
В чём состоит отличие вида от особи?
Подробней что такое вид
Это группа организмов с аналогичными особенностями. Все организмы одного конкретного вида обладают таким же количеством хромосом, что означает, что они имеют сходные морфологические, анатомические и физиологические характеристики. Поэтому экологические ниши более или менее одинаковы в каждом отдельном случае. Обычно конкретный вид обладает исключительными видоспецифическими особенностями, которые не встречаются у других видов.
Вид может быть далее разделен на подвиды, но между подвидами нет большой разницы. Согласно таксономии, может быть любое количество видов под одним родом, который действительно является предком вида.
Несколько слов про особь
Биологическое определение термина «Особь» представляет собой лицо одного и того же вида, живущих в одном и том же месте. Поскольку эти индивидуумы одной особи. Как правило, в определенной популяции скрещиваются, чтобы поддерживать численность населения, которая обеспечивает успешное развитие будущих поколений, и их вид сохраняется.
Виды подвергаются изменению со временем в соответствии с изменениями окружающей среды. Эти изменения происходят с точки зрения численности видов. Когда условия благоприятствуют организму, размер популяции увеличивается и уменьшается в противном случае. Успех конкретного вида можно было бы определить путем изучения изменения численности видов в течение периода времени.
В чем разница между видом и особью
- Виды формируются с разделения определенной популяции, которые делают его уникальным, тогда как особь формируется из человека или животных, с конкретными условиями среды обитания.
- Особь подвергается изменениям, но виды не меняются; если это так, образуется новый вид.
- Особь имеет временные и пространственные ограничения, но виды нет.

- Виды указываются с использованием определенной методологии, но нет конкретного набора правил для указания населения.
перевод в Русско-казахском терминологическом словаре «Биология» онлайн
Смотреть что такое ОСОБЬ в других словарях:
ОСОБЬ
(индивид) — реальная единица или существо, имеющее собственную действительность, в отличие от общих понятий, существующих только в мысли, и от групп, и… смотреть
ОСОБЬ
индивид, индивидуум (от лат. individuum — неделимое), неделимая единица жизни на Земле (разделить О. на части без потери «индивидуальности» нев… смотреть
ОСОБЬ
ОСОБЬ, -и, ж. (книжн.). Самостоятельно существующий организм,индивидуум. Многообразие особей растительного мира. О. женского пола.
ОСОБЬ
особь ж. Отдельный живой организм; индивид.
ОСОБЬ
особь Iж.individual IIособь статья разг. — quite another matter
ОСОБЬ
особь
См. единица, существо…
Словарь русских синонимов и сходных по смыслу выражений. — под. ред. Н. Абрамова, М.: Русские словари,1999.
особь
единица, существо; индивидуум, индивид; особняком, организм, отдельно, экземпляр
Словарь русских синонимов.
Особь
экземпляр
Словарь синонимов русского языка. Практический справочник. — М.: Русский язык.З. Е. Александрова.2011.
особь
сущ.
• индивидуум
• индивид
отдельно рассматриваемый живой организм)
Словарь русских синонимов. Контекст 5.0 — Информатик.2012.
особь
сущ., кол-во синонимов: 9
• индивид (13)
• индивидуум (15)
• организм (67)
• особняком (28)
• отдельно (59)
• самик (4)
• существо (37)
• человек (86)
• экземпляр (19)
Словарь синонимов ASIS.В.Н. Тришин.2013.
.
Синонимы:
индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр… смотреть
— под. ред. Н. Абрамова, М.: Русские словари,1999.
особь
единица, существо; индивидуум, индивид; особняком, организм, отдельно, экземпляр
Словарь русских синонимов.
Особь
экземпляр
Словарь синонимов русского языка. Практический справочник. — М.: Русский язык.З. Е. Александрова.2011.
особь
сущ.
• индивидуум
• индивид
отдельно рассматриваемый живой организм)
Словарь русских синонимов. Контекст 5.0 — Информатик.2012.
особь
сущ., кол-во синонимов: 9
• индивид (13)
• индивидуум (15)
• организм (67)
• особняком (28)
• отдельно (59)
• самик (4)
• существо (37)
• человек (86)
• экземпляр (19)
Словарь синонимов ASIS.В.Н. Тришин.2013.
.
Синонимы:
индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр… смотреть
ОСОБЬ
ОСОБЬ, индивид, индивидуум (от лат.
individuum — неделимое), неделимая единица жизни на Земле (разделить О.
на части без потери «индивидуальности» не. .. смотреть
.. смотреть
ОСОБЬ
Особь (индивид) — реальная единица или существо, имеющее собственную действительность, в отличие от общих понятий, существующих только в мысли, и от гр… смотреть
ОСОБЬ
ОСОБЬ (79) пр. и нар. I. Пр. нескл. Отдельный: тъ ѥсть дѣвьствьнъ. не иже тѣло своѥ хранить. ѿ съвъкѹплени˫а несквьрньно. нъ стыдѧисѧ ѥгда особь бываѥ… смотреть
ОСОБЬ
ОСОБЬ индивидуум, отдельный экземпляр (особь живого), элементарная единица жизни. В эволюционном смысле особь — существо, происходящее от одной зиго… смотреть
ОСОБЬ
1) о́собь -и, ж. Каждый отдельный, самостоятельно существующий организм; индивидуум.Человеческая особь. □ Каждый вид птиц и животных представлен [в Арк… смотреть
ОСОБЬ
ОСОБЬ индивид, индивидуум (от лат. individuum — неделимое), неделимая единица жизни. Самый существенный признак О.— строгая взаимозависимость отд. … смотреть
ОСОБЬ
I о/собь(3 ж), Р., Д., Пр. о/соби; мн. о/соби, Р. о/собей
II осо/бьосо/бь статья/
Синонимы:
индивид, индивидуум, организм, особняком, отдельно, сущес… смотреть
о/собей
II осо/бьосо/бь статья/
Синонимы:
индивид, индивидуум, организм, особняком, отдельно, сущес… смотреть
ОСОБЬ
Синонимы: индивид, индивидуум, рамет организм, образующийся в результате полового или вегетативного размножения, взаимосвязь частей которого обусловлен… смотреть
ОСОБЬ
I`особьж.individuo mмногообразие особей растительного мира — variedad de plantas del reino vegetalII ос`обьэто особь статья разг. — es un problema apar… смотреть
ОСОБЬ
1) Орфографическая запись слова: особь2) Ударение в слове: `особь3) Деление слова на слоги (перенос слова): особь4) Фонетическая транскрипция слова осо… смотреть
ОСОБЬ
Особь(индивид, индивидуум) — наименьшая ед. биол. вида, подверженная действию факторов эволюции. Термин О. в микробиологии используют в значении одиноч… смотреть
ОСОБЬ
(индивидуум, от лат. individuum — неделимое), самостоятельно существующий организм; наименьшая неделимая единица биологической) вида, подверженная дейс. .. смотреть
.. смотреть
ОСОБЬ
— каждое отдельное животное или растение какого-либо вида. Син.: индивидум, экземпляр. Геологический словарь: в 2-х томах. — М.: Недра.Под редакцией К…. смотреть
ОСОБЬ
— отдельный живой организм, обладающий всеми признаками, свойственными виду, к которому он принадлежит, и в то же время обладающий морфологическими и физиологическими особенностями, отличающими его от других организмов того же вида. <br><b>Синонимы</b>: <div> индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр </div><br><br>… смотреть
ОСОБЬ
— англ. individual; нем. Individuum. Индивид, каждый отдельный организм. О. свойственны все признаки вида, к к-рому она принадлежит, а также свои сооственные морфологические и физиолог, особенности, отличающие ее от других О. того же вида.
Antinazi.Энциклопедия социологии,2009
Синонимы:
индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр. .. смотреть
.. смотреть
ОСОБЬ
корень — ОСОБЬ; нулевое окончание;Основа слова: ОСОБЬВычисленный способ образования слова: Бессуфиксальный или другой∩ — ОСОБЬ; ⏰Слово Особь содержит с… смотреть
ОСОБЬ
1. о́собь, -и; мн. -и, -ей2. осо́бь; осо́бь статья́Синонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
ж. биол.individu m; exemplaire m (экземпляр)Синонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
(индивидуум) (от лат. individuum — неделимое), неделимая единица биол. вида. Для колониальных, вегетативно размножающихся и симбиотич. организмов понят… смотреть
ОСОБЬ
Особь второго сорта с неполным набором хромосом. Жарг. мол. Неодобр. Об очень глупом или умственно неполноценном человеке. Максимов, 290.Синонимы: инди… смотреть
ОСОБЬ
особьפּרָט ז’ [ר’ פּרָטִים, פּרָטֵי-/פִּרטֵי-]* * *אדםאישייחידייחודיפרטפרטניСинонимы:
индивид, индивидуум, организм, особняком, отдельно, существо, э.
ОСОБЬ
См. особыйВ. В. Виноградов.История слов,2010Синонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
1. о́собь, о́соби, о́соби, о́собей, о́соби, о́собям, о́собь, о́собей, о́собью, о́собями, о́соби, о́собях 2. особь (Источник: «Полная акцентуированная парадигма по А. А. Зализняку») . Синонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр… смотреть
ОСОБЬ
жIndividuum n, pl -duenСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
ж. биол. individu m; exemplaire m (экземпляр)
ОСОБЬ
(καθ᾽ ἑαυτόν) сам по себе, отдельно/Синонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
— — англ. individual; нем. Individuum. Индивид , каждый отдельный организм . О. свойственны все признаки вида, к к-рому она принадлежит, а также свои сооственные морфологические и физиолог, особенности, отличающие ее от других О.
ОСОБЬ
особь, ′особь, -и, ж. (книжн.). Самостоятельно существующий организм, индивидуум. Многообразие особей растительного мира. О. женского пола.II. ОС′ОБЬ (… смотреть
ОСОБЬ
I `особь ж. биол. individuo m, organismo m II ос`обь •• особь статья прост. — è un altro paio di maniche Итальяно-русский словарь.2003. Синонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр… смотреть
ОСОБЬ
элементарная, неделимая единица живого. Начала современного естествознания. Тезаурус. — Ростов-на-Дону.В.Н. Савченко, В.П. Смагин.2006. Синонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр… смотреть
ОСОБЬ
ОСОБЬ (индивидуум) (от лат . individuum — неделимое), наименее неделимая единица биологического вида. Для колониальных, вегетативно размножающихся и симбиотических организмов понятие «особь» относительно.<br><br><br>.
ОСОБЬ
ОСОБЬ нескл. прил. (старин.). Только в выражении: особь статья (разг.) — совсем иное, то, что не следует смешивать с остальным. Дело об лесе. сказал секретарь на ушко покупателю, особь статья. Даль.<br><br><br>… смотреть
ОСОБЬ
ОСОБЬ (индивидуум) (от лат. individuum — неделимое) — наименее неделимая единица биологического вида. Для колониальных, вегетативно размножающихся и симбиотических организмов понятие «особь» относительно.<br>… смотреть
ОСОБЬ
жindivíduo m, tipo mСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
— (индивидуум) (от лат. individuum — неделимое) — наименее неделимаяединица биологического вида. Для колониальных, вегетативно размножающихсяи симбиотических организмов понятие «»особь»» относительно…. смотреть
ОСОБЬ
биологич.egyedСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
особь ж Individu|um ( — » v i : — ] n 1, pl -du|enСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
жbireyСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
‘особь, -иСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
enkeltvesenСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
personСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
ОСОБЬ особи, ж. (науч.). Каждый самостоятельно существующий организм, индивид. Белуга очень крупная рыба: отдельные особи достигают 1.200 кг.
(науч.). Каждый самостоятельно существующий организм, индивид. Белуга очень крупная рыба: отдельные особи достигают 1.200 кг.
ОСОБЬ
Ударение в слове: `особьУдарение падает на букву: оБезударные гласные в слове: `особь
ОСОБЬ
особь статьяСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
个体 gètǐСинонимы: индивид, индивидуум, организм, особняком, отдельно, существо, экземпляр
ОСОБЬ
ОСОБЬ, -и, ж. (книжное). Самостоятельно существующий организм, индивидуум. Многообразие особей растительного мира. Особь женского пола.
ОСОБЬ
I [c darkred]жен. асобіна, жен. каліва, ср.II [c darkred]особь статья разг. — справа асобая
ОСОБЬ
(син.: индивид, индивидуум)отдельный живой организм.
ОСОБЬ
1) individual 2) specimen 3) animal unit
ОСОБЬ
неделимая единица жизни, морфофизиологическая единица, происходящая у видов с половым размножением из одной зиготы.
ОСОБЬ
ж. биол. individual— гетерозиготная особь — гомозиготная особь
биол. individual— гетерозиготная особь — гомозиготная особь
ОСОБЬ
о’собь, о’соби, о’соби, о’собей, о’соби, о’собям, о’собь, о’собей, о’собью, о’собями, о’соби, о’собях
ОСОБЬ
особь единица, существо, индивидуум, индивид, особняком, организм, отдельно, экземпляр
ОСОБЬ
ОСОБЬ: особь статья (прост.) — что-нибудь совсем особое, совсем иное. Это дело — особь статья.
ОСОБЬ
особь (син.: индивид, индивидуум) — отдельный живой организм.
ОСОБЬ
I о’собь асобіна, каліва, II осо’бь особь статья разг. — справа асобая
ОСОБЬ
Начальная форма — Особь, неизменяемое, неодушевленное, одушевленное
ОСОБЬ
I. особень (-бня), особина. II. Это особь статья це инша річ.
ОСОБЬ
— Каждое отдельное животное или растение какого-либо вида.
ОСОБЬ
(син.: индивид, индивидуум) отдельный живой организм.
ОСОБЬ
Dan.: особь статья ayrıca, xüsusi, məxsus.
ОСОБЬ
ж. биол.
особь (өзүнчө жашоочу организм).
ОСОБЬ
особь см. единица, существо
ОСОБЬ
особьСм. единица, существо…
ОСОБЬ
Достойный представитель вида
Биологический диктант по теме «Вид и популяция»
Биология. 9 класс
Биологический диктант по теме «Вид и популяция»
Вариант I
1. Физиологический критерий вида проявляется в том, что у всех его особей …
2. Относительность морфологического критерия вида состоит в том, что …
3. При определении принадлежности организма к тому или иному виду необходимо учитывать …
4. Особей в одну популяцию объединяет …
5. Диплоидный набор хромосом – это … критерий вида.
6. Соотнесите критерии вида с их характеристиками (ответ записывать в форме: 1 – г и т. д.):
|
Критерий вида |
Характеристика |
|
1. 2. Экологический 3. Географический 4. Морфологический 5. Физиологический |
а) Определенная область распространения б) Сходство процессов жизнедеятельности в) Внешнее строение г) Строение геномного материала д) Факторы внешней среды е) История развития вида ж) Определенные условия обитания |
 Генетический
Генетический
Биология. 9 класс
Биологический диктант по теме «Вид и популяция»
Вариант II
1. Приспособленность вида к жизни в разных условиях в пределах своего ареала обеспечивает его существование в форме …
2. Что является морфологическим критерием вида?
3. Для определения вида недостаточно использовать только морфологический критерий, так как …
4. Элементарной единицей существования и адаптации вида является …
5. Наследственная изменчивость,
борьба за существование, естественный отбор проявляются в популяции, поэтому ее
считают …
Наследственная изменчивость,
борьба за существование, естественный отбор проявляются в популяции, поэтому ее
считают …
6. Соотнесите демографические показатели популяции с их характеристиками (ответ записывать в форме: 1 – г и т. д.):
|
Демографический показатель |
Характеристика |
|
1. Рождаемость 2. Численность 3. Возрастной состав 4. Плотность 5. Смертность |
а) Общее количество особей б) Скорость прибавления особей в) Соотношение самцов и самок г) Скорость сокращения численности д) Количество особей на определенную территорию е) Соотношение старых и молодых особей ж) Прибавление особей других популяций |
Ответы
Вариант I
1) наблюдается сходство всех процессов жизнедеятельности;
2) самцы и самки одного вида различаются внешне, есть виды-двойники;
3) весь комплекс критериев вида;
4) свободное скрещивание, относительная изоляция от других популяций этого же вида;
5) генетический;
6) 1 – г, 2 – ж, 3 – а, 4 –
в, 5 – б.
Вариант II
1) популяций;
2) особенности внешнего и внутреннего строения;
3) существуют виды-двойники;
4) популяция;
5) единицей эволюции;
6) 1 – б, 2 – а, 3 – е, 4 – д, 5 – г.
Вид. Признаки вида
Цель урока: формирование у учащихся общих представлений о понятии «биологический вид».
Задачи урока: изучить основные признаки (критерии) вида и сформировать у учащихся представление о понятии «Вид»; продолжить развитие у учащихся умений обобщать и систематизировать знания; продолжить развитие навыков самостоятельной работы и самоанализа у учащихся.
ХОД УРОКА
1. Активизация познавательной деятельности учащихся
Учитель: В мире существует огромное множество различных видов животных, растений, грибов и бактерий. Какие виды животных и растений известны вам?
Дети называют известные им виды.
Учитель: Вы назвали много различных видов. А теперь давайте поиграем с вами в мини-игру. (Приложение 1. Слайд 1.)
Учитель: Как вы думаете, эти насекомые относятся к одному виду? Что это за насекомые? (Слайды 2 и 3).
Дети высказывают свои предположения.
Учитель: Это два разных вида. Оса и муха-журчалка, «маскирующаяся» под осу. Они похожи, не так ли?
Учитель: А эти две змеи относятся к одному виду? (Слайды 4-5).
Дети отвечают.
Учитель: Эти ядовитый коралловый аспид и безвредная молочная змея. Чем они похожи, а чем отличаются?
Дети высказываются.
Учитель: А эти две птицы относятся к одному виду? Они выглядят такими разными. (Слайды 6 и 7).
Учитель: В мире существует много
различных видов похожих между собой. Например
ястреб тетеревятник и ястреб перепелятник (Слайд
8). Или же наоборот, самцы и самки одного вида
могут сильно отличаться внешне (Слайд 9).
Например
ястреб тетеревятник и ястреб перепелятник (Слайд
8). Или же наоборот, самцы и самки одного вида
могут сильно отличаться внешне (Слайд 9).
Так как же учены определяют, какие животные
относятся к одному виду, а какие к разным? (Слайд
10).
2. Новая тема: Вид. Признаки вида
Учитель: Так что же такое «Вид»?
Давайте попытаемся разобраться. (Слайд 11).
Существуют несколько признаков – критериев, по
которым следует сравнивать особей прежде, чем
относить их к одному биологическому виду.
Первый критерий – Морфологический. (Слайд
12).
Морфологический критерий – это внешний облик
животного или растения. Особи, относящиеся к
одному виду, обычно похожи между собой. Например,
тигры или волки, или одуванчики внешне схожи с
особями своего вида.
Второй критерий (признак) –
Физиолого-биохимический. (Слайд 13).
Он говорит о физиологических процессах в
организмах. У особей одного вида они обычно
протекают с одинаковой скоростью. Хотя бывают и
исключения. Например, головастик и взрослая
лягушка относятся к одному виду, но
физиологически процессы в их организмах
протекают с различной скоростью.
У особей одного вида они обычно
протекают с одинаковой скоростью. Хотя бывают и
исключения. Например, головастик и взрослая
лягушка относятся к одному виду, но
физиологически процессы в их организмах
протекают с различной скоростью.
Биохимический признак рассказывает о химическом
составе клеток животных и растений. У особей
одного вида, химический состав сходен.
Третий признак – Географический. (Слайд 14).
Ареал – это определенная территория, на которой
обитает тот или иной вид. Взглянув на
географическую карту, мы всегда можем указать
области, в которых встречается тот или иной вид
животных или растений. На слайде у нас
представлен ареал обитания бурого медведя. Где
же обитает этот крупный хищник?
Дети отвечают на вопрос.
Учитель: Четвертый признак –
Экологический.(Слайд 15)
Этот критерий рассматривает условия в которых
обитает особь, а так же его образ жизни. Некоторые
виды предпочитают влажные джунгли, другие
приспособились к обитанию в суровых условиях
пустынь. Одни животные активны днем, другие
ночью. Назовите несколько видов растений и
животных, обитающих в пустыне?
Некоторые
виды предпочитают влажные джунгли, другие
приспособились к обитанию в суровых условиях
пустынь. Одни животные активны днем, другие
ночью. Назовите несколько видов растений и
животных, обитающих в пустыне?
Каких дневных и ночных хищников вы знаете?
Дети дают ответы.
Учитель: Пятый признак –
генетический (Слайд 16).
Исследователи сравнивают количество хромосом в
клетках растений и животных, а так же общее
строение молекул ДНК. У Особей одного вида
наследственный материал сходный.
