
Каковы различия между клеточной стенкой и клеточной мембраной? — Наука и Техника — Каталог статей
Клеточные стенки и клеточные мембраны служат самой внешней частью клетки. Роль каждого из них очень похожа с точки зрения транспорта и межсотового общения. Как клеточная мембрана, так и клеточная стенка существуют от образования клетки путем деления до ее гибели.
Существует 10 различных различий между клеточной мембраной и клеточной стенкой. Клеточная стенка присутствует только в клетках растений и клеток бактерий; животные клетки не имеют клеточной стенки, но вместо этого имеют только клеточную мембрану.
Клеточная стенка заключает в себе плазматическую мембрану и обеспечивает клетке достаточную жесткость, чтобы сохранить свою форму. Однако клеточная мембрана животной клетки более гибкая. Клетка у животного все еще имеет форму, но мембрана не такая жесткая, как клеточная стенка.
Клеточная стенка растения состоит из пектина, хитина и лигнина, тогда как в бактериальных клетках она состоит из гликолипидов и гликопротеинов.
Клеточная стенка жесткая, а также имеет фиксированную и четкую форму; давление, приложенное к клеточной стенке, может сломать ее. Клеточная мембрана является гибкой и обладает способностью сокращаться, изменяя свою форму по мере необходимости и удаляясь от силы.
Клеточная стенка не имеет каких-либо рецепторов, но клеточная мембрана имеет рецепторы, которые используются для получения сигналов от внешних химических сообщений других клеток.
Клеточная стенка включает жгутики и пилюли в виде маленьких волосоподобных предметов снаружи. В клеточной мембране бактериальной клетки есть жгутики, чтобы инициировать движение, и пилюли, чтобы помочь в репродуктивных функциях деления клетки.
Клеточная стенка на самом деле увеличивается в толщине с течением времени и присутствует со времени, когда клетка развивается посредством клеточного деления, пока клетка не умрет. Клеточная мембрана остается той же толщины со времени создания и на протяжении всей своей жизни в организме.
Клеточная стенка является полупроницаемой, что позволяет проходить веществам как внутрь клетки, так и из нее. Клеточная мембрана также проницаема и контролирует движение веществ внутрь и наружу клетки, но она избирательна в отношении веществ, которые она пропускает.
Клеточная стенка защищает клетку от сил во внешней резкой среде, в то время как клеточная мембрана выполняет функции проницаемости, получения химических сигналов, проводимости нервных клеток, деления клеток и размножения половым путем.
Клеточная стенка не нуждается в питании, кроме случаев, когда она повреждена и нуждается в ремонте. Клеточная мембрана требует постоянного питания от клетки, потому что она может сжиматься из-за потери питания и воды.
Учёные создали бактерии без стенок, способные к размножению
Бациллы, которых пенициллин лишает клеточных стенок, обычно не способны жить и размножаться. Английским учёным с помощью «противоестественного» отбора удалось вывести бактерии, которые прекрасно живут и делятся без стенки.
Клетки животных прячутся от внешнего мира за жиденькой двухслойной мембраной. Клетки грибов, растений и бактерий защищены куда лучше – помимо мембраны у них есть клеточная стенка, защищающая от резких колебаний температуры, кислотности и «солёности». Именно ею прокариоты и некоторые грибы обязаны своей феноменальной живучестью.
Эта же структура становится определяющей, когда речь заходит о размножении бактерий – именно с неё начинается «перетяжка» бактериальной клетки, заканчивающаяся появлением пары клеточных потомков.
Джеффу Эррингтону из британского Университета Ньюкасла и его коллегам удалось создать штаммы сенной палочки Bacillus subtilis, лишенные клеточной стенки, но при том сохранившие способность к размножению.
L-формы
бактерии, частично или полностью лишённые клеточной стенки, но сохранившие способность к развитию. L – первая буква названия Листеровского института в Лондоне, где Эмми Кляйнебергер-Нобель обратила внимание на развитие морфологически.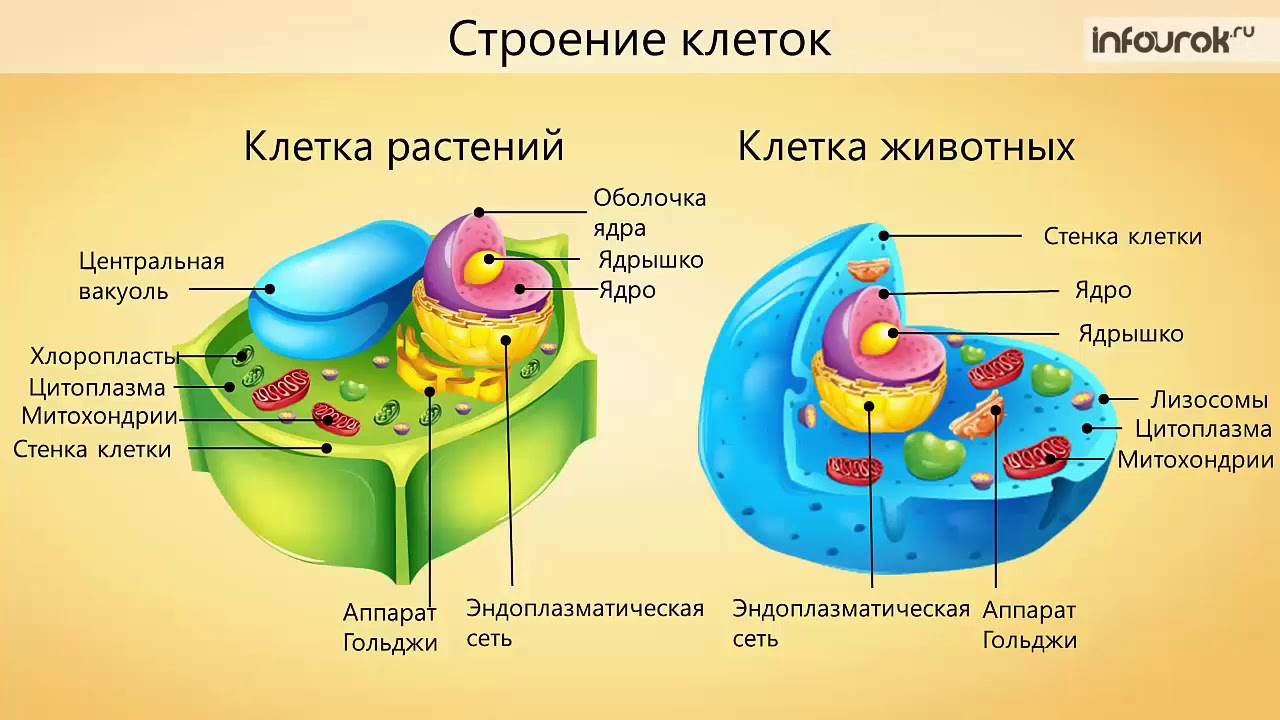 ..
..
Сам факт существования таких организмов не противоречит законам природы – Николай Федорович Гамалея ещё в 1894 году описал бактерии без стенки, позже получившие название «L-формы». Литера L к контурам прокариот не имеет никакого отношения, даже наоборот – в отсутствие жесткой стенки, играющей роль наружного скелета, бактерии просто не способны поддерживать свою форму, и «палочки» превращаются» в шарики, периодически образующие выросты.
Позже были открыты и бактерии, существующие только в L-форме, – это некоторые археи, а также спироплазмы, микоплазмы и уреаплазмы, ставшие настоящей головной болью для ученых и врачей. И хотя у всех у них сохранена липидная мембрана, отграничивающая внутреннее содержимое от внешней среды, защитной роли она не несёт.
Штамм
выделенный в чистой культуре изолят вируса или организма, размножающегося бесполым путём.
Отсутствующая у нас самих клеточная стенка – отличная мишень для антибиотиков: остальные методы «вытравливания» заразы более токсичны для эукариотических клеток. Отчасти этим и обусловлен интерес Эррингтона и соавторов публикации в Nature к образованию и делению L-форм.
Отчасти этим и обусловлен интерес Эррингтона и соавторов публикации в Nature к образованию и делению L-форм.
Чтобы ответить на вопрос, как это происходит, ученые взяли штамм сенной палочки M96. В этих «экспериментальных» бактериях синтез важнейшего компонента клеточной стенки пептидогликана происходит только при наличии в среде «древесного сахара» ксилозы. Без неё не запускается процесс считывания гена, кодирующего одно из веществ-предшественников пептидогликана. Микробиологи ксилозу добавлять не стали, наслаждаясь спонтанным появлением шарообразных L-форм. Но эти клетки практически не делились и были слишком разнообразны, так что говорить о воспроизводимости результатов не представлялось возможным.
Пришлось встроить в геном ещё один репрессор синтеза пептидогликана и включить механизм искусственного отбора, добавив в среду пенициллин, тоже ограничивающий сборку клеточной стенки.
Выжили только те, кто в результате случайных мутаций «научился» обходиться без стенки.
Собственно, этот штамм и стал основным достижением Эррингтона и главным объектом для дальнейших исследований. Результатами учёные могут гордиться:
им удалось сфотографировать размножение L-форм, больше похожее на почкование, нежели на традиционное равноценное деление на две части.
Обычно деление бактерии, уже «удвоившей» свой геном, начинается с Z-кольца, опоясывающего клеточную стенку. То есть, в отличие от тех же животных, процесс запускается снаружи, а не изнутри. «Дефектные» герои этой работы пошли другим путём: после увеличения в размерах до 3–4 микрометров они начинали «выпускать» многочисленные нестабильные выросты, пока один из них просто не отпочковывался, рос и снова давал жизнеспособное поколение. Полное наблюдение этого цикла под микроскопом заняло у ученых 7 часов.
«Дефектные» герои этой работы пошли другим путём: после увеличения в размерах до 3–4 микрометров они начинали «выпускать» многочисленные нестабильные выросты, пока один из них просто не отпочковывался, рос и снова давал жизнеспособное поколение. Полное наблюдение этого цикла под микроскопом заняло у ученых 7 часов.
Сам же механизм образования выростов пока остается неизвестным. В случае с животными подобное контролируемое выпячивание мембраны – обычное явление, необходимое для движения клеток или фагоцитоза. Но бактерии лишены развитого цитоскелета, способного самостоятельно регулировать форму клетки. Возможно, эти выросты, которых за раз образуется до 5 штук, формируются под действием белков, «растаскивающих» делящуюся ДНК, хотя детали феномена ещё предстоит установить.
В любом случае у микробиологов появилась новая модель для отработки перспектив в лечении уреаплазмоза, не поддающегося традиционным антибиотикам.
Да и на эволюцию прокариот можно взглянуть с другой стороны.
Не исключено, что именно подобным «дефектным» бесстеночным формам, отказавшимся от мощного железного занавеса ради большей восприимчивости и мобильности, животные обязаны своим возникновением.
что это такое, как применяются
Cостояние печени играет жизненно важную роль для здоровья человека. Этот орган весом более килограмма выполняет множество задач. В одной клетке печени, гепатоците, протекает около 500 различных биохимических процессов1. В печени осуществляется распад и/или детоксикация вредных веществ, а также выведение их из организма. Кроме того, орган синтезирует важнейшие составные части биологических мембран — фосфолипиды2.
Что такое клеточная мембрана?
Клетки — основные строительные блоки человеческого организма. Они, в свою очередь, не могут существовать без жиров и фосфолипидов, образующих наружную мембрану, «стенку» клетки, которая удерживает внутри нее цитоплазму.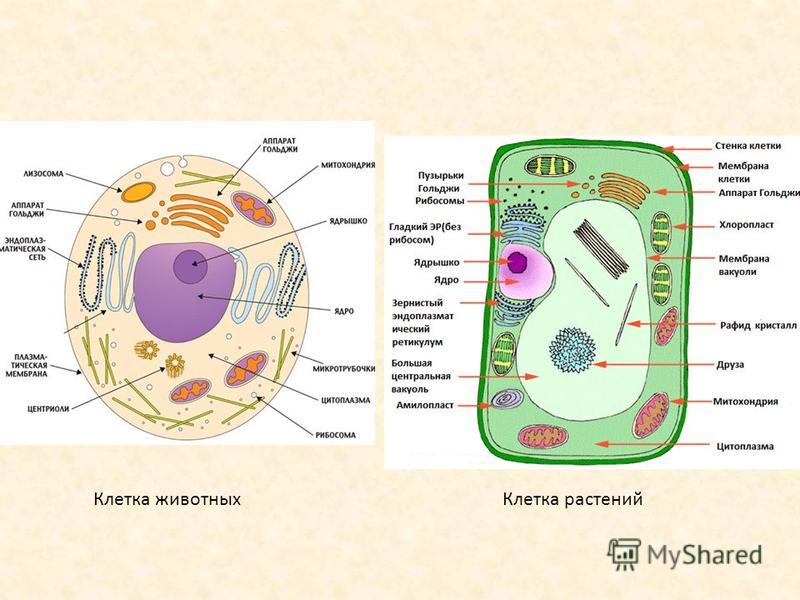 Мембрана представляет собой два слоя фосфолипидов, которые состоят из гидрофильной «головки», притягивающейся к воде, и гидрофобного, то есть водоустойчивого «хвоста». «Головки» двух рядов фосфолипидов обращены наружу, к жидкости, а «хвосты» скрепляются друг с другом, обеспечивая клеточным стенкам высокую прочность
2.
Мембрана представляет собой два слоя фосфолипидов, которые состоят из гидрофильной «головки», притягивающейся к воде, и гидрофобного, то есть водоустойчивого «хвоста». «Головки» двух рядов фосфолипидов обращены наружу, к жидкости, а «хвосты» скрепляются друг с другом, обеспечивая клеточным стенкам высокую прочность
2.
Фосфолипиды выполняют структурную функцию, поддерживая клеточный каркас, участвуют в процессах молекулярного транспорта, ферментативных и других, не менее значимых процессах. Любое нарушение их деятельности может иметь самые серьезные последствия2.
