
(PDF) Стрекательные клетки Polypodium hydriforme (Cnidaria, Polypodiozoa)
ЦИТОЛОГИЯ том 61 № 3 2019
СТРЕКАТЕЛЬНЫЕ КЛЕТКИ POLYPODIUM HYDRIFORME 201
ные двуядерными клетками – инвазивной стадией
развития полиподия, заражающей рыбу (рис. 1г).
Стрекательные клетки служат для прикрепления га-
метофоров к хозяину (Смольянов, Райкова, 1961).
У P. h yd rif or me обнаружены три размерных катего-
рии стрекательных капсул (книдоцист или немато-
цист) с диаметром 12, 7 и 5 мкм (в среднем). Капсулы
размером 7 мкм сосредоточены в области рта и на
сенсорных щупальцах (рис. 1а, 1б), а капсулы разме-
ром 12 и 5 мкм в основном усеивают концы опорных
щупалец (рис. 1а, 1в). Поскольку свободноживущие
особи активно расходуют стрекательные клетки, об-
разование новых капсул (книдогенез) не прекраща-
ется, и развивающиеся книдобласты мигрируют к
местам их использования (Lipin, 1911). Развитие всех
стрекательных капсул сходно (Lipin, 1911; Райкова,
1978) и типично для книдарий. Будущая капсула раз-
Будущая капсула раз-
вивается из последней большой цистерны аппарата
Гольджи. Стрекательная нить закладывается вне-
капсулярно и состоит из чередующихся светлых и
темных фрагментов одинакового размера. Затем
нить втягивается внутрь капсулы, в стенке которой к
этому времени различаются внутренний и внешний
слои, и капсула замыкается крышечкой, к которой
крепится стрекательная нить (Raikova, 1990).
ТИПЫ СТРЕКАТЕЛЬНЫХ
КАПСУЛ P. HY D R I F OR ME
Зрелые стрекательные капсулы P. hy dr if o r m e по
классификации Шульце (Schultze, 1917) относятся к
большим и малым глютинантам, а по классифика-
ции Вейла (Weill,1934) – к типу atrichous и ho-
lotrichous isorhiza.
Atrichous isorhiza (атрихи) – большие капсулы
диаметром около 12 мкм – устилают концы опор-
ных щупалец (рис. 1а, 1в, 2а). Свернутая стрекатель-
ная нить в них занимает все пространство капсулы
(рис. 1в). Нить уложена по принципу двойной
складки и на срезах демонстрирует сложные профили
(Raikova, 1990), но не “профиль пропеллера”, как в
капсулах большинства книдарий, где прослеживается
трехскладчатость (Mariscal, 1974). Все книдоцили та-
Все книдоцили та-
ких капсул прямые и одинаковой длины (рис. 1в, 2а).
Эти капсулы используются при передвижении поли-
подия по субстрату и при заякоривании на нем. При
этом выстреливающая стрекательная нить приклеи-
вается к песчинкам, а капсула может легко отделять-
ся от щупальца полиподия. Выстрелившая стрека-
тельная нить длинная и ровная, на ней часто заметен
обвивающий ее слизистый тяж (рис. 3в, 3г) (Ибраги-
мов, 1999, 2002; Ibragimov, Raikova, 2004).
Holotrichous isorhiza (холотрихи) – средние и мелкие
капсулы, диаметром соответственно 7 и 5 мкм, лока-
лизуются вокруг ротового отверстия и на сенсорных
щупальцах, а также единично встречаются на по-
верхности опорных щупалец. Капсулы диаметром
7 мкм окрашиваются гистологическими красителя-
ми. В капсулах вокруг рта (7 мкм) стрекательная
нить отходит от крышечки капсулы под прямым уг-
лом, делает 2–3 витка, а затем укладывается на дне
капсулы (рис. 1б). На электронограммах нить в кап-
1б). На электронограммах нить в кап-
суле демонcтрирует S-образные профили, т.е. двух-
складчатость (Ibragimov, Raikova, 2004). Капсулы же
размером 5 мкм при гистологической окраске обыч-
но остаются бесцветными, а нить в них, как и у ат-
рих, заполняет все пространство капсулы (рис. 1в).
Выстрелившие нити капсул холотрих отличаются
друг от друга длиной, они покрыты мелкими шипа-
ми, расположенными в два ряда (рис. 3д), что явля-
ется их основной характеристикой, тогда как у дру-
гих книдарий стрекательные нити имеют три ряда
шипов. Некоторые нити суживаются к концу, поэто-
му их предлагается именовать holotrichous anisorhiza.
Но внешне капсулы, ещe не выстрелившие, неразли-
чимы (Ибрагимов, 1999). Главное же отличие холо-
трих от атрих состоит в том, что холотрихи у полипо-
дия обладают уникальными пенетрантными свой-
ствами: их нить пронзает покровы тубифекса и
парамеции (Raikova, 1990; Ibragimov, Raikova, 2004).
Стрекательные клетки у P. hy dr if or me (и атрихи, и
холотрихи) относятся к самому примитивному типу
нематоцист (Werner, 1965; Боженова, 1988; Östman,
1999, 2000; David et al., 2008; Fautin, 2009). Эволюция
стрекательных клеток в настоящее время интенсив-
но разрабатывается, но книдоциты нашего объекта
изучаются очень слабо, хотя благодаря своим не-
обычным признакам интересны как определенная
ступень эволюции стрекательных клеток, как воз-
можное связующее звено между книдоцитами сво-
бодноживущих книдарий и полярными капсулами
миксоспоридий.
ТОНКОЕ СТРОЕНИЕ СТРЕКАТЕЛЬНОЙ
КЛЕТКИ И КНИДОЦИЛЬНОГО АППАРАТА
Каждая зрелая стрекательная клетка P. hy dr if or me
имеет книдоцильный аппарат (рис. 1б, 1в; 2а–2д; 3а),
состоящий из книдоциля – чувствительного механо-
сенсорного неподвижного жгутика, окруженного коль-
цом выростов апикальной поверхности стрекательной
клетки, называемых стереоцилиями (рис. 2д; 3а). Все
2д; 3а). Все
стрекательные капсулы у полиподия имеют округ-
лую форму, что не типично ни для глютинант, опи-
санных в литературе (обычно продолговатых капсул)
(Fautin, 2009), ни для других категорий книдоцитов
большинства книдарий. Вторая их особенность –
положение книдоциля непосредственно над кры-
шечкой капсулы (рис. 1б, 1в; 2а–2д; 3а). Эта особен-
|
 doc), 1 курс
doc), 1 курс
|
Дисциплина «Биология» относится к базовой части Блока 1 специальностей «Педиатрия», «Лечебное дело», «Стоматология» и «Сестринское дело», дисциплина «Биология, экология» – специальности «Медико-профилактическое дело», дисциплина «Биология, эволюционная биология» – по специальности «Медицинская биофизика». Эти дисциплины направлены на формирование у обучающихся естественнонаучного мировоззрения на базе общетеоретических знаний в области биологии, имеющих фундаментальное значение для научной и практической медицины Студенты, обучающиеся по специальностям «Педиатрия», «Лечебное дело», Медико-профилактическое дело»и «Стоматология» изучают биологию на первом курсе в 1 и 2 семестрах и по окончании курса сдают экзамен. Согласно учебному плану специальности «Сестринское дело» студенты изучают биологию в 1 семестре и сдают зачет. Студенты, обучающиеся по специальности «Медицинская биофизика», изучают дисциплину «Биология, эволюционная биология» базовой части Блока 1 на первом и втором курсах (1 – 4 семестры) и сдают экзамен в 4 семестре.
Эти дисциплины направлены на формирование у обучающихся естественнонаучного мировоззрения на базе общетеоретических знаний в области биологии, имеющих фундаментальное значение для научной и практической медицины Студенты, обучающиеся по специальностям «Педиатрия», «Лечебное дело», Медико-профилактическое дело»и «Стоматология» изучают биологию на первом курсе в 1 и 2 семестрах и по окончании курса сдают экзамен. Согласно учебному плану специальности «Сестринское дело» студенты изучают биологию в 1 семестре и сдают зачет. Студенты, обучающиеся по специальности «Медицинская биофизика», изучают дисциплину «Биология, эволюционная биология» базовой части Блока 1 на первом и втором курсах (1 – 4 семестры) и сдают экзамен в 4 семестре.
Дисциплина «Биология клетки» относится к вариативной части дисциплин Блока 1 специальностей «Педиатрия», «Лечебное дело», «Медицинская биофизика», «Медико-профилактическое дело» и «Стоматология». Дисциплина «Биология клетки» предполагает углубленное изучение молекулярно-клеточных процессов для более эффективного усвоения ряда последующих дисциплин. Дисциплина направлена на формирование у обучающихся представления о молекулярных механизмах клеточных процессов в норме и патологии и инновационных биологических методах в медицине. Зачет по дисциплине «Биология клетки» проводится в форме компьютерного тестирования.
Дисциплина направлена на формирование у обучающихся представления о молекулярных механизмах клеточных процессов в норме и патологии и инновационных биологических методах в медицине. Зачет по дисциплине «Биология клетки» проводится в форме компьютерного тестирования.
Дисциплина «Концепции современного естествознания» относится к вариативной части дисциплин Блока 1 специальности «Сестринское дело», изучается в 3 семестре.
Рабочие программы для всех специальностей соответствуют государственным образовательным стандартам. Изучаемые дисциплины способствуют формированию у будущих врачей общекультурных и профессиональных компетенций.
Календарные планы лекций и практических занятий
Календарные планы лекций и практических занятий
Малярия
Малярия (Средние века итал. mala aria — «плохой воздух», ранее известная как «болотная лихорадка» ) — группа трансмиссивных инфекционных заболеваний, передаваемых человеку при укусах комаров рода Anopheles («малярийных комаров») и сопровождающихся лихорадкой, ознобами, спленомегалией (увеличением размеров селезёнки), гепатомегалией (увеличением размеров печени), анемией.
Ежегодно фиксируется 350—500 миллионов случаев заражения людей малярией, из них 1,3—3 миллиона заканчиваются смертью. 85—90 % случаев заражения приходится на районы Африки южнее Сахары, в подавляющем большинстве инфицируются дети в возрасте до 5 лет.
История
Есть предположение, что люди болеют малярией уже в течение 50 000 лет. Считается, что родиной малярии является Западная Африка (P. falciparum) и Центральная Африка (P. vivax). Молекулярно-генетические данные свидетельствуют, что предпаразитический предок плазмодия был свободноживущим простейшим, способным к фотосинтезу, который приспособился жить в кишечнике водных беспозвоночных. Также он мог жить в личинках первых кровососущих насекомых отряда Diptera, которые появились 150—200 миллионов лет назад, быстро приобретя возможность иметь двух хозяев. Древнейшие найденные окаменелости комаров с остатками малярийных паразитов имеют возраст 30 миллионов лет.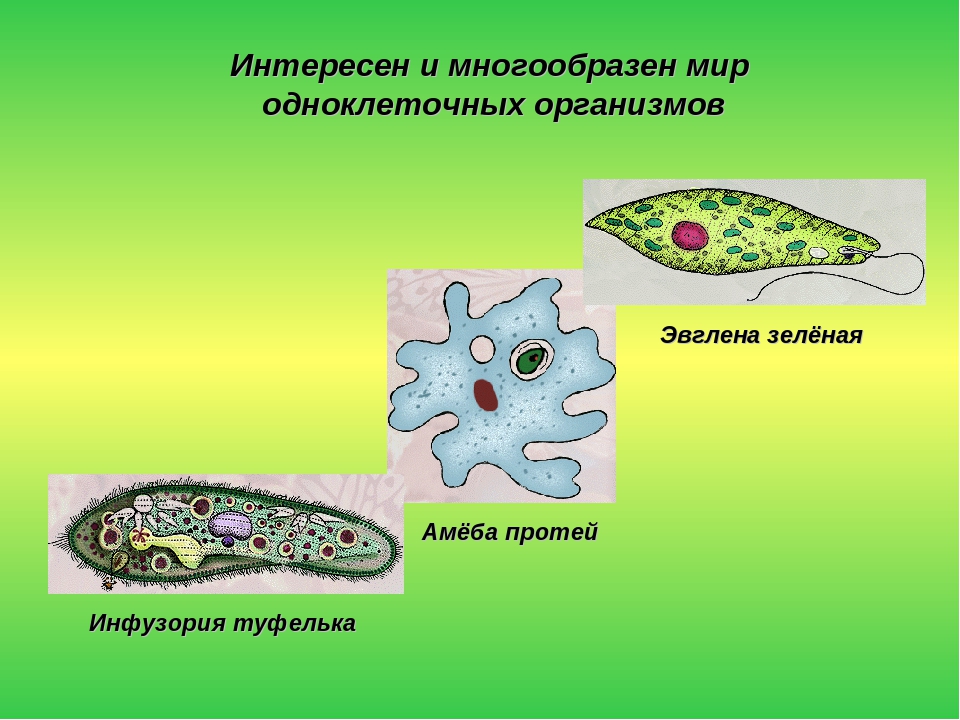
Выяснение причины заболевания
В 1880 году французский военный врач Шарль Луи Альфонс Лаверан, работавший в Алжире, обнаружил в кровяных шариках больного малярией живой одноклеточный организм. Спустя год ученый опубликовал в медицинской печати статью «Паразитарный характер заболевания малярией: описание нового паразита, найденного в крови больных малярией». Это был первый случай, когда простейшие были идентифицированы как причина болезни. За это и другие открытия он был награждён Нобелевской премией по физиологии и медицине 1907 года. Название рода паразита Plasmodium было предложено в 1895 году итальянскими учеными Этторе Маркиафава (на англ.) (итал. Ettore Marchiafava) и Анджело Челли (на англ.) (итал. Angelo Celli)[10]. В 1894 году паразитолог Патрик Мэнсон впервые предположил, что малярия может передаваться человеку комарами. В 1896 г.
 Гордасом для оздоровительных мероприятий, осуществлённых на строительстве Панамского канала.
Гордасом для оздоровительных мероприятий, осуществлённых на строительстве Панамского канала.
В начале XX века, до открытия антибиотиков, практиковалось умышленное заражение малярией пациентов, больных сифилисом. Малярия обеспечивала повышенную температуру тела, при которой сифилис если и не проходил полностью, то во всяком случае снижал свою активность и переходил в латентную стадию. Контролируя течение лихорадки с помощью хинина, врачи таким образом пытались минимизировать негативные эффекты сифилиса. Несмотря на то, что некоторые пациенты умирали, это считалось предпочтительнее неизбежной смерти от сифилитической инфекции.
Открытие покоящейся стадии паразита
Хотя стадии жизненного цикла паразита, проходящие в кровяном русле человека и в теле комара, были описаны ещё в конце XIX — начале XX века, только в 1980-е годы стало известно о существовании покоящейся стадии. Открытие этой формы паразита окончательно объяснило, как люди, вылечившиеся от малярии, могли вновь заболевать спустя годы после исчезновения клеток плазмодия из кровяного русла.
Ареал
Малярийные комары живут почти во всех климатических зонах, за исключением субарктического, арктического поясов и пустынь. В России они обитают на всей европейской территории страны и в Западной Сибири, кроме полярных и приполярных широт. В Восточной Сибири не обитают: зимы там слишком суровые, и комары не выживают.
Однако для того, чтобы существовал риск заражения малярией, требуются, помимо малярийных комаров, условия для быстрого размножения их и переноса малярийного плазмодия. Такие условия достигаются в тех районах, где не бывает низких температур, имеются болота и выпадает много осадков. Поэтому малярия шире всего распространена в экваториальной и субэкваториальной зонах.
Также к естественному ареалу малярии относят влажный субтропический пояс: так, в районе Сочи малярия в начале XX века была большой проблемой до принятия мер по осушению заболоченной местности, нефтеванию водоёмов и проведению других мероприятий, приведших в итоге к уничтожению мест размножения малярийных комаров в курортной зоне.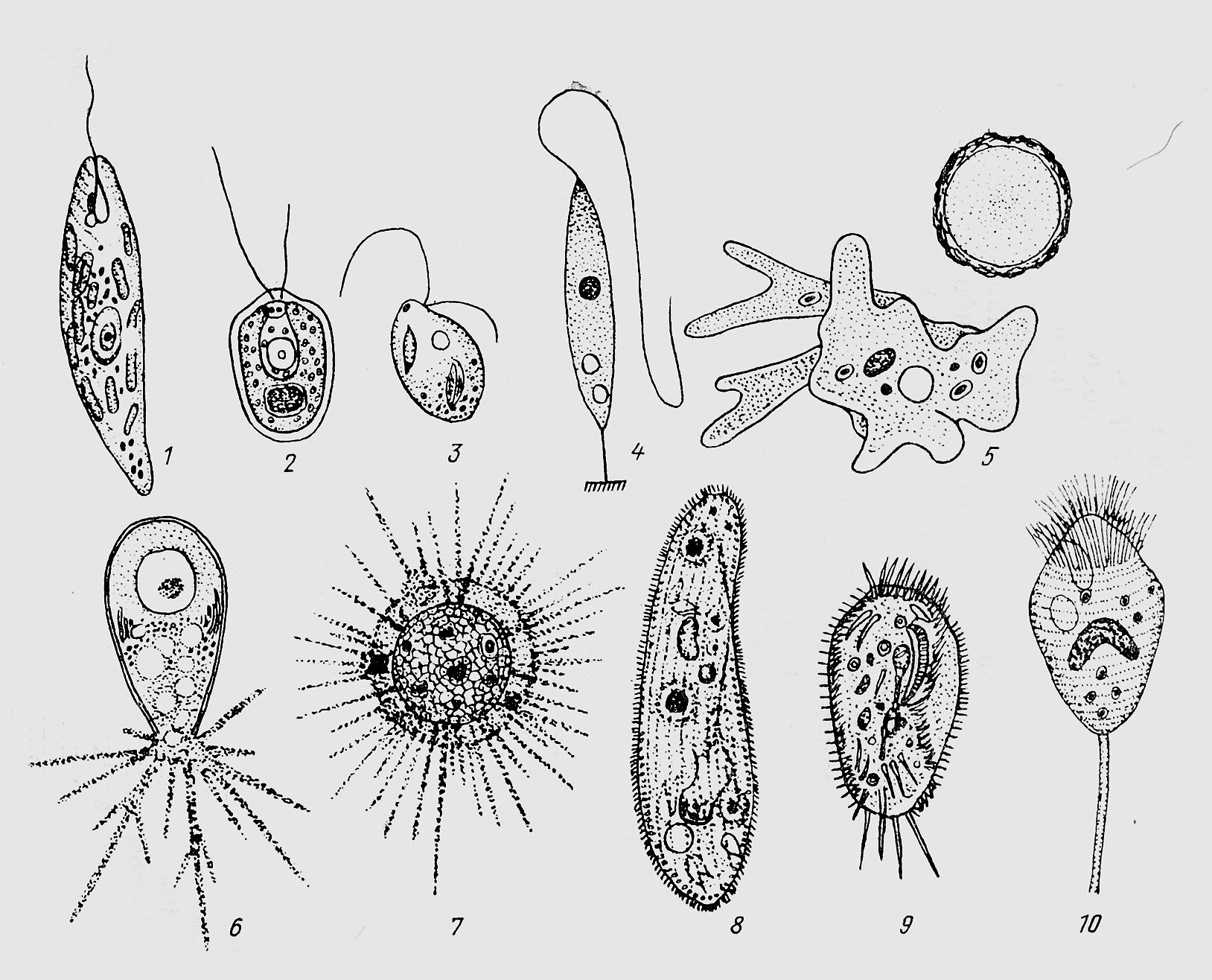
В России и в СССР до начала 1950-х годов заболеваемость малярией была массовой, причем не только на Кавказе, в Закавказье и в Средней Азии, но и средней полосе европейской части (Поволжье и другие районы). Пик абсолютного числа заболевших пришелся на 1934—1935 годы, когда было зарегистрировано более 9 млн случаев малярии.
Этиология
Возбудители малярии — простейшие рода Plasmodium (плазмодии). Для человека патогенны четыре вида этого рода: P.vivax (англ.), P.ovale (англ.), P.malariae (англ.) и P.falciparum. В последние годы установлено, что малярию у человека в Юго-Восточной Азии вызывает также пятый вид — Plasmodium knowlesi. Человек заражается ими в момент инокуляции (впрыскивания) самкой малярийного комара одной из стадий жизненного цикла возбудителя (так называемых спорозоитов) в кровь или лимфатическую систему, которое происходит при кровососании.
Эритроцитарная, или клиническая стадия малярии начинается с прикрепления попавших в кровь мерозоитов к специфическим рецепторам на поверхности мембраны эритроцитов. Эти рецепторы, служащие мишенями для заражения, по-видимому, различны для разных видов малярийных плазмодиев.
Эти рецепторы, служащие мишенями для заражения, по-видимому, различны для разных видов малярийных плазмодиев.
Плазмодии, попадая в жертву, стимулируют выбросы ей веществ, являющихся привлекательными для комаров. К такому выводу пришли исследователи, когда провели серию экспериментов на мышах. Малярийные паразиты изменяли запах тела мышей, и особенно «привлекательным» этот запах становился в период их (паразитов) полного созревания.
Симптомы и диагностика
Симптомы малярии обычно следующие: лихорадка, ознобы, артралгия (боль в суставах), рвота, анемия, вызванная гемолизом, гемоглобинурия (выделение гемоглобина в моче) и конвульсии. Возможно также ощущение покалывания в коже, особенно в случае малярии, вызванной P. falciparum. Также могут наблюдаться спленомегалия (увеличенная селезенка), нестерпимая головная боль, ишемия головного мозга. Малярийная инфекция смертельно опасна. Особенно уязвимы дети и беременные женщины.
