Черви плоские
Плоские черви. Плоские черви — это животные с двусторонней симметрией тела. Тело сплюснуто в спинно-брюшном направлении, поэтому внешне они похожи на лист, пластинку или ленту. Значительная часть плоских червей — это свободноживущие обитатели морских и пресных водоемов, влажных почв, но есть и большое количество паразитических форм.[ …]
Плоские черви являются первичноротыми животными. Они характеризуются также отсутствием метамерии, скелетной, сосудистой и дыхательной систем.[ …]
Продукты выделения поступают в цитоплазму пламенных клеток, а затем в канальцы, объединяющиеся и открывающиеся наружу в виде пор. Через эти поры продукты выделения и выходят в окружающую среду.[ …]
У плоских червей концентрация нервных клеток завершилась формированием парных головных нервных узлов с отходящими от них нервными волокнами к органам чувств и парных нервных стволов, проходящих по телу от головной части к каудальной. У круглых червей уже встречается окологлоточное нервное кольцо, образуемое за счет слияния головных нервных узлов, а у кольчатых червей даже развивается нервная цепочка за счет образования парных нервных узлов в сегментах тела.
Тип Плоские черви (Р1а1Ье1т1п1Ье8). К этому типу относят животных, характеризующихся вытянутой уплощенной билатерально-симметричной формой и обитающих в воде, почве, организме растений, животных и человека. Они составляют один из наиболее больших по численности типов животных (около 9000 видов).[ …]
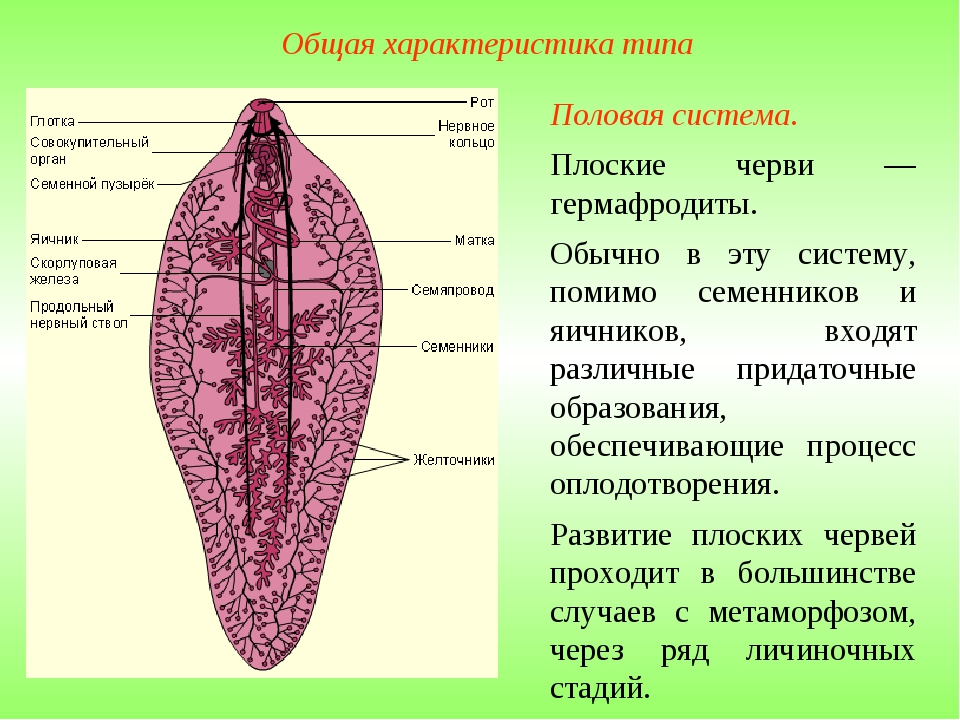
Все плоские черви — трехслойные животные (рис. 75). У них имеется кожно-мускульный мешок, который образует покров и мускулатуру тела. Появляются выделительная и пищеварительная системы. Нервная система состоит из двух нервных узлов и нервных стволов. У свободноживущих червей есть глаза и осязательные лопасти. Все плоские черви — гермафродиты, откладывают яйца в кокон. Плоских червей подразделяют на ресничных, ленточных и сосальщиков.[ …]
Тип Плоские черви — Plathelmintes представлен одним видом реснитчатых червей только в Красной книге Республики Татарстан (1995 г.).[ …]
Для плоских червей характерно развитие с неполным метаморфозом. Их личиночные стадии являются мелкими организмами, не способными к восприятию пищи, а также с ограниченной локомоторной способностью и временем жизни. У некоторых видов лишь один экземпляр на 1 млн особей является удачливым в переживании. Поэтому переживание паразитических видов обеспечивается, возможно, лишь их гигантской репродуктивной способностью.[ …]
У некоторых видов лишь один экземпляр на 1 млн особей является удачливым в переживании. Поэтому переживание паразитических видов обеспечивается, возможно, лишь их гигантской репродуктивной способностью.[ …]
Размеры плоских червей в длину чрезвычайно варьируются — от 0,5 мм до 30 м. У одних плоских червей поверхностный (покровный) эпителий снабжен ресничками или даже жгутиками, у других он характеризуется тем, что ядра и цитоплазма смещаются в глубь тела, формируя так называемый погружной эпителий (те-гумент). Покровный эпителий и располагающаяся под ним кожная мускулатура формируют кожно-мускульный мешок. Пространство между мешком и внутренними органами заполнено паренхимой, являющейся производной мезодермы. Пищеварительная система у некоторых плоских червей представлена ротовым отверстием, глоткой, пищеводом и кишкой, а у некоторых (у ленточных) ее совсем нет. Все плоские черви гермафродиты.[ …]
Дождевой червь. Водится в садах, парках, на огородах, чаще всего в жирном, хорошо обработанном грунте. Вырастают выползки до 20 см длиной и 9—12 мм толщиной. Окраска головной части у них обычно бурая или красная, хвостовой — более светлого оттенка. Хвост у выползка плоский. Их можно собирать ночью, вооружившись фонарем. Иногда выползки не покидают своих нор, а лишь высовывают из норы туловище до половины длины. При попытках вытащить червя из норы он обрывается. В этом случае следует ухватить червя и, слегка потянув к себе, выждать несколько секунд. После этого, ослабевший, он легко вытаскивается. Собранных червей лучше держать с тем навозом, из которого они были добыты.[ …]
Вырастают выползки до 20 см длиной и 9—12 мм толщиной. Окраска головной части у них обычно бурая или красная, хвостовой — более светлого оттенка. Хвост у выползка плоский. Их можно собирать ночью, вооружившись фонарем. Иногда выползки не покидают своих нор, а лишь высовывают из норы туловище до половины длины. При попытках вытащить червя из норы он обрывается. В этом случае следует ухватить червя и, слегка потянув к себе, выждать несколько секунд. После этого, ослабевший, он легко вытаскивается. Собранных червей лучше держать с тем навозом, из которого они были добыты.[ …]
Тегумент плоских червей, главным образом цестод, играет исключительную роль в физиологии питания и имеет большое значение для изучения биофизических и биохимических взаимоотношений паразит — хозяин.[ …]
У ресничных и плоских червей строение пищеварительной системы еще сохраняет сходство с пищеварительной системой кишечнополостных. Анального отверстия нет, поэтому выброс пищеварительных остатков происходит через рот. Но у круглых червей пищеварительная система уже несколько усложняется, поскольку пищеварительная трубка заметно подразделяется на передний, средний и задний отделы, из которых передний и задний отделы имеют эктодермальное происхождение, средний — энтодермальное. Впервые развивается анальное отверстие за счет впячивания эктодермы в каудальную часть тела и соединения ее с полостью первичной кишки. У кольчатых червей пищеварительный тракт уже представлен глоткой, пищеводом, зобом, желудком, средней и задней кишками.[ …]
Но у круглых червей пищеварительная система уже несколько усложняется, поскольку пищеварительная трубка заметно подразделяется на передний, средний и задний отделы, из которых передний и задний отделы имеют эктодермальное происхождение, средний — энтодермальное. Впервые развивается анальное отверстие за счет впячивания эктодермы в каудальную часть тела и соединения ее с полостью первичной кишки. У кольчатых червей пищеварительный тракт уже представлен глоткой, пищеводом, зобом, желудком, средней и задней кишками.[ …]
Один из двух видов плоских червей обитает только в верховьях рек при температуре воды от 6,5 до 16°С, а другой вид, выносящий температуры в пределах от 6,5 до 23°С, может жить на всем протяжении реки. Когда оба вида симпатрические, то стенотермный вид вытесняет эвритермный из верховьев. и последний сохраняется только в нижнем течении реки, где значения температуры воды колеблются между 13 и 23°С.[ …]
Твистеры, имитируя различных червей» представляют собой кольчатое тело с плоским изогнутым хвостом, который при движении в воде совершает волнообразные колебания, чем придает приманке сходство с живым организмом и привлекает внимание рыбы.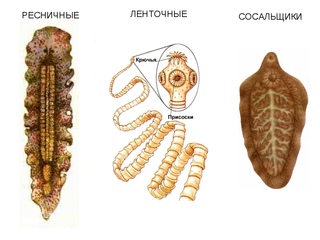
Анатомия и биология трематод. Это плоские черви листовидной формы. Тело обычно удлиненно-овальное, размером от 1 мм до 15 см. Оно покрыто тегументом (гладкое, с шипами или с щетинками), состоящим из наружного и внутреннего слоев (рис. 61). У большинства трематод две присоски: ротовая (на переднем конце тела) и брюшная (чаще на середине тела). У некоторых трематод, например у представителей Бап шшсоШае, паразитирующих в крови рыб, присоски рудиментированы или отсутствуют. От ротового отверстия отходит глотка, затем пищевод, переходящий в кишечник. Последний в виде двух стволов, достигнув заднего конца тела, слепо заканчивается. В очень редких случаях кишечные стволы открываются наружу. Сосальщики питаются тканевой жидкостью, слизью, некоторые — кровью. Продукты обмена выделяются через экскреторную систему протонефридиального типа. По всему телу размещены канальцы, объединяющиеся в два выделительных канала, открывающихся на заднем конце тела.[ …]
 Это плоские черви листовидной формы. Тело обычно удлиненно-овальное, размером от 1 мм до 15 см. Оно покрыто тегументом (гладкое, с шипами или с щетинками), состоящим из наружного и внутреннего слоев (рис. 61). У большинства трематод две присоски: ротовая (на переднем конце тела) и брюшная (чаще на середине тела). У некоторых трематод, например у представителей Бап шшсоШае, паразитирующих в крови рыб, присоски рудиментированы или отсутствуют. От ротового отверстия отходит глотка, затем пищевод, переходящий в кишечник. Последний в виде двух стволов, достигнув заднего конца тела, слепо заканчивается. В очень редких случаях кишечные стволы открываются наружу. Сосальщики питаются тканевой жидкостью, слизью, некоторые — кровью. Продукты обмена выделяются через экскреторную систему протонефридиального типа. По всему телу размещены канальцы, объединяющиеся в два выделительных канала, открывающихся на заднем конце тела.[ …]
Это плоские черви листовидной формы. Тело обычно удлиненно-овальное, размером от 1 мм до 15 см. Оно покрыто тегументом (гладкое, с шипами или с щетинками), состоящим из наружного и внутреннего слоев (рис. 61). У большинства трематод две присоски: ротовая (на переднем конце тела) и брюшная (чаще на середине тела). У некоторых трематод, например у представителей Бап шшсоШае, паразитирующих в крови рыб, присоски рудиментированы или отсутствуют. От ротового отверстия отходит глотка, затем пищевод, переходящий в кишечник. Последний в виде двух стволов, достигнув заднего конца тела, слепо заканчивается. В очень редких случаях кишечные стволы открываются наружу. Сосальщики питаются тканевой жидкостью, слизью, некоторые — кровью. Продукты обмена выделяются через экскреторную систему протонефридиального типа. По всему телу размещены канальцы, объединяющиеся в два выделительных канала, открывающихся на заднем конце тела.[ …]
У многих беспозвоночных животных, например, у кишечнополостных, плоских и круглых червей еще нет специальных органов дыхания.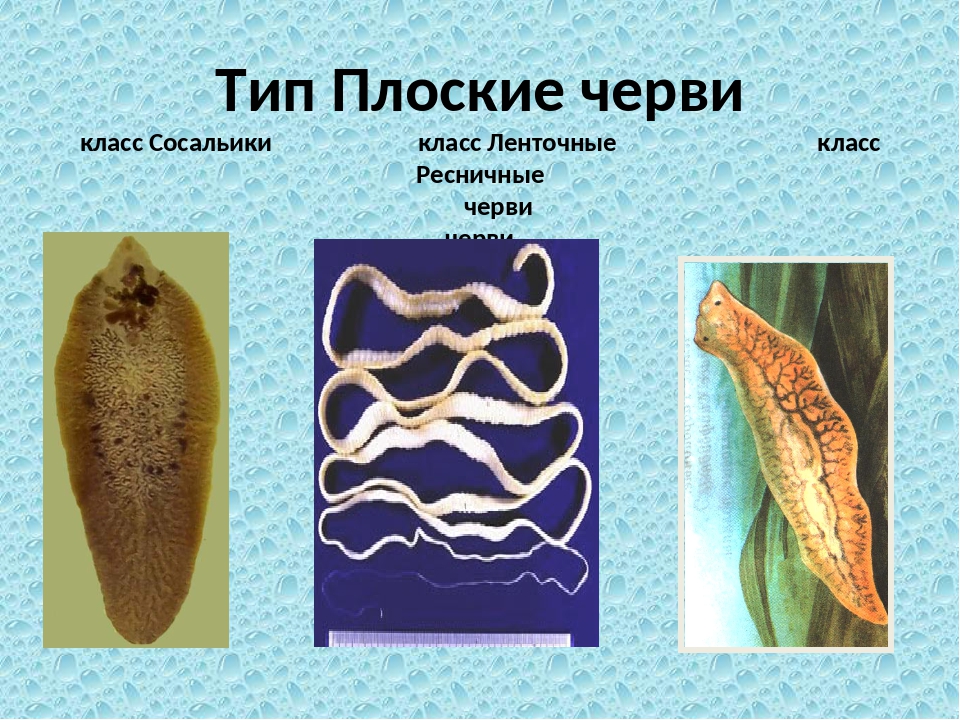 У них газообмен со средой осуществляется всей поверхностью тела (рис. 189). Впервые органы дыхания встречаются у морских кольчатых червей и водных членистоногих в виде пористых жабер, располагающихся по обеим сторонам тела и обильно снабжаемыми кровью. У моллюсков в мантийной полости развиваются пластинчатые жабры. Однако уже у наземных членистоногих в углублениях тела имеются трахеи или листовидные легкие.[ …]
У них газообмен со средой осуществляется всей поверхностью тела (рис. 189). Впервые органы дыхания встречаются у морских кольчатых червей и водных членистоногих в виде пористых жабер, располагающихся по обеим сторонам тела и обильно снабжаемыми кровью. У моллюсков в мантийной полости развиваются пластинчатые жабры. Однако уже у наземных членистоногих в углублениях тела имеются трахеи или листовидные легкие.[ …]
Эндотрофные и эктотрофные П. Эндотрофные П. живут внутри тканей организма-хозяина (плоские и круглые черви— в кишечнике позвоночных, круглые черви нематоды — в стеблях хлебных злаков), а эктотрофные «пасутся» на поверхности организма-хозяина (блохи, вши, ржавчинные грибы и многое др.). Особый случай эктотрофного паразитизма — образ жизни карликовых самцов глубоководных удильщиков (см. Взаимоотношения «паразит — хозяин»). Существуют суперпаразиты от первого до четвертого порядка (их можно представить в виде матрешки), на верхнем уровне — это бактерии или вирусы.[ …]
Эндотрофные и эктотрофные П. Эндотрофные П. живут внутри тканей организма-хозяина (плоские и круглые черви— в кишечнике позвоночных, круглые черви нематоды — в стеблях хлебных злаков), а эктотрофные «пасутся» на поверхности организма-хозяина (блохи, вши, ржавчинные грибы и многое др.). Особый случай эктотрофного паразитизма — образ жизни карликовых самцов глубоководных удильщиков (см. Взаимоотношения «паразит — хозяин»). Существуют суперпаразиты от первого до четвертого порядка (их можно представить в виде матрешки), на верхнем уровне — это бактерии или вирусы.[ …]
Эндотрофные П. живут внутри тканей организма-хозяина (плоские и круглые черви— в кишечнике позвоночных, круглые черви нематоды — в стеблях хлебных злаков), а эктотрофные «пасутся» на поверхности организма-хозяина (блохи, вши, ржавчинные грибы и многое др.). Особый случай эктотрофного паразитизма — образ жизни карликовых самцов глубоководных удильщиков (см. Взаимоотношения «паразит — хозяин»). Существуют суперпаразиты от первого до четвертого порядка (их можно представить в виде матрешки), на верхнем уровне — это бактерии или вирусы.[ …]
Одни из главных макропаразитов животных (фото 10) — глисты (гельминты), к которым относятся плоские черви (например, ленточные черви и трематоды), скребни и нематоды. Кроме того, животных поражают вши, блохи, клещи, а также грибы. К макропаразитам растений (рис. 12.6) относятся грибы, вызывающие настоящую и ложную мучнистую росу, ржавчину и головню; а также галлообразующие и минирующие насекомые и такие цветковые растения как повилика и заразиха.
Шистозоматоз, часто называемый биларзиазом, — паразитическое заболевание, вызываемое небольшими плоскими червями, попадающими во внутренние органы человека: сердце, легкие, вены и печень. Яйца шистозомных червей из кишечника человека вместе с фекальными массами попадают в воду. Это тяжелое заболевание, от которого не найдено иммунизации, представляет очень серьезную проблему для сельскохозяйственных районов Азии, Африки и Южной Америки. Шистозоматоз не встречается в континентальной части США, так как промежуточное звено — особая порода змей, в которых поселяются личинки этого паразита, — здесь не водится. Утиный шистозоматоз, передающийся аналогично человеческому, встречается в озерах США, куда часто прилетают дикие утки. Если инфицированные змеи( поселяются на пляжах у этих озер, выделенные ими церкарии могут проникнуть под кожу купающихся людей.