Гепатоциты — «кирпичики» печени
Клетки печени, гепатоциты, составляют до 70 — 85% массы органа. Они несут основную ответственность за деятельность печени, участвуя в таких процессах, как:
- Синтез и хранение протеинов
- Расщепление углеводов
- Синтез холестерина, желчных солей и фосфолипидов
- Детоксикация, расщепление и выведение веществ
- Инициирование образования и, собственно, выработка желчи.

Клеточная стенка гепатоцитов, как и любых других клеток, состоит из фосфолипидов, обеспечивающих ее полноценное функционирование. Однако, к сожалению, она уязвима. Воздействие негативных факторов, например, некоторых лекарственных препаратов, токсичных веществ и особенно алкоголя и даже несбалансированного рациона приводит к нарушению внутриклеточного обмена и гибели гепатоцитов. Так развиваются различные заболевания печени1.
Когда печень «шалит»?
Проблемы с печенью прежде всего связаны с хронической интоксикацией, которая, в свою очередь, может быть вызвана различными заболеваниями и состояниями. К ним относится хронический прием алкоголя, сахарный диабет 2 типа, экологическая интоксикация, «химизация» пищи и быта, неблагоприятное действие лекарств и другие факторы. Все они способствуют развитию оксидативного стресса вследствие нарушения адекватной работы антиоксидантных механизмов. Постепенно на фоне хронического негативного влияния происходит деструкция клеточных мембран, белков и ДНК, нарушается работа клетки.
Постепенно на фоне хронического негативного влияния происходит деструкция клеточных мембран, белков и ДНК, нарушается работа клетки.
Итогом длительной интоксикации является триада: перекисное окисление липидов (окислительная их деградация, происходящая под действием свободных радикалов), накопление в клетках печени жира более 5% от массы органа (стеатоз) и хроническое воспаление1.
Эссенциальный — значит, необходимый
Для лечения поражений печени различного происхождения широко применяются эссенциальные фосфолипиды (ЭФЛ). Их принципиальным отличием от обычных фосфолипидов является наличие дополнительной молекулы линолевой кислоты. Это позволяет ЭФЛ с легкостью восполнять дефекты клеточной мембраны, что увеличивает ее гибкость и нормализует функции. Именно наличие линолевой кислоты считается наиболее важным отличием ЭФЛ от классических фосфолипидов, например, лецитина, которое лежит в основе лечебных преимуществ эссенциальных фосфолипидов2.
В каком-то смысле назначение ЭФЛ можно назвать мембранной терапией, ведь их активность связана именно с клеточными стенками. Возможным же такое лечение стало благодаря соевым бобам, из которых и получают ценное вещество3.
Соевые бобы: из глубины веков до наших дней
Однолетнее растение семейства Бобовые на протяжении тысячелетий используется человеком. Упоминания о нем есть в книгах времен императора Шэн Нунг, царствовавшего в 2838 году до нашей эры1. Тогда соевые бобы считались одни из пяти «святых зерновых», без которых была невозможна жизнь на земле.
Сегодня известно около 800 видов соевых бобов. Они содержат 35 — 40% белков, 20 — 30% углеводов, 5 — 10% сопутствующих веществ (витамины, тритерпеновые сапонины, флавоноиды и т.д.), а также 12 — 18% жиров. Масла, входящие в состав бобов, на 90 — 95% состоят из глицеридов жирных кислоты, в частности, олеиновой и линолевой. В процессе переработки сырого масла удается получить 30 — 45% соевого лецитина (фосфатидилхолина), который и является «целевым продуктом», используемым в фармацевтической промышленности для создания препаратов, проявляющих гепатопротекторный эффект.
Фосфатидилхолины в действии
Получаемые из соевых бобов фосфатидилхолины представляют собой типичный липидный двойной слой, состоящий из гидрофильной «головки» и гидрофобного «хвоста» и являющийся основным структурным компонентом биологических мембран. Эссенциальные фосфолипиды легко заменяют эндогенные, то есть, «собственные» фосфолипиды организма, которые оказались по тем или иным причинам повреждены, встраиваясь в клеточную мембрану. При этом ЭФЛ могут поступать в организм как перорально, в виде твердых лекарственных форм (капсулы), так и внутривенно, с инъекционным раствором высокой степени очистки4.
Кстати, название «эссенциальные фосфолипиды» зарегистрировано только для препаратов, где содержится не менее 75% фосфатилхолина. Примеромлекарственного средства, содержащего ЭФЛ в высокой концентрации — Эссенциале Форте Н. В 1 капсуле Эссенциале форте Н содержится 76% фосфатидилхолина3.
На защиту мембран!
Встраивание эссенциальных фосфолипидов в поврежденные мембраны гепатоцитов обеспечивает восстановление нормальных мембранных структур, что, в свою очередь приводит к комплексному терапевтическому эффекту. Какое же действие оказывают ЭФЛ? Прежде всего, они проявляют протективные (защитные) и регенеративные свойства в отношении клеток печени1. При этом поражение печени может быть вызвано самыми различными факторами, среди которых токсические, воспалительные, аллергические, обменные и иммунологические реакции1.
Какое же действие оказывают ЭФЛ? Прежде всего, они проявляют протективные (защитные) и регенеративные свойства в отношении клеток печени1. При этом поражение печени может быть вызвано самыми различными факторами, среди которых токсические, воспалительные, аллергические, обменные и иммунологические реакции1.
Доказано, что ЭФЛ защищают гепатоциты при повреждениях, связанных с действием различных химических веществ, алкоголя, наркотических препаратов, цитостатиков, которые применяются для лечения онкологических заболеваний, ионизирующего излучения и так далее1.
ЭФЛ и полиненасыщенные жирные кислоты
Механизм действия ЭФЛ сродни действию омега 3-6-полиненасыщенных жирных кислот (ПНЖК), поскольку первые представляют собой по сути природную форму существования вторых. Омега 3-6-ПНЖК — эссенциальные жиры, снижающие риск ряда заболеваний, в том числе сердечно-сосудистых и диабета. Они не синтезируются в организме, и должны ежедневно поступать в организм в количестве 2 г омега-3 и 6 граммов омега-6 ПНЖК. Однако красные сорта рыбы, грецкие орехи, растительные масла, соевые бобы, где содержатся в большом количестве эти кислоты, высококалорийны, что ограничивает их употребление5.
Однако красные сорта рыбы, грецкие орехи, растительные масла, соевые бобы, где содержатся в большом количестве эти кислоты, высококалорийны, что ограничивает их употребление5.
Препараты, содержащие ЭФЛ, например, Эссенциале форте Н, могут широко применяться для восполнения диетического дефицита полиненасыщенных жирных кислот, не увеличивая калорийность рациона6. А какую важную роль они играют при заболеваниях, связанных с нарушением обмена жиров!
Холестерин — стоп!
Эссенциальные фосфолипиды принимают участие в транспорте холестерина в плазме и тканях, а также образовании липопротеинов высокой и низкой плотности (ЛПВП и ЛПНП) 7. Напомним, что именно с повышением уровня ЛПНП и триглицеридов и снижением содержания ЛПВП связано одно из самых опасных заболеваний — атеросклероз.
ЭФЛ обеспечивают так называемую системную мобилизацию холестерина и его утилизацию на всех этапах метаболизма за счет ряда процессов, в том числе:
- Повышение синтеза ЛПНП и мобилизация холестерина из плазмы крови
- Обеспечение захвата ЛПНП гепатоцитами
- Повышение секреции холестерина, фосфолипидов и жиров в желчь, что, соответственно, снижает их накопление в печени.
 8
8
В состав желчи входят желчные кислоты (около 70%) и ЭФЛ (фосфатидилхолин, составляет 22% желчи)9. У здоровых людей ЭФЛ обеспечивают растворимость холестерина. Если же соотношение желчных кислот и ЭФЛ нарушается, кристаллы холестерина могут выпадать в осадок, вследствие чего развивается желчнокаменная болезнь (ЖКБ).
Камни в желчном пузыре: легче предотвратить, чем лечить
Как известно, до 90% желчных камней являются преимущественно холестериновыми, состоящими как минимум на 70% из холестерина10. Увеличение выброса в желчь холестерина, приводящее к камнеобразованию, может происходить вследствие несбалансированности рациона и его насыщения животными жирами. Важным фактором риска ЖКБ считается и экологическое загрязнение, приводящее к хронической интоксикации организма и, как следствие, увеличению потребности в антиоксидантах и ЭФЛ, необходимых для связывания токсических веществ. Если запасы ЭФЛ и антиоксидантов не пополняются, функция клеточных мембран нарушается, и организм начинает накапливать холестерин, чтобы сохранить структуру клеточной стенки, запуская процесс камнеобразования11, 12.
Если запасы ЭФЛ и антиоксидантов не пополняются, функция клеточных мембран нарушается, и организм начинает накапливать холестерин, чтобы сохранить структуру клеточной стенки, запуская процесс камнеобразования11, 12.
К группе повышенного риска ЖКБ относятся лица, злоупотребляющие алкоголем. Профилактическое назначение препаратов ЭФЛ тем, кто испытывает высокую нагрузку токсинами или алкоголем, компенсирует дефицит фосфолипидов и препятствует камнеобразованию12.
Неалкогольная жировая болезнь печени
Важный эффект ЭФЛ достигается благодаря их способности улучшать обмен липидов. Он особенно актуален при лечении неалкогольной жировой болезни печени (НАЖБП), которая возникает вследствие избыточного накопления жиров в органе. Необходимо отметить, что распространенность этого заболевания постоянно растет, в том числе и у детей и подростков, особенно живущих в городах6. В РФ только у пациентов общей практики распространенность НАЖБП достигает 27%6!
В РФ только у пациентов общей практики распространенность НАЖБП достигает 27%6!
На фоне заболевания развивается комплекс патологических нарушений, среди которых снижение чувствительности тканей к инсулину, повышение содержания инсулина в крови. Увеличивается масса висцерального жира, что приводит к ожирению и артериальной гипертензии. У больных часто развивается сахарный диабет 2 типа и гиперлипидемия — повышение уровня холестерина и триглицеридов в крови. Люди, страдающие НАЖБП, должны контролировать калорийность рациона, минимизировать поступление транс-жиров, холестерина и увеличить потребление полиненасыщенных липидов (более 10 граммов в сутки)7. В связи с высокой калорийностью содержащих ПНЖК продуктов особое значение приобретают препараты ЭФЛ, оказывающие выраженный терапевтический эффект.
Фосфолипиды при НАЖБП
При жировой инфильтрации гепатоцитов, которая наблюдается у больных НАЖБП, препараты, содержащие эссенциальные фосфолипиды, способствуют13 снижению стресса митохондрий клеток, обусловленного избытком жирных кислот, уменьшению выраженности воспалительного процесса и нормализации жирового обмена за счет коррекции дислипидемии14. Назначение этих препаратов позволяет улучшить липидный состав крови, показатели перекисного окисления жиров и восстановить систему антиоксидантной защиты в целом6.
Назначение этих препаратов позволяет улучшить липидный состав крови, показатели перекисного окисления жиров и восстановить систему антиоксидантной защиты в целом6.
Клинически доказано, что эссенциальные фосфолипиды, например, в составе препарата Эссенциале форте Н, при НАЖБП улучшают течение и прогноз жировой инфильтрации печени, повышают чувствительность тканей к инсулину, нормализуют липидный профиль (то есть, способствуют снижению уровня «плохого» холестерина и триглицеридов) и к тому же уменьшают выраженность симптомов нарушений пищеварения.
Испытание алкоголем
Не менее важным показанием к назначению эссенциальных фосфолипидов является алкогольная болезнь печени (АБП). Высокому риску развития заболевания подвергаются люди, которые ежедневно принимают 40-80 мл чистого этанола на протяжении 4-6 лет15. Алкоголь богат «легкими» углеводными калориями, которые постепенно вызывают жировую дегенерацию внутренних органов. Кроме того, он способствует развитию нарушения кровообращения органа. При хронической алкогольной интоксикации на фоне оксидативного стресса происходит некроз гепатоцитов, развивается хронический воспалительный процесс и фиброз печени. Механизм развития последнего во многом обусловлен активацией так называемых звездчатых клеток. В норме они находятся в состоянии покоя, а при повреждении гепатоцитов «просыпаются» и становятся способными к интенсивному делению в участках воспаления16. В результате ткань печени разрастается, и в дальнейшем этот процесс часто заканчивается циррозом.
Кроме того, он способствует развитию нарушения кровообращения органа. При хронической алкогольной интоксикации на фоне оксидативного стресса происходит некроз гепатоцитов, развивается хронический воспалительный процесс и фиброз печени. Механизм развития последнего во многом обусловлен активацией так называемых звездчатых клеток. В норме они находятся в состоянии покоя, а при повреждении гепатоцитов «просыпаются» и становятся способными к интенсивному делению в участках воспаления16. В результате ткань печени разрастается, и в дальнейшем этот процесс часто заканчивается циррозом.
Действие ЭФЛ при алкогольном поражении печени
Назначение эссенциальных фосфолипидов при алкогольном поражении печени позволяет связать активные формы алкоголя, которые образуются при поступлении высоких доз алкоголя и «запускают» механизм оксидативного стресса. Благодаря высокой биодоступности и возможности внедряться в клеточные мембраны гепатоцитов ЭФЛ способствуют удалению свободных радикалов и восстанавливают структуру клеточной стенки. Кроме того, ЭФЛ оказывают антифибротическое действие, подавляя активацию звездчатых клеток печени и останавливая патологический процесс развития фиброза17.
Кроме того, ЭФЛ оказывают антифибротическое действие, подавляя активацию звездчатых клеток печени и останавливая патологический процесс развития фиброза17.
ЭФЛ можно назвать универсальным средством, оказывающим противовоспалительное, антифибротическое действие, а также предотвращающим гибель клеток печени18. Эти возможности обеспечивают эффективность при хронических гепатитах, циррозе, жировой дистрофии печени, алкогольном гепатите и других нарушениях работы печени.
