деккерс, Дж.CM (2007). Прогнозирование реакции на маркерную и геномную селекцию с использованием теории индекса селекции. Дж. Аним. Порода. Гене . 124, 331–341. doi: 10.1111/j.1439-0388.2007.00701.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дошл-Уилсон, А.Б., Вильянуэва, Б., и Кириазакис, И. (2012). Первый шаг к генетической селекции на толерантность хозяина к инфекционным патогенам: получение фенотипа толерантности посредством групповых оценок. Перед.Жене. 3:265. doi: 10.3389/fgene.2012.00265
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Элгерсма Г.Г., Де Йонг Г., Ван дер Линде Р. и Малдер Х.А. (2018). Колебания надоев являются наследственными и могут использоваться в качестве индикатора устойчивости для разведения здоровых коров. Дж. Молочная. Наука . 101, 1240–1250. doi: 10.3168/jds.2017-13270
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фальконер, Д. С., и Маккей, Т.ФК (1996). Введение в количественную генетику (четвертое издание) . Харлоу: Longman Group Ltd.
Академия Google
Финли, К.В., и Уилкинсон, Г.Н. (1963). Анализ адаптации в селекционной программе. австр. Дж. Агрик. Рез . 14, 742–754. дои: 10.1071/AR9630742
Полнотекстовая перекрестная ссылка | Академия Google
Ge, L., Anten, N.P.R., Van Dixhoorn, I.D.E., Feindt, P.H., Kramer, K., Leemans, R., et al. (2016). Почему нам нужно мышление устойчивости для решения социальных проблем в производственных системах на основе биологии. Курс. мнение Окружающая среда. Поддержать . 23, 17–27. doi: 10.1016/j.cosust.2016.11.009
Полнотекстовая перекрестная ссылка | Академия Google
Gijzel, S.M.W., Van de Leemput, I.A., Scheffer, M., Roppolo, M., Olde Rikkert, MGM, and Melis, RJF (2017). Показатели динамической устойчивости во временном ряду самооценки здоровья соответствуют уровням слабости у пожилых людей. Дж. Геронтол. биол. науч. Мед. Наука . 72, 991–996. doi: 10.1093/gerona/glx065
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гримсруд, К.М., Нильсен, Х.М., Навруд, С., и Олесен, И. (2013). Готовность домохозяйств платить за улучшение благосостояния рыбы в программах разведения атлантического лосося, выращиваемого на фермах. Аквакультура 372, 19–27. doi: 10.1016/j.aquaculture.2012.10.009
Полнотекстовая перекрестная ссылка | Академия Google
Herrero-Medrano, J.M., Mathur, P.K., Ten Napel, J., Rashidi, H., Alexandri, P., Knol, E.F., et al. (2015). Оценка генетических параметров и племенной ценности в сложных условиях для отбора крепких свиней. Дж. Аним. Наука . 93, 1494–1502 гг. doi: 10.2527/jas.2014-8583
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Холлинг, К.С. (1973). Устойчивость и устойчивость экологических систем. Анну. Преподобный Экол. Эвол. Сист . 4, 1–23. doi: 10.1146/annurev.es.04.110173.000245
Полнотекстовая перекрестная ссылка | Академия Google
Ибаньес-Эскриче, Н., Варона, Л., Соренсен, Д., и Ногера, Дж. Л. (2008). Исследование неоднородности окружающей среды для убойного веса свиней. Животное 2, 19–26. дои: 10.1017/S1751731107001000
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Юнг, Л.Х.С., Невес, Х.Х.Р., Малдер, Х.А., и Карвальейро, Р. (2017). Генетический контроль остаточной дисперсии годового веса у мясного скота Неллор. Дж. Аним. Наука . 95, 1425–1433. doi: 10.2527/jas.2016.1326
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Янхунен М., Каузе А., Вехвиляйнен Х. и Ярвисало О.(2012). Генетика микросредовой чувствительности веса тела у радужной форели ( Oncorhynchus mykiss ), выбранной для улучшения роста. PLoS ONE 7:e38766. doi: 10.1371/journal.pone.0038766
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Капелл, Д.Н.Р.Г., Эшворт, С.Дж., Кнап, П.В., и Роэ, Р. (2011). Генетические параметры выживаемости поросят, размер помета и вес при рождении или его вариации в помете у линий производителей и маток с использованием байесовского анализа. Прямой эфир. Наука . 135, 215–224. doi: 10.1016/j.livsci.2010.07.005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кнап, П. В. (2009a). «Надежность», в Теория распределения ресурсов в применении к сельскохозяйственному животноводству , изд. В.М. Rauw (Уоллингфорд: CABI Publishing), 288–301.
