
Таблицa по биологии Деление клетки. Митоз
Издательство: Ксеноксс
Вид УМК: Таблицa
| Интерфаза | Происходит подготовка к делению. Важнейшим процессом в хромосомах является удвоение ДНК, после которого каждая хромосома состоит из двух идентичных половинок – хроматид. Удвоение ДНК получило название «редупликация». При редупликации водородные связи между комплементарными азотистыми основаниями аденином – тимином и гуанином – цитозином разрываются специальным ферментом. Нити, составляющие двойную спираль ДНК, расходятся, и к каждому нуклеотиду обеих нитей подстраиваются комплементарные нуклеотиды. Также при интерфазе увеличивается число многих органоидов, происходит запасание АТФ |
| Профазы | Процесс митотического деления. Ядро увеличивается в объёме, хромосомы начинают скручиваться. Удвоившиеся в интерфазу центриоли клеточного центра расходятся к полюсам клетки. Начинает формироваться веретено деления.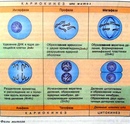 |
| Метафаза | Хромосомы максимально скручены. Каждая из них имеет форму и представляет собой вытянутое тельце, состоящее из двух хроматид. Хроматиды соединены в единую хромосому в области (центромеры). Во время метафазы хромосомы состоят из двух дочерних хроматид, располагаются в экваториальной плоскости клетки. Нити веретена деления, идущие от центриолей, прикрепляются к каждой хромосоме в области центромеры. |
| Анафаза | Дочерние хроматиды отделяются друг от друга и расходятся к полюсам клетки. Движение хроматид происходит, потому что нити веретена деления укорачиваются и тащат хроматиды к полюсам клетки. |
| Телофаза | Хроматиды достигают полюсов клетки и раскручиваются. Вокруг них формируются ядерные оболочки, и в результате оформляются два ядра. Одновременно происходит деление цитоплазмы, органоиды распределяются между двумя клетками. Две одинаковые клетки отделяются друг от друга.
|
Адрес: 1069, Курземес проспект 106/45, Рига, Латвия.
Тел.: +371 29-851-888 E-mail: [email protected]
Каковы стадии деления клетки в мейозе? Кратко — Медицина — Наука — Каталог статей
Процесс клеточного деления при мейозе встречается у всех организмов, которые могут размножаться половым путем, включая людей, растения и животных. Мейоз — это деление клеток на две части с образованием четырех дочерних клеток с половиной числа хромосом, как у исходной или родительской клетки. Процесс деления на две части называется мейозом I и мейозом II. Так, телофазный мейоз характеризуется телофазой I и телофазой II, так же как все другие стадии происходят дважды в процессе деления при мейозе.
Межфазная стадия — это когда клетка находится в состоянии покоя и получает предметы, необходимые для предстоящего деления клетки. На этой стадии клетки остаются большую часть своей жизни. Интерфаза разбита на три фазы: G1, S и G2. В фазе G1 клетка увеличивается в массе, чтобы подготовиться к делению. G представляет собой разрыв, а первая представляет собой фазу, что означает, что фаза G1 является первой фазой разрыва в делении клеток мейоза.
Интерфаза разбита на три фазы: G1, S и G2. В фазе G1 клетка увеличивается в массе, чтобы подготовиться к делению. G представляет собой разрыв, а первая представляет собой фазу, что означает, что фаза G1 является первой фазой разрыва в делении клеток мейоза.
S-фаза — это следующая стадия синтеза ДНК. S обозначает синтез. Фаза G2 является второй фазой разрыва, в которой клетка синтезирует свои белки, и она продолжает увеличиваться в размере. В конце интерфазы в клетке присутствуют ядрышки, а ядро связано ядерной оболочкой. Клеточные хромосомы делятся и находятся в форме хроматина. В клетках животных и человека две пары центриолей образуются и располагаются снаружи ядра.
Фаза I — это этап, когда несколько изменений в клетке вступают в силу. Хромосомы сгущаются в размере, и затем они прикрепляются к ядерной оболочке. Пара идентичных или гомологичных хромосом выстраиваются в линию близко друг к другу, образуя тетраду, которая состоит из четырех хроматид. Это известно как синапсис.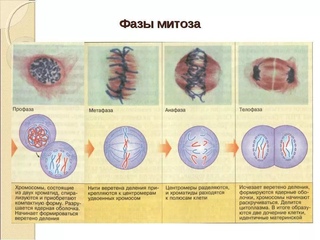 Пересечение может произойти, чтобы создать новые генетические комбинации, которые отличаются от любой из родительских клеток.
Пересечение может произойти, чтобы создать новые генетические комбинации, которые отличаются от любой из родительских клеток.
Хромосомы утолщаются, а затем они отделяются от ядерной оболочки. Центриоли отодвигаются друг от друга и начинают мигрировать к противоположным сторонам или полюсам клетки. Ядра и ядерная оболочка разрушаются, и хромосомы начинают перемещаться в метафазную пластинку.
Метафаза I — это следующая стадия, на которой тетрады выравниваются на метафазной пластинке в клетке, и идентичные пары хромосом или центромеры теперь находятся на противоположных сторонах клетки.
В анафазе I волокна развиваются от противоположных полюсов клетки, чтобы тянуть хромосомы к двум полюсам. Две идентичные копии хромосомы, которые связаны центромерой или сестринскими хроматидами, остаются вместе после того, как хромосомы перемещаются к противоположным полюсам.
Следующим этапом является телофаза I, в которой волокна веретена продолжают тянуть гомологичные хромосомы к противоположным полюсам.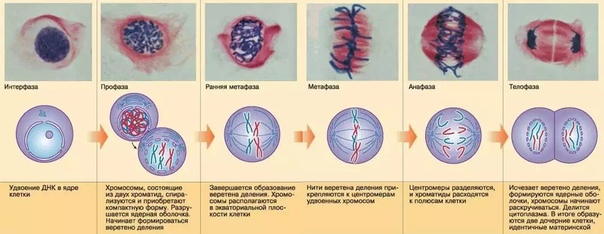 После того, как они достигают полюсов, каждый из двух полюсов содержит гаплоидную клетку, которая содержит вдвое меньше хромосом, чем родительская клетка. Деление цитоплазмы обычно происходит в телофазе I. В конце телофазы I и в процессе цитокинеза, когда клетка делится, каждая клетка будет иметь половину хромосом родительской клетки. Генетический материал больше не дублируется, и клетка переходит в мейоз II.
После того, как они достигают полюсов, каждый из двух полюсов содержит гаплоидную клетку, которая содержит вдвое меньше хромосом, чем родительская клетка. Деление цитоплазмы обычно происходит в телофазе I. В конце телофазы I и в процессе цитокинеза, когда клетка делится, каждая клетка будет иметь половину хромосом родительской клетки. Генетический материал больше не дублируется, и клетка переходит в мейоз II.
В профазе II ядра и ядерная мембрана распадаются по мере появления веретенной сети волокон. Хромосомы снова начинают мигрировать в пластинку метафазы II, которая находится в центре или на экваторе клеток.
Метафаза II — это стадия, на которой хромосомы клетки выравниваются на метафазной пластине II в центре клетки, а волокна сестринских хроматид указывают на два противоположных полюса на противоположных сторонах клетки.
Анафаза II — это следующая стадия деления клеток при мейозе, когда сестринские хроматиды отделяются друг от друга и начинают двигаться к противоположным концам клетки.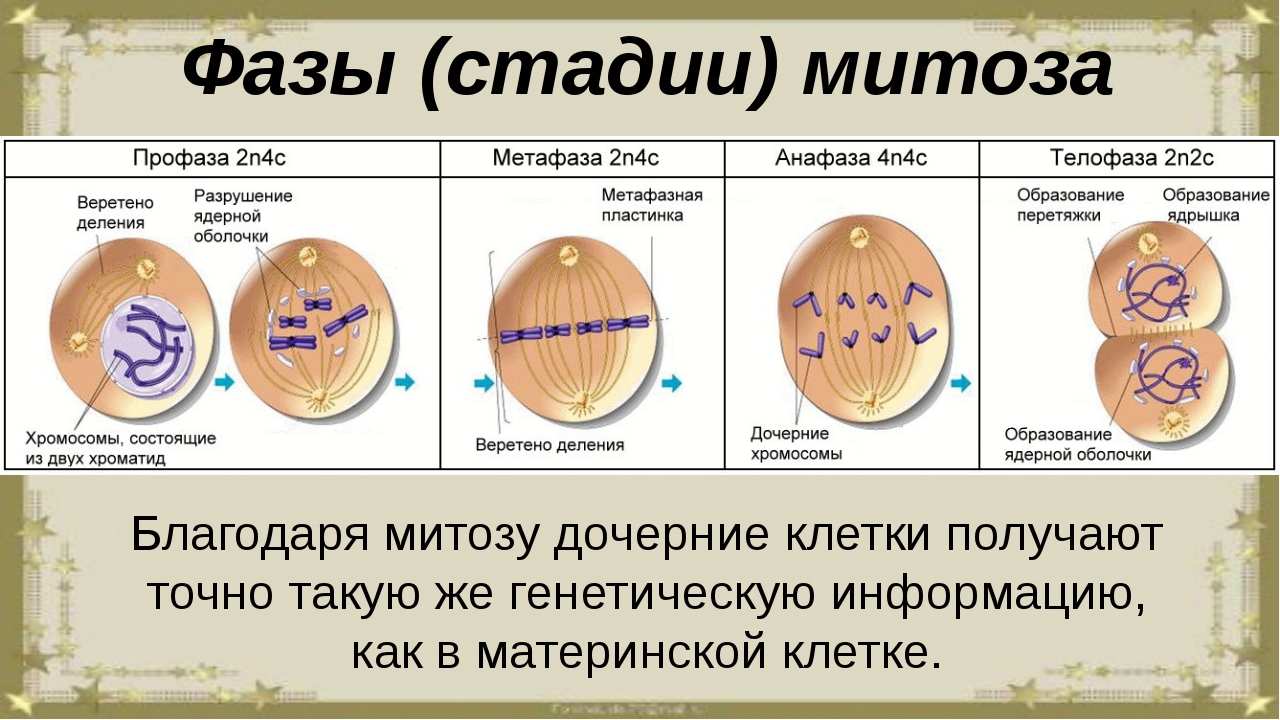 Волокна веретена, которые не связаны с двумя хроматидами, удлиняются, и это удлиняет клетку. Разделение сестринских хроматид в паре — это точка, когда хроматиды становятся хромосомами, называемыми дочерними хромосомами. По мере удлинения ячейки полюса смещаются дальше друг от друга. В конце этой стадии каждый полюс содержит полный набор хромосом.
Волокна веретена, которые не связаны с двумя хроматидами, удлиняются, и это удлиняет клетку. Разделение сестринских хроматид в паре — это точка, когда хроматиды становятся хромосомами, называемыми дочерними хромосомами. По мере удлинения ячейки полюса смещаются дальше друг от друга. В конце этой стадии каждый полюс содержит полный набор хромосом.
В телофазе II два разных ядра начинают формироваться на противоположных полюсах клетки. Цитоплазма делится через цитокинез с образованием двух отдельных клеток, которые называются дочерними клетками, каждая с половиной числа хромосом в качестве родительской клетки. Конечным продуктом после мейоза I и II стадии являются четыре гаплоидные дочерние клетки. Когда гаплоидные клетки объединяются во время оплодотворения сперматозоида и яйцеклетки, они становятся диплоидной клеткой, так же, как исходная родительская клетка находилась в начале клетки перед делением.
Деление клетки. Митоз. — биология, уроки
Класс: 10
Дата: 08.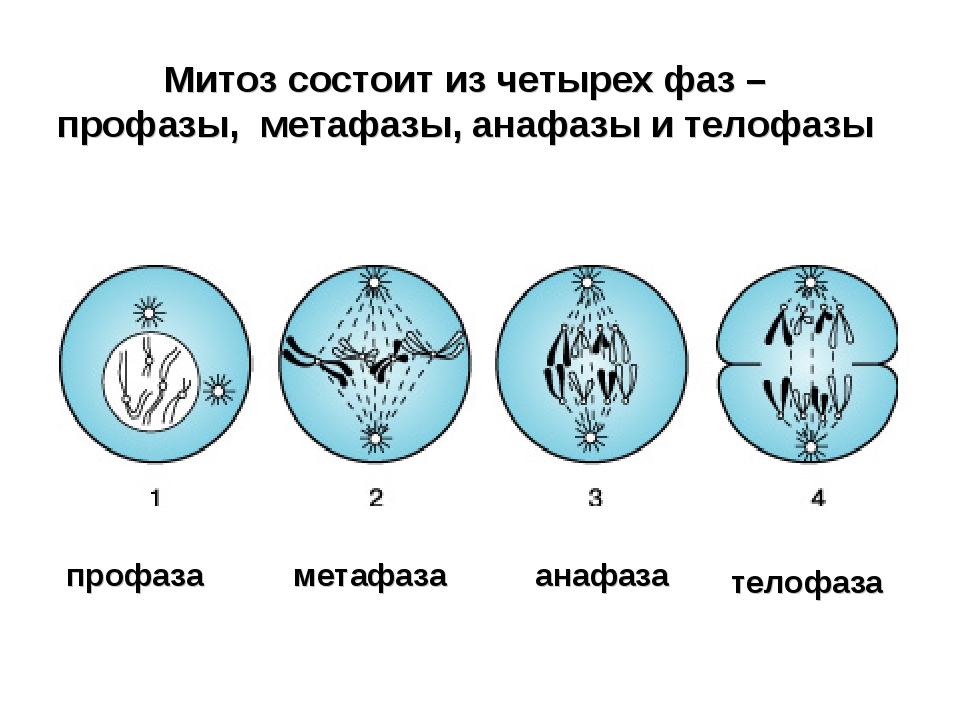 02.2017 г.
02.2017 г.
Тема: Деление клетки. Митоз
Цель: углубить знания учащихся о наследственности организмов, ознакомить со строением и функциями хромосом; дать представление о клеточном цикле деления клеток. Продолжить формирование у учащихся научного мировоззрения, развитие мышления, памяти, внимания, научной биологической речи, обще учебных умений и навыков. Воспитание ценностей личного отношения к изучаемым знаниям. Формирование информационных, коммуникативных, обще учебных компетентностей.
Оборудование и материалы: учебник, иллюстрации по теме урока
Понятия и термины: хромосома, центромера, клеточный цикл, фазы митоза (профаза, метафаза, анафаза, телофаза), интерфаза, диплоидный набор хромосом
Тип урока: изучение нового материала
Ход урока
I Организационный этап
II Проверка домашнего задания
1. Объяснить, как происходит конъюгация у прокариот.
Объяснить, как происходит конъюгация у прокариот.
2. Объяснить, как происходит оплодотворение у животных.
3. Объяснить, как происходит оплодотворение у покрытосеменных растений (двойное оплодотворение).
IIІ Актуализация опорных знаний
- Беседа (фронтально)
- Что такое ядро?
- Какие органеллы находятся в ядре клетки?
- Что такое хромосомы?
IV Мотивация учебной деятельности
- Слово учителя
Сегодня на уроке мы будем говорить с вами о клеточном цикле и таком делении клеток как митоз.
Цикл жизни клетки – интерфаза и митоз,
А как он протекает? – это главный вопрос.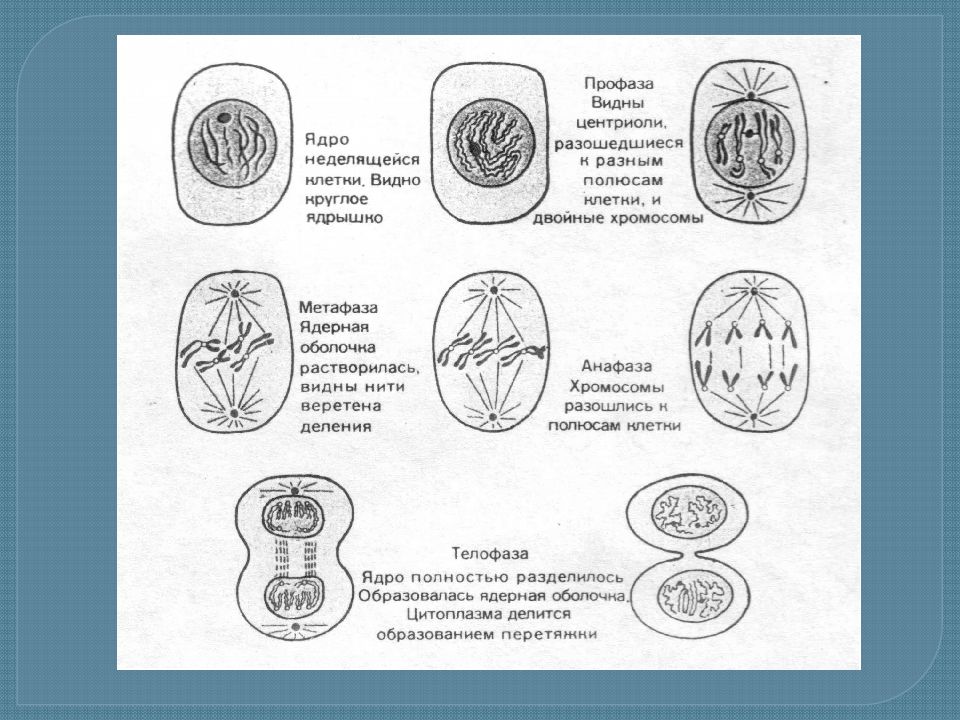
Об этом не скажешь ведь в двух словах,
Процесс жизни клетки рассмотрим в стихах.
Интерфаза длится дольше, чем само деление,
Очень быстро происходит ДНК удвоение.
Идет биосинтез, активны ферменты.
Клетка растет, образует органоиды и элементы
Затем следует митотическое деление,
Фазы его легко запомнить — и в этом нет сомнения.
Внимательно на них ты посмотри.
Каждая фаза как член большой и дружной семьи.
Глава семьи – папа (всем ясно сразу),
И первая фаза митоза — профаза.
Исчезло ядрышко и ядерная оболочка,
Но на этом рано еще ставить точку.
Хромосомы укорачиваются, утолщаются,
В компактные формы превращаются.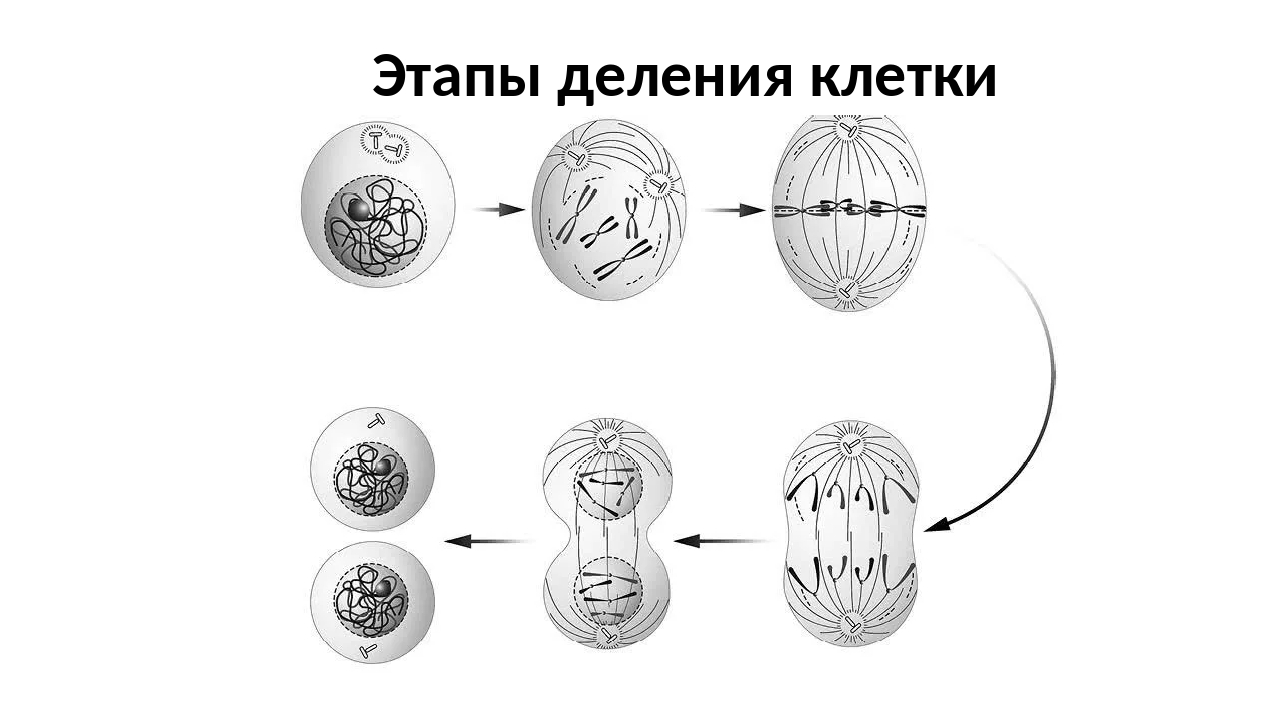
И затем без промедления —
Появляются нити веретена деления.
Мама — солнышко наше, тепло, доброта.
Метафаза – вторая фаза митоза всегда.
Дети для мамы равны без дозатора,
Хромосомы лежат в области экватора
Дочка — Аня в семье — просто принцесса.
Анафаза – третья фаза процесса.
Убедиться в этом ты можешь сам —
Нити веретена деления оттягивают
хроматиды к различным полюсам.
Сынок в семье Толя – ну, как по заказу
Четвертая фаза митоза – телофаза.
Хромосомы раскручиваются, у них выход один —
Снова превратиться в хроматин.
После деления цитоплазмы и органоидов клетки,
Появляются две прелестные, чудные детки.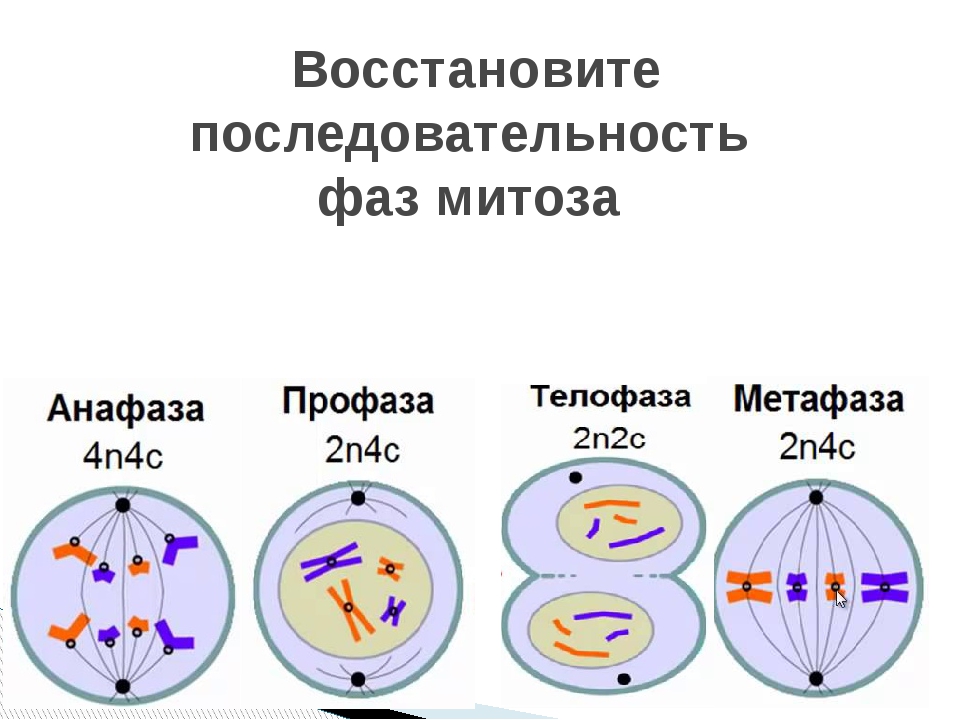
Имеют диплоидный набор дочерние клетки
и в точности похожи на материнскую клетку.
V Изучение нового материала
1. Клеточный цикл
Клеточный цикл – период существования клетки между началами её двух последовательных делений или же от начала деления до ее гибели.
Клеточный цикл состоит из периодов деления и интерфазы.
Интерфаза – это период между двумя последовательными делениями клетки.
Интерфаза состоит из 3-х этапов: предсинтетического, синтетического и послесинтетического. На синтетическом этапе происходит рост клетки, удвоение молекул ДНК, синтез белков и других органических соединений, деление митохондрий и пластид, … . Активно запасается энергия, необходимая для деления клетки.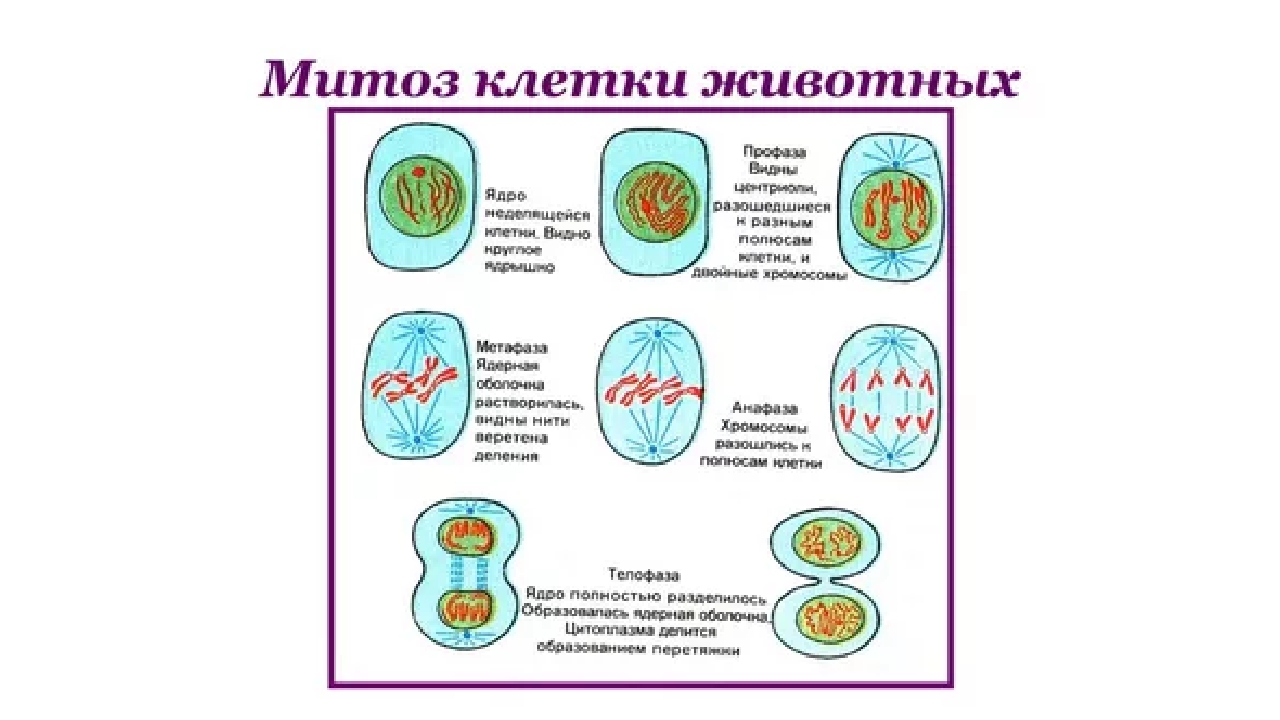
Этап между завершеним предыдущего деления и синтетическим этапом называют предсинтетическим, а между завершеним синтетического этапа и началом следующего деления – послесинтетическим.
