
Синтез углеводов происходит при фотосинтезе
Синтез углеводов происходит при фотосинтезе и сопровождается формированием не только простых моносахаридов, но и молекул сложных сахаров – крахмала, сахарозы и клетчатки. В растениях происходит разложение углеводов до простых составляющих, и их перетекание в органы, нуждающиеся в питании. Уже здесь происходит новое образование сложных углеводов, регулирующих процессы прорастания, распада или увеличения корневой системы.
Процессы перехода углеводов из одной формы в другую проходят при участии фосфорных эфиров полисахаридов или их уридиндифосфатов (УДФ-доноры), которые представляют собой определённый полисахарид, соединённый с уридином при помощи двух фосфорных остатков.
Сахароза. Синтез углевода происходит при фотосинтезе из фруктозы и глюкозы. Сначала молекула глюкозы фосфилируется в присутствии АТФ до образования глюкозы-6-фосфата и молекулы АДФ (аденозиндифосфат) – переносчика фосфатной группы.
Одновременно проходит похожая реакция с участием фруктозы и образованием фруктозо-6-фосфата, соединяемого с УДФ-глюкозой в присутствии фермента сахарозофосфат-УДФ-глюкозилтрансферазы. Образуется сахарозо-6-фосфат, который под воздействием фосфатазы, контролирующей гидролиз, переходит в свободную сахарозу.
Крахмал. Синтезируемый углевод состоит из амилозы и амилопектина – двух полисахаридов. Амилоза образуется удлинением глюкозных остатков уридиндифосфатглюкозы (УДФК) с помощью акцептора в присутствии фермента крахмалглюкозилтрансферазы. Аналогичная реакция происходит при синтезе амилопектина, только в качестве носителей глюкозных остатков используют аденозиндифосфатглюкозу.
Целлюлоза. Синтез углевода происходит при фотосинтезе, и молекула состоит из остатков β-глюкозы. В качестве исходного материала используется гуанозиндифосфатглюкоза, преобразующаяся в присутствии фермента целлюлозосинтетазы в целлюлозу.
Фармконтракт-Синтез является структурным подразделением ГК «Фармконтракт», разрабатывает и реализует проекты биохимического синтеза органических соединений, ферментов и молекул при помощи высокоточных систем в условиях приближённых к натуральным. Мы самостоятельно устанавливаем приборы и комплексы, обеспечиваем сервисное обслуживание и быструю доставку оборудования.
Синтез углеводов и других веществ в растениях
В противоположность только что описанным процессам существует другой путь распада крупных молекул органических веществ на более мелкие частицы — анаэробное разложение. Он происходит лишь в отсутствии кислорода воздуха. Этот процесс известен также под названием гниения . В нем принимают участие живые организмы. Однако, в то время как в процессе аэробного разложения участвуют многие виды организмов, процессы гниения осуществляются лишь некоторыми видами бактерий. При поступлении воздуха в достаточном количестве бактерии гниения подавляются другими организмами, поглощающими кислород.
В связи с важной ролью углеводов в обмене веществ и в жизнедеятельности растений изучение их синтеза и обмена постоянно находилось в центре внимания биохимиков.
 Некоторые из этих процессов были изучены довольно давно, другие — лишь Б последнее время, а детали химизма ряда сложных процессов еще неясны. [c.141]
Некоторые из этих процессов были изучены довольно давно, другие — лишь Б последнее время, а детали химизма ряда сложных процессов еще неясны. [c.141] Синтез крахмала. Крахмал — основное запасное вещество больщинства растений, в которых он очень легко образуется как в процессе ассимиляции СО2, так и из других углеводов. Как мы видели, крахмал состоит из амилозы, представляющей собой неразветвленную цепь остатков а-глюкозы, соединенных
Биосинтез начинается с фотосинтеза [1]. Вся жизнь на Земле зависит от способности некоторых организмов (зеленых растений, водорослей и фотосинтезирующих бактерий), содержащих характерные фотосинтезирующие пигменты, использовать энергию солнечной радиации для синтеза органических молекул из неорганических веществ — диоксида углерода, азота и серы. Продукты фотосинтеза служат затем не только исходными веществами, но и источником химической энергии для всех последующих биосинтетических реакций. Обычно принято описывать фотосинтез только как процесс образования углеводов в некоторых случаях основными продуктами фотосинтеза, действительно, являются исключительно крахмал, целлюлоза и сахароза, однако в других организмах на синтез углеводов идет, быть может, всего лишь третья часть углерода, связываемого и восстанавливаемого в процессе фотосинтеза.
СИНТЕЗ УГЛЕВОДОВ И ДРУГИХ ВЕЩЕСТВ В РАСТЕНИЯХ [c.234]
Стереохимическая специфичность действия ферментов проявляется не только в реакциях гидролитического расщепления, но и в любых реакциях. При искусственном синтезе простейших углеводов, аминокислот и некоторых других веществ образуется смесь изомеров (рацематов), тогда как при ферментативных синтезах всегда образуется только один из оптических изомеров вешества. Растения и животные из -кислот и аммиака синтезируют аминокислоты -ряда. При фотосинтезе из углекислого газа и воды образуется >-глюкоза и т. д. [c.132]
Синтез углеводов.

Калий, натрий и кальций находятся в растениях в виде солей органических и неорганических кислот. Соли калия оказывают влияние на синтез углеводов, белков и других веществ и способствуют оттоку образовавшихся в листе углеводов. При недостатке калия плохо развиваются опорные ткани и на растениях появляются бурые пятна.
Магний входит в состав хлорофилла, усиливает синтез белков, углеводов, липидов и других веществ. Железо участвует в образовании хлорофилла, а также содержится в ряде дыхательных ферментов. Микроэлементы (например, молибден, марганец, медь) играют очень важную роль в жизни растений, так как входят в состав ферментов, катализирующих многие процессы обмена веществ. [c.9]
Например, в живых организмах из веществ относительно простого состава (вода, углекислый газ и сахар), в которых упорядоченность очень низка, образуются многие сложно построенные соединения (белки и т. д.), имеющие весьма высокую упорядоченность атомов. Такие процессы (например, синтез белков) идут с уменьшением энтропии. Зато одновременно другие вещества (молекулы сахара, жиры и т. д.) в результате окисления распадаются на более простые соединения (в конечном счете на углекислый газ и воду), а при этом снова происходит значительный рост энтропии. Конечно, в растениях из углекислого газа и воды в процессе фотосинтеза образуются также сахар и другие углеводы, но этот процесс идет не изолированно, а с поглощением энергии излучения Солнца.
В результате работ И. П. Бородина выяснилось, что синтез аспарагина протекает в растениях с большой скоростью в условиях недостатка углеводов, когда происходит интенсивный окислительный распад белков. Д. Н. Прянишников выполнил очень важные исследования по обмену амидов (аспарагина и глютамина) в растениях. В исследованиях Д. Н. Прянишникова было показано, что синтез аспарагина и глютамина в растениях является процессом, аналогичным синтезу мочевины в животном мире. В обоих случаях достигается обезвреживание аммиака, но при этом все же имеется существенная разница. Мочевина является неактивным веществом в отношении дальнейшего участия в процессах обмена она выводится из организма без изменений, являясь типичным примером конечного продукта обмена. Аспарагин же и глютамин способны к дальнейшим превращениям и могут вовлекаться в процессы синтеза белка и других азотистых соединений (стр. 378). В. Л. Кретович показал, что в обмене аспарагина и глютамина в растениях имеется существенное различие.
Фосфорорганические соединения, так же как и углеводы, в обработанных дефолиантами листьях распадаются на более подвижные соединения и интенсивно оттекают в другие органы растений. Это говорит о том, что дефолианты сильно нарушают ход физиологических и биохимических процессов в листьях растений, в частности ослабляют процесс синтеза органических веществ и усиливают процесс распада. Такой характер обмена веществ способствует образованию отделительного слоя и опадению листа. При-этом отделительный слой образуется вследствие деления и обособления клеток указанной зоны.
Одновременно с распадом органического азотсодержащего вещества в почве происходят и вторичные процессы синтеза, когда минеральные соединения азота вновь переходят в органические, не усвояемые для растений. Эти процессы носят биологический характер. Микроорганизмы строят белок своих тел, используя углеводы и азот. Различные бактерии, актиномицеты и плесневые грибы разлагают клетчатку, пентозаны и другие вещества растительных и животных остатков. В качестве азотной пищи им необходимы в первую очередь легкорастворимые соединения азота почвы, которые они переводят в белок плазмы клеток. [c.181]
В качестве азотной пищи им необходимы в первую очередь легкорастворимые соединения азота почвы, которые они переводят в белок плазмы клеток. [c.181]
Распад нуклеиновых кислот под влиянием нуклеаз происходит и в животном, и в растительном организмах. Образовавшиеся пуриновые и пиримидиновые основания, углевод и фосфорная кислота подверга.ются в организме дальнейшим превращениям. Рибоза и дезоксирибоза могут окисляться до углекислоты и воды. Пуриновые и пиримидиновые основания превращаются в конечные продукты обмена, которые у животных выделяются из организма, а у растений могут быть использованы для синтеза других веществ. [c.372]
Одним из методов синтеза меченых соединений является биосинтез. Биосинтез широко используется при введении метки в сложные органические вещества природного происхождения (белки, углеводы и т. п.). Простейшим примером является получение меченой глюкозы и других углеводов в процессе фотосинтеза.
 Освещенные зеленые листья живого растения по мещают в атмосферу меченной по углероду (С) углекислоты. Затем сахар экстрагируют из растений и очищают, используя обычные химические и биохимические процедуры. [c.176]
Освещенные зеленые листья живого растения по мещают в атмосферу меченной по углероду (С) углекислоты. Затем сахар экстрагируют из растений и очищают, используя обычные химические и биохимические процедуры. [c.176] Превращения углеводов, жиров и белков, их распад и синтез в организме теснейшим образом связаны друг с другом. Нельзя представить себе изолированно превращение отдельных органических, а также и неорганических веществ в организме. Только как исключение можно наблюдать преимущественный синтез углеводов (у зеленых растений на свету), распад углеводов с образованием этилового спирта и углекислого газа (в дрожжевых клетках при спиртовом брожении) и молочной кислоты (при работе мышц), синтез жиров (при откорме животных), синтез белков (при усиленном росте). Но даже и в этих случаях обмен веществ не сводится к превра[це-пиям т0JПзK0 одной какой-либо группы веществ. Обмен веществ между любым живым организмом п окружающей его средой является чрезвычайно слюж-ным процессом, в который вовлекаются химические составные части организма и вещества, поступающие в пего извне (пищевые вещества, включая кпс лород и воду). Обмен веществ у человека и животных регулируется централыюй нервной системой. При изучении превращений углеводов, жиров и белков приводились данные о регуляторной деятельности центральной нервной системы. Было бы ошибочным полагать суп.1,ествование в центральной нервной системе отдельных механизмов, регулирующих превращения отдельных групп веществ. Процесс обмена веществ между организмом и внешней средой, лежащей в основе проявления жизни,— единый биологический процесс и если его расчленяют на процессы превращения отдельных веществ, то это делают только с целью более глубокого его познания и изучения. [c.459]
Обмен веществ у человека и животных регулируется централыюй нервной системой. При изучении превращений углеводов, жиров и белков приводились данные о регуляторной деятельности центральной нервной системы. Было бы ошибочным полагать суп.1,ествование в центральной нервной системе отдельных механизмов, регулирующих превращения отдельных групп веществ. Процесс обмена веществ между организмом и внешней средой, лежащей в основе проявления жизни,— единый биологический процесс и если его расчленяют на процессы превращения отдельных веществ, то это делают только с целью более глубокого его познания и изучения. [c.459]
Если представить себе работу живой клетки в целом, то можно сказать, что клетки организмов животных, растений и даже одноклеточные микроорганизмы представляют собой поразительные по своему совершенству химические заводы. В них вырабатываются, например, сложнейшие полимерные частицы с самыми разнообразными свойствами, различного состава, разной прочности, эластичности, различной термоустойчивости и окраски. В клетках очень постоянно, одновременно, или, если это нужно, то в необходимой последовательности, происходят тонкие синтезы таких сложных и разнохарактерных веществ, как нуклеиновые кислоты, белки и в том числе ферменты, сложные углеводы, жиры, витамины, гормоны и множество других сложных соединений. Эти заводы в клетках смонтированы очень компактно, все процессы в них точно и четко регулируются, режим оптимальный. В основе работы их лежит согласованное действие ферментных систем, осуществляющих цепи управляемых химических реакций. Естественно, что в будущем подобные принципы работы должны быть использованы и химической, и биохимической промышленностью. [c.337]
В клетках очень постоянно, одновременно, или, если это нужно, то в необходимой последовательности, происходят тонкие синтезы таких сложных и разнохарактерных веществ, как нуклеиновые кислоты, белки и в том числе ферменты, сложные углеводы, жиры, витамины, гормоны и множество других сложных соединений. Эти заводы в клетках смонтированы очень компактно, все процессы в них точно и четко регулируются, режим оптимальный. В основе работы их лежит согласованное действие ферментных систем, осуществляющих цепи управляемых химических реакций. Естественно, что в будущем подобные принципы работы должны быть использованы и химической, и биохимической промышленностью. [c.337]
Промышленный органический синтез (получение топлива, растворителей, высокополимеров и т. д.) основывается на минеральном углеродистом сырье нефти, каменном угле и природных газах. Кроме того, многие органические веш,ества могут быть получены из природного сырья, т. е. из органических соединений, синтезируемых растениями и животными.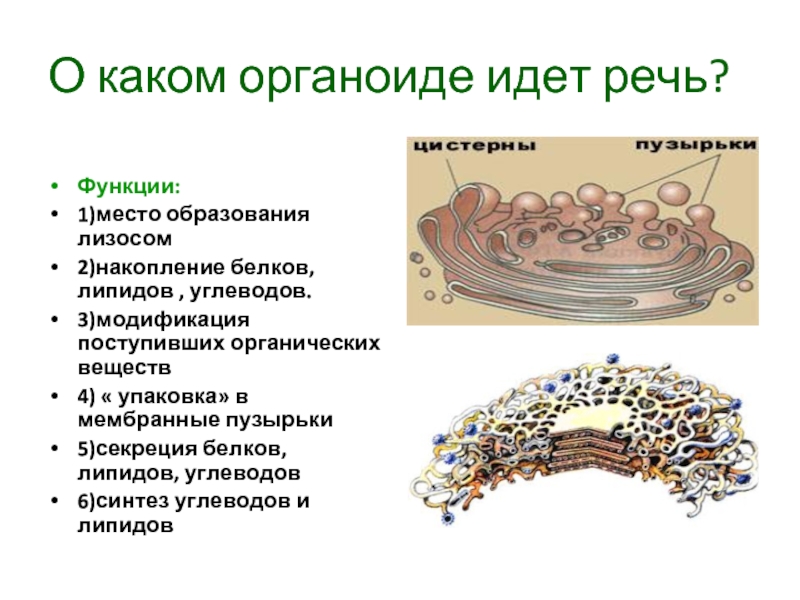 Такие соединения являются представителями трех основных групп природных веществ углеводов, жиров и белков. Одни из них, а именно углеводы, весьма широко используются в химической и пищевой промышленности (переработка древесины различными способами, добывание и переработка крахмалистых и сахаристых продуктов), другие — жиры — занимают несравненно меньшее, но все же значительное место в промышленности (главным образом пищевой и мыловаренной), третьи — белки — широко используются в пищевой промышленности и в незначительной степени в других производствах (некоторые виды пластмасс, кожевенное производство). [c.5]
Такие соединения являются представителями трех основных групп природных веществ углеводов, жиров и белков. Одни из них, а именно углеводы, весьма широко используются в химической и пищевой промышленности (переработка древесины различными способами, добывание и переработка крахмалистых и сахаристых продуктов), другие — жиры — занимают несравненно меньшее, но все же значительное место в промышленности (главным образом пищевой и мыловаренной), третьи — белки — широко используются в пищевой промышленности и в незначительной степени в других производствах (некоторые виды пластмасс, кожевенное производство). [c.5]
Растение и животное в создании органического вещества. Основная задача сельскохозяйственного производства— получение органических веществ растительного и животного происхождения углеводов, белков, жиров и других органических соединений. Синтез органического вещества осуществляется в природных условиях в организме растений и животных. [c.20]
Медь участвует в углеводном и белковом обмене в растениях. Под влиянием меди повышается как активность пероксидазы, так и синтез белков, углеводов и жиров. Недостаток меди вызывает у растений понижение активности синтетических процессов и ведет к накоплению растворимых углеводов, аминокислот и других продуктов распада сложных органических веществ. [c.21]
Под влиянием меди повышается как активность пероксидазы, так и синтез белков, углеводов и жиров. Недостаток меди вызывает у растений понижение активности синтетических процессов и ведет к накоплению растворимых углеводов, аминокислот и других продуктов распада сложных органических веществ. [c.21]
Уничтожая главные составные части растений — лигнин, целлюлозу и другие углеводы, сапрофиты синтезируют вещества другой природы — белки, жиры, хитин. Поэтому вопрос об исходном материале горючих ископаемых не так прост, как это казалось лет 20 тому назад. Горючие ископаемые образуются не непосредственно из растительных остатков и не из продуктов их химического распада, а из продуктов сложного биохимического синтеза, осуществляемого микробами. Характер продуктов этого синтеза зависит не столько от химического состава первичного растительного материала, сколько от условий, в которые попадает этот материал после его отмирания. [c.372]
При недостатке или отсутствии углеводов для дыхания используются другие органические соединения, обладающие необходи-мым. запасом энергии. Однако в этом случае синтез всех других веществ будет в значительной степени нарушен, а рост и развитие растений будут подавлены. [c.12]
запасом энергии. Однако в этом случае синтез всех других веществ будет в значительной степени нарушен, а рост и развитие растений будут подавлены. [c.12]
Продукты распада сине-зеленых водорослей. Сине-зеленые водоросли относятся к группе низших, наиболее примитивных растений. В большинстве случаев это одноклеточные организмы, обычно соединяющиеся в кс-лонии. У некоторых клетки при помощи слизи и выростов соединены в ценобии в виде нитей, давая внешнюю картину многоклеточности (рис. 9.1). Размножаются они преимущественно путем деления клеток. Живут сине-зеленые водоросли не только в воде, но и на суше (на берегах водоемов, в почвах и на их поверхности). Это самые распространенные растения земного шара. Они первыми заселяют бесструктурные почвы и совместно с бактериями подготавливают их для освоения другими растениями. Эти водоросли вообще аэробные организмы. Они способны к синтезу углеводов, но используют и распадающиеся органические вещества. [c.253]
Переаминирование имеет большое значение для синтеза белков, а также для дезаминирования аминокислот. Дезаминирование — это отщепление аминогруппы от аминокислоты, в результате образуются аммиак и кетокис-лота. Кетокислота используется растением для переработки в углеводы, жиры и другие вещества аммиак же вступает в реакцию прямого аминиро-вания кетокислот, возникающих из углеводов, и дает аминокислоты. Кроме [c.184]