У многих беспозвоночных животных сосудистой системы как таковой нет. В частности, у губок, кишечнополостных и плоских червей транспорт питательных веществ и кислорода в разные части тела осуществляется путем диффузии их тканевых жидкостей. Однако у организмов других групп появляются сосуды, обеспечивающие циркуляцию тканевой жидкости в определенных направлениях. В начале развиваются примитивные сосуды, но в результате развития в сосудах мышечной ткани они в дальнейшем оказываются способными к сокращению. Параллельно развивается в кровь жидкость, заполняющая сосуды.[ …]
Осушение заболоченных пастбищ — надежный метод борьбы с фасциолезом — заболеванием сельскохозяйственных животных. Болезнь вызывается плоскими червями фасциолами, паразитирующими в желчных ходах печени хозяев. Цикл биологического развития фасциол связан с промежуточными хозяевами — прудовиками (моллюсками), предпочитающими влажные местообитания. Мелиоративные мероприятия по осушению пастбищ разрывают эпизоотическую цепь: лишенные необходимой влажности прудовики погибают, цикл развития фасциол нарушается. Циркуляция фасциол в БГЦ прекращается, что служит эффективным методом профилактики фасциолеза.[ …]
Болезнь вызывается плоскими червями фасциолами, паразитирующими в желчных ходах печени хозяев. Цикл биологического развития фасциол связан с промежуточными хозяевами — прудовиками (моллюсками), предпочитающими влажные местообитания. Мелиоративные мероприятия по осушению пастбищ разрывают эпизоотическую цепь: лишенные необходимой влажности прудовики погибают, цикл развития фасциол нарушается. Циркуляция фасциол в БГЦ прекращается, что служит эффективным методом профилактики фасциолеза.[ …]
Общий уровень метаболизма у паразитических видов существенно не изменен. Однако, живя в кишечнике животных или человека, где количество кислорода ограничено, многие плоские черви развили анаэробный тип дыхания, при котором кислород они получают путем восстановления углеводов (по типу брожения).[ …]
Значительный вклад в систематику принадлежит французскому ученому Ж. Ламарку (1744-1829), который разделил животных на беспозвоночных и позвоночных, а также определил основные группы (типы) червей (плоские, круглые и кольчатые). [ …]
[ …]
У беспозвоночных покровы тела не достигают значительного развития, будучи представленными эктодермой и ее производными. Эволюция покровов у них шла в направлении развития мерцательного эпителия в плоский эпителий, что отмечается у турбелля-рий и других плоских червей, а также у круглых и кольчатых червей. Мерцательный эпителий служит примитивным органом движения, тогда как плоские этой способностью не обладают.[ …]
У многоклеточных животных вегетативное размножение происходит путем фрагментации их тела на части, после чего каждая часть развивается в новое животное. Такое размножение характерно для губок, кишечнополостных (гидр), немертин, плоских червей, иглокожих (морских звезд) и некоторых других организмов. Близкой формой к вегетативному размножений животных фрагментацией является полиэмбриония животных, заключающаяся в том, что на определенной стадии развития эмбрион разделяется на несколько частей, каждая из которых развивается в самостоятельный организм. Полиэмбриония встречается у броненосцев. Однако последние размножаются половым путем. Поэтому полиэмбриония является скорее своеобразной стадией в половом размножении, а потомство, возникающее в результате полиэмбрионии, представлено монозиготными близнецами.[ …]
Однако последние размножаются половым путем. Поэтому полиэмбриония является скорее своеобразной стадией в половом размножении, а потомство, возникающее в результате полиэмбрионии, представлено монозиготными близнецами.[ …]
В нескольких экспериментах с вмешательством в природные сообщества было показано конкурентное исключение для ассамблей мелких млекопитающих [80], гильднй саламандр рода Plethodon [81] и ассамблей муравьев и грызунов [64]. Аналогичные эксперименты с популяциями плоских червей, морских звезд, морских желудей, блюдечек и воробьев дали сходные результаты [51, 54, 83, 296].[ …]
Другой тип — паразитизм. В самом обычном случае организм-паразит постоянно обитает на поверхности или внутри тела другого животного или растения (т. е. «хозяина») и живет за счет его питательных веществ. Примерами могут служить обычные вши, кишечные гельминты (плоские и круглые черви), клещи, простейшие, вызывающие заболевания, а из растений — повилика или полупаразит — омела. Такой паразитизм носит название истинного, при котором паразит не убивает своего хозяина. [ …]
[ …]
У простейших специальной репродуктивной системы нет. У губок и кишечнополостных такой системы тоже нет, их половые клетки выходят наружу через стенки тела. Репродуктивная система в виде половых желез и протоков для выведения половых клеток впервые развивается у плоских червей. Вопреки гермафродитизму, у плоских червей формируются семяпроводы и семяизвергатель-ные каналы для мужских половых клеток, а также яйцеводы, матка и влагалище для женских половых клеток. Половые клетки развиваются в семенниках и яичниках.[ …]
Вегетативное размножение — это размножение отдельными органами, частями органов или тела. Оно основано на способности организмов восстанавливать недостающие части тела — регенерации. Встречается у растений (размножение стеблями, листьями, побегами), у низших беспозвоночных животных (кишечнополостных, плоских и кольчатых червей).[ …]
При тесном контакте паразита с хозяином преимущество получают организмы, способные длительное время использовать хозяина, не приводя его к слишком ранней гибели и обеспечивая тем самым наилучшее существование. К числу постоянных паразитов относятся простейшие (малярийный плазмодий, дизентерийная амеба), плоские черви (сосальщики, цепни), круглые черви (аскарида, трихина и многие другие), членистоногие (вши, блохи, чесоточный клещ). Паразиты могут поселяться в крови, в тканях и полостях тела. Поскольку при постоянном паразитизме организм хозяина — единственное местообитание для паразита, с гибелью хозяина погибает и паразит.[ …]
К числу постоянных паразитов относятся простейшие (малярийный плазмодий, дизентерийная амеба), плоские черви (сосальщики, цепни), круглые черви (аскарида, трихина и многие другие), членистоногие (вши, блохи, чесоточный клещ). Паразиты могут поселяться в крови, в тканях и полостях тела. Поскольку при постоянном паразитизме организм хозяина — единственное местообитание для паразита, с гибелью хозяина погибает и паразит.[ …]
В пустом вегетационном сосуде, эмалированном ведре или кастрюле для каждого сосуда опыта отвешивают одно и то же количество почвы, пересыпают ее в эмалированный противень или большой эмалированный таз (блюдо) и тщательно перетирают между ладонями рук, отбрасывая механические примеси (корешки, камешки), насекомых и червей; особенно тщательно следят, чтобы не попались проволочники. Затем приливают, согласно схеме опыта, раствор удобрения или добавляют порошок с удобрением, или то и другое вместе и снова тщательно смешивают с почвой руками. После этого берут сосуд, на дне которого встряхиванием образуют для дренажа горку из битого стекла или гравия, покрывают ее кусочком марли и на дне дренажа укрепляют стеклянную трубку. В подготовленный таким образом сосуд переносят горстями почву, смешанную с удобрениями, и равномерно уплотняют ее легким нажатием руки. При набивке почвы в сосуды необходимо первый ее слой, лежащий над дренажем, уплотнять довольно сильно, а верхний слой — гораздо слабее. После того как все сосуды той или иной схемы опыта набиты, поверхность почвы в них выравнивают с небольшим уплотнением плоской стеклянной пробкой или деревянным кружком.[ …]
В подготовленный таким образом сосуд переносят горстями почву, смешанную с удобрениями, и равномерно уплотняют ее легким нажатием руки. При набивке почвы в сосуды необходимо первый ее слой, лежащий над дренажем, уплотнять довольно сильно, а верхний слой — гораздо слабее. После того как все сосуды той или иной схемы опыта набиты, поверхность почвы в них выравнивают с небольшим уплотнением плоской стеклянной пробкой или деревянным кружком.[ …]
Длина животного вместе со стрелкой соответствует продолжительности периода, в течение которого в пруду обнаруживаются взрослые представители каждого из пяти видов. Время пересыхания пруда и появления сухопутных организмов обозначено рисунком ювенильной стадии кузнечика (VI), / — тигровая амбистома; // — циклоп; ///— жаброиог; IV — днаптомус; V — плоский червь.[ …]
Паразитизм — явление столь всеобщее, что единственные живые существа, не подверженные нападению паразитов, это те паразиты, «оторые представляют собой последнее звено длинной цепи питания. Однако чем ниже на эволюционной лестнице находится та или иная группа живых организмов, тем больше она включает видов паразитов. Некоторые группы низших животных, особенно это касается плоских червей, нематод и некоторых членистоногих, состоят исключительно из паразитических форм. У позвоночных паразитизм как способ существования встречается крайне редко. В царстве растений паразиты широко распространены среди грибов (рис. 5.86).[ …]
Однако чем ниже на эволюционной лестнице находится та или иная группа живых организмов, тем больше она включает видов паразитов. Некоторые группы низших животных, особенно это касается плоских червей, нематод и некоторых членистоногих, состоят исключительно из паразитических форм. У позвоночных паразитизм как способ существования встречается крайне редко. В царстве растений паразиты широко распространены среди грибов (рис. 5.86).[ …]
Вторым крупнейшим этапом эволюции была трансформация одноклеточных организмов в многоклеточные. Считают, что первые эукариоты появились 1,5 млрд лет назад. Их появление есть крупнейший эволюционный переход, ибо в основе появления эукариотов лежит формирование митоза и развитие клеточных структур. Последние разветвились на пять линий, одна из которых повела к иглокожим и хордовым. Остальные линии повели к большинству других типов животного мира.[ …]
В Нижегородской области распоряжением губернатора от 13.05.97 № 574-р утверждено Положение о Красной книге Нижегородской области. Распоряжением от 22.07.98 № 1153-р утвержден Перечень видов позвоночных животных (118 видов, в том числе 30 — млекопитающих, 70 — птиц, 3 — пресмыкающихся, 1 -земноводных, 15 — рыб, 2 — круглоротых), внесенных в Красную книгу Нижегородской области, от 15.03.99 № 377-р — Перечни (отдельные) видов одноклеточных и беспозвоночных животных (138 видов, в том числе 4 — инфузорий, 1 — плоских червей, 1 — моллюсков, 2 — ракообразных, 4 — пауков, 126 — насекомых), грибов (34 вида), лишайников (11) и водорослей (2), высших растений (178 видов). Издание сводного тома Красной книги Нижегородской области планируется в конце 2003 г.[ …]
Распоряжением от 22.07.98 № 1153-р утвержден Перечень видов позвоночных животных (118 видов, в том числе 30 — млекопитающих, 70 — птиц, 3 — пресмыкающихся, 1 -земноводных, 15 — рыб, 2 — круглоротых), внесенных в Красную книгу Нижегородской области, от 15.03.99 № 377-р — Перечни (отдельные) видов одноклеточных и беспозвоночных животных (138 видов, в том числе 4 — инфузорий, 1 — плоских червей, 1 — моллюсков, 2 — ракообразных, 4 — пауков, 126 — насекомых), грибов (34 вида), лишайников (11) и водорослей (2), высших растений (178 видов). Издание сводного тома Красной книги Нижегородской области планируется в конце 2003 г.[ …]
Одним из главных преимуществ паразитов является обильное снабжение пищей за счет содержимого клеток, соков и тканей тела хозяина или содержимого его кишечника. Обильная и легкодоступная пища служит условием быстрого роста паразитов. Так, в кишечном тракте позвоночных паразиты достигают больших размеров по сравнению с их свободноживущими родственниками.[ …]
Рис. 14.17 иллюстрирует такие различия, наблюдаемые при выращивании зонтичного смирнии овощной (Smyr-nium olusatrum) при нормальных условиях и при недостатке питательных веществ (Lovett-Doust, 1980). Рис. 14.5 показывает одновременное влияние конкурентоспособности (а следовательно, и уровня питания) на скорость развития и размер отдельных экземпляров кобылок, а рис. 14.18 — различные реакции двух видов плоских червей на разные рационы (Woolhead, 1983). Все эти примеры говорят о том, что жизненные циклы часто отражают непосредственную реакцию организма на прошлые и настоящие условия существования.[ …]
14.17 иллюстрирует такие различия, наблюдаемые при выращивании зонтичного смирнии овощной (Smyr-nium olusatrum) при нормальных условиях и при недостатке питательных веществ (Lovett-Doust, 1980). Рис. 14.5 показывает одновременное влияние конкурентоспособности (а следовательно, и уровня питания) на скорость развития и размер отдельных экземпляров кобылок, а рис. 14.18 — различные реакции двух видов плоских червей на разные рационы (Woolhead, 1983). Все эти примеры говорят о том, что жизненные циклы часто отражают непосредственную реакцию организма на прошлые и настоящие условия существования.[ …]
Следующая за археем протерозойская эра — огромный по продолжительности период исторического развития Земли, в котором произошло значительное перераспределение площадей суши и моря на планете и развитие многоклеточных организмов. В течение этой эры бактерии и водоросли достигли своего расцвета. Господство эукариот (плавающих в толще воды растений) привело к появлению форм, прикрепленных ко дну. Постепенно тело многоклеточных растений расчленялось на части, выполнявшие разные функции (прикрепления, фотосинтеза и т.д.). В этот период сформировались губки, кишечнополостные, плоские и кольчатые черви, появились первые членистоногие, моллюски, иглокожие.[ …]
Постепенно тело многоклеточных растений расчленялось на части, выполнявшие разные функции (прикрепления, фотосинтеза и т.д.). В этот период сформировались губки, кишечнополостные, плоские и кольчатые черви, появились первые членистоногие, моллюски, иглокожие.[ …]
В лаборатории содержимое пробоотборника и контейнера укладывают в сито с размером отверстий 4 мм и промывают их через сито с отверстиями 250 мкм. Необходимо убедиться, что все существа целы и невредимы. Крупные беспозвоночные, застрявшие в сетке, хорошо уда-ляються из нее промывкой струей воды. Если это не удается, то их вынимают пинцетом. Моллюсков обычно извлекают при встряхивании пробоотборника. Труднее всего извлечь пиявок, но если поместить пробоотборник в 0,4%-ный раствор формалина или в теплую воду, то их можно быстро извлечь без повреждений. При сортировке в первую очередь удаляют попадающих в пробоотборник плоских червей.[ …]
Перенос заразного начала от больных животных к здоровым может быть либо механическим, либо биологическим. При механическом переносе микроорганизмы находятся на (в) теле переносчика, не вступая с ним в биологические отношения. Перемещаясь с больного животного на здоровое, переносчик при укусе заражает его. Примером может служить перенос кровососущими насекомыми бациллы сибирской язвы — возбудителя тяжелой болезни животных и людей. Слепни, мухи-жигалки, другие насекомые-кровососы, паразитируя на заболевших сибирской язвой животных, высасывают кровь и находящиеся в ней бациллы. Инфицированные сибиреязвенными бациллами насекомые-кровососы перелетают на других животных, в том числе здоровых, и, паразитируя, заражают их сибирской язвой. При биологической передаче возбудитель болезни, будучи связанным с переносчиком, вступает с ним в сложные биологические взаимоотношения, проходит определенный путь индивидуального развития и становится болезнетворным для сельскохозяйственных животных. В качестве примера можно привести заражение животных плоскими червями — фасциолами — возбудителями заболевания, называемого фасциолезом (рис.
При механическом переносе микроорганизмы находятся на (в) теле переносчика, не вступая с ним в биологические отношения. Перемещаясь с больного животного на здоровое, переносчик при укусе заражает его. Примером может служить перенос кровососущими насекомыми бациллы сибирской язвы — возбудителя тяжелой болезни животных и людей. Слепни, мухи-жигалки, другие насекомые-кровососы, паразитируя на заболевших сибирской язвой животных, высасывают кровь и находящиеся в ней бациллы. Инфицированные сибиреязвенными бациллами насекомые-кровососы перелетают на других животных, в том числе здоровых, и, паразитируя, заражают их сибирской язвой. При биологической передаче возбудитель болезни, будучи связанным с переносчиком, вступает с ним в сложные биологические взаимоотношения, проходит определенный путь индивидуального развития и становится болезнетворным для сельскохозяйственных животных. В качестве примера можно привести заражение животных плоскими червями — фасциолами — возбудителями заболевания, называемого фасциолезом (рис. 18). Фасциолы поражают животных всех видов и человека. Половозрелые фасциолы, паразитирующие в желчных ходах печени животных, выделяют яйца, которые попадают в кишечник, а оттуда с фекалиями во внешнюю среду. При благоприятных условиях из яйца выходит зародыш (мирацидий). Он плавает в воде, но сохраняется в ней недолго. Дальнейшее его развитие возможно только в теле промежуточного хозяина — биологического переносчика фасциол — моллюска (малого прудовика), обитающего в лужах, заросших травой. Плавая в воде, мирацидий контактирует с моллюском, внедряется в его тело, проходя дальнейшие стадии своего индивидуального развития. Затем зародыш выходит из тела промежуточного хозяина, превращается в адолескария, прикрепляется к листьям и стеблям растений, плавает в воде. Когда животные поедают траву и пьют воду, они заглатывают адолескариев и заражаются фасциолезом. Интенсивность распространения энзоотий фасциолеза на пастбищах определяется ареалом промежуточных хозяев фасциол — малых прудовиков.
