
Мышцы и их функции
Сокращение мышц обеспечивает движение тела и удержание его в вертикальном положении. Вместе со скелетом мышцы придают телу форму. С деятельностью мышц связана функция отдельных органов: дыхания, пищеварения, кровообращения; мышцы гортани и языка участвуют н воспроизведении членораздельной речи.
В организме человека различают три вида мышечной ткани: скелетная, сердечная и стенок внутренних органов. В зависимости от строения мышцы, подразделяются на гладкие (непроизвольные) и поперечнополосатые (произвольные).
Сокращение поперечнополосатой ткани подчинено сознанию. В теле человека насчитывается около 600 скелетных мышц, что составляет 2/5 общей массы тела.
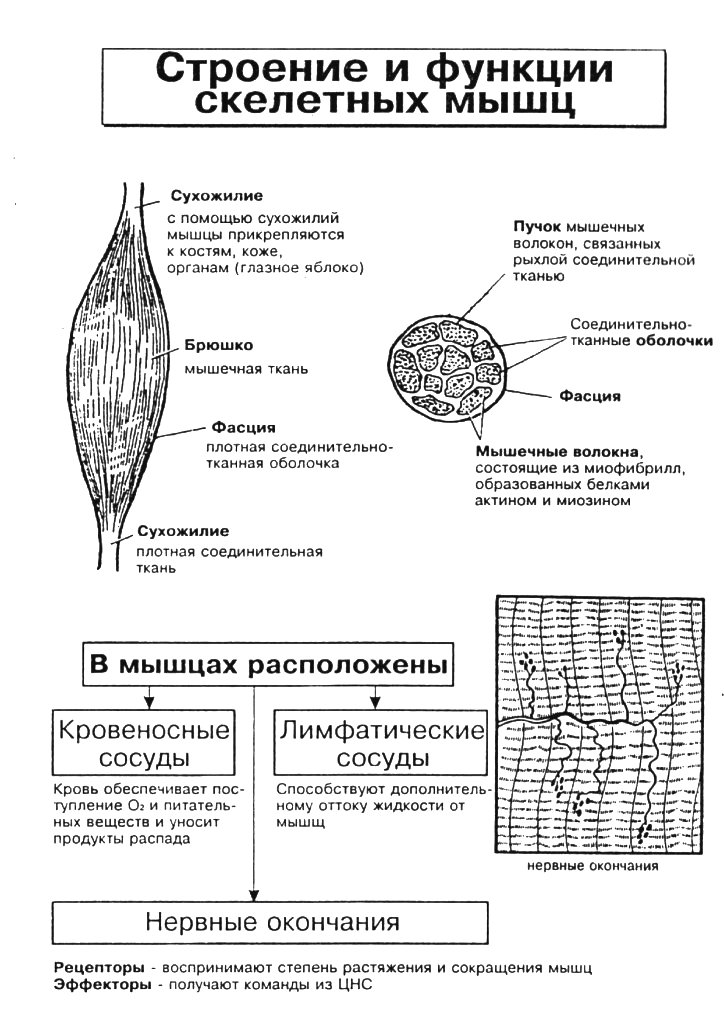
Скелетная мышца покрыта плотной соединительнотканной оболочкой, плотно соединенной с мышечной тканью и препятствующей ее чрезмерному растяжению. Между пучками волокон в мышце расположены кровеносные сосуды и нервы. На концах мышца переходит в сухожилие, обладающее большой прочностью, но в отличие от мышц не обладающее сокращением.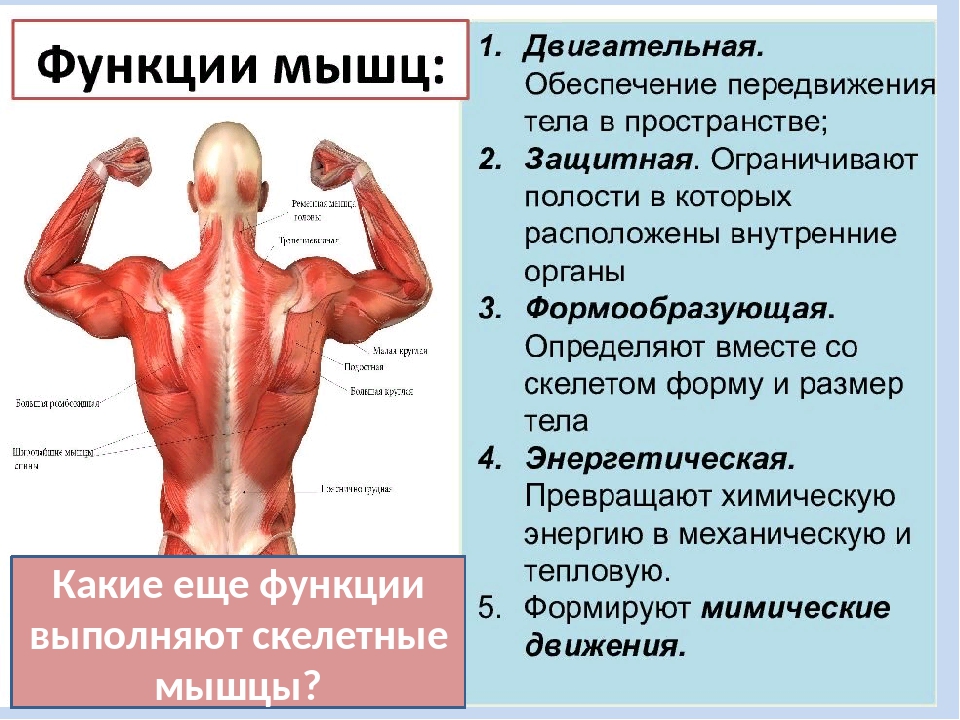
Строение мышцы
Особый вид мышечной ткани — сердечная мышца, образованная поперечнополосатыми мышечными волокнами, но сокращается она непроизвольно. Следовательно, функциональные особенности, строение отличают мышцу сердца от других мышц.
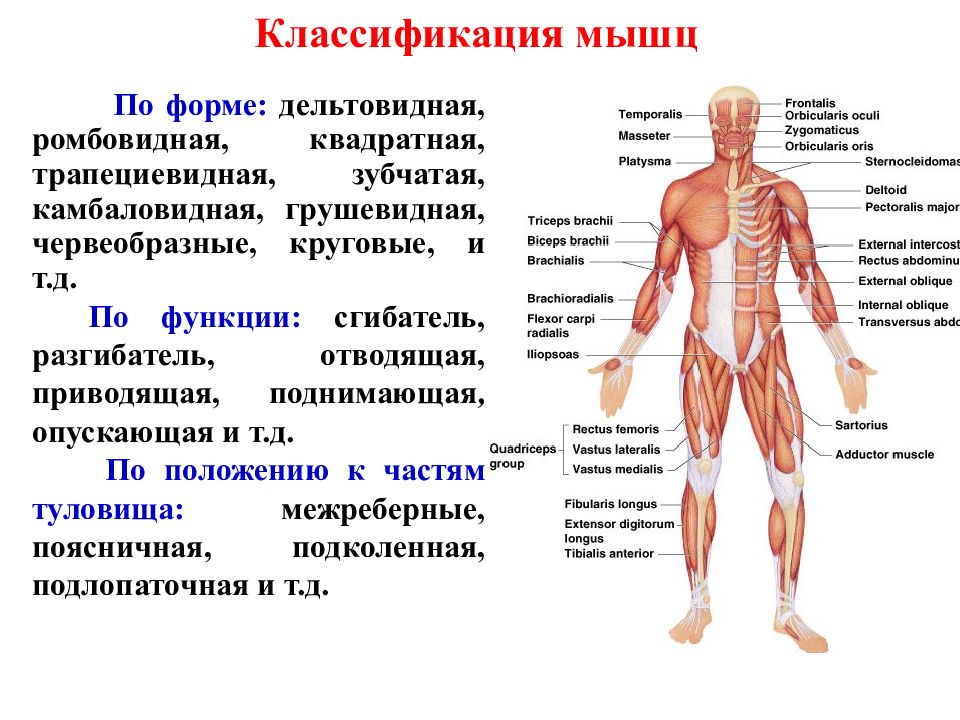
Различают мышцы короткие и толстые, находящиеся преимущественно в глубоких слоях около позвоночного столба; длинные и тонкие, расположенные на конечностях; широкие и плоские, сосредоточенные в основном на туловище.
По функциям мышцы делятся на сгибатели, разгибатели, приводящие, отводящие, вращатели. При сокращении мышц сгибателей одновременно расслабляются разгибатели, что обеспечивает согласованность движений.
Мышцы, сокращение которых вызывает движение конечности от тела, называются отводящими, а приближающие конечность к телу, — приводящими. Мышцы вращатели при своем сокращении вращают ту или иную часть тела (голову, плечо, предплечье).
В организме человека различают мышцы туловища, головы, верхних и нижних конечностей. Мышцы туловища подразделяются на мышцы груди, спины и живота. К мышцам груди относятся наружные и внутренние межреберные мышцы и диафрагма, или грудобрюшная преграда, с помощью которых осуществляется дыхание. Большая и малая грудные мышцы, передняя зубчатая и подключичная мышцы приводят в движение плечевой пояс и руки.
Мышцы туловища подразделяются на мышцы груди, спины и живота. К мышцам груди относятся наружные и внутренние межреберные мышцы и диафрагма, или грудобрюшная преграда, с помощью которых осуществляется дыхание. Большая и малая грудные мышцы, передняя зубчатая и подключичная мышцы приводят в движение плечевой пояс и руки.
Мышцы живота вызывают сгибание позвоночника вперед, в сторону и поворот его вокруг продольной оси, Образуют брюшной пресс, сокращение которого способствует глубокому выдоху, выведению кала, мочи, а также родовому акту у женщин.
Поверхностные мышцы спины (трапециевидная и широчайшая) обеспечивают укрепление и движение плечевого пояса и рук. Глубокие мышцы спины фиксируют позвоночник, вызывают его разгибание, сгибшие, наклоны в сторону и вращение, разгибание и вращение головы, участвуют в дыхательных движениях. Самая крупная мышца шеи — грудино-ключично-сосцевидная.
Мышцы головы подразделяются на две группы: жевательные и мимические. Собственно жевательная мышца начинается от нижнего края скуловой кости и прикрепляется к нижней челюсти. Сокращаясь, она поднимает нижнюю челюсть, участвуя в пережевывании пищи.
Сокращаясь, она поднимает нижнюю челюсть, участвуя в пережевывании пищи.
Мимические мышцы прикрепляются одним концом к костям черепа, другим — к коже лица. Благодаря им лицо человека выражает те или иные эмоции: гнев, горе, радость. Кроме того, они участвуют в акте речи, дыхания.
На лбу расположены лобные мышцы, вокруг глазницы — круговая мышца (способствует закрыванию век). Вокруг ротового отверстия находится круговая мышца рта.
Мускулатура верхних конечностей подразделяется на мышцы плечевого пояса (дельтовидная, большая и малая грудная), которые обеспечивают его подвижность, и мышцы свободной конечности. Важнейшие мышцы свободной конечности — двуглавая мышца (сгибает предплечье) и трехглавая (на задней поверхности плечевой кости), разгибающая плечо и предплечье. На передней поверхности предплечья находятся мышцы — сгибатели предплечья, кисти и пальцев, на задней — мышцы — разгибатели предплечья, кисти и пальцев.
Мышцы нижних конечностей подразделяются на мышцы тазового пояса и свободной конечности. К мышцам таза относятся подвздошно-поясничная мышца и три ягодичные, самая крупная, разгибающая бедро, — большая ягодичная. На задней поверхности бедра выделяются полусухожильная, полуперепончатая и двуглавая мышцы, при сокращении которых происходит сгибание голени в коленном суставе и разгибание бедра. На передней поверхности бедра лежит четырехглавая мышца, при сокращении которой разгибается голень. На передней поверхности голени находятся мышцы — разгибатели стопы и пальцев, на задней — их сгибатели. Важнейшие из них — икроножная и камбалообразная. Обе мышцы заканчиваются ахилловым сухожилием, которое прикрепляется к пяточному бугру. Икроножная мышца поднимает пятку при ходьбе и принимает участие в поддержании тела в вертикальном положении.
К мышцам таза относятся подвздошно-поясничная мышца и три ягодичные, самая крупная, разгибающая бедро, — большая ягодичная. На задней поверхности бедра выделяются полусухожильная, полуперепончатая и двуглавая мышцы, при сокращении которых происходит сгибание голени в коленном суставе и разгибание бедра. На передней поверхности бедра лежит четырехглавая мышца, при сокращении которой разгибается голень. На передней поверхности голени находятся мышцы — разгибатели стопы и пальцев, на задней — их сгибатели. Важнейшие из них — икроножная и камбалообразная. Обе мышцы заканчиваются ахилловым сухожилием, которое прикрепляется к пяточному бугру. Икроножная мышца поднимает пятку при ходьбе и принимает участие в поддержании тела в вертикальном положении.
Мышцы верхней конечности осуществляют разнообразные и многочисленные движения руки. Так как нижние конечности человека выдерживают всю тяжесть тела и целиком принимают на себя функцию его передвижения, то их мышцы значительно массивнее, и, следовательно, сильнее, чем мышцы рук, но вместе с тем обладают более ограниченным размахом движений.
Работа мышц.
Движения в суставах — сгибание и разгибание конечностей — совершаются благодаря поочередному сокращению и расслаблению мышц сгибателей и разгибателей, действующих согласованно вследствие иннервации их нервных центров, последовательно переходящих из состояния возбуждения в состояние торможения.
Работа мышц связана с расходом энергии, которую дает аденозинтрифосфорная кислота (АТФ), ее запасы в мышцах небольшие и израсходуются за доли секунды. Синтезируется АТФ за счет энергии, освобождающейся при окислении глюкозы, которую приносит к мышцам кровь вместе с питательными веществами и уносит продукты распада и углекислый газ. Таким образом, эффективность работы мышц зависит от кровоснабжения мышц и, следовательно, от работы сердечно-сосудистой системы.
Различают работу статическую и динамическую. При статической работе мышцы находятся в постоянном напряжении, но не сокращаются (поднятие тяжести, удержание груза). Такая работа очень утомительна, особенно для детей и подростков.
Динамическая работа мышц сопровождается поочередными сокращениями и расслаблениями мышц (бег, хождение, плавание, различные игры), она менее утомительна, потребует много энергии.
Показателем эффективности работы мышц является коэффициент полезного действия — КПД, измеряемый по формуле (известной из физики) КПД = A/Q. то есть соотношение выполненной работы к общему количеству затраченной энергии. КПД мышц человека в среднем равно 25-30%, то есть 30% всей энергии затрачивается на сокращение мышц, остальные 70% — преобразуются в тепло.
Утомляемость — это временное понижение работоспособности, наступающее в результате труда и исчезающее после отдыха. Для борьбы с утомлением необходимо чередовать разнообразную деятельность.
Ахиллово сухожилие — анатомия и функция
Анатомия и функция ахиллова сухожилия
Ахиллово сухожилие является самым толстым и мощным сухожилием в теле человека.
Функция ахиллова сухожилия
Основная функция ахиллова сухожилия —
 е. передний отдел стопы опускается вниз. Без этого движения невозможны полноценная ходьба и бег. Ахиллово сухожилие также участвует в супинации стопы (вращение внутрь).
е. передний отдел стопы опускается вниз. Без этого движения невозможны полноценная ходьба и бег. Ахиллово сухожилие также участвует в супинации стопы (вращение внутрь). Ахиллово сухожилие испытывает колоссальные нагрузки, особенно при прыжках и беге. Например, при беге на ахиллово сухожилие действует сила, равная восьмикратному весу Вашего тела.1
Ахиллово сухожилие — это печально известное слабое место, особенно у бегунов, ведь очень многие из них на себе испытали ахиллодинию — боли в области ахиллова сухожилия.
Интересный факт: источником названия сухожилия считают древнегреческий миф об Ахиллесе (Ахилле). По преданию, его тело было неуязвимо за исключением одного места – пятки. По легенде, во время Троянской войны Ахиллес погиб от стрелы, выпущенной в его пятку. Ахиллесова пята стала синонимом уязвимого места и дала название ахиллову сухожилию человека.
Источник
1 Klein, C.: Orthopädie für Patienten [Orthopaedics for Patients]. Publisher: Michels-Klein, Remagen 2014.
Publisher: Michels-Klein, Remagen 2014.
При болях в спине и суставах обязательно надо лечить еще и мышцы Medical On Group Тюмень
Возникновение болей является тревожным сигналом организма о том, что в определенной области развивается проблема. Множество причин может повлечь возникновение такого дискомфорта. Самыми частыми виновниками становятся дегенеративно-дистрофические процессы (остеохондроз, артроз), а так же различные травмы, будь то ушибы, разрывы связок, или даже просто растяжения или перенапряжение не тренированных мышц.Чрезмерные нагрузки на элементы опорно-двигательной системы или их физиологическое старение провоцирует развитие болезней. Поэтому даже незначительные боли могут оказаться предвестниками или причинами серьезных проблем. Своевременное обращение к врачу и правильное лечение позволит: предотвратить прогрессирование патологии, снять болевой синдром, предотвратить возникновение осложнений, а так же улучшит прогноз лечения и, в большинстве случаев, удастся излечиться или добиться стойкой ремиссии и не дать болезни развиваться дальше.
Болевой синдром любой локализации, возникший по любой причине, может запустить «цепную реакцию» под названием миофасциальный болевой синдром. Человек интуитивно при болях старается выбрать наиболее комфортное положение, и даже привычные движения совершаются по-другому – начинают активно задействоваться те мышцы, которые раньше использовались в меньшей мере. Организм стремится создать мышечный каркас для того чтобы обездвижить болезненную область. В последующем именно те мышцы, которые взяли на себя дополнительную нагрузку, обуславливают возникновение миофасциального болевого синдрома.
Миофасциальный болевой синдром (МБС) — хроническое, патологическое состояние, обусловленное формированием локальных мышечных спазмов или уплотнений, представленных болевыми точками (триггерами). Боли могут провоцироваться нажатием на эти точки, или движением мышц вовлеченных в патологический процесс. Данный синдром приводит к ограничению объема движений, и повышенной мышечной утомляемости.
При возникновении МБС боль распространяется по мышцам от первоисточника в окружающие мягкие ткани, что проявляется возникновением новых болезненных уплотнений (болевых точек). Эти участки уплотнения мышц не большие и имеют диаметр 0,1-0,3 см, но если их появляется большое количество, то они группируются, тем самым создавая триггерную зону диаметром от 1 см и более. Появление болевых точек происходит при перенапряжении и микротравматизации мышцы. МБС может вызывать нарушение нормального распределения нагрузок в ближайших суставах и приводить к раннему износу и даже деформации (искривлению).
Предрасполагающими факторами к появлению МБС являются:
- Патология позвоночника. Дегенеративно-дистрофические заболевания (остеохондроз любой локализации, артроз межпозвонковых суставов), травмы позвоночника являются источником боли, которая провоцирует мышцы спины к поддержанию повышенного тонуса. Так же фактором провоцирующим возникновение МСБ становится вынужденное, из-за боли, статичное положение, обуславливающее мышечное перенапряжение.

- Патология опорно-двигательной системы. деформация позвоночника, асимметрия нижних конечностей (одностороннее укорочение), асимметричность (перекос) таза, поперечное и продольное плоскостопие могут приводить к неправильному, не равномерному распределению нагрузки на мышцы, а так же хрящи и кости тела, что определят возникновение не только МБС , но и таких заболеваний как артроз, артрит, синовит, лигаментит. У 70% населения имеется сколиотическая деформация позвоночника и у 50% плоскостопие различных степеней.
- Вынужденная поза. Работа в однообразном положении тела, фиксации конечностей, статичное положение приводят к мышечной перегрузке.
- Стереотипные движения. Одинаковые повторяющиеся движения происходят с сокращением одних и тех же мышц, приводящее к их перенапряжению и неправильному распределению тонуса.
-
Не адекватная нагрузка, в особенности нетренированных мышц. В результате возникает мышечное перенапряжение и микро надрывы.

- Ушибы. Непосредственная травматизация самой мышцы обуславливает нарушение структуры ее волокон. Следствием этого является нарушение функции, если функция одних волокон уменьшается, то функция других должна увеличиться, компенсируя недостаток первых, что и ведет к перегрузке последних.
- Соматические заболевания. Болезненность внутренних органов, провоцирует тоническое напряжение определенных групп мышц. При длительном сохранении болевого синдрома могут формироваться триггерные точки и хронический выраженный болевой синдром.
- Эмоциональное перенапряжение. Различные психоэмоциональные реакции, такие как стресс, тревога могут побуждать мышцы к тоническому напряжению. В свою очередь возникновение мышечно-тонических состояний, и их сохранение после перенесенного эмоционального переживания, способны формировать МБС и постепенное развитие хронической боли в спине (остеохондроз) отдающиеся в конечности.
 Происходит высвобождение ионов кальция, что ведет к повреждению на клеточном уровне. Избыточное количество кальция провоцирует сокращение миоцита (мышечная клетка). А мышца, находящаяся в спазмированном состоянии, обеспечивает повышение внутримышечного давления, что обуславливает ухудшение микроциркуляции. При длительном сокращении, выполняется большой объем работы, на которую затрачивается энергия, для восполнения энергии требуется период расслабления. Если периода расслабления нет длительное время, срабатывают механизмы компенсации: энергия в клетке восполняется не за счет вновь прибывших питательных веществ из кровотока, а за счет имеющихся запасов внутри себя, что приводит низкоэффективной работе. Еще более длительная нагрузка, превышающая возможности мышцы, вызывает срыв компенсаторных механизмов — накоплению шлаков в мышечной ткани, и как результат образованию триггерной точки. Появившаяся боль провоцирует мышечное сокращение. Формируется порочный круг: боль — мышечное напряжение— боль.
Происходит высвобождение ионов кальция, что ведет к повреждению на клеточном уровне. Избыточное количество кальция провоцирует сокращение миоцита (мышечная клетка). А мышца, находящаяся в спазмированном состоянии, обеспечивает повышение внутримышечного давления, что обуславливает ухудшение микроциркуляции. При длительном сокращении, выполняется большой объем работы, на которую затрачивается энергия, для восполнения энергии требуется период расслабления. Если периода расслабления нет длительное время, срабатывают механизмы компенсации: энергия в клетке восполняется не за счет вновь прибывших питательных веществ из кровотока, а за счет имеющихся запасов внутри себя, что приводит низкоэффективной работе. Еще более длительная нагрузка, превышающая возможности мышцы, вызывает срыв компенсаторных механизмов — накоплению шлаков в мышечной ткани, и как результат образованию триггерной точки. Появившаяся боль провоцирует мышечное сокращение. Формируется порочный круг: боль — мышечное напряжение— боль. Так же к этому кругу может присоединяться нарушение структуры хряща сустава и кости. При распространение боли по мышцам могут вовлекаться нервы, по которым болевой синдром, может передаваться на удалённые области (например — отдавать в руки или ноги при болях в области позвоночника).
Так же к этому кругу может присоединяться нарушение структуры хряща сустава и кости. При распространение боли по мышцам могут вовлекаться нервы, по которым болевой синдром, может передаваться на удалённые области (например — отдавать в руки или ноги при болях в области позвоночника).Бывают активные и латентные триггерные точки. Активные болевые точки могут проявляться при движениях и даже в покое. Латентные точки в покое не болят, но при нажатии на них возникает резкая боль. Выделяются 3 формы миофасциального болевого синдрома: острая, подострая и хроническая. При острой форме болевой синдром поддерживается постоянно и усиливается при движениях. Подострая форма характеризуется болью только при движениях, и при покое не проявляется. При хронической форме МБС боли возникаю только при непосредственном нажатии на саму триггерную точку, а также при движениях может проявляться чувство дискомфорта.