Диагноз ставится на основе выявления паразитов в мазках крови.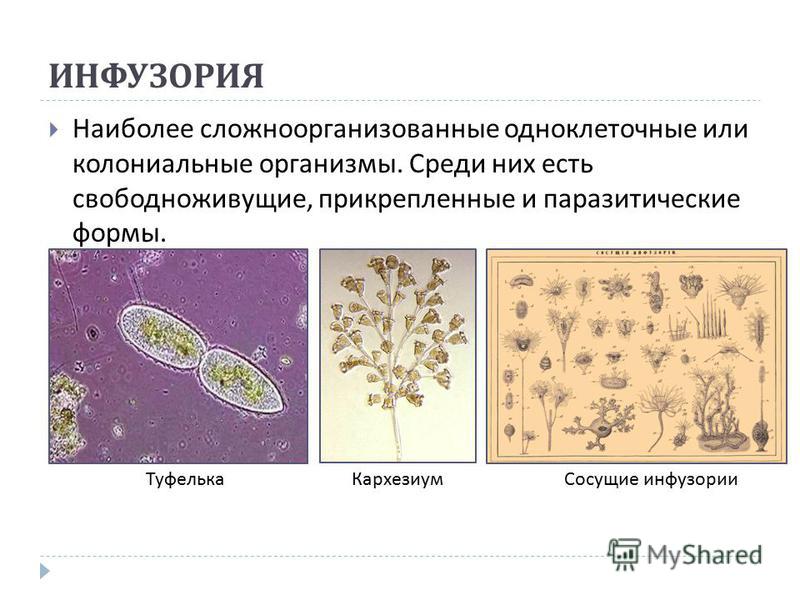 Традиционно используют два типа мазков — тонкий и толстый (или так называемую «толстую каплю»). Тонкий мазок позволяет с большей надёжностью определить разновидность малярийного плазмодия, поскольку внешний вид паразита (форма его клеток) при данном типе исследования лучше сохраняется. Толстый мазок позволяет микроскописту просмотреть больший объём крови, поэтому этот метод чувствительнее, но внешний вид плазмодия при этом изменяется, что не позволяет легко различать разновидности плазмодия. Поставить диагноз на основе микроскопического исследования зачастую бывает затруднительно, так как незрелые трофозоиты разных видов малярийного плазмодия плохо различимы, и обычно необходимо несколько плазмодиев, находящихся на разных стадиях созревания, для надёжной дифференциальной диагностики.
Традиционно используют два типа мазков — тонкий и толстый (или так называемую «толстую каплю»). Тонкий мазок позволяет с большей надёжностью определить разновидность малярийного плазмодия, поскольку внешний вид паразита (форма его клеток) при данном типе исследования лучше сохраняется. Толстый мазок позволяет микроскописту просмотреть больший объём крови, поэтому этот метод чувствительнее, но внешний вид плазмодия при этом изменяется, что не позволяет легко различать разновидности плазмодия. Поставить диагноз на основе микроскопического исследования зачастую бывает затруднительно, так как незрелые трофозоиты разных видов малярийного плазмодия плохо различимы, и обычно необходимо несколько плазмодиев, находящихся на разных стадиях созревания, для надёжной дифференциальной диагностики.
В настоящее время используются также быстрые диагностические тесты (RDT, Rapid Diagnostic Tests) с использованием иммунохимических наборов (более дорогие, но дающие результат через 5—15 минут и не требующие использования микроскопа) и тесты с помощью ПЦР (наиболее дорогие, но наиболее надежные)
Виды (формы) малярии
Симптоматика, течение и прогноз заболевания отчасти зависят от вида плазмодия, который является возбудителем данной формы болезни.
- Возбудитель тропической малярии — P. falciparum. Вызывает наиболее опасную форму, часто протекающую с осложнениями и имеющую высокую смертность. Эта же форма наиболее широко распространена (91 % всех случаев малярии в 2006 г.).
- Возбудитель четырехдневной малярии — Plasmodium malariae. Приступы происходят обычно через 72 часа.
- Возбудители трехдневной малярии и похожей на неё овале-малярии — соответственно, Plasmodium vivax и Plasmodium ovale. Приступы происходят через каждые 40—48 часов.
Эти формы малярии различаются также по длительности инкубационного периода, продолжительности разных стадий жизненного цикла плазмодиев, симптоматике и течению.
Противомалярийный иммунитет
Иммунный ответ против малярийной инфекции развивается медленно. Он характеризуется малой эффективностью и практически не защищает от повторного инфицирования. Приобретённый иммунитет развивается после нескольких заболеваний малярией за несколько лет. Этот иммунитет специфичен к стадии заболевания, к виду и даже к конкретному штамму малярийного плазмодия. Но клинические проявления и симптомы уменьшаются с развитием специфического противомалярийного иммунитета.
Этот иммунитет специфичен к стадии заболевания, к виду и даже к конкретному штамму малярийного плазмодия. Но клинические проявления и симптомы уменьшаются с развитием специфического противомалярийного иммунитета.
Среди возможных объяснений такого слабого иммунного ответа называют нахождение малярийного плазмодия в клетках на протяжении большей части его жизненного цикла, общее угнетение иммунной системы, присутствие антигенов, которые не опознаются T-клетками, подавление пролиферации B-клеток, значительный полиморфизм малярийного плазмодия и быстрая смена потенциальных антигенов на его поверхности.
Лечение
Самым распространенным медикаментом для лечения малярии сегодня, как и раньше, является хинин. На некоторое время он был заменен хлорохином, но ныне снова приобрел популярность. Причиной этому стало появление в Азии и затем распространение по Африке и другим частям света Plasmodium falciparum с мутацией устойчивости к хлорохину.
Также существует несколько других веществ, которые используются для лечения и иногда для профилактики малярии. Многие из них могут использоваться для обеих целей. Их использование зависит преимущественно от устойчивости к ним паразитов в области, где используется тот или другой препарат.
Многие из них могут использоваться для обеих целей. Их использование зависит преимущественно от устойчивости к ним паразитов в области, где используется тот или другой препарат.
В настоящее время наиболее эффективными лекарственными средствами для лечения являются комбинированные с артемезинином лекарственные средства. Резолюция ВОЗ WHA60.18 (май 2007) настаивает на использовании этих препаратов, однако в РФ они до сих пор не зарегистрированы и не применяются.
| Основные антималярийные препараты | ||||
| Препарат | Англ. название | Профилактика | Лечение | Примечания |
| Артеметер-люмефантрин | Artemether-lumefantrine | — | + | коммерческое название Коартем |
| Артезунат-амодиахин | Artesunate-amodiaquine | + | — | |
| Атовакуон-прогуанил | Atovaquone-proguanil | + | + | коммерческое название Маларон |
| Хинин | Quinine | — | + | |
| Хлорохин | Chloroquine | + | + | после появления резистентности использование ограничено коммерческое название Делагил |
| Котрифазид | Cotrifazid | + | + | |
| Доксициклин | Doxycycline | + | + | |
| Мефлохин | Mefloquine | + | + | коммерческое название Лариам |
| Прогуанил | Proguanil | — | + | |
| Примахин | Primaquine | + | — | |
| Сульфадоксин-пириметамин | Sulfadoxine-pyrimethamine | + | + | коммерческое название Фансидар |
Экстракты растения Artemisia annua (Полынь однолетняя), которые содержат вещество артемизинин и его синтетические аналоги, имеют высокую эффективность, но их производство дорого. В настоящее время (2006) изучаются клинические эффекты и возможность производства новых препаратов на основе артемизинина. Другая работа команды французских и южноафриканских исследователей разработала группу новых препаратов, известных как G25 и TE3, успешно испытанных на приматах.
В настоящее время (2006) изучаются клинические эффекты и возможность производства новых препаратов на основе артемизинина. Другая работа команды французских и южноафриканских исследователей разработала группу новых препаратов, известных как G25 и TE3, успешно испытанных на приматах.
Хотя противомалярийные лекарства присутствуют на рынке, болезнь представляет угрозу для людей, которые живут в эндемичных областях, где нет надлежащего доступа к эффективным препаратам. Согласно данным организации «Врачи без границ», средние затраты на лечение человека, зараженного малярией, в некоторых африканских странах составляют всего 0,25—2,40 $.
Профилактика
Методы, которые используются для предотвращения распространения болезни или для защиты в областях, эндемичных для малярии, включают профилактические лекарственные средства, уничтожение комаров и средства для предотвращения укусов комаров. В настоящий момент нет вакцины против малярии, но ведутся активные исследования для её создания.
Разработка вакцины
Ведутся разработки и начаты клинические испытания вакцин от малярии.
В марте 2013 года, после серии неудачных экспериментов, ученые из США успешно испытали на мышах быстродействующее средство против малярии, новый препарат готовят к испытанию на людях.
Малярия всегда была и остается одной из опаснейших болезней человека. К известным личностям, которые умерли от малярии, принадлежат: Александр Македонский, Аларих (король вестготов), Чингисхан, святой Августин, как минимум 5 римских пап, итальянский поэт Данте, император Священной Римской империи Карл V, Христофор Колумб, Оливер Кромвель, Микеланджело Меризи Караваджо, лорд Байрон и многие другие.
Современные данные о малярии:
- В районах распространения малярии проживает 2,4 миллиарда человек, или 34 % населения мира.
- Ежегодно 300—500 миллионов человек заражается малярией, и, согласно ВОЗ, это число ежегодно возрастает на 16 %.
 90 % случаев регистрируется в Африке, из остальных — 70 % случаев приходится на Индию, Бразилию, Шри-Ланку, Вьетнам, Колумбию и Соломоновы Острова.
90 % случаев регистрируется в Африке, из остальных — 70 % случаев приходится на Индию, Бразилию, Шри-Ланку, Вьетнам, Колумбию и Соломоновы Острова. - Ежегодно 1,5—3 миллиона человек умирает от малярии (в 15 раз больше, чем от ВИЧ/СПИД).
- За последнее десятилетие с третьего места по числу смертельных случаев за год (после пневмонии и туберкулёза) малярия вышла на первое среди инфекционных заболеваний.
- Каждый год около 30 000 человек, посещающих опасные районы, заболевают малярией, 1 % из них умирает.
Вторичный метаболизм высших растений и клеток вне организма
78-е Тимирязевское чтение, д.б.н., проф. А.М.Носов, ИФР РАН, Москва
ИФР РАН, Москва
5 июня 2017 г. в 13:00
По словам академика А.Т. Мокроносова, растения кардинально отличают от представителей других царств живых организмов только два процесса – фотосинтез и вторичный метаболизм..
К настоящему времени в высших растениях обнаружено около 100 000 вторичных метаболитов («веществ специализированного обмена растений») – относительно низкомолекулярных соединений, присутствие которых в растительных клетках в принципе не обязательно, и которые, как правило, являются биологически-активными веществами (БАВ). В результате разноплановых исследований установлено, что образование вторичных метаболитов в растении – сложный, четко спланированный в пространстве и во времени процесс, в котором задействовано множество (несколько тысяч) ферментов и регуляторных белков. Расшифровка растительных геномов показала, что в процессах вторичного метаболизма задействовано от 15 до 25% от общего количества генов организма.
В результате разноплановых исследований установлено, что образование вторичных метаболитов в растении – сложный, четко спланированный в пространстве и во времени процесс, в котором задействовано множество (несколько тысяч) ферментов и регуляторных белков. Расшифровка растительных геномов показала, что в процессах вторичного метаболизма задействовано от 15 до 25% от общего количества генов организма.
Наиболее сложным и принципиально важным для физиологии растений является вопрос о роли вторичных метаболитов в жизнедеятельности растений. На протяжении столетней истории изучения веществ специализированного обмена обсуждались различные варианты – от полного отсутствия их функционального значения до главенствующей роли в защите растений от биотических стрессов. К настоящему времени превалирующей является точка зрения, что вторичные метаболиты – это своеобразные «биохимические инструменты» с помощью которых растение, — организм, лишенный возможности активного перемещения в пространстве – решает большинство своих «экологических» проблем – от привлечения опылителей до защиты от разнообразных стрессов. Однако строго доказать функции вторичных метаболитов достаточно сложно. Серьезным инструментом в изучении закономерностей вторичного метаболизма, механизмов его регулирования и функциональной значимости могут быть культуры клеток высших растений.
Однако строго доказать функции вторичных метаболитов достаточно сложно. Серьезным инструментом в изучении закономерностей вторичного метаболизма, механизмов его регулирования и функциональной значимости могут быть культуры клеток высших растений.
Российские ученые – прежде всего, член-корр. АН СССР Р.Г. Бутенко – стояли у истоков изучения биологии клеток высших растений вне организма. За полвека исследований в созданном Р.Г. Бутенко Отделе биологии клетки и биотехнологии ИФР РАН было показано, что культура клеток высших растений представляет собой экспериментальную, не имеющую природных аналогов, биологическую систему– популяцию свободноживущих дедифференцированных растительных клеток . Эта система имеет свои законы развития и многие процессы в ней значительно отличаются от таковых в интактном растении. В частности, вторичный метаболизм в клетках in vitro происходит в неспециализированных пролиферирующих клетках, тогда как в целом растении этот процесс осуществляется в определенных неделящихся клетках и может быть «распределен» по разным тканям и органом растения. Эти различия обуславливают существенные изменения качественного и количественного состава веществ специализированного обмена в клетках вне организма. Целенаправленными воздействиями на клетки in vitro различными факторами (трофическими, гормональными, стрессовыми) можно регулировать их вторичный метаболизм, что является важнейшим преимуществом использования культур клеток для изучения этого процесса.
Эти различия обуславливают существенные изменения качественного и количественного состава веществ специализированного обмена в клетках вне организма. Целенаправленными воздействиями на клетки in vitro различными факторами (трофическими, гормональными, стрессовыми) можно регулировать их вторичный метаболизм, что является важнейшим преимуществом использования культур клеток для изучения этого процесса.
Таким образом, сопоставление закономерностей образования вторичных метаболитов в двух различных биологических системах – в интактном растении и в популяции дедифференцированных клеток — позволяет не только выяснить механизмы регулирования их образования, но и определить функциональное значение для целого организма и клеток in vitro.
Исследования вторичного метаболизма высших растений и клеток in vitro имеют важное значение не только для фундаментальной науки, но и для создания современных инновационных биотехнологий. В настоящее время более половины всех лекарств, нутрицевтиков и косметических средств содержат вещества растительного происхождения, большинство из которых являются вторичными метаболитам. Для производства этих продуктов используется миллионы тонн дикорастущего растительного сырья, что приводит к его дефициту и резкому сокращению ряда видов растений, вплоть до их полного исчезновения. Биомасса культур клеток высших растений, выращенных в биореакторах промышленного объема, является высококачественным возобновляемым сырьем для получения целевых БАВ растительного происхождения. Однако исходя из специфики культур клеток как биологической системы, получение штамма-продуцента и создание подобной Нigh-Тech биотехнологии возможно только на базе детального исследования механизмов регулирования и управления вторичным метаболизмом в клетках in vitro.
Для производства этих продуктов используется миллионы тонн дикорастущего растительного сырья, что приводит к его дефициту и резкому сокращению ряда видов растений, вплоть до их полного исчезновения. Биомасса культур клеток высших растений, выращенных в биореакторах промышленного объема, является высококачественным возобновляемым сырьем для получения целевых БАВ растительного происхождения. Однако исходя из специфики культур клеток как биологической системы, получение штамма-продуцента и создание подобной Нigh-Тech биотехнологии возможно только на базе детального исследования механизмов регулирования и управления вторичным метаболизмом в клетках in vitro.
Изложению и анализу вышеперечисленных фундаментальных и прикладных проблем, иллюстрированных конкретными примерами из работ Отдела биологии клетки и биотехнологии ИФР РАН, будут посвящены 78 Тимирязевские чтения.
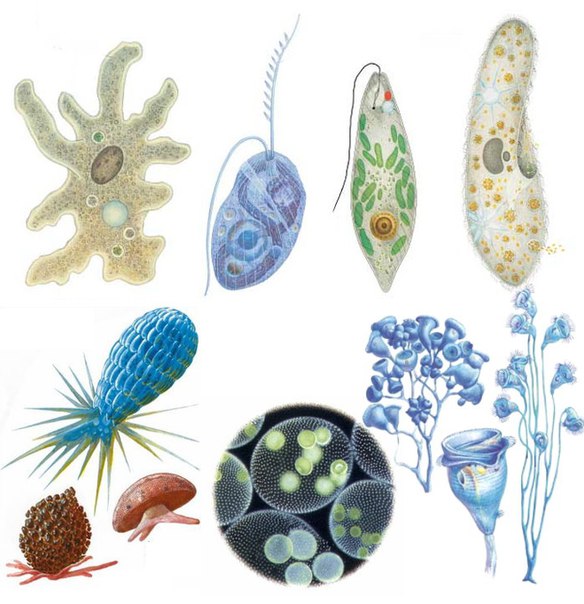
ЦЖ: Кишечнополостные Flashcards | Quizlet
~одиночный полип~В водоемах с чистой водой (
~Это небольшое полупрозрачное животное длиной около 1 см.
~Тело имеет цилиндрическую форму. Нижним концом его (подошвой) она прикрепляется к стеблям и листьям водных растений, корягам и камням. На верхнем конце тела. вокруг рта, имеются 6—12 щупалец. Ее тело и щупальца могут удлиняться и укорачиваться
~Гидра ведет малоподвижный образ жизни.
~В спокойном состоянии
щупальца вытягиваются на несколь-
ко сантиметров. Животное медленно
водит ими из стороны в сторону, подстерегая добычу. При необходимо-
сти гидра может медленно передви-
гаться. При этом она словно шагает
(кувыркается). прикрепляясь к под-
водным предметам попеременно то
верхним, то нижним концами тела.
~Если дотронуться до гидры или уколоть иглой, животное сожмется. Это пример безусловного рефлекса.
~стрекательные клетки В основном расположены на щупальцах гидры.
~Питание. Каждая из пищеварительно—мускульных клеток энтодермы
имеет один — три жгугика. Колеблющиеся жгутики создают ток воды, которым пищевые частички подгоняются к клеткам. Имеющиеся в энтодерме
Имеющиеся в энтодерме
железистые клетки вьщеляют внутрь кишечной полости пищеварительный
сок, который разжижает и частично переваривает пищу.
Пищеварительно-мускульные клетки энтодермы способны образовывать
ложноножки` захватывать и переваривать в пищеварительных вакуолях мелкие пищевые частицы. Таким образом, пищеварение у гидры и всех кишеч-
нополостных внутриклеточное и полостное.
~Питаются гидры мелкими беспозвоночными животными (часто рачками — дафниями и циклопами), которых они ловят щупальцами.
~Органов дыхания у нее нет. и она поглощает кислород всей поверхностью тела
~Размножается гидра половым и бесполым
путем.
Гидры чаще всего размножаются беспо-
лым путем. C приближением холодов, гидры начинают размножаться половым путем
Половые клетки образуются в эктодерме из промежуточных клеток. На теле гидры возникают бугорки двух типов. В одних фоpмируются сперматозоиды, в других — яйцеклетки.
Бывают гидры, у которых сперматозоиды и яйцеклетки образуются на
различных особях.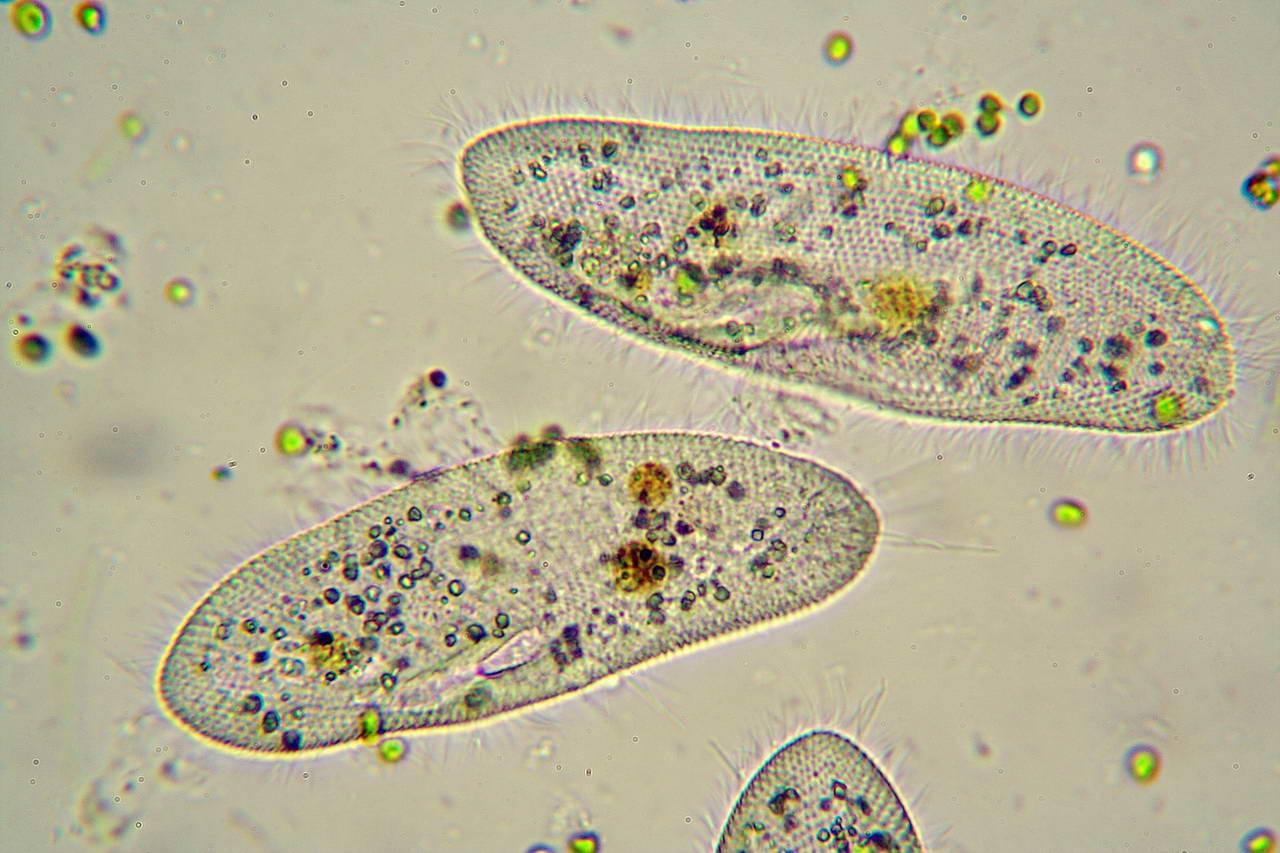 Это раздельнополые животные. У других видов гидр
Это раздельнополые животные. У других видов гидр
и сперматозоиды, и яйцеклетки образуются на теле одного организма. Такие
животные, совмещающие в себе признаки и женского, и мужского пола.
