
Доказательства эволюции — Википедия
Материал из Википедии — свободной энциклопедии
Ископаемый археоптерикс, обнаруженный вскоре после публикации «Происхождения видов». Строение черепа археоптерикса (наличие зубов), длинный хвостовой отдел позвоночника из 20 позвонков, пальцы с развитыми фалангами на передних конечностях, наличие брюшных рёбер, отсутствие клюва и ряд других признаков, характерных для нептичьих теропод. В то же время общая форма тела, сросшиеся ключицы, трёхпалые передние конечности и характер оперения, включающего сложно устроенные маховые перья, характерны для птиц. Тем самым археоптерикс занимает по морфологии промежуточное положение между традиционно помещаемыми в класс рептилий динозаврами и птицами (хотя не все авторы согласны с традиционной систематикой), то есть является переходной формой[~ 1]Доказа́тельства эволю́ции — научные данные и концепции, подтверждающие происхождение всех живых существ на Земле от общего предка[~ 2]. Благодаря этим доказательствам основы эволюционного учения получили признание в научном сообществе, а ведущей системой представлений о процессах видообразования стала синтетическая теория эволюции
Эволюционные процессы наблюдаются как в естественных, так и лабораторных условиях. Известны случаи образования новых
ru.wikipedia.org
Макроэволюция, её доказательства. Направления эволюции.
МАКРОЭВОЛЮЦИЯ, ЕЕ ДОКАЗАТЕЛЬСТВА
Цель: познакомить с макроэволюцией и ее доказательствами — палеонтологическими, эмбриологическими и другими.
Оборудование: рис учебника «Сходство начальных стадий эмбрионального развития позвоночных», рис., образцы, демонстрирующие формы сохранности ископаемых растений и животных.
Ход урокаI. Проверка знаний.
1. Тестирование.
Заполните пробелы в нижеприведенных фразах.
1) Микроэволюцией называют эволюционные изменения, протекающие на … уровне
2) Различают две основные формы видообразования: … и…
3) Репродуктивная изоляция связана с прекращением обмена … между двумя …
4) Генетическая дивергенция означает … генов
5) Форма видообразования, связанная с расширением ареала исходного вида, называется…
6) Современные виды ландыша возникли в результате ….
7) Зарождение в рамках исходной популяции новой формы называется …
8) Симпатрическое видообразование имеет две формы: … и …
9) Пять видов синиц образовались в результате … видообразования, а полиплоидный картофель — в результате … видообразования.
10) Образование новых видов в результате хромосомных перестроек может происходить в популяциях…
2. Письменный ответ. Задание.
Из приведенного перечня выберите факторы, способствующие видообразованию, и обоснуйте свой выбор: малочисленность вида, многочисленность вида, сравнительно однообразные условия существования, разнообразные условия существования, маленький ареал, большой ареал, острая борьба за существование, относительно слабая борьба за существование, изоляция, дивергенция.
3. Письменный ответ. Задание.
Запишите последовательность событий при географическом видообразовании.
1) Расселение на новые территории за пределами ареала.
2) Отбор в новых условиях среды.
3) Географическая изоляция между популяциями.
4) Образование новых видов.
5) Возникновение подвидов.
6) Биологическая изоляция.
4. Ответы по вопросам в конце § 60, № 2, 3, и выполнение задания, отмеченного знаком
II. Изучение нового материала.
1. Макроэволюция.
Этим понятием обозначают происхождение надвидовых таксонов (родов, отрядов, классов, типов, отделов).
В общем смысле макроэволюцией можно назвать развитие жизни на Земле в целом, включая и ее происхождение. Макроэволюционным событием считается также возникновение человека. Между микро- и макроэволюцией нельзя провести резкую границу, потому что процесс микроэволюции, вызвавший дивергенцию, продолжается без какого-либо перерыва и на макро-эволюционном уровне внутри вновь возникших форм. Отсутствие принципиальных различий в протекании микро- и макроэволюционного процессов позволяет рассматривать их как две стороны единого эволюционного процесса.
2. Палеонтологические доказательства макроэволюции.
Палеонтологические данные позволяют узнать о растительном и животном мире прошлого, реконструировать облик вымерших организмов, обнаружить связь между древнейшими и современными представителями флоры и фауны. Сопоставление ископаемых остатков из земных пластов разных геологических эпох позволяет установить последовательность возникновения и развития разных групп организмов. Например, в самых древних пластах находят остатки представителей типов беспозвоночных животных, а в более поздних пластах — уже и остатки хордовых. В еще более молодых геологических пластах содержатся остатки животных и растений, относящихся к видам, похожим на современные.
Палеонтологами были обнаружены формы организмов, сочетающие признаки более древних и более молодых групп. Такие ископаемые переходные формы служат доказательством эволюции, поскольку свидетельствуют об исторической связи разных групп организмов. Например, ископаемая первоптица археоптерикс — связующее звено между рептилиями и птицами. Археоптерикс имеет, как и рептилии, длинный хвост, несросшиеся позвонки, развитые зубы; как птица, он покрыт перьями, имеет крылья, частично пневматичные кости.
Другими примерами переходных форм являются кистеперые рыбы, связывающие рыб с вышедшими на сушу земноводными, семенные папоротники переходная форма между папоротниковидными и голосеменными.
Доказательством эволюции служат и палеонтологические ряды. Палеонтологами были найдены остатки ранее живших видов, которые связаны между собой родством, то есть свидетельствовали о происхождении одних видов от других.
Русский ученый В. О. Ковалевский, исследуя историю развития лошади, показал, что современные однопалые животные происходят от мелких пятипалых всеядных предков, живших в лесах 60-70 млн лет назад. Изменение климата Земли повлекло за собой сокращение площадей лесов и увеличение площадей степей. Животные оказались в новых условиях. Преобразование конечностей — уменьшение числа пальцев от пяти до одного — возникло в связи с необходимостью защиты от хищников и передвижения на большие расстояния в поисках пищи. К настоящему времени установлены палеонтологические ряды в эволюции слонов, носорогов, китов.
3. Эмбриологические доказательства.
В пользу эволюции органического мира говорят данные эмбриологии. Эмбриологами было обнаружено и изучено сходство начальных стадий эмбрионального развития животных. Все многоклеточные животные развиваются из одной оплодотворенной яйцеклетки, В процессе индивидуального развития они проходят стадии дробления, бластулы, гаструлы, образования трехслойного зародыша, формирования органов из зародышевых листков. Сходство зародышевого развития животных свидетельствует о единстве их происхождения.
О сходстве эмбриональных стадий у всех позвоночных свидетельствует закладка жаберных дуг, одинаковая форма тела, наличие хвоста, зачатков конечностей. Во многом аналогично на этих стадиях внутреннее строение зародышей: сначала закладывается хорда, кровеносная система с одним кругом кровообращения (как у рыб), одинаковое строение почек. По мере развития сходство между зародышами уменьшается. Все более четко начинают проявляться черты организации тех классов, к которым животные принадлежат. У наземных животных зарастают жаберные карманы, у зародыша человека особенно сильно развивается головной отдел, включающий мозг, формируются пятипалые конечности.
По ходу эмбрионального развития последовательно идет расхождение признаков зародышей, приобретающих черты, характеризующие класс, отряд, род, вид, к которому они принадлежат. Эта закономерность в развитии зародышей указывает на их родство, происхождение от одного ствола, который в ходе эволюции распался на множество ветвей.
4. Сравнительно-анатомические доказательства.
Для изучения сравнительно-анатомических доказательств эволюции необходимо обратиться к понятиям «гомология» «рудименты», «атавизмы». Гомологичными называются органы имеющие одинаковый план строения, развивающиеся из сходных зачатков и одинаково расположенные, но выполняющие разные функции. Гомология указывает на общность происхождения обладающих ею организмов, различия в строении гомологичных органов — результат дивергенции.
Главный признак аналогии — сходство функций вне связи со строением и происхождением. Аналогичные органы — результат конвергенции. Наличие аналогичных органов не свидетельствует о тесном родстве обладающих ими организмов.
Рудименты недоразвитые органы, утратившие в ходе эволюции свои биологические функции.
У некоторых особей рудименты могут развиваться в органы нормальных размеров. Такой возврат к строению органа предковых форм называют атавизмом. Изучение атавистических признаков позволяет воссоздать строение гомологичных органов у предков.