Например, у человека – 46 хромосом, у волка – 78, у
лисы– 36, у слона – 56, у дельфина – 44, а у комара –
всего 6 хромосом.
Вывод: лишь сравнив особей по всем
признакам, мы можем с уверенностью говорить, что
они относятся к одному виду.
Почему необходимо сравнивать по всем признакам?
(Слайд 17)
Дети отвечают, приводя примеры « исключений»
по каждому из пяти признаков.
Учитель: Так что же такое Вид? Давайте попробуем дать определение этому понятию.
Дети высказывают свои варианты определения
«Вида», учитель корректирует и направляет их
ответы. Наиболее полное определение дети
записывают в тетрадь.
(Слайд 18).
3. Закрепление: дети самостоятельно заполняют представленную таблицу. По завершении работы, они обмениваются тетрадями и с помощью учителя проверяют работы. (Слайд 18) Приложение 2.
Таблица «Признаки Вида».
| Признак Вида | Значение признака |
| Морфологический | |
| Физиолого-Биохимический | |
| Географический | |
| Экологический | |
| Генетический |
4. Рефлексия
Рефлексия
Учитель:
– Так по каким же признакам должны быть сходны
организмы, что бы их отнесли к одному виду?
– Почему необходимо сочетание большинства
признаков? Какие исключения вам известны?
– Что такое Вид?
– Что нового вы узнали на сегодняшнем уроке? Что
вас удивило?
5. Домашнее задание: §30.
Биологический вид — единственно возможная форма существования высших организмов: эволюционное значение полового размножения
Представлен целостный целостный взгляд на половые виды как на высшую форму биологического существования. Идея Вейсмана о том, что пол и рекомбинация обеспечивают вариацию для естественного отбора, преобладает в большинстве дискуссий о биологическом значении полового размножения. Здесь обосновывается идея о том, что главное преимущество пола противоположное: способность противодействовать не только вымиранию, но и дальнейшей эволюции. Живые системы живут долго благодаря своей способности воспроизводить себя с высокой точностью. Простые организмы (например, бактерии) достигают непрерывного существования благодаря высокой точности репликации индивидуального генома. У организмов с большим геномом и сложным развитием достижимой точности репликации ДНК недостаточно для точного воспроизведения генома. Такие виды должны быть способны к выживанию и должны оставаться неизменными, несмотря на постоянные изменения их генов. Эта проблема не имеет решения в рамках асексуальных («гомеогеномных») линий передачи.В ходе безрассудной эволюции они быстро деградируют и вымирают или размываются. Основным результатом перехода к половому размножению стало создание многоорганической сущности — биологического вида. Отдельные организмы утратили способность к автономному воспроизводству. Это означает, что отдельные организмы утратили способность к существенной эволюции. Они развиваются как часть биологического вида. В случае обязательной сексуальности не бывает синхронного многоуровневого отбора.
Живые системы живут долго благодаря своей способности воспроизводить себя с высокой точностью. Простые организмы (например, бактерии) достигают непрерывного существования благодаря высокой точности репликации индивидуального генома. У организмов с большим геномом и сложным развитием достижимой точности репликации ДНК недостаточно для точного воспроизведения генома. Такие виды должны быть способны к выживанию и должны оставаться неизменными, несмотря на постоянные изменения их генов. Эта проблема не имеет решения в рамках асексуальных («гомеогеномных») линий передачи.В ходе безрассудной эволюции они быстро деградируют и вымирают или размываются. Основным результатом перехода к половому размножению стало создание многоорганической сущности — биологического вида. Отдельные организмы утратили способность к автономному воспроизводству. Это означает, что отдельные организмы утратили способность к существенной эволюции. Они развиваются как часть биологического вида. В случае обязательной сексуальности не бывает синхронного многоуровневого отбора. Естественный отбор не может выбрать ничего, кроме единицы воспроизводства. Иерархия в биологии подразумевает функциональное предназначение частей ради целого. Важнейшей особенностью полового размножения является формирование геномов отдельных организмов путем случайного отбора их из постоянно перетасовываемого генофонда вместо прямой репликации генома предка. Явное антиэволюционное последствие сексуальности очевидно из того факта, что генотипы особей с повышенной конкурентоспособностью не передаются следующему поколению.Вместо этого после спаривания с «обычными» особями эти генотипы разбрасываются и перестраиваются в новые комбинации генов, таким образом не позволяя победителю воспользоваться успехом.
Естественный отбор не может выбрать ничего, кроме единицы воспроизводства. Иерархия в биологии подразумевает функциональное предназначение частей ради целого. Важнейшей особенностью полового размножения является формирование геномов отдельных организмов путем случайного отбора их из постоянно перетасовываемого генофонда вместо прямой репликации генома предка. Явное антиэволюционное последствие сексуальности очевидно из того факта, что генотипы особей с повышенной конкурентоспособностью не передаются следующему поколению.Вместо этого после спаривания с «обычными» особями эти генотипы разбрасываются и перестраиваются в новые комбинации генов, таким образом не позволяя победителю воспользоваться успехом.
18.2A: Концепция биологических видов
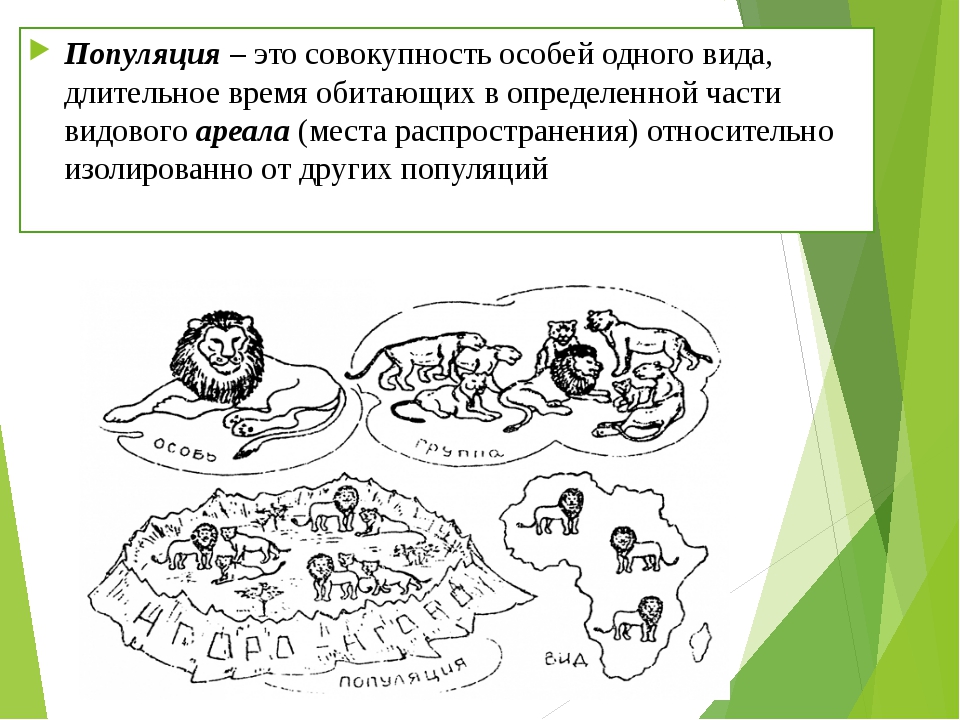
Вид определяется как группа особей, которые в природе способны спариваться и производить жизнеспособное плодородное потомство.
Виды и способность к воспроизводству
Вид — это группа отдельных организмов, которые скрещиваются и производят плодовитое жизнеспособное потомство.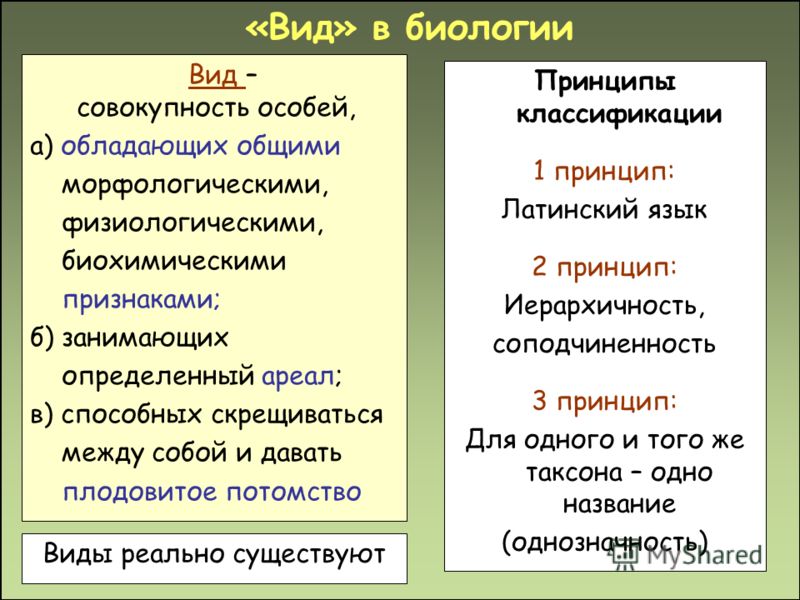 Согласно этому определению, один вид отличается от другого, когда в природе невозможно, чтобы при спаривании между особями каждого вида производилось плодовитое потомство.
Согласно этому определению, один вид отличается от другого, когда в природе невозможно, чтобы при спаривании между особями каждого вида производилось плодовитое потомство.
Члены одного и того же вида обладают как внешними, так и внутренними характеристиками, которые развиваются из их ДНК. Чем ближе отношения между двумя организмами, тем больше у них общего ДНК, как у людей и их семей. ДНК людей, скорее всего, будет больше похожа на ДНК их отца или матери, чем на ДНК их кузена или бабушки или дедушки.Организмы одного вида имеют самый высокий уровень выравнивания ДНК и, следовательно, обладают общими характеристиками и поведением, которые приводят к успешному воспроизводству.
Внешний вид вида может вводить в заблуждение, предполагая способность или неспособность к спариванию. Например, несмотря на то, что домашние собаки ( Canis lupus knownis ) демонстрируют фенотипические различия, такие как размер, телосложение и шерсть, большинство собак могут скрещиваться и производить жизнеспособных щенков, которые могут созревать и воспроизводиться половым путем.
В других случаях особи могут казаться похожими, хотя они не принадлежат к одному виду. Например, хотя белоголовые орланы ( Haliaeetus leucocephalus ) и африканские орлы-рыбы ( Haliaeetus voiceifer ) являются одновременно птицами и орлами, каждый принадлежит к отдельной группе видов. Если бы люди искусственно вмешались и оплодотворили яйцеклетку белоголового орлана спермой африканского рыбного орла, и птенец вылупился, это потомство, названное гибридом (помесь двух видов), вероятно, было бы бесплодным: неспособным успешно выжить. воспроизводятся по достижении зрелости.У разных видов могут быть разные гены, активные в развитии; следовательно, может оказаться невозможным вырастить жизнеспособное потомство с двумя разными наборами направлений. Таким образом, даже несмотря на то, что гибридизация может иметь место, эти два вида по-прежнему остаются отдельными.
Таким образом, даже несмотря на то, что гибридизация может иметь место, эти два вида по-прежнему остаются отдельными.
Популяции видов имеют общий генофонд: совокупность всех вариантов генов вида. Опять же, основа любых изменений в группе или популяции организмов должна быть генетической, поскольку это единственный способ поделиться и передать признаки. Когда изменения происходят внутри вида, они могут передаваться следующему поколению только двумя основными путями: бесполое размножение или половое размножение. Изменение будет передано бесполым путем просто, если воспроизводящая клетка обладает измененным признаком.Чтобы измененный признак передавался половым путем, гамета, такая как сперматозоид или яйцеклетка, должна обладать измененным признаком.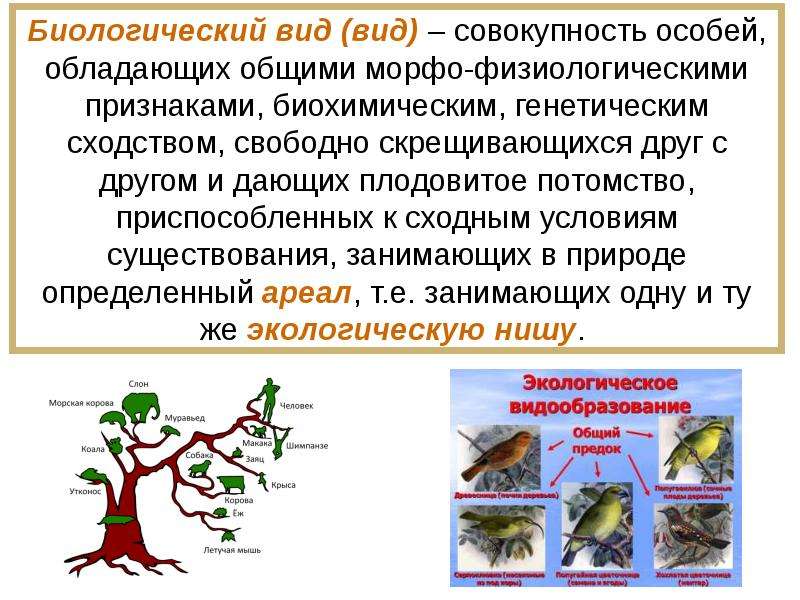 Другими словами, организмы, размножающиеся половым путем, могут испытывать несколько генетических изменений в клетках своего тела, но если эти изменения не происходят в сперматозоиде или яйцеклетке, измененный признак никогда не дойдет до следующего поколения. Только наследственные черты могут развиваться. Следовательно, воспроизводство играет первостепенную роль в том, чтобы генетические изменения укоренились в популяции или виде. Короче говоря, организмы должны иметь возможность размножаться друг с другом, чтобы передавать новые черты потомству.
Другими словами, организмы, размножающиеся половым путем, могут испытывать несколько генетических изменений в клетках своего тела, но если эти изменения не происходят в сперматозоиде или яйцеклетке, измененный признак никогда не дойдет до следующего поколения. Только наследственные черты могут развиваться. Следовательно, воспроизводство играет первостепенную роль в том, чтобы генетические изменения укоренились в популяции или виде. Короче говоря, организмы должны иметь возможность размножаться друг с другом, чтобы передавать новые черты потомству.
естественных видов, особей или что? на JSTOR
Абстрактный Какие биологические виды? Аристотелиане и локкисты согласны с тем, что они являются естественными людьми; но теория эволюции показывает, что ни один из традиционных философских подходов не является по-настоящему адекватным. Недавно Майкл Гизелин и Дэвид Халл заявили, что виды — это индивидуумы. Доказано, что это утверждение противоречит духу большей части современной биологии. Делается вывод, что виды — это своего рода естественные виды, и что любая «объективность», которой они обладают, проистекает из того, что они находятся в фокусе согласованности индукций.
Делается вывод, что виды — это своего рода естественные виды, и что любая «объективность», которой они обладают, проистекает из того, что они находятся в фокусе согласованности индукций.
Текущие выпуски теперь размещены на веб-сайте Chicago Journals. Прочтите последний выпуск. Начиная с 1950 года, The British Journal for the Philosophy of Science (BJPS) представляет лучшие новые работы в этой дисциплине. Международный лидер в области философии науки, BJPS демонстрирует выдающиеся исследования по множеству тем, от природы моделей и симуляций до математического объяснения и фундаментальных проблем в физических, биологических и социальных науках.Журнал, публикуемый от имени Британского общества философии науки, предлагает новаторские и заставляющие задуматься статьи, которые открывают новые области исследований или проливают новый свет на известные проблемы.
Информация об издателе С момента своего основания в 1890 году в качестве одного из трех основных подразделений Чикагского университета, University of Chicago Press взял на себя обязательство распространять стипендии высочайшего стандарта и публиковать серьезные работы, которые способствуют образованию, способствуют общественному пониманию , и обогатить культурную жизнь. Сегодня Отдел журналов издает более 70 журналов и сериалов в твердом переплете по широкому кругу академических дисциплин, включая социальные науки, гуманитарные науки, образование, биологические и медицинские науки, а также физические науки.
Сегодня Отдел журналов издает более 70 журналов и сериалов в твердом переплете по широкому кругу академических дисциплин, включая социальные науки, гуманитарные науки, образование, биологические и медицинские науки, а также физические науки.
ВИДЫ ПОНЯТИЯ
Концепция биологических видов: группы фактически или потенциально скрещивающихся естественных популяций, которые репродуктивно изолированы от других таких групп. Эрнст Майр 1940
Концепция эволюционных видов: родословная (последовательность потомков-предков популяций), развивающаяся отдельно от других и обладающая собственными унитарными эволюционными ролями и тенденциями.
Концепция кладистических видов: эволюционная линия между двумя точками ветвления или между одной точкой ветвления и событием вымирания или современной популяцией.
Концепция видов сплоченности: наиболее обширная группа организмов, обладающих потенциалом генетического и / или демографического обмена. Группа организмов, обладающих потенциалом фенотипической сплоченности за счет внутренних механизмов.
Группа организмов, обладающих потенциалом фенотипической сплоченности за счет внутренних механизмов.
Понятие видов конкуренции: наиболее обширные единицы в натуральном хозяйстве, так что между их частями происходит репродуктивная конкуренция.
Концепция экологических видов: набор организмов, эксплуатируемых или адаптированных к одной нише; линия или тесно связанный набор линий передачи, который занимает адаптивную зону, минимально отличающуюся от зоны любой другой линии в ее диапазоне.
Изоляция Концепция видов: системы популяций, обмен генами между которыми ограничен или предотвращен репродуктивным изолирующим механизмом или, возможно, комбинацией нескольких таких механизмов.
Фенетическая концепция видов (концепция морфологических видов): набор организмов, которые похожи друг на друга и отличаются от других наборов.
Концепция филогенетических видов: наименьшая монофилетическая группа, различаемая по общим производным (синапоморфным) характеристикам.
Концепция распознавания видов: наиболее инклюзивная популяция отдельных двупародительских организмов, которые имеют общую систему оплодотворения.
Концепция типологических видов: группа организмов, соответствующих общему морфологическому плану, с акцентом на вид как на по существу статичную, неизменяемую совокупность.
Концепция генеологических видов: исключительная группа организмов, члены которой больше похожи друг на друга, чем члены какой-либо внешней группы.
Концепция биологических видов: критическая оценка
Abstract
Термин «вид» был центральным принципом биологической веры с первых дней биологии. Но понятия, связанные с этим термином, менялись и часто не получали четких определений. Цель данной статьи — исследовать концепцию биологических видов (BSC): рассмотреть ее теоретические аспекты, как на самом деле разграничить биологические виды в природе, существуют ли такие виды в природе и имеет ли эта концепция какую-либо уникальную ценность для изучение эволюции. Классическое определение BSC разделено на его основные компоненты, и обсуждаются некоторые из их аспектов и проблем. Подробно описаны три основные операции, необходимые для определения границ биологических видов в природе. Это операции, основанные на критериях: (1) географической близости, (2) фенетического сходства и (3) скрещивания. Определяются и обсуждаются два свойства скрещивания: связность и успешность воспроизводства. Блок-схема распознавания биологических видов построена на основе определения, данного Майром.Каждый шаг включает одну из трех операций, упомянутых выше. Приводятся причины включения каждого шага, а также присущие каждому из них трудности. Можно видеть, что большинство шагов в значительной степени или полностью фенетично, даже теоретически. Необходимые фенетические шаги называются «фенетическими узкими местами». Для проверки блок-схемы мы предполагаем нереалистичную, но оптимальную ситуацию полного знания о межпородных отношениях между отобранными организмами. Фенетические узкие места остаются в этом оптимальном случае, и степень зависимости от фенетической информации для определения границ биологических видов возрастает по мере того, как мы отклоняемся от оптимальной ситуации и делаем ее более реалистичной.
