
Эукариотическая клетка. Ядро. Биология 9 класс Мамонтов
Вопрос 1. Опишите строение ядра эукариотической клетки.
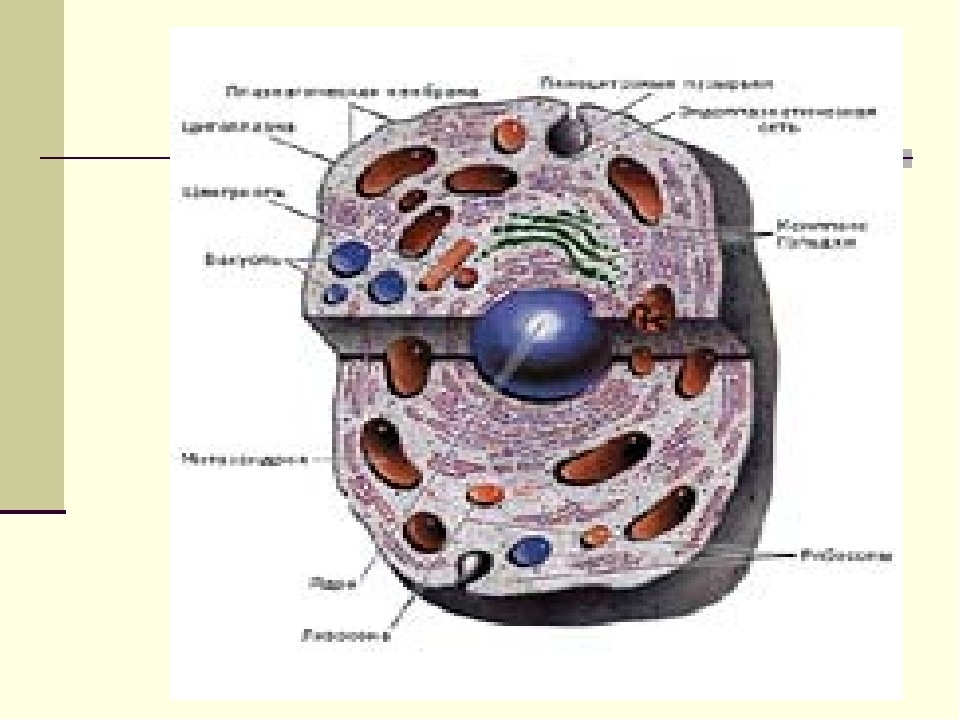
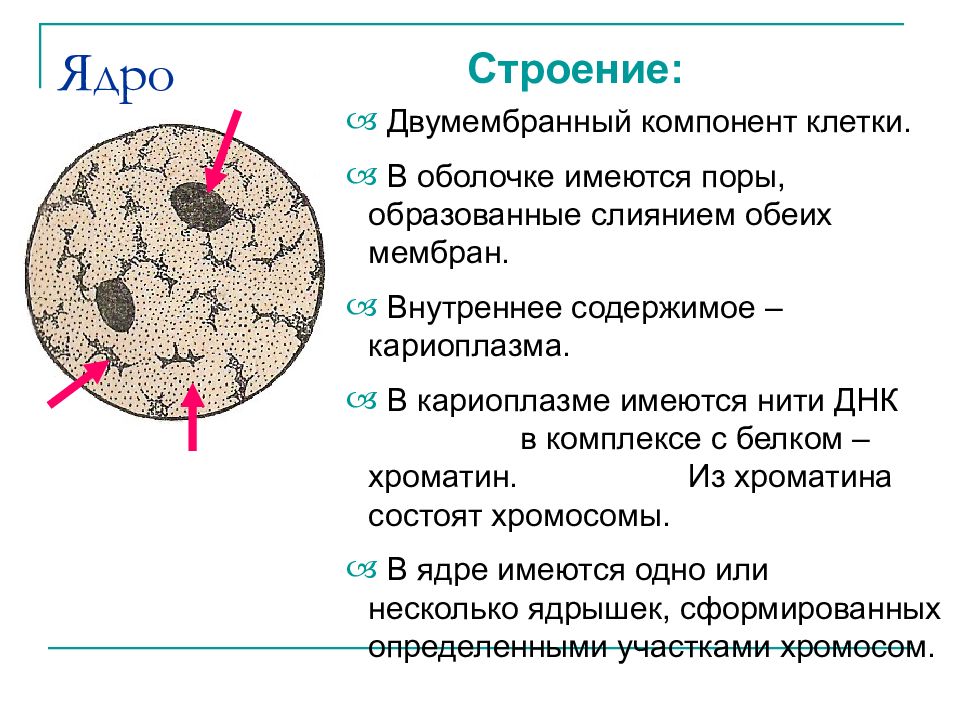
Ядро окружено оболочкой, которая состоит из двух мембран. Ядерная мембрана со стороны, обращённой в цитоплазму, покрыта рибосомами, внутренняя мембрана ядра гладкая. Ядерная оболочка – часть мембранной системы клетки. Выросты внешней ядерной мембраны соединяются с каналами эндоплазматической сети, образуя единую систему сообщающихся каналов. Между ядром и цитоплазмой осуществляется постоянный обмен веществами.
Несмотря на активный обмен между ядром и цитоплазмой, ядерная оболочка отграничивает ядерное содержимое от цитоплазмы, обеспечивая тем самым различия в их химическом составе. Это необходимо для нормального функционирования ядерных структур.
В гелеобразном ядерном соке располагаются хроматин и одно или несколько ядрышек.
В живой клетке ядерный сок выглядит бесструктурной массой, заполняющей промежутки между структурами ядра.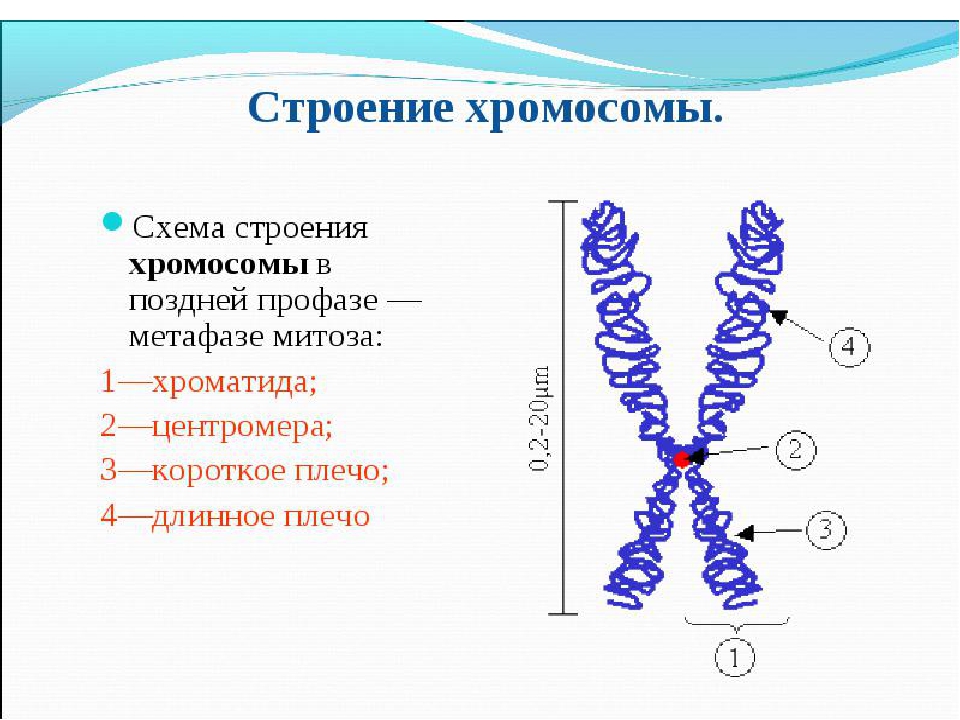
Вопрос 2. Что такое ядрышко? Как вы считаете, можно ли ядрышко выделить из ядра как самостоятельную единую структуру? Объясните свою точку зрения.
Ядрышко – структура, составленная из расположенных рядом участков нескольких различных хромосом. Эти участки представляют собой большие петли ДНК, содержащие гены рибосомальной РНК (рРНК). Такие петли называются ядрышковым организатором.
Ядрышко — не самостоятельная структура или органоид. Оно — производное
хромосомы, один из ее локусов, активно функционирующий в интерфазе.
Ядрышко является центром образования рибосом, т.к. здесь осуществляется синтез рРНК и соединение этих молекул с белками, т.е. происходит формирование субъединиц рибосом, которые затем поступают в цитоплазму, где и завершается сборка рибосом.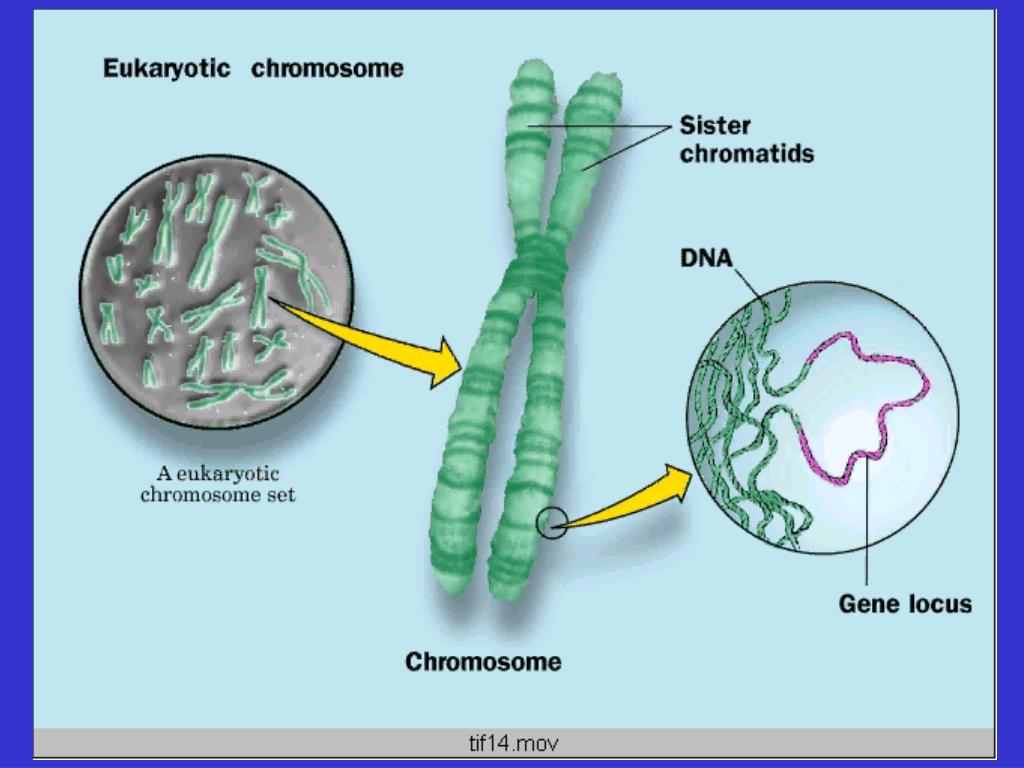
Вопрос 3. Что такое хроматин? Опишите строение и состав хромосомы.
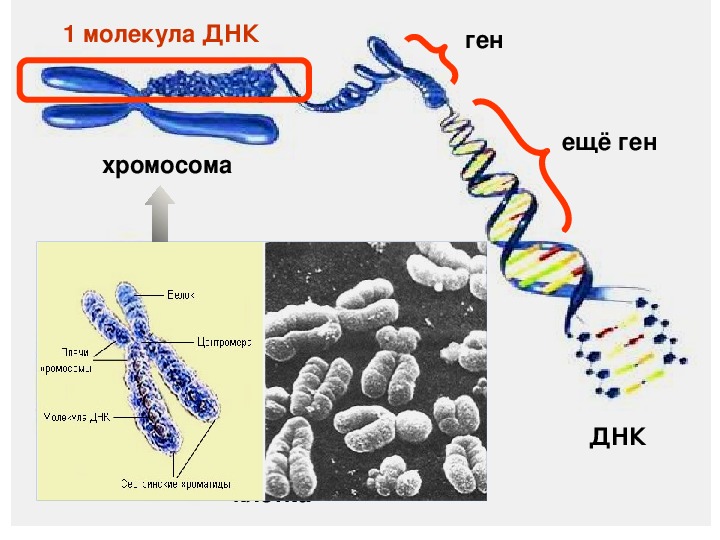
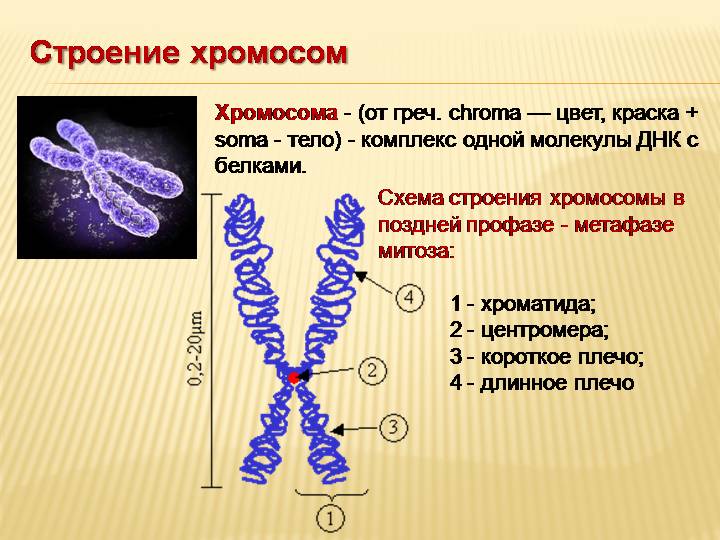
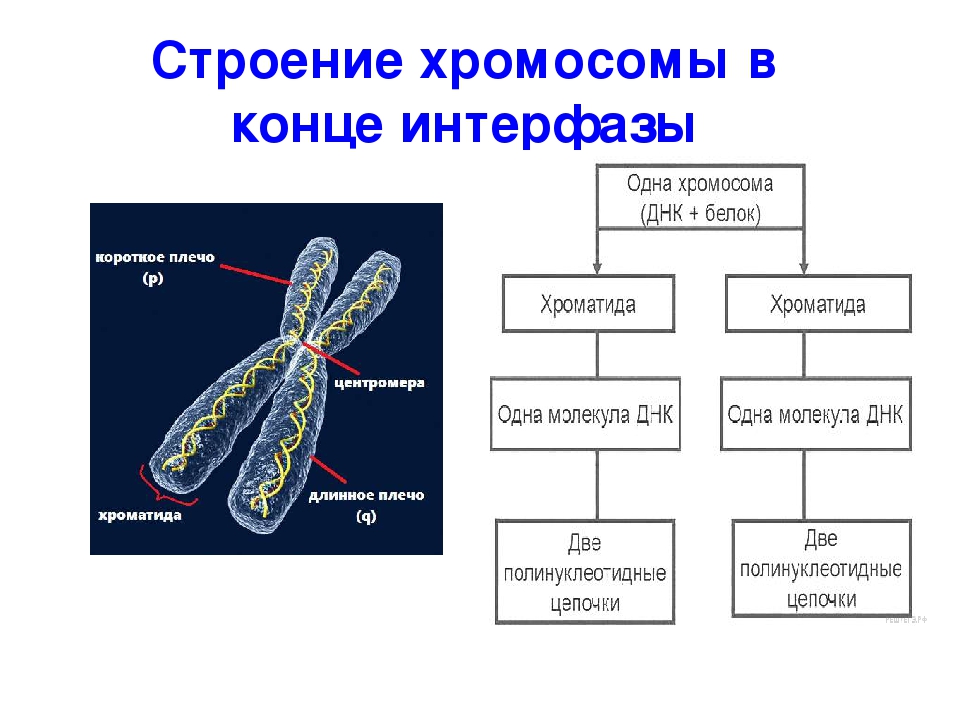
Хроматином (от греч. хрома – окраска, цвет) называют комплекс ДНК и белков, интенсивно окрашивающийся некоторыми красителями и отличающийся по форме от ядрышка. В делящихся клетках молекулы ДНК сильно спирализуются, укорачиваются и приобретают компактные размеры и форму. Такое компактное состояние ДНК называют хромосомами.
Хромосомы – органоиды клеточного ядра, совокупность которых определяет основные наследственные свойства клеток и организмов. Полный набор хромосом в клетке, характерный для данного организма, называется кариотипом. В любой клетке тела большинства животных и растений каждая хромосома представлена дважды: одна из них получена от отца, другая – от матери при слиянии ядер половых клеток в процессе оплодотворения. Такие хромосомы называются гомологичными, набор гомологичных хромосом – диплоидным.
Форма хромосом зависит от положения так называемой первичной перетяжки, или центромеры, – области, к которой во время деления клетки (митоза) прикрепляются нити веретена деления. Центромера делит хромосому на два плеча, которые могут быть одинаковой или разной длины.
Центромера делит хромосому на два плеча, которые могут быть одинаковой или разной длины.
Вопрос 4. Как соотносится число хромосом в соматических и половых клетках? Почему число хромосом в половых клетках должно быть вдвое меньше, чем в соматических?
Число хромосом в кариотипе большинства видов живых организмов чётное. Это объясняется тем, что в каждой соматической клетке находятся две одинаковые по форме и размеру хромосомы: одна – из отцовского организма, вторая – из материнского.
Хромосомы, одинаковые по форме и размеру и несущие одинаковые гены, называют гомологичными. Хромосомный набор соматической клетки, в котором каждая хромосома имеет себе пару, носит название двойного (или диплоидного) и обозначается 2n. Из каждой пары гомологичных хромосом в половые клетки попадает только одна хромосома, поэтому хромосомный набор гамет называют одинарным (или гаплоидным) и обозначают 1n.
Вопрос 5.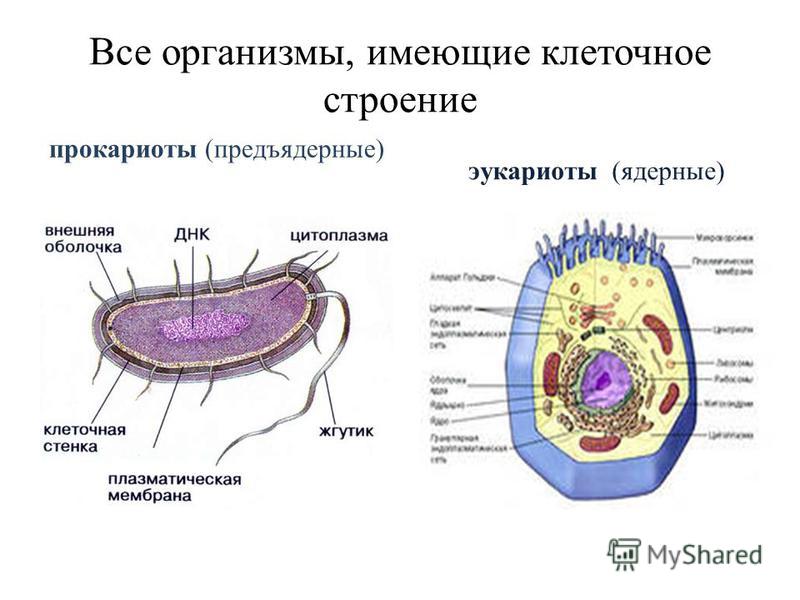 Какие хромосомы называют гомологичными?
Какие хромосомы называют гомологичными?
Хромосомы, одинаковые по форме и размеру и несущие одинаковые гены, называют гомологичными.
Вопрос 6. Что такое кариотип?
Совокупность количественных (число и размеры) и качественных (форма) признаков хромосомного набора соматической клетки называют кариотипом.
Вопрос 7. Вспомните строение ДНК бактерий. Выберите критерии и сравните наследственный материал про- и эукариотических клеток.
Строение ДНК бактерий аналогично таковому клеток эукариотического типа (растений, животных, грибов). В отличие от бактерий у вирусов геном представлен одной нуклеиновой кислотой – ДНК или РНК. Бактериальные клетки, кроме ДНК, могут иметь генетически полноценные образования функционирующие автономно. Необходимо подчеркнуть, что носителями наследственности бактерий кроме ДНК являются плазмиды и эписомы. В этой связи, любая структура бактериальной клетки, способна к саморепликации.
Бактериальная хромосома представлена одной двунитевой молекулой ДНК кольцевидной формы и называется нуклеотидом. Длина нуклеотида в растянутом виде составляет примерно 1 мм. Нуклеотид – эквивалент ядра. Расположен он в центре бактерии. В отличие от эукариот ядро бактерий не имеет ядерной оболочки, ядрышка и основных белков (гистонов).
Геномы прокариот и эукариот, хотя и имеют определенное сходство, но все же существенно различаются по своей структуре. Геномы прокариот практически целиком состоят из генов и регуляторных последовательностей. В генах прокариот нет интронов. Часто функционально родственные гены прокариот находятся под единым транскрипционным контролем, то есть транскрибируются вместе, составляя оперон.
Геномы эукариот существенно больше геномов бактерий, у дрожжей примерно в 2 раза, а у человека – на три порядка, то есть в тысячу раз. Однако прямой зависимости между количеством ДНК и эволюционной сложностью видов не наблюдается.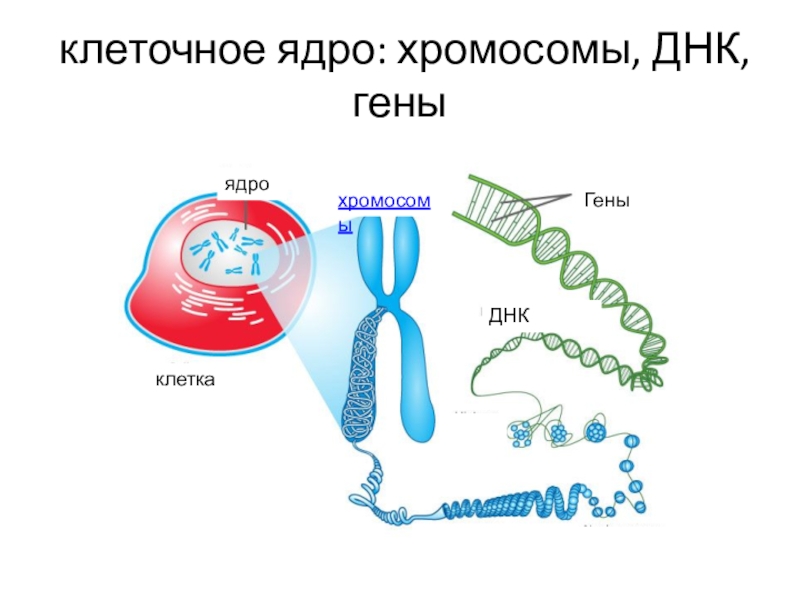 Достаточно сказать, что геномы некоторых видов амфибий или растений в десять или даже в сто раз превосходят по размеру геном человека. В некоторых случаях близкие виды организмов могут существенно различаться по количеству ДНК. Важным обстоятельством является то, что при переходе от прокариот к эукариотам увеличение генома происходит, главным образом, за счет появления огромного количества некодирующих последовательностей. Действительно, в геноме человека кодирующие области, то есть экзоны, суммарно занимают не более 3%, а по некоторым оценкам около 1% от общей длины ДНК.