называются гермафродитами.
Попавший в воду сперматозоид плавает с помощью длинного жгутика
и достигает неподвижных яйцеклеток. Оплодотворение (соединение сперма-
тозоида с яйцеклеткой) происходит в теле материнского организма. После
этого вокруг оплодотворенной яйцеклетки формируется плотная оболочка.
Оплодотворенная яйцеклетка многократно делится — образуется зародыш.
Осенью гидра погибает, & покрытые оболочкой зародыши опускаются на дно.
Весной развитие зародыша продолжается. После того как водоем прогре-
ется, оболочки, покрывающие зародыши, разрушаются и маленькие гидры
выходят наружу.
~Регенерация.
Свободноживущие микроорганизмы, фиксирующие молекулярный азот
К настоящему времени установлено, что многие свободноживущие бактерии — представители около 30 видов — могут фиксировать молекулярный азот. Большое значение в фиксации азота имеет семейство Azotobacteriaceae (бактерия Azotobacter chroococcum, открытая М. Бейеринком и др.).
Большое значение в фиксации азота имеет семейство Azotobacteriaceae (бактерия Azotobacter chroococcum, открытая М. Бейеринком и др.).
Молодые клетки азотобактера представляют собой палочки размером 2—3X4—6 мкм. Позже они превращаются в крупные кокки диаметром до 4 мкм. Кокковидные клетки обычно покрываются капсулой и содержат разные включения (жир, крахмал и т. д.).
Иногда у кокковидных клеток появляется толстая оболочка, и они превращаются в цисты. На одних питательных средах палочки быстро приобретают кокковидную форму, на других — по истечении длительного времени. Палочковидные клетки микроба имеют жгутики и обладают подвижностью. При переходе палочек в кокки жгутики обычно теряются (рис. 34).
Из описанных видов азотобактера наиболее изучены Az. chroococcum, Az. vinelandii и Az. agilis. Эти виды различаются между собой размерами и формой клетки, а также некоторыми другими признаками, в частности пигментацией колоний. Так, колонии Az. chroococcum имеют бурый, почти черный цвет. Для Az. agilis характерны бесцветные колонии, Az. vinelandii дает флюоресцирующую желтовато-зеленоватую окраску среды. Наиболее обычен для почвы Аz. chroococcum.
Для Az. agilis характерны бесцветные колонии, Az. vinelandii дает флюоресцирующую желтовато-зеленоватую окраску среды. Наиболее обычен для почвы Аz. chroococcum.
Все виды азотобактера аэробы. В качестве источника азота могут ассимилировать соли аммония, нитриты, нитраты и аминокислоты. При отсутствии связанных форм азота фиксируют молекулярный азот. Небольшие дозы азотсодержащих соединений не депрессируют фиксацию азота, а иногда даже стимулируют ее. Увеличение дозировки связанного азота в среде полностью подавляет усвоение молекулярного азота.
Энергия усвоения азота у отдельных культур азотобактера колеблется в широком диапазоне. Активные культуры связывают 15—20 мг азота на 1 г потребленного органического вещества.
Азотобактер способен использовать огромный набор органических соединений — моно — и дисахариды, некоторые полисахариды (декстрин, крахмал), многие спирты, органические кислоты, в том числе ароматические. Вообще азотобактер проявляет высокую потребность в органических веществах.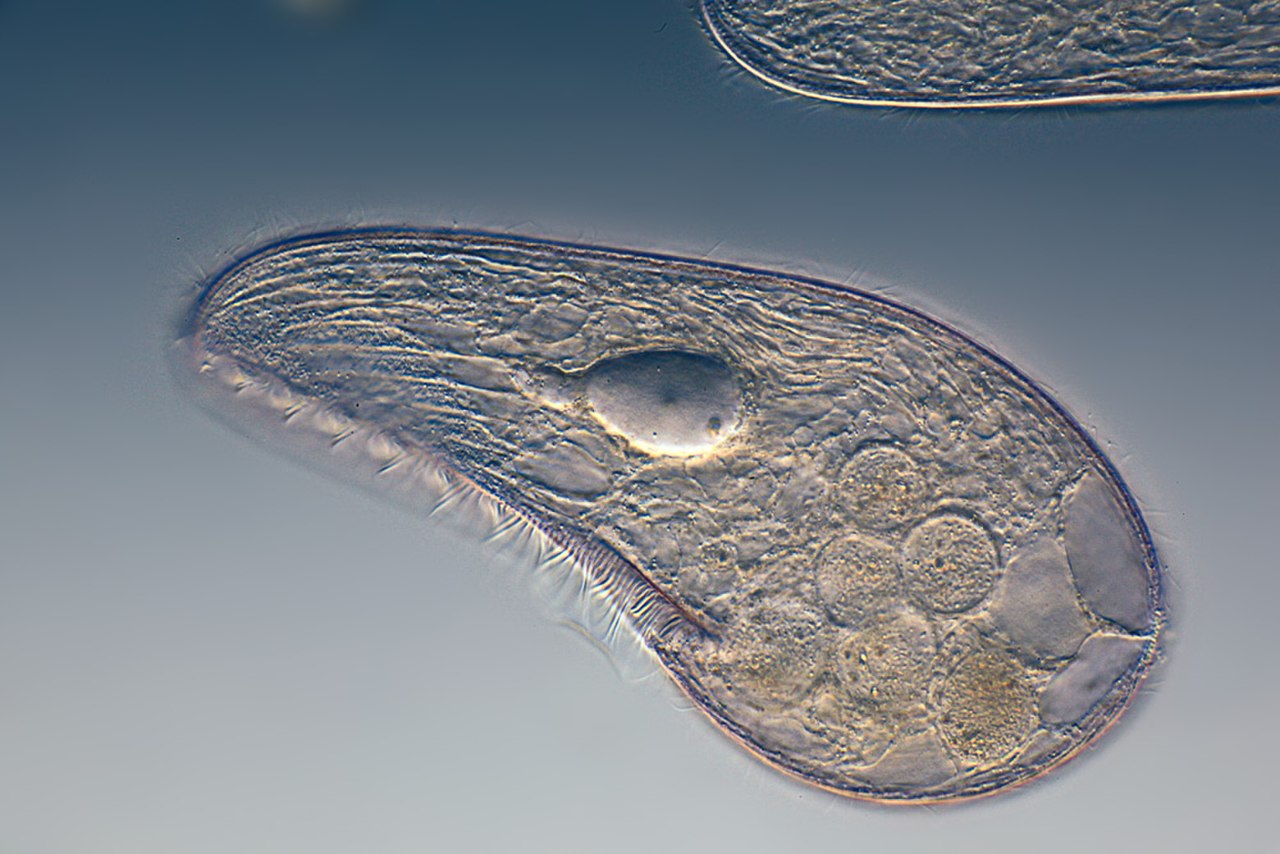 Поэтому он в больших количествах встречается в почвах, хорошо заправленных органическими удобрениями.
Поэтому он в больших количествах встречается в почвах, хорошо заправленных органическими удобрениями.
Для роста азотобактер нуждается в элементах минерального питания, особенно в фосфоре и кальции. Потребность азотобактера в этих элементах столь значительна, что его используют в качестве биологического индикатора на наличие фосфора и кальция в почве.
Для энергичной фиксации молекулярного азота азотобактеру и другим фиксаторам азота нужны микроэлементы. Важное значение имеет молибден, который входит в состав ферментов, катализирующих процесс усвоения азота.
Отмеченные физиологические особенности азотобактера определяют экологию данного организма. Он обитает в высокоплодородных, достаточно влажных почвах с нейтральной или близкой к ней реакцией среды. При дефиците увлажнения большинство клеток этого микроорганизма отмирают. Во многих черноземах, каштановых и сероземных почвах, благоприятных для азотобактера, его обнаруживают в значительных количествах лишь весной. При летнем иссушении почвы в ней остаются немногочисленные его зародыши. В зоне подзолистых и дерново-подзолистых почв азотобактер можно найти в огородных и пойменных почвах, богатых органическими соединениями, с благоприятным значением pH.
При летнем иссушении почвы в ней остаются немногочисленные его зародыши. В зоне подзолистых и дерново-подзолистых почв азотобактер можно найти в огородных и пойменных почвах, богатых органическими соединениями, с благоприятным значением pH.
Отметим, что из ризосферы тропического злака Paspalum notatum был выделен Azotobacter, дающий колонии зеленого тона и имеющий некоторые физиологические отличия от типовой культуры. Этот микроорганизм назван Azotobacter paspali.
К семейству Azotobacteriaceae относятся бактерии рода Beijerinckia, близкие по свойствам к азотобактеру. От азотобактера они отличаются значительной кислотоустойчивостью, кальцифобностью и некоторыми другими свойствами. Они могут расти даже в среде с pH около 3.
Впервые бактерия рода Beijerinckia была получена из кислых почв рисовых полей Индии Р. Старки и П. Де (1939). Ее назвали Azotobacter indicum. Материал, накопленный позднее, показал, что рассматриваемый микроорганизм относится к особому роду, названному в честь известного голландского ученого М.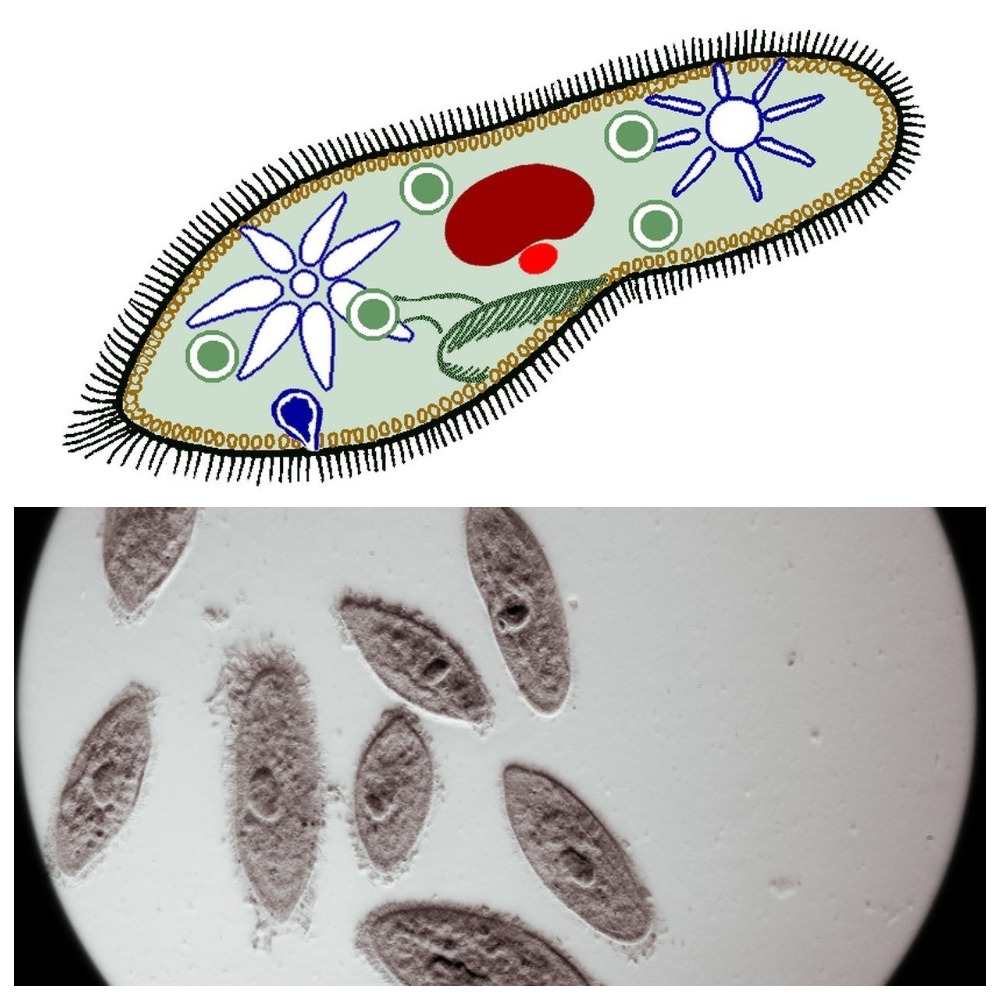 Бейеринка — Beijerinckia. К настоящему времени описан ряд видов бактерий этого рода.
Бейеринка — Beijerinckia. К настоящему времени описан ряд видов бактерий этого рода.
Клетки Beijerinckia имеют различную форму — палочковидную, овальную или круглую. У некоторых видов они подвижны, у других неподвижны. Иногда наблюдается образование капсул. Цисты и эндоспоры отсутствуют.
Большинство культур бактерий рода Beijerinckia дают на безазотной среде с глюкозой выпуклые, блестящие, нередко складчатые слизистые колонии вязкой консистенции. При старении колонии окрашиваются в красноватый или темно-коричневый цвет. В отличие от азотобактера, Beijerinckia не усваивает ароматические соединения и хуже ассимилирует органические кислоты. При развитии на углеводах в среде накапливаются кислые продукты (уксусная и другие органические кислоты). Бактерии рода Beijerinckia менее требовательны по сравнению с азотобактером к концентрации фосфорных соединений в среде. Небольшие дозы соединений кальция тормозят рост Beijerinckia. Значительно менее, чем азотобактер, Beijerinckia чувствительна к повышенной концентрации солей железа и алюминия. Beijerinckia нуждается в молибдене, но довольствуется меньшими его дозами.
Beijerinckia нуждается в молибдене, но довольствуется меньшими его дозами.
Бактерии рода Beijerinckia широко распространены в кислых почвах субтропической и тропической зон. Реже они встречаются в почвах зоны умеренного климата. Бактерии рода Beijerinckia найдены в красноземах Грузии. В окультуренных кислых почвах юга содержится больше клеток Beijerinckia, чем в целинных. Целинные луговые почвы богаче бактериями рода Beijerinckia, чем лесные.
К свободноживущим фиксаторам молекулярного азота семейства Azotobacteriaceae относятся также виды рода Derxia, получившего свое наименование в честь голландского микробиолога Г. Деркса. Это медленно растущие на безазотных средах палочковидные бактерии, обладающие в определенной стадии развития жгутиком. Колонии могут быть пленочными или слизистыми. При старении они становятся желтовато-коричневыми. Derxia обнаружена в почвах тропической зоны, как и бактерии рода Azomonas, также близкие к азотобактеру. Последние имеют округлые клетки и образуют колонии с флюоресцирующим пигментом.
Как показали исследования последнего времени, в почве могут находиться азотфиксирующие бактерии рода Klebsiella (семейство Enterobacteriaceae), представляющие собой грамотрицательные, подвижные с перитрихиальным расположением жгутиков, факультативно-анаэробные палочки. Они выносят довольно низкое значение pH и в большом количестве имеются в лесных подзолистых почвах. В зоне умеренного климата они встречаются и под травянистой растительностью.
К данному семейству относится род Erwinia, представители которого, населяющие надземную часть травянистых растений, могут усваивать атмосферный азот.
К азотфиксаторам относятся и некоторые расы Pseudomonas fluorescens (семейство Pseudomonadaceae). Этот аэробный микроб богаче представлен в почвах северной зоны.
Среди бактерий, фиксирующих молекулярный азот, имеются вибрионы и спириллы. Недавно большой интерес вызвали работы Ж. Доберейнер (Бразилия), показавшей, что в тропических почвах на корнях травянистых растений присутствуют аэробные фиксирующие молекулярный азот спириллы. В то время как большинство сапрофитных азотфиксаторов населяют почву прикорневой зоны (ризосферу), спириллы находятся на самой поверхности корня (ризоплане) и подчас даже проникают в корневую систему. Таким образом, эти микроорганизмы тесно связаны с растениями, и можно ожидать, что они оказывают существенное влияние на их рост.
В то время как большинство сапрофитных азотфиксаторов населяют почву прикорневой зоны (ризосферу), спириллы находятся на самой поверхности корня (ризоплане) и подчас даже проникают в корневую систему. Таким образом, эти микроорганизмы тесно связаны с растениями, и можно ожидать, что они оказывают существенное влияние на их рост.
Изучение физиологических свойств отмеченных спирилл показало, что существует, по крайней мере, две их группы — Azospirillum lipoferum и Azospirillum brasilense, отличающиеся некоторыми физиологическими свойствами. Растения с типом С4 –фотосинтеза (кукуруза, просо, сорго и т. д.) имеют на корневой системе преимущественно A. lipoferum, а растения с типом С3-фотосинтеза (рис, пшеница и др.) —A. brasilense. Исключение составляет сахарный тростник, заражаемый преимущественно A. brasilense.
Достаточно широкие исследования ряда микробов позволяют заключить, что бактерии рода Azospirillum встречаются на корневой системе растений и в более северной зоне, доминируя в зоне южных почв.
Обнаружены и другие аэробные азотфиксирующие бактерии, экология и роль которых в почвенных процессах еще достаточно не изучена. Это представители рода Arthrobacter, относящегося к коринебактериям — микробам, на определенных этапах развития образующим искривленные, неправильной формы клетки и т. д.
Из анаэробных азотфиксаторов довольно хорошо изучены представители рода Clostridium (семейство Bacillaceae).
Как отмечалось выше, первым из свободноживущих фиксаторов азота был открыт Clostridium pasteurianum, Этот микроорганизм имеет палочковидные клетки длиной 1,5—8 мкм и шириной 0,8— 1,3 мкм. Молодые клетки несут перитрихиально расположенные жгутики, старые — образуют споры (см. рис. 29). При спорообразовании клетки утолщаются посередине или на конце. В присутствии кислорода воздуха Cl. pasteurianum может развиваться только при наличии в среде аэробных бактерий, поглощающих кислород, малочувствителен к реакции среды и встречается как в кислых (pH 4,5—5,5), так и в щелочных (pH 8—9) почвах. Споры Cl. pasteurianum довольно устойчивы: они выдерживают нагревание до 96° С в течение нескольких минут.
Споры Cl. pasteurianum довольно устойчивы: они выдерживают нагревание до 96° С в течение нескольких минут.
В качестве источника азотного питания бактерии рода Clostridium могут использовать соли аммония, азотной кислоты и многие содержащие азот органические соединения. При дефиците этого элемента бактерии усваивают молекулярный азот. Источником углерода для Cl. pasteurianum может быть широкий набор углеродсодержащих соединений — моносахариды, дисахариды, некоторые полисахариды (декстрин, крахмал) и органические кислоты. Развиваясь на питательных средах, содержащих углеводы, Cl. pasteurianum разлагает их с образованием масляной и уксусной кислот, углекислого газа и водорода. Освобождающаяся при сбраживании углеводов энергия частично идет на усвоение газообразного азота атмосферы.
Cl. pasteurianum обычно считался слабоактивным фиксатором азота. Пределом его активности было связывание от 1 до 3 мг азота на 1 г сброженного сахара. Однако, используя питательные среды, наиболее отвечающие физиологическим потребностям Cl. pasteurianum, удалось повысить его активность до 10—12 мг азота на 1 г сброженного сахара, а в некоторых случаях и более.
pasteurianum, удалось повысить его активность до 10—12 мг азота на 1 г сброженного сахара, а в некоторых случаях и более.
Способность фиксировать азот атмосферы свойственна и другим видам рода Clostridium (Cl. butyricum, Cl. acetobutylicum, Cl. pectinovorum, Cl. felsineum и т. д.). Эти бактерии близки в систематическом отношении к Cl. pasteurianum, но отличаются от него ферментативными свойствами. Так, Cl. acetobutylicum вызывает ацетонобутиловое брожение, Cl. pectinovorum может разлагать пектин и т. д. Отдельные виды Clostridium приурочены к определенным почвам (см. главу 14).
Из других анаэробных азотфиксирующих бактерий в почвах нередко обнаруживается Bacillus polymyxa, относящаяся к семейству Bacillaceae. Этот микроорганизм в качестве углеродного питания использует простые сахара и некоторые полисахариды, а также пектин.
Небольшие количества молекулярного азота могут фиксировать некоторые олиготрофные бактерии.
Весьма своеобразная обстановка для микроорганизмов вообще и азотфиксаторов в частности создается на затопленных полях под посевами риса. При разложении массы растительных остатков в почве под слоем воды наряду с другими веществами образуются газообразные соединения — водород, метан, СО2. Установлено, что Н2 и СН4 могут быть источниками энергии и питания для некоторых азотфиксирующих бактерий. Имеются корине — и микобактерии, способные жить автотрофно, окисляя водород и ассимилируя углекислоту. Одновременно они фиксируют молекулярный азот.
При разложении массы растительных остатков в почве под слоем воды наряду с другими веществами образуются газообразные соединения — водород, метан, СО2. Установлено, что Н2 и СН4 могут быть источниками энергии и питания для некоторых азотфиксирующих бактерий. Имеются корине — и микобактерии, способные жить автотрофно, окисляя водород и ассимилируя углекислоту. Одновременно они фиксируют молекулярный азот.
Метан используется специфическими азотфиксирующими бактериями из рода Methylomonas (семейство Methylomonadaceae), которые в аэробных условиях могут жить, окисляя только метан или метиловый спирт. Это небольшие монотрихи, не образующие спор.
В воде рисовых полей, а также в Других водоемах можно обнаружить и других азотфиксаторов, представляющих собой анаэробные бактерии. К ним относятся фототрофные пурпурные серобактерии (Thiocapsa, Chromatium, Thiocystis и др.), пурпурные несерные бактерии (Rhodospirillum, Rhodopseudomonas и др.) и зеленые серобактерии (Chlorobium, Pelodiction).
В почвах, загрязненных нефтью, развивается специфический ценоз микроорганизмов, усваивающих молекулярный азот. В основном здесь размножаются ассимилирующие углеводороды коринебактерии (Arthrobacter).
К активным азотфиксаторам относятся аэробные цианобактерии (сине-зеленые водоросли). Можно считать, что все микроорганизмы этой группы, обладающие гетероцистами (клетки с толстой оболочкой), способны фиксировать N2, Они входят в порядки Mastigocladales, Stigonematales и Nostocales класса Hormogoniophyсеае. В почвах СНГ обнаружено около 130 видов и разновидностей гетероцистных форм цианобактерий. Из них к азотусвоителям принадлежат роды Anabaena, Nostoc, Cylindrospermum, Calothris, Tolypothrix и Scytonema, причем наиболее распространены в почвах представители рода Nostoc.