III. Закрепление.
Вспомните и перечислите признаки сходства всех классов позвоночных животных. Чем они объясняются?
Рассмотрите рисунок 153-154; 156-157; 158-159 в учебнике В.К.Шумного. Какие признаки в филогенетическом ряду копытных наиболее сильно подверглись действию естественного отбора? В чем значение палеонтологических исследований.
Какой вклад в эволюционную теорию вносят эмбриологические исследования?
Домашнее задание: § 54,55,56,57.
СИСТЕМА РАСТЕНИЙ И ЖИВОТНЫХ — ОТОБРАЖЕНИЕ ЭВОЛЮЦИИ
Цели: продолжить знакомство с доказательствами макроэволюции на примере современной системы растений и животных; сформировать представление о принципах современной классификации.
Ход урока
I. Проверка знаний.
1. Тестирование.
В нижеприведенных предложениях заполните пробелы.
1) Макроэволюция — -… эволюция
2) Макроэволюция, как и микроэволюция, носит … характер
3) Палеонтология изучает … организмов
4) … находки позволяют реконструировать внешний облик организмов
5) Археоптерикса с пресмыкающимися сближает длинный …
6) Ряды видов, последовательно сменяющихся друг друга,
называются…
7) Ископаемые формы, сочетающие признаки … и более … групп высокого систематического ранга, называются …
8) Все многоклеточные животные развиваются из …….. яйцеклетки
9) Сходство зародышевого развития животных свидетельствует о … происхождения
10) Крыло птицы и бабочки являются примером … органов
2. Письменный ответ по карточке. Задание.
Заполните таблицу «Сравнительная характеристика этапов эволюционного процесса».
|
Этап |
В каких группах организмов осуществляется |
Материал для эволюционного процесса |
Главный эволюционный фактор |
Результаты |
|
Микроэволюция |
|
|
|
|
|
Макроэволюция |
|
|
|
|
3.Письменный ответ. Задание.
1) Внимательно прочтите текст и рассмотрите рисунок (череп саблезубого тигра).
Палеонтологу известно, что размеры тела и длина клыков саблезубых тигров постепенно увеличивались к периоду их вымирания. Учитывая это, можно ли предположить, что:
а) в это время происходило потепление климата и саблезубые тигры широко расселились на Земле;
б) условия существования этих животных ограничивались своеобразными узкими пределами;
в) крупные тигры с длинными клыками уничтожили всех более мелких;
г) одностороннее развитие некоторых органов указывает на крайнюю специализацию, результатом чего может быть вымирание;
д) количество наследственных изменений у саблезубых тигров в сторону увеличения длины клыков постоянно росло?
2) Объясните, почему одни предположения правильны, а другие — нет.
4. Ответы по вопросам в конце §54,55,56,57 Ш.
II. Изучение нового материала.
1. Актуализация знаний учащихся.
Кем разработаны основы современной классификации организмов?
Перечислите систематические группы животных и растений, известных вам из курса зоологии, ботаники.
2. Систематические группы.
Данные систематики используют для доказательства эволюции, так как они устанавливают родство между таксонами.
В современной систематике организмы распределяются по ряду систематических категорий: вид, род, семейство, отряд (порядок для растений), класс, тип (отдел для растений) и др.
Каждая высшая систематическая категория, начиная от рода, объединяет группы, стоящие по рангу ниже и имеющие общего предка. Род объединяет виды, произошедшие от одного предка и оказавшиеся в результате борьбы за существование и естественного отбора способными существовать и успешно размножаться в различных географических и экологических условиях.
Признаки (критерии), на основании которых близкие виды объединяются в роды, хорошо видны на примере дарвиновских вьюрков. На Галапагосских островах вьюрки представлены тремя родами: земляные, древесные и славковые. Земляные вьюрки гнездятся в засушливой зоне и кормятся большей частью на открытых местах, древесные гнездятся в засушливой зоне и питаются на деревьях; славковые занимают разные местообитания.
Главный признак, по которому различают виды вьюрков, -строение клюва, тесно связанное с особенностями питания. На цветках кактуса питается кактусовый земляной вьюрок, у которого длинный клюв и расщепленный язык. У большого земляного вьюрка толстый, массивный клюв, отлично справляющийся с крупными семенами. Дятловый, древесный, вьюрок получил свое название за прямой, как у дятла, клюв, которым он долбит древесную кору, ползая вверх и вниз по стволу. Отсутствие длинного языка он восполняет кактусовой иглой или веточкой, удерживая ее в клюве и выковыривая насекомых из отверстия в коре, которое выдолбил. Все виды дарвиновских вьюрков не скрещиваются; некоторые виды образуют подвиды — — значит, видообразование продолжается. Все виды вьюрков произошли от одного исходного вида.
3. Принципы современной классификации.
Основы научной систематики заложил еще в XVIIIв. К. Линней. Принципы классификации Линнея действуют и ныне.
В любом видовом названии присутствует имя рода. Род объединяет наиболее близкие виды организмов. Выделяют, например, такие роды, как кошки, лошади, дубы и т. д. Первоначально для видового названия к имени рода прибавлялись фразы, которыми описывались характерные видовые признаки. Например, дуб красный назывался «дуб с листьями, имеющими глубокие прорези, оканчивающимися волосоподобными зубчиками». Позднее, после работ Линнея, укоренилось двойное, или биноминальное, название видов. Первое слово представляет собой имя рода, второе название вида. Например, Дуб красный. Собака домашняя.
Современная система классификации учитывает признаки родства видов как с ныне живущими, так и с уже вымершими. Каждая таксономическая категория соответствует группе организмов, которые имеют общего предка. Такая система классификации отражает естественную общность организмов и поэтому называется естественной. Естественные классификации позволяют предсказывать наличие у организмов тех или иных свойств в зависимости от их положения в системе.
Взаимоотношения между основными группами современных организмов, сложившиеся в процессе эволюции, представляют собой подобие ветвей могучего дерева. Родословное древо в целом и его ветвление отчетливо выявляют общий характер макроэволюции: развитие живых существ от менее сложных к более сложным, дивергентный и приспособительный характер эволюции.
III. Закрепление.
Фронтальная письменная работа. Задание.
Заполните таблицу «Развитие кровеносной системы у позвоночных».
|
Особенности строения органов кровообращения |
Рыбы |
Земноводные |
Пресмыкающиеся |
Птицы |
Млекопитающие |
|
Число сердечных камер |
|
|
|
|
|
|
Число кругов кровообращения |
|
|
|
|
|
Домашнее задание: § 75.
ГЛАВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ ОРГАНИЧЕСКОГО МИРА
Цели: сформировать понятие о главных направлениях эволюции, показать соотношение путей эволюции; познакомить с типами эволюционных изменений.
Ход урока
I. Изучение нового материала.
1.Типы эволюционных изменений.
Параллелизм это процесс эволюционного развития в сходном направлении двух или нескольких первоначально дивергировавших групп. Например, палеонтологи очень часто обнаруживают синхронный параллелизм, то есть независимое приобретение сходных черт родственными, но живущими в разное время организмами. Примером может служить развитие саблезубости у представителей разных подсемейств кошачьих.
Конвергенция — процесс эволюционного развития двух и более неродственных групп в сходном направлении. Конвергенция обусловлена одинаковой средой обитания, в которую попадают неродственные организмы. Классическим примером конвергентного развития является возникновение сходных форм тела у акулы, ихтиозавра, дельфина. При конвергентном развитии сходство между неродственными организмами бывает всегда только внешним.
Дивергенция представляет собой независимое образование различных признаков у родственных организмов. В основе дивергенции лежит экологическая дифференциация вида на самостоятельные ветви. При дивергенции сходство между организмами объясняется общностью их происхождения, а различия приспособлением к разным условиям среды.
Примером дивергенции форм является возникновение разнообразных по морфофизиологическим особенностям вьюрков от одного или немногих предковых видов на Галапагосских островах. Расхождение внутривидовых форм и видов по разным местообитаниям определяется конкуренцией в борьбе за одинаковые условия, выход из которых и заключается в расселении по разным экологическим нишам.
2. Главные направления эволюции.
Развитие живой природы шло от простого к сложному и имело прогрессивный характер. Наряду с этим происходило приспособление видов к конкретным условиям жизни, осуществлялась их специализация. Биологический прогресс достигается различными путями. А. Н. Северцов назвал их главными направлениями эволюционного прогресса. В настоящее время выделяют следующие пути биологического прогресса: ароморфозы, идиоадаптации и дегенерации.