Основным способом деления эукариотических клеток является митоз.
2. Фазы митотического деления
Продолжительность митоза составляет от нескольких минут до 2-3 часов.
Митоз включает следующие фазы (рис. 31, с. 107):
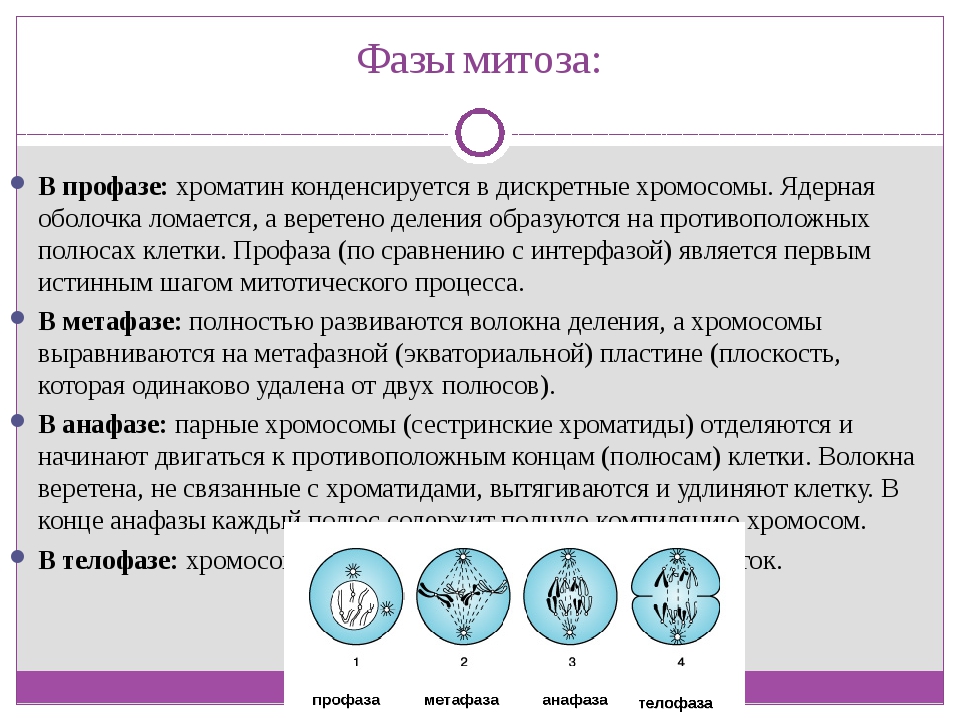
- Профаза: происходит спирализация хромосом, исчезает ядерная оболочка, ядрышко; начинает формироваться веретено деления. Нити этого веретена деления прикрепляются к центромерам, и хромосомы начинают перемещаться к центральной части клетки.
-
Метафаза: завершается спирализация хромосом и формирование веретена деления.
 Хромосомы «выстраиваются» в центральной части клетки.
Хромосомы «выстраиваются» в центральной части клетки.
- Анафаза: самая короткая фаза митоза. Делятся центромеры и хроматиды расходятся к разным полюсам клетки.
- Телофаза: вокруг каждого из двух скоплений хроматид формируется ядерная оболочка, появляются ядрышки. Постепенно исчезает веретено деления. В конце телофазы делится цитоплазма материнской клетки и образуются две дочерние.
3. Биологическое значение митоза
Митоз обеспечивает постоянство числа хромосом во всех клетках организма, вследствие чего все они имеют одну и ту же генетическую информацию
VI Обобщение и закрепление изученного материала
- Тест «Митоз» (фронтально)
VII Итог урока
VIII Домашнее задание
§23 (только митоз, значение митоза), выучить конспект.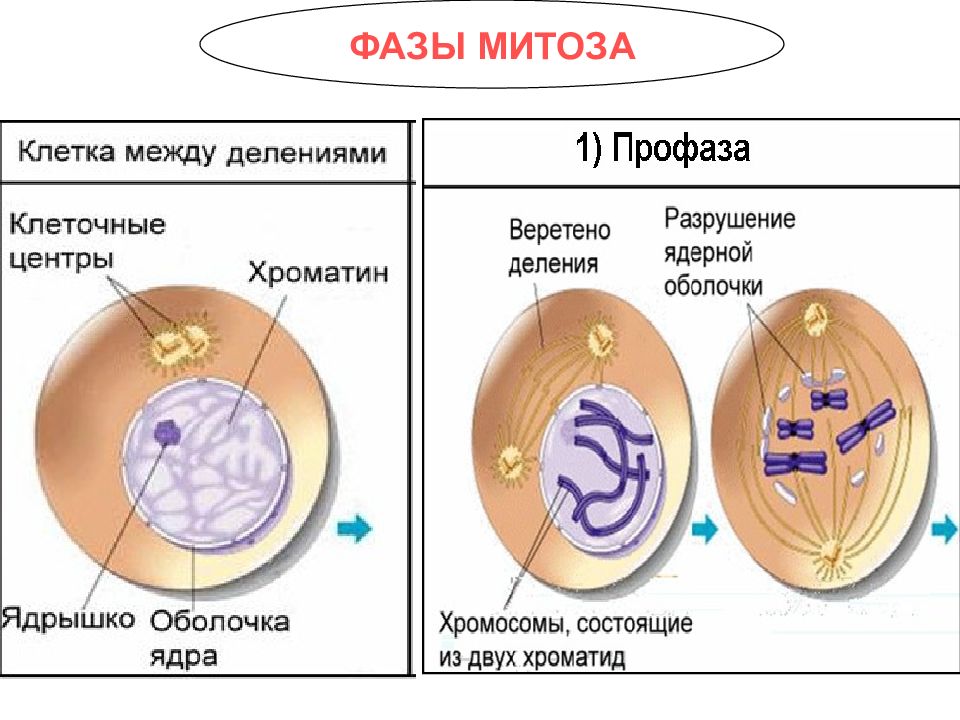
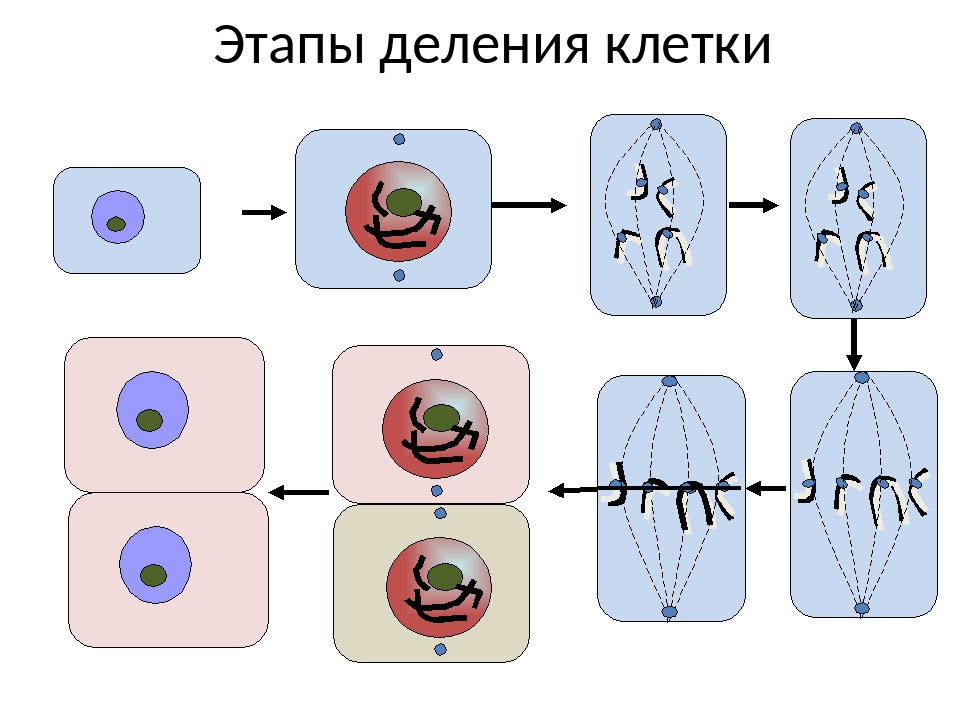
Деление клетки: митоз
Клетки многоклеточного организма чрезвычайно разнообразны по выполняемым функциям. В соответствии со специализацией клетки имеют разную продолжительность жизни. Например, нервные и мышечные клетки после завершения эмбрионального периода развития перестают делиться и функционируют на протяжении всей жизни организма. Клетки же других тканей — костного мозга, эпидермиса, эпителия тонкого кишечника — в процессе выполнения своей функции быстро погибают и замещаются новыми в результате непрерывного клеточного размножения.
Таким образом, жизненный цикл клеток обновляющихся тканей включает функционально активную деятельность и период деления. Деление клеток лежит в основе развития и роста организмов, их размножения, а также обеспечивает самообновление тканей на протяжении жизни организма и восстановление их целостности после повреждения.
Наиболее широко распространенная форма воспроизведения клеток у живых организмов — непрямое деление, или митоз.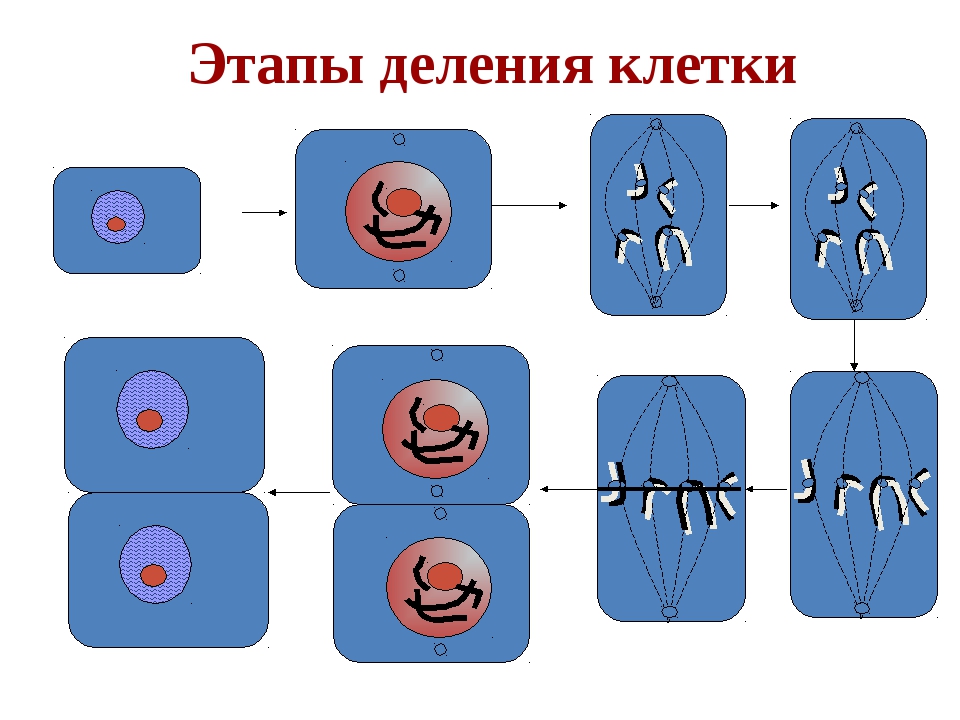 Для митоза характерны сложные преобразования ядра клетки, сопровождающиеся формированием специфических структур-хромосом. Хромосомы постоянно присутствуют в клетке, но в период между двумя делениями — интерфазе — находятся в деспирализованном состоянии и потому не видны в световой микроскоп. В интерфазе осуществляется подготовка к митозу, заключающаяся главным образом в удвоении (редупликации) ДНК. Совокупность процессов, происходящих в период подготовки клетки к делению, а также на протяжении самого митоза, называется митотическим циклом. На рисунке видно, что после завершения деления клетка может вступить в период подготовки к синтезу ДНК, обозначаемый символом G1. В это время в клетке усиленно синтезируются РНК и белки, повышается активность ферментов, участвующих в синтезе ДНК. Затем клетка приступает к синтезу ДНК. Две спирали старой молекулы ДНК расходятся, и каждая становится матрицей для синтеза новых цепей ДНК. В результате каждая из двух дочерних молекул обязательно включает одну старую спираль и одну новую.
Для митоза характерны сложные преобразования ядра клетки, сопровождающиеся формированием специфических структур-хромосом. Хромосомы постоянно присутствуют в клетке, но в период между двумя делениями — интерфазе — находятся в деспирализованном состоянии и потому не видны в световой микроскоп. В интерфазе осуществляется подготовка к митозу, заключающаяся главным образом в удвоении (редупликации) ДНК. Совокупность процессов, происходящих в период подготовки клетки к делению, а также на протяжении самого митоза, называется митотическим циклом. На рисунке видно, что после завершения деления клетка может вступить в период подготовки к синтезу ДНК, обозначаемый символом G1. В это время в клетке усиленно синтезируются РНК и белки, повышается активность ферментов, участвующих в синтезе ДНК. Затем клетка приступает к синтезу ДНК. Две спирали старой молекулы ДНК расходятся, и каждая становится матрицей для синтеза новых цепей ДНК. В результате каждая из двух дочерних молекул обязательно включает одну старую спираль и одну новую.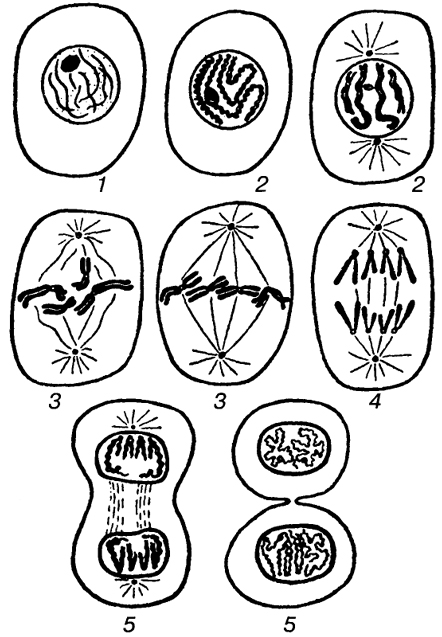 Новая молекула абсолютно идентична старой. В этом заключается глубокий биологический смысл: таким путем в бесчисленных клеточных поколениях сохраняется преемственность генетической информации.
Новая молекула абсолютно идентична старой. В этом заключается глубокий биологический смысл: таким путем в бесчисленных клеточных поколениях сохраняется преемственность генетической информации.
Продолжительность синтеза ДНК в разных клетках неодинакова и колеблется от нескольких минут у бактерий до 6-12 ч в клетках млекопитающих. После завершения синтеза ДНК — фазы S митотического цикла — клетка не сразу начинает делиться. Период от окончания синтеза ДНК и до начала митоза называется фазой G2. В этот период клетка завершает подготовку к митозу: накапливается АТФ, синтезируются белки ахроматинового веретена, удваиваются центриоли.
Процесс собственно митотического деления клетки состоит из четырех фаз: профазы, метафазы, анафазы и телофазы.
В профазе увеличивается объем ядра и клетки в целом, клетка округляется, снижается или прекращается ее функциональная активность (например, амебоидное движение у простейших и у лейкоцитов высших животных). Часто исчезают специфические структуры клетки (реснички и др.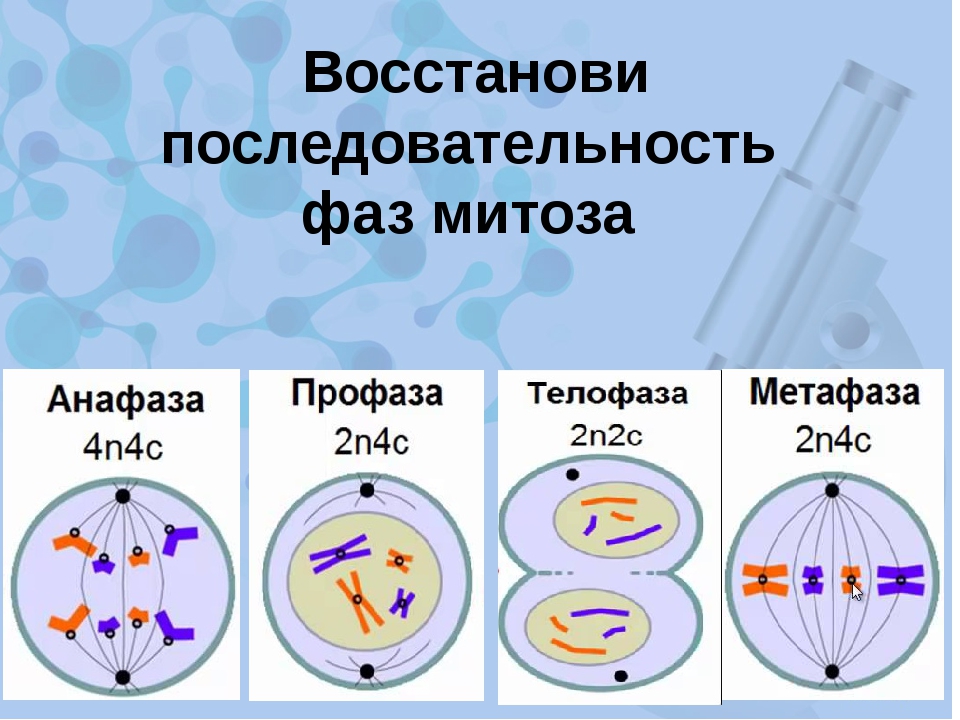 ). Центриоли попарно расходятся к полюсам, хромосомы спирализуются и вследствие этого утолщаются, становятся видимыми. Считывание генетической информации с молекулДНК становится невозможным: синтез РНК прекращается, ядрышко исчезает. Между полюсами клетки протягиваются нити веретена деления — формируется аппарат, обеспечивающий расхождение хромосом к полюсам клетки. На протяжении всей профазы продолжается спирализация хромосом, которые становятся толстыми и короткими. В конце профазы ядерная оболочка распадается и хромосомы оказываются беспорядочно рассеянными в цитоплазме.
). Центриоли попарно расходятся к полюсам, хромосомы спирализуются и вследствие этого утолщаются, становятся видимыми. Считывание генетической информации с молекулДНК становится невозможным: синтез РНК прекращается, ядрышко исчезает. Между полюсами клетки протягиваются нити веретена деления — формируется аппарат, обеспечивающий расхождение хромосом к полюсам клетки. На протяжении всей профазы продолжается спирализация хромосом, которые становятся толстыми и короткими. В конце профазы ядерная оболочка распадается и хромосомы оказываются беспорядочно рассеянными в цитоплазме.
В метафазе спирализация хромосом достигает максимума, и укороченные хромосомы устремляются к экватору клетки, располагаясь на равном расстоянии от полюсов. Образуется экваториальная, или метафазная, пластинка. На этой стадии митоза отчетливо видна структура хромосом, их легко сосчитать и изучить их индивидуальные особенности.
В каждой хромосоме имеется область первичной перетяжки — центромера, к которой во время митоза присоединяются нить веретена деления и плечи.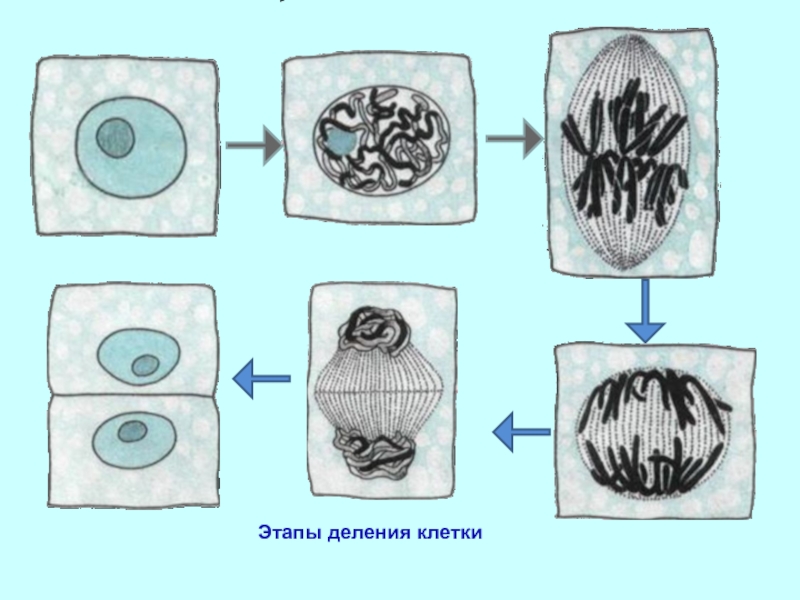 На стадии метафазы хромосома состоит из двух хроматид, соединенных между собой только в области центромеры.
На стадии метафазы хромосома состоит из двух хроматид, соединенных между собой только в области центромеры.
Во всех соматических клетках любого организма содержится строго определенное число хромосом. У всех организмов, относящихся к одному виду, число хромосом в клетках одинаково: у домашней мухи — 12, у дрозофилы — 8, у кукурузы — 20, у земляники садовой — 56, у рака речного — 116, у человека — 46, у шимпанзе, таракана и перца — 48. Как видно, число хромосом не зависит от высоты организации и не всегда указывает на филогенетическое родство. Число хромосом, таким образом, не служит видоспецифическим признаком.Носовокупность признаков хромосомного набора (кариотип) — форма, размеры и число хромосом — свойственна только одному какому-то виду растений или животных.
Число хромосом в соматических клетках всегда парное. Это объясняется тем, что в этих клетках находятся две одинаковые по форме и размерам хромосомы: одна происходит от отцовского, другая — от материнского организма. Хромосомы, одинаковые по форме и размерам и несущие одинаковые гены, называются гомологичными. Хромосомный набор соматической клетки, в котором каждая хромосома имеет себе пару, носит название двойного,или диплоидного набора, и обозначается 2n. Количество ДНК, соответствующее диплоидному набору хромосом, обозначают как 2с. В половые клетки из каждой пары гомологичных хромосом попадает только одна, поэтому хромосомный набор гамет называется одинарным или гаплоидным.
Хромосомы, одинаковые по форме и размерам и несущие одинаковые гены, называются гомологичными. Хромосомный набор соматической клетки, в котором каждая хромосома имеет себе пару, носит название двойного,или диплоидного набора, и обозначается 2n. Количество ДНК, соответствующее диплоидному набору хромосом, обозначают как 2с. В половые клетки из каждой пары гомологичных хромосом попадает только одна, поэтому хромосомный набор гамет называется одинарным или гаплоидным.
Изучение деталей строения хромосом метафазной пластинки имеет очень большое значение для диагностики заболеваний человека, обусловленных нарушениями строения хромосом.
В анафазе вязкость цитоплазмы уменьшается, центромеры разъединяются, и с этого момента хроматиды становятся самостоятельными хромосомами. Нити веретена деления, прикрепленные к центромерам, тянут хромосомы к полюсам клетки, а плечи хромосом при этом пассивно следуют за центромерой. Таким образом, в анафазе хроматиды удвоенных еще в интерфазе хромосом точно расходятся к полюсам клетки.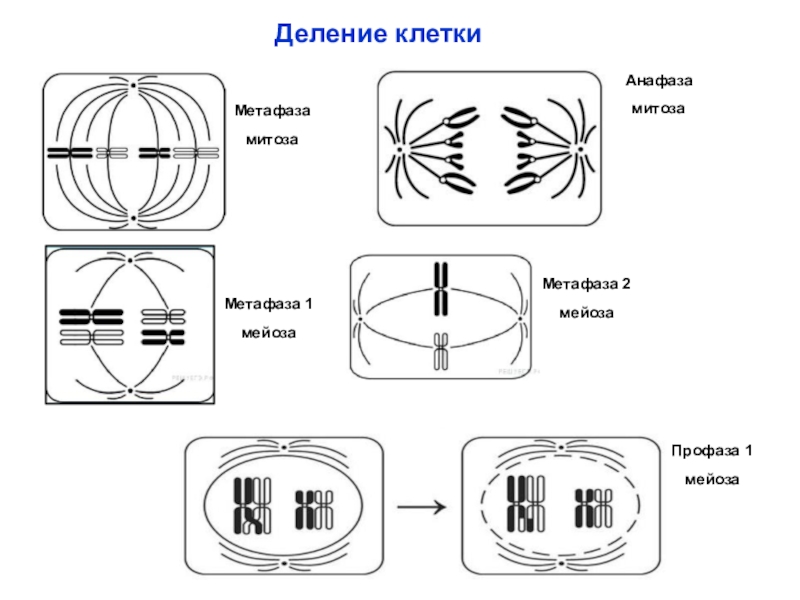 В этот момент в клетке находятся два диплоидных набора хромосом (4n4с).
В этот момент в клетке находятся два диплоидных набора хромосом (4n4с).
В заключительной стадии — телофазе — хромосомы раскручиваются, деспирализуются. Из мембранных структур Цитоплазмы образуется ядерная оболочка. У животных клетка делится на две меньших размеров путем образования перетяжки. У растений цитоплазматическая мембрана возникает в середине клетки и распространяется к периферии, разделяя клетку пополам. После образования поперечной цитоплазматической мембраны у растительных клеток появляется целлюлозная стенка. Так из одной клетки формируются две дочерние, в которых наследственная информация точно копирует информацию, содержавшуюся в материнской клетке. Начиная с первого митотического деления оплодотворенной яйцеклетки (зиготы) все дочерние клетки, образовавшиеся в результате митоза, содержат одинаковый набор хромосом и одни и те же гены. Следовательно, митоз — это способ деления клеток, заключающийся в точном распределении генетического материала между дочерними клетками.
В результате митоза обе дочерние клетки получают диплоидный набор хромосом.
Митоз тормозитсявысокой температурой, высокими дозами ионизирующей радиации, действием растительных ядов. Один из таких ядов — колхицин — применяют в цитогенетике: с его помощью можно остановить митоз на стадии метафазной пластинки, что позволяет подсчитать число хромосом и дать каждой из них индивидуальную характеристику, т. е. провести кариотипирование.