Достаточно сказать, что геномы некоторых видов амфибий или растений в десять или даже в сто раз превосходят по размеру геном человека. В некоторых случаях близкие виды организмов могут существенно различаться по количеству ДНК. Важным обстоятельством является то, что при переходе от прокариот к эукариотам увеличение генома происходит, главным образом, за счет появления огромного количества некодирующих последовательностей. Действительно, в геноме человека кодирующие области, то есть экзоны, суммарно занимают не более 3%, а по некоторым оценкам около 1% от общей длины ДНК.
Вопрос 8. Используя рисунок 14, расскажите, как осуществляется обмен веществами между ядром и цитоплазмой.
Обмен веществ между ядром и цитоплазмой осуществляется двумя путями. Во-первых, ядерная оболочка пронизана многочисленными порами, через которые происходит обмен молекулами между ядром и цитоплазмой. Во-вторых, поступление веществ из ядра в цитоплазму и обратно может происходить в результате отделения выростов и впячиваний ядерной оболочки.
Пути обмена веществ между ядром и цитоплазмой.
1 — обмен веществ через ядерные поры,
2 — впячивание цитоплазмы внутрь ядра,
3 — впячивание ядерной оболочки,
4 — продвижение ядерной мембраны в эндоплазматическую сеть;
5 — выведение части каналов во внешнее межклеточное пространство.
Вопрос 9. Используя дополнительные источники информации, приведите примеры числа хромосом у разных видов живых организмов. Сделайте вывод, зависит ли степень сложности организации вида от числа хромосом.
Человек Homo sapiens 46
Гориллы Gorilla 48
Волк Canis lupus 78
Кошка Felis catus 38
Осёл Equus asinus 62
Ананас Ananas comosus 50
Картофель Solanum tuberosum 48
Комар Aedes aegypti 6
Наименьшее число хромосом: самки подвида муровьев Myrmecia pilosula имеют пару хромосом на клетку.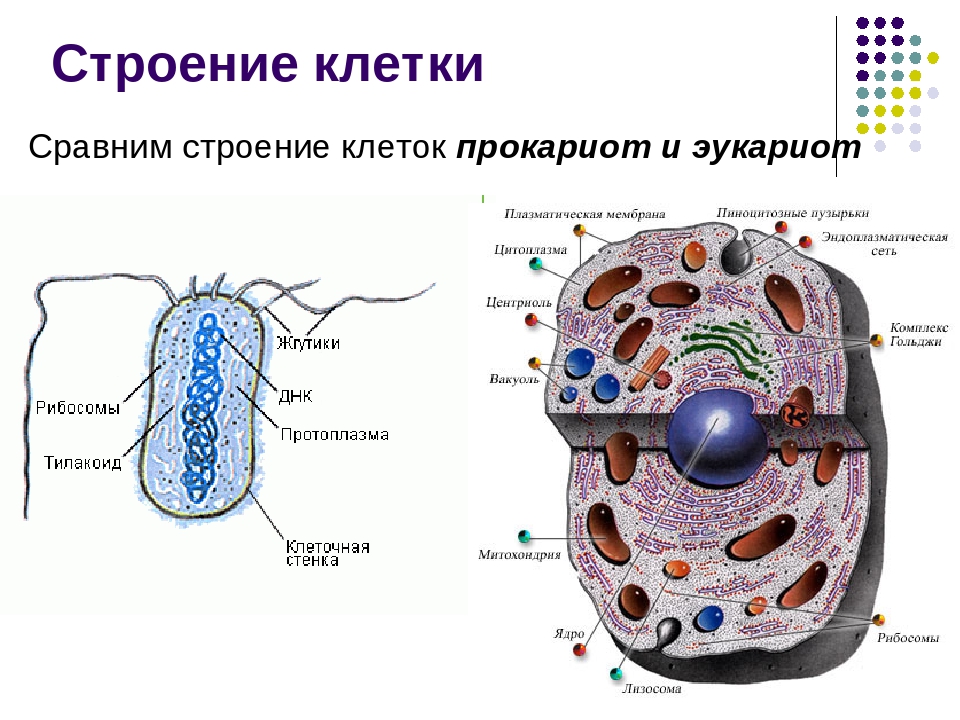 Самцы имеют только 1 хрососому в каждой клетке.
Самцы имеют только 1 хрососому в каждой клетке.
Наибольшее число: вид папоротников Ophioglossum reticulatum имеет около 630 пар хромосом, или 1260 хромосом на клетку
Верхний предел числа хромосом не зависит от количества ДНК которое в них входит: у американской амфибии Amphiuma ДНК в ~30 раз больше, чем у человека, которая помещается в 14 хромосомах.
Число хромосом не зависит от уровня организации и не всегда указывает на родство: одно и тоже число их может быть у очень далёких друг от друга систематических групп и может сильно отличаться у близких по происхождению видов.
Например величина генома у эукариот обычно гораздо больше, чем у прокариот. Отклонения в величине генома у эукариот гораздо больше, чем у бактерий: от 8.8*10 в 6 степени нуклеиновых пар до 6.9*10 в 11 степени нуклеиновых пар, т. е. приблизительно в 80 тысяч раз. Огромная межвидовая вариация в размере генома среди эукариот не имеет отношения ни к сложности организма, ни к вероятному числу генов, которые этот организм имеет.
а) размеры генома большинства эукариот настолько велики, что их потенциальная информационная емкость намного превышает реальное число генов;
б) виды одного и того же рода могут существенно (в несколько раз) отличаться по величине генома;
в) так называемые «эволюционно примитивные» реликтовые формы («живые ископаемые») по содержанию ДНК на клетку зачастую превосходят представителей эволюционно преуспевающих таксономических групп: почти 35-кратное превышение генома двоякодышаших рыб над геномом человека.
Таким образом величина геномов у представителей разных таксонов вовсе не согласуется с нашими интуитивными представлениями о том, кто из них «выше», а кто «ниже» на эволюционной лестнице. «Судить о степени эволюционной продвинутости по размерам генома столь же правомочно, как оценивать общественное положение человека по его весу». ))
«Судить о степени эволюционной продвинутости по размерам генома столь же правомочно, как оценивать общественное положение человека по его весу». ))
Избыточность величины генома конкретно выражается в наличии многочисленных семейств повторяющейся ДНК. Разнообразие семейств повторяющейся ДНК с трудом поддается систематизации.
Вопрос 10. Согласны ли вы с утверждением, что ядро является важнейшей частью клетки? Ответ обоснуйте.
Ядро – основной компонент клетки, несущей генетическую информации Ядро – располагается в центре, оно является важнейшей частью клетки. Значение ядра: участвует в образовании белка, РНК, рибосом; регуляция формообразования процессов и функции клеток; хранение генетического кода и его точное воспроизведение в ряду клеточного поколения.
Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал функционирует и воспроизводится. Поэтому выпадение или нарушение любой из перечисленных выше функций губительно для клетки в целом. Так нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не будет обеспечивать клеточные функции, в результате чего клетка погибает. Нарушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что также губительно для клеток. К такому же результату приведет нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток. Выпадение в результате поражения ядра или в случае нарушений каких-либо регуляторных процессов синтеза любой формы РНК автоматически приведет к остановке синтеза белка в клетке или к грубым его нарушениям.
Поэтому выпадение или нарушение любой из перечисленных выше функций губительно для клетки в целом. Так нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не будет обеспечивать клеточные функции, в результате чего клетка погибает. Нарушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что также губительно для клеток. К такому же результату приведет нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток. Выпадение в результате поражения ядра или в случае нарушений каких-либо регуляторных процессов синтеза любой формы РНК автоматически приведет к остановке синтеза белка в клетке или к грубым его нарушениям.
Никогда не сдавайтесь!
Никогда не сдавайтесь!«Никогда не сдавайтесь.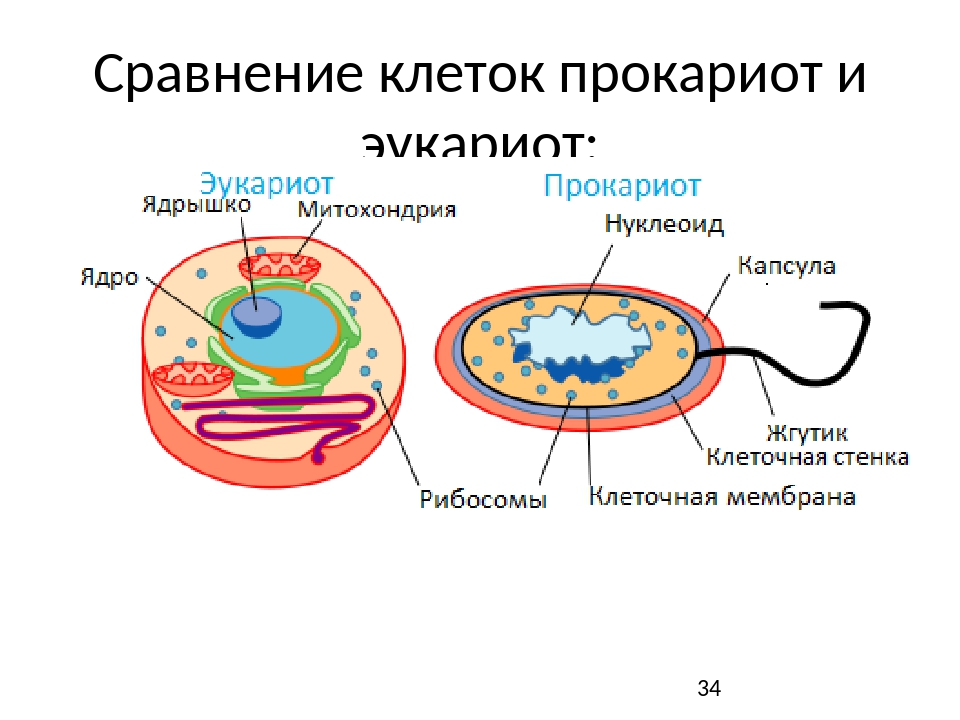 Никогда не сдавайтесь. Никогда, никогда, никогда, никогда — и ни в чем, в великом или малом, большом или незначительном — из этого правила нет исключения…»
Никогда не сдавайтесь. Никогда, никогда, никогда, никогда — и ни в чем, в великом или малом, большом или незначительном — из этого правила нет исключения…»
Уинстон Черчилль.
Изучив данный урок, Вы узнаете как «перемалывать» проблемы, ошибки и «внезапно навалившиеся неудачи», которые бывают в любом виде деятельности. Особенно, в новых для Вас областях.
Вот что говорит о своём понимании неудач и проблем мультимиллионер Марк Аллен в своей книге «Успех для ленивых» :
«Я был ленивым, неорганизованным, неопытным, подавленным и финансово-зависимым до 30 лет. Я до сих пор ленивый и неорганизованный, но больше не неопытный, не подавленный, и, слава Богу, финансово независимый.
Ты можешь быть ленивым и в то же время настойчивым. И как только ты научишься этому — ты во многом достигнешь совершенства… Одна простая фраза может перевернуть твой образ мышления, изменить поток твоих мыслей и жизненный путь.
Вот эта фантастическая фраза, которая изменила мои мысли и мою жизнь: «В каждой проблеме есть равносильная или большая польза. В каждой проблеме заключены огромные возможности. Даже с ударами судьбы мы получаем замечательные дары.»
Человек, воспитанный в нашей, «цивилизованной» школьной и экономической системе воспринимают любую ошибку как неудачу или вообще трагедию. Отсюда и боязнь сделать ошибку, которая просто парализует Волю, сковывает творческое мышление и вызывает негативные эмоции.
А давайте посмотрим на проблемы и ошибки с логической точки зрения. Ну вот, случилось что-то…
Прежде всего, успокойтесь, расслабьтесь, подумайте о чем-то другом. Сделайте волевое усилие, нарисуйте в своей голове радостную картинку хотя бы на 10 секунд.
Смогли? А теперь — решаем проблему по следующему алгоритму:
- Каковы причины этой проблемы и что нужно изменить, чтобы такой проблемы больше не возникло? Какие действия, убеждения, страхи привели именно к этой проблеме?
- Каким наихудшим образом может закончиться эта проблема? Подробно опишите самый негативный исход.
 Спокойно, отрешенно. Возможно, он не такой уж и ужасный, как Вам показалось сначала.
Спокойно, отрешенно. Возможно, он не такой уж и ужасный, как Вам показалось сначала.
- Какой наилучший выход из этой проблемы? Представьте, что Вы достойно справились с данной проблемой. Какой должен быть идеальный результат.
- Возможно ли решить проблему нестандартным образом, не «как все» и не «как всегда»? От каких старых привычек, убеждений, и навыков придётся при этом отказаться?
- Какие ресурсы, сколько времени и какие действия понадобятся, чтобы решить эту проблему? Просто составляете реальный, пошаговый план.
- Какую пользу можно извлечь из данной проблемы? Обычно над этим никто не задумывается. А ведь именно этот пункт и есть самый важный.
Отдельно надо сказать вот ещё о какой беде, которая может свести на нет все Ваши успешные начинания. Негативное эмоциональное отношение к трудностям и ошибкам, которые неизбежно были, есть и будут в любой человеческой деятельности.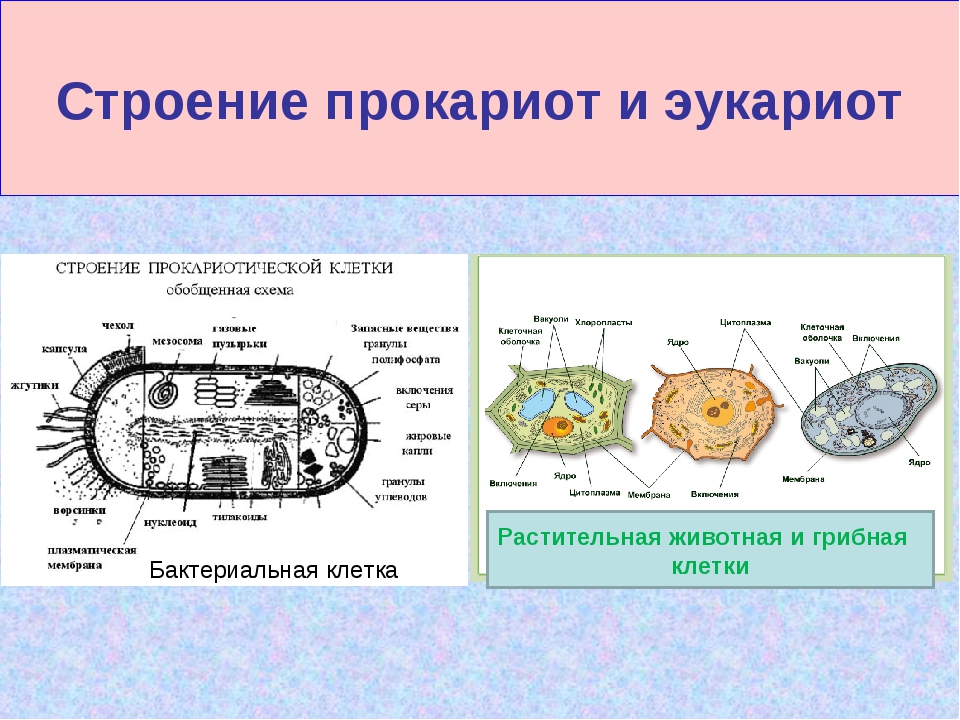 Негативные эмоции, которые не только парализуют любые порывы к движению вперед, но и часто сказываются даже на физическом здоровье человека. Это плохая новость.
Негативные эмоции, которые не только парализуют любые порывы к движению вперед, но и часто сказываются даже на физическом здоровье человека. Это плохая новость.
А вот хорошая — включение негативных эмоций это обычная человеческая привычка. И, как любую привычку её можно заменить на противоположную. А ещё очень важно просто «выдавить» такие «убийственные» для дела и для здоровья мысли из подсознания.
Вот несколько советов:
- Любите себя. Относитесь к себе с любовью. Вы достойны всего в этой жизни. Почувствуйте это. Запомните это состояние и вызывайте его тогда, когда Вам плохо.
- Любите жизнь. Такой, какая она есть. Это Ваша жизнь!
- Не жалуйтесь! Никогда! Нытикам всегда не везёт.
- Высыпайтесь, расслабляйтесь, отдыхайте…
- Если Вы постоянно тревожитесь, то, во-первых снизьте важность всех дел (позвольте себе делать дело неидеально, а иногда и плохо), во вторых — старайтесь делать только самые важные дела.

- Постарайтесь не сравнивать себя с другими, особенно с теми, которые кажутся Вам более преуспевающими в чём-то. У них свои проблемы, просто Вы об этом не догадываетесь.
- Хвалите себя. За каждое выполненное до конца дело, каким бы незначительным оно не было.
- Воспитайте в себе привычку радоваться проблемам, ошибкам и неудачам. Да, это очень трудно, но такое отношение превращает неудачу в жизненный опыт позволит легко превратить неудачи в победы уже в ближайшем будущем.
- Страх — это всего лишь боязнь неизведанного. Враг любого страха — действие. Если Вы не действуете, Вы теряете контроль над ситуацией. Если Вы действуете, то скоро увидите, что 90% страха — это Ваша неинформированность, а оставшиеся 10% — это проблемы, которые можно решить. Отдавайте энергию решениям, а не переживаниям!
- Программируйте подсознание на позитив. Повторяйте про себя «Я смогу», «У меня всегда всё хорошо получается» и т.
 д.
д.
- Негативные переживания — это привычка. Замените эту привычку на противоположную. Дайте волю чувствам энтузиазма, подъёма, уверенности в своих силах и другим позитивным эмоциям.
И главное! Никогда, никогда, никогда не сдавайтесь!
Вы и только Вы можете позволить сломать себя.
Выбирать Вам!
Материальный носитель наследственной информации называется. ДНК – носитель наследственной информации
1) и- РНК 2) т-РНК 3) ДНК 4) хромосома
А2. В дочерние клетки кожи человека при их размножении поступает от материнской клетки:
полная генетическая информация
половина информации
четверть информации
нет верного ответа
А3. Репликация ДНК сопровождается разрывом химических связей:
пептидных, между аминокислотами
ковалентных, между углеводом и фосфатом
водородных, между азотистыми основаниями
ионных, внутри структуры молекулы
А4. При реплткации молекулы ДНК образуется:
нить, распавшаяся на отдельные фрагменты дочерних молекул
молекула, состаящая из двух новых цепей ДНК
молекула, половина которой состоит из нити и-РНК
дочерняя молекула, состоящая из одной старой и одной новой цепи ДНК
А5. Транскрипция – это процесс:
Транскрипция – это процесс:
1)репликации ДНК
2) синтеза и-РНК
3) синтеза белка
4) присоединения т-РНК к аминокислоте
А6. Если аминокислота кодируется кодоном УГГ, то в ДНК ему соответствует триплет:
ТЦЦ 2) АГГ 3) УЦЦ 4) АЦЦ
А7. Один триплет ДНК несет информацию о:
Последовательности аминокислот в молекуле белка
Месте определенной АК в белковой цепи
Признаке конкретного организма
Аминокислоте, включаемой в белковую цепь
А8. Количество т-РНК, участвующих в трансляции, равно количеству:
Кодонов и-РНК, шифрующих аминокислоты
Молекул и-РНК
Генов, входящих в молекулу ДНК
Белков, синтезируемых на рибосомах
А9. Период жизни клетки от деления до деления называется:
Интерфаза 3) мейоз
Митоз 4) клеточный цикл
А10. Сколько хроматид содержится в 8 видимых в метафазе митоза хромосомах:
1) 6 2) 8 3) 12 4) 16
А11. Количество хромосом в соматических клетках человека после митоза равно:
1) 23 2) 46 3) 92 4) 44
Каждый белок представлен одной или несколькими полипептидными цепями.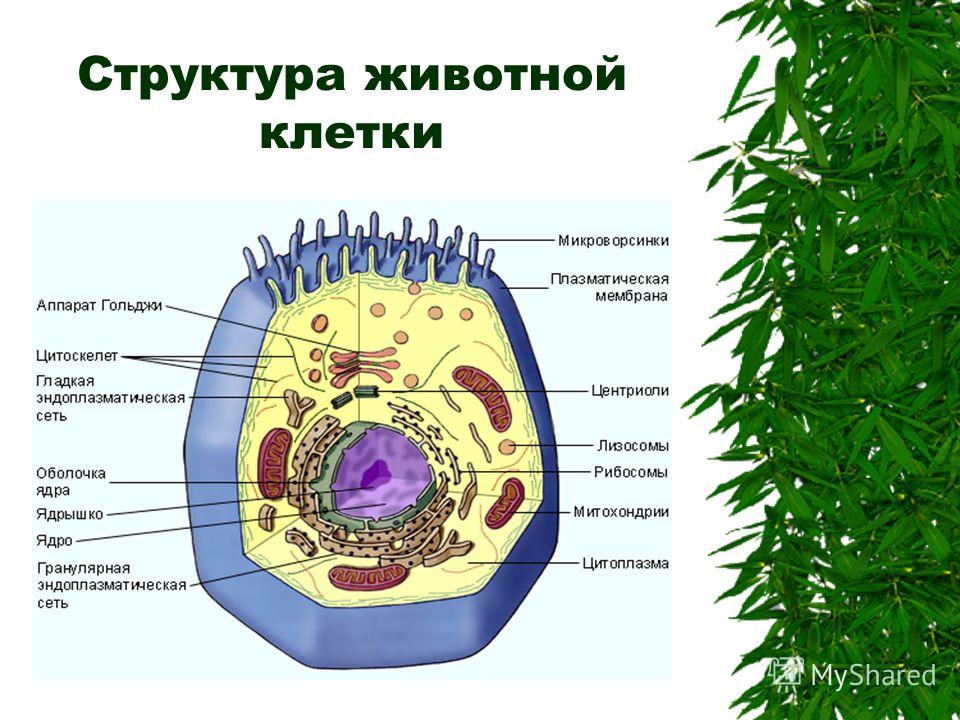 Участок ДНК, несущий информацию об одной полипептидной цепи, называется геном
. Каждая молекула ДНК содержит множество разных генов.
Совокупность молекул ДНК клетки выполняет функцию носителя генетической
информации. Благодаря уникальному свойству — способности к удвоению,
которым не обладает ни одна другая из известных молекул, ДНК могут
копироваться. При делении «копии» ДНК расходятся по двум дочерним
клеткам, каждая из которых вследствие этого будет иметь ту же информацию,
которая содержалась в материнской клетке. Так как гены — это участки
молекул ДНК, то две клетки, образующиеся при делении, имеют одинаковые
наборы генов. Каждая клетка многоклеточного организма при половом
размножении возникает из одной оплодотворенной яйцеклетки в результате
многократных делений. Значит, случайно возникшая ошибка в гене одной клетки
будет воспроизведена в генах миллионов ее потомков. Вот почему все
эритроциты больного серповидноклеточной анемией
имеют одинаково испорченный гемоглобин. Ошибка произошла в гене, несущем
информацию о бета-цепи белка.
Участок ДНК, несущий информацию об одной полипептидной цепи, называется геном
. Каждая молекула ДНК содержит множество разных генов.