Праймер для микроскопии молекулярных выражений: методы специализированной микроскопии — Галерея цифровых изображений флуоресценции
Галерея цифровых изображений флуоресценции
Клетки в культуре
Серьезные попытки культивирования целых тканей и изолированных клеток были впервые предприняты в начале 1900-х годов как метод исследования поведения животных клеток в изолированной и строго контролируемой среде. Термин культура ткани возник потому, что большинство ранних клеток было получено из первичных тканевых эксплантатов, метод, который доминировал в этой области более 50 лет. По мере появления установленных клеточных линий применение четко определенных нормальных и трансформированных клеток в биомедицинских исследованиях стало важным элементом развития клеточной и молекулярной биологии. В этой галерее флуоресцентных изображений исследуется более 30 наиболее распространенных клеточных линий, меченных различными флуорофорами с использованием как традиционных методов окрашивания, так и методов иммунофлуоресценции.
По мере появления установленных клеточных линий применение четко определенных нормальных и трансформированных клеток в биомедицинских исследованиях стало важным элементом развития клеточной и молекулярной биологии. В этой галерее флуоресцентных изображений исследуется более 30 наиболее распространенных клеточных линий, меченных различными флуорофорами с использованием как традиционных методов окрашивания, так и методов иммунофлуоресценции.
Клетки эмбрионального фибробласта швейцарской мыши-альбиноса ( 3T3 ), представленные на цифровом изображении выше, были резидентными в прилипшей культуре, окрашенной на F-актин с помощью Alexa Fluor 488, конъюгированной с фаллоидином (зеленая флуоресценция), и на ДНК с бис-бензимидазолом. краситель Hoechst 33258 (псевдоцветный голубой).Кроме того, культуру иммунофлуоресцентно метили с помощью Alexa Fluor 568 (псевдоцветный красный) и Alexa Fluor 750 (псевдоцветный желтый), конъюгированных с козьими вторичными антителами, которые нацелены на мышиный антивиментин (промежуточные филаменты) и кроличий антигигантин (нацеленный на комплекс Гольджи). первичные антитела соответственно. Одновременно митохондриальную сеть визуализировали с помощью MitoTracker Deep Red 633 (псевдо-синий). Изображения были записаны в оттенках серого с помощью системы камеры QImaging Retiga Fast-EXi, соединенной с микроскопом Olympus BX-51, оснащенным оптическими блоками полосно-эмиссионного флуоресцентного фильтра, предоставленными Omega Filters.На этапе обработки отдельные каналы изображения были окрашены в псевдоцвет со значениями RGB, соответствующими каждому из спектральных профилей излучения флуорофора.
первичные антитела соответственно. Одновременно митохондриальную сеть визуализировали с помощью MitoTracker Deep Red 633 (псевдо-синий). Изображения были записаны в оттенках серого с помощью системы камеры QImaging Retiga Fast-EXi, соединенной с микроскопом Olympus BX-51, оснащенным оптическими блоками полосно-эмиссионного флуоресцентного фильтра, предоставленными Omega Filters.На этапе обработки отдельные каналы изображения были окрашены в псевдоцвет со значениями RGB, соответствующими каждому из спектральных профилей излучения флуорофора.
Клеточные линии почек африканской зеленой обезьяны — Африканская зеленая мартышка была обычным объектом научных исследований в течение многих лет, и клетки тканей этого вида, Cercopithecus aethiops , наряду с клетками макаки-резуса, имеют используется для производства вакцин против полиомиелита с 1950-х годов.В последнее время африканские зеленые мартышки вызвали значительный интерес из-за высокого уровня вируса обезьяньего иммунодефицита типа 1 ( SIV-1 ), который часто встречается в их крови, что может быть отдаленно связано с вирусом иммунодефицита человека ( HIV ). ), вызывающий СПИД. Кроме того, несколько нормальных и трансформированных линий клеток почек африканских зеленых мартышек являются отличными кандидатами для исследований трансфекции рекомбинантными плазмидами.
), вызывающий СПИД. Кроме того, несколько нормальных и трансформированных линий клеток почек африканских зеленых мартышек являются отличными кандидатами для исследований трансфекции рекомбинантными плазмидами.
-
Нормальные клетки фибробластов почек африканской зеленой обезьяны (CV-1) — Клеточная линия CV-1 была создана в марте 1964 года F.К. Дженсен и его коллеги использовали срез ткани, вырезанный из почки нормального взрослого самца африканской зеленой обезьяны ( Cercopithecus aethiops ). Популярная линия фибробластов первоначально использовалась в исследованиях, направленных на трансформацию вызывающего рак вируса саркомы Рауса ( RSV ), но теперь она является очень полезным хозяином для исследований приобретенного иммунодефицита ( AIDS ), а также экспериментов по трансфекции. с обезьяньим вирусом 40 и рекомбинантными плазмидными векторами.
-
Нормальные эпителиальные клетки почек африканской зеленой обезьяны (Vero) — Линия эпителиальных клеток Vero была создана в 1962 году Y.
 Yasumura и Y. Kawakita в университете Chiba в Тибе, Япония. Ткань, из которой была получена линия, была получена из почек здоровой взрослой африканской зеленой обезьяны. Хотя клетки Vero широко используются для трансфекций и производства вакцин, они также часто используются для обнаружения веротоксинов, группы взаимосвязанных токсинов, продуцируемых некоторыми штаммами Escherichia coli , которые являются ключевой причиной геморрагического колита и гемолитико-уремического синдрома у люди.
Yasumura и Y. Kawakita в университете Chiba в Тибе, Япония. Ткань, из которой была получена линия, была получена из почек здоровой взрослой африканской зеленой обезьяны. Хотя клетки Vero широко используются для трансфекций и производства вакцин, они также часто используются для обнаружения веротоксинов, группы взаимосвязанных токсинов, продуцируемых некоторыми штаммами Escherichia coli , которые являются ключевой причиной геморрагического колита и гемолитико-уремического синдрома у люди. -
Трансформированные (обезьяний вирус 40) клетки фибробластов почек африканской зеленой обезьяны (COS-1) — Трансформированная линия клеток COS-1 была получена из линии фибробластов CV-1 , начало которой было положено в марте 1964 г. Дженсен и его коллеги из нормальной почки взрослой африканской зеленой обезьяны. Клеточная линия COS-1, разработанная Яковом Глузманом, отличается от CV-1 из-за трансформации более ранней линии мутантом обезьяньего вируса 40 ( SV40 ), который кодирует Т-антиген дикого типа.
 Две другие трансформированные линии, COS-3 и COS-7, также были созданы Глузманом, но клетки COS-1 уникальны тем, что содержат единственную интегрированную копию полной ранней области ДНК SV40. Однако все три линии полностью поддерживают литический рост SV40 (штамм tsA209 ) при 40 градусах Цельсия и репликацию мутантов SV40 с делециями в ранней области.
Две другие трансформированные линии, COS-3 и COS-7, также были созданы Глузманом, но клетки COS-1 уникальны тем, что содержат единственную интегрированную копию полной ранней области ДНК SV40. Однако все три линии полностью поддерживают литический рост SV40 (штамм tsA209 ) при 40 градусах Цельсия и репликацию мутантов SV40 с делециями в ранней области. -
Трансформированные (обезьяний вирус 40) клетки фибробластов почек африканской зеленой обезьяны (COS-7) — Клеточная линия COS-7 была получена Яковом Глузманом в начале 1980-х годов из ранее установленной почки африканской зеленой мартышки CV-1 линии путем трансформации нормальных клеток дефектным по происхождению мутантом обезьяньего вируса 40 ( SV40 ), который кодирует Т-антиген вируса дикого типа.Линия фибробластов растет в культуре на стекле и пластике и обычно используется в качестве хозяина для трансфекции вирусных геномов и рекомбинантных плазмид.
Клетки фибробластов кожи африканского водяного мангуста (APM) — Линия клеток APM была создана в Лаборатории морских биологических наук ( NBL ) в Окленде, штат Калифорния, из кожи африканского водяного мангуста ( Atilax paludinosus ) . Неуловимое животное, африканский водяной мангуст ведет уединенный ночной образ жизни и хорошо плавает, хотя часто задерживается на мелководье, где ловит моллюсков, крабов, лягушек и других водных и полуводных животных для пропитания.Клетки мангуста APM обладают морфологией фибробластов и, как и другие линии фибробластов, относятся к числу клеток, наиболее легко выращиваемых в культуре. Клеточные биологи предполагают, что способность фибробластов так быстро расти за пределами тела связана с их центральной ролью в заживлении ран, что требует их размножения при травмах или других менее чем оптимальных условиях. Также обычно считается, что фибробласты ведут относительно одиночный образ жизни, что, по мнению некоторых, также может быть фактором их благоприятного роста в культуре.
Неуловимое животное, африканский водяной мангуст ведет уединенный ночной образ жизни и хорошо плавает, хотя часто задерживается на мелководье, где ловит моллюсков, крабов, лягушек и других водных и полуводных животных для пропитания.Клетки мангуста APM обладают морфологией фибробластов и, как и другие линии фибробластов, относятся к числу клеток, наиболее легко выращиваемых в культуре. Клеточные биологи предполагают, что способность фибробластов так быстро расти за пределами тела связана с их центральной ролью в заживлении ран, что требует их размножения при травмах или других менее чем оптимальных условиях. Также обычно считается, что фибробласты ведут относительно одиночный образ жизни, что, по мнению некоторых, также может быть фактором их благоприятного роста в культуре.
Клетки фибробластов почек детенышей хомяка (BHK-21) — Линия клеток фибробластов BHK-21 была создана в марте 1961 г. И. А. Макферсоном и М. Г. П. Стокером. Широко используемая линия представляет собой субклон (клон 13) родительской линии, полученной из почек пяти бесполых однодневных хомяков. Эти хомяки принадлежали к виду Mesocricetus auratus и широко известны как сирийские золотые хомяки. После 84 дней непрерывного культивирования, прерываемого только на 8-дневное сохранение путем замораживания, клон 13 был инициирован путем выделения одиночных клеток.Клетки BHK-21 чувствительны к аденовирусу D человека, реовирусу 3 и вирусу везикулярного стоматита (штамм Indiana), но устойчивы к полиовирусу 2. Кроме того, клетки отрицательны по обратной транскриптазе, что указывает на отсутствие целостных геномов ретровируса. Линия BHK-21 использовалась в качестве хозяина для трансформации экспрессионными векторами, содержащими селектируемые и амплифицируемые маркерные ДНК, а также полезна для трансфекций.
Эти хомяки принадлежали к виду Mesocricetus auratus и широко известны как сирийские золотые хомяки. После 84 дней непрерывного культивирования, прерываемого только на 8-дневное сохранение путем замораживания, клон 13 был инициирован путем выделения одиночных клеток.Клетки BHK-21 чувствительны к аденовирусу D человека, реовирусу 3 и вирусу везикулярного стоматита (штамм Indiana), но устойчивы к полиовирусу 2. Кроме того, клетки отрицательны по обратной транскриптазе, что указывает на отсутствие целостных геномов ретровируса. Линия BHK-21 использовалась в качестве хозяина для трансформации экспрессионными векторами, содержащими селектируемые и амплифицируемые маркерные ДНК, а также полезна для трансфекций.
Эндотелиальные клетки легочной артерии крупного рогатого скота (BPAE) — Клеточная линия BPAE была создана в январе 1979 г.Del Vecchio из главного ствола легочной артерии молодой коровы ( Bos taurus ). Легочные артерии, которые простираются от сердца к легким, являются единственными артериями в организме млекопитающих, по которым течет темная неоксигенированная кровь.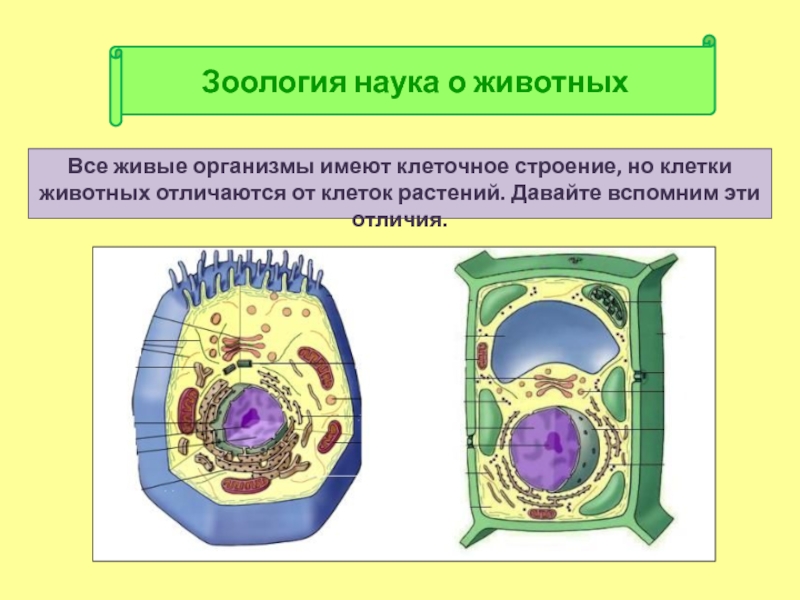 Линия эндотелиальных клеток BPAE положительна в отношении вируса диареи крупного рогатого скота, одного из наиболее важных известных вирусных патогенов крупного рогатого скота, вызывающего широкий спектр клинических синдромов, которые ежегодно приводят к значительным потерям в мясной промышленности.Клетки BPAE также положительны в отношении фермента, преобразующего ангиотензин ( ACE ), фермента, который принимает активное участие в поддержании кровяного давления и объема. По этой причине клетки BPAE часто используются в исследованиях гипертонии, а также в исследованиях атеросклероза и ишемической болезни сердца.