Академия Google
Кнап, П. В. (2009b). «Добровольное потребление корма и свиноводство », в Добровольное потребление корма свиньями , ред. D.Торраллардона и Э.А. Рура (Вагенинген: Академическое издательство Вагенинген).
Марьянович, Дж., Малдер, Х.А., Рённегард, Л., и Бийма, П. (2018). Моделирование коэволюции косвенных генетических эффектов и наследственной изменчивости. Наследственность 121, 631–647. doi: 10.1038/s41437-018-0068-z
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мильор, Ф., Флеминг, А., Мальчиоди, Ф., Брито, Л. Ф., Мартин, П., и Баес, К. Ф. (2017). 100-летний обзор: идентификация и генетический отбор экономически важных признаков молочного скота. Дж. Молочная. Наука . 100, 10251–10271. doi: 10.3168/jds.2017-12968
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Miglior, F., Muir, B.L., and Van Doormaal, B.J. (2005). Селекционные показатели голштинского скота разных стран. J. Dairy Sci . 88, 1255–1263. doi: 10.3168/jds.S0022-0302(05)72792-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Малдер, HA (2017). GxE — бремя или благословение? Возможности для геномной селекции и больших данных.Дж. Аним. Порода. Гене . 134, 435–436. doi: 10.1111/jbg.12303
Полнотекстовая перекрестная ссылка | Академия Google
Малдер, Х. А., Бийма, П., и Хилл, В. Г. (2007). Прогнозирование племенной ценности и селекционных реакций при генетической неоднородности изменчивости окружающей среды. Генетика 175, 1895–1910. doi: 10.1534/genetics.106.063743
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Малдер, Х. А., Бийма, П., и Хилл, В. Г. (2008). Отбор на однородность домашнего скота путем использования генетической неоднородности остаточной дисперсии. Жен. Сел . Эвол . 40, 37–59. doi: 10.1051/gse:2007034
Полнотекстовая перекрестная ссылка | Академия Google
Малдер, Х. А., Эрреро-Медрано, Дж. М., Селл-Кубиак, Э., Мартур, П. К., и Кнол, Э. Ф. (2015a). «Статистические модели для повышения устойчивости к болезням и однородности в животноводстве», в EAAP — 66 th Annual Meeting (Варшава)
Академия Google
Малдер, Х. А., Хилл, В. Г., и Кнол, Э.Ф. (2015б). Наследуемая изменчивость окружающей среды вызывает нелинейные отношения между признаками: приложение к весу при рождении и мертворождение свиней. Генетика 199, 1255–1269. doi: 10.1534/genetics.114.173070
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Малдер, Х. А., Рённегард, Л., Фиксе, В. Ф., Веркамп, Р. Ф., и Страндберг, Э. (2013). Оценка генетической дисперсии макро- и микросредовой чувствительности с использованием двойных иерархических обобщенных линейных моделей. Жен. Сел. Эвол . 45:23. дои: 10.1186/1297-9686-45-23
Полнотекстовая перекрестная ссылка | Академия Google
Малдер, Х. А., Вишер, Дж., и Фаблет, Дж. (2016). Оценка чистопородной генетической корреляции однородности окраски яичной скорлупы у кур-несушек. Жен. Сел. Эвол . 48:39. doi: 10.1186/s12711-016-0212-2
Полнотекстовая перекрестная ссылка | Академия Google
Нитесон-Ван Ньювенховен, А.-М., Кнап, П., и Авенданьо, С. (2013). Роль устойчивого коммерческого свиноводства и птицеводства для обеспечения продовольственной безопасности. Аним. Передний . 3, 52–57. doi: 10.2527/af.2013-0008
Полнотекстовая перекрестная ссылка | Академия Google
Невес, Х.Х.Р., Карвальейро, Р., Розо, В.М., и Кейрос, С.А. (2011). Генетическая изменчивость остаточной дисперсии продуктивных признаков у мясного скота Неллор. Прямой эфир. Наука . 142, 164–169. doi: 10.1016/j.livsci.2011.07.010
Полнотекстовая перекрестная ссылка | Академия Google
Нильсен, Х.М., и Амер, П.Р. (2007). Подход к получению экономических весов в целях разведения с использованием экспериментов по выбору частичного профиля. Животное 1, 1254–1262. дои: 10.1017/S1751731107000729
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нильсен Х.М., Олесен И., Навруд С., Колстад К. и Амер П. (2011). Как учитывать ценность сельскохозяйственных животных в племенных целях. Обзор текущего состояния и будущих задач . Дж. Агрик. Окружающая среда. Этика 24, 309–330. doi: 10.1007/s10806-010-9264-4