Понимание причины заболевания требуется для подбора правильного лечения. Поэтому, в практике используется классификация МБС по этиологии: это может быть первичный миофасциальный болевой синдром, первопричиной которого, является непосредственное повреждение мышечного волокна – перенапряжение, микроразрывы; а так же вторичный МБС, первопричиной в этом случае служит боль в суставах, во внутренних органах, в позвоночнике.
Для жизни МБС не опасен, но в значительной степени может ухудшить ее качество, снизить трудоспособность и ускорить износ костных и хрящевых структур опорно-двигательной системы. Хронический болевой синдром изматывает человека физически, плохо отражается на психоэмоциональном состоянии, и может нарушать сон. Недосып усиливает усталость, негативно сказываясь на жизнедеятельности и трудовой активности.
Большую часть болей при дегенеративных патологиях опорно-двигательной системы обусловливает именно миофасциальный болевой синдром. Даже если убрать первоисточник болевых ощущений, при уже сформированном МБС, боль не уйдет, более того, неправильное распределение нагрузок в соседних, от пораженных мышц, областях будет провоцировать микротравматизацию и воспаление. Поэтому важно лечить не только артрозы суставов, протрузии и грыжи межпозвонковых дисков, бурситы, синовиты, но и окружающие мышцы.
Передовыми методами лечения миофасциального синдрома считается комплекс мер, воздействующий на все звенья цепочки патологических процессов, и обеспечивающий купирование первоисточника боли, улучшение микроциркуляции в, спазмированных, мышцах, за счет расслабления мышечного тонуса и непосредственного расширения просвета сосудов, восстановления процессов внутриклеточного обмена веществ и транспорта через мембрану клетки, а так же купирования воспаления.
Хороший эффект дают комбинации некоторых методов физиолечения, (Hil-терапия высокоинтенсивным лазером, ударно-волновая терапия, ультразвуковое воздействие с медицинскими препаратами) с массажем, различными вариантами блокад, лечебной физкультурой и медикаментозной терапией, а также использованием различных ортопедических изделий.
Сердечная недостаточность — Кардиология | Хирсланден Швейцария
Слабость сердечной мышцы, как правило, развивается незаметно. Сердечная мышца при этом медленно слабеет до состояния, когда, в конечном счете, больше не в состоянии закачивать нужное количество крови в систему кровообращения. Как следствие пониженной насосной функции снижается и количество кислорода, необходимого для питания таких важных органов, как головной мозг, почки или мышцы. В зависимости от того, какие камеры сердца страдают от недостаточности — левые или правые, говорят о сердечной недостаточности правых или левых отделов сердца. Если недостаточность затронула обе камеры, то речь идёт о глобальной сердечной недостаточности. К ослаблению сердечной мышцы могут приводить разные причины. Самая частая из них – это нарушения коронарного кровообращения (ИБС, ишемическая болезнь сердца). Но также и клапанные болезни сердца, воспаление сердечной мышцы или заболевания лёгких могут приводить к сердечной недостаточности.
Если недостаточность затронула обе камеры, то речь идёт о глобальной сердечной недостаточности. К ослаблению сердечной мышцы могут приводить разные причины. Самая частая из них – это нарушения коронарного кровообращения (ИБС, ишемическая болезнь сердца). Но также и клапанные болезни сердца, воспаление сердечной мышцы или заболевания лёгких могут приводить к сердечной недостаточности.
Слабость сердечной мышцы сначала протекает малосимптоматично, либо вообще не даёт о себе знать. Тревожными признаками, которые могут указывать на начинающуюся слабость сердечной мышцы, это прогрессирование чувства усталости, снижение выносливости к нагрузкам, отёчность ног и щиколоток, а также ночной кашель. Однако также и одышка с нехваткой воздуха, приступы головокружения, боли в груди или неясные колебания веса могут указывать на сердечную недостаточность.
Чем раньше будет распознана сердечная недостаточность и назначено лечение, тем лучше прогнозы. Диагноз ставят на основании клинической картины жалоб и на основании данных различных исследований. Самое важное исследование здесь – это измерение насосной функции сердца методом эхокардиография. В зависимости от степени тяжести сердечной недостаточности, она подразделяется на 4 стадии (ФК), которые определены Классификацией Нью-Йоркской Ассоциации сердца (NYHA). На I стадии ещё возможна нормальная физическая нагрузка. На II стадии жалобы появляются при интенсивной физической нагрузке, а на III стадии эти жалобы уже появляются при лёгкой нагрузке. Для стадии IV, в конечном итоге, эти симптомы характерны уже даже в состоянии покоя.
Самое важное исследование здесь – это измерение насосной функции сердца методом эхокардиография. В зависимости от степени тяжести сердечной недостаточности, она подразделяется на 4 стадии (ФК), которые определены Классификацией Нью-Йоркской Ассоциации сердца (NYHA). На I стадии ещё возможна нормальная физическая нагрузка. На II стадии жалобы появляются при интенсивной физической нагрузке, а на III стадии эти жалобы уже появляются при лёгкой нагрузке. Для стадии IV, в конечном итоге, эти симптомы характерны уже даже в состоянии покоя.
Для лечения сердечной недостаточности применяют различные медикаменты. Цель медикаментозного лечения состоит в разгрузке сердца (снижении артериального давления) и усилении насосной функции. В отдельных случаях, когда сердечная недостаточность связана с нарушением проводимости электрического импульса, может применяться кардиальная ресинхронизационная терапия.
Если, несмотря на все усилия по лечению сердечной недостаточности, она продолжает демонстрировать тенденцию роста, то в некоторых случаях в качестве последней опции лечения остается только трансплантация сердца.
Сильная и здоровая спина: когда тренажерного зала недостаточно
Практически каждый человек, посещающий фитнес-клуб, преследует вполне определенную цель – быть здоровым и иметь красивое тело. В нашем представлении стройная фигура – это рельефный торс, «кубики» пресса, подтянутые ягодицы. Да, это действительно выглядит эффектно! Однако есть и другой аспект красоты – здоровье. Даже люди, регулярно посещающие тренажерный зал, к сожалению, очень часто жалуются на напряжение или ноющие боли в пояснице, шейном отделе, снижение роста на 2-3 см. Почему так происходит – разберемся в этой статье.
Наш позвоночник удерживает не широчайшая мышца спины и не «кубики» пресса, а глубокие (аутохтонные) мышцы, прилегающие непосредственно к позвоночному столбу. Среди них есть короткие мышцы, которые крепятся попарно к соседним позвонкам, есть длинные — тянутся вдоль всего позвоночного столба от крестца до затылочной кости. Основная функция этих околопозвоночных мышц — выпрямление туловища. Даже тогда когда мы наклоняемся вперед, эти мышцы сильно сокращаются (а не растягиваются!), противодействуя силе тяжести. Именно аутохтонные мышцы спины отвечают за стабильность позвоночных сегментов и помогают удлинить (вытянуть) позвоночник. Они так и называются – мышцы-стабилизаторы позвоночника.
Основная функция этих околопозвоночных мышц — выпрямление туловища. Даже тогда когда мы наклоняемся вперед, эти мышцы сильно сокращаются (а не растягиваются!), противодействуя силе тяжести. Именно аутохтонные мышцы спины отвечают за стабильность позвоночных сегментов и помогают удлинить (вытянуть) позвоночник. Они так и называются – мышцы-стабилизаторы позвоночника.
Глубокие мышцы спины, кроме стабилизации позвоночника, обеспечивают подвижность каждого его отдела. Именно благодаря им мы можем наклоняться вперед, поворачивать корпус вокруг собственной оси, совершать боковые наклоны. Когда мы двигаемся таким образом, происходит улучшение кровотока в спине, и в частности – в самих позвонках. Это очень важный момент, на который следует обратить внимание каждому. Здоровая подвижность спины и полноценное кровоснабжение каждого позвонка – залог долголетия межпозвонковых дисков. Диски участвуют в выполнении основных функций позвоночника: защита спинного мозга и отходящих от его столба нервных корешков, амортизация при ходьбе и других вертикальных нагрузках. Необходимо, чтобы каждый из 24 дисков позвоночника получал в полном объеме питательные вещества и воду. В противном случае происходит их постепенное разрушение: диски теряют влагу и упругость, уменьшаются в высоту (именно из-за этого происходит «возрастное снижение роста»), ткани дисков разрушаются, что может привести к грыже и защемлению нервного корешка. Большую вероятность получения таких последствий несут в себе долгие неподвижные осевые нагрузки, например, пребывание в положении сидя на стуле.
Необходимо, чтобы каждый из 24 дисков позвоночника получал в полном объеме питательные вещества и воду. В противном случае происходит их постепенное разрушение: диски теряют влагу и упругость, уменьшаются в высоту (именно из-за этого происходит «возрастное снижение роста»), ткани дисков разрушаются, что может привести к грыже и защемлению нервного корешка. Большую вероятность получения таких последствий несут в себе долгие неподвижные осевые нагрузки, например, пребывание в положении сидя на стуле.
Как же происходит питание межпозвонкового диска, если известно, что его ткани не имеют собственного кровоснабжения, т.е. к ним не подходят кровеносные сосуды? Только путем всасывания питательных веществ и воды из капилляров прилежащих позвонков. Соответственно, чем больше двигательная активность глубоких мышц (именно их, а не крупных поверхностных мышц спины!), тем выше кровоток в позвонках.
Если глубокие мышцы спины у человека слабые и обладают невысокой выносливостью, мы испытываем к концу дня усталость в спине и пояснице, нам трудно держать ровную осанку, появляется «удобная» сутулость.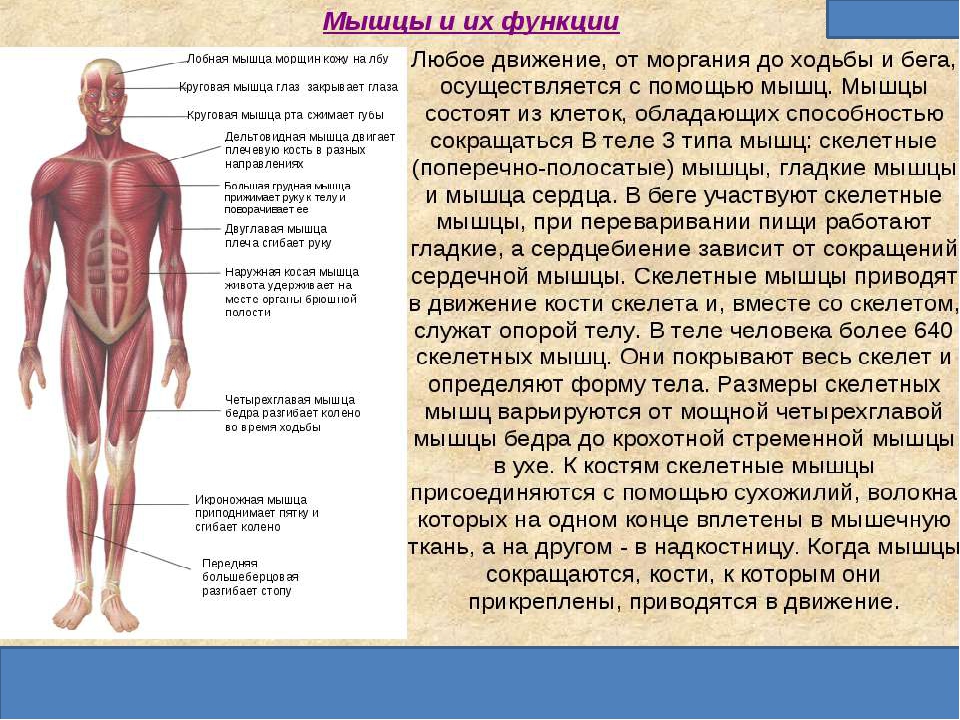 Все это говорит о том, что глубокие мышцы спины не справляются со своей прямой функцией. Если мы не обращаем внимания на такие явления и, придя вечером в зал, кладем себе на плечи штангу, мы не способствуем этими действиями укреплению глубоких мышц спины. Наоборот, из-за слабости удерживающих позвоночник мышц и недостаточного питания межпозвонковых дисков мы можем получить смещения позвонков, протрузии, грыжи.
Все это говорит о том, что глубокие мышцы спины не справляются со своей прямой функцией. Если мы не обращаем внимания на такие явления и, придя вечером в зал, кладем себе на плечи штангу, мы не способствуем этими действиями укреплению глубоких мышц спины. Наоборот, из-за слабости удерживающих позвоночник мышц и недостаточного питания межпозвонковых дисков мы можем получить смещения позвонков, протрузии, грыжи.
Доказано, что укрепление любой мышцы и поддержание ее работоспособности происходит только при регулярных силовых тренировках. Это же относится и к глубоким мышцам спины, даже к самым коротким из них. Важно понимать, что тренировка этих мышц требует особенного подхода: в тренажерах и тем более при работе со штангой они не прорабатываются в должной степени. На таких занятиях задействуются поверхностные, крупные мышцы. Тренировки, направленные на построение «мышечного корсета», базируются на упражнениях в статике и удержании баланса. Чем дольше вы держите неустойчивое и неудобное (по началу) положение, тем глубже идет проработка мышц-стабилизаторов.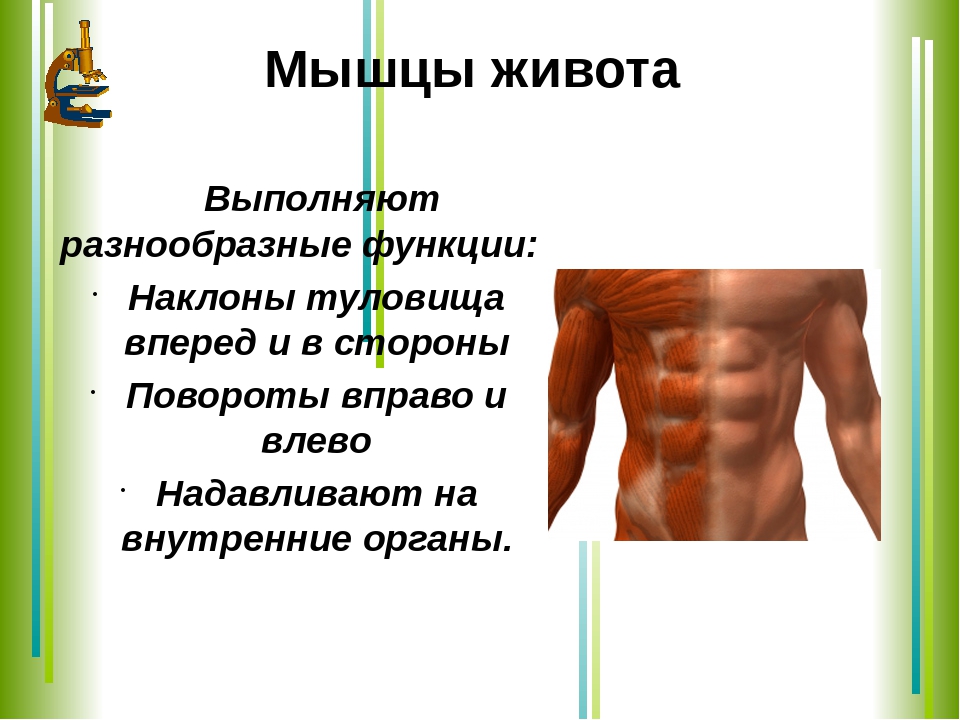
Качественная и безопасная проработка глубоких мышц позвоночника и всех суставов тела происходит на занятиях хатха-йогой в силовом режиме тренировки. Широкий арсенал упражнений, возможность модификации каждого положения с учетом текущей физической подготовки, надлежащий режим дыхания при статичных нагрузках – все это позволяет максимально эффективно тренировать все мышечные слои нашего тела – от глубоких до поверхностных. Всем известное положение «Планка» (вариация асаны «Чатуранга» из хатха-йоги) считается одним из универсальных упражнений, которое тренирует сразу весь «мышечный корсет». При его выполнении практически все мышцы тела одновременно сопротивляются силе тяжести, стараясь предотвратить падение. Причем основная работа по удержанию тела в заданном положении ложится на стабилизаторы спины и таза (аутохтонные мышцы спины и глубокие мышцы живота), а не на мышцы рук, как многие предполагают в начале.
Если вы хотите иметь не только красивое, рельефное, но и здоровое тело, обязательно уделяйте время тренировкам глубоких мышц. Это обеспечит вам здоровье позвоночника и всего опорно-двигательного аппарата, сделает ваше тело не только сильным, но и гармонично развитым и выносливым!
Это обеспечит вам здоровье позвоночника и всего опорно-двигательного аппарата, сделает ваше тело не только сильным, но и гармонично развитым и выносливым!
| 1. |
Прикрепление мышц к костям
Сложность: среднее |
1 |
2.
|
Восстановление работоспособности мышц
Сложность: среднее |
1 |
| 3. |
Боль в мышцах
Сложность: среднее |
1 |
4.
|
Определи мышцу по рисунку
Сложность: среднее |
1 |
| 5. |
Особенности гладкой мышечной ткани
Сложность: среднее |
1 |
6.
|
Согласованная работа мышц
Сложность: среднее |
1 |
| 7. |
Мышцы локтевого сустава
Сложность: среднее |
1 |
8.
|
Динамическая и статическая работа мышц
Сложность: среднее |
1 |
| 9. |
Дополни текст «Мышечные ткани человека»
Сложность: среднее |
1 |
Физиологические причины потери контроля над мочеиспусканием у мужчин и женщин
Временное недержание.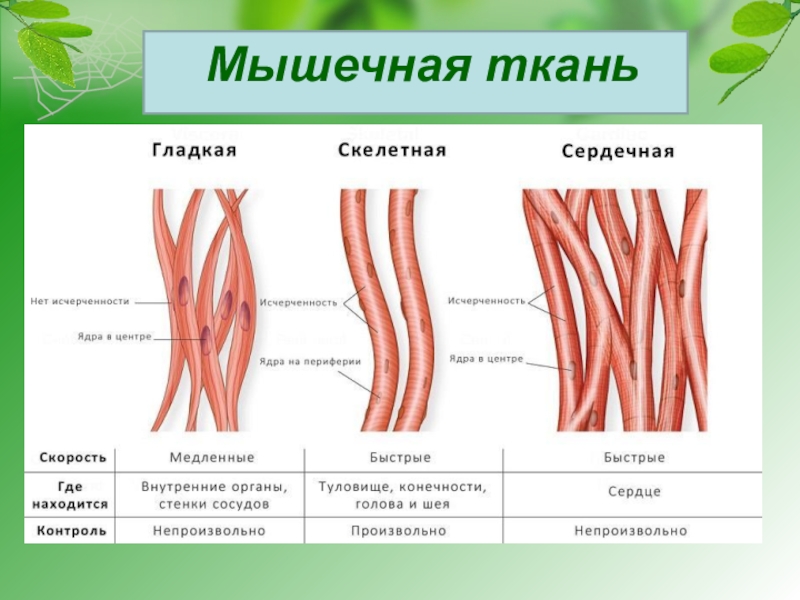
Функциональное недержание – когда нормальной регуляции мочеиспускания препятствуют психические или физические (невозможность быстро дойти до туалета) проблемы, притом, что сама выделительная система в порядке. К функциональному недержанию приводят болезнь Паркинсона, болезнь Альцгеймера и другие формы деменции.
Ургентное недержание. Люди, у которых происходит утечка мочи после внезапного, острого желания помочиться страдают ургентным недержанием. Это происходит из-за повышения тонуса мышц мочевого пузыря, что может явиться следствием большого числа причин, таких как: психическое напряжение, повреждение нервов (при диабете, инсульте), инфекция, камни в мочевом пузыре, опухоль. В этой ситуации необходимо полное обследование, чтобы выявить первопричину и назначить адекватное лечение. Острое недержание чаще всего начинается с острых позывов к мочеиспусканию при невозможности его осуществить. Это состояние прогрессирует до той точки, когда контролировать процесс мочеиспускания становится невозможно. При ургентном недержании показана терапия лекарственными препаратами, которые помогают регулировать тонус мочевого пузыря, а также упражнения Кегеля для укрепления мышц тазового дна и «приучение» мочевого пузыря к работе с определенным интервалом.
При ургентном недержании показана терапия лекарственными препаратами, которые помогают регулировать тонус мочевого пузыря, а также упражнения Кегеля для укрепления мышц тазового дна и «приучение» мочевого пузыря к работе с определенным интервалом.
Стрессовое недержание. Каждая пятая женщина в возрасте после 40 знает, что такое стрессовое недержание. Это когда при кашле, смехе, чихании, физических нагрузках (особенно – повышающих внутрибрюшное давление) выделяется небольшая порция мочи. Стрессовое недержание мочи в подавляющем большинстве случаев свойственно женщинам, но не зависит от возраста – с этим знакомы и молодые женщины, и профессиональные спортсменки. Речь не об эмоциональном стрессе, а об ослабленных мышцах сфинктера и тазового дна, что ведет к тому, что стрессовое воздействие, будь то резкое движение или увеличение внутрибрюшного давления, приводит к выдавливанию порции мочи из мочевого пузыря в уретру. В данной ситуации могут помочь упражнения для укрепления мышц тазового дна и гимнастика Кегеля.
Недержание от переполнения. Этот вид недержания распространен среди пожилых мужчин. Причиной в большинстве случаев является доброкачественное разрастание тканей предстательной железы, которое сжимает уретру и влияет на отток мочи. Симптомы этой патологии редко встречаются у мужчин до 40 лет, но уже к 60 более половины сильной половины человечества познакомятся с симптомами недержания от переполнения, а к 80 годам их количество увеличится до 90%. Из-за закупорки уретры моча постоянно скапливается в пузыре, растягивая его стенки. Симптомы варьируются, но наиболее распространенные из них – затрудненное мочеиспускание (невозможность помочиться, слабая струя), подтекание мочи, частые позывы к мочеиспусканию, особенно в ночное время. Проблемы с мочеиспусканием не обязательно вызваны увеличением простаты. Лечение чаще всего хирургическое или катетеризация мочевого пузыря.
Смешанный тип недержания. Представляет собой комбинацию симптомов стрессового и ургентного недержания.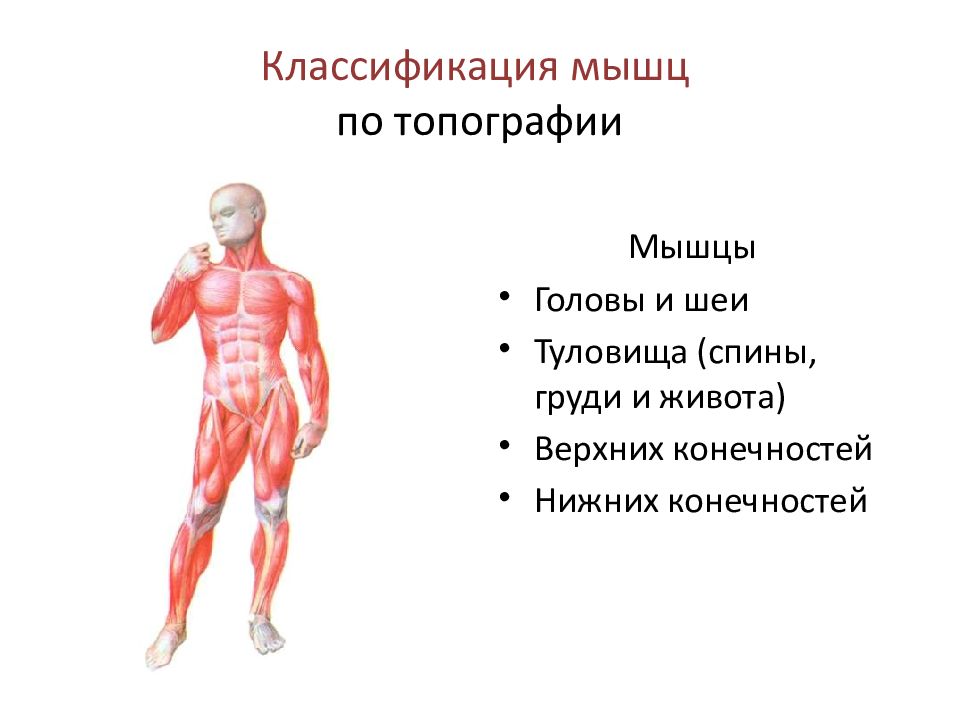 Встречается у значительного количества пациентов. Для выявления причин необходимо полное обследование.