Линия эндотелиальных клеток BPAE положительна в отношении вируса диареи крупного рогатого скота, одного из наиболее важных известных вирусных патогенов крупного рогатого скота, вызывающего широкий спектр клинических синдромов, которые ежегодно приводят к значительным потерям в мясной промышленности.Клетки BPAE также положительны в отношении фермента, преобразующего ангиотензин ( ACE ), фермента, который принимает активное участие в поддержании кровяного давления и объема. По этой причине клетки BPAE часто используются в исследованиях гипертонии, а также в исследованиях атеросклероза и ишемической болезни сердца.
Клетки миобластов среднего слоя грудной аорты эмбриона крысы (A-10) — Клональная линия клеток A-10 была получена Б. Кимесом и Б. Брандтом из грудной аорты эмбриональной крысы ( Rattus norvegicus ). ) от установленного штамма DB1X .Грудная аорта — это ветвь нисходящей аорты, по которой кровь транспортируется от сердца к другим органам и частям тела. Обычно считается, что это произвольное анатомическое образование простирается от дуги аорты до диафрагмы. Клетки срослись и демонстрируют морфологию миобластов, обладая многими свойствами, характерными для гладкомышечных клеток. Клеточные продукты включают миокиназу, креатинфосфокиназу и миозин. Клетки A-10 производят спонтанные потенциалы действия в стационарной фазе цикла роста и демонстрируют повышение активности ферментов миокиназы и креатинфосфокиназы.
Обычно считается, что это произвольное анатомическое образование простирается от дуги аорты до диафрагмы. Клетки срослись и демонстрируют морфологию миобластов, обладая многими свойствами, характерными для гладкомышечных клеток. Клеточные продукты включают миокиназу, креатинфосфокиназу и миозин. Клетки A-10 производят спонтанные потенциалы действия в стационарной фазе цикла роста и демонстрируют повышение активности ферментов миокиназы и креатинфосфокиназы.
Эмбриональные клетки фибробластов швейцарской мыши (3T3) — Созданная Джорджем Тодаро и Ховардом Грином в 1962 году из дезагрегированной ткани эмбриона швейцарской мыши ( Mus musculus ) клеточная линия 3T3 представляет собой стандартную линию клеток фибробластов, используемую в широкий спектр исследовательских и промышленных биомедицинских приложений. Варианты исходной клеточной линии были протестированы и оказались отрицательными на вирус эктромелии (оспа мышей), но большинство из них чувствительны к полиоме и обезьяньему вирусу 40 ( SV40 ). Кроме того, клетки 3T3 отрицательны для вирусного фермента обратной транскриптазы, что указывает на отсутствие целостных геномов ретровирусов. Внутри цитоплазмы лизофосфатидилхолин ( lyso-PC ) индуцирует активность AP-1 и активность N-концевой киназы c-jun ( JNK1 ) посредством протеинкиназного C-независимого пути. Контакт подавлен, сливающийся монослой клеток 3T3 дает примерно 40 000 клеток на квадратный сантиметр.
Кроме того, клетки 3T3 отрицательны для вирусного фермента обратной транскриптазы, что указывает на отсутствие целостных геномов ретровирусов. Внутри цитоплазмы лизофосфатидилхолин ( lyso-PC ) индуцирует активность AP-1 и активность N-концевой киназы c-jun ( JNK1 ) посредством протеинкиназного C-независимого пути. Контакт подавлен, сливающийся монослой клеток 3T3 дает примерно 40 000 клеток на квадратный сантиметр.
Эпителиальные клетки почек кенгуру самок крысы (PtK1) — Линия клеток PtK1 была получена в 1962 году из почек нормальной взрослой самки Potorous tridactylus , вида крысиного кенгуру, распространенного в Австралии.Эпителиальная линия считается первой установленной постоянной клеточной линией сумчатого происхождения. Клетки PtK1 положительны на кератин при иммунопероксидазе и флуоресцентном окрашивании и подвержены везикулярному стоматиту (штамм Indiana). Однако эпителиальные клетки PtK1 устойчивы к полиовирусу 2, а также отрицательны в отношении обратной транскриптазы, что указывает на отсутствие целостных геномов ретровирусов.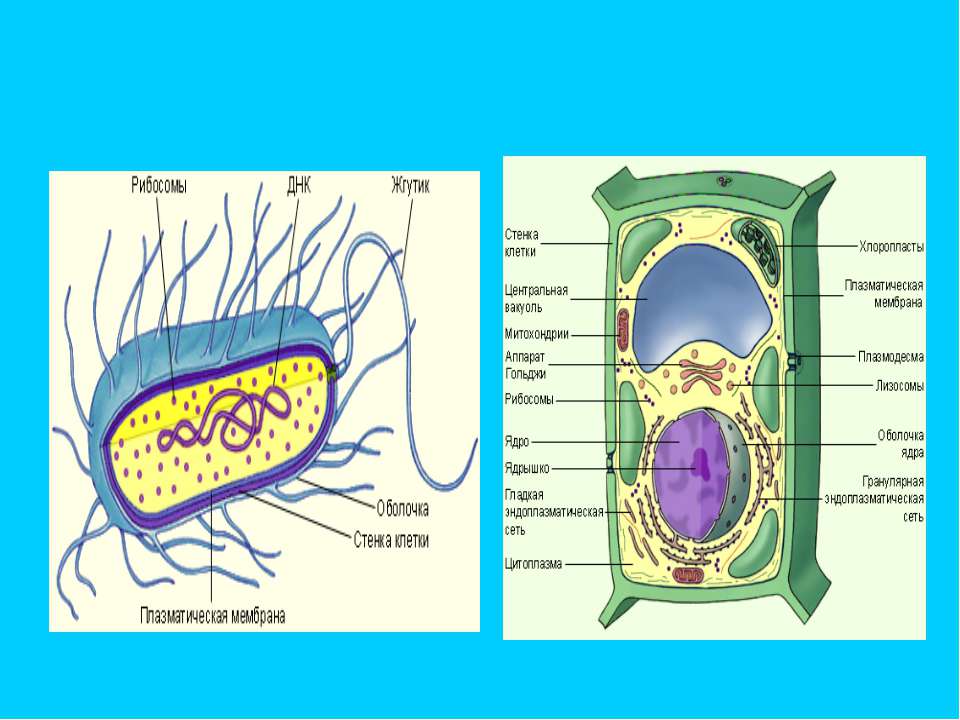 Клеточная линия в основном использовалась в митотических исследованиях из-за малого количества, большого размера и отличной морфологии хромосом того вида, из которого она была создана.
Клеточная линия в основном использовалась в митотических исследованиях из-за малого количества, большого размера и отличной морфологии хромосом того вида, из которого она была создана.
Клетки дермальных фибробластов лошади (NBL-6) — Линия клеток NBL-6 , также известная как E. Derm , была получена из дермы 4-летней самки лошади ( Equus caballus ) породы четвероногая лошадь. Клетки, которые, по-видимому, стареют примерно после 40 пассажей, восприимчивы к простому герпесу, реовирусу 3, везикулярному стоматиту (штамм Огден) и коровьей оспе, но устойчивы к аденовирусу 5, вирусу Коксаки А9 и В5 и полиовирусу 2.Клетки NBL-6, которые отрицательны в отношении обратной транскриптазы и демонстрируют типичную морфологию фибробластов, использовались для различных исследований, а также для размножения вирусов в вакцинах для лошадей. Эта клеточная линия сыграла особенно важную роль в исследованиях вирусного артериита лошадей ( EVA ), заразного вирусного заболевания, поражающего популяции лошадей во всем мире и, по-видимому, заболеваемости растущим.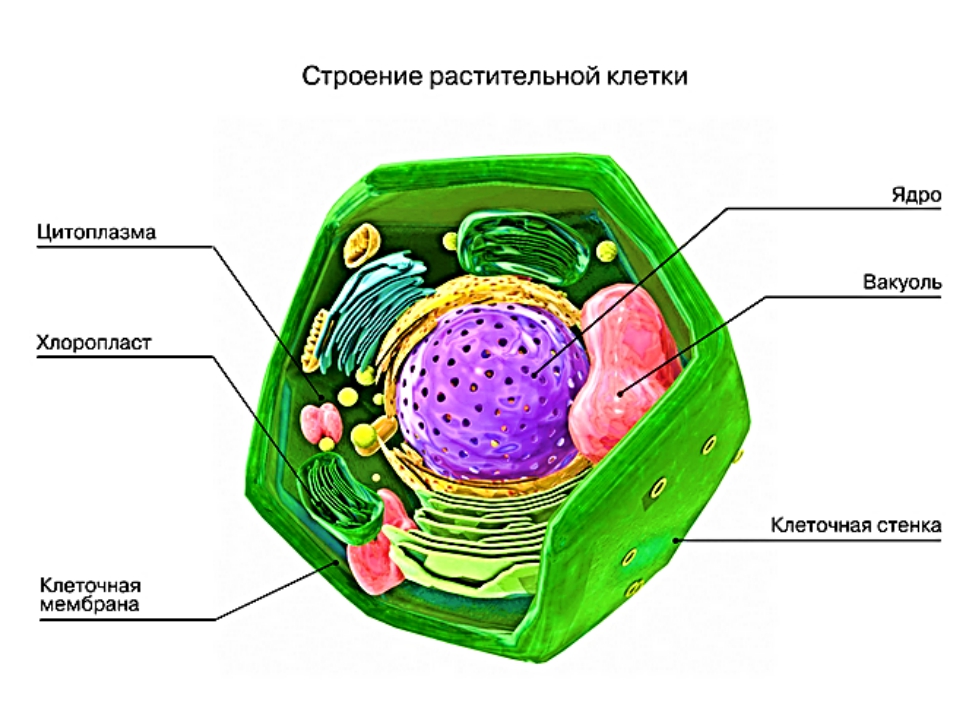
Клетки костной остеосаркомы человека (U-2 OS) — Клеточная линия U-2 OS , первоначально известная как линия 2T, была получена из костной ткани пятнадцатилетней женщины, страдающей остеосаркомой.Основанная J. Ponten и E. Saksela в 1964 г., исходные клетки были взяты из умеренно дифференцированной саркомы большеберцовой кости. Клетки U-2 OS демонстрируют типичную морфологию эпителия, и вирусы не были обнаружены в линии во время совместного культивирования с клетками WI-38 или в тестах CF против обезьяньего вируса 40 ( SV40 ), респираторно-синцитиального вируса ( RSV ) или аденовирусы. Заражение микоплазмами основных запасов линии U-2 OS было обнаружено и впоследствии ликвидировано в 1972 году.Клетки являются положительными по рецепторам инсулиноподобного фактора роста I ( IGF-I ) и инсулиноподобного фактора роста II ( IGF II ) и экспрессируют ряд антигенов, включая группу крови A, Rh +, HLA A2, Aw30, B12, Bw35 и B40 (+/-).
Клетки глиомы головного мозга человека (U-118 MG) — Клеточная линия U-118 MG является одной из нескольких клеточных линий, полученных из злокачественных глиом Дж.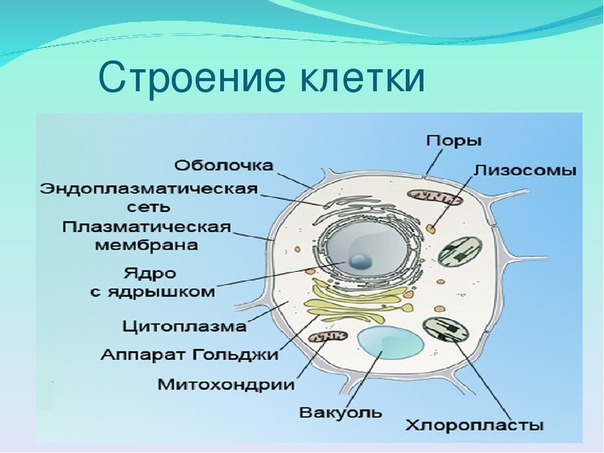 Понтеном и соавторами в конце 1960-х годов. Источником клеток этой конкретной линии был 50-летний мужчина европеоидной расы.Морфология линии U-118 MG смешанная, присутствуют как клетки глиобластомы, так и клетки астроцитомы. Клетки MG U-118 очень похожи на линию клеток MG глиобластомы U-138, хотя предполагается, что эта линия была получена из другого источника. Исследования показали, что обе эти линии имеют идентичные паттерны VNTR (переменное количество тандемных повторов) и похожие паттерны STR (короткие тандемные повторы) в экспериментах по анализу ДНК. У них также есть общие по крайней мере шесть производных маркерных хромосом.Загрязнение микоплазмой было обнаружено и устранено из клеточной линии U-118 MG в 1987 году путем обработки культур BM-циклином в течение шестинедельного периода. Было продемонстрировано, что клетки являются канцерогенными у мышей nude, подкожно инокулированных.
Понтеном и соавторами в конце 1960-х годов. Источником клеток этой конкретной линии был 50-летний мужчина европеоидной расы.Морфология линии U-118 MG смешанная, присутствуют как клетки глиобластомы, так и клетки астроцитомы. Клетки MG U-118 очень похожи на линию клеток MG глиобластомы U-138, хотя предполагается, что эта линия была получена из другого источника. Исследования показали, что обе эти линии имеют идентичные паттерны VNTR (переменное количество тандемных повторов) и похожие паттерны STR (короткие тандемные повторы) в экспериментах по анализу ДНК. У них также есть общие по крайней мере шесть производных маркерных хромосом.Загрязнение микоплазмой было обнаружено и устранено из клеточной линии U-118 MG в 1987 году путем обработки культур BM-циклином в течение шестинедельного периода. Было продемонстрировано, что клетки являются канцерогенными у мышей nude, подкожно инокулированных.
Клетки аденокарциномы шейки матки человека (HeLa) — Линия HeLa является одной из самых известных клеточных линий в мире. Полученные в 1951 году из аденокарциномы шейки матки, обнаруженной у 31-летней женщины (Генриетта Лакс), клетки HeLa были первыми человеческими клетками, которые выжили в лаборатории на неопределенный срок.Клетки демонстрируют морфологию эпителия и прикрепляются к ним, воспроизводя целое поколение примерно каждые 24 часа. Клеточные продукты линии HeLa включают кератин и лизофосфатидилхолин ( lyso-PC ), которые индуцируют активность AP-1 и активность N-концевой киназы c-jun ( JNK1 ) посредством протеинкиназы C-независимого пути. Сообщалось, что клетки HeLa содержат последовательности вируса папилломы человека 18 ( HPV-18, ). Экспрессия P53 в клетках была описана как низкая, хотя уровни pRB (супрессора ретинобластомы), по-видимому, нормальны.