Полнотекстовая перекрестная ссылка
Олесен И., Джедрем Т., Бентсен Х.Б., Гьерде Б. и Рай М. (2003). Программы разведения для устойчивой аквакультуры. J. Appl. Аква. 13, 179–204. дои: 10.1300/J028v13n03_01
Полнотекстовая перекрестная ссылка | Академия Google
Putz, A.M., Harding, J.C., Dyck, M.K., Canada, P., Fortain, F., Plastow, G., et al. (2018). Новые фенотипы устойчивости с использованием данных о потреблении корма из модели заражения естественным заболеванием у свиней в период отъема до откорма. Перед. Жене. 9:660. doi: 10.3389/fgene.2018.00660
Полнотекстовая перекрестная ссылка | Академия Google
Рашиди, Х., Малдер, Х.А., Матур, П., Ван Арендонк, Дж.А.М., и Кнол, Э.Ф. (2014). Изменчивость среди свиноматок в ответ на репродуктивно-респираторный синдром свиней. Дж. Аним. Наука . 92, 95–105. doi: 10.2527/jas.2013-6889
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Руттен, М. Дж. М., Бийма, П., Вуллиамс, Дж. А., и Ван Арендонк, Дж. А. М. (2002). SelAction: программное обеспечение для прогнозирования реакции селекции и уровня инбридинга в программах разведения скота. Дж.Здесь . 93, 456–458. doi: 10.1093/jhered/93.6.456
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сае-Лим, П., Каузе, А., Янхунен, М., Вехвиляйнен, Х., Коскинен, Х., Гьерде, Б., и другие. (2015). Генетическая (ко)изменчивость массы тела радужной форели ( Oncorhynchus mykiss ) и ее однородность в производственных условиях. Жен. Сел. Эвол . 47:46. doi: 10.1186/s12711-015-0122-8
Полнотекстовая перекрестная ссылка | Академия Google
Сае-Лим, П., Каузе А., Лиллехаммер М. и Малдер Х.А. (2017). Оценка племенной ценности однородности роста атлантического лосося ( Salmo salar ) с использованием родословных или одноэтапной геномной оценки. Жен. Сел. Эвол . 49:33. doi: 10.1186/s12711-017-0308-3
Полнотекстовая перекрестная ссылка | Академия Google
Scheffer, M., Bolhuis, J.E., Borsboom, D., Buchman, T.G., Gijzel, S.M.W., Goulson, D., et al. (2018). Количественная оценка устойчивости человека и других животных. Проц. Натл. акад. науч. США . 115, 11883–11890. doi: 10.1073/pnas.1810630115
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шеффер М., Карпентер С. Р., Дакос В. и Ван Нес Э. Х. (2015). Общие индикаторы экологической устойчивости: определение вероятности критического перехода. Анну. Преподобный Экол. Эвол. Сист . 46, 145–167. doi: 10.1146/annurev-ecolsys-112414-054242
Полнотекстовая перекрестная ссылка | Академия Google
Селл-Кубяк, Э., Bijma, P., Knol, E.F., and Mulder, H.A. (2015). Сравнение методов изучения однородности признаков: приложение к весу свиней при рождении. Дж. Аним. Наука . 93, 900–911. doi: 10.2527/jas.2014-8313
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ван дер Ваайдж, Э. Х., Бийма, П., Бишоп, С. К., и Ван Арендонк, Дж. А. М. (2000). Моделирование отбора по производственным признакам в условиях постоянного инфекционного давления. Дж. Аним. Наука . 78, 2809–2820. дои: 10.2527/2000.78112809x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Westneat, D.F., Wright, J., and Dingemanse, N.J. (2015). Биология скрыта внутри остаточной внутрииндивидуальной фенотипической изменчивости. биол. Преподобный Кэмб. Филос. Соц . 90, 729–743. doi: 10.1111/brv.12131
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
лигеров, тигонов и гроларов, о боже! Гибридизация и ее влияние на биоразнообразие · Frontiers for Young Minds
Аннотация
Вы когда-нибудь слышали о лигерах и тигонах, помесях львов и тигров? Волкособы, помесь волков и собак? Или, может быть, помесь гризли и белого медведя? Когда организмы двух разных видов смешиваются или размножаются вместе, это называется гибридизацией.Потомство, полученное от этих смесей, известно как гибриды. Гибриды встречаются в естественном мире и представляют собой мощную эволюционную силу. Они также важны в нашей повседневной жизни — вы, вероятно, едите гибридные растения каждый день. В этой статье мы погружаемся в захватывающий мир гибридизации, описывая, как это происходит и что может произойти, когда у гибридов появляются дети.