Классическое определение BSC разделено на его основные компоненты, и обсуждаются некоторые из их аспектов и проблем. Подробно описаны три основные операции, необходимые для определения границ биологических видов в природе. Это операции, основанные на критериях: (1) географической близости, (2) фенетического сходства и (3) скрещивания. Определяются и обсуждаются два свойства скрещивания: связность и успешность воспроизводства. Блок-схема распознавания биологических видов построена на основе определения, данного Майром.Каждый шаг включает одну из трех операций, упомянутых выше. Приводятся причины включения каждого шага, а также присущие каждому из них трудности. Можно видеть, что большинство шагов в значительной степени или полностью фенетично, даже теоретически. Необходимые фенетические шаги называются «фенетическими узкими местами». Для проверки блок-схемы мы предполагаем нереалистичную, но оптимальную ситуацию полного знания о межпородных отношениях между отобранными организмами. Фенетические узкие места остаются в этом оптимальном случае, и степень зависимости от фенетической информации для определения границ биологических видов возрастает по мере того, как мы отклоняемся от оптимальной ситуации и делаем ее более реалистичной. BSC оказывается произвольным (sensu Simpson), когда предпринимаются попытки применить его к реальным данным в природе, и не только потому, что произвольные фенетические решения являются необходимой частью определения границ биологических видов в природе. Задавая некоторые важные вопросы о значении BSC для таксономии и эволюции, мы обнаруживаем, что BSC не является необходимым для практической таксономии, не является ни необходимым, ни особенно полезным для эволюционной таксономии, а также не является уникальной или эвристической концепцией, необходимой для генерации гипотез. в эволюционной теории.Большинство важных эволюционных принципов, обычно связанных с BSC, могут быть легко применены к локализованным биологическим популяциям, что часто приводит к более глубокому пониманию эволюции. Решив, что BSC не является ни функциональным, ни эвристическим, ни какой-либо практической ценностью, мы заключаем, что фенетический вид, как обычно описывается, является желательной концепцией вида, которая должна быть связана с таксономической категорией «виды», и что локализованная биологическая популяция может быть самый полезный блок для изучения эволюции.
BSC оказывается произвольным (sensu Simpson), когда предпринимаются попытки применить его к реальным данным в природе, и не только потому, что произвольные фенетические решения являются необходимой частью определения границ биологических видов в природе. Задавая некоторые важные вопросы о значении BSC для таксономии и эволюции, мы обнаруживаем, что BSC не является необходимым для практической таксономии, не является ни необходимым, ни особенно полезным для эволюционной таксономии, а также не является уникальной или эвристической концепцией, необходимой для генерации гипотез. в эволюционной теории.Большинство важных эволюционных принципов, обычно связанных с BSC, могут быть легко применены к локализованным биологическим популяциям, что часто приводит к более глубокому пониманию эволюции. Решив, что BSC не является ни функциональным, ни эвристическим, ни какой-либо практической ценностью, мы заключаем, что фенетический вид, как обычно описывается, является желательной концепцией вида, которая должна быть связана с таксономической категорией «виды», и что локализованная биологическая популяция может быть самый полезный блок для изучения эволюции.
ВАРИАНТЫ ДОСТУПА
Войдите, чтобы получить доступ к подписке
Войдите в систему, если у вас есть индивидуальная подписка на этот журнал или вы уже приобрели этот журнал. статья или обзор.
Подпишитесь на этот журнал прямо сейчас, используя раскрывающееся меню «Подписаться» или нажав здесь.
Войдите, чтобы получить доступ к своей подписке
Чтобы получить доступ к этому журналу через свое членство в спонсирующей организации, нажмите «Обзор проблем» в раскрывающемся меню выберите «Доступ для участников».’
Войдите в систему через свое учреждение
Посетите веб-сайт вашей институциональной библиотеки, чтобы войти в систему
или обратитесь к своему библиотекарю, чтобы получить доступ к этому журналу.
Некоторые учреждения могут предоставить
Доступ к системе единого входа здесь.
Использование сетей спаривания для определения границ видов
Abstract
Хотя интерфертильность является ключевым критерием, на котором основана концепция биологических видов Майра, она никогда не применялась напрямую для определения границ видов в естественных условиях.Наше исследование восполняет этот пробел. Мы использовали критерий интерфертильности, чтобы разграничить два близкородственных вида дуба в лесном насаждении, проанализировав сеть естественных брачных событий между особями. Результаты выявили две группы интерфертильных особей, связанных всего несколькими событиями спаривания. Эти две группы в значительной степени соответствовали группам, определенным с использованием других критериев (морфологическое сходство, генотипическое сходство и индивидуальное родство). Таким образом, наше исследование показывает, что анализ сетей спаривания является эффективным методом определения границ видов на основе критерия интерфертильности при условии, что могут быть собраны адекватные сетевые данные. Наше исследование также показывает, что, хотя границы видов в значительной степени совпадают между методами определения границ видов, они не совсем совпадают. Большинство различий связано с отнесением людей к промежуточной категории. Расхождения между методами могут отражать биологическую реальность. Действительно, критерий интерфертильности является критерием, зависящим от окружающей среды, поскольку численность видов обычно влияет на скорость гибридизации в естественных условиях. Таким образом, ожидается, что методы определения границ видов на основе критерия интерфертильности дадут результаты, несколько отличающиеся от результатов, основанных на критериях, не зависящих от среды (таких как критерии генотипического сходства).Однако, какой бы критерий ни был выбран, задача, с которой мы сталкиваемся при разграничении видов, состоит в том, чтобы обобщить непрерывные, но неоднородные изменения в биологическом разнообразии. Модель степени членства, которую мы используем в этом исследовании, является подходящим инструментом.
Наше исследование также показывает, что, хотя границы видов в значительной степени совпадают между методами определения границ видов, они не совсем совпадают. Большинство различий связано с отнесением людей к промежуточной категории. Расхождения между методами могут отражать биологическую реальность. Действительно, критерий интерфертильности является критерием, зависящим от окружающей среды, поскольку численность видов обычно влияет на скорость гибридизации в естественных условиях. Таким образом, ожидается, что методы определения границ видов на основе критерия интерфертильности дадут результаты, несколько отличающиеся от результатов, основанных на критериях, не зависящих от среды (таких как критерии генотипического сходства).Однако, какой бы критерий ни был выбран, задача, с которой мы сталкиваемся при разграничении видов, состоит в том, чтобы обобщить непрерывные, но неоднородные изменения в биологическом разнообразии. Модель степени членства, которую мы используем в этом исследовании, является подходящим инструментом.
Образец цитирования: Lagache L, Leger J-B, Daudin J-J, Petit RJ, Vacher C. (2013) Проверка концепции биологических видов: использование сетей спаривания для определения границ видов. PLoS ONE 8 (6): e68267. https: // doi.org / 10.1371 / journal.pone.0068267
Редактор: Энрико Скалас, Университет дель Пьемонте, Италия
Поступила: 17 января 2013 г .; Принята к печати: 28 мая 2013 г .; Опубликован: 20 июня 2013 г.
Авторские права: © 2013 Lagache et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Финансирование было предоставлено проектом LinkTree (ANR BIODIVERSA), Европейской сетью передового опыта EvolTree, ANR-10-EQPX-16 XYLOFOREST и INRA (грант CJS). Генотипирование финансировалось за счет грантов от Conseil Régional d’Aquitaine n ° 20030304002FA, n ° 20040305003FA и Европейского Союза, FEDER n ° 2003227. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Генотипирование финансировалось за счет грантов от Conseil Régional d’Aquitaine n ° 20030304002FA, n ° 20040305003FA и Европейского Союза, FEDER n ° 2003227. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Согласно концепции биологического вида, способность к скрещиванию (т.е. интерфертильность) является определяющим свойством вида [1]. Тем не менее, насколько нам известно, критерий интерфертильности никогда не использовался для определения границ видов на основе событий спаривания, наблюдаемых в естественных условиях. Для этой цели использовались только искусственные скрещивания, в том числе у грибов (например, [2]), растений [3] или насекомых [4]. Однако этот подход подвергался критике (например, [5,6]), потому что искусственные скрещивания обходят некоторые пре-спаривающие барьеры для гибридизации: события спаривания, наблюдаемые в искусственных условиях, могут не отражать то, что могло бы произойти естественным образом. Следовательно, на сегодняшний день нет удовлетворительного примера использования критерия интерфертильности для определения границ видов. Фактически, методы, наиболее часто используемые для определения границ видов, не являются производными от хорошо известной концепции биологических видов, а являются производными от других концепций, таких как концепция филогенетических видов, концепция генотипических видов и концепция морфологических видов. Определения видов в соответствии с этими концепциями и возможные связанные критерии для определения границ видов перечислены в таблице 1.
Следовательно, на сегодняшний день нет удовлетворительного примера использования критерия интерфертильности для определения границ видов. Фактически, методы, наиболее часто используемые для определения границ видов, не являются производными от хорошо известной концепции биологических видов, а являются производными от других концепций, таких как концепция филогенетических видов, концепция генотипических видов и концепция морфологических видов. Определения видов в соответствии с этими концепциями и возможные связанные критерии для определения границ видов перечислены в таблице 1.
| Концепция видов | Определение видов в соответствии с этой концепцией | Возможный критерий разграничения видов, полученный из этого определения | Возможный метод разграничения видов с использованием этого критерия | Первое применение участок исследования |
|---|---|---|---|---|
| Концепция биологического вида | Виды — это « групп фактически или потенциально скрещивающихся природных популяций, которые репродуктивно изолированы от других таких групп » [1]. Согласно Хаусдорфу [17], «естественные популяции» могут быть заменены в этом утверждении на «индивидуумы» без изменения смысла. Согласно Хаусдорфу [17], «естественные популяции» могут быть заменены в этом утверждении на «индивидуумы» без изменения смысла. | Более высокая естественная интерферируемость между особями внутри, чем между видами | Кластеризация сети естественных брачных событий между особями с использованием непрерывной стохастической блочной модели (C-SBM) [24]. | это исследование |
| Филогенетическая концепция видов | Вид — это « диагностируемый кластер индивидуумов, внутри которого существует родительский образец происхождения и происхождения, за которым его нет, и который демонстрирует образец филогенетического происхождения и происхождения». спуск среди единиц одного вида »[29]. | Более высокое генетическое родство между индивидами внутри, чем между видами | Кластеризация сети родственных отношений между индивидами с использованием C-SBM [24]. | это исследование |
| Концепция генотипа видов | Вид — это « генотипический кластер особей, которые могут перекрываться, не сливаясь со своими братьями и сестрами » [17,52] | Более высокое генотипическое сходство между особями внутри вида | Кластеризация особей на основе их мультилокусного генотипа со СТРУКТУРОЙ [50] | Guichoux et al. 2012 [23] |
| Концепция морфологического вида | Виды — это « наименьших обнаруженных образцов самовоспроизводящихся организмов с уникальными наборами признаков » [53,54]. | Более высокое морфологическое сходство между особями внутри, чем между видами | Кластеризация особей на основе нескольких морфологических признаков с помощью факторного дискриминантного анализа [55]. | Bacilieri et al. 1996 [26] |
Таблица 1.Основные концепции видов с соответствующими возможными критериями разграничения видов.
Одним из возможных методов разграничения видов на основе критерия интерфертильности является анализ сетей спаривания. Брачные сети представляют собой брачные события между людьми [7]. Узлы сети представляют людей, а ссылки связывают людей, между которыми произошли брачные события. Применение методов сетевой кластеризации [8–10] к сетям сопряжения должно позволить идентифицировать подмножества сильно взаимосвязанных узлов, которые соответствуют видам.Если концепция биологического вида интерпретируется строго, то вид должен соответствовать связанному компоненту сети спаривания (рис. 1А). Связанный компонент — это подмножество узлов в сети, которые прямо или косвенно связаны, но не связаны с узлами, не входящими в подмножество. Согласно упрощенной концепции биологических видов, которая допускает некоторый уровень гибридизации между видами [11–13], вид должен соответствовать сообществу в сети спаривания (рис. 1B).Сообщества — это подмножества узлов с высокой плотностью связей внутри группы и более низкой плотностью связей между различными группами [8]. Именно в этом последнем случае, когда виды гибридизируются, разграничение видов на основе критерия интерфертильности становится особенно сложной задачей, и сетевой анализ может быть особенно полезным.
1B).Сообщества — это подмножества узлов с высокой плотностью связей внутри группы и более низкой плотностью связей между различными группами [8]. Именно в этом последнем случае, когда виды гибридизируются, разграничение видов на основе критерия интерфертильности становится особенно сложной задачей, и сетевой анализ может быть особенно полезным.
Рис. 1. Пример сетей спаривания с видовыми границами.
Каждый узел сети, представленный черной звездой или белым кружком, является индивидуальным.Каждое звено сети, представленное тонкой черной линией, соответствует брачному событию между двумя людьми. В A не происходит спаривания между двумя группами особей, тогда как в B между группами происходит несколько событий спаривания. Границы видов в соответствии со строгим применением концепции биологических видов обозначены сплошной толстой черной линией. Границы видов в соответствии с упрощенной интерпретацией концепции биологических видов, допускающей межвидовую гибридизацию, обозначены пунктирной красной линией. В теории сетей непрерывная черная линия ограничивает соединенные компоненты сети, а пунктирная красная линия ограничивает сообщества.
В теории сетей непрерывная черная линия ограничивает соединенные компоненты сети, а пунктирная красная линия ограничивает сообщества.
https://doi.org/10.1371/journal.pone.0068267.g001
Идея анализа сетей спаривания для определения границ видов в соответствии с концепцией биологических видов была предложена более 40 лет назад Сокалом и Кровелло [14], но она похоже, не было реализовано на практике. Построение сети спаривания — действительно сложная задача, поскольку для этого требуется очень большой набор данных о событиях спаривания, собранных в естественных условиях.Виды должны быть симпатрическими и иметь полупроницаемые репродуктивные барьеры, чтобы вопрос о разграничении видов был актуален. Более того, виды должны быть не только ауткроссинговыми (с низким уровнем самоопыления), но и высоко полигамными и иметь несколько потомков на поколение, чтобы фактические события спаривания представляли потенциальные события спаривания между особями в данный момент [15-17].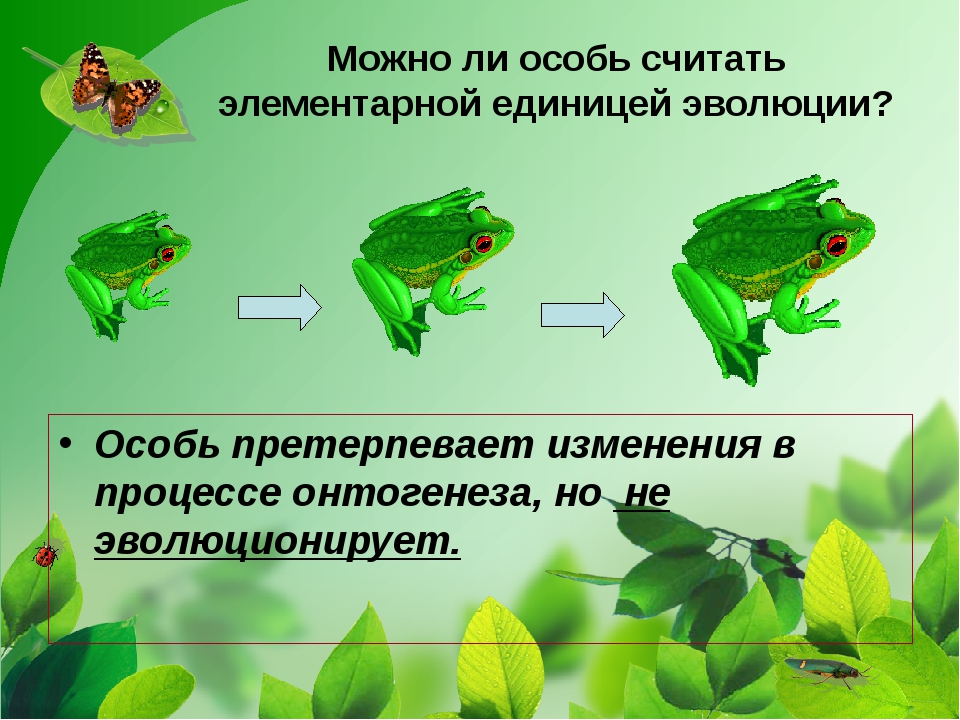 Если бы такие данные были доступны, стал бы анализ сетей спаривания эффективным методом определения границ видов на основе критерия интерфертильности? Будут ли границы между видами такими же, как и границы, полученные с использованием других критериев разграничения видов?
Если бы такие данные были доступны, стал бы анализ сетей спаривания эффективным методом определения границ видов на основе критерия интерфертильности? Будут ли границы между видами такими же, как и границы, полученные с использованием других критериев разграничения видов?
Чтобы ответить на эти вопросы, мы исследуем соответствие между четырьмя методами разграничения видов, основанными на концепциях биологических, морфологических, генотипических и филогенетических видов (таблица 1), применяя их к двум гибридизирующимся древесным видам, живущим в симпатрии.Исследуемый участок представляет собой смешанный насаждение площадью 5 га Quercus robur и Q . petraea , состоящий из 298 взрослых деревьев, возникших в результате естественного возобновления [18]. Как и многие другие близкородственные виды растений [19], эти два вида дуба гибридизуются в естественных условиях [20], в том числе в изученном насаждении [21–23]. Чтобы разграничить виды в соответствии с критерием интерфертильности, мы анализируем сеть наблюдаемых естественных спариваний между парами взрослых деревьев с помощью метода сетевой кластеризации. Каждый узел сети спаривания соответствует взрослому дереву, и каждая ссылка соответствует по крайней мере одному событию спаривания между двумя деревьями. Для кластеризации индивидуумов мы выбрали среди доступных методов сетевой кластеризации [8–10] Непрерывную стохастическую блочную модель (C-SBM), недавно представленную Даудином и др. . [24]. C-SBM синтезирует неоднородность реальной сети, создавая упрощенную версию сети, состоящую из нескольких виртуальных узлов, называемых экстремальными гипотетическими узлами (EHN).В отличие от многих методов сетевой кластеризации, которые предполагают, что каждый узел принадлежит только одной группе, C-SBM позволяет узлам демонстрировать смешанное поведение связности, предполагая, что каждый узел реальной сети представляет собой смесь EHN. Таким образом, этот метод особенно подходит для нашего исследования. Действительно, поскольку известно, что два ранее идентифицированных вида дуба [23,25] гибридизуются [22], мы ожидали найти некоторых особей со смешанным репродуктивным поведением, т.
Каждый узел сети спаривания соответствует взрослому дереву, и каждая ссылка соответствует по крайней мере одному событию спаривания между двумя деревьями. Для кластеризации индивидуумов мы выбрали среди доступных методов сетевой кластеризации [8–10] Непрерывную стохастическую блочную модель (C-SBM), недавно представленную Даудином и др. . [24]. C-SBM синтезирует неоднородность реальной сети, создавая упрощенную версию сети, состоящую из нескольких виртуальных узлов, называемых экстремальными гипотетическими узлами (EHN).В отличие от многих методов сетевой кластеризации, которые предполагают, что каждый узел принадлежит только одной группе, C-SBM позволяет узлам демонстрировать смешанное поведение связности, предполагая, что каждый узел реальной сети представляет собой смесь EHN. Таким образом, этот метод особенно подходит для нашего исследования. Действительно, поскольку известно, что два ранее идентифицированных вида дуба [23,25] гибридизуются [22], мы ожидали найти некоторых особей со смешанным репродуктивным поведением, т. е. размножаться с обоими видами. Тот же метод использовался для определения границ видов на основе генетического родства между особями.В этом случае каждый узел сети соответствует взрослому дереву, а ссылки соединяют людей, которые считаются родственными на основании их генотипа. Наконец, мы сравниваем индивидуальные назначения, полученные путем анализа сети спаривания и сети родства, с теми, которые были ранее получены в том же исследовательском центре, с использованием критериев морфологического и генотипического сходства [23,25]. Затем мы обсуждаем, как суммировать непрерывные, но неоднородные изменения в биологическом разнообразии.
е. размножаться с обоими видами. Тот же метод использовался для определения границ видов на основе генетического родства между особями.В этом случае каждый узел сети соответствует взрослому дереву, а ссылки соединяют людей, которые считаются родственными на основании их генотипа. Наконец, мы сравниваем индивидуальные назначения, полученные путем анализа сети спаривания и сети родства, с теми, которые были ранее получены в том же исследовательском центре, с использованием критериев морфологического и генотипического сходства [23,25]. Затем мы обсуждаем, как суммировать непрерывные, но неоднородные изменения в биологическом разнообразии.
Результаты
Определение границ видов на основе интерфертильности
Согласно критерию AIC, лучшей моделью для сопряженной сети была модель с четырьмя EHN, за которой следовали модели с пятью и тремя EHN (рисунок S1 в файле S1).Мы выбрали модель с тремя EHN, потому что две другие модели подчеркнули структуру плана выборки (текст S1 в файле S1). Согласно матрице связности для EHN (рис. 2A), EHN 0 соответствует виртуальному узлу, не подключенному ко всей сети. Эта EHN, которая систематически присутствует в сетевых моделях, производимых C-SBM [24], позволяет учитывать изменение количества ссылок, прикрепленных к узлам реальной сети. Две другие EHN, названные EHN B1 и EHN B2 , были прочно связаны между собой и не были связаны с другими EHN.
Согласно матрице связности для EHN (рис. 2A), EHN 0 соответствует виртуальному узлу, не подключенному ко всей сети. Эта EHN, которая систематически присутствует в сетевых моделях, производимых C-SBM [24], позволяет учитывать изменение количества ссылок, прикрепленных к узлам реальной сети. Две другие EHN, названные EHN B1 и EHN B2 , были прочно связаны между собой и не были связаны с другими EHN.