Таблица Митотический цикл и митоз (Т.Л. Богданова. Биология. Задания и упражнения. Пособие для поступающих в ВУЗы. М.,1991)
|
Фазы |
Процесс, происходящий в клетке |
|
|
Интерфаза (фаза между делениями клеток) |
Пресинтетический период |
Синтез белка. |
|
Синтетический период |
Синтез ДНК — самоудвоение молекулы ДНК. Построение второй хроматиды, в которую переходит вновь образовавшаяся молекула ДНК: получаются двухроматидные хромосомы |
|
|
Постсинтетический период |
Синтез белка, накопление энергии, подготовка к делению |
|
|
Профаза (первая фаза деления) |
Двухроматидные хромосомы спирализуются, ядрышки растворяются, центриоли расходятся, ядерная оболочка растворяется, образуются нити веретена деления |
|
|
Фазы митоза |
Метафаза (фаза скопления хромосом) |
Нити веретена деления присоединяются к центромерам хромосом, двухроматидные хромосомы сосредоточиваются на экваторе клетки |
|
Анафаза (фаза расхождения хромосом) |
Центромеры делятся, однохроматидные хромосомы растягиваются нитями веретена деления к полюсам клетки |
|
|
Телофаза (фаза окончания деления) |
Однохроматидные хромосомы деспирализуются, сформировывается ядрышко, восстанавливается ядерная оболочка, на экваторе начинает закладываться перегородка между клетками, растворяются нити веретена деления |
|
 На деспирализованных молекулах ДНК синтезируется РНК
На деспирализованных молекулах ДНК синтезируется РНКОсобенности митоза у растений и у животных
|
Растительная клетка |
Животная клетка |

Тест по биологии на тему «Деление клетки. Митоз. Мейоз» (9 класс)
Деление клетки. Митоз. Мейоз
Вариант 1
1. Что такое митоз?
1) деление всех клеток
2) деление клетки одноклеточного организма
3) деление прокариотической клетки
4) деление эукариотической клетки, при котором образуются две дочерние клетки с идентичным родительскому набором хромосом
2. Какие клетки в организме человека не способны к делению?
Какие клетки в организме человека не способны к делению?
1) остеобласты
2) фибробласты
3) нейроны
4) росткового слоя эпидермиса
3. Из чего состоит хромосома?
1) из центромер
2) из хроматид
3) из микротрубочек
4) из веретен деления
4. В какой фазе деления клетки хроматиды расходятся к противоположным полюсам клетки?
1) в анафазе 3) в телофазе
2)в профазе 4)в метафазе
5. В ядре клетки листа томата 24 хромосомы. Сколько хромосом будет в ядре клетки корня томата после ее деления?
1) 12
2) 48
3) 36
4) 24
6.В приведённой ниже таблице между позициями первого и второго столбца имеется взаимосвязь.
Часть
Целое
Хромосома
…..
Кровеносная система
Сердце
Какое понятие следует вписать на место пропуска в этой таблице?
1) ткань
2) клетка
3) ядро
4) ген
7. В метафазе митоза происходит
В метафазе митоза происходит
1) расхождение хроматид
2) удвоение хромосом
3)размещение хромосом в плоскости экватора клетки
4)формирование ядерной оболочки и ядрышек
8.Мейоз происходит в клетках
1) крови лягушки
2) половых желез крысы
3) камбия тополя
4) эпителия человека
9.Конъюгация и обмен участками гомологичных хромосом происходит в
1) профазе II мейоза
2) профазе митоза
3) метафазе II мейоза
4) профазе I мейоза
10..Какая фаза деления клетки изображена на рисунке?
1) профаза
2) анафаза
3) метафаза
4) телофаза
11.По каким признакам можно узнать телофазу митоза?
беспорядочному расположению
1) спирализованных хромосом в цитоплазме
выстраиванию хромосом в
2) экваториальной плоскости клетки
расхождению дочерних
3) хроматид к противоположным полюсам клетки
деспирализации хромосом и
4) образованию ядерных оболочек вокруг двух ядер
12. Какие процессы протекают во время мейоза?
Какие процессы протекают во время мейоза?
1) транскрипция
2) редукционное деление
3) денатурация
4) трансляция
5) конъюгация
6) кроссинговер
13.Установите, в какой последовательности происходят фазы митоза.
А) расхождение сестринских хроматид
Б) удвоение молекулы ДНК
В) образование метафазной пластинки
Г) деление цитоплазмы
14.Верны ли следующие суждения о митозе?
А. Митоз — способ деления клеток, в результате которого образуются клетки с редуцированным набором хромосом.
Б. Образующиеся в результате митоза клетки содержат наследственную информацию, идентичную информации материнской клетки.
1) верно только А
2) верно только Б
3) верны оба суждения
4) оба суждения неверны
15.Установите соответствие между характеристикой процесса и способом деления клетки, который она иллюстрирует.
ОСОБЕННОСТЬ ДЕЛЕНИЯ
А)образуются две диплоидные дочерние клетки
Б)обеспечивает созревание гамет у животных
В) сохраняет постоянство числа хромосом в клетках
Г) происходит перекомбинация генов в хромосомах
Д) служит способом бесполого размножения простейших
СПОСОБ ДЕЛЕНИЯ
1) митоз
2) мейоз
Деление клетки. Митоз. Мейоз
Митоз. Мейоз
Вариант 2
1. Для каких организмов характерно деление клеток?
1)эукариот
2) прокариот
3) всех организмов
4) многоклеточных организмов
2. Процесс деления клеток в живом организме заканчивается:
1) вместе с его ростом
2) с его смертью
3) после полового созревания
4) после его размножения
3. Укажите вариант ответа, где стадии митоза даны в правильной последовательности.
1) телофаза — анафаза — метафаза — профаза
2) метафаза—профаза — телофаза—анафаза
3) профаза — метафаза — анафаза — телофаза
4) анафаза—метафаза — профаза—телофаза
4. Что происходит в телофазе митоза ?
1) формирование веретена деления
2) формирование новых ядер и цитокинез
3) разделение хромосом
4) перемещение хромосом в центр клетки
5.Сколько хромосом будет содержаться в клетках кожи у четвертого поколения собаки Герды, если у неё в этих клетках содержится 78 хромосом?
1) 39
2) 325
3) 156
4) 78
6. В приведённой ниже таблице между позициями первого и второго столбца имеется взаимосвязь.
В приведённой ниже таблице между позициями первого и второго столбца имеется взаимосвязь.
Объект
Процесс
ядро
хранение информации
…
деление клетки
Какое понятие следует вписать на место пропуска в этой таблице?
1) митохондрия
2) клеточный центр
3) рибосома
4) вакуоль
7.Расхождение хроматид к полюсам клетки происходит в
1) метафазе
2) телофазе
3) профазе
4) анафазе
8.Какие признаки характерны для метафазы митоза?
1) спирализация хромосом
2) выстраивание хромосом в экваториальной плоскости клетки
3) деление центромеры и расхождение хромосом к полюсам клетки
4) деспирализация хромосом, образование двух ядер
9.Обмен между участками гомологичных хромосом происходит в процессе
1) синтеза иРНК
2) кроссинговера
3) редупликации ДНК
4) образования двух хроматид
10. Расхождение хромосом происходит в
Расхождение хромосом происходит в
1) анафазе-1 мейоза
2) метафазе-1 мейоза
3) анафазе-2 мейоза
4) метафазе-2 мейоза
11.Верны ли следующие суждения о мейозе?
А.Мейоз — способ деления клеток, приводящий к уменьшению вдвое числа хромосом.
Б. Мейоз состоит из двух последовательных делений, которым предшествует однократное удвоение ДНК в интерфазе.
1) верно только А
2) верно только Б
3) верны оба суждения
4) оба суждения неверны
12.Какая фаза деления клетки изображена на рисунке?
1) телофаза
2) метафаза
3) анафаза
4) профаза
13 .Биологическое значение мейоза заключается в
1)предотвращении удвоения числа хромосом в каждом новом поколении
2) образовании мужских и женских гамет
3) образовании соматических клеток
4)создании возможностей возникновения новых генных комбинаций
5) увеличении числа клеток в организме
6) кратном увеличении набора хромосом
14. Установите последовательность изменений, происходящих с хромосомами в процессе митоза.
Установите последовательность изменений, происходящих с хромосомами в процессе митоза.
А) деление центромеры и образование из хроматид хромосом
Б) расхождение гомологичных хроматид к разным полюсам клетки
В) расположение хромосом в плоскости экватора
Г) свободное расположение хромосом в цитоплазме
15.Установите соответствие между характеристикой процесса и способом деления клетки, который она иллюстрирует.
ХАРАКТЕРИСТИКА
А) расхождение к полюсам гомологичных хромосом
Б) конъюгация гомологичных хромосом
В) образование четырёх гаплоидных дочерних клеток
Г) образование двух дочерних клеток с числом хромосом, равным материнской клетке
Д) обмен генами между хроматидами гомологичных хромосом
СПОСОБ ДЕЛЕНИЯ
1) мейоз
2) митоз
МАТРИЦА ОТВЕТОВ
Задание
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
Ответ
ВАРИАНТ 1
Задание
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
Ответ
4
3
2
1
4
4
3
2
4
2
4
256
БВАГ
2
12121
ВАРИАНТ 2
Задание
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
Ответ
3
2
3
2
4
2
4
1
2
1
3
2
124
ГВАБ
11121
Деление клетки.
 Митоз | Параграф 2.14
Митоз | Параграф 2.14
«Введение в общую биологию и экологию. 9 класс». А.А. Каменский (гдз)
Вопрос 1. В чем биологическое значение митоза?
Биологический смысл митоза.
В результате митоза образуется две дочерние клетки с таким же набором хромосом, как и материнская клетка. Значение митоза:
1. Генетическая стабильность, т.к. хроматиды образуются в результате репликации, т.е. наследственная информация их идентична материнской.
2. Рост организмов, т.к. в результате митоза число клеток увеличивается.
3. Бесполое размножение – многие виды растений и животных размножаются в результате митотического деления.
4. Регенерация и замещение клеток идет за счет митоза.
Вопрос 2. Какие фазы включает в себя митоз?
Митоз (кариокинез) – это непрямое деление клетки, в котором выделяют фазы: профаза, метафаза, анафаза и телофаза.
1. Профаза характеризуется:
1) хромонемы спирализуются, утолщаются и укорачиваются.
2) ядрышки исчезают, т.е. хромонема ядрышка упаковывается к хромосомам, имеющим вторичную перетяжку, которую называют ядрышковый организатор.
3) в цитоплазме образуется два клеточных центра (центриолей) и формируются нити веретена деления.
4) в конце профазы, распадается ядерная оболочка и хромосомы оказываются в цитоплазме. Набор хромосом профазы составляет — 2п4с.
2. Метафаза характеризуется:
1) к центромерам хромосом прикрепляются нити веретена деления и хромосомы начинают двигаться и выстраиваются на экваторе клетки.
2) метафазу называют «паспортом клетки», т.к. хорошо видно, что хромосома состоит из двух хроматид. Хромосомы максимально спирализованы, хроматиды начинают отталкиваться друг от друга, но еще соединены в области центромера. На этой стадии изучают кариотип клеток, т.к. четко видно число и форма хромосом. Фаза очень короткая.
Фаза очень короткая.
Набор хромосом метафазы составляет — 2п4с.
3. Анафаза характеризуется:
1) центромеры хромосом делятся и сестринские хроматиды расходятся к полюсам клетки и становятся самостоятельными хроматидами, которые называют дочерними хромосомами. На каждом полюсе в клетке находится по диплоидному набору хромосом.
Набор хромосом анафазы составляет — 4п4с.
4. Телофаза характеризуется:
Однохроматидные хромосомы деспирализуются у полюсов клетки, образуются ядрышки, восстанавливается ядерная оболочка.
Набор хромосом телофазы составляет — 2п2с.
4. Телофаза заканчивается цитокинезом. Цитокинез – процесс разделения цитоплазмы между двумя дочерними клетками. Цитокинез происходит по разному у растений и животных.
В животной клетке. На экваторе клетки появляется кольцевидная перетяжка, которая углубляется и полностью перешнуровывает тело клетки. В результате образуется две новые клетки вдвое меньше материнской клетки.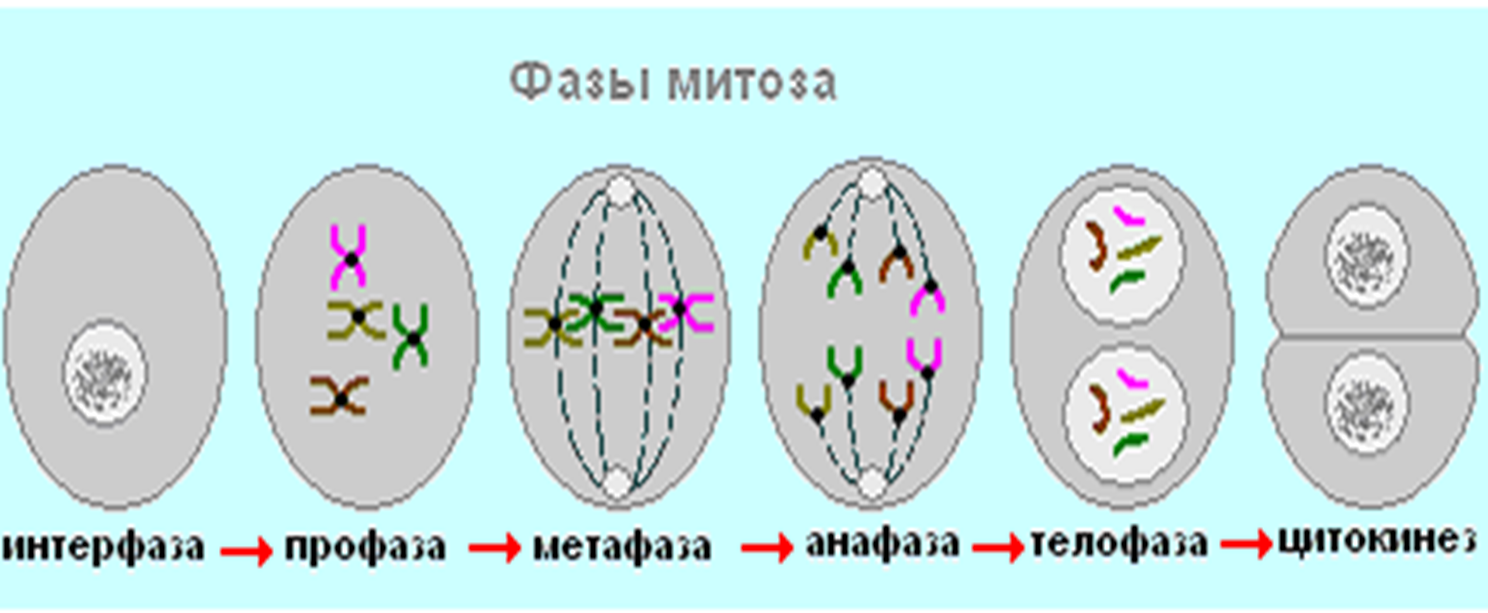 В области перетяжки много актина, т.е. в движении играют роль микрофиламенты. Цитокинез идет путем перетяжки.
В области перетяжки много актина, т.е. в движении играют роль микрофиламенты. Цитокинез идет путем перетяжки.
В растительной клетке. На экваторе, в центре клетки в результате скопления пузырьков диктиосом комплекса Гольджи, образуется клеточная пластинка, которая разрастается от центра к периферии и приводит к разделению материнской клетки на две клетки. В дальнейшем перегородка утолщается, за счет отложения целлюлозы, образуя клеточную стенку. Цитокинез идет путем перегородки.
Вопрос 3. Что такое редупликация ДНК?
Редупликацией называется удвоение молекулы ДНК в период интерфазы. Под воздействием фермента водородные связи между комплементарными азотистыми основаниями разрываются. Нити, составляющие двойную спираль ДНК, расходятся. Из свободных нуклеотидов согласно принципу комплементарности достраиваются вторые цепи образовавшихся нитей ДНК. В результате из одной материнской молекулы возникают две идентичные дочерние молекулы ДНК.
Вопрос 4. Что происходит в интерфазу для подготовки деления клетки?
Что происходит в интерфазу для подготовки деления клетки?
Во время интерфазы идет интенсивная подготовка клетки к делению, которая заключается в следующем:
• происходит редупликация ДНК;
• увеличивается число многих органоидов, в том числе митохондрий, центриолей и других;
• синтезируется и запасается АТФ, которая необходима для процессов последующего деления.
Вопрос 5. В какой фазе происходит деление цитоплазмы клетки?
Цитокинез – процесс разделения цитоплазмы между двумя дочерними клетками происходит в последнюю фазу митоза – телофазу.
Эукариотический клеточный цикл — Клетка
Цикл деления большинства клеток состоит из четырех скоординированных процессов: клеточного роста, репликации ДНК, распространения дуплицированных хромосом в дочерние клетки и клеточного деления. У бактерий клеточный рост и репликация ДНК происходят на протяжении большей части клеточного цикла, а дублированные хромосомы распределяются между дочерними клетками вместе с плазматической мембраной.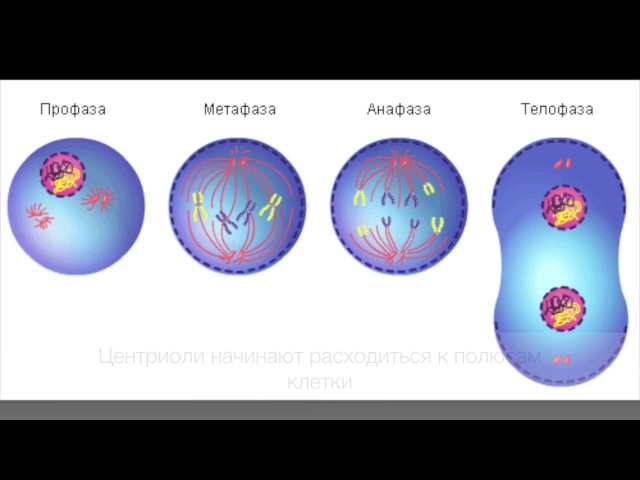 Однако у эукариот клеточный цикл более сложен и состоит из четырех дискретных фаз.Хотя клеточный рост обычно представляет собой непрерывный процесс, ДНК синтезируется только во время одной фазы клеточного цикла, а затем реплицированные хромосомы распределяются по дочерним ядрам в результате сложной серии событий, предшествующих клеточному делению. Переход между этими стадиями клеточного цикла контролируется консервативным регуляторным аппаратом, который не только координирует различные события клеточного цикла, но также связывает клеточный цикл с внеклеточными сигналами, которые контролируют клеточную пролиферацию.
Однако у эукариот клеточный цикл более сложен и состоит из четырех дискретных фаз.Хотя клеточный рост обычно представляет собой непрерывный процесс, ДНК синтезируется только во время одной фазы клеточного цикла, а затем реплицированные хромосомы распределяются по дочерним ядрам в результате сложной серии событий, предшествующих клеточному делению. Переход между этими стадиями клеточного цикла контролируется консервативным регуляторным аппаратом, который не только координирует различные события клеточного цикла, но также связывает клеточный цикл с внеклеточными сигналами, которые контролируют клеточную пролиферацию.
Фазы клеточного цикла
Типичный эукариотический клеточный цикл иллюстрируют человеческие клетки в культуре, которые делятся примерно каждые 24 часа.С точки зрения микроскопа клеточный цикл делится на две основные части: митоз и интерфазу. Митоз (деление ядра) — наиболее драматичная стадия клеточного цикла, соответствующая расхождению дочерних хромосом и обычно заканчивающаяся клеточным делением (цитокинез).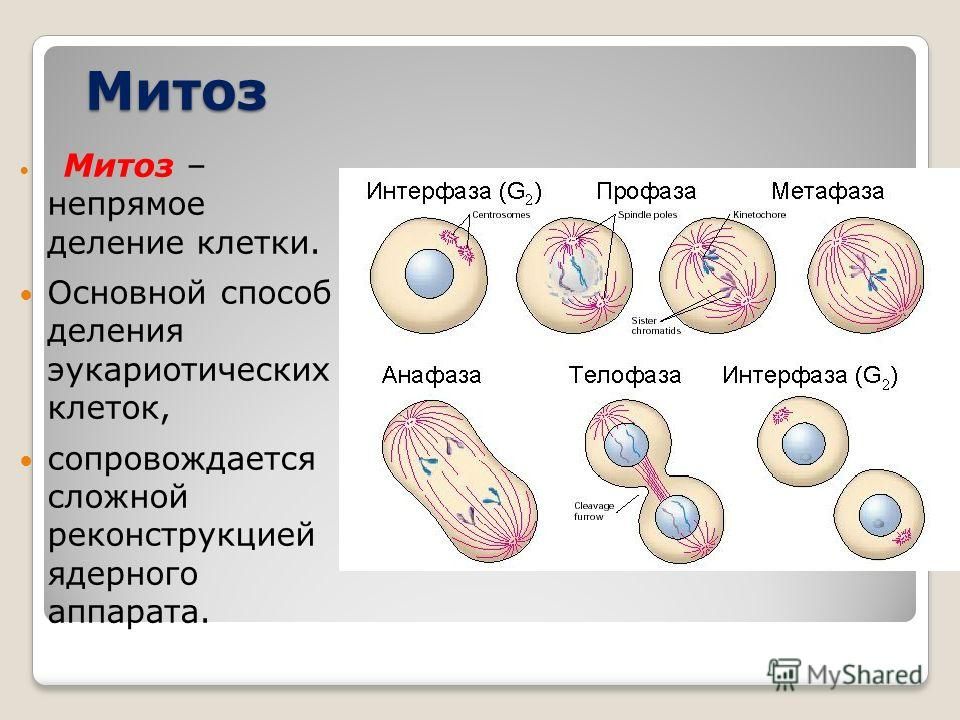 Однако митоз и цитокинез длятся всего около часа, поэтому примерно 95 % клеточного цикла приходится на интерфазу — период между митозами. Во время интерфазы хромосомы деконденсируются и распределяются по ядру, поэтому ядро выглядит морфологически однородным.Однако на молекулярном уровне интерфаза — это время, в течение которого и рост клеток, и репликация ДНК происходят упорядоченным образом при подготовке к клеточному делению.
Однако митоз и цитокинез длятся всего около часа, поэтому примерно 95 % клеточного цикла приходится на интерфазу — период между митозами. Во время интерфазы хромосомы деконденсируются и распределяются по ядру, поэтому ядро выглядит морфологически однородным.Однако на молекулярном уровне интерфаза — это время, в течение которого и рост клеток, и репликация ДНК происходят упорядоченным образом при подготовке к клеточному делению.
Клетка растет с постоянной скоростью на протяжении всей интерфазы, при этом большинство делящихся клеток удваиваются в размере между одним митозом и другим. Напротив, ДНК синтезируется только в течение части интерфазы. Таким образом, синхронизация синтеза ДНК делит цикл эукариотических клеток на четыре дискретные фазы (4). Фаза М цикла соответствует митозу, за которым обычно следует цитокинез.За этой фазой следует фаза G 1 (пробел 1), которая соответствует интервалу (пробелу) между митозом и инициацией репликации ДНК. Во время G 1 клетка метаболически активна и постоянно растет, но не реплицирует свою ДНК. За G 1 следует S-фаза (синтез), во время которой происходит репликация ДНК. За завершением синтеза ДНК следует фаза (пробел 2), во время которой продолжается рост клеток и происходит синтез белков для подготовки к митозу.
Во время G 1 клетка метаболически активна и постоянно растет, но не реплицирует свою ДНК. За G 1 следует S-фаза (синтез), во время которой происходит репликация ДНК. За завершением синтеза ДНК следует фаза (пробел 2), во время которой продолжается рост клеток и происходит синтез белков для подготовки к митозу.
Рисунок 14.1
Фазы клеточного цикла. Цикл деления большинства эукариотических клеток делится на четыре отдельные фазы: M, G 1 , S и G 2 . Фаза М (митоз) обычно сопровождается цитокинезом. S-фаза – это период, в течение которого происходит репликация ДНК. Клетка растет (подробнее…)
Продолжительность этих фаз клеточного цикла значительно различается у разных типов клеток. Для типичной быстро пролиферирующей клетки человека с общим временем цикла 24 часа фаза G 1 может длиться около 11 часов, фаза S — около 8 часов, G 2 — около 4 часов и М — около 1 часа.Однако другие типы клеток могут делиться намного быстрее.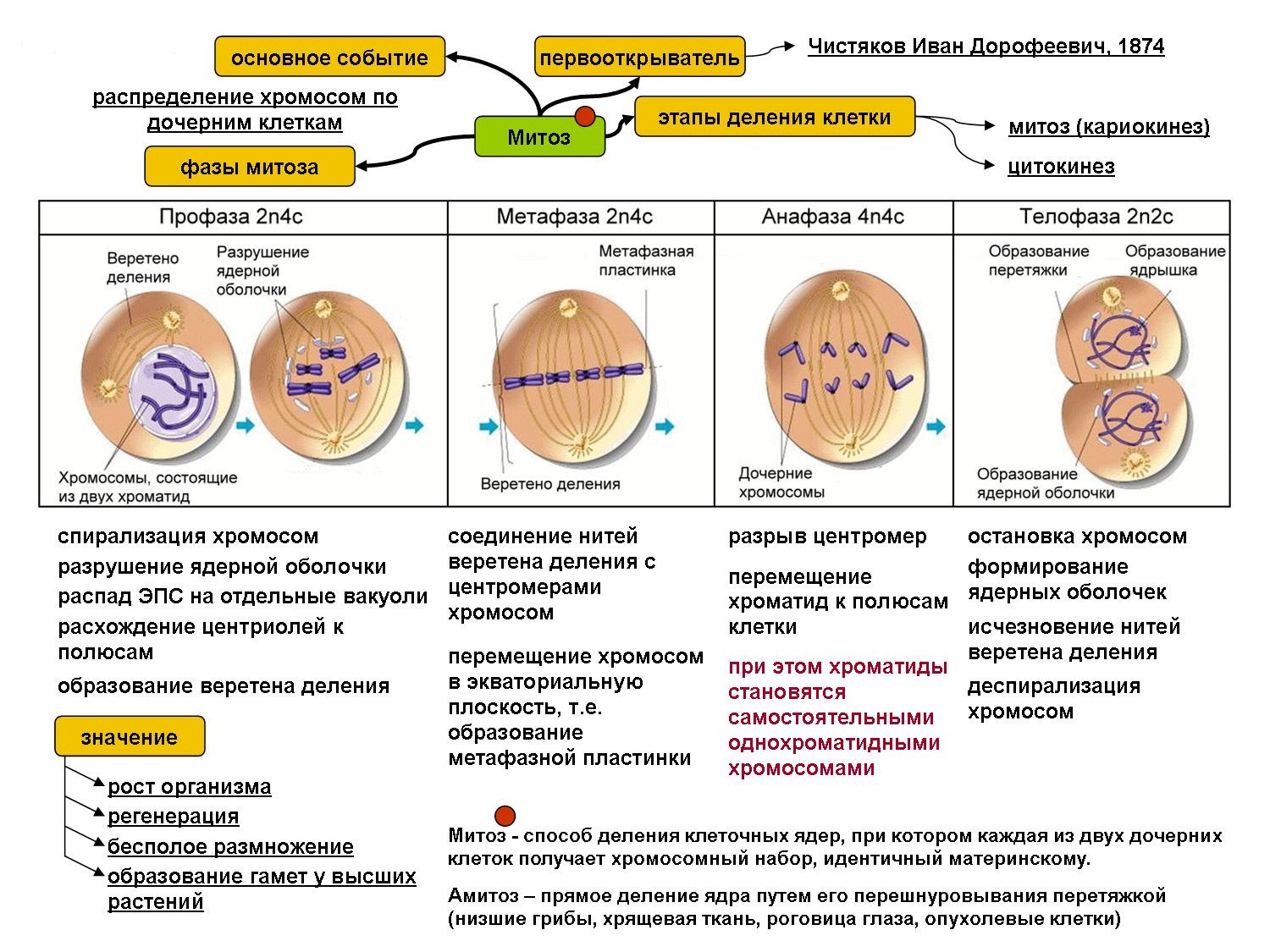 Почкующиеся дрожжи, например, могут пройти все четыре стадии клеточного цикла всего за 90 минут. Еще более короткие клеточные циклы (30 минут или меньше) встречаются в ранних эмбриональных клетках вскоре после оплодотворения яйцеклетки (14). Однако в этом случае роста клеток не происходит. Вместо этого эти ранние эмбриональные клеточные циклы быстро делят цитоплазму яйца на более мелкие клетки. Фаза G 1 или G 2 отсутствует, и репликация ДНК происходит очень быстро в этих ранних эмбриональных клеточных циклах, которые, следовательно, состоят из очень коротких S фаз, чередующихся с M фазами.