Полученные в 1951 году из аденокарциномы шейки матки, обнаруженной у 31-летней женщины (Генриетта Лакс), клетки HeLa были первыми человеческими клетками, которые выжили в лаборатории на неопределенный срок.Клетки демонстрируют морфологию эпителия и прикрепляются к ним, воспроизводя целое поколение примерно каждые 24 часа. Клеточные продукты линии HeLa включают кератин и лизофосфатидилхолин ( lyso-PC ), которые индуцируют активность AP-1 и активность N-концевой киназы c-jun ( JNK1 ) посредством протеинкиназы C-независимого пути. Сообщалось, что клетки HeLa содержат последовательности вируса папилломы человека 18 ( HPV-18, ). Экспрессия P53 в клетках была описана как низкая, хотя уровни pRB (супрессора ретинобластомы), по-видимому, нормальны.
Кортикальные нейрональные клетки человека (HCN-1A) — Источником линии клеток HCN-1A была кортикальная ткань, удаленная у пациента, перенесшего полушарийэктомию по поводу трудноизлечимых приступов. Пациентка — 18-месячная женщина, страдающая односторонней мегалэнцефалией.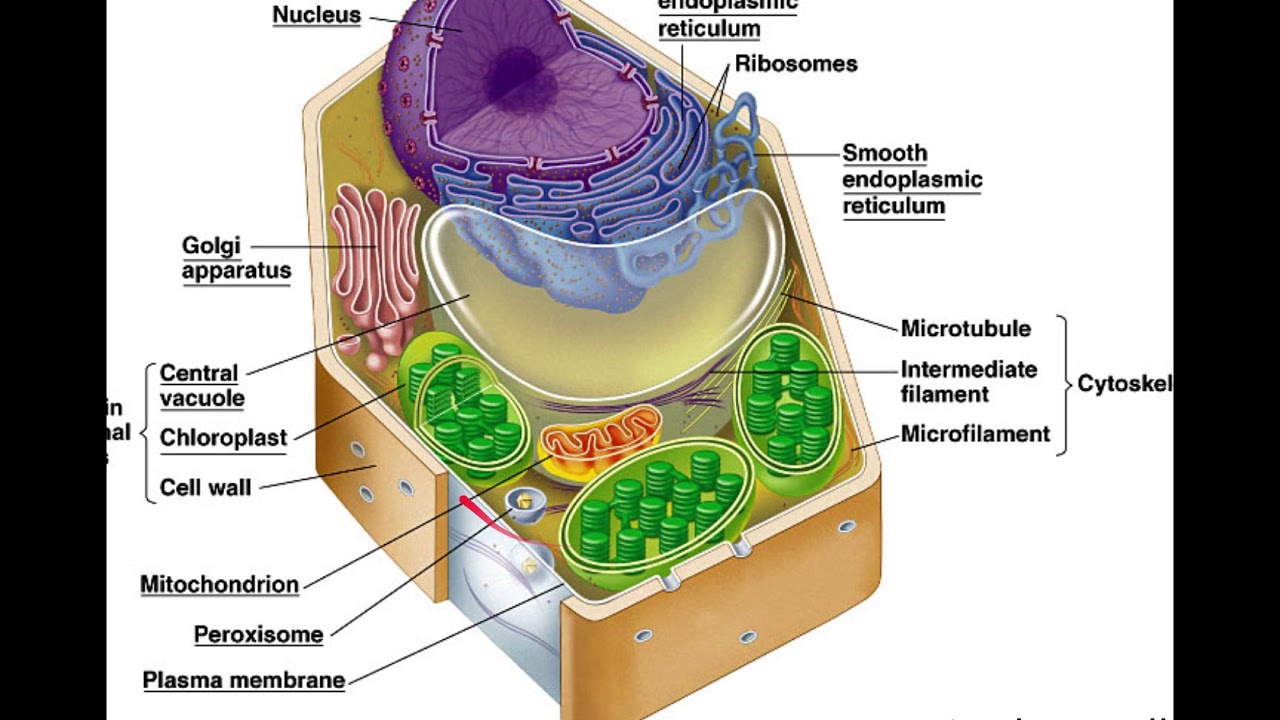 Это состояние, также известное как гемимегаленцефалия, характеризуется разрастанием всего или части одного из полушарий головного мозга. Клетки HCN-1A окрашиваются положительно по ряду нейрональных маркеров, включая нейрон-специфичную энолазу.Они также положительны в отношении тубулина, виментина, соматостатина, глутамата, гамма-аминомасляной кислоты, холецистокинина-8 и вазоактивного кишечного пептида. Однако клетки отрицательны в отношении глиального фибриллярного кислого белка и белка миелиновой основы. Клетки HCN-1A могут быть индуцированы к дифференцировке при культивировании со смесью фактора роста нервов, дибутирилциклического аденозинмонофосфата и 1-изобутил-3-метилксантина. Дифференциация сопровождается зрелой морфологией и замедлением роста.
Это состояние, также известное как гемимегаленцефалия, характеризуется разрастанием всего или части одного из полушарий головного мозга. Клетки HCN-1A окрашиваются положительно по ряду нейрональных маркеров, включая нейрон-специфичную энолазу.Они также положительны в отношении тубулина, виментина, соматостатина, глутамата, гамма-аминомасляной кислоты, холецистокинина-8 и вазоактивного кишечного пептида. Однако клетки отрицательны в отношении глиального фибриллярного кислого белка и белка миелиновой основы. Клетки HCN-1A могут быть индуцированы к дифференцировке при культивировании со смесью фактора роста нервов, дибутирилциклического аденозинмонофосфата и 1-изобутил-3-метилксантина. Дифференциация сопровождается зрелой морфологией и замедлением роста.
Клетки фибробластов легких плода человека (MRC-5) — Линия клеток MRC-5 обычно используется при разработке вакцин, в качестве хозяина для трансфекции в вирусологических исследованиях, а также для тестирования цитотоксичности in vitro . Основанная в сентябре 1966 года Дж. П. Джейкобсом, эта клеточная линия была получена из ткани легких человека 14-недельного плода мужского пола, прерванного от 27-летней женщины. Клетки MRC-5, которые прикрепляются к культуре и демонстрируют морфологию фибробластов, могут удвоиться в размере от 42 до 46 раз до начала старения.Они восприимчивы к полиовирусу 1, простому герпесу и везикулярному стоматиту (штамм Indiana). Однако линия отрицательна по обратной транскриптазе, что указывает на отсутствие целостных геномов ретровирусов.
Основанная в сентябре 1966 года Дж. П. Джейкобсом, эта клеточная линия была получена из ткани легких человека 14-недельного плода мужского пола, прерванного от 27-летней женщины. Клетки MRC-5, которые прикрепляются к культуре и демонстрируют морфологию фибробластов, могут удвоиться в размере от 42 до 46 раз до начала старения.Они восприимчивы к полиовирусу 1, простому герпесу и везикулярному стоматиту (штамм Indiana). Однако линия отрицательна по обратной транскриптазе, что указывает на отсутствие целостных геномов ретровирусов.
Клетки карциномы легкого человека (A-549) — Клеточная линия A-549 была первоначально культивирована в 1972 г. Д. Дж. Джардом вместе с несколькими сотрудниками из карциномы легкого человека 58-летнего мужчины европеоидной расы. Линия обычно используется для исследования широкого спектра респираторных заболеваний, таких как вирусные инфекции, вызывающие астму, повреждение тканей, связанное с воздействием асбеста, и эмфизему, связанную с курением. Адгезивные и эпителиальные клетки A-549 положительны по кератину при окрашивании иммунопероксидазой, но отрицательны по обратной транскриптазе, что указывает на отсутствие целостных геномов ретровирусов. Исследования группы под руководством М. Либера показали, что клетки A-549 способны синтезировать лецитин с высоким процентом ненасыщенных жирных кислот, используя путь цитидиндифосфохолина.
Адгезивные и эпителиальные клетки A-549 положительны по кератину при окрашивании иммунопероксидазой, но отрицательны по обратной транскриптазе, что указывает на отсутствие целостных геномов ретровирусов. Исследования группы под руководством М. Либера показали, что клетки A-549 способны синтезировать лецитин с высоким процентом ненасыщенных жирных кислот, используя путь цитидиндифосфохолина.
Эпителиальные клетки эпидермоидной карциномы кожи человека (A-431) — Линия клеток A-431 , полученная из эпидермоидной карциномы, вырезанной из кожной ткани 85-летней женщины, является одной из последовательных клеточных линий, созданных из солидных опухолей группой исследователей под руководством Д.J. Giard. Эпителиальная линия канцерогенна, образуя быстрорастущие подкожные опухоли у мышей с подавленным иммунитетом и колонии в мягком агаре. Эпидермоидная карцинома кожи, также известная как плоскоклеточная карцинома кожи, представляет собой злокачественную опухоль эпидермальных кератиноцитов, заболевание, частота которого значительно возросла за последнее столетие, возможно, отчасти из-за продолжающегося истощения защитного озонового слоя.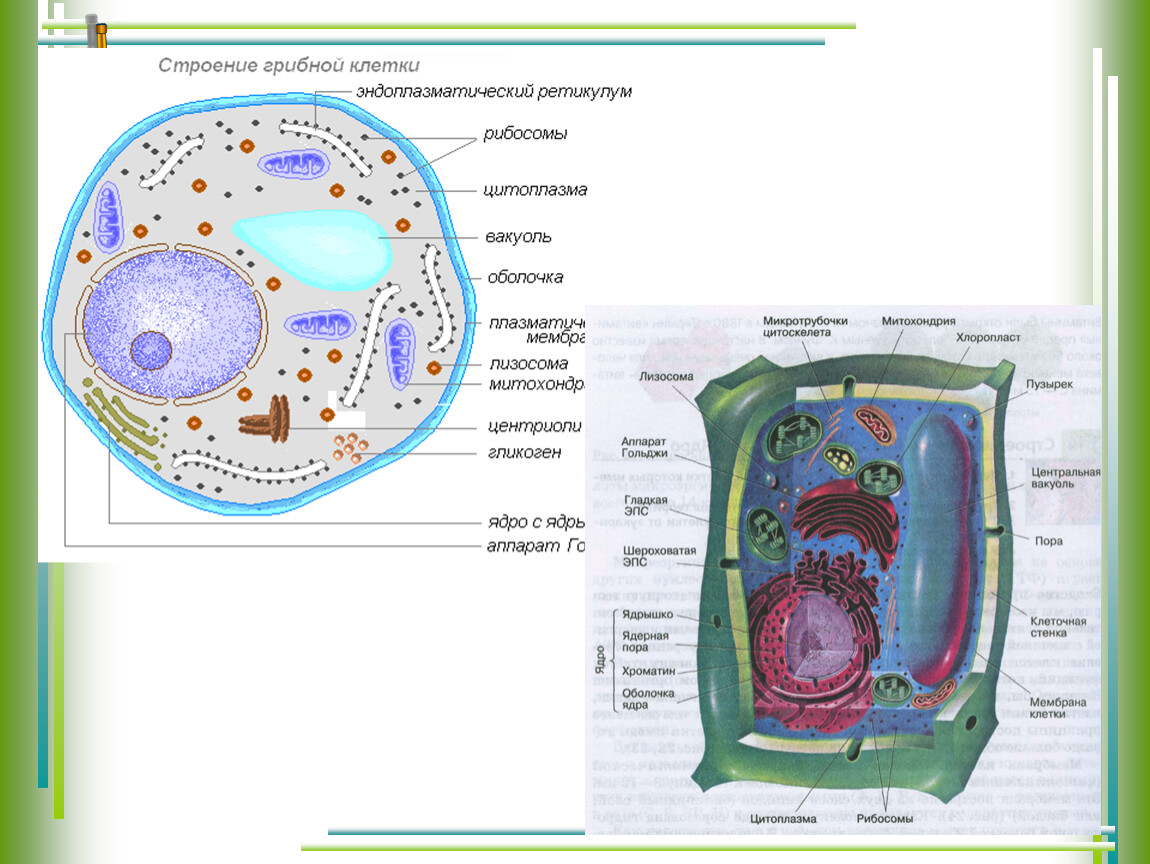
Клетки фибробластов кожи индийского оленя мунтжак — Линия клеток фибробластов, полученная при биопсии кожи взрослого самца, линия эпидермиса индийского оленя мунтжак обычно используется в лабораториях по всему миру, особенно для исследований хромосом.Члены семейства Cervidae , Muntjacs — лают олени, которые издают свой характерный звук, когда чувствуют угрозу или тревогу. Нормальная (нетрансформированная) линия клеток Indian Muntjac чувствительна к вирусу простого герпеса, вирусу осповакцины и вирусу везикулярного стоматита (штамм Indiana), но устойчива к полиовирусу 1. Недавние тесты показали, что клетки продуцируют оба обнаруживаемых вируса крупного рогатого скота. антигены вируса диареи ( BVDV ) и инфекционные вирионы BVDV.Клетки Muntjac отрицательны по обратной транскриптазе, что указывает на отсутствие целостных геномов ретровирусов.
Madin-Darby Canine Kidney Epithelial Cells (MDCK) — Полученная SH Madin и NB Darby из ткани почек взрослой самки кокер-спаниеля, клеточная линия MDCK возникла в сентябре 1958 года. широко использовался для исследования процессинга белка-предшественника амилоида beta , а также сортировки его протеолитических продуктов.Морфология клеточной линии MDCK является эпителиальной, и клетки положительны на кератин по окрашиванию иммунопероксидазой. К вирусам, которым подвержены клетки MDCK, относятся везикулярный стоматит (штамм Indiana), коровьей оспы, вирус Коксаки B5, реовирус 2 и 3, аденовирус 4 и 5, везикулярная экзантема свиней и инфекционный гепатит собак. Клетки проявляют устойчивость к вирусу Коксаки B3 и B4, а также к полиовирусу 2, и отрицательны в отношении обратной транскриптазы.