Рис. 2. Треугольное представление узлов (A) сети сопряжения и (B) сети взаимосвязей, показывающее смесь EHN (т.е. экстремальных гипотетических узлов) для каждого узла в соответствии с C-SBM.
В A узлы, которые находятся на границе между EHN 0 и EHN B1 , классифицируются в группе B1, в то время как узлы на границе между EHN 0 и EHN B2 классифицируются в группе B2. Остальные индивиды классифицируются как промежуточные (группа Bi).В B узлы, которые находятся на границе между EHN 0 и EHN P1 , классифицируются в группе P1, в то время как узлы на границе между EHN 0 и EHN P2 классифицируются в группу P2. Остальные индивиды классифицируются как промежуточные (группа Pi). Матрицы связности для EHN представлены рядом с каждым треугольным представлением. Ненулевые значения выделены жирным шрифтом.
Остальные индивиды классифицируются как промежуточные (группа Pi). Матрицы связности для EHN представлены рядом с каждым треугольным представлением. Ненулевые значения выделены жирным шрифтом.
https://doi.org/10.1371/journal.pone.0068267.g002
Затем узлы сопрягающей сети (каждый соответствует отдельному человеку) были представлены в виде треугольника с одной EHN в каждой точке (рисунок 2A).Чем выше доля данной EHN в смеси узлов, тем ближе узел к этой EHN в треугольнике. Согласно матрице связности для EHN (рис. 2A), узлы с высокой долей EHN 0 в их смеси были слабо подключены к сопряженной сети. Узлы с высокой долей EHN B1 в их смеси принадлежали к группе узлов, сильно связанных друг с другом и слабо связанных с узлами с высокой долей EHN B2 .И наоборот, узлы с высокой долей EHN B2 в их смеси принадлежали к группе узлов, сильно связанных друг с другом и слабо связанных с узлами с высокой долей EHN B1 . Таким образом, в сети спаривания было две группы взрослых деревьев, внутри которых спаривания происходили часто, а между которыми спаривания происходили редко. Графическое представление сети подтвердило этот результат (рис. 3A). Согласно упрощенной интерпретации концепции биологического вида, эти две группы людей должны соответствовать двум биологическим видам (рис. 1B).
Таким образом, в сети спаривания было две группы взрослых деревьев, внутри которых спаривания происходили часто, а между которыми спаривания происходили редко. Графическое представление сети подтвердило этот результат (рис. 3A). Согласно упрощенной интерпретации концепции биологического вида, эти две группы людей должны соответствовать двум биологическим видам (рис. 1B).
Рисунок 3. Графическое представление (A) сети сопряжения и (B) сети взаимосвязи с использованием программного обеспечения PAJECK со следующими параметрами: Draw / Layout / Energy / Kamada-Kawaï / Separate Components.
Лица, отнесенные к группе B1 (в A) или группе P1 (в B), показаны зеленым, лица, принадлежащие к группе B2 (в A) или группе P2 (в B), показаны желтым, а промежуточные особи показаны черным.
https: // doi.org / 10.1371 / journal.pone.0068267.g003
Чтобы отнести особей к двум видам, мы классифицировали узлы брачной сети в соответствии с их относительными пропорциями: EHN B1 и EHN B2 . Мы предположили, что особь принадлежала к виду B1, если соответствующий узел был смесью EHN 0 и EHN B1 и только этих двух узлов. Напротив, мы предположили, что особь принадлежала к виду B2, если соответствующий узел был смесью EHN 0 и EHN B2 .Остальные были отнесены к промежуточным в репродуктивном отношении (группа Bi). В треугольном представлении (рис. 2A) особи, отнесенные к виду B1, находились на границе между EHN 0 и EHN B1 (n = 78 особей), в то время как особи, отнесенные к виду B2, находились на границе между EHN 0 и EHN B2 (n = 121 человек). Промежуточные особи находились внутри треугольника (n = 7 человек). Эти три группы показаны разными цветами в представлении сети (рис. 3A).
Мы предположили, что особь принадлежала к виду B1, если соответствующий узел был смесью EHN 0 и EHN B1 и только этих двух узлов. Напротив, мы предположили, что особь принадлежала к виду B2, если соответствующий узел был смесью EHN 0 и EHN B2 .Остальные были отнесены к промежуточным в репродуктивном отношении (группа Bi). В треугольном представлении (рис. 2A) особи, отнесенные к виду B1, находились на границе между EHN 0 и EHN B1 (n = 78 особей), в то время как особи, отнесенные к виду B2, находились на границе между EHN 0 и EHN B2 (n = 121 человек). Промежуточные особи находились внутри треугольника (n = 7 человек). Эти три группы показаны разными цветами в представлении сети (рис. 3A).
Определение границ видов на основе родства
Согласно критерию AIC, оптимальное количество EHN в сети взаимосвязи составляло шесть. Модели с тремя, четырьмя, пятью и семью EHN также были хорошими моделями (рисунок S2 в файле S1). Поскольку мы не нашли удовлетворительного способа определить лучшую модель (текст S2 в файле S1), мы выбрали модель с тремя EHN, чтобы облегчить сравнение между структурой сети взаимосвязи и структурой сети сопряжения. Согласно матрице связности для EHN (рисунок 2B), EHN 0 соответствует виртуальному узлу, не подключенному ко всей сети.Две другие EHN, названные EHN P1 и EHN P2 , были прочно связаны между собой и не были подключены к другим EHN. Таким образом, как и в брачной сети, особи были разделены на три группы, названные P1, P2 и Pi. Группа P1 (n = 70 человек, расположенных на границе между EHN 0 и EHN P1 в треугольном представлении; рис. 2B) и группа P2 (n = 108 человек, расположенных на границе между EHN 0 и EHN P2. ; Рисунок 2B) включали людей с высокой степенью родства внутри группы и низкой степенью межгруппового родства.Третья группа Pi (n = 28 особей, расположенных внутри треугольника; рис.
Поскольку мы не нашли удовлетворительного способа определить лучшую модель (текст S2 в файле S1), мы выбрали модель с тремя EHN, чтобы облегчить сравнение между структурой сети взаимосвязи и структурой сети сопряжения. Согласно матрице связности для EHN (рисунок 2B), EHN 0 соответствует виртуальному узлу, не подключенному ко всей сети.Две другие EHN, названные EHN P1 и EHN P2 , были прочно связаны между собой и не были подключены к другим EHN. Таким образом, как и в брачной сети, особи были разделены на три группы, названные P1, P2 и Pi. Группа P1 (n = 70 человек, расположенных на границе между EHN 0 и EHN P1 в треугольном представлении; рис. 2B) и группа P2 (n = 108 человек, расположенных на границе между EHN 0 и EHN P2. ; Рисунок 2B) включали людей с высокой степенью родства внутри группы и низкой степенью межгруппового родства.Третья группа Pi (n = 28 особей, расположенных внутри треугольника; рис. 2В) включала деревья, относящиеся как к особям P1, так и к P2, и деревья с несколькими родственниками. Эти три группы показаны разными цветами в представлении сети (рис. 3B).
2В) включала деревья, относящиеся как к особям P1, так и к P2, и деревья с несколькими родственниками. Эти три группы показаны разными цветами в представлении сети (рис. 3B).
Определение границ видов на основе морфологии и мультилокусных генотипов
Критерий морфологического сходства ранее использовался Bacilieri et al. [26], чтобы идентифицировать все деревья с места исследования. Основываясь на их результатах, мы разделили людей на две чистые морфологические группы (в этом исследовании они называются M1 и M2 и соответствуют Q .robur и Q . petraea соответственно) и морфологически промежуточному классу (называемому Mi). Guichoux et al. [23] использовал генотипическое сходство в качестве критерия для отнесения деревьев исследуемого участка к видам. Основываясь на их результатах, мы классифицировали взрослые деревья на две чистокровные группы (далее G1 и G2) и один генетически промежуточный класс (Gi).
Соответствие четырех методов определения границ видов
Чтобы оценить соответствие между четырьмя методами определения границ видов, мы сравнили пространственное распределение трех групп особей, идентифицированных с помощью каждого метода.Границы видов очень похожи (рис. 4). Среди 206 взрослых деревьев, включенных в сеть спаривания и в сеть родства, было 97 деревьев, последовательно классифицированных в группы B1, P1, G1 и M1, и 63 дерева, последовательно классифицированных в группы B2, P2, G2 и M2. Поэтому мы переименовали группы B1, P1, G1 и M1 Q . robur и группы B2, P2, G2 и M2 Q . petraea . Основываясь на этой классификации, было всего четыре инверсии видов, связанных с методами разграничения (таблица S1 в файле S1).Среди 206 взрослых деревьев 42 были классифицированы как промежуточные по крайней мере по одному методу. Удивительно, но ни один человек не был классифицирован как промежуточный по всем четырем методам. Таким образом, 91% расхождений между четырьмя методами был вызван назначением промежуточному классу (рисунок S3 и таблица S1 в файле S1).
Рисунок 4. Границы видов на основе критериев интерфертильности ( A ) , родства ( B ) , генотипического сходства ( C ) и морфологического сходства ( D ) критериев, представленных на карте стенда.
В A, B и C особи, отнесенные к видам B1, P1 или G1, соответственно, представлены желтыми треугольниками. Особи, отнесенные к видам B2, P2 или G2, представлены зелеными ромбами. Промежуточные особи представлены черными крестами. В D особи, отнесенные к M1, показаны красным, индивидуумы, отнесенные к M2, синим, а морфологически промежуточные особи обозначены черными крестами. Лица группы M1 отнесены к Q.robur и отдельные лица группы M2 до Q. petraea на основе текущей таксономической практики.
https://doi.org/10.1371/journal.pone.0068267.g004
Было обнаружено девять расхождений между отдельными отнесениями по критериям генотипического и морфологического сходства, с одной стороны, и критерию интерфертильности, с другой. Мы исследовали, может ли биотическая среда людей объяснить эти несоответствия.Наша гипотеза состоит в том, что соседство каждого дерева влияет на его систему спаривания и, таким образом, может влиять на его отнесение к видам на основе критерия интерфертильности, тогда как оно вряд ли повлияет на его отнесение к видам на основе генотипических и морфологических критериев. Поэтому мы исследовали окрестности каждого дерева, для которых отнесение к видам на основе критериев генотипического и морфологического сходства было конгруэнтным (N = 192). Для каждого дерева мы рассчитали долю аллоспецифических соседей в радиусе 69 м (что соответствует среднему расстоянию распространения пыльцы в пределах древостоя для Q .petraea , вид с наименьшей способностью к расселению [22]). Мы обнаружили, выполнив логистическую регрессию, что доля аллоспецифических соседей оказала существенное влияние на соответствие между отдельными отнесениями в соответствии с критериями генотипического и морфологического сходства, с одной стороны, и критерием интерфертильности, с другой (χ 2).
Мы исследовали, может ли биотическая среда людей объяснить эти несоответствия.Наша гипотеза состоит в том, что соседство каждого дерева влияет на его систему спаривания и, таким образом, может влиять на его отнесение к видам на основе критерия интерфертильности, тогда как оно вряд ли повлияет на его отнесение к видам на основе генотипических и морфологических критериев. Поэтому мы исследовали окрестности каждого дерева, для которых отнесение к видам на основе критериев генотипического и морфологического сходства было конгруэнтным (N = 192). Для каждого дерева мы рассчитали долю аллоспецифических соседей в радиусе 69 м (что соответствует среднему расстоянию распространения пыльцы в пределах древостоя для Q .petraea , вид с наименьшей способностью к расселению [22]). Мы обнаружили, выполнив логистическую регрессию, что доля аллоспецифических соседей оказала существенное влияние на соответствие между отдельными отнесениями в соответствии с критериями генотипического и морфологического сходства, с одной стороны, и критерием интерфертильности, с другой (χ 2). = 6,5, df = 1, значение p = 0,01). У лиц с совпадающими назначениями в среднем было меньше аллоспецифических соседей (29% по сравнению с 51% для лиц с неконгруэнтными назначениями).Следовательно, определение отдельных видов на основе критерия интерфертильности зависело от окружающей среды.
= 6,5, df = 1, значение p = 0,01). У лиц с совпадающими назначениями в среднем было меньше аллоспецифических соседей (29% по сравнению с 51% для лиц с неконгруэнтными назначениями).Следовательно, определение отдельных видов на основе критерия интерфертильности зависело от окружающей среды.
Обсуждение
Насколько нам известно, это первый случай, когда критерий интерфертильности успешно используется для определения границ видов в естественных условиях. Анализ сети событий спаривания между парами взрослых деревьев, построенный на основе мощного анализа отцовства большого количества сеянцев, полученных в естественных условиях, позволил нам идентифицировать две группы интерфертильных особей с лишь несколькими событиями спаривания между ними. группы.Две группы, которые были разграничены, соответствующие двум видам в соответствии с расслабленной интерпретацией концепции биологических видов (рис. 1B), были тесно связаны с группами, полученными ранее с использованием морфологического и генотипического сходства в качестве критериев для определения границ видов [23,26]. Действительно, 88% индивидуумов были последовательно классифицированы в соответствии с критериями интерфертильности, морфологического сходства и генотипического сходства. Наши результаты не подтверждают более ранние утверждения о том, что критерий интерферируемости не может быть применен в полевых условиях (например,г. [14,15]), особенно в роду Quercus [27]. Вместо этого они показывают, что анализ сетей спаривания можно использовать для определения границ видов в соответствии с концепцией биологических видов, как впервые было предложено Сокалом и Кровелло [14].
Действительно, 88% индивидуумов были последовательно классифицированы в соответствии с критериями интерфертильности, морфологического сходства и генотипического сходства. Наши результаты не подтверждают более ранние утверждения о том, что критерий интерферируемости не может быть применен в полевых условиях (например,г. [14,15]), особенно в роду Quercus [27]. Вместо этого они показывают, что анализ сетей спаривания можно использовать для определения границ видов в соответствии с концепцией биологических видов, как впервые было предложено Сокалом и Кровелло [14].
Однако этот метод определения границ видов имеет два основных недостатка. Во-первых, сложно собрать адекватные сетевые данные. В нашем исследовании мы провели анализ отцовства на 3046 потомках, произведенных 51 матерью, чтобы построить сеть спаривания для взрослых деревьев.Несмотря на очень большое количество потомства, наши сетевые данные не позволили нам отнести всех особей в древостоях к видам. Не все люди дали потомство, а некоторые дали слишком мало потомков, чтобы их можно было надежно подключить к сети. Например, трое из особей, назначение которых на основе критерия интерфертильности отличалось от распределения на основе трех других критериев, были представлены одним потомством в тесте на потомство. Таким образом, они были подключены к брачной сети всего одним звеном.Во-вторых, план выборки может вызвать некоторую неоднородность в сетевой структуре, которая стирает биологическую неоднородность, вызванную существованием различных видов. Это произошло в наших сетевых данных, потому что мы собрали потомство только с 20% деревьев в насаждении. Таким образом, собранные деревья (то есть материнские деревья) имели больше связей в сети спаривания, чем другие деревья. Чтобы решить обе проблемы, нужно было собрать семена со всех особей в насаждении, предполагая, что все они дали семена.В принципе, эта цель может быть достигнута с помощью нашей биологической системы, если отобрать образцы на несколько лет, потому что виды дуба многолетние и однодомные. Однако для однолетних и раздельнополых видов это невозможно.
Например, трое из особей, назначение которых на основе критерия интерфертильности отличалось от распределения на основе трех других критериев, были представлены одним потомством в тесте на потомство. Таким образом, они были подключены к брачной сети всего одним звеном.Во-вторых, план выборки может вызвать некоторую неоднородность в сетевой структуре, которая стирает биологическую неоднородность, вызванную существованием различных видов. Это произошло в наших сетевых данных, потому что мы собрали потомство только с 20% деревьев в насаждении. Таким образом, собранные деревья (то есть материнские деревья) имели больше связей в сети спаривания, чем другие деревья. Чтобы решить обе проблемы, нужно было собрать семена со всех особей в насаждении, предполагая, что все они дали семена.В принципе, эта цель может быть достигнута с помощью нашей биологической системы, если отобрать образцы на несколько лет, потому что виды дуба многолетние и однодомные. Однако для однолетних и раздельнополых видов это невозможно. Другой возможностью уменьшить шум, вызванный выборкой, может быть введение структуры выборки в качестве ковариаты в статистической модели (например, [28]). К сожалению, непрерывная стохастическая блочная модель [24], которая была выбрана для этого исследования, поскольку она позволяет моделировать непрерывные изменения в свойствах связности узлов, в настоящее время не позволяет включать ковариаты.
Другой возможностью уменьшить шум, вызванный выборкой, может быть введение структуры выборки в качестве ковариаты в статистической модели (например, [28]). К сожалению, непрерывная стохастическая блочная модель [24], которая была выбрана для этого исследования, поскольку она позволяет моделировать непрерывные изменения в свойствах связности узлов, в настоящее время не позволяет включать ковариаты.
Наши результаты также показывают, что анализ сети современных отношений родства является подходящим методом для определения границ видов. Две группы, обнаруженные в нашем исследовании, могут быть интерпретированы как соответствующие двум различным «филогенетическим видам» [29], если филогенетические отношения рассматриваются в широком смысле, включая современные родословные. Методы разграничения видов, основанные на концепции филогенетических видов, почти исключительно сосредоточены на глубоком происхождении с использованием филогенетических методов, основанных на деревьях (рассмотрено в 30, но см. 31).Эти методы не подходят для разграничения гибридизующихся видов, потому что горизонтальный перенос генов между видами, вызванный гибридизацией и последующими событиями обратного скрещивания, вызывает конфликты между деревьями генов и деревьями видов [32,33]. По сравнению с данными о спаривании, данные о родстве было легче получить, и проблем с выборкой не возникало. Анализ сети родства выявил две группы людей с высокой степенью родства внутри группы и низкой степенью межгруппового родства. Эти две группы очень соответствовали группам, полученным с использованием интерфертильности, морфологического сходства и генотипического сходства в качестве критериев, что указывает на то, что анализ сетей родства может иметь потенциал для определения границ видов.Однако у этого метода есть и недостатки: лучшая модель имела пять групп связанных лиц, и мы не нашли никакой гипотезы, объясняющей их происхождение; Таким образом, количество видов должно быть известно заранее, чтобы применить этот метод.
31).Эти методы не подходят для разграничения гибридизующихся видов, потому что горизонтальный перенос генов между видами, вызванный гибридизацией и последующими событиями обратного скрещивания, вызывает конфликты между деревьями генов и деревьями видов [32,33]. По сравнению с данными о спаривании, данные о родстве было легче получить, и проблем с выборкой не возникало. Анализ сети родства выявил две группы людей с высокой степенью родства внутри группы и низкой степенью межгруппового родства. Эти две группы очень соответствовали группам, полученным с использованием интерфертильности, морфологического сходства и генотипического сходства в качестве критериев, что указывает на то, что анализ сетей родства может иметь потенциал для определения границ видов.Однако у этого метода есть и недостатки: лучшая модель имела пять групп связанных лиц, и мы не нашли никакой гипотезы, объясняющей их происхождение; Таким образом, количество видов должно быть известно заранее, чтобы применить этот метод.
Сравнивая полученные результаты с четырьмя критериями, используемыми для определения границ видов (т.е. интерфертильность, родственные отношения, морфологическое или генотипическое сходство), мы показали, что границы видов в значительной степени совпадали между методами определения границ видов.Наш анализ подтвердил существование двух групп индивидуумов, которые были морфологически и генетически дифференцированы. Мы также показали, что особи каждой группы преимущественно спаривались и были более связаны друг с другом, чем с особями из другой группы. Таким образом, в изученном насаждении было две «эволюционных линии». Концепция родовых видов, введенная Симпсоном [34,35], затем принятая Уайли [36] и де Кейросом [16,37,38], фокусируется на вопросе соответствия между методами разграничения видов.По мнению этих авторов, современные концепции видов (например, морфологические, филогенетические, генотипические и биологические) явно или неявно ассимилируют виды «с отдельно развивающимися (сегментами) метапопуляционных линий» и, таким образом, все являются побочными продуктами концепции линейных видов [16, 17]. Это должно объяснить высокую степень соответствия между методами определения границ видов.
Это должно объяснить высокую степень соответствия между методами определения границ видов.