Совокупность молекул ДНК клетки выполняет функцию носителя генетической
информации. Благодаря уникальному свойству — способности к удвоению,
которым не обладает ни одна другая из известных молекул, ДНК могут
копироваться. При делении «копии» ДНК расходятся по двум дочерним
клеткам, каждая из которых вследствие этого будет иметь ту же информацию,
которая содержалась в материнской клетке. Так как гены — это участки
молекул ДНК, то две клетки, образующиеся при делении, имеют одинаковые
наборы генов. Каждая клетка многоклеточного организма при половом
размножении возникает из одной оплодотворенной яйцеклетки в результате
многократных делений. Значит, случайно возникшая ошибка в гене одной клетки
будет воспроизведена в генах миллионов ее потомков. Вот почему все
эритроциты больного серповидноклеточной анемией
имеют одинаково испорченный гемоглобин. Ошибка произошла в гене, несущем
информацию о бета-цепи белка. Копией гена является и-РНК
. По ней, как по матрице, в каждом эритроците тысячи раз
«печатается» неправильный белок. Дети получают испорченные гены
от родителей через их половые клетки. Генетическая информация передается
как от одной клетки в дочерние клетки, так и от родителей детям. Ген
является единицей генетической, или наследственной, информации.
Копией гена является и-РНК
. По ней, как по матрице, в каждом эритроците тысячи раз
«печатается» неправильный белок. Дети получают испорченные гены
от родителей через их половые клетки. Генетическая информация передается
как от одной клетки в дочерние клетки, так и от родителей детям. Ген
является единицей генетической, или наследственной, информации.
Каждый белок представлен одной или несколькими полипиптидными цепями. Участок ДНК, несущий информацию об одной полипиптидной цепи, называют геном . Совокупность молекул ДНК клетки выполняет функцию носителя генетической информации. Генетическая информация передается как от материнской клетки дочерним клеткам, так и от родителей детям. Ген является единицей генетической , или наследственной, информации.
ДНК – носитель генетической информаци в клетке
– непосредственного участия в синтезе белков не принимает. В клетках эукариот молекулы ДНК содержатся в хромосомах ядра и отделены ядерной оболочкой от цитоплазмы, где происходит синтез белков. К рибосомам – местам сборки белков – высылается из ядра несущий информацию посредник, способный пройти через поры ядерной оболочки. Таким посредником является информационная РНК (иРНК). По принципу комплементарности она синтезируется на ДНК при участие фермента, называемого РНК-полимеразой
.
К рибосомам – местам сборки белков – высылается из ядра несущий информацию посредник, способный пройти через поры ядерной оболочки. Таким посредником является информационная РНК (иРНК). По принципу комплементарности она синтезируется на ДНК при участие фермента, называемого РНК-полимеразой
.
1) Процесс синтеза РНК, в котором в качестве матрицы используется одна из цепей молекулы ДНК, называют транскрипцией .
2) Транскрипция – это механизм, с помощью которого нуклеотидная поледовательность одной из цепей ДНК переписыватся комплементарной ей последовательность молекулы иРНК.
Информационная РНК – это однонитевая молекула, и транскрипция идет с одной цепи двунитевой молекулы ДНК. Она является копией не всей молекулы ДНК, а только части ее – одного гена у эукариот или группы рядом расположенных генов, несущих информацию о структуре белков, необходимых для выполнения одной функции, у прокариот. Такую группу генов называют опероном
. В начале каждого оперона находится своего рода посадочная площадка для РНК-полимеразы, называемая промотором
.это специфическая последовательность нуклеотидов ДНК, которую фермент «узнает» благодаря химическому сродству. Только присоединившись к промотору, РНК-полиммераза способна начать интез РНК. Доядя до конца оперона, фермент встречает сигнал (в виде определенной последоватльности нуклеотидов), означающий конец считывания. Готовая иРНК отходит от ДНК и направляется к месту синтеза белков.
В начале каждого оперона находится своего рода посадочная площадка для РНК-полимеразы, называемая промотором
.это специфическая последовательность нуклеотидов ДНК, которую фермент «узнает» благодаря химическому сродству. Только присоединившись к промотору, РНК-полиммераза способна начать интез РНК. Доядя до конца оперона, фермент встречает сигнал (в виде определенной последоватльности нуклеотидов), означающий конец считывания. Готовая иРНК отходит от ДНК и направляется к месту синтеза белков.
В процессе транскрипции можно выделить четыре стадии: 1) связывание РНК
-полимеразы с промотором; 2) инициация
– начало синтеза. Оназаключается в образовании первой фосфодиэфирной связи между АТФ или ГТФ и вторым нуклеотидом синтезирующейся молекулы РНК; 3) элонгация
– рост цепи РНК; т.е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные им нуклеотиды в транскрибируемой нити ДНК. Скорость элонгации 50 нуклеотидов в секунду; 4) терминация
– завершение синтеза РНК.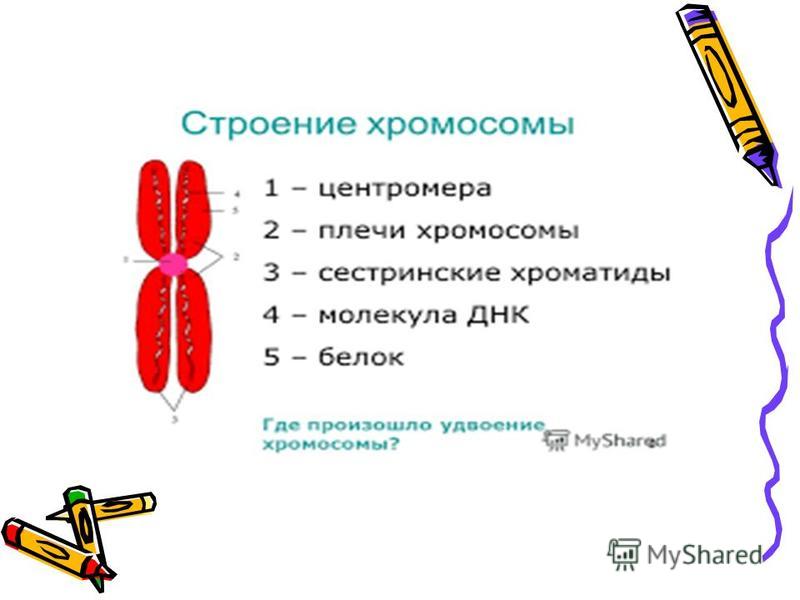
Пройдя через поры ядерной оболочки, иРНК направляется к рибосомам, где осуществляется расшифровка генетической информации – перевод ее с «языка» нуклеотидов на «язык» аминокислот. Синтез полипептидных цепей по матрице иРНК, происходящий в рибосомах, называют трансляцией (лат. translation – перевод).
Аминокислоты, из котрых синтезируютсябелки, доставляются к рибосомам с помощью специальных РНК, называемых транспортными (тРНК). В клетке имеется столько же разных тРНК, сколько кодонов, шифрующих аминокислоты. На вершине «листа» каждой тРНК имеется последовательность трех нуклеотидов, комплементарных нуклеотидам кодона в иРНК. Ее называют антикодоном. Специальный фермент – кодаза – опознает тРНК и присоединяет к «черешку листа» аминокислоту – только ту, которая кодируется триплетом, комплементарным антикодону. На образование ковалентной связи между тРНК и «своей» аминокислотой затрачивается энергия одной молекулы АТФ.
Для того чтобы аминокислота включилась в полипептидную цепь, она должна оторваться от тРНК.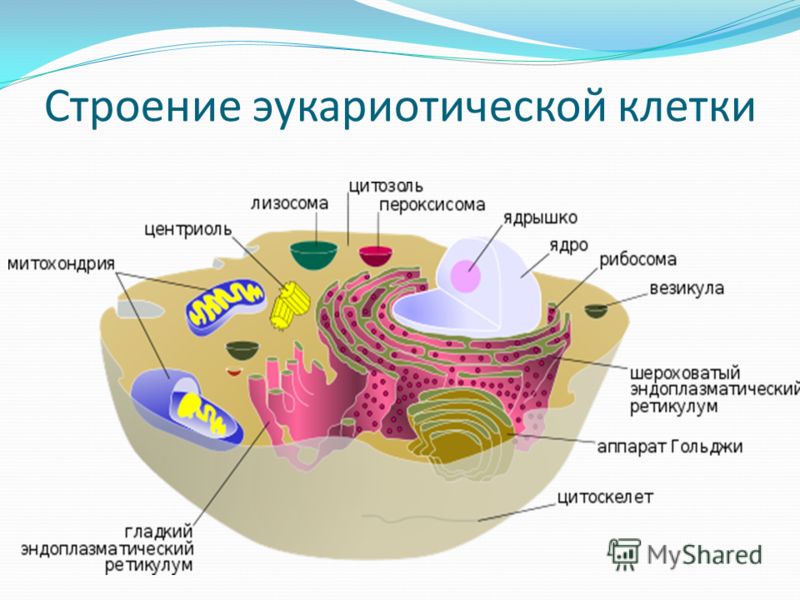 Это становится возможным, когда тРНК поступает на рибососму и антикодон узнает свой кодон в иРНК. В рибосоме имеется два участка для связывания двух молекул тРНК. В один из этих участков, называемый акцепторным
, поступает тРНК с аминокислотой и присоединяется к своему кодону (I). Эта аминокислота присоединяет к себе (акцептирует) растущую цепь белка (II)? Между ними образуется пептидная связь. тРНК, к которой теперь присоединяется вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся акцепторный участок приходит новая тРНК, связанная с аминокислотой, которая шифруется очередным кодоном (III). Из донорного участка сюда вновь переносится оторвавшаяся полипептидная цепь и удлинняется еще на одно звено. Аминокислоты в растущей цепи соединены в той последовательности, в которой расположены шифрующие их кодоны в иРНК.
Это становится возможным, когда тРНК поступает на рибососму и антикодон узнает свой кодон в иРНК. В рибосоме имеется два участка для связывания двух молекул тРНК. В один из этих участков, называемый акцепторным
, поступает тРНК с аминокислотой и присоединяется к своему кодону (I). Эта аминокислота присоединяет к себе (акцептирует) растущую цепь белка (II)? Между ними образуется пептидная связь. тРНК, к которой теперь присоединяется вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся акцепторный участок приходит новая тРНК, связанная с аминокислотой, которая шифруется очередным кодоном (III). Из донорного участка сюда вновь переносится оторвавшаяся полипептидная цепь и удлинняется еще на одно звено. Аминокислоты в растущей цепи соединены в той последовательности, в которой расположены шифрующие их кодоны в иРНК.
Когда на рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА
), являющиеся «знаками препинания» между генами, ни одна тРНК не может занять место в акцепторном участке.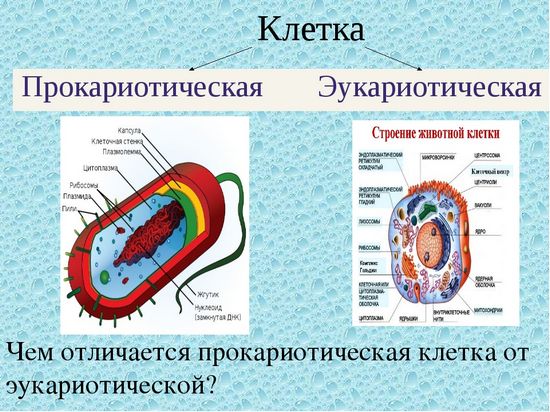 Дело в том, что не существует антикодонов, комплементарных последовательностям нуклеотидов «знаков препинания». Оторвавшейся цепи не к чему присоединиться в акцепторном участке, и она покидает рибосому. Синтез белка завершен.
Дело в том, что не существует антикодонов, комплементарных последовательностям нуклеотидов «знаков препинания». Оторвавшейся цепи не к чему присоединиться в акцепторном участке, и она покидает рибосому. Синтез белка завершен.
У прокариот синтез белков начинается с того, что кодонАУГ , расположенный на первом месте в копии с каждого гена, занимае в рибосоме такую позицию, что с ним взаимодействует антикодон особой тРНК, оединенной с формилментионином . Эта измененная форма аминокислоты метионина сразу попадает в донорный участок и выполняет роль заглавной буквы во фразе – с нее в бактериальной клетке начинается синтез любой полипептидной цепи. Когда триплет АУГ стоит не на первом месте, а внутри копии с гена, он кодирует аминокислоту метионин. После завершения синтеза полипептидной цепи формилметионин отщепляется от нее и в готовом белке отсуствует.
Для увеличения производства белков иРНК часто проходит одновременно не по одной, а по нескольким рибосомам.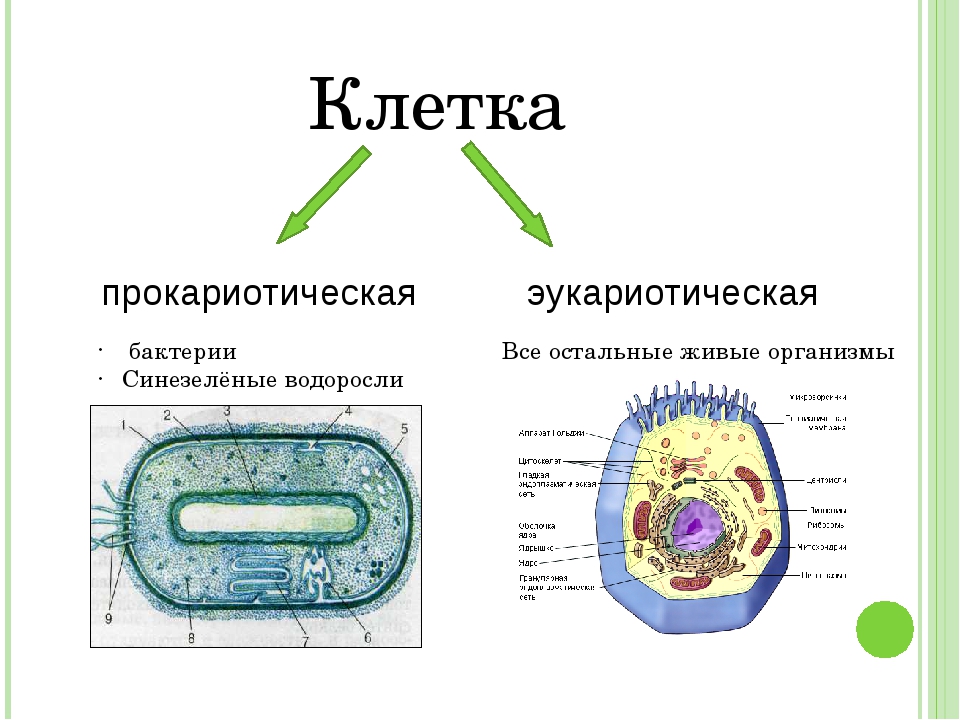 Акую структуру, объединенную одной молекулой иРНК, называют полисомой
. На каждой рибосоме вэтом похожем на нитку бус конвейере синтезируются одинаковые белки.
Акую структуру, объединенную одной молекулой иРНК, называют полисомой
. На каждой рибосоме вэтом похожем на нитку бус конвейере синтезируются одинаковые белки.
Аминокислоты бесперебойно поставляются к рибосомам с помощью тРНК. Отдав аминокислоту, тРНК покидает рибосому и с помощью кодазы соединяется. Высокая слаженность всех «служб комбината» по производсву белов позволяет в течении нескольких секунд синтезировать полипептидные цепи, состоящие из сотен аминокислот.
Свойства генетического кода. Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку
ДНК → иРНК → белок
Генетическая информация, содержащаяся в ДНК и в иРНК, заключена в последовательности расположения нуклеотидов в молекулах.
Каким же образом происходит перевод информации с «языка» нуклеотидов на «язык» аминокислот? Такой перевод осуществляется с помощью генетического кода. Код, или шифр
, — это система символов для перевода одной формы информации в другую. Генетический код
–это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в иРНК.
Генетический код
–это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в иРНК.
Какими же свойствами обладает генетический код?
1. Код триплетен . В состав РНК входят четыре нуклеотида: А, Г, Ц, У. Если бы мы пытались обозначить одну аминокислоту одним нуклеотидом, то 16 из 20 аминокислот остались бы не зашифрованы. Двух буквенный код позволил бы зашифровать 16 аминокислот. Природа создала трехбуквенный, или триплетный, код. Это означает, что каждая из 20 аминокислот зашифрована последовательностью из трех нуклеотидов, называемой триплетом или кодоном.
2. Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном. Исключения: метеонин и триптофан, каждая из которых кодируется одним триплетом.
3.
Код однозначен. Каждый кодон шифрует только одну аминокислоту.
4. Между генами имеется «знаки препинания». В печатном тексте в конце каждой фразы стоит точка. Несколько связанных по смыслу фраз составляют абзац. На языке генетической информации таким абзацем являетсяоперон и комплементарная ему иРНК. Каждый ген в опероне прокариот или отдельный ген эукариот кодирует одну полипептидную цепочку – фразу. Так как в ряде случаев по матрице иРНК последовательно создается несколько разных полипептидных цепей, они должны быть отделены друг от друга. Для этого в генетическом годе имеются три специальных триплета – УАА, УАГ, УГА, каждый из которых обозначает прекращение синтеза одной полипептидной цепи. Таким образом, эти триплеты выполняют функцию знаков препинания. Они находятся в конце каждого гена.
5. Внутри гена нет «знаков препинания».
6.
Код универсален.
Генетический код един для всех живущих на Земле существ. У бактерий и грибов, пшеницы и хлопка, рыб и червей, лягушек и человека одни и те же триплеты кодируют одни и те же аминокислоты.
Принципы репликации ДНК. Преемственность генетического материала в поколениях клеток и организмов обеспечивается процессом репликации – удвоения молекул ДНК. Этот сложный процесс осуществляется комплексом нескольких ферментов и не обладающих каталитической активностью белов, необходимых для придания полинуклеотидным цепям нужной конформации. В результате репликации образуются две идентичные двойные спирали ДНК. Эти так называемые дочерние молекулы ничем не отличаются друг от друга и от исходной материнской молекулы ДНК. Репликация происходит в клетке перед делением, поэтому каждая дочерняя клетка получает точно такие же молекулы ДНК, какие имела материнская клетка. Процесс репликации основан на ряде принципов:
1. Комплементарность . Каждая из двух цепей материнской молекулы ДНК служит матрицей для синтеза дополняющей ее, т.е. комплементарной, дочерней цепи.
2.
Полуконсервативность
. В результате репликации образуются две двойные дочерние спирали, каждая из которых сохраняет (консервирует) в неизменном виде одну из цепей материнской ДНК.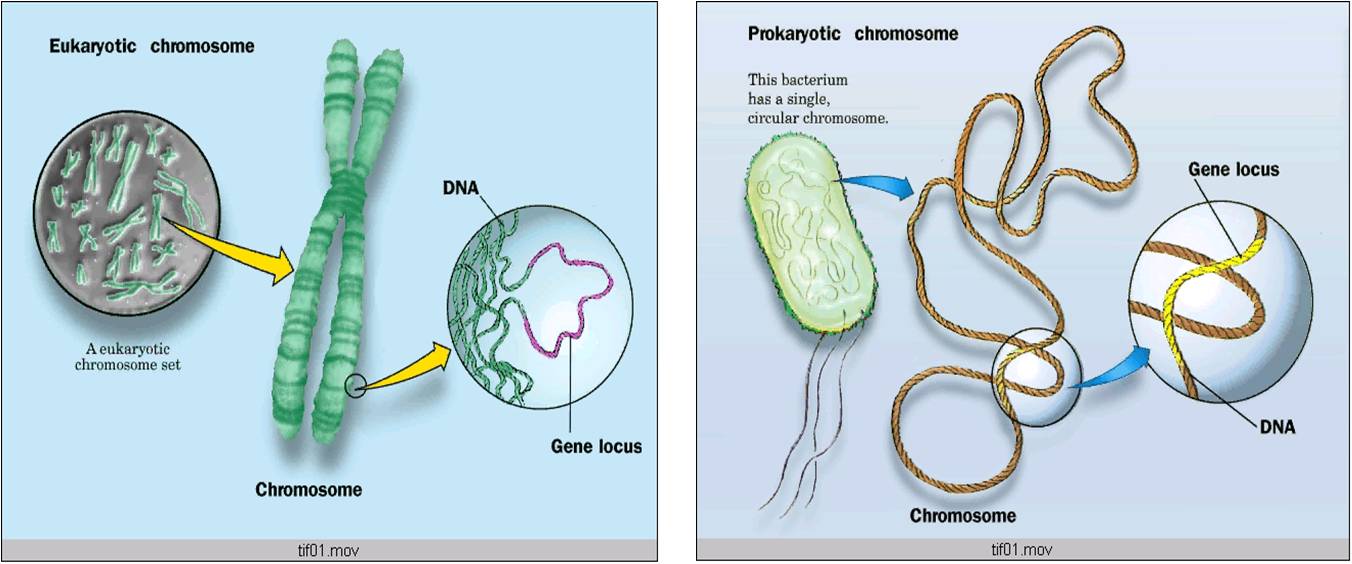 Вторые цепи дочерних молекул синтезируются из нуклеотидов заново по прицепу комплементарности к нитям материнской ДНК. Дочерние ДНК ничем не отличаются друг от друга и от материнской двойной спирали.
Вторые цепи дочерних молекул синтезируются из нуклеотидов заново по прицепу комплементарности к нитям материнской ДНК. Дочерние ДНК ничем не отличаются друг от друга и от материнской двойной спирали.
3.
Антипараллельность
. Каждая цепь ДНК имеет определенную ориентацию. Один конец несет гидроксильную группу (-ОН),присоединенную к 3’-углероду в сахаре дезоксирибозе, на другом конце цепи находится остаток фосфорной кислоты в положении 5’-положении сахара. Две комплементарные цепи в молекуле ДНК ориентированы в противоположных направлениях – антипараллельно. Ферменты, синтезирующие новые нити ДНК и называемые ДНК-полимеразами, могут передвигаться вдоль матричных цепей лишь в одном направлении – от их 3’-концов к 5’-концам. При этом синтез комплементарных нитей всегда ведется в 5’→3’ направлении, т.е. униполярно. Поэтому в процессе репликации одновременный синтез новых цепей идет антипараллельно.
4.
Прерывитость
. Для того чтобы новые нити ДНК были построены по принципу комплементарности, двойная спираль должна быть раскручена и между материнскими цепями должны отсутствовать водородные связи.
Только в этом случае ДНК-полимеразы способна двигаться по материнским нитям и использовать их в качестве матриц для безошибочного синтеза дочерних цепей. Но полное раскручивание спиралей, состоящих из многих миллионов пар нуклеотидов, сопряжено со столь значительным числом вращений и такими энергетическими затратами, которые невозможны в условиях клетки. Поэтому репликация у эукариот начинается одновременно в некоторых местах молекулы ДНК. Участок между двумя точками, в которых начинается синтез дочерних цепей, называют репликоном
. Он является единицей репликации.