Дезаминирование — это отщепление аминогруппы от аминокислоты, в результате образуются аммиак и кетокис-лота. Кетокислота используется растением для переработки в углеводы, жиры и другие вещества аммиак же вступает в реакцию прямого аминиро-вания кетокислот, возникающих из углеводов, и дает аминокислоты. Кроме [c.184]
Все эмпирически известное по поводу энзимов в клетках растений не имеет непосредственного отношения к этим гипотетическим катализаторам. Данные, имеющиеся в нашем распоряжении, касаются хорошо известных энзимов, вроде каталазы, карбоангидразы, фос-форилазы, амилазы, мальтазы и инвертазы, которые или вообще не имеют отношения к синтезу углеводов, или участвуют лишь в его заключительных стадиях (образование и разлоягение сахарозы и крахмала). О наличии энзимов, превращающих углеводы в зеленых листьях, кратко говорилось в главе П1. Здесь мы добавим немногие данные по другим энзималг, найденным в выделенном веществе хлоропластов [97, 98, 105]. [c. 381]
381]
Во время дыхания растительный организм, постепенно расщепляя молекулу глюкозы, извлекает ее энергию для накоплепия во вновь образующихся фосфатйых связях АТФ. Молекулы ад но-зинтрифосфата (АТФ) переносят полученную за счет питательных веществ или солнечного света свободную энергию от центров дыхания (митохондрий) или фотосинтеза (пластид) во все участки клетки, обеспечивая прохождение всех процессов, связанных с поглощением энергии. Углеводы при дыхании окисляются не сразу до углекислого газа и воды, а постепенно, образуя десятки промежуточных соединений, которые растения используют для синтеза всех необходимых им других органических соединений. Получающиеся при дыхании органические кислоты, спирты и другие соединения служат, как и при фотосинтезе, для образования белков, жиров и других веществ. Суммарно и схематически процесс дыхания растений можно представить уравнением, как бы обратным уравнению фотосинтеза [c.11]
Переаминирование имеет большое значение для синтеза белков, а также для дезаминирования аминокислот. Дезаминирование — это отщепление аминогруппы от аминокислоты, в результате образуются аммиак и кетокис-лота. Кетокислота используется растением для переработки в углеводы, жиры и другие вещества аммиак же вступает в реакцию прямого аминирова-ния кетокислот, возникающих из углеводов, и дает аминокислоты. Кроме того, аммиак реагирует с аспарагиновой и глютаминово кислотами, способными связать еще но одной его молекуле, давая таким образом амиды амино дикарбоновых кислот [c.173]
Дезаминирование — это отщепление аминогруппы от аминокислоты, в результате образуются аммиак и кетокис-лота. Кетокислота используется растением для переработки в углеводы, жиры и другие вещества аммиак же вступает в реакцию прямого аминирова-ния кетокислот, возникающих из углеводов, и дает аминокислоты. Кроме того, аммиак реагирует с аспарагиновой и глютаминово кислотами, способными связать еще но одной его молекуле, давая таким образом амиды амино дикарбоновых кислот [c.173]
Фотохимические (фотокаталитически е) реакции происходят в природе и сравнительно давно используются промышленностью. Фотохимическими называют реакции, вызываемые и ускоряемые действием света их элементарный механизм состоит в активации молекул при поглощении фотонов. Большинство промышленных фотохимических реакций происходит по цепному механизму, т. е. молекулы, поглотившие фотон, диссоциируют, и активированные атомы или группы атомов служат инициаторами вторичных реакций. По такому типу протекают галогенирование углеводородов и других веществ, синтез полистирола, сульфохлорирование парафинов и т. п. Природный фотосинтез требует непрерывного подвсда световой энергии синтез углеводов из диоксида углерода воздуха совершается под действием солнечного света, поглощаемого пигментом растений хлорофиллом (аналог гемоглобина крови). При фотокаталитических реакциях фотоны поглощаются не реагентами, а катализаторами, ускоряющими химическую реакцию, т. е. реакция ускоряется в результате суммарного действия катализатора и световой энергии. [c.150]
п. Природный фотосинтез требует непрерывного подвсда световой энергии синтез углеводов из диоксида углерода воздуха совершается под действием солнечного света, поглощаемого пигментом растений хлорофиллом (аналог гемоглобина крови). При фотокаталитических реакциях фотоны поглощаются не реагентами, а катализаторами, ускоряющими химическую реакцию, т. е. реакция ускоряется в результате суммарного действия катализатора и световой энергии. [c.150]
ФОТОСИНТЕЗ — синтез растениями органических веществ (углеводов, белков, жиров) из диоксида углерода, воды, азота, ( юсфора, минеральных солей и других компонентов с помощью солнечной энергии, поглощаемой пигментом хлорофиллом. Ф.— основной процесс образования органических веществ на Земле, определяющий круговорот углерода, кислорода и других элементов, а также основной механизм трансформации солнечной энергии на нашей планете. В процессе Ф, растения усваивают вгод4 101 туглерода, разлагают 1,2 х X 10 т воды, выделяют 1 10 т кислорода и запасают 4-102° кал солнечной энергии в виде химической энергии продуктов Ф. Это количество энергии намного превышает годовую потребность человечества в ней. Ф.—сложный окис-лительно-восстановительный процесс, сочетающий фотохимические реакции с ферментативными. Вследствие Ф. происходит окисление воды с выделением молекулярного кислорода и восстановление диоксида углерода, что выражается [c.268]
Это количество энергии намного превышает годовую потребность человечества в ней. Ф.—сложный окис-лительно-восстановительный процесс, сочетающий фотохимические реакции с ферментативными. Вследствие Ф. происходит окисление воды с выделением молекулярного кислорода и восстановление диоксида углерода, что выражается [c.268]
В организмах человека и животных происходят процессы синтеза различных органических веществ. Следует, однако, отметить, что процессы синтеза в этих организмах не столь разнообразны, как в зеленых растениях, и известным образом ограничены. Следует подчеркнуть, что животные организмы неспособны использовать энергию солнечных лучей для синтеза органических соединений. Из этого отнюдь не вытекает, что в организме животных не используется для синтеза углекислый газ, вода и аммиак. Уже с давних пор известно, что выделяющаяся нз организмов человека, млекопитающих животных, амфибий и рыб мочевина синтезируется из углекислого газа, воды и аммиака применение метода меченых молекул позволило выявить участие воды, углекислого газа и аммиака в процессах синтеза сложных органических веществ — составных частей организма. После введения в организм животных тяжелой воды можно обнаружить тяжелый водород (дейтерий) в составе жиров, углеводов, белков и других веществ организма. Введение в организм животных карбонатов, меченных i позволяет проследить, как различные органические вещества приобретают радиоактивную метку, благодаря включению в их состав углерода углекислого газа. После введения в организм животных аммонийных солей, меченных стабильным изотопом азота (N ), появляется в составе белков и других азотистых веществ. Все эти данные с несомненностью показывают, что в организме животных для синтеза органических веществ используются минеральные вещества — аммиак, вода и углекислый газ. Было бы, однако, ошибочным считать, что в организмах животных и в зеленых растениях отсутствуют различия в использовании минеральных веществ для синтетических целей. Различия эти прежде всего количественного характера. Объем использования углекислого газа, воды и аммиака для синтеза органических веществ в организмах животных, но сравнению с организмами зеленых растений, незначителен.
После введения в организм животных тяжелой воды можно обнаружить тяжелый водород (дейтерий) в составе жиров, углеводов, белков и других веществ организма. Введение в организм животных карбонатов, меченных i позволяет проследить, как различные органические вещества приобретают радиоактивную метку, благодаря включению в их состав углерода углекислого газа. После введения в организм животных аммонийных солей, меченных стабильным изотопом азота (N ), появляется в составе белков и других азотистых веществ. Все эти данные с несомненностью показывают, что в организме животных для синтеза органических веществ используются минеральные вещества — аммиак, вода и углекислый газ. Было бы, однако, ошибочным считать, что в организмах животных и в зеленых растениях отсутствуют различия в использовании минеральных веществ для синтетических целей. Различия эти прежде всего количественного характера. Объем использования углекислого газа, воды и аммиака для синтеза органических веществ в организмах животных, но сравнению с организмами зеленых растений, незначителен. Далее обращают на себя внимание и различия качественного характера-, ряд веществ, синтезирующихся в растениях, вовсе не синтезируются в организмах человека и животных, и эти вещества должны доставляться человеку и животным в готовом виде с продуктами питания. Так, оргагшзмы человека и животных не способны к синтезу ряда аминокислот, входящих в состав белков, не могут синтезировать различные витамины и т. д. Отсутствие этих веществ в пище приводит к их гибели. [c.235]
Далее обращают на себя внимание и различия качественного характера-, ряд веществ, синтезирующихся в растениях, вовсе не синтезируются в организмах человека и животных, и эти вещества должны доставляться человеку и животным в готовом виде с продуктами питания. Так, оргагшзмы человека и животных не способны к синтезу ряда аминокислот, входящих в состав белков, не могут синтезировать различные витамины и т. д. Отсутствие этих веществ в пище приводит к их гибели. [c.235]
Известно, что при фотосинтезе и минеральном питании растений, этих двух взаимосвязанных автотрофных процессах, происходят поглощение и ассимиляция органогенов С, О, Н, Ы, Р, 5 и других элементов (металлов и неметаллов) и синтез углеводов, белков, липидов и веществ вторичного происхождения (органические кислоты, гликозиды, фенольные соединения, алкалоиды, каучук и др.), т, е. синтез органических веществ. Установленные закономерности н корреляционные зависимости между световым и углеродным питанием (фотосинтез), транспирацией, дыханием, минеральным питанием и урож айность10 составляют основу теории объединения воздушного и минерального питания растений как интегрального процесса. Поэтому для протекания в растениях эффективного продукционного процесса необходимы следующие условия [c.350]
Поэтому для протекания в растениях эффективного продукционного процесса необходимы следующие условия [c.350]
Для исследования процессов образования полисахаридов большое значение имеет использование меченых соединений, например Oq, меченых сахаров и их производных. Предполагается, что биосинтезирующая система растений не различает соединений, содержащих С и С, и в обоих случаях реакции протекают одинаково, Включение СОз в состав воздуха, окружающего растущее растение, и анализ образцов, собранных через определенное время после начала эксперимента, позволили сделать ряд интересных выводов. За очень короткие промежутки времени в процессе фотосинтеза образуется большое количество меченых соединений. Тем не менее у большинства растений основная масса С уже в первые минуты концентрируется в углеводах. Когда в питательную среду вводится радиоактивная глюкоза, радиоактивность прежде всего обнаруживается в полисахаридах и только потом — в иротеинах, липидах и других полимерах. Если учесть, что глюкоза и ее соединения являются исходным веществом для синтеза большинства биополимеров, то можно полагать, что путь иревра- [c.24]
Если учесть, что глюкоза и ее соединения являются исходным веществом для синтеза большинства биополимеров, то можно полагать, что путь иревра- [c.24]
Заканчивая рассмотрение аминокислотного обмена, следует сказать, что обычно в растениях в свободном состоянии содержится 20—30 различных аминокислот, которые подвергаются непрерывным превращениям используются для синтеза белков, нуклеиновых кислот, алкалоидов и других азотистых веществ, превращаются в безазотистые соединения — органические кислоты, углеводы, жиры. Содержание аминокислот в растениях может резко меняться в зависимости от возраста растений, от ряда внешних условий (температуры, длины дня, увлажнения и т. д.), а также от питания. При этом изменяется ке только концентрация, но и качественный состав аминокислот. Различные внешние воздействия, нарушая течение азотного обмена, часто направляют его по другим путям, что приводит к уменьшению или даже к исчезновению ряда аминокислот, характерных для данного растения, или, наоборот, к повышенпю общего содержания аминокислот, или появлению ряда нехарактерных продуктов азотного обмена. При обычных условиях выращивания количество свободных аминокислот с возрастом растений понижается. В вегетативных органах растений свободных аминокислот обычно больше, чем в репродуктивных, в то время как для белков наблюдается обратная зависимость. При различных условиях минерального питания содержание индивидуальных аминокислот в растениях и соотношение между ими могут быть резко различными. Увеличение общего количества свободных аминокислот в растениях и усиленное накопление отдельных аминокислот наблюдается при пониженном питании растений калием, фосфором, серой, кальцием и магнием, а также при недостатке ряда микроэлементов цинка, меди, марганца, железа. Увеличение содержания аминокислот наблюдалось также при лучших условиях азотного питания. При недостатке молибдена количество свободных аминокислот и амидов в растениях уменьшалось вследствие ослабления восстановления нитратов. В настоящее время проводятся широкие исследования [c.264]
При обычных условиях выращивания количество свободных аминокислот с возрастом растений понижается. В вегетативных органах растений свободных аминокислот обычно больше, чем в репродуктивных, в то время как для белков наблюдается обратная зависимость. При различных условиях минерального питания содержание индивидуальных аминокислот в растениях и соотношение между ими могут быть резко различными. Увеличение общего количества свободных аминокислот в растениях и усиленное накопление отдельных аминокислот наблюдается при пониженном питании растений калием, фосфором, серой, кальцием и магнием, а также при недостатке ряда микроэлементов цинка, меди, марганца, железа. Увеличение содержания аминокислот наблюдалось также при лучших условиях азотного питания. При недостатке молибдена количество свободных аминокислот и амидов в растениях уменьшалось вследствие ослабления восстановления нитратов. В настоящее время проводятся широкие исследования [c.264]
Одновременно с минерализацией органического азотсодержащего вещества в почве происходят и вторичные процессы синтеза, когда минеральные соединения азота вновь переходят в органические, не усвояемые для растений. Эти процессы носят биологический характер. Микроорганизмы строят белок своих тел, используя углеводы и азот. Различные бактерии, лктиномицеты и плесневые грибы разлагают клетчатку, пентозаны и другие [c.192]
Эти процессы носят биологический характер. Микроорганизмы строят белок своих тел, используя углеводы и азот. Различные бактерии, лктиномицеты и плесневые грибы разлагают клетчатку, пентозаны и другие [c.192]
При. попытке выделения продуктов метаболизма 2,4-Д в растениях обнаружено вещество кислого характера, извлекаемое эфиром [936], в которое пе >еходит до 60% радиоактивного углерода. Предполагали, что это может быть один из изомеров окси-2,4-Д [936]. Синтез и изучение некоторых таких соединений показали, что они физиологически неактивны [937, 938]. Обработка растений фасоли малыми дозами 2,4-Д, меченной С , показала, что в результате метаболизма 2,4-Д превращаег-ся в весьма разл1ичные продукты, так как С обнаружен в белках, углеводах, кислотах и других соединениях [939, 940]. [c.375]
В настоящее время следовало бы попытаться произвести синтез более сложных азотистых соединений, исходя из формамида и формальдегида или других углеводов. Получаясь при соединении двух характерных групп СНО и Nh3 и являясь типичным альдегидамидом, формамид должен обладать способностью вступать в соединения посредством либо одной, либо другой из своих групп или посредством обеих. Если бы можно было достигнуть таким образом синтеза аспарагина или аналогичного вещества, то вопрос о синтезе белковых веществ в растениях существенно продвинулся бы. Я сделал в этом направлении некоторые опыты, которые до настоящего времени не дали удовлетворительных результатов я предполагаю продолжить эти исследования. [c.11]
Получаясь при соединении двух характерных групп СНО и Nh3 и являясь типичным альдегидамидом, формамид должен обладать способностью вступать в соединения посредством либо одной, либо другой из своих групп или посредством обеих. Если бы можно было достигнуть таким образом синтеза аспарагина или аналогичного вещества, то вопрос о синтезе белковых веществ в растениях существенно продвинулся бы. Я сделал в этом направлении некоторые опыты, которые до настоящего времени не дали удовлетворительных результатов я предполагаю продолжить эти исследования. [c.11]
Основные закономерности метаболических процессов в организме человека. Часть 2.
Рассматривая обмен веществ в условиях нормального функционирования организма, следует остановиться на безусловно взаимосвязанных, но в то же время достаточно специфичных составляющих метаболизма, а именно на углеводном, белковом, липидном и водно-электролитном обмене.
Очевидно, что основная роль углеводов в метаболизме определяется их энергетической функцией. Именно глюкоза крови вследствие наличия простого и быстрого пути гликолитической диссимиляции и последующего окисления в цикле трикарбоновых кислот, а также возможности максимально быстрого извлечения ее из депо гликогена, обеспечивающей экстренную мобилизацию энергетических ресурсов, является наиболее востребованным источником энергии в организме. Использование циркулирующей в плазме глюкозы разными органами неодинаково: мозг задерживает 12% глюкозы, кишечник— 9%, мышцы — 7%, почки — 5%. При этом уровень глюкозы плазмы крови является одной из важнейших гомеостатических констант организма, составляя 3, 3—5, 5 ммоль/л. Как известно снижение уровня глюкозы ниже допустимого передела имеет своим незамедлительным следствием дискоординацию деятельности ЦНС, проявляющуюся соответствующей клинической симптоматикой: головной мозг содержит небольшие резервы углеводов и нуждается в постоянном поступлении глюкозы, поскольку энергетические расходы мозга покрываются исключительно за счет углеводов. Глюкоза в тканях мозга преимущественно окисляется, а небольшая часть ее превращается в молочную кислоту.