Почкующиеся дрожжи, например, могут пройти все четыре стадии клеточного цикла всего за 90 минут. Еще более короткие клеточные циклы (30 минут или меньше) встречаются в ранних эмбриональных клетках вскоре после оплодотворения яйцеклетки (14). Однако в этом случае роста клеток не происходит. Вместо этого эти ранние эмбриональные клеточные циклы быстро делят цитоплазму яйца на более мелкие клетки. Фаза G 1 или G 2 отсутствует, и репликация ДНК происходит очень быстро в этих ранних эмбриональных клеточных циклах, которые, следовательно, состоят из очень коротких S фаз, чередующихся с M фазами.
Рисунок 14.2
Эмбриональные клеточные циклы. Ранние эмбриональные клеточные циклы быстро делят цитоплазму яйца на более мелкие клетки. Клетки не растут во время этих циклов, в которых отсутствуют G 1 и G 2 и которые состоят просто из коротких S фаз, чередующихся с M фазами.
В отличие от быстрой пролиферации эмбриональных клеток, некоторые клетки у взрослых животных полностью прекращают деление (например, нервные клетки), а многие другие клетки делятся только изредка, по мере необходимости, чтобы заменить клетки, которые были потеряны из-за повреждения или гибели клеток.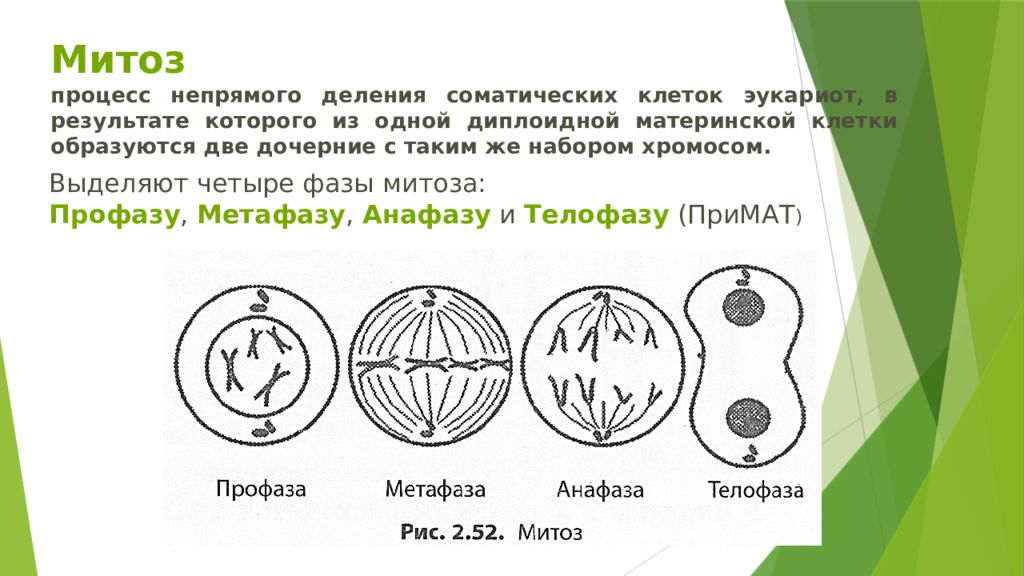 К клеткам последнего типа относятся кожные фибробласты, а также клетки многих внутренних органов, таких как печень, почки и легкие. Как обсуждается далее в следующем разделе, эти клетки выходят из G 1 и вступают в покоящуюся стадию цикла, называемую G 0 , где они остаются метаболически активными, но больше не размножаются, если к этому не призывают соответствующие меры. внеклеточные сигналы.
К клеткам последнего типа относятся кожные фибробласты, а также клетки многих внутренних органов, таких как печень, почки и легкие. Как обсуждается далее в следующем разделе, эти клетки выходят из G 1 и вступают в покоящуюся стадию цикла, называемую G 0 , где они остаются метаболически активными, но больше не размножаются, если к этому не призывают соответствующие меры. внеклеточные сигналы.
Анализ клеточного цикла требует идентификации клеток на различных стадиях, описанных выше.Хотя митотические клетки можно различить под микроскопом, клетки в других фазах цикла (G 1 , S и G 2 ) необходимо идентифицировать по биохимическим критериям. Клетки в S-фазе можно легко идентифицировать, поскольку они содержат радиоактивный тимидин, который используется исключительно для синтеза ДНК (4). Например, если популяция быстро пролиферирующих клеток человека в культуре подвергается воздействию радиоактивного тимидина в течение короткого периода времени (например, 15 минут), а затем анализируется с помощью авторадиографии, то обнаруживается, что примерно треть клеток имеет радиоактивную метку. соответствует доле клеток в S-фазе.
соответствует доле клеток в S-фазе.
Рисунок 14.3
Идентификация клеток S фазы путем включения радиоактивного тимидина. Клетки подвергали воздействию радиоактивного тимидина и анализировали авторадиографией. Помеченные ячейки указаны стрелками. (Из DW Stacey et al., 1991. Mol. Cell Biol. 11: 4053.) (подробнее…)
Вариации таких экспериментов по мечению клеток также можно использовать для определения длины различных стадий клетки. цикл. Например, рассмотрим эксперимент, в котором клетки подвергают воздействию радиоактивного тимидина в течение 15 минут, после чего радиоактивный тимидин удаляют и клетки культивируют в течение различных периодов времени перед авторадиографией.Радиоактивно меченые интерфазные клетки, находившиеся в S-фазе во время воздействия радиоактивного тимидина, будут наблюдаться в течение нескольких часов по мере прохождения оставшейся части S и G 2 . Напротив, радиоактивно меченные митотические клетки не будут наблюдаться до 4 часов после мечения. Это 4-часовое запаздывание соответствует длине G 2 — минимальному времени, необходимому для того, чтобы клетка, включившая радиоактивный тимидин в конце S-фазы, вступила в митоз.
Это 4-часовое запаздывание соответствует длине G 2 — минимальному времени, необходимому для того, чтобы клетка, включившая радиоактивный тимидин в конце S-фазы, вступила в митоз.
Клетки на разных стадиях клеточного цикла также можно отличить по содержанию ДНК ().Например, клетки животных в G 1 являются диплоидными (содержат две копии каждой хромосомы), поэтому содержание их ДНК обозначается как 2 n ( n обозначает гаплоидное содержание ДНК в геноме). Во время фазы S репликация увеличивает содержание ДНК в клетке с 2 n до 4 n , поэтому клетки в S имеют содержание ДНК в диапазоне от 2 n до 4 n . Затем содержание ДНК остается на уровне 4 н для клеток в G 2 и М, снижаясь до 2 н после цитокинеза.Экспериментально содержание клеточной ДНК можно определить путем инкубации клеток с флуоресцентным красителем, который связывается с ДНК, с последующим анализом интенсивности флуоресценции отдельных клеток в проточном цитометре или флуоресцентно-активированном клеточном сортере , тем самым различая клетки в G 1 , S и G 2 /M фазы клеточного цикла.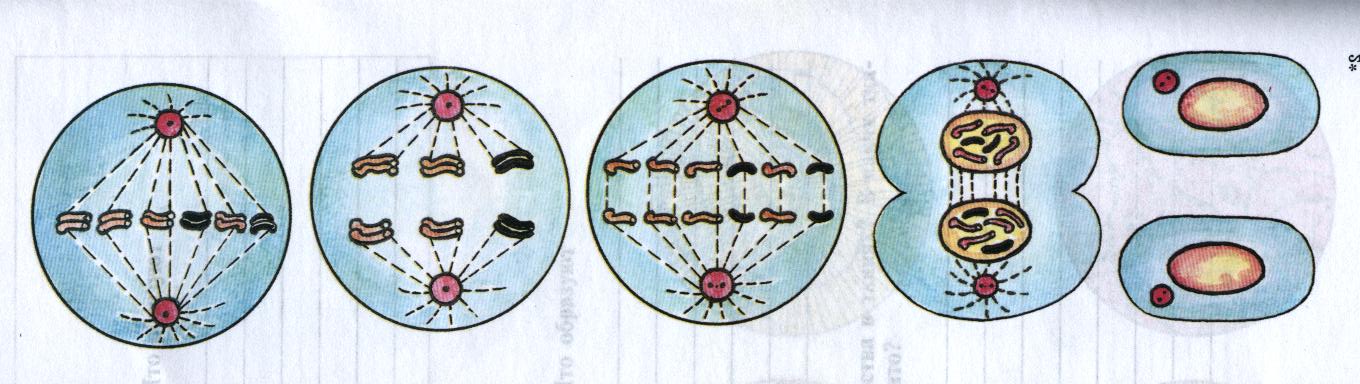
Рисунок 14.4
Определение содержания клеточной ДНК. Популяцию клеток метят флуоресцентным красителем, который связывает ДНК.Затем клетки пропускают через проточный цитометр, который измеряет интенсивность флуоресценции отдельных клеток. Данные представлены в виде клеток (подробнее…)
Регуляция клеточного цикла с помощью роста клеток и внеклеточных сигналов
Прохождение клеток через цикл деления регулируется внеклеточными сигналами из окружающей среды, а также внутренними сигналами которые контролируют и координируют различные процессы, происходящие на разных фазах клеточного цикла. Примером регуляции клеточного цикла внеклеточными сигналами может служить влияние факторов роста на пролиферацию клеток животных.Кроме того, различные клеточные процессы, такие как клеточный рост, репликация ДНК и митоз, должны координироваться во время развития клеточного цикла. Это достигается серией контрольных точек, которые регулируют прохождение через различные фазы клеточного цикла.
Основная регуляторная точка клеточного цикла во многих типах клеток возникает поздно в G 1 и контролирует переход от G 1 к S. Эта регуляторная точка была впервые определена в исследованиях почкующихся дрожжей ( Saccharomyces cerevisiae ), где он известен как СТАРТ ().После того, как клетки прошли СТАРТ, они готовы вступить в S-фазу и пройти один цикл клеточного деления. Однако прохождение через START является строго регулируемым событием в клеточном цикле дрожжей, где оно контролируется внешними сигналами, такими как доступность питательных веществ, а также размером клетки. Например, если дрожжи сталкиваются с нехваткой питательных веществ, они останавливают свой клеточный цикл в момент НАЧАЛА и переходят в состояние покоя, а не в S-фазу. Таким образом, СТАРТ представляет собой точку принятия решения, в которой клетка определяет, доступно ли достаточное количество питательных веществ для поддержки прохождения оставшейся части цикла деления.Полипептидные факторы, которые сигнализируют о спаривании дрожжей, также останавливают клеточный цикл в START, позволяя гаплоидным дрожжевым клеткам сливаться друг с другом вместо перехода в S-фазу.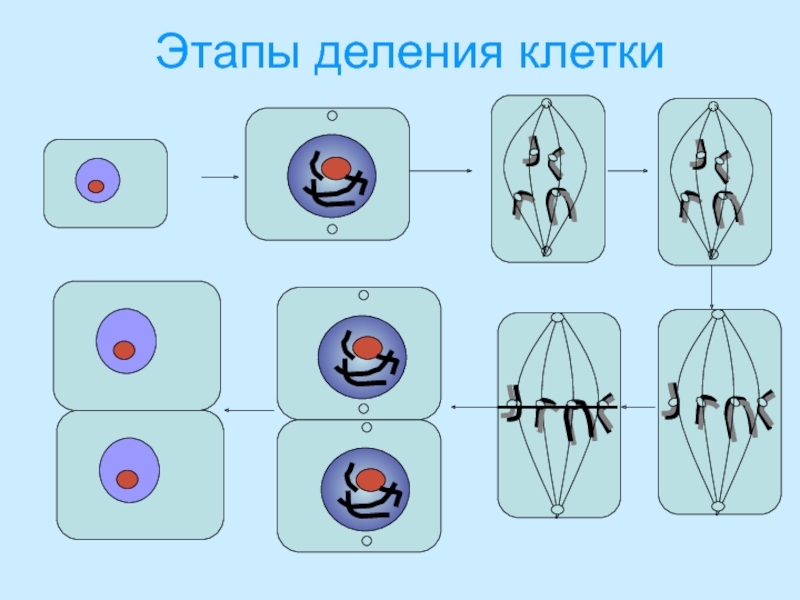
Рисунок 14.5
Регуляция клеточного цикла почкующихся дрожжей. (A) Клеточный цикл Saccharomyces cerevisiae регулируется преимущественно в конце G 1 , называемом START. Прохождение через START контролируется наличием питательных веществ, факторами спаривания и размером клеток. (более…)
Помимо использования в качестве точки принятия решения для мониторинга внеклеточных сигналов, START является точкой, в которой клеточный рост координируется с репликацией ДНК и клеточным делением. Важность этой регуляции особенно очевидна у почкующихся дрожжей, у которых при делении клеток образуются клетки-потомки самых разных размеров: большая материнская клетка и маленькая дочерняя клетка. Чтобы дрожжевые клетки сохраняли постоянный размер, маленькая дочерняя клетка должна расти больше, чем крупная материнская клетка, прежде чем снова делиться.Таким образом, необходимо контролировать размер клеток, чтобы координировать рост клеток с другими событиями клеточного цикла. Это регулирование осуществляется с помощью механизма управления, который требует, чтобы каждая ячейка достигла минимального размера, прежде чем она сможет пройти START. Следовательно, маленькая дочерняя клетка дольше находится в G 1 и растет больше, чем материнская клетка.
Это регулирование осуществляется с помощью механизма управления, который требует, чтобы каждая ячейка достигла минимального размера, прежде чем она сможет пройти START. Следовательно, маленькая дочерняя клетка дольше находится в G 1 и растет больше, чем материнская клетка.
Пролиферация большинства клеток животных аналогично регулируется в фазе G 1 клеточного цикла. В частности, точка принятия решения в конце G 1 , называемая точкой рестрикции в клетках животных, функционирует аналогично START у дрожжей (4).Однако, в отличие от дрожжей, прохождение животных клеток через клеточный цикл регулируется главным образом внеклеточными факторами роста, которые сигнализируют о пролиферации клеток, а не наличием питательных веществ. В присутствии соответствующих факторов роста клетки проходят точку рестрикции и вступают в S-фазу. После прохождения точки рестрикции клетка обязуется пройти через S-фазу и остальную часть клеточного цикла, даже в отсутствие дальнейшей стимуляции фактором роста.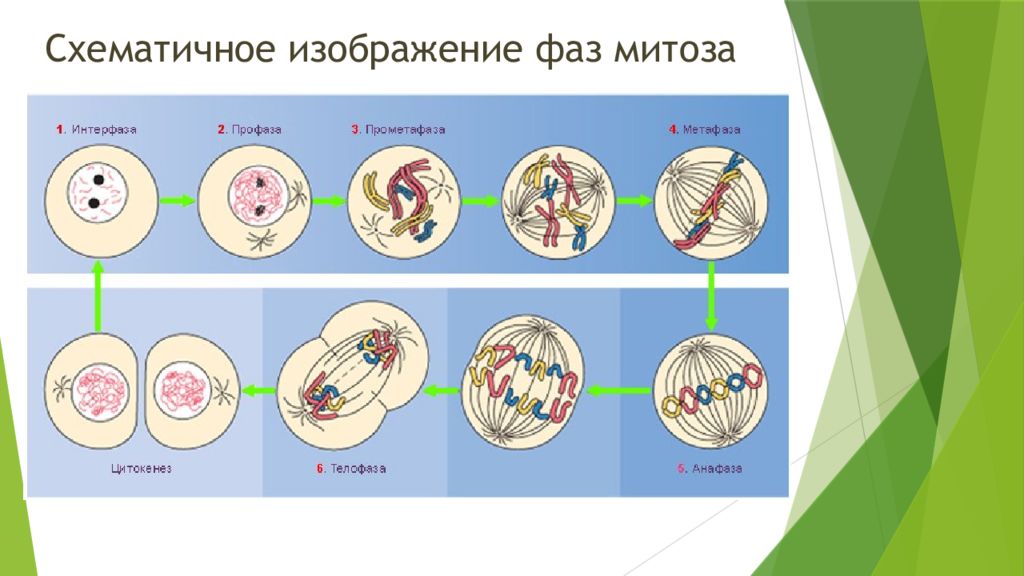 С другой стороны, если соответствующие факторы роста недоступны в G 1 , продвижение по клеточному циклу останавливается в точке рестрикции. Такие задержанные клетки затем вступают в стадию покоя клеточного цикла, называемую G 0 , в которой они могут оставаться в течение длительного периода времени без пролиферации. Клетки G 0 метаболически активны, хотя прекращают рост и снижают скорость синтеза белка. Как уже отмечалось, многие клетки у животных остаются в G 0 , если только соответствующие факторы роста или другие внеклеточные сигналы не призывают к пролиферации.Например, фибробласты кожи задерживаются в G 0 до тех пор, пока они не будут стимулированы к делению, необходимому для восстановления повреждений, полученных в результате раны. Пролиферация этих клеток запускается тромбоцитарным фактором роста, который высвобождается из тромбоцитов во время свертывания крови и сигнализирует о пролиферации фибробластов вблизи поврежденной ткани.
С другой стороны, если соответствующие факторы роста недоступны в G 1 , продвижение по клеточному циклу останавливается в точке рестрикции. Такие задержанные клетки затем вступают в стадию покоя клеточного цикла, называемую G 0 , в которой они могут оставаться в течение длительного периода времени без пролиферации. Клетки G 0 метаболически активны, хотя прекращают рост и снижают скорость синтеза белка. Как уже отмечалось, многие клетки у животных остаются в G 0 , если только соответствующие факторы роста или другие внеклеточные сигналы не призывают к пролиферации.Например, фибробласты кожи задерживаются в G 0 до тех пор, пока они не будут стимулированы к делению, необходимому для восстановления повреждений, полученных в результате раны. Пролиферация этих клеток запускается тромбоцитарным фактором роста, который высвобождается из тромбоцитов во время свертывания крови и сигнализирует о пролиферации фибробластов вблизи поврежденной ткани.
Рисунок 14.6
Регуляция клеточных циклов животных факторами роста. Доступность факторов роста контролирует клеточный цикл животных в точке в конце G 1 , называемой точкой рестрикции.Если факторы роста недоступны во время G 1 , клетки переходят в стадию покоя (подробнее…)
Хотя пролиферация большинства клеток регулируется главным образом в G 1 , некоторые клеточные циклы контролируются главным образом в G 2 . Одним из примеров является клеточный цикл делящихся дрожжей Schizosaccharomyces pombe (). В отличие от Saccharomyces cerevisiae , клеточный цикл S . pombe регулируется главным образом путем контроля перехода от G 2 к M, что является основной точкой, в которой отслеживаются размер клеток и доступность питательных веществ.У животных основным примером контроля клеточного цикла в G 2 являются ооциты. Ооциты позвоночных могут оставаться задержанными в G 2 в течение длительных периодов времени (несколько десятилетий у человека), пока их переход в М-фазу не будет вызван гормональной стимуляцией.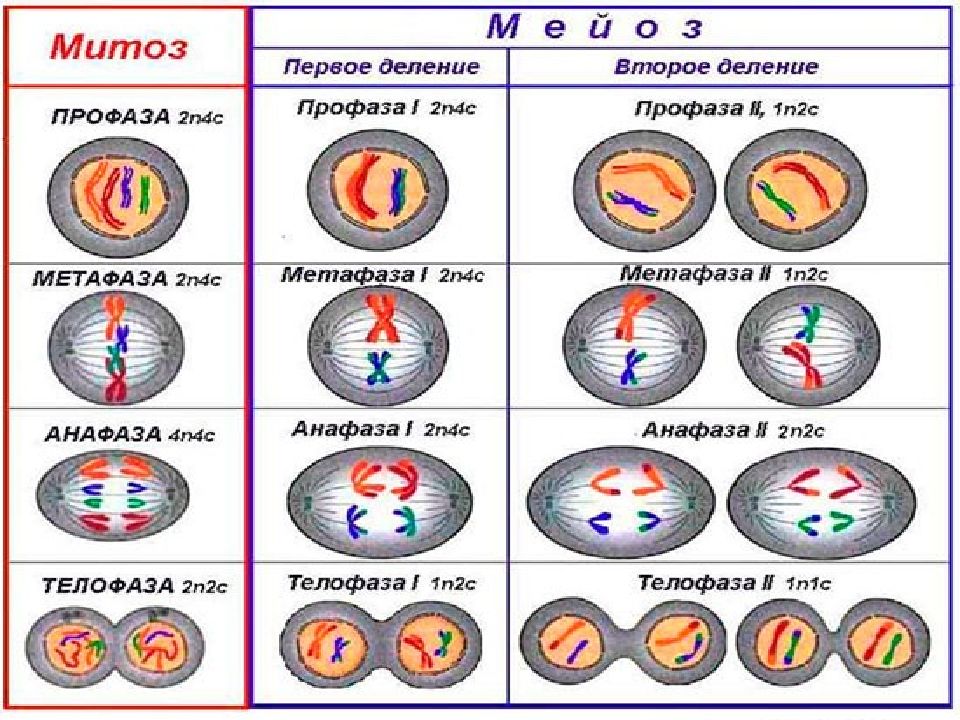 Таким образом, внеклеточные сигналы могут контролировать пролиферацию клеток, регулируя переход от G 2 к M, а также от G 1 к S фазам клеточного цикла.
Таким образом, внеклеточные сигналы могут контролировать пролиферацию клеток, регулируя переход от G 2 к M, а также от G 1 к S фазам клеточного цикла.
Рисунок 14.7
Клеточный цикл делящихся дрожжей.(A) Делящиеся дрожжи растут, удлиняясь с обоих концов, и делятся, образуя стенку через середину клетки. В отличие от цикла почкующихся дрожжей клеточный цикл делящихся дрожжей имеет нормальные фазы G 1 , S, G 2 и М. (подробнее…)
Контрольные точки клеточного цикла
Элементы управления, описанные в предыдущем разделе, регулируют ход клеточного цикла в зависимости от размера клеток и внеклеточных сигналов, таких как питательные вещества и факторы роста. Кроме того, события, происходящие на разных стадиях клеточного цикла, должны быть согласованы друг с другом, чтобы они происходили в соответствующем порядке.Например, критически важно, чтобы клетка не начинала митоз до завершения репликации генома. Альтернативой могло бы стать катастрофическое деление клетки, при котором дочерние клетки не смогли бы унаследовать полные копии генетического материала.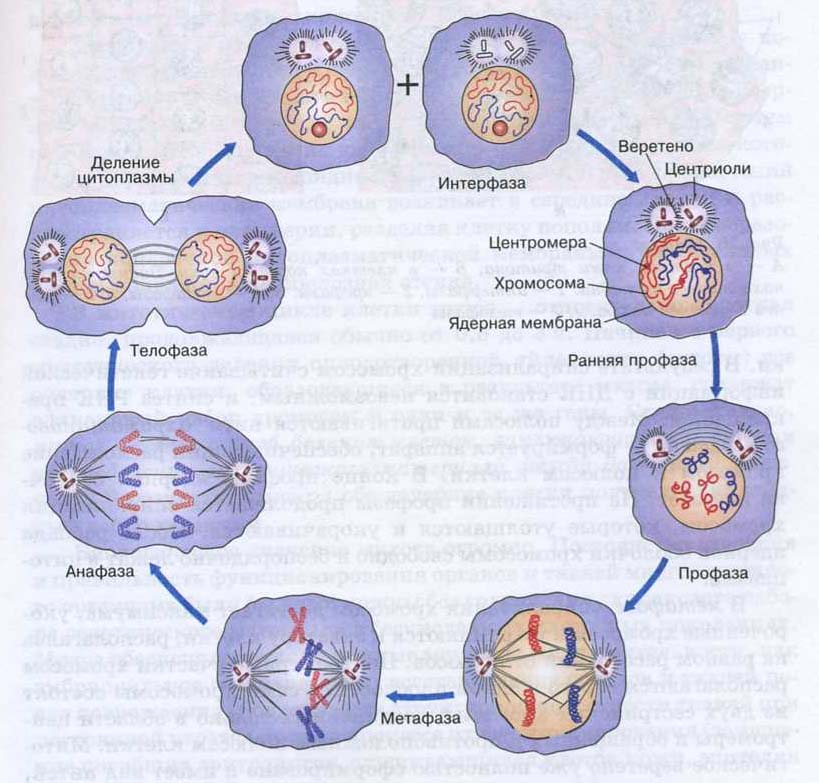 В большинстве клеток эта координация между различными фазами клеточного цикла зависит от системы контрольных точек и контроля с обратной связью, которые предотвращают переход к следующей фазе клеточного цикла до тех пор, пока не будут завершены события предыдущей фазы.
В большинстве клеток эта координация между различными фазами клеточного цикла зависит от системы контрольных точек и контроля с обратной связью, которые предотвращают переход к следующей фазе клеточного цикла до тех пор, пока не будут завершены события предыдущей фазы.