Ароморфоз — главный путь достижения биологического прогресса. Ароморфоз — возникновение в ходе эволюции признаков, повышающих уровень организации живых существ. Ароморфозы формируются на основе наследственной изменчивости и естественного отбора и являются приспособлениями широкого значения. Они дают преимущества в борьбе за существование и открывают возможности освоения новой, прежде недоступной среды обитания. Путем ароморфоза возникают в процессе эволюции крупные систематические группы рангом выше семейства.
К ароморфозам у животных можно отнести появление живорождения, способности к поддержанию постоянной температуры тела, возникновение замкнутой системы кровообращения, а у растений — появление цветка, сосудистой системы, способности к поддержанию и регулированию газообмена в листьях.
Ароморфозам А. Н. Северцов противопоставлял идиоадаптации — частные приспособления видов, позволяющие освоить специфические условия среды. В отличие от ароморфозов идиоадаптации открывают перед организмами возможность биологического прогресса без повышения уровня биологической организации. Например, благодаря формированию различных идиоадаптации млекопитающие смогли распространиться не только в различных географических зонах (от тропиков до ледяных пустынь), но и освоить самые разнообразные условия среды (на поверхности суши, в воде, почве, частично в воздухе). Это существенно снизило конкуренцию между видами за пищу, места обитания, причем уровень организации остался тем же.
Обычно мелкие систематические группы виды, роды, семейства в процессе эволюции возникают путем идиоадаптации.
Идиоадаптация так же, как и ароморфоз, приводит к увеличению численности вида, расширению ареала, ускорению видообразования, то есть к биологическому прогрессу.
Типичные идиоадаптации у животных особенности строения конечностей (например у крота, копытных, ластоногих), особенности клюва (у хищных птиц, куликов, попугаев), приспособления придонных рыб (у скатов, камбаловых), покровительственная окраска у насекомых. Примерами идиоадаптации у растений могут служить многообразные приспособления к опылению, распространению плодов и семян.
Общая дегенерация.
Ч. Дарвин отмечал, что способность организмов выживать в борьбе за существование вовсе не обязательно должна быть связана с более высокой организацией. Какие преимущества, например, могли бы получить инфузории или земляной червь из более высокой организации, чем они имеют? Условия жизни этих организмов относительно постоянны, они хорошо приспособлены каждый к своей среде. Вот почему естественный отбор не совершенствовал их в сторону прогрессивного усложнения. Более того, при упрощении условий среды организмы утрачивают часть признаков развиваются по пути общей дегенерации, ведущей к упрощению организации. Это соответствует дарвиновскому учению, согласно которому эволюция заключается в выживании наиболее приспособленных, а не более высокоорганизованных.
Дивергенция часто связана с переходом к пещерному, сидячему, паразитическому образу жизни. Упрощения организации обычно сопровождается возникновением различных приспособлений к специфическим условиям жизни.
Особенно наглядно это прослеживается на паразитических организмах. У свиного цепня, лентеца широкого и др. ленточных червей паразитов человека и животных, нет кишечника, слабо развита нервная система. Однако они отличаются огромной плодовитостью благодаря сильно развитым органам размножения, обладают присосками и крючками, при помощи которых держатся на стенках кишечника своего хозяина.
Переход некоторых растений к паразитизму сопровождался снижением активности аппарата фотосинтеза, редукцией листьев до чешуи, преобразованием корней в присоски. Одновременно развилась сложная система приспособлений к хозяину (химическая сигнализация при поиске растения-хозяина, химический механизм внедрения в ткани хозяина и др.).
Общая дегенерация не исключает процветания вида. Многие группы паразитов процветают, хотя организация их претерпевает значительное упрощение. Следовательно, и дивергенция может приводить к биологическому прогрессу.
Многие современные виды охвачены биологическим прогрессом. Например, заяц-русак. Сейчас известно около 20 его видов.
В природе наблюдается и биологический регресс. Он характеризуется чертами, противоположными биологическому прогрессу: уменьшением численности, сужением ареала, уменьшением числа видов, популяций. В итоге регресс часто ведет к вымиранию вида. В процессе эволюции исчезли древние папоротникообразные, многие группы растений и животных.
Деятельность человека является мощным фактором биологического прогресса одних видов, нередко вредных для него, и биологического регресса других, нужных и полезных ему. Например, появление насекомых, устойчивых к ядохимикатам, болезнетворных микробов, устойчивых к действию лекарств, бурное развитие сине-зеленых водорослей в сточных водах. При посевах человек вторгается в живую природу, уничтожает на больших площадях множество диких популяций, заменяя их искусственными. Усиленное истребление человеком многих видов ведет к их биологическому регрессу, который грозит им вымиранием.
3. Соотношение путей эволюции.
Из всех рассмотренных путей достижения биологического прогресса наиболее редки ароморфозы, поднимающие ту или иную систематическую группу на качественно новый, более высокий уровень развития.
Ароморфозы можно рассматривать как переломные пункты развития жизни. Для групп, подвергнувшихся соответствующим морфофизиологическим преобразованиям, открываются новые возможности в освоении внешней среды.
За каждым ароморфозом следует множество идиоадаптаций, которые обеспечивают более полное использование всех имеющихся ресурсов и освоение новых местообитаний.
Есть систематические группы, которые развиваются по пути общей дегенерации. Этот путь эволюции может осуществляться при попадании организмов в постоянную, сравнительно однородную среду, например при паразитическом образе жизни.
II. Практическая работа. Задание.
Используя учебники ботаники или зоологии, выполни в тетрадях предложенные задания, указанные в карточках.
Карточка 1.
Заполните таблицу «Главные направления органической эволюции».
|
Направление эволюции |
Примеры |
|
Биологический прогресс процветание, определенное относительно высокими темпами эволюции, при которых приспособленность организмов возрастает. Объективным показателем биологического прогресса является увеличение численности, ведущее к расселению и расширению ареала. (И. Шмалъгаузен.)
Биологический регресс снижение приспособленности организма. Организм отстает в темпах эволюции от изменений во внешней среде и в особенности от темпов эволюции и распространения экологически близких форм. Численность вида уменьшается. Вид или группа видов идет навстречу вымиранию. (И. Шмалъгаузен.) |
1 2 3
1 2 3
|
Карточка 2.
Заполните таблицу «Сравнительная характеристика главных направлений эволюции органического мира».
|
Главные направления эволюции |
Краткая характеристика |
Примеры |
|
1 2 3 |
|
1 2 3 1 2 3 1 2 3 |
|
|
||
|
|
Карточка 3.
Используя знания из курса зоологии, приведите убедительные доводы против теории Ж.-Б. Ламарка о «врожденном стремлении к прогрессу».
Карточка 4.
Приведите примеры, подтверждающие, что прогресс в общей организации всегда оказывался связанным с регрессом (недоразвитием отдельных органов или частей тела).
Карточка 5.
Почему повышение уровня организации — главный, но не единственный путь эволюции? Ответ подтвердите примерами.
Домашнее задание: § 73,74 повторить § 52-69.
mplmurmansk.ru
Макроэволюция, ее доказательства. Виды древних растений и животных
- ГДЗ
-
1 Класс
- Математика
- Английский язык
- Русский язык
- Немецкий язык
- Информатика
- Природоведение
- Основы здоровья
- Музыка
- Литература
- Окружающий мир
- Человек и мир
-
2 Класс
- Математика
- Английский язык
- Русский язык
- Немецкий язык
- Белорусский язык
- Украинский язык
- Информатика
- Природоведение
- Основы здоровья
- Музыка
- Литература
- Окружающий мир
- Человек и мир
- Технология
-
3 Класс
- Математика
- Английский язык
- Русский язык
- Немецкий язык
- Белорусский язык
- Украинский язык
- Информатика
- Музыка
- Литература
- Окружающий мир
- Человек и мир
- Испанский язык
-
4 Класс
- Математика
- Английский язык
- Русский язык
- Немецкий язык
- Белорусский язык
- Украинский язык
- Информатика
- Основы здоровья
resheba.me
Доказательства эволюции органического мира
Об эволюционном развитии органического мира свидетельствуют многие факты, накопленные разными науками о природе, в первую очередь палеонтологией, морфологией и анатомией, цитологией, эмбриологией, биогеографией и др.