Встречается у значительного количества пациентов. Для выявления причин необходимо полное обследование.
Что это такое, функции, расположение и анатомия
Обзор
Что такое скелетная мышца?
Большинство мышц вашего тела — скелетные. Они составляют от 30 до 40% от общей массы вашего тела. Сухожилия (жесткие полосы соединительной ткани) прикрепляют ткань скелетных мышц к костям по всему телу. Мышцы плеча, подколенного сухожилия и мышцы живота являются примерами скелетных мышц.
В чем разница между скелетными, сердечными и гладкими мышцами?
В вашем теле есть три типа мышц:
- Скелетные мышцы: Скелетные мышцы — это произвольные мышцы, то есть вы контролируете, как и когда они двигаются и работают.Нервы в вашей соматической нервной системе посылают сигналы, чтобы заставить их функционировать. Если вы тянетесь за книгой на полке, вы задействуете скелетные мышцы шеи, рук и плеч.
- Сердечная мышца: Сердечная мышца находится только в вашем сердце.
 Они помогают сердцу перекачивать кровь по всему телу. Это непроизвольные мышцы, которые контролирует вегетативная нервная система. Это означает, что они работают, даже если вам не нужно об этом думать.
Они помогают сердцу перекачивать кровь по всему телу. Это непроизвольные мышцы, которые контролирует вегетативная нервная система. Это означает, что они работают, даже если вам не нужно об этом думать. - Гладкая мышца: Гладкая мышца образует ваши органы, кровеносные сосуды, пищеварительный тракт, кожу и другие области.Гладкие мышцы тоже непроизвольны. Итак, ваша вегетативная нервная система тоже контролирует их. Например, мышцы мочевыделительной системы помогают избавиться от шлаков и токсинов.
Функция
Для чего нужны скелетные мышцы?
Скелетные мышцы — жизненно важная часть вашего опорно-двигательного аппарата. Они выполняют множество функций, в том числе:
- Жевание и глотание — первые этапы пищеварения.
- Расширение и сжатие грудной клетки, чтобы вы могли дышать и выдыхать по своему желанию.
- Поддержание осанки.
- Перемещение костей в разных частях тела.
- Защита суставов и удержание их на месте.

Анатомия
Где расположены скелетные мышцы?
В вашем теле есть скелетные мышцы. Они находятся между костями.
Из чего состоят скелетные мышцы?
Скелетные мышцы состоят из гибких мышечных волокон, диаметр которых колеблется от менее полдюйма до чуть более трех дюймов.Эти волокна обычно охватывают длину мышцы. Волокна сокращаются (сжимаются), что позволяет мышцам перемещать кости, поэтому вы можете выполнять множество различных движений.
Как устроены скелетные мышцы?
Каждая мышца может содержать тысячи волокон. Волокна окружают различные типы оболочек или покрытий:
- Эпимизий: Самый внешний слой ткани, окружающий всю мышцу.
- Perimysium: Средний слой, окружающий пучки мышечных волокон.
- Endomysium: Самый внутренний слой, окружающий отдельные мышечные волокна.
Как выглядят скелетные мышцы?
Волокна скелетных мышц красные и белые. Они выглядят полосатыми или полосатыми, поэтому их часто называют поперечно-полосатыми мышцами. Сердечные мышцы также имеют поперечнополосатую форму, а гладкие — нет.
Они выглядят полосатыми или полосатыми, поэтому их часто называют поперечно-полосатыми мышцами. Сердечные мышцы также имеют поперечнополосатую форму, а гладкие — нет.
Насколько тяжелы скелетные мышцы?
Хотя скелетные мышцы обычно составляют примерно 35% веса вашего тела, это может варьироваться от человека к человеку.У мужчин масса скелетных мышц примерно на 36% больше, чем у женщин. Люди с высоким или избыточным весом также имеют более высокую мышечную массу. Мышечная масса уменьшается с возрастом как у мужчин, так и у женщин.
Состояния и расстройства
Какие состояния и нарушения влияют на скелетные мышцы?
Скелетные мышцы могут поражать широкий спектр состояний, от легких травм до серьезных или даже опасных для жизни миопатий (заболеваний, поражающих скелетные мышцы). Некоторые из них:
- Мышечные дистрофии : Эта группа заболеваний вызывает прогрессирующую дегенерацию волокон скелетных мышц.Они являются результатом аномального гена и могут передаваться по наследству (передаваться через семьи).
 Есть много разных мышечных дистрофий.
Есть много разных мышечных дистрофий. - Myasthenia gravis (MG) : Это аутоиммунное заболевание препятствует тому, чтобы мышцы и нервы взаимодействовали должным образом. Это приводит к сильной мышечной слабости и утомляемости. MG может затруднить движение, ходить, говорить, жевать, видеть, держать голову вверх или держать веки открытыми. Это может даже привести к серьезным проблемам с дыханием.
- Рабдомиолиз : Это опасное для жизни состояние вызывает разрушение мышечной ткани.Поврежденные мышцы выделяют в кровь белки, электролиты и другие вещества. Это может привести к серьезному повреждению органов. Травматические повреждения, тепловой удар или сильное перенапряжение могут вызвать рабдомиолиз.
- Саркопения : С возрастом мы постепенно теряем массу скелетных мышц. Саркопения начинается примерно в 40 лет. К 80 годам мы теряем около 50% нашей мышечной массы. Саркопения может привести к потере функции, подвижности, проблемам с равновесием и падению.
 Ожирение, гормональные изменения и другие состояния здоровья могут ускорить потерю мышечной массы.
Ожирение, гормональные изменения и другие состояния здоровья могут ускорить потерю мышечной массы. - Штаммы : Мышечные деформации или растяжения мышц возникают, когда вы чрезмерно растягиваете мышечные волокна. Эти травмы обычно возникают в результате чрезмерного использования. Сильное напряжение может привести к частичному или полному разрыву мышц.
- Тендинит : Сухожилия соединяют скелетные мышцы с костями. Воспаление сухожилий может затруднить работу этих мышц. Как и деформации, тендинит обычно вызывается переутомлением сухожилий.
Насколько распространены состояния скелетных мышц?
Некоторые состояния скелетных мышц, такие как растяжения и возрастная дегенерация, действительно встречаются.Мышечные травмы составляют от 10 до 55% всех спортивных травм, и около 90% из них связаны с растяжением. Остальные встречаются довольно редко. Например, миастения поражает от 14 до 40 человек из каждых 100000 в США
.
уход
Как сохранить здоровье скелетных мышц?
Позаботьтесь о своих скелетных мышцах:
- Выполнение регулярных силовых тренировок и упражнений на сопротивление.
- Соблюдайте полноценную и сбалансированную диету.
- Поддержание здоровой массы тела.
- Растяжка и разминка мышц перед физической нагрузкой.
Часто задаваемые вопросы
Когда мне следует позвонить своему врачу?
Обратитесь к врачу прямо сейчас, если вы:
- Не может пошевелить ни одной частью тела.
- Ощущение онемения лица или конечностей.
- Сильная внезапная мышечная боль или судороги.
- Очень темная моча или низкий диурез.
- Чувство чрезмерной слабости или усталости.
Записка из клиники Кливленда
Скелетные мышцы — самые распространенные мышцы вашего тела. Вы используете их для движения костей, поэтому они играют жизненно важную роль в повседневной деятельности. Травмы или заболевания скелетных мышц могут сильно повлиять на вашу жизнь. Важно, чтобы ваши мышцы были максимально сильными и здоровыми.
Травмы или заболевания скелетных мышц могут сильно повлиять на вашу жизнь. Важно, чтобы ваши мышцы были максимально сильными и здоровыми.
Типы мышц, функции и общие состояния
Обзор
Что такое мышцы?
Мышцы — это мягкие ткани.Мышцы состоят из множества эластичных волокон. В вашем теле более 600 мышц. У разных типов мышц разная работа. Некоторые мышцы помогают бегать, прыгать или выполнять такие деликатные задачи, как заправка нити в иглу. Другие мышцы позволяют дышать или переваривать пищу. Ваше сердце — это трудолюбивая мышца, которая бьется тысячи раз в день.
Многие расстройства, травмы и заболевания могут повлиять на работу мышц. Эти состояния могут вызвать мышечную боль, мышечные спазмы или мышечную слабость. Более серьезные нарушения могут привести к параличу.Кардиомиопатия и другие сердечные заболевания затрудняют перекачку крови через тело сердца.
Здоровый образ жизни помогает вашим мышцам работать так, как они должны. Вы можете сохранить свои мышцы сильными, поддерживая здоровый вес, соблюдая сбалансированную диету и много упражняясь. Обязательно регулярно посещайте своего врача для проверки на наличие заболеваний и состояний, которые могут привести к проблемам с мышцами.
Вы можете сохранить свои мышцы сильными, поддерживая здоровый вес, соблюдая сбалансированную диету и много упражняясь. Обязательно регулярно посещайте своего врача для проверки на наличие заболеваний и состояний, которые могут привести к проблемам с мышцами.
Какие бывают типы мышц?
Вы произвольно управляете некоторыми мышцами с помощью своей нервной системы (командного центра вашего тела).Вы заставляете их двигаться, думая о том, чтобы переместить их.
Другие мышцы работают непроизвольно, а это значит, что вы не можете их контролировать. Они делают свою работу автоматически. Чтобы работать, они принимают сигналы от других систем организма, таких как пищеварительная система или сердечно-сосудистая система.
В теле есть три типа мышечной ткани. Их:
- Скелетные: Как часть опорно-двигательного аппарата, эти мышцы работают с вашими костями, сухожилиями и связками.Сухожилия прикрепляют скелетные мышцы к костям по всему телу.
 Вместе они поддерживают вес вашего тела и помогают двигаться. Вы контролируете эти произвольные мышцы. Некоторые мышечные волокна сокращаются быстро и используют короткие всплески энергии (быстро сокращающиеся мышцы). Другие двигаются медленно, например мышцы спины, которые помогают сохранять осанку.
Вместе они поддерживают вес вашего тела и помогают двигаться. Вы контролируете эти произвольные мышцы. Некоторые мышечные волокна сокращаются быстро и используют короткие всплески энергии (быстро сокращающиеся мышцы). Другие двигаются медленно, например мышцы спины, которые помогают сохранять осанку. - Сердечный: Эти мышцы выстилают стенки сердца. Они помогают сердцу перекачивать кровь, которая проходит через сердечно-сосудистую систему. Вы не контролируете сердечные мышцы.Ваше сердце подсказывает им, когда нужно сжиматься.
- Гладкие: Эти мышцы выстилают внутренние поверхности таких органов, как мочевой пузырь, желудок и кишечник. Гладкие мышцы играют важную роль во многих системах организма, включая женскую репродуктивную систему, мужскую репродуктивную систему, мочевыделительную систему и дыхательную систему. Эти типы мышц работают, даже если вам не нужно о них думать. Они выполняют важную работу, например перемещают отходы через кишечник и помогают легким расширяться, когда вы дышите.

Функция
Что делают мышцы?
Мышцы играют роль почти во всех системах и функциях тела. Помогают разные виды мышц:
- Дыхание, речь и глотание.
- Переваривание пищи и избавление от мусора.
- Двигаться, сидеть неподвижно и стоять прямо.
- Перекачивание крови через сердце и сосуды.
- Толкает ребенка через родовые пути, когда мышцы матки сокращаются и расслабляются.
- Зрение и слух.
Анатомия
Как выглядят мышцы?
Все типы мышечной ткани похожи. Но есть небольшие отличия во внешнем виде:
- Скелетные мышцы: Многие отдельные волокна составляют скелетные мышцы. Актин и миозин — это белки, из которых состоят волокна. Пучки волокон образуют веретенообразную форму (длинные и прямые с заостренными концами). Мембрана окружает каждый шпиндель. Провайдеры описывают скелетные мышцы как полосатые (полосатые) из-за полосатого рисунка, который образуют веретена.

- Сердечные мышцы: Эти поперечно-полосатые мышцы похожи на скелетные мышцы. Специальные клетки, называемые кардиомиоцитами, составляют волокна сердечных мышц. Кардиомиоциты помогают сердцу биться.
-
Гладкие мышцы: Белки актин и миозин также образуют гладкие мышечные волокна. В скелетных мышцах эти белки объединяются, образуя веретенообразную форму. В гладких мышцах эти белки появляются в виде листов. Листы придают этой мышечной ткани гладкий вид.
У вас есть мускулы любого размера. Самая большая мышца — это большая ягодичная мышца (мышца, составляющая вашу ягодицу). Самая маленькая мышца — это стременик, который находится глубоко внутри уха. Эта крошечная мышца помогает вам слышать, контролируя вибрацию и движение мелких костей в ухе.
Состояния и расстройства
Какие состояния и нарушения влияют на мышцы?
Различные расстройства, заболевания, лекарства и травмы могут вызвать проблемы с работой мышц. В их числе:
В их числе:
- Рак и другие заболевания: Множественные типы рака (например, саркома) и другие заболевания могут приводить к проблемам с мышцами. К ним относятся нервно-мышечные заболевания, такие как боковой амиотрофический склероз (БАС), аутоиммунные нарушения, такие как миастения (MG), и многие типы миопатий (мышечные заболевания). Заболевание, называемое полимиозитом, вызывает воспаление в мышцах, что приводит к мышечной слабости.
- Сердечно-сосудистые заболевания: Некоторые виды заболеваний вен и сердечно-сосудистых заболеваний, включая ишемическую болезнь сердца, могут вызывать проблемы с сердцем и кровеносными сосудами.Сердечный приступ может произойти, когда мышцы кровеносных сосудов ослабнут.
- Хронические болевые расстройства: Фибромиалгия и другие расстройства вызывают хроническую боль в мышцах по всему телу.
- Генетические нарушения: Мышечная дистрофия — это наследственное заболевание (передается от семьи).
 Существует более 30 видов мышечной дистрофии. Заболевание вызывает стойкую мышечную слабость.
Существует более 30 видов мышечной дистрофии. Заболевание вызывает стойкую мышечную слабость. - Инфекции: Бактериальные и вирусные инфекции могут повреждать мышечные волокна.Эти инфекции включают болезнь Лайма, малярию и пятнистую лихорадку Скалистых гор.
- Травмы: Многие травмы могут привести к разрыву мышц или слишком сильному растяжению (растяжение мышц). Растяжение спины — очень распространенная травма. Несчастные случаи, травмы и травмы, вызванные чрезмерным использованием, могут вызвать мышечные судороги или мышечные спазмы. В тяжелых случаях эти травмы могут привести к параличу.
- Лекарства: Некоторые лекарства, например химиотерапевтические препараты, могут вызывать мышечную боль. Боль в мышцах также может быть результатом приема лекарств, которые лечат высокое кровяное давление.У некоторых людей мышечная слабость развивается после тяжелой аллергической реакции на лекарство или токсичное вещество.
Что происходит с мышцами во время и после тренировки?
У многих людей после тренировки болят мышцы. Болезненность возникает из-за крошечных разрывов (микротрещин), которые возникают при нагрузке на мышцу. Обычно болезненность мышц наступает через день или два после интенсивных упражнений. Вот почему врачи называют это состояние отсроченной болезненностью мышц (DOMS).
По мере того, как мышцы восстанавливаются и заживают крошечные разрывы, мышечная ткань воспаляется.В течение нескольких дней ваши мышцы восстанавливаются, и воспаление проходит. При продолжении упражнений мышечная ткань разрывается и восстанавливается снова и снова. Этот процесс заставляет мышцы становиться больше.
Каковы общие признаки или симптомы состояний, поражающих мышцы?
К наиболее частым признакам мышечных проблем относятся:
Многие из этих симптомов не обязательно означают, что что-то не так. Боль в мышцах или слабость часто проходят после отдыха и гидратации.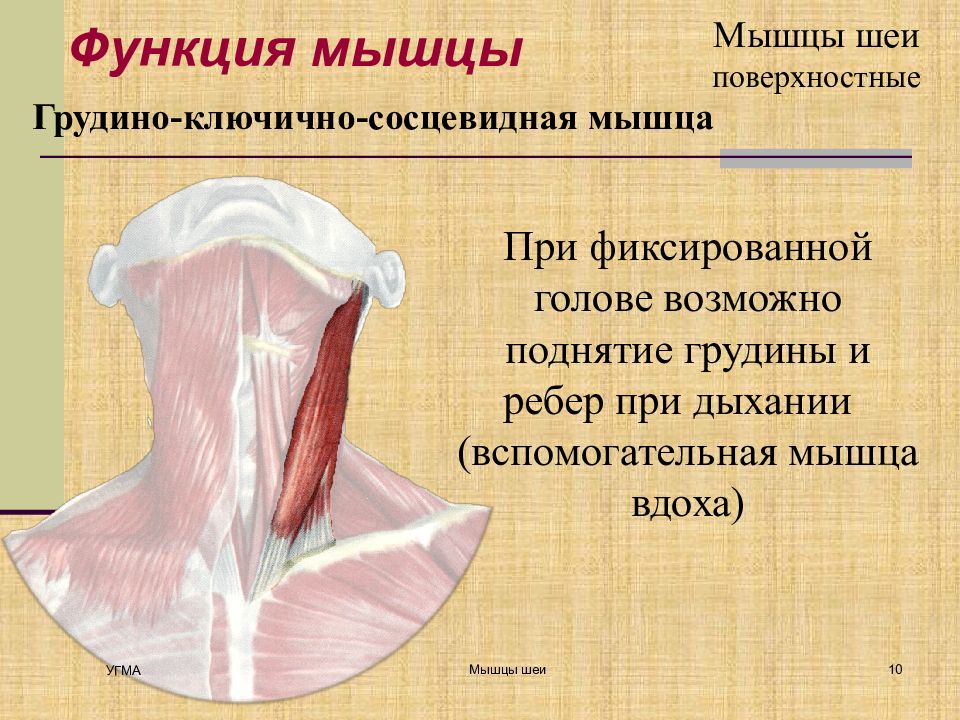 Если какой-либо из этих симптомов возникнет внезапно, немедленно поговорите со своим врачом.Внезапная мышечная слабость или боль могут быть признаками серьезного состояния здоровья.
Если какой-либо из этих симптомов возникнет внезапно, немедленно поговорите со своим врачом.Внезапная мышечная слабость или боль могут быть признаками серьезного состояния здоровья.
Какие общие тесты для проверки здоровья мышц?
В зависимости от ваших симптомов ваш врач может порекомендовать:
- Полный анализ крови (CBC), серия анализов крови, позволяющих оценить ваше общее состояние здоровья и выявить наличие инфекции.
- Электромиография (ЭМГ) для измерения работы нервов и мышц.
- Визуальные исследования, такие как МРТ, которые показывают изображения повреждений мышц.
- Биопсия мышцы для проверки образца мышечной ткани на наличие заболевания.
уход
Как сохранить здоровье мускулов?
Чтобы ваши мышцы оставались здоровыми, вам следует сосредоточиться на том, чтобы оставаться здоровым в целом:
- Делайте много упражнений: Сохранение активности может сохранить все ваши мышцы сильными, в том числе сердце.
 Попробуйте сочетать сердечно-сосудистую деятельность и упражнения с отягощениями. Поговорите со своим врачом о программе упражнений, которая подходит именно вам.Во избежание травм обязательно хорошенько разминайтесь перед тренировкой. У вас меньше шансов повредить теплые мышцы.
Попробуйте сочетать сердечно-сосудистую деятельность и упражнения с отягощениями. Поговорите со своим врачом о программе упражнений, которая подходит именно вам.Во избежание травм обязательно хорошенько разминайтесь перед тренировкой. У вас меньше шансов повредить теплые мышцы. - Правильно питайтесь и делайте правильный выбор: Обязательно соблюдайте сбалансированную диету, чтобы ваши мышцы оставались сильными. Избегайте употребления натрия и трансжиров (например, в жареной пище), которые могут привести к сердечным заболеваниям. Если вы курите, поговорите со своим поставщиком медицинских услуг о плане, который поможет вам бросить курить.
- Поддерживайте здоровый вес: Перенос лишних килограммов может привести к травмам. Это также увеличивает риск проблем со здоровьем, включая высокое кровяное давление.Поговорите со своим врачом о наиболее подходящем весе для вашего тела и образа жизни. Если вы несете лишний вес, спросите своего врача о плане контроля веса.

- Отдыхайте, когда вам нужно: Дайте мышцам время восстановиться после напряжения. Вам также следует отдохнуть, если вы почувствуете болезненность после тяжелых упражнений. Если дать мышцам время на восстановление и восстановление, это поможет вам избежать травм.
- Запланировать регулярные осмотры: Регулярно посещайте своего провайдера. Пройдите обследование на наличие заболеваний, повышающих риск мышечных проблем.Постоянное наблюдение за своим здоровьем позволяет вашему провайдеру обнаруживать проблемы на ранней стадии. Вот тогда лечение более эффективно.
Часто задаваемые вопросы
Когда мне следует позвонить своему врачу по поводу моих мышц?
Если у вас внезапно возникла мышечная слабость или мышечная боль, немедленно позвоните своему врачу. Получите неотложную медицинскую помощь, если у вас проблемы с дыханием или глотанием, или если у вас есть изменения зрения, боль в груди или проблемы с равновесием. Это могут быть признаки серьезного состояния здоровья.
Это могут быть признаки серьезного состояния здоровья.
Записка из клиники Кливленда
Ваши мышцы играют важную роль в поддержании вашей жизни и в взаимодействии с окружающим миром. Некоторые мышцы помогают вам видеть, слышать и двигаться. Другие отвечают за то, чтобы помочь вам дышать или переваривать пищу. Каждый человек с возрастом теряет некоторую мышечную массу. Чтобы ваши мышцы работали должным образом, вы должны поддерживать здоровый вес, много заниматься спортом и соблюдать сбалансированную диету. Посещайте своего поставщика медицинских услуг для регулярных обследований, которые могут выявить проблемы со здоровьем, которые могут привести к проблемам с мышцами.
Потеря мышечной функции | UF Health, University of Florida Health
Определение
Потеря мышечной функции — это когда мышца не работает или не двигается нормально. Медицинский термин, обозначающий полную потерю мышечной функции, — паралич.
Альтернативные имена
Паралич; Парез; Потеря движения; Двигательная дисфункция
Соображения
Потеря мышечной функции может быть вызвана:
- Заболеванием самой мышцы (миопатия)
- Заболеванием области, где встречаются мышца и нерв (нервно-мышечное соединение)
- Заболеванием нервная система: повреждение нервов (невропатия), повреждение спинного мозга (миелопатия) или повреждение головного мозга (инсульт или другое повреждение головного мозга)
Утрата мышечной функции после событий такого типа может быть серьезной. В некоторых случаях мышечная сила может не полностью восстановиться даже после лечения.
В некоторых случаях мышечная сила может не полностью восстановиться даже после лечения.
Паралич может быть временным или постоянным. Он может поражать небольшую область (локализованный или очаговый) или быть широко распространенным (генерализованным). Это может повлиять на одну сторону (одностороннее) или на обе стороны (двустороннее).
Если паралич поражает нижнюю половину тела и обе ноги, это называется параплегией. Если поражаются обе руки и ноги, это называется квадриплегией. Если паралич поражает мышцы, вызывающие дыхание, это быстро становится опасным для жизни.