Несколько контрольных точек клеточного цикла служат для того, чтобы неполные или поврежденные хромосомы не реплицировались и не передавались дочерним клеткам (). Одна из наиболее четко определенных контрольных точек встречается в G 2 и предотвращает инициацию митоза до завершения репликации ДНК. Эта контрольная точка G 2 обнаруживает нереплицированную ДНК, которая генерирует сигнал, приводящий к остановке клеточного цикла. Таким образом, работа контрольной точки G 2 предотвращает инициацию M-фазы до завершения S-фазы, поэтому клетки остаются в G 2 до тех пор, пока геном не будет полностью реплицирован.Только тогда ингибирование прогрессии G 2 снимается, позволяя клетке инициировать митоз и распределять полностью реплицированные хромосомы по дочерним клеткам.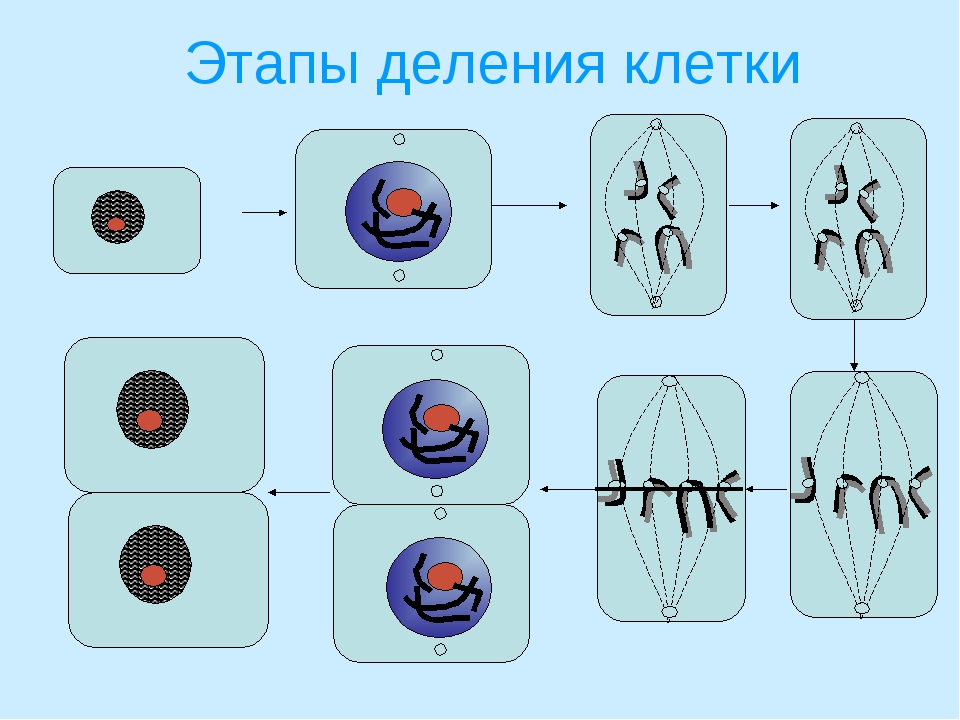
Рисунок 14.8
Контрольные точки клеточного цикла. Несколько контрольных точек обеспечивают передачу полных геномов дочерним клеткам. Одна из основных контрольных точек арестовывает клетки в G 2 в ответ на поврежденную или нереплицированную ДНК. Присутствие поврежденной ДНК также приводит к клеточному (подробнее…)
Прохождение клеточного цикла также останавливается на контрольной точке G 2 в ответ на повреждение ДНК, например, в результате облучения.Эта остановка дает время для устранения повреждений, а не для передачи их дочерним клеткам. Исследования мутантов дрожжей показали, что одна и та же контрольная точка клеточного цикла отвечает за остановку G 2 , вызванную либо нереплицированной, либо поврежденной ДНК, обе из которых сигнализируют об остановке клеточного цикла через родственные пути.
Повреждение ДНК не только останавливает клеточный цикл в G 2 , но также замедляет продвижение клеток через S фазу и останавливает прогрессирование клеточного цикла в контрольной точке в G 1 .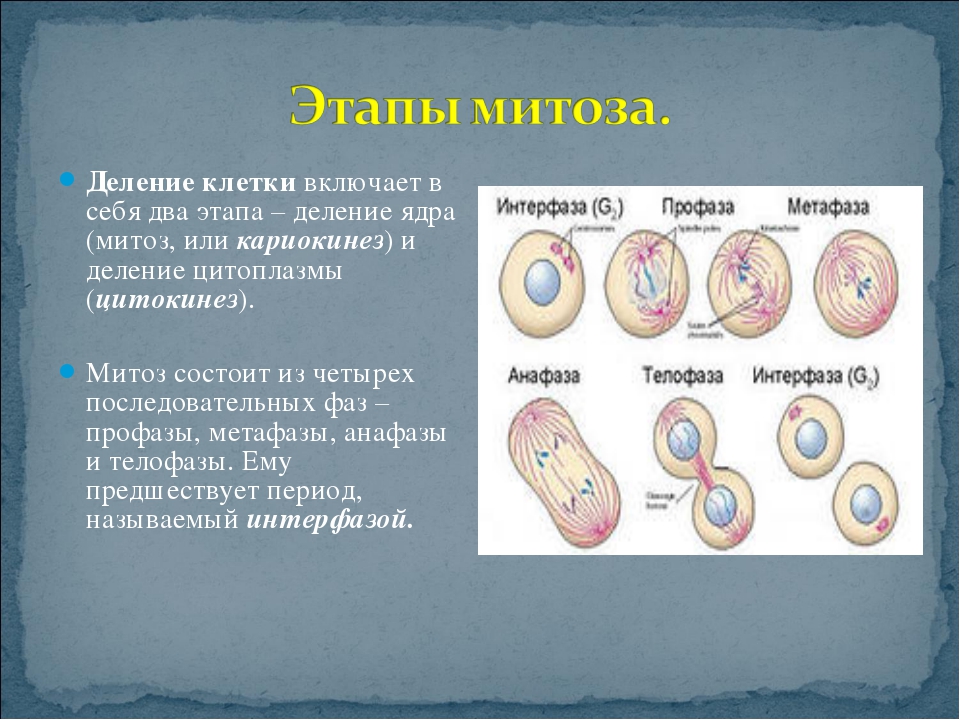 Эта остановка G 1 может позволить восстановить повреждение до того, как клетка войдет в S-фазу, где будет реплицироваться поврежденная ДНК. В клетках млекопитающих остановка в контрольной точке G 1 опосредована действием белка, известного как p53, который быстро индуцируется в ответ на поврежденную ДНК (4). Интересно, что ген, кодирующий p53, часто мутирует при раке человека. Потеря функции p53 в результате этих мутаций предотвращает остановку G 1 в ответ на повреждение ДНК, поэтому поврежденная ДНК реплицируется и передается дочерним клеткам вместо восстановления.Это наследование поврежденной ДНК приводит к повышенной частоте мутаций и общей нестабильности клеточного генома, что способствует развитию рака. Мутации в гене p53 являются наиболее распространенными генетическими изменениями при раке человека (см. главу 15), иллюстрируя критическую важность регуляции клеточного цикла в жизни многоклеточных организмов.
Эта остановка G 1 может позволить восстановить повреждение до того, как клетка войдет в S-фазу, где будет реплицироваться поврежденная ДНК. В клетках млекопитающих остановка в контрольной точке G 1 опосредована действием белка, известного как p53, который быстро индуцируется в ответ на поврежденную ДНК (4). Интересно, что ген, кодирующий p53, часто мутирует при раке человека. Потеря функции p53 в результате этих мутаций предотвращает остановку G 1 в ответ на повреждение ДНК, поэтому поврежденная ДНК реплицируется и передается дочерним клеткам вместо восстановления.Это наследование поврежденной ДНК приводит к повышенной частоте мутаций и общей нестабильности клеточного генома, что способствует развитию рака. Мутации в гене p53 являются наиболее распространенными генетическими изменениями при раке человека (см. главу 15), иллюстрируя критическую важность регуляции клеточного цикла в жизни многоклеточных организмов.
Рис. 14.9
Роль p53 в G 1 остановке, вызванной повреждением ДНК.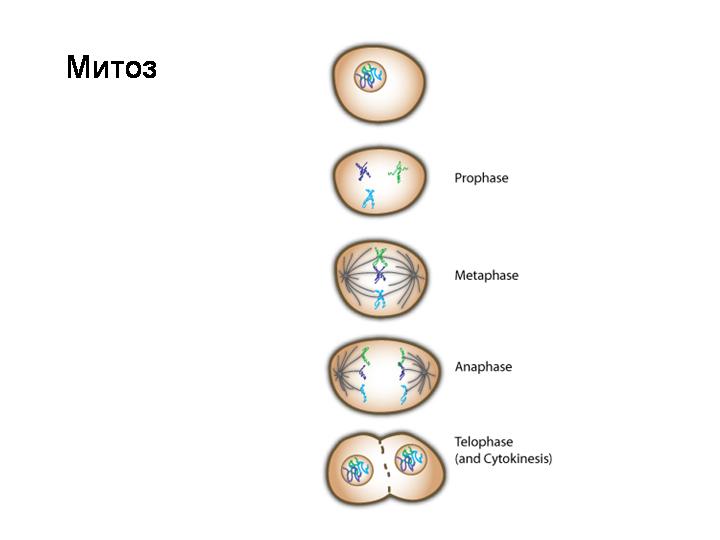 Повреждение ДНК, например, в результате облучения, приводит к быстрому повышению уровня р53.Затем белок p53 сигнализирует об остановке клеточного цикла в контрольной точке G 1 .
Повреждение ДНК, например, в результате облучения, приводит к быстрому повышению уровня р53.Затем белок p53 сигнализирует об остановке клеточного цикла в контрольной точке G 1 .
Другая важная контрольная точка клеточного цикла, которая поддерживает целостность генома, происходит ближе к концу митоза (см. ). Эта контрольная точка отслеживает выравнивание хромосом в митотическом веретене, гарантируя тем самым точное распределение полного набора хромосом по дочерним клеткам. Например, неспособность одной или нескольких хромосом правильно выровняться на веретене приводит к остановке митоза в метафазе до сегрегации вновь реплицированных хромосом в дочерние ядра.В результате этой контрольной точки хромосомы не разделяются до тех пор, пока не будет организован полный набор хромосом для распределения по каждой дочерней клетке.
Связывание S-фазы с M-фазой
Контрольная точка G 2 предотвращает инициацию митоза до завершения S-фазы, тем самым гарантируя, что не полностью реплицированная ДНК не будет распределена по дочерним клеткам.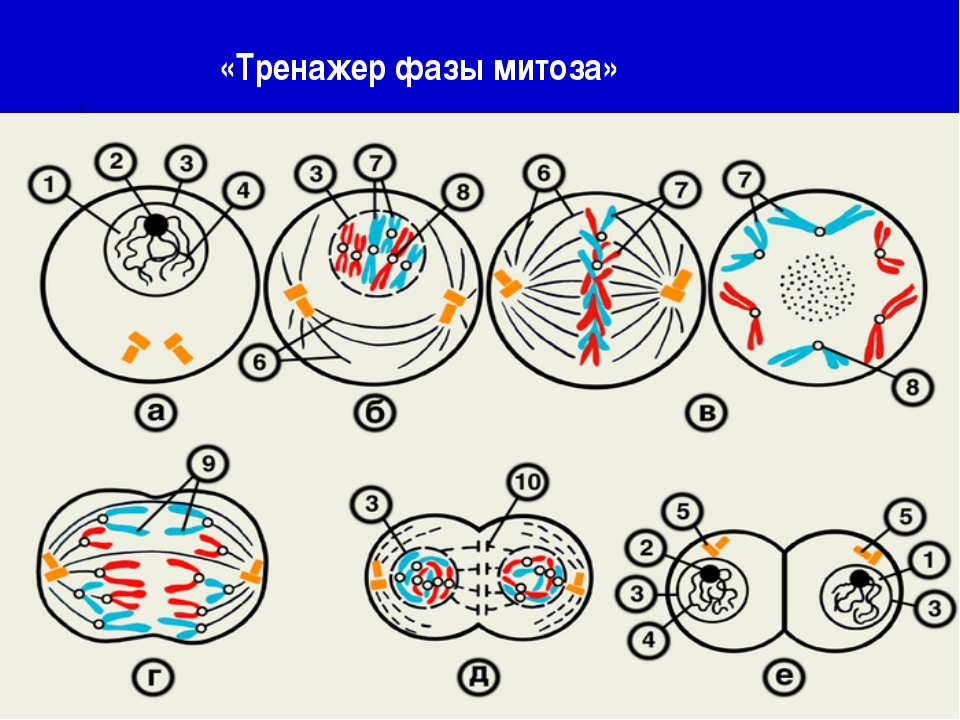 Не менее важно обеспечить, чтобы геном реплицировался только один раз за клеточный цикл.Таким образом, после репликации ДНК должны существовать механизмы контроля, чтобы предотвратить инициацию новой S-фазы до митоза. Эти элементы управления предотвращают повторный вход клеток в S-фазе G 2 и блокируют инициацию другого раунда репликации ДНК до окончания митоза, после чего клетка переходит в фазу G 1 следующего клеточного цикла.
Не менее важно обеспечить, чтобы геном реплицировался только один раз за клеточный цикл.Таким образом, после репликации ДНК должны существовать механизмы контроля, чтобы предотвратить инициацию новой S-фазы до митоза. Эти элементы управления предотвращают повторный вход клеток в S-фазе G 2 и блокируют инициацию другого раунда репликации ДНК до окончания митоза, после чего клетка переходит в фазу G 1 следующего клеточного цикла.
Первоначальное понимание этой зависимости S-фазы от M-фазы пришло из экспериментов по слиянию клеток Поту Рао и Роберта Джонсона в 1970 году ().Эти исследователи изолировали клетки в разных фазах цикла, а затем сливали эти клетки друг с другом, чтобы сформировать клеточные гибриды. При слиянии клеток G 1 с клетками S фазы ядро G 1 немедленно начинало синтезировать ДНК. Таким образом, цитоплазма клеток S фазы содержала факторы, инициирующие синтез ДНК в ядре G 1 . Однако слияние клеток G 2 с клетками S фазы дало совершенно другой результат: ядро G 2 не могло инициировать синтез ДНК даже в присутствии цитоплазмы S фазы.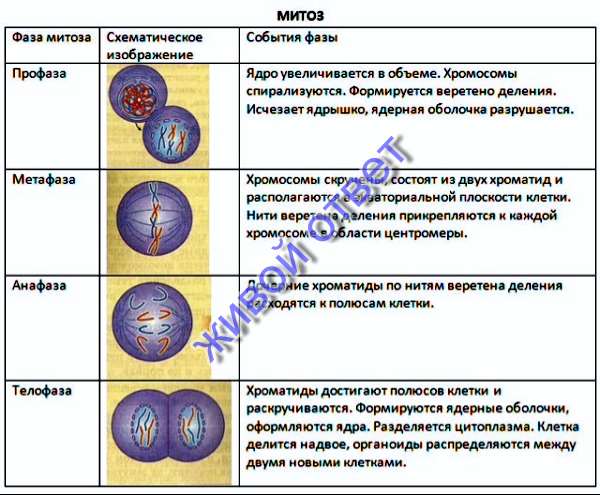 Таким образом, оказалось, что синтез ДНК в ядре G 2 предотвращается механизмом, который блокирует повторную репликацию генома до тех пор, пока не произойдет митоз.
Таким образом, оказалось, что синтез ДНК в ядре G 2 предотвращается механизмом, который блокирует повторную репликацию генома до тех пор, пока не произойдет митоз.
Рис. 14.10
Эксперименты по слиянию клеток, демонстрирующие зависимость S-фазы от M-фазы. Клетки в S-фазе сливали либо с клетками G 1 , либо с клетками G 2 . Когда клетки G 1 были слиты с клетками S фазы, ядро G 1 немедленно начало реплицировать ДНК.Напротив, (подробнее…)
Молекулярный механизм, который ограничивает репликацию ДНК до одного раза за клеточный цикл, включает действие семейства белков (называемых белками MCM), которые связываются с точками начала репликации вместе с комплексом репликации начала (ORC). белки (см. ). Белки MCM действуют как «факторы лицензирования», которые позволяют инициировать репликацию (4). Их связывание с ДНК регулируется во время клеточного цикла, так что белки MCM способны связываться с точками начала репликации только во время G 1 , что позволяет инициировать репликацию ДНК, когда клетка входит в S-фазу.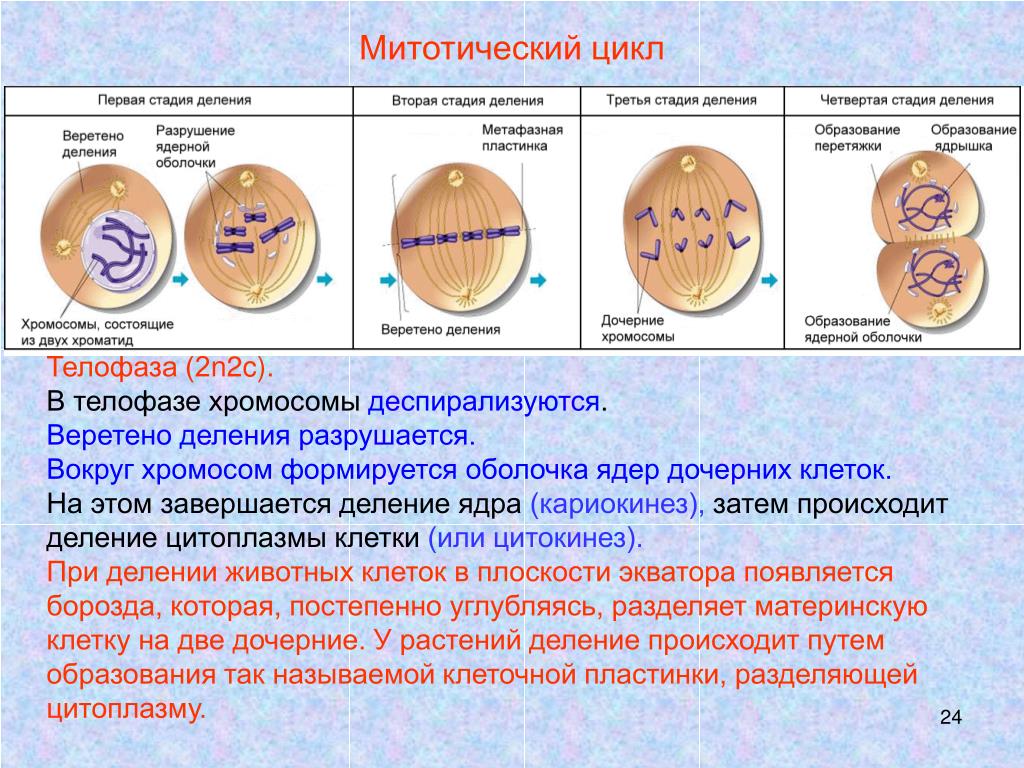 Однако, как только происходит инициация, белки MCM смещаются из источника, поэтому репликация не может снова инициироваться до тех пор, пока клетка не пройдет митоз и не войдет в фазу G 1 следующего клеточного цикла.
Однако, как только происходит инициация, белки MCM смещаются из источника, поэтому репликация не может снова инициироваться до тех пор, пока клетка не пройдет митоз и не войдет в фазу G 1 следующего клеточного цикла.
Рисунок 14.11
Ограничение репликации ДНК. Репликация ДНК ограничена одним разом за клеточный цикл белками MCM, которые связываются с точками начала репликации вместе с белками ORC (комплекс репликации происхождения) и необходимы для инициации репликации ДНК.MCM (подробнее…)
Фаза M клеточного цикла – обзор
3 волны G2–M
Фаза G2–M у почкующихся дрожжей представляет собой основной переход клеточного цикла в этом организме и, соответственно, большую группу клеток. гены, регулируемые циклом, иногда известные как кластер CLB2, экспрессируются исключительно в это время. Первым геном в этом классе был SWI5 , где Mcm1p был идентифицирован как фактор транскрипции, контролирующий его экспрессию, а остальная часть группы (Althoefer et al.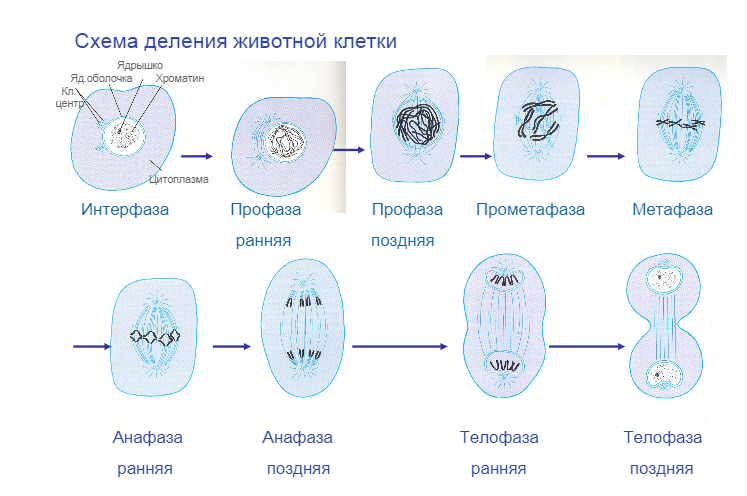 , 1995; Лидалл и др. , 1991; Махер и др. , 1995). Mcm1p является членом класса так называемых факторов транскрипции MADS box , которые обнаруживаются во всех эукариотических организмах, от дрожжей до человека, где он известен как фактор ответа сыворотки (Shore and Sharrocks, 1995; Treisman, 1994; Wynne). и Трейсман, 1992). Mcm1p был хорошо изучен у почкующихся дрожжей, и было показано, что он играет много ролей в контроле экспрессии ряда генов, включая те, которые отвечают за переключение типа спаривания (Gelli, 2002).Способность Mcm1p контролировать различные группы экспрессии генов возникает благодаря тому, что он имеет разных партнеров по связыванию, которые придают специфичность, и является способом действия белков-боксов MADS во всех организмах (Acton et al. , 1997; Messenguy and Dubois, 2003; Tuch и др. , 2008a).
, 1995; Лидалл и др. , 1991; Махер и др. , 1995). Mcm1p является членом класса так называемых факторов транскрипции MADS box , которые обнаруживаются во всех эукариотических организмах, от дрожжей до человека, где он известен как фактор ответа сыворотки (Shore and Sharrocks, 1995; Treisman, 1994; Wynne). и Трейсман, 1992). Mcm1p был хорошо изучен у почкующихся дрожжей, и было показано, что он играет много ролей в контроле экспрессии ряда генов, включая те, которые отвечают за переключение типа спаривания (Gelli, 2002).Способность Mcm1p контролировать различные группы экспрессии генов возникает благодаря тому, что он имеет разных партнеров по связыванию, которые придают специфичность, и является способом действия белков-боксов MADS во всех организмах (Acton et al. , 1997; Messenguy and Dubois, 2003; Tuch и др. , 2008a).
Ассоциированный партнер Mcm1p по связыванию, который контролирует транскрипцию G2-M, был впервые охарактеризован биохимически и назван Swi Five Factor (SFF) с двумя факторами транскрипции, подобными вилке, Fkh2p и Fkh3p, и другим белком Ndd1p, которые впоследствии были идентифицированы как важные компоненты. (Jorgensen and Tyers, 2000, 2004; Knapp et al., 1996 г.; Коранда и др. , 2000; Рис и др. , 2000; Чжу и др. , 2000). Два транскрипционных фактора forkhead необходимы для обеспечения правильной экспрессии генов в интервале G2-M (Борос и др. , 2003; Дариева и др. , 2003; Холленхорст и др. , 2000, 2001; Кумар ). и др. , 2000; Лим и др. , 2003; Рейнольдс и др. , 2003). Оба связываются с мотивами ДНК в виде вилки, присутствующими в промоторах генов в кластере CLB2.Эти два белка частично избыточны, хотя, по-видимому, они играют отдельные и разные роли. Fkh3p связывается с Mcm1p на промоторах генов, тогда как Fkh2p связывается в отсутствие Mcm1p, при этом изменения в конформации ДНК считаются важными для рекрутирования SFF на промоторную ДНК.
(Jorgensen and Tyers, 2000, 2004; Knapp et al., 1996 г.; Коранда и др. , 2000; Рис и др. , 2000; Чжу и др. , 2000). Два транскрипционных фактора forkhead необходимы для обеспечения правильной экспрессии генов в интервале G2-M (Борос и др. , 2003; Дариева и др. , 2003; Холленхорст и др. , 2000, 2001; Кумар ). и др. , 2000; Лим и др. , 2003; Рейнольдс и др. , 2003). Оба связываются с мотивами ДНК в виде вилки, присутствующими в промоторах генов в кластере CLB2.Эти два белка частично избыточны, хотя, по-видимому, они играют отдельные и разные роли. Fkh3p связывается с Mcm1p на промоторах генов, тогда как Fkh2p связывается в отсутствие Mcm1p, при этом изменения в конформации ДНК считаются важными для рекрутирования SFF на промоторную ДНК.
Активация транскрипции не передается через изменения в связывании Mcm1p или Fkh3p с промоторной ДНК, поскольку оба остаются в контакте на протяжении всего клеточного цикла.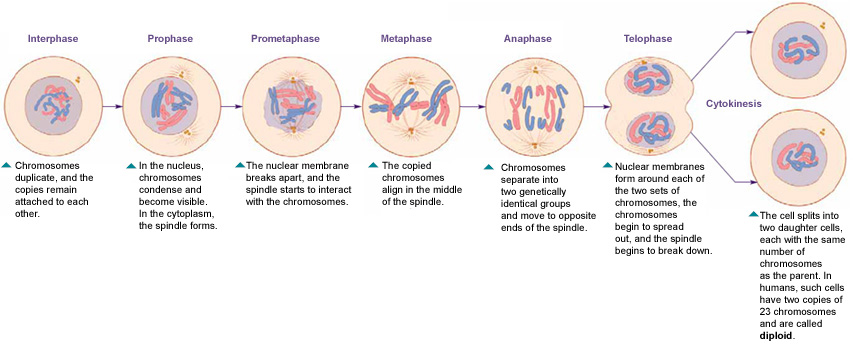 Вместо этого важную роль в этом процессе играет другой активатор транскрипции, Ndd1p (Koranda et al., 2000; Лой и др. , 1999). Ndd1p необходим для транскрипции G2-M, при этом он связывается с промоторами кластерных генов CLB2, зависящими от Fkh3p и Mcm1p. Уровни белка Ndd1p достигают пика в фазах S и G2 как за счет регуляции его транскрипции, так и за счет стабильности белка. Важно отметить, что связывание Ndd1p с промоторами контролируется регулируемым фосфорилированием Fkh3p с помощью комплекса Clb5p-Cdc28p CDK (Pic-Taylor et al. , 2004). Как только Ndd1p связывается с Fkh3p, это взаимодействие стабилизируется другим комплексом CDK, Clb2p-Cdc28p, фосфорилирующим Ndd1p по треонину 319 и приводящим к его специфическому связыванию с доменом FHA Fkh3p.Комплекс Ndd1p-Fkh3p-Mbx1p приводит к транскрипции CLB1 и CLB2 , создавая таким образом петлю положительной обратной связи. Ndd1p также непосредственно фосфорилируется по серину 85 другой протеинкиназой клеточного цикла, polo-подобной киназой, кодируемой геном CDC5 , которая необходима для нормальной временной экспрессии кластера CLB2 (Darieva et al.