18). Фасциолы поражают животных всех видов и человека. Половозрелые фасциолы, паразитирующие в желчных ходах печени животных, выделяют яйца, которые попадают в кишечник, а оттуда с фекалиями во внешнюю среду. При благоприятных условиях из яйца выходит зародыш (мирацидий). Он плавает в воде, но сохраняется в ней недолго. Дальнейшее его развитие возможно только в теле промежуточного хозяина — биологического переносчика фасциол — моллюска (малого прудовика), обитающего в лужах, заросших травой. Плавая в воде, мирацидий контактирует с моллюском, внедряется в его тело, проходя дальнейшие стадии своего индивидуального развития. Затем зародыш выходит из тела промежуточного хозяина, превращается в адолескария, прикрепляется к листьям и стеблям растений, плавает в воде. Когда животные поедают траву и пьют воду, они заглатывают адолескариев и заражаются фасциолезом. Интенсивность распространения энзоотий фасциолеза на пастбищах определяется ареалом промежуточных хозяев фасциол — малых прудовиков. [ …]
[ …]
ЦЖ: Тип Плоские черви Flashcards
~обитают в кишечнике животных и человека.~больше приспособлен к паразитическому образу жизни чем сосальщики
~Тело поделено на членики. На переднем конце тела расположена головка (сколекс), которая имеет органы прикрепления — присоски и крючья (За эти крючья червя называют также вооруженным цепнем), с помощью которых паразит закрепляется в кишечнике животного-хозяина.
За головкой располагается шейка. Новые членики образуются только в области шейки, поэтому передние, молодые членики небольшие, а удаленные от головки — более крупные.
Членики, находящиеся на заднем конце тела, периодически отрываются и с испражнениями хозяина попадают наружу.
~ как и у сосальщиков, тело покрыто кутикулой,
~Нервная система развита слабо, а органы чувств отсутствуют.
~Ленточные черви полностью утратили пищеварительную систему, а питательные вещества всасывают всей поверхностью тела.
~Ленточные черви гермафродиты. В каждом членике содержатся мужские и женские половые органы. Оплодотворение перекрёстное, но бывает и самооплодотворение.
В каждом членике содержатся мужские и женские половые органы. Оплодотворение перекрёстное, но бывает и самооплодотворение.
Обычно в членикох сначало развиваются оргоны мужской половой системы, а затем
в них образуются органы женской половой системы.
После оплодотворения мужская половая система постепенно исчезает, а весь членик наполняется яйцами. Членики, которые находятся на заднем конце тела, полностью заполнены зрелыми яйцами. Эти членики отрываются и выводятся из организма хозяина наружу с испражнениями. Один червь может продуцировать огромное число яиц — до сотен миллионов в год. При этом живут цепни несколько лет.
~Жизненный цикл бычьего цепня
Бычий цепень паразитирует в кишечнике человека (основной хозяин), где может достигать до 10 метров в длину.
вместе с испражнениями хозяина наружу выводятся яйца. Они попадают на траву, и поглощаются крупным рогатым скотом (промежуточный хозяин). В кишечнике животных из яйца выходит личинка, которая укореняется в мышцах и там превращается в следующую личиночную стадию — финну.
Финна растет, достигая величины крупной горошины. Ее тело представляет собой пузырь, наполненный жидкостью. Внутрь финны вдается зачаток головки цепня.
Человек (окончательный хозяин) заражается цепнем, когда употребляет в пищу недостаточно приготовленное мясо. В кишечнике человека головка финны выворачивается наружу. С помощью присосок и крючьев паразит прикрепляется к стенке кишечника, где он питается, растёт, и на его теле образуются членики.
~Жизненный цикл широкого лентеца
сменa двух промежуточных хозяев (рачка циклопа и рыбы). Основным (окончательным) хозяином широкого лентеца является человек.
Заразиться этим паразитом можно съев сырую или непрожаренную рыбу.
~Жизненный цикл эхинококка
Эхинококк — мелкий червь, живущий во взрослом состоянии в кишечнике собак, волков, шакалов, кошек (окончательные хозяева).
Пузырчатая стадия эхинококка (разновидность финны), развивающаяся в печени, лёгких, мышцах, костях промежуточных хозяев (овец, коров, свиней, человека), достигает величины головы ребёнка. В пузыре развивается большое число вторичных и даже третичных пузырей с головками внутри.
В пузыре развивается большое число вторичных и даже третичных пузырей с головками внутри.
Заражение окончательных хозяев происходит при поедании мяса с эхинококковыми пузырями. Промежуточные хозяева заражаются эхинококком при употреблении пищи, загрязненной яйцами паразита, которые попадают во внешнюю среду из кишечника больных собак, шакалов и волков.
~~Для свиного цепня человек -окончательный хозяин, промежуточным хозяином cлужит свинья. Свиньи заражаются цепнем, поедая нечистоты и эоглотывоя его яйцо. У попавших в кишечник свиньи яиц оболочка разрушается. Из яйцо выходит
личинка, имеющая облик маленького шарико с шестью хитиновыми крючками. С их помощью личинка вбуровливоется в стенки желудка или
кишки, проникает в кровеносные сосуды и током крови
разносится по телу, попадая в печень, сердце, легкие,
мозг, мышцы. В этих органах личинка превращается
в следующую личиночную стадию — финну. Фиино ростет, достигая величины крупной горошины. Ее тело представляет собой пузырь, наполненный жидкостью. Внутрь
Внутрь
финны вдоется зачаток головки цепня. Однако в теле
свиньи (промежуточного хозяина) дальнейшего развития
паразита не происходит. Для продолжения развития
финна должно попасть в организм человека. Это случается, когда человек употребляет в пищу непроверенную, непрожоренную или непросоленную свинину. В кишечнике человека головка финнывыворачивается наружу. Посредством присосок и крючь-
ев поразит прикрепляется к стенке кишечника.
Он питается, растет, и но его теле образуются членики.
Ленточные глисты у кошек, гельминтоз: симптомы и лечение
Гельминтоз у кошек — ужасное явление, по-другому и не скажешь. К сожалению, это довольно распространенная проблема у домашних питомцев, в частности у кошек. Что представляют собой ленточные черви? Заразны ли ленточные черви у кошек? И самый главный вопрос: как избавиться от ленточных червей?
Что представляют собой ленточные черви?
Ленточные черви — это длинные плоские черви. Во рту у них имеются крючки, с помощью которых они закрепляются в тонком кишечнике животного. Питаются они нутриентами, поступающими в организм кошки. Способны достигать 50 см в длину, но большинство взрослых червей вырастает приблизительно до 20 см. По мере роста от тела ленточного червя начинают отрываться отдельные сегменты, которые ученые называют проглоттидами. Проглоттиды размером с рисовое зернышко отделяются от задней части тела червя и попадают в фекалии кошки.
Питаются они нутриентами, поступающими в организм кошки. Способны достигать 50 см в длину, но большинство взрослых червей вырастает приблизительно до 20 см. По мере роста от тела ленточного червя начинают отрываться отдельные сегменты, которые ученые называют проглоттидами. Проглоттиды размером с рисовое зернышко отделяются от задней части тела червя и попадают в фекалии кошки.
Заражение кошки ленточными червями происходит несколькими путями. Самый распространенный — через блох. Крошечные личинки блох могут быть заражены ленточными червями. Если кошка проглотит зараженную блоху во время вылизывания шерсти, то вместе с блохой в ее организм попадет крошечный паразит, который вскоре вырастет до размеров зрелого червя. Кошка также может заразиться ленточными червями, съев мелкое животное, например белку или мышь.
Какой вред ленточные черви причиняют кошке?
Хотя ленточные черви у кошек и достигают больших размеров, ветеринарные врачи не считают их опасными. Все дело в том, что они не способны причинить здоровью животного стойкий вред, как считают специалисты из Ветеринарного центра Дрейка (Drake Center for Veterinary Care). Итак, если ваша кошка заражена ленточными червями, например глистами, она начнет терять вес, потому что паразиты будут съедать питательные вещества, поступающие из корма. Иногда ленточные черви выбираются из тонкого кишечника и попадают в желудок. Тогда у питомца может начаться рвота, и живой паразит выйдет наружу вместе с рвотными массами, вызвав испуг у владельца кошки, не подозревавшего о ее заражении.
Все дело в том, что они не способны причинить здоровью животного стойкий вред, как считают специалисты из Ветеринарного центра Дрейка (Drake Center for Veterinary Care). Итак, если ваша кошка заражена ленточными червями, например глистами, она начнет терять вес, потому что паразиты будут съедать питательные вещества, поступающие из корма. Иногда ленточные черви выбираются из тонкого кишечника и попадают в желудок. Тогда у питомца может начаться рвота, и живой паразит выйдет наружу вместе с рвотными массами, вызвав испуг у владельца кошки, не подозревавшего о ее заражении.
Как определить, что кошка заражена ленточными червями?
Естественно, однозначным признаком паразитов служат сегменты тела ленточных червей в рвотных массах домашнего питомца. К прочим симптомам гельминтоза у кошек относится необъяснимая потеря веса, но наиболее распространенным признаком являются проглоттиды. Трудно не заметить белые, рисообразные, наполненные яйцами сегменты тела червя в фекалиях кошки и на шерсти возле заднего прохода.![]() Также вы можете заметить, как животное будто бы чешет заднюю часть туловища о пол, так как паразиты раздражают кожу в области заднего прохода, хотя такое поведение более свойственно собакам.
Также вы можете заметить, как животное будто бы чешет заднюю часть туловища о пол, так как паразиты раздражают кожу в области заднего прохода, хотя такое поведение более свойственно собакам.
Как лечить гельминтоз у кошек?
К счастью, гельминтоз лечится довольно просто и эффективно. Если ваша кошка заразилась, ветеринарный врач предложит вам противоглистные лекарства. Обычно они выпускаются в виде препаратов для приема внутрь, но иногда — в форме инъекций.
После приема противоглистного препарата гельминты погибают Соответственно, в кошачьем лотке признаков их присутствия вы больше не увидите. Противоглистные препараты обычно не вызывают у кошки никаких побочных эффектов, таких как рвота или диарея.
Конечно, лучше всего вообще не допускать заражения кошки ленточными червями. Риск гельминтоза существенно снижается при регулярном применении средств для защиты от блох и домашнем содержании питомца. Сами по себе ленточные черви не заразны, как, например, простуда, но могут передаваться (через блох) другим животным и изредка человеку. Аналогичным образом при проглатывании зараженной блохи заражается гельминтозом и собака. Если вы или ваши дети случайно проглотите блоху, вы тоже можете заразиться.
Аналогичным образом при проглатывании зараженной блохи заражается гельминтозом и собака. Если вы или ваши дети случайно проглотите блоху, вы тоже можете заразиться.
Сколько видов ленточных червей существует?
Существует два вида ленточных червей. Наиболее распространен так называемый Dipylidium Caninum, как пояснили специалисты из Центра контроля и предотвращения болезней (Centers for Disease Control and Prevention, CDC), которому посвящена данная статья.
Второй вид, представляющий более серьезную угрозу, называется эхинококк (Echinococcus). Согласно CDC, эхинококкоз пузырный развивается в результате заражения личиночной стадией ленточных червей эхинококков (Echinococcus granulosus), переносчиками которых являются собаки, овцы, крупный рогатый скот, козы и свиньи.
«Несмотря на то что большая часть заболевания протекает бессимптомно, при пузырном эхинококкозе развиваются опасные, постепенно увеличивающиеся в размерах кисты в печени, легких и прочих органах, которые больные не замечают годами», — говорят специалисты из CDC.
Еще одной разновидностью эхинококка является эхинококк многокамерный, вызывающий заболевание под названием эхинококкоз альвеолярный. Переносчиками данного вида паразитов являются лисы, собаки, кошки и мелкие грызуны. Случаи заболевания у человека крайне редки, но очень серьезны и характеризуются развитием паразитарных опухолей в печени, легких, мозге и других органах. Согласно информации CDC, при отсутствии лечения альвеолярный эхинококкоз может закончиться летальным исходом. Но, к счастью, такие случаи редки.
Другие паразитарные черви у кошек
Ленточные черви — всего лишь один из часто встречающихся видов паразитарных червей, заражающих животных. Международная организация по уходу за кошками (International Cat Care) выделяет еще несколько видов паразитарных червей, встречающихся у животных:
- Круглые черви. Наиболее часто обнаруживаются у кошек. Котята заражаются ими через молоко матери. Взрослое животное заражается, съедая инфицированного грызуна.

- Нематоды. Наиболее распространены у собак, но встречаются и у кошек. Они мелкие и, подобно ленточным червям, живут в тонком кишечнике животного. Питаются кровью животного, что может привести к анемии. Заражение происходит при проглатывании яиц или личинок нематод.
- Некишечные черви. Легочные, сердечные и глазные, живущие в соответствующих частях тела животного.
Разговор о червях-паразитах, живущих в организме животного, может вызвать тошноту у владельцев даже с самым крепким желудком. К счастью, даже несмотря на их большой размер, от паразитных червей довольно просто избавиться, и не стоит опасаться каких-то долговременных последствий для здоровья питомца. Лучшее, что можно сделать для кошки, — внимательно следить за ее поведением. Внезапные изменения в ее поведении могут говорить о проблемах со здоровьем. Именно поэтому так важен регулярный ветеринарный осмотр.
Об авторе
Кара Мёрфи
Кара Мёрфи — писатель-фрилансер из города Эри, штат Пенсильвания.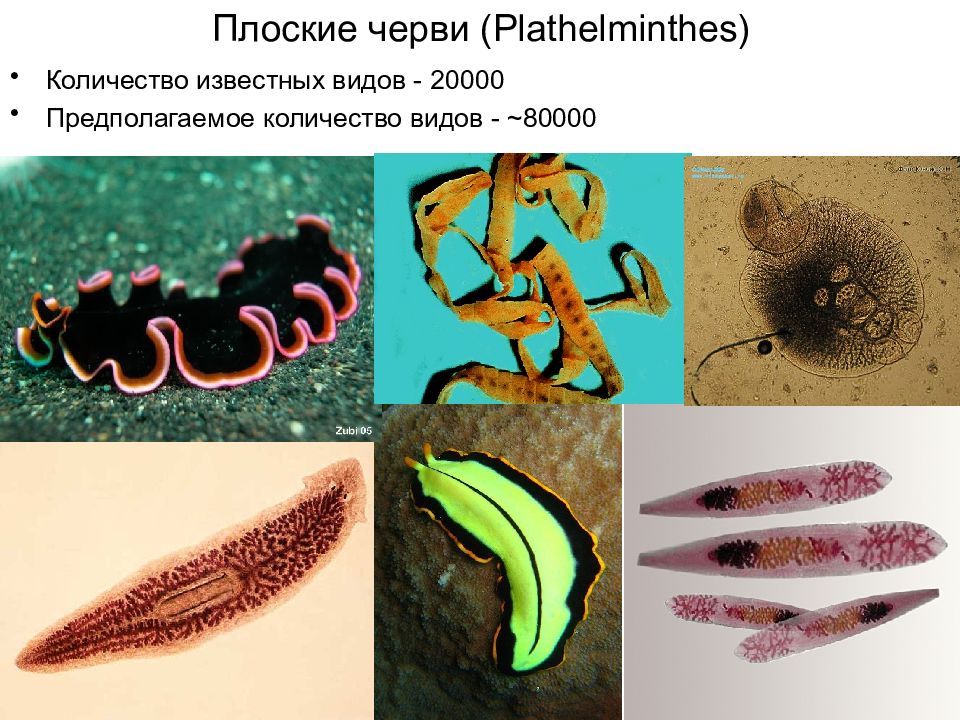 Хозяйка голдендудля по кличке Мэдди.
Хозяйка голдендудля по кличке Мэдди.
Купаться запрещено: опасности стоячей воды
Красные пятна по всему телу появились у Татьяны Аникиной и ее сына. Она подумала, что это комары, затем – ветрянка, и только врач установил: это последствия купания в озере – укусы плоских червей церкариев. Это не опасно, но крайне мучительно.
«Температуры не было, но единственное – был очень сильный зуд, прямо чуть ли не до слез», – говорит Татьяна Аникина.
Сразу два соседних озера в Нижнем Новгороде оказались заражены личинками паразитов. Несмотря на наличие рядом проточной реки Оки, до которой всего 5 километров, в жару люди предпочитали купаться в заболоченном озере у дома. Результат – массовое заражение в районе.
Интернет пестрит жалобами на укусы и жуткую чесотку в регионах, где стояла аномальная жара. Все водоемы Нижегородской, Томской, Челябинской и Московской областей, где заразились люди, объединяет одно.
«Зонами повышенного риска заражения людей церкариозом являются заболоченные заросшие водоемы, где присутствуют дикие водоплавающие птицы, моллюски», – рассказала замначальника отдела надзора за объектами коммунально-бытового назначения и средой обитания человека управления Роспотребнадзора по Москве Любовь Чердакова.
Стоячая вода – рай для паразитов и уток. И те и другие там прекрасно себя чувствуют из-за обилия пропитания.
«Плоские черви паразитируют у основных хозяев – уток, водоплавающих. С фекалиями эти черви попадают в воду. Промежуточный хозяин – это ракушки, улитки, из которых выходят церкарии и попадают на кожный покров», – объяснила заведующая филиалом Юго-Западный Московского научно-практического центра дерматовенерологии и косметологии Наталья Заторская.
В московский центр дерматовенерологии этим знойным летом с укусами церкариев обращаются буквально каждый день. Даже коллега на днях заболел, поделились врачи. Специфического лечения нет, только симптоматическое, чтобы облегчить зуд. Помогают растворы, содержащие ментол, анестезин.
«Хорошо протирать кожу салициловым спиртом, можно какой-то подкисленной водой – с лимоном или слабым раствором уксуса. Травы используют – отвар череды, ромашки, чай», – говорит Заторская.
Плюс специальные мази. Выздоровление – от недели до трех, если без осложнений. Врачи настоятельно рекомендуют не купаться в заболоченных местах со стоячей водой, водорослями, утками и улитками. А если вы все же зашли в воду, то тут же нужно смыть болотную воду чистой и насухо вытереться жестким полотенцем.
Врачи настоятельно рекомендуют не купаться в заболоченных местах со стоячей водой, водорослями, утками и улитками. А если вы все же зашли в воду, то тут же нужно смыть болотную воду чистой и насухо вытереться жестким полотенцем.