Что такое вид?
Гибридизация – это скрещивание двух разных видов [1]. Итак, чтобы заглянуть в мир гибридизации, мы должны сначала понять, что такое вид.Организмы одного вида больше похожи друг на друга, чем на организмы разных видов. Некоторые виды легко отличить друг от друга, например бегемот явно отличается от гепарда (см. рисунок 1). Но в чем разница между гепардом и леопардом? Это тоже разные виды, но леопард внешне очень похож на гепарда. Оба живут в Африке, хищники, кошки, и у обоих даже есть пятна. Наиболее часто используемые правила деления организмов на виды называются концепцией биологических видов [2].Эти правила считают животных разными видами, если они не могут размножаться вместе или если они размножаются вместе и производят бесплодное потомство, то есть потомство, которое не может иметь собственных детенышей. Поскольку гепард и леопард не могут размножаться вместе, мы считаем их двумя разными видами. Другие правила, разделяющие сходных животных или растения на разные виды, вызывают споры. Некоторые ученые ищут физические различия, например, различия в форме клюва, цвете тела, поведении, среде обитания или географическом положении.Другие ученые используют различия в генах, чтобы найти разные виды. У каждого живого организма есть гены, которые содержатся в ДНК и содержат информацию, которая сообщает телу, как работать. Внутри вида будут небольшие различия в генах, называемые мутациями. Именно такие мутации вызывают небольшие различия внутри вида, например, разные цвета глаз у людей. Мутации даже определяют, можете ли вы сворачивать язык или нет! Между видами гораздо больше мутаций между генами.Именно мутации вызывают различия в размере клюва или в поведении, которые мы видим. Если ученые не уверены, являются ли два организма разными видами, они могут сравнить и подсчитать мутации, чтобы проверить.
- Рис. 1. Гепард и бегемот — два разных вида.
- Оба живут на африканском континенте, но гиппопотамы живут в воде и болотах, а гепарды ненавидят сырость и живут на африканских лугах. Бегемот — травоядное, а гепард — хищник.Эти два вида не могут скрещиваться.
Что такое гибриды?
Когда два животных одного вида спариваются, их потомство получает 50% своих генов от каждого родителя. Это то, что делает вас похожим на смесь ваших родителей. Гибриды представляют собой скрещивания двух разных видов, поэтому они содержат 50% генов от каждого родительского вида [1]. Известным гибридом является мул, помесь осла и лошади. Пятьдесят процентов генов мула исходят от лошади и 50 процентов — от осла.Из-за этого смешения мулы имеют черты каждого родительского вида и сильны, как ослы, а также умны, как лошади [3]. Фермеры разводят мулов, потому что эта комбинация делает мулов превосходными для перевозки припасов. Использование гибридизации для объединения желаемых аспектов каждого родительского вида очень полезно для людей, и гибриды часто используются в сельском хозяйстве. Многие из вкусных фруктов, которые вы покупаете в продуктовом магазине, были созданы путем гибридизации! Бананы, грейпфруты, морковь и огурцы — гибридные виды.На самом деле существуют сотни сортов бананов, но большинство из нас знакомы с бананом-гибридом. Фермеры продолжали смешивать сорта бананов, чтобы создать идеальное сочетание мягких, вкусных фруктов без слишком большого количества семян [4].