Вместо этого важную роль в этом процессе играет другой активатор транскрипции, Ndd1p (Koranda et al., 2000; Лой и др. , 1999). Ndd1p необходим для транскрипции G2-M, при этом он связывается с промоторами кластерных генов CLB2, зависящими от Fkh3p и Mcm1p. Уровни белка Ndd1p достигают пика в фазах S и G2 как за счет регуляции его транскрипции, так и за счет стабильности белка. Важно отметить, что связывание Ndd1p с промоторами контролируется регулируемым фосфорилированием Fkh3p с помощью комплекса Clb5p-Cdc28p CDK (Pic-Taylor et al. , 2004). Как только Ndd1p связывается с Fkh3p, это взаимодействие стабилизируется другим комплексом CDK, Clb2p-Cdc28p, фосфорилирующим Ndd1p по треонину 319 и приводящим к его специфическому связыванию с доменом FHA Fkh3p.Комплекс Ndd1p-Fkh3p-Mbx1p приводит к транскрипции CLB1 и CLB2 , создавая таким образом петлю положительной обратной связи. Ndd1p также непосредственно фосфорилируется по серину 85 другой протеинкиназой клеточного цикла, polo-подобной киназой, кодируемой геном CDC5 , которая необходима для нормальной временной экспрессии кластера CLB2 (Darieva et al.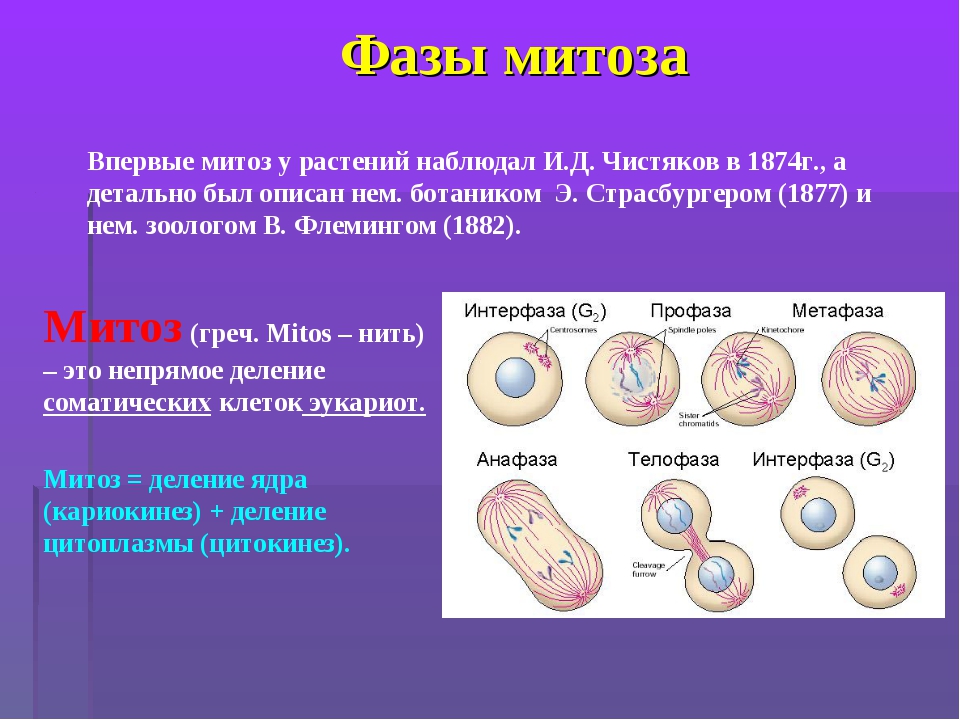 , 2006) . Часть этого механизма происходит за счет связывания Cdc5p специфически с промоторной ДНК в то время, когда она активирует Ndd1p.Поскольку CDC5 является частью кластера CLB2, этот процесс предлагает еще один механизм положительной обратной связи для контроля его экспрессии.
, 2006) . Часть этого механизма происходит за счет связывания Cdc5p специфически с промоторной ДНК в то время, когда она активирует Ndd1p.Поскольку CDC5 является частью кластера CLB2, этот процесс предлагает еще один механизм положительной обратной связи для контроля его экспрессии.
По-видимому, также действует отдельный уровень контроля, включающий стабильность белка, по крайней мере, для одного гена в кластере CLB2, CDC20 . Это происходит за счет того, что Cks1p рекрутирует протеасому на свой промотор, что приводит к экспрессии гена в G2-M (Morris et al. , 2003; Tempé et al. , 2007; Yu et al. , 2005).
Клеточный цикл, его фазы и контрольные точки – обзор
Клеточный цикл и его фазы являются одним из центральных понятий клеточной биологии. Он включает в себя серию четко определенных событий, через которые проходит клетка, чтобы скопировать свое генетическое содержимое и разделить на две идентичные дочерние клетки.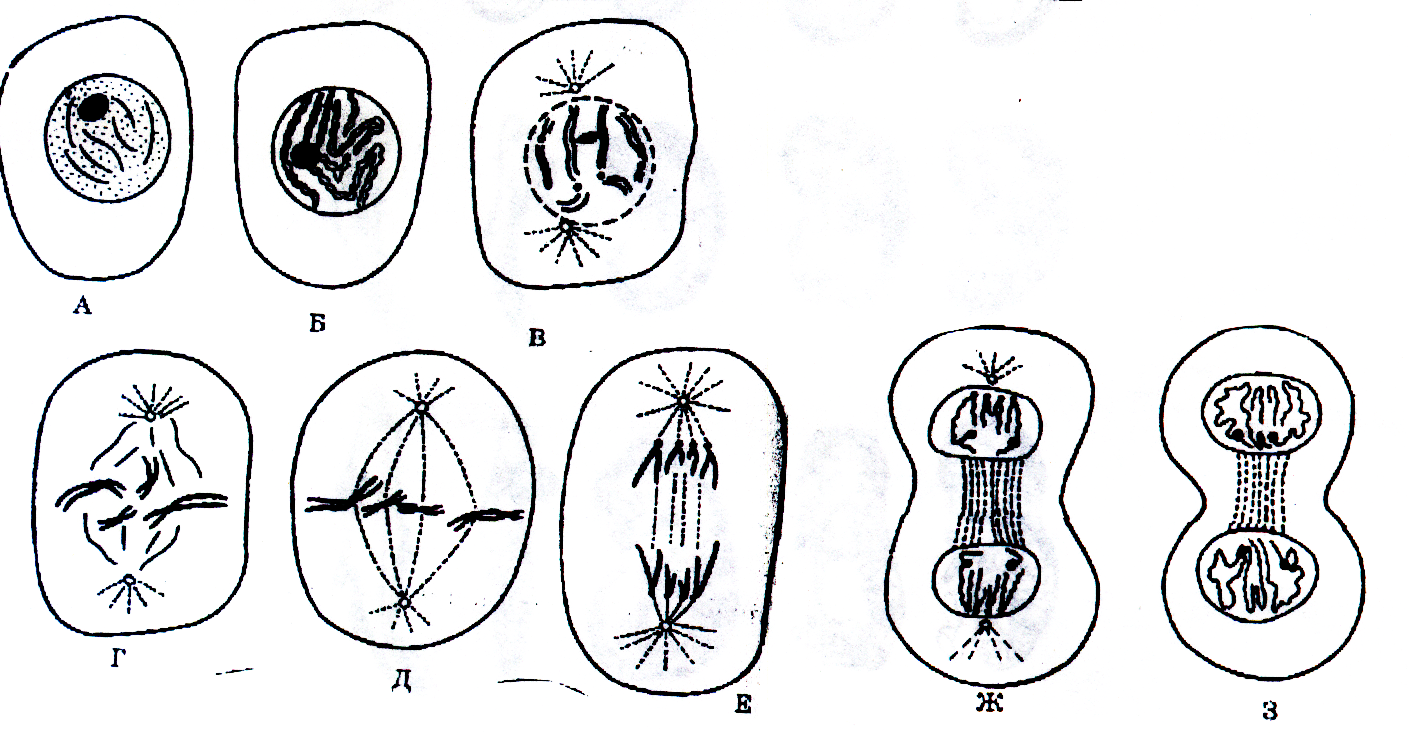 Это деление клеток является основной особенностью жизни. Все эукариотические клетки, то есть клетки с ядрами, проходят клеточный цикл, чтобы делиться. Несмотря на то, что в некоторых аспектах клетки животных, растительных клеток и грибов различаются, общая процедура для них одинакова.
Это деление клеток является основной особенностью жизни. Все эукариотические клетки, то есть клетки с ядрами, проходят клеточный цикл, чтобы делиться. Несмотря на то, что в некоторых аспектах клетки животных, растительных клеток и грибов различаются, общая процедура для них одинакова.
Основные фазы клеточного цикла
Клеточный цикл обычно делится на четыре основные фазы. Первые три фазы часто называют интерфазой, и они подготавливают клетку к четвертой фазе — митозу.
Схематическое изображение клеточного цикла с указанием основных фаз и контрольных точек.G
1 фаза – рост клетокКлетка увеличивается в размерах и увеличивает производство мРНК, белка и клеточных органелл, чтобы подготовиться к более поздним стадиям.
S фаза – синтез ДНК
Производство мРНК и белка снижается, и клетка фокусируется на репликации всей своей ДНК, создавая по одной копии каждой хромосомы. Две идентичные копии, называемые сестринскими хроматидами, прикрепляются друг к другу в точке, называемой центромерой.
Классическое изображение хромосомы. ДНК была реплицирована, поэтому хромосома состоит из двух идентичных сестринских хроматид, соединенных центромерой.
G
2 фаза – подготовка к делению клетокКлетка продолжает расти и синтезировать белки и органеллы, необходимые для последней фазы.
М-фаза – образование дочерних клеток путем митоза
Ядерная мембрана разрывается, освобождая хромосомы. Сестринские хроматиды разделяются и образуют два идентичных ядра, оттянутых к противоположным сторонам клетки. Затем он разделяет клеточную мембрану и цитоплазму на две равные части, разделяя между ними ДНК и клеточные органеллы. Были созданы две генетически идентичные дочерние клетки.
Клетка во время М-фазы. (1) Ядерная мембрана разрушается, освобождая хромосомы.(2) Хромосомы выстраиваются в линию и прикрепляются к микротрубочкам веретена. (3) Хромосомы разрываются на центромере, и микротрубочки тянут сестринские хроматиды к противоположным сторонам клетки.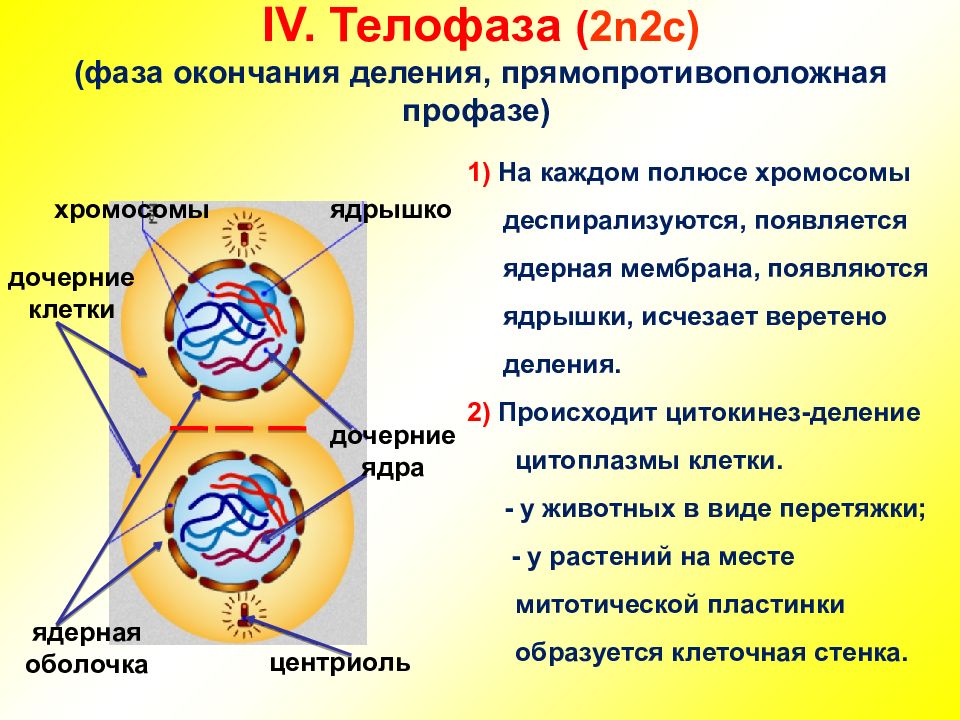 (4) Образуются два новых полных ядра, и клетка может делиться на две дочерние клетки.
(4) Образуются два новых полных ядра, и клетка может делиться на две дочерние клетки.
Выход из клеточного цикла – G
0 фазаПосле того, как клетка завершила фазу M, обе дочерние клетки снова переходят в фазу G 1 . Отсюда они могут либо продолжить клеточный цикл и снова делиться, либо выйти из него и перестать делиться.Клетка, выходящая из клеточного цикла, переходит в фазу G 0 , где она все еще полностью функциональна, но больше не делится. Некоторые клетки никогда больше не покидают фазу G 0 , как и большинство нейронных клеток, тогда как другие могут повторно входить в фазу G 1 и при необходимости продолжать клеточный цикл. Как и большинство эпителиальных клеток, некоторые клетки никогда не вступают в фазу G 0 , но продолжают делиться на протяжении всей своей жизни.
Контрольные точки клеточного цикла
Чтобы убедиться, что каждая фаза клеточного цикла выполняется правильно, за основными этапами следует контрольная точка. Это механизм контроля, в котором должны быть выполнены определенные условия, прежде чем клетка сможет перейти к следующему шагу. Сбой контрольной точки вызывает остановку клеточного цикла, не позволяя ячейке продолжать работу до тех пор, пока она не решит проблемы.
Это механизм контроля, в котором должны быть выполнены определенные условия, прежде чем клетка сможет перейти к следующему шагу. Сбой контрольной точки вызывает остановку клеточного цикла, не позволяя ячейке продолжать работу до тех пор, пока она не решит проблемы.
Клеточный цикл включает несколько контрольных точек, основными из которых являются G 1 , G 2 и M контрольная точка.
G
1 Контрольный пункт – проверка размера и питательных веществ Эта контрольная точка, также называемая контрольной точкой ограничения, имеет место между G 1 и фазой S.Клетка проверяет, достаточно ли она велика для деления, не повреждена ли ее ДНК и есть ли достаточный доступ к питательным веществам и стимулирующим факторам роста. Если он проходит контрольную точку G 1 , он необратимо завершает весь клеточный цикл и делится. Если нет, то пытается исправить сбой или переходит в фазу G 0 и ждет более благоприятных условий.
G
2 контрольно-пропускной пункт – контроль качества ДНКПосле второй фазы роста клетка проверяет, полностью ли и правильно реплицировалась ДНК во время S-фазы.В случае успеха он переходит к фазе М, а в случае неудачи пытается исправить ошибки. Если клетка не может восстановить ДНК, она подвергается апоптозу. Это предотвращает передачу поврежденной ДНК дочерним клеткам.
Контрольно-пропускной пункт М – проверка установки ядерной дивизии
Эта контрольная точка происходит во время M-фазы. Его также называют контрольной точкой веретена, потому что клетка проверяет, правильно ли все сестринские хроматиды прикреплены к микротрубочкам веретена, которые их разделяют.Если нет, клетка приостанавливает митоз до тех пор, пока все сестринские хроматиды не будут прикреплены должным образом.
Пролиферация клеток – деление клеток
и рост клеток Клеточная пролиферация — это термин, тесно связанный с клеточным циклом, означающий процесс, в ходе которого увеличивает количество клеток за счет клеточного деления и роста клеток . В то время как деление клеток необходимо для увеличения числа клеток, рост клеток также важен, поскольку родительская клетка теряет половину своего материала, когда образует дочернюю клетку.
В то время как деление клеток необходимо для увеличения числа клеток, рост клеток также важен, поскольку родительская клетка теряет половину своего материала, когда образует дочернюю клетку.
Целью пролиферации клеток является обеспечение роста и развития тканей, а затем поддержание здоровой клеточной популяции в зрелом органе. Когда старые и поврежденные клетки умирают и уничтожаются, оставшиеся клетки размножаются строго регулируемым образом, чтобы заменить их. Это гарантирует, что общее количество функциональных ячеек останется прежним. Сбой в этом регулируемом процессе, приводящий к тому, что клетки продолжают бесконтрольно размножаться, является причиной опухолей и рака.
Клеточный цикл
Рост и деление клетки организованы в строго контролируемый и упорядоченный процесс, называемый клеточным циклом.
Фото: mirela377/Shutterstock.com
Клеточный цикл состоит из 4 стадий; Фаза разрыва 1 (G1), фаза синтеза (S), фаза разрыва 2 (G2) и фаза митоза (M).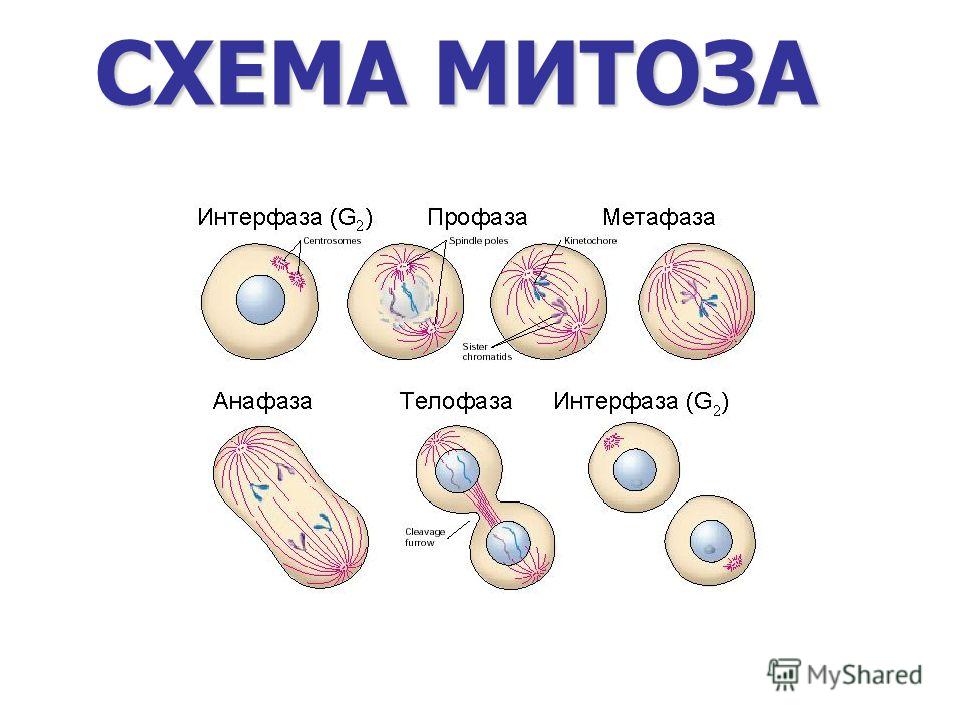 Для большинства клеток человека один клеточный цикл занимает примерно 24 часа. Однако в тканях, где существует постоянная потребность в обновлении и замене клеток, таких как слизистая оболочка кишечника, этот процесс намного короче и занимает всего 9 часов.
Для большинства клеток человека один клеточный цикл занимает примерно 24 часа. Однако в тканях, где существует постоянная потребность в обновлении и замене клеток, таких как слизистая оболочка кишечника, этот процесс намного короче и занимает всего 9 часов.
Интерфаза
Фазы G1, S и G2 в совокупности называются интерфазой, включающей рост клетки и репликацию ее ДНК.Первоначально в фазе G1 клетка растет физически и увеличивает объем как белка, так и органелл. В S-фазе клетка копирует свою ДНК с образованием двух сестринских хроматид и реплицирует свои нуклеосомы. Наконец, фаза G2 включает дальнейший рост клеток и организацию клеточного содержимого.
Митоз
Во время М-фазы клетка делится на две дочерние клетки. Первоначально ДНК конденсируется с образованием хромосом, которые разделяются митотическим веретеном. Эта М-фаза далее делится на 4 стадии; профаза, метафаза, анафаза и телофаза.
Профаза : ДНК конденсируется с образованием хромосом, и между двумя нуклеосомами начинают формироваться митотические веретена.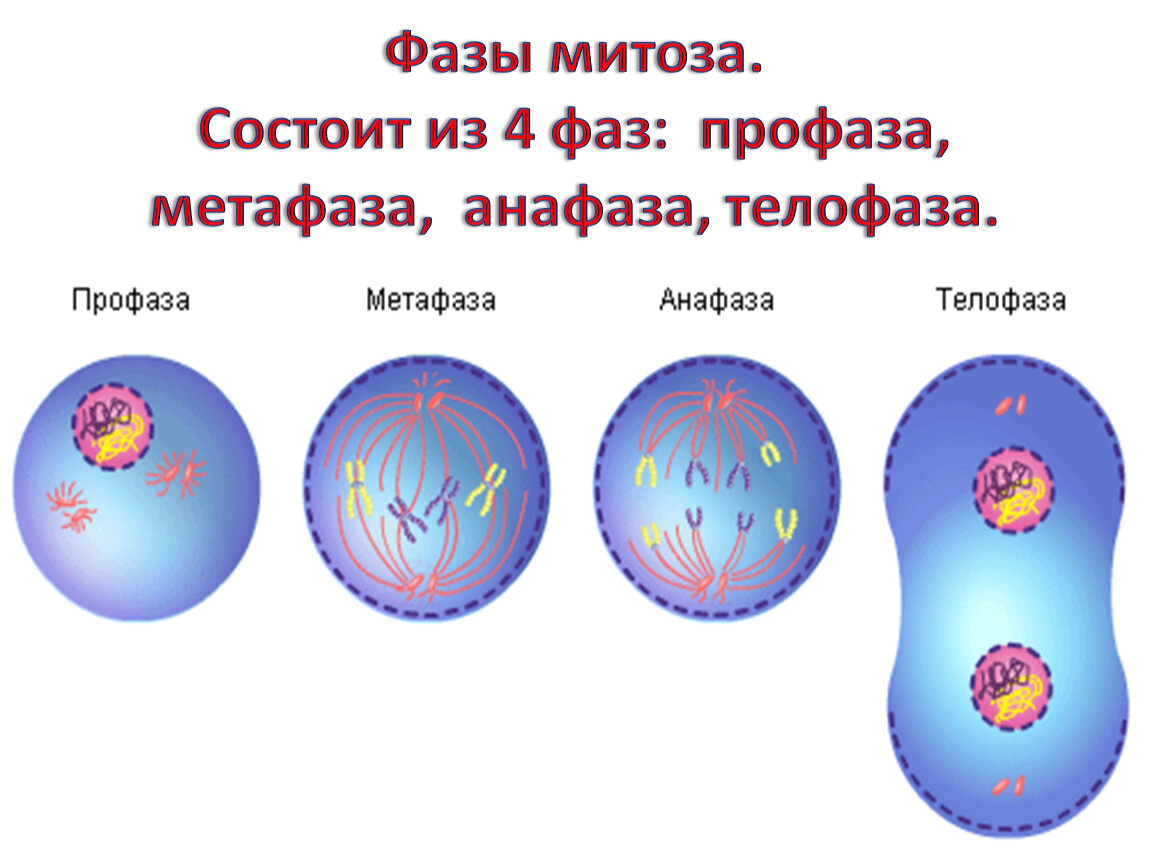 Затем эти веретена начинают связываться с кинетохорами на хромосомах и организовывать их в центре клетки.
Затем эти веретена начинают связываться с кинетохорами на хромосомах и организовывать их в центре клетки.
Метафаза : Хромосомы, связанные своими кинетохорами, втягиваются в центр клетки, образуя линию, называемую метафазной пластинкой. Затем клетка гарантирует, что все хромосомы связаны на двух отдельных кинетохорах, по одной на каждой сестринской хроматиде, в фазе, называемой контрольной точкой веретена.Это подтверждает, что каждая новая клетка будет содержать четное количество материала ДНК после деления клетки.
Анафаза: После успешного прохождения этой контрольной точки ячейка войдет в анафазу. Когезины, скрепляющие хромосомы, расщепляются, и связанные микротрубочки начинают укорачиваться. Этот процесс раздвигает сестринские хроматиды к противоположным полюсам клетки. Оставшиеся микротрубочки, которые не связаны с хромосомами, затем удлиняются и отдаляют две половины клетки дальше друг от друга.
Телофаза: К этому моменту клетка практически полностью делится.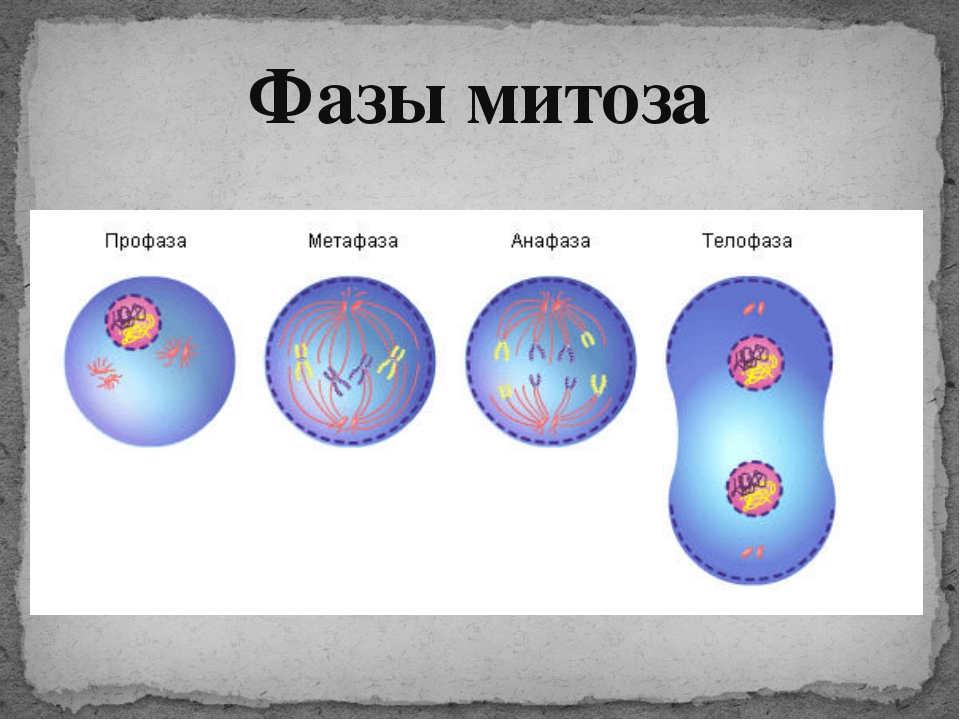 В телофазе происходит деконденсация ДНК и разрушение митотического веретена до того, как разовьются два отдельных ядра.
В телофазе происходит деконденсация ДНК и разрушение митотического веретена до того, как разовьются два отдельных ядра.
Цитокинез
После расщепления ДНК физическая клетка делится в процессе, называемом цитокинезом. Вначале в центре образуется сократительное кольцо, разделяющее и сжимающее клетку пополам. Это образует углубление, называемое бороздой деления, которая в конечном итоге разделяет клетку на две идентичные дочерние клетки.