Причины
Заболевания мышц, вызывающие потерю мышечной функции, включают:
Заболевания нервной системы, вызывающие потерю мышечной функции, включают:
Уход на дому
Внезапная потеря мышечной функции требует неотложной медицинской помощи. Немедленно обратитесь за медицинской помощью.
После того, как вы прошли курс лечения, ваш лечащий врач может порекомендовать некоторые из следующих мер:
- Соблюдайте предписанную терапию.

- Если нервы на лице или голове повреждены, у вас могут возникнуть трудности с жеванием и глотанием или с закрытием глаз.В этих случаях может быть рекомендована мягкая диета. Вам также понадобятся средства защиты глаз, например повязка на глаз во время сна.
- Длительная неподвижность может вызвать серьезные осложнения. Часто меняйте позу и берегите кожу. Упражнения на диапазон движений могут помочь поддерживать некоторый мышечный тонус.
- Шины могут помочь предотвратить мышечные контрактуры — состояние, при котором мышца постоянно укорачивается.
Когда обращаться к медицинскому работнику
Мышечный паралич всегда требует немедленной медицинской помощи.Если вы заметили постепенное ослабление или проблемы с мышцами, как можно скорее обратитесь за медицинской помощью.
Чего ожидать при посещении вашего офиса
Врач проведет медицинский осмотр и задаст вопросы о вашей истории болезни и симптомах, в том числе:
Местоположение:
- Какие части вашего тела затронуты?
- Это влияет на одну или обе стороны тела?
- Происходило ли оно по схеме «сверху вниз» (нисходящий паралич) или по схеме «снизу вверх» (восходящий паралич)?
- Вам трудно встать со стула или подняться по лестнице?
- Вам трудно поднять руку над головой?
- У вас есть проблемы с разгибанием или поднятием запястья (запястье опускается)?
- Есть ли у вас трудности с захватом?
Симптомы:
- Боль?
- Есть ли у вас онемение, покалывание или потеря чувствительности?
- Есть ли у вас трудности с контролем над мочевым пузырем или кишечником?
- У вас одышка?
- Какие еще симптомы у вас есть?
Временной график:
- Повторяются ли эпизоды (повторяющиеся)?
- Как долго они служат?
- Утрата мышечной функции ухудшается (прогрессирует)?
- Медленно или быстро?
- Стало ли хуже с течением дня?
Отягчающие и успокаивающие факторы:
- Что усугубляет паралич?
- Стало ли хуже после приема добавок калия или других лекарств?
- А после отдыха лучше?
Можно провести следующие тесты:
В тяжелых случаях может потребоваться внутривенное кормление или зонд для кормления. Могут быть рекомендованы физиотерапия, трудотерапия или логопедия.
Могут быть рекомендованы физиотерапия, трудотерапия или логопедия.
Изображений
Источников
Эволи А., Винсент А. Нарушения нервно-мышечной передачи. В: Goldman L, Schafer AI, ред. Гольдман-Сесил Медицина . 26-е изд. Филадельфия, Пенсильвания: Эльзевьер; 2020: глава 394.
Селцен Д. Заболевания мышц. В: Goldman L, Schafer AI, ред. Гольдман-Сесил Медицина . 26-е изд. Филадельфия, Пенсильвания: Эльзевьер; 2020: глава 393.
Warner WC, Sawyer JR.Нервно-мышечные расстройства. В: Azar FM, Beaty JH, Canale ST, ред. Оперативная ортопедия Кэмпбелла . 13-е изд. Филадельфия, Пенсильвания: Эльзевьер; 2017: глава 35.
MACF1 контролирует функцию скелетных мышц через микротрубочко-зависимую локализацию внесинаптических миоядер и биогенез митохондрий
[Примечание редакции: авторы повторно представили исправленную версию статьи на рассмотрение. Ниже приводится ответ авторов на первый раунд проверки.]
Рецензент №1:
В этой рукописи Ghasemizadeh et al.
исследовали роль MACF1 скелетных мышц, белка, связанного с сетью микротрубочек, в поддержании миоядер и локализации митохондрий в зрелых мышечных клетках. Основные выводы: (1) MACF1 регулирует расположение миоядер на этапах созревания миофибрилл, но не на ранних этапах их формирования, (2) MACF1 контролирует локализацию и структурную целостность митохондрий мышц, (3) нокдаун MACF1 в мышечных трубках с shRNA снижает образование агрин-индуцированных кластеров AChR и увеличивает движения миоядер, (4) потеря функции MACF1, в частности, в мышцах (условная мышца-KO) вызывает неправильную локализацию миоядер у взрослых, но не у молодых мышей, изменяется структурная целостность НМС у более молодых мышей и увеличивает содержание митохондрий у взрослых мышей, (7) мышцы условных мышей Macf1 демонстрируют более высокое сопротивление усталости по сравнению с мышцами контрольных мышей.
В целом, это интересное исследование, направленное на ответ на важный вопрос и обеспечивающее важное механистическое понимание роли мышцы MACF1 в расположении миоядер и структурной целостности митохондрий и нервно-мышечных соединений.
Представленные данные кажутся убедительными и хорошо обсуждаются в свете предыдущей литературы. Однако необходимо решить некоторые проблемы, чтобы сделать рукопись более заметной.
Мы благодарим рецензента за его положительные комментарии и надеемся, что теперь все его опасения устранены.
1. Было бы полезно узнать о субклеточном расположении MACF1 в мышечных клетках (если антитело действительно работает в иммуноцитохимии).
Чтобы определить субклеточную локализацию MACF1, мы выполнили иммуноокрашивание образцов мышц (миофибры in vitro и ex vivo) с использованием кроличьих поликлональных антител от Novus Biology.
Как представлено на Фигуре 1B, иммуноокрашивание показало, что MACF1 сконцентрирован на периферии зрелых первичных миофибрилл in vitro.Кроме того, он выглядит как точки вокруг миоядер, следующие за линиями, которые колокализуются с сетью микротрубочек (в основном в коре миофибриллярных волокон, а также внутри миофибрилл). Интересно, что не наблюдается совместной локализации MACF1 с актиновой сетью. Подтверждая специфичность антитела, иммуноокрашивание MACF1 было сильно снижено в миофибриллах, обработанных Macf1 -shRNA (рис. 1G).
Интересно, что не наблюдается совместной локализации MACF1 с актиновой сетью. Подтверждая специфичность антитела, иммуноокрашивание MACF1 было сильно снижено в миофибриллах, обработанных Macf1 -shRNA (рис. 1G).
В соответствии с этими данными, выделенные ex vivo миофибриллы Tibialis Anterior от мышей дикого типа показали четкое перинуклеарное накопление MACF1 (рис. 3D, верхние панели).Кроме того, MACF1 колокализуется как с продольной, так и с поперечной сетью микротрубочек, но не с сетью актина (Figure 3D, верхние панели). Как и ожидалось, маркировка MACF1 была почти полностью потеряна в мышечно-специфической условной модели мыши MACF1 с нокаутом (рис. 3D, нижние панели).
Благодаря этому подходу с иммуноокрашиванием мы теперь смогли указать на субклеточную локализацию MACF1, , то есть вокруг миоядер и в ассоциации с микротрубочками в волокнах скелетных мышц.
2. На рис. 1 D и E, MHC-положительные клетки, по-видимому, встречаются с низкой частотой, что указывает на то, что большинство клеток остаются недифференцированными миоблатами или мертвыми клетками.
Это поднимает вопрос о том, являются ли миобласты или мышечные трубки основным источником экспрессии белка MACF1, наблюдаемого в экстрактах общего белка с помощью вестерн-блоттинга. Авторы должны решить эту проблему.
Чтобы ответить на этот вопрос, мы проанализировали присутствие белка MACF1 в первичных мышечных клетках, когда они дифференцируются в мышечные трубки (рис. 1C).В растущих (и недифференцированных) миобластах (GM) MACF1 не обнаружен. Однако мы четко наблюдали присутствие MACF1 через 3 и 5 дней дифференцировки (рис. 1C). Таким образом, эти результаты показывают, что мышечные трубки являются основным источником MACF1 во время процесса дифференцировки. Эта панель теперь перенесена на дополнительный рисунок 1E.
3. Авторы утверждают, что MACF1 прогрессивно экспрессируется и накапливается во время процессов дифференцировки и созревания мышц.Однако данные, подтверждающие это утверждение, отсутствуют. Напротив, уровень экспрессии MACF1, по-видимому, менее выражен через 10 дней дифференцировки мышц (Рисунок 1 C).
Было бы полезно изучить уровни экспрессии MACF1 во время процессов дифференцировки и созревания мышечных клеток как in vivo, так и in vitro.
Чтобы изучить уровни экспрессии MACF1 in vitro, мы выполнили вестерн-блоттинг как первичных мышечных клеток (рисунок 1C и дополнительный рисунок 1C), так и линии миогенных клеток C2C12 (дополнительный рисунок 1D).Очевидно, мы наблюдали всплеск экспрессии MACF1 при сравнении пролиферативных миобластов и 3-дневных дифференцированных мышечных трубок (рисунок 1C и дополнительный рисунок 1C, 1D). Однако уровень белка MACF1 снижался в первичных клетках от 3 до 5 дней дифференцировки. in vivo, мы провели подход кПЦР на образцах из Tibialis Anterior мышей контрольной (Cre-) и Macf1 -cKO (Cre +) мышей, подтвердив, что экспрессия мРНК Macf1 относительно стабильна в зрелых мышцах (изображение ответа автора 1).
Учитывая эти наблюдения, рукопись была исправлена, выделив всплеск экспрессии MACF1 по мере дифференциации мышечных клеток (строки 120-123).
QRT-PCR, представляющие относительный уровень экспрессии генов (по отношению к генам домашнего хозяйства) для MACF1 в
Macf1cKO Cre- и Cre + в каждом данном возрасте.4.На рисунке 2 авторы показали, что количество точек AChR было значительно увеличено в мышечных трубках, обработанных кшРНК MAFC1. Однако необычная форма и размер флуоресцентного BTX вызвали некоторые опасения по поводу специфичности окрашивания. Этого можно исключить, пропитав все AChR немеченым BTX, а затем добавив флуоресцентно меченый BTX. В этом случае вы не должны ожидать появления пятен.
Α-Бунгаротоксин (aBTX) — это змеиный яд, который нацелен на рецептор ацетилхолина (AchR).Когда aBTX наносится на зрелые миофибриллы, экстрагированные ex vivo, он связывает и специфически окрашивает постсинаптическую зону НМС (Tzartos SJ & Changeux JP, 1983, PMID: 11894953).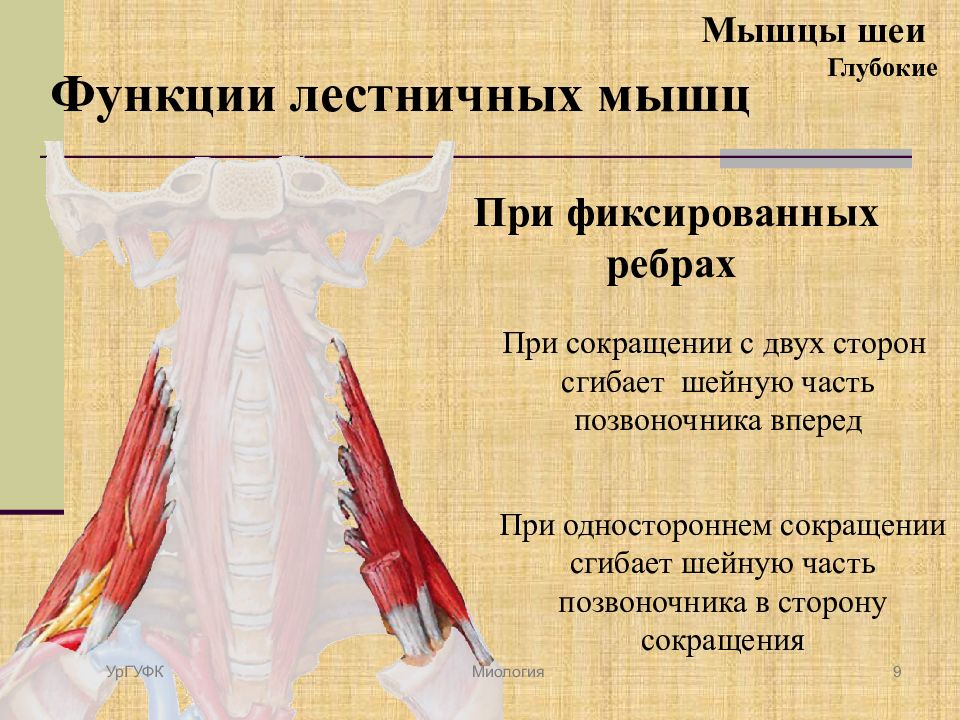 На рисунке 2 aBTX непосредственно применялся на зрелых миофибриллах. В этом состоянии при окрашивании aBTX выявляются длинные (более 10 мкм) или точечные кластеры AChR (Vilmont et al., 2016, PMID: 27283349). Чтобы еще больше подтвердить специфичность нашего окрашивания, разработанные in vitro зрелые миофибриллы, обработанные scramble или Macf1 shRNA, сначала были помечены BTX, соединенным с Alexa-мукой 555 (BTX-AF555, разведение 1/500), промыты, а затем помечены BTX, связанный с Alexa-мукой 647 (BTX-AF647, разведение 1/500).
На рисунке 2 aBTX непосредственно применялся на зрелых миофибриллах. В этом состоянии при окрашивании aBTX выявляются длинные (более 10 мкм) или точечные кластеры AChR (Vilmont et al., 2016, PMID: 27283349). Чтобы еще больше подтвердить специфичность нашего окрашивания, разработанные in vitro зрелые миофибриллы, обработанные scramble или Macf1 shRNA, сначала были помечены BTX, соединенным с Alexa-мукой 555 (BTX-AF555, разведение 1/500), промыты, а затем помечены BTX, связанный с Alexa-мукой 647 (BTX-AF647, разведение 1/500).
Как показано на изображении ответа автора 2, для aBTX-AF555 был обнаружен сильный сигнал, тогда как для aBTX-AF647 сигнал почти не был виден. Это подтвердило надежность нашего окрашивания aBTX.
ОкрашиваниеaBTX в первичных миофибриллах, трансфицированных scramble или MACF1 shRNA.
5.
Вызывает недоумение то, что неправильная локализация миоядер в условных мышцах MACF1-KO происходит только через год, в то время как в НМС количество расположенных субсинаптических ядер было изменено намного раньше (4 месяца). Авт. Должны исследовать идею, зависит ли расположение несинаптических и субсинаптических миоядер от активности MCF1. Один из возможных экспериментов — денервировать мышцы мутантного и дикого типов и увидеть расположение / количество миоядер.
Мы благодарим рецензента за этот комментарий, который позволил нам лучше различать, какие миоядра находятся под контролем MACF1 в миофибриллах.
Наши предыдущие данные показали, что миоядра были интернализованы у 12-месячных мышей. Чтобы определить, повлияла ли внесинаптическая локализация миоядер, мы сначала извлекли мышечных волокон Tibialis Anterior у мышей MACF1 cKO и количественно определили расстояние между миоядрами у мышей в возрасте 2, 6 и 12 месяцев. Этот подход показал, что внесинаптическое позиционирование миоядер затронуто у мышей MACF1 cKO по сравнению с контролем. Действительно, мы наблюдали уменьшение расстояния между ближайшими миоядрами на 20% через 2 месяца, на 40% через 6 месяцев и до 70% через 12 месяцев (Рисунок 4B).Таким образом, как предполагают наши результаты in vitro (Рис. 1J), изменение внесинаптических миоядер, распространяющихся на периферию миофибрилл, происходит преждевременно. Напротив, интернализация миоядер является поздним событием, сопутствующим дезорганизации микротрубочек и промежуточных филаментов (Рис. 3 и 7G).
Действительно, мы наблюдали уменьшение расстояния между ближайшими миоядрами на 20% через 2 месяца, на 40% через 6 месяцев и до 70% через 12 месяцев (Рисунок 4B).Таким образом, как предполагают наши результаты in vitro (Рис. 1J), изменение внесинаптических миоядер, распространяющихся на периферию миофибрилл, происходит преждевременно. Напротив, интернализация миоядер является поздним событием, сопутствующим дезорганизации микротрубочек и промежуточных филаментов (Рис. 3 и 7G).
Во втором подходе мы количественно оценили количество синаптических миоядер (ниже НМС) во время старения. Интересно, что мы обнаружили, что количество синаптических миоядер увеличивается у контрольных мышей, с 7 у молодых мышей до 11 у старых мышей, в то время как это число относительно стабильно (около 7) у мышей MACF1 cKO (рисунок 5D).Поскольку НМС у мышей MACF1 KO меньше и представляют собой кластеры фрагментированных AChR по сравнению с контролем (рис. 5B-C), мы предположили, что сбой в рекрутировании синаптических миоядер, наблюдаемый у мышей MACF1 cKO, преимущественно вызван сбоями в кластеризации AChR ( Рисунок 5B-C).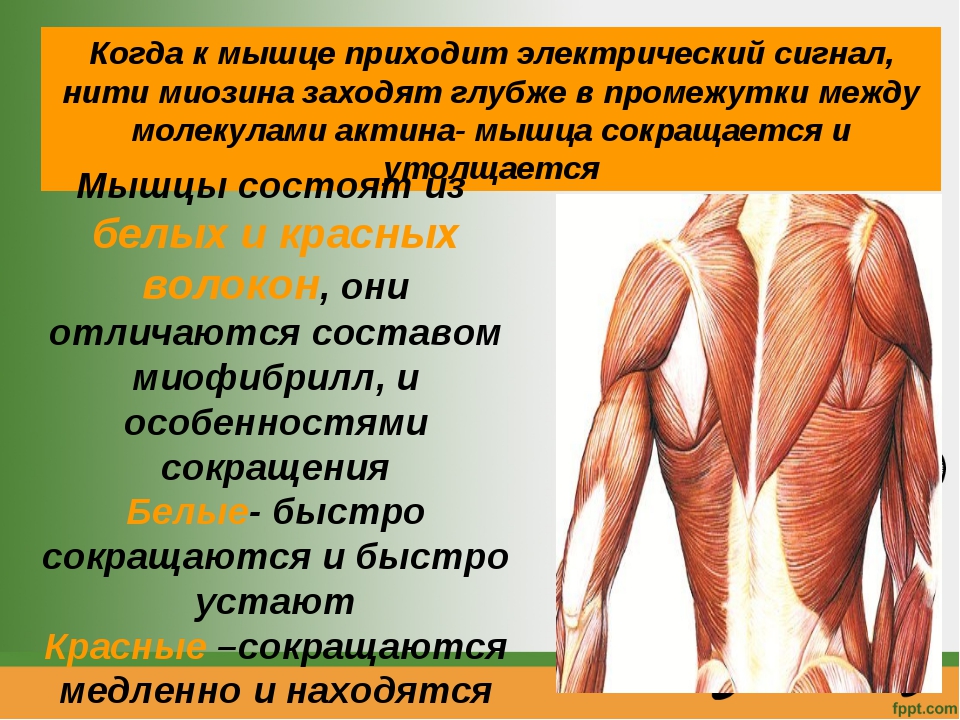 Чтобы проверить эту гипотезу, мы провели эксперимент по денервации мышц. Раздавление седалищного нерва, как было опубликовано ранее (Milanič et al., 1999, PMID: 10421467), уменьшает как миофибриллы (площадь поперечного сечения, рис. 5H), так и площадь НМС (рис. 5I) через 15 дней.Эти эффекты были сходными у мышей дикого типа и у мышей MACF1 cKO (фиг. 5H-I). Мы также наблюдали аналогичную потерю синаптических миоядер в НМС после эксперимента по денервации (1 синаптическое миоядро), предполагая, что MACF1 не принимает непосредственного участия в позиционировании синаптических миоядер в НМС.
Чтобы проверить эту гипотезу, мы провели эксперимент по денервации мышц. Раздавление седалищного нерва, как было опубликовано ранее (Milanič et al., 1999, PMID: 10421467), уменьшает как миофибриллы (площадь поперечного сечения, рис. 5H), так и площадь НМС (рис. 5I) через 15 дней.Эти эффекты были сходными у мышей дикого типа и у мышей MACF1 cKO (фиг. 5H-I). Мы также наблюдали аналогичную потерю синаптических миоядер в НМС после эксперимента по денервации (1 синаптическое миоядро), предполагая, что MACF1 не принимает непосредственного участия в позиционировании синаптических миоядер в НМС.
Следовательно, эти данные показывают, что MACF1 регулирует непосредственно динамику внесинаптических миоядер на периферии мышечных волокон, а не контролирует синаптические миоядра.
6. Авторам следует также посмотреть, чувствительны ли синаптические митохондрии к изменениям у более молодых мышей MACF1 с условными мышцами-KO.
Чтобы ответить на этот вопрос, мы исследовали организацию митохондрий на изолированных миофибриллах, выделенных из старых мышей дикого типа и MACF1 cKO. Как показано на рис. 7 G и H, общая организация митохондрий изменена у взрослых мышей MACF1 cKO по сравнению с контролем. Однако мы не наблюдали изменений в плотности митохондрий в НМС у мышей, лишенных MACF1, по сравнению с контрольными мышами (изображение ответа автора 3).
Как показано на рис. 7 G и H, общая организация митохондрий изменена у взрослых мышей MACF1 cKO по сравнению с контролем. Однако мы не наблюдали изменений в плотности митохондрий в НМС у мышей, лишенных MACF1, по сравнению с контрольными мышами (изображение ответа автора 3).
Следовательно, эти данные показывают, что MACF1 изменяет организацию несинаптических митохондрий.
Репрезентативные изображения конфокальной микроскопии (63X) плотности митохондрий (отраженные окрашиванием цитохромом-C) в НМС (отраженные окрашиванием BTX) извлеченных
Tibialis anterior 12-месячных мышей MACF1 Creand Cre +.В наложении митохондрии представлены зеленым, постсинаптические НМС — красным, а синаптические ядра — синим.Шкала шкалы 10 мкм.
Рецензент № 2:
В этой рукописи Ghasemizadeh et al.
Проанализировали функцию MACF1, белка, перекрестно связывающего микротрубочки с актином, в мышцах. Авторы использовали нокаут shRNA в условиях культивирования, а также специфический нокаут мышечной ткани на мышиной модели для своего анализа. Они показывают, что позиционирование миоядер in vitro и in vivo нарушено в условно нокаутированных мышцах мышей, и что позиционирование и морфология митохондрий нарушены.Оба фенотипа могут быть связаны с дестабилизированной динамикой микротрубочек в мышцах, как наблюдают авторы. Кроме того, наблюдалась аномальная кластеризация рецепторов ацетилхолина в нервно-мышечном соединении.
Описанные результаты интересны, однако они могут не быть связаны друг с другом, и их новизна ограничена. Авторы не предоставляют нового механистического понимания какого-либо из описанных процессов, а скорее предоставляют набор наблюдений и фенотипов, наблюдаемых после нокаута MACF1 в мышцах.
Мы уважительно не согласны с этим рецензентом, поскольку наша работа представляет ранее неисследованную роль MACF1 в контроле динамики / стабильности микротрубочек и, следовательно, внесинаптического позиционирования миоядер, кластеризации AchRs и биогенеза митохондрий. Теперь мы предоставили новые механистические данные, которые дополнительно подтверждают важную роль MACF1 в поддержании локализации миоядер и общей функции волокон скелетных мышц.
Теперь мы предоставили новые механистические данные, которые дополнительно подтверждают важную роль MACF1 в поддержании локализации миоядер и общей функции волокон скелетных мышц.