Именно глюкоза крови вследствие наличия простого и быстрого пути гликолитической диссимиляции и последующего окисления в цикле трикарбоновых кислот, а также возможности максимально быстрого извлечения ее из депо гликогена, обеспечивающей экстренную мобилизацию энергетических ресурсов, является наиболее востребованным источником энергии в организме. Использование циркулирующей в плазме глюкозы разными органами неодинаково: мозг задерживает 12% глюкозы, кишечник— 9%, мышцы — 7%, почки — 5%. При этом уровень глюкозы плазмы крови является одной из важнейших гомеостатических констант организма, составляя 3, 3—5, 5 ммоль/л. Как известно снижение уровня глюкозы ниже допустимого передела имеет своим незамедлительным следствием дискоординацию деятельности ЦНС, проявляющуюся соответствующей клинической симптоматикой: головной мозг содержит небольшие резервы углеводов и нуждается в постоянном поступлении глюкозы, поскольку энергетические расходы мозга покрываются исключительно за счет углеводов. Глюкоза в тканях мозга преимущественно окисляется, а небольшая часть ее превращается в молочную кислоту.
Единственной формой углеводов, которая может всасываться в кишечнике, являются моносахара. Они всасываются главным образом в тонкой кишке, током крови переносятся в печень и к тканям. Основная часть поступающей с пищей глюкозы (около 70%) окисляется в тканях до воды и углекислого газа, около 25—28% пищевой глюкозы превращается в жир и только 2—5% ее синтезируется в гликоген. Гликоген печени представляет собой основной резерв углеводов в организме, достигая по своей массе у взрослого человека 150—200 г. Синтез гликогена происходит достаточно быстро, что, наряду с быстрой мобилизацией гликогена и поступлением глюкозы в кровь в процессе гликогенолиза, является одним из механизмов поддержания гликемии в константных пределах. Помимо печени в качестве депо гликогена выступают также мышцы. Однако запас гликогена в мышечной массе по отношению к всему гликогену организма составляет всего 1 — 2%. В мышцах под влиянием фермента фосфорилазы, которая активируется в начале мышечного сокращения, происходит усиленное расщепление гликогена, являющегося одним из источников энергии мышечного сокращения. При распаде мышечного гликогена процесс идет до образования пировиноградной и молочной кислот. Этот процесс называют гликолизом. В фазе отдыха из молочной кислоты в мышечной ткани происходит ресинтез гликогена.
При распаде мышечного гликогена процесс идет до образования пировиноградной и молочной кислот. Этот процесс называют гликолизом. В фазе отдыха из молочной кислоты в мышечной ткани происходит ресинтез гликогена.
При полном отсутствии углеводов в пище они образуются в организме из продуктов трансформации жиров и белков. В печени возможно новообразование углеводов как из собственных продуктов их распада (пировиноградной или молочной кислоты), так и из продуктов диссимиляции жиров и белков (кетокислот и аминокислот), что обозначается как глюконеогенез. В результате трансформации аминокислот образуется пировиноградная кислота, при окислении жирных кислот — ацетилкоэнзим А, который может превращаться в пировиноградную кислоту — предшественник глюкозы. Это наиболее важный общий путь биосинтеза углеводов. Между двумя основными источниками энергии — углеводами и жирами — существует тесная физиологическая взаимосвязь. Повышение содержания глюкозы в крови увеличивает биосинтез триглицеридов и уменьшает распад жиров в жировой ткани. Поступление в кровь свободных жирных кислот уменьшается. В случае возникновения гипогликемии процесс синтеза триглицеридов тормозится, ускоряется распад жиров и в кровь в большом количестве поступают свободные жирные кислоты. Гликогенез, гликогенолиз и глюконеогенез являются тесно взаимосвязанными процессами, обеспечивающими оптимальный уровень глюкозы крови сообразно степени функционального напряжения организма.
Поступление в кровь свободных жирных кислот уменьшается. В случае возникновения гипогликемии процесс синтеза триглицеридов тормозится, ускоряется распад жиров и в кровь в большом количестве поступают свободные жирные кислоты. Гликогенез, гликогенолиз и глюконеогенез являются тесно взаимосвязанными процессами, обеспечивающими оптимальный уровень глюкозы крови сообразно степени функционального напряжения организма.
Центральным звеном регуляции углеводного и других видов обмена и местом формирования сигналов, управляющих уровнем глюкозы, является гипоталамус. Отсюда регулирующие влияния реализуются вегетативными нервами и гуморальным путем, включающим эндокринные железы. Единственным гормоном, снижающим уровень гликемии, является инсулин — гормон, вырабатываемый β-клетками островков Ланхгерганса. Снижение гликемии происходит за счет усиления инсулином синтеза гликогена в печени и мышцах и повышения потребления глюкозы тканями организма. Увеличение уровня глюкозы в крови возникает при действии нескольких гормонов. Это глюкагон, продуцируемый α-клетками островков Ланхгерганса, адреналин — гормон мозгового слоя надпочечников, глюкокортикоиды — гормоны коркового слоя надпочечников, соматотропный гормон гипофиза, тироксин и трийодтиронин — гормоны щитовидной железы. Данные гормоны в связи с однонаправленностью их влияния на углеводный обмен и функциональным антагонизмом по отношению к эффектам инсулина часто объединяют понятием «контринсулярные гормоны».
Это глюкагон, продуцируемый α-клетками островков Ланхгерганса, адреналин — гормон мозгового слоя надпочечников, глюкокортикоиды — гормоны коркового слоя надпочечников, соматотропный гормон гипофиза, тироксин и трийодтиронин — гормоны щитовидной железы. Данные гормоны в связи с однонаправленностью их влияния на углеводный обмен и функциональным антагонизмом по отношению к эффектам инсулина часто объединяют понятием «контринсулярные гормоны».
Таким образом биологическая роль углеводов для организма человека определяется прежде всего их энергетической функцией. Обладая энергетической ценностью в 16, 7 кДж (4, 0 ккал) на 1 грамм вещества, углеводы являются основным источником энергии для всех клеток организма, при этом выполняя еще пластическую и опорную функции. Суточная потребность взрослого человека в углеводах составляет около 500 г.
Характерной особенностью белкового обмена является его чрезвычайная разветвленность. Достаточно указать, что в обмене 20 аминокислот, входящих в состав белковых молекул, в организме животных участвуют сотни промежуточных метаболитов, тесно связанных с обменом углеводов и липидов. Число ферментов, катализирующих химические реакции азотистого обмена, также исчисляется сотнями. Собственно белки (протеины и протеиды), высокомолекулярные соединения, построенные из мономеров — аминокислот, занимают ведущее место среди органических элементов организма, составляя более 50 % сухой массы клетки. Как известно, белки в организме выполняют ряд важнейших биологических функций, а именно:
Число ферментов, катализирующих химические реакции азотистого обмена, также исчисляется сотнями. Собственно белки (протеины и протеиды), высокомолекулярные соединения, построенные из мономеров — аминокислот, занимают ведущее место среди органических элементов организма, составляя более 50 % сухой массы клетки. Как известно, белки в организме выполняют ряд важнейших биологических функций, а именно:
— пластическая (структурная) функция заключается в том, что белки являются главной составной частью всех клеточных и межклеточных структур тканей;
— ферментная (каталитическая, энзимная) функция состоит в обеспечении всех химических реакций, протекающих в ходе обмена веществ в организме (дыхание, пищеварение, выделение), деятельностью ферментов, являющихся по своей структуре белками;
— транспортная функция белков заключается в их способности к соединению с целым рядом метаболитов и переносе последних в связанном состоянии в межтканевой жидкости и плазме крови к области их утилизации;
— защитная функция белков проявляется реализацией иммунного ответа образованием иммуноглобулинов (антител) и системы комплемента при поступлении в организм чужеродного белка, а также способностью к непосредственному связыванию экзогенных токсинов; белки системы гемостаза обеспечивают свертывание крови и остановку кровотечения при повреждении кровеносных сосудов;
— регуляторная функция, направленная на сохранение гомеостаза с поддержанием биологических констатнт организма, реализуется буферными свойствами молекулы протеинов, белковой структурой клеточных рецепторов, активируемых в свою очередь регуляторными полипептидами и гормонами, также имеющими белковую структуру;
— двигательная функция, обеспечивается взаимодействием сократительных белков мышечной ткани актина и миозина;
— энергетическая роль белков состоит в обеспечении организма энергией, образующейся при диссимиляции белковых молекул; при окислении 1 г белка в среднем освобождается энергия, равная 16, 7 кДж (4, 0 ккал).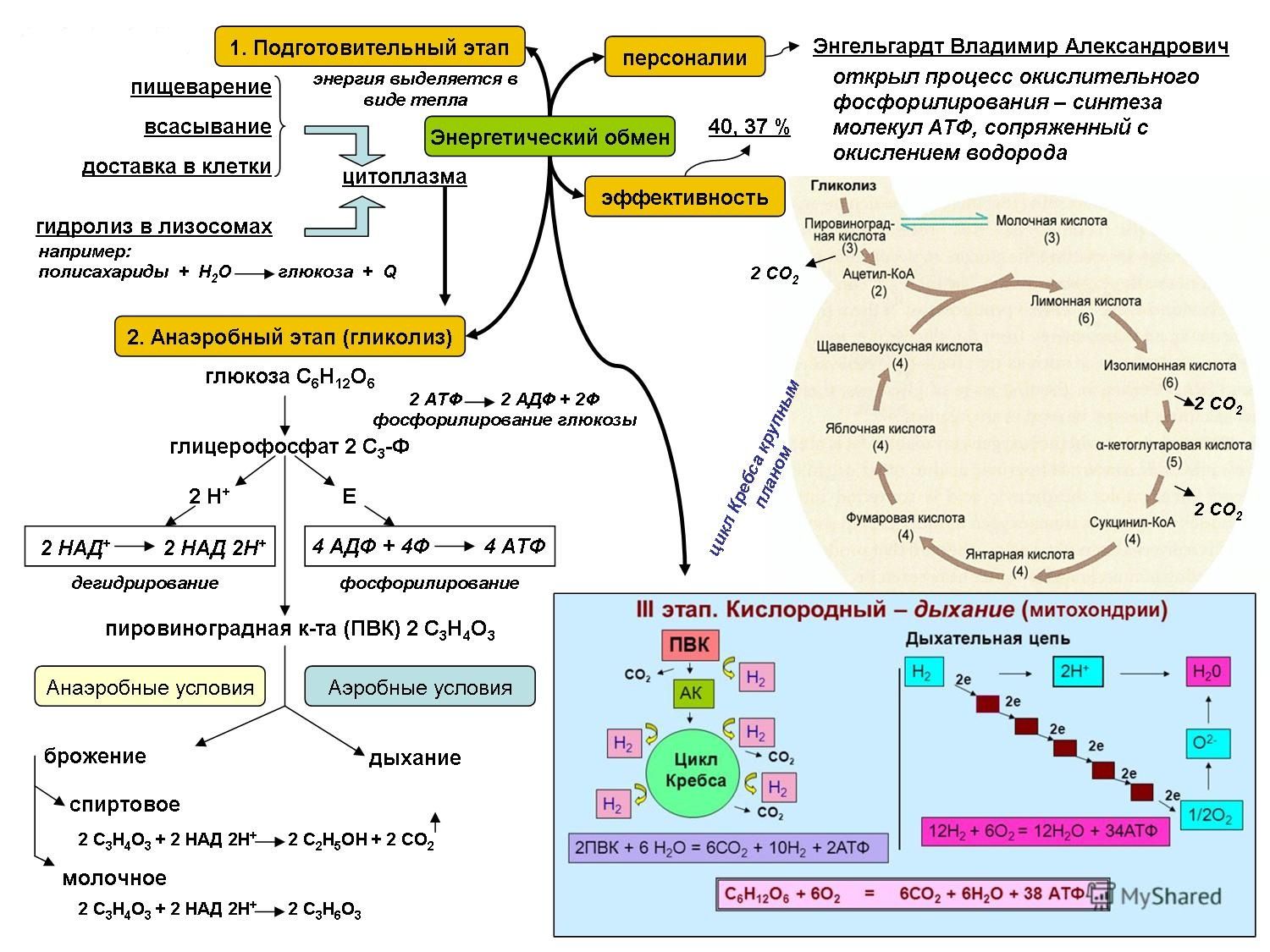
В организме постоянно происходит распад и синтез белков. Единственным источником синтеза нового белка являются белки пищи. В пищеварительном тракте белки ферментативно расщепляются ферментами до аминокислот и абсорбируются в тонкой кишке. Транспорт их осуществляется двумя путями: через воротную систему печени, ведущую прямо в печень, и по лимфатическим сосудам, сообщающимся с кровью через грудной лимфатический проток. Максимальная концентрация аминокислот в крови достигается через 30 — 50 мин после приёма белковой пищи (углеводы и жиры замедляют всасывание аминокислот). Всасывание L-аминокислот (но не D-изомеров) — активный процесс, требующий затраты энергии. Аминокислоты переносятся через кишечную стенку от слизистой её поверхности в кровь. Перенос через щеточную кайму осуществляется целым рядом переносчиков, многие из которых действуют при участии Na+-зависимых механизмов симпорта, подобно переносу глюкозы.
Из аминокислот и простейших пептидов клетки тканей синтезируют собственный белок, который характерен только для данного организма. Белки не могут быть заменены другими пищевыми веществами, так как их синтез в организме возможен только из аминокислот. Вместе с тем белок может замещать собой жиры и углеводы, то есть использоваться для синтеза этих соединений. В тканях постоянно протекают процессы распада белка с последующим выделением из организма неиспользованных продуктов белкового обмена и параллельно с этим — синтез белков. Катаболизм большинства аминокислот начинается с отщепления α-аминогруппы результате реакций трансаминирования и дезаминирования. Чаще всего в реакциях трансаминирования участвуют аминокислоты, содержание которых в тканях значительно выше остальных — глутамат, аланин, аспартат и соответствующие им кетокислоты — α—кетоглутарат, пируват и оксалоацетат. Основным донором аминогруппы служит глутамат. Реакции трансаминирования играют большую роль в обмене аминокислот. Поскольку этот процесс обратим, ферменты аминотрансферазы функционируют как в процессах катаболизма, так и биосинтеза аминокислот.
Белки не могут быть заменены другими пищевыми веществами, так как их синтез в организме возможен только из аминокислот. Вместе с тем белок может замещать собой жиры и углеводы, то есть использоваться для синтеза этих соединений. В тканях постоянно протекают процессы распада белка с последующим выделением из организма неиспользованных продуктов белкового обмена и параллельно с этим — синтез белков. Катаболизм большинства аминокислот начинается с отщепления α-аминогруппы результате реакций трансаминирования и дезаминирования. Чаще всего в реакциях трансаминирования участвуют аминокислоты, содержание которых в тканях значительно выше остальных — глутамат, аланин, аспартат и соответствующие им кетокислоты — α—кетоглутарат, пируват и оксалоацетат. Основным донором аминогруппы служит глутамат. Реакции трансаминирования играют большую роль в обмене аминокислот. Поскольку этот процесс обратим, ферменты аминотрансферазы функционируют как в процессах катаболизма, так и биосинтеза аминокислот. Трансаминирование — заключительный этап синтеза заменимых аминокислот из соответствующих α-кетокислот, если они в данный момент необходимы клеткам. В результате происходит перераспределение аминного азота в тканях организма. Трансаминирование — первая стадия дезаминирования большинства аминокислот, то есть начальный этап их катаболизма. Образующиеся при этом кетокислоты окисляются в ЦТК или используются для синтеза глюкозы и кетоновых тел. При трансаминировании общее количество аминокислот в клетке не меняется. В свою очередь дезаминирование аминокислот — реакция отщепления α-аминогруппы от аминокислоты, в результате чего образуется соответствующая α-кетокислота (безазотистый остаток) и выделяется молекула аммиака. Аммиак токсичен для ЦНС, поэтому в организме человека и млекопитающих он превращается в нетоксичное хорошо растворимое соединение — мочевину. В виде мочевины, а также в виде солей аммония аммиак выводится из организма. Безазотистый остаток используется для образования аминокислот в реакциях трансаминирования.
Трансаминирование — заключительный этап синтеза заменимых аминокислот из соответствующих α-кетокислот, если они в данный момент необходимы клеткам. В результате происходит перераспределение аминного азота в тканях организма. Трансаминирование — первая стадия дезаминирования большинства аминокислот, то есть начальный этап их катаболизма. Образующиеся при этом кетокислоты окисляются в ЦТК или используются для синтеза глюкозы и кетоновых тел. При трансаминировании общее количество аминокислот в клетке не меняется. В свою очередь дезаминирование аминокислот — реакция отщепления α-аминогруппы от аминокислоты, в результате чего образуется соответствующая α-кетокислота (безазотистый остаток) и выделяется молекула аммиака. Аммиак токсичен для ЦНС, поэтому в организме человека и млекопитающих он превращается в нетоксичное хорошо растворимое соединение — мочевину. В виде мочевины, а также в виде солей аммония аммиак выводится из организма. Безазотистый остаток используется для образования аминокислот в реакциях трансаминирования.
При катаболизме почти все природные аминокислоты сначала передают аминогруппу на а-кетоглутарат в реакции трансаминирования с образованием глутамата и соответствующей кетокислоты. Затем глутамат подвергается прямому окислительному дезаминированию под действием глутаматдегидрогеназы, в результате чего получаются а-кетоглутарат и аммиак. При необходимости синтеза аминокислот и наличии необходимых а-кетокислот обе стадии непрямого дезаминирования протекают в обратном направлении. В результате восстановительного аминирования а-кетоглутарата образуется глутамат, который вступает в трансаминирование с соответствующей а-кетокислотой, что приводит к синтезу новой аминокислоты. В случае использования белков в качестве источника энергии большинство аминокислот окисляются в конечном счёте через цикл лимонной кислоты до углекислого газа и воды. Прежде, чем эти вещества вовлекаются в заключительный этап катаболизма, их углеродный скелет превращается в двухуглеродный фрагмент в форме ацетил-КоА. Именно в этой форме большая часть молекул аминокислот включается в цикл лимонной кислоты.