Рассмотрим некоторые из этих доказательств.
Цитологические доказательства
Цитология — наука о строении и функциях клетки. Она дала доказательства единого клеточного строения всех организмов на земле — от одноклеточных растений и животных до многоклеточных организмов. Это свидетельствует об общности происхождения органического мира.
Морфологические доказательства
Морфология и анатомия — две близко связанные науки, изучающие внешнее и внутреннее строение организмов (растений и животных). Было установлено определенное сходство строения разных групп организмов и выявлены переходные формы между ними.
Большую роль для понимания процессов и направлений эволюции сыграло обнаружение рудиментов и атавизмов.
Атавизмы — возврат к признакам или появление органов, которые существовали у отдаленных предков, но были полностью утрачены в процессе эволюции. Например, появление хвоста, нескольких сосков на груди и животе или густого волосяного покрова у человека. Случаи появления атавизмов свидетельствуют о том, что гены, кодирующие их образование, не исчезли из генома, а находятся в нем в заблокированном состоянии. Если этот блок по каким-то причинам не срабатывает, то появляются атавизмы.
Рудиментами называются органы, имеющиеся у организмов, но давно утратившие свое исходное значение и поэтому находящиеся в недоразвитом состоянии. Эти органы были в активном состоянии у предков, но в связи с изменением условий жизни перестали быть необходимыми у потомков. Они закладываются на стадии эмбриогенеза, но не получают полного развития у взрослых форм растений и животных. Примерами могут быть ушные мышцы, отросток слепой кишки (аппендикс) и «третье веко»у человека (всего у человека более 90 рудиментарных органов). Рудиментами являются неразвитые кости задних конечностей у китообразных, глаза у пещерных и роющих животных (слепышей, кротов и др.) и т. п. В отличие от атавизмов, рудиментарные органы всегда присутствуют у организмов.
Изучение жизненных форм (или биоморф) растений и животных убедительно доказало возможность перехода от одних из них к другим. Например, у близких видов растений древесные формы могут заменяться на кустарниковые или стелющиеся в зависимости от условий обитания.
Палеонтологические доказательства
Палеонтология — наука, изучающая ископаемые остатки разных групп организмов или их отпечатки, следы и т. п., а также целые палеоценозы территорий. Изучение этих остатков обнаружило факты безусловного изменения растительного и животного мира во времени — в разных геологических пластах, различающихся по времени образования, присутствуют неодинаковые формы вымерших организмов. Показано, что и сами природные ландшафты целых регионов сильно изменялись во времени: моря наступали на сушу и отступали на обширных территориях, равнины сменялись горами, леса — степями или наоборот, и т. п. Ученым удалось также найти большое число переходных форм между ныне живущими и ископаемыми организмами (например, археоптерикс, сочетающий признаки птиц и рептилий; зверозубые ящеры, имеющие признаки млекопитающих; группа семенных папоротников, давшая начало голосеменным и т. д.).
Палеонтологам удалось установить ряд филогенетических рядов некоторых животных (например, прослежена эволюция лошади от мелкого по размерам эогиппуса с четырехпалыми передними и трехпалыми задними конечностями до современной лошади с однопалыми конечностями).
Эмбриологические доказательства
Эмбриология — наука о зародышевом (или эмбриональном) развитии организмов. Установлено, что все многоклеточные организмы, способные к половому размножению, развиваются из одного оплодотворенного яйца (яйцеклетки). При этом К. Бэром в 1825-1828 гг. было обнаружено большое сходство развития зародышей (эмбрионов) у животных, относящихся к одному типу, описанное им как закон зародышевого сходства. Дальнейшие исследования подтвердили справедливость наблюдений К.Бэра. Сходство развития зародышей у животных разных систематических групп безусловно свидетельствует об общности их происхождения. При этом сначала проявляются признаки более древних предков (у хордовых — это зачатки хорды, наличие жаберных щелей), а затем — черты более поздних прародителей. По мере развития зародыша он приобретает все более заметные черты строения, характерные для класса, отряда, рода и, наконец, вида, к которому он принадлежит. Такое расхождение признаков зародышей по мере их развития носит название эмбриональной дивергенции.
Обобщая эти данные, немецкие ученые Ф. Мюллер и Э. Геккель (1864-1866 гг.) сформулировали биогенетический закон: индивидуальное развитие (онтогенез) всякого организма есть краткое и сжатое повторение пути исторического развития (филогенеза) того вида, к которому данный организм относится.
Возврат к признакам предков получил в теории эволюции название рекапитуляции. Данный закон был развит и уточнен крупным российским (советским) ученым академиком А. Н. Северцовым, показавшим, что в индивидуальном развитии происходит повторение форм развития не взрослых предков, а только их эмбриональных стадий. Поэтому в целом соотношение между онтогенезом и филогенезом носит значительно более сложный характер, чем это постулировали Ф. Мюллер и Э. Геккель. Филогенез следует рассматривать как исторический ряд отобранных в процессе естественного отбора онтогенезов.
Биогенетический закон применим не только к хордовым, но и к другим группам животных и к растениям. Например, у многих насекомых личиночные стадии похожи на червей (гусеницы бабочек, личинки мух и т. д.), что свидетельствует о возможной близости предков этих животных. У ряда мохообразных (например, кукушкин лен) спора при прорастании образует нитчатое образование — протонему, похожую на нитчатые водоросли. В целом биогенетический закон сыграл огромную роль в выяснении филогенетических связей между разными группами организмов.
Доказательства биогеографии
Биогеография — наука о закономерностях распространения растений, животных, грибов, бактерий на нашей планете. Она изучает пути и последствия распределения в природе и миграций организмов на формирование современных флор и фаун регионов. В путях расселения могут возникать различные препятствия или же новые связи между регионами (островами, материками и т. п.). Это отражается на сходстве или непохожести флор и фаун друг с другом. Например, раннее отделение Австралии, Океании и Южной Америки привело к формированию уникальных форм растительного и животного мира этих регионов (сохранение многих форм сумчатых и яйцекладущих млекопитающих, реликтовых растений, исчезнувших на других материках). Напротив, длительно существовавшая связь Северной Америки и Евразии привела к высокой степени сходства их живого мира.
Доказательства из области генетики и молекулярной биологии
Генетика и молекулярная биология — науки о молекулярных основах наследственности и закономерностях их проявления в популяциях организмов. Эти науки позволяют уточнить филогенетическую близость или отдаленность разных групп растений и животных и таким образом дополнить данные, получаемые другими науками. Сведения, подтверждающие современные представления об эволюции живого мира, имеются и во многих других биологических науках — селекции растений, животных, микроорганизмов, сравнительной физиологии и биохимии разных групп организмов, систематике и др.
www.polnaja-jenciklopedija.ru
Доказательства эволюции | Info-Farm.RU
Доказательства эволюции — научные данные и концепции, которые подтверждают происхождение всех живых существ на Земле от общего предка. Благодаря этим доказательствам основы эволюционной теории получили признание научного сообщества, а ведущей системой представлений о процессах видообразования стала синтетическая теория эволюции.
Эволюционные процессы наблюдаются как в естественных, так и в лабораторных условиях. Известны случаи образования новых видов. Описаны также случаи развития новых свойств благодаря случайным мутациям. Факт эволюции на внутривидовом уровне доказано экспериментально, а процессы видообразования наблюдались непосредственно в природе.
С целью получения сведений об эволюционной истории жизни, палеонтологи анализируют ископаемые остатки организмов. Степень родства между современными видами можно определить сравнивая их строение, геномы, развитие эмбрионов (эмбриогенез). Дополнительный источник информации об эволюции — закономерности географического распространения животных и растений, которые изучает биогеография. Все эти данные складываются в единую картину — эволюционное древо жизни.
Видимая эволюция
Изменения в популяциях, которые наблюдаются сегодня, доказывают не только существование эволюции, а также и ряда механизмов, необходимых для эволюционного происхождения всех видов от общего предка. Установлено, что геномы подвержены разнообразным мутациям, среди которых перемещение интронов, Дупликация, рекомбинации, транспозиции, ретровирусные вставки (горизонтальный перенос генов), замена, удаление и вставка отдельных нуклеотидов, а также хромосомные перестройки. Известны такие хромосомные перестройки как дупликация генома (полиплоидия), неравный кроссинговер, хромосомная инверсия, транслокация, деление, слияние, дупликация и удаление хромосом. Также наблюдаются изменения строения организмов и функциональные изменения — разнообразные адаптации, появление способности усваивать новый вид пищи (в том числе — нейлон и пентахлорфенол, производство которых началось в 1930-х годах) и т. Д. Кроме того, были обнаружены различные промежуточные этапы возникновения новых видов, что свидетельствует о плавный характер видообразования.