Усвоение молекулярного азота происходит в гетероцистах, то есть в клетках, куда ограничен доступ кислорода. Тем не менее, ферментный аппарат, связывающий N2, обнаружен и в вегетативных клетках гетероцистных форм цианобактерий. Это послужило основанием для поиска негетероцистных азотфиксирующих форм. В последнее время они найдены — это представители родов Plectonema и Phormidium, которые, не обладая гетероцистами, могут связывать N2. В СНГ подобные формы найдены Е. М. Панкратовой.
Это послужило основанием для поиска негетероцистных азотфиксирующих форм. В последнее время они найдены — это представители родов Plectonema и Phormidium, которые, не обладая гетероцистами, могут связывать N2. В СНГ подобные формы найдены Е. М. Панкратовой.
Цианобактерии распространены во всех почвенно-климатических зонах. Однако они предпочитают нейтральную среду, и. поэтому их численность и видовой состав существенно возрастают в нейтральных почвах южной зоны. Отдельные их виды приурочены к определенным местам обитания. Многие цианобактерии живут в оимбиозе с другими растительными организмами, например,— с грибами, образуя при этом лишайники. В результате адаптации к местным условиям они приобрели способность фиксировать азот при температуре, близкой к 0°С, иногда азотфиксация происходит даже при —5°С, оптимальная температура для этого процесса 15—20°С. В тропиках цианобактерии находятся в симбиозе с саговниками и печеночниками, а также с водным папоротником Azolla и т. д.
В природной обстановке цианобактерии всегда сожительствуют с другими микроорганизмами — бактериями и грибами. В местах разрастания водорослей особенно много олиготрофных бактерий. Массовое развитие цианобактерий отмечается в сильно увлажненных почвах, где они нередко образуют «цветение» почв. Аналогичное явление имеет место в водоемах при обильном размножении водорослей. В неорошаемых окультуренных почвах наиболее благоприятные условия для роста цианобактерий бывают весной и осенью, то есть в периоды увлажнения почвы, а в поливных, кроме того, и после орошения пашни.
В местах разрастания водорослей особенно много олиготрофных бактерий. Массовое развитие цианобактерий отмечается в сильно увлажненных почвах, где они нередко образуют «цветение» почв. Аналогичное явление имеет место в водоемах при обильном размножении водорослей. В неорошаемых окультуренных почвах наиболее благоприятные условия для роста цианобактерий бывают весной и осенью, то есть в периоды увлажнения почвы, а в поливных, кроме того, и после орошения пашни.
Вклад свободноживущих азотфиксаторов в азотный фонд почвы весьма существен. Вопрос о роли свободноживущих и симбиотических азотфиксаторов в азотном балансе почв рассмотрен в главе 16.
Свободноживущие клетки человека реконфигурируют свои хромосомы в процессе эволюции обратно к одноклеточности
Существенные изменения:
1) То, что вы называете линиями нераковых клеток, на самом деле увековечено либо теломеразой, либо EBV; Таким образом, они являются плохими заменителями «нормальных клеток», но ближе к нормальным, чем раковые клетки, которые уже претерпели кариотипические изменения.
Требуется таблица, в которой перечислены все линии клеток, включая обсуждение связанных предостережений.
Мы рассмотрим проблему далее в начале Обсуждения. Действительно, «нормальные клеточные линии» означают «клеточные линии, полученные из незлокачественных клеток». Хотя они не являются действительно нормальными (клеточные линии, по определению, не являются нормальными), они могут служить ориентиром, по которому может быть измерена эволюция линий раковых клеток.
Мы добавили две дополнительные таблицы, которые включают запрашиваемую информацию. Дополнительный файл 4 содержит информацию о линиях раковых клеток (подраздел «Сбор данных», второй абзац).Дополнительный файл 5 содержит информацию о линиях нераковых клеток (см. Последний абзац вышеупомянутого подраздела).
2) Каково обоснование ожидания того, что Х-хромосомы должны быть потеряны только на 15% — если это основано на размере, это можно лучше объяснить с помощью рисунка 1A.

Да, ожидаемая потеря хромосомы Y / X рассчитывается из регрессии процента LOH на размер хромосомы.
Редакция: «Учитывая его положение как 7-ю по величине хромосому, не ожидается, что X будет потеряна более чем в 15% клеточных линий на основе регрессионного анализа на Рисунке 1A»
3) Модели, которые вызывают или не вызывают выбор, могут быть более полезными там, где есть средства для получения селективных коэффициентов.Похоже, это то, к чему вы направились с моделью перехода, но не оправдывает ожиданий и вместо этого просто приходит к выводу, что выбор должен играть роль для достижения соотношения X: A = 2: 3. (см. также пункт 10 ниже).
Этот момент заслуживает более подробного рассмотрения. Мы добавили новый раздел в Материалы и методы.
Редакция: Следующее описание воспроизводится из новых материалов и методов.
«В этом исследовании мы используем модель для сравнения наблюдений с нейтральным ожиданием. Хотя мы могли отклонить нейтральную модель и сделать вывод о направлении отбора, мы воздерживаемся от оценки силы отбора по двум причинам — как биологическим, так и техническим. […] Учитывая различную историю этих клеточных линий, мы подозреваем, что значение может находиться в диапазоне от 0,1 до 10. Различия качественно согласуются, но фактические значения потребуют знания точной истории культивирования каждой клеточной линии ».
Хотя мы могли отклонить нейтральную модель и сделать вывод о направлении отбора, мы воздерживаемся от оценки силы отбора по двум причинам — как биологическим, так и техническим. […] Учитывая различную историю этих клеточных линий, мы подозреваем, что значение может находиться в диапазоне от 0,1 до 10. Различия качественно согласуются, но фактические значения потребуют знания точной истории культивирования каждой клеточной линии ».
4) Как были культивированы все клетки? Если в условиях отсутствия сыворотки или богатой сывороткой, то объясняет ли этот фактор некоторое расхождение (разброс) наблюдаемых соотношений X: A.Таблица 1, в которой перечислены клеточные линии, может прояснить это, и это также позволит опровергнуть гипотезу о том, что на самом деле клетки конвергентно эволюционируют к условиям среды с высоким содержанием сыворотки, а не к « одноклеточности » (в общем). Скудное описание сред и условий выращивания — одно из вопиющих упущений в настоящей рукописи.

Вопрос очень резонный. Поскольку в целом верно, что для культивирования клеток используется ограниченное количество рецептов, как мы пришли к выводу, что эволюция клеточной линии определяется скорее «одноклеточностью», чем «условиями культивирования».Этот вопрос заслуживает полного ответа (см. Ниже).
Версия: в разделе «Материалы и методы» мы даем более полный ответ, который воспроизводится ниже.
«Когда мы приписываем наблюдаемые генетические изменения, связанные с одноклеточным существованием клеток, мы включаем все факторы окружающей среды, которые делают возможным одноклеточное существование. […] Также важно отметить, что эти факторы окружающей среды часто антагонистичны многоклеточной жизни ».
5) Недопустимо, чтобы критическая часть данных из ячеек HeLa была ссылкой на другую неопубликованную рукопись.По крайней мере, эта рукопись должна быть предоставлена рецензентам или соответствующие данные должны быть включены в эту рукопись (см.
Также пункт 8 ниже).
В настоящее время исследование находится на рассмотрении, но было опубликовано на BioRxiv, поэтому оно доступно для широкой читательской аудитории (https://www.biorxiv.org/content/early/2017/10/05/193482).
6) Предположение о том, что аутосомы приобретаются индивидуально, кажется необоснованным (даже если это только для аргументации) — более вероятно, что неспособность пройти надлежащую хромосомную сегрегацию привела к изменениям плоидности после отдельных аутосом, которые могли быть потеряны.
Комментарий 6 позже будет расширен и будет включен в комментарий 9 и комментарий 10. Поэтому мы ответим на 3 вопроса вместе, отвечая на комментарии 9–10.
В то время как обоснование сосредоточения внимания только на крупных аутосомах кажется разумным, данные о более мелких аутосомах и их дисперсии должны быть доступны читателю, даже если только в виде приложений.
Число копий хромосом меньше chr13 добавлено в дополнительный файл 2.
7) Описание женских клеточных линий немного сбивает с толку. Общее утверждение или вывод состоит в том, что для женских линий с дополнительными Х-хромосомами получается активная (Ха) хромосома. Это важно, потому что усиление неактивной Х-хромосомы (замалчиваемое) не должно приводить к большому отбору. Однако на рисунке 1F экспрессия XIST (связанная с инактивацией Х-хромосомы) наблюдается в большинстве (N = 103) клеточных линий. Кроме того, на рис. 1E проиллюстрировано только 20% женских клеточных линий — каково состояние остальных 80% клеточных линий? Для оценки требуется больше ясности.
Мы согласны с тем, что описания и рисунок 1E-1F сбивают с толку. Новое описание, воспроизводимое ниже, должно прояснить проблему.
Редакция: Рисунки 1E и 1F были изменены следующим образом:
«Мы используем только женские линии, которые показывают LOH всей Х-хромосомы (~ 37% женских линий) при подсчете Ха по следующей причине. […] Подобно мужским линиям на Рисунке 1B, Рисунок 1F также показывает, что примерно половина женских линий получила дополнительный Ха ».
И условные обозначения фигур также были переработаны.
8) Исследования линии клеток HeLa немного сбивают с толку. Рисунок 2D используется, чтобы заявить, что «внутриклеточная неоднородность, по-видимому, не опровергает наших выводов». Затем были выделены 6 отдельных клеток HeLa и выращены до миллиона клеток (подраздел «Согласованная эволюция аутосом как набора», второй абзац). WGS был использован для оценки плоидности с выводом, что более крупные (chr 1-13) развиваются «сплоченно» как множество (см. Вышеупомянутый подраздел, последний абзац). Это очень интересное наблюдение и вывод, но цифры (N = 6) небольшие.
Рисунок 2D и рисунок 4B-C на самом деле представляют два связанных, но несколько разных аспекта эволюции. Оба предполагают, что внутристрочные вариации демонстрируют существенную сплоченность, которая больше, чем ожидалось. Разница в том, что на рисунке 2D исследуются вариации внутри линий (с помощью FISH с фокусом на X), а на рисунке 4 показаны средние изменения в эволюции (с помощью секвенирования ДНК, в основном на аутосомах). Рисунок 2D похож на исследование различий от человека к человеку, а рисунок 4 рассматривает средние различия между людьми и шимпанзе.
Рисунок 2D похож на исследование различий от человека к человеку, а рисунок 4 рассматривает средние различия между людьми и шимпанзе.
Согласны, что N = 6 немного мало, но оказалось достаточно. Поскольку лежащее в основе распределение Пуассона является широким, каждая линия фактически обеспечивает более чем скромную статистическую поддержку. В совокупности есть достаточная мощность, чтобы отклонить нулевую модель.
Однако, если это правда, должна быть возможность кариотипировать отдельные клетки, чтобы увидеть, соблюдаются ли ожидаемые соотношения 1: 2, 2: 3 X: A, как и прогнозировалось. Эти данные уже доступны (рис. 2B), и их можно просто систематизировать и представить.
Мы используем FISH в сочетании с X-специфическим окрашиванием для подсчета X каждой отдельной клетки. С другой стороны, подсчет аутосом одной клетки является сложной задачей, потому что аутосомы не имеют индивидуальной маркировки. Некоторые аутосомы могут находиться вне поля или происходить из соседних клеток.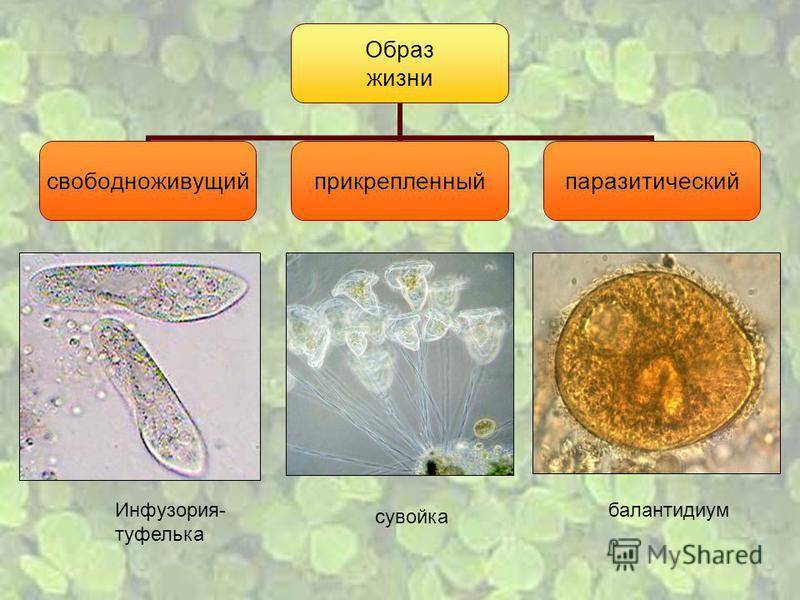 При анализе нам все еще необходимо усреднить клетки одной и той же клеточной линии. В этом случае секвенирование ДНК или устаревшие микроматрицы более точны в обеспечении среднего отношения X: A в пределах линии.(Мы также знаем, что разброс внутри строки невелик.)
При анализе нам все еще необходимо усреднить клетки одной и той же клеточной линии. В этом случае секвенирование ДНК или устаревшие микроматрицы более точны в обеспечении среднего отношения X: A в пределах линии.(Мы также знаем, что разброс внутри строки невелик.)
Эти данные могут быть уже представлены на Рисунке 4 — дополнения к рисунку 1 и 2, но на нем есть мужские и женские пики. Рисунок 4 — дополнение к рисунку 2, но Рисунок 4 — дополнение к рисунку 1 касается линии клеток HeLa (женской), поэтому источники мужских пиков неясны. Требуются разъяснения.
На рис. 4 — дополнительный рисунок 2 представлены подсчеты для нескольких клеточных линий (не HeLa). Вот почему есть мужские и женские линии.
Следующие три вопроса образуют набор:
[(6) здесь повторяется: предположение о том, что аутосомы приобретаются индивидуально, кажется необоснованным (даже если это только ради аргумента) — более вероятно, что неспособность пройти надлежащую сегрегацию хромосом привела к изменениям плоидности после отдельных аутосом, которые могли быть потеряны.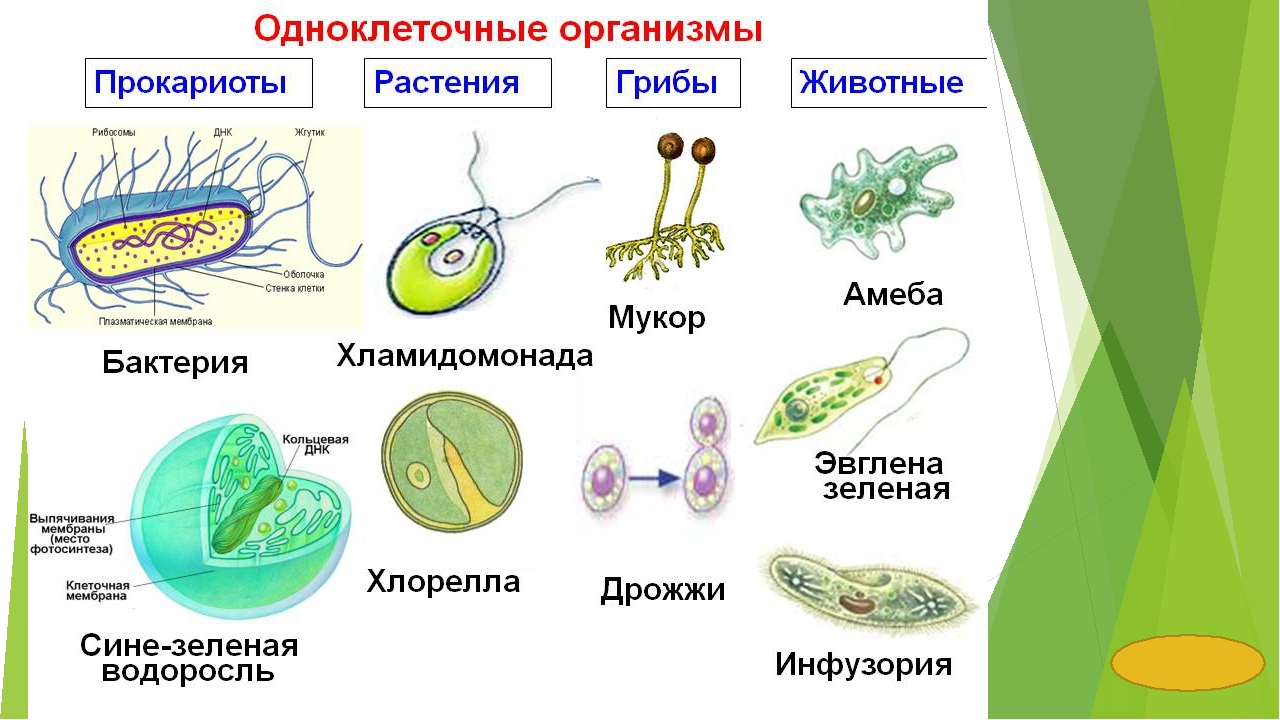 ]
]
9) Обоснование рисунка 4D также должно учитывать потенциальную погрешность в том смысле, что для отношений X; A, как правило, X = 1 или 2 хромосомы и A = 22 или более хромосом.То, что X может измениться, когда одна хромосома изменяет плоидность, но A требует, чтобы 22 хромосомы изменялись равномерно, что менее вероятный сценарий, который следует обсудить или прояснить.
10) В соответствии с этими линиями моделирование на Рисунке 5 иллюстрирует «А» как единый объект с целочисленными перемещениями. Учитывая большое количество аутосомных хромосом, возможность этой модели требует дальнейшего уточнения.
Следуя предложениям рецензентов, мы уточняем описание в расширенном подразделе «Результаты» «Модель эволюции свободноживущих клеток».
Ревизия:
Как показано в разделе «Материалы и методы».
«Предположение модели
В модели аутосомы рассматриваются как интегрированный набор, помечены буквой «A» и учитываются как набор. Для этого могут быть две причины. […] Основная цель работы по моделированию (см. Рис. 5B и 5C) — проверить пригодность этих «целых» генотипов ».
Для этого могут быть две причины. […] Основная цель работы по моделированию (см. Рис. 5B и 5C) — проверить пригодность этих «целых» генотипов ».
В целом, отношения X: A сложны, потому что, хотя можно вычислить среднее значение «A», сложная перестроенная природа многих хромосом раковых клеток (где хромосомы могут быть гибридами многих разных аутосом) может передать значение «A» немного неуверенно.Было бы полезно представить, сколько именно клеточных линий на самом деле X: A1: 2 против 2: 3, а не ни то, ни другое.
В ответе на Q7 выложим процедуру. Вот почему можно было использовать менее половины линий, так как нам пришлось отбросить те линии с частичными анеуплоидами (полиплоидия для частей хромосом). Отметим, что транслокации среди аутосом с низкой анеуплоидией подходят для анализа. С другой стороны, транслокации X: аутосомы (которые встречаются нечасто) не могут быть использованы.
https://doi.org/10.7554/eLife.28070. 027
027
Наименьшая свободноживущая клетка, малый геном
Государственный университет Орегона и корпорация Diversa обнаружили, что самая маленькая свободноживущая клетка, известная как SAR11, также имеет самый маленький геном.
SAR11, группа бактерий, настолько доминирующая, что их совокупный вес превышает вес всех рыб в Мировом океане, процветает там, где погибло бы большинство других клеток, и играет огромную роль в круговороте углерода на Земле.
«Океан — очень конкурентная среда, и эти бактерии, очевидно, выиграли гонку», — сказал Стивен Джованнони, профессор микробиологии Университета штата Орегон. «Наш анализ генома SAR11 показывает, что они стали доминирующей формой жизни в океанах, в основном, будучи простейшими».
SAR11 обитает в морской среде с низким содержанием питательных веществ и других ресурсов, но он способен выживать и размножаться в огромных количествах — миллилитр морской воды у побережья Орегона может содержать 500 000 таких клеток.
Результаты опубликованы в Science.
Авторские права, 2005 г., United Press International
SAR11, самый многочисленный организм в Мировом океане, обладает способностью вырабатывать метан.
Ссылка : Наименьшая свободноживущая клетка, малый геном (20 августа 2005 г.) получено 6 января 2022 г. с https: // физ.org / news / 2005-08-smallest-free-living-cell-small-genome.html
Этот документ защищен авторским правом. За исключением честных сделок с целью частного изучения или исследования, никакие
часть может быть воспроизведена без письменного разрешения. Контент предоставляется только в информационных целях.
За исключением честных сделок с целью частного изучения или исследования, никакие
часть может быть воспроизведена без письменного разрешения. Контент предоставляется только в информационных целях.
Кто открыл свободные живые клетки в воде пруда для класса 9 по биологии CBSE
Подсказка: Теория клеток подчеркивает единство в разнообразии.Это организация форм жизни. У всех живых организмов есть основная единица, называемая клеткой. Все организмы состоят из клеток. Некоторые состоят из одной клетки и называются одноклеточными организмами, а другие состоят из многоклеточных организмов. Клетки выполняют физиологические процессы, такие как пищеварение, выделение, память, защита, распознавание и т. Д. Полное объяснение:
Антон ван Левенгук был первым человеком, который увидел и описал живую клетку. Он был ученым, первым обнаружившим свободноживущие клетки водоросли Spirogyra в воде пруда в \ [1674 \].Он наблюдал за клетками в усовершенствованный электронный микроскоп, который выявил все структурные детали клетки.
Он был ученым, первым обнаружившим свободноживущие клетки водоросли Spirogyra в воде пруда в \ [1674 \].Он наблюдал за клетками в усовершенствованный электронный микроскоп, который выявил все структурные детали клетки.