Контроль клеточного цикла
В целом этот процесс строго контролируется различными белками, которые стимулируют и ингибируют клеточный цикл. Циклин и циклинзависимые киназы (CDK) являются одними из наиболее важных белков, участвующих в стимуляции цикла. Уровни циклина повышаются и понижаются на разных стадиях цикла, что стимулирует функцию CDK, присутствие которых стабильно, но может функционировать только в присутствии циклинов. CDK функционируют для фосфорилирования многих различных белков, которые необходимы для прохождения важных точек клеточного цикла, называемых контрольными точками.
Эти контрольные точки присутствуют в конце G1 и в начале G2, функционируя для обеспечения того, чтобы жизненно важные процессы каждой стадии выполнялись до того, как клетка перейдет в S-фазу или покинет ее. Существует также контрольная точка М-фазы (контрольная точка веретена), которая гарантирует, что хромосомы правильно выровнены, как описано ранее.
В целом эти контрольные точки обеспечивают целостность генома и предотвращают повреждение ДНК. Если клетка не соответствует требованиям контрольной точки, то клеточный цикл останавливается и ДНК можно восстановить, или если ДНК не подлежит восстановлению, то можно стимулировать апоптоз.
Если эти контрольные точки потеряны из-за мутировавших белков, то клеточный цикл больше не контролируется и может привести к неуправляемой репликации, например, мутации в гене TP53 приводят ко многим типам рака.
Этот белок участвует в остановке клеточного цикла и транскрипции белков, участвующих в репарации ДНК или апоптозе.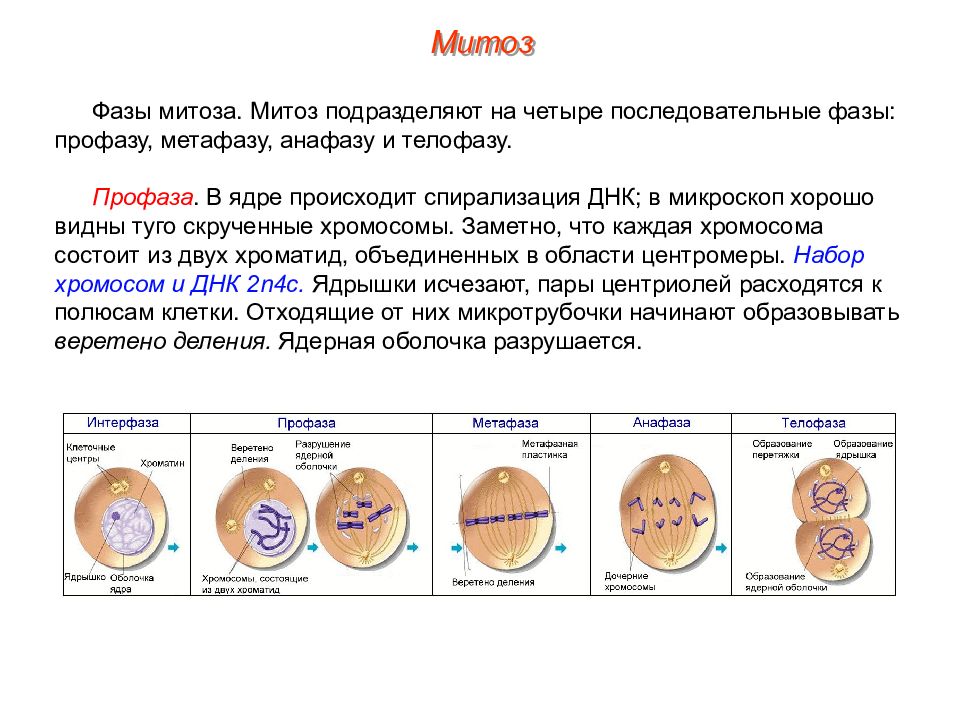 Следовательно, потеря этого гена означает, что клеточный цикл не может быть остановлен, а ДНК не может быть восстановлена, что приводит к опухолевому росту.
Следовательно, потеря этого гена означает, что клеточный цикл не может быть остановлен, а ДНК не может быть восстановлена, что приводит к опухолевому росту.
В целом клеточный цикл является важным процессом для роста и восстановления тканей.Он разделен на 4 отдельных этапа; Фаза G1, фаза S, фаза G2 и фаза M, и контролируется наличием контрольно-пропускных пунктов. Потеря контроля связана с раком, например, с мутациями, приводящими к потере остановки цикла и репарации ДНК, что демонстрирует важность правильной регуляции.
Как делятся клетки — Фазы митоза — Деление клеток и клеточный цикл — Деление клеток Играть
Дополнительное чтение
3.2 Клеточный цикл | Деление клетки
Клеточный цикл представляет собой серию событий, происходящих в клетке, которые приводят к репликации ДНК и клеточному делению.В клеточном цикле выделяют две основные стадии. Первая стадия — интерфаза , во время которой клетка растет и реплицирует свою ДНК. Вторая фаза — это митотическая фаза (М-фаза), во время которой клетка делится и передает одну копию своей ДНК двум идентичным дочерним клеткам. На рис. 3.1 представлен краткий обзор того, что происходит во время каждого из ключевых событий клеточного цикла.
Вторая фаза — это митотическая фаза (М-фаза), во время которой клетка делится и передает одну копию своей ДНК двум идентичным дочерним клеткам. На рис. 3.1 представлен краткий обзор того, что происходит во время каждого из ключевых событий клеточного цикла.
Межфазный (ESG5K)
Интерфаза — самая продолжительная фаза клеточного цикла.Во время этой фазы клетка вырастает до максимального размера, выполняет свои нормальные клеточные функции, реплицирует свою ДНК и готовится к делению клетки. Этот этап разделен на три части: G 1 , G 2 и S фазы .
Некоторым клеткам больше не нужно делиться и выходить из клеточного цикла. Эти клетки могут выйти из клеточного цикла навсегда, например, нейроны, или они могут выйти из клеточного цикла временно. Говорят, что эти ячейки находятся в G 0 . G 0 не является стадией клеточного цикла.
В клетках без ядра (прокариотических клетках, например, бактериях) существует множество копий ДНК, плавающих по всей клетке.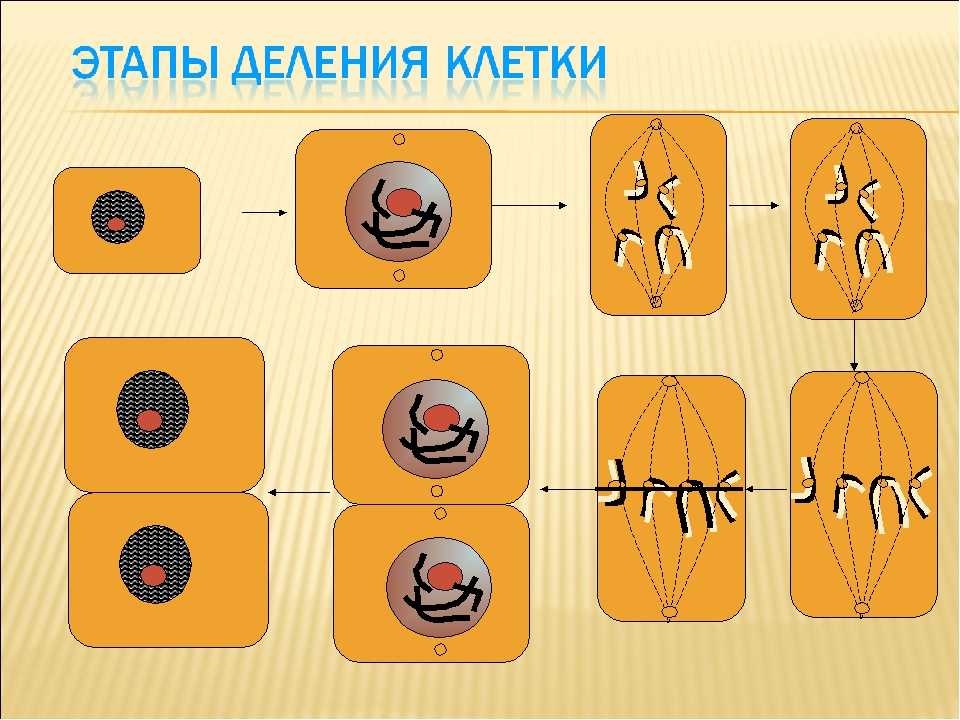 Прокариотический клеточный цикл происходит посредством процесса, называемого бинарным делением. В клетках с ядром (эукариоты) вся ДНК находится внутри ядра, поэтому для репликации требуется более сложный клеточный цикл.
Прокариотический клеточный цикл происходит посредством процесса, называемого бинарным делением. В клетках с ядром (эукариоты) вся ДНК находится внутри ядра, поэтому для репликации требуется более сложный клеточный цикл.
G 1 фаза : Происходит сразу после разделения двух дочерних клеток, когда клетки имеют только одну копию своей ДНК.Клетки на этой стадии синтезируют белки и увеличиваются в размерах. Клетки могут оставаться в этой стадии в течение длительного времени.
S-фаза : Стадия, во время которой происходит репликация ДНК. Клетка делает идентичную копию каждой из своих хромосом . Хромосомы находятся внутри ядра клетки и состоят из длинных нитей ДНК, содержащих генетическую информацию клетки.
G 2 фаза : Происходит после удвоения ДНК в S фазе.Во время этой фазы клетка может продолжать расти и выполнять нормальные клеточные функции. К концу этой фазы клетка начнет реплицировать свои органеллы, готовясь к митозу.
(G 1 , S и G 2 фазы) составляет примерно \(\text{90}\%\) клеточного цикла, а другая \(\text{10}\%\) составляет поглощается митозом.
Митотическая фаза (ESG5M)
Митотическая фаза (М-фаза) состоит из двух тесно связанных процессов: митоза и цитокинеза .Во время митоза хромосомы в клеточном ядре разделяются на два одинаковых набора в двух ядрах. Затем следует цитокинез , при котором цитоплазма, органеллы и клеточная мембрана расщепляются на две клетки, содержащие примерно равные доли этих клеточных компонентов. Теперь мы опишем, что происходит на стадиях М-фазы, которая включает четыре широкие фазы митоза (профаза, метафаза, анафаза, телофаза) и пятую фазу цитокинеза:
- профаза
- метафаза
- анафаза
- телофаза
- цитокинез
1.Профаза
Во время профазы материал хроматина укорачивается и утолщается в отдельные хромосомы, которые видны под световым микроскопом. Каждая хромосома состоит из двух нитей или хроматид , соединенных центромерой (рис. 3.2).
Каждая хромосома состоит из двух нитей или хроматид , соединенных центромерой (рис. 3.2).
Клетки человека имеют 46 хромосом. (23 от матери и 23 от отца).
По мере развития профазы ядерная оболочка и ядрышко разрушаются. В клетках животных центриолей расходятся и расходятся к противоположным полюсам. Центриоли дают начало веретенообразным волокнам, образующимся между полюсами. В растительных клетках нет центриолей для движения к полюсам, поэтому в цитоплазме образуются веретеновидные волокна.
| Схематическая диаграмма : животная клетка | Микрофотография : растительная клетка |
2.Метафаза
Во время метафазы хромосомы выстраиваются на экваторе клетки.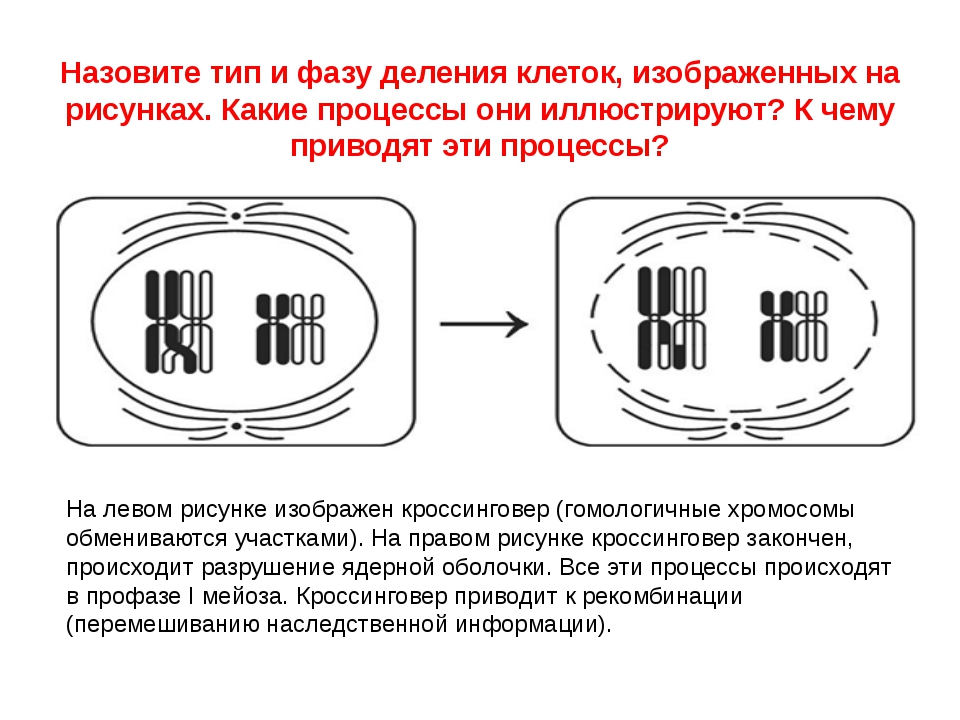 Хромосомы располагаются прямой линией посередине клетки. Каждая хромосома прикреплена к нитям веретена деления своей центромерой.
Хромосомы располагаются прямой линией посередине клетки. Каждая хромосома прикреплена к нитям веретена деления своей центромерой.
Стадии клеточного цикла ( i интерфаза, p рофаза, m этафаза, a нафаза, t элофаза) можно запомнить с помощью мнемоники IPMAT .
3.Анафаза
Во время анафазы хроматиды оттягиваются к противоположным полюсам клетки за счет укорочения волокон веретена деления. Хроматиды теперь называются дочерними хромосомами .
| Схематическая диаграмма : животная клетка | Микрофотография : растительная клетка |
В растительных клетках нет центриолей для движения к полюсам, поэтому в цитоплазме образуются веретеновидные волокна.
4. Телофаза
Во время телофазы ядерная мембрана восстанавливается вокруг дочерних хромосом, собравшихся на каждом из полюсов.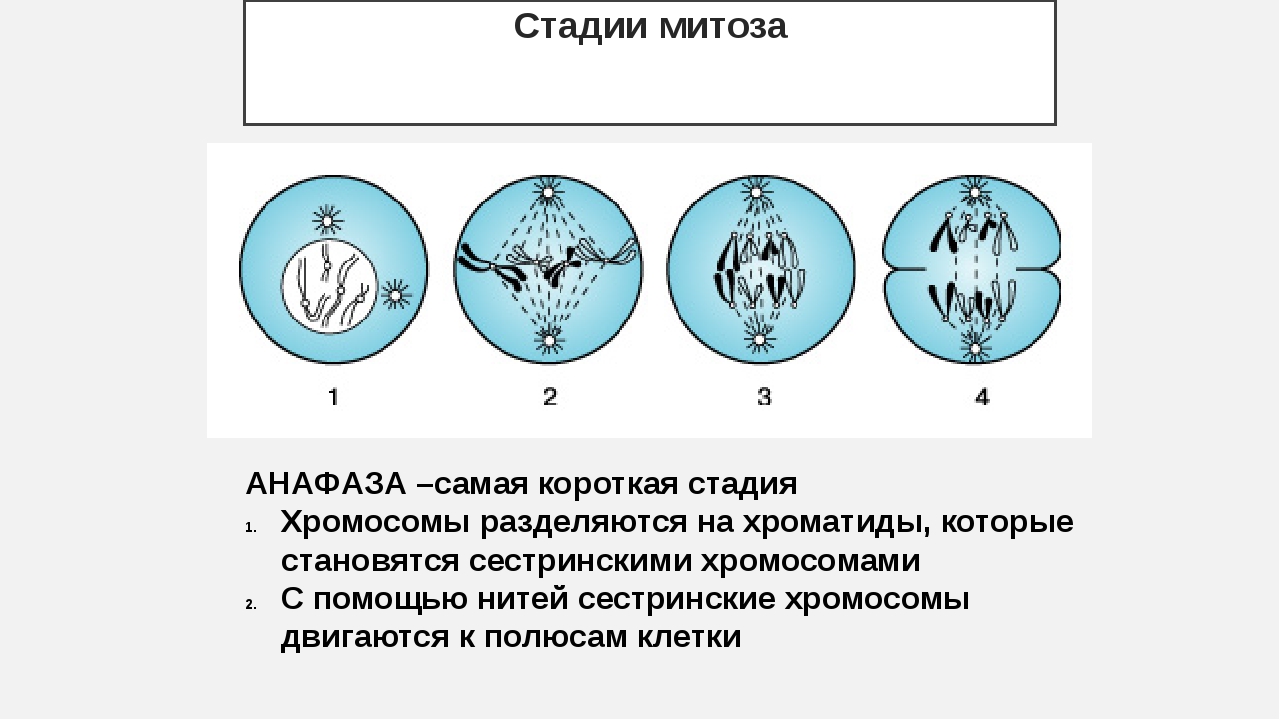 Дочерние хромосомы снова разворачиваются, образуя хроматин. Реформируется ядерная мембрана.
Дочерние хромосомы снова разворачиваются, образуя хроматин. Реформируется ядерная мембрана.
| Схематическая диаграмма : животная клетка | Микрофотография : растительная клетка |
5. Цитокинез
Затем цитоплазма делится в ходе процесса, называемого цитокинезом .Цитокинез — это не стадия митоза, а процесс деления цитоплазмы на две части. В животной клетке сужается клеточная мембрана. Эта инвагинация или складчатость цитоплазмы делит клетку надвое. В растительной клетке поперечная стенка образована клеточной пластинкой , разделяющей цитоплазму надвое.
| Схематическая диаграмма : животная клетка | Микрофотография | |

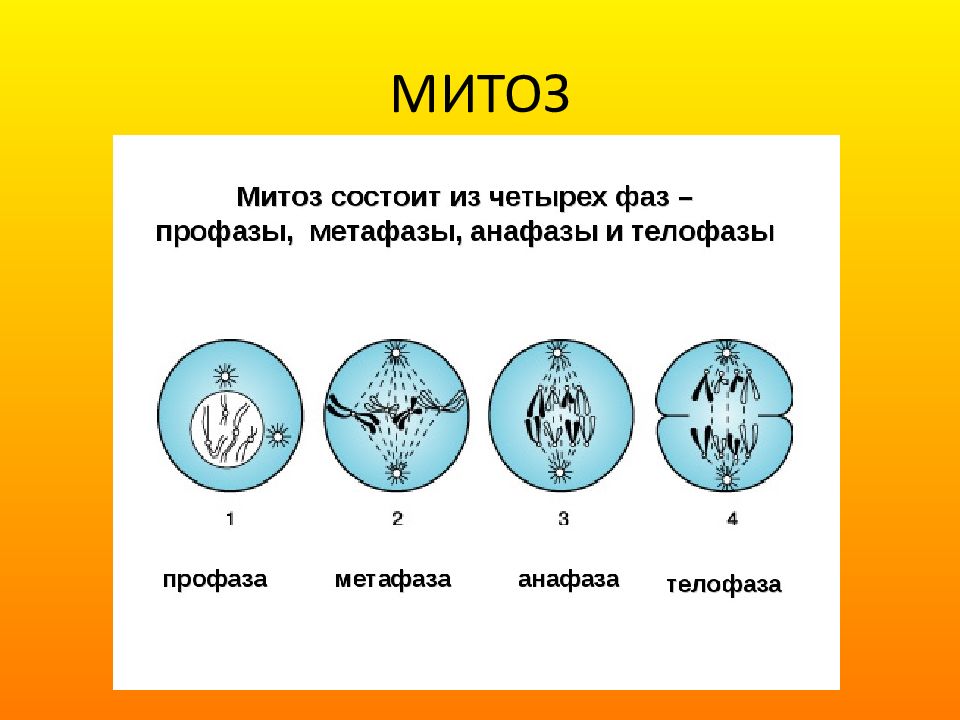 Цитоплазма также обычно делится, в результате чего образуются две дочерние клетки
Цитоплазма также обычно делится, в результате чего образуются две дочерние клетки  Клетки увеличиваются в размерах и образуют органеллы.
Клетки увеличиваются в размерах и образуют органеллы. 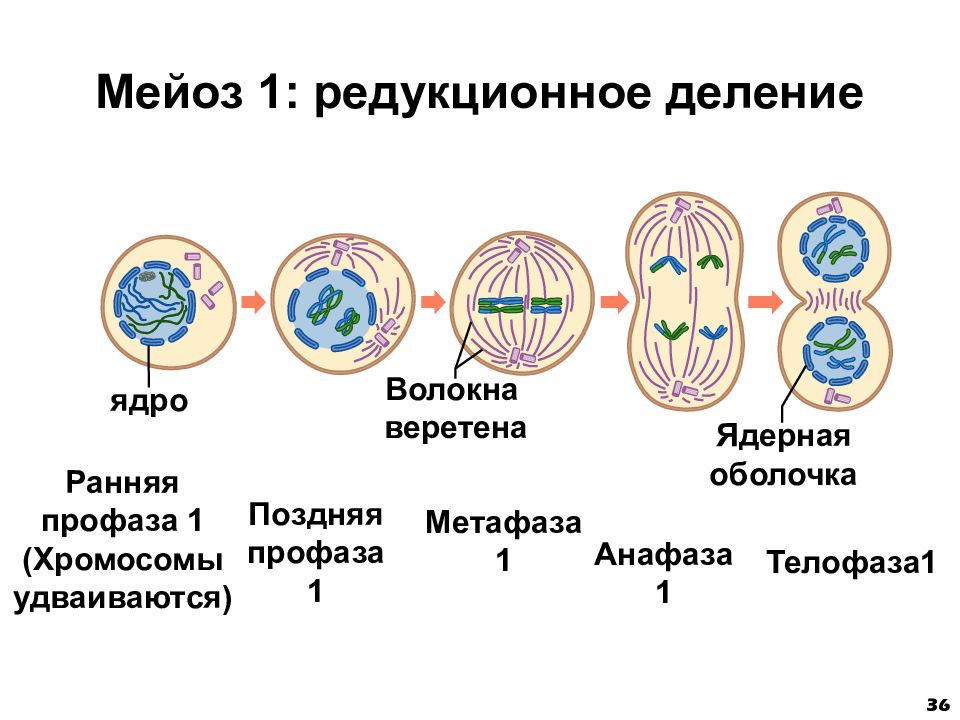 На этом этапе клетка готовится к делению. Здесь клетка имеет двойную ДНК и снова увеличивается в размерах.Некоторые клеточные органеллы дублируются, а цитоскелет демонтируется, чтобы обеспечить ресурсы для митотической фазы. Во время G 2 может наблюдаться дополнительный рост клеток.
На этом этапе клетка готовится к делению. Здесь клетка имеет двойную ДНК и снова увеличивается в размерах.Некоторые клеточные органеллы дублируются, а цитоскелет демонтируется, чтобы обеспечить ресурсы для митотической фазы. Во время G 2 может наблюдаться дополнительный рост клеток. 

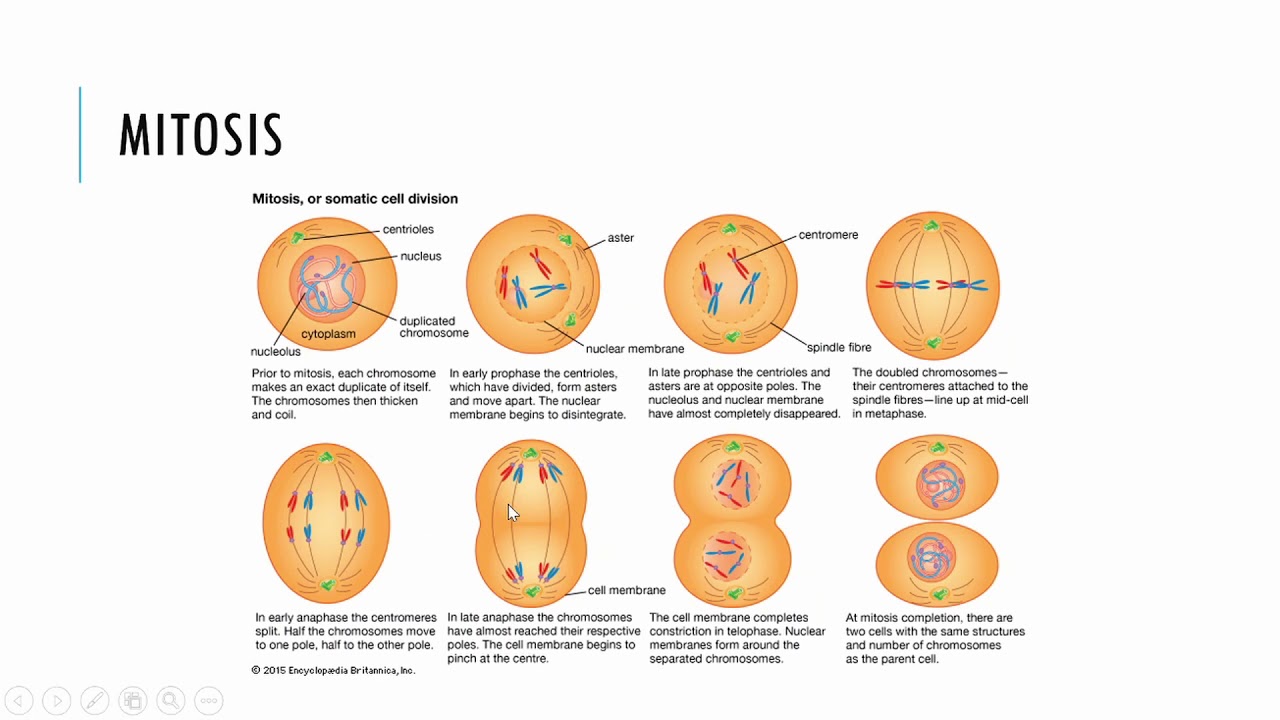 Во время этого цикла двойная спираль ДНК клетки реплицируется с образованием двух идентичных дочерних молекул ДНК. За этим следует митоз, чтобы разделить клетку на две дочерние клетки.