Эволюция кишечной палочки в лаборатории
Группе биологов из Мичиганского университета под руководством Ричарда Ленски удалось смоделировать в лаборатории процесс эволюции живых организмов на примере бактерии кишечной палочки (Escherichia coli).
За большой скорости размножения смена поколений кишечной палочки происходит крайне быстро, поэтому ученые ожидали, что длительное наблюдение продемонстрирует механизмы эволюции в действии. На первом этапе эксперимента, в 1988 году, 12 колоний бактерий были помещены в идентичные условия: изолированную питательную среду, в которой присутствовало только один источник питательных веществ — глюкоза. Кроме того, в среде был цитрат, который в присутствии кислорода эти бактерии не могут использовать в качестве источника пищи. За 20 лет в колониях изменилось более 44 тысяч поколений бактерий.
Ученые наблюдали за изменениями, которые происходили с бактериями. Большинство из них имели одинаковый характер во всех популяциях — например, размер бактерий увеличивался, хотя и разными темпами. Однако примерно между поколениями номер 31 000 и 32 000 в одной из популяций произошли кардинальные изменения, которые не наблюдались в других. Бактерии приобрели способность усваивать цитрат. Сохраненных образцы бактерий из различных поколений, исследователям удалось установить, что начало серии изменений, которые привели к образованию новой разновидности бактерий, произошло в пределах поколения номер 20000 и только в этой колонии.
Опыты Григория Шапошникова
В конце 1950-х — начале 1960-х годов известный советский биолог Георгий Шапошников провел ряд опытов, в ходе которых проводилась смена кормовых растений у разных видов тлей. Во время опытов впервые наблюдалась репродуктивная изоляция использованных в эксперименте особей от исходной популяции, что свидетельствует об образовании нового вида.
Эволюция вирусов
Частота возникновения мутаций у вирусов намного выше, чем у многоклеточных организмов, поэтому их эволюция происходит в десятки раз быстрее. Наиболее заметным для людей эволюционным приспособлением вирусов является приобретение устойчивости к лекарственным препаратам. Когда в 80-х появились первые средства для лечения ВИЧ, у большинства пациентов эффективное лечение продолжалось недостаточно долго, поскольку вирус приобретал устойчивости против антиретровирусных препаратов в течение нескольких лет, что делало лечение ими неэффективным. Сейчас, чтобы замедлить эволюцию вируса ВИЧ в форм нечувствительных к лекарствам, используют для лечения больных СПИДом используют смеси препаратов с различными механизмами действия, в этом случае возникновения у одного из экземпляров вируса мутации, защищает от действия определенного препарата, не гарантирует этом экземпляру вируса выживания, что замедляет закрепление такой мутации.
Быстрая эволюция ящериц в новых условиях
Согласно опубликованному в 2008 году исследования в популяции стенных ящериц Podarcis sicula всего за 36 лет (очень короткий для эволюции период) изменились размер и форма головы, увеличилась сила укуса и развились новые структуры в пищеварительном тракте. Эти физические изменения произошли одновременно с серьезными изменениями в популяционной плотности и социальной структуре.
В 1971 году пять пар взрослых особей Podarcis sicula с острова Под Кописте были перевезены на соседний остров Под Маркар (эти хорватские острова расположены на юге Адриатического моря, недалеко от Ластово). Условия на новом месте были близки к привычной среды, наземные хищники отсутствовали, и за три десятилетия ящерицы естественным образом распространились на острове (вытеснив при этом местную популяцию Podarcis melisellensis). Когда исследователи вернулись на Под Маркар, они обнаружили, что ящерицы на этом острове существенно отличаются от ящериц на Под Кописте. Сравнение митохондриальной ДНК показало, что популяции генетически не отличаются, но в то же время в популяции с Под Маркар средний размер особей увеличен, задние конечности короче, максимальная скорость бега меньше, а также отличается реакция на нападение хищников. Эти изменения строения и поведения объясняются меньшей активностью хищников на Под Маркар и наличием плотной растительности, что способствовало маскировке. Кроме того, в новой популяции существенно изменились размер и форма головы, она стала более массивной, увеличилась сила укуса. Эти физические изменения соответствуют изменению рациона: P. sicula с Под Кописте преимущественно насекомоядные, а диета ящериц на Под Маркар в зависимости от сезона почти на две трети состоит из растительной пищи. Растения на новом месте жительства жесткие и волокнистые, а мощные челюсти позволяют качественнее откусывать и измельчать растительную пищу.
Наиболее неожиданным отличием оказалась наличие в новой популяции в кишечнике илеоцекальных клапанов, которые предназначены для замедления прохода пищи за счет создания камер брожения, в которых микробы могут разбивать трудно перевариваемые кусочки растительности. Эти клапаны никогда раньше не были обнаружены в этой разновидности ящериц, включая начальную популяцию с острова Под Кописте. Специалисты называют их «эволюционным новшествам, новым свойством, которая отсутствовала в предковой популяции и ранее в ходе эволюции в этих ящериц». Кроме того, в кишечнике новой популяции присутствуют нематоды, которые отсутствуют в исходной популяции.
Изменения в диете также вызвали изменения в плотности популяции и социальной структуре. Так что растения — это гораздо больше, предсказуемое и надежный источник пищи, чем насекомые, количество ящериц увеличилось. Кроме того, пропала необходимость гоняться за добычей, а это привело к тому, что ящерицы перестали агрессивно защищать свои территории.
Развитие устойчивости к антибиотикам и пестицидов
Развитие и распространение устойчивости к антибиотикам у бактерий и устойчивости к пестицидам у растений и насекомых является доказательством эволюции видов. Появление устойчивых к ванкомицину форм золотистого стафилококка и опасность, которую они представляют для пациентов больниц, — это прямой результат эволюции путем естественного отбора. Еще один пример — развитие штаммов шигеллы, устойчивых к антибиотикам из группы сульфаниламидов. Появление малярийных комаров, устойчивых к ДДТ, и развитие в австралийских популяций кроликов устойчивости к миксоматоза демонстрируют действие эволюции в условиях давления отбора при быстрой смене поколений.
За последние 50 лет зарегистрировано более 2500 случаев адаптации насекомых-вредителей к различным пестицидов. Поэтому для борьбы с вредителями вместо обычных ядов пытаются использовать биологические пестициды — Энтомопатогенные вирусы. В частности для борьбы с яблонной плодожоркой Cydia pomonella (личинки которой — это те самые «черви» в червивых яблоках) активно используется Cydia pomonella грануровирус (CpGV). Но в 2003 году были зафиксированы первые случаи снижения эффективности вируса, и к 2005 году появились популяции яблонной плодожорки в которых устойчивость к вирусу выросла в 100 тыс. Раз по сравнению с первоначальной. Установлено, что причиной устойчивости является мутация в половой Z-хромосоме.
Примеры видообразования
Согласно синтетической теории эволюции, образование новых видов происходит в результате разделения особей одного вида на группы, которые не скрещиваются между собой. Такое разделение может быть вызван различными причинами, такими как наличие непреодолимых географических барьеров (алопатричне видообразования) или разделение по экологическим нишам внутри одного ареала (Симпатрические видообразования). После возникновения между популяциями различных типов изоляций, популяций начинают эволюционировать независимо, в результате чего между популяциями постепенно накапливаются генетические различия, и через некоторое время достигается генетическая несовместимость — скрещивание оказывается невозможным.
Кроме того, новые виды могут возникать практически мгновенно (в геологическом масштабе времени) путем полиплоидии (кратного увеличения числа хромосом в клетке) и гибридизации (скрещивания разных видов).
Наблюдение за современными видами показывают, что видообразование непрерывно происходит в существующих популяциях.
Промежуточные этапы видообразования
В определении понятия биологического вида существенное значение имеет практически полная репродуктивная изоляция в природных условиях. Если видообразование происходит постепенно, как следствие незначительных последовательных изменений, тогда и сегодня мы должны наблюдать все возможные стадии видообразования, а это значит и все возможные стадии репродуктивной изоляции: свободное скрещивание между популяциями, затруднено скрещивания, снижение плодовитости потомства, бесплодное потомство и полную генетическую изоляцию . Если ничего подобного среди современных видов обнаружить не удалось, то тем самым основы эволюционной теории были бы опровергнуты.