В каждой молекуле ДНК эукариотической клетки имеется много репликонов. В каждом репликоне можно видеть репликативную вилку – ту часть молекулы ДНК, которая под действием специальных ферментов уже расплелась. Каждая нить в вилке служит матрицей для синтеза комплементарной дочерней цепи. В ходе репликации вилка перемещается вдоль материнской молекулы, при этом расплетаются новые участки ДНК.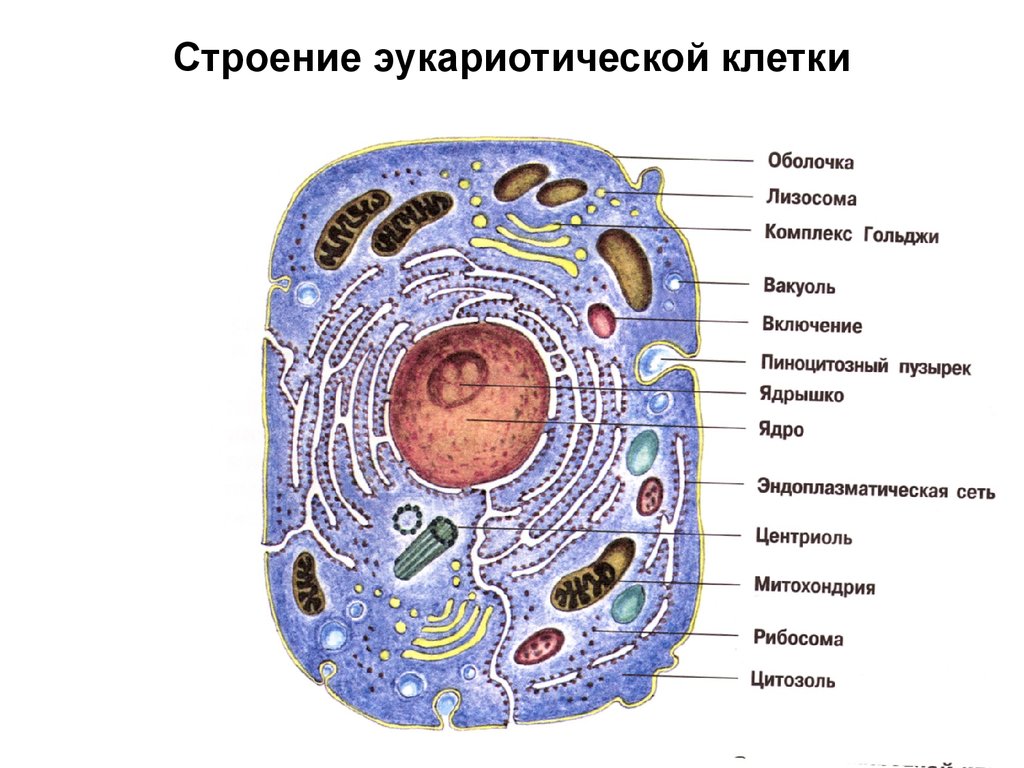 Так как ДНК-полимеразы могут двигаться лишь в одном направлении вдоль матричных нитей, а нити ориентированы антипараллельно, то в каждой вилке одновременно ведут синтез два разных ферментативных комплекса. Причем в каждой вилке одна дочерняя (лидирующая) цепь растет непрерывно, а другая (отстающая) синтезируется отдельными фрагментами длинной в несколько нуклеотидов. Такие ферменты, названые в честь открывшего их японского ученого фрагментами Оказаки
, сшиваются ДНК-лигазой, образуя непрерывную цепь. Механизм образования дочерних цепей ДНК фрагментами называют прерывистыми.
Так как ДНК-полимеразы могут двигаться лишь в одном направлении вдоль матричных нитей, а нити ориентированы антипараллельно, то в каждой вилке одновременно ведут синтез два разных ферментативных комплекса. Причем в каждой вилке одна дочерняя (лидирующая) цепь растет непрерывно, а другая (отстающая) синтезируется отдельными фрагментами длинной в несколько нуклеотидов. Такие ферменты, названые в честь открывшего их японского ученого фрагментами Оказаки
, сшиваются ДНК-лигазой, образуя непрерывную цепь. Механизм образования дочерних цепей ДНК фрагментами называют прерывистыми.
5.
Потребность в затравке ДНК-полимераза не способна начинать синтез лидирующей цепи, ни синтез фрагментов Оказаки отстающей цепи. Она может лишь наращивать уже имеющуюся полинуклеотидную нить, последовательно присоединяя дезоксирибонуклеотиды к ее 3’-ОН концу. Откуда же берется начальный 5’-концевой участок растущей цепи ДНК? Его синтезирует на матрице ДНК особая РНК-полимераза, называемая праймазой
(англ. Primer – затравка). Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-поимеразой. Выполнившая сво. Функци. РНК-затравка удаляется специальным ферментом, а образованная при эом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3’-ОН конец соседнего фрагмента Оказаки.
Primer – затравка). Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-поимеразой. Выполнившая сво. Функци. РНК-затравка удаляется специальным ферментом, а образованная при эом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3’-ОН конец соседнего фрагмента Оказаки.
Проблема недорепликации концов линейных молекул ДНК. Удаление крайних РНК-праймеров,комплементрных 3’-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние цепи оказываются короче 10-20 нуклеотидов. В этом и заключается проблема недорепликации концов линейных молекул.
Проблема недорепликации 3’-концов линейных молекул ДНК решается эукариотическими клетками с помощью специального фермента – теломеразы .
Теломераза является ДНК-полимеразой, достраивающей 3’-концылинейных молекул ДНК хромосом короткими повторяющимися последовательностями. Они, располагаясь друг за другом, образуют регулярную концевую структуру длинной до 10 тыс. нуклеотидов. Помимо белковой части, теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами.
нуклеотидов. Помимо белковой части, теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами.
Схема удлинения концов молекул ДНК. Сначала происходит комплементарное связывание выступающего конца ДНК с матричным участком теломеразной РНК, затем теломераза наращивает ДНК, используя в качестве затравки ее 3’-ОН конец, а в качестве матрицы – РНК, входящую в состав фермента. Эта стадия называется элонгацией. После этого происходит транслокация, т.е. перемещение ДНК, удлиненной на один повтор, относительно фермента. Следом идет элонгация и очередная транслокация.
В результате образуются специализированные концевые структуры хромосом. Они состоят из многократно повторенных коротких последовательностей ДНК и специфических белков.
Краткие итоги.
Урок по общей биологии.
Тема: «ДНК – носитель наследственной информации.
Генетический код».
Цель урока
: закрепить знания о строении ДНК и РНК, изучить понятие ген, генетический код, его свойства.
Оборудование: таблица “Строение животной клетки”, “Белки”, модель ДНК, мультимедийная установка, презентация в Power Point.
Ход урока
1. Орг. момент ……………………………………………………………………1-2 мин.
2. Основная часть: ………………………………………………………………. 30 мин.
2.1 Повторение раннее изученного: ………………………………………….…. 12 мин
2.2 Изучение нового материала: ……………………………….…………………18 мин
3. Закрепление …………………………………………………………………….8 мин
2.1. Повторение ранее изученного
Вопросы к ученикам:
- Что такое белки?
- Что является мономерами всех природных белков? (20 аминокислот).
- Вспомните, какие функции выполняют белки? (Назовите особенности строения нуклеиновых кислот)
- Вспомните, где содержатся молекулы ДНК в клетках растений и животных?
- Что такое комплементарность?
- Назовите виды РНК.
2.2. Изучение нового материала
Все свойства любого организма определяются его белковым составом. Причем структура каждого белка определяется последовательностью аминокислотных остатков. Следовательно, в итоге наследственная информация, которая передается из поколения в поколение, должна содержать сведения о первичной структуре белков.
Следовательно, в итоге наследственная информация, которая передается из поколения в поколение, должна содержать сведения о первичной структуре белков.
Генетическая информация – это информация о строении всех белков организма заключенная в молекулах ДНК.
Ген – это участок молекулы ДНК, кодирующий первичную структуру одной полипептидной цепи. В ДНК заложена информация о первичной структуре белка.
Генетический код – набор сочетаний трех нуклеотидов, кодирующих 20 типов аминокислот, входящих в состав белков.
Свойства генетического кода:
- Код триплетен. Каждой АК (аминокислоте) соответствует участок цепи ДНК, и соответственно, и-РНК из трех рядом стоящих нуклеотидов. В настоящее время генетический код полностью расшифрован и составлена карта, то есть известно, какие триплеты соответствуют той или иной аминокислоте из 20, входящих в состав белков.
- Код однозначен. Каждый кодон шифрует только одну АК.
- Код избыточен (специфичен). Это означает, что каждая АК шифруется более чем одним кодоном (за исключением метионина и триптофана).
 ДНК состоит из 4 разных видов нуклеотидов, а наименьшей структурной единицей гена является триплет нуклеотидов. Поэтому число возможных комбинаций равно 43 = 64. Разных же аминокислот только 20. Таким образом, различных триплетов нуклеотидов с избытком хватает для кодирования всех аминокислот.
ДНК состоит из 4 разных видов нуклеотидов, а наименьшей структурной единицей гена является триплет нуклеотидов. Поэтому число возможных комбинаций равно 43 = 64. Разных же аминокислот только 20. Таким образом, различных триплетов нуклеотидов с избытком хватает для кодирования всех аминокислот.
- Код не перекрывается. Любой нуклеотид может входить в состав только одного триплета.
- Между генами имеются “знаки препинания”. Из 64 триплетов –У-А-А, У-А-Г, У-Г-А не кодируют АК (рассмотреть в учебнике таблицу генетического кода). Эти триплеты – сигналы окончания синтеза полипептидной цепи. Необходимость в наличии данных триплетов объясняется тем, что в ряде случаев на и-РНК осуществляется синтез нескольких полипептидных цепей, и для отделения их друг от друга используются эти триплеты.
- Код универсален. Генетический код един для всех живущих на Земле живых организмов.
3. Закрепление:
Выполнение упражнений по рабочей тетради. (Рабочая тетрадь к учебникам Захарова В.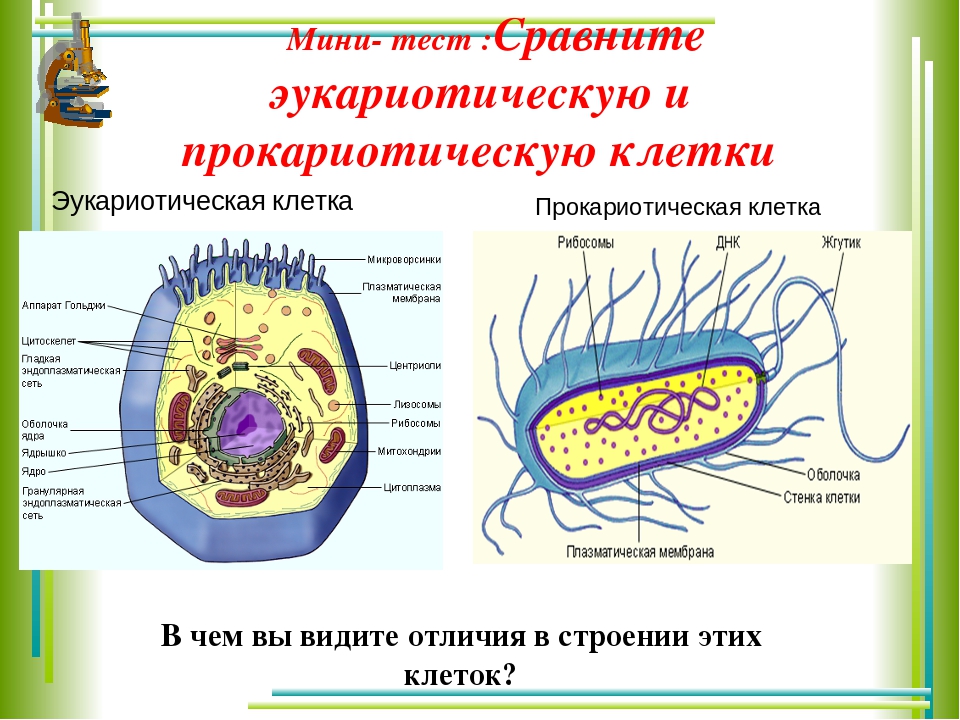 Б., Сухова Т.С. и др.)
Б., Сухова Т.С. и др.)
Домашнее задание. § 2.10 с. 73–75, учебника В. Б. Захарова, С. Г. Мамонтова, Н. И. Сонина, Е. Т. Захаровой 10 класс «Биология. Общая биология», конспект урока.
Предварительный просмотр:
Чтобы пользоваться предварительным просмотром презентаций создайте себе аккаунт (учетную запись) Google и войдите в него: https://accounts.google.com
Подписи к слайдам:
Тема: «ДНК – носитель наследственной информации. Генетический код»
Структурная Каталитическая (Б-ферменты) Регуляторная (Б-гормоны) Сократительная Транспортная Защитная Запасная Энергетическая Ф У Н К Ц И И Б Е Л К А
Строение НК РНК ________________________________ ДНК Азотистое основание (А, Г, Ц, У) Остаток ФК Углевод – рибоза Азотистое Основание (А, Г, Ц, Т) Углевод – дезоксирибоза Остаток ФК
В хромосомах ядра
Комплементарность — пространственная взаимодополняемость молекул или их частей, приводящая к образованию водородных связей. Комплементарные структуры подходят друг к другу как «ключ с замком» (А+Т)+(Г+Ц)=100%
Генетическая информация – это информация о строении всех белков организма заключенная в молекулах ДНК 1 ген = 1 молекула белка
Виды РНК В клетке имеется несколько видов РНК. Все они участвуют в синтезе белка. Транспортные РНК (т-РНК) — это самые маленькие по размерам РНК. Они связывают АК и транспортируют их к месту синтеза белка. Информационные РНК (и-РНК) — они в 10 раз больше тРНК. Их функция состоит в переносе информации о структуре белка от ДНК к месту синтеза белка. Рибосомные РНК (р-РНК) — имеют наибольшие размеры молекулы, входят в состав рибосом.
Все они участвуют в синтезе белка. Транспортные РНК (т-РНК) — это самые маленькие по размерам РНК. Они связывают АК и транспортируют их к месту синтеза белка. Информационные РНК (и-РНК) — они в 10 раз больше тРНК. Их функция состоит в переносе информации о структуре белка от ДНК к месту синтеза белка. Рибосомные РНК (р-РНК) — имеют наибольшие размеры молекулы, входят в состав рибосом.
Ген – это участок молекулы ДНК, кодирующий первичную структуру одной полипептидной цепи Генетический код – набор сочетаний трех нуклеотидов, кодирующих 20 типов аминокислот, входящих в состав белков.
Одна аминокислота закодирована тремя нуклеотидами (один кодон). АЦТ АГЦ ГАТ Триплет, кодон ген АК1 АК2 АК3 белок Свойства генетического кода: Код триплетен. Каждой АК соответствует участок цепи ДНК, и соответственно, и-РНК из трех рядом стоящих нуклеотидов.
Код однозначен. Каждый кодон шифрует только одну АК. Код избыточен. Это означает, что каждая АК шифруется более чем одним кодоном (за исключением метионина и триптофана). Код неперекрывающийся. Любой нуклеотид может входить в состав только одного триплета. Между генами имеются «знаки препинания» (полярность). Из 64 триплетов –У-А-А, У-А-Г, У-Г-А не кодируют АК. Код универсален. Генетический код един для всех живущих на Земле живых организмов.
Код неперекрывающийся. Любой нуклеотид может входить в состав только одного триплета. Между генами имеются «знаки препинания» (полярность). Из 64 триплетов –У-А-А, У-А-Г, У-Г-А не кодируют АК. Код универсален. Генетический код един для всех живущих на Земле живых организмов.
Домашнее задание Конспект урока Подготовить сообщение: «Генетический код».
Решение задач 1) Пользуясь таблицей генетического кода ДНК, определите, какие АК кодируются триплетами: ЦАТ, ТТТ, ГАТ. 2) Используя таблицу генетического кода, нарисуйте участок ДНК, в котором закодирована информация о следующей последовательности аминокислот в белке: — аланин – аргинин – валин – глицин – лизин.
После открытия принципа молекулярной организации такого вещества, как ДНК в 1953 году, начала развиваться молекулярная биология. Далее в процессе исследований ученые выяснили как рекомбенируется ДНК, ее состав и как устроен наш человеческий геном.
Каждый день на молекулярном уровне происходят сложнейшие процессы. Как устроена молекула ДНК, из чего она состоит? И какую роль играют в клетке молекулы ДНК? Расскажем подробно обо всех процессах, происходящих внутри двойной цепи.
Как устроена молекула ДНК, из чего она состоит? И какую роль играют в клетке молекулы ДНК? Расскажем подробно обо всех процессах, происходящих внутри двойной цепи.
Что такое наследственная информация?
Итак, с чего все начиналось? Еще в 1868 нашли в ядрах бактерий. А в 1928 г. Н. Кольцов выдвинул теорию о том, что именно в ДНК зашифрована вся генетическая информация о живом организме. Затем Дж. Уотсон и Ф. Крик нашли модель всем теперь известной спирали ДНК в 1953 году, за что заслужено получили признание и награду — Нобелевскую премию.
Что такое вообще ДНК? Это вещество состоит из 2 объединенных нитей, точнее спиралей. Участок такой цепочки с определенной информацией называется геном.
В ДНК хранится вся информация о том, что за белки будут формироваться и в каком порядке. Макромолекула ДНК — это материальный носитель невероятно объемной
информации, которая записана строгой последовательностью отдельных кирпичиков — нуклеотидов. Всего нуклеотидов 4, они дополняют друг друга химически и геометрически. Этот принцип дополнения, или комплементарности, в науке будет описан позже. Это правило играет ключевую роль в кодировке и декодировании генетической информации.
Этот принцип дополнения, или комплементарности, в науке будет описан позже. Это правило играет ключевую роль в кодировке и декодировании генетической информации.
Так как нить ДНК невероятно длинная, повторений в этой последовательности не бывает. У каждого живого существа собственная уникальная цепочка ДНК.
Функции ДНК
К функциям относятся хранение наследственной информации и ее
передача потомству. Без этой функции геном вида не мог бы сохраняться и развиваться на протяжении тысячелетий.
Организмы, которые претерпели серьезные
мутации генов, чаще не выживают или теряют способность производить потомство. Так происходит природная защита от вырождения вида.
Еще одна существенно важная функция — реализация хранимой информации. Клетка не может создать ни одного жизненно важного белка без тех инструкций, которые хранятся в двойной цепочке.
Состав нуклеиновых кислот
Сейчас уже достоверно известно, из чего состоят сами нуклеотиды — кирпичики ДНК.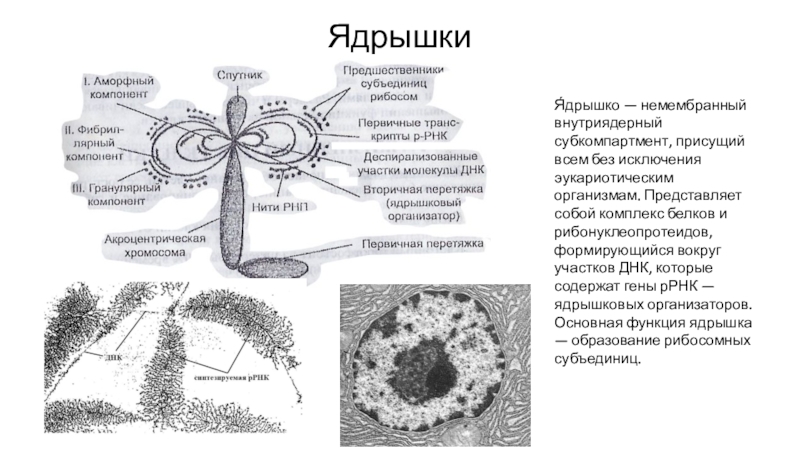 В их состав входят 3 вещества:
В их состав входят 3 вещества:
- Ортофосфорная кислота.
- Азотистое основание. Пиримидиновые основания — которые имеют только одно кольцо. К ним относят тимин и цитозин. Пуриновые основания, в составе которых присутствуют 2 кольца. Это гуанин и аденин.
- Сахароза. В составе ДНК — дезоксирибоза, В РНК — рибоза.
Число нуклеотидов всегда равно числу азотистых оснований. В специальных лабораториях расщепляют нуклеотид и выделяют из него азотистое основание. Так изучают отдельные свойства этих нуклеотидов и возможные мутации в них.
Уровни организации наследственной информации
Разделяют 3 уровня организации: генный, хромосомный и геномный. Вся информация, нужная для синтеза нового белка, содержится на небольшом участке цепочки — гене. То есть ген считается низший и самый простой уровень кодировки информации.
Гены, в свою очередь, собраны в хромосомы. Благодаря такой организации носителя наследственного материала группы признаков по определенным
законам чередуются и передаются от одного поколения к другому. Надо заметить, генов в организме невероятно много, но информация не теряется, даже когда много раз рекомбенируется.
Надо заметить, генов в организме невероятно много, но информация не теряется, даже когда много раз рекомбенируется.
Разделяют несколько видов генов:
- по функциональному назначению выделяют 2 типа: структурные и регуляторные последовательности;
- по влиянию на процессы, протекающие в клетке, различают: супервитальные, летальные, условно летальные гены, а также гены мутаторы и антимутаторы.
Располагаются гены вдоль хромосомы в линейном порядке. В хромосомах информация сфокусирована не вразброс, существует определенный порядок. Существует даже карта, в которой отображены позиции, или локусы генов. Например, известно, что в хромосоме № 18 зашифрованы данные о цвете глаз ребенка .
А что же такое геном? Так называют всю совокупность нуклеотидных последовательностей в клетке организма. Геном характеризует целый вид, а не отдельную особь.
Каков генетический код человека?
Дело в том, что весь огромнейший потенциал человеческого развития заложен уже в период зачатия. Вся наследственная информация, которая необходима для развития зиготы и роста ребенка
уже после рождения, зашифрована в генах. Участки ДНК и есть самые основные носители наследственной информации.
Вся наследственная информация, которая необходима для развития зиготы и роста ребенка
уже после рождения, зашифрована в генах. Участки ДНК и есть самые основные носители наследственной информации.
У человека 46 хромосом, или 22 соматические пары плюс по одной определяющей пол хромосоме от каждого родителя. Этот диплоидный набор хромосом кодирует весь физический облик человека, его умственные и физические способности и предрасположенность к заболеваниям. Соматические хромосомы внешне неразличимы, но несут они разную информацию, так как одна из них от отца, другая — от матери.
Мужской код отличается от женского последней парой хромосом — ХУ. Женский диплоидный набор — это последняя пара, ХХ. Мужчинам достается одна Х-хромосома от биологической матери, и затем она передается дочерям. Половая У-хромосома передается сыновьям.
Хромосомы человека значительно разнятся по размеру. Например, самая маленькая пара хромосом — №17. А самая большая пара — 1 и 3.
Диаметр двойной спирали у человека — всего 2 нм.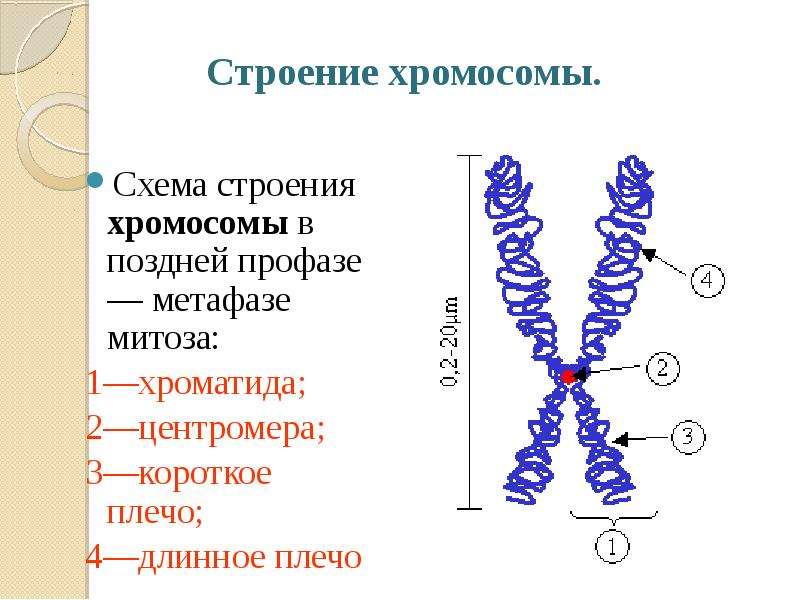 ДНК настолько плотно закручена, что вмещается в маленьком ядре клетки, хотя ее
длина будет достигать 2 метров, если ее
раскрутить. Длина спирали — это сотни миллионов нуклеотидов.