Именно в этой форме большая часть молекул аминокислот включается в цикл лимонной кислоты.
Белки организма находятся в динамическом состоянии: из-за непрерывного процесса их разрушения и образования происходит обновление белков, скорость которого неодинакова для различных тканей. С наибольшей скоростью обновляются белки печени, слизистой оболочки кишечника, а также других внутренних органов и плазмы крови. Медленнее обновляются белки, входящие в состав клеток мозга, сердца, половых желез и еще медленнее — белки мышц, кожи и особенно опорных тканей (сухожилий, костей и хрящей). Важнейшими азотистыми продуктами распада белков, которые выделяются с мочой и потом, являются мочевина, мочевая кислота и аммиак. Преобладание в организме в данный момент времени синтеза или распада белка отражается понятием азотистого баланса — разностью между количеством азота, содержащегося в пище человека, и его уровнем в выделениях. Азотистым равновесием называют состояние, при котором количество выведенного азота равно количеству поступившего в организм. Азотистое равновесие наблюдается у здорового взрослого человека, если минимальное количество белков в пище соответствует 30-50 г/сут. Оптимальное количество поступления белка с пищей при средней физической нагрузке составляет около 100-120 г/сут. При положительном азотистом балансе количество азота в выделениях организма значительно меньше, чем содержание его в пище, то есть наблюдается задержка азота в организме. Положительный азотистый баланс отмечается у детей в связи с усиленным ростом, у женщин во время беременности, при усиленной спортивной тренировке, приводящей к увеличению мышечной массы, при заживлении обширных ран и при разрешении патологического процесса, связанного с выраженными системными нарушениями. Отрицательный азотистый баланс отмечается тогда, когда количество выделяющегося азота больше содержания его в пище, поступающей в организм. Отрицательный азотистый баланс наблюдается при белковом голодании, лихорадочных состояниях, нарушениях нейроэндокринной регуляции белкового обмена.
Азотистое равновесие наблюдается у здорового взрослого человека, если минимальное количество белков в пище соответствует 30-50 г/сут. Оптимальное количество поступления белка с пищей при средней физической нагрузке составляет около 100-120 г/сут. При положительном азотистом балансе количество азота в выделениях организма значительно меньше, чем содержание его в пище, то есть наблюдается задержка азота в организме. Положительный азотистый баланс отмечается у детей в связи с усиленным ростом, у женщин во время беременности, при усиленной спортивной тренировке, приводящей к увеличению мышечной массы, при заживлении обширных ран и при разрешении патологического процесса, связанного с выраженными системными нарушениями. Отрицательный азотистый баланс отмечается тогда, когда количество выделяющегося азота больше содержания его в пище, поступающей в организм. Отрицательный азотистый баланс наблюдается при белковом голодании, лихорадочных состояниях, нарушениях нейроэндокринной регуляции белкового обмена.
Некоторые аминокислоты не могут синтезироваться в организме человека и должны обязательно поступать с пищей в готовом виде. Эти аминокислоты принято называть незаменимыми, или эссенциальными. Экспериментально установлено, что из 20 входящих в состав белков аминокислот 12 синтезируются в организме (заменимые аминокислоты), а 8 не синтезируются (незаменимые аминокислоты) . К незаменимым аминоксилотам относятся: валин, метионин, треонин, лейцин, изолейцин, фенилаланин, триптофан и лизин. Две аминокислоты — аргинин и гистидин — у взрослых образуются в достаточных количествах, однако детям для нормального роста организма необходимо дополнительное поступление этих аминокислот с пищей. Поэтому их называют частично заменимыми. Две другие аминокислоты — тирозин и цистеин — условно заменимые, так как для их синтеза необходимы незаменимые аминокислоты. Тирозин синтезируется из фенилаланина, а для образования цистеина необходим атом серы метионина. Белки, содержащие весь необходимый набор аминокислот, называют биологически полноценными (табл. 1. 1. ). Наиболее высока биологическая ценность белков молока, яиц, рыбы, мяса. Биологически неполноценными называют белки, в составе которых отсутствует хотя бы одна аминокислота, которая не может быть синтезирована в организме. Неполноценными белками являются белки кукурузы, пшеницы, ячменя.
1. 1. ). Наиболее высока биологическая ценность белков молока, яиц, рыбы, мяса. Биологически неполноценными называют белки, в составе которых отсутствует хотя бы одна аминокислота, которая не может быть синтезирована в организме. Неполноценными белками являются белки кукурузы, пшеницы, ячменя.
Таблица 1. 1. Аминокислоты, входящие в состав белков человека.
|
1. Незаменимые Валин Лейцин Изолейцин Треонин Метионин Фенилаланин Триптофан Лизин |
2. Частично заменимые Гистидин Аргинин 3. Условно заменимые Цистеин Тирозин |
4. Заменимые Аланин Аспарагиновая кислота Аспарагин Глутаминовая кислота Глутамин Пролин Глицин Серин |
Жиры (липиды) по своей химической структуре представляют собой триглицериды — сложные эфиры глицерина и жирных кислот (табл. 1. 2). Изначально эти соединения были объединены в одну химическую группу по общему признаку растворимости: они не растворяются в воде, но растворяются в органических растворителях (эфир, спирт, бензол). Жиры делят на простые липиды (нейтральные жиры, воски), сложные липиды (фосфолипиды, гликолипиды, сульфолипиды) и стероиды (холестерин). Основная масса липидов представлена в организме человека нейтральными жирами — триглицеридами олеиновой, пальмитиновой, стеариновой, линолевой и линоленовой жирных кислот.
1. 2). Изначально эти соединения были объединены в одну химическую группу по общему признаку растворимости: они не растворяются в воде, но растворяются в органических растворителях (эфир, спирт, бензол). Жиры делят на простые липиды (нейтральные жиры, воски), сложные липиды (фосфолипиды, гликолипиды, сульфолипиды) и стероиды (холестерин). Основная масса липидов представлена в организме человека нейтральными жирами — триглицеридами олеиновой, пальмитиновой, стеариновой, линолевой и линоленовой жирных кислот.
Таблица 1. 2. Классификация липидов организма человека.
|
1. Гликолипиды. |
Содержат углеводный компонент. |
|
2. Жиры. |
Эфиры глицерина и высших жирных кислот. Химическое название — ацилглицерины. Преобладают триацилглицерины. |
|
3. |
Свободные жирные кислоты, жирорастворимые витамины, биологически активные вещества липидной природы — простагландины и др. |
|
4. Стероиды. |
В основе строения — полициклическая структура циклопентанпергидрофенантрен-стеран. |
|
А. Стерины (спирты). |
Наиболее важен холестерин. |
|
В. Стериды. |
Эфиры стеринов и высших жирных кислот. Наиболее распространены эфиры холестерина. |
|
5. Фосфолипипы. |
Отличительная особенность — остаток фосфорной кислоты в составе молекулы. |
 Минорные липиды.
Минорные липиды. Жиры растительного и животного происхождения имеют различный состав жирных кислот, определяющий их физические свойства и физиолого-биохимические эффекты. Жирные кислоты подразделяются на два основных класса — насыщенные и ненасыщенные. Насыщенность жира определяется количеством атомов водорода, которое содержит каждая жирная кислота (или, иначе, количеством двойных связей С=С). Жирные кислоты со средней длиной цепи (С8-С14) способны усваиваться в пищеварительном тракте без участия желчных кислот и панкреатической липазы, не депонируются в печени и подвергаются β-окислению. Животные жиры могут содержать насыщенные жирные кислоты с длиной цепи до двадцати и более атомов углерода, они имеют твердую консистенцию и высокую температуру плавления. Как известно высокое потребление насыщенных жирных кислот является важнейшим фактором риска развития диабета, ожирения, атеросклероза. К мононенасыщенным жирным кислотам относятся миристолеиновая и пальмитолеиновая кислоты (жиры рыб и морских млекопитающих), олеиновая (оливковое, сафлоровое, кунжутное, рапсовое масла). Мононенасыщенные жирные кислоты помимо их поступления с пищей в организме синтезируются из насыщенных жирных кислот и частично из углеводов.
Жирные кислоты подразделяются на два основных класса — насыщенные и ненасыщенные. Насыщенность жира определяется количеством атомов водорода, которое содержит каждая жирная кислота (или, иначе, количеством двойных связей С=С). Жирные кислоты со средней длиной цепи (С8-С14) способны усваиваться в пищеварительном тракте без участия желчных кислот и панкреатической липазы, не депонируются в печени и подвергаются β-окислению. Животные жиры могут содержать насыщенные жирные кислоты с длиной цепи до двадцати и более атомов углерода, они имеют твердую консистенцию и высокую температуру плавления. Как известно высокое потребление насыщенных жирных кислот является важнейшим фактором риска развития диабета, ожирения, атеросклероза. К мононенасыщенным жирным кислотам относятся миристолеиновая и пальмитолеиновая кислоты (жиры рыб и морских млекопитающих), олеиновая (оливковое, сафлоровое, кунжутное, рапсовое масла). Мононенасыщенные жирные кислоты помимо их поступления с пищей в организме синтезируются из насыщенных жирных кислот и частично из углеводов. Жирные кислоты с двумя и более двойными связями между углеродными атомами называются полиненасыщенными – ПНЖК. Особое значение для организма человека имеют такие ПНЖК как линолевая, линоленовая, являющиеся структурными элементами клеточных мембран и обеспечивающие нормальное развитие и адаптацию организма человека к неблагоприятным факторам окружающей среды. ПНЖК являются предшественниками образующихся из них биорегуляторов – эйкозаноидов. Двумя основными группами ПНЖК являются кислоты семейств ω-6 и ω-3. Жирные кислоты ω-6 содержатся практически во всех растительных маслах и орехах. ω-3 жирные кислоты также содержатся в ряде масел (льняном, из семян крестоцветных, соевом). Основным пищевым источником ω-3 жирных кислот являются жирные сорта рыб и некоторые морепродукты. Из ПНЖК ω — 6 особое место занимает линолевая кислота, которая является предшественником наиболее физиологически активной кислоты этого семейства — арахидоновой. Арахидоновая кислота является преобладающим представителем ПНЖК в организме человека и служит субстратом для синтеза простагландинов и лейкотриенов.
Жирные кислоты с двумя и более двойными связями между углеродными атомами называются полиненасыщенными – ПНЖК. Особое значение для организма человека имеют такие ПНЖК как линолевая, линоленовая, являющиеся структурными элементами клеточных мембран и обеспечивающие нормальное развитие и адаптацию организма человека к неблагоприятным факторам окружающей среды. ПНЖК являются предшественниками образующихся из них биорегуляторов – эйкозаноидов. Двумя основными группами ПНЖК являются кислоты семейств ω-6 и ω-3. Жирные кислоты ω-6 содержатся практически во всех растительных маслах и орехах. ω-3 жирные кислоты также содержатся в ряде масел (льняном, из семян крестоцветных, соевом). Основным пищевым источником ω-3 жирных кислот являются жирные сорта рыб и некоторые морепродукты. Из ПНЖК ω — 6 особое место занимает линолевая кислота, которая является предшественником наиболее физиологически активной кислоты этого семейства — арахидоновой. Арахидоновая кислота является преобладающим представителем ПНЖК в организме человека и служит субстратом для синтеза простагландинов и лейкотриенов.
Источниками жира в организме являются экзогенный жир, поступающий с пищей, и эндогенный жир, синтезируемый в печени из углеводов. Жир, всасывающийся из кишечника, поступает преимущественно в лимфу и в меньшем количестве — непосредственно в кровь. Большая часть жиров в организме находится в жировой ткани, меньшая часть входит в состав клеточных структур. В жировой ткани жир, находящийся в клетке в виде включений, легко выявляется при микроскопическом и гистохимическом исследованиях. Жировые вакуоли в клетках — это резервный жир, используемый для обеспечения прежде всего энергетических потребностей клетки. Больше всего запасного жира содержится в жировой ткани, а также в некоторых органах, например в печени и мышцах. Количество запасного жира зависит от характера питания, количества пищи, конституциональных особенностей, а также от величины расхода энергии при мышечной деятельности; количество же протоплазматического жира является устойчивым и постоянным. В жировой ткани нейтральный жир депонируется виде триглицеридов. Сложные липиды — фосфолипиды и гликолипиды — входят в состав всех клеток, но в большей степени в состав клеток нервной ткани. Общее количество жира в организме человека колеблется в широких пределах и в среднем составляет 10—20% от массы тела, а в случае патологического ожирения может достигать 50%. Суточная потребность взрослого человека в нейтральном жире составляет 70—80 г. У человека состав и свойства жира относительно постоянны. При употреблении пищи, содержащей даже небольшое количество жира, в теле человека жир все же откладывается в депо. При этом эндогенный жир имеет некоторые видовые особенности, однако видовая специфичность жиров выражена несравнимо меньше, чем видовая специфичность белков.
Сложные липиды — фосфолипиды и гликолипиды — входят в состав всех клеток, но в большей степени в состав клеток нервной ткани. Общее количество жира в организме человека колеблется в широких пределах и в среднем составляет 10—20% от массы тела, а в случае патологического ожирения может достигать 50%. Суточная потребность взрослого человека в нейтральном жире составляет 70—80 г. У человека состав и свойства жира относительно постоянны. При употреблении пищи, содержащей даже небольшое количество жира, в теле человека жир все же откладывается в депо. При этом эндогенный жир имеет некоторые видовые особенности, однако видовая специфичность жиров выражена несравнимо меньше, чем видовая специфичность белков.
Основная биологическая роль жиров — обеспечение пластического и энергетического обмена в организме. Пластическая роль липидов состоит в том, что они входят в состав клеточных мембран, в значительной мере определяя их свойства. Фосфатиды и стерины входят в состав клеточных структур, в частности клеточных мембран, а также ядерного вещества и цитоплазмы. Исключительно важное физиологическое значение имеют стерины, в частности холестерин. Это вещество входит в состав клеточных мембран, является источником образования желчных кислот, а также гормонов коры надпочечников и половых желез, витамина D. Печень является практически единственным органом, поддерживающим уровень фосфолипидов в крови и местом синтеза эндогенного холестерина. В плазме крови холестерин находится в составе липопротеидных комплексов, с помощью которых и осуществляется его транспорт. У взрослых людей 67—70% холестерина плазмы крови находится в составе липопротеидов низкой плотности (ЛПНП), 9—10% — в составе липопротеидов очень низкой плотности (ЛПОНП) и 20—24% — в составе липопротеидов высокой плотности (ЛПВП). Давно доказано, что именно липопротеиды определяют уровень холестерина и динамику его обмена.
Исключительно важное физиологическое значение имеют стерины, в частности холестерин. Это вещество входит в состав клеточных мембран, является источником образования желчных кислот, а также гормонов коры надпочечников и половых желез, витамина D. Печень является практически единственным органом, поддерживающим уровень фосфолипидов в крови и местом синтеза эндогенного холестерина. В плазме крови холестерин находится в составе липопротеидных комплексов, с помощью которых и осуществляется его транспорт. У взрослых людей 67—70% холестерина плазмы крови находится в составе липопротеидов низкой плотности (ЛПНП), 9—10% — в составе липопротеидов очень низкой плотности (ЛПОНП) и 20—24% — в составе липопротеидов высокой плотности (ЛПВП). Давно доказано, что именно липопротеиды определяют уровень холестерина и динамику его обмена.
Энергетическая роль жиров определяется их максимальной среди всех биологических молекул энергоемкостью, более чем в два раза превышающую таковую углеводов или белков. При окислении 1 г жира выделяется 37, 7 кДж (9, 0 ккал) энергии. В отличие от углеводов жиры составляют энергетический резерв организма. Преимущество жира в качестве энергетического резерва заключается в том, что жиры являются более восстановленными веществами по сравнению с углеводами (в молекулах углеводов при каждом углеродном атоме есть кислород — группы -CHOH-; у жира имеются длинные углеводородные радикалы, в которых преобладают группы -Ch3- — в них нет кислорода). От жира можно отнять больше водорода, который затем проходит по цепи митохондриального окисления с образованием АТФ. Еще одним преимуществом жира как энергетического резерва, в отличие от углеводов, является гидрофобность — он не связан с водой. Это обеспечивает компактность жировых запасов — они хранятся в безводной форме, занимая малый объем. В среднем, у человека запас чистых триацилглицеринов составляет примерно 13 кг. Этих запасов могло бы хватить на 40 дней голодания в условиях умеренной физической нагрузки. Для сравнения: общие запасы гликогена в организме — примерно 400 г; при голодании этого количества не хватает даже на одни сутки.
При окислении 1 г жира выделяется 37, 7 кДж (9, 0 ккал) энергии. В отличие от углеводов жиры составляют энергетический резерв организма. Преимущество жира в качестве энергетического резерва заключается в том, что жиры являются более восстановленными веществами по сравнению с углеводами (в молекулах углеводов при каждом углеродном атоме есть кислород — группы -CHOH-; у жира имеются длинные углеводородные радикалы, в которых преобладают группы -Ch3- — в них нет кислорода). От жира можно отнять больше водорода, который затем проходит по цепи митохондриального окисления с образованием АТФ. Еще одним преимуществом жира как энергетического резерва, в отличие от углеводов, является гидрофобность — он не связан с водой. Это обеспечивает компактность жировых запасов — они хранятся в безводной форме, занимая малый объем. В среднем, у человека запас чистых триацилглицеринов составляет примерно 13 кг. Этих запасов могло бы хватить на 40 дней голодания в условиях умеренной физической нагрузки. Для сравнения: общие запасы гликогена в организме — примерно 400 г; при голодании этого количества не хватает даже на одни сутки.