широко использовался для исследования процессинга белка-предшественника амилоида beta , а также сортировки его протеолитических продуктов.Морфология клеточной линии MDCK является эпителиальной, и клетки положительны на кератин по окрашиванию иммунопероксидазой. К вирусам, которым подвержены клетки MDCK, относятся везикулярный стоматит (штамм Indiana), коровьей оспы, вирус Коксаки B5, реовирус 2 и 3, аденовирус 4 и 5, везикулярная экзантема свиней и инфекционный гепатит собак. Клетки проявляют устойчивость к вирусу Коксаки B3 и B4, а также к полиовирусу 2, и отрицательны в отношении обратной транскриптазы.
Madin-Darby Ovine Kidney Epithelial Cells (MDOK) — Клеточная линия Madin-Darby овечьих почек ( MDOK ) была получена из почечной ткани самца овцы.Эта линия демонстрирует типичную морфологию эпителия и чувствительна к нескольким вирусам, включая везикулярный стоматит (штаммы Индианы и Нью-Джерси), инфекционный ринотрахеит крупного рогатого скота и вирус блютанга овец. Эпителиальные клетки обычно существуют в теле в виде листов, покрывающих органы и другие внутренние и внешние поверхности, которые могут контактировать с инородными материалами. Обычно клетки содержат относительно большое количество цитоплазмы и значительное количество гранул. Функции клеток различны, некоторые из них выполняют абсорбционную или защитную роль, а другие в основном действуют как секреторные клетки.
Эпителиальные клетки обычно существуют в теле в виде листов, покрывающих органы и другие внутренние и внешние поверхности, которые могут контактировать с инородными материалами. Обычно клетки содержат относительно большое количество цитоплазмы и значительное количество гранул. Функции клеток различны, некоторые из них выполняют абсорбционную или защитную роль, а другие в основном действуют как секреторные клетки.
Эпителиальные клетки почек кенгуру самцов крыс (PtK2) — Широко используемая линия клеток PtK2 была получена из ткани почек взрослого самца кенгуру крысы ( Potorous tridactylus ), длинноносого сумчатого животного довольно распространен в Австралии. Эпителиальная линия положительна для жесткого, нерастворимого белка промежуточного филамента кератина при окрашивании иммунопероксидазой, но отрицательна для обратной транскриптазы, что указывает на отсутствие целостных геномов ретровирусов.Клетки PtK2 чувствительны к вирусу Коксаки А9, вирусу простого герпеса, осповакцине и везикулярному стоматиту (штамм Огдена).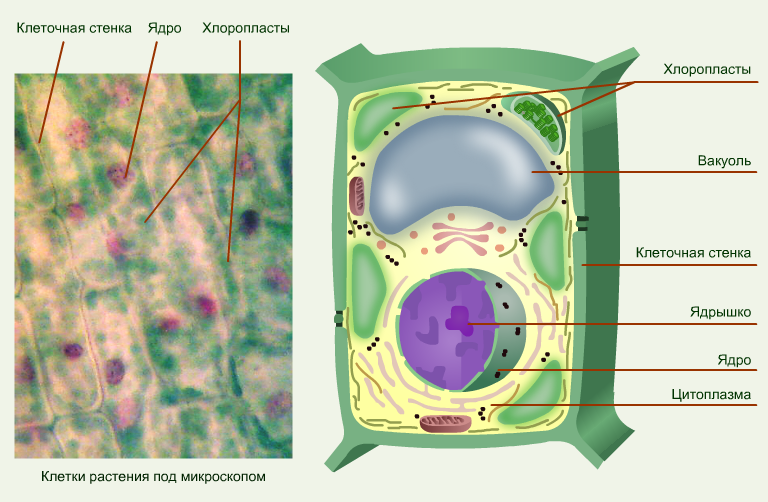 Однако эти клетки устойчивы к аденовирусу 5, вирусу Коксаки B5 и полиовирусу 2. Линия эпителиальных клеток PtK2 используется для множества применений, но в первую очередь для исследований в области митоза.
Однако эти клетки устойчивы к аденовирусу 5, вирусу Коксаки B5 и полиовирусу 2. Линия эпителиальных клеток PtK2 используется для множества применений, но в первую очередь для исследований в области митоза.
Эпителиальные клетки эндометрия матки норки (GMMe) — Клеточная линия GMMe была создана путем стабильной трансфекции ткани эндометрия взрослой самки норки ( Mustela vison ) с использованием плазмидного вектора, кодирующего обезьяний вирус 40 ( SV40 ) большой Т-антиген, управляемый промотором бета -актина человека.Клетки котрансфицировали вторым плазмидным вектором для придания устойчивости к неомицину и отбирали в среде, содержащей G418. Кубовидные клетки GMMe, которые проявляют эпителиальные характеристики, сильно положительны в отношении цитокератина и слабо положительны в отношении виментина. Линия также положительна для щелочной фосфатазы, но отрицательна для десмина. Клетки GMME использовались в совместном культивировании с эмбрионами норки в облигатной диапаузе для увеличения длины и частоты выживаемости эмбрионов in vitro , хотя стромальная линия норки GMMs использовалась таким же образом с более высокой степенью выживаемости. скорость успеха.
скорость успеха.
Эпителиальные клетки тератокарциномы мышиных эмбрионов (P19) — Линия клеток P19 была получена из эмбриональной карциномы, индуцированной у линии мышей C3H / He вида Mus musculus . Подобно другим клеткам эмбриональной карциномы, клетки линии P19 способны дифференцироваться в множество типов клеток и поэтому описаны как плюрипотентные. Хотя они могут поддерживаться и размножаться в тканевой культуре в недифференцированном состоянии, при воздействии 500 нМ ретиноевой кислоты клетки P19 могут быть индуцированы к дифференцировке в нейральные и глиальные клетки.Однако в присутствии от 0,5 до 1,0 процента диметилсульфоксида ( DMSO ) клетки дифференцируются с образованием элементов, подобных сердечным и скелетным мышцам, но не образуют нервные или глиальные клетки. При одновременном введении и ДМСО, и ретиноевой кислоты клетки P19 дифференцируются, как если бы они подвергались воздействию только ретиноевой кислоты.
Эндотелиальные клетки гемангиоэндотелиомы мыши (EOMA) — Линия клеток EOMA была получена в 1980 году из смешанной гемангиоэндотелиомы, присутствующей у взрослой мыши ( Mus musculus ). Гемангиоэндотелиома — это термин, используемый для описания разнообразной группы сосудистых опухолей, которые обычно выглядят как красные или синие узелки и имеют тенденцию биологически вести себя таким образом, чтобы их можно было классифицировать как попадающие между доброкачественной гемангиомой и злокачественной ангиосаркомой. Клетки EOMA являются онкогенными у сингенных мышей и проявляют характерные свойства эндотелиальных клеток. Клетки синтезируют ряд клеточных продуктов, включая ангиотензин-превращающий фермент ( ACE ), тромбоспондин, катепсин L, эндостатин и интерлейкин-6.Поверхностные рецепторы для ацетилированного липопротеина низкой плотности экспрессируются клетками EOMA, как и сосудистый адресин, тканеспецифическая молекула адгезии эндотелиальных клеток, идентифицированная антителом MECA-99.
Гемангиоэндотелиома — это термин, используемый для описания разнообразной группы сосудистых опухолей, которые обычно выглядят как красные или синие узелки и имеют тенденцию биологически вести себя таким образом, чтобы их можно было классифицировать как попадающие между доброкачественной гемангиомой и злокачественной ангиосаркомой. Клетки EOMA являются онкогенными у сингенных мышей и проявляют характерные свойства эндотелиальных клеток. Клетки синтезируют ряд клеточных продуктов, включая ангиотензин-превращающий фермент ( ACE ), тромбоспондин, катепсин L, эндостатин и интерлейкин-6.Поверхностные рецепторы для ацетилированного липопротеина низкой плотности экспрессируются клетками EOMA, как и сосудистый адресин, тканеспецифическая молекула адгезии эндотелиальных клеток, идентифицированная антителом MECA-99.
Нормальные эпителиальные клетки почек крысы (NRK) — Линия клеток NRK была получена из почек крысы вида Rattus norvegicus . Клетки срослись и демонстрируют морфологию эпителия. Эпителиальные клетки — это клетки, которые существуют в теле в виде листов, покрывающих органы и другие внутренние и внешние поверхности, которые могут контактировать с инородными материалами.Обычно эпителиальные клетки содержат относительно большое количество цитоплазмы и значительное количество гранул. Функции клеток различны, некоторые из них выполняют абсорбционную или защитную роль, а другие в основном действуют как секреторные клетки. Эпителиальные клетки почек в основном относятся к этой последней категории, играя важную роль в хранении и последующей секреции различных экскреторных материалов.
Эпителиальные клетки — это клетки, которые существуют в теле в виде листов, покрывающих органы и другие внутренние и внешние поверхности, которые могут контактировать с инородными материалами.Обычно эпителиальные клетки содержат относительно большое количество цитоплазмы и значительное количество гранул. Функции клеток различны, некоторые из них выполняют абсорбционную или защитную роль, а другие в основном действуют как секреторные клетки. Эпителиальные клетки почек в основном относятся к этой последней категории, играя важную роль в хранении и последующей секреции различных экскреторных материалов.
Эпителиальные клетки почек свиней (LLC-PK1) — Клеточная линия LLC-PK1 была получена из почек нормального здорового самца свиньи ( Sus scrofa ) возрастом от 3 до 4 недель. .Свинья была представителем породы Хэмпшир, которая первоначально была импортирована в Америку из графства Хэмпшир, Англия, в начале 1800-х годов. Линия LLC-PK1, обычно используемая в лабораториях по всему миру, демонстрирует типичную морфологию эпителия. Клеточные продукты линии почек свиньи LLC-PK1 включают активатор плазминогена, вещество, стимулирующее фибринолиз. В последние годы активатор плазминогена был включен в препараты, используемые при лечении тромбозов, поскольку он способствует растворению небольших сгустков крови.Клетки также производят большое количество цитокератина. Линия LLC-PK1 часто используется в качестве модели эпителиальной ткани, а также в широком спектре фармакологических и метаболических исследований.
Клеточные продукты линии почек свиньи LLC-PK1 включают активатор плазминогена, вещество, стимулирующее фибринолиз. В последние годы активатор плазминогена был включен в препараты, используемые при лечении тромбозов, поскольку он способствует растворению небольших сгустков крови.Клетки также производят большое количество цитокератина. Линия LLC-PK1 часто используется в качестве модели эпителиальной ткани, а также в широком спектре фармакологических и метаболических исследований.
Энтероглиальные клетки тонкой кишки и кишечного сплетения крысы (EGC / PK060399egfr) — Энтероглиальные клетки (известные под аббревиатурой: EGC ) считаются важной частью кишечной нервной системы, но функции этих клеток еще не решены. неизвестный.Чтобы облегчить будущие исследования клеток, А. Рул и его коллеги разработали в 2001 году новый метод выделения и очистки энтероглиальных клеток из миэнтериального сплетения, сети нервных волокон, расположенных в стенке кишечника. Метод Руля включал ферментативную диссоциацию образцов мышечно-кишечного сплетения, очистку кишечных глиальных клеток посредством комплемент-опосредованного цитолиза контаминирующих клеток и трансформацию путем переноса ретровирусного гена. Затем группа охарактеризовала полученные клоны как иммуногистохимически, так и с помощью дот-блот-анализа.В результате их усилий был установлен ряд трансформированных линий EGC, которые сохраняют свои глиальные и функциональные характеристики.
Затем группа охарактеризовала полученные клоны как иммуногистохимически, так и с помощью дот-блот-анализа.В результате их усилий был установлен ряд трансформированных линий EGC, которые сохраняют свои глиальные и функциональные характеристики.
Мезангиальные клетки почек крысы (RMC) — Линия клеток RMC была получена из ткани почек трехмесячного самца крысы ( Rattus norvegicus ), принадлежащего штамму Sprague-Dawley. Мезангиальные клетки были иммортализованы на восьмом пассаже плазмидой pSV3-Neo, которая кодирует большой Т-антиген обезьяньего вируса 40 ( SV40 ), и поддерживаются в присутствии антибиотика G-418.Клетки почек крысы экспрессируют нормальные гены мезангиальных клеток дикого типа и прилипают к стеклянным и полимерным поверхностям в монослойной культуре. Клетки RMC положительны для десмина и виментина, но отрицательны для цитокератина 8. Мезангиальные клетки — это специализированные клетки, обычно связанные с клубочками, которые имеют решающее значение для функции почек. Таким образом, линия RMC и другие линии мезангиальных клеток широко используются в исследованиях почек.
Таким образом, линия RMC и другие линии мезангиальных клеток широко используются в исследованиях почек.