ДНК настолько плотно закручена, что вмещается в маленьком ядре клетки, хотя ее
длина будет достигать 2 метров, если ее
раскрутить. Длина спирали — это сотни миллионов нуклеотидов.
Как передается генетический код?
Итак, какую роль играют в клетке молекулы ДНК при делении? Гены — носители наследственной информации — находятся внутри каждой клетки организма. Чтобы передать свой код дочернему организму, многие существа делят свое ДНК на 2 одинаковые спирали. Это называется репликацией. В процессе репликации ДНК расплетается и специальные «машины» дополняют каждую цепочку. После того как раздвоится генетическая спираль, начинает делиться ядро и все органеллы, а затем и вся клетка.
Но у человека другой процесс передачи генов — половой. Признаки отца и матери перемешиваются, в новом генетическом коде содержится информация от обоих родителей.
Хранение и передача наследственной информации возможны благодаря сложной организации спирали ДНК. Ведь как мы говорили, структура белков зашифрована именно в генах. Раз создавшись во время зачатия, этот код на протяжении всей жизни будет копировать сам себя. Кариотип (личный набор хромосом) не изменяется во время обновления клеток органов. Передача же информации осуществляется с помощью половых гамет — мужских и женских.
Раз создавшись во время зачатия, этот код на протяжении всей жизни будет копировать сам себя. Кариотип (личный набор хромосом) не изменяется во время обновления клеток органов. Передача же информации осуществляется с помощью половых гамет — мужских и женских.
Передавать свою информацию потомству не способны только вирусы, содержащие одну цепочку РНК. Поэтому, чтобы воспроизводиться, им нужны клетки человека или животного.
Реализация наследственной информации
В ядре клетки постоянно происходят важные процессы. Вся информация, записанная в хромосомах, используется для построения белков из аминокислот. Но цепочка ДНК никогда не покидает ядро, поэтому здесь нужна помощь другого важного соединения = РНК. Как раз РНК способно проникнуть через мембрану ядра и взаимодействовать с цепочкой ДНК.
Посредством взаимодействия ДНК и 3 видов РНК происходит реализация всей закодированной информации. На каком уровне происходит реализация наследственной информации? Все взаимодействия происходят на уровне нуклеотидов.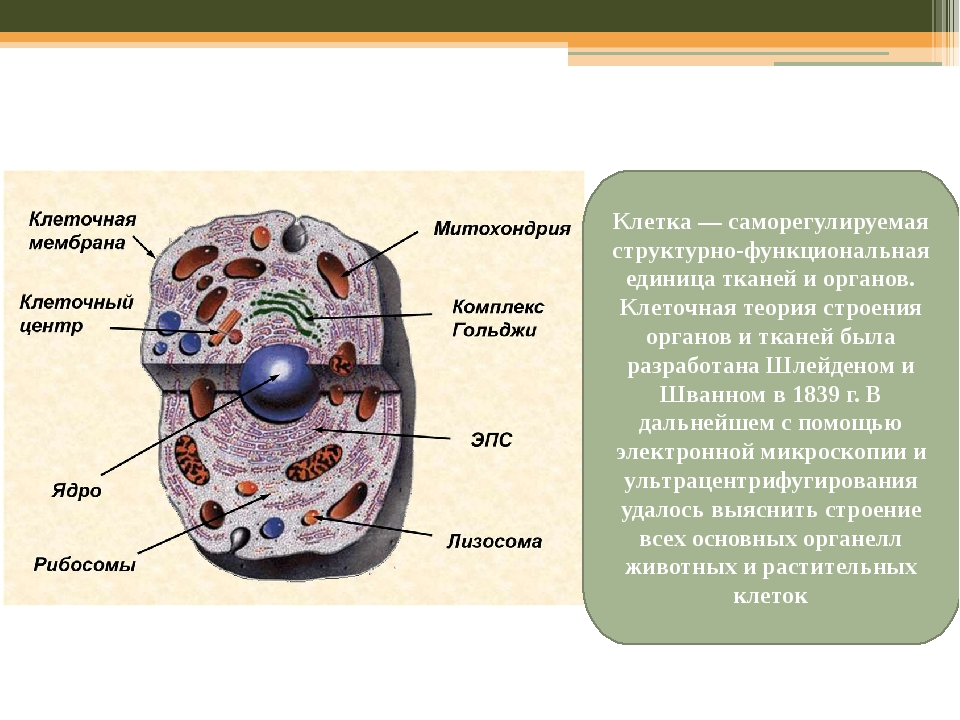 Информационная РНК копирует участок цепи ДНК и приносит эту копию в рибосому. Здесь начинается синтез из нуклеотидов новой молекулы.
Информационная РНК копирует участок цепи ДНК и приносит эту копию в рибосому. Здесь начинается синтез из нуклеотидов новой молекулы.
Для того чтобы иРНК могла скопировать необходимую часть цепи, спираль разворачивается, а затем, по завершении процесса перекодировки, снова восстанавливается. Причем этот процесс может происходить одновременно на 2 сторонах 1 хромосомы.
Принцип комплементарности
Состоят из 4 нуклеотидов — это аденин (А), гуанин (G), цитозин (С), тимин (T). Соединены они водородными связями по правилу комплементарности. Работы Э. Чаргаффа помогли установить это правило, так как ученый заметил некоторые закономерности в поведении этих веществ. Э. Чаргафф открыл, что молярное отношение аденина к тимину равно единице. И точно так же отношение гуанина к цитозину всегда равно единице.
На основе его работ генетики сформировали правило взаимодействия нуклеотидов. Правило комплементарности гласит, что аденин соединяется только с тимином, а гуанин — с цитозином. Во время декодирования спирали и синтеза нового белка в рибосоме такое правило чередования помогает быстро найти необходимую аминокислоту, которая прикреплена к транспортной РНК.
Во время декодирования спирали и синтеза нового белка в рибосоме такое правило чередования помогает быстро найти необходимую аминокислоту, которая прикреплена к транспортной РНК.
РНК и его виды
Что такое наследственная информация? нуклеотидов в двойной цепи ДНК. А что такое РНК? В чем заключается ее работа? РНК, или рибонуклеиновая кислота, помогает извлекать информацию из ДНК, декодировать ее и на основе принципа комплементарности создавать необходимые клеткам белки.
Всего выделяют 3 вида РНК. Каждая из них выполняет строго свою функцию.
- Информационная (иРНК) , или еще ее называют матричная. Она заходит прямо в центр клетки, в ядро. Находит в одной из хромосом необходимый генетический материал для постройки белка и копирует одну из сторон двойной цепи. Копирование происходит снова по принципу комплементарности.
- Транспортная
— это небольшая молекула, у которой на одной стороне декодеры-нуклеотиды, а на другой стороне соответствующие основному коду аминокислоты.
 Задача тРНК — доставить в «цех», то есть в рибосому, где синтезирует необходимую аминокислоту.
Задача тРНК — доставить в «цех», то есть в рибосому, где синтезирует необходимую аминокислоту. - рРНК — рибосомная. Она контролирует количество белка, который продуцируется. Состоит из 2 частей — аминокислотного и пептидного участка.
Единственное отличие при декодировании — у РНК нет тимина. Вместо тимина тут присутствует урацил. Но потом, в процессе синтеза белка, при ТРНК все равно правильно устанавливает все аминокислоты. Если же происходят какие-то сбои в декодировании информации, то возникает мутация.
Репарация поврежденной молекулы ДНК
Процесс восстановления поврежденной двойной цепочки называется репарацией. В процессе репарации поврежденные гены удаляются.
Затем необходимая последовательность элементов в точности воспроизводиться и врезается обратно в то же место на цепи, откуда было извлечено. Все это происходит благодаря специальным химическим веществам — ферментам.
Почему происходят мутации?
Почему некоторые гены начинают мутировать и перестают выполнять свою функцию — хранение жизненно необходимой наследственной информации? Это происходит из-за ошибки при декодировании.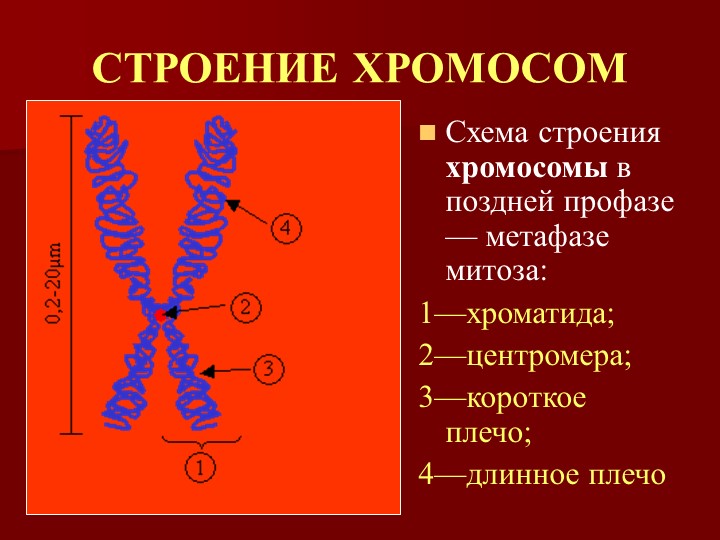 Например, если аденин случайно заменен
на тимин.
Например, если аденин случайно заменен
на тимин.
Существуют также хромосомные и геномные мутации. Хромосомные мутации случаются, если участки наследственной информации выпадают, удваиваются либо вообще переносятся и встраиваются в другую хромосому.
Геномные мутации наиболее серьезны . Их причина — это изменение числа хромосом. То есть когда вместо пары — диплоидного набора присутствует в кариотипе триплоидный набор.
Наиболее известный пример триплоидной мутации — это синдром Дауна, при котором личный набор хромосом 47. У таких детей образуется 3 хромосомы на месте 21-й пары.
Известна также такая мутация, как полиплодия. Но полиплодия встречается только у растений.
эукариотических и прокариотических хромосом
Поскольку прокариотические клетки обычно имеют только одну кольцевую хромосому, они могут реплицироваться быстрее, чем эукариотические клетки. Фактически, прокариотическая клетка может пройти два цикла репликации ДНК, прежде чем сама клетка разделится. Это означает, что репликация ДНК может происходить во время клеточного деления у прокариот.
Это означает, что репликация ДНК может происходить во время клеточного деления у прокариот.
Поскольку эукариотические клетки обычно имеют несколько линейных хромосом, покрытых теломерами, репликация эукариотической ДНК и деление клеток (митоз и мейоз) немного сложнее.В эукариотических клетках репликация ДНК происходит до начала митоза и не может происходить во время деления клетки. Кроме того, теломеры — повторяющиеся последовательности ДНК на концах каждой хромосомы — ограничивают число делений клетки, прежде чем она умрет или станет стареющей. Каждый раз, когда типичная или соматическая эукариотическая клетка делится, теломеры становятся короче.
2. Хромосомы эукариот расположены внутри ядра, тогда как хромосомы прокариот расположены в нуклеоиде.
Ключевое различие между прокариотическими и эукариотическими клетками заключается в том, что эукариотические клетки имеют мембраносвязанное ядро (и мембраносвязанные органеллы), тогда как прокариотические клетки лишены ядра.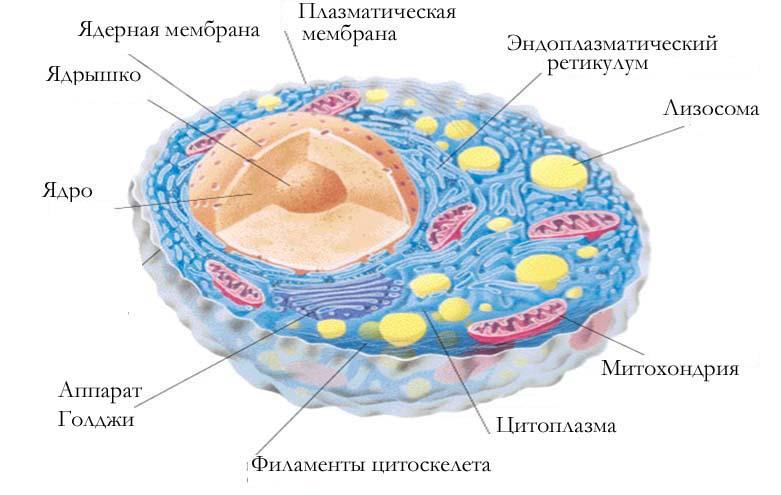 В эукариотических клетках все хромосомы содержатся в ядре. В прокариотических клетках хромосома расположена в области цитоплазмы, называемой нуклеоидом, в котором отсутствует мембрана.
В эукариотических клетках все хромосомы содержатся в ядре. В прокариотических клетках хромосома расположена в области цитоплазмы, называемой нуклеоидом, в котором отсутствует мембрана.
Одним интересным следствием этой разницы в расположении эукариотических и прокариотических хромосом является то, что транскрипция и трансляция — процессы создания молекулы РНК и использования этой молекулы для синтеза белка — могут происходить одновременно у прокариот. Это возможно, потому что у прокариотических клеток отсутствует ядерная мембрана, поэтому транскрипция и трансляция происходят в одном и том же регионе.Когда РНК транскрибируется, рибосомы могут начать процесс трансляции, связывая вместе аминокислоты. Напротив, в эукариотических клетках транскрипция всегда происходит первой и происходит внутри ядра. Молекула РНК должна пройти редактирование, прежде чем она покинет ядро. Затем трансляция осуществляется рибосомой в цитоплазме.
3. В эукариотических хромосомах ДНК наматывается на гистоновые белки, а затем дополнительно уплотняется за счет суперспирализации и фолдинга.
 В прокариотических хромосомах ДНК суперскручена и уплотнена белками, ассоциированными с нуклеоидами.
В прокариотических хромосомах ДНК суперскручена и уплотнена белками, ассоциированными с нуклеоидами.
Структура ДНК, репликация и структура эукариотического хроматина
Длина ДНК в ядре намного превышает размер компартмента. в котором он содержится. Чтобы поместиться в этот отсек, ДНК должна быть каким-то образом сконденсирован. Степень конденсации ДНК выражается как коэффициент упаковки.
Коэффициент упаковки — длина ДНК, деленная на длину, на которую он упакован
Например, самая короткая хромосома человека содержит 4.6 x 10 7 п.н. ДНК (примерно в 10 раз больше размера генома E. coli ). Это эквивалентно 14 000 мкм расширенной ДНК. В наиболее конденсированном состоянии во время митоза длина хромосомы около 2 мкм. Это дает коэффициент упаковки 7000 (14 000/2).
Для достижения общего коэффициента упаковки ДНК не упаковывается непосредственно в конечный продукт.
строение хроматина. Вместо этого он содержит несколько иерархий организации. Первый уровень упаковки достигается за счет наматывания ДНК на белок.
ядро для создания «шаровидной» структуры, называемой нуклеосомой .Этот
дает коэффициент упаковки около 6. Эта структура инвариантна как в
эухроматин и гетерохроматин всех хромосом. Второй уровень упаковки
представляет собой скручивание шариков в спиральную структуру, называемую волокном 30 нм .
обнаружен как в интерфазном хроматине, так и в митотических хромосомах. Этот
структура увеличивает коэффициент упаковки примерно до 40. Окончательная упаковка происходит
когда волокно организовано в виде петель, каркасов и доменов, которые дают окончательную
коэффициент упаковки около 1000 в интерфазных хромосомах и около 10000 в
митотических хромосом.
Первый уровень упаковки достигается за счет наматывания ДНК на белок.
ядро для создания «шаровидной» структуры, называемой нуклеосомой .Этот
дает коэффициент упаковки около 6. Эта структура инвариантна как в
эухроматин и гетерохроматин всех хромосом. Второй уровень упаковки
представляет собой скручивание шариков в спиральную структуру, называемую волокном 30 нм .
обнаружен как в интерфазном хроматине, так и в митотических хромосомах. Этот
структура увеличивает коэффициент упаковки примерно до 40. Окончательная упаковка происходит
когда волокно организовано в виде петель, каркасов и доменов, которые дают окончательную
коэффициент упаковки около 1000 в интерфазных хромосомах и около 10000 в
митотических хромосом.
Хромосомы эукариот состоят из ДНК-белкового комплекса, организованного компактно, что позволяет хранить большое количество ДНК в ядро клетки. Обозначение субъединицы хромосомы – хроматин. Основной единицей хроматина является нуклеосома.
Хроматин — единица анализа хромосомы; хроматин отражает общая структура хромосомы, но не уникальна для какой-либо конкретной хромосома
Нуклеосома — простейшая упаковочная структура ДНК, обнаруженная в
все эукариотические хромосомы; ДНК обернута вокруг октамера небольших основных
белки, называемые гистонами; 146 п. н. обернуты вокруг ядра, а остальные
основания связываются со следующей нуклеосомой; эта структура вызывает отрицательную суперспирализацию
н. обернуты вокруг ядра, а остальные
основания связываются со следующей нуклеосомой; эта структура вызывает отрицательную суперспирализацию
Нуклеосома состоит примерно из 200 п.н., обернутых вокруг гистонового октамера. который содержит две копии гистоновых белков h3A, h3B, h4 и h5.Эти известные как коровые гистоны. Гистоны – это основные белки, обладающие сродством для ДНК и являются наиболее распространенными белками, связанными с ДНК. Амино кислотная последовательность этих четырех гистонов сохраняется, что предполагает аналогичную функцию для всех.
Длина ДНК, связанной с нуклеосомной единицей, колеблется в пределах
разновидность. Но независимо от размера задействованы два компонента ДНК. Ядро
ДНК — это ДНК, которая фактически связана с октамером гистонов.Это значение является инвариантным и составляет 146 пар оснований. Ядро ДНК образует две петли
вокруг октамера, и это позволяет двум областям, отстоящим друг от друга на 80 п.н.,
оказаться в непосредственной близости. Таким образом, две последовательности, находящиеся далеко друг от друга, могут
взаимодействуют с одним и тем же регуляторным белком, чтобы контролировать экспрессию генов. То
ДНК, которая находится между каждым октамером гистонов, называется линкерной ДНК .
и может варьироваться по длине от 8 до 114 пар оснований. Эта вариация является видом
специфичны, но вариации длины линкерной ДНК также связаны с
стадии развития организма или отдельных участков генома.
Таким образом, две последовательности, находящиеся далеко друг от друга, могут
взаимодействуют с одним и тем же регуляторным белком, чтобы контролировать экспрессию генов. То
ДНК, которая находится между каждым октамером гистонов, называется линкерной ДНК .
и может варьироваться по длине от 8 до 114 пар оснований. Эта вариация является видом
специфичны, но вариации длины линкерной ДНК также связаны с
стадии развития организма или отдельных участков генома.
Следующим уровнем организации хроматина является волокно 30 нм. Этот
по-видимому, представляет собой соленоидную структуру с примерно 6 нуклеосомами на виток. Этот
дает коэффициент упаковки 40, что означает, что каждый 1 мкм вдоль оси
содержит 40 мкм ДНК. Стабильность этой структуры требует
присутствие последнего члена семейства генов гистонов, гистона h2. Так как
эксперименты, которые удаляют h2 из хроматина, сохраняют нуклеосому, но не
структуры 30 нм был сделан вывод, что h2 важен для стабилизации
структуры 30 нм.
Конечный уровень упаковки характеризуется видимой структурой 700 нм. в метафазной хромосоме. Конденсированный участок хроматина имеет характерная каркасная структура, которую можно обнаружить в метафазе хромосомы. Это, по-видимому, является результатом обширного зацикливания ДНК. в хромосоме.
Последние определения, которые необходимо представить, это эухроматин и гетерохроматин . При окраске хромосом красителями они выглядят как чередование светло- и темноокрашенных участков.Слабо окрашенные участки являются эухроматиновыми и содержат однокопийную генетически активную ДНК. То темные участки представляют собой гетерохроматин и содержат повторяющиеся последовательности которые генетически неактивны.
Центромеры и теломеры
Центромеры и теломеры являются двумя важными характеристиками всех эукариотических хромосом. Каждый из них обеспечивает уникальную функцию, абсолютно необходимую для стабильности хромосомы. Центромеры необходимы для сегрегации центромер во время
йоз и митоз, а телеомеры обеспечивают терминальную стабильность хромосомы и обеспечивают ее выживание.
Центромеры — это те конденсированные области внутри хромосомы, которые отвечают за точную сегрегацию реплицированной хромосомы во время митоза и мейоза. Когда хромосомы окрашиваются, они обычно показывают темную окрашенную область, которая является центромерой. Во время митоза центромера, общая для сестринских хроматид, должна делиться, чтобы хроматиды могли мигрировать в противоположные полюса клетки. С другой стороны, во время первого мейотического деления центромера сестринских хроматид должна оставаться интактной, тогда как при мейозе II они должны действовать так же, как во время митоза.Следовательно, центромера представляет собой важный компонент структуры хромосом и сегрегации.
В районе центромеры у большинства видов есть несколько мест, где прикрепляются волокна веретена деления, и эти участки состоят из ДНК и белка. Фактическое место, где происходит прикрепление, называется кинетохорой и состоит из ДНК
и белок. Последовательность ДНК в этих участках называется
CEN ДНК . Поскольку ДНК CEN может быть перемещена из одной хромосомы в другую и при этом обеспечить способность хромосомы к сегрегации, эти последовательности не должны выполнять какую-либо другую функцию.
Поскольку ДНК CEN может быть перемещена из одной хромосомы в другую и при этом обеспечить способность хромосомы к сегрегации, эти последовательности не должны выполнять какую-либо другую функцию.
Обычно ДНК CEN имеет длину около 120 пар оснований и состоит из нескольких субдоменов, CDE-I , CDE-II и CDE-III . Мутации в первых двух субдоменах не влияют на сегрегацию, но точечная мутация в субдомене CDE-III com полностью устраняет способность центромеры функционировать при сегрегации хромосом. Следовательно, CDE-III д. активно участвовать в связывании волокон веретена деления с центромерой.
Белковый компонент кинетохора только сейчас охарактеризован.Комплекс из трех белков, названный Cbf-III , связывается с нормальными областями CDE-III, но не может связываться с областью CDE-III с точечной мутацией, предотвращающей митотическое расщепление. Шерсть
Кроме того, мутации генов, кодирующих белки Cbf-III, также устраняют способность хромосом к сегрегации во время митоза. Для полного понимания механики необходимы дополнительные анализы ДНК и белковых компонентов центромеры.
сегрегации хромосом.
Для полного понимания механики необходимы дополнительные анализы ДНК и белковых компонентов центромеры.
сегрегации хромосом.