Еще перед купанием могут помочь репелленты, но постоянно использовать мазь от комаров на пляже – удовольствие весьма сомнительное.
Определение плоские черви общее значение и понятие. Что это такое плоские черви
Плательминто (слово, образованное от слова Platyhelminthes ) — это организм, лишенный позвонков, также известный как плоский червь, который обладает характеристиками протомаста и трибластического ацеломадо. Большая часть этого клана является паразитической и гермафродитной, и не имеет дыхательной или кровеносной системы. С другой стороны, следует сказать, что многие из двадцати тысяч видов плоских червей не имеют пищеварительной системы или заднего прохода, поскольку пищеварительный тракт функционирует как желудочно-сосудистая полость (то есть выполняет пищеварительные функции и распределяет питательные вещества).
Плоские черви обычно обитают во флювиальных, морских и наземных влажных средах. В зависимости от их особенностей их также можно разделить на четыре класса: турбелария ( Turbellaria ), моногенный ( Monogenea ), цестоды ( Cestoda ) и трематоды ( Trematoda ). Эти последние три комплекта являются паразитами и не имеют головы.
У турбелариев есть свободная жизнь, они — плотоядные животные, и они копают в поисках пищи . Моногены — это паразиты рыб и амфибий, а цестоды и трематоды могут жить у разных млекопитающих, в том числе у человека .
Следует отметить, что плоские черви не имеют двигательных придатков; поэтому они двигаются благодаря вибрациям реснитчатого эпителия.
В случае плоских червей, которые могут повлиять на человека, они также известны как ленточные черви или одиночные. Люди могут быть заражены этими путями, такими как употребление в пищу определенных продуктов (овощей, фруктов или сырой свинины или плохо приготовленных) с яйцами таэнии.
Среди наиболее частых симптомов, с которыми сталкиваются граждане, страдающие этим заболеванием, включают потерю веса, трудности с засыпанием, тошноту, легкие желудочно-кишечные боли, нервозность или отсутствие аппетита.
Факт воздействия этих ленточных червей влечет за собой несколько последствий, наиболее серьезными из которых являются различные травмы головного мозга, повреждения зрения, такие как слепота, а также судороги различных диапазонов. В этом случае лучше всего обратиться к медицинскому работнику, который назначит необходимое лечение, чтобы покончить с этой инфекцией.
Однако следует подчеркнуть, что, как и во многих других заболеваниях или патологиях, существует ряд домашних средств для той же цели. В частности, в этом случае рекомендуется употреблять в пищу свежие фрукты, такие как клубника и другие продукты, такие как лук, грецкие орехи или чеснок.
Фундаментальный совет, которому необходимо следовать, чтобы предотвратить воздействие этого «животного», не ешьте сырое мясо или не приготовленное должным образом, всегда мойте руки после посещения туалета, а также держите их в идеальном состоянии и мойте после Я занимался свининой или говядиной.
Плоские черви позволили Джеймсу Макконнеллу провести различные эксперименты с целью доказать, что существует химическая основа для памяти . Для этого он сочетал яркий свет с электрическим током и изучал реакцию плоских червей. После нескольких испытаний он обнаружил, что платигельминты начали реагировать на свет так же, как если бы они подвергались поражению электрическим током.
Эксперимент пошел дальше: разрезав червей пополам и дав им возможность регенерировать, можно было увидеть, как у обеих половин развивалась реакция на свет и поражение электрическим током.
Плоские черви могут размножаться, разрывая себя пополам
Когда планарии хотят размножаться, некоторые из них занимаются сексом. Другие, более прямолинейно, разрывают себя надвое.
Последний вариант быстрый и жестокий. Планария начинается как маленькое, приплюснутое, похожее на слизня существо с лопатообразной головой и двумя вытаращенными глазами. После нескольких минут растяжения и разрыва он разделяется на две половины — голову и хвост. В течение нескольких дней у головы вырастает хвост. И что еще более чудесно, у хвоста отрастает голова.«Это просто сногсшибательно», — сказала мне Ева-Мария Коллинз из Суортморского колледжа, изучающая этих животных. Разводить их несложно: при наличии достаточного количества пищи планарии многократно удваивают себя, деля себя пополам. И если Коллинз быстро понадобится больше животных, она может сделать с помощью скальпеля то, что черви делают своими мускулами. Как писал натуралист Джон Грэм Дайелл в 1814 году, планарий можно «почти назвать бессмертными под лезвием ножа».
В течение нескольких дней у головы вырастает хвост. И что еще более чудесно, у хвоста отрастает голова.«Это просто сногсшибательно», — сказала мне Ева-Мария Коллинз из Суортморского колледжа, изучающая этих животных. Разводить их несложно: при наличии достаточного количества пищи планарии многократно удваивают себя, деля себя пополам. И если Коллинз быстро понадобится больше животных, она может сделать с помощью скальпеля то, что черви делают своими мускулами. Как писал натуралист Джон Грэм Дайелл в 1814 году, планарий можно «почти назвать бессмертными под лезвием ножа».
Существуют тысячи видов планарий, и все они сильно отличаются от более привычных червей, таких как дождевые черви.Их тела представляют собой переплетение мышц и соединительной ткани, без внутренних полостей, заполненных мягкими органами. Рот находится посередине нижней стороны и служит анальным отверстием. Они выделяют жидкие отходы через поры на спине. Они получают кислород путем диффузии, и у них отсутствуют легкие, жабры, сердце и кровеносные сосуды. У них действительно есть своего рода мозг — два скопления нейронов в голове. Они ведут к нервной системе в форме лестницы, состоящей из двух нервных тяжей, спускающихся вниз по телу и соединенных поперечными перекладинами.
У них действительно есть своего рода мозг — два скопления нейронов в голове. Они ведут к нервной системе в форме лестницы, состоящей из двух нервных тяжей, спускающихся вниз по телу и соединенных поперечными перекладинами.
Прочтите: Самый неловкий орган тела — чудо эволюции
Эта необычная анатомия еще более странна, потому что она может выдержать разрезание пополам. Этот подвиг интриговал людей по крайней мере с девятого века, но его трудно наблюдать. Планарии самофрагментируются всего раз в месяц, и этот процесс завершается в течение нескольких минут. Они также предпочитают разделяться в темноте и останавливаются, если их потревожить. Чтобы изучить их, Коллинз и ее команда снимали один вид, Dugesia japonica , непрерывно в течение нескольких месяцев.Они увидели, что существо начинает свое саморасчленение, сокращая свою среднюю часть, чтобы создать талию, изменяя свою форму с сигары на песочные часы. Затем он прикрепляет голову и хвост — к чашке Петри в лаборатории, но обычно к подводному камню в дикой природе — и сокращает промежуточные мышцы, неоднократно растягивая плоть на талии, пока она не порвется. (Процесс различается у разных видов; у Schmidtea mediterranea чем крупнее червь, тем больше кусков он может оторвать от хвоста.)
(Процесс различается у разных видов; у Schmidtea mediterranea чем крупнее червь, тем больше кусков он может оторвать от хвоста.)
После того, как червь раскололся, его фрагменты не остаются без дела. Как только они освобождаются друг от друга, обе части кажутся «обладающими свойствами совершенного животного, передвигающегося в воде так же скользяще, как и до разделения», — писал натуралист Джеймс Роулинз Джонсон в 1822 году. Столетия спустя Коллинз показал, что эта автономия более глубока, чем кто-либо подозревал. Ее команда (включая студентов бакалавриата Дилана Ле и Зиада Сабри и старшеклассника Аарава Чандра) показала, что неповрежденная планария поворачивается, если ее ткнуть в голову, растягивается, если ее ткнуть в середину, и сжимается, если ее ткнуть в хвост.Но если эту планарию разрезать на три части — голову, туловище и хвост — каждая часть ведет себя как целое существо. Передний конец туловища повернется, как будто это голова, а задний конец сожмется, как если бы это был хвост.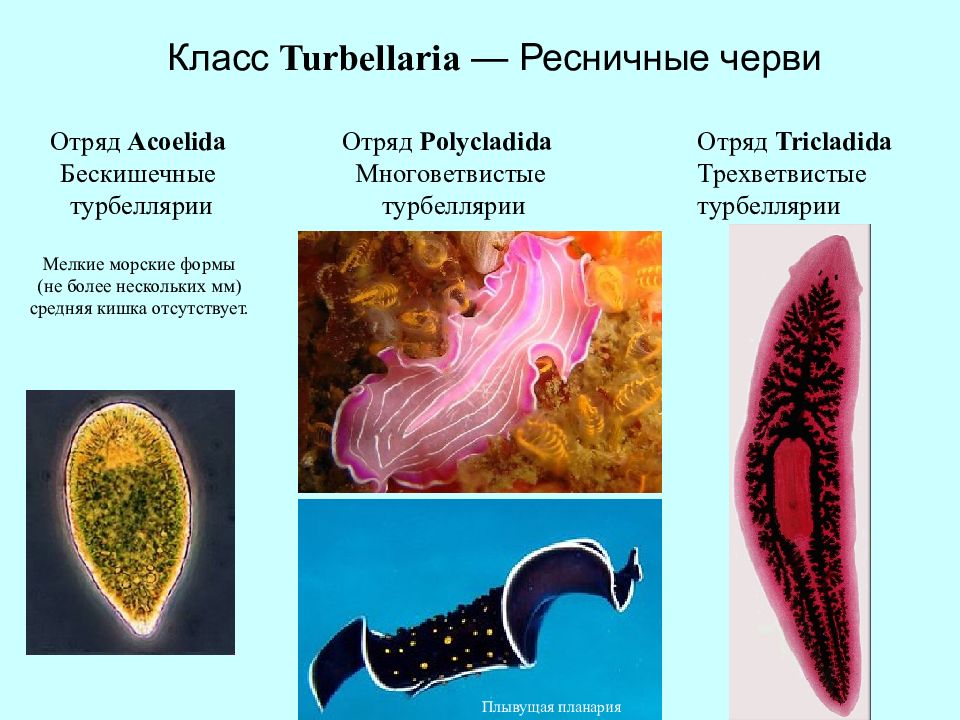
Прочтите: Обезглавленные черви выздоравливают, посмотрите еще раз
Коллинз считает, что нервные цепи, контролирующие такое поведение, повторяются по всей длине планарии, так что каждая часть тела способна действовать как голова, туловище или хвостик.Присутствие фактической головы обычно не позволяет нижестоящим областям действовать как одна. Но после обезглавливания самая передняя часть оставшейся планарии может взять на себя обязанности ныне отсутствующей головы. Коллинз рассматривает эту крайнюю приспособляемость как стратегию выживания. Это означает, что каждый фрагмент может убежать от опасности, давая ему достаточно времени для того, чтобы сработали его экстраординарные регенеративные способности.
Не все планарии могут регенерировать, но те, которые могут, имеют тенденцию быть эффектными в этом. Когда еды не хватает, они могут «вырастать», разрушая свои собственные клетки, только чтобы снова набрать вес, когда условия улучшатся. Они бросают вызов процессу старения, регулярно заменяя свои старые ткани и органы. Они могут оправиться практически от любой физической травмы (хотя некоторые из них сложнее; надрез между глазами может побудить планарию регенерировать две головы). Биолог Томас Хант Морган однажды подсчитал, что полная планария может регенерировать всего из одной 279 части своего тела. «Немногие животные могут регенерировать свою нервную систему, и я не знаю ни , ни других животных, способных заново вырастить мозг», — говорит Оне Паган из Уэст-Честерского университета в Пенсильвании, написавший книгу «Первый мозг: неврология планарий».
Они бросают вызов процессу старения, регулярно заменяя свои старые ткани и органы. Они могут оправиться практически от любой физической травмы (хотя некоторые из них сложнее; надрез между глазами может побудить планарию регенерировать две головы). Биолог Томас Хант Морган однажды подсчитал, что полная планария может регенерировать всего из одной 279 части своего тела. «Немногие животные могут регенерировать свою нервную систему, и я не знаю ни , ни других животных, способных заново вырастить мозг», — говорит Оне Паган из Уэст-Честерского университета в Пенсильвании, написавший книгу «Первый мозг: неврология планарий».
Читайте: Мозг, который не должен был зажить
Когда планарии делятся естественным путем, фрагмент головы обычно больше и содержит мозг, глаза, сенсоры обоняния и вкуса, а также рот. «Наконечнику просто нужно залечить рану и продолжить путь», — сказал мне Коллинз. Тем временем хвост должен регенерировать все остальное. Без рта у него нет возможности получать питательные вещества. Вместо этого некоторые из его клеток самоуничтожаются, чтобы обеспечить сырье для создания новой плоти. Медленно изолированный хвост претерпевает «масштабную реконструкцию», — сказал мне Алехандро Санчес Альварадо, эксперт по планариям из Института Стоуэрса, — и в итоге вы получаете крошечную версию исходного животного.«Хвостовые части погибают примерно в 10 раз чаще, чем головные, — добавил Коллинз, — но все же примерно семь из восьми выживают».
Без рта у него нет возможности получать питательные вещества. Вместо этого некоторые из его клеток самоуничтожаются, чтобы обеспечить сырье для создания новой плоти. Медленно изолированный хвост претерпевает «масштабную реконструкцию», — сказал мне Алехандро Санчес Альварадо, эксперт по планариям из Института Стоуэрса, — и в итоге вы получаете крошечную версию исходного животного.«Хвостовые части погибают примерно в 10 раз чаще, чем головные, — добавил Коллинз, — но все же примерно семь из восьми выживают».
Эти способности зависят от особых клеток, называемых необластами, которые были обнаружены только у планарий. Они распределены по всему телу существа, составляя от 25 до 30 процентов его клеток. В 2014 году команда под руководством Питера Реддиена из Массачусетского технологического института бомбардировала планарий смертельной дозой радиации и пересадила один необласт — всего один — от второй особи на хвост обреченного животного.Когда реципиент умер от головы вниз, пересаженный необласт начал производить новые ткани от хвоста вверх. Новые клетки в конечном итоге заменили все умирающие, как если бы планария-донор через единственную клетку завладела и оживила труп реципиента. Через две недели уползло полное и здоровое животное — планария Тесея.
Новые клетки в конечном итоге заменили все умирающие, как если бы планария-донор через единственную клетку завладела и оживила труп реципиента. Через две недели уползло полное и здоровое животное — планария Тесея.
Читайте: Выращивание органов на яблоках
Конечно, большинство животных вырастают из одной оплодотворенной яйцеклетки. Но по мере того, как это яйцо становится эмбрионом, клетки внутри него становятся более настроенными по-своему.Клетка кожи не превращается в нейрон. Стволовые клетки более гибкие, но у взрослых животных даже они имеют свои пределы: стволовая клетка крови не может создавать клетки печени или сердца. Необласты взрослых планарий не имеют таких ограничений. Это кусочки неограниченной возможности, способные производить любую ткань или орган.
Необласты не работают по отдельности. Тот, что пересадил Реддиен, не начал делать глаза или мозг; он создал ткани, соответствующие его местонахождению. Это связано с тем, что концентрации определенных молекул меняются по длине планарии, спереди назад и сверху вниз.Это создает своего рода систему координат, которая сообщает ячейкам в каждой секции, где они находятся в общем плане тела. Они могут использовать эту информацию во время процесса регенерации, чтобы заново вырастить то, что необходимо. Головы регенерируют хвосты, а не лишние головы (хотя возможны ошибки). Хвосты делают головы. Стволы прорастают головками и хвостиками. И независимо от их происхождения, новые животные, кажется, помнят что-то из своего прошлого существования.
Это связано с тем, что концентрации определенных молекул меняются по длине планарии, спереди назад и сверху вниз.Это создает своего рода систему координат, которая сообщает ячейкам в каждой секции, где они находятся в общем плане тела. Они могут использовать эту информацию во время процесса регенерации, чтобы заново вырастить то, что необходимо. Головы регенерируют хвосты, а не лишние головы (хотя возможны ошибки). Хвосты делают головы. Стволы прорастают головками и хвостиками. И независимо от их происхождения, новые животные, кажется, помнят что-то из своего прошлого существования.
В 1950-х и 1960-х годах биолог Джеймс В. Макконнелл показал, что безголовые планарии, которые были вынуждены заново вырастить свой мозг, все еще могут помнить поведение, которому они научились до обезглавливания.Он даже опубликовал результаты, свидетельствующие о том, что нетренированные планарии могут вести себя так, как обученные, если первые съели измельченных частей последних. Скептики критиковали эти эксперименты и утверждали, что МакКоннелл просто видел поведение, которое хотел видеть. Но десятилетия спустя Майк Левин и Тал Шомрат из Университета Тафтса разработали машину, которая могла автоматически обучать и отслеживать планарий без какого-либо вмешательства или предвзятости человека. Они показали, что черви, обученные распознавать текстуру шероховатой чашки Петри, все еще могут делать это после обезглавливания и отращивания новых голов.
Но десятилетия спустя Майк Левин и Тал Шомрат из Университета Тафтса разработали машину, которая могла автоматически обучать и отслеживать планарий без какого-либо вмешательства или предвзятости человека. Они показали, что черви, обученные распознавать текстуру шероховатой чашки Петри, все еще могут делать это после обезглавливания и отращивания новых голов.
Прочтите: Безмозглая слизь, которая разделяет воспоминания путем слияния
Как минимум, утверждает Левин, это показывает, что воспоминания действительно могут храниться вне мозга. Это также укрепляет его ощущение того, что взгляд из учебника на память — что она закодирована силой синаптических связей между различными нейронами — неверен. Вместо этого Левин подозревает, что нервная система могла эволюционировать, чтобы интерпретировать воспоминаний, а не кодировать их; они хранятся в другом месте, в каком-то аспекте наших клеток, который еще никто не определил. Это, если быть точным, весьма спекулятивно. «У нас есть только одно исследование, и оно далеко не окончательное, — сказал мне Левин. «Но это один из многих фрагментов данных, предполагающих, что мы вообще не понимаем память».