Другим важным результатом этого сравнения является то, что независимо от критерия, используемого для определения границ видов, мы обнаружили промежуточных особей, которые обладали чертами обоих видов.Интересно, что люди, классифицируемые как промежуточные, часто различались в зависимости от метода. В частности, ни один человек не был последовательно классифицирован как промежуточный по всем четырем методам. Эти расхождения можно объяснить пороговыми значениями, которые были выбраны эмпирически для определения границ чистопородных видов, а также проблемами качества данных. Как упоминалось выше, изучение большего количества потомков на родительском дереве может улучшить разграничение видов на основе критерия интерфертильности. Точно так же большее количество молекулярных маркеров [39] может улучшить методы разграничения видов на основе критериев генотипа и родства.Точно так же большее количество морфологических маркеров [26] может улучшить морфологическое разграничение видов. Однако мы полагаем, что эти расхождения также могут отражать биологическую реальность. Действительно, как показано в других исследованиях [40–42], в том числе в дубах [22,43], относительная численность видов влияет на скорость гибридизации. Человек имеет тенденцию к размножению со своими соседями. Если он окружен многочисленными аллоспецифическими и немногими аналогами (например, [22,42]), это может привести к значительной гибридизации. Такой человек будет иметь тенденцию быть отнесенным к другому виду или к классу с промежуточным воспроизводством в соответствии с методами определения границ, основанными на интерфертильности.Следовательно, мы ожидаем некоторых расхождений в отнесении видов между методами, основанными на критериях, зависящих от среды (например, на основе критерия интерфертильности), и методами, основанными на критериях, не зависящих от среды (например, на основе критерия генотипического сходства). Из-за этих фундаментальных различий между методами невозможно вычислить эталонный набор данных, который дал бы правильное назначение для каждого человека.
Однако мы полагаем, что эти расхождения также могут отражать биологическую реальность. Действительно, как показано в других исследованиях [40–42], в том числе в дубах [22,43], относительная численность видов влияет на скорость гибридизации. Человек имеет тенденцию к размножению со своими соседями. Если он окружен многочисленными аллоспецифическими и немногими аналогами (например, [22,42]), это может привести к значительной гибридизации. Такой человек будет иметь тенденцию быть отнесенным к другому виду или к классу с промежуточным воспроизводством в соответствии с методами определения границ, основанными на интерфертильности.Следовательно, мы ожидаем некоторых расхождений в отнесении видов между методами, основанными на критериях, зависящих от среды (например, на основе критерия интерфертильности), и методами, основанными на критериях, не зависящих от среды (например, на основе критерия генотипического сходства). Из-за этих фундаментальных различий между методами невозможно вычислить эталонный набор данных, который дал бы правильное назначение для каждого человека. Таким образом, наши результаты не могут быть использованы для определения одного метода разграничения видов, который дал бы более надежные определения, чем другие.Вместо этого наши результаты показывают, что разные методы разграничения видов дают несколько разные результаты в применении к реальным биологическим данным.
Таким образом, наши результаты не могут быть использованы для определения одного метода разграничения видов, который дал бы более надежные определения, чем другие.Вместо этого наши результаты показывают, что разные методы разграничения видов дают несколько разные результаты в применении к реальным биологическим данным.
Заключение
Наши результаты подтвердили существование двух дифференцированных групп особей на участке исследования, соответствующих двум видам: Quercus robur и Q . petraea . Однако, в зависимости от критерия, используемого для отнесения особей к видам (т. Е. Интерфертильности, родства, морфологического или генотипического сходства), граница между видами не была точно такой же.Большинство различий связано с отнесением людей к промежуточной категории. Это открытие иллюстрирует непрерывный характер изменений между видами. Используемая нами модель, которая относится к категории, называемой «моделями степени членства» (рассмотренная в 10), подходит для синтеза непрерывных (но не однородных) вариаций биологического разнообразия. Однако, чтобы приблизиться к концепциям видов, которые обычно определяют виды как группы индивидуумов, мы наконец классифицировали индивидуумов на неперекрывающиеся группы.Таким образом, наш подход иллюстрирует влияние концепций на наше (неверное) представление видов и на наше понимание биологического разнообразия. Фрост и Хиллис [44], а также Майр [45] предложили определять виды как «целое», а не как группу особей. Согласно нашему исследованию, виды можно также определить как «крайнюю точку», к которой более или менее близки особи, что допускает возможность того, что особь представляет собой смесь двух разных видов.
Однако, чтобы приблизиться к концепциям видов, которые обычно определяют виды как группы индивидуумов, мы наконец классифицировали индивидуумов на неперекрывающиеся группы.Таким образом, наш подход иллюстрирует влияние концепций на наше (неверное) представление видов и на наше понимание биологического разнообразия. Фрост и Хиллис [44], а также Майр [45] предложили определять виды как «целое», а не как группу особей. Согласно нашему исследованию, виды можно также определить как «крайнюю точку», к которой более или менее близки особи, что допускает возможность того, что особь представляет собой смесь двух разных видов.
Материалы и методы
Определение границ видов на основе интерференции
Чтобы построить сеть спаривания для взрослых деревьев, мы использовали тест потомства с участием 3046 потомков, полученных в результате открытого опыления, собранных с 51 материнского дерева, разбросанного по всему насаждению (рис. S4 в файле S1).Анализ отцовства был проведен [22] путем генотипирования всех потомков из теста и всех взрослых деревьев, для которых была доступна ДНК, с использованием 12 мультиплексированных микросателлитных (SSR) маркеров, разработанных Guichoux et al. [46]. Согласно анализу отцовства, у 1575 потомков был только один возможный отец в стае, у 54 потомков было несколько потенциальных отцов в стае, а у 1417 потомков не было отца в стае [22]. По потомству, у которого был найден только отец-одиночка, мы идентифицировали 198 отцовских деревьев в насаждении.Среди этих деревьев 43 дерева, которые также были матерями, потому что дубы однодомны. На основании этих результатов мы реконструировали 1629 случаев спаривания между 206 взрослыми деревьями в насаждении. Эти события спаривания позволили нам идентифицировать 751 пару деревьев, которые спаривались хотя бы один раз, что указывает на то, что они были интерфертильными в естественных условиях. Эти данные были представлены неориентированной и невзвешенной сетью, в которой каждый из 206 узлов соответствовал взрослому дереву, а каждая из 751 ссылки соответствовала по крайней мере одному событию спаривания между двумя деревьями.
[46]. Согласно анализу отцовства, у 1575 потомков был только один возможный отец в стае, у 54 потомков было несколько потенциальных отцов в стае, а у 1417 потомков не было отца в стае [22]. По потомству, у которого был найден только отец-одиночка, мы идентифицировали 198 отцовских деревьев в насаждении.Среди этих деревьев 43 дерева, которые также были матерями, потому что дубы однодомны. На основании этих результатов мы реконструировали 1629 случаев спаривания между 206 взрослыми деревьями в насаждении. Эти события спаривания позволили нам идентифицировать 751 пару деревьев, которые спаривались хотя бы один раз, что указывает на то, что они были интерфертильными в естественных условиях. Эти данные были представлены неориентированной и невзвешенной сетью, в которой каждый из 206 узлов соответствовал взрослому дереву, а каждая из 751 ссылки соответствовала по крайней мере одному событию спаривания между двумя деревьями.
Затем сеть была смоделирована с помощью C-SBM [24]. Параметры модели — это коэффициенты связности между EHN и коэффициенты смеси EHN для каждого узла реальной сети. Для каждого возможного количества EHN параметры модели были выведены методом максимального правдоподобия, полученным с использованием программы MATLAB C-Mixnet (доступной по адресу http://www.agroparistech.fr/mia/doku.php?id=productions : logiciels /). Затем было определено оптимальное количество EHN в сети с использованием критерия AIC [24].Результаты визуализировали с помощью программного обеспечения pajek [47].
Для каждого возможного количества EHN параметры модели были выведены методом максимального правдоподобия, полученным с использованием программы MATLAB C-Mixnet (доступной по адресу http://www.agroparistech.fr/mia/doku.php?id=productions : logiciels /). Затем было определено оптимальное количество EHN в сети с использованием критерия AIC [24].Результаты визуализировали с помощью программного обеспечения pajek [47].
Определение границ видов на основе родства
Чтобы построить сеть родства, мы оценили родство 206 взрослых деревьев, включенных в сеть спаривания. Оценка проводилась с помощью программного обеспечения COANCESTRY [48], которое предлагает семь различных оценок родства. В соответствии с рекомендациями Ванга [48] мы использовали 1629 потомков, для которых оба родителя, как известно, определили наиболее подходящую оценку.Была выбрана триадная оценка правдоподобия (обозначенная как TrioML в COANCESTRY [49]), потому что она давала значения родства, наиболее близкие к нулю для неродственных потомков, наиболее близкие к 0,25 для полусибсов и наиболее близкие к 0,5 для полных братьев-сестер. С помощью этого оценщика максимальное значение родства между двумя неродственными потомками составило 0,22. Поэтому мы рассматривали 0,22 как порог: деревья, значение родства которых было выше, считались родственными особями, а другие деревья считались несвязанными.Затем отношения родства были представлены невзвешенной и ненаправленной сетью с 206 узлами, каждый из которых соответствует взрослому дереву, и 1078 звеньями, соединяющими людей, считающихся связанными. Как и в случае стыковочной сети, мы смоделировали структуру сети с помощью C-SBM [24] и визуализировали результаты с помощью программного обеспечения pajek [47].
С помощью этого оценщика максимальное значение родства между двумя неродственными потомками составило 0,22. Поэтому мы рассматривали 0,22 как порог: деревья, значение родства которых было выше, считались родственными особями, а другие деревья считались несвязанными.Затем отношения родства были представлены невзвешенной и ненаправленной сетью с 206 узлами, каждый из которых соответствует взрослому дереву, и 1078 звеньями, соединяющими людей, считающихся связанными. Как и в случае стыковочной сети, мы смоделировали структуру сети с помощью C-SBM [24] и визуализировали результаты с помощью программного обеспечения pajek [47].
Определение границ видов на основе морфологии
Критерий морфологического сходства ранее использовался Bacilieri et al. [26], чтобы идентифицировать все деревья с места исследования. Эти авторы выполнили факторный дискриминантный анализ (FDA) на основе 31 морфологического признака листа для определения границ вида. Их исследование выявило наличие двух групп особей, различающихся по морфологии. На первую ось FDA приходилось 33% общей дисперсии, и она сильно коррелировала с морфологическими маркерами, традиционно используемыми систематиками для различения Q . robur из Q .petraea . Распределение особей по этой оси было использовано для графического отнесения особей к двум чистым морфологическим группам (в данном исследовании они называются M1 и M2 и соответствуют Q . Robur и Q . Petraea соответственно) и к морфологически промежуточному классу (так называемому Mi). Среди 206 взрослых деревьев, включенных в сети спаривания и родства, 123 дерева были отнесены к M1, 80 — к M2 и 3 — к Mi (рисунок S5 в файле S1).
На первую ось FDA приходилось 33% общей дисперсии, и она сильно коррелировала с морфологическими маркерами, традиционно используемыми систематиками для различения Q . robur из Q .petraea . Распределение особей по этой оси было использовано для графического отнесения особей к двум чистым морфологическим группам (в данном исследовании они называются M1 и M2 и соответствуют Q . Robur и Q . Petraea соответственно) и к морфологически промежуточному классу (так называемому Mi). Среди 206 взрослых деревьев, включенных в сети спаривания и родства, 123 дерева были отнесены к M1, 80 — к M2 и 3 — к Mi (рисунок S5 в файле S1).
Определение границ видов на основе мультилокусных генотипов
Guichoux et al. [23] использовал генотипическое сходство в качестве критерия для отнесения деревьев исследуемого участка к видам. Эти авторы генотипировали взрослые деревья с помощью мультиплекса из 12 SSR, разработанного Guichoux et al. [46] и чип из 262 однонуклеотидных полиморфизмов (SNP), обогащенный маркерами, сильно дифференцированными между видами [23]. Они использовали программную структуру [50] для группировки особей в генотипические кластеры, но не определили формально оптимальное количество генотипических кластеров в насаждении перед выполнением кластеризации.Здесь мы использовали статистику ΔK [51], чтобы определить количество генетически различных групп. Оптимальное количество кластеров было два (рисунок S6 в файле S1), как ранее предполагали Guichoux et al. [23]. Таким образом, взрослые деревья были разделены на две чистокровные группы и один генетически промежуточный класс. Среди 206 взрослых деревьев, включенных в сети спаривания и родства, 78 деревьев были отнесены к первой чистокровной группе (далее называемой G1), 118 — ко второй чистокровной группе (G2) и 10 — к генетически промежуточному классу (Gi) (Рисунок S7. в файле S1).
[46] и чип из 262 однонуклеотидных полиморфизмов (SNP), обогащенный маркерами, сильно дифференцированными между видами [23]. Они использовали программную структуру [50] для группировки особей в генотипические кластеры, но не определили формально оптимальное количество генотипических кластеров в насаждении перед выполнением кластеризации.Здесь мы использовали статистику ΔK [51], чтобы определить количество генетически различных групп. Оптимальное количество кластеров было два (рисунок S6 в файле S1), как ранее предполагали Guichoux et al. [23]. Таким образом, взрослые деревья были разделены на две чистокровные группы и один генетически промежуточный класс. Среди 206 взрослых деревьев, включенных в сети спаривания и родства, 78 деревьев были отнесены к первой чистокровной группе (далее называемой G1), 118 — ко второй чистокровной группе (G2) и 10 — к генетически промежуточному классу (Gi) (Рисунок S7. в файле S1).
Благодарности
Мы благодарны Алексису Дукуссо, который организовал тест потомства Petite Charnie и поделился информацией о стенде, а также за его помощь вместе со Стефани Вагнер во время отбора проб. Благодарим ОНФ за организацию стенда. Патрик Леже очень помог с микросателлитным генотипированием. Генотипирование было выполнено на предприятии Genome-Transcriptome в Бордо. Мы особенно благодарны Франсуа Юберу за полезные обсуждения филогении и Бастьену Кастагнейролу, Вирджилу Фьеве, Сирилу Дутеху, Антуану Кремеру и двум анонимным рецензентам за их конструктивные комментарии к рукописи.
Благодарим ОНФ за организацию стенда. Патрик Леже очень помог с микросателлитным генотипированием. Генотипирование было выполнено на предприятии Genome-Transcriptome в Бордо. Мы особенно благодарны Франсуа Юберу за полезные обсуждения филогении и Бастьену Кастагнейролу, Вирджилу Фьеве, Сирилу Дутеху, Антуану Кремеру и двум анонимным рецензентам за их конструктивные комментарии к рукописи.
Вклад авторов
Эксперимент задумал и спроектировал: LL RJP. Проведены эксперименты: LL. Проанализированы данные: LL JBL JJD CV. Предоставленные реагенты / материалы / инструменты анализа: JBL JJD. Написал рукопись: LL JBL JJD RJP CV.
Ссылки
- 1. Майр Э. (1942) Систематика и происхождение видов. Нью-Йорк: издательство Колумбийского университета.
- 2.
Деттман Дж. Р., Якобсон Д. Д., Тернер Е., Прингл А., Тейлор Дж. В. (2003) Репродуктивная изоляция и филогенетическая дивергенция в Neurospora : сравнение методов распознавания видов в модельном эукариоте.
 Evolution 57: 2721-2741. DOI: https: //doi.org/10.1554/03-074. PubMed: 14761052.
Evolution 57: 2721-2741. DOI: https: //doi.org/10.1554/03-074. PubMed: 14761052. - 3. Маркуссен Т., Борген Л. (2011) Определение границ понто-кавказского комплекса Viola sieheana на основе данных аллозимов, морфологии, уровней плоидности и экспериментов по скрещиванию. Plant Syst Evol 291: 183-196. DOI: https: //doi.org/10.1007/s00606-010-0377-z.
- 4. Марин Дж., Кроу-Рой Б., Хемптин Дж. Л., Лекомпте Э, Магро А (2010) Coccinella septempunctata (Coleoptera, Coccinellidae): комплекс видов? Zool Scripta 39: 591-602.DOI: https: //doi.org/10.1111/j.1463-6409.2010.00450.x.
- 5. Hibbett DS, Fukumasa-Nakai Y, Tsuneda A, Donoghue MJ (1995) Филогенетическое разнообразие шиитаке, выведенное из последовательностей ядерной рибосомной ДНК. Mycologia 87: 618-638. DOI: https://doi.org/10.2307/3760806.
- 6.
Тейлор Дж. У., Якобсон Д. Д., Крокен С., Касуга Т., Гейзер Д. М. и др. (2000) Филогенетическое распознавание видов и видовые концепции грибов.
 Fungal Genet Biol 31: 21-32. DOI: https: //doi.org/10.1006/fgbi.2000.1228. PubMed: 11118132.
Fungal Genet Biol 31: 21-32. DOI: https: //doi.org/10.1006/fgbi.2000.1228. PubMed: 11118132. - 7. Fortuna MA, García C, Guimarães PR Jr, Bascompte J (2008) Пространственные сети спаривания у растений, опыляемых насекомыми. Ecol Lett 11: 490-498. DOI: https: //doi.org/10.1111/j.1461-0248.2008.01167.x. PubMed: 18318718.
- 8. Ньюман М (2003) Структура и функции сложных сетей. Сиам Ред. 45: 167-256. DOI: https://doi.org/10.1137/S003614450342480.
- 9. Fortunato S (2010) Обнаружение сообществ в графах.Phys Rep 486: 75-174. DOI: https://doi.org/10.1016/j.physrep.2009.11.002.
- 10. Leger J-B, Vacher C, Daudin J-J (2013) Обнаружение структурно однородных подмножеств в графах. Statist Comput (в печати).
- 11. Орр Х.А. (2001) Некоторые сомнения относительно (еще одного) взгляда на виды. J Evol Biol 14: 870-871. DOI: https: //doi.org/10.1046/j.1420-9101.2001.00340.x.
- 12.
Noor MAF (2002) Показывает ли концепция биологических видов свой возраст? Тенденции Ecol Evol 17: 153-154.
 DOI: https: //doi.org/10.1016/S0169-5347 (02) 02452-7.
DOI: https: //doi.org/10.1016/S0169-5347 (02) 02452-7. - 13. Coyne JA, Orr HA (2004) Видообразование. Сандерленд, Массачусетс, США: Sinauer Associates.
- 14. Сокал Р.Р., Кровелло Т.Дж. (1970) Концепция биологических видов: критическая оценка. Am Nat 104: 127-153. DOI: https: //doi.org/10.1086/282646.
- 15. de Meeûs T, Durand P, Renaud F (2003) Видовые концепции: зачем? Trends Parasitol 19: 425-427. DOI: https: //doi.org/10.1016/S1471-4922 (03) 00195-8.PubMed: 14519574.
- 16. de Queiroz (2005) Эрнст Майр и современная концепция видов. Proc Natl Acad Sci U S A 102: 6600-6607. DOI: https: //doi.org/10.1073/pnas.0502030102. PubMed: 15851674.
- 17. Хаусдорф Б. (2011) Прогресс в направлении общей концепции видов. Evolution 65: 923-931. DOI: https: //doi.org/10.1111/j.1558-5646.2011.01231.x. PubMed: 21463293.
- 18.
Streiff R, Ducousso A, Kremer A (1998) Пространственная генетическая структура и поток генов пыльцы в смешанном дубовом насаждении.
 Genet Sel Evol 30: S137-S152. DOI: https: //doi.org/10.1186/1297-9686-30-S1-S137.
Genet Sel Evol 30: S137-S152. DOI: https: //doi.org/10.1186/1297-9686-30-S1-S137. - 19. Ризеберг Л.Х., Карни С.Е. (1998) Гибридизация растений. Новый Phytol 140: 599-624. DOI: https: //doi.org/10.1046/j.1469-8137.1998.00315.x.
- 20. Petit RJ, Bodénès C, Ducousso A, Roussel G, Kremer A (2003) Гибридизация как механизм вторжения в дубы. Новый Фитол. 161: 151-164. DOI: https: //doi.org/10.1046/j.1469-8137.2003.00944.x.
- 21. Lepais O, Gerber S (2011) Репродуктивные паттерны формируют динамику интрогрессии и сукцессию видов в пределах комплекса видов европейского белого дуба.Evolution 65: 156-170. DOI: https: //doi.org/10.1111/j.1558-5646.2010.01101.x. PubMed: 20722727.
- 22. Lagache L, Klein EK, Guichoux E, Petit RJ (2013) Мелкомасштабный экологический контроль гибридизации дубов. Мол Экол 22: 423-436. DOI: https: //doi.org/10.1111/mec.12121. PubMed: 23173566.
- 23.
Guichoux E, Garnier-Géré P, Lagache L, Lang T, Bourry C et al.