Маттиас Шлейден был немецким ботаником, который изучил огромное количество растений в \ [1838 \] и обнаружил, что все растения состоят из нескольких типов клеток, которые создают ткани растения. Примерно в то же время Теодор Шванн, британский зоолог, изучал различные типы клеток животных и заметил, что клетки имеют тонкий внешний слой, который сегодня известен как плазматическая мембрана.Шлейден и Шванн вместе сформулировали клеточную теорию.
Итак, клеточная теория, которая вводится в мир, состоит в том, что все живые организмы состоят из клеток и продуктов клеток, и все клетки возникают из уже существующих клеток. В организмах клетка — это фундаментальная единица структуры и организации. Такого общепринятого определения жизни не существует. Некоторые биологи считают неклеточные образования, такие как вирусы, живыми организмами, поэтому обоснованно не согласны с первым принципом.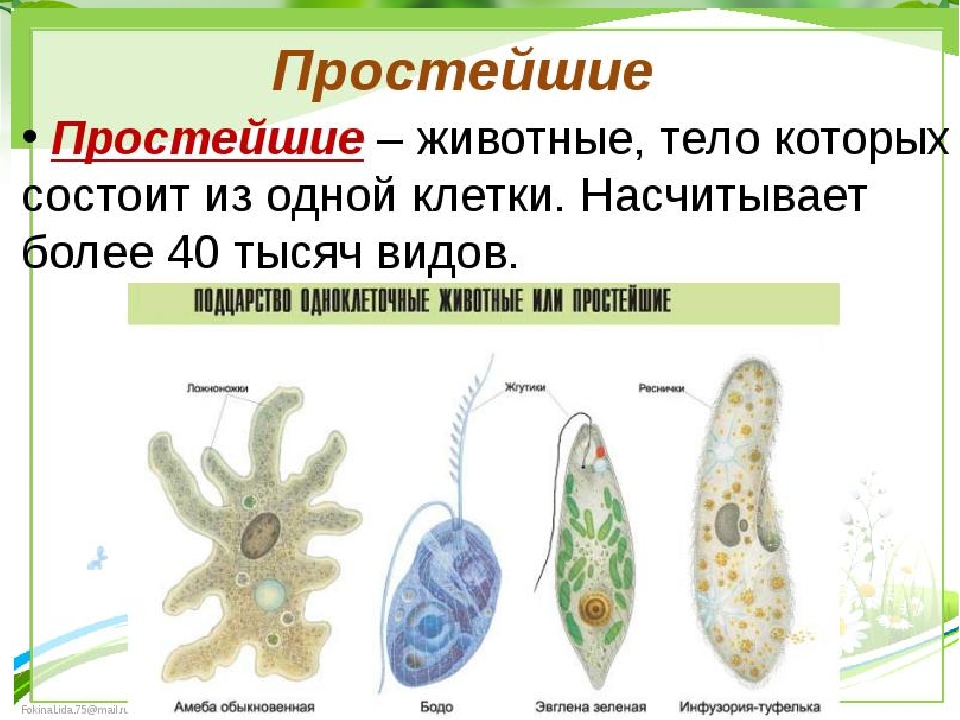
Есть также много ограничений клеточной теории, против которых выступают другие ученые. В теории клетки было много противоположных концепций. Клетка была впервые обнаружена Робертом Гуком в [1665] с помощью микроскопа. В клеточной теории внутренние компоненты клеток назывались протоплазмой и характеризовались как желеобразное вещество, которое также упоминалось как живое желе. Так много других теорий, таких как броуновское движение, полупроницаемая мембрана.
Примечание:
Как мы уже говорили, теория клеток была предложена двумя известными учеными.Они открыли много вещей, и их вклад является важным в теории клеток, и они получили множество наград за свой вклад. Они получили признание в этой области клеточной биологии после публикации своих статей. Это по-прежнему одно из величайших открытий в биологическом мире. Поскольку клетки обладают центром жизни. Жизнь можно развить, размножить из крошечной единственной клетки. Следовательно, это величайшее из известных на сегодняшний день открытий.
Теория эндосимбиотиков | Спросите у биолога
Слизневые плесени — отличный пример сотрудничества между клетками.Щелкните для получения более подробной информации.
Хорошо дружить с соседями, правда? Отдельные люди и сообщества добиваются большего успеха, если они помогают друг другу. Сотрудничество важно не только для людей; без небольшого взаимодействия с соседями жизнь в том виде, в каком мы ее знаем, не существовала бы.
Самые ранние живые соседи на нашей планете были одноклеточными существами. Некоторые из соседних одиночных клеток соединились и начали жить вместе как один организм, одна внутри другого. Это партнерство было настолько успешным, что привело к эволюции многих форм жизни на нашей планете, включая человека.
Что такое ячейка?
Все живые существа состоят из клеток. Несмотря на то, что на Земле существует много миллионов форм жизни, все они состоят только из двух основных типов клеток: прокариот и эукариот.
Клетки содержат ДНК. Прокариоты (про-кэрри-овсы) маленькие и простые, они имеют кольца кольцевой ДНК, свободно плавающие внутри клетки. Эукариоты (овсяные хлопья) большие и более сложные. У них есть ядро, которое удерживает цепочки линейной ДНК внутри липидной мембраны.Все формы жизни, которые вы привыкли видеть — животные (включая людей), растения и грибы — состоят из эукариотических клеток. Бактерии, которые слишком малы, чтобы их можно было увидеть без микроскопа, состоят из прокариотических клеток.
Прокариоты (про-кэрри-овсы) маленькие и простые, они имеют кольца кольцевой ДНК, свободно плавающие внутри клетки. Эукариоты (овсяные хлопья) большие и более сложные. У них есть ядро, которое удерживает цепочки линейной ДНК внутри липидной мембраны.Все формы жизни, которые вы привыкли видеть — животные (включая людей), растения и грибы — состоят из эукариотических клеток. Бактерии, которые слишком малы, чтобы их можно было увидеть без микроскопа, состоят из прокариотических клеток.
Прокариотическая клетка. Щелкните для получения более подробной информации.
Прокариотические клетки были одними из самых ранних форм жизни на Земле. Впервые они появляются в летописи окаменелостей около 4 миллиардов лет назад. Прокариоты существовали долгое время, прежде чем появились эукариотические клетки около 1 года.8 миллиардов лет назад. Это привело нас к мысли, что предком всех эукариотических клеток был прокариот.
Но чтобы превратиться из прокариота в эукариот, клетка должна была стать намного более сложной. Эукариотические клетки питаются от специальных органелл, которые работают как батарейки. У всех эукариот есть органелла, называемая митохондрией, которая вырабатывает энергию для питания клетки. У растительных клеток есть органеллы другого типа, называемые пластидами. Пластиды могут собирать энергию из солнечного света, как солнечная батарея.Хлоропласты — это разновидность пластид.
Эукариотические клетки питаются от специальных органелл, которые работают как батарейки. У всех эукариот есть органелла, называемая митохондрией, которая вырабатывает энергию для питания клетки. У растительных клеток есть органеллы другого типа, называемые пластидами. Пластиды могут собирать энергию из солнечного света, как солнечная батарея.Хлоропласты — это разновидность пластид.
Что такое теория эндосимбиотиков?
Как эукариоты стали такими сложными? А откуда взялись эти батарейчатые органеллы?
Мы думаем, что знаем часть ответа. Эукариотические клетки могли развиться, когда несколько клеток объединились в одну. Они начали жить в так называемых симбиотических отношениях. Теория, объясняющая, как это могло произойти, называется эндосимбиотической теорией. Эндосимбионт — это один организм, живущий внутри другого.Все эукариотические клетки, как и ваша собственная, являются существами, состоящими из частей других существ.
Митохондрии, важные генераторы энергии наших клеток, произошли от свободноживущих клеток. Щелкните для получения более подробной информации.
Щелкните для получения более подробной информации.
Митохондрия и хлоропласт — это органеллы, которые когда-то были свободноживущими клетками. Это были прокариоты, попавшие в другие клетки (клетки-хозяева). Они могли присоединиться к другой клетке, будучи съеденными (процесс, называемый фагоцитозом), или, возможно, они были паразитами этой клетки-хозяина.
Вместо того, чтобы перевариваться или убивать клетку-хозяина, внутренняя клетка выживала и вместе они процветали. Это вроде как домовладелец и арендатор. Клетка-хозяин обеспечивает удобное и безопасное место для жизни, а органелла платит ренту, производя энергию, которую клетка-хозяин может использовать. Это произошло очень давно, и со временем органелла и клетка-хозяин эволюционировали вместе. Теперь одно не могло существовать без другого. Сегодня они функционируют как единый организм, но мы все еще можем найти доказательства свободноживущего прошлого органелл, если присмотреться.
Какие доказательства поддерживают теорию эндосимбиотиков?
Еще в 1883 году ботаник Андреас Шимпер изучал пластидные органеллы растительных клеток с помощью микроскопа. Он смотрел, как делятся пластиды, и заметил кое-что странное. Процесс выглядел очень похоже на то, как делятся некоторые свободноживущие бактерии.
Он смотрел, как делятся пластиды, и заметил кое-что странное. Процесс выглядел очень похоже на то, как делятся некоторые свободноживущие бактерии.
В 1950-х и 60-х годах ученые обнаружили, что и митохондрии, и пластиды внутри растительных клеток имеют собственную ДНК. Он отличался от остальной ДНК растительной клетки.Когда ученые внимательно изучили гены митохондриальной и пластидной ДНК, они обнаружили, что гены больше похожи на гены прокариот. Это говорит нам о том, что органеллы более тесно связаны с прокариотами.
Зеленые хлоропласты в этой клетке сейчас являются важной частью растительных клеток, но они произошли от совершенно другого организма, нежели растительная клетка. Считается, что хлоропласт произошел от цианобактериальной клетки, которая сумела выжить благодаря клеточной защите.
Мы знаем, что органеллы также окружены множеством мембран.Если мы посмотрим на молекулы этих мембран, они выглядят как мембраны, которые окружают современных свободноживущих прокариот.
Итак, органеллы имеют собственную ДНК, и их гены очень похожи на гены современных прокариот. У них есть мембраны, похожие на мембраны прокариот, и, похоже, они также делятся и размножаются аналогичным образом. Если эукариотическая клетка теряет органеллу, она не может ее переделать. Каждая эукариотическая клетка должна унаследовать по крайней мере одну копию органеллы от своей родительской клетки, чтобы выжить.Это означает, что генетическая информация, необходимая для создания органелл, не содержится в ДНК эукариотической клетки. Все эти данные подтверждают теорию о том, что органеллы пришли извне эукариотической клетки. Мы думаем, что это говорит нам о том, что когда-то они были свободноживущими прокариотами.
Эукариотические клетки имеют множество структур, которых нет в прокариотических клетках.
Ученый по имени Линн Маргулис собрал всю эту информацию и опубликовал ее в 1967 году. Ее статья называется «О происхождении митозирующих клеток».Митозирующие клетки — эукариоты. Сегодня ученые знают, что ее статья очень важна, но прошло много лет, прежде чем они приняли ее теорию.
Сегодня ученые знают, что ее статья очень важна, но прошло много лет, прежде чем они приняли ее теорию.
Но наша история эволюции эукариотических клеток далека от завершения. Мы вообще не говорили о других структурах, которые мы можем найти в эукариотических клетках, но не в прокариотических клетках, и о том, как они развивались. К ним относятся ядро, аппарат Гольджи, эндоплазматический ретикулум, лизосомы и цитоскелет.
Откуда они взялись? По правде говоря, мы все еще не уверены.Они могли со временем развиться внутри эукариотических клеток. Или они также могли быть результатом других древних эндосимбиотических событий. Как они развивались — проблема, которую еще предстоит решить.
Дополнительные изображения через Wikimedia Commons. Нитчатые цианобактерии через Салли Уорринг.
Линн Маргулис и гипотеза эндосимбионта: 50 лет спустя
Резюме
Статья 1967 года Линн Маргулис (тогда Линн Саган) «О происхождении митозирующих клеток» в журнале Journal of Theoretical Biology широко известна как стимулирующая и обновленная. интерес к гипотезе происхождения органелл о длительно спящих эндосимбионтах.В своей статье Маргулис не только отстаивала эндосимбиотическое происхождение митохондрий и пластид от бактериальных предков, но также утверждала, что эукариотический жгутик (у нее ундулиподиум) и митотический аппарат произошли от эндосимбиотического, спирохетоподобного организма. По сути, она представила всеобъемлющий симбиотический взгляд на эволюцию эукариотических клеток (эукариогенез). Не все идеи в ее статье были приняты из-за отсутствия убедительных доказательств, но ее энергичное продвижение роли симбиоза в эволюции клеток, несомненно, оказало большое влияние на то, как последующие исследователи рассматривали происхождение и эволюцию митохондрий, пластид и эукариотическая клетка как таковая.
интерес к гипотезе происхождения органелл о длительно спящих эндосимбионтах.В своей статье Маргулис не только отстаивала эндосимбиотическое происхождение митохондрий и пластид от бактериальных предков, но также утверждала, что эукариотический жгутик (у нее ундулиподиум) и митотический аппарат произошли от эндосимбиотического, спирохетоподобного организма. По сути, она представила всеобъемлющий симбиотический взгляд на эволюцию эукариотических клеток (эукариогенез). Не все идеи в ее статье были приняты из-за отсутствия убедительных доказательств, но ее энергичное продвижение роли симбиоза в эволюции клеток, несомненно, оказало большое влияние на то, как последующие исследователи рассматривали происхождение и эволюцию митохондрий, пластид и эукариотическая клетка как таковая.
В 1967 году Линн Маргулис (тогда Линн Саган) опубликовала статью под названием «О происхождении митозирующих клеток» в журнале Journal of Theoretical Biology (Sagan, 1967). У этой публикации не было благоприятного начала: по сообщениям, она была отвергнута более чем дюжиной журналов, прежде чем в конечном итоге нашла свой дом (Archibald, 2014). Сейчас это широко считается знаменателем современного возрождения эндосимбиотической теории происхождения органелл.
Сейчас это широко считается знаменателем современного возрождения эндосимбиотической теории происхождения органелл.
В своей статье Маргулис выдвинула гипотезу, что «три фундаментальные органеллы: митохондрии, фотосинтетические пластиды и (9 + 2) базальные тела жгутиков когда-то сами были свободноживущими (прокариотическими) клетками.«То, что митохондрии и пластиды могли происходить эндосимбиотическим путем от прокариотических предшественников, не было в то время новой идеей, впервые появившись в различных формах в конце 19-го и начале 20-го веков, прежде чем исчезнуть с основной биологической точки зрения (Sapp, 1994). Однако статья Маргулиса примечательна тем, что в ней излагается всеобъемлющий взгляд на (эндо) симбиоз как на конечную цель эукариотической клетки: это была, возможно, первая единая теория эукариогенеза. Статья включала новое предположение о том, что третья субклеточная структура, эукариотический жгутик («ундулиподиум» в ее использовании), возникла в результате «поглощения определенных подвижных прокариот», «возможно, подобных спирохетам», которые в конечном итоге «стали симбиотическими у своих хозяев». Этот общий сценарий позже был назван теорией серийного эндосимбиоза (Taylor, 1974).
Этот общий сценарий позже был назван теорией серийного эндосимбиоза (Taylor, 1974).
Хотя обсуждение происхождения митоза, изложенное Маргулис, составляет значительную часть ее статьи, нет никаких доказательств, подтверждающих это, в отличие от предполагаемого эндосимбиотического происхождения митохондрий и пластид. Причина проста: ни один геном не был связан с жгутиковым аппаратом эукариот, несмотря на попытки его найти (Johnson and Rosenbaum, 1991), и это происходит через геномы, содержащиеся в митохондрии и пластиде — гены, которые они несут и как они устроены. организованы и выражены — что мы знаем с высокой степенью уверенности, откуда произошли эти органеллы: бактериальные клады α-Proteobacteria и Cyanobacteria, соответственно (Gray and Doolittle, 1982; Gray, 1992).
Активная пропаганда Маргулисом роли симбиоза в эволюции эукариотических клеток (Margulis, 1970) вызвала оживленные дискуссии на протяжении 1970-х и в 1980-е годы между сторонниками автогенного происхождения («происхождение изнутри») и ксеногенного происхождения («происхождение изнутри»). без ») теории эволюции органелл. Хотя различные авторы отвергают эндосимбионтный сценарий как для митохондрий, так и для пластид (Uzzell and Spolsky, 1974), разногласия в этот период были сосредоточены особенно на митохондриях (Raff and Mahler, 1972).Особенно тревожным вопросом, ранее отмеченным Малером (1981), был тот факт, что «митохондриальная генетическая система демонстрирует безошибочные признаки большого меж- и внутривидового разнообразия», предполагая, что «эта система уникальна и ее особенности отличаются от как его прокариотические, так и эукариотические аналоги ». Последующий сравнительный анализ митохондриальных геномов и их экспрессии только укрепил мнение о том, что в митохондриях все работает (Burger et al. , 2003). Тем не менее, совокупность данных — биохимических, молекулярных и клеточно-биологических, в сочетании с характеристиками в группе эукариотических микробов (жгутиконосцев якобид) богатого генами митохондриального генома, сильно напоминающего уменьшившийся бактериальный геном (Burger et al.
без ») теории эволюции органелл. Хотя различные авторы отвергают эндосимбионтный сценарий как для митохондрий, так и для пластид (Uzzell and Spolsky, 1974), разногласия в этот период были сосредоточены особенно на митохондриях (Raff and Mahler, 1972).Особенно тревожным вопросом, ранее отмеченным Малером (1981), был тот факт, что «митохондриальная генетическая система демонстрирует безошибочные признаки большого меж- и внутривидового разнообразия», предполагая, что «эта система уникальна и ее особенности отличаются от как его прокариотические, так и эукариотические аналоги ». Последующий сравнительный анализ митохондриальных геномов и их экспрессии только укрепил мнение о том, что в митохондриях все работает (Burger et al. , 2003). Тем не менее, совокупность данных — биохимических, молекулярных и клеточно-биологических, в сочетании с характеристиками в группе эукариотических микробов (жгутиконосцев якобид) богатого генами митохондриального генома, сильно напоминающего уменьшившийся бактериальный геном (Burger et al. , 2013) — теперь это убедительный аргумент в пользу единственного эндосимбиотического α-протеобактериального происхождения митохондрий (Gray et al. , 1999; Gray, 2012).
, 2013) — теперь это убедительный аргумент в пользу единственного эндосимбиотического α-протеобактериального происхождения митохондрий (Gray et al. , 1999; Gray, 2012).
Убедительные аргументы в пользу эндосимбиотического происхождения всегда было легче представить для пластиды, чем для митохондрии. Во-первых, пластида эволюционно моложе митохондрии: тогда как последний общий предок эукариот (LECA) уже имел функциональную митохондрию, приближающуюся к своему современному аналогу (Koumandou et al., 2013), несколько основных эукариотических линий (например, содержащие животных и грибы) явно примитивно апластидны и происходят от предков, у которых никогда не было пластид. Как следствие, у большинства (хотя и не у всех) пластид-несущих эукариот сходство между пластидной и цианобактериальной структурой и биохимией значительно более выражено, чем при сравнении митохондрий / α-протеобактерий. Кроме того, пластидные геномы обычно содержат значительно больше генов, на которых можно основывать такое сравнение, чем митохондриальные геномы, а система трансляции пластид демонстрирует явно более бактериальный характер, чем ее аналог в большинстве митохондриальных систем (Gray, 1992).
Обращение Маргулис к пластиде в ее статье 1967 года на удивление кратко: она просто заявила (стр. 244), что «клетки эукариотических растений не развивают фотосинтез, устраняющий кислород»; вместо этого «они приобрели его в результате симбиоза» (от сине-зеленых водорослей, т. е. цианобактерий). Далее она предположила, что «разные фотосинтезирующие эукариоты (протопластиды) в разное время поедались гетеротрофными простейшими» в ходе эволюции, становясь «обязательно симбиотическими пластидами, сохраняя свои характерные фотосинтетические пигменты и пути метаболизма.Эта тема множественного происхождения пластид была позже подхвачена другими (например, Raven, 1970). Однако в настоящее время консенсус заключается в том, что митохондрия и пластида имеют единое, отдельное эндосимбиотическое происхождение, причем первичное происхождение последних (от эндосимбиотической цианобактерии) происходит от предка Archaeplastida, эукариотической линии, содержащей наземные растения и зеленые, красные и зеленые растения.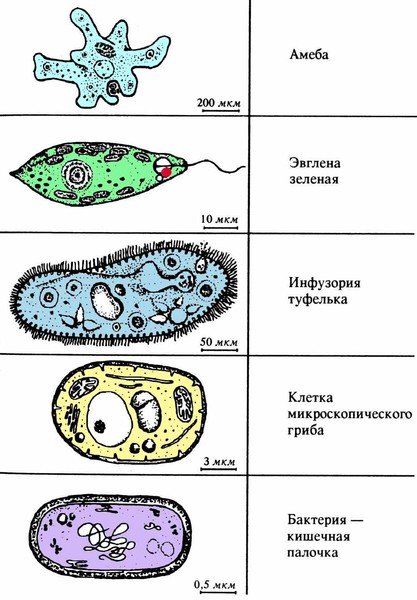 и цианофитные водоросли. Пластиды впоследствии вошли в другие клады водорослей посредством процесса вторичного симбиоза, в котором эукариотический хозяин принимает эукариотического симбионта (зеленую или красную водоросль) (Archibald and Keeling, 2002).
и цианофитные водоросли. Пластиды впоследствии вошли в другие клады водорослей посредством процесса вторичного симбиоза, в котором эукариотический хозяин принимает эукариотического симбионта (зеленую или красную водоросль) (Archibald and Keeling, 2002).
Несмотря на общепринятое мнение о том, что митохондрии и хлоропласты произошли от свободноживущих бактериальных предков в результате процесса эндосимбиоза, то, как на самом деле произошел этот симбиогенез, до сих пор не совсем понятно. В частности, в случае митохондрии, вопросы, которые все еще обсуждаются, включают: когда произошло инициирующее событие (как в рамках общей временной шкалы биологической эволюции, так и относительно происхождения эукариотической клетки как таковой), как долго процесс превращения бактериального эндосимбионта в полную интегрированная органелла, в чем вовлечен этот процесс преобразования и с помощью каких эволюционных механизмов он происходил.Действительно, за пять десятилетий с момента публикации статьи Маргулиса 1967 г. (см., Например, Martin et al. , 2015) было предложено множество моделей симбиогенеза, которые задействуют разные хозяева и процессы.