Теломеры представляют собой участок ДНК на конце линейного эукариотического хромосомы, которые необходимы для репликации и стабильности хромосомы.МакКлинток узнала их особые черты, когда заметила, что если двое хромосомы были разорваны в клетке, конец одной мог прикрепиться к другой и наоборот. Чего она никогда не замечала, так это прикрепления сломанного конец к концу неразорванной хромосомы. Таким образом, концы сломанных хромосом липкие, тогда как нормальный конец не липкий, что предполагает концы хромосомы имеют уникальные особенности. Обычно, но не всегда, теломерная ДНК гетерохроматична и содержит прямые тандемно повторяющиеся последовательности.То В следующей таблице показаны повторяющиеся последовательности нескольких видов. Часто они имеют вид (T/A) x G y , где x находится в диапазоне от 1 до 4, а y больше 1.
теломер повторяющихся последовательностей
| Виды | Повтор последовательности |
|---|---|
| арабидопсиса | TTTAGGG |
| Человека | TTAGGG |
| Oxytricha | TTTTGGGG |
| слизевики | Taggg |
| Tetrahymena | TTGGGG | Trypanosome | Taggg |
| Дрожжи 7 | (TG) 1-3 TG 2-3 |
Обратите внимание, что количество последовательностей TG и количество цитозинов в последовательности дрожжей варьируется.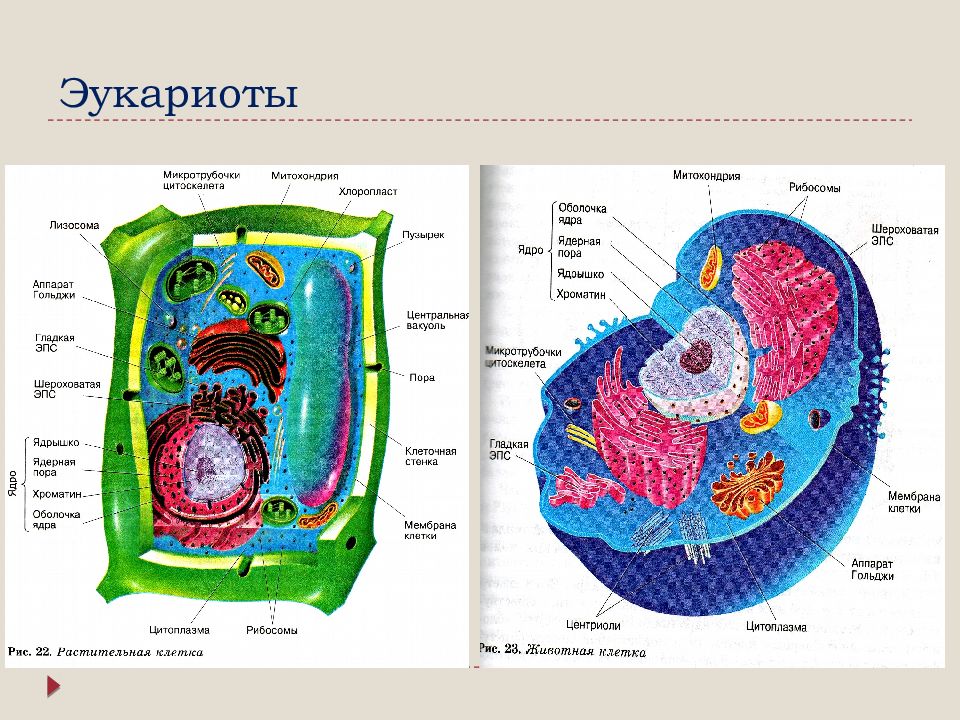 По крайней мере, для дрожжей было показано, что разные штаммы содержат телеомеры разной длины и что эта длина находится под генетическим контролем.
По крайней мере, для дрожжей было показано, что разные штаммы содержат телеомеры разной длины и что эта длина находится под генетическим контролем.
Основная трудность с теломерами заключается в репликации отстающей нити. Поскольку для синтеза ДНК требуется матрица РНК (которая обеспечивает свободный 3′-ОН группа) для запуска репликации ДНК, и эта матрица в конечном итоге деградирует, на конце хромосомы останется короткая одноцепочечная область. Этот регион будет восприимчив к ферментам, разрушающим одноцепочечную ДНК.То результатом будет то, что длина хромосомы будет укорачивается после каждого деления. Но этого не видно.
Действие ферментов теломеразы обеспечивает правильную репликацию концов отстающих цепей. Хорошо изученная система включает простейший организм Tetrahymena . Теломеры этого организма заканчиваются последовательностью 5′-TTGGGG-3′. Т
Теломераза добавляет серию 5′-TTGGGG-3′ повторов к концам отстающей нити. Шпилька возникает, когда между остатками гуанина в повторяющейся форме образуются необычные пары оснований. Затем РНК-праймер удаляется, и можно использовать 5′-конец отстающей цепи.
для синтеза ДНК. Перевязка происходит между готовой отстающей прядью и шпилькой. Наконец, шпилька удаляется на повторе 5′-TTGGGG-3′. Таким образом, конец хромосомы точно реплицируется. На следующем рисунке показаны эти шаги.
Затем РНК-праймер удаляется, и можно использовать 5′-конец отстающей цепи.
для синтеза ДНК. Перевязка происходит между готовой отстающей прядью и шпилькой. Наконец, шпилька удаляется на повторе 5′-TTGGGG-3′. Таким образом, конец хромосомы точно реплицируется. На следующем рисунке показаны эти шаги.
Репликация теломер
Анализ последовательностей ДНК в геномах эукариот
Метод, который используется для определения сложности последовательности любого генома.
включает денатурацию и ренатурацию ДНК.ДНК денатурирована
путем нагревания, которое плавит Н-связи и делает ДНК одноцепочечной. Если
ДНК быстро охлаждается, ДНК остается одноцепочечной. Но если ДНК
дают медленно остыть, комплементарные последовательности найдут каждую
другой и, в конечном итоге, снова пара оснований. Скорость повторного отжига ДНК
(другой термин для ренатурации) является функцией вида, ДНК которого
был изолирован. Ниже представлена кривая, полученная из простого генома.
Ось Y представляет собой процент ДНК, которая остается одноцепочечной.Это выражается как отношение концентрации одноцепочечной ДНК ( C ) к общей концентрации исходной ДНК ( Co ). Ось X логарифмическая шкала произведения исходной концентрации ДНК (в молях/литр) умножить на время протекания реакции (в секундах). Обозначение для этого значения Детская кроватка и называется значением «Детская кроватка». Сама кривая называется кривой «Кот». Как видно, кривая довольно гладкая, что указывает на то, что повторный отжиг происходит медленно, но постепенно в течение время.Одним из полезных значений является Cot½ , Значение Cot , при котором половина ДНК подверглась повторному отжигу.
шагов, связанных с экспериментами по денатурации и ренатурации ДНК
1. Разрезать ДНК до размера около 400 п.н.
2. Денатурировать ДНК нагреванием до 100 o C.
3. Медленно охладить и отобрать образцы через разные промежутки времени.
4. Определите % одноцепочечной ДНК в каждый момент времени.
Форма кривой «Cot» для данного вида зависит от двух факторов:
- размер или сложность генома; и
- количество повторяющихся ДНК в геноме
Если мы построим кривые «Кот» генома трех видов, таких как бактериофаг
лямбда, Е.coli и дрожжей мы увидим, что они имеют одинаковую форму,
но Cot½ дрожжей будет самым большим, E. coli следующим
и лямбда наименьшая. Физически, чем больше размер генома, тем дольше он
потребуется, чтобы любая последовательность встретила свою комплементарную последовательность в
решение. Это связано с тем, что две комплементарные последовательности должны встретиться
друг друга, прежде чем они смогут соединиться. Чем сложнее геном, тем
чем больше уникальных последовательностей доступно, тем больше времени потребуется для любого
две дополнительные последовательности, чтобы встретиться друг с другом и спариться.Учитывая подобное
концентрации в растворе, то для более сложных видов потребуется больше времени
чтобы достичь Cot½ .
Повторяющиеся последовательности ДНК, последовательности ДНК, которые встречаются более одного раза в геном вида , оказывает заметное влияние на кривые «Кот». Если конкретная последовательность дважды представлена в геноме, она будет иметь два комплементарные последовательности для сопряжения и, как таковые, будут иметь половинное значение Cot такой большой, как последовательность, представленная только один раз в геноме.
Эукариотические геномы на самом деле имеют широкий спектр последовательностей, которые представлены
на разных уровнях повторения. Однокопийные последовательности найдены один раз
или несколько раз в геноме. Многие из последовательностей, которые кодируют функциональные
гены попадают в этот класс. Средние повторяющиеся ДНК обнаружены из 10 —
1000 раз в геноме. Примеры из них включают рРНК и тРНК
гены и запасные белки в растениях, таких как кукуруза. Средняя повторяющаяся ДНК
может варьироваться от 100-300 п.н. до 5000 п. н. и может быть разбросан по всему геному.
Наиболее распространенные последовательности обнаружены в высоко повторяющихся ДНК
класс.Эти последовательности встречаются в геноме от 100 000 до 1 миллиона раз.
и может иметь размер от нескольких до нескольких сотен оснований в длину. Эти
последовательности находятся в областях хромосомы, таких как гетерохроматин,
центромеры и теломеры и, как правило, располагаются в виде тандемных повторов. То
Ниже приведен пример тандемно повторяющейся последовательности:
н. и может быть разбросан по всему геному.
Наиболее распространенные последовательности обнаружены в высоко повторяющихся ДНК
класс.Эти последовательности встречаются в геноме от 100 000 до 1 миллиона раз.
и может иметь размер от нескольких до нескольких сотен оснований в длину. Эти
последовательности находятся в областях хромосомы, таких как гетерохроматин,
центромеры и теломеры и, как правило, располагаются в виде тандемных повторов. То
Ниже приведен пример тандемно повторяющейся последовательности:
АТТАТА АТТАТА АТТАТА // АТТАТА
Геномы, которые содержат эти разные классы последовательностей, повторно отжигаются в другом
образом, чем геномы только с одной копией последовательностей.Вместо того, чтобы иметь
единая гладкая кривая «Детская кроватка», можно увидеть три отдельные кривые, каждая из которых представляет
другой класс повторения. Первыми повторными отжигами являются высоко
повторяющиеся последовательности, потому что в геноме существует так много их копий,
и потому что они имеют низкую сложность последовательности.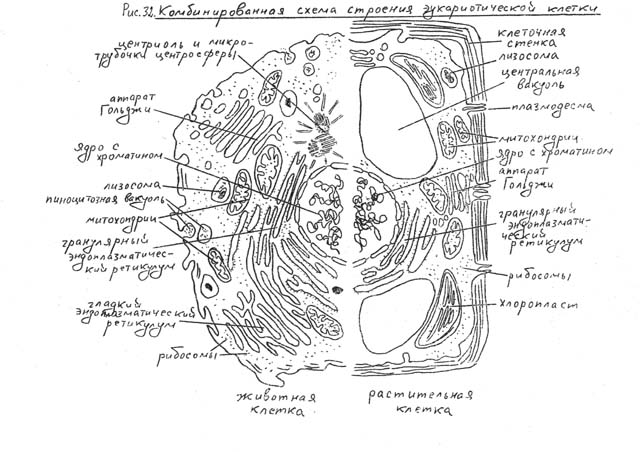 Вторая часть
геном для повторного отжига — это средняя повторяющаяся ДНК, а последняя часть — для повторного отжига.
reanneal — это единственная копия ДНК. На следующей схеме изображена «Детская кроватка».
кривая для «типичного» эукариотического генома
Вторая часть
геном для повторного отжига — это средняя повторяющаяся ДНК, а последняя часть — для повторного отжига.
reanneal — это единственная копия ДНК. На следующей схеме изображена «Детская кроватка».
кривая для «типичного» эукариотического генома
В следующей таблице показано распределение последовательностей для выбранных видов.
| Виды | Распределение последовательности |
|---|---|
| Бактерии | 99,7% Один экземпляр |
| Мышь | 60 % Один экземпляр 25 % Среднее повторение 10 % Многократное повторение |
| Человек | 70 % Однократное копирование 13 % Среднее повторение 8 % Многократное повторение |
| Хлопок | 61 % Однократное копирование 27 % Среднее повторение 8 % Многократное повторение |
| Кукуруза | 30 % Один экземпляр 40 % Среднее повторение 20 % Многократное повторение |
| Пшеница | 10 % Однократное копирование 83 % Среднее повторение 4 % Многократное повторение |
| Арабидопсис | 55 % Однократное копирование 27 % Среднее повторение 10 % Многократное повторение |
Вставка последовательности
Несмотря на то, что геномы высших организмов содержат одну копию, средние
повторяющиеся и сильно повторяющиеся последовательности ДНК, эти последовательности не упорядочены
одинаково у всех видов. Выдающееся расположение называется короткий период
вставка . Это расположение характеризуется повторяющимися последовательностями
100-200 п.н. в длину, вкрапленные среди последовательностей с одной копией, которые составляют 1000-2000
б.п. в длину. Такое расположение встречается у животных, грибов и растений.
Выдающееся расположение называется короткий период
вставка . Это расположение характеризуется повторяющимися последовательностями
100-200 п.н. в длину, вкрапленные среди последовательностей с одной копией, которые составляют 1000-2000
б.п. в длину. Такое расположение встречается у животных, грибов и растений.
Второй тип расположения — длиннопериодные вкрапления . Этот характеризуется участками 5000 п.н. повторяющихся последовательностей, перемежающихся в областях из 35 000 п.н. однокопийной ДНК. Дрозофила является примером вида с таким необычным расположением последовательностей. В обоих случаях повторяющиеся последовательности обычно относятся к среднему повторяющемуся классу. Мы обсудили выше, где обнаружены часто повторяющиеся последовательности.
Кариотип эукариотической хромосомы
В то время как бактерии имеют только одну хромосому, эукариотические виды имеют
хотя бы одну пару хромосом. Большинство из них имеют более одной пары. Другой актуальный
Дело в том, что эукариотические хромосомы обнаруживаются только во время клеточного
деления, а не на всех стадиях клеточного цикла.Они в своих самых
конденсированная форма во время метафазы, когда сестринские хроматиды присоединяются.
Это первичный этап, когда выполняется цитогенетический анализ.
Другой актуальный
Дело в том, что эукариотические хромосомы обнаруживаются только во время клеточного
деления, а не на всех стадиях клеточного цикла.Они в своих самых
конденсированная форма во время метафазы, когда сестринские хроматиды присоединяются.
Это первичный этап, когда выполняется цитогенетический анализ.
Каждый вид характеризуется кариотипом . Кариотип – это описание числа хромосом в нормальной диплоидной клетке, а также как их распределение по размерам. Например, в хромосоме человека 23 пары. хромосом, 22 соматические пары и одна пара половых хромосом. Один важный Одним из аспектов генетического исследования является сопоставление изменений кариотипа с изменениями в фенотипе особи.
Одним из важных аспектов генетики является корреляция изменений кариотипа с
изменения фенотипа. Например, люди, у которых есть дополнительная хромосома 21.
есть синдром Дауна. Инсерции, делеции и изменения числа хромосом
могут быть обнаружены опытным цитогенетиком, но коррелируя их с
конкретных фенотипов трудно.
Первым отличительным параметром при разработке кариотипа является размер и число хромосом. Хотя это и полезно, но не дает достаточно подробно, чтобы начать разработку корреляции между структурой и функция (фенотип).Чтобы еще больше различать хромосомы, они обрабатывают красителем, окрашивающим ДНК воспроизводимым образом. После окрашивание, некоторые области окрашены слабо, а другие сильно окрашенный. Как описано выше, слабоокрашенные участки называются эухроматин , а окрашенная в темный цвет область называется гетерохроматин . В настоящее время выбран краситель по Гимзе , и полученный шаблон называется шаблоном G-полосы.
Парадокс C-значения
Помимо описания генома организма по количеству хромосом,
он также описывается количеством ДНК в гаплоидной клетке.Обычно это
выражается как количество ДНК на гаплоидную клетку (обычно выражается в пикограммах)
или количество тысяч оснований на гаплоидную клетку и называется значением C. Один
непосредственная особенность эукариотических организмов выдвигает на первый план специфическую аномалию, которая
был обнаружен в начале молекулярных исследований. Несмотря на то, что эукариотические организмы
имеют в 2-10 раз больше генов, чем прокариоты, у них много порядков
на порядок больше ДНК в клетке. Кроме того, количество ДНК на геном
коррелирует не с предполагаемой эволюционной сложностью вида.Это сформулировано как парадокс значения C: количество ДНК в гаплоидном
клетка организма не связана с его эволюционной сложностью. (Другой
Важно помнить, что нет никакой связи между
число хромосом и предполагаемая эволюционная сложность
организм.)
Один
непосредственная особенность эукариотических организмов выдвигает на первый план специфическую аномалию, которая
был обнаружен в начале молекулярных исследований. Несмотря на то, что эукариотические организмы
имеют в 2-10 раз больше генов, чем прокариоты, у них много порядков
на порядок больше ДНК в клетке. Кроме того, количество ДНК на геном
коррелирует не с предполагаемой эволюционной сложностью вида.Это сформулировано как парадокс значения C: количество ДНК в гаплоидном
клетка организма не связана с его эволюционной сложностью. (Другой
Важно помнить, что нет никакой связи между
число хромосом и предполагаемая эволюционная сложность
организм.)
C Значение организмов, используемых в генетических исследованиях
| Разновидность | Килобазы / гаплоидный геном |
|---|---|
| Э.кишечная палочка |
4,5 x 10 3 |
| Человек |
3,0 x 10 6 |
| Дрозофила |
1,7 x 10 5 |
| Кукуруза |
2,0 x 10 6 |
| Арибидопсис |
7. Геном — полный набор хромосом, унаследованный от одного родителя; полный компонент ДНК человека; определение часто исключает органеллы Copyright © 1997. Phillip McClean Структура эукариотической хромосомы | Science Primer Хромосомы * s представляют собой длинные нити ДНК Во время клеточного деления хромосомы эукариот конденсируются в высокоскрученные структуры. Плотная спираль и высокая степень организации этой сверхспиральной ДНК способствуют правильной сегрегации во время митоза и клеточного деления.На следующем рисунке показаны форма, классификация и особенности эукариотической хромосомы. Состояние: Номер: Тип: Этикетка: Видимые характеристики:
Самые ранние попытки описать хромосомы были основаны на видимых характеристиках.
В конце клеточного цикла, до деления клетки, происходит суперспирализация ДНК. На этом этапе клеточного цикла клетка содержит две полные копии каждой хромосомы. Каждая сверхскрученная копия хромосомы называется хроматидой К концу клеточного цикла сестринские хроматиды сходятся и прикрепляются к центромере, образуя характерную Х-образную структуру. Когда клетки делятся во время митоза, каждая дочерняя клетка получает по одной сестринской хроматиде от каждой хромосомы. Положение центромеры является основой системы классификации хромосом. Различные классы:
 Он играет важную роль в делении клеток как место расположения кинетохора. Кинетохора является точкой прикрепления волокон, которые раздвигают сестринские хроматиды во время митоза. Он играет важную роль в делении клеток как место расположения кинетохора. Кинетохора является точкой прикрепления волокон, которые раздвигают сестринские хроматиды во время митоза.
Линейная природа эукариотических хромосом создает проблемы для репликации, потому что ДНК-полимераза * * требует праймирующей последовательности для удлинения цепи ДНК. Без каких-либо механизмов сохранения концов линейные нити ДНК в хромосомах эукариот становились бы короче каждый раз, когда клетка делится, что приводило бы к потере генетической информации.
ДНК эукариотических клеток становится суперскрученной во время клеточного деления и должна быть раскручена (или расслаблена) для осуществления транскрипции и репликации.