Эпителиальные клетки яичников Tahr (HJ1.Ov) — Клеточная линия HJ1.Ov была получена из ткани яичника нормальной и здоровой самки гималайского тара ( Hemitragus jemlahicus ), которая является родственником дикой козы, специально адаптированной к жизни в суровых условиях. , горная среда Гималаев. Разработанные в The Naval Biosciences Laboratory ( NBL ) в Окленде, Калифорния, непрерывные культуры клеток HJ1.Ov демонстрируют типичную морфологию эпителия и прилипают к стеклянным и полимерным поверхностям в монослойной культуре.Эпителиальные клетки включают бессосудистые ткани, выстилающие как внутренние, так и внешние поверхности тела и его органов. Также секреторные части желез и их протоки образованы инвагинированными эпителиальными клетками. Хотя в организме существует множество различных типов эпителиальных клеток, которые могут быть расположены по-разному, клетки всегда прилегают друг к другу, так что они создают непрерывный барьер.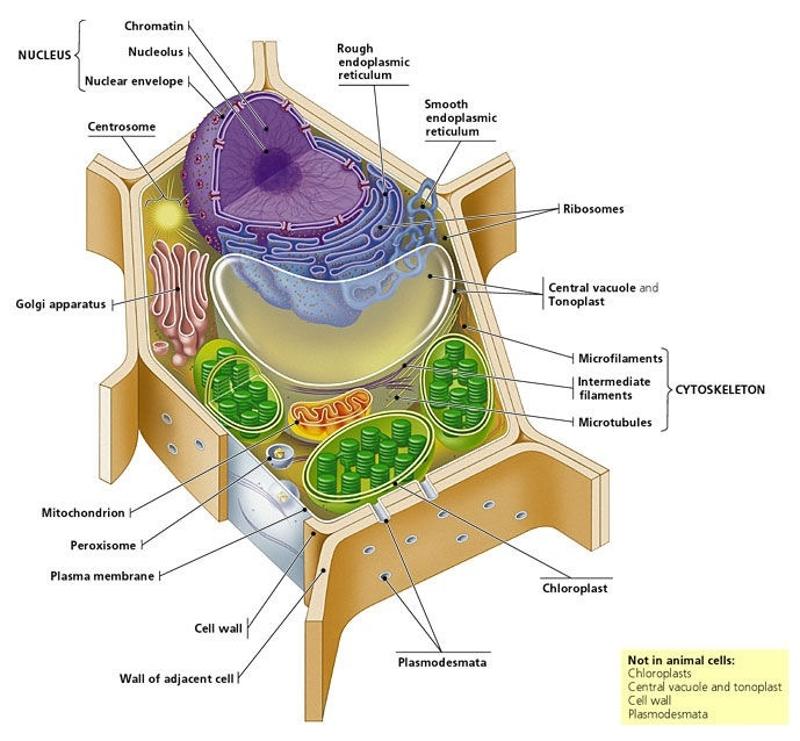
Трансформированные клетки фибробластов куриных эмбрионов (UMNSAH / DF-1) — UMNSAH / DF-1 — это спонтанно иммортализованная линия клеток курицы ( Gallus gallus ), полученная из 10-дневного штамма (9000 East Lansing). ELL-0 ) яйца.Для создания линии первичные куриные эмбриональные фибробласты диссоциировали и выращивали в культуре. Фибробласты пассировали до тех пор, пока они не начали стареть. Линия фибробластов куриных эмбрионов UMNSAH / DF-1 полезна в качестве субстрата для размножения вируса, экспрессии рекомбинантного белка и продукции рекомбинантного вируса. Линия чувствительна к ряду вирусов, включая вирус герпеса 1 Meleagrid, вирус оспы птиц, реовирус, вирус лейкемии саркомы птиц и вирус саркомы Рауса. Однако эти клетки не являются канцерогенными у мышей с подавленным иммунитетом, но действительно образуют колонии в полутвердой среде.Клетки UMNSAH / DF-1 отрицательны по обратной транскриптазе, что указывает на отсутствие целостных геномов ретровирусов.
Трансформированные микроглиальные клетки мозжечка мыши (C8-B4) — Микроглия — это специализированные макрофаги , которые очень важны для их роли в защите центральной нервной системы. C8-B4 представляет собой спонтанно трансформированный клон микроглии линии клеток, первоначально полученной из мозжечка 8-дневной мыши ( Mus musculus ) в 1984 году.Эту исходную культуру органов использовали для создания нескольких различных линий клеток астроглии. Клон C8-B4 был создан в 1996 году, и нейронные клетки прочно растут в культуре. Классические микроглиальные маркеры, включая MAC1, F4 / 80 и 2-4G2, экспрессируются клоном C8-B4, который, по-видимому, происходит из предшественника микроглии, поскольку он, как сообщается, не экспрессирует антигены дифференцировки, присутствующие на ранней стадии моноцитарной происхождение. Клетки C8-B4 производят и выделяют большое количество глутамата, вещества, которое обычно функционирует как нейромедиатор.
Соавторы
Джон Д. Гриффин , Натан С. Клэкстон , Джон Д. Хоман , Афердита Ишмаку , Уильям С. Фэрхерст , Ричард Л. Ладлоу , Синди Монкриф , Б. Boles , Lionel Parsons, Jr. , Ryan R. Spindell , Shannon H. Neaves и Michael W. Davidson — Национальная лаборатория сильных магнитных полей, 1800 East Paul Dirac Dr., Государственный университет Флориды, Таллахасси, Флорида, 32310.
Гриффин , Натан С. Клэкстон , Джон Д. Хоман , Афердита Ишмаку , Уильям С. Фэрхерст , Ричард Л. Ладлоу , Синди Монкриф , Б. Boles , Lionel Parsons, Jr. , Ryan R. Spindell , Shannon H. Neaves и Michael W. Davidson — Национальная лаборатория сильных магнитных полей, 1800 East Paul Dirac Dr., Государственный университет Флориды, Таллахасси, Флорида, 32310.
НАЗАД В ГАЛЕРЕЮ ФЛУОРЕСЦЕНЦИИ
Вопросы или комментарии? Отправить нам письмо.
© 1998-2021, автор — Майкл В. Дэвидсон и Государственный университет Флориды. Все права защищены. Никакие изображения, графика, сценарии или апплеты не могут быть воспроизведены или использованы каким-либо образом без разрешения правообладателей. Использование этого веб-сайта означает, что вы соглашаетесь со всеми юридическими положениями и условиями, изложенными владельцами.
Этот веб-сайт поддерживается командой
Graphics & Web Programming Team
.
 в сотрудничестве с оптической микроскопией в Национальной лаборатории сильного магнитного поля
в сотрудничестве с оптической микроскопией в Национальной лаборатории сильного магнитного поля .
Последнее изменение: пятница, 13 ноября 2015 г., 13:19
Счетчик доступа с 16 июля 2004 г .: 264201
Микроскопы, флуоресцентные фильтры и оборудование для цифровой обработки изображений, предоставленные:
Молекулярные выражения Биология клетки: Структура клеток животных
центриолей
Обнаруженные только в клетках животных, эти парные органеллы обычно расположены вместе около ядра в центросоме , гранулированной массе, которая служит организующим центром для микротрубочек.Внутри центросомы центриоли расположены под прямым углом друг к другу, как показано на рисунке 1. Каждая центриоль состоит из девяти пучков микротрубочек (по три на пучок), расположенных в кольцо.
Центриоли играют заметную роль в делении клеток. Во время интерфазы животной клетки центриоли и другие компоненты центросомы дублируются, хотя ученые еще не уверены, как это дублирование происходит. Сначала две пары центриолей остаются в непосредственной близости друг от друга, но по мере инициации митоза исходная центросома делится, и пары разделяются, так что один набор центриолей располагается в каждом из новых центров организации микротрубочек.Эти новые центры излучают микротрубочки в звездных кластерах, известных как asters . Когда звездочки перемещаются к противоположным полюсам клеток, микротрубочки с помощью центриолей организуются в веретенообразное образование, охватывающее клетку (см. Рис. 2). Эти волокна веретена действуют как направляющие для выравнивания хромосом, поскольку они отделяются позже в процессе деления клеток.
Хотя центриоли играют роль в митозе клеток животных, клетки растений могут воспроизводиться без них.Поэтому исследователи были очень заинтересованы в том, чтобы точно определить, насколько действительно важны органеллы. Исследования показали, что определенные клетки животных, особенно женские гаметы (ооциты), могут успешно делиться, даже если их центриоли разрушены. Однако некоторые исследователи также обнаружили, что отсутствие центриолей в клетках животных связано с повышенным числом ошибок деления и существенными задержками митотического процесса, особенно перед сегрегацией хромосом.Следовательно, было высказано предположение, что центриоли эволюционировали как уточнение клетки, что сделало митоз гораздо более эффективным и менее подверженным ошибкам процессом.
Однако некоторые исследователи также обнаружили, что отсутствие центриолей в клетках животных связано с повышенным числом ошибок деления и существенными задержками митотического процесса, особенно перед сегрегацией хромосом.Следовательно, было высказано предположение, что центриоли эволюционировали как уточнение клетки, что сделало митоз гораздо более эффективным и менее подверженным ошибкам процессом.
В клетках, которые имеют реснички или жгутики, присутствует базальных тел , которые имеют ту же структурную форму, что и центриоли. Эти сборки расположены, однако, около поверхности клетки у основания каждой реснички или жгутика, а не в центросоме рядом с ядром. Базальные тельца закреплены в своих цитоплазматических местах с помощью того, что называется системой корней в клетке.У некоторых организмов, таких как одноклеточные Chlamydomonas , базальные тельца меняют свое положение и функционально преобразуются в центриоли перед митотическим процессом.
НАЗАД К СТРУКТУРЕ ЖИВОТНЫХ
Вопросы или комментарии? Отправить нам письмо.
© 1995-2021, автор — Майкл В. Дэвидсон и Государственный университет Флориды. Все права защищены. Никакие изображения, графика, программное обеспечение, сценарии или апплеты не могут быть воспроизведены или использованы каким-либо образом без разрешения правообладателей.Использование этого веб-сайта означает, что вы соглашаетесь со всеми юридическими положениями и условиями, изложенными владельцами.
Этот веб-сайт поддерживается нашим
Команда разработчиков графики и веб-программирования
в сотрудничестве с оптической микроскопией в Национальной лаборатории сильного магнитного поля
.
Последнее изменение: пятница, 13 ноября 2015 г., 13:18
Счетчик доступа с 1 октября 2000 г .: 532473
Микроскопы предоставлены:
границ | Эволюция и разнообразие клеточной стенки
Введение
Стенки растительных клеток представляют собой многофункциональные волокнистые композиты, богатые полисахаридами, в которых полимеры взаимодействуют с образованием несущих структур, встроенных в полисахаридную матрицу (Bacic et al. , 1988; Фрай, 2004). Клетки в растущих частях растений связаны «первичными стенками», в которых несущую функцию выполняют в основном микрофибриллы целлюлозы. Модели клеточной стенки растений обычно изображают микрофибриллы, поперечно сшитые гемицеллюлозами, включая маннаны, ксиланы, глюканы со смешанными связями (MLG) и ксилоглюканы. Затем эта сеть встраивается в матрицу пектиновых полисахаридов, включая гомогалактуронан (HG), рамногалактуронан-I (RG-I), рамногалактуронан-II (RG-II) и ксилогалактуронан (Fry, 2004; Mohnen, 2008; Caffall and Mohnen). , 2009; Harholt et al., 2010). Однако это обычное описание первичных стенок, которое подчеркивает, что гликаны связывания являются незаменимыми «несущими» конструкциями, возможно, потребует пересмотра, как это обсуждается в Scheller and Ulvskov (2010). Первичные клеточные стенки создают основу для формы клеток и противостоят растягивающим силам, оказываемым тургорным давлением. Они также должны быть способны к контролируемой экспансии для обеспечения роста клеток.
, 1988; Фрай, 2004). Клетки в растущих частях растений связаны «первичными стенками», в которых несущую функцию выполняют в основном микрофибриллы целлюлозы. Модели клеточной стенки растений обычно изображают микрофибриллы, поперечно сшитые гемицеллюлозами, включая маннаны, ксиланы, глюканы со смешанными связями (MLG) и ксилоглюканы. Затем эта сеть встраивается в матрицу пектиновых полисахаридов, включая гомогалактуронан (HG), рамногалактуронан-I (RG-I), рамногалактуронан-II (RG-II) и ксилогалактуронан (Fry, 2004; Mohnen, 2008; Caffall and Mohnen). , 2009; Harholt et al., 2010). Однако это обычное описание первичных стенок, которое подчеркивает, что гликаны связывания являются незаменимыми «несущими» конструкциями, возможно, потребует пересмотра, как это обсуждается в Scheller and Ulvskov (2010). Первичные клеточные стенки создают основу для формы клеток и противостоят растягивающим силам, оказываемым тургорным давлением. Они также должны быть способны к контролируемой экспансии для обеспечения роста клеток. В тканях нерастущих растений некоторые клетки обычно окружены «вторичными стенками», основная роль которых заключается в сопротивлении силе сжатия, и, поскольку расширение клеток не требуется, эти стенки часто укрепляются лигнином (Hepler et al., 1970; Карпита и Жибо, 1993; Boerjan et al., 2003; Косгроув, 2005). Хотя эти описания служат для описания многих клеточных стенок растений в общих чертах, они являются обобщениями и в основном основаны на исследованиях клеточных стенок цветковых растений. Однако клеточные стенки демонстрируют удивительное разнообразие на многих уровнях, а составляющие их полисахариды различаются по тонкой структуре, относительному содержанию и молекулярным ассоциациям (Burton et al., 2010). Огромная сложность и неоднородность гликомов клеточной стенки является продуктом совместной деятельности огромного количества биосинтетических ферментов.Из секвенирования генома ясно, что сотни гликозилтрансфераз (GT) катализируют образование гликозидных связей в полисахаридах — более 50 для одних только пектиновых полимеров (Scheible and Pauly, 2004; Mohnen, 2008; Yin et al.