(см., Например, Martin et al. , 2015) было предложено множество моделей симбиогенеза, которые задействуют разные хозяева и процессы.
Особенно спорным вопросом является природа хоста. В описаниях эндосимбиоза в учебниках хозяин часто изображается как примитивный (т. Е. Амитохондриат) эукариот, поглощающий прокариотический симбионт посредством фаготрофии — процесса захвата и интернализации других организмов (фагоцитоз).Другие изображения показывают хозяина в виде прокариота, проглатывающего другой прокариот, что выглядит как фагоцитоз, хотя фаготрофия у свободноживущих прокариот пока неизвестна.
В своей статье 1967 года Маргулис предположила, что «первый шаг в происхождении эукариот от прокариот был связан с выживанием в новой кислородсодержащей атмосфере: аэробный прокариотический микроб (т. Е. Протомитохондрия) попал в цитоплазму человека. гетеротрофный анаэроб. Этот эндосимбиоз стал облигатным и привел к появлению первых аэробных амитотических амебоидных организмов. Из этого описания неясно, был ли предполагаемый хозяин сам прокариотом или чем-то более «продвинутым»: Маргулис не дает точного ответа на этот вопрос. Намек на «амебоид» и «прием пищи» действительно предполагает тип протоэукариот, хотя и без многих определяющих черт современной эукариотической клетки, в частности ядра и митотического аппарата. Позже, однако, Маргулис (1981) прояснила, что она предпочитает прокариотического хозяина, заявив: «вполне вероятно, что протомитохондрии вторглись в их хозяев так же, как современные хищные бактерии Bdellovibrio вторгаются в хищные бактерии»: «удивительный пример прокариот-прокариот. «эмбоитмент» без фагоцитоза.Имейте в виду, что, учитывая, что Bdellovibrio очень эффективно уничтожает свою «хозяйскую» бактерию в процессе вторжения в нее, этот тип сценария действительно предлагает особенно многообещающий путь к стабильному симбиозу прокариот-прокариот.
Из этого описания неясно, был ли предполагаемый хозяин сам прокариотом или чем-то более «продвинутым»: Маргулис не дает точного ответа на этот вопрос. Намек на «амебоид» и «прием пищи» действительно предполагает тип протоэукариот, хотя и без многих определяющих черт современной эукариотической клетки, в частности ядра и митотического аппарата. Позже, однако, Маргулис (1981) прояснила, что она предпочитает прокариотического хозяина, заявив: «вполне вероятно, что протомитохондрии вторглись в их хозяев так же, как современные хищные бактерии Bdellovibrio вторгаются в хищные бактерии»: «удивительный пример прокариот-прокариот. «эмбоитмент» без фагоцитоза.Имейте в виду, что, учитывая, что Bdellovibrio очень эффективно уничтожает свою «хозяйскую» бактерию в процессе вторжения в нее, этот тип сценария действительно предлагает особенно многообещающий путь к стабильному симбиозу прокариот-прокариот.
Природа хозяина, по сути, является центральной для широко различающихся моделей симбиогенеза митохондриального происхождения и эволюции, которые делятся примерно на две широкие категории: ранние митохондрии (мито-ранние или митохондриальные) и поздние митохондрии (митохондриальные). late, или mito-last), различающиеся по времени — в пределах перехода от первого общего предка эукариот к LECA — и имеющие разные последствия для общего происхождения эукариотической клетки (Poole and Gribaldo, 2014).Сравнительная геномика и другие анализы подчеркивают, что LECA уже был сложным организмом с полностью функционирующей митохондрией (Koumandou et al. , 2013) и что все предположительно амитохондриальные эукариотические линии (за одним недавно описанным исключением; Karnkowska et al. , 2016) содержат связанные с митохондриями органеллы и происходят от митохондрий-содержащих предков. Таким образом, первоначальное приобретение бактериального симбионта, которому суждено было стать митохондрией, не могло произойти очень близко к появлению LECA, учитывая многочисленные и сложные изменения, которые, очевидно, должны были произойти при переходе от симбионта к органелле, хотя это свидетельствует о том, что Позднее приобретение митохондрии было недавно опубликовано (Pittis and Gabaldón, 2016).
late, или mito-last), различающиеся по времени — в пределах перехода от первого общего предка эукариот к LECA — и имеющие разные последствия для общего происхождения эукариотической клетки (Poole and Gribaldo, 2014).Сравнительная геномика и другие анализы подчеркивают, что LECA уже был сложным организмом с полностью функционирующей митохондрией (Koumandou et al. , 2013) и что все предположительно амитохондриальные эукариотические линии (за одним недавно описанным исключением; Karnkowska et al. , 2016) содержат связанные с митохондриями органеллы и происходят от митохондрий-содержащих предков. Таким образом, первоначальное приобретение бактериального симбионта, которому суждено было стать митохондрией, не могло произойти очень близко к появлению LECA, учитывая многочисленные и сложные изменения, которые, очевидно, должны были произойти при переходе от симбионта к органелле, хотя это свидетельствует о том, что Позднее приобретение митохондрии было недавно опубликовано (Pittis and Gabaldón, 2016).
Пожалуй, самой известной мито-ранней моделью является водородная гипотеза (Martin and Müller, 1998), в которой хозяин, анаэробный водородзависимый археон, принимает α-протеобактерию, которая способна дышать, но генерирует молекулярный водород как побочный продукт анаэробного гетеротрофного метаболизма. Здесь отбор на эндосимбиоз обусловлен метаболической синтрофией между двумя партнерами: продукт жизнедеятельности (водород) одного используется в качестве важного метаболического ресурса другим.В этом сценарии происхождение митохондрии и происхождение эукариотической клетки одновременны, с последующим появлением определяющих субклеточных свойств последней, напрямую зависящих от выраженного увеличения клеточной энергии, обеспечиваемой последней.
Напротив, в моделях мито-лат основным механизмом симбиогенеза является фаготрофия, отличительный признак эукариотических клеток и широко распространенный в эукариотическом домене. Фаготрофия включает эндоцитоз, при котором ограничивающая мембрана одного организма (хозяина) окружает другой организм (симбионт), интернализируя его в мембраносвязанную фагосому. В некоторых мито-поздних моделях хозяином фактически является амитохондриальный эукариот, способный к фагоцитозу (Cavalier-Smith, 1987).
В некоторых мито-поздних моделях хозяином фактически является амитохондриальный эукариот, способный к фагоцитозу (Cavalier-Smith, 1987).
Недавние данные подтверждают идею, что эукариоты специфически связаны с недавно описанной кладой архей, суперфилумом Асгарда (Zaremba-Niedzwiedzka et al. , 2017). Эта группа архей кодирует ряд белков, гомологи которых ранее были обнаружены только у эукариот, что позволяет предположить, что линия архей, которая уже имела особенности, характерные для эукариот, включая, возможно, фагоцитоз, могла быть хозяином для митохондриального эндосимбиоза.Эти наблюдения обеспечивают основу для модели эукариогенеза с теорией фагоцитозирующего архея (PhAT), при этом митохондриальный эндосимбионт был приобретен временно сложным фагоцитирующим археоном (Martijn and Ettema, 2013).
Моя цель здесь — не перечислить и критически оценить все различные симбиогенетические модели эволюции органелл, а скорее проиллюстрировать на нескольких примерах расходящиеся (и меняющиеся) мнения по этому поводу.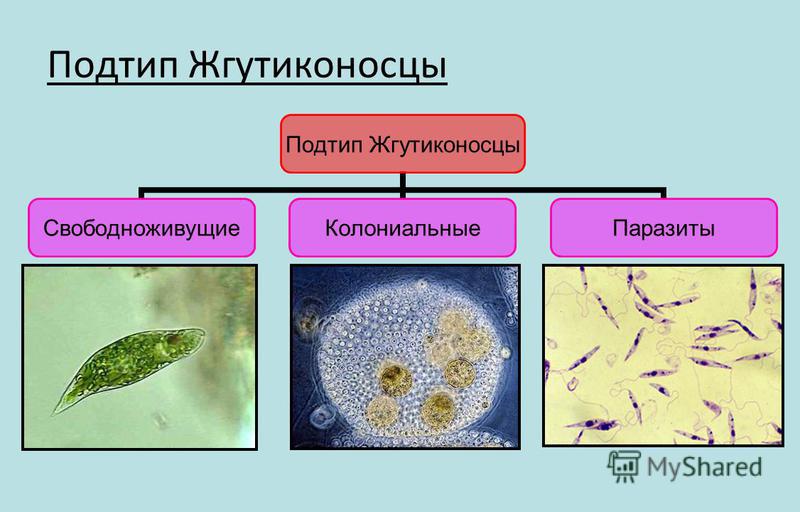 Переход от симбионта к органелле, очевидно, включал много шагов: потеря стенки бактериальной клетки; раннее приобретение симбионтом переносчиков основных метаболитов; массивное (и непостоянное) сокращение генома симбионта за счет потери генов или их переноса в ядро; функциональная активация перенесенных генов в ядре и перенацеливание их цитоплазматически синтезированных белковых продуктов обратно в развивающуюся органеллу — через специфические нацеленные на органеллы последовательности — или в другое место в клетке; и массовое привлечение многих дополнительных органелларных белков, происхождение которых неясно.Мы делаем вывод об этом процессе ремоделирования, сравнивая современные органеллы с их ближайшими бактериальными родственниками, и заключаем, что это был постепенный процесс, происходящий в течение длительного периода времени.
Переход от симбионта к органелле, очевидно, включал много шагов: потеря стенки бактериальной клетки; раннее приобретение симбионтом переносчиков основных метаболитов; массивное (и непостоянное) сокращение генома симбионта за счет потери генов или их переноса в ядро; функциональная активация перенесенных генов в ядре и перенацеливание их цитоплазматически синтезированных белковых продуктов обратно в развивающуюся органеллу — через специфические нацеленные на органеллы последовательности — или в другое место в клетке; и массовое привлечение многих дополнительных органелларных белков, происхождение которых неясно.Мы делаем вывод об этом процессе ремоделирования, сравнивая современные органеллы с их ближайшими бактериальными родственниками, и заключаем, что это был постепенный процесс, происходящий в течение длительного периода времени.
Хотя мы многое узнали о происхождении митохондрий и пластид за пять десятилетий, прошедших с момента публикации статьи Маргулиса 1967 года, и об их роли в общем процессе эукариогенеза, очевидно, что еще многое предстоит выяснить.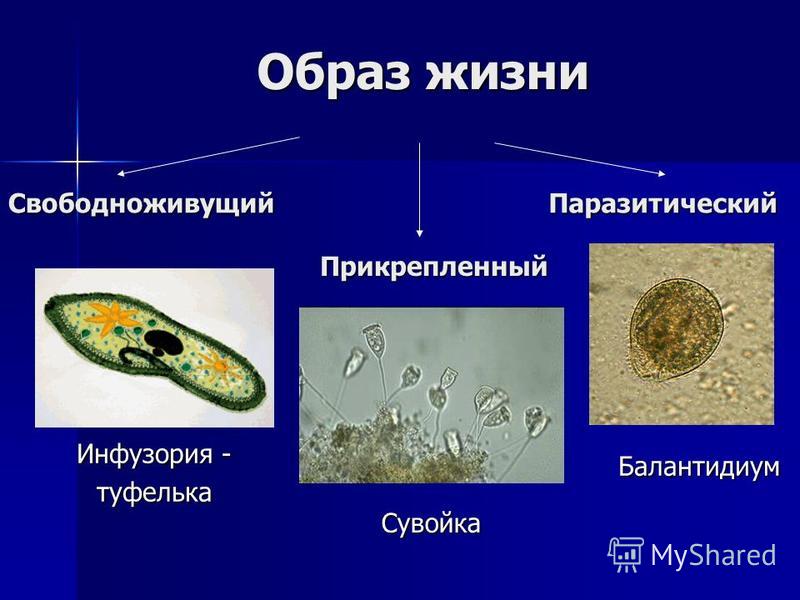 Особенно сложной задачей является расшифровка происхождения большей части митохондриальных и пластидных протеомов, которые, по-видимому, не имеют α-протеобактериального или цианобактериального происхождения соответственно.Мы принимаем, что митохондрии и пластиды являются продуктами прямого эволюционного развития бактериального эндосимбиоза, но в то же время мы признаем, что это мозаичные органеллы, компоненты и функции которых имеют более чем одно происхождение (Cavalier-Smith, 1987). Как утверждала Маргулис на протяжении всей своей карьеры, симбиоз сыграл решающую роль в происхождении органелл и общем эукариогенезе, хотя, возможно, не в той всеобъемлющей и всеобъемлющей роли, которую она изначально предполагала.
Особенно сложной задачей является расшифровка происхождения большей части митохондриальных и пластидных протеомов, которые, по-видимому, не имеют α-протеобактериального или цианобактериального происхождения соответственно.Мы принимаем, что митохондрии и пластиды являются продуктами прямого эволюционного развития бактериального эндосимбиоза, но в то же время мы признаем, что это мозаичные органеллы, компоненты и функции которых имеют более чем одно происхождение (Cavalier-Smith, 1987). Как утверждала Маргулис на протяжении всей своей карьеры, симбиоз сыграл решающую роль в происхождении органелл и общем эукариогенезе, хотя, возможно, не в той всеобъемлющей и всеобъемлющей роли, которую она изначально предполагала.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера на прием файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.

Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в cookie-файлах может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
границ | Остерегайтесь «живых мертвецов»: бесклеточные ферменты и их судьба
Важность микробов и их внеклеточной ферментативной активности (EEA)
Морская среда играет решающую роль в глобальных биогеохимических циклах (Daily, 2003; Harley et al., 2006; Хатчинс и Фу, 2017). Микробы являются двигателями биогеохимических циклов Земли (Falkowski et al., 2008). У этих крошечных организмов есть набор основных генов, кодирующих ферменты основных реакций, ответственных за преобразование энергии и материи в (пригодные для использования) субстраты, необходимые для жизни (Falkowski et al., 2008). Мы живем во времена перемен, и антропогенные воздействия могут изменить структуру и функционирование морских микробных сообществ и, как следствие, роль океана в глобальных биогеохимических циклах.Таким образом, если мы стремимся понять, каким будет будущее морского биогеохимического круговорота, нам необходимо понять, какова будет судьба микробов и их ферментов.
Когда дело доходит до потребления органического вещества для преобразования и переработки, микробы, похоже, отдают предпочтение определенным типам органических веществ. Согласно модели «размер-реактивность», гетеротрофные микробы предпочтительно разлагают высокомолекулярное растворенное органическое вещество (РОВ), потому что оно имеет тенденцию быть более биодоступным, чем РОВ с низкой молекулярной массой (Benner and Amon, 2015).Но за эту пищевую селективность приходится платить: гетеротрофным прокариотам необходимо будет гидролизовать большинство этих молекул до субъединиц, достаточно малых для включения, потому что большинство молекул должны быть меньше 600 Да, чтобы проходить через стенку прокариотических клеток (Weiss et al. , 1991). Для этого они используют внеклеточные ферменты; поэтому из-за центральной роли этих ферментов их называют «привратниками» цикла C (Arnosti, 2011). Однако недавно был обнаружен альтернативный механизм поглощения полисахаридов бактериями, который позволяет им напрямую включать соединения РОВ с большой молекулярной массой (Cuskin et al., 2015; Reintjes et al., 2017). Тем не менее, не весь высокомолекулярный РОВ может переноситься с помощью этого механизма (т.
Согласно модели «размер-реактивность», гетеротрофные микробы предпочтительно разлагают высокомолекулярное растворенное органическое вещество (РОВ), потому что оно имеет тенденцию быть более биодоступным, чем РОВ с низкой молекулярной массой (Benner and Amon, 2015).Но за эту пищевую селективность приходится платить: гетеротрофным прокариотам необходимо будет гидролизовать большинство этих молекул до субъединиц, достаточно малых для включения, потому что большинство молекул должны быть меньше 600 Да, чтобы проходить через стенку прокариотических клеток (Weiss et al. , 1991). Для этого они используют внеклеточные ферменты; поэтому из-за центральной роли этих ферментов их называют «привратниками» цикла C (Arnosti, 2011). Однако недавно был обнаружен альтернативный механизм поглощения полисахаридов бактериями, который позволяет им напрямую включать соединения РОВ с большой молекулярной массой (Cuskin et al., 2015; Reintjes et al., 2017). Тем не менее, не весь высокомолекулярный РОВ может переноситься с помощью этого механизма (т. Е. Биохимические и микробиологические исследования показывают, что связывание / гидролиз / транспорт очень селективны для конкретных полисахаридов), и этот механизм все еще включает внеклеточный гидролиз — но связывающие белки зависают. на части, так что они транспортируются в клетку без потерь для внешней среды (Cuskin et al., 2015). Тем не менее внеклеточная ферментативная активность (EEA) обнаруживается в эпипелагических и батипелагических водах, обычно наблюдая увеличение отношения EEA к численности клеток с глубиной (Hoppe and Ullrich, 1999; Hoppe et al., 2002; Балтар и др., 2009).
Е. Биохимические и микробиологические исследования показывают, что связывание / гидролиз / транспорт очень селективны для конкретных полисахаридов), и этот механизм все еще включает внеклеточный гидролиз — но связывающие белки зависают. на части, так что они транспортируются в клетку без потерь для внешней среды (Cuskin et al., 2015). Тем не менее внеклеточная ферментативная активность (EEA) обнаруживается в эпипелагических и батипелагических водах, обычно наблюдая увеличение отношения EEA к численности клеток с глубиной (Hoppe and Ullrich, 1999; Hoppe et al., 2002; Балтар и др., 2009).
Живые и «живые мертвецы»: прикрепленные к клеткам или бесклеточные EEA
Внеклеточные ферменты существуют в двух формах; связанные с клетками (т. е. прикрепленные к клеткам) и растворенные (т. е. бесклеточные, оперативно определяемые как прохождение через фильтр 0,22 мкм). Общий EEA является результатом комбинации клеточно-ассоциированных и внеклеточных ферментов (рис. 1). Используя Интернет в качестве аналогии, мы могли бы сказать, что EEA с привязкой к ячейке будет эквивалентом «проводного» подключения к Интернету, тогда как EEA без сотовой связи будет «беспроводным» подключением (т. е.е., по-прежнему будет обеспечивать конечный продукт — передачу данных / гидролизат — даже если физически не подключен к оборудованию / ячейке). Были предложены различные источники бесклеточных ферментов, включая активное высвобождение бактериями в ответ на соответствующий субстрат (Alderkamp et al., 2007) и бактериальное голодание (Albertson et al., 1990), а также изменения проницаемости клеток (Chrost , 1990). Эти исследования показывают, что морские бактерии могут выделять ферменты в окружающую среду не только для облегчения гидролиза определенных субстратов во время экспоненциального роста на определенных субстратах, но также что голодные клетки во время стационарной фазы показали большее высвобождение внеклеточных ферментов, чем в начале голодания.Фактически, даже во время этого периода голодания происходит синтез белка de novo для производства и / или высвобождения бесклеточных ферментов в окружающую среду, что подчеркивает важность этого процесса для некоторых морских бактерий (Albertson et al.
е.е., по-прежнему будет обеспечивать конечный продукт — передачу данных / гидролизат — даже если физически не подключен к оборудованию / ячейке). Были предложены различные источники бесклеточных ферментов, включая активное высвобождение бактериями в ответ на соответствующий субстрат (Alderkamp et al., 2007) и бактериальное голодание (Albertson et al., 1990), а также изменения проницаемости клеток (Chrost , 1990). Эти исследования показывают, что морские бактерии могут выделять ферменты в окружающую среду не только для облегчения гидролиза определенных субстратов во время экспоненциального роста на определенных субстратах, но также что голодные клетки во время стационарной фазы показали большее высвобождение внеклеточных ферментов, чем в начале голодания.Фактически, даже во время этого периода голодания происходит синтез белка de novo для производства и / или высвобождения бесклеточных ферментов в окружающую среду, что подчеркивает важность этого процесса для некоторых морских бактерий (Albertson et al. , 1990). Помимо прямого высвобождения, существуют и другие косвенные источники внеклеточных ферментов, включая выпас на бактериальных сообществах (Bochdansky et al., 1995) и вирусный лизис (Karner and Rassoulzadegan, 1995).
, 1990). Помимо прямого высвобождения, существуют и другие косвенные источники внеклеточных ферментов, включая выпас на бактериальных сообществах (Bochdansky et al., 1995) и вирусный лизис (Karner and Rassoulzadegan, 1995).