Могут ли гибриды иметь детей?
Мулы и бананы являются примерами бесплодных гибридов, поэтому они не могут иметь собственных детенышей. Но удивительно, что есть много примеров гибридов, которые действительно могут иметь детей. Это происходит, когда гибрид спаривается с другим гибридом или с тем же видом, что и один из его родителей.Например, когда львы и тигры скрещиваются, получается лигр. Лигры плодовиты и могут спариваться с другими лигерами, львами или тиграми. Плодовитые гибриды создают очень сложную проблему в науке, поскольку это нарушает правило Концепции биологических видов, согласно которому два отдельных вида не должны иметь возможности размножаться и давать плодовитое потомство. Означает ли это, что родители этих плодовитых гибридов не являются отдельными видами? Нет, это просто означает, что концепция биологического вида подходит не для каждого вида.Благодаря открытию того, что некоторые гибриды способны к размножению, ученые продолжают спорить о том, что такое вид, и, вероятно, будут делать это еще много лет. Именно это делает гибридизацию такой интересной — она бросает вызов некоторым из наших основных научных идей [1].
Когда гибриды спариваются с любым из их родительских видов, их потомство известно как гибриды обратного скрещивания [1]. На рисунках 2A, B мы видим лигра, гибрид льва и тигра, который спаривался с тигром. Ребенок от этой смеси, гибрид обратного скрещивания, все еще имеет некоторые львиные гены.Если обратное скрещивание продолжается в течение многих поколений (обратное скрещивание спаривается с тигром, затем его потомство делает то же самое), процент львиных генов будет становиться все меньше и меньше, но они не теряются полностью. Это означает, что гены льва могут в конечном итоге стать частью генофонда тигра. Когда один вид содержит некоторые гены другого вида, это называется интрогрессией. Это мощная эволюционная сила, потому что эти новые гены могут кодировать новые черты или поведение, которые могут помочь родительским видам [5].
- Рис. 2. (A) В левом верхнем углу лев и тигр спариваются для получения гибрида.
- Этот гибрид льва и тигра имеет 50% генов от льва и 50% от тигра. Если затем гибрид спаривается с тигром (внизу слева), это потомство, называемое гибридом обратного скрещивания, будет иметь меньший процент львиных генов. (A) Это мультфильм, и в реальности лигеры и тигоны смешаны по всему телу, как мы видим в (B) . Это пример двух лигров, живущих в зоопарке.
Какое влияние гибридизация оказывает на мир природы?
До сих пор мы говорили только о гибридах, созданных людьми. Львы и тигры никогда не встречаются естественным образом в дикой природе, но другие гибриды встречаются в природе. На самом деле в мире природы существуют сотни гибридов. Считается, что каждый четвертый вид растений и каждый десятый вид животных гибридизуются [6]. Гибридизация может помочь родительским видам путем передачи новых генов посредством интрогрессии и даже может привести к созданию новых видов [5].Например, южноамериканские бабочки Heliconius приобрели часть своих красивых рисунков крыльев в результате гибридизации (рис. 3) [7]. Бабочки Heliconius используют узоры своих крыльев, чтобы привлекать партнеров, а также избегать хищников, которые интерпретируют узоры как предупреждающие сигналы. [7]. Древняя гибридизация видов подсолнечника также привела к появлению новых видов в Северной Америке. Эти подсолнухи гибридного происхождения могут жить в более экстремальных условиях, где почва бедна или токсична.Гибридизация объединила черты двух родительских видов, образовав у гибрида новую комбинацию генов, которая позволила ему жить в этой новой среде обитания [8].
- Рисунок 3. Крылья трех бабочек Heliconious.
- На верхней панели показан гибрид двух видов бабочек на панелях под ней, поэтому узоры крыльев гибрида представляют собой смесь двух родителей. Эта гибридизация может быть выгодной, потому что новые узоры крыльев могут привлекать партнеров, но она также может быть и невыгодной, потому что некоторые узоры крыльев могут сделать бабочек более заметными для хищников.Изображения взяты с сайта heliconius.ecdb.io.