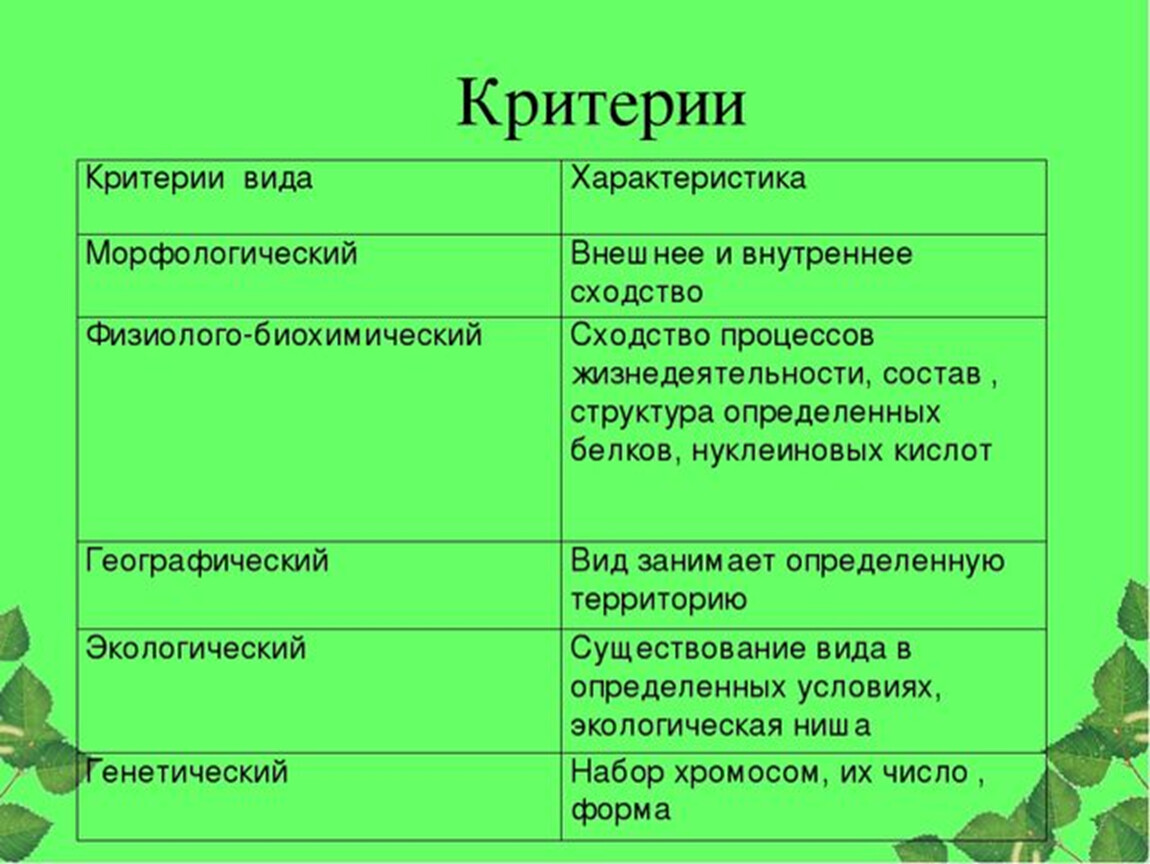 (2013) Outlier loci подчеркивают направление интрогрессии у дубов. Мол Экол 22: 450-462. doi: https: // doi.org / 10.1111 / mec.12125. PubMed: 231
(2013) Outlier loci подчеркивают направление интрогрессии у дубов. Мол Экол 22: 450-462. doi: https: // doi.org / 10.1111 / mec.12125. PubMed: 231.
- 24. Даудин Дж. Дж., Пьер Л., Вашер С. (2010) Модель гетерогенных случайных сетей с использованием непрерывных скрытых переменных и приложение к сети «дерево-гриб». Биометрия 66: 1043-1051. DOI: https: //doi.org/10.1111/j.1541-0420.2009.01378.x. PubMed: 20105159.
- 25. Bacilieri R, Ducousso A, Petit RJ, Kremer A (1996) Система спаривания и асимметричная гибридизация в смешанном насаждении европейских дубов. Evolution 50: 900-908.DOI: https: //doi.org/10.2307/2410861.
- 26. Bacilieri R, Ducousso A, Kremer A (1996) Сравнение морфологических признаков и молекулярных маркеров для анализа гибридизации скального и черешчатого дуба. Ann Sci Forestières 53: 79-91. DOI: https: //doi.org/10.1051/forest: 19960106.
- 27.
Донохью М.Дж. (1985) Критика концепции биологических видов и рекомендации по филогенетической альтернативе.
 Бриолог 88: 172-181. doi: https: //doi.org/10.2307/3243026.
Бриолог 88: 172-181. doi: https: //doi.org/10.2307/3243026. - 28. Мариадассу М., Робин С., Вашер С. (2010) Выявление скрытой структуры в значных графах: вариационный подход. Annals Appl Statistics 4: 715-742. DOI: https: //doi.org/10.1214/10-AOAS361.
- 29. Элдридж Н., Крафт Дж. (1980) Филогенетические закономерности и эволюционный процесс. Нью-Йорк: издательство Колумбийского университета.
- 30. Сайты JJW, Jonathon CM (2004) Рабочие критерии для определения границ видов. Annu Rev Ecol Evol Syst 35: 199-227.DOI: https: //doi.org/10.1146/annurev.ecolsys.35.112202.130128.
- 31.
Moalic Y, Arnaud-Haond S, Perrin C, Pearson GA, Serrao EA (2011) Путешествие во времени с сетями: выявление современной гибридизации и предкового полиморфизма между двумя видами бурых водорослей, Fucus vesiculosus и F. spiralis . BMC Evol Biol 11: 33. DOI: https: //doi.org/10.1186/1471-2148-11-33. PubMed: 21281515.

- 32. Эдвардс С.В. (2009) Возникает ли новая общая теория молекулярной систематики? Эволюция 63: 1-19.DOI: https: //doi.org/10.1111/j.1558-5646.2008.00549.x. PubMed: 19146594.
- 33. Мэддисон В.П. (1997) Генные деревья в деревьях видов. Syst Biol 46: 523-536. DOI: https: //doi.org/10.1093/sysbio/46.3.523.
- 34. Симпсон Г.Г. (1951) Видовое понятие. Evolution 5: 285-298. DOI: https: //doi.org/10.2307/2405675.
- 35. Симпсон Г.Г. (1961) Принципы систематики животных. Нью-Йорк: издательство Колумбийского университета.
- 36. Wiley EO (1978) Пересмотр концепции эволюционных видов.Syst Biol 27: 17-26.
- 37. de Queiroz K (2007) Видовые концепции и разграничение видов. Syst Biol 56: 879-886. DOI: https://doi.org/10.1080/10635150701701083. PubMed: 18027281.
- 38.
de Queiroz K (1998) Общая концепция происхождения видов, критерии видов и процесс видообразования.
 Издательство Оксфордского университета.
Издательство Оксфордского университета. - 39. Vähä J-P, Primmer CR (2006) Эффективность основанных на моделях байесовских методов для обнаружения гибридных особей при различных сценариях гибридизации и с различным количеством локусов.Мол Экол 15: 63-72. PubMed: 16367830.
- 40. Field DL, Ayre DJ, Whelan RJ, Young AG (2008) Относительная частота симпатрических видов влияет на скорость межвидовой гибридизации, производство семян и производительность всходов у необычного Eucalyptus aggregata . Дж. Экол 96: 1198-1210. DOI: https: //doi.org/10.1111/j.1365-2745.2008.01434.x.
- 41. Фокке В.О. (1881) Die Pflanzenmischlinge. Берлин: Bornträger.
- 42. Хаббс К.Л. (1955) Гибридизация между видами рыб в природе.Систематика зоологии 4: 1-20.
- 43.
Lepais O, Petit RJ, Guichoux E, Lavabre JE, Alberto F et al. (2009) Относительное обилие видов и направление интрогрессии дубов. Мол Экол 18: 2228-2242.
 DOI: https: //doi.org/10.1111/j.1365-294X.2009.04137.x. PubMed: 19302359.
DOI: https: //doi.org/10.1111/j.1365-294X.2009.04137.x. PubMed: 19302359. - 44. Frost DR, Hillis DM (1990) Виды в концепции и на практике: герпетологические приложения. Herpetologica 46: 86-104.
- 45. Майр Э. (1992) Местная флора и концепция биологических видов.Am J Bot 79: 222-238. DOI: https: //doi.org/10.2307/2445111.
- 46. Guichoux E, Lagache L, Wagner S, Léger P, Petit RJ (2011) Два высоко проверенных мультиплекса (12-сплетение и 8-сплетение) для определения границ видов и анализа происхождения дубов ( Quercus spp.). Мол Экол Ресур 11: 578-585. DOI: https: //doi.org/10.1111/j.1755-0998.2011.02983.x. PubMed: 21481218. ,
- , 47. Батагель В., Мрвар А. (1998) PAJEK — Программа для анализа больших сетей.
- 48.Ван Дж. Л. (2010) Coancestry: программа для моделирования, оценки и анализа коэффициентов родства и инбридинга. Мол Экол Ресур 11: 141-145. PubMed: 21429111.
- 49.
Ван Дж.
 Л. (2007) Триадные коэффициенты IBD и приложения к оценке попарного родства. Genet Res 89: 135-153. PubMed: 17894908.
Л. (2007) Триадные коэффициенты IBD и приложения к оценке попарного родства. Genet Res 89: 135-153. PubMed: 17894908. - 50. Притчард Дж. К., Стивенс М., Доннелли П. (2000) Вывод структуры популяции с использованием данных мультилокусного генотипа. Генетика 155: 945-959.PubMed: 10835412.
- 51. Эванно Г., Регнаут С., Годе Дж. (2005) Определение количества групп людей с использованием структуры программного обеспечения: исследование моделирования. Мол Экол 14: 2611-2620. DOI: https: //doi.org/10.1111/j.1365-294X.2005.02553.x. PubMed: 15969739.
- 52. Маллет Дж. (1995) Определение видов для современного синтеза. Trends Ecol Evol 10: 294-299. DOI: https: //doi.org/10.1016/0169-5347 (95) -4. PubMed: 21237047.
- 53. Нельсон Г., Плантик Н. (1981) Систематика и биогеография: кладистика и викариантность.Нью-Йорк: издательство Колумбийского университета.
- 54.
Мишлер Б.Д. (1985) Морфологические, онтогенетические и филогенетические основы видовых представлений у мохообразных.
 Бриолог 88: 207-214. DOI: https: //doi.org/10.2307/3243030.
Бриолог 88: 207-214. DOI: https: //doi.org/10.2307/3243030. - 55. Legendre G, Legendre P (1984) Ecologie numérique. Париж, Франция: Массон.
ПРОГРЕСС НА ПУТИ К ОБЩЕЙ КОНЦЕПЦИИ ВИДОВ — Хаусдорф — 2011 — Эволюция
Хотя «вид» является одной из наиболее важных единиц в биологии, определение этого термина все еще остается спорным.Большая часть биологической информации дается со ссылкой на вид. Существует целая дисциплина, таксономия, посвященная разграничению видов, которая требует концепции вида. Понятия видов не только определяют, что такое вид, но, определяя, что такое вид, они также определяют, что такое видообразование. Таким образом, исследовательские программы, посвященные условиям и факторам, приводящим к видообразованию, зависят от концепции вида. Многие другие биологические исследования также зависят от определения границ видов. Например, многие экологические и поведенческие исследования исследуют взаимодействие видов или взаимосвязь видового богатства с абиотическими и биотическими факторами. Большинство усилий в области природоохранной биологии сосредоточено на видах. Даже в законодательстве этот вид является одной из наиболее часто используемых биологических единиц не только с точки зрения сохранения, но и с точки зрения сельского хозяйства или медицины. Таким образом, четкое определение термина «вид» важно для более широкой аудитории, а не только для академических биологов.
Большинство усилий в области природоохранной биологии сосредоточено на видах. Даже в законодательстве этот вид является одной из наиболее часто используемых биологических единиц не только с точки зрения сохранения, но и с точки зрения сельского хозяйства или медицины. Таким образом, четкое определение термина «вид» важно для более широкой аудитории, а не только для академических биологов.
Обсуждение концепций различных видов — это не просто философский спор, но различные концепции видов могут фактически привести к разным выводам (Isaac et al.2004 г.). Например, обзор литературы выявил на 49% большее количество видов в исследованиях, использующих концепцию филогенетических видов, по сравнению с исследованиями, в которых применялись нефилогенетические концепции на тех же организмах, и связанное с этим уменьшение размера и ареала популяции (Agapow et al. 2004).
Из-за важности определения термина «вид» было предложено множество концепций видов (обзор см. В Mayden 1997; de Queiroz 1998; Harrison 1998; Coyne and Orr 2004).Однако до сих пор нет единого мнения о концепции вида. Бесконечные споры о концепции вида привели к разочарованию и снижению интереса к развитию концепции. Тем не менее, новое понимание процесса видообразования и природы «видов», накопленное за последнее десятилетие, требует корректировки концепции вида.
В Mayden 1997; de Queiroz 1998; Harrison 1998; Coyne and Orr 2004).Однако до сих пор нет единого мнения о концепции вида. Бесконечные споры о концепции вида привели к разочарованию и снижению интереса к развитию концепции. Тем не менее, новое понимание процесса видообразования и природы «видов», накопленное за последнее десятилетие, требует корректировки концепции вида.
Одно из наиболее важных новых открытий в отношении концепции вида заключается в том, что репродуктивные барьеры полупроницаемы для потока генов и что виды могут дифференцироваться, несмотря на продолжающееся скрещивание (Harrisson 1998; Rieseberg 2001; Wu 2001; Rieseberg et al.2003; Койн и Орр 2004; Ву и Тинг 2004; Молот 2005, 2008; Лексер и Видмер 2008). Хотя известно, что по крайней мере 25% видов растений и 10% видов животных гибридизуются с другими видами с потенциальной интрогрессией генов (Mallet 2005), в большинстве концепций видов не учитывается адекватный обмен генами между видами. Сохранение дифференцированных видов и продолжающаяся дифференциация, несмотря на обмен генами, ставят под сомнение эти концепции.
Дальнейшее понимание природы «видов», которое до сих пор не рассматривалось в контексте представлений о видах, было получено путем изучения параллельного видообразования.Параллельное видообразование — это случай параллельной эволюции, в которой сходные черты, обеспечивающие репродуктивную изоляцию, происходят независимо от отдельных близкородственных линий, что приводит к полифилетическому образованию, которое развивается независимо от предковых видов (Schluter and Nagel 1995; Rundle et al. 2000; Johannesson 2001; Носил и др., 2002). Черты, определяющие репродуктивную изоляцию, могут развиваться как побочный продукт адаптации к разным условиям, но они также могут возникать в результате других процессов, таких как независимая полиплоидизация.Фактически, большинство полиплоидных образований возникло в результате повторяющейся полиплоидизации, часто в различных популяциях предковых видов (Soltis and Soltis 1999, 2000). Независимо полученные полиплоидные особи могут скрещиваться друг с другом и образовывать согласованные полифилетические образования, обычно признаваемые видами.
Еще одно важное открытие заключается в том, что однопородные организмы на самом деле организованы в единицы, которые напоминают виды двупородных организмов (Holman 1987; Gevers et al.2005; Коэн и Перри 2007; Fontaneto et al. 2007; Cohan and Koeppel 2008). Термин «вид» используется для всех групп организмов. Таким образом, вопрос заключается в том, могут ли все явления в континууме репродуктивных систем от однокоренных до двупородных, которые называются видами, быть определены в общей видовой концепции.
Далее я буду обсуждать влияние этих идей на некоторые из наиболее широко принятых или наиболее инновационных концепций видов и попытаюсь разработать синтез существующих концепций и новых идей.
Понятие вида и критерии вида
de Queiroz (1998) подчеркнул различие между концепцией видов и критериями видов. Однако его определение «концепции вида» как «идеи о виде сущности, представленной категорией видов» недостаточно отчетливо, чтобы быть полезным. Концепция вида должна определять условия, которые необходимы и достаточны для идентификации группы особей как вида (см. Также Pigliucci 2003). Основываясь на этом определении, я выступаю против его точки зрения, согласно которой большинство ранее сформулированных концепций видов являются только видовыми критериями.Критерий вида — это условие, которое необходимо или достаточно для идентификации группы особей как вида. Условия, указанные в понятиях видов, являются видовыми критериями. Однако не все действительные критерии должны быть указаны непосредственно в концепции вида. Различные концепции видов часто относятся к разным стадиям видообразования (de Queiroz 1998, 2005a, b, 2007). Если понятие вида определяет условия, которые обычно достигаются на более поздней стадии дифференциации, чем условия другого понятия, то понятия вложены, и условия первого понятия являются достаточными критериями второго.
Концепция вида должна определять условия, которые необходимы и достаточны для идентификации группы особей как вида (см. Также Pigliucci 2003). Основываясь на этом определении, я выступаю против его точки зрения, согласно которой большинство ранее сформулированных концепций видов являются только видовыми критериями.Критерий вида — это условие, которое необходимо или достаточно для идентификации группы особей как вида. Условия, указанные в понятиях видов, являются видовыми критериями. Однако не все действительные критерии должны быть указаны непосредственно в концепции вида. Различные концепции видов часто относятся к разным стадиям видообразования (de Queiroz 1998, 2005a, b, 2007). Если понятие вида определяет условия, которые обычно достигаются на более поздней стадии дифференциации, чем условия другого понятия, то понятия вложены, и условия первого понятия являются достаточными критериями второго.
Представления о видах не являются правильными или неправильными. В принципе, можно использовать практически любую концепцию вида. Однако принятие концепции вида влечет за собой соответствующую адаптацию использования термина «вид» и разграничения видов. Если, например, адаптируется видовая концепция, которая не подразумевает, что группы полиплоидных особей должны рассматриваться как виды, отличные от других подобных диплоидных особей, текущая классификация должна быть соответствующим образом изменена.Таким образом, концепции видов должны оцениваться тем выше, чем лучше они соответствуют нынешнему использованию термина «виды».
В принципе, можно использовать практически любую концепцию вида. Однако принятие концепции вида влечет за собой соответствующую адаптацию использования термина «вид» и разграничения видов. Если, например, адаптируется видовая концепция, которая не подразумевает, что группы полиплоидных особей должны рассматриваться как виды, отличные от других подобных диплоидных особей, текущая классификация должна быть соответствующим образом изменена.Таким образом, концепции видов должны оцениваться тем выше, чем лучше они соответствуют нынешнему использованию термина «виды».
Понятия видов
ПОНЯТИЯ ВИДОВ НА ОСНОВЕ ЛИНИИ
де Кейрос (1998, 2005a, b, 2007) утверждал, что все современные концепции видов имеют общий элемент, а именно они явно или неявно приравнивают виды к « отдельно развивающимся (сегментам) метапопуляционных линий » (de Queiroz 2005).Эта «общая концепция происхождения», которая представляет собой переформулировку концепции эволюционного вида (Simpson 1951, 1961; Wiley 1978), стала популярной среди систематиков (Wiens 2007).
Чтобы понять концепции видов, основанные на происхождении, сначала необходимо дать определение терминам «происхождение» и «популяция». Линия определяется как «последовательность потомков-предков популяций» (Simpson, 1961). Термин «популяция» обычно определяется как группа организмов одного вида, которые занимают более или менее четко определенный географический регион и демонстрируют непрерывность воспроизводства от поколения к поколению (Futuyma 1998).Есть несколько вариантов этого определения (Waples and Gaggiotti 2006). В любом случае необходимо указать, какие из особей, которые совместно встречаются на территории, образуют популяцию. В некоторых определениях «популяции» более обычные фразы «особи одного вида» или «особи одного вида» заменены на «скрещивающиеся особи» (например, Mayr 1942, 1963; de Queiroz and Donoghue 1988). Однако такое определение популяции вводит понятие биологического вида через черный ход, включая его в определение популяции. Это определение не использовалось де Кейросом (1998), когда он вводил общую концепцию линии передачи. Скорее он подчеркивал, что использует популяцию «в общем смысле организационного уровня выше уровня организма, а не в конкретном смысле репродуктивного сообщества половых организмов» (de Queiroz 1998). Он не уточнил никаких свойств этого «организационного уровня выше, чем у организма», оставив неясным точное определение популяции и, таким образом, происхождения и вида. Более позднее добавление термина «метапопуляция» к общей концепции происхождения и его определение как «инклюзивная популяция, состоящая из набора связанных субпопуляций» де Кейросом (2005a, b) не помогло прояснить концепцию, а скорее ввело дополнительные неопределенные термины. .Не было объяснено, как определяются «субпопуляции» и как они «связаны».