Это, если быть точным, весьма спекулятивно. «У нас есть только одно исследование, и оно далеко не окончательное, — сказал мне Левин. «Но это один из многих фрагментов данных, предполагающих, что мы вообще не понимаем память».
Планарианцы усложняют и другие, казалось бы, простые понятия. Рассмотрим вопрос, который Левин и его коллеги задали в 2016 году: после того, как разделенная пополам планарий регенерирует в двух новых животных, будет ли планария, выросшая из головы, считать ту, которая выросла из хвоста, своим близнецом , своим братом , его потомок или сам ? Ответ не очевиден, потому что эти слова были определены людьми — видом, который, как я проверял в последний раз, не может размножаться, разрывая себя на части.«Странные вещи — это именно то, на что вам нужно обращать внимание», — сказал мне Левин. «Они говорят вам, что ваша модель мира несовершенна в важных аспектах. Вы должны дорожить исключениями».
Прочтите: Решение проблемы регенерации человека, возможно, зависло в давно забытой области науки
Планарии, безусловно, исключительны, но они не уникальны в своих талантах. Многие другие животные могут регенерировать недостающие части тела, включая саламандр, ящериц и морских звезд.Некоторые из них могут размножаться, разделившись на две части, в том числе морские анемоны и щупальцевидная гидра . Многие ученые изучают этих существ в надежде найти медицинские открытия, которые смогут восстановить поврежденные органы и потерянные конечности. Но более неотложной наградой является осознание того, насколько неполным является наше понимание природы и насколько наш язык и понятия ограничены нашими собственными негибкими, неделимыми телами.
Многие другие животные могут регенерировать недостающие части тела, включая саламандр, ящериц и морских звезд.Некоторые из них могут размножаться, разделившись на две части, в том числе морские анемоны и щупальцевидная гидра . Многие ученые изучают этих существ в надежде найти медицинские открытия, которые смогут восстановить поврежденные органы и потерянные конечности. Но более неотложной наградой является осознание того, насколько неполным является наше понимание природы и насколько наш язык и понятия ограничены нашими собственными негибкими, неделимыми телами.
11.6: Плоские черви — Биология LibreTexts
Поверите ли вы, что это существо с золотыми точками — плоский червь?
Нет? Что ж, это так.Существует более 25 000 различных видов плоских червей, поэтому они могут сильно различаться по внешнему виду. И многие даже не похожи на вашего типичного червя.
Плоские черви
Плоские черви относятся к типу Platyhelminthes. Примеры плоских червей показаны на рисунке ниже. Тип плоских червей насчитывает более 25 000 видов.
Примеры плоских червей показаны на рисунке ниже. Тип плоских червей насчитывает более 25 000 видов.
Платигельминты. К платигельминтам относятся плоские черви, ленточные черви и двуустки.
Структура и функции плоских червей
Длина плоских червей варьируется примерно от 1 миллиметра (0.04 дюйма) до более чем 20 метров (66 футов). У них плоское тело, потому что у них нет целома или даже псевдоцелома. У них также отсутствует дыхательная система. Вместо этого их клетки обмениваются газами путем диффузии непосредственно с окружающей средой. У них несовершенная пищеварительная система.
Плоские черви отражают несколько основных эволюционных достижений беспозвоночных. Они имеют три зародышевых клеточных слоя, включая мезодерму. Слой мезодермы позволяет им развивать системы органов. Например, у них есть мышечная и выделительная системы.Мышечная система позволяет им передвигаться с места на место по твердым поверхностям. Выделительная система позволяет им поддерживать правильный баланс воды и солей. Плоские черви также демонстрируют цефализацию и двустороннюю симметрию.
Плоские черви также демонстрируют цефализацию и двустороннюю симметрию.
Размножение плоских червей
Плоские черви размножаются половым путем. У большинства видов одни и те же особи производят как яйцеклетки, так и сперматозоиды. После оплодотворения оплодотворенные яйца выходят из тела взрослой особи и превращаются в личинок. Может быть несколько разных личиночных стадий.Последняя личиночная стадия развивается во взрослую форму, и жизненный цикл повторяется.
Экология плоских червей
Как трематоды, так и ленточные черви паразитируют на позвоночных, включая человека. Сосальщики живут в кровеносной системе или печени хозяина. Ленточные черви живут в пищеварительной системе хозяина. Обычно для завершения жизненного цикла паразита требуется более одного типа хозяина. Посмотрите на жизненный цикл печеночной двуустки на рисунке ниже. Во взрослом возрасте у двуустки есть позвоночный хозяин.Как личинка, у него есть беспозвоночный хозяин. Если вы проследите за жизненным циклом, вы увидите, как заражается каждый хозяин, чтобы двуустка могла продолжить свой жизненный цикл.
Жизненный цикл двуустки овечьей печени. У овечьей двуустки сложный жизненный цикл с двумя хозяевами. Как мог сложиться такой сложный образ жизни?
Ленточные черви и двуустки имеют присоски и другие структуры для питания хозяина. Ленточные черви также имеют сколекс , кольцо крючков на голове, чтобы прикрепляться к хозяину (см. Рисунок ниже).В отличие от других беспозвоночных, у ленточных червей нет рта и пищеварительной системы. Вместо этого они поглощают питательные вещества прямо из пищеварительной системы хозяина своими присосками.
Присоски и крючки для ленточных червей. Головка цепня имеет несколько присосок. На самом верху головы находится «корона» из крючьев, называемая сколексом.
Не все плоские черви являются паразитами. Некоторые из них являются свободноживущими хищниками. Они поедают других мелких беспозвоночных и разлагающихся животных. Большинство свободноживущих видов живут в водной среде обитания, но некоторые обитают во влажной почве.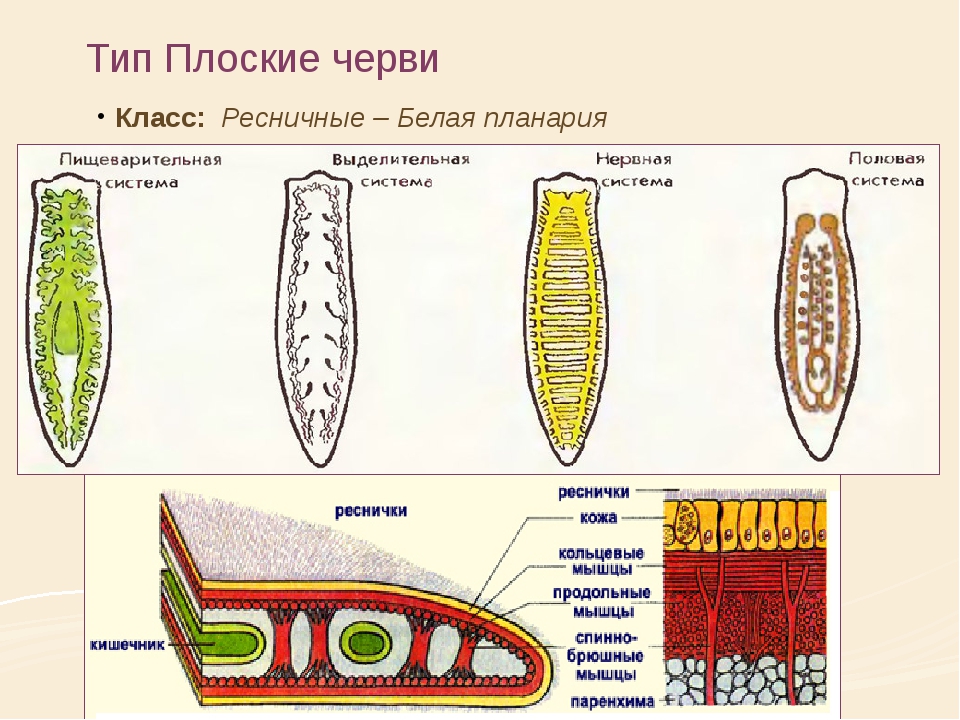
Плоские черви — Всплеск
Хотя их называют плоскими червями, эти беспозвоночные не являются плоскими. Они имеют круглое тело с плоским животом. Они длиннее, чем в ширину. Они заострены на каждом конце. У большинства плоских червей есть по крайней мере два глазных пятна рядом с головным концом тела.
Не менее 20 видов плоских червей обитают в весенних лужах. Три вида легко найти в весенних прудах Сакраменто. Первый плоский червь, Mesostoma, полупрозрачный и белый.Второй плоский червь полупрозрачный и зеленый. В его теле живут зеленые водоросли. Третий плоский червь темно-коричневый, с пятнами или пятнами на верхней стороне.
Научное название: Многие видыТип: Platyhelminthes
Класс: Турбеллярия
Место обитания: Весенние заводи, весенние болота и пресноводные болота
Размер: 0.длиной от 5 мм до 1 см
Забавные факты:
Плоский червь имеет единственное отверстие в желудке, расположенное посередине его нижней части. Плоские черви в весенних лужах могут втыкать эту глотку (горло/рот) в свою добычу. Подобно короткому шлангу пылесоса, он высасывает внутренности их добычи. Они также используют его, чтобы выталкивать отходы из своего тела.
Плоские черви в весенних лужах могут втыкать эту глотку (горло/рот) в свою добычу. Подобно короткому шлангу пылесоса, он высасывает внутренности их добычи. Они также используют его, чтобы выталкивать отходы из своего тела.
Жизненный цикл:
Плоские черви — одни из первых тварей, которые вылупляются, когда бассейны начинают наполняться дождевой водой.Молодые вылупляются из яиц и внешне похожи на взрослых. Повзрослев, плоские черви спариваются и откладывают темные круглые яйца. Большинство плоских червей являются гермафродитами, что означает, что они являются как самцами, так и самками. Каждый плоский червь может спариваться с любым другим плоским червем, если он принадлежит к тому же виду.
Экология:
Плоские черви поедаются взрослыми особями и личинками водяных жуков. Плоские черви в весенних лужицах питаются бактериями, водорослями, простейшими и мелкими беспозвоночными, такими как сказочные креветки.Некоторые плоские черви питаются мертвыми и умирающими беспозвоночными.
Белый плоский червь, Mesostoma, подвешивает нить слизи в воде, чтобы ловить водяных блох. Он возвращается, чтобы проверить свою «линию ловушек» и собрать всех водяных блох, попавших в слизь.
Плоский червь передвигается, расправляя множество крошечных, тонких, похожих на волоски структур, называемых ресничками, на внешней стороне тела. Плоский живот позволяет ему скользить по поверхности воды, а реснички мягко перемещают его в поисках пищи или партнера.
Расследовать:
Следите за коричневыми плоскими червями, которые скользят по поверхности весенних луж.Они реагируют на колебания поверхности воды, чтобы выследить добычу. Если насекомое упадет (или вы коснетесь) на поверхность воды, плоские черви будут двигаться в сторону беспокойства, надеясь поймать какую-нибудь добычу, терпящую бедствие.
Исследуйте тварей Исследуйте растения
Плоские черви Acoel не являются платигельминтами: данные филогеномики
Abstract
Плоские черви Acoel — это мелкие морские черви, традиционно считающиеся принадлежащими к типу Platyhelminthes. Однако молекулярный филогенетический анализ предполагает, что акоэли не являются представителями Platyhelminthes, а скорее являются существующими представителями самых ранних дивергирующих Bilateria. Этот результат был поставлен под сомнение из-за подозрений в артефакте притяжения длинных ветвей (LBA). Здесь мы повторно изучаем эту проблему с помощью филогеномного подхода, используя 68 различных кодирующих белок генов из acoel Convoluta pulchra и 51 многоклеточного вида, принадлежащего к 15 различным типам. Мы используем смешанную модель, названную CAT, которая, как было установлено ранее, преодолевает артефакты LBA там, где классические модели терпят неудачу.Наши результаты однозначно показывают, что акоэли не являются частью классически определенных Platyhelminthes, что делает последних полифилетическими. Более того, они указывают на родство вторичноротых с акоэлями, потенциально как сестринскую группу со всеми вторичноротыми, с Xenoturbellida, Ambulacraria или даже с хордовыми.
Однако молекулярный филогенетический анализ предполагает, что акоэли не являются представителями Platyhelminthes, а скорее являются существующими представителями самых ранних дивергирующих Bilateria. Этот результат был поставлен под сомнение из-за подозрений в артефакте притяжения длинных ветвей (LBA). Здесь мы повторно изучаем эту проблему с помощью филогеномного подхода, используя 68 различных кодирующих белок генов из acoel Convoluta pulchra и 51 многоклеточного вида, принадлежащего к 15 различным типам. Мы используем смешанную модель, названную CAT, которая, как было установлено ранее, преодолевает артефакты LBA там, где классические модели терпят неудачу.Наши результаты однозначно показывают, что акоэли не являются частью классически определенных Platyhelminthes, что делает последних полифилетическими. Более того, они указывают на родство вторичноротых с акоэлями, потенциально как сестринскую группу со всеми вторичноротыми, с Xenoturbellida, Ambulacraria или даже с хордовыми. Тем не менее, слабая поддержка, обнаруженная для большинства узлов вторичноротых, вместе с очень быстрой скоростью эволюции acoel Convoluta pulchra , требуют дополнительных данных о медленно развивающихся acoel (или из его родственной группы, Nemertodermatida) для решения этой сложной филогенетической проблемы. проблема.
Тем не менее, слабая поддержка, обнаруженная для большинства узлов вторичноротых, вместе с очень быстрой скоростью эволюции acoel Convoluta pulchra , требуют дополнительных данных о медленно развивающихся acoel (или из его родственной группы, Nemertodermatida) для решения этой сложной филогенетической проблемы. проблема.
Образец цитирования: Филипп Х., Бринкманн Х., Мартинес П., Риуторт М., Багунья Дж. (2007) Плоские черви Acoel не являются платигельминтами: данные филогеномики. ПЛОС ОДИН 2(8): е717. https://doi.org/10.1371/journal.pone.0000717
Академический редактор: Жан-Николя Вольф, Высшая нормальная школа Лиона, Франция
Поступила в редакцию: 4 мая 2007 г.; Принято: 4 июля 2007 г .; Опубликовано: 8 августа 2007 г.
Авторское право: © 2007 Philippe et al.Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: HP выражает благодарность Génome Québec, Канадскому научно-исследовательскому председателю и Университету Монреаля за финансовую поддержку, PM, MR и JB поддерживаются грантами от ICREA, Generalitat de Catalunya и Ministryio de Educación y Ciencia.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Плоские черви Acoelomorph (Acoela+Nemertodermatida) составляют небольшую группу билатеральных морских червей, которые недавно привлекли внимание. Несколько морфологических сходств позволяют предположить, что акоеломорфы принадлежат Platyhelminthes [1]–[3]; однако эти признаки часто плохо определены или представляют собой простые симплезиоморфии [4], что оставляет неясным статус филума Platyhelminthes.Молекулярные филогении, основанные на SSU рРНК [5], [6], комбинированных SSU и LSU рРНК [7], миозине II [8] и митохондриальных геномах [9], обеспечивают сильную статистическую поддержку исключения акоеломорфов из Platyhelminthes. Скорее, эти маркеры определяют их как сестринскую группу для всех оставшихся Bilateria.
Скорее, эти маркеры определяют их как сестринскую группу для всех оставшихся Bilateria.
Однако, поскольку акоеломорфы эволюционируют с очень высокой скоростью (за исключением, возможно, миозина II), их базальное появление может быть легко объяснено артефактом притяжения длинных ветвей (LBA) [10], запускаемым отдаленно родственной внешней группой (Cnidaria).Хотя осторожные подходы (например, выбор наиболее медленно эволюционирующей последовательности рРНК среди 18 акоэлей [5]) использовались в попытке избежать артефактов LBA, положение акоэломорфов остается неясным. В частности, акоеломорфы часто рассматриваются как вторично упрощенные организмы [11], [12], что потенциально объясняет их высокую скорость эволюции. Однако обнаружение ацеломорфов у основания Bilateria или рядом с ним вместо того, чтобы принадлежать к Platyhelminthes, позволило бы поляризовать несколько ключевых билатеральных признаков, включая мозг, целом, нефридий и наличие первичной личиночной стадии.
Поэтому мы решили повторно проанализировать этот важный вопрос, применяя мощный одновременный анализ нескольких ортологичных генов, кодирующих ядерный белок [13], и используя современные методы реконструкции деревьев для усиления филогенетического сигнала [14].
Результаты и обсуждение
Мы секвенировали 2304 EST из acoel Convoluta pulchra , число, которое было предложено для обеспечения достаточного количества гомологичных положений [15]. В общей сложности 68 различных генов, кодирующих белок, были однозначно отнесены к набору данных консервативных однокопийных генов, ранее клонированных из других многоклеточных животных и родственных таксонов [14], [16].Богатая таксонная выборка из 51 вида, содержащая основные линии билатеральных (Deuterostomia, Ecdysozoa и Lophotrochozoa) и несколько близких и медленно развивающихся видов внешней группы (четыре Cnidaria, три Porifera, три Choanoflagellata и три Ichthyosporea), была отобрана для максимально возможного снижения потенциала Артефакты LBA. После удаления неоднозначно выровненных позиций и частей, которые не были секвенированы в Convoluta , было получено выравнивание 11 959 позиций.
Последовательности из Convoluta чрезвычайно расходятся, что делает его размещение, вероятно, подверженным артефакту LBA. Действительно, при использовании метода экономии, высокочувствительного к этому артефакту [10], Convoluta неизменно привлекает самая длинная непрерывная ветвь: оболочник Oikopleura с Bootstrap Support (BS) 96% (рис. S1). , или Platyhelminthes (BS = 72%), если исключить Oikopleura (рис. S2). Поэтому мы обратились к вероятностным методам, менее чувствительным к LBA [17], [18]. Кроме того, мы использовали модель CAT [19], поскольку было показано, что она позволяет преодолеть артефакт LBA, когда другие модели не работают [14], [20].Результирующее дерево (рис. 1) прекрасно согласуется с текущими знаниями [12] и недавними крупномасштабными анализами [14], [16], [21]–[24]: монофилия всех типов животных и большинства супер- phyla (Bilateria, Protostomia, Ecdysozoa и Lophotrochozoa) восстанавливается при сильной поддержке (BS>99%). Важно отметить, что Convoluta не группируется ни с Platyhelminthes, ни с Oikopleura , хотя эти две группы имеют самые длинные ветви.