Катаболизм жира включает в себя три этапа: 1) гидролиз жира до глицерина и жирных кислот (липолиз) ; 2) трансформация глицерина с последующим вступлением продуктов в гексозобифосфатный путь, а также окисление жирных кислот до ацетил-КоА; 3) вступление вышеуказанных продуктов в цикл трикарбоновых кислот. Кроме указанных этапов к катаболизму жиров относят также окисление кетоновых тел и перекисное окисление липидов. Обмен полученного в результате липолиза глицерина может осуществляться несколькими путями. Значительная часть образовавшегося при гидролизе липидов глицерина используется для ресинтеза триглицеридов. Второй путь обмена глицерина — включение продукта его окисления в гликолиз или в глюконеогенез. Окисление жирных кислот осуществляется различными путями, наиболее значимым из них является β-окисление. В ходе β-окисления последовательно происходит активация жирной кислоты на мембране митохондрии и ее связывание с молекулой карнитина, прохождение комплекса нв внутреннюю поверхность мембраны митохондрии, внутримитохондриальное окисление жирной кислоты с образованием ацетил-КоА и АТФ.
Одним из продуктов катаболизма жиров, имеющем важное значения для метаболизма в целом являются кетоновые тела. Кетоновые тела — группа органических соединений, являющихся промежуточными продуктами жирового, углеводного и белкового обменов. К кетоновым телам относят β-оксимасляную и ацетоуксусную кислоты и ацетон, имеющие сходное строение и способные к взаимопревращениям. Главным путем синтеза кетоновых тел, происходящего в основном в печени, считается реакция конденсации между двумя молекулами ацетил-КоА, образовавшегося при β-окислении жирных кислот или при окислительном декарбоксилировании пирувата (пировиноградной кислоты) в процессе обмена глюкозы и ряда аминокислот. Данный путь синтеза кетоновых тел более других зависит от характера питания и в большей степени страдает при патологических нарушениях обмена веществ. Из печени кетоновые тела поступают в кровь и с нею во все остальные органы и ткани, где они включаются в универсальный энергообразующий цикл — цикл трикарбоновых кислот, в котором окисляются до углекислоты и воды. Кетоновые тела используются также для синтеза холестерина, высших жирных кислот, фосфолипидов и заменимых аминокислот. При голодании, однообразном безуглеводистом питании и при недостаточной секреции инсулина использование ацетил-КоА в цикле трикарбоновых кислот подавляется, так как все метаболически доступные ресурсы организма превращаются в глюкозу крови. В этих условиях увеличивается синтез кетоновых тел. Следует подчеркнуть важную роль кетоновых тел в поддержании энергетического баланса. Кетоновые тела – поставщики «топлива» для мышц, почек и действуют, возможно, как часть регуляторного механизма с обратной связью, предотвращая чрезвычайную мобилизацию жирных кислот из жировых депо. Печень в этом смысле является исключением, она не использует кетоновые тела в качестве энергетического материала.
Кетоновые тела используются также для синтеза холестерина, высших жирных кислот, фосфолипидов и заменимых аминокислот. При голодании, однообразном безуглеводистом питании и при недостаточной секреции инсулина использование ацетил-КоА в цикле трикарбоновых кислот подавляется, так как все метаболически доступные ресурсы организма превращаются в глюкозу крови. В этих условиях увеличивается синтез кетоновых тел. Следует подчеркнуть важную роль кетоновых тел в поддержании энергетического баланса. Кетоновые тела – поставщики «топлива» для мышц, почек и действуют, возможно, как часть регуляторного механизма с обратной связью, предотвращая чрезвычайную мобилизацию жирных кислот из жировых депо. Печень в этом смысле является исключением, она не использует кетоновые тела в качестве энергетического материала.
Процесс образования, отложения и мобилизации из депо жира регулируется нервной и эндокринной системами, а также тканевыми механизмами и тесно связаны с углеводным обменом. Так, повышение концентрации глюкозы в крови уменьшает распад триглицеридов и активизирует их синтез. Понижение концентрации глюкозы в крови, наоборот, тормозит синтез триглицеридов и усиливает их расщепление. Таким образом, взаимосвязь жирового и углеводного обменов направлена на обеспечение энергетических потребностей организма. При избытке углеводов в пище триглицериды депонируются в жировой ткани, при нехватке углеводов происходит расщепление триглицеридов с образованием неэтерифицнрованных жирных кислот, служащих источником энергии. В обмене жиров одна из важнейших ролей принадлежит печени. Печень — основной орган, в котором происходит образование кетоновых тел (бета-оксимасляная, ацетоуксусная кислоты, ацетон), используемых как альтернативный глюкозе источник энергии.
Понижение концентрации глюкозы в крови, наоборот, тормозит синтез триглицеридов и усиливает их расщепление. Таким образом, взаимосвязь жирового и углеводного обменов направлена на обеспечение энергетических потребностей организма. При избытке углеводов в пище триглицериды депонируются в жировой ткани, при нехватке углеводов происходит расщепление триглицеридов с образованием неэтерифицнрованных жирных кислот, служащих источником энергии. В обмене жиров одна из важнейших ролей принадлежит печени. Печень — основной орган, в котором происходит образование кетоновых тел (бета-оксимасляная, ацетоуксусная кислоты, ацетон), используемых как альтернативный глюкозе источник энергии.
При обильном углеводном питании и отсутствии жиров в пище синтез жира в организме может происходить из углеводов. Источником углерода для синтеза жирных кислот служит ацетил-КоА, образующийся при распаде глюкозы в абсорбтивном периоде. В норме у человека 25—30% углеводов пищи превращается в жиры. Превращение белка в жирные кислоты происходит, вероятнее всего, также через образование углеводов. С другой стороны и нейтральные жиры в энергетическом отношении могут быть заменены углеводами. Тем не менее жиры необходимы для нормальной жизнедеятельности. Известно, что длительное исключение жиров из пищевого рациона может явиться причиной возникновения целого ряда тяжелых метаболических нарушений. Отчасти это связано с отсутствием поступления в организм жирорастворимых витаминов (A, D, E, K). Но основная причина метаболических нарушений кроется в возникновении в организме дефицита незаменимых жирных кислот. Некоторые ненасыщенные жирные кислоты (с числом двойных связей более 1), например линолевая, линоленовая и арахидоновая, в организме человека и некоторых животных не образуются из других жирных кислот и поэтому являются незаменимыми. Особенно остро реагирует организм на дефицит незаменимой линолевой кислоты СН3- (СН2) 4 — СН = СН — СН2 — СН = СН — (СН2) 7 — СООН. Возможно это связано с тем, что эта ненасыщенная жирная кислота в организме человека служит предшественником арахидоновой кислоты, которая в свою очередь необходима для синтеза универсальных биорегуляторов — простагландинов.
С другой стороны и нейтральные жиры в энергетическом отношении могут быть заменены углеводами. Тем не менее жиры необходимы для нормальной жизнедеятельности. Известно, что длительное исключение жиров из пищевого рациона может явиться причиной возникновения целого ряда тяжелых метаболических нарушений. Отчасти это связано с отсутствием поступления в организм жирорастворимых витаминов (A, D, E, K). Но основная причина метаболических нарушений кроется в возникновении в организме дефицита незаменимых жирных кислот. Некоторые ненасыщенные жирные кислоты (с числом двойных связей более 1), например линолевая, линоленовая и арахидоновая, в организме человека и некоторых животных не образуются из других жирных кислот и поэтому являются незаменимыми. Особенно остро реагирует организм на дефицит незаменимой линолевой кислоты СН3- (СН2) 4 — СН = СН — СН2 — СН = СН — (СН2) 7 — СООН. Возможно это связано с тем, что эта ненасыщенная жирная кислота в организме человека служит предшественником арахидоновой кислоты, которая в свою очередь необходима для синтеза универсальных биорегуляторов — простагландинов. Основными пищевыми источниками полиненасыщенных жирных кислот, в том числе линолевой, являются растительные масла.
Основными пищевыми источниками полиненасыщенных жирных кислот, в том числе линолевой, являются растительные масла.
Как указывалось выше метаболизм жиров контролируется нервной и эндокринной системами. Мобилизация жиров из депо происходит под влиянием гормонов мозгового слоя надпочечников — адреналина и норадреналина. Соматотропный гормон гипофиза также обладает жиромобилизирующим действием. Аналогично действует тироксин — гормон щитовидной железы. Тормозят мобилизацию жира глюкокортикоиды — гормоны коркового слоя надпочечника, вероятно, вследствие того, что они несколько повышают уровень глюкозы в крови. Действие инсулина связано с повышением активности внутриклеточной фосфодиэстеразы, что приводит к снижению концентрации цАМФ и угнетению липолиза. Таким образом, инсулин усиливает синтез жира и уменьшает скорость его мобилизации. Имеются данные, свидетельствующие о возможности прямых нервных влияний на обмен жиров. Симпатические влияния тормозят синтез триглицеридов и усиливают их распад. Парасимпатические влияния, напротив, способствуют отложению жира в депо.
Парасимпатические влияния, напротив, способствуют отложению жира в депо.
Статья добавлена 31 мая 2016 г.
ЧАСТЬ 1 / КонсультантПлюс
ЧАСТЬ 1
|
Ответом к заданиям 1 — 36 является одна цифра, которая соответствует номеру правильного ответа. Запишите эту цифру в поле ответа в тексте работы. |
|
1 |
Развитие организма животного от момента образования зиготы до рождения изучает наука |
1) генетика
2) физиология
3) экология
4) эмбриология
|
2 |
Организмы растений, животных, грибов и бактерий состоят из клеток, что свидетельствует о |
1) единстве органического мира
2) разнообразии строения живых организмов
3) связи организмов со средой обитания
4) сложном строении живых организмов
|
3 |
На мембранах каких органоидов клетки располагаются рибосомы? |
1) хлоропластов
2) комплекса Гольджи
3) лизосом
4) эндоплазматической сети
|
4 |
Одна интерфаза и два следующих друг за другом деления характерны для процесса |
1) оплодотворения
2) дробления зиготы
3) мейоза
4) митоза
|
5 |
Какая форма жизни занимает промежуточное положение между телами живой и неживой природы? |
1) вирусы
2) бактерии
3) лишайники
4) грибы
|
6 |
Каково значение вегетативного размножения в природе? |
1) способствует быстрому увеличению численности особей
2) ведет к появлению комбинативной изменчивости
3) увеличивает численность гетерозиготных особей
4) приводит к разнообразию особей в популяции
|
7 |
Парные гены, расположенные в гомологичных хромосомах и определяющие окраску цветков гороха, называют |
1) сцепленными
2) рецессивными
3) аллельными
4) доминантными
|
8 |
Гомозиготные доминантные серые овцы при переходе на грубые корма гибнут, а гетерозиготные выживают. |
 Определите генотип жизнеспособной особи по данному признаку.
Определите генотип жизнеспособной особи по данному признаку.1) AA
2) Aa
3) aa
4) AB
|
9 |
Полиплоидия — одна из форм изменчивости |
1) модификационной
2) мутационной
3) комбинативной
4) соотносительной
|
10 |
Грибы, по сравнению с бактериями, имеют более высокий уровень организации, так как |
1) их клетки имеют плотную оболочку
2) по способу питания они являются гетеротрофами
3) их клетки имеют оформленное ядро
4) их споры переносят неблагоприятные условия
|
11 |
Через устьица растений происходит |
1) газообмен
2) транспорт минеральных солей
3) транспорт органических веществ
4) выделение тепла
|
12 |
Главный признак, по которому цветковые растения относят к одному классу, |
1) строение плода
2) строение семени
3) способ размножения
4) совместное обитание
|
13 |
Какую функцию выполняют органоиды, обозначенные вопросительным знаком на схеме строения инфузории-туфельки? |
Рисунок (не приводится) |
1) переваривание пищевых частиц
2) удаление остатков непереваренной пищи
3) образование органических веществ на свету
4) удаление избытка воды и растворенных в ней ненужных веществ
|
14 |
У животных какого класса появился второй круг кровообращения? |
1) Костные рыбы
2) Птицы
3) Пресмыкающиеся
4) Земноводные
|
15 |
Сходство нервной и мышечной тканей состоит в том, что они обладают свойством |
1) сократимости
2) проводимости
3) возбудимости
4) воспроизведения
|
16 |
Давление крови на стенки сосудов создается силой сокращения |
1) предсердий сердца
2) желудочков сердца
3) створчатых клапанов
4) популунных клапанов
|
17 |
Способностью захватывать и переваривать инородные вещества и микроорганизмы в организме человека обладают |
1) антитела
2) фагоциты
3) эритроциты
4) тромбоциты
|
18 |
Какой орган в организме человека одновременно выполняет функции желез внешней и внутренней секреции? |
1) печень
2) гипофиз
3) щитовидная железа
4) поджелудочная железа
|
19 |
Одна из причин близорукости — |
1) помутнение хрусталика
2) повреждение зрительного нерва
3) уменьшение способности хрусталика изменять кривизну
4) нарушение в зрительной зоне коры больших полушарий
|
20 |
Микроэволюция приводит к изменению |
1) видов
2) родов
3) семейств
4) отрядов
|
21 |
Фактор эволюции, увеличивающий неоднородность особей в популяции, — это |
1) мутационная изменчивость
2) модификационная изменчивость
3) борьба за существование
4) массовый отбор
|
22 |
Виды-двойники отличаются друг от друга, главным образом, по критерию |
1) морфологическому
2) экологическому
3) генетическому
4) географическому
|
23 |
Череп современного человека отличается от черепа человекообразных обезьян наличием |
1) носовых отверстий
2) подбородочного выступа
3) височных костей
4) лицевого отдела
|
24 |
Какой фактор в наибольшей степени ограничивает распространение растений в Мировом океане? |
1) недостаток кислорода
2) скорость течения
3) недостаток света
4) высокая концентрация солей
|
25 |
Водоросли — важный компонент водной экосистемы, так как они |
1) способствуют очищению воды
2) выполняют роль продуцентов
3) способствуют зарастанию водоема
4) используют органические вещества в процессе дыхания
|
26 |
Благодаря какому процессу количество кислорода в биосфере остается относительно постоянной величиной? |
1) обмен веществ
2) круговорот веществ
3) смена экосистем
4) гниение
|
27 |
Синтез молекул АТФ происходит в |
1) эндоплазматической сети и рибосомах
2) комплексе Гольджи и лизосомах
3) хлоропластах и митохондриях
4) клеточном центре и вакуолях
|
28 |
Расщепление макромолекул, их подготовка к дальнейшим химическим превращениям в процессе метаболизма происходят в |
1) лизосомах
2) рибосомах
3) хлоропластах
4) митохондриях
|
29 |
Для какой фазы мейоза характерно расположение в экваториальной плоскости клетки спаренных гомологичных хромосом? |
1) профазы первого деления
2) профазы второго деления
3) метафазы первого деления
4) метафазы второго деления
|
30 |
Если исследуемые гены расположены в разных парах гомологичных хромосом, то проявляется |
1) закон независимого наследования признаков
2) единообразие гибридов
3) закон сцепленного наследования
4) неполное доминирование
|
31 |
Значение клеточной инженерии, в отличие от классических методов селекции растений, состоит в том, что она |
1) сокращает срок создания сорта
2) значительно ускоряет рост вегетативных органов
3) усиливает пластический обмен
4) повышает жизнеспособность потомства
|
32 |
Древние пресмыкающиеся смогли широко распространиться по суше, так как в процессе эволюции у них впервые сформировались |
1) два круга кровообращения и трехкамерное сердце
2) яйца с большим запасом питательных веществ
3) пять отделов головного мозга
4) рычажные конечности
|
33 |
У человека кровь, насыщенная углекислым газом, поступает в сердце по |
1) легочным артериям
2) легочным венам
3) артериям большого круга
4) венам большого круга
|
34 |
В организме человека нервные импульсы возникают в клетках |
1) скелетных мышц
2) эпидермиса кожи
3) эндокринных желез
4) чувствительных нейронов
|
35 |
Чтобы ограничить рост численности популяции растительноядных рыб в водоеме, нужно |
1) подкармливать растения минеральными удобрениями
2) провести очистку водоема
3) привлечь растительноядных птиц
4) заселить водоем хищными рыбами
|
36 |
Верны ли следующие суждения об особенностях клеточной организации? |
А.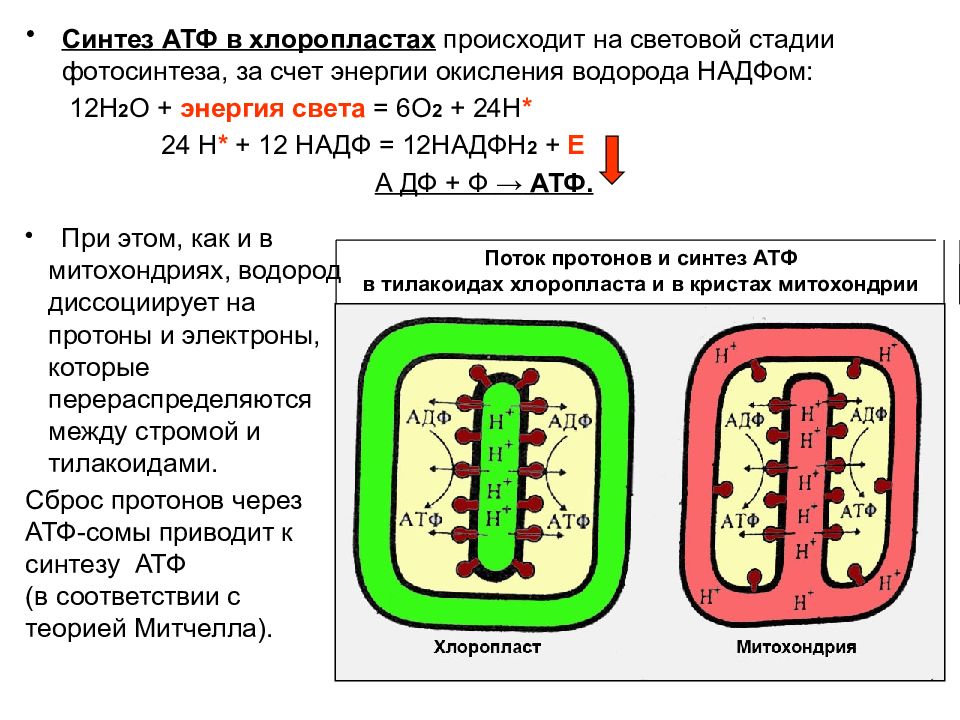 Отличительной особенностью эукариотической клетки служит наличие в ее цитоплазме нуклеоидов.