1. Авторы использовали мышей с условным нокаутом MACF1, описанных в Goryunov et al., 2010. В этой модели авт. Удалили экзоны 6 и 7, кодирующие C-концевую половину актин-связывающего домена MACF1 (ABD). MACF1 — это большой белок с отдельными доменами, включая домены ABD, Plakin, микротрубочек и EB1. Авторы предполагают, что основной вклад MACF-1 в мышцы связан с его функцией в качестве белка, связывающего микротрубочки. Однако усеченный белок (продуцируемый у мышей KO) лишен только части домена ABD и все еще содержит функциональные домены связывания плакина и микротрубочек.Возможно, что наблюдаемый слабый фенотип не соответствует основной функции MACF1 в мышцах. Авторы не рассматривали и не обсуждали эту возможность.
Действительно, Горюнов и др. заметил, что условная делеция экзонов 6 и 7 из Macf1 не устраняет присутствие изоформы MACF1c. Насколько нам известно, эта изоформа уникальна для нервной системы (Cusseddu et al. , 2021, PMID: 33816492). Подтверждая эти данные, мы не наблюдали присутствия такой изоформы или усеченного белка в нашей мышечной системе (рисунок 1c и дополнительный рисунок 1c).Следовательно, изоформа MACF1c не может быть ответственной за наблюдаемый мягкий фенотип.
Насколько нам известно, эта изоформа уникальна для нервной системы (Cusseddu et al. , 2021, PMID: 33816492). Подтверждая эти данные, мы не наблюдали присутствия такой изоформы или усеченного белка в нашей мышечной системе (рисунок 1c и дополнительный рисунок 1c).Следовательно, изоформа MACF1c не может быть ответственной за наблюдаемый мягкий фенотип.
У мышечно-специфичных условных мышей KO MACF1 анализ qPCR показал, что экспрессия мРНК Macf1 была снижена на 50% у 2-месячных мышей и на 70% у 12-месячных мышей (дополнительная фигура 3A). Частичное снижение экспрессии Macf1 может быть связано с несколькими параметрами. Во-первых, эксперименты qPCR были выполнены на всех мышечных экстрактах, которые не отражают только экспрессию Macf1 в миофибриллах.В самом деле, скелетные мышцы содержат несколько типов клеток, в которых экспрессия Macf1 и не изменена у мышечно-специфичных MACF1 условных KO мышей. Затем мышечные стволовые клетки вносят вклад в неповрежденный гомеостаз миофибрилл на протяжении всей жизни (Keefe et al. , 2015, PMID: 25971691, Pawlikowski et al. , 2015, PMID: 26668715), тем самым обеспечивая новые миоядра, в которых MACF1 еще не истощен. . Кроме того, мРНК Macf1 может быть доставлена в зрелое миофибриллярное волокно после слияния мышечных стволовых клеток.Вместе это может частично ослабить действие рекомбиназы Cre и позволить поддерживать небольшие количества белка MACF1 в миофибриллах.
, 2015, PMID: 25971691, Pawlikowski et al. , 2015, PMID: 26668715), тем самым обеспечивая новые миоядра, в которых MACF1 еще не истощен. . Кроме того, мРНК Macf1 может быть доставлена в зрелое миофибриллярное волокно после слияния мышечных стволовых клеток.Вместе это может частично ослабить действие рекомбиназы Cre и позволить поддерживать небольшие количества белка MACF1 в миофибриллах.
2. Многие из фенотипов, описанных авторами, уже были проиллюстрированы на других модельных организмах, включая дефекты миоядерного позиционирования, миоядерную сферичность и дезорганизацию микротрубочек (например, Wang et al., 2015; Kim et al., 2010, Bottenberg et al. ., 2009). Фенотип фрагментации митохондрий является новым и интересным, однако авт. Не предоставляют механистического объяснения этого наблюдения, а только предполагают, что это происходит из-за аберрантной организации микротрубочек.
Как показано в первой версии нашей рукописи, мы наблюдали увеличение количества митохондриальных частиц после подавления MACF1. Это явление, которое было названо «фрагментацией митохондрий», привело нас к гипотезе о том, что потеря MACF1 нарушает целостность митохондриальной сети. Дезорганизуя сеть микротрубочек (MT), подавление MACF1 будет влиять на экспрессию или уровень активности MT-ассоциированных белков, таких как DNM1 или DNM2, которые, как известно, участвуют в делении или слиянии митохондрий.
Это явление, которое было названо «фрагментацией митохондрий», привело нас к гипотезе о том, что потеря MACF1 нарушает целостность митохондриальной сети. Дезорганизуя сеть микротрубочек (MT), подавление MACF1 будет влиять на экспрессию или уровень активности MT-ассоциированных белков, таких как DNM1 или DNM2, которые, как известно, участвуют в делении или слиянии митохондрий.
Чтобы исследовать эту гипотезу, мы провели следующие эксперименты. Во-первых, экспрессия мРНК акторов деления / слияния митохондрий была изучена как in vitro, так и in vivo. Никаких различий не наблюдали в уровнях экспрессии Dnm1 или Dnm2 , когда MACF1 подавляется либо in vitro (изображение ответа автора 4), либо in vivo у мыши MACF1 cKO (фигура 7F). Следовательно, потеря MACF1 не влияет на процессы деления / слияния митохондрий.
Результаты QRT-PCR, представляющие относительный уровень экспрессии генов DNM1 (слева) и DNM2 (справа) по отношению к генам домашнего хозяйства в коротких дифференцированных мышечных трубках или долгосрочно дифференцированных миофибриллах, трансфицированных скремблированной si / shRNA или пулом si / shRNA против MACF1.
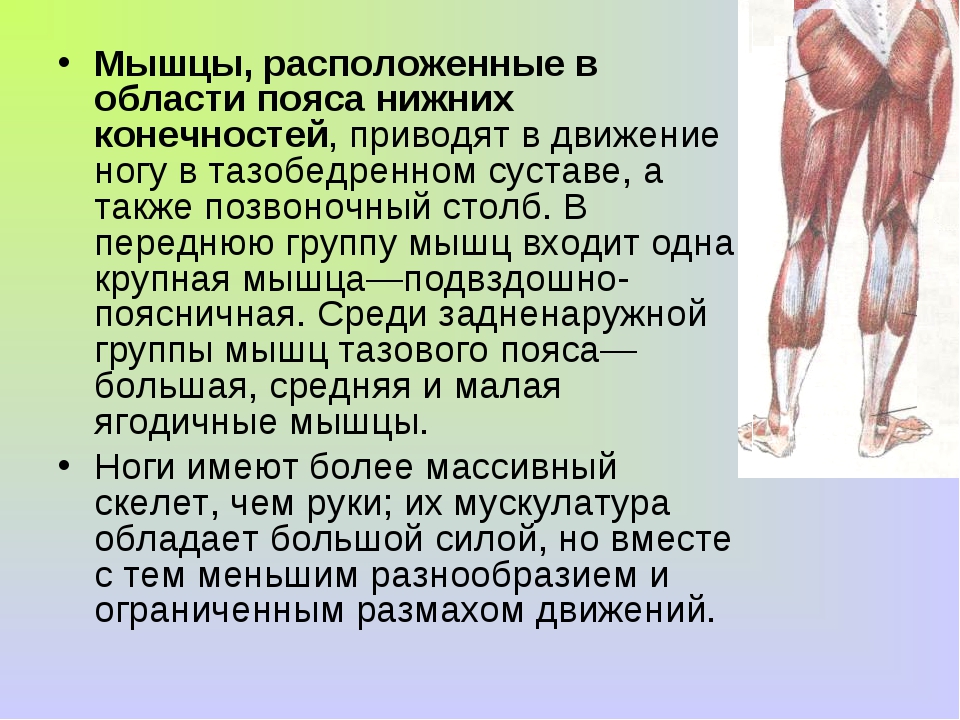
Однако гисто-энзимологическое окрашивание SDH in vivo (фиг. 7B) и q-PCR анализ количества митохондриальной ДНК (фиг. 7E) выявили увеличение количества митохондрий у взрослых мышей MACF1 KO по сравнению с контрольными мышами.Это предполагает возможное усиление митохондриального биогенеза.
Интересно, что различные исследования связывают повышение уровня сарколипина (белка, связанного с саркоплазматическим ретикулумом в скелетных мышцах) с биогенезом митохондрий (Santosh K. Maurya et al., 2015, PMID: 25713078; Santosh K. Maurya et al., 2018, PMID : 30208317). Когда мы оценили уровень экспрессии сарколипина в мышиной модели MACF1 cKO, мы обнаружили, что мРНК сарколипина демонстрирует 40-кратное увеличение в мышцах взрослых мышей MACF1 KO по сравнению с контролем (фигура 7F).
В целом, эти данные показывают, что потеря MACF1 увеличивает митохондриальный биогенез, а не модифицирует слияние, деление или фрагментацию митохондрий.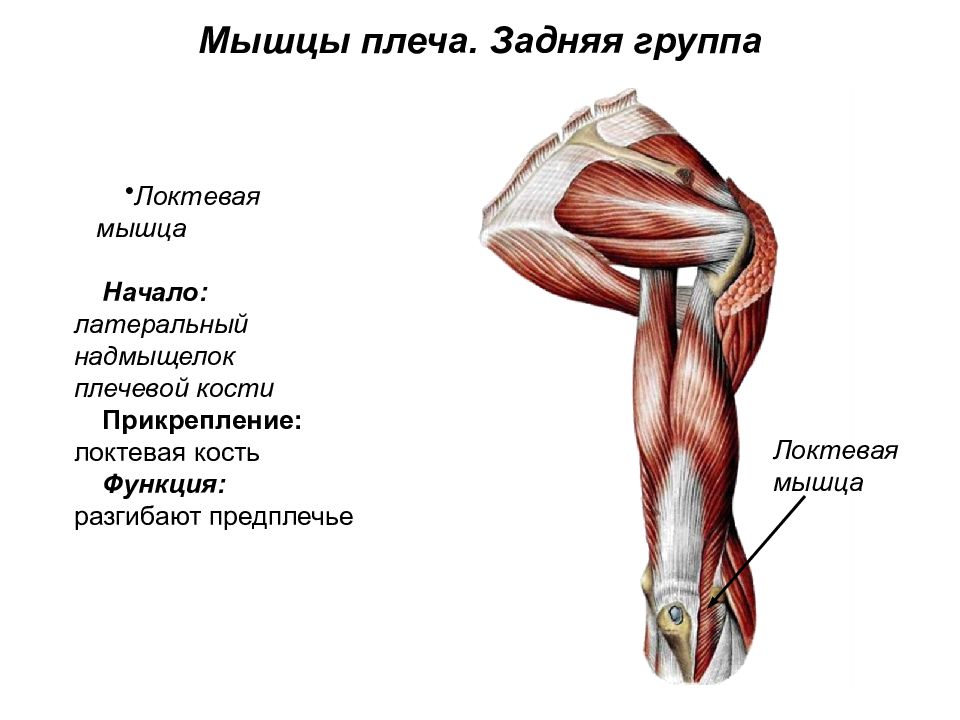 Мы уточнили наш вывод и соответствующим образом изменили текст. Увеличение количества митохондриальных частиц in vitro теперь вводится как увеличение содержания митохондриальных частиц.
Мы уточнили наш вывод и соответствующим образом изменили текст. Увеличение количества митохондриальных частиц in vitro теперь вводится как увеличение содержания митохондриальных частиц.
3. Некоторые изображения неубедительны: Рисунок 1 — разница между контролем и миРНК MACF1 минимальна и может быть связана с разной длиной миофибрилл.Рисунок 2 — актин не кажется нормальным. Различия в организации микротрубочек недостаточно ясны. ЭМ-микрофотографии среднего качества и не позволяют читателю видеть детали митохондрий. Требуются изображения с большим увеличением.
Мы согласны с рецензентом и теперь предоставили изображения с большим увеличением, которые подтверждают наши выводы.
Для уменьшенного расстояния между соседними миоядрами в миофибриллах MACF1-KD по сравнению с контрольными миофибрами (рис. 1J) нет явной разницы в длине и / или форме миофибрилл.Мы добавили увеличенное изображение клеточной культуры, чтобы лучше оценить неизмененную культуру миофибрилл (рис. 1D). Чтобы исследовать организацию актина в волокнах скелетных мышц, длину саркомера измеряли на изображениях с использованием окрашивания F-актина в контроле и миофибр MACF1-KD ( Рисунок 1F). Никаких существенных различий не наблюдалось, показывая, что MACF1 не участвует в формировании паттерна актиновой сети в миофибриллах. Эти результаты согласуются с нашими результатами in vivo, представляющими одинаковую длину саркомера у мышей дикого типа и у мышей MACF1 cKO (рисунок 3E и дополнительный рисунок 3F).
1D). Чтобы исследовать организацию актина в волокнах скелетных мышц, длину саркомера измеряли на изображениях с использованием окрашивания F-актина в контроле и миофибр MACF1-KD ( Рисунок 1F). Никаких существенных различий не наблюдалось, показывая, что MACF1 не участвует в формировании паттерна актиновой сети в миофибриллах. Эти результаты согласуются с нашими результатами in vivo, представляющими одинаковую длину саркомера у мышей дикого типа и у мышей MACF1 cKO (рисунок 3E и дополнительный рисунок 3F).
Чтобы проанализировать изменения в сети микротрубочек вокруг миоядер, интенсивность микротрубочек измеряли в перинуклеарной области. Как показано на рисунке 1H, отсутствие MACF1 снижает интенсивность микротрубочек вокруг миоядер до 50%. Панели на Рисунке 1G были добавлены, чтобы визуально выделить разницу.
Для электронной микроскопии изображения с более высоким разрешением представляют скопление митохондрий круглой формы между миофибриллами у мышей MACF1 KO по сравнению с контрольными мышами (рис. 7J-K).
7J-K).
4. Авторы должны показать, где находится MACF1 в нормальной мышце. Перекрывается ли это с МП? Он локализован на ядерной мембране? перекрывается ли он с тирозином или нетирозином? Это перекрывается с EB1?
См. Комментарий №1 рецензента №1.
Иммуноокрашивание проводили для определения локализации MACF1 в миофибриллах in vitro и in vivo. in vitro MACF1 концентрируется вокруг миоядер и совместно с кортикальными микротрубочками в зрелых первичных миофибриллах in vitro (рис. 1B).Кроме того, мы показали, что домен связывания микротрубочек MACF1, помеченный GFP, колокализуется с тирозинированным тубулином в незрелых мышечных трубках в виде кометоподобного концевого связывания белка (рис. 2А).
in vivo, иммуноокрашивание выявило явное накопление MACF1 вокруг миоядер в изолированных миофибриллах Tibialis Anterior мышей WT (рис. 3D, верхние панели). Кроме того, MACF1 колокализуется как с продольными, так и с поперечными микротрубочками в дополнение к перинуклеарной области.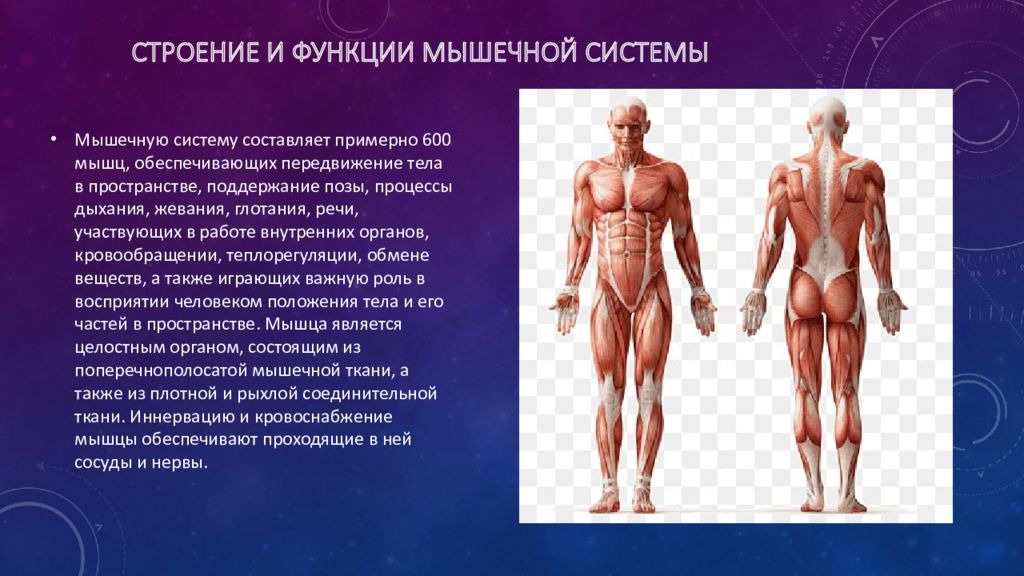 Эти результаты указывают на специфическую локализацию MACF1 вокруг миоядер и в ассоциации с микротрубочками в волокнах скелетных мышц.
Эти результаты указывают на специфическую локализацию MACF1 вокруг миоядер и в ассоциации с микротрубочками в волокнах скелетных мышц.
Рецензент № 3:
За исключением сгруппированных миоядер в нервно-мышечном соединении (НМС), многоядерные клетки скелетных мышц позиционируют большую часть своих ядер, чтобы оптимизировать расстояния между ними. Более того, клетки скелетных мышц разработали механизмы для поддержания этих положений, несмотря на сокращение мышц. В рукописи основное внимание уделяется определению роли связывающего микротрубочки белка MACF1 в мышечных клетках мышей с помощью серии экспериментов in vitro и in vivo.Исследователи сообщают, что MACF1 связан с миоядерным позиционированием в НМС и по всему волокну, позиционированием / структурой митохондрий, организацией Т-канальцев и кластеризацией рецепторов ацетилхолина в НМС. Авторы предполагают, что ключевая роль MACF1 заключается в регуляции посттрансляционных модификаций микротрубочек и, как следствие, динамики микротрубочек.
Они утверждают, что изменения в динамике микротрубочек, обнаруживаемые при потере MACF1, связаны с дефицитом постсинаптического ядерного позиционирования и функции, миоядерного и митохондриального позиционирования и, в конечном итоге, мышечной функции.
Авт. Предоставляют большое количество данных in vivo и in vitro, чтобы развить свое утверждение, что MACF1 играет критическую роль в созревании и функционировании мышц.
Мы благодарим рецензента за то, что он оценил объем данных, предоставленных в поддержку нашего вывода.
Они обнаружили, что потеря / уменьшение MACF1 приводит к ряду нарушений в зрелом волокне: к ним относятся морфологические дефекты, включая аберрантное миоядерное положение, особенно постсинаптических ядер, снижение кластеризации ACHR в синапсе, неправильное расположение и морфологию митохондрий, T- аномалии канальцев и т. д. и функциональные дефекты мышц (уменьшенная сила, но большая выносливость).Авторы пытаются связать мириады дефектов мышечных волокон с одним лежащим в основе структурным изменением волокон: аномальной динамикой микротрубочек.
Их доказательства подтверждают это: изменения в количестве тирозинированных / детирозированных микротрубочек (которые должны быть количественно определены, но не являются) и изменения скорости кометы EB1 (также должны быть включены число и числа траектории). Оба они указывают на высокодинамичные микротрубочки в ko / kd миофибриллах и некоторую роль MACF1 в контроле динамичности микротрубочек (прямо или косвенно).Принимая во внимание, что основные выводы основываются на заключении о динамике контрольных микротрубочек MACF1, необходимо предоставить некоторую дополнительную поддержку, которая соединяет MACF1 с микротрубочками с динамикой микротрубочек и впоследствии эти измененные MTs с др. Описанными фенотипами. Например, приводит ли избыточная экспрессия MACF1 к увеличению стабильных микротрубочек? Как это влияет на кластеризацию AChR, кластеризацию миоядер в синапсе или даже на структуры Т-канальцев? Привлекает ли MACF1 ферменты, которые посттрансляционно модифицируют MT в MT? Где MACF1 экспрессируется или обогащается миофибром мыши в разном возрасте?
Мы провели ряд дополнительных экспериментов, чтобы укрепить наш вывод о том, что MACF1 контролирует динамику микротрубочек и его потеря неизбежно приводит к функциональным нарушениям зрелого миофибра.
Что касается локализации MACF1 в мышечных волокнах, аналогичные вопросы были заданы рецензентом №1 / вопрос №1 и рецензентом №2 / вопросом №4. Мы благодарим всех трех наших рецензентов за то, что они указали на отсутствие такой важной информации. Таким образом, использование кроличьих поликлональных антител от Novus bios, также подтвержденных на предмет их специфичности с помощью вестерн-блоттинга, позволило нам определить локализацию MACF1 в мышечных волокнах in vitro и ex vivo (рисунки 1 и 3). Используя этот подход к иммуноокрашиванию, мы подтвердили MACF1 как эксклюзивный белок, связывающий микротрубочки, который колокализуется с сетью микротрубочек и накапливается преимущественно в перинуклеарной области.Поскольку MACF1 представляет собой огромный белок с двумя основными изоформами на 830 и 630 кДа, сверхэкспрессия полноразмерного MACF1 in vitro с использованием плазмиды экспериментально невозможна. Однако мы сверхэкспрессировали домен связывания микротрубочек (MTBD) MACF1 в нашей системе in vitro. MACF1-MTBD сформировал пучки с предпочтительным расположением вокруг миоядер (рис. 2В). Это указывает на то, что MACF1-MTBD отвечает за перинуклеарное позиционирование эндогенного MACF1.
MACF1-MTBD сформировал пучки с предпочтительным расположением вокруг миоядер (рис. 2В). Это указывает на то, что MACF1-MTBD отвечает за перинуклеарное позиционирование эндогенного MACF1.
Кроме того, мы проанализировали скорость комет EB3 по мере созревания миофибрилл в контроле и в миофибриллах, истощенных по MACF1 (Рисунок 2C).Эти данные показывают (1), что во время формирования миофибрилл скорость кометы EB3 снижается и (2) что истощение MACF1 поддерживает скорость комет EB3 на высоком уровне. Все вместе эти данные демонстрируют, что MACF1 контролирует динамическую скорость микротрубочек по мере созревания миофибрилл.
Также важно: попросите носителя английского языка отредактировать рукопись, так как есть много проблем с грамматикой, выбором слов и структурой предложения. См. Также некоторые комментарии, перечисленные ниже для каждого рисунка.
Наша рукопись проверена всеми нашими сотрудниками.
https://doi. org/10.7554/eLife.70490.sa2
org/10.7554/eLife.70490.sa2
Что такое мышечная система? — Функции и как мышцы работают в группах — Видео и стенограмма урока
Анатомия мышц
Сначала давайте кратко рассмотрим сами мышцы. Как и многое другое, мышцы состоят из множества более мелких частей. Начнем с уровня мышечного волокна. Мышечное волокно — это просто мышечная клетка или строительные блоки мышц. Понимаете, точно так же, как остальная часть вашего тела состоит из множества отдельных клеток, ваши мышцы также состоят из клеток.
Множественные волокна соединяются вместе, образуя следующий уровень организации — пучок . Пучок — это группа или пучок мышечных волокон. И подобно тому, как многие мышечные волокна соединяются вместе, образуя пучок, многие пучки соединяются вместе, образуя мышцу.