РИСУНОК 1. Общая внеклеточная ферментативная активность (EEA) является результатом комбинации клеточно-ассоциированных и внеклеточных ферментов. Используя аналогию с подключением к Интернету; EEA с привязкой к ячейке будет эквивалентом «проводного» соединения, тогда как EEA без ячейки будет «беспроводным» подключением. Более того, связанный с клеткой EEA будет тесно связан с поведением живой клетки, тогда как внеклеточные ферменты не обязательно будут связаны с факторами, влияющими на микробные клетки, которые их первоначально продуцировали, и будут оставаться активными в течение длительного периода времени. , вдали от камеры.Таким образом, используя другую аналогию, мы могли бы сказать, что присоединенный к клетке EEA будет более тесно связан с «живой» активностью, тогда как бесклеточный EEA будет в сфере «живых мертвецов».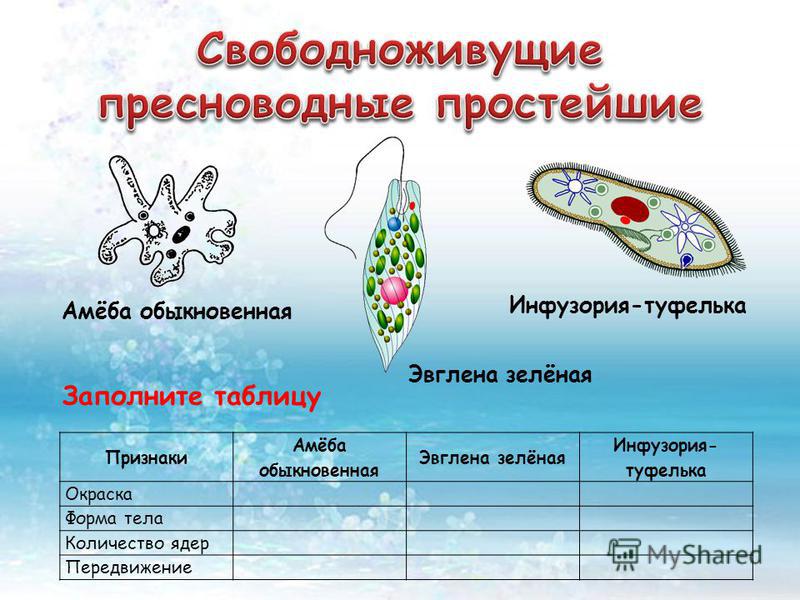
Тогда возникает вопрос, является ли стратегия высвобождения ферментов в водную среду энергетически и экологически эффективной. Было показано, что внеклеточные ферменты, выделяемые бактериями, производят из твердых частиц органического углерода достаточное количество продуктов для поддержки роста бактерий в отсутствие какого-либо другого значительного источника органического углерода и без прямого контакта между клеткой и твердым субстратом (Vetter and Deming, 1999). ).Эти эмпирические результаты, полученные Веттером и Демингом (1999), подтверждают прогнозы, сформулированные ими в модели (Веттер и др., 1998), предполагая, что растворенные внеклеточные ферменты имеют преимущество, когда бактерии прикреплены к частицам и когда субстрат находится внутри лунки. определенное расстояние от источника фермента. Более того, другие моделирование позже показало, что, когда микробы, продуцирующие ферменты, конкурировали с непродуцентами: (1) непродуцентам благоприятствовала более высокая стоимость ферментов, (2) производителям благоприятствовала более низкая скорость диффузии ферментов и (3) не производились -производители и продуценты сосуществовали в высокоорганизованных пространственных структурах при промежуточных затратах на ферменты и скорости распространения (Allison, 2005). Эти исследования также согласуются с недавно предложенными решениями «дилеммы общественного блага» в бактериальных биопленках (Drescher et al., 2014). В этом исследовании секреция общественных благ (например, хитиназ), которые потенциально могут использоваться непродуцирующими клетками, объяснялась двумя механизмами: клетки могут производить толстые биопленки, которые ограничивают товары производителями, или поток жидкости может удалять растворимые продукты переваривание хитина, закрывая доступ непроизводителям. Совсем недавно другая модель предположила, что внеклеточные ферменты не только могут быть прибыльной стратегией для микробов, живущих рядом с частицами или прикрепленных к ним, но пошла дальше, чтобы указать, что бесклеточные ферменты могут быть жизнеспособной стратегией даже для свободноживущих клеток, если использование субстрата рассматривается как совместная работа (Traving et al., 2015). Эти авторы также предположили, что даже небольшое количество долгоживущих бесклеточных ферментов, выделяемых бесклеточными микробами, может вносить значительный вклад в пул активности растворенных ферментов.
Эти исследования также согласуются с недавно предложенными решениями «дилеммы общественного блага» в бактериальных биопленках (Drescher et al., 2014). В этом исследовании секреция общественных благ (например, хитиназ), которые потенциально могут использоваться непродуцирующими клетками, объяснялась двумя механизмами: клетки могут производить толстые биопленки, которые ограничивают товары производителями, или поток жидкости может удалять растворимые продукты переваривание хитина, закрывая доступ непроизводителям. Совсем недавно другая модель предположила, что внеклеточные ферменты не только могут быть прибыльной стратегией для микробов, живущих рядом с частицами или прикрепленных к ним, но пошла дальше, чтобы указать, что бесклеточные ферменты могут быть жизнеспособной стратегией даже для свободноживущих клеток, если использование субстрата рассматривается как совместная работа (Traving et al., 2015). Эти авторы также предположили, что даже небольшое количество долгоживущих бесклеточных ферментов, выделяемых бесклеточными микробами, может вносить значительный вклад в пул активности растворенных ферментов.
Когда эти ферменты высвобождаются, они могут сохранять активность (т.е.способны выполнять свои функции) до тех пор, пока не достигнут определенного субстрата, а затем гидролизуют его. Существует не так много исследований, изучающих время жизни этих бесклеточных ферментов в морской воде, но имеющиеся показывают, что время их полужизни может достигать 20 дней (Ziervogel et al., 2010; Стин и Арности, 2011; Baltar et al., 2013), и что глубоководные ферменты, по-видимому, имеют более продолжительный срок жизни, чем поверхностные (Baltar et al., 2013). Эти наблюдения особенно интересны, потому что, хотя изначально предполагалось, что только связанная с клетками активность будет иметь значение в морской среде (Rego et al., 1985; Chrost and Rai, 1993), накапливались данные, ясно указывающие на то, что фракция растворенных EEA сопоставим со связанной фракцией (Karner and Rassoulzadegan, 1995; Li et al., 1998; Балтар и др., 2010, 2013, 2016; Duhamel et al., 2010; Allison et al., 2012).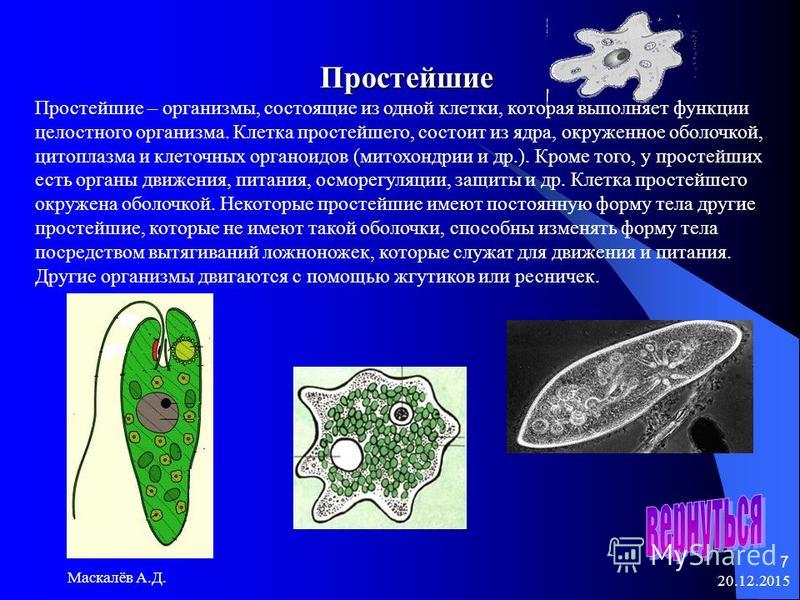 Фактически, во многих случаях было замечено, что доля растворенного EEA может достигать значения 100%, что означает, что вся ферментативная активность в этих водах во время отбора проб осуществлялась бесклеточными ферментами ( например, Karner and Rassoulzadegan, 1995; Baltar et al., 2010, 2016). Таким образом, несоответствие между показателями EEA и составом сообщества неудивительно (D’ambrosio et al., 2014). Когда внеклеточные ферменты ответственны за высокую долю общего EEA, возникает разрыв между микробами и ферментативной активностью; то есть отделение скоростей гидролиза на месте от реальной микробной динамики.Таким образом, высокая доля растворенного EEA может указывать на большее значение истории водной массы, чем реальных процессов, происходящих во время отбора проб (Karner and Rassoulzadegan, 1995; Baltar et al., 2010, 2016; Arnosti, 2011 ).
Фактически, во многих случаях было замечено, что доля растворенного EEA может достигать значения 100%, что означает, что вся ферментативная активность в этих водах во время отбора проб осуществлялась бесклеточными ферментами ( например, Karner and Rassoulzadegan, 1995; Baltar et al., 2010, 2016). Таким образом, несоответствие между показателями EEA и составом сообщества неудивительно (D’ambrosio et al., 2014). Когда внеклеточные ферменты ответственны за высокую долю общего EEA, возникает разрыв между микробами и ферментативной активностью; то есть отделение скоростей гидролиза на месте от реальной микробной динамики.Таким образом, высокая доля растворенного EEA может указывать на большее значение истории водной массы, чем реальных процессов, происходящих во время отбора проб (Karner and Rassoulzadegan, 1995; Baltar et al., 2010, 2016; Arnosti, 2011 ).
Возможные различия во временных масштабах активности для прикрепленных к клеткам и внеклеточных ферментов поднимают вопрос о контроле над этими пулами ферментов. Связанный с клеткой EEA будет тесно связан с поведением живой клетки (т.е. на него будут влиять те же факторы, которые контролируют рост, активность и разнообразие живых клеток).Напротив, бесклеточные ферменты не обязательно будут подвержены влиянию одних и тех же факторов, влияющих на клетки, но будут оставаться активными в течение длительного периода времени и, вероятно, будут подвержены влиянию разных факторов (или по-разному в ответ на одни и те же факторов), чем связанный с клеткой EEA. Имея это в виду, используя другую аналогию, мы могли бы сказать, что связанный с клеткой EEA является активностью «живого», поскольку эта активность выполняется прикрепленной к живой клетке, и, таким образом, эта активность будет изменяться в ответ на потребности клетки и в ответ на динамику клеток / сообщества (рис. 1).В то время как бесклеточный EEA ближе к деятельности «живых мертвецов» в том смысле, что он больше не «живой», потому что отделен от живой клетки, но все же остается каким-то образом «живым» в смысле что он все еще может выполнять свою функцию, когда сталкивается с правильным субстратом.
Связанный с клеткой EEA будет тесно связан с поведением живой клетки (т.е. на него будут влиять те же факторы, которые контролируют рост, активность и разнообразие живых клеток).Напротив, бесклеточные ферменты не обязательно будут подвержены влиянию одних и тех же факторов, влияющих на клетки, но будут оставаться активными в течение длительного периода времени и, вероятно, будут подвержены влиянию разных факторов (или по-разному в ответ на одни и те же факторов), чем связанный с клеткой EEA. Имея это в виду, используя другую аналогию, мы могли бы сказать, что связанный с клеткой EEA является активностью «живого», поскольку эта активность выполняется прикрепленной к живой клетке, и, таким образом, эта активность будет изменяться в ответ на потребности клетки и в ответ на динамику клеток / сообщества (рис. 1).В то время как бесклеточный EEA ближе к деятельности «живых мертвецов» в том смысле, что он больше не «живой», потому что отделен от живой клетки, но все же остается каким-то образом «живым» в смысле что он все еще может выполнять свою функцию, когда сталкивается с правильным субстратом.
Однако важно понимать, что столкновение с частицами не обязательно является концом линии для фермента, и большая часть бесклеточных ферментов может быть захвачена частицами, включая коллоиды или липосомы.Фактически, время жизни бесклеточных ферментов можно продлить, если они связаны с частицами (Gianfreda and Scarfi, 1991; Ziervogel et al., 2007), поскольку поверхностные ассоциации могут обеспечить повышенную устойчивость к физико-химической деградации (Lähdesmäki and Piispanen, 1992) и защиты от реминерализации (Lozzi et al., 2001). Сообщалось о наличии бактериальных бесклеточных ферментов, встроенных в экзополимерный матрикс (Decho, 1990), где бесклеточные EE, прикрепленные к этой матрице, могут образовывать комплекс, подобный ферментно-гуминовым комплексам в почвах (Chrost, 1990).Кроме того, некоторые ферменты могут быть связаны с частицами из-за захвата пищеварительных ферментов частично разрушенными бактериальными мембранами, которые действуют как мицеллы (липосомы) (Nagata and Kirchman, 1992).
Судьба «живых мертвецов» без клеток EEA
Активность внеклеточных ферментов контролирует скорость переработки органических веществ в океане. Учитывая свидетельства высокой активности внеклеточных ферментов, важно понимать, как контролировать время жизни и активность этих внеклеточных ферментов.
Исследований по этой теме очень мало, но они уже начинают выявлять некоторые ключевые факторы, контролирующие относительную важность бесклеточного EEA, и температура, по-видимому, является основным из них. В сезонном исследовании Балтийского моря (продолжительностью 1,5 года) была обнаружена значимая обратная зависимость между долей растворенных веществ по отношению к общему количеству EEA (Baltar et al., 2016). В лабораторном эксперименте инкубация микробных сообществ из вод Большого Барьерного рифа (Австралия) при трех различных температурах (т.e., in situ , +3 и -3 ° C), снова была обнаружена значимая обратная зависимость между температурой и относительной долей растворенных веществ в общем EEA (Baltar et al. , 2017). Эти результаты согласуются с наблюдаемым увеличением доли растворенных веществ по отношению к общему количеству EEA с глубиной по мере того, как температура падает по всей толще воды на (суб) тропическом разрезе Атлантического океана (Baltar et al., 2010). Эти исследования указывают на то, с разных сторон (например, сезонные исследования, исследования в лаборатории по изменению климата и круизы по трансектам), температура как главный фактор, влияющий на относительную важность и активность бесклеточного EEA.Все эти исследования, кажется, указывают на то, что чем выше температура, тем ниже доля растворенного EEA. Это также согласуется с более длительным сроком жизни бесклеточных ферментов, обнаруженным для глубинных вод по сравнению с поверхностными (Baltar et al., 2013).
, 2017). Эти результаты согласуются с наблюдаемым увеличением доли растворенных веществ по отношению к общему количеству EEA с глубиной по мере того, как температура падает по всей толще воды на (суб) тропическом разрезе Атлантического океана (Baltar et al., 2010). Эти исследования указывают на то, с разных сторон (например, сезонные исследования, исследования в лаборатории по изменению климата и круизы по трансектам), температура как главный фактор, влияющий на относительную важность и активность бесклеточного EEA.Все эти исследования, кажется, указывают на то, что чем выше температура, тем ниже доля растворенного EEA. Это также согласуется с более длительным сроком жизни бесклеточных ферментов, обнаруженным для глубинных вод по сравнению с поверхностными (Baltar et al., 2013).
Еще одним фактором, который, по-видимому, в значительной степени способствует контролю продолжительности жизни бесклеточных ферментов, является ультрафиолетовое излучение (УФР). На эту тему также имеется очень мало исследований. Воздействие УФИ на бесклеточные ферменты непосредственно было протестировано в морской воде Арктики, и было обнаружено, что, хотя естественное освещение не вызывает значительных эффектов фотодеградации, снижение активности бесклеточных ферментов было обнаружено при искусственно высоких дозах УФИ (т.е., интенсивность УФ-B в 5–10 раз выше, чем у in situ ) (Steen, Arnosti, 2011). Интересно, что эти авторы обнаружили значительный эффект УФИ на лейцинаминопептидазу и щелочную фосфатазу, но не на бета-глюкозидазу при любом уровне лечения. Недавнее исследование бесклеточных ферментов из вод Новой Зеландии показало, что экологически значимое УФ-излучение снижает активность бесклеточных ферментов до 87% за 36 часов по сравнению с темным контролем, что, вероятно, является следствием фотодеградации (Thomson et al., 2017). Это исследование также показало, что величина воздействия УФИ на бесклеточные ферменты варьировалась в зависимости от фракции УФИ. Интересно, что в соответствии с выводами Steen and Arnosti (2011) эффект УФИ различается в зависимости от фермента; значительно снижая активность внеклеточной лейцинаминопептидазы и щелочной фосфатазы, но не влияя на β-глюкозидазу.
Воздействие УФИ на бесклеточные ферменты непосредственно было протестировано в морской воде Арктики, и было обнаружено, что, хотя естественное освещение не вызывает значительных эффектов фотодеградации, снижение активности бесклеточных ферментов было обнаружено при искусственно высоких дозах УФИ (т.е., интенсивность УФ-B в 5–10 раз выше, чем у in situ ) (Steen, Arnosti, 2011). Интересно, что эти авторы обнаружили значительный эффект УФИ на лейцинаминопептидазу и щелочную фосфатазу, но не на бета-глюкозидазу при любом уровне лечения. Недавнее исследование бесклеточных ферментов из вод Новой Зеландии показало, что экологически значимое УФ-излучение снижает активность бесклеточных ферментов до 87% за 36 часов по сравнению с темным контролем, что, вероятно, является следствием фотодеградации (Thomson et al., 2017). Это исследование также показало, что величина воздействия УФИ на бесклеточные ферменты варьировалась в зависимости от фракции УФИ. Интересно, что в соответствии с выводами Steen and Arnosti (2011) эффект УФИ различается в зависимости от фермента; значительно снижая активность внеклеточной лейцинаминопептидазы и щелочной фосфатазы, но не влияя на β-глюкозидазу. Это указывает на то, что УФИ (при уровне радиации в окружающей среде) может быть ключевым фактором, снижающим активность (и время жизни) бесклеточных ферментов (Thomson et al., 2017). Кроме того, тот факт, что эффекты УФИ различаются для разных ферментов, указывает на то, что УФИ может изменить спектр ЕЭП и, таким образом, состав образующегося пула органических веществ. Более того, этот эффект УФИ на бесклеточный EEA может помочь объяснить, почему доля растворенного EEA имеет тенденцию быть ниже в поверхностных водах и почему доля бесклеточного EEA выше зимой и ниже летом.
Это указывает на то, что УФИ (при уровне радиации в окружающей среде) может быть ключевым фактором, снижающим активность (и время жизни) бесклеточных ферментов (Thomson et al., 2017). Кроме того, тот факт, что эффекты УФИ различаются для разных ферментов, указывает на то, что УФИ может изменить спектр ЕЭП и, таким образом, состав образующегося пула органических веществ. Более того, этот эффект УФИ на бесклеточный EEA может помочь объяснить, почему доля растворенного EEA имеет тенденцию быть ниже в поверхностных водах и почему доля бесклеточного EEA выше зимой и ниже летом.
Необходимы дополнительные исследования, чтобы полностью ограничить факторы и механизмы, контролирующие активность и время жизни бесклеточного EEA, поскольку другие факторы, такие как pH, могут иметь значение.Более того, активности, измеренные с использованием добавленных извне субстратов, таких как лейцин-MCA (7-амидо-4-метилкумарин) или MUF (4-метилумбеллиферил) -β-глюкоза в качестве субстратов, отражают активность неизвестного количества / типа различных ферментов. которые расщепляют тот же субстрат (Steen et al., 2015). Кроме того, ферменты с разной первичной и третичной структурой могут гидролизовать один и тот же субстрат, но эти ферменты, вероятно, будут в разной степени восприимчивы к УФ-излучению, тепловой инактивации и т. Д. Тем не менее, на основании имеющихся на данный момент данных и с учетом прогнозируемого Из-за потепления окружающей среды океана и переменного светового режима ультрафиолетового излучения в ближайшем будущем, похоже, могут произойти серьезные изменения в активности бесклеточных EEA и их вкладе в реминерализацию органического вещества.
которые расщепляют тот же субстрат (Steen et al., 2015). Кроме того, ферменты с разной первичной и третичной структурой могут гидролизовать один и тот же субстрат, но эти ферменты, вероятно, будут в разной степени восприимчивы к УФ-излучению, тепловой инактивации и т. Д. Тем не менее, на основании имеющихся на данный момент данных и с учетом прогнозируемого Из-за потепления окружающей среды океана и переменного светового режима ультрафиолетового излучения в ближайшем будущем, похоже, могут произойти серьезные изменения в активности бесклеточных EEA и их вкладе в реминерализацию органического вещества.
Заключение и перспективы на будущее
Теперь ясно, что в любом данном морском месте в любой момент времени EEA без ячеек может быть не менее важным, чем EEA с привязкой к ячейкам. Это сильно влияет на то, как мы смотрим и интерпретируем многие микробные параметры, которые мы измеряем, поскольку это может означать разделение между составом / функцией сообщества и фактическими скоростями гидролиза, которые мы измеряем. Например, это усложняет изучение функциональной избыточности в морской среде, если оно основано на этих внеклеточных ферментах, поскольку изменения в составе микробного сообщества могут происходить в другом временном масштабе по сравнению с изменениями в EEA.Таким образом, важно учитывать необходимость определения бесклеточной фракции EEA (а не только общей фракции), если наша цель — связать EEA с другими микробными параметрами.
Например, это усложняет изучение функциональной избыточности в морской среде, если оно основано на этих внеклеточных ферментах, поскольку изменения в составе микробного сообщества могут происходить в другом временном масштабе по сравнению с изменениями в EEA.Таким образом, важно учитывать необходимость определения бесклеточной фракции EEA (а не только общей фракции), если наша цель — связать EEA с другими микробными параметрами.
В ближайшем будущем развитие технологий (например, методов на основе генов и белков, а также инструментов для определения характеристик органического вещества и т. Д.) Позволит глубже понять функцию и судьбу бесклеточного EEA. Например, многочисленные периплазматические белки были недавно обнаружены с помощью метапротеомики на концентратах бесклеточной фракции морской воды (Xie et al., 2017). Основываясь на этом, эти авторы предположили, что свободные белки, выделяемые микробами, могут иметь важное значение для функционирования экосистемы (Xie et al., 2017).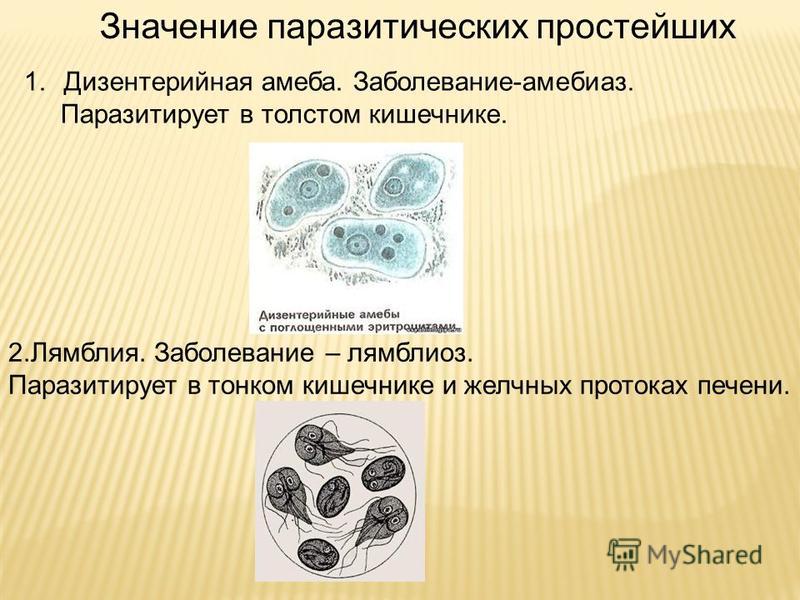 Они признали, что у них нет четкого объяснения высокого присутствия периплазматических белков в «небактериальном» мире. Но, имея в виду то, что обсуждалось в этой статье, возможно, что многие из этих белков так называемого «небактериального мира» Xie et al. (2017) действительно могли быть внеклеточными (здесь описаны как «живые мертвецы») внеклеточными ферментами.Это пример того, как использование новых технологий может помочь сделать новые открытия в этой области. Однако мы должны не только сосредоточиться на разработке новых технологий, но и дополнить их, используя классические измерения скорости. Более точная характеристика пула DOM (например, ЖХ – МС / МС) в сочетании с комбинированным определением скоростей гидролиза и экспрессии генов / белков может открыть новые возможности и открытия в этой области исследований. Кроме того, подобные эксперименты, проводимые в условиях различных антропогенных факторов стресса, также могут помочь выяснить, как роль этих ферментов может измениться в ответ на различные климатические сценарии.