Действительно, при использовании метода экономии, высокочувствительного к этому артефакту [10], Convoluta неизменно привлекает самая длинная непрерывная ветвь: оболочник Oikopleura с Bootstrap Support (BS) 96% (рис. S1). , или Platyhelminthes (BS = 72%), если исключить Oikopleura (рис. S2). Поэтому мы обратились к вероятностным методам, менее чувствительным к LBA [17], [18]. Кроме того, мы использовали модель CAT [19], поскольку было показано, что она позволяет преодолеть артефакт LBA, когда другие модели не работают [14], [20].Результирующее дерево (рис. 1) прекрасно согласуется с текущими знаниями [12] и недавними крупномасштабными анализами [14], [16], [21]–[24]: монофилия всех типов животных и большинства супер- phyla (Bilateria, Protostomia, Ecdysozoa и Lophotrochozoa) восстанавливается при сильной поддержке (BS>99%). Важно отметить, что Convoluta не группируется ни с Platyhelminthes, ни с Oikopleura , хотя эти две группы имеют самые длинные ветви..jpg) Хотя точное положение Convoluta точно не определено, два хорошо поддерживаемых узла (Protostomia и Lophotrochozoa) отделяют acoels от Platyhelminthes.Поэтому отказ от традиционной морфологической гипотезы очень силен.
Хотя точное положение Convoluta точно не определено, два хорошо поддерживаемых узла (Protostomia и Lophotrochozoa) отделяют acoels от Platyhelminthes.Поэтому отказ от традиционной морфологической гипотезы очень силен.
Рисунок 1. Филогенетический анализ геномных данных решительно опровергает группировку Convoluta и Platyhelminthes.
Байесовское дерево, полученное в результате анализа 11 959 выровненных позиций аминокислот с помощью модели CAT. Полученные значения начальной загрузки указываются, если <100%, в противном случае на узле присутствует маркер. Масштабная линейка показывает количество изменений на сайте.
https://дои.org/10.1371/journal.pone.0000717.g001
Однако использование стандартной модели WAG обеспечивает сильную поддержку группировки двух длинных ветвей платигельминтов и акоэлей (рис. S3). Чтобы еще раз подтвердить, что эта группировка является артефактом как экономии, так и модели WAG, мы искали гены, которые присутствуют у Convoluta и у вторичноротых, но отсутствуют у первичноротых (подробности см.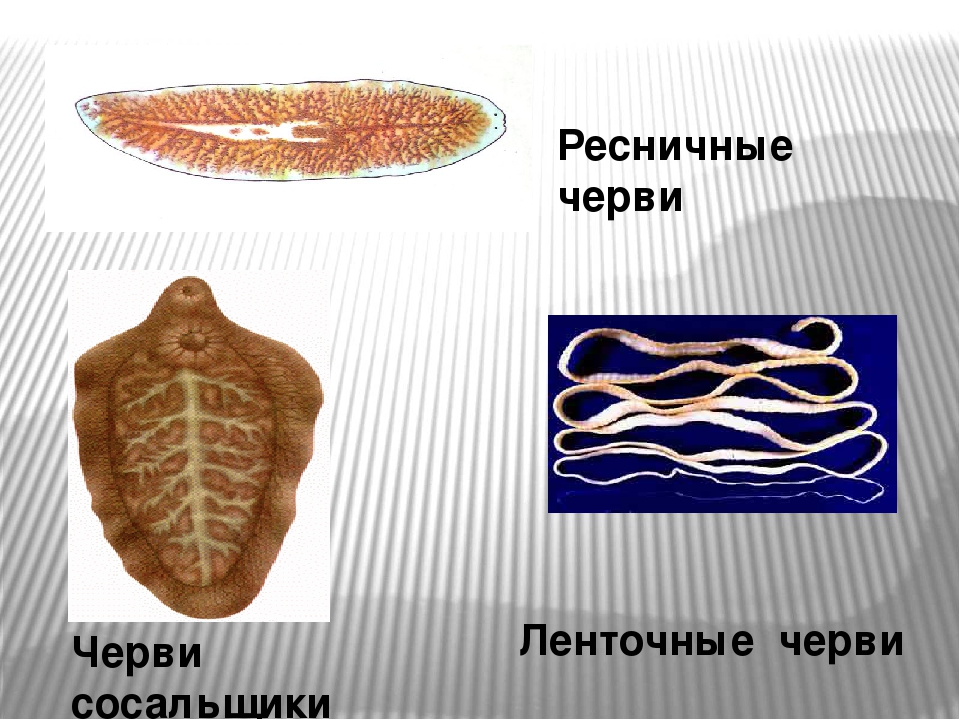 В дополнении). Несмотря на то, что мы проанализировали только 1664 контига из Convoluta (в основном кодирующих универсальные белки, такие как рибосомные белки), мы обнаружили один ген, гуанидиноацетат N-метилтрансферазу, который присутствует у губок, книдарий, вторичноротых и Convoluta , но отсутствует у всех первичноротых, кроме базальных щетинкочелюстных [24].Дерево модели WAG (рис. S3), в котором акоэли отнесены к сестринской группе платигельминтов, предполагает как минимум три независимые потери, тогда как дерево на рис. 1 подразумевает одну потерю первичноротых после появления щетинкочелюстных. Совпадение дерева, полученного с помощью CAT-модели, и распределения гуанидиноацетат-N-метилтрансферазы убедительно доказывает, что дерево, полученное с использованием модели WAG, смещено из-за артефакта LBA, указывая на пониженную чувствительность CAT-модели к этому артефакту [14]. ], [20].
В дополнении). Несмотря на то, что мы проанализировали только 1664 контига из Convoluta (в основном кодирующих универсальные белки, такие как рибосомные белки), мы обнаружили один ген, гуанидиноацетат N-метилтрансферазу, который присутствует у губок, книдарий, вторичноротых и Convoluta , но отсутствует у всех первичноротых, кроме базальных щетинкочелюстных [24].Дерево модели WAG (рис. S3), в котором акоэли отнесены к сестринской группе платигельминтов, предполагает как минимум три независимые потери, тогда как дерево на рис. 1 подразумевает одну потерю первичноротых после появления щетинкочелюстных. Совпадение дерева, полученного с помощью CAT-модели, и распределения гуанидиноацетат-N-метилтрансферазы убедительно доказывает, что дерево, полученное с использованием модели WAG, смещено из-за артефакта LBA, указывая на пониженную чувствительность CAT-модели к этому артефакту [14]. ], [20].
Хотя наше филогеномное древо Bilateria в целом хорошо разрешено, три узла внутри первичноротых, положения хетогнат, тихоходок и платигельминтов, плохо подтверждены. Что еще более поразительно, большинство отношений между вторичноротыми не поддерживаются. За исключением монофилии Olfactores [16] и Ambulacraria [21], остальные узелки, включая вторичноротых и хордовых, получают BS ниже 55%. Очень быстро развивающийся Convoluta появляется в этой части дерева как сестринская группа вторичноротых (рис. 1, BS = 14%), хордовых (BS = 33%) или Xenoturbella (BS = 20). %). Convoluta проявляет сходство с вторичноротыми в 90% бутстрепных повторов, будучи базальным по отношению к Bilateria только в 7% повторений и к Protostomia только в 3%. Следует отметить, что отсутствие поддержки внутри вторичноротых не связано с наличием длинной ветви Convoluta , поскольку BS остается низким (между 42% и 73%) при удалении Convoluta (рис. S4). Как следствие, надежному местоположению Convoluta препятствуют две трудности: (i) его очень высокая скорость эволюции и (ii) расположение в вторичноротых или рядом с ними, область, для которой мощный филогеномный подход дает только плохо подкрепленные результаты [16], [21], [23].
Что еще более поразительно, большинство отношений между вторичноротыми не поддерживаются. За исключением монофилии Olfactores [16] и Ambulacraria [21], остальные узелки, включая вторичноротых и хордовых, получают BS ниже 55%. Очень быстро развивающийся Convoluta появляется в этой части дерева как сестринская группа вторичноротых (рис. 1, BS = 14%), хордовых (BS = 33%) или Xenoturbella (BS = 20). %). Convoluta проявляет сходство с вторичноротыми в 90% бутстрепных повторов, будучи базальным по отношению к Bilateria только в 7% повторений и к Protostomia только в 3%. Следует отметить, что отсутствие поддержки внутри вторичноротых не связано с наличием длинной ветви Convoluta , поскольку BS остается низким (между 42% и 73%) при удалении Convoluta (рис. S4). Как следствие, надежному местоположению Convoluta препятствуют две трудности: (i) его очень высокая скорость эволюции и (ii) расположение в вторичноротых или рядом с ними, область, для которой мощный филогеномный подход дает только плохо подкрепленные результаты [16], [21], [23].
Чтобы усилить филогенетический сигнал [25], мы удалили внешнюю группу (т.е. небилатериальные виды). Удивительно, но происходит единственное топологическое изменение (рис. S5): Xenoturbella становится сестринской группой для Convoluta , хотя и с низкой поддержкой (41%). Кроме того, слабая поддержка недавно предложенного Xenoturbella + Ambulacraria [21] не связана с присутствием Convoluta (рис. S6). Таким образом, наше большое сопоставление из 11 959 позиций решительно отвергает объединение акоэлей с платигельминтами и, в более общем смысле, с первичноротыми.Это способствует сродству вторичноротых с акоэлями, потенциально как сестринская группа по отношению к ксенотурбеллидам. Однако для решения этой сложной филогенетической проблемы необходимо больше данных, особенно о медленно развивающихся акоэлях.
Отказ от объединения акоэлей с платигельминтами sensu stricto (подробное таксономическое обсуждение см. в [4]) имеет несколько морфологических последствий. Во-первых, отсутствие протонефридиев у акоэлей, традиционно считавшееся следствием утраты их предком-платигельминтом, теперь может рассматриваться как сохранение примитивного состояния, общего с диплобластами.Аналогичный аргумент можно было бы применить к мешкообразному кишечнику ацелей (и его родственной группы, Nemertodermatida; [26], [27]), которая теперь может быть симплезиоморфией, разделяемой с аналогичным состоянием диплобластов и независимой от сходного состояния. мешкообразное состояние Platyhelminthes sensu stricto внутри Lophotrochozoa. Кроме того, своеобразное дуэтно-спиральное дробление акоэлей, также традиционно считавшееся производным от четверного спирального дробления других платигельминтов, теперь можно рассматривать как происходящее от формы радиального или бирадиального дробления, характерного для более примитивных многоклеточных животных.Интересно, что группировка Acoelomorpha и Xenoturbellida (рис. S5) была предложена на основе ультраструктур эпидермальных двигательных цилиарных корешков [28] и заслуживает будущих исследований.
Во-первых, отсутствие протонефридиев у акоэлей, традиционно считавшееся следствием утраты их предком-платигельминтом, теперь может рассматриваться как сохранение примитивного состояния, общего с диплобластами.Аналогичный аргумент можно было бы применить к мешкообразному кишечнику ацелей (и его родственной группы, Nemertodermatida; [26], [27]), которая теперь может быть симплезиоморфией, разделяемой с аналогичным состоянием диплобластов и независимой от сходного состояния. мешкообразное состояние Platyhelminthes sensu stricto внутри Lophotrochozoa. Кроме того, своеобразное дуэтно-спиральное дробление акоэлей, также традиционно считавшееся производным от четверного спирального дробления других платигельминтов, теперь можно рассматривать как происходящее от формы радиального или бирадиального дробления, характерного для более примитивных многоклеточных животных.Интересно, что группировка Acoelomorpha и Xenoturbellida (рис. S5) была предложена на основе ультраструктур эпидермальных двигательных цилиарных корешков [28] и заслуживает будущих исследований.
Наши результаты противоречат не только морфологической точке зрения, но и предыдущим молекулярным филогениям, которые сильно поддерживают появление акоэлей у основания билатерий [5]–[9], поскольку эта позиция получает здесь бутстрап-поддержку только на 7%. Но ожидается, что такое базальное появление будет усилено артефактом LBA между длинной ветвью Convoluta и дальней родственной внешней группой.Использование модели CAT, которая менее чувствительна к LBA [14], [20], частично объясняет наблюдаемое отсутствие поддержки базального выхода, поскольку последняя поддержка увеличивается при использовании модели WAG (рис. S7). Однако основная причина расхождений с предыдущими молекулярными исследованиями [5]–[9], вероятно, заключается в том, что внешняя группа менее отдалена в нашем наборе данных, тем самым уменьшая артефакт LBA: расстояние от книдарий до последнего общего предка билатерий составляет 55,5% расстояния между книдариями и позвоночными для наших ядерных белков (рис. 1), но 63.9% для митохондриальных белков [9] и 73,7% для 18S рРНК [5].
1), но 63.9% для митохондриальных белков [9] и 73,7% для 18S рРНК [5].
В более концептуальных терминах положение акоэлей среди Platyhelminthes должно предостеречь нас от наивного взгляда, который рассматривает некоторые признаки как «потерянные», «отсутствующие» или «уменьшенные» в кладах (например, акоэлях), чем они могли бы никогда не иметь. на первом месте. Действительно, возможное положение акоэлей как сестринской группы по отношению к Xenoturbellida, другой группе, состоящей из простых организмов, в основании вторичноротых оставляет открытым вопрос об эволюционном происхождении их морфологической простоты.
Материалы и методы
Клонирование генов
Библиотеку кДНК конструировали из взрослой ткани acoel Convoluta pulchra с использованием стандартных методологий. кДНК клонировали в плазмидный вектор pSPORT1. Была сгенерирована библиотека из нескольких тысяч клонов со средним размером вставки 1,5 кб. Из созданной коллекции было отобрано 2304 клона, которые были секвенированы с использованием праймера Т7. Все последовательности были сгруппированы, и для филогенетического анализа были отобраны унигены плюс одиночные.
Все последовательности были сгруппированы, и для филогенетического анализа были отобраны унигены плюс одиночные.
Блок данных
Каждое из выравниваний генов, использованных в предыдущих исследованиях [14], [16], [23], [29], было обновлено, в дополнение к последовательностям Convoluta , новыми доступными последовательностями, загруженными из Trace Archive (http:// www.ncbi.nlm.nih.gov/Traces/) и базу данных EST (http://www.ncbi.nlm.nih.gov/dbEST/) GenBank в Национальном центре биотехнологической информации (http://www. .ncbi.nlm.nih.gov/), используя новые возможности программы ED из пакета MUST [30] (список видов см. в таблице S1).Неоднозначно выровненные области были автоматически обнаружены и удалены с помощью программы Gblocks [31], и этот выбор был уточнен вручную с помощью программы ED.
Конкатенация 68 генов, для которых было доступно последовательностей Convoluta , была выполнена программой ScaFos [32]. ScaFos позволяет выбирать медленно развивающиеся последовательности в соответствии с их степенью дивергенции, используя расстояния ML, рассчитанные по модели WAG+F с помощью Tree-Puzzle [33]. Это также позволяет уменьшить процент отсутствующих данных по таксону за счет создания химерных последовательностей из видов, принадлежащих к одной и той же предопределенной таксономической группе (см. Таблицу S2).Были рассмотрены только гены, представленные не менее чем 42 видами. Полученное выравнивание состоит из 68 генов, 52 видов и 15 554 однозначно выровненных позиций. Далее мы удалили все позиции, для которых Convoluta отсутствовала (из-за частичных последовательностей генов), в результате чего было получено окончательное выравнивание 11 959 позиций. Выравнивание доступно по запросу в компании H.P. ([email protected]).
Это также позволяет уменьшить процент отсутствующих данных по таксону за счет создания химерных последовательностей из видов, принадлежащих к одной и той же предопределенной таксономической группе (см. Таблицу S2).Были рассмотрены только гены, представленные не менее чем 42 видами. Полученное выравнивание состоит из 68 генов, 52 видов и 15 554 однозначно выровненных позиций. Далее мы удалили все позиции, для которых Convoluta отсутствовала (из-за частичных последовательностей генов), в результате чего было получено окончательное выравнивание 11 959 позиций. Выравнивание доступно по запросу в компании H.P. ([email protected]).
Химерические операционные таксономические единицы (OTU)
Чтобы увеличить объем информации, мы создали химерические последовательности, используя близкородственные таксоны в тех случаях, когда отсутствовали полноразмерные последовательности.Список химерных видов показан в Таблице S2. OTU выше видового уровня были названы в честь наиболее часто представленных видов в наборе данных для этой инклюзивной таксономической группы. Список 68 генов, использованных в этом исследовании, а также виды, отсутствующие для каждого гена, приведены в таблице S3.
Список 68 генов, использованных в этом исследовании, а также виды, отсутствующие для каждого гена, приведены в таблице S3.
Филогенетическая реконструкция
анализов PhyloBayes были выполнены с моделью смеси CAT, которая объясняет неоднородность между сайтами в процессе замены аминокислот [19].Было выполнено два независимых прогона общей продолжительностью 15 000 циклов (250 топологических ходов на цикл) с теми же операторами, что и в Lartillot et al. [20]. Первые 500 точек были отброшены как прожиг, а апостериорный консенсус был рассчитан для оставшихся 14 500 деревьев. Пропорции начальной загрузки были получены после 100 псевдоповторений, созданных с помощью SeqBoot [34]. Из соображений времени вычислений мы выполнили только 2000 циклов. Мы убедились, что 2000 циклов дали практически те же результаты, что и 14 500 циклов для полного набора данных.Кроме того, мы использовали консервативное значение приработки, равное 1000 (ручная проверка нескольких повторов показывает, что пригорание составляет менее 500). Деревья были собраны после сжигания, и для каждой повторности консенсус этих 1000 деревьев был рассчитан с помощью филобей. Эти 100 деревьев консенсуса передаются в CONSENSE [34] для вычисления значений поддержки начальной загрузки для каждого узла.