Существует множество примеров того, как различные виды могут скрещиваться в исключительных условий. Например, в черной вороны и серой вороны разные ареалы, но на пересечении этих ареалов виды могут образовывать гибриды. Еще один пример — различные виды рыб рода чукучан из реки Платт, которые живут рядом, но редко скрещиваются.
Неполное видообразования можно наблюдать в различных кольцевых видов: представители вида живут вокруг гор, водоемов и т. П., При этом соседние популяции скрещиваются между собой и незначительно отличаются, а крайние формы различаются значительно и не скрещиваются. Пример кольцевого вида — безлегочных саламандра (Ensatina eschscholtzii), живущий на склонах гор вблизи тихоокеанского побережья Северной Америки. В зависимости от района проживания вокруг гор саламандры образуют различные формы, постепенно меняют свои морфологические и экологические характеристики. Крайние формы Ensatina eschscholtzii eschscholtzii и Ensatina eschscholtzii klauberi, первая из которых имеет красноватую окраску, а вторая черно-белое, сосуществуют в узкой зоне гор на юге Калифорнии, однако между собой не скрещиваются. Другие примеры круговых видов — зеленый пеночка (Phylloscopus trochiloides), оленья мышь (Peromyscus maniculatus), наземный улитка (Partula olympia), дрозофила (Drosophila paulistorum).
Во многих гибридов снижена фертильность. У гибридов землероек из рода бурозубок размножения затруднено вследствие разницы хромосомных наборов. Тот же эффект наблюдался в лаборатории при скрещивании дрозофил Drosophila pseudoobscura с Юты и Калифорнии. Фертильность также снижена у гибридов лягушек из семьи бомбинових краснобрюхая жерлянка и кумки желтобрюхой.
Многие другие виды могут давать бесплодное гибридное потомство. Среди них — различные земноводные (например — некоторые виды лягушек из рода Rana) и млекопитающие (например, ил — гибрид от скрещивания лошади и осла — как правило, бесплодный). Гибриды гребенчатого тритона и мраморного тритона бесплодны вследствие наличия напорных хромосом.
При скрещивании ряда других видов достигается оплодотворение, но позднее зародыш погибает. В качестве примера можно привести леопардовую лягушку (Rana pipiens) и лесную лягушку (Rana sylvatica) из рода настоящих лягушек. Аналогичная ситуация наблюдается у дрозофил и у некоторых растений, таких как хлопчатник обыкновенный (Gossypium hirsutum) и хлопчатник барбадосский (Gossypium barbadense).
- Белый медведь
Конкретным примером масштабных эволюционных изменений при неполном видообразовании является белый медведь, который, несмотря на родство с бурым медведем (Ursus arctos), очевидную через тот факт, что эти виды могут скрещиваться и давать плодовитое потомство, приобрел значительные физиологических различий с бурым медведем. Эти различия позволяют белому медведю комфортно жить в условиях, в которых бурый медведь не выжил бы. В частности, белый медведь способен проплыть десятки километров в ледяной воде, его окраска сливается со снегом, он не замерзает в Арктике. Все это возможно благодаря конкретным изменениям: белая окраска способствует маскировке хищника при охоте на тюленей; пустотелые волоски увеличивают плавучесть и сохраняют тепло; слой подкожного жира, толщина которого к зиме доходит до 10 сантиметров, обеспечивает дополнительную теплоизоляцию; удлиненная, по сравнению с другими медведями, шея позволяет легче держать голову над водой во время плавания; увеличены лапы с перепонками действуют как весла; небольшие бугорки и полости-присоски на подошвах уменьшают опасность поскользнуться на льду, а плотная шерсть на подошвах защищает лапы от сильного холода и обеспечивает трения; уши меньше, чем в других медведей, и уменьшают потери тепла; веки действуют как солнечные очки; зубы острые, чем у других медведей, и больше подходят для полностью мясного рациона; увеличен объем желудка позволяет голодному хищнику съесть сразу целого тюленя, кроме того, белый медведь способен обойтись без еды до девяти месяцев за счет переработки мочевины.
Наблюдаемое видообразование
Судя по палеонтологической летописи и по измерениям скорости мутаций, полная несовместимость геномов, которая делает невозможным скрещивание, достигается в природе в среднем за 3 млн лет. Это означает, что наблюдение за образованием нового вида в природных условиях в принципе возможно, но это — редкое событие. В то же время, в лабораторных условиях скорость эволюционных изменений может быть увеличена, поэтому есть основания надеяться на наблюдение видообразования у лабораторных животных.
Известно много случаев видообразования через гибридизацию и полиплоидизация в таких растений, как конопля, крапива, первоцвет, редька, капуста, а также в различных видов папоротника. В ряде случаев видообразования у растений происходило без гибридизации и полиплоидизации (кукуруза, Stephanomeria malheurensis с семейства астровых).
Дрозофилы, также известные как плодовые мушки, относятся к наиболее изученных организмов. С 1970-х годов зафиксировано много случаев видообразования у дрозофил. Видообразования происходило, в частности, за счет пространственного разделения, разделения по экологическим нишами в одном ареале, изменения поведения в спаривании, дизруптивного отбора, а также за счет сочетания эффекта основателя с эффектом бутылочного горла.
Видообразования наблюдалось в лабораторных популяциях комнатных мух, мухи-осетници Eurosta solidaginis, яблоневой плодовой мухи Rhagoletis pomonella, мучных жуков, комаров и других насекомых.
Известны случаи, когда в результате давления отбора (в присутствии хищников) одноклеточные зеленые водоросли из рода хлорелла образовывали многоклеточные колониальные организмы, а бактерии при аналогичных условиях меняли строение и увеличивали размеры (с 1,5 до 20 микрометров за 8-10 недель). Являются ли эти случаи примерами видообразования, зависит от того, какое определение вида используется (по бесполого размножения невозможно включить критерий репродуктивной изоляции).
Видообразования также наблюдалось и у млекопитающих. Шесть случаев видообразования в домашних мышей на острове Мадейра за последние 500 лет были следствием исключительно географической изоляции, генетического дрейфа и слияния хромосом. Слияние двух хромосом — это заметная разница геномов человека и шимпанзе, а в некоторых популяций мышей на Мадейре за 500 лет было девять подобных слияний.
- Яблоневая плодовая муха
Яблоневая плодовая муха Rhagoletis pomonella является примером наблюдаемого симпатрического видообразования (то есть видообразования в результате разделения по экологическим нишами). Сначала вид жил в восточной части США. До появления европейцев личинки этих мух развивались только в плодах боярышника. Однако с завозом в Америку яблонь (первое упоминание яблонь в Америке — 1647 год), открылась новая экологическая ниша. В 1864 году личинок Rhagoletis pomonella было обнаружено в яблоках, и тем самым зафиксировано яблоневую расу этого вида. За полтора века наблюдений расы очень сильно разошлись. Они почти не скрещиваются друг с другом (уровень гибридизации не превышает 4-6%). Яблоневая раса спаривается почти исключительно на яблонях, а Глодова — на боярышнике, что, учитывая разное время созревания плодов, приводит к репродуктивной изоляции. Вскоре возможно преобразование этих рас в самостоятельные виды.
У плодовых мух, или же мух-осетниць известно еще несколько видов-двойников, которые живут на разных видах растений, — вероятно, видообразование у них происходило именно по описанной схеме.
Эволюционное дерево
Общепринятая в научном сообществе классификация живых организмов представляет собой многоуровневую иерархическую структуру: организмы делятся на царства, царства делятся на типы, типы — на классы, классы — на ряды, и так далее. Вследствие такого разветвления образуется филогенетическое дерево. Существуют противоречия между биологами по поводу отнесения отдельных видов к конкретным группам, но эти противоречия носят единичный характер. На практике биологические классификации, построенные на основе различных признаков, в тенденции приближаются к одной и той же древовидной иерархической схемы — естественной классификации. А значит, возможно, в основе этой классификации существует закономерность.
Именно такой результат можно ожидать при евлюцийному происхождении животных от общего предка. Разветвления филогенетического дерева соответствует разделению популяций в процессе видообразования.