Они признали, что у них нет четкого объяснения высокого присутствия периплазматических белков в «небактериальном» мире. Но, имея в виду то, что обсуждалось в этой статье, возможно, что многие из этих белков так называемого «небактериального мира» Xie et al. (2017) действительно могли быть внеклеточными (здесь описаны как «живые мертвецы») внеклеточными ферментами.Это пример того, как использование новых технологий может помочь сделать новые открытия в этой области. Однако мы должны не только сосредоточиться на разработке новых технологий, но и дополнить их, используя классические измерения скорости. Более точная характеристика пула DOM (например, ЖХ – МС / МС) в сочетании с комбинированным определением скоростей гидролиза и экспрессии генов / белков может открыть новые возможности и открытия в этой области исследований. Кроме того, подобные эксперименты, проводимые в условиях различных антропогенных факторов стресса, также могут помочь выяснить, как роль этих ферментов может измениться в ответ на различные климатические сценарии. Эти исследования, вероятно, подтвердят, что «двигателями» морских биогеохимических циклов являются не только микробные клетки, но и другие процессы, протекающие вне клеток, которые также являются важной частью этого двигателя.
Эти исследования, вероятно, подтвердят, что «двигателями» морских биогеохимических циклов являются не только микробные клетки, но и другие процессы, протекающие вне клеток, которые также являются важной частью этого двигателя.
В заключение, результаты, обсуждаемые в этой статье, отстаивают необходимость выйти за рамки «живых существ» и изучить не только то, как живые клетки / организмы будут реагировать на антропогенные возмущения, но и то, как «живые мертвые» клетки — свободные активные молекулы будут, если мы действительно стремимся полностью ограничить будущее морских биогеохимических циклов.
Взносы авторов
Автор подтверждает, что является единственным соавтором данной работы, и одобрил ее к публикации.
Заявление о конфликте интересов
Автор заявляет, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
FB был поддержан Rutherford Discovery Fellowship (Королевское общество Новой Зеландии).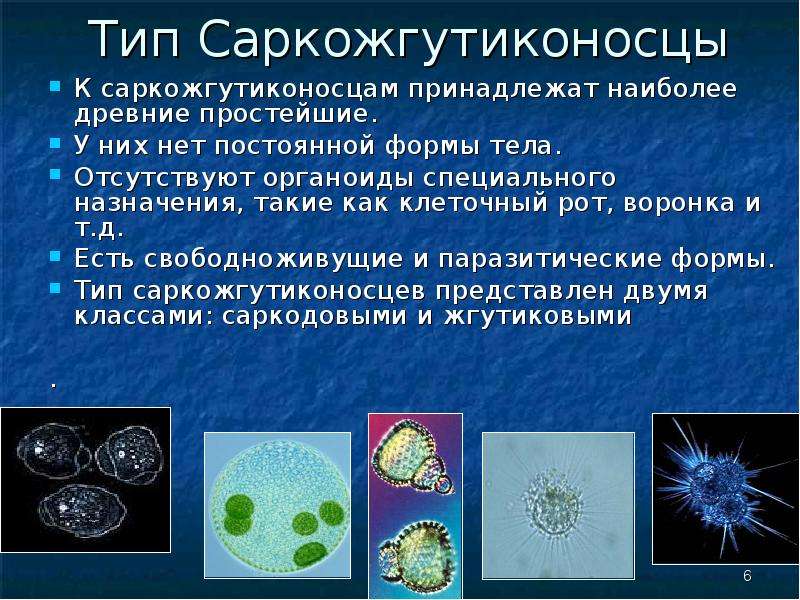 Автор хотел бы поблагодарить рецензентов за поддержку и проницательные комментарии, которые явно помогли улучшить общее качество рукописи.
Автор хотел бы поблагодарить рецензентов за поддержку и проницательные комментарии, которые явно помогли улучшить общее качество рукописи.
Список литературы
Альбертсон, Н. Х., Нистрем, Т., и Кьеллеберг, С. (1990). Экзопротеазная активность двух морских бактерий при голодании. Заявл. Environ. Microbiol. 56, 218–223.
PubMed Аннотация | Google Scholar
Альдеркамп, А. К., Рейссел, М. В., и Болхуис, Х. (2007). Характеристика морских бактерий и активности их ферментных систем, участвующих в деградации запаса глюкана ламинарина в водорослях. FEMS Microbiol. Ecol. 59, 108–117. DOI: 10.1111 / j.1574-6941.2006.00219.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эллисон, С. Д. (2005). Обманщики, диффузия и питательные вещества сдерживают разложение микробными ферментами в пространственно структурированной среде. Ecol. Lett. 8, 626–635. DOI: 10.1111 / j.1461-0248.2005.00756.x
CrossRef Полный текст | Google Scholar
Эллисон, С. Д., Чао, Ю., Фаррара, Дж. Д., Хатоси, С., и Мартини, А. (2012). Мелкомасштабные временные вариации морских внеклеточных ферментов прибрежной южной Калифорнии. Фронт. Microbiol. 3: 301. DOI: 10.3389 / fmicb.2012.00301
Д., Чао, Ю., Фаррара, Дж. Д., Хатоси, С., и Мартини, А. (2012). Мелкомасштабные временные вариации морских внеклеточных ферментов прибрежной южной Калифорнии. Фронт. Microbiol. 3: 301. DOI: 10.3389 / fmicb.2012.00301
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Балтар, Ф., Аристеги, Дж., Газоль, Дж. М., Синтес, Э., Акен, Х. М. В. и Херндл, Г. Дж. (2010). Высокая активность растворенных внеклеточных ферментов в глубинах центральной части Атлантического океана. Aquat. Microb. Ecol. 58, 287–302. DOI: 10.3354 / ame01377
CrossRef Полный текст | Google Scholar
Балтар, Ф., Аристеги, Дж., Газоль, Дж. М., Йококава, Т., и Херндл, Г. Дж. (2013). Бактериальное и архейное происхождение внеклеточной ферментативной активности в глубоких водах Северо-Восточной Атлантики. Microb. Ecol. 65, 277–288. DOI: 10.1007 / s00248-012-0126-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Балтар, Ф. , Аристеги, Дж., Синтес, Э., ван Акен, Х. М., Газоль, Дж. М., и Херндл, Г.Дж. (2009). Внеклеточная ферментативная активность прокариот в отношении производства биомассы и дыхания в мезо- и батипелагических водах (суб) тропической Атлантики. Environ. Microbiol. 11, 1998–2014. DOI: 10.1111 / j.1462-2920.2009.01922.x
, Аристеги, Дж., Синтес, Э., ван Акен, Х. М., Газоль, Дж. М., и Херндл, Г.Дж. (2009). Внеклеточная ферментативная активность прокариот в отношении производства биомассы и дыхания в мезо- и батипелагических водах (суб) тропической Атлантики. Environ. Microbiol. 11, 1998–2014. DOI: 10.1111 / j.1462-2920.2009.01922.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Балтар, Ф., Легран, К., Пиньясси, Дж. (2016). Внеклеточная ферментативная активность вне клеток связана с сезонными изменениями температуры: пример из Балтийского моря. Biogeosciences 13, 2815–2821. DOI: 10.5194 / bg-13-2815-2016
CrossRef Полный текст | Google Scholar
Балтар, Ф., Моран, X.А.Г., и Лёнборг, К. (2017). Источники потепления и органических веществ влияют на долю растворенных в общей активности морских внеклеточных ферментов. Биогеохимия 133, 307–316. DOI: 10.1007 / s10533-017-0334-9
CrossRef Полный текст | Google Scholar
Бочданский, А. Б., Пушкарич, С., и Херндл, Г.Дж. (1995). Влияние выпаса зоопланктона на свободно растворенные ферменты в море. Mar. Ecol. Прог. Сер. 121, 53–63. DOI: 10.3354 / meps121053
Б., Пушкарич, С., и Херндл, Г.Дж. (1995). Влияние выпаса зоопланктона на свободно растворенные ферменты в море. Mar. Ecol. Прог. Сер. 121, 53–63. DOI: 10.3354 / meps121053
CrossRef Полный текст | Google Scholar
Хрост, Р. Дж. (1990). «Микробные эктоферменты в водной среде», в Aquatic Microbial Ecology. Биохимические и молекулярные подходы , ред. Дж. Овербек и Р. Дж. Хрост (Нью-Йорк, штат Нью-Йорк: Springer), 47–78.
PubMed Аннотация | Google Scholar
Хрост, Р.Дж. И Рай Х. (1993). Эктоферментная активность и вторичная продукция бактерий в мезокосмах, обогащенных и обогащенных питательными веществами. Microb. Ecol. 25, 131–150. DOI: 10.1007 / BF00177191
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кускин, Ф., Лоу, Э. К., Темпл, М. Дж., Чжу, Ю., Камерон, Э. А., Пудло, Н. А. и др. (2015). Бактероиды кишечника человека могут использовать дрожжевой маннан эгоистичным образом. Nature 517, 165–169. DOI: 10.1038 / природа13995
DOI: 10.1038 / природа13995
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Daily, Г. (2003). «Что такое экосистемные услуги», в Глобальные экологические вызовы для XXI века: ресурсы, потребление и устойчивые решения, , изд. Д. Э. Лори (Лэнхэм, Мэриленд: Роуман и Литтлфилд), 227–231.
Google Scholar
Д’амброзио, Л., Зервогель, К., Макгрегор, Б., Теске, А., и Арности, К. (2014). Состав и ферментативная функция ассоциированных с частицами и свободноживущих бактерий: сравнение прибрежных и морских районов. ISME J. 8, 2167–2179. DOI: 10.1038 / ismej.2014.67
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дечо, А. В. (1990). Секреции микробных экзополимеров в окружающей среде океана: их роль (и) в пищевых сетях и морских процессах. Oceanogr. Mar. Biol. Аня. Rev. 28, 73–153.
Google Scholar
Дрешер К., Наделл К. Д., Стоун Х. А., Вингрин Н. С. и Басслер Б. Л. (2014). Решения дилеммы общественных благ в бактериальных биопленках. Curr. Биол. 24, 50–55. DOI: 10.1016 / j.cub.2013.10.030
Решения дилеммы общественных благ в бактериальных биопленках. Curr. Биол. 24, 50–55. DOI: 10.1016 / j.cub.2013.10.030
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дюамель, С., Дирман, С. Т., и Карл, Д. М. (2010). Активность и регуляция щелочной фосфатазы в субтропическом круговороте северной части Тихого океана. Лимнол. Oceanogr. 55, 1414–1425. DOI: 10.4319 / lo.2010.55.3.1414
CrossRef Полный текст | Google Scholar
Джанфреда Л. и Скарфи М. Р. (1991). Ферментная стабилизация: современное состояние. Мол. Клетка. Биохим. 100, 97–108. DOI: 10.1007 / BF00234161
CrossRef Полный текст | Google Scholar
Харли К. Д., Рэндалл Хьюз А., Халтгрен К. М., Майнер Б. Г., Сорте К. Дж., Торнбер К. С. и др. (2006). Воздействие изменения климата на прибрежные морские системы. Ecol. Lett. 9, 228–241. DOI: 10.1111 / j.1461-0248.2005.00871.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хоппе, Х. -Г., Арности, К., и Херндл, Г.J. (2002). «Экологическое значение бактериальных ферментов в морской среде», в журнале Enzymes in the Environment: Activity, Ecology, and Applications , ред. Р. Г. Бернс и Р. П. Дик (Нью-Йорк. Нью-Йорк: Марсель Деккер, Инк.), 73–108.
-Г., Арности, К., и Херндл, Г.J. (2002). «Экологическое значение бактериальных ферментов в морской среде», в журнале Enzymes in the Environment: Activity, Ecology, and Applications , ред. Р. Г. Бернс и Р. П. Дик (Нью-Йорк. Нью-Йорк: Марсель Деккер, Инк.), 73–108.
Google Scholar
Хоппе, Х.-Г., и Ульрих, С. (1999). Профили эктоферментов в Индийском океане: явления активности фосфатазы в мезопелагиали. Aquat. Microb. Ecol. 19, 139–148. DOI: 10.3354 / ame019139
CrossRef Полный текст | Google Scholar
Карнер, М., и Расулзадеган, Ф. (1995). Активность внеклеточных ферментов: признаки высокой краткосрочной изменчивости в прибрежной морской экосистеме. Microb. Ecol. 30, 143–156. DOI: 10.1007 / BF00172570
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Lähdesmäki, P., and Piispanen, R. (1992). Энзимология почвы: роль защитных коллоидных систем в сохранении активности экзоферментов в почве. Soil Biol.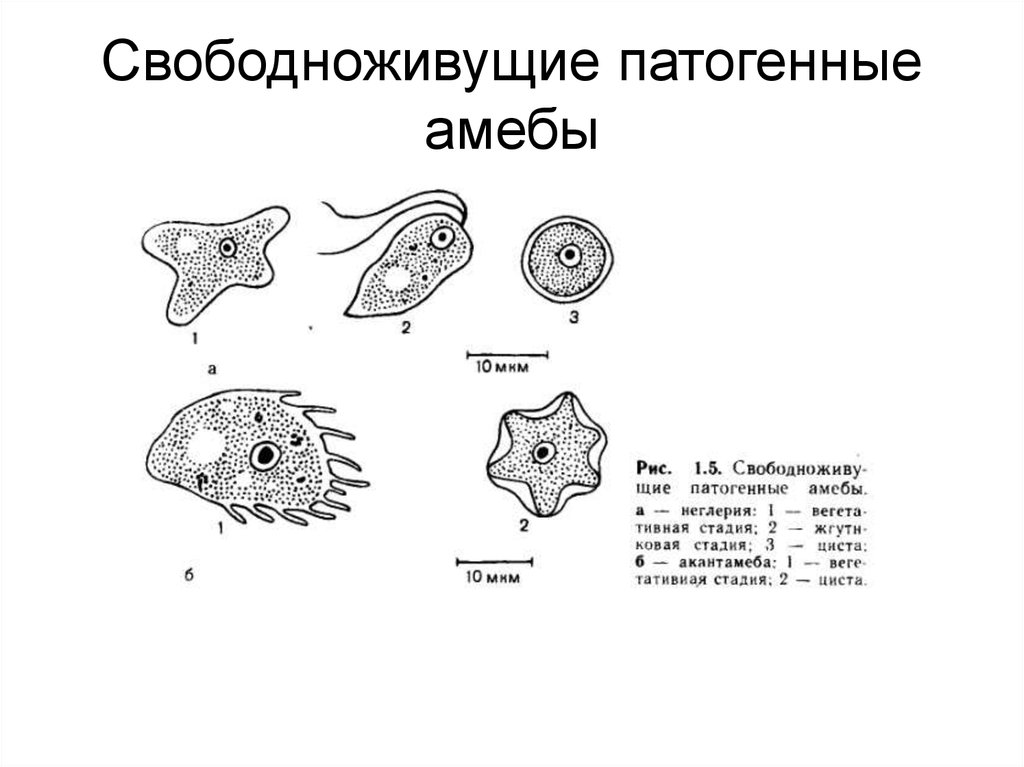 Биохим. 24, 1173–1177. DOI: 10.1016 / 0038-0717 (92)
Биохим. 24, 1173–1177. DOI: 10.1016 / 0038-0717 (92)
-9
CrossRef Полный текст | Google Scholar
Ли, Х., Велдхейс, М. Дж. У. и Пост, А. Ф. (1998). Активность щелочной фосфатазы среди планктонных сообществ в северной части Красного моря. Mar. Ecol. Прог. Сер. 173, 107–115. DOI: 10.3354 / meps173107
CrossRef Полный текст | Google Scholar
Lozzi, I., Calami, L., Fusi, P., Bosetto, M., and Stotzky, G. (2001). Взаимодействие пероксидазы хрена с монтмориллонитом, гомоионным к Na + и Ca2 +: влияние на ферментативную активность и микробную деградацию. Soil Biol.Биохим. 33, 1021–1028. DOI: 10.1016 / S0038-0717 (01) 00005-0
CrossRef Полный текст | Google Scholar
Нагата Т. и Кирчман Д. Л. (1992). Высвобождение высокомолекулярных органических комплексов гетеротрофными морскими жгутиконосцами. Mar. Ecol. Прогр. Сер. 83, 233–240. DOI: 10.3354 / meps083233
CrossRef Полный текст | Google Scholar
Рего, Дж. В., Биллен, Г., Фонтиньи, А., и Сомвилл, М. (1985). Свободная и присоединенная протеолитическая активность в водных средах. Mar. Ecol. Прогр. Сер. 21, 245–249. DOI: 10.3354 / meps021245
В., Биллен, Г., Фонтиньи, А., и Сомвилл, М. (1985). Свободная и присоединенная протеолитическая активность в водных средах. Mar. Ecol. Прогр. Сер. 21, 245–249. DOI: 10.3354 / meps021245
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рейнтьес, Г., Арности, К., Фукс, Б. М., и Аманн, Р. (2017). Альтернативный механизм поглощения полисахаридов морскими бактериями. ISME J. 11, 1640–1650. DOI: 10.1038 / ismej.2017.26
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стин, А. Д., и Арности, К. (2011). Длительное время жизни бета-глюкозидазы, лейцинаминопептидазы и фосфатазы в арктической морской воде. Mar. Chem. 123, 127–132. DOI: 10.1016 / j.marchem.2010.10.006
CrossRef Полный текст | Google Scholar
Стин, А. Д., Вазин, Дж. П., Хаген, С. М., Маллиган, К. Х., Вильгельм, С. В. (2015). Субстратная специфичность водных внеклеточных пептидаз оценивается с помощью анализов конкурентного ингибирования с использованием синтетических субстратов. Aquat. Microb. Ecol. 75, 271–281. DOI: 10.3354 / ame01755
Aquat. Microb. Ecol. 75, 271–281. DOI: 10.3354 / ame01755
CrossRef Полный текст | Google Scholar
Томсон, Б., Хепберн, К. Д., Ламар, М., и Балтар, Ф. (2017). Температура и ультрафиолетовое излучение влияют на активность бесклеточных ферментов морских организмов. Biogeosciences 14, 3971–3977. DOI: 10.5194 / bg-14-3971-2017
CrossRef Полный текст | Google Scholar
Трэвинг, С. Дж., Тайгесен, У. Х., Риман, Л., Стедмон, К. А. (2015). Модель внеклеточных ферментов в свободноживущих микробах: какая стратегия окупается? Заявл. Environ. Microbiol. 81, 7385–7393. DOI: 10.1128 / AEM.02070-15
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Веттер, Ю.А. и Деминг Дж. У. (1999). Скорость роста изолятов морских бактерий на твердых органических субстратах, солюбилизированных свободно высвобождаемыми внеклеточными ферментами. Microb. Ecol. 37, 86–94. DOI: 10.1007 / s002489
3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Веттер Ю. А., Деминг Дж. У., Джумарс П. А. и Кригер-Брокетт Б. Б. (1998). Прогностическая модель поиска пищи бактериями с помощью свободно высвобождаемых внеклеточных ферментов. Microb. Ecol. 36, 75–92. DOI: 10.1007 / s002489
А., Деминг Дж. У., Джумарс П. А. и Кригер-Брокетт Б. Б. (1998). Прогностическая модель поиска пищи бактериями с помощью свободно высвобождаемых внеклеточных ферментов. Microb. Ecol. 36, 75–92. DOI: 10.1007 / s002489
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вайс, М., Абеле, У., Векессер, Дж., Велте, В., Шильц, Э. и Шульц, Г. Э. (1991). Молекулярная архитектура и электростатические свойства бактериального порина. Наука 254, 1627–1630. DOI: 10.1126 / science.1721242
CrossRef Полный текст | Google Scholar
Xie, Z.-X., Chen, F., Zhang, S.-F., Wang, M.-H., Zhang, H., Kong, L.-F., et al.(2017). Метапротеомика концентратов морских вирусов выявляет ключевые вирусные популяции и обильные периплазматические белки в олиготрофном глубинном максимуме хлорофилла в Южно-Китайском море. Environ. Microbiol. DOI: 10.1111 / 1462-2920.13937 [Epub перед печатью].
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ziervogel, K. , Karlsson, E., and Arnosti, C. (2007). Поверхностные ассоциации ферментов и органического вещества: последствия для гидролитической активности и реминерализации органического вещества в морских системах. Mar. Chem. 104, 241–252. DOI: 10.1016 / j.marchem.2006.12.001
, Karlsson, E., and Arnosti, C. (2007). Поверхностные ассоциации ферментов и органического вещества: последствия для гидролитической активности и реминерализации органического вещества в морских системах. Mar. Chem. 104, 241–252. DOI: 10.1016 / j.marchem.2006.12.001
CrossRef Полный текст | Google Scholar
Ziervogel, K., Steen, A. D., and Arnosti, C. (2010). Изменения в спектре и скорости активности внеклеточных ферментов в морской воде после образования агрегатов. Биогеонауки 7, 1007–1015. DOI: 10.5194 / bg-7-1007-2010
CrossRef Полный текст | Google Scholar
.