Это определение не использовалось де Кейросом (1998), когда он вводил общую концепцию линии передачи. Скорее он подчеркивал, что использует популяцию «в общем смысле организационного уровня выше уровня организма, а не в конкретном смысле репродуктивного сообщества половых организмов» (de Queiroz 1998). Он не уточнил никаких свойств этого «организационного уровня выше, чем у организма», оставив неясным точное определение популяции и, таким образом, происхождения и вида. Более позднее добавление термина «метапопуляция» к общей концепции происхождения и его определение как «инклюзивная популяция, состоящая из набора связанных субпопуляций» де Кейросом (2005a, b) не помогло прояснить концепцию, а скорее ввело дополнительные неопределенные термины. .Не было объяснено, как определяются «субпопуляции» и как они «связаны».
Если принято обычное определение популяции как группы организмов того же вида, которые занимают более или менее четко определенный географический регион, популяция (и, следовательно, суб- или метапопуляция) может быть разграничена только в том случае, если мы можем решить, какие организмы сородичи. Таким образом, виды должны быть определены до того, как можно будет определить границы популяций. Определение вида как последовательности (мета) популяций является циклическим, поскольку часть определения «популяции» состоит в том, что организмы, образующие популяцию, принадлежат к одному и тому же виду.Термин «популяция» не должен использоваться в определениях «вида», если он не определен без ссылки на организмы того же вида. Обратите внимание, что это необходимо, чтобы избежать округлости, и не указывает на какие-либо сомнения в эволюционной важности популяций.
Таким образом, виды должны быть определены до того, как можно будет определить границы популяций. Определение вида как последовательности (мета) популяций является циклическим, поскольку часть определения «популяции» состоит в том, что организмы, образующие популяцию, принадлежат к одному и тому же виду.Термин «популяция» не должен использоваться в определениях «вида», если он не определен без ссылки на организмы того же вида. Обратите внимание, что это необходимо, чтобы избежать округлости, и не указывает на какие-либо сомнения в эволюционной важности популяций.
Понятие «отдельно развивающихся» единиц, несомненно, является стержнем всех современных концепций видов. Однако, чтобы быть полезным критерием в концепции вида, необходимо указать, что означает «отдельно эволюционирующий».Неясно, как следует классифицировать популяции, связанные ограниченным потоком генов. В результате интрогрессии таксоны с двумя родителями могут не развиваться отдельно в некоторых локусах, но развиваются в большинстве других (Harrison 1998; Wu 2001; Coyne and Orr 2004; Wu and Ting 2004; Mallet 2005). Более того, когерентные полифилетические сущности, которые возникли в результате параллельного видообразования и эволюционируют отдельно от предковых видов, не могут быть классифицированы как отдельные виды с помощью концепций видов, основанных на родословной.
Более того, когерентные полифилетические сущности, которые возникли в результате параллельного видообразования и эволюционируют отдельно от предковых видов, не могут быть классифицированы как отдельные виды с помощью концепций видов, основанных на родословной.
Концепции видов, основанные на происхождении, переносят проблему определения термина «вид» на проблему определения термина «популяция» и приводят к сложной переформулировке концепции биологического вида или к круговым рассуждениям.Более того, важный критерий «отдельно эволюционирующий» в общей концепции родословной оставался неясным, поскольку не уточнялось, что означает это понятие. Таким образом, концепции видов, основанные на происхождении, представляют лишь ограниченный прогресс в направлении общеприменимой концепции видов.
КОНЦЕПЦИЯ БИОЛОГИЧЕСКОГО ВИДА
Наиболее влиятельной концепцией видов по-прежнему является концепция биологических видов, которая определяет виды как « группу фактически или потенциально скрещивающихся естественных популяций, репродуктивно изолированных от других таких групп » (Mayr 1942, 1963; «естественные популяции» в этой формулировке можно заменить словом «индивидуумы» без изменения смысла). Однако два новых открытия бросают вызов концепции биологического вида. Открытие того, что репродуктивные барьеры полупроницаемы для потока генов и что виды могут дифференцироваться, несмотря на продолжающееся скрещивание (Harrisson 1998; Rieseberg 2001; Wu 2001; Rieseberg et al. 2003; Coyne and Orr 2004; Wu and Ting 2004; Mallet 2005, 2008; Lexer) и Widmer 2008) указывает на то, что концепция биологических видов не соответствует нынешнему использованию термина «виды». Принятие этой концепции потребует объединения многих хорошо дифференцированных и общепринятых видов, которые, тем не менее, регулярно скрещиваются.Некоторые авторы (Орр, 2001; Нур, 2002; Койн, Орр, 2004) выступали за смягчение интерпретации концепции биологических видов, чтобы сохранить ее, несмотря на новые идеи. Однако смысл концепции вида состоит в том, что она определяет условия, при которых группа особей должна быть классифицирована как вид. Если игнорировать условия, указанные в концепции вида, она становится бесполезной.
Однако два новых открытия бросают вызов концепции биологического вида. Открытие того, что репродуктивные барьеры полупроницаемы для потока генов и что виды могут дифференцироваться, несмотря на продолжающееся скрещивание (Harrisson 1998; Rieseberg 2001; Wu 2001; Rieseberg et al. 2003; Coyne and Orr 2004; Wu and Ting 2004; Mallet 2005, 2008; Lexer) и Widmer 2008) указывает на то, что концепция биологических видов не соответствует нынешнему использованию термина «виды». Принятие этой концепции потребует объединения многих хорошо дифференцированных и общепринятых видов, которые, тем не менее, регулярно скрещиваются.Некоторые авторы (Орр, 2001; Нур, 2002; Койн, Орр, 2004) выступали за смягчение интерпретации концепции биологических видов, чтобы сохранить ее, несмотря на новые идеи. Однако смысл концепции вида состоит в том, что она определяет условия, при которых группа особей должна быть классифицирована как вид. Если игнорировать условия, указанные в концепции вида, она становится бесполезной.
Более того, концепция биологического вида изначально была сформулирована исключительно для двупародительских организмов.Принятие концепции биологических видов повлечет за собой, что термин виды не следует использовать для однопородных организмов, хотя растущие данные указывают на то, что они организованы в единицы, которые напоминают виды двупародительских организмов (Holman 1987; Gevers et al.2005; Cohan and Perry 2007 ; Fontaneto et al.2007; Cohan and Koeppel 2008). Было высказано предположение, что модификация концепции биологического вида может быть применена к монородительским прокариотам (Dykhuizen and Green 1991). Это было основано на понимании того, что прокариоты не являются простыми клональными организмами, но, по крайней мере, в некоторых группах, происходит частый обмен генами.Однако обмен генами между прокариотами не ограничивается близкородственными видами (Ochman et al. 2005), и, таким образом, применение модификации концепции биологических видов потребует объединения многих хорошо дифференцированных и общепринятых видов. Более того, существуют однопородные организмы без обмена генами, такие как партеногенетические виды, для которых не применима даже модификация концепции биологических видов.
Более того, существуют однопородные организмы без обмена генами, такие как партеногенетические виды, для которых не применима даже модификация концепции биологических видов.
КОНЦЕПЦИЯ ФИЛОГЕНЕТИЧЕСКИХ ВИДОВ
Одной из важных альтернатив концепции биологических видов, которая может принципиально применяться также и к однопородным организмам, является концепция филогенетических видов.Согласно диагностируемой версии концепции филогенетических видов, вид — это « диагностируемый кластер индивидов, внутри которого существует родительский образец происхождения и происхождения, вне которого нет, и который демонстрирует образец филогенетического происхождения и потомков среди подобные единицы »(Элдридж и Крафт, 1980). Эта концепция также была рекомендована для прокариот (Staley 2006) и недавно упоминалась в нескольких исследованиях штрих-кодирования ДНК / таксономии ДНК (например,г., Kelly et al. 2007; Саркар и др. 2008; Monaghan et al. 2009 г.).
2009 г.).
Однако с этим понятием есть проблемы как у однопородных, так и у двуногих. У монородителей, у которых обмен генами отсутствует или незначителен, каждый клон с мутацией будет классифицироваться как отдельный вид (Coyne and Orr 2004). У монородителей с более высокими уровнями обмена генами и у двойных родителей каждая замена будет иметь свое собственное конкретное распределение, и между наборами индивидуумов, у которых диагностированы независимо полученные мутации, может существовать небольшая или отсутствующая согласованность, за исключением тех, которые ограничены барьерами для потока генов (Avise and Ball 1990) .Элдридж и Крафт (1980) ограничили группы индивидуумов, которые классифицируются как «виды», включив в концепцию условие, «внутри которого существует родительский образец происхождения и происхождения, за которым нет». Это условие имеет функцию ограничить группу особей, которые классифицируются как «виды», теми, которые ограничены препятствиями для потока генов. Однако понимание того, что гибридизация между близкородственными видами происходит часто и что виды могут дифференцироваться, несмотря на продолжающееся скрещивание, означает, что многие признанные в настоящее время виды не демонстрируют отдельных «родительских моделей происхождения и происхождения», но что некоторые потомки, принадлежащие к одному виду, могут иметь предков. принадлежность к другому одновременно существующему виду и наоборот.Таким образом, эти идеи имеют такие же последствия для применения концепции филогенетических видов, как и для концепции биологических видов. Принятие концепции филогенетических видов также потребует объединения многих хорошо дифференцированных и общепринятых видов, которые, тем не менее, регулярно скрещиваются.
Однако понимание того, что гибридизация между близкородственными видами происходит часто и что виды могут дифференцироваться, несмотря на продолжающееся скрещивание, означает, что многие признанные в настоящее время виды не демонстрируют отдельных «родительских моделей происхождения и происхождения», но что некоторые потомки, принадлежащие к одному виду, могут иметь предков. принадлежность к другому одновременно существующему виду и наоборот.Таким образом, эти идеи имеют такие же последствия для применения концепции филогенетических видов, как и для концепции биологических видов. Принятие концепции филогенетических видов также потребует объединения многих хорошо дифференцированных и общепринятых видов, которые, тем не менее, регулярно скрещиваются.
Более того, полифилетические виды, происходящие в результате параллельного видообразования, также не будут демонстрировать «родительские образцы происхождения и происхождения», отличные от таковых у предковых видов, и должны быть также включены в понятие филогенетических видов.
ОПРЕДЕЛЕНИЕ ГЕНОТИПИЧЕСКОГО КЛАСТЕРА
Маллет (1995) признал, что концепция биологических видов несостоятельна перед лицом потока генов между независимо эволюционирующими единицами, и предложил концепцию видов, основанных на паттернах. Добавив генетику к концепции фенетических видов, которая определяет виды как группы индивидуумов с небольшим количеством промежуточных звеньев или без них, он сформулировал определение генотипического кластера, согласно которому вид — это «генотипический кластер , который может перекрываться, не сливаясь со своим братом ».”
Так же, как и концепция филогенетических видов, эта концепция в принципе применима и к однопородным организмам. Однако принятие определения генотипического кластера приведет к нежелательным последствиям, когда каждый генетически отличающийся клон будет идентифицирован как отдельный вид (Coyne and Orr 2004).
Предсказание о том, что виды рано или поздно образуют генотипические и фенотипические кластеры, может быть получено из большинства представлений о видах. Таким образом, это, несомненно, полезный критерий для определения границ условных видов. Однако зарождающиеся виды могут еще не распознаваться как отдельные кластеры на основе случайной выборки генетических маркеров. В случае перипатрического видообразования периферические виды сначала часто образуют кластер с соседними популяциями более широко распространенных видов, так что более распространенные виды не образуют генотипический кластер, отличный от периферических видов. По крайней мере, на начальных стадиях когерентные сущности, происходящие в результате параллельного видообразования из разных популяций предкового вида, не могут быть распознаны как отдельные генотипические кластеры и должны быть объединены с предковыми видами в соответствии с определением генотипического кластера.
Таким образом, это, несомненно, полезный критерий для определения границ условных видов. Однако зарождающиеся виды могут еще не распознаваться как отдельные кластеры на основе случайной выборки генетических маркеров. В случае перипатрического видообразования периферические виды сначала часто образуют кластер с соседними популяциями более широко распространенных видов, так что более распространенные виды не образуют генотипический кластер, отличный от периферических видов. По крайней мере, на начальных стадиях когерентные сущности, происходящие в результате параллельного видообразования из разных популяций предкового вида, не могут быть распознаны как отдельные генотипические кластеры и должны быть объединены с предковыми видами в соответствии с определением генотипического кластера.
КОНЦЕПЦИЯ ВИДОВ СПОСОБНОСТИ
Темплтон (1989) признал, что существует целый континуум репродуктивных систем от однопородных до двупородных и что любое априорное ограничение объема концепции вида в отношении режима воспроизводства является в некоторой степени искусственным. Кроме того, он признал, что эволюционные процессы, помимо репродуктивной изоляции, также способствуют формированию и поддержанию целостных образований, особенно у однопородных.Темплтон (1989) собрал эти процессы и классифицировал эти «механизмы сплоченности» на механизмы, влияющие на генетический обмен, определяя пределы распространения новых генетических вариантов через поток генов, и механизмы, влияющие на демографический обмен, определяя фундаментальную нишу и пределы распространения новых генетических вариантов. варианты через генетический дрейф и естественный отбор. Однако термин «механизм сплоченности» может вводить в заблуждение, потому что большинство биологических свойств, которые придают сплоченность, вероятно, возникли не для этой цели (Harrison 1998).Таким образом, сплоченность — это эффект, а не механизм. Основываясь на «механизмах сплоченности», Темплтон (1989) сформулировал концепцию видов сплоченности, согласно которой вид — это « наиболее инклюзивная группа организмов, обладающая потенциалом к генетическому и / или демографическому обмену ».
Кроме того, он признал, что эволюционные процессы, помимо репродуктивной изоляции, также способствуют формированию и поддержанию целостных образований, особенно у однопородных.Темплтон (1989) собрал эти процессы и классифицировал эти «механизмы сплоченности» на механизмы, влияющие на генетический обмен, определяя пределы распространения новых генетических вариантов через поток генов, и механизмы, влияющие на демографический обмен, определяя фундаментальную нишу и пределы распространения новых генетических вариантов. варианты через генетический дрейф и естественный отбор. Однако термин «механизм сплоченности» может вводить в заблуждение, потому что большинство биологических свойств, которые придают сплоченность, вероятно, возникли не для этой цели (Harrison 1998).Таким образом, сплоченность — это эффект, а не механизм. Основываясь на «механизмах сплоченности», Темплтон (1989) сформулировал концепцию видов сплоченности, согласно которой вид — это « наиболее инклюзивная группа организмов, обладающая потенциалом к генетическому и / или демографическому обмену ».
Койн и Орр (2004) утверждали, что концепция сплоченных видов может не дать решения о статусе видов двух групп, когда критерии генетической и демографической взаимозаменяемости противоречат друг другу, например, если две группы двупародительских особей генетически неизменяемы, но демографически обменный.Это не тот случай. «И / или» между генетической и демографической взаимозаменяемостью в формулировке концепции сплоченности Темплтона (1989) — это логическое «или», включающая дизъюнкция. Таким образом, все люди, связанные каким-либо «механизмом сплоченности», классифицируются как один вид в соответствии с концепцией сплоченности. Если есть, например, группы двух родительских особей, которые генетически не подлежат обмену (т. Е. Репродуктивно изолированы), но демографически обмениваются, наиболее инклюзивная группа включает всех индивидуумов.Таким образом, в рамках концепции сплоченности группы индивидов должны рассматриваться как особые, если они занимают одну и ту же фундаментальную нишу, даже если они репродуктивно изолированы. Принятие концепции сплоченности потребовало бы объединения многих общепринятых репродуктивно изолированных видов. Более того, концепция сплоченности не определяет, какой вид потока генов влияет на генетический обмен и, таким образом, также приведет к объединению видов, которые дифференцируются, несмотря на продолжающийся обмен генами.
Принятие концепции сплоченности потребовало бы объединения многих общепринятых репродуктивно изолированных видов. Более того, концепция сплоченности не определяет, какой вид потока генов влияет на генетический обмен и, таким образом, также приведет к объединению видов, которые дифференцируются, несмотря на продолжающийся обмен генами.
КОНЦЕПЦИЯ РОДОВЫХ ВИДОВ
Ву (2001) разработал новую модель видообразования, основанную на рассмотрении генетических процессов, происходящих во время видообразования, которая объясняет, почему и какой вид обмена генами не влияет на сохранение дифференцированных видов и дальнейшую дифференциацию видов.Согласно этой модели, весь процесс видообразования зависит в первую очередь от генов, ответственных за дифференциальную адаптацию к разным естественным или половым условиям, «генов видообразования» (см. Также Wu and Ting 2004). Ву (2001) определил дифференциальную адаптацию как форму дивергенции, при которой альтернативные аллели гена имеют противоположные эффекты приспособленности в двух группах людей. В процессе видообразования гены видообразования могут составлять лишь небольшую часть генома.Обмен генов ограничен в этих локусах, тогда как обмен генов в других локусах, «маркерных локусах», может сохраняться в течение длительного периода времени даже после видообразования. Видообразование — это стадия, на которой группы особей не теряют своего расхождения при контакте и могут продолжать расходиться. Основываясь на этой модели, Wu (2001) определил виды как « группы, которые дифференцированно адаптированы и при контакте не способны делиться генами, контролирующими эти адаптивные признаки, путем прямого обмена или через промежуточные гибридные популяции .«Хотя изначально она была сформулирована для двупородных видов, эта концепция в принципе применима и к монородителям.
В процессе видообразования гены видообразования могут составлять лишь небольшую часть генома.Обмен генов ограничен в этих локусах, тогда как обмен генов в других локусах, «маркерных локусах», может сохраняться в течение длительного периода времени даже после видообразования. Видообразование — это стадия, на которой группы особей не теряют своего расхождения при контакте и могут продолжать расходиться. Основываясь на этой модели, Wu (2001) определил виды как « группы, которые дифференцированно адаптированы и при контакте не способны делиться генами, контролирующими эти адаптивные признаки, путем прямого обмена или через промежуточные гибридные популяции .«Хотя изначально она была сформулирована для двупородных видов, эта концепция в принципе применима и к монородителям.
Концепция генных видов Wu (2001) подверглась критике, поскольку она сосредоточена исключительно на дифференциальной адаптации, вызванной мутациями в генах (Britton-Davidian 2001; Orr 2001; Rundle et al. 2001; Noor 2002). Другие генетические особенности, которые могут привести к репродуктивной изоляции, такие как хромосомные изменения, были классифицированы Wu (2001) как «особые случаи».Однако критики предпочитают классифицировать сущности, которые развиваются отдельно, потому что они репродуктивно изолированы из-за хромосомных изменений или других негенных мутаций, как отдельные виды. Даже если дифференциальная адаптация будет наиболее частым процессом, приводящим к видообразованию, другие процессы, которые также могут вызывать образование видов, такие как генетический дрейф, не должны быть исключены априори в концепции вида (Britton-Davidian 2001; Orr 2001; Rundle et al. 2001; Нур 2002).
2001; Noor 2002). Другие генетические особенности, которые могут привести к репродуктивной изоляции, такие как хромосомные изменения, были классифицированы Wu (2001) как «особые случаи».Однако критики предпочитают классифицировать сущности, которые развиваются отдельно, потому что они репродуктивно изолированы из-за хромосомных изменений или других негенных мутаций, как отдельные виды. Даже если дифференциальная адаптация будет наиболее частым процессом, приводящим к видообразованию, другие процессы, которые также могут вызывать образование видов, такие как генетический дрейф, не должны быть исключены априори в концепции вида (Britton-Davidian 2001; Orr 2001; Rundle et al. 2001; Нур 2002).
КОНЦЕПЦИЯ ДИФФЕРЕНЦИАЛЬНЫХ ВИДОВ ФИТНЕСА
По сравнению с другими концепциями видов, способность концепции генных видов У (2001) указывать, в каких случаях группы особей могут быть классифицированы как виды, несмотря на поток генов между этими группами, представляет собой важный концептуальный прогресс. Таким образом, стоит задуматься о том, можно ли отменить два критикуемых ограничения концепции генных видов, согласно которым в ней рассматриваются только виды, возникшие в результате (1) мутаций в генах, приводящих к (2) дифференциальной адаптации. Ву (2001) определил дифференциальную адаптацию как «противоположные эффекты приспособленности» альтернативных аллелей. Однако признаки, вызывающие видообразование, не обязательно должны быть альтернативными аллелями, и их эффекты приспособленности не обязательно должны быть противоположными у разных видов, что положительно для видов, которые они характеризуют, и отрицательно для других видов.Чтобы ограничить поток генов, достаточно, чтобы такие особенности, которые могут быть наследственными признаками, отличными от генов, имели негативные эффекты приспособляемости у других видов. Они могут быть нейтральными по отношению к описываемым ими видам. Например, инфекция Wolbachia может быть почти нейтральной для вида-хозяина, но иметь отрицательный эффект приспособленности для других, неинфицированных видов.