Как правило, объекты, возникшие не в ходе эволюции, не имеют таких свойств. Невозможно объективно построить иерархию элементарных частиц, химических элементов, планет Солнечной системы. Также не существует объективной иерархии таких сознательно созданных объектов как книги в библиотеке, дома, мебель, машины. Можно при желании объединить эти объекты в различные иерархии, но не существует единой объективной иерархии, принципиально лучше, чем все остальные.
Существует ряд статистических методов для точной оценки того, насколько объекты с разными качествами вкладываются в ту или иную иерархию. Эти методы измеряют так называемый «филогенетический сигнал», позволяя отличить мнимые закономерности от объективных. Например, у любого «генеалогического древа» автомобилей будет низкий уровень филогенетического сигнала. В эволюционного дерева и в различных его частей, наоборот, стабильно четкий филогенетический сигнал.
Есть несколько источников данных, на основе который можно делать выводы о степени родства между видами. Если существует единственное эволюционное дерево, которое объединяет все виды в объективную генеалогию, то все данные должны подтверждать эту генеалогию. Филогенетические деревья, построенные независимо, должны соответствовать друг другу. Наиболее простой, хотя не очень точный способ построить такое дерево — сравнение строения организмов животных: чем меньше разногласий между видами, тем меньше поколений отделяют их от общего предка. Палеонтологическая летопись позволяет установить родство между классами животных. Например, найдены останки пернатых динозавров свидетельствуют о родстве между рептилиями и птицами. Биогеография и эмбриология также дают информацию о эволюционную близость видов. Точнее источник данных, которое было недоступно во времена Дарвина — сравнительный анализ геномов различных организмов. Эволюционное дерево можно построить по каждому отдельному гену, исследователи также рассматривают различные последовательности, которые не кодируют белки.
Все эти источники информации дают одинаковую картину с точностью до погрешности используемых методов. Тот факт, что эволюционные деревья, построенные по разным данным, соответствуют друг другу, объясняется эволюционной теорией. Другие объяснения отсутствуют: например, нет никаких причин, почему организмы, сходные по строению, должны иметь подобные некодирующие последовательности в геноме, если они не происходят от одного предка.
Палеонтологические доказательства
Как правило, остатки растений и животных разлагаются и исчезают без следа. Но иногда биологические ткани замещаются минеральными веществами, и образуются окаменелости. Обычно находят окаменелые кости или раковины, то есть твердые части живых организмов. Иногда находят отпечатки следов животных или следы их жизнедеятельности. Еще реже находят животное целиком — фасованное во льду в районах современной вечной мерзлоты, или которая попала в окаменевшую позже смолу древних растений — янтарь, или в другую природную смолу — асфальт.
Изучением ископаемых останков занимается палеонтология. Обычно, осадочные породы залегают слоями, поэтому более глубокие слои содержат окаменелости с более раннего периода (принцип суперпозиции в геологии). А значит, сравнивая ископаемые формы из последовательных наслоений, можно делать выводы об основных направлениях эволюции живых организмов. Для оценки возраста окаменелостей используются многочисленные методы геохронологии.
При взгляде на палеонтологическая летопись можно сделать вывод, что жизнь на Земле существенно менялось. Чем глубже в прошлое мы смотрим, тем меньше видим общего с современной биосферой. Первые прокариоты (простейшие одноклеточные, не обладающих оформленным клеточным ядром) появляются примерно 3500000000 лет назад. Первые одноклеточные эукариоты появляются 2,7-1,75 млрд лет назад. Примерно через миллиард лет, 840 млн лет назад, в палеонтологической летописи появляются первые многоклеточные животные, представители хайнаньского фауны. Согласно опубликованному в 2009 году исследование, вероятно, больше 635 млн лет назад уже существовали многоклеточные, относящиеся к одному из современных типов — губки. В период «кембрийського взрыва», 540-530 млн лет назад, за геологически короткое время, в геологической летописи появляются остатки представителей большинства современных типов, имеющих скелеты, а через 15 млн лет — первые примитивные позвоночные, похожие на современных миноги. Челюстные рыбы появляются 410 млн лет назад, насекомые — 400 млн лет назад, и еще 100 млн лет на суше господствуют папоротникообразные, а основными группами наземной фауны остаются насекомые и земноводные. С 250 по 65 млн лет назад на Земле господствующее положение «верховных хищников» и крупных травоядных занимали динозавры и другие пресмыкающиеся, наиболее распространенными растениями были саговники и другие группы голосеменных. Первые ископаемые остатки цветочных появляются 140-130 млн лет назад, а начало их широкого распространения относится к середине мелового периода (около 100 млн лет назад). Эта картина соответствует происхождению всех видов от одноклеточных организмов и не имеет других научных объяснений.
Известный доказательство эволюции — наличие так называемых переходных форм, то есть организмов, сочетающих в себе характерные признаки различных видов. Как правило, говоря о промежуточных (или «переходные») формы подразумевают ископаемые виды, хотя промежуточные виды не всегда вымирают. На основе филогенетического дерева теория эволюции предсказывает, промежуточные формы могут быть найдены, а какие — нет. Согласно научных методов, прогнозирования, которые сбываются, подтверждают теорию. Например, зная строение организмов пресмыкающихся и птиц, можно предположить некоторые особенности переходной формы между ними. Можно прогнозировать возможность найти останки животных, подобных рептилиям, но с перьями, или останки животных, подобных птицам, но с зубами или с длинными хвостами со скелетом с несросшихся позвонков. При этом можно предположить, не будут найдены переходные формы между птицами и млекопитающими, например — ископаемые млекопитающие с перьями или подобные птицам ископаемые с костями среднего уха как у млекопитающих.
Вскоре после публикации «Происхождения видов» были обнаружены останки археоптерикса — промежуточной формы между рептилиями и птицами. Для археоптерикса свойственно дифференцированное оперение (типичная черта птиц), а по строению позвоночника он мало отличался от динозавров из группы Компсогнат. у него были когти на передних конечностях, зубы и длинный хвост с позвоночником из позвонков, не срослись, а вероятные уникальные «птичьи» особенности скелета позже были обнаружены в других рептилий. Позже были найдены и другие переходные формы между рептилиями и птицами.
Известно немало других переходных форм, в том числе — от беспозвоночных до рыб, от рыб к четвероногим, от земноводных к рептилиям и от рептилий к млекопитающим.
В некоторых случаях ископаемые переходные формы обнаружить не удалось, например — нет следов эволюции шимпанзе (вероятно, это объясняется отсутствием условий для образования окаменелостей в лесах, где они живут), нет следов ресничных червей, а этот класс объединяет более 3500 видов . Разумеется, чтобы возразить спростовуванисть теории эволюции, недостаточно указать на подобные пробелы в палеонтологической летописи. Чтобы опровергнуть эволюционную теорию, надо было бы предъявить скелет, который не соответствует филогенетическом дереву или не укладывается в хронологической последовательности. Так, в ответ на вопрос о том, какая находка могла бы опровергнуть эволюционную теорию, Джон Холдейн ответил: «Ископаемые кролики в докембрии!» Были найдены миллионы окаменелостей у 250000 ископаемых видов, и каждый находка — это проверка теории эволюции, а пройденная проверка подтверждает теорию.
В тех случаях, когда палеонтологическая летопись оказывается особенно полным, появляется возможность построить так называемые филогенетические ряды, то есть такие ряды видов (родов и т. Д.), Которые последовательно сменяли друг друга в процессе эволюции. Наиболее известные филогенетические ряды человека и лошади, также в качестве примера можно привести эволюцию китообразных.
Эволюция человека
Первый экземпляр неандертальца был найден в 1856 году, за три года до издания «Происхождение видов». На момент выхода книги не было известно никаких других ископаемых остатков, подтверждающих эволюционное происхождение человека и человекообразных обезьян от общего предка. Хотя неандерталец является независимым видом из рода людей, с тех пор было найдено множество скелетов промежуточных форм между давними антропоидами и человеком.
Поскольку общий предок человека и шимпанзе передвигался на четырех конечностях, а мозг у него был больше чем у шимпанзе, согласно теории Дарвина в процессе эволюции должно было развиться прямохождение, а также имел увеличиться объем мозга. Таким образом, должен был существовать один из трех вариантов переходных форм:
- Промежуточный размер мозга, прямохождение, которое развивается.