Обзор видео: Хромосомы и хроматин — Клетка Геномы большинства эукариот не только намного сложнее, чем у прокариот, но и ДНК эукариотических клеток организована иначе, чем у прокариотических клеток. Геномы прокариот содержатся в одиночных хромосомах, которые обычно представляют собой кольцевые молекулы ДНК. Напротив, геномы эукариот состоят из множества хромосом, каждая из которых содержит линейную молекулу ДНК.Хотя количество и размеры хромосом значительно различаются у разных видов (), их основная структура одинакова у всех эукариот. Таблица 4.2Числа хромосом эукариотических клеток. Хроматин Комплексы между эукариотической ДНК и белками называются хроматином, который обычно содержит примерно в два раза больше белка, чем ДНК. Основными белками хроматина являются гистоны — небольшие белки, содержащие большое количество основных аминокислот (аргинин и лизин), которые облегчают связывание с отрицательно заряженной молекулой ДНК.Существует пять основных типов гистонов, называемых h2, h3A, h3B, h4 и h5, которые очень похожи у разных видов эукариот (12). Основная структурная единица хроматина, нуклеосома, была описана Роджером Корнбергом в 1974 г. (). Два типа экспериментов привели Корнберга к предложению нуклеосомной модели.Во-первых, было обнаружено, что частичное расщепление хроматина микрококковой нуклеазой (ферментом, расщепляющим ДНК) дает фрагменты ДНК длиной примерно 200 пар оснований. Рисунок 4.8 Организация хроматина в нуклеосомах. (A) ДНК обернута вокруг гистонов в ядрах нуклеосом и запечатана гистоном h2.Негистоновые белки связываются с линкерной ДНК между частицами ядра нуклеосомы. (B) Гель-электрофорез фрагментов ДНК (подробнее. Было обнаружено, что более интенсивное расщепление хроматина микрококковой нуклеазой приводит к образованию частиц (называемых частицами ядра нуклеосомы ), которые соответствуют гранулам, видимым с помощью электронной микроскопии. Детальный анализ этих частиц показал, что они содержат 146 пар оснований ДНК, обернутых 1,65 раза вокруг гистонового ядра, состоящего из двух молекул h3A, h3B, h4 и h5 (коровые гистоны) (2).Одна молекула пятого гистона, h2, связывается с ДНК, когда входит в каждую частицу ядра нуклеосомы. Это формирует субъединицу хроматина, известную как хроматосома, которая состоит из 166 пар оснований ДНК, обернутых вокруг ядра гистона и удерживаемых на месте с помощью h2 (линкерный гистон). Рисунок 4.9 Структура хромосомы. (A) Частица ядра нуклеосомы состоит из 146 пар оснований ДНК, обернутых на 1,65 витка вокруг гистонового октамера, состоящего из двух молекул h3A, h3B, h4 и h5.Хроматосома содержит два полных витка ДНК (166 оснований (подробнее. Упаковка ДНК гистонами дает хроматиновое волокно диаметром примерно 10 нм, состоящее из хроматосом, разделенных линкерными сегментами ДНК, в среднем около 80 пар оснований в длина (). В электронном микроскопе это 10-нм волокно имеет вид бусинок, что наводит на мысль о нуклеосомной модели. Упаковка ДНК в такое 10-нм хроматиновое волокно сокращает его длину примерно в шесть раз. Затем хроматин может быть дополнительно конденсирован путем скручивания в 30-нм волокна, структуру которых еще предстоит определить.Взаимодействия между молекулами гистона h2, по-видимому, играют важную роль на этой стадии конденсации хроматина. Рисунок 4.10Хроматиновые волокна. Упаковка ДНК в нуклеосомы дает хроматиновое волокно диаметром около 10 нм. Хроматин далее конденсируется путем скручивания в волокно длиной 30 нм, содержащее около шести нуклеосом на виток. (Фотографии предоставлены Адой (подробнее…) Степень конденсации хроматина меняется в течение жизненного цикла клетки. Рисунок 4.11 Интерфазный хроматин. Электронная микрофотография интерфазного ядра. Эухроматин распределен по всему ядру. Гетерохроматин обозначен стрелками, а ядрышко стрелкой. (С любезного разрешения Ады Л. Олинс и Дональда Э. Олинса, Дуб (подробнее.. В отличие от эухроматина около 10% интерфазного хроматина (называемого гетерохроматином) находится в очень сильно конденсированном состоянии, которое напоминает хроматин клеток подвергается митозу.Гетерохроматин транскрипционно неактивен и содержит сильно повторяющиеся последовательности ДНК, такие как те, которые присутствуют в центромерах и теломерах. Когда клетки вступают в митоз, их хромосомы становятся сильно конденсированными, чтобы их можно было распределить по дочерним клеткам. Считается, что петли 30-нм хроматиновых волокон складываются дальше друг от друга, образуя компактные метафазные хромосомы митотических клеток, в которых ДНК конденсируется почти в 10 000 раз (11). Такой конденсированный хроматин больше не может использоваться в качестве матрицы для синтеза РНК, поэтому транскрипция прекращается во время митоза.Электронные микрофотографии показывают, что ДНК в метафазных хромосомах организована в виде больших петель, прикрепленных к белковому каркасу (), но в настоящее время мы не понимаем ни детальной структуры этого высококонденсированного хроматина, ни механизма конденсации хроматина. Рисунок 4.12Конденсация хроматина во время митоза. Сканирующая электронная микрофотография метафазных хромосом. Добавлен искусственный цвет. (Biophoto Associates/Photo Researchers Inc.) Рисунок 4.13Строение метафазных хромосом. Электронная микрофотография петель ДНК, прикрепленных к белковому каркасу метафазных хромосом, лишенных гистонов. (Из J. R. Paulson and U. K. Laemmli, 1977. Cell 12: 817.) Метафазные хромосомы настолько сильно конденсированы, что их морфологию можно изучать с помощью светового микроскопа (). Несколько методов окрашивания дают характерные образцы чередующихся светлых и темных полос хромосом, которые являются результатом предпочтительного связывания красителей или флуоресцентных красителей с последовательностями ДНК, богатыми AT, а не GC.Эти полосы специфичны для каждой хромосомы и, по-видимому, представляют отдельные участки хромосомы. Гены могут быть локализованы в определенных участках хромосом с помощью гибридизации in situ , что указывает на то, что упаковка ДНК в метафазные хромосомы является высокоупорядоченным и воспроизводимым процессом. Рисунок 4.14Метафазные хромосомы человека. (A) Световая микрофотография хромосом человека, распространяющихся из метафазной клетки. (B) Хромосомы человека расположены парами, пронумерованными от наибольшей (хромосома 1) к наименьшей.Показанные хромосомы принадлежат женщине, поэтому (далее…) Центромеры Центромера — это специализированный участок хромосомы, играющий критическую роль в обеспечении правильного распределения дуплицированных хромосом по дочерним клеткам во время митоза () . Клеточная ДНК реплицируется во время интерфазы, что приводит к образованию двух копий каждой хромосомы до начала митоза. Когда клетка вступает в митоз, конденсация хроматина приводит к образованию метафазных хромосом, состоящих из двух идентичных сестринских хроматид.Эти сестринские хроматиды удерживаются вместе на центромере, которая рассматривается как суженный участок хромосомы. По мере митоза микротрубочки митотического веретена прикрепляются к центромере, а две сестринские хроматиды расходятся и расходятся к противоположным полюсам веретена. Рис. 4.15Хромосомы во время митоза.Поскольку ДНК реплицируется во время интерфазы, клетка содержит две идентичные дублированные копии каждой хромосомы до вступления в митоз. Таким образом, центромеры служат как местами ассоциации сестринских хроматид, так и местами прикрепления микротрубочек митотического веретена. Они состоят из специфических последовательностей ДНК, с которыми связывается ряд белков, связанных с центромерами, образуя специализированную структуру, называемую кинетохорой (4). Связывание микротрубочек с кинетохорными белками опосредует прикрепление хромосом к митотическому веретену.Белки, связанные с кинетохорой, затем действуют как «молекулярные моторы», которые управляют движением хромосом вдоль волокон веретена деления, разделяя хромосомы на дочерние ядра. Рис. 4.16Центромера метафазной хромосомы. Центромера — это область, в которой две сестринские хроматиды остаются прикрепленными в метафазе. Специфические белки связываются с центромерной ДНК, образуя кинетохору, которая является местом прикрепления нитей веретена деления. (подробнее…) Последовательности центромерной ДНК лучше всего определены у дрожжей, где их функцию можно оценить, проследив расхождение плазмид при митозе ().Плазмиды, содержащие функциональные центромеры, сегрегируют подобно хромосомам и равномерно распределяются между дочерними клетками после митоза. Однако в отсутствие функциональной центромеры плазмида не сегрегирует должным образом, и многие дочерние клетки не могут наследовать плазмидную ДНК. Анализы этого типа позволили определить последовательности, необходимые для функции центромер. Такие эксперименты впервые показали, что центромерные последовательности хорошо изученных дрожжей Saccharomyces cerevisiae содержат примерно 125 пар оснований, состоящих из трех элементов последовательности: двух коротких последовательностей из 8 и 25 пар оснований, разделенных 78-86 парами оснований очень АТ. Рисунок 4.17Анализ центромеры дрожжей. Обе показанные плазмиды содержат селектируемый маркер ( LEU2 ) и последовательности ДНК, которые служат источником репликации в дрожжах (ARS, что означает автономно реплицирующуюся последовательность). Однако в плазмиде I отсутствует центромера и (далее…) Рис. (A) Последовательностицентромер S. cerevisiae (CEN) состоят из двух коротких консервативных последовательностей (CDEI и CDEIII), разделенных 78–86 парами оснований (bp) богатой AT ДНК (CDEII).Последовательности (подробнее…) Короткие центромерные последовательности, определенные у S. cerevisiae , однако, похоже, не отражают ситуацию у других эукариот. Более поздние исследования определили центромеры делящихся дрожжей Schizosaccharomyces pombe с помощью аналогичного функционального подхода. Хотя S. cerevisiae и S. pombe оба являются дрожжами, они, по-видимому, так же отличаются друг от друга, как и от человека, и сильно отличаются во многих аспектах своей клеточной биологии. Исследования хромосомы дрозофилы предоставили первую характеристику центромеры у высших эукариот (). Центромера Drosophila охватывает 420 т.п.н., большая часть которых (более 85%) состоит из двух сильно повторяющихся сателлитных ДНК с последовательностями AATAT и AAGAG. Остальная часть центромеры состоит из вкраплений мобильных элементов, которые также обнаруживаются в других участках генома Drosophila , в дополнение к неповторяющейся области AT-богатой ДНК. Центромеры человека и других млекопитающих еще не определены функциональными исследованиями, но они были идентифицированы по связыванию белков, ассоциированных с центромерами. Центромеры млекопитающих характеризуются обширными областями гетерохроматина, состоящими из высоко повторяющихся последовательностей сателлитной ДНК.У людей и других приматов первичной центромерной последовательностью является α-сателлитная ДНК, которая представляет собой последовательность из 171 пары оснований, организованную в виде тандемных повторов, охватывающих до миллионов пар оснований. α-сателлитная ДНК, по-видимому, играет роль в структуре и функции центромер, поскольку было обнаружено, что она связывает белки, ассоциированные с центромерами. Теломеры Последовательности на концах эукариотических хромосом, называемые теломерами, играют важную роль в репликации и поддержании хромосом. Первоначально теломеры были признаны отдельными структурами, потому что сломанные хромосомы были очень нестабильны в эукариотических клетках, что означает, что на нормальных концах хромосом необходимы специфические последовательности. Впоследствии это было продемонстрировано в опытах, в которых к концам линейных молекул плазмидной ДНК дрожжей добавлялись теломеры простейших Tetrahymena .Добавление этих последовательностей теломерной ДНК позволило этим плазмидам реплицироваться в виде линейных хромосомоподобных молекул у дрожжей, прямо демонстрируя, что теломеры необходимы для репликации линейных молекул ДНК. Последовательности ДНК теломер различных эукариот сходны и состоят из повторов простой последовательности ДНК, содержащей кластеры остатков G на одной цепи (). Например, последовательность теломерных повторов у человека и других млекопитающих — AGGGTT, а теломерный повтор Tetrahymena — GGGGTT.Эти последовательности повторяются сотни или тысячи раз, охватывая, таким образом, несколько тысяч оснований, и заканчиваются выступом одноцепочечной ДНК. Недавние результаты свидетельствуют о том, что повторяющиеся последовательности теломерной ДНК образуют петли на концах хромосом, тем самым защищая концы хромосом от деградации (4). Рисунок 4.19Структура теломеры. Теломерная ДНК закручивается сама на себя, образуя кольцевую структуру, которая защищает концы хромосом. Теломеры играют решающую роль в репликации концов линейных молекул ДНК (см. главу 5).ДНК-полимераза способна удлинять растущую цепь ДНК, но не может инициировать синтез новой цепи на конце линейной молекулы ДНК. Структура и функция хроматина Хроматин представляет собой комплекс макромолекул, состоящий из ДНК, РНК и белка, который находится внутри ядра эукариотических клеток. Хроматин существует в двух формах: гетерохроматин (конденсированный) и эухроматин (расширенный). Первичными белковыми компонентами хроматина являются гистоны, которые помогают организовать ДНК в «шаровидные» структуры, называемые нуклеосомами, обеспечивая основу, на которую может быть намотана ДНК. Нуклеосома состоит из 147 пар оснований ДНК, которые обернуты вокруг набора из 8 гистонов, называемых октомерами. Хроматин, хромосомы и хроматиды Люди часто путают эти три термина: хроматин, хромосома и хроматида. Хотя все эти три структуры состоят из ДНК и белков внутри ядра, каждая из них имеет уникальное определение. Как упоминалось выше, хроматин состоит из ДНК и гистонов, упакованных в тонкие нитевидные волокна. Хроматин подвергается дальнейшей конденсации с образованием хромосомы. Таким образом, хроматин представляет собой более низкий порядок организации ДНК, а хромосомы — более высокий порядок организации ДНК. Хромосомы представляют собой одноцепочечные группы конденсированного хроматина. Во время процессов клеточного деления митоза и мейоза хромосомы реплицируются, чтобы гарантировать, что каждая новая дочерняя клетка получит правильное количество хромосом. Хроматида представляет собой одну из двух нитей реплицированной хромосомы. Хроматиды, соединенные центромерой, называются сестринскими хроматидами. В конце клеточного деления сестринские хроматиды расходятся и становятся дочерними хромосомами во вновь образованных дочерних клетках. Функция хроматина Упаковка ДНК Это самая фундаментальная функция хроматина: компактизация длинных цепей ДНК.Длина ДНК в ядре намного больше, чем размер компартмента, в котором она хранится. Чтобы поместиться в этом отсеке, ДНК должна быть каким-то образом конденсирована. Коэффициент упаковки используется для описания степени конденсации ДНК. Для достижения общего коэффициента упаковки ДНК не упаковывается непосредственно в структуру хроматина. Вместо этого он содержит несколько иерархий организации. Первый уровень упаковки достигается путем намотки ДНК вокруг нуклеосомы, что дает коэффициент упаковки около 6.Эта структура инвариантна как в эухроматине, так и в гетерохроматине всех хромосом. Второй уровень упаковки — это обертывание шариков волокном 30 нм, которое встречается как в интерфазном хроматине, так и в митотических хромосомах. Эта структура увеличивает коэффициент упаковки примерно до 40. Окончательная упаковка происходит, когда волокно организовано в виде петель, каркасов и доменов, которые дают окончательный коэффициент упаковки около 1000 в интерфазном хроматине и около 10000 в митотических хромосомах. Регламент транскрипции Транскрипция — это процесс, при котором генетическая информация, хранящаяся в ДНК, считывается белками, а затем транскрибируется в РНК, а затем РНК транслируется в функциональные белки.Если хроматин укрепляется и ограничивает доступ к считываемым белкам, транскрипция не происходит. Эухроматин, расширенный тип хроматина, может проводить процесс транскрипции. Колебания между открытым и закрытым хроматином могут способствовать прерывности транскрипции или разрыву транскрипции. Вероятно, могут быть задействованы и другие факторы, такие как ассоциация и диссоциация комплексов факторов транскрипции с хроматином.Это явление, в отличие от простых вероятностных моделей транскрипции, может объяснить высокую изменчивость экспрессии генов между клетками в изогенной популяции Восстановление хроматина и ДНК Упаковка ДНК в хроматин представляет собой барьер для всех процессов, связанных с ДНК. Благодаря высокодинамичной организации белков и ДНК хроматин может легко изменять свою форму и структуру. Релаксация хроматина быстро происходит в месте повреждения ДНК, что позволяет белкам репарации связываться с ДНК и восстанавливать ее. Артикул: 1. Комингс Д. 2. Widom J. Структура, динамика и функция хроматина in vitro [J]. Ежегодный обзор биофизики и биомолекулярной структуры , 1998, 27(1): 285-327. 3. Мерсер Т. Р., Маттик Дж. С. Структура и функция длинных некодирующих РНК в эпигенетической регуляции [J]. Природа структурная и молекулярная биология , 2013, 20(3): 300-307. Основной механизм сегрегации хромосом эукариот на JSTORАбстрактный Теперь у нас есть убедительные доказательства того, что основной механизм сегрегации хромосом одинаков у различных эукариот, поскольку используются одни и те же гены. Даже у прокариот самая основная черта расхождения хромосом имеет сходство с таковой у эукариот. Многие аспекты сегрегации хромосом тесно связаны с контролем клеточного цикла, который включает стадийно-специфическую модификацию белков и протеолиз. С самого начала своей истории Королевское общество уделяло большое внимание публикации сообщений своих членов и других лиц. В течение трех лет после выдачи первой Хартии Генри Ольденбург, первый секретарь, в марте 1665 года начал публиковать «Философские труды», и с тех пор это продолжается. Королевское общество является самоуправляемым объединением многих самых выдающихся ученых мира, представляющих все области науки, техники и медицины, и является старейшей непрерывно существующей научной академией. Основная цель Общества, отраженная в его учредительных Уставах 1660-х годов, состоит в том, чтобы признавать, продвигать и поддерживать достижения в науке и поощрять развитие и использование науки на благо человечества.Общество сыграло роль в некоторых из самых фундаментальных, значительных и судьбоносных открытий в истории науки, и ученые Королевского общества продолжают вносить выдающийся вклад в науку во многих областях исследований. Что такое клеточный цикл эукариот?Цикл эукариотической клетки делится на две части: интерфаза и митоз . Интерфаза разделена на три части:
Знание интерфазы на самом деле не требуется, даже на уровне A, поэтому я не буду вдаваться в подробности здесь; все, что вам нужно знать, это то, что во время интерфазы клетка растет , а дублирует свои внутренние структуры , готовясь к митозу. Митоз делится на три части:
Поскольку митоз — очень длительный процесс, я дам его лишь в общих чертах. Но вот видео, объясняющее это подробно. В профазе ядерная мембрана и ядрышко растворяются, в то время как ДНК конденсируется с образованием видимых хромосом , которые представляют собой две сестринские хроматиды , присоединенные к центромере .Также формируется митотическое веретено . Во время метафазы митотическое веретено тянет каждую хромосому к центру клетки , и каждая из них выстраивается вдоль экватора клетки . Во время анафазы митотическое веретено удлиняется , притягивая хромосомы к концам клетки . В этот момент сестринские хроматиды разделены на 90 466 90 467 на центромере. Удлинение митотического веретена также заставляет клетку удлиняться . Во время телофазы ядерные мембраны и ядрышки реформируются вокруг хроматид. Хромосомы также раскручивают и ослабляют . 
|
 8 т.п.н./гаплоид
геном) вершина сложности. В пересчете на вес это 0,07 пикограмма.
на гаплоидный геном Arabidopsis и 100 пикограмм на гаплоидную лилию
геном.
8 т.п.н./гаплоид
геном) вершина сложности. В пересчете на вес это 0,07 пикограмма.
на гаплоидный геном Arabidopsis и 100 пикограмм на гаплоидную лилию
геном.
 «> * в клетках, которые несут генетическую информацию.Большинство прокариотических клеток содержат одну кольцевую хромосому. Эукариотические клетки с их гораздо более крупными геномами имеют множественные линейные хромосомы. Длина и линейный характер эукариотических хромосом усложняют задачу поддержания организованности генетического материала и передачи надлежащего количества ДНК каждой дочерней клетке во время митоза.
«> * в клетках, которые несут генетическую информацию.Большинство прокариотических клеток содержат одну кольцевую хромосому. Эукариотические клетки с их гораздо более крупными геномами имеют множественные линейные хромосомы. Длина и линейный характер эукариотических хромосом усложняют задачу поддержания организованности генетического материала и передачи надлежащего количества ДНК каждой дочерней клетке во время митоза.
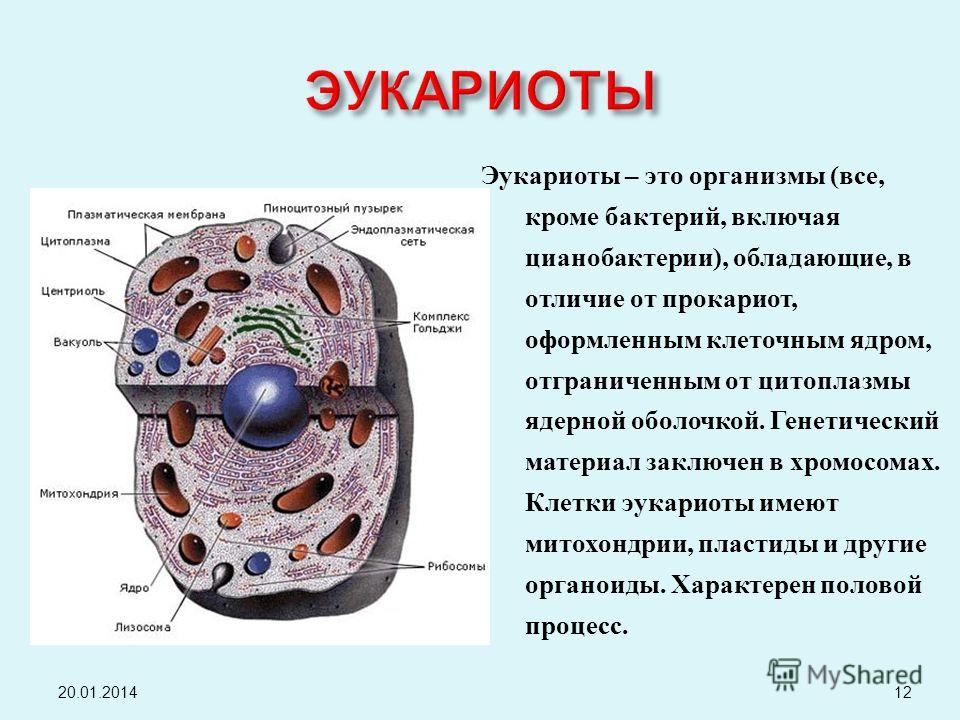 Наиболее заметными особенностями являются центральная точка сужения и ответвления, отходящие от этой точки сужения.Точка сужения называется центромерой, ответвления называются рукавами. Каждая хромосома имеет два или четыре плеча, отходящих от центромеры *.
Наиболее заметными особенностями являются центральная точка сужения и ответвления, отходящие от этой точки сужения.Точка сужения называется центромерой, ответвления называются рукавами. Каждая хромосома имеет два или четыре плеча, отходящих от центромеры *.
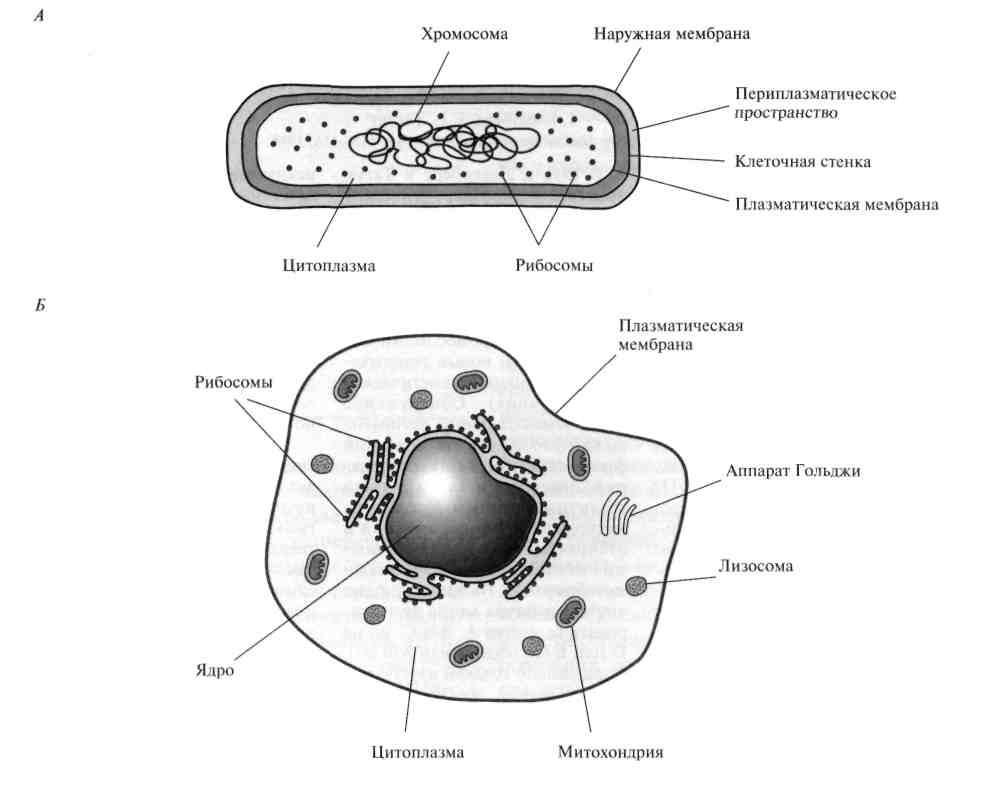 «>*. Пары совпадающих хромосом называются сестринскими хроматидами.
«>*. Пары совпадающих хромосом называются сестринскими хроматидами. Предотвращение потери генетического материала достигается наличием участков повторяющейся некодирующей ДНК на обоих концах хромосомы. Эти повторяющиеся последовательности называются теломерами **s, и они защищают остальную часть нити от деградации во время репликации.
Предотвращение потери генетического материала достигается наличием участков повторяющейся некодирующей ДНК на обоих концах хромосомы. Эти повторяющиеся последовательности называются теломерами **s, и они защищают остальную часть нити от деградации во время репликации.
 В течение большого периода **с клеточного цикла хромосомы находятся в нескрученном, диффузном состоянии.