Отличительной особенностью эукариотической клетки служит наличие в ее цитоплазме нуклеоидов.
Б. У эукариотических и прокариотических клеток имеется цитоплазма, содержащая митохондрии.
1) верно только А
2) верно только Б
3) верны оба суждения
4) оба суждения неверны
|
В заданиях 37 — 39 выберите три верных ответа из шести. Запишите в таблицу цифры, соответствующие выбранным ответам. |
|
37 |
Какие методы используют при изучении наследственности человека? |
1) близнецовый
2) гибридологический
3) эволюционный
4) генеалогический
5) цитогенетический
6) экспериментальный
|
38 |
Какие из перечисленных признаков характерны как для голосеменных, так и для покрытосеменных растений? |
1) образование триплоидного эндосперма
2) развитая проводящая система
3) наличие плодов
4) наличие семян
5) наличие травянистых форм
6) формирование пыльцы
|
39 |
Какие из перечисленных видов претерпевают в настоящее время биологический прогресс? |
1) зубр
2) лошадь Пржевальского
3) домовый воробей
4) амурский тигр
5) пырей ползучий
6) одуванчик обыкновенный
|
В заданиях 40, 41 к каждому элементу первого столбца подберите соответствующий элемент из второго столбца и запишите в таблицу выбранные цифры под соответствующими буквами. |
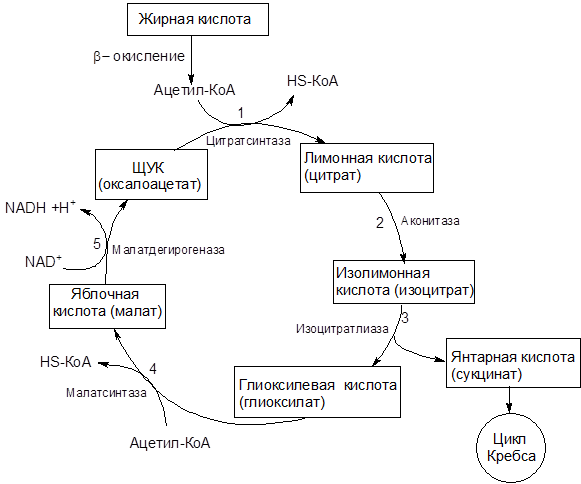
|
40 |
Установите соответствие между признаком животного и классом позвоночных животных, к которому он принадлежит. |
|
ПРИЗНАК ЖИВОТНОГО |
КЛАСС ПОЗВОНОЧНЫХ |
|||
|
А) |
наличие клоаки |
1) |
Птицы |
|
|
Б) |
появление диафрагмы |
2) |
Млекопитающие (плацентарные) |
|
|
В) |
альвеолярное строение легких |
|||
|
Г) |
наличие крупных яйцеклеток |
|||
|
Д) |
образование потовых желез |
|||
|
Е) |
отсутствие мочевого пузыря |
|||
|
Ответ: |
А |
Б |
В |
Г |
Д |
Е |
|
|
41 |
Установите соответствие между функцией и форменными элементами крови, которые ее выполняют. |

|
ФУНКЦИЯ |
ФОРМЕННЫЕ ЭЛЕМЕНТЫ КРОВИ |
|||
|
А) |
участвуют в свертывании крови |
1) |
эритроциты |
|
|
Б) |
транспортируют кислород из легких в ткани |
2) |
лейкоциты |
|
|
В) |
защищают организм от инфекций |
3) |
тромбоциты |
|
|
Г) |
участвуют в синтезе антител |
|||
|
Д) |
транспортируют оксигемоглобин |
|||
|
Ответ: |
А |
Б |
В |
Г |
Д |
|
|
В задании 42 установите последовательность биологических процессов, явлений, практических действий. |
 Запишите цифры, которыми обозначены биологические процессы, явления, практические действия, в правильной последовательности в таблицу.
Запишите цифры, которыми обозначены биологические процессы, явления, практические действия, в правильной последовательности в таблицу.|
42 |
Установите последовательность процессов круговорота углерода в биосфере, начиная с поглощения растениями углекислого газа. |
1) окисление органических веществ при дыхании до углекислого газа и воды
2) поглощение углекислого газ из атмосферы продуцентами
3) синтез углеводов в процессе фотосинтеза
4) использование первичного органического вещества всеми организмами
Синтез углеводов — Биохимия
Объяснение: Пентозофосфатный путь (ППФ) — это метаболический путь в клетках, который используется для выработки НАДФН и/или рибозо-5-фосфата для использования в клетке, в зависимости от потребностей клетки. НАДФН используется в первую очередь для обеспечения восстановительной способности для нескольких биосинтетических реакций, но он также служит средством сохранения глутатиона преимущественно в его восстановленной форме в клетке. Это, в свою очередь, помогает поддерживать восстановительную среду внутри клеток.Кроме того, рибозо-5-фосфат используется в качестве основного предшественника для синтеза нуклеотидов.
Это, в свою очередь, помогает поддерживать восстановительную среду внутри клеток.Кроме того, рибозо-5-фосфат используется в качестве основного предшественника для синтеза нуклеотидов.
NADH и FADH 2 вырабатываются не PPP, а путем окисления глюкозы через аэробный дыхательный путь. Эти две молекулы являются переносчиками высокоэнергетических электронов, которые используются для генерации АТФ через цепь переноса электронов.
Глутатион, как упоминалось ранее, не производится PPP; однако он использует NADPH, продуцируемый PPP, для поддержания своей восстановленной формы внутри клетки, что, в свою очередь, поддерживает преимущественно восстанавливающую среду внутри клетки.2,3-бисфосфоглицерат является промежуточным продуктом гликолиза, а не PPP. Одной из основных функций 2,3-BPG является связывание гемоглобина и снижение его сродства к O 2 . Это позволяет эритроцитам легче высвобождать O 2 в ткани, которые в этом нуждаются.
Фруктозо-2,6-бисфосфат не является продуктом PPP. Скорее, он образуется в результате побочной реакции гликолитического промежуточного продукта фруктозо-6-фосфата. Фруктозо-2,6-бисфосфат служит аллостерическим регулятором фермента фруктозо-1,6-бисфосфатазы, который является важным регуляторным ферментом гликолиза и глюконеогенеза.Гормоны, такие как инсулин и глюкагон, могут стимулировать клетки к изменению концентрации фруктозо-2,6-бисфосфата, который, в свою очередь, регулирует активность гликолиза и глюконеогенеза. Глицерин-3-фосфат также не производится из PPP. Скорее, он может быть получен в результате фосфорилирования глицерина или восстановления дигидроксиацетонфосфата, промежуточного продукта гликолиза. Он используется в качестве основы для образования триглицеридов и фосфолипидов.
Скорее, он образуется в результате побочной реакции гликолитического промежуточного продукта фруктозо-6-фосфата. Фруктозо-2,6-бисфосфат служит аллостерическим регулятором фермента фруктозо-1,6-бисфосфатазы, который является важным регуляторным ферментом гликолиза и глюконеогенеза.Гормоны, такие как инсулин и глюкагон, могут стимулировать клетки к изменению концентрации фруктозо-2,6-бисфосфата, который, в свою очередь, регулирует активность гликолиза и глюконеогенеза. Глицерин-3-фосфат также не производится из PPP. Скорее, он может быть получен в результате фосфорилирования глицерина или восстановления дигидроксиацетонфосфата, промежуточного продукта гликолиза. Он используется в качестве основы для образования триглицеридов и фосфолипидов.
Ацетоацетат и бета-гидроксибутират являются кетоновыми телами, образующимися не в результате ППС, а в результате конденсации двух молекул ацетил-КоА плюс дополнительные модификации.Как правило, когда организм находится в состоянии голодания и ему необходимо резервировать уровень глюкозы в крови, могут вырабатываться кетоновые тела, которые действуют как альтернативный источник энергии, что позволяет в основном экономить глюкозу.
Биосинтез компонентов клетки — Клетка
В предыдущем разделе были рассмотрены основные метаболические реакции, с помощью которых клетка получает и сохраняет энергию в форме АТФ. Затем эта метаболическая энергия используется для выполнения различных задач, включая синтез макромолекул и других компонентов клетки.Таким образом, энергия, полученная в результате распада органических молекул (катаболизм), используется для запуска синтеза других необходимых для клетки компонентов. Большинство катаболических путей включают окисление органических молекул в сочетании с выработкой как энергии (АТФ), так и восстановительной способности (НАДН). Напротив, биосинтетические (анаболические) пути обычно включают использование как АТФ, так и восстановительной способности (обычно в форме НАДФН) для производства новых органических соединений. В предыдущем разделе обсуждался один из основных путей биосинтеза — синтез углеводов из СО 2 и Н 2 О во время темновых реакций фотосинтеза. Дополнительные пути, ведущие к биосинтезу основных клеточных компонентов (углеводов, липидов, белков и нуклеиновых кислот), рассматриваются в следующих разделах.
Дополнительные пути, ведущие к биосинтезу основных клеточных компонентов (углеводов, липидов, белков и нуклеиновых кислот), рассматриваются в следующих разделах.
Углеводы
Глюкоза может быть получена непосредственно из пищи или получена в результате фотосинтеза и может быть синтезирована из других органических молекул. В клетках животных синтез глюкозы (глюконеогенез) обычно начинается с лактата (образуется в результате анаэробного гликолиза), аминокислот (образуется при распаде белков) или глицерина (образуется при распаде липидов).Растения (но не животные) также способны синтезировать глюкозу из жирных кислот — процесс, который особенно важен во время прорастания семян, когда энергия, накопленная в виде жиров, должна быть преобразована в углеводы для поддержания роста растения. Как в животных, так и в растительных клетках простые сахара полимеризуются и запасаются в виде полисахаридов.
Глюконеогенез включает превращение пирувата в глюкозу, по сути, обратное гликолизу.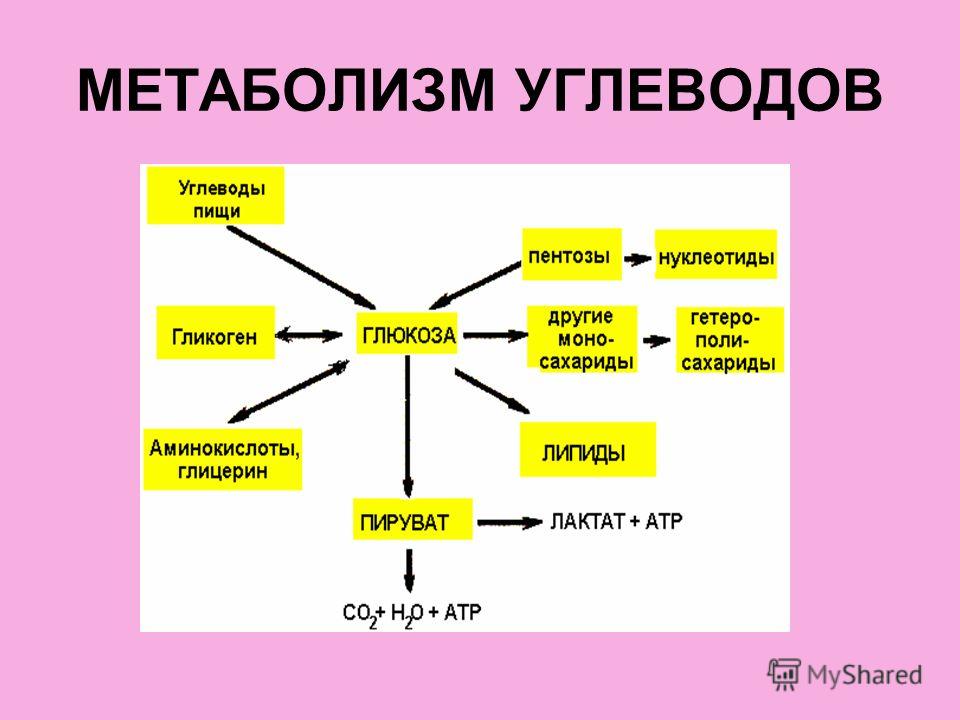 Однако, как обсуждалось ранее, гликолитическое превращение глюкозы в пируват представляет собой энергетический путь, при котором образуется по две молекулы АТФ и НАДН.Хотя некоторые реакции гликолиза легко обратимы, другие будут протекать только в направлении распада глюкозы, поскольку связаны с большим уменьшением свободной энергии. Эти энергетически выгодные реакции гликолиза обходят во время глюконеогенеза другие реакции (катализируемые различными ферментами), которые связаны с расходом АТФ и НАДН, чтобы направить их в направлении синтеза глюкозы. В целом, для образования глюкозы из двух молекул пирувата требуется четыре молекулы АТФ, две молекулы ГТФ и две молекулы НАДН.Этот процесс значительно дороже, чем простой реверс гликолиза (для которого потребовались бы две молекулы АТФ и две молекулы НАДН), что свидетельствует о дополнительной энергии, необходимой для направления пути в направлении биосинтеза.
Однако, как обсуждалось ранее, гликолитическое превращение глюкозы в пируват представляет собой энергетический путь, при котором образуется по две молекулы АТФ и НАДН.Хотя некоторые реакции гликолиза легко обратимы, другие будут протекать только в направлении распада глюкозы, поскольку связаны с большим уменьшением свободной энергии. Эти энергетически выгодные реакции гликолиза обходят во время глюконеогенеза другие реакции (катализируемые различными ферментами), которые связаны с расходом АТФ и НАДН, чтобы направить их в направлении синтеза глюкозы. В целом, для образования глюкозы из двух молекул пирувата требуется четыре молекулы АТФ, две молекулы ГТФ и две молекулы НАДН.Этот процесс значительно дороже, чем простой реверс гликолиза (для которого потребовались бы две молекулы АТФ и две молекулы НАДН), что свидетельствует о дополнительной энергии, необходимой для направления пути в направлении биосинтеза.
Как в растительных, так и в животных клетках глюкоза хранится в форме полисахаридов (соответственно крахмала и гликогена).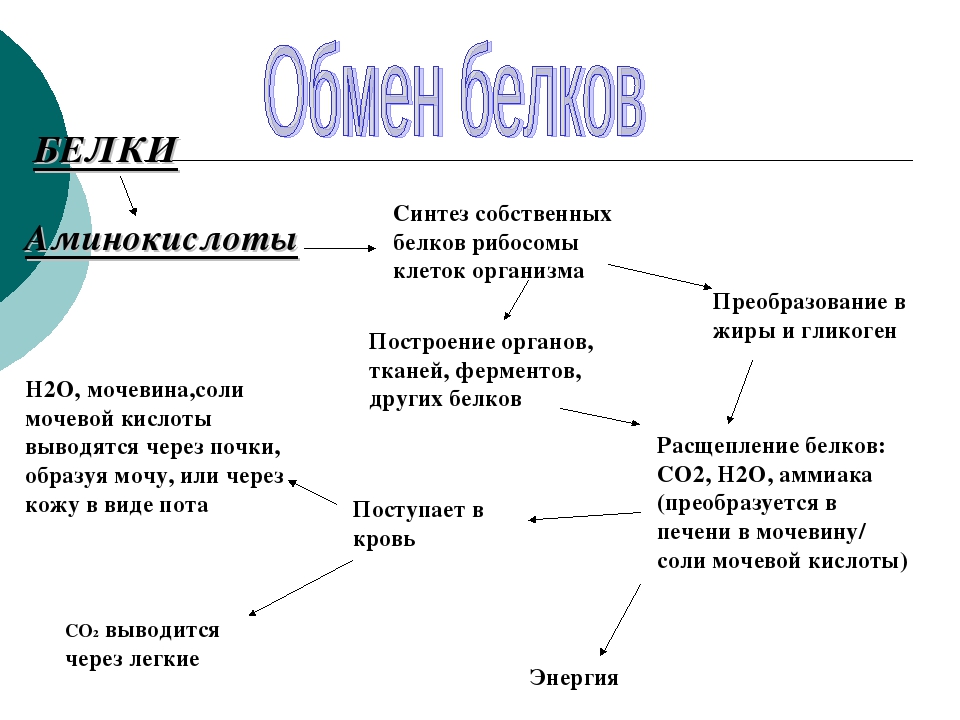 Синтез полисахаридов, как и всех других макромолекул, является энергоемкой реакцией. Как отмечалось ранее, связывание двух сахаров гликозидной связью можно представить как реакцию дегидратации, в которой удаляется H 2 O (см. ).Однако такая реакция энергетически невыгодна и поэтому не может протекать в прямом направлении. Следовательно, образование гликозидной связи должно быть сопряжено с реакцией с выделением энергии, которая достигается за счет использования нуклеотидных сахаров в качестве промежуточных продуктов в синтезе полисахаридов. Глюкоза сначала фосфорилируется в реакции, управляемой АТФ, до глюкозо-6-фосфата, который затем превращается в глюкозо-1-фосфат. Глюкозо-1-фосфат реагирует с UTP (трифосфатом уридина), давая UDP-глюкозу плюс пирофосфат, который гидролизуется до фосфата с выделением дополнительной свободной энергии.УДФ-глюкоза представляет собой активированный промежуточный продукт, который затем отдает свой остаток глюкозы растущей полисахаридной цепи в энергетически благоприятной реакции.