Хотя многие из естественных гибридов, о которых мы говорили, произошли от современных видов, есть и примеры древних гибридизаций, произошедших десятки тысяч лет назад. Эти гибриды могут быть идентифицированы даже тогда, когда родительские виды вымерли. Это связано с тем, что некоторые гены родительского вида все еще будут присутствовать в небольшом проценте в гибриде. Сравнивая генные мутации между близкородственными видами, мы можем найти потенциальных гибридов, ища очень разные гены или мутации, которые произошли от одного из родительских видов древнего гибрида.С помощью этого метода было обнаружено, что древний гибрид является предком многих видов рыб-клоунов (например, Немо из В поисках Немо ). Как и у подсолнуха, сочетание адаптаций у этого древнего гибрида позволило предку рыбы-клоуна жить в новой среде обитания [9]. В результате этот древний гибрид является предком многих современных видов рыб-клоунов.
Иногда гибриды могут быть вредны для родительских видов и для мира природы. Если гибриды будут очень успешными, их может быть так много, что они будут конкурировать со своими родительскими видами за пищу и жизненное пространство, что может привести к исчезновению родительских видов.Потеря вида плохо сказывается на биоразнообразии и может повлиять на другие виды в этой среде обитания. Когда исчезновение этого вида происходит естественным образом, ученые не пытаются его остановить, потому что это естественный процесс. Потеря родительского вида из-за гибридного потомства проблематична только тогда, когда гибрид создан людьми и завезен в район, где родительские виды не встречались естественным путем. В таких случаях мы должны действовать, чтобы предотвратить исчезновение родительских видов. Но не волнуйтесь, гибриды, которые мы покупаем в продуктовом магазине, вряд ли нанесут серьезный вред окружающей среде, потому что существуют правила, гарантирующие их выращивание с большой осторожностью.
Заключение
Гибридизация — это сложный процесс, включающий смешение двух видов. Гибридизация является важной частью эволюции из-за передачи генов посредством интрогрессии и ее роли в создании новых видов. Он также является частью нашей повседневной жизни и используется для улучшения продуктов питания и домашнего скота.
Глоссарий
Гибридизация : ↑ Скрещивание двух разных видов.
Виды : ↑ Организмы, которые похожи друг на друга и могут размножаться вместе, чтобы произвести плодовитое потомство.Не все ученые согласны с тем, как определить, что такое отдельные виды.
Гены : ↑ Участки ДНК, содержащие инструкции для процессов и характеристик организма (например, цвет глаз).
Мутации : ↑ Небольшие различия в генах, которые делают людей уникальными. Между видами происходит множество мутаций.
Плодородный : ↑ Способен воспроизводить потомство и иметь детей. Бесплодие — это наоборот, что означает неспособность иметь детей.
Интрогрессия : ↑ Когда гены одного вида передаются другому виду путем гибридизации и обратного скрещивания.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Каталожные номера
[1] ↑ Аллендорф, Ф. В., и Люкарт, Г.(ред.) 2007. «Гибридизация», в Сохранение и генетика популяций (Малден, Массачусетс: Паб Блэквелл). 421–48.
[2] ↑ Mayr, E. 2000. «Концепция биологических видов», в Species Concepts and Phylogenetic Theory: A Debate , eds QD Wheeler and R. Meier (New York, NY: Columbia University Press). 17–20.