Таким образом, стоит задуматься о том, можно ли отменить два критикуемых ограничения концепции генных видов, согласно которым в ней рассматриваются только виды, возникшие в результате (1) мутаций в генах, приводящих к (2) дифференциальной адаптации. Ву (2001) определил дифференциальную адаптацию как «противоположные эффекты приспособленности» альтернативных аллелей. Однако признаки, вызывающие видообразование, не обязательно должны быть альтернативными аллелями, и их эффекты приспособленности не обязательно должны быть противоположными у разных видов, что положительно для видов, которые они характеризуют, и отрицательно для других видов.Чтобы ограничить поток генов, достаточно, чтобы такие особенности, которые могут быть наследственными признаками, отличными от генов, имели негативные эффекты приспособляемости у других видов. Они могут быть нейтральными по отношению к описываемым ими видам. Например, инфекция Wolbachia может быть почти нейтральной для вида-хозяина, но иметь отрицательный эффект приспособленности для других, неинфицированных видов. Группы людей становятся эволюционно независимыми только в том случае, если поток генов ограничен в обоих направлениях. Таким образом, каждая группа должна иметь особенности, которые отрицательно влияют на приспособленность других видов.Основываясь на этих идеях, виды можно определить как группу особей, которые взаимно характеризуются чертами, которые могут иметь отрицательные эффекты приспособленности в других группах и которыми нельзя регулярно обмениваться между группами при контакте .
Группы людей становятся эволюционно независимыми только в том случае, если поток генов ограничен в обоих направлениях. Таким образом, каждая группа должна иметь особенности, которые отрицательно влияют на приспособленность других видов.Основываясь на этих идеях, виды можно определить как группу особей, которые взаимно характеризуются чертами, которые могут иметь отрицательные эффекты приспособленности в других группах и которыми нельзя регулярно обмениваться между группами при контакте .
Эта концепция различных видов учитывает не только мутации в генах, но и любые различия, включая, например, хромосомные изменения (White 1954; Rieseberg 2001), инфекции Wolbachia (Hurst and Schilthuizen 1998; Telschow et al.2005; Werren et al. 2008), другие эгоистичные генетические элементы, такие как мобильные элементы (Hurst and Schilthuizen 1998; Hurst and Werren 2001) или определяющие нишу гены или наборы генов, приобретенные путем горизонтального переноса у прокариот (Ochman et al. 2005; Cohan and Koeppel 2008). Различия могут быть результатом дифференциальной адаптации из-за естественного или полового отбора, но также могут быть результатом генетического дрейфа или других часто неадаптивных процессов, таких как полиплоидизация или заражение симбионтами.Признаки могут быть специфически адаптивными к нише одной группы, но неадекватными для другой группы или могут быть несовместимы в контексте генетического фона другой группы (например, различные инфекции Wolbachia ). Такая несовместимость может привести к снижению фертильности или несбалансированной физиологической функции. Как и концепция генных видов, концепция дифференциальной приспособленности допускает обмен генами, поскольку они не важны для свойств, которые имеют отрицательные эффекты приспособленности у других видов.
2005; Cohan and Koeppel 2008). Различия могут быть результатом дифференциальной адаптации из-за естественного или полового отбора, но также могут быть результатом генетического дрейфа или других часто неадаптивных процессов, таких как полиплоидизация или заражение симбионтами.Признаки могут быть специфически адаптивными к нише одной группы, но неадекватными для другой группы или могут быть несовместимы в контексте генетического фона другой группы (например, различные инфекции Wolbachia ). Такая несовместимость может привести к снижению фертильности или несбалансированной физиологической функции. Как и концепция генных видов, концепция дифференциальной приспособленности допускает обмен генами, поскольку они не важны для свойств, которые имеют отрицательные эффекты приспособленности у других видов.
Концепция видов с дифференциальной приспособленностью отличается от концепции биологических видов тем, что считается, что обмен видоспецифическими особенностями может быть ограничен не только репродуктивной изоляцией, но также и дивергентным отбором. В этом отношении концепция дифференциальной приспособленности видов ближе к пониманию видов Дарвином (1859), чем к концепции биологических видов. Дарвин (1859: 485) утверждал, что «единственное различие между видами и хорошо выраженными разновидностями состоит в том, что последние, как известно, или, как полагают, в настоящее время связаны промежуточными градациями, тогда как раньше виды были связаны таким образом.Отсутствие промежуточных градаций в первых различиях, характеризующих возникающие виды, является следствием неспособности обмениваться этими признаками между группами и не обязательно связано с репродуктивной изоляцией. Одним из недостатков чисто фенотипической оценки способности обмениваться потенциально видоспецифическими признаками является то, что группы особей, характеризующиеся разными дискретными полиморфизмами, могут быть приняты за отдельные виды из-за отсутствия промежуточных градаций.Однако это не влияет на концепцию различных видов приспособленности, которая не сосредотачивается на отсутствии промежуточных градаций как таковых, а основана на неспособности обмена видоспецифическими особенностями между группами.
В этом отношении концепция дифференциальной приспособленности видов ближе к пониманию видов Дарвином (1859), чем к концепции биологических видов. Дарвин (1859: 485) утверждал, что «единственное различие между видами и хорошо выраженными разновидностями состоит в том, что последние, как известно, или, как полагают, в настоящее время связаны промежуточными градациями, тогда как раньше виды были связаны таким образом.Отсутствие промежуточных градаций в первых различиях, характеризующих возникающие виды, является следствием неспособности обмениваться этими признаками между группами и не обязательно связано с репродуктивной изоляцией. Одним из недостатков чисто фенотипической оценки способности обмениваться потенциально видоспецифическими признаками является то, что группы особей, характеризующиеся разными дискретными полиморфизмами, могут быть приняты за отдельные виды из-за отсутствия промежуточных градаций.Однако это не влияет на концепцию различных видов приспособленности, которая не сосредотачивается на отсутствии промежуточных градаций как таковых, а основана на неспособности обмена видоспецифическими особенностями между группами. Пока группы особей, характеризующиеся разными дискретными полиморфизмами, способны обмениваться этими полиморфизмами, то есть лежащими в основе генами, эти группы не считаются видами в соответствии с концепцией видов с дифференциальной приспособленностью. Но когда эта способность исчезает, группы индивидуумов, характеризующиеся разными чертами, становятся видами в соответствии с этой концепцией.
Пока группы особей, характеризующиеся разными дискретными полиморфизмами, способны обмениваться этими полиморфизмами, то есть лежащими в основе генами, эти группы не считаются видами в соответствии с концепцией видов с дифференциальной приспособленностью. Но когда эта способность исчезает, группы индивидуумов, характеризующиеся разными чертами, становятся видами в соответствии с этой концепцией.
Концепция видов с дифференциальной приспособленностью может применяться ко всему спектру организмов от однокоренных до двуногих. Концепция видов с дифференциальной приспособленностью классифицирует группы как виды, если они характеризуются характеристиками, которые могут иметь отрицательные эффекты приспособленности в других таких группах и которыми нельзя регулярно обмениваться между этими группами. У двойных родителей эти особенности включают в себя, помимо адаптации к разным условиям окружающей среды, также модификации репродуктивной системы, которые определяют, с какими другими особями индивидуум может производить потомков, способных к зачатию. Например, полиплоидия имеет отрицательный эффект приспособленности в группе диплоидных особей, потому что это может привести к репродуктивной несовместимости. Точно так же диплоидия отрицательно влияет на приспособленность группы полиплоидных индивидуумов. Таким образом, эти группы классифицируются как разные виды в соответствии с концепцией дифференциальной приспособленности, даже если они занимают одну и ту же нишу. Однородительные группы, характеризующиеся особенностями, которые могут иметь негативное влияние на приспособленность других групп, в большинстве случаев занимают другие ниши.Таким образом, у однопородных видов, как определено концепцией дифференциальной приспособленности, обычно соответствуют экотипы, которые фактически считались эквивалентами двупородных видов прокариот (Gevers et al. 2005; Cohan and Perry 2007; Cohan and Koeppel 2008). Однако мутации в разных генах в разных штаммах, каждый из которых сам по себе избирательно нейтрален, могут приводить к генной несовместимости, то есть могут иметь отрицательные эффекты приспособленности в штаммах с несовместимыми мутациями в других генах, так что такие штаммы квалифицируются как разные.
Например, полиплоидия имеет отрицательный эффект приспособленности в группе диплоидных особей, потому что это может привести к репродуктивной несовместимости. Точно так же диплоидия отрицательно влияет на приспособленность группы полиплоидных индивидуумов. Таким образом, эти группы классифицируются как разные виды в соответствии с концепцией дифференциальной приспособленности, даже если они занимают одну и ту же нишу. Однородительные группы, характеризующиеся особенностями, которые могут иметь негативное влияние на приспособленность других групп, в большинстве случаев занимают другие ниши.Таким образом, у однопородных видов, как определено концепцией дифференциальной приспособленности, обычно соответствуют экотипы, которые фактически считались эквивалентами двупородных видов прокариот (Gevers et al. 2005; Cohan and Perry 2007; Cohan and Koeppel 2008). Однако мутации в разных генах в разных штаммах, каждый из которых сам по себе избирательно нейтрален, могут приводить к генной несовместимости, то есть могут иметь отрицательные эффекты приспособленности в штаммах с несовместимыми мутациями в других генах, так что такие штаммы квалифицируются как разные. виды согласно концепции дифференциальной приспособленности, даже если они не являются экологически дифференцированными.Сомнительно, что нейтральные мутации часто сохраняются у прокариот достаточно долго, чтобы могла возникнуть такая генная несовместимость, или же они обычно устраняются периодическим отбором до того, как может возникнуть генная несовместимость.
виды согласно концепции дифференциальной приспособленности, даже если они не являются экологически дифференцированными.Сомнительно, что нейтральные мутации часто сохраняются у прокариот достаточно долго, чтобы могла возникнуть такая генная несовместимость, или же они обычно устраняются периодическим отбором до того, как может возникнуть генная несовместимость.
Сравнение представлений о видах
Для облегчения сравнения последствий, вытекающих из различных концепций видов, в таблице 1 перечислены некоторые иллюстративные группировки и их классификация в рамках различных концепций видов.Общая концепция происхождения не могла быть рассмотрена из-за проблем, рассмотренных выше.
Таблица 1. Классификация групп особей как видов (+), обычно видов (±) или не отдельных видов (-) с помощью различных видовых концепций (н.о., концепция неприменима). BSC = концепция биологических видов; PSC = концепция филогенетических видов; GCD = определение генотипического кластера; CSC = видовая концепция сплоченности; GSC = концепция генных видов; DFSC = концепция различных видов приспособленности.
| BSC | PSC | GCD | CSC | GSC | DFSC | |
|---|---|---|---|---|---|---|
| Группы однородных родителей, для которых характерны особенности, которые могут иметь отрицательный эффект приспособленности в других группах | на | + | ± | + | + | + |
| Поддающиеся диагностике группы, которые не характеризуются признаками, которыми нельзя обмениваться с другими группами, но не обмениваются генами с другими группами из-за географической изоляции | ||||||
| Поддающиеся диагностике группы однородных родителей, для которых не характерны черты, которые могли бы иметь отрицательные последствия для пригодности в других группах | на | + | ± | – | – | – |
| Группы двойных родителей, репродуктивно изолированные, но занимающие одну и ту же нишу | ||||||
| Дифференциально адаптированные группы, не способные разделять гены, контролирующие эти адаптивные признаки, хотя они скрещиваются между собой | – | – | ± | – | + | + |
| Группы двойных родителей, репродуктивно изолированные из-за негенных мутаций или процессов, отличных от дифференциальной адаптации |
Концепция биологического вида отличается от других обсуждаемых концепций тем, что она сформулирована исключительно для двупородных родителей и требует внутренней репродуктивной изоляции. Филогенетическая концепция видов отличается от других концепций тем, что классифицирует группы, которые изолированы только внешне (например, географически), как виды. Это приводит к значительной таксономической инфляции (Agapow et al. 2004; Isaac et al. 2004). Более того, он классифицирует также диагностируемые группы монородителей, которые не характеризуются особенностями, которые могли бы иметь отрицательные эффекты приспособленности в других группах, как виды, что привело бы к еще большей инфляции в таксономии прокариот. Такие группы часто можно классифицировать как виды, также используя определение генотипического кластера.Что касается классификации других групп, определение генотипического кластера наиболее похоже на концепцию видов с дифференциальной приспособленностью. Однако его результат не может быть однозначно предсказан на основе условий, указанных в таблице 1, поскольку ожидается, что генотипические кластеры возникнут при всех указанных условиях, но могут появиться только на более поздних стадиях дифференцировки.
Филогенетическая концепция видов отличается от других концепций тем, что классифицирует группы, которые изолированы только внешне (например, географически), как виды. Это приводит к значительной таксономической инфляции (Agapow et al. 2004; Isaac et al. 2004). Более того, он классифицирует также диагностируемые группы монородителей, которые не характеризуются особенностями, которые могли бы иметь отрицательные эффекты приспособленности в других группах, как виды, что привело бы к еще большей инфляции в таксономии прокариот. Такие группы часто можно классифицировать как виды, также используя определение генотипического кластера.Что касается классификации других групп, определение генотипического кластера наиболее похоже на концепцию видов с дифференциальной приспособленностью. Однако его результат не может быть однозначно предсказан на основе условий, указанных в таблице 1, поскольку ожидается, что генотипические кластеры возникнут при всех указанных условиях, но могут появиться только на более поздних стадиях дифференцировки. Обратите внимание, что генотипические кластеры могут возникать также только в изолированных внешне группах, но такие кластеры не квалифицируются как виды в соответствии с определением генотипического кластера, потому что они будут сливаться при контакте.Самая необычная особенность концепции сплоченности состоит в том, что группы двух родительских особей считаются конвидовыми, если они занимают одну и ту же фундаментальную нишу, даже если они репродуктивно изолированы. Концептуальный прогресс концепций видов генетической и дифференциальной приспособленности состоит в том, что они не требуют полной репродуктивной изоляции, но допускают обмен чертами, за исключением тех, которые являются причинными для дифференциации. Группы, которые классифицируются как виды в соответствии с концепцией генных видов, также считаются видами в рамках более общей концепции видов с дифференциальной приспособленностью.То же самое верно для групп, которые классифицируются как виды в соответствии с концепцией биологического вида, потому что особенности, которые вызывают репродуктивную изоляцию у двупородных родителей, имеют негативные эффекты приспособленности в других группах и не могут регулярно обмениваться.
Обратите внимание, что генотипические кластеры могут возникать также только в изолированных внешне группах, но такие кластеры не квалифицируются как виды в соответствии с определением генотипического кластера, потому что они будут сливаться при контакте.Самая необычная особенность концепции сплоченности состоит в том, что группы двух родительских особей считаются конвидовыми, если они занимают одну и ту же фундаментальную нишу, даже если они репродуктивно изолированы. Концептуальный прогресс концепций видов генетической и дифференциальной приспособленности состоит в том, что они не требуют полной репродуктивной изоляции, но допускают обмен чертами, за исключением тех, которые являются причинными для дифференциации. Группы, которые классифицируются как виды в соответствии с концепцией генных видов, также считаются видами в рамках более общей концепции видов с дифференциальной приспособленностью.То же самое верно для групп, которые классифицируются как виды в соответствии с концепцией биологического вида, потому что особенности, которые вызывают репродуктивную изоляцию у двупородных родителей, имеют негативные эффекты приспособленности в других группах и не могут регулярно обмениваться.
Концепция вида и определение границ
Формулирование концепции вида и разработка подходов к определению границ видов — две разные задачи (de Queiroz 2005b, 2007). Концепция вида должна быть сформулирована таким образом, чтобы охватывались также самые ранние стадии после видообразования.Собственно, понятие вида определяет, что такое видообразование. Самые ранние стадии после видообразования могут отличаться только некоторыми свойствами. После дивергенции дифференциация образовавшихся видов продолжается, так что становится легче различать более поздние стадии.
Подходы к определению границ видов в идеале должны основываться на условиях, указанных в концепции благоприятных видов. Тогда отграниченные виды должны точно соответствовать видовой концепции. Однако часто свойства, приобретенные позже во время дифференциации, легче наблюдать, чем те немногие свойства, которые приобретаются непосредственно на стадии видообразования. Таким образом, подходы к разграничению часто основываются на таких свойствах. Если подход к определению границ видов основан на свойствах, которые были приобретены на более поздних стадиях, этот подход может привести к ложноотрицательным результатам, то есть к видам на ранних стадиях видообразования, которые не распознаются как таковые, но не к ложноположительным. Если свойства, на которых основан подход, могут быть приобретены также до спецификации, также могут возникнуть ложные срабатывания. Поскольку методы, которые не основаны непосредственно на условиях, указанных в концепции вида, иногда не могут правильно определить границы видов, необходим эклектичный подход к разграничению видов (Sites and Marshall 2004).
Таким образом, подходы к разграничению часто основываются на таких свойствах. Если подход к определению границ видов основан на свойствах, которые были приобретены на более поздних стадиях, этот подход может привести к ложноотрицательным результатам, то есть к видам на ранних стадиях видообразования, которые не распознаются как таковые, но не к ложноположительным. Если свойства, на которых основан подход, могут быть приобретены также до спецификации, также могут возникнуть ложные срабатывания. Поскольку методы, которые не основаны непосредственно на условиях, указанных в концепции вида, иногда не могут правильно определить границы видов, необходим эклектичный подход к разграничению видов (Sites and Marshall 2004).
Если разграничение видов должно основываться непосредственно на условиях, указанных в концепции дифференциальной приспособленности, необходимо продемонстрировать, что группы особей взаимно характеризуются признаками, которые (1) имеют отрицательные эффекты приспособленности в других группах и что (2) не могут быть регулярно обмениваются между группами при контакте. Во многих случаях особенности, которые могут иметь негативное влияние на приспособленность других групп, например, разные размеры или формы клюва у птиц, позволяющие особям, принадлежащим к разным видам, использовать разную пищу, копулятивные органы различного образования, которые препятствуют скрещиванию между особями разных видов, или разные формы клюва. уровни плоидности известны и уже обычно используются для определения границ видов.Прерывистое изменение таких характеристик обычно указывает на невозможность регулярного обмена ими между группами. Таким образом, такие группы можно рассматривать как виды в рамках концепции дифференциальных видов. Более подробные проверки условий, указанных в концепции дифференциальной приспособленности, будут возможны только в меньшинстве случаев, когда генетика хорошо изучена, так же как были проведены явные проверки условий, указанных, например, в концепции биологического вида. только в меньшинстве случаев.Проверка того, что виды характеризуются признаками, которые «будут» иметь отрицательные эффекты приспособленности в других группах, может быть трудной, особенно в однопородных группах.
Во многих случаях особенности, которые могут иметь негативное влияние на приспособленность других групп, например, разные размеры или формы клюва у птиц, позволяющие особям, принадлежащим к разным видам, использовать разную пищу, копулятивные органы различного образования, которые препятствуют скрещиванию между особями разных видов, или разные формы клюва. уровни плоидности известны и уже обычно используются для определения границ видов.Прерывистое изменение таких характеристик обычно указывает на невозможность регулярного обмена ими между группами. Таким образом, такие группы можно рассматривать как виды в рамках концепции дифференциальных видов. Более подробные проверки условий, указанных в концепции дифференциальной приспособленности, будут возможны только в меньшинстве случаев, когда генетика хорошо изучена, так же как были проведены явные проверки условий, указанных, например, в концепции биологического вида. только в меньшинстве случаев.Проверка того, что виды характеризуются признаками, которые «будут» иметь отрицательные эффекты приспособленности в других группах, может быть трудной, особенно в однопородных группах. Однако это не дисквалифицирует концепцию как таковую. В принципе, эффекты приспособленности генов других видов могут быть проверены экспериментальной трансформацией также у монородителей.
Однако это не дисквалифицирует концепцию как таковую. В принципе, эффекты приспособленности генов других видов могут быть проверены экспериментальной трансформацией также у монородителей.
Если ничего не известно об эффектах приспособленности признаков и их возможности обмена, стандартный подход к разграничению видов — определение фенотипических или генотипических кластеров (Сайты и Маршалл 2004; Хаусдорф и Хенниг 2010).Такой подход оправдан, по крайней мере, для двупородных видов нашим пониманием процессов дифференциации. После видообразования все больше и больше аллелей ограничиваются одним из видов-потомков, так что особи, принадлежащие к одному виду, рано или поздно сформируют генотипические и фенотипические кластеры, даже если они изначально были парафилетическими (например, в результате перипатрического видообразования) или полифилетическими. (например, из-за параллельного видообразования).
Заместитель редактора: Д. Фэйрбэрн
БЛАГОДАРНОСТИ
Благодарю Р.