Деревья были собраны после сжигания, и для каждой повторности консенсус этих 1000 деревьев был рассчитан с помощью филобей. Эти 100 деревьев консенсуса передаются в CONSENSE [34] для вычисления значений поддержки начальной загрузки для каждого узла.
MP был проведен с использованием Paup* [35] с 10 случайными добавлениями видов, заменой ветвей TBR и MAXTREES = 1000.Проценты начальной загрузки MP были получены после 1000 повторений с использованием той же стратегии эвристического поиска с использованием Paup*.
Основанные на правдоподобии тесты альтернативных топологий были рассчитаны с использованием Consel [36]. Топология, полученная с помощью CAT-модели, использовалась в качестве основы, на которую были добавлены все возможные позиции Convoluta , что дало 99 (рисунок S3) и 91 дерево, когда Platyhelminthes были отброшены (рисунок S7). Длины ветвей ML альтернативных топологий были сначала выведены, предполагая конкатенированную модель WAG+F+Γ 4 с использованием Tree-Puzzle [33], затем значения логарифмического правдоподобия по сайтам были вычислены с помощью Codeml [37] и p-значения различные тесты на основе правдоподобия были рассчитаны с помощью Consel.
Взрывной поиск генной сигнатуры
Мы искали гены, которые могли бы быть информативными для филогенетического положения Convoluta , т.е. присутствующих у всех животных, включая Convoluta , но отсутствующих или сильно отличающихся от Platyhelminthes. Учитывая отсутствие полных геномов Lophotrochozoa, мы искали гены, отсутствующие у первичноротых. Мы использовали пять полных геномов из Deuterostomes ( Xenopus Tropicalis , Homo Sapiens , Takifugu Rubripes , CIONA Intestinalis , Shipa Cionalocentrotus Purpuratus ) и четыре из протокуров ( Caenorhabditis elegans , APIS MELIFERA , Anopheles gambiae и Drosophila melanogaster ).Мы провели поиск сходства с помощью blastx для 1664 контигов из Convoluta в сравнении с этими геномами и сохранили 80 последовательностей, которые демонстрируют на 10% больше сходства с вторичноротыми, чем с первичноротыми, и десять последовательностей, которые демонстрируют на 10% больше сходства с первичноротыми, чем с вторичноротыми. Разница в 10% гарантирует, что сходство последовательностей является хорошим индикатором филогенетической близости (неопубликованное наблюдение). Эти 90 последовательностей подвергали взрыву (blastx) против nr-секции NCBI. Ручная проверка вывода позволила исключить многочисленные ложные срабатывания (т.г. гены, потерянные у насекомых или нематод, но присутствующие у Platyhelminthes или моллюсков), что дает 40 генов для более тщательного анализа. Затем мы добавили последовательности дополнительных видов (все виды, используемые для нашего филогеномного дерева). Только один ген присутствовал у Convoluta и только у повторноротых, но не у первичноротых: гуанидиноацетат-N-метилтрансфераза. Мы провели аналогичный анализ для проверки базального положения Convoluta [5], [8], [9]. Мы искали гены, которые присутствуют у Convoluta и у Nematostella (http://genome.jgi-psf.org/Nemve1/Nemve1.home.html), но отсутствует в Bilateria. Однако не было обнаружено генов, которые благоприятствовали бы сестринской группе Convoluta для всех Bilateria.
Разница в 10% гарантирует, что сходство последовательностей является хорошим индикатором филогенетической близости (неопубликованное наблюдение). Эти 90 последовательностей подвергали взрыву (blastx) против nr-секции NCBI. Ручная проверка вывода позволила исключить многочисленные ложные срабатывания (т.г. гены, потерянные у насекомых или нематод, но присутствующие у Platyhelminthes или моллюсков), что дает 40 генов для более тщательного анализа. Затем мы добавили последовательности дополнительных видов (все виды, используемые для нашего филогеномного дерева). Только один ген присутствовал у Convoluta и только у повторноротых, но не у первичноротых: гуанидиноацетат-N-метилтрансфераза. Мы провели аналогичный анализ для проверки базального положения Convoluta [5], [8], [9]. Мы искали гены, которые присутствуют у Convoluta и у Nematostella (http://genome.jgi-psf.org/Nemve1/Nemve1.home.html), но отсутствует в Bilateria. Однако не было обнаружено генов, которые благоприятствовали бы сестринской группе Convoluta для всех Bilateria.
Дополнительная информация
Рисунок S1.
Дерево максимальной экономии, полученное из 11 959 однозначно выровненных аминокислотных позиций с PAUP. Надежность филогенетического вывода оценивалась по 1000 бутстреп-репликациям. Узлы, поддерживаемые 100% начальной загрузкой, обозначены черными кружками, а более низкие значения указаны явно.Масштабная линейка показывает количество изменений.
https://doi.org/10.1371/journal.pone.0000717.s004
(0,02 МБ PDF)
Рисунок S2.
Максимальное экономное дерево, полученное из 11 959 однозначно выровненных аминокислотных позиций без быстро развивающейся оболочника Oikopleura. Надежность филогенетического вывода оценивалась по 1000 бутстреп-репликациям. Узлы, поддерживаемые 100% начальной загрузкой, обозначены черными кружками, а более низкие значения указаны явно.Масштабная линейка показывает количество изменений.
https://doi.org/10.1371/journal.pone.0000717.s005
(0,02 МБ PDF)
Рисунок S3.

Сравнение всех возможных размещений Convoluta внутри Holozoa. Возможные положения Convoluta были проверены с помощью программы CONSEL (Shimodaira & Hasegawa 2001) на выравнивании 11 959 положений с моделью WAG+Γ. Из 99 возможных позиций только три не были отклонены тестом AU. Номер топологии, указанный во втором первом столбце, сообщается в топологии, чтобы указать положение Convoluta.
https://doi.org/10.1371/journal.pone.0000717.s006
(0,02 МБ PDF)
Рисунок S4.
Байесовское дерево, полученное из 11 959 однозначно выровненных аминокислотных позиций без Convoluta с использованием CAT-модели. Надежность филогенетического вывода оценивалась по 100 бутстрепным повторам. Узлы, поддерживаемые значениями начальной загрузки 100%, обозначены черными кружками, а более низкие значения указаны явно. Масштабная линейка показывает количество изменений на сайте.
https://doi.org/10.1371/journal.pone.0000717.s007
(0,02 МБ PDF)
Рисунок S5.

Байесовское дерево, полученное из 11 959 однозначно выровненных позиций аминокислот без внешней группы (Ichthyosporea, Choanoflageltata, Porifera и Cnidaria) с использованием модели CAT. Надежность филогенетического вывода оценивалась по 100 бутстрепным повторам. Узлы, поддерживаемые значениями начальной загрузки 100%, обозначены черными кружками, а более низкие значения указаны явно.Масштабная линейка показывает количество изменений на сайте.
https://doi.org/10.1371/journal.pone.0000717.s008
(0,01 МБ PDF)
Рисунок S6.
Байесовское дерево, полученное из 11 959 однозначно выровненных аминокислотных позиций без Convoluta и внешней группы (Ichthyosporea, Choanoflagellata, Porifera и Cnidaria) с использованием CAT-модели. Надежность филогенетического вывода оценивалась с помощью 100 бутстрепных повторов. Узлы, поддерживаемые значениями начальной загрузки 100%, обозначены черными кружками, а более низкие значения указаны явно.Масштабная линейка показывает количество изменений на сайте.
https://doi.org/10.1371/journal.pone.0000717.s009
(0,01 МБ PDF)
Рисунок S7.
Сравнение всех возможных мест размещения Convoluta внутри Holozoa при удалении Platyhelminthes. Возможные положения Convoluta были проверены с помощью программы CONSEL (Shimodaira & Hasegawa 2001) на выравнивании 11 959 положений с моделью WAG+Γ. Из 91 возможной позиции шесть не были отклонены тестом АС.Номер топологии, указанный во втором столбце, сообщается о топологии, чтобы указать положение Convoluta.
https://doi.org/10.1371/journal.pone.0000717.s010
(0,02 МБ PDF)
Благодарности
Мы хотели бы поблагодарить Peter Ladurner (Университет Инсбрука, Австрия) за предоставление образцов Convoluta pulchra и Ketil Malde (Университет Бергена, Норвегия) за помощь в анализе данных EST. Réseau Québecois de Calcul de Haute Performance, Суперкомпьютерный центр Барселоны (Mare nostrum) и Центр суперкомпьютеров Каталонии получили признание за вычислительные ресурсы.
Вклад авторов
Идея и разработка экспериментов: HP JB. Выполняли опыты: ПМ МР. Проанализированы данные: HP HB. Предоставленные реагенты/материалы/инструменты для анализа: HP JB PM MR. Написал газету: HP JB.
Каталожные номера
- 1. Хайман Л. (1951) Беспозвоночные. Том. II. Platyhelminthes и Rhynchocoela. Нью-Йорк: Макгроу-Хилл.
- 2. Петерсон К.Дж., Эрнисс Д.Дж. (2001)Филогения животных и происхождение билатеральных: выводы на основе морфологии и последовательностей генов 18S рДНК.Эволюция и развитие 3: 170–205.
- 3. Smith JPS, Tyler S, Rieger RM (1986) Является ли Turbellaria полифилетическим? Гидробиология 132: 71–78.
- 4. Baguñà J, Riutort M (2004) Рассвет билатеральных животных: случай ацеломорфных плоских червей. Биоэссе 26: 1046–1057.
- 5.
Руис-Трилло I, Риуторт М., Литтлвуд Д.Т., Херниу Э.А., Багунья Дж. (1999) Плоские черви Acoel: самые ранние из сохранившихся билатеральных многоклеточных животных, а не представители Platyhelminthes.
 Наука 283: 1919–1923.
Наука 283: 1919–1923. - 6. Jondelius U, Ruiz-Trillo I, Baguñà J, Riutort M (2002) Nemertodermatida являются базальными билатериями, не входящими в Platyhelminthes. Zool Scr 31: 201–215.
- 7. Telford MJ, Lockyer AE, Cartwright-Finch C, Littlewood DT (2003)Филогения комбинированных больших и малых субъединиц рибосомной РНК подтверждают базальное положение ацеломорфных плоских червей. Proc R Soc Lond B Biol Sci 270: 1077–1083.
- 8. Руис-Трильо И., Папс Дж., Лукота М., Рибера С., Йонделиус У. и др.(2002) Филогенетический анализ последовательностей тяжелой цепи миозина типа II подтверждает, что Acoela и Nemertodermatida являются базальными билатериями. Proc Natl Acad Sci U S A 99: 11246–11251.
- 9. Ruiz-Trillo I, Riutort M, Fourcade HM, Baguñà J, Boore JL (2004) Данные митохондриального генома подтверждают базальное положение Acoelomorpha и полифилию Platyhelminthes. Мол Филогенет Эвол 33: 321–332.
- 10.
 Felsenstein J (1978) Случаи, когда методы экономии или совместимости будут явно вводить в заблуждение.Сист Зоол 27: 401–410.
Felsenstein J (1978) Случаи, когда методы экономии или совместимости будут явно вводить в заблуждение.Сист Зоол 27: 401–410. - 11. Adoutte A, Balavoine G, Lartillot N, Lespinet O, Prud’homme B, et al. (2000) Новая филогения животных: достоверность и последствия. Proc Natl Acad Sci U S A 97: 4453–4456.
- 12. Галаныч К.М. (2004) Новый взгляд на филогению животных. Ежегодный обзор экологической эволюции и систематики 35: 229–256.
- 13. Делсук Ф., Бринкманн Х., Филипп Х. (2005) Филогеномика и реконструкция древа жизни.Nat Rev Genet 6: 361–375.
- 14. Baurain D, Brinkmann H, Philippe H (2007) Отсутствие разрешения в филогении животных: близко расположенные кладогенезы или необнаруженные систематические ошибки? Мол Биол Эвол 24: 6–9.
- 15. Филипп Х., Телфорд М.Дж. (2006) Крупномасштабное секвенирование и новая филогения животных. Тенденции Ecol Evol 21: 614–620.
- 16.
Delsuc F, Brinkmann H, Chourrout D, Philippe H (2006)Оболочечные, а не головохордовые, являются ближайшими живыми родственниками позвоночных.
 Природа 439: 965–968.
Природа 439: 965–968. - 17. Филипп Х., Чжоу Ю., Бринкманн Х., Родриг Н., Делсук Ф. (2005) Гетеротахия и притяжение длинных ветвей в филогенетике. БМС Эвол Биол 5: 50.
- 18. Felsenstein J (1981) Эволюционные деревья из последовательностей ДНК: подход максимального правдоподобия. Дж Мол Эвол 17: 368–376.
- 19. Lartillot N, Philippe H (2004)Байесовская смешанная модель межсайтовой неоднородности в процессе замены аминокислот. Мол Биол Эвол 21: 1095–1109.
- 20. Lartillot N, Brinkmann H, Philippe H (2007)Подавление артефактов притяжения длинных ветвей в филогении животных с использованием гетерогенной модели сайта. BMC Evol Biol 7: Приложение 1S4.
- 21. Бурла С.Дж., Юлиусдоттир Т., Лоу С.Дж., Фриман Р., Ароновиц Дж. и др. (2006) Филогения вторичноротых выявляет монофилетических хордовых и новый тип Xenoturbellida. Природа 444: 85–88.
- 22.
Matus DQ, Copley RR, Dunn CW, Hejnol A, Eccleston H, et al.
 (2006) Обширная выборка таксонов и генов указывает на то, что хетогнаты являются первичноротыми. Курр Биол 16: R575–576.
(2006) Обширная выборка таксонов и генов указывает на то, что хетогнаты являются первичноротыми. Курр Биол 16: R575–576. - 23. Philippe H, Lartillot N, Brinkmann H (2005) Мультигенный анализ двусторонних животных подтверждает монофилию Ecdysozoa, Lophotrochozoa и Protostomia. Мол Биол Эвол 22: 1246–1253.
- 24. Marletaz F, Martin E, Perez Y, Papillon D, Caubit X и другие. (2006) Филогеномика Chaetognath: первичноротый с вторичноротым развитием.Курр Биол 16: R577–578.
- 25. Brinkmann H, Giezen M, Zhou Y, Raucourt GP, Philippe H (2005) Эмпирическая оценка артефактов притяжения длинных ветвей в глубокой эукариотической филогеномике. Сист биол 54: 743–757.
- 26. Axe P (1996) Многоклеточные животные. Новый подход к филогенетическому порядку в природе. Берлин: Спрингер.
- 27. Элерс У (1985) Das Phylogenetische System der Plathelminthes. Штутгарт, Нью-Йорк: Г. Фишер.
- 28.Лундин К. (1998) Новый взгляд на эпидермальные ресничные корешки Xenoturbella bocki (Xenoturbellida): новая поддержка возможного родства с Acoelomorpha (Platyhelminthes).
 Zoologica Scripta 27: 263–270.
Zoologica Scripta 27: 263–270. - 29. Philippe H, Snell EA, Bapteste E, Lopez P, Holland PW, et al. (2004) Филогеномика эукариот: влияние недостающих данных на большие выравнивания. Мол Биол Эвол 21: 1740–1752.
- 30. Philippe H (1993) MUST, компьютерный пакет утилит управления последовательностями и деревьями.Нуклеиновые кислоты Рез. 21: 5264–5272.
- 31. Castresana J (2000) Выбор консервативных блоков из нескольких выравниваний для их использования в филогенетическом анализе. Мол Биол Эвол 17: 540–552.
- 32. Рур Б., Родригес-Эспелета Н., Филипп Х. (2007) SCaFoS: инструмент для выбора, объединения и слияния последовательностей для филогеномики. BMC Evol Biol 7: Приложение 1S2.
- 33. Schmidt HA, Strimmer K, Vingron M, von Haeseler A (2002) TREE-PUZZLE: филогенетический анализ максимального правдоподобия с использованием квартетов и параллельных вычислений.Биоинформатика 18: 502–504.
- 34.
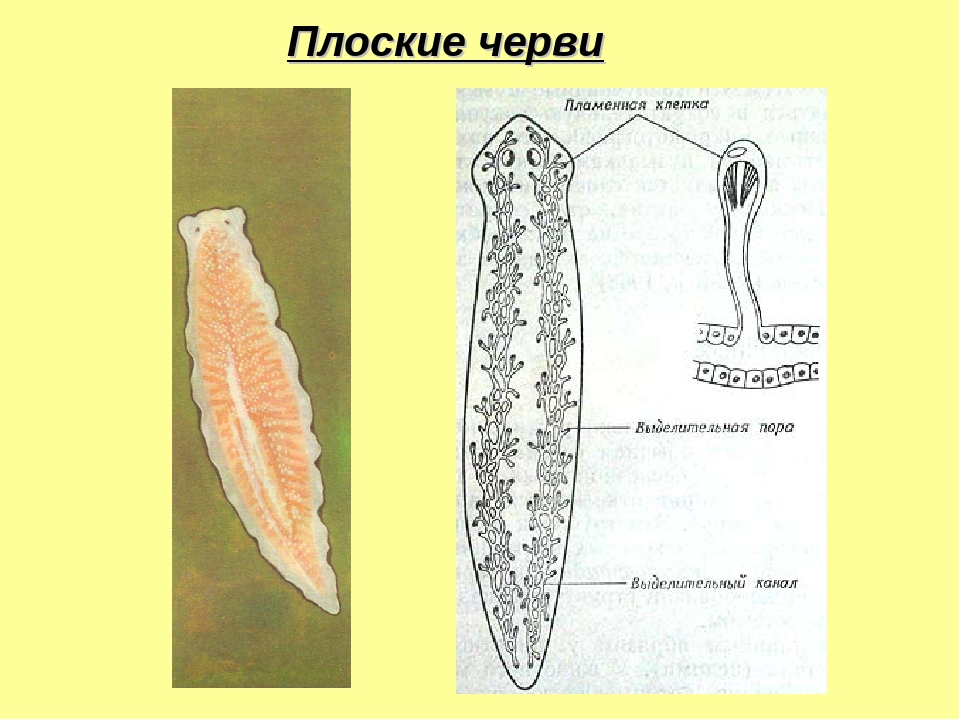 Felsenstein J (2001) PHYLIP (Phylogene Inference Package). Сиэтл: Факультет генетики Вашингтонского университета. 3.6 изд.: Распространяется автором.