Во время этого цикла двойная спираль ДНК клетки реплицируется с образованием двух идентичных дочерних молекул ДНК. За этим следует митоз, чтобы разделить клетку на две дочерние клетки. 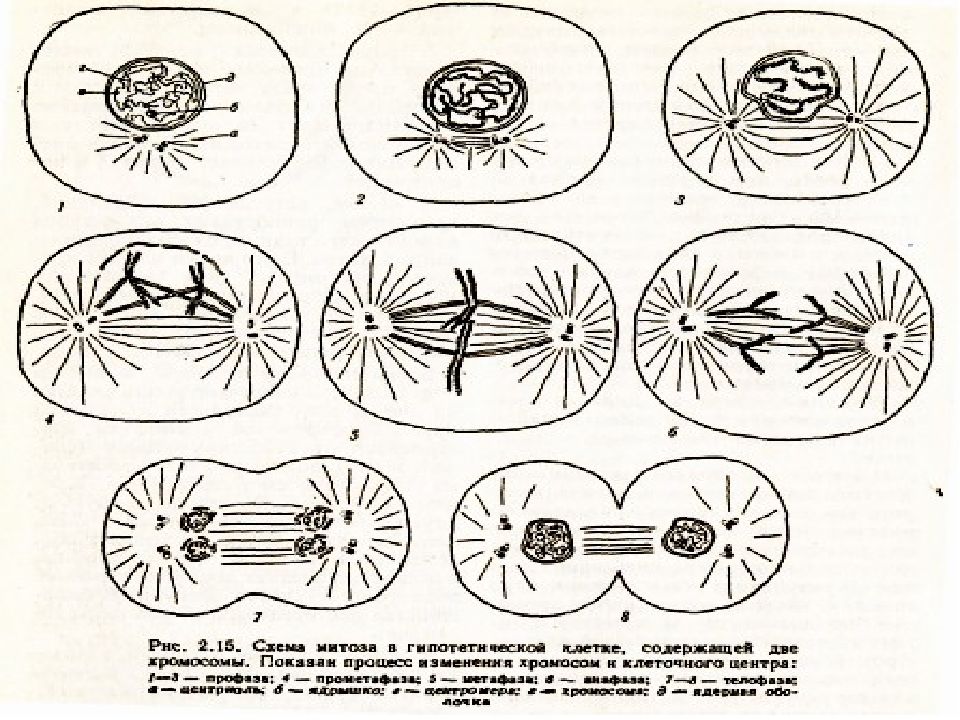



 Зазор 2 фазы (G2)
Зазор 2 фазы (G2)

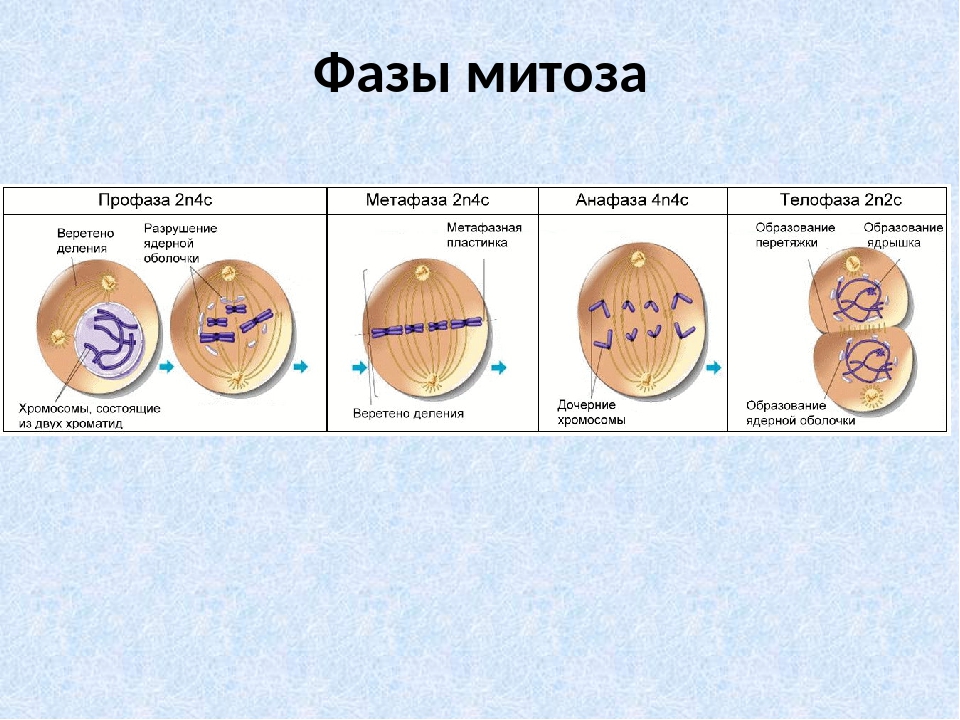
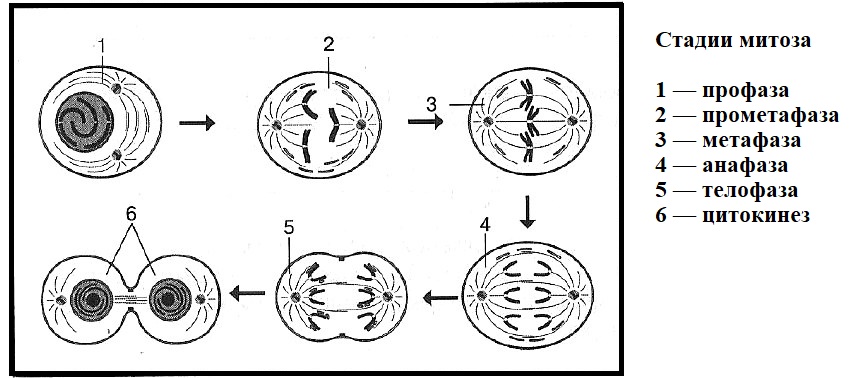 Во время этой фазы вокруг хромосом формируется ядерная оболочка, в результате которой образуются два отдельных дочерних ядра.
Во время этой фазы вокруг хромосом формируется ядерная оболочка, в результате которой образуются два отдельных дочерних ядра.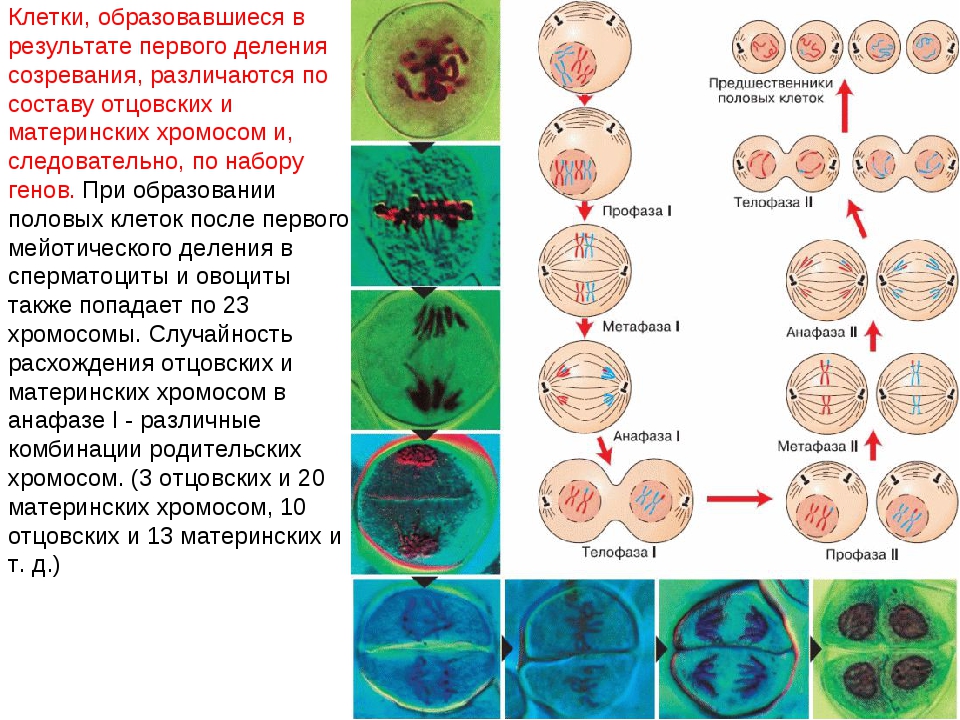

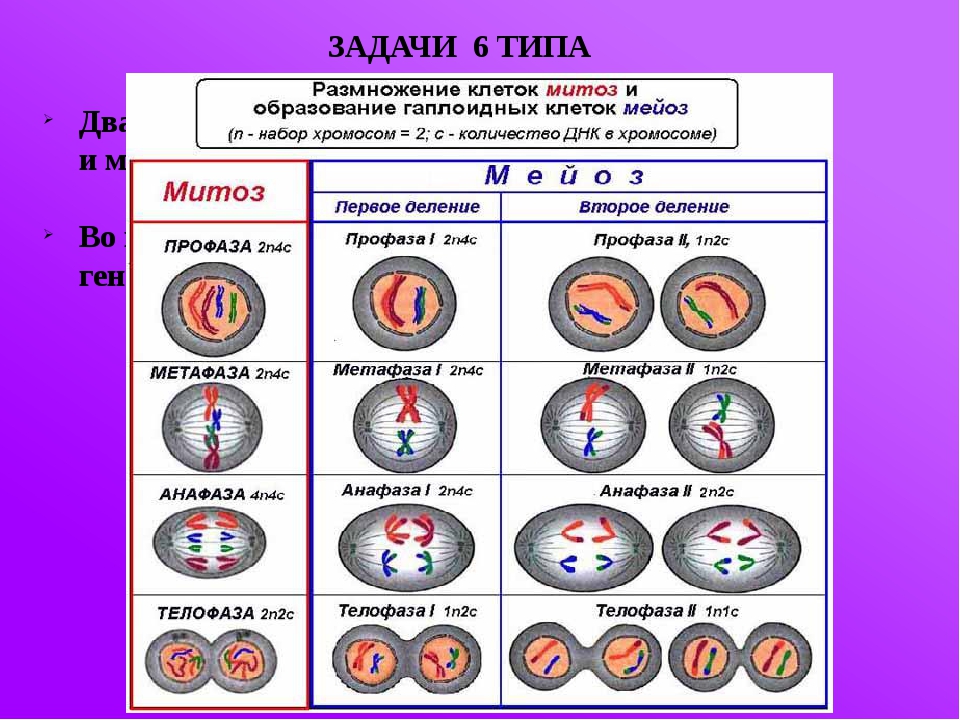 Затем комплекс отвечает за регуляцию клеточного цикла.
Затем комплекс отвечает за регуляцию клеточного цикла. 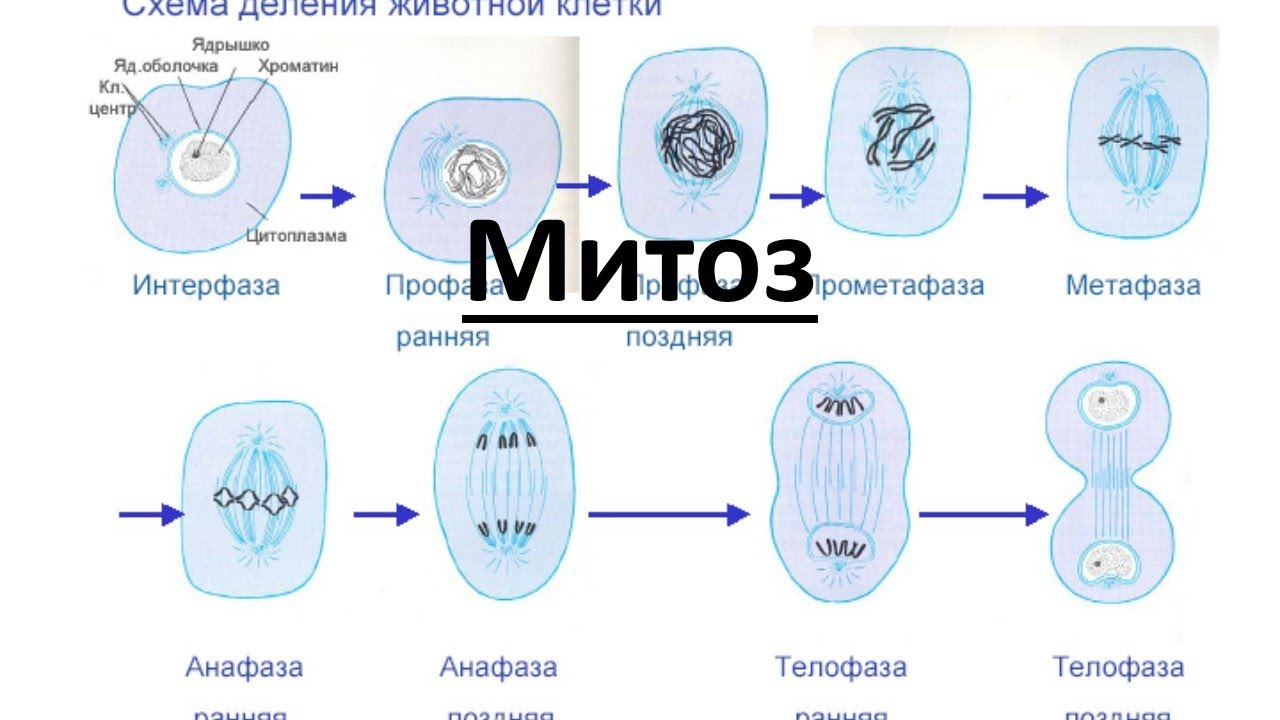

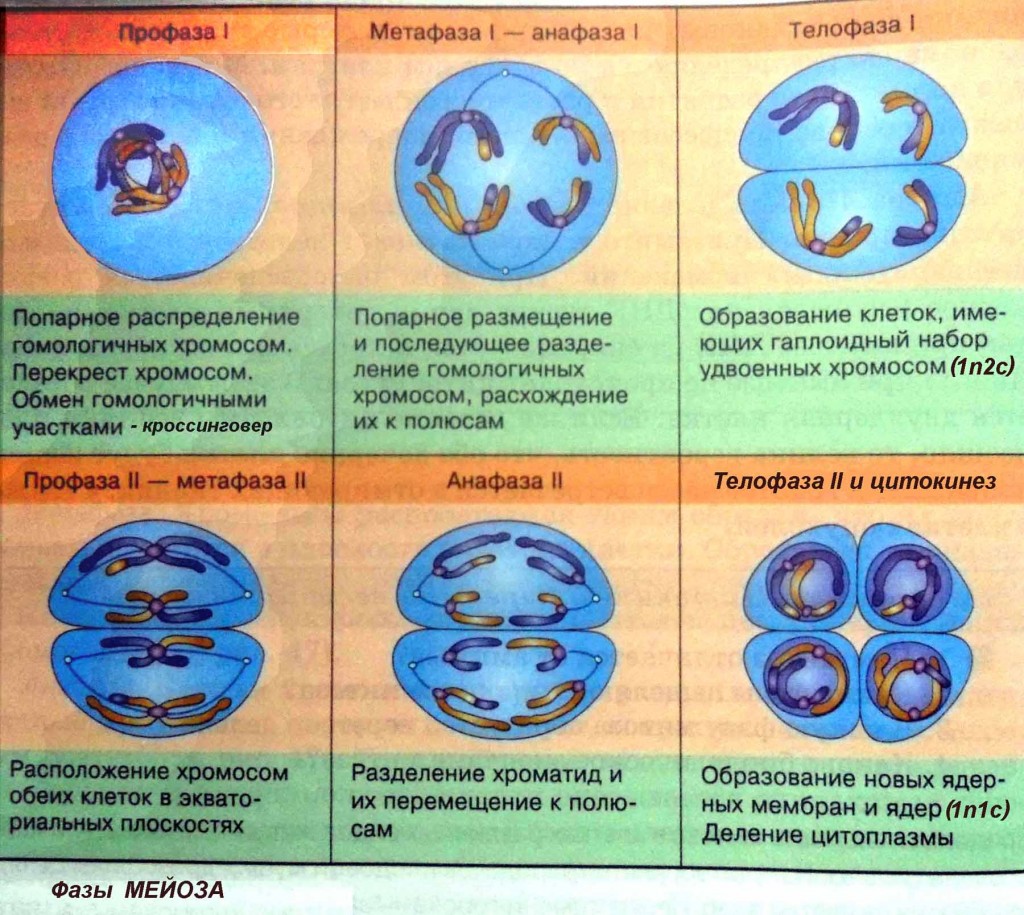
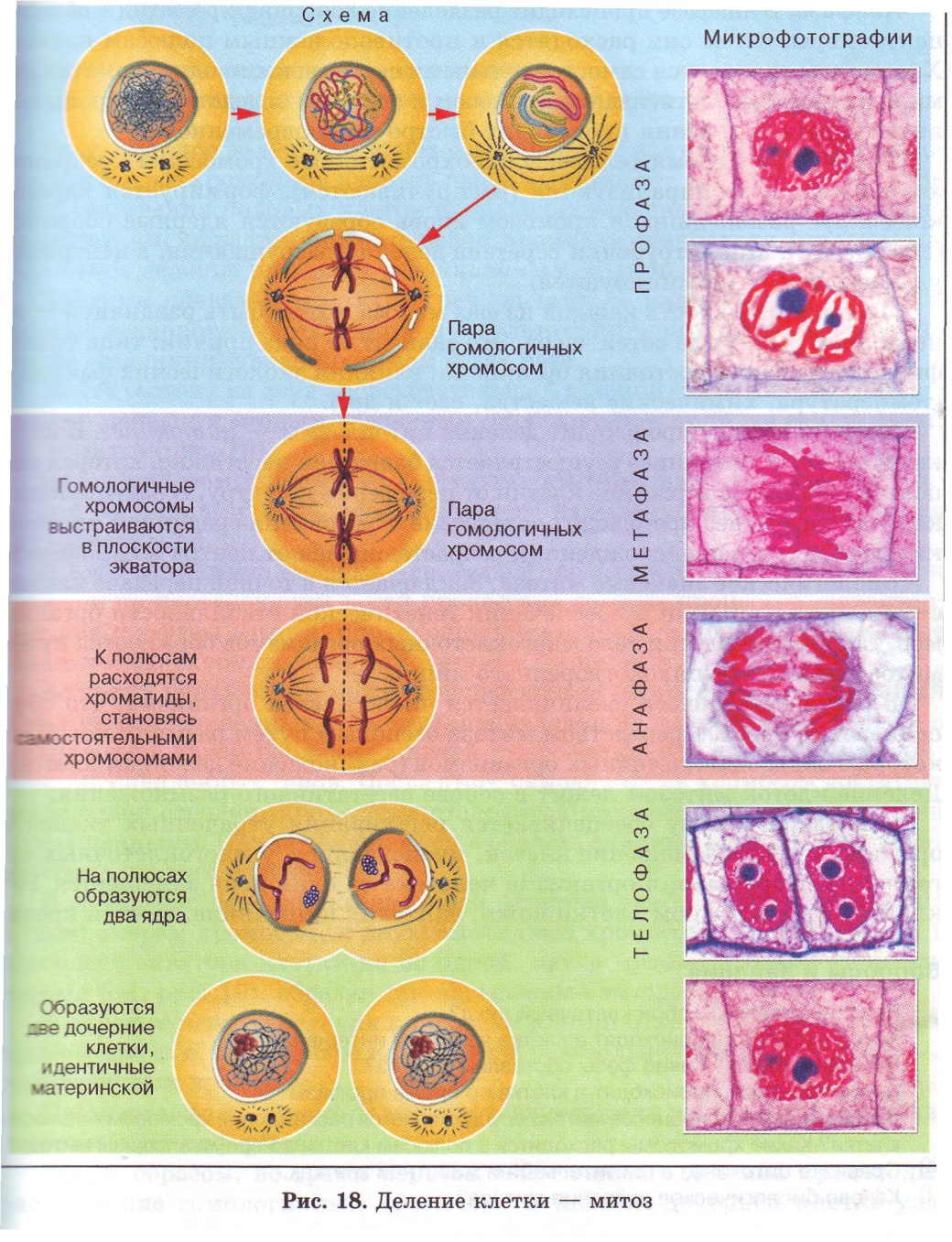


 Эти факторы регулируют время и уровни экспрессии различных генов, участвующих в процессе клеточного цикла.
Эти факторы регулируют время и уровни экспрессии различных генов, участвующих в процессе клеточного цикла. 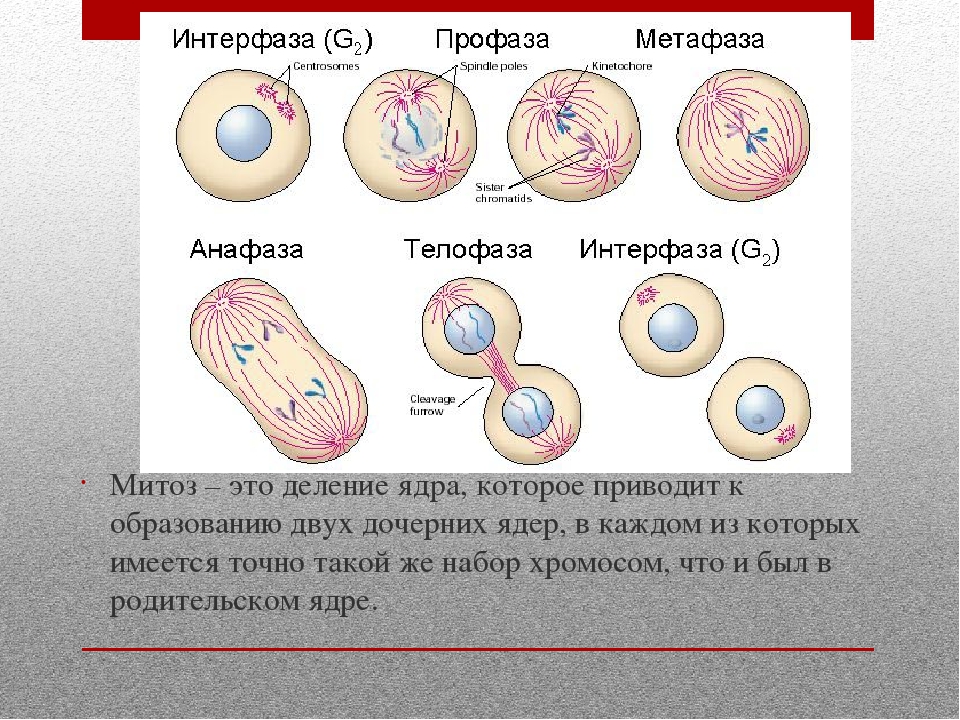
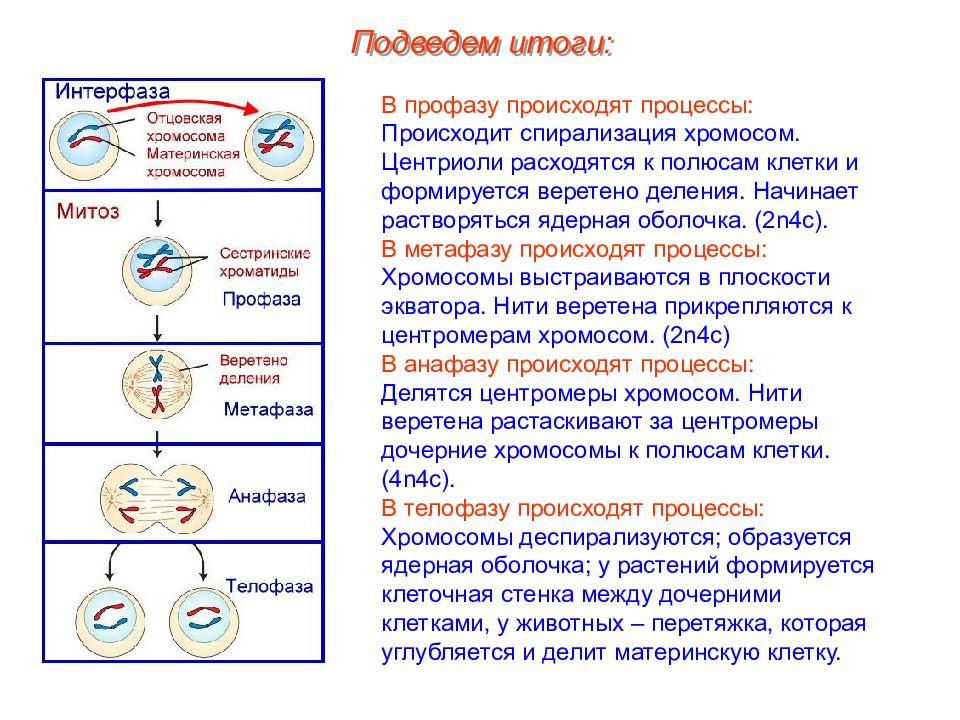


 и др. Молекулярная биология клетки.4-е издание. Нью-Йорк: Гарланд Наука; 2002. Обзор M Phase. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK26931/
и др. Молекулярная биология клетки.4-е издание. Нью-Йорк: Гарланд Наука; 2002. Обзор M Phase. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK26931/ ” Биохимический журнал том. 308 (часть 3), часть 3 (1995): 697-711. дои: 10.1042/bj3080697
” Биохимический журнал том. 308 (часть 3), часть 3 (1995): 697-711. дои: 10.1042/bj3080697 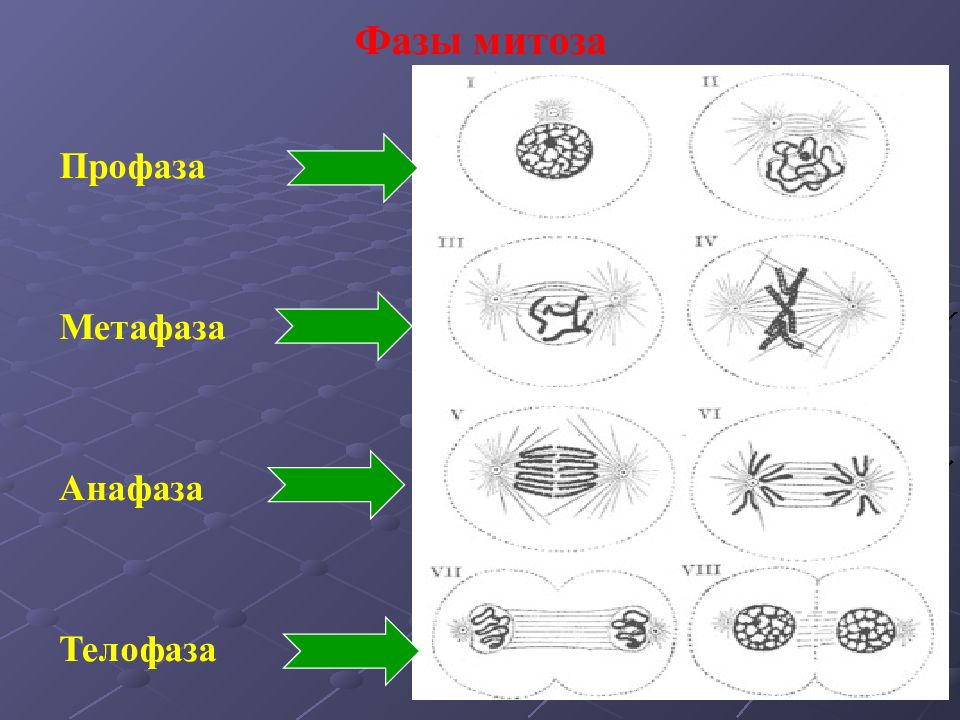 Патол Res Pract. 1996 г., июль; 192 (7): 669–75. doi: 10.1016/S0344-0338(96)80088-4. PMID: 8880867.
Патол Res Pract. 1996 г., июль; 192 (7): 669–75. doi: 10.1016/S0344-0338(96)80088-4. PMID: 8880867. 