[3] ↑ Proops, L., Burden, F., and Osthaus, B. 2009. Познание мула: случай гибридной силы? Аним.Познан. 12:75–84. doi: 10.1007/s10071-008-0172-1
[4] ↑ Perrier, X., De Langhe, E., Donohue, M., Lentfer, C., Vrydaghs, L., Bakry, F., et al. 2011. Междисциплинарные взгляды на одомашнивание бананов ( Musa spp.). Проц. Натл. акад. науч. США 108:11311–8. doi: 10.1073/pnas.1102001108
[5] ↑ Арнольд М.Л., Сепир Ю. и Мартин Н.Х. 2008. Генетический обмен и происхождение адаптаций: от прокариот к приматам. Филос. Транс. Р. Соц. Лонд. Б биол. науч. 363:2813–20. doi: 10.1098/rstb.2008.0021
[6] ↑ Маллет, Дж. 2005. Гибридизация как вторжение в геном. Тренды Экол. Эвол. 20: 229–37. doi: 10.1016/j.tree.2005.02.010
[7] ↑ Кронфорст, М. Р., Янг, Л. Г., Блюм, Л. М., и Гилберт, Л. Э. 2006. Мультилокусный анализ примеси и интрогрессии среди гибридизирующихся бабочек Heliconius . Эволюция 60:1254–68.дои: 10.1554/06-005.1
[8] ↑ Ризеберг Л. Х., Раймонд О., Розенталь Д. М., Лай З., Ливингстон К., Наказато Т. и др. 2003. Основные экологические изменения дикорастущих подсолнухов, которым способствует гибридизация. Наука 301:1211–6. doi: 10.1126/science.1086949
[9] ↑ Лициос Г. и Саламин Н. 2014. Гибридизация и диверсификация адаптивной радиации рыб-клоунов. BMC Evol. биол. 14:245. дои: 10.1186/с12862-014-0245-5
|
Селекционеры животных и растений в современном мире стремятся получить организмы, которые будут обладать желаемыми характеристиками, такими как высокая урожайность, устойчивость к болезням, высокая скорость роста и многие другие фенотипические характеристики. Этот результат обычно достигается путем скрещивания двух представителей одного и того же вида, обладающих доминантными аллелями определенных генов, таких как долголетие и быстрый метаболизм, в одном организме, скрещенных с другим организмом, обладающим генами быстрого роста и высокой продуктивности.Поскольку оба этих организма имеют доминантные гены для этих желательных характеристик, при скрещивании они дадут по крайней мере некоторое потомство, которое проявит ВСЕ эти желательные характеристики. Когда происходит такое скрещивание, потомство называется гибридом, полученным от двух генетически разных родителей, которые обычно производят потомство с более желательными качествами. Потомство гетерозиготно, то есть ген каждой характеристики будет иметь один доминантный и один рецессивный аллель.У большинства профессиональных селекционеров имеется настоящее селекционное скрещивание (т. е. AAbb с AAbb ), так что они создают банк генов этих качеств, который можно скрещивать с aaBB для получения гетерозиготного потомства. Таким образом, доминантные признаки сохраняются в первой племенной группе и могут быть переданы потомству. Этот процесс выбора родителей называется искусственным отбором или селекционным разведением. Нижняя сторона Программы селективного разведения, которые концентрируются на небольшом подмножестве генов, определяющих ограниченную группу желаемых характеристик, часто используют небольшое количество особей в качестве «популяции-основателя» для всех потомков.Несмотря на то, что большое внимание уделяется получению особей, гомозиготных по желаемым характеристикам, существует множество других генов, которые также оказываются гомозиготными в небольшой группе инбредных основателей. Некоторые из этих генов имеют пагубные последствия, и их неблагоприятные условия, как правило, непропорционально часто встречаются у членов популяций, происходящих от небольшой группы основателей. В больших популяциях перекрестное скрещивание и естественная гибридизация приводят к гетерозиготам, в которых мутантный вредный рецессивный аллель маскируется нормальным доминантным аллелем. Примеры среди пород собак включают рак собак, болезни сердца, дисплазию тазобедренного сустава, аномалии зрения и слуха. Евреи-ашкенази демонстрируют большую долю заболеваемости болезнью Тея-Сакса и муковисцидозом, среди других состояний, чем население в целом. Считается, что высокий уровень инбридинга или кровнородственных браков в некоторых арабских группах способствует возникновению врожденных пороков развития, которые являются второй по значимости причиной младенческой смертности в Бахрейне, Кувейте, Омане и Катаре и являются основной причиной младенческой смертности (40.3%) в Объединенных Арабских Эмиратах. Прослеживание родословных до групп древних людей, вышедших из Африки и затем распространившихся по миру, также зависит от наличия определенных мутаций у всех членов группы и их потомков. Пакт с Дьяволом Искусственный отбор/селекционное разведение привело к повышению эффективности животных и растений, которых мы разводим, например, к увеличению надоев молока от коров за счет постоянного скрещивания отобранных коров друг с другом для получения гибридный. К сожалению, многие из новых пород сильно зависят от человеческой деятельности для повышения уровня продуктивности.Им требуется много ископаемого топлива, искусственных удобрений, орошения, гербицидов и пестицидов. Воздействие на окружающую среду чрезвычайно велико. Кроме того, обращение с некоторыми сельскохозяйственными животными в условиях промышленного производства значительно меняет качество жизни одомашненных видов. Нынешнее население и его неумолимый рост зависят от улучшения сельского хозяйства для борьбы с голодом и голодом. 
|