Синтез полисахаридов, как и всех других макромолекул, является энергоемкой реакцией. Как отмечалось ранее, связывание двух сахаров гликозидной связью можно представить как реакцию дегидратации, в которой удаляется H 2 O (см. ).Однако такая реакция энергетически невыгодна и поэтому не может протекать в прямом направлении. Следовательно, образование гликозидной связи должно быть сопряжено с реакцией с выделением энергии, которая достигается за счет использования нуклеотидных сахаров в качестве промежуточных продуктов в синтезе полисахаридов. Глюкоза сначала фосфорилируется в реакции, управляемой АТФ, до глюкозо-6-фосфата, который затем превращается в глюкозо-1-фосфат. Глюкозо-1-фосфат реагирует с UTP (трифосфатом уридина), давая UDP-глюкозу плюс пирофосфат, который гидролизуется до фосфата с выделением дополнительной свободной энергии.УДФ-глюкоза представляет собой активированный промежуточный продукт, который затем отдает свой остаток глюкозы растущей полисахаридной цепи в энергетически благоприятной реакции. Таким образом, химическая энергия в форме АТФ и УТФ запускает синтез полисахаридов из простых сахаров.
Таким образом, химическая энергия в форме АТФ и УТФ запускает синтез полисахаридов из простых сахаров.
Рисунок 2.40
Синтез полисахаридов. Глюкоза сначала превращается в активированную форму, УДФ-глюкозу, за счет одной молекулы АТФ и УДФ. Остаток глюкозы затем может быть перенесен с УДФ-глюкозы на растущую полисахаридную цепь в энергетически (более..)
Липиды
Липиды являются важными молекулами-аккумуляторами энергии и основным компонентом клеточных мембран. Они синтезируются из ацетил-КоА, образующегося при расщеплении углеводов, в ходе ряда реакций, которые напоминают обратную реакцию окисления жирных кислот. Однако, как и в случае биосинтеза углеводов, реакции, ведущие к синтезу жирных кислот, отличаются от реакций, участвующих в их расщеплении, и направлены в сторону биосинтеза, поскольку сопряжены с расходом как энергии в виде АТФ, так и восстановительной способности в виде НАДФН.Жирные кислоты синтезируются путем ступенчатого добавления двухуглеродных звеньев, полученных из ацетил-КоА, к растущей цепи. Присоединение каждого из этих двухуглеродных звеньев требует затрат одной молекулы АТФ и двух молекул НАДФН.
Присоединение каждого из этих двухуглеродных звеньев требует затрат одной молекулы АТФ и двух молекул НАДФН.
Основным продуктом биосинтеза жирных кислот, происходящим в цитозоле эукариотических клеток, является пальмитат 16-углеродной жирной кислоты. Основные составляющие клеточных мембран (фосфолипиды, сфингомиелин и гликолипиды) затем синтезируются из свободных жирных кислот в эндоплазматическом ретикулуме и аппарате Гольджи (см. главу 9).
Белки
В отличие от углеводов и липидов белки (а также нуклеиновые кислоты) содержат азот в дополнение к углероду, водороду и кислороду. Азот включается в органические соединения из разных источников в разных организмах (). Некоторые бактерии могут использовать атмосферный N 2 в процессе, называемом фиксацией азота, в котором N 2 восстанавливается до NH 3 за счет энергии в виде АТФ. Хотя относительно немногие виды бактерий способны фиксировать азот, большинство бактерий, грибов и растений могут использовать нитраты (NO 3 — ), которые являются обычным компонентом почвы, восстанавливая их до NH 3 с помощью электронов. полученный из НАДН или НАДФН.Наконец, все организмы способны включать аммиак (NH 3 ) в органические соединения.
полученный из НАДН или НАДФН.Наконец, все организмы способны включать аммиак (NH 3 ) в органические соединения.
Рисунок 2.41
Ассимиляция азота в органические соединения. Аммиак включается в органические соединения всеми организмами. Некоторые бактерии способны превращать атмосферный азот в аммиак, а большинство бактерий, грибов и растений могут утилизировать нитраты из почвы. (подробнее…)
NH 3 встраивается в органические молекулы главным образом во время синтеза аминокислот глутамата и глутамина, которые образуются из промежуточного соединения цикла лимонной кислоты α-кетоглутарата.Эти аминокислоты затем служат донорами аминогрупп во время синтеза других аминокислот, которые также образуются в результате центральных метаболических путей, таких как гликолиз и цикл лимонной кислоты (12). Таким образом, сырье для синтеза аминокислот получают из глюкозы, а аминокислоты синтезируются за счет как энергии (АТФ), так и восстановительной способности (НАДФН). Многие бактерии и растения могут синтезировать все 20 аминокислот. Однако люди и другие млекопитающие могут синтезировать только около половины необходимых аминокислот; остальное должно быть получено из диетических источников ().
Многие бактерии и растения могут синтезировать все 20 аминокислот. Однако люди и другие млекопитающие могут синтезировать только около половины необходимых аминокислот; остальное должно быть получено из диетических источников ().
Рисунок 2.42
Биосинтез аминокислот. Углеродные скелеты аминокислот образуются из промежуточных продуктов гликолиза и цикла лимонной кислоты.
Таблица 2.2
Пищевые потребности человека в аминокислотах.
Полимеризация аминокислот с образованием белков также требует энергии. Подобно синтезу полисахаридов, образование пептидной связи можно рассматривать как реакцию дегидратации, которая должна быть направлена в направлении синтеза белка за счет соединения с другим источником метаболической энергии.В биосинтезе полисахаридов это связывание осуществляется за счет превращения сахаров в активированные промежуточные продукты, такие как УДФ-глюкоза. Аминокислоты аналогичным образом активируются перед использованием для синтеза белка.
Важнейшее различие между синтезом белков и синтезом полисахаридов заключается в том, что аминокислоты включаются в белки в уникальном порядке, определяемом геном. Порядок нуклеотидов в гене определяет аминокислотную последовательность белка посредством трансляции, в которой информационная РНК (мРНК) действует как матрица для синтеза белка (см. главу 3).Каждая аминокислота сначала присоединяется к специфической молекуле транспортной РНК (тРНК) в реакции, связанной с гидролизом АТФ. Затем аминоацил-тРНК выравниваются на матрице мРНК, связанной с рибосомами, и каждая аминокислота добавляется к С-концу растущей пептидной цепи посредством ряда реакций, которые будут подробно обсуждаться в главе 7. В ходе этого процесса две дополнительные молекулы ГТФ гидролизуются, поэтому включение каждой аминокислоты в белок сопряжено с гидролизом одной молекулы АТФ и двух молекул ГТФ.
Рисунок 2.43
Образование пептидной связи. Аминокислота сначала активируется путем присоединения к ее тРНК в двухстадийной реакции, включающей гидролиз АТФ до АМФ.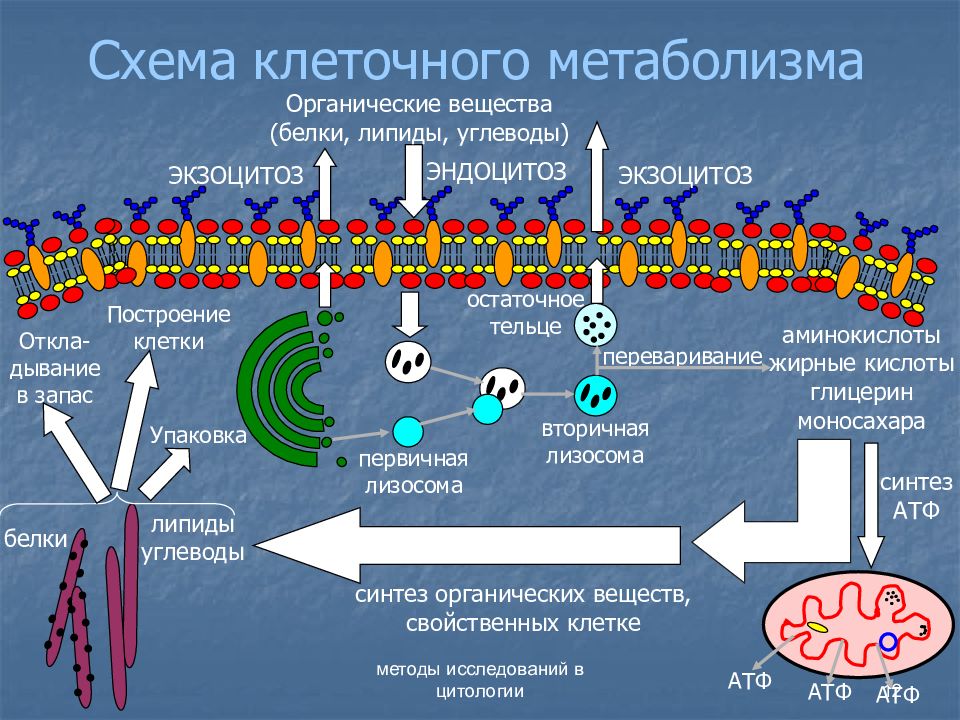 ТРНК служат адаптерами для выравнивания аминокислот на матрице мРНК, связанной с рибосомами. Каждый (подробнее…)
ТРНК служат адаптерами для выравнивания аминокислот на матрице мРНК, связанной с рибосомами. Каждый (подробнее…)
Нуклеиновые кислоты
Предшественники нуклеиновых кислот, нуклеотиды, состоят из фосфорилированных пятиуглеродных сахаров, соединенных с основаниями нуклеиновых кислот. Нуклеотиды могут быть синтезированы из углеводов и аминокислот; их также можно получать из пищевых источников или повторно использовать после распада нуклеиновых кислот.Отправной точкой для биосинтеза нуклеотидов является фосфорилированный сахар рибозо-5-фосфат, который является производным глюкозо-6-фосфата. Затем расходящиеся пути приводят к синтезу пуриновых и пиримидиновых рибонуклеотидов, которые являются непосредственными предшественниками синтеза РНК. Эти рибонуклеотиды превращаются в дезоксирибонуклеотиды, которые служат мономерными строительными блоками ДНК.
РНК и ДНК представляют собой полимеры нуклеозидмонофосфатов. Однако, что касается других макромолекул, прямая полимеризация нуклеозидмонофосфатов энергетически невыгодна, а синтез полинуклеотидов вместо этого использует нуклеозидтрифосфаты в качестве активированных предшественников (). Нуклеозид 5′-трифосфат присоединяется к 3′ гидроксильной группе растущей полинуклеотидной цепи с высвобождением и последующим гидролизом пирофосфата, направляя реакцию в направлении синтеза полинуклеотидов.
Нуклеозид 5′-трифосфат присоединяется к 3′ гидроксильной группе растущей полинуклеотидной цепи с высвобождением и последующим гидролизом пирофосфата, направляя реакцию в направлении синтеза полинуклеотидов.
Рисунок 2.44
Синтез полинуклеотидов. Нуклеозидтрифосфаты присоединяются к 3′-концу растущей полинуклеотидной цепи с высвобождением пирофосфата.
Коробка
Молекулярная медицина: Фенилкетонурия.
Какие технологические установки используют для производства углеводов? | Главная Руководства
Автор SF Gate Contributor Обновлено 2 декабря 2020 г.
Зеленые растения сами создают углеводы в процессе фотосинтеза.Функция углеводов в растениях и животных заключается в том, что они являются источником топлива для роста. Если хотя бы один основной ингредиент, необходимый растению для фотосинтеза, отсутствует или отсутствует, здоровье растения может пострадать, что приведет к задержке роста или снижению продуктивности. Садовник, который обеспечивает количество воды и света, необходимое растению для производства пищи, вознаграждается здоровым растением.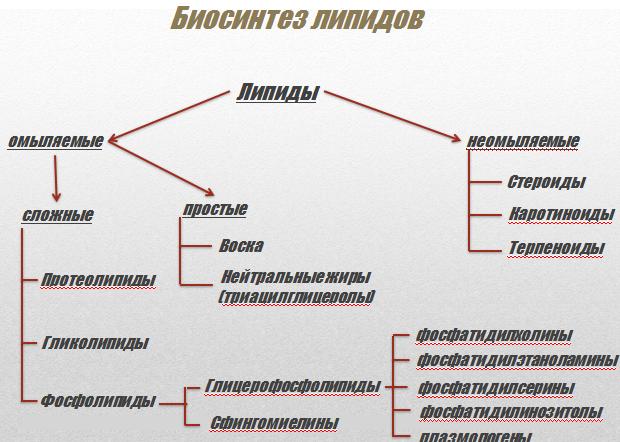
Совет
Растения используют процесс фотосинтеза для создания углеводов.
Листья, которые дышат
Люди вдыхают кислород и выделяют углекислый газ, но для того, чтобы растение производило углеводы, листья поглощают углекислый газ через устьица на поверхности листа.Из устьиц клетки мезофилла используют углекислый газ, чтобы соединить его с водой и светом для фотосинтеза. После того, как растение использует углекислый газ для фотосинтеза, побочным продуктом является кислород, который выделяется через устьица на листьях.
Транспортировка воды от корней
Для движения воды против силы тяжести вверх по растению от корней растение использует транспирацию, то есть испарение воды через устьица на листьях.В этом процессе используется до 90 процентов воды растения, и менее 10 процентов остается для фотосинтеза и роста.
Солнечный свет в фотосинтезе
Электроны, высвобождаемые при разрыве этих связей, направляются в клеточный процесс, называемый цепью переноса электронов. Эта система образует АТФ, тип молекулы энергии, которая требуется клетке для соединения водорода из воды и углерода и кислорода из углекислого газа для производства глюкозы.
Эта система образует АТФ, тип молекулы энергии, которая требуется клетке для соединения водорода из воды и углерода и кислорода из углекислого газа для производства глюкозы.
Фотосинтез создает углеводы
Фотосинтез требует солнечного света, углекислого газа и воды для производства глюкозы.Этот простой сахар представляет собой углевод, который в сочетании с другими сахарами образует структуру растения и сохраняет энергию для будущего использования.
Глюкоза, полученная в результате фотосинтеза, поступает к остальным частям растения через флоэму, которая представляет собой небольшие трубки, проходящие по всему растению. Растение хранит излишки углеводов в кроне и корнях, где они будут оставаться до тех пор, пока они не потребуются растению в темное время суток или зимой, когда лиственным деревьям не хватает листьев для производства большего количества пищи посредством фотосинтеза.
Использование и хранение углеводов
Помимо использования сложных углеводов для создания структуры растения, растения запасают углеводы или используют их для получения энергии для роста. Чтобы использовать накопленные углеводы, растения берут глюкозу, образующуюся во время фотосинтеза, и объединяют углеводы с кислородом — процесс, называемый дыханием — для высвобождения энергии. В отличие от фотосинтеза, который происходит только в зеленых частях растения, дыхание происходит во всех живых тканях, включая подземные корни.
Чтобы использовать накопленные углеводы, растения берут глюкозу, образующуюся во время фотосинтеза, и объединяют углеводы с кислородом — процесс, называемый дыханием — для высвобождения энергии. В отличие от фотосинтеза, который происходит только в зеленых частях растения, дыхание происходит во всех живых тканях, включая подземные корни.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка браузера для приема файлов cookie
Существует множество причин, по которым файл cookie не может быть установлен правильно. Ниже приведены наиболее распространенные причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки браузера, чтобы принять файлы cookie, или спросить вас, хотите ли вы принимать файлы cookie.

- Ваш браузер спрашивает, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файл cookie.
- Ваш браузер не поддерживает файлы cookie. Попробуйте другой браузер, если вы подозреваете это.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы это исправить, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Предоставить доступ без файлов cookie
потребует от сайта создания нового сеанса для каждой посещаемой вами страницы, что замедляет работу системы до неприемлемого уровня.
Предоставить доступ без файлов cookie
потребует от сайта создания нового сеанса для каждой посещаемой вами страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в файле cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только та информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, если вы не решите ввести его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступ к остальной части вашего компьютера, и только сайт, создавший файл cookie, может его прочитать.
Синтез сахаров: химики UM разработали метод, упрощающий создание углеводов
ANN ARBOR—Углеводы представляют собой сложные молекулы, которые трудно синтезировать в лаборатории, но это полезно для изучения полезных сахаров, таких как содержащиеся в человеческом грудном молоке, или позволяет исследователям адаптировать химическую структуру потенциальных лекарств, вакцин и натуральных продуктов.
Химики Мичиганского университета разработали простой метод синтеза углеводов, который расширяет круг лабораторий, которые могут использовать синтетическую химию для создания и изучения новых структур углеводов.Их результаты опубликованы в Журнале Американского химического общества.
«Нуклеиновые кислоты, белки и углеводы — это три основных строительных блока, о которых вы узнаете на уроках биологии», — сказал ведущий автор Джон Монтгомери, профессор химии и медицинской химии. «Химия смогла автоматизировать получение нуклеиновых кислот и белков там, где доступ к этим структурам является рутинным, но углеводы приготовить на порядки сложнее. В автоматизации синтеза углеводов достигнуты невероятные успехи, но факт остается фактом: это жесткая химия, которая сдерживает прогресс в гликобиологии.
Углеводы обладают большим структурным разнообразием, сложными схемами ветвления и трехмерной архитектурой, которые мешают ученым синтезировать соединения, говорит Монтгомери. Используя кремниевую группу для предварительного проектирования того, как углеводы будут реагировать друг с другом, метод команды UM позволяет контролировать схему ветвления углеводов и сокращать количество шагов, необходимых для доступа к более сложным структурам.
Используя кремниевую группу для предварительного проектирования того, как углеводы будут реагировать друг с другом, метод команды UM позволяет контролировать схему ветвления углеводов и сокращать количество шагов, необходимых для доступа к более сложным структурам.
Например, молекулы углеводов обычно состоят из атомов углерода, водорода, кислорода и иногда азота и имеют разную длину, причем наиболее распространены пяти- и шестиуглеродные сахара.Большинство атомов углерода имеют спиртовую группу, которая может быть связана со следующим углеводом бесчисленным множеством возможных моделей.
«Эта особенность ветвления делает синтетическую химию очень утомительной, потому что вы должны контролировать селективность между тем, какая часть молекулы будет реагировать со следующей», — сказал Монтгомери.
Чтобы контролировать, как эти спиртовые группы объединяют углеводы, Монтгомери и его команда поместили кремниевые «защитные группы» на отдельные спирты в углеводах.