Мышцы состоят из групп пучков.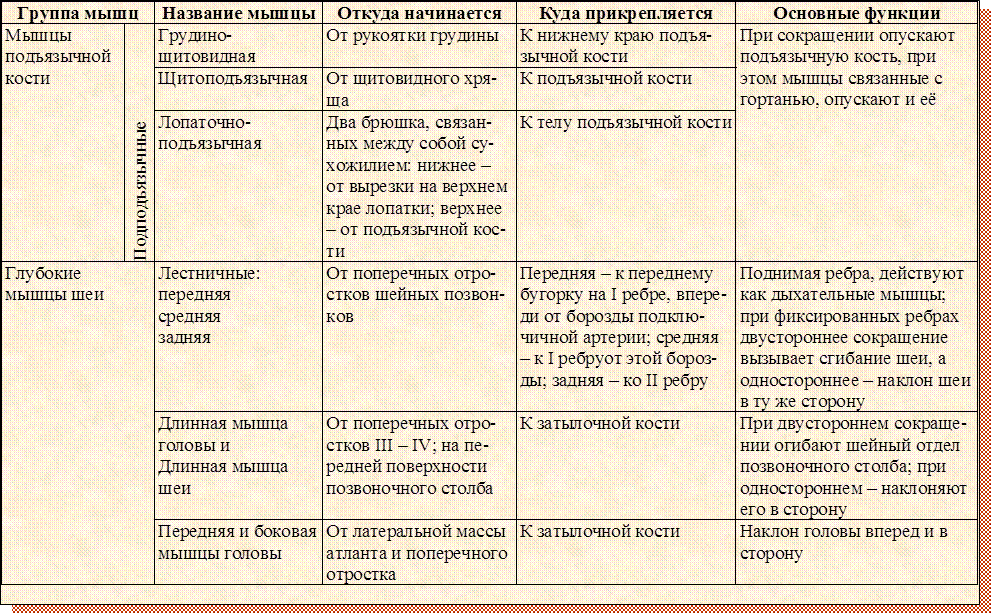 При стимуляции они сокращаются, чтобы произвести движение.Без мышц ваше тело не могло бы функционировать. Больше никаких бегов, разговоров, ходьбы и, ну, вы поняли!
При стимуляции они сокращаются, чтобы произвести движение.Без мышц ваше тело не могло бы функционировать. Больше никаких бегов, разговоров, ходьбы и, ну, вы поняли!
Организация мышцы позволяет всем волокнам и, следовательно, пучкам сокращаться и расслабляться как группа. Сокращение стимулируется нервными импульсами и запускает движение мышцы, а релаксация происходит, когда импульс прекращается и мышца расслабляется, возвращаясь в свое естественное состояние. Этот паттерн сокращения и расслабления отвечает за все движения вашего тела.
Часть вашего тела, которая движется в ответ на сокращение мышц, зависит от расположения и исходной точки самих мышц. Чтобы упростить задачу, мы сосредоточимся на скелетных мышцах тела. Большинство скелетных мышц прикреплены к кости, хрящу или соединительной ткани, что ограничивает или направляет их движение. Например, мышца, прикрепленная к кости руки, будет двигать кость руки только при стимуляции. Он не может двигать костью ноги; следовательно, его движение определяется его точками крепления.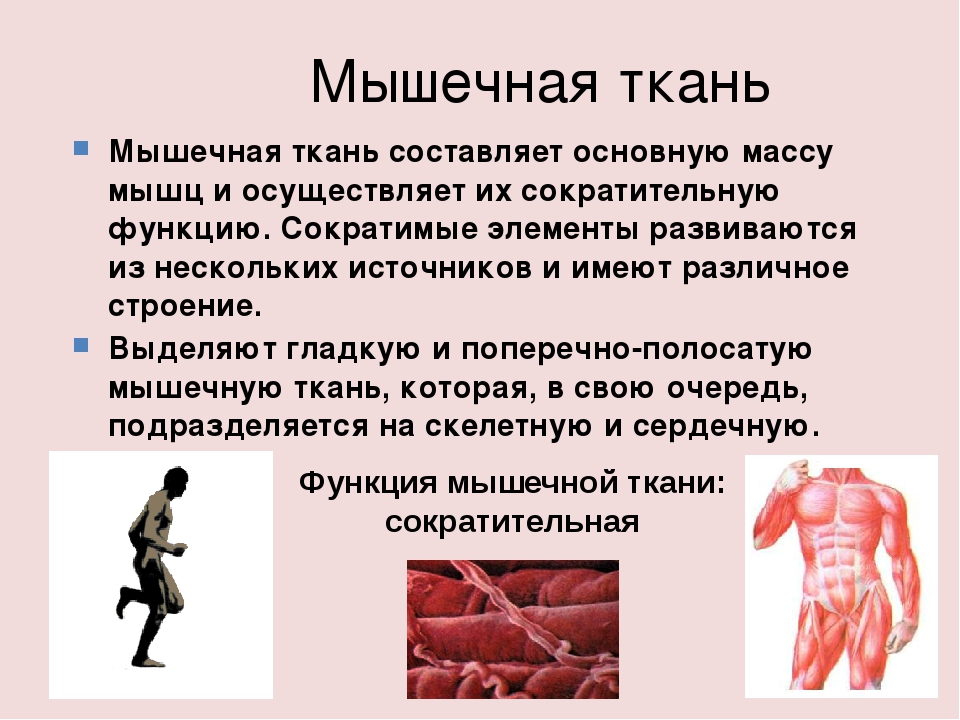
Исходная точка и точка вставки
Каждая мышца имеет две основные точки прикрепления: исходную точку и точку вставки. Начало мышцы — это неподвижный конец, конец, который не движется. Этот конец обычно представляет собой кость, хрящ или соединительную ткань. Противоположный конец мышцы — точка прикрепления . Это подвижный конец мышцы, прикрепленный к перемещаемой структуре.
Вернемся к нашему примеру с мышцами руки.Если мы посмотрим на кость плеча, называемую плечевой костью, то увидим большую мышцу поверх нее. Эта мышца, называемая biceps brachii , имеет точку начала на лопатке, а точку прикрепления — на радиусе предплечья. Когда мышца сокращается, она перемещает радиус вверх по направлению к лопатке, но лопатка и плечо не перемещаются. Вот почему радиус называется точкой вставки, а лопатка — точкой отсчета.
Если вы все еще не знаете, как определить разницу, вы можете попробовать самостоятельно.Когда вы двигаете рукой или ногой, обратите внимание, какой конец мышцы движется, а какой остается. Это поможет вам выяснить, какая вставка (та, которая выполняет перемещение), а какая — начало координат.
Агонисты и антагонисты
Когда мышца движется, это происходит потому, что нервная система стимулирует ее сокращение, но что происходит после этого? Как он возвращается в исходное положение? Для этого ему нужна помощь другой мышцы, которая тянет его назад.
Чтобы одна мышца сократилась, противоположная мышца должна растянуться или расслабиться. Вы можете думать об этом как об игре в перетягивание каната. Когда одна команда тянет к себе веревку, сторона другой команды тянется, и наоборот. Это как взаимные уступки, и большая часть наших скелетных мышц работает в этих парах. Подобно инь и янь мышечной системы, они уравновешивают друг друга.
Официально эти пары состоят из агониста и антагониста. Агонист является основным двигателем, мышца, сокращение которой приводит к определенному движению, в то время как антагонист выполняет действие, противоположное агонисту.
Агонист является основным двигателем, мышца, сокращение которой приводит к определенному движению, в то время как антагонист выполняет действие, противоположное агонисту.
Возвращаясь к нашему примеру с рукой, сокращение бицепса приводит к движению нижней части руки в локтевом суставе. Поскольку бицепс вызывает сгибание руки, он считается агонистом. Но чтобы сократиться, противоположная мышца должна растянуться. Это трицепс.
И наоборот. Когда трицепс сокращается, опуская нижнюю руку обратно в исходное положение, бицепс расслабляется. Большинство, но не все скелетные мышцы вашего тела работают в этих парах агонист-антагонист.Если агонист сгибает мышцу, то его антагонист растягивает мышцу. Если агонист перемещает кость к вашему телу, его антагонист отодвигает ее, и так далее.
Synergists
Хорошо, а как насчет скелетных мышц, которые не являются частью этих пар? Кто они такие? Что ж, они принадлежат к другой группе мышц под названием синергистов . Эти мышцы работают с агонистами, помогая им сокращаться более эффективно. Они делают это, создавая дополнительную тяговую силу в месте введения, помогая начать движение мышцы или помогая стабилизировать мышцу в исходном месте, чтобы помочь контролировать движение.Это своего рода помощник, помогающий боссу выполнять свою работу, или приятель, помогающий супергерою бороться с преступностью. Они играют меньшую, но все же важную роль в работе мышцы.
Эти мышцы работают с агонистами, помогая им сокращаться более эффективно. Они делают это, создавая дополнительную тяговую силу в месте введения, помогая начать движение мышцы или помогая стабилизировать мышцу в исходном месте, чтобы помочь контролировать движение.Это своего рода помощник, помогающий боссу выполнять свою работу, или приятель, помогающий супергерою бороться с преступностью. Они играют меньшую, но все же важную роль в работе мышцы.
Например, под двуглавой мышцей, нашей мышцей-агонистом, находится меньшая мышца, называемая плечевой мышцей. Бицепс прикрепляется только к одной из костей нижней части руки, а плечевая мышца — к другой. Итак, бицепс нуждается в помощи синергиста, плечевой мышцы, чтобы согнуть нижнюю часть руки.
Краткое содержание урока
Итак, хотя запоминание всех мышц мышечной системы может быть непосильной задачей, понять, как они работают вместе, немного легче.Каждая мышца состоит из нескольких пучков, которые работают вместе, чтобы сокращаться одновременно, и каждый пучок состоит из нескольких волокон, которые также работают вместе.
Точно так же, как отдельные части мышцы должны работать вместе, разные отдельные мышцы также должны работать вместе. У вас не может быть агониста без антагониста, как не может быть верха без низа или света без тьмы. Каждый работает вместе с другим. Агонист мышцы — это мышца, которая выполняет основное движение, а его антагонист выполняет противоположное движение.Движение мышцы инициируется сигналом нервной системы, который приказывает мышце сокращаться. Конец мышцы, которая движется во время сокращения, называется точкой прикрепления , а конец мышцы, который остается неподвижным, — точкой начала отсчета .
Надеюсь, теперь, когда вы знаете некоторые основы того, как работает мышечная система и как она устроена, остальное не будет казаться таким устрашающим.
Результаты обучения
После этого видеоурока вы сможете:
- Описывать структуру мышцы
- Обобщите, как сокращаются мышцы
- Различие между исходной точкой и точкой вставки
- Объясните взаимосвязь между мышцами-агонистами, антагонистами и синергистами
Функция мышечного веретена в здоровой и больной мышце | Скелетная мышца

Шеррингтон CS. О проприоцептивной системе, особенно в ее рефлекторном аспекте. Головной мозг. 1907; 29 (4): 467–82.
Артикул Google Scholar
Дитц В. Проприоцепция и двигательные нарушения. Nature Rev Neurosci. 2002; 3 (10): 781–90.
CAS Статья Google Scholar
Кин О. Расшифровка организации контуров позвоночника, управляющих движением.Nature Rev Neurosci. 2016; 17 (4): 224–38.
CAS Статья Google Scholar
Kröger S. Proprioception 2.0: новые функции для мышечных веретен. Curr Opin Neurol. 2018; 31 (5): 592–8.
PubMed Статья CAS Google Scholar
Коул Дж. Д., Седжвик EM. Восприятие силы и движения у мужчины без крупных миелинизированных сенсорных афферентов ниже шеи.J Physiol. 1992; 449: 503–15.
CAS PubMed PubMed Central Статья Google Scholar
Chesler AT, Szczot M, Bharucha-Goebel D, Ceko M, Donkervoort S, Laubacher C, et al. Роль PIEZO2 в механочувствительности человека. New Eng J Med. 2016; 375 (14): 1355–64.
CAS PubMed Статья Google Scholar
Делле Ведове А., Сторбек М., Хеллер Р., Холкер И., Хеббар М., Шукла А. и др.Биаллельная потеря связанного с проприоцепцией PIEZO2 вызывает мышечную атрофию с перинатальным респираторным расстройством, артрогрипозом и сколиозом. Am J Hum Genet. 2016; 99 (6): 1406–8.
CAS PubMed PubMed Central Статья Google Scholar
Халилоглу Г., Беккер К., Темуцин С., Талим Б., Кучуксахин Н., Перганд М. и др. Рецессивная стоп-мутация PIEZO2 вызывает дистальный артрогрипоз со слабостью дистальных мышц, сколиозом и дефектами проприоцепции. J Hum Genet. 2017; 62 (4): 497–501.
J Hum Genet. 2017; 62 (4): 497–501.
CAS PubMed Статья Google Scholar
Махмуд А.А., Нахид Н.А., Нассиф К., Сайид М.С., Ахмед М.Ю., Парвин М. и др. Утрата проприоцепции и канала сенсорных ощущений PIEZO2 у братьев и сестер с прогрессирующей формой контрактуры. Clin Genet. 2017; 91 (3): 470–5.
CAS PubMed Статья Google Scholar
Smith LJ, Norcliffe-Kaufmann L, Palma JA, Kaufmann H, Macefield VG. Нарушение сенсомоторного контроля руки при врожденном отсутствии функциональных мышечных веретен. J Neurophysiol. 2018; 120 (6): 2788–95.
PubMed PubMed Central Статья Google Scholar
Masingue M, Faure J, Sole G, Stojkovic T., Leonard-Louis S. Новая бессмысленная мутация PIEZO2 в семье со сколиозом и проприоцептивным дефектом. Нейромышечный дисорд.2019; 29 (1): 75–9.
PubMed Статья Google Scholar
Бехунова Дж., Герикова Буялкова М., Грас Дж., Тейлор Т., Ихм Ю., Кирхер С. и др. Дистальный артрогрипоз с нарушением проприоцепции и осязания: описание раннего фенотипа у мальчика со сложной гетерозиготностью мутаций PIEZO2 и обзор литературы. Мол Синдромол. 2019; 9 (6): 287–94.
PubMed Статья CAS Google Scholar
Blecher R, Heinemann-Yerushalmi L, Assaraf E, Konstantin N, Chapman JR, Cope TC, Bewick GS, Banks RW, Zelzer E. Новые функции проприоцептивной системы в биологии скелета. Фил Транс Р. Соц Б. 2018; 373: 20170327. http://dx.doi.org/10.1098/rstb.2017.0327.
Блечер Р., Криф С., Галили Т., Ассараф Е., Стерн Т., Анекштейн Ю. и др. Проприоцептивная система регулирует морфологическое восстановление сломанных костей. Cell Rep., 2017; 20 (8): 1775–83.
CAS PubMed PubMed Central Статья Google Scholar

Блечер Р., Криф С., Галили Т., Битон И.Е., Стерн Т., Ассараф Э. и др. Проприоцептивная система вдохновляет выравнивание позвоночника: понимание механизма сколиоза. Dev Cell. 2017; 42 (4): 388–99 e3.
CAS PubMed Статья Google Scholar
Takeoka A, Vollenweider I, Courtine G, Arber S. Обратная связь с мышечным веретеном направляет восстановление опорно-двигательного аппарата и реорганизацию цепи после травмы спинного мозга. Клетка. 2014. 159 (7): 1626–39.
CAS PubMed Статья Google Scholar
Такеока А., Арбер С. Функциональные местные цепи проприоцептивной обратной связи инициируют и поддерживают восстановление опорно-двигательного аппарата после травмы спинного мозга. Cell Rep. 2019; 27 (1): 71–85 e3.
CAS PubMed Статья Google Scholar
Ассараф Э., Блехер Р. , Хайнеман-Йерушалми Л., Криф С., Кармель Винсток Р., Битон И. Э. и др.Piezo2, экспрессируемый в проприоцептивных нейронах, необходим для целостности скелета. Nature Communicat. 2020; 11 (1): 3168.
, Хайнеман-Йерушалми Л., Криф С., Кармель Винсток Р., Битон И. Э. и др.Piezo2, экспрессируемый в проприоцептивных нейронах, необходим для целостности скелета. Nature Communicat. 2020; 11 (1): 3168.
CAS Статья Google Scholar
Мэтьюз ПБ. Там, где шла анатомия, за ней последовала физиология: обзор нашего развивающегося понимания мышечного веретена, того, что оно делает и как работает. J Anat. 2015; 227 (2): 104–14.
PubMed PubMed Central Статья Google Scholar
Proske U, Gandevia SC. Проприоцептивные органы чувств: их роль в передаче сигналов о форме тела, положении и движении тела, а также о мышечной силе. Physiol Rev.2012; 92 (4): 1651–97.
CAS PubMed Статья Google Scholar
Бэнкс Р. У., Баркер Д. В: Engel AG, Францини-Армстронг К., редакторы. Мышечное веретено.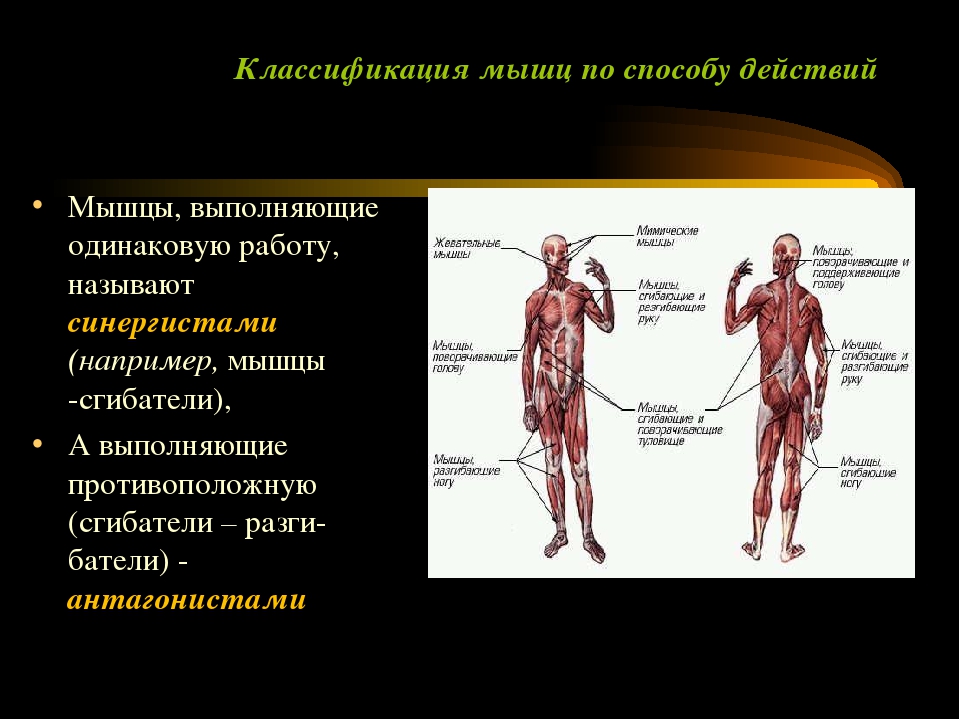 Миология (Макгроу-Хилл). 3-е изд; 2004. с. 489–509.
Миология (Макгроу-Хилл). 3-е изд; 2004. с. 489–509.
Google Scholar
Cobo JL, Abbate F, de Vicente JC, Cobo J, Vega JA. Поиск проприорецепторов в лицевых мышцах человека. Neurosci Lett. 2017; 640: 1–5.
CAS PubMed Статья Google Scholar
Блумер Р., Лукас Дж. Р., Эйгнер М., Биттнер Р., Баумгартнер И., Майр Р. Анализ тонкой структуры веретен экстраокулярных мышц двухлетнего младенца. Инвестируйте Ophthalmol Vis Sci. 1999. 40 (1): 55–64.
CAS PubMed Google Scholar
Баттнер-Энневер Дж.А., Конакчи К.З., Блумер Р. Сенсорный контроль экстраокулярных мышц. Prog Brain Res. 2006; 151: 81–93.
CAS PubMed Статья Google Scholar
Лиенбахер К., Мустари М., Хесс Б., Баттнер-Энневер Дж., Хорн А.К. Есть ли смысл в палисадных окончаниях глазных мышц? Ann N Y Acad Sci. 2011; 1233: 1–7.
2011; 1233: 1–7.
PubMed PubMed Central Статья Google Scholar
Wilkinson KA, Kloefkorn HE, Hochman S. Характеристика афферентов мышечного веретена у взрослых мышей с использованием препарата мышцы-нерв in vitro. ПлоС один. 2012; 7 (6): e39140.
CAS PubMed PubMed Central Статья Google Scholar
Кеннеди WR. Иннервация нормальных мышечных веретен человека. Неврология. 1970. 20 (5): 463–75.
CAS PubMed Статья Google Scholar
Lionikas A, Smith CJ, Smith TL, Bunger L, Banks RW, Bewick GS. Анализ мышечных веретен камбаловидной мышцы шести инбредных линий мышей. J Anat. 2013. 223 (3): 289–96.
PubMed PubMed Central Статья Google Scholar
Macefield VG, Knellwolf TP. Функциональные свойства мышечных веретен человека. J Neurophysiol. 2018; 120 (2): 452–67.
J Neurophysiol. 2018; 120 (2): 452–67.
CAS PubMed Статья Google Scholar
Osterlund C, Liu JX, Thornell LE, Eriksson PO. Состав и распределение мышечного веретена в молодых жевательных мышцах и двуглавой мышце плеча человека обнаруживают ранний рост и созревание. Анат Рек. 2011; 294 (4): 683–93.
Артикул Google Scholar
Kuffler SW, Hunt CC, Quilliam JP. Функция мозговых волокон мелких нервов в вентральных корешках млекопитающих; иннервация эфферентного мышечного веретена. J Neurophysiol. 1951; 14 (1): 29–54.
CAS PubMed Статья Google Scholar
Эдман К.А., Радзюкевич Т., Кронборг Б. Сократительные свойства изолированных мышечных веретен лягушки. Журнал Physiol. 2002. 541 (Pt 3): 905–16.
CAS Статья Google Scholar

Бойд И.А. Структура и иннервация системы мышечных волокон ядерной сумки и системы мышечных волокон ядерной цепи в веретенах мышц млекопитающих. Фил Trans Royal Soc Series B, Biol Sci. 1962. 245: 81–136.
Google Scholar
Банки RW. Иннервация мышечного веретена: личный анамнез. J Anat. 2015; 227 (2): 115–35.
PubMed PubMed Central Статья Google Scholar
Gerwin L, Haupt C, Wilkinson KA, Kröger S. Рецепторы ацетилхолина в экваториальной области интрафузальных мышечных волокон модулируют чувствительность мышечного веретена мыши. J Physiol. 2019; 597 (7): 1993–2006.
CAS PubMed PubMed Central Статья Google Scholar
Оливер К.М., Флорез-Пас Д.М., Бадеа ТЦ, Ментис Г.З., Менон В., де Нойдж Дж. Молекулярное развитие сенсорных афферентов мышечного веретена и органа сухожилия Гольджи, выявленное с помощью анализа транскриптома одного проприорецептора. bioRxiv. 2020. https://doi.org/10.1101/2020.04.03.023986.
bioRxiv. 2020. https://doi.org/10.1101/2020.04.03.023986.
Шредер Дж. М., Бодден Х., Хамахер А., Веррес С. Сканирующая электронная микроскопия раздробленных интрафузальных мышечных волокон из веретен мышц крысы. Мышечный нерв. 1989. 12 (3): 221–32.
PubMed Статья Google Scholar
Sonner MJ, Walters MC, Ковш DR. Анализ проприоцептивной сенсорной иннервации камбаловидной мышцы мыши: комплексный мышечный подход. ПлоС один. 2017; 12 (1): e0170751.
PubMed PubMed Central Статья CAS Google Scholar
Кеннеди В.Р., Вебстер Х.Ф., Юн К.С. Веретена мышц человека: тонкая структура первичного сенсорного окончания. J Neurocytol. 1975. 4 (6): 675–95.
CAS PubMed Статья PubMed Central Google Scholar
Бэнкс Р. У., Халлигер М, Саед Х. Х., Стейси МД. Сравнительный анализ инкапсулированных концевых органов скелетных мышц млекопитающих и их сенсорных нервных окончаний. J Anat. 2009. 214 (6): 859–87.
Сравнительный анализ инкапсулированных концевых органов скелетных мышц млекопитающих и их сенсорных нервных окончаний. J Anat. 2009. 214 (6): 859–87.
CAS PubMed PubMed Central Статья Google Scholar
Ли Дж., Фризе А., Милих М., Сигрист М., Арбер С. Масштабирование транскрипции гена проприорецептора с помощью ретроградной передачи сигналов NT3. ПлоС один. 2012; 7 (9): e45551.