Felsenstein J (2001) PHYLIP (Phylogene Inference Package). Сиэтл: Факультет генетики Вашингтонского университета. 3.6 изд.: Распространяется автором. - 35. Swofford DL (2000) PAUP*: Филогенетический анализ с использованием экономных и других методов. Сандерленд, Массачусетс: Синауэр. 4б10 изд:.
- 36. Shimodaira H, Hasegawa M (2001) CONSEL: для оценки достоверности выбора филогенетического дерева. Биоинформатика 17: 1246–1247.
- 37. Yang Z (1997) PAML: пакет программ для филогенетического анализа по максимальному правдоподобию. Comput Appl Biosci 13: 555–556.
Гигантские плоские черви вторглись во Францию и вышли из-под контроля в течение 2 десятилетий, прежде чем ученые это поняли
Оглядываясь назад, можно было заметить признаки.
В 2013 году группу детсадовцев напугали сотни «маленьких змей» на траве в их школе. Женщина, несомненно, в ужасе обнаружив длинного червя на шерсти своей кошки, отнесла его к местному ветеринару, но тот сказал, что это ленточный червь.
Другие люди обнаружили странных червей, которых они показывали местным университетам, но от них отмахивались, называя их пиявками или другими «простыми, неинтересными животными». А еще в 2005 году один ученый действительно опубликовал отчет о странной новой группе червей во Франции, но сделал это в малоизвестном микологическом журнале. Биологи-беспозвоночные их не читают (в их защиту, как и все остальные).
Тем не менее, есть некоторые вещи, которые, как вы ожидаете, ученые заметят довольно быстро.Скажем… хищные плоские черви-молоты длиной в фут, выделяющие токсины, в месте, где раньше не было плоских червей-молотов.
Экспонат А:
Кредит: Жюстин и др. 2018Это не то животное, которое легко спутать с любым другим.
Но каким-то образом именно такое существо пробиралось из поля зрения французских ученых более двух десятков лет, несмотря на вышеупомянутые многочисленные сообщения граждан, которые были чертовски уверены, что видят что-то новое.
«Мы до сих пор поражены полным отсутствием реакции со стороны научных органов на присутствие этих червей», — написали авторы нового исследования PeerJ .
Как это произошло — комментарий грустный, на мой взгляд, но об этом позже.
Сначала пикантные подробности.
Наземные плоские черви, как следует из названия, имеют плоскую форму и обладают уникальной «ползучей подошвой» на нижней стороне. На этой подошве лежит толстый ковер из мощных ресничек, которые бьются взад-вперед по слизистой полоске, отложенной особыми железами. Иногда они могут использовать эту слизь, чтобы спуститься на землю на веревке, как это делают слизни.
Они охотятся и выслеживают добычу по запаху и используют липкие, вероятно, ядовитые выделения, чтобы усмирить ее.Как только еда получена, они поедают свою добычу, вводя пищеварительные ферменты, а затем высасывая полученные соки. Их предпочтительными блюдами являются обитающие в почве животные, такие как дождевые черви, слизни, протисты, круглые черви и личинки насекомых.
В Европе обитают наземные плоские черви, но их длина обычно не превышает одного сантиметра. С другой стороны, инвазивные плоские черви-молоты из Юго-Восточной Азии измеряются в дюймах, а не в миллиметрах. Некоторые — по-видимому, даже во Франции — измеряются в футах.
В новой статье в журнале PeerJ группа французских и австралийских ученых проанализировала четырехлетние отчеты гражданских ученых о червях. Они также сравнили собранные гены цитохромоксидазы I червей, чтобы увидеть, сколько видов присутствует и насколько разнообразен каждый вид.
Собрав все вместе, они определили, что во Франции существует по крайней мере пять видов инвазивных плоских червей-молотов, три из названных видов и два безымянных.
Внутри видов плоские черви представляют собой клоны, поскольку их последовательности цитохромоксидазы показали 0% внутривидовой изменчивости. То, как молотоголовые клонируют себя, носит восхитительно причудливое название «ножницы». За этим напыщенным названием процесс сводится к тому, что червь склеивает свой хвост, тянет и отрывает еще извивающийся кусок, который продолжает отращивать голову и ладить с ней. Знаешь, как и ты.
То, как молотоголовые клонируют себя, носит восхитительно причудливое название «ножницы». За этим напыщенным названием процесс сводится к тому, что червь склеивает свой хвост, тянет и отрывает еще извивающийся кусок, который продолжает отращивать голову и ладить с ней. Знаешь, как и ты.
В своем доме в тропической и субтропической Юго-Восточной Азии молотоголовые плоские черви размножаются половым путем, хотя по причинам, которые мне не удалось обнаружить, они этого не делают в местах, где они вторглись.Это отсутствие пола объясняет отсутствие генетического разнообразия, но, кажется, это совершенно не беспокоит плоских червей (что интересно, то же самое верно и для короля инвазивных видов, одуванчика*). Нулевое генетическое разнообразие внутри видов также, по-видимому, означает, что каждый вид был завезен во Францию один раз.
Как черви попали во Францию? Это легко. Они попали туда тем же путем, что и во всем мире, благодаря великому способу проникновения инвазивных видов — торговле питомниками.Они путешествовали в почве горшечных растений, перевозимых между странами.
Ранее ученые думали, что инвазивные плоские черви могут выжить только в Европе в теплицах, где они были впервые зарегистрированы в 1878 году в теплице Королевского ботанического сада в Кью. Но видео, снятое во французском городе Уркюит в 1999 году и полученное этими учеными, ясно показывает, что Bipalium kewense смывает эту теорию в унитаз. С тех пор количество сообщений увеличилось.
Хотя черви в настоящее время зарегистрированы на большей части территории Франции, они, похоже, предпочитают юго-западный регион под названием Атлантические Пиренеи, который авторы называют «маленьким раем для инвазивных наземных планарий».В этом регионе мягкие зимы и влажная погода, благоприятные для плоских червей. Один житель наблюдал несколько видов плоских червей-молотов в сентябре 2017 г. , декабре 2017 г. и январе 2018 г. живыми на открытом воздухе в почве сада. Понятно, что европейская зима им там не помеха.
, декабре 2017 г. и январе 2018 г. живыми на открытом воздухе в почве сада. Понятно, что европейская зима им там не помеха.
Оглядываясь назад, не должно быть ничего удивительного в том, что черви были обнаружены во Франции, учитывая, что на протяжении десятилетий было известно, что четыре вида вторглись в субтропические штаты США: Флориду, Джорджию, Северную и Южную Каролину, Луизиану, Техас и даже Калифорнию. .Южная Франция также считается субтропической.
Я сам видел молотоголового плоского червя, когда жил в Техасе несколько лет назад, и, озадаченный его странной головой и характерным внешним видом, я смог выяснить, что это такое, с помощью быстрого поиска в Google. Никто во Франции не делал этого в течение 20 лет, а если и делал, то не думал и не знал, что это достойно внимания или публикации?
Неспособность французского научного сообщества распознать этих червей на протяжении более двух десятилетий является печальным комментарием к состоянию естествознания и образования в области разнообразия среди биологов не только во Франции — историческом оплоте исследований разнообразия (одна из моих любимых прерий). цветы, Gaillardia , был назван в честь французского покровителя ботаники) — но везде.
цветы, Gaillardia , был назван в честь французского покровителя ботаники) — но везде.
Авторы списывают это на «невежество», но я пойду дальше. У меня сильные чувства на эту тему.
В течение 20 лет биологам неоднократно не хватало знаний в области естественной истории даже для того, чтобы знать, что они видят что-то новое и необычное , такой поворот событий я нахожу постыдным. Однако не специалистам должно быть стыдно. Именно образовательные учреждения и финансирующие агентства пожертвовали образованием и исследованиями разнообразия на алтарь молекулярной биологии.
Я буду первым, кто признает важность молекулярной биологии — даже жизненно важную. Я был ассистентом преподавателя по вводной биохимии в колледже и постоянно включаю молекулярную и биохимическую информацию в сообщения в этом блоге. Без него невозможно понять биологию.
Но разнообразие и естественная история также важны. Я уже подробно писал здесь ранее о том, почему, но достаточно сказать, что, когда биологи не могут определить, что большой ядовитый хищный плоский червь является чем-то шокирующе новым, мы пришли к печальному положению вещей. Необходимо уделить больше внимания биологическому образованию и финансированию исследований по этой важной теме. Это должно.
Необходимо уделить больше внимания биологическому образованию и финансированию исследований по этой важной теме. Это должно.
Прибытие плоских червей во Францию вызывает тревогу, говорят авторы, потому что, как мы видели, плоские черви-молоты являются крупными хищниками. Они могут быть крупнейшими беспозвоночными хищниками в тех районах, куда они вторгаются, что говорит о том, что их влияние на местных беспозвоночных «не пренебрежимо мало», хотя никто еще не знает наверняка. Зловещая французская команда сфотографировала плоского червя, нападающего на местного дождевого червя (в Северной Америке около трети групп дождевых червей сами являются инвазивными, так что….. маленькая серебряная подкладка). Какими еще местными беспозвоночными они охотно лакомятся?
В конце концов, спасением Франции могут стать сами плоские черви. Они «свои злейшие враги», сообщает Университет Флориды IFAS Extension, по очень простой причине: они полные энтузиазма каннибалы.
______________
*сильный аргумент можно привести и в пользу человека, хотя в пользу одуванчиков мы еще не приобрели устойчивость к Раундапу или переносимому ветром потомству
№ по каталогу
Жюстин Дж. , Винсор Л., Гей Д., Грос П., Тевено Дж.(2018) «Гигантские черви chez moi! Плоские черви-молоты (Platyhelminthes, Geoplanidae, Bipalium spp., Diversibipalium spp.) в метрополии Франции и заморских французских территориях». PeerJ 6:e4672 https://doi.org/10.7717/peerj.4672
, Винсор Л., Гей Д., Грос П., Тевено Дж.(2018) «Гигантские черви chez moi! Плоские черви-молоты (Platyhelminthes, Geoplanidae, Bipalium spp., Diversibipalium spp.) в метрополии Франции и заморских французских территориях». PeerJ 6:e4672 https://doi.org/10.7717/peerj.4672
Неместные плоские черви / RHS Садоводство
Наземные плоские черви (Platyhelminthes) представляют собой группу (класс животных) свободноживущих червей. Это маленькие, сплюснутые животные, часто с лентовидным внешним видом и несегментированным телом.Они покрыты слизью и обычно оставляют след. Они встречаются по всему миру, в Великобритании и Ирландии насчитывается около четырех видов, считающихся местными, и, вероятно, 17 неместных видов, прижившихся из южного полушария.
Большинство неместных видов плоских червей питаются дождевыми червями, слизнями, улитками и другими почвенными организмами. Австралийские и новозеландские плоские черви — это два вида, которые прижились и широко распространились в Великобритании и Ирландии. Оба вида специализируются на дождевых червях, и они могут серьезно сократить популяции некоторых видов дождевых червей и, следовательно, повлиять на экосистему почвы. Однако несколько других видов были случайно интродуцированы, в том числе два вида 90 626 Kontikia , и есть свидетельства того, что неместные плоские черви продолжают интродуцироваться, включая плоских червей Обамы
Оба вида специализируются на дождевых червях, и они могут серьезно сократить популяции некоторых видов дождевых червей и, следовательно, повлиять на экосистему почвы. Однако несколько других видов были случайно интродуцированы, в том числе два вида 90 626 Kontikia , и есть свидетельства того, что неместные плоские черви продолжают интродуцироваться, включая плоских червей Обамы
Австралийский плоский червь ( Australplana sanguinea ) лососево-розовый, заостренный с обоих концов и достигает 2-8 см в длину.Впервые он был зарегистрирован на островах Силли в 1980-х годах и с тех пор получил широкое распространение в южной Англии и Уэльсе.
Новозеландский плоский червь ( Arthurdendyus triangulates ) достигает 20 см (8 дюймов) в длину и имеет темно-коричневый цвет с более светлыми краями. Он прибыл в Великобританию, вероятно, с импортированными растениями в 1960-х годах и с тех пор получил широкое распространение. Он питается исключительно дождевыми червями и способен сокращать популяцию дождевых червей. Это оказывает нежелательное воздействие на структуру почвы, а также лишает дождевых червей источника пищи для тех местных животных, которые ими питаются.Этот плоский червь происходит из Новой Зеландии и в настоящее время процветает в Шотландии, Северной Англии и Северной Ирландии.
Он питается исключительно дождевыми червями и способен сокращать популяцию дождевых червей. Это оказывает нежелательное воздействие на структуру почвы, а также лишает дождевых червей источника пищи для тех местных животных, которые ими питаются.Этот плоский червь происходит из Новой Зеландии и в настоящее время процветает в Шотландии, Северной Англии и Северной Ирландии.
Плоский червь Обамы ( Обама nungara ), впервые был зарегистрирован в 2014 году, он произрастает в Южной Америке и распространяется по Европе. Его название означает « листовое животное ». Его длина составляет до 7 см, цвет тела от светло-коричневого до черного с бледной средней линией и небольшими черными отметинами по всему телу. Питается дождевыми червями и улитками. Этот вид считается чрезвычайно инвазивным и характеризуется как наиболее опасный вид плоских червей для почвенной экосистемы и местных почвенных организмов в настоящее время в Европе.
Черно-коричневые плоские черви Kontikia ( Kontikia ventrolineta и Kontikia andersoni ) маленькие (1-2,5 см в длину и 1-2 мм в ширину), родом из Австралии и Новой Зеландии. Kontikia ventrolineata имеет черный цвет с двумя узкими серыми линиями, первое сообщение в Британии было получено из frlomLIverpool в 1994 г. Известно, что она питается мелкими улитками и, возможно, слизнями. Kontikia andersoni имеет бледно-коричневый цвет с тремя линиями более темных пятен вдоль тела.
Kontikia ventrolineata имеет черный цвет с двумя узкими серыми линиями, первое сообщение в Британии было получено из frlomLIverpool в 1994 г. Известно, что она питается мелкими улитками и, возможно, слизнями. Kontikia andersoni имеет бледно-коричневый цвет с тремя линиями более темных пятен вдоль тела.
Из-за их потенциального воздействия на популяции дождевых червей и других почвенных животных интродукция неместных плоских червей представляет большой риск для сельского хозяйства и садоводства. Таким образом, все неместные плоские черви включены в Приложение 9 Закона о дикой природе и сельской местности 1981 года, в котором говорится, что интродукция или выпуск их в дикую природу является правонарушением. Кроме того, новозеландский плоский червь включен в список инвазивных чужеродных видов Европейского союза.
Если вы обнаружите неместных плоских червей, о них можно сообщить в Секретариат неместных.
Аквариум Вайкики » Плоские черви
Плоские черви принадлежат к отдельному типу. Для них характерно чрезвычайно плоское несегментированное тело с простым головным концом. Их тела настолько плоские, что кислород может проникать в ткани без жабр или других дыхательных структур. У них также отсутствует система кровообращения, но кишечник сильно разветвлен, поэтому переваренные питательные вещества циркулируют по всем частям тела. Кишечник имеет только одно отверстие, поэтому то, что не может быть переварено, должно быть удалено через ротовое отверстие.
Для них характерно чрезвычайно плоское несегментированное тело с простым головным концом. Их тела настолько плоские, что кислород может проникать в ткани без жабр или других дыхательных структур. У них также отсутствует система кровообращения, но кишечник сильно разветвлен, поэтому переваренные питательные вещества циркулируют по всем частям тела. Кишечник имеет только одно отверстие, поэтому то, что не может быть переварено, должно быть удалено через ротовое отверстие.
Плоские черви могут достигать только 1-2 дюймов в длину, но являются ненасытными охотниками и падальщиками на рифе. Большинство питается более мелкими червями, улитками или двустворчатыми моллюсками. Они обнаруживают добычу, используя химические датчики на головном конце тела, либо специальные сенсорные пятна, либо складки вдоль переднего края тела и крошечные антенноподобные выступы сразу за краем. Они подчиняют добычу, обвивая ее своим гибким телом, нагнетая пищеварительные ферменты на жертву (или в ее раковину, если это улитка или устрица) с помощью трубчатого выступа рта (глотки). После того, как мясо предварительно переварено и разжижено, оно перекачивается в кишечник для дальнейшего переваривания и распределения.
После того, как мясо предварительно переварено и разжижено, оно перекачивается в кишечник для дальнейшего переваривания и распределения.
Плоские черви ползают или скользят по дну, ударяя микроскопическими волосовидными выступами, называемыми ресничками, по следу слизи, который они откладывают. Тела некоторых червей имеют мускулистые края, которые они могут волновать, чтобы двигаться быстрее, и они могут даже плавать на короткие расстояния по течению, волнообразно двигая краем своего тела.
Хотя плоских червей иногда путают с морскими слизнями, они гораздо проще.Они очень плоские, у них нет хорошо развитой головы со сложными щупальцами и сенсорными структурами, а на верхней поверхности нет перистых жабр. Плоские черви гермафродиты, но оплодотворяются перекрестно. Оба партнера могут откладывать плоские спирали крошечных яиц на поверхности камней.
Редко встречающиеся на открытом воздухе плоские черви живут на нижней поверхности скал или в расщелинах приливных заводей и рифов.