«Обычно защитные группы необходимо включать и выключать во время синтеза, что увеличивает время и стоимость процедуры. Наша стратегия разработана таким образом, чтобы защитная группа естественным образом отпадала во время реакции связывания и умножала последовательность, в которой спиртовая группа реагирует, когда это необходимо», — сказал Монтгомери. «С помощью этого подхода мы можем изменить реакционную способность двух спиртов или взять два спирта, которые обычно имеют одинаковую реакционную способность, и заставить один реагировать избирательно по отношению к другому без дополнительных шагов.
Наша стратегия разработана таким образом, чтобы защитная группа естественным образом отпадала во время реакции связывания и умножала последовательность, в которой спиртовая группа реагирует, когда это необходимо», — сказал Монтгомери. «С помощью этого подхода мы можем изменить реакционную способность двух спиртов или взять два спирта, которые обычно имеют одинаковую реакционную способность, и заставить один реагировать избирательно по отношению к другому без дополнительных шагов.
Затем исследователи могут контролировать стереохимию углевода — трехмерное расположение молекулы углевода — путем проведения межмолекулярных или внутримолекулярных реакций между углеводами. Если реакция межмолекулярная, это означает, что реакция происходит между двумя разными молекулами. Если реакция внутримолекулярная, это означает, что перед реакцией собирается одна молекула, в которой два сахара соединены через кремний.
И структура ветвления, и стереохимия углеводов влияют на общую трехмерную архитектуру сахара, но этот структурный аспект также создает трудности для синтеза сахаров.
Поскольку молекулы сахара очень сложны, существует «огромное количество статистически возможных комбинаций, например, если бы у вас было пять сахаров», — сказал Монтгомери. «Число может исчисляться миллиардами комбинаций даже для этих относительно коротких углеводных цепей. Именно это разнообразие делает углеводы такими особенными для молекулярного распознавания в природе, но также делает их синтез чрезвычайно трудным».
Метод команды особенно полезен, потому что использование функции контроля кремния позволяет исследователям сократить количество отдельных реакций для создания цепочки углеводов за один шаг.В этом подходе элемент управления кремнием позволяет протекать реакциям с тремя различными сахарами в правильной последовательности, так что трисахарид может быть синтезирован за одну стадию синтеза. Затем эти небольшие группы могут быть дополнительно связаны для очень быстрого доступа к более сложным цепочкам.
«Вы можете настроить несколько реактивных возможностей и сделать их каскадными в нужной последовательности», — сказал Монтгомери. «Вещи, которые вы обычно запускаете в серии отдельных реакций, мы можем использовать этот кремниевый элемент управления, чтобы в основном все это происходило в реакционной колбе одновременно.Мы все еще улучшаем этот аспект с помощью новых каталитических структур, но я думаю, что эта стратегия может стать очень мощной».
«Вещи, которые вы обычно запускаете в серии отдельных реакций, мы можем использовать этот кремниевый элемент управления, чтобы в основном все это происходило в реакционной колбе одновременно.Мы все еще улучшаем этот аспект с помощью новых каталитических структур, но я думаю, что эта стратегия может стать очень мощной».
Вычислительный сотрудник Пол Циммерман, соавтор исследования и профессор химии, дает представление о том, как работает процесс и как его улучшить.
Наконец, традиционные методы связывания углеводов очень чувствительны к воде и температуре. В методе команды UM используются коммерческие реагенты, которые можно использовать без очистки или сушки для индукции химических реакций, и все процедуры выполняются при комнатной температуре.
Исследование Монтгомери было поддержано программой Glycoscience Общего фонда Национального института здравоохранения. Общий фонд был создан для обеспечения междисциплинарного внимания к научным проблемам в области биомедицинских исследований. Montgomery и UM подали предварительную патентную заявку на эту работу.
Montgomery и UM подали предварительную патентную заявку на эту работу.
Дополнительная информация:
Влияние апвеллинговых импульсов на избыточный синтез углеводов, выведенное из профилей питательных веществ, углекислого газа и кислорода в JSTOR
Абстрактный Связь между интенсивностью апвеллинга у северо-западного побережья Пиренейского моря и дифференцированным синтезом и использованием углеводов в толще воды была исследована с использованием консервативных химических параметров «NO», «CO» и «NCO».Различия между наблюдаемым вертикальным распределением «NO» и «CO» и ожидаемым по стехиометрии Редфилда указывают на то, что избыточный синтез углеводов происходит в поверхностном слое, когда питательные вещества истощаются в периоды релаксации апвеллинга. Избыток углеводов, синтезируемых в верхних слоях, расходуется на подповерхностных уровнях. Это потребление можно объяснить наличием автотрофных мигрирующих организмов, таких как фотосинтезирующие инфузории Mesodinium rubrum и несколько динофлагеллят, которые производят углеводы в верхних, хорошо освещенных слоях и поглощают питательные вещества в субфотической зоне. И наоборот, ситуации интенсивного апвеллинга связаны с образованием органического вещества в соответствии с коэффициентом Редфилда и снижением синтеза углеводов. Отклонения от стехиометрии Редфилда, обнаруженные в других морских районах, могут быть объяснены процессами, сравнимыми с процессами, происходящими в северо-западном иберийском апвеллинге, что позволяет предположить, что вертикальная миграция фотосинтезирующих организмов может играть большую роль в вертикальном переносе азота и углерода, чем обычно считается.
И наоборот, ситуации интенсивного апвеллинга связаны с образованием органического вещества в соответствии с коэффициентом Редфилда и снижением синтеза углеводов. Отклонения от стехиометрии Редфилда, обнаруженные в других морских районах, могут быть объяснены процессами, сравнимыми с процессами, происходящими в северо-западном иберийском апвеллинге, что позволяет предположить, что вертикальная миграция фотосинтезирующих организмов может играть большую роль в вертикальном переносе азота и углерода, чем обычно считается.
Ведущий журнал в своей области, MEPS освещает все аспекты морской экологии, как фундаментальные, так и прикладные.Охваченные темы включают микробиологию, ботанику, зоологию, исследования экосистем, биологическую океанографию, экологические аспекты рыболовства и аквакультуры, загрязнение, охрану окружающей среды, сохранение и управление ресурсами. MEPS стремится к высочайшему качеству научных материалов, быстрой публикации и высокому техническому стандарту презентации.
Inter-Research — это самостоятельный международный научный центр — небольшой, но с большим глобальным влиянием.Он издает ведущие журналы в области водной экологии и исследований климата.
Цикл Кальвина – Концепции биологии – 1-е канадское издание
Цели обучения
К концу этого раздела вы сможете:
- Опишите цикл Кальвина
- Определение фиксации углерода
- Объясните, как работает фотосинтез в энергетическом цикле всех живых организмов
После того, как энергия солнца преобразована и упакована в АТФ и НАДФН, клетка получает топливо, необходимое для производства пищи в виде молекул углеводов.Образовавшиеся молекулы углеводов будут иметь основу из атомов углерода. Откуда берется углерод? Атомы углерода, используемые для построения молекул углеводов, образуются из углекислого газа, газа, который животные выдыхают при каждом вдохе. Цикл Кальвина — это термин, используемый для реакций фотосинтеза, в которых используется энергия, накопленная в результате реакций, зависящих от света, для образования глюкозы и других молекул углеводов.
Цикл Кальвина — это термин, используемый для реакций фотосинтеза, в которых используется энергия, накопленная в результате реакций, зависящих от света, для образования глюкозы и других молекул углеводов.
У растений углекислый газ (CO 2 ) проникает в хлоропласт через устьица и диффундирует в строму хлоропласта — место реакций цикла Кальвина, где синтезируется сахар.Реакции названы в честь ученого, который их открыл, и ссылаются на тот факт, что реакции функционируют как цикл. Другие называют его циклом Кальвина-Бенсона, чтобы включить имя другого ученого, участвовавшего в его открытии (рис. 5.14).
Рис. 5.14. Светозависимые реакции используют энергию солнца для производства АТФ и НАДФН. Эти молекулы, несущие энергию, попадают в строму, где происходят реакции цикла Кальвина. Реакции цикла Кальвина (рис. 5.15) можно разделить на три основные стадии: фиксация, редукция и регенерация.В строме, помимо CO 2 , присутствуют два других химических вещества, инициирующих цикл Кальвина: фермент, сокращенно RuBisCO, и молекула рибулозобисфосфата (RuBP). RuBP имеет пять атомов углерода и фосфатную группу на каждом конце.
RuBP имеет пять атомов углерода и фосфатную группу на каждом конце.
RuBisCO катализирует реакцию между CO 2 и RuBP, в результате которой образуется шестиуглеродное соединение, которое немедленно превращается в два трехуглеродных соединения. Этот процесс называется фиксацией углерода, потому что CO 2 «фиксируется» из неорганической формы в органические молекулы.
АТФ и НАДФН используют накопленную энергию для преобразования трехуглеродного соединения 3-PGA в другое трехуглеродное соединение, называемое G3P. Этот тип реакции называется реакцией восстановления, потому что он включает в себя приобретение электронов. Редукция – это приобретение электрона атомом или молекулой. Молекулы АДФ и НАД + , образующиеся в результате реакции восстановления, возвращаются к светозависимым реакциям для перезарядки.
Одна из молекул G3P выходит из цикла Кальвина, чтобы участвовать в образовании молекулы углевода, обычно это глюкоза (C 6 H 12 O 6 ). Поскольку молекула углевода состоит из шести атомов углерода, для образования одной молекулы углевода требуется шесть оборотов цикла Кальвина (по одному на каждую фиксированную молекулу углекислого газа). Остальные молекулы G3P регенерируют RuBP, что позволяет системе подготовиться к стадии фиксации углерода. АТФ также используется в регенерации RuBP.
Поскольку молекула углевода состоит из шести атомов углерода, для образования одной молекулы углевода требуется шесть оборотов цикла Кальвина (по одному на каждую фиксированную молекулу углекислого газа). Остальные молекулы G3P регенерируют RuBP, что позволяет системе подготовиться к стадии фиксации углерода. АТФ также используется в регенерации RuBP.
Таким образом, требуется шесть оборотов цикла Кальвина, чтобы зафиксировать шесть атомов углерода из CO 2 . Эти шесть оборотов требуют ввода энергии от 12 молекул АТФ и 12 молекул НАДФН на стадии восстановления и 6 молекул АТФ на стадии регенерации.
Концепция в действии
Ниже приведена ссылка на анимацию цикла Кальвина. Нажмите «Этап 1», «Этап 2», а затем «Этап 3», чтобы увидеть регенерацию G3P и АТФ с образованием RuBP.
Фотосинтез
Общая эволюционная история всех фотосинтезирующих организмов бросается в глаза, поскольку основной процесс мало изменился с течением времени. Даже между гигантскими тропическими листьями в тропических лесах и крошечными цианобактериями процесс и компоненты фотосинтеза, использующие воду в качестве донора электронов, остаются в основном одинаковыми. Фотосистемы поглощают свет и используют цепи переноса электронов для преобразования энергии. Реакции цикла Кальвина собирают молекулы углеводов с этой энергией.
Однако, как и во всех биохимических путях, различные условия приводят к различным адаптациям, влияющим на основной паттерн. Фотосинтез у растений, живущих в засушливом климате (рис. 5.16), развился с приспособлениями, сохраняющими воду. В суровой сухой жаре каждая капля воды и драгоценной энергии должны быть использованы, чтобы выжить. У таких растений развились две адаптации. В одной форме более эффективное использование CO 2 позволяет растениям фотосинтезировать даже при дефиците CO 2 , например, когда устьица закрыты в жаркие дни.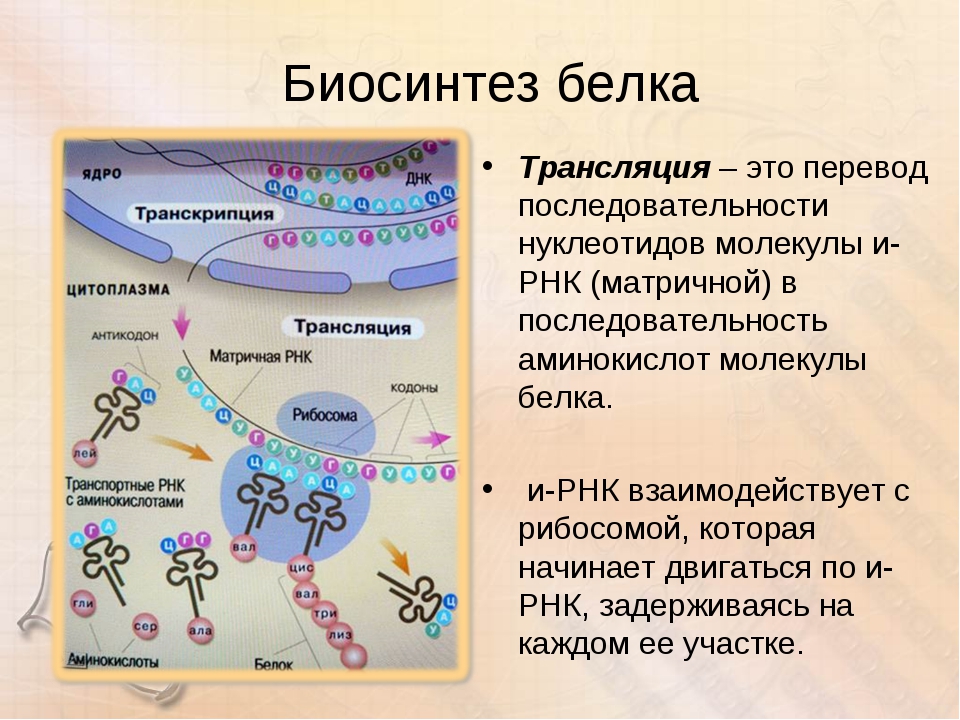 Другое приспособление выполняет предварительные реакции цикла Кальвина ночью, потому что открытие устьиц в это время сохраняет воду из-за более низких температур. Кроме того, эта адаптация позволила растениям осуществлять низкий уровень фотосинтеза, вообще не открывая устьиц, что является экстремальным механизмом для периодов чрезвычайной засухи.
Другое приспособление выполняет предварительные реакции цикла Кальвина ночью, потому что открытие устьиц в это время сохраняет воду из-за более низких температур. Кроме того, эта адаптация позволила растениям осуществлять низкий уровень фотосинтеза, вообще не открывая устьиц, что является экстремальным механизмом для периодов чрезвычайной засухи.
Два этапа фотосинтеза — светозависимые реакции и цикл Кальвина — были описаны в том виде, в каком они происходят в хлоропластах. Однако у прокариот, таких как цианобактерии, отсутствуют мембраносвязанные органеллы. Прокариотические фотосинтезирующие автотрофные организмы имеют складки плазматической мембраны для прикрепления хлорофилла и фотосинтеза (рис. 5.17). Именно здесь такие организмы, как цианобактерии, могут осуществлять фотосинтез.
Живые существа получают энергию, расщепляя молекулы углеводов. Однако, если растения производят молекулы углеводов, зачем им их расщеплять? Углеводы — это запасные молекулы для энергии во всех живых существах. Хотя энергия может храниться в таких молекулах, как АТФ, углеводы являются гораздо более стабильными и эффективными резервуарами для химической энергии.Фотосинтезирующие организмы также осуществляют реакции дыхания, чтобы собрать энергию, которую они запасли в углеводах, например, растения имеют митохондрии в дополнение к хлоропластам.
Вы могли заметить, что общая реакция фотосинтеза:
6CO 2 + 6H 2 + 6H 2 O → C 612 O 6 + 6O 2 6CO 2 + 6H 2 O → C 6 H 12 O 6 + 6О
— обратная общая реакция клеточного дыхания:
6O 2 + C 6 H 12 O 6 → 6000 o 2 + 6H 2 + 6H 2 O 6 O 2 + C 6 H 12 O 6 →6CO 2 +6H 2 O
Фотосинтез производит кислород в качестве побочного продукта, а дыхание производит углекислый газ в качестве побочного продукта.
В природе отходов не бывает. Каждый отдельный атом материи сохраняется, перерабатываясь бесконечно. Вещества изменяют форму или переходят из одного типа молекулы в другой, но никогда не исчезают (рис. 5.18).
CO 2 является формой отходов, образующихся при дыхании, не больше, чем кислород является побочным продуктом фотосинтеза. Оба являются побочными продуктами реакций, которые переходят в другие реакции. Фотосинтез поглощает энергию для создания углеводов в хлоропластах, а аэробное клеточное дыхание высвобождает энергию, используя кислород для расщепления углеводов.Обе органеллы используют цепи переноса электронов для выработки энергии, необходимой для запуска других реакций. Фотосинтез и клеточное дыхание функционируют в биологическом цикле, позволяя организмам получать доступ к поддерживающей жизнь энергии, которая берет свое начало в звезде, находящейся за миллионы километров.
Рис. 5.18. В круговороте углерода реакции фотосинтеза и клеточного дыхания имеют общие реагенты и продукты. (кредит: модификация работы Стюарта Бэссила)
(кредит: модификация работы Стюарта Бэссила)
Используя энергоносители, образующиеся на первой стадии фотосинтеза, реакции цикла Кальвина фиксируют CO 2 из окружающей среды для построения молекул углеводов.Фермент RuBisCO катализирует реакцию фиксации путем объединения CO 2 с RuBP. Полученное шестиуглеродное соединение расщепляется на два трехуглеродных соединения, и энергия АТФ и НАДФН используется для преобразования этих молекул в G3P. Одна из трехуглеродных молекул G3P выходит из цикла и становится частью молекулы углевода. Оставшиеся молекулы G3P остаются в цикле, чтобы сформироваться обратно в RuBP, который готов к реакции с большим количеством CO 2 . Фотосинтез образует сбалансированный энергетический цикл с процессом клеточного дыхания.Растения способны как к фотосинтезу, так и к клеточному дыханию, поскольку содержат как хлоропласты, так и митохондрии.
Глоссарий
Цикл Кальвина: реакции фотосинтеза, использующие энергию, накопленную в результате светозависимых реакций, для образования глюкозы и других молекул углеводов
фиксация углерода: процесс преобразования неорганического CO 2 газа в органические соединения
.