CAS PubMed PubMed Central Статья Google Scholar
Усоскин Д., Фурлан А., Ислам С., Абдо Х., Лоннерберг П., Лу Д. и др. Беспристрастная классификация типов сенсорных нейронов с помощью крупномасштабного секвенирования одноклеточной РНК. Nat Neurosci. 2015; 18 (1): 145–53.
CAS PubMed Статья Google Scholar
Де-Донкер Л., Пике Ф., Пети Дж., Фалемпин М. Характеристика афферентов веретена в камбаловидной мышце крысы с использованием наклонно-удерживаемых и синусоидальных растяжений. J Neurophysiol. 2003. 89 (1): 442–9.
J Neurophysiol. 2003. 89 (1): 442–9.
PubMed Статья Google Scholar
Johnson MI, Ovalle WK. Сравнительное исследование мышечных веретен в медленных и быстрых мышцах новорожденных нормальных и дистрофических мышей. Am J Anat. 1986. 175 (4): 413–27.
CAS PubMed Статья Google Scholar
Bewick GS, Banks RW. Механотрансдукция в мышечном веретене. Pflugers Archiv: Eur J Physiol.2015; 467 (1): 175–90.
CAS Статья Google Scholar
Franco JA, Kloefkorn HE, Hochman S, Wilkinson KA. Мышечно-нервный препарат взрослых мышей in vitro для изучения активирующих свойств мышечных афферентов. J Vis Exp. 2014; 91: 51948.
Google Scholar
Банки РВ. Моторная иннервация мышечных веретен млекопитающих. Prog Neurobiol. 1994. 43 (4-5): 323–62.
1994. 43 (4-5): 323–62.
CAS PubMed Статья Google Scholar
Мануэль М., Зитницки Д. Альфа, бета и гамма мотонейроны: функциональное разнообразие в конечном пути двигательной системы. J Integr Neurosci. 2011; 10 (3): 243–76.
PubMed Статья Google Scholar
Zhang Y, Lin S, Karakatsani A, Rüegg MA, Kröger S. Дифференциальная регуляция кластеризации AChR в полярной и экваториальной области мышечных веретен мышей.Eur J Neurosci. 2015; 41 (1): 69–78.
PubMed Статья Google Scholar
Проске Ю. Мышечное веретено млекопитающих. Новости Physiol Sci. 1997; 12: 37–42.
Google Scholar
Прзедпельска-Обер Э. Развитие мышечных веретен у плодов человека. Анат Анз. 1982; 152 (4): 371–80.
CAS PubMed PubMed Central Google Scholar

Мавринская Л.Ф. Развитие мышечных веретен у человека. Neurosci Translat. 1968; 2: 529–35.
Артикул Google Scholar
Дериу Ф., Тедде Пирас А., Монтелла А. Раннее развитие мышечного веретена у плода человека. Ital J Anat Embryol. 1996. 101 (3): 163–72.
CAS PubMed PubMed Central Google Scholar
Козека К., Онтелл М. Трехмерная цитоархитектура развивающихся мышечных веретен мышей.Deve Biol. 1981. 87 (1): 133–47.
CAS Статья Google Scholar
Zelena J. Нервы и механорецепторы — роль иннервации в развитии и поддержании механорецепторов млекопитающих. Нью-Йорк: Чепмен и Холл; 1994. с. 38–137.
Google Scholar
Кучера Дж., Вальро Дж. М.. Происхождение интрафузальных волокон из подмножества первичных мышечных трубок крысы. Анат Эмбриол. 1995. 192 (2): 149–58.
Анат Эмбриол. 1995. 192 (2): 149–58.
CAS Google Scholar
Barbet JP, Thornell LE, Butler-Browne GS. Иммуноцитохимическая характеристика двух поколений волокон во время развития четырехглавой мышцы человека. Mech Dev. 1991; 35 (1): 3–11.
CAS PubMed Статья PubMed Central Google Scholar
Ландон DN. Тонкая структура экваториальных областей развивающихся мышечных веретен крысы.J Neurocytol. 1972. 1 (2): 189–210.
CAS PubMed Статья PubMed Central Google Scholar
Кучера Дж., Вальро Дж. М.. Последовательности формирования интрафузальных волокон в задних конечностях крысы зависят от мышц. Анат Эмбриол. 1994; 190 (3): 273–86.
CAS Статья Google Scholar
Кучера Дж., Вальро Дж. М., Райхлер Дж. Моторная и сенсорная иннервация мышечных веретен у новорожденных крыс.Анат Эмбриол. 1988. 177: 427–36.
Моторная и сенсорная иннервация мышечных веретен у новорожденных крыс.Анат Эмбриол. 1988. 177: 427–36.
CAS Статья Google Scholar
Кучера Дж., Вальро Дж. М., Райхлер Дж. Роль нервных и мышечных факторов в развитии веретен мышц крыс. Am J Anat. 1989; 186: 144–60.
CAS PubMed Статья PubMed Central Google Scholar
Маеда Н., Осава К., Масуда Т., Хакеда Ю., Кумегава М. Постнатальное развитие анулоспиральных окончаний волокон Ia в мышечных веретенах мышей.Acta Anat. 1985. 124 (1-2): 42–6.
CAS PubMed Статья PubMed Central Google Scholar
Осава К., Маэда Н., Сато М., Кавасаки Т., Масуда Т., Ямамото И. и др. Постнатальное развитие аннулоспиральных окончаний волокон Ia в мышечных веретенах височной мышцы мыши. Анат Анз. 1988. 167 (4): 253–7.
CAS PubMed PubMed Central Google Scholar

Holst-Wolf JM, Yeh IL, Konczak J. Развитие проприоцептивной остроты у типично развивающихся детей: нормативные данные о чувстве положения предплечья. Front Hum Neurosci. 2016; 10: 436.
PubMed PubMed Central Статья Google Scholar
Vejsada R, Hnik P, Payne R, Ujec E, Palecek J. Постнатальное функциональное развитие рецепторов растяжения мышц у крыс. Somatosens Res. 1985. 2 (3): 205–22.
CAS PubMed Статья PubMed Central Google Scholar
Кучера Дж., Вальро Дж. М., Райхлер Дж. Дифференциальные эффекты неонатальной денервации на интрафузальные мышечные волокна у крыс. Анат Эмбриол. 1993. 187 (4): 397–408.
CAS Статья Google Scholar
Кучера Дж., Вальро Дж. М.. Постнатальное созревание веретен в глухих камбаловидных мышцах крыс. Анат Эмбриол. 1987. 176 (4): 449–61.
CAS Статья Google Scholar
Зелена Ю. Роль сенсорной иннервации в развитии механорецепторов. Prog Brain Res. 1976; 43: 59–64.
CAS PubMed Статья PubMed Central Google Scholar
Hippenmeyer S, Shneider NA, Birchmeier C, Burden SJ, Jessell TM, Arber S. Роль передачи сигналов нейрегулина1 в дифференцировке мышечного веретена. Нейрон. 2002. 36 (6): 1035–49.
CAS PubMed Статья PubMed Central Google Scholar
Cheret C, Willem M, Fricker FR, Wende H, Wulf-Goldenberg A, Tahirovic S и др. Bace1 и neuregulin-1 взаимодействуют, чтобы контролировать формирование и поддержание мышечных веретен. EMBO J. 2013; 32 (14): 2015–28.
CAS PubMed PubMed Central Статья Google Scholar
Андречек Э.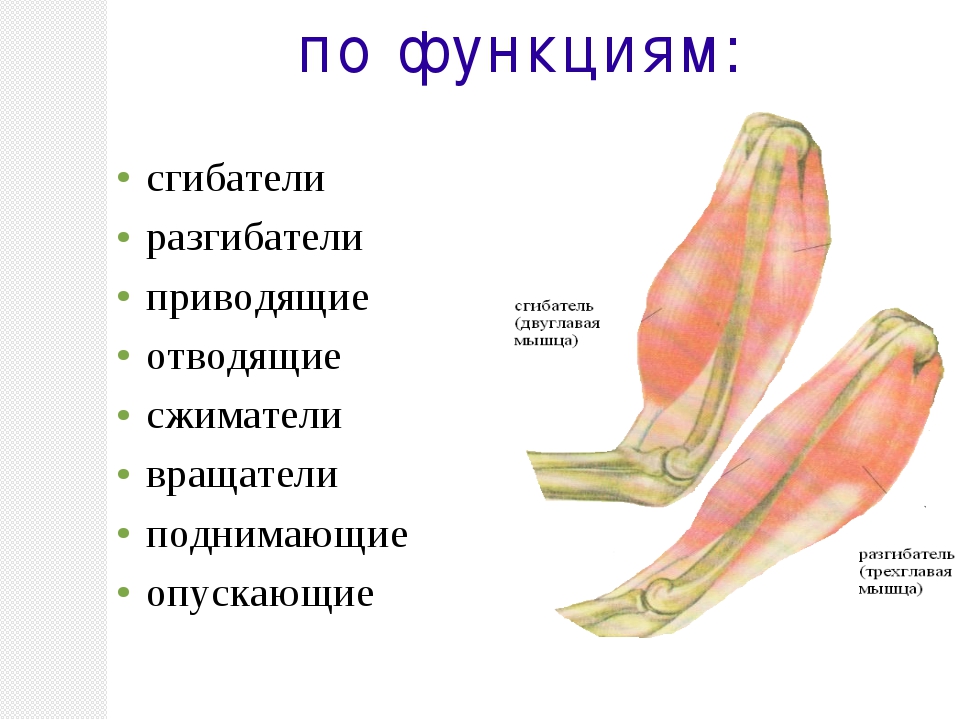 Р., Харди В. Р., Гиргис-Габардо А. А., Перри Р. Л., Батлер Р., Грэм Флорида и др. ErbB2 необходим для выживания мышечного веретена и миобластных клеток.Mol Cell Biol. 2002. 22 (13): 4714–22.
Р., Харди В. Р., Гиргис-Габардо А. А., Перри Р. Л., Батлер Р., Грэм Флорида и др. ErbB2 необходим для выживания мышечного веретена и миобластных клеток.Mol Cell Biol. 2002. 22 (13): 4714–22.
CAS PubMed PubMed Central Статья Google Scholar
Leu M, Bellmunt E, Schwander M, Farinas I, Brenner HR, Muller U. Erbb2 регулирует формирование нервно-мышечных синапсов и важен для развития мышечного веретена. Разработка. 2003. 130 (11): 2291–301.
CAS PubMed Статья PubMed Central Google Scholar
Tourtellotte WG, Keller-Peck C, Milbrandt J, Kucera J. Фактор транскрипции Egr3 модулирует сенсорные взаимодействия аксона и мышечной трубки во время морфогенеза мышечного веретена. Dev Biol. 2001. 232 (2): 388–99.
CAS PubMed Статья PubMed Central Google Scholar

Oliveira Fernandes M, Tourtellotte WG. Egr3-зависимый рецептор растяжения мышечного веретена, интрафузальная дифференцировка мышечных волокон и гомеостаз фузимоторной иннервации.J Neurosci. 2015; 35 (14): 5566–78.
PubMed PubMed Central Статья CAS Google Scholar
Tourtellotte WG, Milbrandt J. Сенсорная атаксия и агенез мышечного веретена у мышей, лишенных фактора транскрипции Egr3. Нат Жене. 1998. 20 (1): 87–91.
CAS PubMed Статья PubMed Central Google Scholar
Arber S, Ladle DR, Lin JH, Frank E, Jessell TM.Ген Er81 ETS контролирует формирование функциональных связей между сенсорными афферентами группы Ia и двигательными нейронами. Клетка. 2000. 101 (5): 485–98.
CAS PubMed Статья PubMed Central Google Scholar
Пак Си, Чан Си, Шин Ю.К., Юн Б.А., Ли Х.Дж., Пак Х.Т. Связующее с Grb2 связующее-1 необходимо для экстрафузального и интрафузального развития мышечных волокон. Нейроотчет. 2017; 28 (10): 604–9.
CAS PubMed Статья PubMed Central Google Scholar
Альберт Ю., Уайтхед Дж., Элдридж Л., Картер Дж., Гао Х, Туртеллотт В.Г. Транскрипционная регуляция спецификации судьбы мышечной трубки и морфогенеза интрафузальных мышечных волокон. J Cell Biol. 2005. 169 (2): 257–68.
CAS PubMed PubMed Central Статья Google Scholar
Смэйн Р.Дж., Кляйн Р., Шнапп А., Лонг Л.К., Брайант С., Левин А. и др. Тяжелые сенсорные и симпатические нейропатии у мышей, несущих нарушенный ген рецептора Trk / NGF.Природа. 1994. 368 (6468): 246–9.
CAS PubMed Статья PubMed Central Google Scholar
Эрнфорс П., Ли К.Ф., Кучера Дж., Яениш Р. Недостаток нейротрофина-3 приводит к недостаточности периферической нервной системы и потере проприоцептивных афферентов конечностей. Клетка. 1994. 77 (4): 503–12.
CAS PubMed Статья Google Scholar
Klein R, Silos-Santiago I, Smeyne RJ, Lira SA, Brambilla R, Bryant S, et al.Нарушение гена рецептора нейротрофина-3 trkC устраняет афференты la мышцы и приводит к аномальным движениям. Природа. 1994. 368 (6468): 249–51.
CAS PubMed Статья Google Scholar
Kucera J, Fan G, Walro J, Copray S, Tessarollo L, Jaenisch R. Нейротрофин-3 и trkC в мышцах не являются необходимыми для развития мышечных веретен мыши. Нейроотчет. 1998. 9 (5): 905–9.
CAS PubMed Статья Google Scholar
Райт Д.Э., Чжоу Л., Кучера Дж., Снайдер В.Д. Введение трансгена нейротрофина-3 в мышцу избирательно спасает проприоцептивные нейроны у мышей, лишенных эндогенного нейротрофина-3. Нейрон. 1997. 19 (3): 503–17.
CAS PubMed Статья Google Scholar
Тейлор, доктор медицины, Ванкура Р., Паттерсон К.Л., Уильямс Дж. М., Рикхоф Дж. Т., Райт Д.Е. Постнатальная регуляция проприоцепции конечностей нейротрофином-3 мышечного происхождения. J Comp Neurol.2001. 432 (2): 244–58.
CAS PubMed Статья Google Scholar
Тейлор, доктор медицины, Холдеман А.С., Велтмер С.Г., Риалс Дж. М., Райт, Делавэр. Модуляция иннервации мышечного веретена нейротрофином-3 после повреждения нерва. Exp Neurol. 2005; 191 (1): 211–22.
CAS PubMed Статья Google Scholar
Пател Т.Д., Крамер И., Кучера Дж., Нидеркофлер В., Джессел Т.М., Арбер С. и др.Периферическая передача сигналов NT3 необходима для экспрессии белка ETS и формирования центрального паттерна проприоцептивных сенсорных афферентов. Нейрон. 2003. 38 (3): 403–16.
CAS PubMed Статья Google Scholar
de Nooij JC, Doobar S, Jessell TM. Инактивация Etv1 выявляет подклассы проприорецепторов, которые отражают уровень экспрессии NT3 в мышечных мишенях. Нейрон. 2013. 77 (6): 1055–68.
PubMed PubMed Central Статья CAS Google Scholar
Аппель Е., Вайсманн С., Зальцберг Ю., Орловский К., Негреану В., Цури М. и др. Ансамбль регуляторных элементов контролирует пространственно-временную экспрессию Runx3 в подмножествах проприоцептивных нейронов ганглиев задних корешков. Genes Dev. 2016; 30 (23): 2607–22.
CAS PubMed PubMed Central Статья Google Scholar
Леванон Д., Беттун Д., Харрис-Черрути С., Вульф Е., Негреану В., Эйлам Р. и др. Фактор транскрипции Runx3 регулирует развитие и выживание нейронов ганглиев дорсальных корешков TrkC.EMBO J. 2002; 21 (13): 3454–63.
CAS PubMed PubMed Central Статья Google Scholar
Verschueren SM, Brumagne S, Swinnen SP, Cordo PJ. Влияние старения на динамическое чувство положения в голеностопном суставе. Behav Brain Res. 2002. 136 (2): 593–603.
CAS PubMed Статья Google Scholar
Landelle C, El Ahmadi A, Kavounoudias A. Возрастное нарушение восприятия движений рук, основанное на проприоцепции мышц и прикосновении.Неврология. 2018; 381: 91–104.
CAS PubMed Статья Google Scholar
Swash M, Fox KP. Влияние возраста на скелетные мышцы человека. Исследования морфологии и иннервации мышечных веретен. J Neurol Sci. 1972. 16 (4): 417–32.
CAS PubMed Статья Google Scholar
Liu JX, Eriksson PO, Thornell LE, Pedrosa-Domellof F.Содержание клетчатки и состав тяжелых цепей миозина в мышечных веретенах двуглавой мышцы плеча человека в возрасте. J Histochem Cytochem. 2005. 53 (4): 445–54.
CAS PubMed Статья Google Scholar
Ким Г. Х., Сузуки С., Канда К. Возрастные физиологические и морфологические изменения мышечных веретен у крыс. J Physiol. 2007. 582 (Pt 2): 525–38.
CAS PubMed PubMed Central Статья Google Scholar
Десаки Дж., Нисида Н. Новые мышечные веретена, содержащие мышечные волокна, лишенные сенсорной иннервации в длинном разгибателе пальцев старых крыс. J Electron Microsc. 2008. 57 (2): 77–82.
Артикул Google Scholar
Воан С.К., Стэнли О.Л., Вальдес Г. Влияние старения на проприоцептивные сенсорные нейроны и интрафузальные мышечные волокна у мышей. Дж. Геронтол А, Биол. Наук. Мед. Наук. 2017; 72 (6): 771–9.
CAS Google Scholar
Conte A, Khan N, Defazio G, Rothwell JC, Berardelli A. Патофизиология соматосенсорных аномалий при болезни Паркинсона. Nat Rev Neurol. 2013; 9 (12): 687–97.
CAS PubMed Статья Google Scholar
Abbruzzese G, Berardelli A. Сенсомоторная интеграция при двигательных расстройствах. Расстройство движения. 2003. 18 (3): 231–40.
PubMed Статья Google Scholar
Кэмерон М. Х., Хорак Ф. Б., Херндон Р. Р., Бурдетт Д. Дисбаланс при рассеянном склерозе: результат замедленной спинальной соматосенсорной проводимости. Somatosens Mot Res. 2008. 25 (2): 113–22.
PubMed PubMed Central Статья Google Scholar
Вильялон Э., Джонс М.Р., Сибигтрот С., Зино С.Дж., Дейл Дж.М., Ландаян Д.С. и др. Изменения мышечного веретена предшествуют развитию сенсомоторной недостаточности при типе 2E Шарко-Мари-Тута. Гены поведения мозга.2017; 16 (2): 260–70.
CAS Статья Google Scholar
Винсент Дж. А., Нарделли П., Габриэль Х. М., Дирдорф А.С., Коуп ТК. Комплексное нарушение проприорецепторов IA-мышц после травматического или нейротоксического повреждения. J Anat. 2015; 227 (2): 221–30.
CAS PubMed PubMed Central Статья Google Scholar
Mentis GZ, Blivis D, Liu W., Drobac E, Crowder ME, Kong L, et al.Раннее функциональное нарушение сенсорно-моторной связи на мышиной модели спинальной мышечной атрофии. Нейрон. 2011; 69 (3): 453–67.
CAS PubMed PubMed Central Статья Google Scholar
van Deursen RW, Sanchez MM, Ulbrecht JS, Cavanagh PR. Роль мышечных веретен в восприятии движений голеностопного сустава у людей с диабетической невропатией. Exp Brain Res. 1998. 120 (1): 1–8.
PubMed Статья Google Scholar
Muller KA, Ryals JM, Feldman EL, Wright DE. Аномальная иннервация мышечного веретена и нейропатия крупных волокон у мышей с диабетом. Сахарный диабет. 2008. 57 (6): 1693–701.
CAS PubMed Статья Google Scholar
Swash M, Fox KP. Патология мышечного веретена при миастении. J Neurol Sci. 1975. 26 (1): 39–47.
CAS PubMed Статья Google Scholar
Cazzato G, Walton JN. Патология мышечного веретена. Исследование биоптата при различных мышечных и нервно-мышечных заболеваниях. J Neurol Sci. 1968; 7 (1): 15–70.
CAS PubMed Статья Google Scholar
Воан С.К., Кемп З., Хаципетрос Т., Виейра Ф., Вальдес Г. Дегенерация окончаний проприоцептивных сенсорных нервов у мышей, несущих мутации, вызывающие амиотрофический боковой склероз. J Comp Neurol. 2015; 523 (17): 2477–94.
CAS PubMed PubMed Central Статья Google Scholar
Сангари С., Иглесиас С., Эль Мендили М.М., Бенали Х., Прадат П.Ф., Маршан-Пауэрт В. Нарушение сенсорно-моторной интеграции на уровне позвоночника при боковом амиотрофическом склерозе. Clin Neurophysiol. 2016; 127 (4): 1968–77.
PubMed Статья Google Scholar
Сабадо Дж., Казановас А., Тарабал О, Хереу М., Пьедрафита Л., Кальдеро Дж. И др.Накопление неправильно свернутой SOD1 в дегенерирующих проприоцептивных сенсорных нейронах ганглия дорсального корешка трансгенных мышей с боковым амиотрофическим склерозом. Biomed Res Int. 2014; 2014: 852163.
PubMed PubMed Central Статья Google Scholar
Секи С., Ямамото Т., Куинн К., Спигельман И., Пантазис А., Олсезе Р. и др. Конструктивно-специфическое раннее нарушение проприоцептивных сенсорных нейронов в мышиной модели SOD1 (G93A) для ALS.J Neurosci. 2019; 39 (44): 8798–815.
PubMed PubMed Central Статья Google Scholar
Powis RA, Gillingwater TH. Избирательная потеря альфа-мотонейронов с сохранением гамма-мотонейронов и холинергических нейронов спинного мозга на мышиной модели спинальной мышечной атрофии. J Anat. 2016; 228 (3): 443–51.
PubMed Статья PubMed Central Google Scholar
Лаланцет-Эбер М, Шарма А, Лященко А.К., Шнейдер Н.А. Гамма-мотонейроны выживают и усугубляют дегенерацию альфа-мотонейронов при БАС. Proc Natl Acad Sci U S. A. 2016; 113 (51): E8316 – E25.
CAS PubMed PubMed Central Статья Google Scholar
Falgairolle M, O’Donovan MJ. Мотонейрональные цепи спинного мозга при дегенеративной болезни мотонейронов. Front Mol Neurosci. 2020; 13:74.
PubMed PubMed Central Статья Google Scholar
Mercuri E, Muntoni F. Мышечные дистрофии. Ланцет. 2013. 381 (9869): 845–60.
CAS PubMed Статья PubMed Central Google Scholar
Кэмпбелл К.П., Каль SD. Ассоциация дистрофина и интегрального мембранного гликопротеина. Природа. 1989. 338 (6212): 259–62.
CAS PubMed Статья PubMed Central Google Scholar
Gao QQ, McNally EM.Дистрофиновый комплекс: структура, функция и значение для терапии. Compr Physiol. 2015; 5 (3): 1223–39.
PubMed PubMed Central Статья Google Scholar
Mah JK, Korngut L, Dykeman J, Day L., Pringsheim T., Jette N. Систематический обзор и метаанализ эпидемиологии мышечной дистрофии Дюшенна и Беккера. Нейромышечный дисорд. 2014; 24 (6): 482–91.
PubMed Статья Google Scholar
Blake DJ, Kröger S. Нейробиология мышечной дистрофии Дюшенна: уроки мышечной массы? Trends Neurosci. 2000. 23 (3): 92–9.
CAS PubMed Статья Google Scholar
Ervasti JM. Дистрофин, его взаимодействия с другими белками и последствия для мышечной дистрофии. Biochimica Biophys Acta. 2007. 1772 (2): 108–17.