В течение большого периода **с клеточного цикла хромосомы находятся в нескрученном, диффузном состоянии.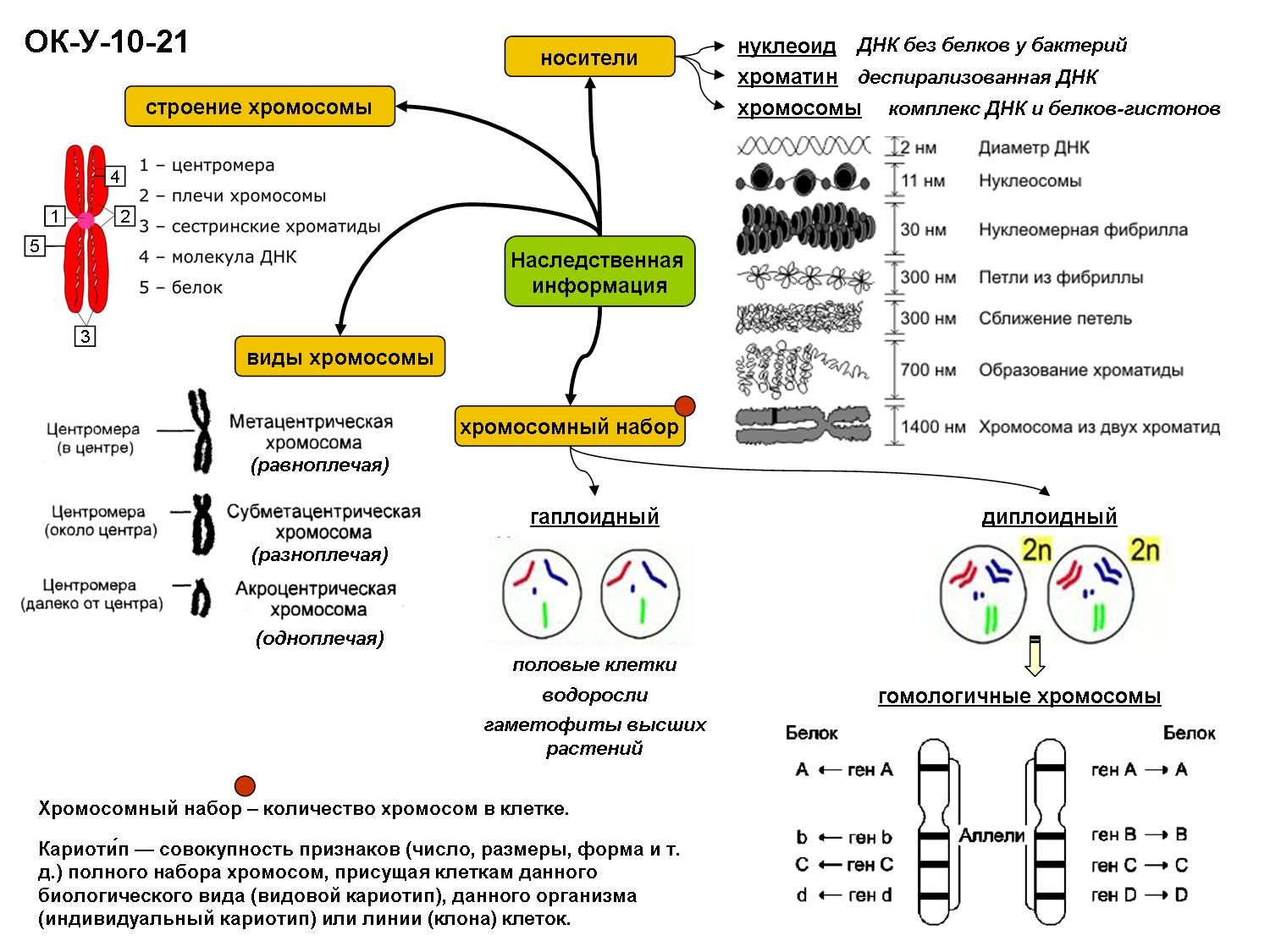 ДНК эукариотических клеток прочно связана с небольшими основными белками (гистонами), которые упорядоченно упаковывают ДНК в ядре клетки. Эта задача является существенной, учитывая содержание ДНК большинства эукариот. Например, общая растянутая длина ДНК в клетке человека составляет почти 2 м, но эта ДНК должна умещаться в ядре диаметром всего от 5 до 10 мкм.Хотя упаковка ДНК также является проблемой для бактерий, механизм, с помощью которого прокариотическая ДНК упаковывается в клетку, отличается от такового у эукариот и недостаточно изучен.
ДНК эукариотических клеток прочно связана с небольшими основными белками (гистонами), которые упорядоченно упаковывают ДНК в ядре клетки. Эта задача является существенной, учитывая содержание ДНК большинства эукариот. Например, общая растянутая длина ДНК в клетке человека составляет почти 2 м, но эта ДНК должна умещаться в ядре диаметром всего от 5 до 10 мкм.Хотя упаковка ДНК также является проблемой для бактерий, механизм, с помощью которого прокариотическая ДНК упаковывается в клетку, отличается от такового у эукариот и недостаточно изучен.  Гистоны чрезвычайно распространены в эукариотических клетках; вместе их масса примерно равна массе клеточной ДНК. Кроме того, хроматин содержит примерно равные массы самых разнообразных негистоновых хромосомных белков. Существует более тысячи различных типов этих белков, которые участвуют в различных процессах, включая репликацию ДНК и экспрессию генов.Гистоны не обнаружены у эубактерий (например, E. coli ), хотя ДНК этих бактерий связана с другими белками, которые предположительно функционируют как гистоны, упаковывая ДНК внутри бактериальной клетки. Архебактерии, однако, содержат гистоны, которые упаковывают их ДНК в структуры, сходные с эукариотическим хроматином.
Гистоны чрезвычайно распространены в эукариотических клетках; вместе их масса примерно равна массе клеточной ДНК. Кроме того, хроматин содержит примерно равные массы самых разнообразных негистоновых хромосомных белков. Существует более тысячи различных типов этих белков, которые участвуют в различных процессах, включая репликацию ДНК и экспрессию генов.Гистоны не обнаружены у эубактерий (например, E. coli ), хотя ДНК этих бактерий связана с другими белками, которые предположительно функционируют как гистоны, упаковывая ДНК внутри бактериальной клетки. Архебактерии, однако, содержат гистоны, которые упаковывают их ДНК в структуры, сходные с эукариотическим хроматином.  Напротив, аналогичное расщепление голой ДНК (не связанной с белками) давало непрерывный мазок из фрагментов произвольного размера. Эти результаты позволяют предположить, что связывание белков с ДНК в хроматине защищает участки ДНК от расщепления нуклеазами, так что фермент может атаковать ДНК только в участках, разделенных примерно 200 парами оснований.В соответствии с этим представлением электронная микроскопия показала, что хроматиновые волокна имеют вид бусинок, причем бусины расположены с интервалом примерно в 200 пар оснований. Таким образом, как нуклеазное расщепление, так и электронно-микроскопические исследования показали, что хроматин состоит из повторяющихся единиц из 200 пар оснований, которые были названы нуклеосомами.
Напротив, аналогичное расщепление голой ДНК (не связанной с белками) давало непрерывный мазок из фрагментов произвольного размера. Эти результаты позволяют предположить, что связывание белков с ДНК в хроматине защищает участки ДНК от расщепления нуклеазами, так что фермент может атаковать ДНК только в участках, разделенных примерно 200 парами оснований.В соответствии с этим представлением электронная микроскопия показала, что хроматиновые волокна имеют вид бусинок, причем бусины расположены с интервалом примерно в 200 пар оснований. Таким образом, как нуклеазное расщепление, так и электронно-микроскопические исследования показали, что хроматин состоит из повторяющихся единиц из 200 пар оснований, которые были названы нуклеосомами.
 ..)
..)  ..)
..)  В интерфазных (неделящихся) клетках большая часть хроматина (называемого эухроматином) относительно деконденсирована и распределена по всему ядру (4). В этот период клеточного цикла происходит транскрипция генов и репликация ДНК для подготовки к делению клеток. Большая часть эухроматина в интерфазных ядрах, по-видимому, находится в форме 30-нм волокон, организованных в большие петли, содержащие приблизительно от 50 до 100 т.п.о. ДНК. Около 10% эухроматина, содержащего активно транскрибируемые гены, находится в более деконденсированном состоянии (конформация 10 нм), что делает возможной транскрипцию.Таким образом, структура хроматина тесно связана с контролем экспрессии генов у эукариот, что будет обсуждаться в главе 6.
В интерфазных (неделящихся) клетках большая часть хроматина (называемого эухроматином) относительно деконденсирована и распределена по всему ядру (4). В этот период клеточного цикла происходит транскрипция генов и репликация ДНК для подготовки к делению клеток. Большая часть эухроматина в интерфазных ядрах, по-видимому, находится в форме 30-нм волокон, организованных в большие петли, содержащие приблизительно от 50 до 100 т.п.о. ДНК. Около 10% эухроматина, содержащего активно транскрибируемые гены, находится в более деконденсированном состоянии (конформация 10 нм), что делает возможной транскрипцию.Таким образом, структура хроматина тесно связана с контролем экспрессии генов у эукариот, что будет обсуждаться в главе 6.
 .)
.) 

 В конце митоза происходит переформирование ядерных мембран и деконденсация хромосом, что приводит к образованию дочерних ядер, содержащих по одной копии каждой родительской хромосомы.
В конце митоза происходит переформирование ядерных мембран и деконденсация хромосом, что приводит к образованию дочерних ядер, содержащих по одной копии каждой родительской хромосомы.

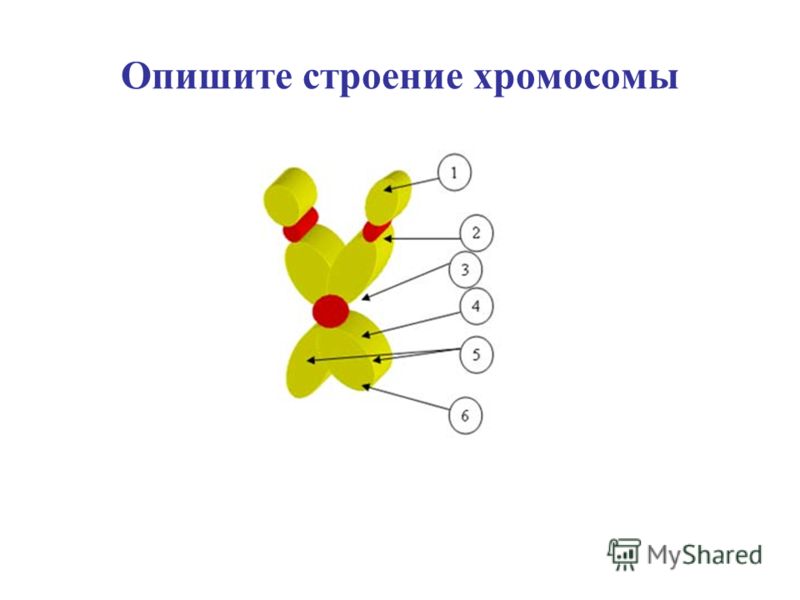 -богатая ДНК ().
-богатая ДНК (). Таким образом, эти два вида дрожжей представляют собой дополнительные модели простых и легко изучаемых эукариотических клеток. Центромеры S. pombe охватывают от 40 до 100 т.п.о. ДНК; они примерно в тысячу раз больше, чем у S. cerevisiae . Они состоят из центрального ядра из 4-7 т.п.н. однокопийной ДНК, окруженного повторяющимися последовательностями (10). Для функции центромеры необходимы не только центральное ядро, но и фланкирующие повторяющиеся последовательности, поэтому центромеры S. pombe кажутся значительно более сложными, чем у S.cerevisiae .
Таким образом, эти два вида дрожжей представляют собой дополнительные модели простых и легко изучаемых эукариотических клеток. Центромеры S. pombe охватывают от 40 до 100 т.п.о. ДНК; они примерно в тысячу раз больше, чем у S. cerevisiae . Они состоят из центрального ядра из 4-7 т.п.н. однокопийной ДНК, окруженного повторяющимися последовательностями (10). Для функции центромеры необходимы не только центральное ядро, но и фланкирующие повторяющиеся последовательности, поэтому центромеры S. pombe кажутся значительно более сложными, чем у S.cerevisiae .  Делеция сателлитных последовательностей и мобильных элементов, а также неповторяющейся ДНК снижала активность центромер в функциональных тестах. Т.о., как повторяющиеся, так и неповторяющиеся последовательности, по-видимому, вносят вклад в формирование кинетохор и функцию центромер.
Делеция сателлитных последовательностей и мобильных элементов, а также неповторяющейся ДНК снижала активность центромер в функциональных тестах. Т.о., как повторяющиеся, так и неповторяющиеся последовательности, по-видимому, вносят вклад в формирование кинетохор и функцию центромер. 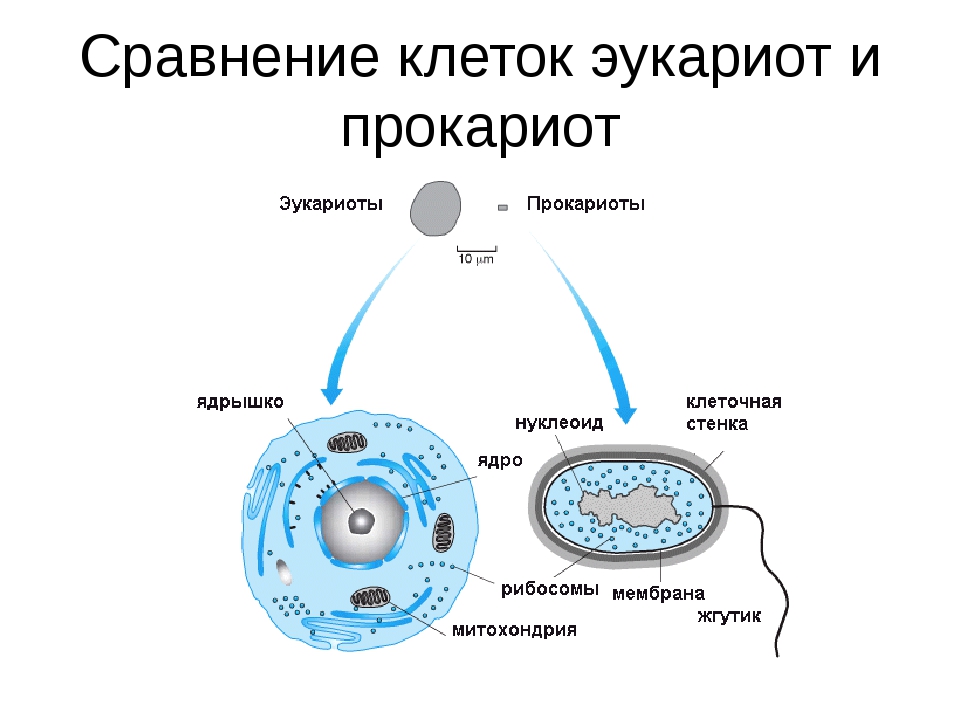 Однако точная функция α-сателлитной ДНК, а также потенциальные роли др. последовательностей в центромерах млекопитающих еще предстоит установить. В соответствии с их большим размером центромеры млекопитающих образуют большие кинетохоры, которые связывают от 30 до 40 микротрубочек, тогда как только отдельные микротрубочки связываются с центромерами S.cerevisiae .
Однако точная функция α-сателлитной ДНК, а также потенциальные роли др. последовательностей в центромерах млекопитающих еще предстоит установить. В соответствии с их большим размером центромеры млекопитающих образуют большие кинетохоры, которые связывают от 30 до 40 микротрубочек, тогда как только отдельные микротрубочки связываются с центромерами S.cerevisiae . 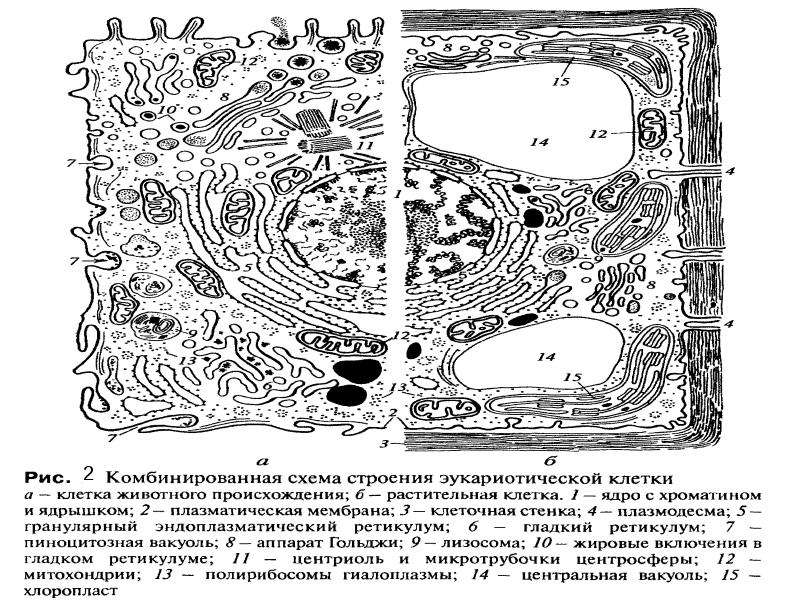
 Следовательно, концы линейных хромосом не могут быть реплицированы обычным действием ДНК-полимеразы. Эта проблема была решена путем эволюции специального механизма, включающего активность обратной транскриптазы, для репликации последовательностей теломерной ДНК. Поддержание теломер, по-видимому, является важным фактором, определяющим продолжительность жизни и репродуктивную способность клеток, поэтому исследования теломер и теломеразы обещают дать новое понимание таких состояний, как старение и рак.
Следовательно, концы линейных хромосом не могут быть реплицированы обычным действием ДНК-полимеразы. Эта проблема была решена путем эволюции специального механизма, включающего активность обратной транскриптазы, для репликации последовательностей теломерной ДНК. Поддержание теломер, по-видимому, является важным фактором, определяющим продолжительность жизни и репродуктивную способность клеток, поэтому исследования теломер и теломеразы обещают дать новое понимание таких состояний, как старение и рак. Нуклеосома может быть дополнительно свернута с образованием хроматинового волокна. Волокна хроматина скручиваются и конденсируются, образуя хромосомы. Хроматин делает возможным протекание ряда клеточных процессов, включая репликацию ДНК, транскрипцию, репарацию ДНК, генетическую рекомбинацию и деление клеток.
Нуклеосома может быть дополнительно свернута с образованием хроматинового волокна. Волокна хроматина скручиваются и конденсируются, образуя хромосомы. Хроматин делает возможным протекание ряда клеточных процессов, включая репликацию ДНК, транскрипцию, репарацию ДНК, генетическую рекомбинацию и деление клеток.  Дублированная хромосома является двухцепочечной и имеет знакомую X-образную форму. Две нити идентичны и соединены в центральной области, называемой центромерой.
Дублированная хромосома является двухцепочечной и имеет знакомую X-образную форму. Две нити идентичны и соединены в центральной области, называемой центромерой. 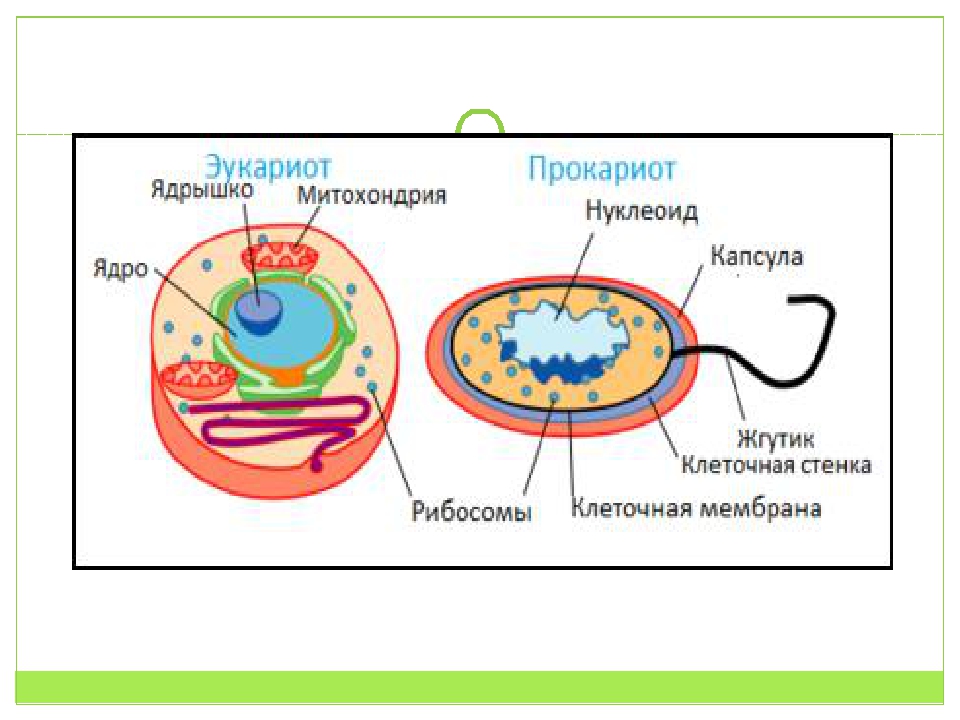
 В то время как гетерохроматин, конденсированный тип хроматина, упакован слишком плотно, чтобы ДНК могла быть прочитана белками.
В то время как гетерохроматин, конденсированный тип хроматина, упакован слишком плотно, чтобы ДНК могла быть прочитана белками.  Э. Строение и функции хроматина [М]. Успехи генетики человека. Springer US , 1972: 237-431.
Э. Строение и функции хроматина [М]. Успехи генетики человека. Springer US , 1972: 237-431.  Разрушение митотического циклина и секурина приводит к выходу из митоза и активации сепаразы соответственно. Ключевыми игроками в сегрегации хромосом являются SMC-содержащие когезин и конденсин, ДНК-топоизомераза II, убиквитинлигаза APC/C, комплекс секурин-сепараза, пассажиры полярных сияний и дестабилизаторы или регуляторы микротрубочек кинетохор. Кроме того, абсолютно необходимо образование митотических кинетохор и веретенообразного аппарата. Обсуждаются роли основных игроков в базовой сегрегации хромосом: большинство игроков выполняют как интерфазные, так и митотические функции.Описан взгляд на то, как формируется центромера/кинетохора.
Разрушение митотического циклина и секурина приводит к выходу из митоза и активации сепаразы соответственно. Ключевыми игроками в сегрегации хромосом являются SMC-содержащие когезин и конденсин, ДНК-топоизомераза II, убиквитинлигаза APC/C, комплекс секурин-сепараза, пассажиры полярных сияний и дестабилизаторы или регуляторы микротрубочек кинетохор. Кроме того, абсолютно необходимо образование митотических кинетохор и веретенообразного аппарата. Обсуждаются роли основных игроков в базовой сегрегации хромосом: большинство игроков выполняют как интерфазные, так и митотические функции.Описан взгляд на то, как формируется центромера/кинетохора.  С 1887 г. и далее, начиная с тома 178, «Транзакции» были разделены на две серии: серию A (математика и физические науки) и серию B (биология).Транзакции публикуются ежемесячно и теперь включают документы, представленные на дискуссионных встречах, а также конкретные темы и обзоры.
С 1887 г. и далее, начиная с тома 178, «Транзакции» были разделены на две серии: серию A (математика и физические науки) и серию B (биология).Транзакции публикуются ежемесячно и теперь включают документы, представленные на дискуссионных встречах, а также конкретные темы и обзоры. 
 А вот и анимация.
А вот и анимация. 