- Размер мозга примерно такой как у шимпанзе, развитое прямохождение.
- Больший мозг, прямохождение не развито.
В 1920-х годах в Африке были обнаружены останки существа, которое Раймонд Дарт назвал австралопитеком. Позже были найдены и другие остатки австралопитеков, в том числе — известная Люси и череп AL 444-2. Австралопитеки жили в восточной и северной Африке 4-2 млн лет назад. Объем мозга австралопитека был немного больше, чем у шимпанзе. Кости таза по строению наиболее близки к костям таза человека. Строение черепа характерно для прямоходящих животных, можно определить в частности по foramen magnum — отверстием в затылочной кости, соединяющий полость черепа с позвоночным каналом. Более того, в Танзании в неподвижном вулканическом пепле были обнаружены следы, оставленные двуногими гоминидами. Возраст следов составляет 3600000 лет. Таким образом, австралопитеки является «переходной формой номер два»: в них было развито частичное прямохождение, но мозг был примерно как у шимпанзе.
Позже были найдены остатки ардипитека, возраст которых составляет 4500000 лет. Анализ скелета показал, что по земле ардипитека передвигались на двух задних конечностях, а по деревьям лазили на всех четырех. Прямохождения у ардипитека было слабо развито по сравнению со следующими гоминидами, они не могли преодолевать большие расстояния. Ардипитека представляет собой переходную форму между общим предком человека и шимпанзе и австралопитеком.
Итак, в результате эволюции в гоминид сначала развилось прямохождение, и только потом началось существенное увеличение объема мозга. В австралопитеков, живших 4-2 млн лет назад, объем мозга был около 400 см³, примерно как у шимпанзе. Человек умелый (Homo habilis) жила 2,4-1,4 млн лет назад, размер мозга составлял в нее 500-640 см³. Человек работающий (Homo ergaster) жила 1,9-1,4 млн лет назад, размер мозга — 700-850 см³. Человек прямоходящий (Homo erectus) жила 1,4-0,2 млн лет назад, размер мозга составлял от 850 см³ в ранней особи до 1100 см³ в поздних. Гейдельбергский человек (Homo heidelbergensis) жила 600-350 тысяч лет назад, размер мозга составлял 1100-1400 см³. Неандертальцы жили 350-25 тысяч лет назад, размер мозга составлял 1200-1900 см³. 200000 лет назад появился человек разумный (Homo sapiens), размер мозга людей составляет 1000-1850 см³. Согласно современным представлениям, не все перечисленные виды были непосредственными предками современных людей. В частности, неандертальцы развивались параллельно с Homo sapiens. Тем не менее, на примере этих видов можно наблюдать направление эволюции гоминид. Эти виды хорошо представлены в палеонтологической летописи.
Согласно Дарвину, «в ряду форм, которые незаметно переходят одна в другую от любой мавпоподибои существа к человеку в его современном состоянии, было бы невозможно точно указать, какой именно из этих форм следует впервые дать наименование» человека «». Именно с этой проблемой столкнулись современные палеонтологи, которые вынуждены делить найдены останки гоминид на разные роды. Например, черепа KNM ER 1813, KNM ER 1470 и OH 24 («Твигги») в разное время классифицировались как Australopithecus habilis и как Homo habilis, только потому, что они оказались на искусственном «границе», произвольно проведенном между практически неразличимыми родами.
Эволюция лошади
Эволюция лошади достаточно полно прослеживается в Северной Америке. Древнейший представитель лошади
info-farm.ru
Доказательства эволюции
Важными доказательствами единства происхождения всех живых организмов служат: их клеточное строение, принципы генетического кодирования наследственной информации, биосинтеза белков , основные механизмы энергетического обмена, использование АТФ как универсального носителя энергии.
Еще в XIX в. в эмбриологии были получены многочисленные факты, свидетельствующие в пользу эволюционной идеи. Благодаря трудам А. О. Ковалевского и И. И. Мечникова было создано «учение о зародышевых листках» вскрывшее общие закономерности эмбрионального развития беспозвоночных и позвоночных животных. В это же время Ф. Мюллером и Э. Геккелем был сформулирован биогенетический закон: онтогенез есть краткое повторение филогенеза (т. е. исторического развития вида). Иными словами, в процессе индивидуального развития организм на стадии зародыша (или личинки) повторяет признаки своих предков. Например, при развитии человека его зародыш на определенных стадиях имеет жаберные щели, хвост, сплошной волосяной покров и т. д. (рис. 1).
Сравнительно-анатомические исследования также наглядно иллюстрируют общность происхождения разных групп животных.
Наиболее существенную роль сыграло выявление гомологии органов разных организмов. Например, передние конечности у позвоночных — конечность лягушки, лапа собаки, крыло птицы, ласт кита, рука обезьяны — сходны по своему строению (составу костей), хотя и выполняют различные функции. Гомологичные органы имеются и у растений: части цветка (пестик, тычинки, лепестки) гомологичны листьям; усики гороха, колючки кактуса — это видоизмененные листья. Гомологичные органы имеют сходное строение и развиваются из одинаковых эмбриональных зачатков.
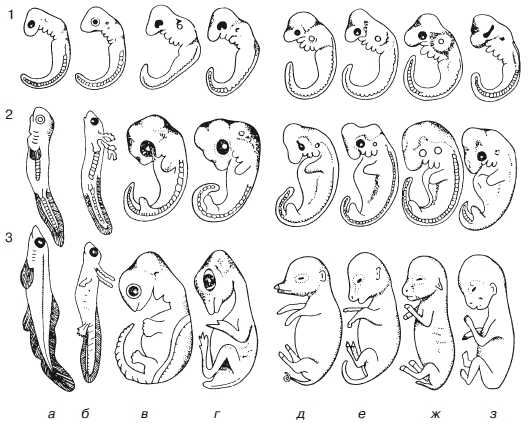
Рис. 1. Сходство эмбриональных стадий как проявление «биогенетического закона»: 1-3 — стадии развития; а — рыба; б — саламандра; в — черепаха; г — птица; л — свинья; е — корова; ж — кролик; з — человек
Яркие доказательства эволюции — атавизмы и рудименты. Атавизм — это появление признака, присущего далекому предку и в норме не встречающегося у современных форм. В качестве примера можно привести появление хвоста у человека, наличие более одной пары молочных желез, волосатость и т. д.
Рудиментарными называются органы, утратившие свою функцию, но сохраняющиеся у особей данного вида в зачаточном состоянии. Например, остатки тазовых костей у кита,
копчика у человека (остаток хвостовых позвонков), зачаточные ушные мышцы (предки человека обладали подвижной ушной раковиной).
Очень важными свидетельствами эволюции, конечно, выступают и палеонтологические данные.
Палеонтология — это наука об организмах, когда-то живших на нашей планете, но затем исчезнувших в ходе эволюции.
Изучение этих организмов ведется по их остаткам в виде окаменелостей и отпечатков. Приуроченность таких находок к определенным геологическим пластам позволяет убедиться в том, что по мере перехода от более древних слоев к новым наблюдается появление организмов, более высоко организованных. Очень ценно обнаружение переходных форм. Так, были обнаружены остатки древней вымершей птицы археоптерикса (рис. 2), в строении которой сочетаются признаки рептилий (зубы, брюшные ребра, длинный хвост с 20 позвонками) и птиц (тело и крылья покрыты перьями, имеются клюв, полые кости).
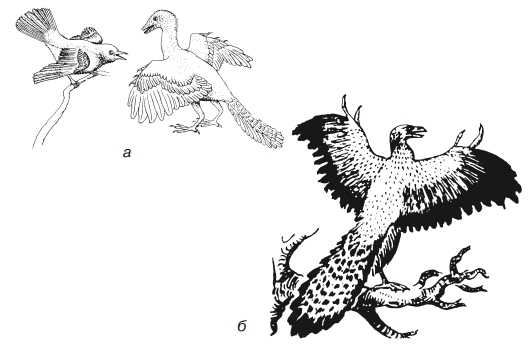
Рис. 2. Существование переходных форм: а — примитивная птица археоптерикс (справа) и современная птица (слева). Обратите внимание на различие в строении хвоста; б — археоптерикс (реконструкция З. Буриана)
Источник: Краснодембский Е. Г.»Общая биология: Пособие для старшеклассников и поступающих в вузы»
xn--90aeobapscbe.xn--p1ai