CAS Статья Google Scholar
Waite A, Brown SC, Blake DJ. Комплекс дистрофин-гликопротеин в развитии и болезнях мозга. Trends Neurosci. 2012. 35 (8): 487–96.
CAS PubMed Статья Google Scholar
Гольдштейн Дж. А., МакНалли Е. М.. Механизмы мышечной слабости при мышечной дистрофии. J Gen Physiol. 2010. 136 (1): 29–34.
CAS PubMed PubMed Central Статья Google Scholar
Le Rumeur E, Winder SJ, Hubert JF. Дистрофин: больше, чем просто сумма его компонентов. Biochim Biophys Acta. 2010; 1804 (9): 1713–22.
PubMed Статья CAS Google Scholar
Wallace GQ, McNally EM. Механизмы дегенерации, регенерации и восстановления мышц при мышечных дистрофиях. Annu Rev Physiol. 2009. 71: 37–57.
CAS PubMed Статья Google Scholar
Mosqueira M, Zeiger U, Forderer M, Brinkmeier H, Fink RH. Сердечная и респираторная дисфункция при мышечной дистрофии Дюшенна и роль вторичных посредников. Med Res Rev.2013; 33 (5): 1174–213.
CAS PubMed Статья Google Scholar
Hsu JD, Furumasu J. Походка и изменения позы у ребенка с мышечной дистрофией Дюшенна. Clin Orthopaed Rel Res. 1993; 288: 122–5.
Google Scholar
Mahjneh I, Marconi G, Bushby K, Anderson LV, Tolvanen-Mahjneh H, Somer H. Дисферлинопатия (LGMD2B): 23-летнее последующее исследование 10 пациентов, гомозиготных по таким же мутациям дисферлина со сдвигом рамки считывания. Нервно-мышечное расстройство. 2001. 11 (1): 20–6.
CAS PubMed Статья Google Scholar
Прадхан С., Гош Д., Шривастава Н.К., Кумар А., Миттал Б., Пандей С.М. и др. Преднизолон при мышечной дистрофии Дюшенна с неизбежной потерей способности передвигаться.J Neurol. 2006. 253 (10): 1309–16.
CAS PubMed Статья Google Scholar
Troise D, Yoneyama S, Resende MB, Reed U, Xavier GF, Hasue R. Влияние визуального и тактильного восприятия на контроль рук у детей с мышечной дистрофией Дюшенна. Dev Med Child Neurol. 2014. 56 (9): 882–7.
PubMed Статья Google Scholar
Kararizou EG, Manta P, Kalfakis N, Gkiatas KA, Vassilopoulos D.Морфолого-морфометрическое исследование мышечного веретена при мышечной дистрофии. Analyt Quantitat Cytol Hstol. 2007. 29 (3): 148–52.
Google Scholar
Swash M, Fox KP. Патология мышечного веретена при мышечной дистрофии Дюшенна. J Neurol Sci. 1976. 29 (1): 17–32.
CAS PubMed Статья Google Scholar
Skuk D, Goulet M, Tremblay JP.Сохранение мышечных веретен у 27-летнего пациента с мышечной дистрофией Дюшенна: важность для стратегий регенеративной медицины. Мышечный нерв. 2010. 41 (5): 729–30.
PubMed Статья PubMed Central Google Scholar
Ovalle WK, Dow PR. Изменения морфологии мышечного веретена на поздних стадиях мышечной дистрофии мышей. Анат Рек. 1986. 216 (2): 111–26.
CAS PubMed Статья PubMed Central Google Scholar
Bulfield G, Siller WG, Wight PA, Moore KJ. Х-хромосомно-связанная мышечная дистрофия (MDX) у мышей. Proc Natl Acad Sci USA. 1984. 81 (4): 1189–92.
CAS PubMed Статья PubMed Central Google Scholar
Willmann R, Possekel S, Dubach-Powell J, Meier T., Ruegg MA. Модели млекопитающих на животных мышечной дистрофии Дюшенна. Нервно-мышечное расстройство. 2009. 19 (4): 241–9.
PubMed Статья PubMed Central Google Scholar
Yellin H. Индукция и дифференциация веретена при мышиной дистрофии. Experientia. 1974. 30 (3): 286–7.
CAS PubMed Статья PubMed Central Google Scholar
Гервин Л., Россманит С., Хаупт С., Шультейс Дж., Бринкмайер Х., Биттнер Р. Э. и др. Нарушение функции мышечного веретена на мышиных моделях мышечной дистрофии. J Physiol. 2020; 598 (8): 1591–609.
CAS PubMed Статья PubMed Central Google Scholar
Хурана Т.С., Прендергаст Р.А., Аламеддин Х.С., Том FM, Фардо М., Арахата К. и др. Отсутствие патологии экстраокулярных мышц при мышечной дистрофии Дюшенна: роль гомеостаза кальция в сохранении экстраокулярных мышц. J Exp Med. 1995. 182 (2): 467–75.
CAS PubMed Статья PubMed Central Google Scholar
Карпати Г., Карпентер С. Волокна скелетных мышц малого калибра не страдают пагубными последствиями экспрессии дистрофических генов.Am J Med Genet. 1986. 25 (4): 653–8.
CAS PubMed Статья PubMed Central Google Scholar
Nahirney PC, Ovalle WK. Распределение дистрофина и белка нейрофиламентов в мышечных веретенах нормальных и Mdx-дистрофических мышей: иммуноцитохимическое исследование. Анат Рек. 1993. 235 (4): 501–10.
CAS PubMed Статья PubMed Central Google Scholar
Тинсли Дж. М., Блейк Д. Д., Рош А., Фэйрбротер У., Рис Дж., Байт BC и др. Первичная структура белка, родственного дистрофину. Природа. 1992; 360: 591–3.
CAS PubMed Статья PubMed Central Google Scholar
Хурана Т.С., Уоткинс С.К., Чафей П., Челли Дж., Том Ф.М., Фардо М. и др. Иммунолокализация и онтогенетическая экспрессия белка, связанного с дистрофином, в скелетных мышцах. Нервно-мышечное расстройство. 1991; 1 (3): 185–94.
CAS PubMed Статья PubMed Central Google Scholar
Lin S, Gaschen F, Burgunder JM. Утрофин представляет собой связанный с регенерацией белок, временно присутствующий в сарколемме регенерирующих волокон скелетных мышц при гипертрофической мышечной дистрофии кошек с дефицитом дистрофина. J Neuropathol Exp Neurol. 1998. 57: 780–90.
CAS PubMed Статья PubMed Central Google Scholar
Ohlendieck K, Ervasti JM, Matsumura K, Kahl SD, Leveille CJ, Кэмпбелл К.П. Белок, связанный с дистрофином, локализован в нервно-мышечных соединениях скелетных мышц взрослого человека. Нейрон. 1991. 7 (3): 499–508.
CAS PubMed Статья PubMed Central Google Scholar
Шофилд Дж., Хузельштейн Д., Дэвис К., Бэкингем М., Эдвардс Ю. Х. Экспрессия гена белка, связанного с дистрофином (утрофина), во время эмбриогенеза мышей.Dev Dyn. 1993. 198 (4): 254–64.
CAS PubMed Статья PubMed Central Google Scholar
Nguyen TM, Ellis JM, Love DR, Davies KE, Gatter KC, Dickson G, et al. Локализация белка, кодируемого дистрофином, связанного с геном DMDL, с использованием панели из девятнадцати моноклональных антител: присутствие в нервно-мышечных соединениях, в сарколемме дистрофических скелетных мышц, в сосудистых и других гладких мышцах и в линиях пролиферирующих клеток мозга.J Cell Biol. 1991. 115 (6): 1695–700.
CAS PubMed Статья PubMed Central Google Scholar
Бьюик Г.С., Николсон Л.В.Б., Янг К., О’Доннелл Е., Слейтер С.Р. Различное распределение дистрофина и родственных белков в нервно-мышечных соединениях. Нейроотчет. 1992; 3: 857–60.
CAS PubMed Статья PubMed Central Google Scholar
Понс Ф, Николсон Л.В., Роберт А., Войт Т., Леже Дж. Дж. Распределение дистрофина и белка, связанного с дистрофином (утрофин), в нормальных скелетных мышцах и скелетных мышцах с дефицитом дистрофина. Нервно-мышечное расстройство. 1993; 3 (5-6): 507–14.
CAS PubMed Статья PubMed Central Google Scholar
Hirst RC, McCullagh KJ, Davies KE. Активация утрофина при мышечной дистрофии Дюшенна. Acta Myologica. 2005. 24 (3): 209–16.
CAS PubMed PubMed Central Google Scholar
Тинсли Дж., Деконинк Н., Фишер Р., Кан Д., Фелпс С., Гиллис Дж. М. и др. Экспрессия полноразмерного утрофина предотвращает мышечную дистрофию у мышей mdx . Nat Med. 1998; 4: 1441–4.
CAS PubMed Статья PubMed Central Google Scholar
Гилберт Р., Налбантоглу Дж., Петроф Б.Дж., Эбихара С., Гибинга Г.Х., Тинсли Дж. М. и др. Опосредованный аденовирусом перенос гена утрофина смягчает дистрофический фенотип мускулов мышей mdx.Hum Gene Ther. 1999. 10 (8): 1299–310.
CAS PubMed Статья PubMed Central Google Scholar
Тинсли JM, Fairclough RJ, Storer R, Wilkes FJ, Potter AC, Squire SE, et al. Ежедневное лечение SMTC1100, новым активатором низкомолекулярного атрофина, резко снижает симптомы дистрофии у мышей MDX. ПлоС один. 2011; 6 (5): e19189.
CAS PubMed PubMed Central Статья Google Scholar
Гиро С., Эдвардс Б., Бэббс А., Сквайр С.Е., Берг А., Мойр Л. и др. Возможности комбинированной терапии утрофином и дистрофином при мышечной дистрофии Дюшенна. Hum Mol Genet. 2019; 28 (13): 2189–200.
CAS PubMed PubMed Central Статья Google Scholar
Дэвис К.Э., Чемберлен Дж. С.. Суррогатная генная терапия мышечной дистрофии. Nat Med. 2019; 25 (10): 1473–4.
CAS PubMed Статья Google Scholar
Биттнер Р.Э., Андерсон Л.В.Б., Буркхардт Э., Башир Р., Вафиадаки Э., Иванова С. и др. Делеция дисферлина у мышей SJL (SJL-Dysf) определяет естественную модель мышечной дистрофии 2B пояса конечностей. Нат Жене. 1999; 23 (2): 141–2.
CAS PubMed Статья Google Scholar
Хорнси М.А., Лаваль С.Х., Баррези Р., Лохмюллер Х., Бушби К. Мышечная дистрофия в моделях мышей с дефицитом дисферлина. Нервно-мышечное расстройство. 2013. 23 (5): 377–87.
PubMed Статья Google Scholar
Cardenas AM, Gonzalez-Jamett AM, Cea LA, Bevilacqua JA, Caviedes P. Функция дисферлина в скелетных мышцах: возможные патологические механизмы и терапевтические цели при дисферлинопатиях. Exp Neurol. 2016; 283 (Pt A): 246–54.
CAS PubMed Статья Google Scholar
Башир Р., Бриттон С., Страчан Т., Кирс С., Вафиадаки Е., Лако М. и др.Ген, относящийся к Caenorhabditis elegans фактор сперматогенеза fer-1, мутирован при мышечной дистрофии конечностей типа 2B. Нат Жене. 1998. 20 (1): 37–42.
CAS PubMed Статья Google Scholar
Лю Дж., Аоки М., Илла И., Ву С., Фардо М., Анджелини С. и др. Дисферлин, новый ген скелетных мышц, мутирует при миопатии Миёси и мышечной дистрофии пояса конечностей. Нат Жене. 1998. 20 (1): 31–6.
CAS PubMed Статья Google Scholar
Аоки М., Лю Дж., Ричард I, Башир Р., Бриттон С., Кеерс С. М. и др. Геномная организация гена дисферлина и новые мутации при миопатии Миёси. Неврология. 2001. 57 (2): 271–8.
CAS PubMed Статья Google Scholar
Андерсон Л.В., Дэвисон К., Мосс Дж. А., Янг К., Каллен М. Дж., Уолш Дж. И др. Дисферлин является белком плазматической мембраны и экспрессируется на ранних этапах развития человека. Hum Mol Genet. 1999. 8 (5): 855–61.
CAS PubMed Статья Google Scholar
Бартелеми Ф., Дефур А., Леви Н., Кран М., Бартоли М. Мышечные клетки устраняют нарушения, организуя балет по восстановлению мембран. J Neuromuscul Dis. 2018; 5 (1): 21–8.
PubMed PubMed Central Статья Google Scholar
Мацуда С., Киёсуэ К., Нишино И., Гото Ю., Хаяси Ю.К. Дисферлинопатические фибробласты нарушают восстановление плазматической мембраны.PLoS токи. 2015; 7. https://doi.org/10.1371/currents.md.5865add2d766f39a0e0411d38a7ba09c.
Хан Р., Кэмпбелл К.П. Дисферлин и восстановление мышечной мембраны. Curr Opin Cell Biol. 2007. 19 (4): 409–16.
CAS PubMed PubMed Central Статья Google Scholar
Kerr JP, Ziman AP, Mueller AL, Muriel JM, Kleinhans-Welte E, Gumerson JD, et al. Дисферлин стабилизирует индуцированную стрессом передачу сигналов Са2 + в мембране поперечных канальцев.Proc Natl Acad Sci U S. A. 2013; 110 (51): 20831–6.
CAS PubMed PubMed Central Статья Google Scholar
Ribot-Ciscar E, Trefouret S, Aimonetti JM, Attarian S, Pouget J, Roll JP. Сохраняется ли проприоцептивная функция мышечного веретена при мышечных дистрофиях? Исследование вибрации сухожилий мышц. Мышечный нерв. 2004. 29 (6): 861–6.
PubMed Статья PubMed Central Google Scholar
Aimonetti JM, Ribot-Ciscar E, Rossi-Durand C, Attarian S, Pouget J, Roll JP. Функциональное сохранение интрафузионных мышечных волокон при мышечных дистрофиях. Мышечный нерв. 2005. 32 (1): 88–94.
PubMed Статья Google Scholar
Мейнард Дж. А., Купер Р. Р., Ионаеску В. В.. Исследование ультраструктуры интрафузальных мышечных волокон при миотонической дистрофии. Арка Вирхова. 1977; 373 (1): 1–13.
CAS Статья Google Scholar
Swash M, Фокс КП. Аномальные интрафузальные мышечные волокна при миотонической дистрофии: исследование с использованием серийных срезов. J Neurol Neurosurg Psychiatry. 1975. 38 (1): 91–9.
CAS PubMed PubMed Central Статья Google Scholar
Messina C, Tonali P, Scoppetta C. Отсутствие глубоких рефлексов при миотонической дистрофии. J Neurol Sci. 1976; 30 (2-3): 303–11.
CAS PubMed Статья Google Scholar
Pouget J, Gastaut JL, Pellissier JF, Cros D, Serratrice G. Арефлексия сухожилий при врожденных миопатиях, сопровождающихся атрофией волокон I типа. Электрофизиологическое исследование. Rev Electroencephalogr Neurophysiol Clin. 1984. 13 (4): 323–8.
CAS PubMed Статья Google Scholar
Занотели Э., Оливейра А.С., Шмидт Б., Габбай А.А. Центроядерная миопатия: клинические аспекты у десяти бразильских пациентов с дебютом в детстве.J Neurol Sci. 1998. 158 (1): 76–82.
CAS PubMed Статья Google Scholar
Удд Б., Харавуори Х., Калимо Х., Партанен Дж., Пулккинен Л., Паэтау А. и др. Мышечная дистрофия большеберцовой кости — от клинического описания до сцепления на хромосоме 2q31. Нервно-мышечное расстройство. 1998. 8 (5): 327–32.
CAS PubMed Статья Google Scholar
Woo SH, Lukacs V, de Nooij JC, Zaytseva D, Criddle CR, Francisco A, et al.Пьезо2 является основным каналом механотрансдукции проприоцепции. Nat Neurosci. 2015. 18 (12): 1756–62.
CAS PubMed PubMed Central Статья Google Scholar
Сяо Б. Использование механически активированных пьезоканалов для потенциального фармакологического вмешательства. Annu Rev Pharmacol Toxicol. 2020; 60: 195–218.
CAS PubMed Статья Google Scholar
Alcaino C, Knutson K, Gottlieb PA, Farrugia G, Beyder A. Механочувствительный ионный канал Piezo2 ингибируется D-GsMTx4. Каналы. 2017; 11 (3): 245–53.
PubMed PubMed Central Статья Google Scholar
Del Rosario JS, Yudin Y, Su S, Hartle CM, Mirshahi T., Rohacs T. Активация Gi-сопряженного рецептора потенцирует токи Piezo2 через Gbetagamma. EMBO Rep.2020; 21 (5): e49124.
PubMed Google Scholar
Аман Дж. Э., Элангован Н., Йе И. Л., Кончак Дж. Эффективность проприоцептивной тренировки для улучшения двигательной функции: систематический обзор. Front Hum Neurosci. 2014; 8: 1075.
PubMed Google Scholar
Тибоне Дж. Э., Фехтер Дж., Као Дж. Т.. Оценка пути проприоцепции у пациентов со стабильными и нестабильными плечами с соматосенсорными корковыми вызванными потенциалами. J Shoulder Elbow Surg. 1997. 6 (5): 440–3.
CAS PubMed Статья Google Scholar
Юн М.С., Ли Ю.С. Влияние проприоцептивных упражнений на голеностопный сустав на статическое и динамическое равновесие у здоровых взрослых. J Phys Ther Sci. 2017; 29 (2): 242–4.
PubMed PubMed Central Статья Google Scholar
Ribot-Ciscar E, Milhe-De Bovis V, Aimonetti JM, Lapeyssonnie B, Campana-Salort E, Pouget J, et al. Функциональное влияние вибрационной проприоцептивной помощи у пациентов с фациоскапуло-плечевой мышечной дистрофией.Мышечный нерв. 2015; 52 (5): 780–7.
PubMed Статья Google Scholar
Ribot-Ciscar E, Aimonetti JM, Azulay JP. Сенсорная тренировка с использованием кинестетических иллюзий, вызванных вибрацией, улучшает проприоцептивную интеграцию у пациентов с болезнью Паркинсона. J Neurol Sci. 2017; 383: 161–5.
PubMed Статья Google Scholar
Акерли Р., Аймонетти Дж. М., Рибо-Сискар Э.Эмоции изменяют мышечное проприоцептивное кодирование движений у людей. Научный доклад 2017; 7 (1): 8465.
PubMed PubMed Central Статья CAS Google Scholar
Папайоанну С., Димитриу М. Функция мышечного веретена при мышечной дистрофии: потенциальная цель для терапевтического вмешательства. J Physiol. 2020; 598 (8): 1433–4.
CAS PubMed Статья Google Scholar
38.4A: Структура и функции мышечной системы
Мышечная система контролирует множество функций, что возможно при значительной дифференциации морфологии и способности мышечной ткани.
Задачи обучения
- Описать три типа мышечной ткани
Ключевые моменты
- Мышечная система отвечает за такие функции, как поддержание осанки, передвижения и управление различными системами кровообращения.
- Мышечная ткань может быть разделена функционально (произвольно или непроизвольно) и морфологически (поперечно-полосатая или не полосатая).
- Эти классификации описывают три различных типа мышц: скелетные, сердечные и гладкие. Скелетные мышцы являются произвольными и поперечнополосатыми, сердечная мышца — непроизвольными и поперечнополосатыми, а гладкие мышцы — непроизвольными и не имеют поперечных полос.
Ключевые термины
- миофибрилла : волокно, состоящее из нескольких миофиламентов, которое способствует возникновению напряжения в миоците.
- миофиламент : филамент, состоящий из множественных миозиновых или актиновых белков, которые скользят друг по другу для создания напряжения.
- миозин : моторный белок, который образует миофиламенты, которые взаимодействуют с актиновыми филаментами для создания напряжения.
- актин : белок, который образует миофиламенты, которые взаимодействуют с миозиновыми волокнами для создания напряжения.
- полосатый : полосатый вид определенных типов мышц, в которых миофибриллы выровнены для создания постоянного направленного напряжения.
- произвольное : движение мышцы под сознательным контролем (например,г. решив переместить предплечье).
- непроизвольное : движение мышцы, не контролируемое сознанием (например, биение сердца).
- миоцит : мышечная клетка.
Опорно-двигательный аппарат
Мышечная система состоит из мышечной ткани и отвечает за такие функции, как поддержание осанки, передвижения и контроль различных систем кровообращения. Это включает сердцебиение и движение пищи по пищеварительной системе.Мышечная система тесно связана с системой скелета в облегчении движения. Как произвольные, так и непроизвольные функции мышечной системы контролируются нервной системой.
Рисунок \ (\ PageIndex {1} \): Мышечная система : Скелетные мышцы мышечной системы тесно связаны со скелетной системой и действуют для поддержания позы и контроля произвольных движений.Мышца — это узкоспециализированная мягкая ткань, которая создает напряжение, которое приводит к возникновению силы.Мышечные клетки или миоциты содержат миофибриллы, состоящие из миофиламентов актина и миозина, которые скользят мимо друг друга, создавая напряжение, которое изменяет форму миоцита. Многочисленные миоциты составляют мышечную ткань, и контролируемое производство напряжения в этих клетках может генерировать значительную силу.
Мышечную ткань можно функционально классифицировать как произвольную или непроизвольную, а морфологически — как поперечно-полосатую или не поперечнополосатую. Произвольный относится к тому, находится ли мышца под сознательным контролем, в то время как полосатость относится к присутствию видимых полос внутри миоцитов, вызванных организацией миофибрилл для создания постоянного напряжения.
Типы мышц
Приведенная выше классификация описывает три формы мышечной ткани, которые выполняют широкий спектр разнообразных функций.
Скелетная мышца
Скелетные мышцы в основном прикрепляются к скелетной системе через сухожилия, чтобы поддерживать осанку и контролировать движения. Например, сокращение двуглавой мышцы, прикрепленной к лопатке и лучевой кости, поднимет предплечье. Некоторые скелетные мышцы могут прикрепляться непосредственно к другим мышцам или к коже, как показано в
лице, где многочисленные мышцы контролируют выражение лица.
Скелетные мышцы находятся под произвольным контролем, хотя это может быть подсознательным при поддержании позы или равновесия. Морфологически скелетные миоциты имеют удлиненную форму, трубчатую и поперечно-полосатую с множеством периферических ядер.
Ткань сердечной мышцы
Ткань сердечной мышцы находится только в сердце, где сердечные сокращения перекачивают кровь по всему телу и поддерживают кровяное давление.
Как и скелетная мышца, сердечная мышца поперечно-полосатая; однако он не контролируется сознательно и поэтому классифицируется как непроизвольный.Сердечная мышца может быть дополнительно дифференцирована от скелетной мышцы по наличию вставных дисков, которые контролируют синхронизированное сокращение сердечных тканей. Сердечные миоциты короче скелетных эквивалентов и содержат только одно или два ядра, расположенных в центре.
Гладкая мышечная ткань
Гладкая мышечная ткань связана с многочисленными органами и тканевыми системами, такими как пищеварительная система и дыхательная система. Он играет важную роль в регуляции потока в таких системах, например, помогает перемещению пищи через пищеварительную систему через перистальтику.
Гладкая мышца не имеет поперечно-полосатой и непроизвольной формы. Гладкомышечные миоциты имеют веретенообразную форму с одним центрально расположенным ядром.
Рисунок \ (\ PageIndex {1} \): Типы мышц : Тело содержит три типа мышечной ткани: скелетные мышцы, гладкие мышцы и сердечные мышцы, визуализированные здесь с помощью световой микроскопии. Видны полосы в скелетных и сердечных мышцах, что отличает их от более рандомизированного вида гладких мышц.