В тканях нерастущих растений некоторые клетки обычно окружены «вторичными стенками», основная роль которых заключается в сопротивлении силе сжатия, и, поскольку расширение клеток не требуется, эти стенки часто укрепляются лигнином (Hepler et al., 1970; Карпита и Жибо, 1993; Boerjan et al., 2003; Косгроув, 2005). Хотя эти описания служат для описания многих клеточных стенок растений в общих чертах, они являются обобщениями и в основном основаны на исследованиях клеточных стенок цветковых растений. Однако клеточные стенки демонстрируют удивительное разнообразие на многих уровнях, а составляющие их полисахариды различаются по тонкой структуре, относительному содержанию и молекулярным ассоциациям (Burton et al., 2010). Огромная сложность и неоднородность гликомов клеточной стенки является продуктом совместной деятельности огромного количества биосинтетических ферментов.Из секвенирования генома ясно, что сотни гликозилтрансфераз (GT) катализируют образование гликозидных связей в полисахаридах — более 50 для одних только пектиновых полимеров (Scheible and Pauly, 2004; Mohnen, 2008; Yin et al. , 2010; Dhugga , 2012). Большинство GT действуют в аппарате Гольджи, и их продукты транспортируются к клеточным стенкам в секреторных пузырьках. Напротив, целлюлозо- и каллозосинтазы и, возможно, класс «D» целлюлозосинтазоподобных GT встраиваются в плазматическую мембрану, и их продукты экструдируются непосредственно в клеточные стенки (Endler and Persson, 2011; Park et al., 2011). Большое количество генов, кодирующих GT, и их различные временные и пространственные профили экспрессии создают огромные возможности для изменчивости клеточной стенки. Дальнейшая гетерогенность обусловлена доступностью широкого диапазона доноров активированных сахаров (Feingold and Avigad, 1980), метилированием и ацетилированием, различными энантиомерами и разнообразием и количеством возможных гликозильных связей, а также полисахаридов в модификации muro , например путем этерификации / деэтерификации пектинов и трансгликозилирования между некоторыми гемицеллюлозами (Fry et al., 2008; Бертон и др., 2010).
, 2010; Dhugga , 2012). Большинство GT действуют в аппарате Гольджи, и их продукты транспортируются к клеточным стенкам в секреторных пузырьках. Напротив, целлюлозо- и каллозосинтазы и, возможно, класс «D» целлюлозосинтазоподобных GT встраиваются в плазматическую мембрану, и их продукты экструдируются непосредственно в клеточные стенки (Endler and Persson, 2011; Park et al., 2011). Большое количество генов, кодирующих GT, и их различные временные и пространственные профили экспрессии создают огромные возможности для изменчивости клеточной стенки. Дальнейшая гетерогенность обусловлена доступностью широкого диапазона доноров активированных сахаров (Feingold and Avigad, 1980), метилированием и ацетилированием, различными энантиомерами и разнообразием и количеством возможных гликозильных связей, а также полисахаридов в модификации muro , например путем этерификации / деэтерификации пектинов и трансгликозилирования между некоторыми гемицеллюлозами (Fry et al., 2008; Бертон и др., 2010). В совокупности эти динамические процессы позволяют растениям генерировать клеточные стенки, которые идеально подходят для преобладающих функциональных требований и могут реагировать на биотические и абиотические стрессы, а также на сигналы развития (Sarkar et al., 2009; Sørensen et al., 2010).
В совокупности эти динамические процессы позволяют растениям генерировать клеточные стенки, которые идеально подходят для преобладающих функциональных требований и могут реагировать на биотические и абиотические стрессы, а также на сигналы развития (Sarkar et al., 2009; Sørensen et al., 2010).
Зачем изучать разнообразие клеточной стенки?
Изучение гликомов клеточной стенки в царстве растений важно для развития нашего понимания структур и функций клеточной стенки, для понимания клеточной стенки и эволюции растений, а также для оптимизации использования крупнейшего источника биомассы на Земле.Растения появились на суше около 470 миллионов лет назад и с тех пор колонизировали значительную часть поверхности Земли (Kenrick and Crane, 1997; Waters, 2003; Niklas and Kutschera, 2010). Переход на сушу стал поворотным событием в истории жизни, которое привело к формированию новых мест обитания и экосистем и оказало глубокое влияние на химию атмосферы. Клеточные стенки сыграли значительную роль в этих эпохальных эволюционных событиях, но наше текущее понимание многих аспектов структур клеточных стенок и их эволюции ограничено (Niklas, 2004; Popper and Tuohy, 2010; Sørensen et al. , 2010). Улучшение нашего понимания будет способствовать более широкому пониманию эволюции растений и филогенетических отношений и может предоставить знания о прошлой окружающей среде и понимание того, как растения могут реагировать на предсказанные сценарии изменения климата.
, 2010). Улучшение нашего понимания будет способствовать более широкому пониманию эволюции растений и филогенетических отношений и может предоставить знания о прошлой окружающей среде и понимание того, как растения могут реагировать на предсказанные сценарии изменения климата.
Изучение эволюции клеточной стенки в основном основано на изучении разнообразия клеточной стенки (Popper, 2008; Sørensen et al., 2010; Popper et al., 2011). Только так можно сопоставить изменения в физиологии, морфологии или привычках растений с соответствующими нововведениями в биологии клеточной стенки.Изучение разнообразия клеточных стенок в царстве растений также имеет и другие преимущества. Полисахариды клеточной стенки являются чрезвычайно ценным возобновляемым биоресурсом и находят множество промышленных применений. Древесина, волокна, бумага, функциональные ингредиенты (например, пектины цветущих растений и альгинаты и каррагинаны из водорослей), а также нутрицевтики и биотопливо первого и второго поколения в основном основаны на клеточных стенках (Bacic et al.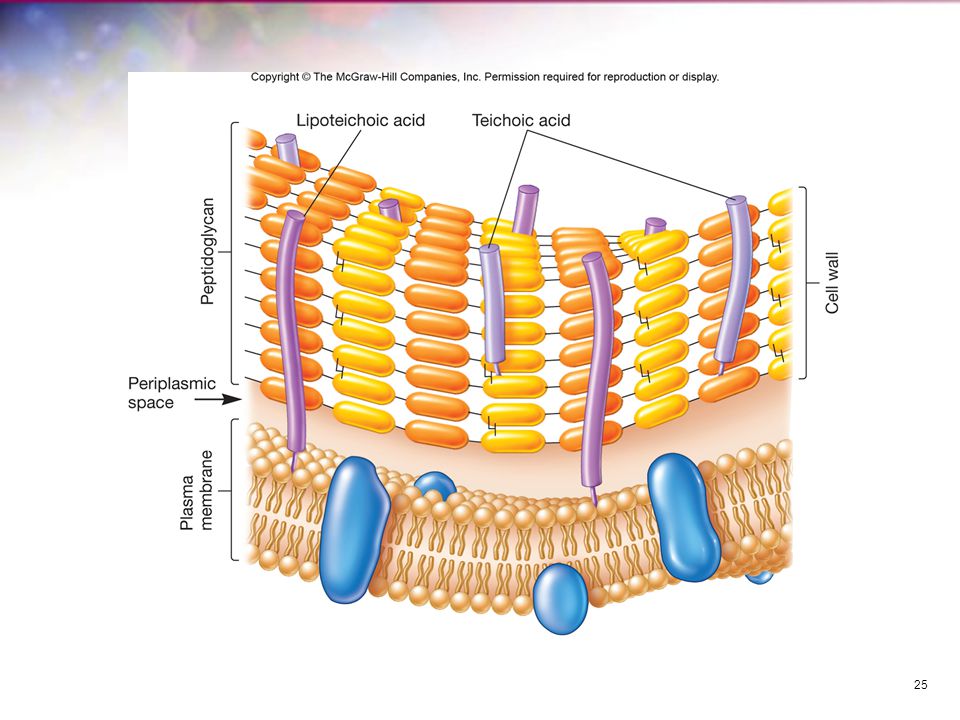 , 1988; Willats et al. , 2006; Pauly, Keegstra, 2010). В отличие от нуклеотидных последовательностей и белков, полисахариды не могут быть легко синтезированы, поэтому их необходимо получать от природы.В настоящее время мы используем только незначительную долю глобального гликома клеточной стенки, и полный перечень доступных полисахаридов может выявить новые ценные молекулы и материалы с новым применением. Анализ разнообразных составов и архитектур клеточных стенок также может послужить источником вдохновения для текущих усилий, направленных на целевую модификацию клеточных стенок, особенно для энергетических культур. Однако исследование клеточных стенок всего царства растений — сложная задача, которая, как описано ниже, влечет за собой множество серьезных проблем и требует междисциплинарного подхода.Это в значительной степени связано с тем, что полисахариды не кодируются напрямую последовательностью генома; Для синтеза активированных сахарных остатков требуется множество ферментов, связи и многие компоненты стенки претерпевают обширные модификации, включая метилирование, этерификацию / деэтерификацию и ацетилирование, а также добавление одного или блоков остатков сахара.
, 1988; Willats et al. , 2006; Pauly, Keegstra, 2010). В отличие от нуклеотидных последовательностей и белков, полисахариды не могут быть легко синтезированы, поэтому их необходимо получать от природы.В настоящее время мы используем только незначительную долю глобального гликома клеточной стенки, и полный перечень доступных полисахаридов может выявить новые ценные молекулы и материалы с новым применением. Анализ разнообразных составов и архитектур клеточных стенок также может послужить источником вдохновения для текущих усилий, направленных на целевую модификацию клеточных стенок, особенно для энергетических культур. Однако исследование клеточных стенок всего царства растений — сложная задача, которая, как описано ниже, влечет за собой множество серьезных проблем и требует междисциплинарного подхода.Это в значительной степени связано с тем, что полисахариды не кодируются напрямую последовательностью генома; Для синтеза активированных сахарных остатков требуется множество ферментов, связи и многие компоненты стенки претерпевают обширные модификации, включая метилирование, этерификацию / деэтерификацию и ацетилирование, а также добавление одного или блоков остатков сахара.
Проблемы изучения разнообразия клеточных стенок
Конкретные гены и ферменты, которые приводят к синтезу определенных компонентов клеточной стенки, еще предстоит полностью выяснить.Кроме того, в большинстве случаев требуется несколько ферментов для синтеза определенного компонента клеточной стенки, который может дополнительно подвергаться последующей модификации в muro . Следовательно, мы еще не находимся на стадии, когда можно определить состав и разнообразие клеточной стенки с помощью сравнительного геномного подхода, и большая часть полученных на сегодняшний день знаний опирается на анализ полимеров. Одна из фундаментальных трудностей, связанных с этим, заключается в том, что полисахариды не поддаются легкому секвенированию.Их структуры можно определить с помощью нескольких хорошо зарекомендовавших себя химических методов, которые были разработаны и применялись для исследований клеточных стенок за последние 50 или около того лет. Каждый метод имеет как ограничения, так и достоинства, но их можно применять совместно для выявления и определения сложности и разнообразия клеточной стенки. Немногие из разработанных к настоящему времени методов поддаются высокопроизводительному скринингу, поэтому широкие исследования должны изначально полагаться на частичную характеристику. Инфракрасная спектроскопия с преобразованием Фурье (FTIR), требующая небольшой подготовки образца, может иметь высокую пропускную способность и полезна для определения различий в составе клеточной стенки в разных образцах, но редко бывает эффективной для точного анализа состава, поскольку не дает информации о последовательности (Mouille et al., 2003). Недавно были разработаны методы, основанные на углеводных микрочипах, зондированных моноклональными антителами (mAb) и углеводсвязывающими модулями (CBM) со специфичностью к эпитопам полисахаридов клеточной стенки. Эта технология может позволить анализ появления 100 эпитопов за 2–3 дня и может многое рассказать о составе клеточной стенки, несмотря на некоторые ограничения, в значительной степени проистекающие из текущей, хотя и увеличивающейся, доступности охарактеризованных mAb и CBM (Moller et al.
Немногие из разработанных к настоящему времени методов поддаются высокопроизводительному скринингу, поэтому широкие исследования должны изначально полагаться на частичную характеристику. Инфракрасная спектроскопия с преобразованием Фурье (FTIR), требующая небольшой подготовки образца, может иметь высокую пропускную способность и полезна для определения различий в составе клеточной стенки в разных образцах, но редко бывает эффективной для точного анализа состава, поскольку не дает информации о последовательности (Mouille et al., 2003). Недавно были разработаны методы, основанные на углеводных микрочипах, зондированных моноклональными антителами (mAb) и углеводсвязывающими модулями (CBM) со специфичностью к эпитопам полисахаридов клеточной стенки. Эта технология может позволить анализ появления 100 эпитопов за 2–3 дня и может многое рассказать о составе клеточной стенки, несмотря на некоторые ограничения, в значительной степени проистекающие из текущей, хотя и увеличивающейся, доступности охарактеризованных mAb и CBM (Moller et al. , 2007; Соренсен и др., 2009; Pattathil et al., 2010). Применение FTIR и углеводных микрочипов может быть использовано для обозначения клеточных стенок, состав которых может потребовать дальнейшего и подробного анализа, поскольку они, по-видимому, значительно отличаются от охарактеризованных клеточных стенок. Таким образом, использование этих методов облегчает разработку гипотез относительно состава клеточной стенки, которая может быть дополнительно исследована путем более подробного анализа подмножества набора образцов. Электрофорез в полисахаридном геле (PACE) (Goubet et al., 2002), профилирование массы олигосахаридов (OLIMP; Lerouxel et al., 2002), бумажная хроматография и тонкослойная хроматография (Fry, 2001) и родственные подходы — мощные инструменты для следующего уровня, работающего с подмножеством исходного набора образцов. Анализы гликозильных связей и ЯМР составляют последний уровень, поскольку эти методы требуют много времени, а для ЯМР также довольно нечувствительны. Таким образом, эти методы не подходят для широкомасштабного отбора проб, но часто незаменимы для углубленного анализа отобранных проб.
, 2007; Соренсен и др., 2009; Pattathil et al., 2010). Применение FTIR и углеводных микрочипов может быть использовано для обозначения клеточных стенок, состав которых может потребовать дальнейшего и подробного анализа, поскольку они, по-видимому, значительно отличаются от охарактеризованных клеточных стенок. Таким образом, использование этих методов облегчает разработку гипотез относительно состава клеточной стенки, которая может быть дополнительно исследована путем более подробного анализа подмножества набора образцов. Электрофорез в полисахаридном геле (PACE) (Goubet et al., 2002), профилирование массы олигосахаридов (OLIMP; Lerouxel et al., 2002), бумажная хроматография и тонкослойная хроматография (Fry, 2001) и родственные подходы — мощные инструменты для следующего уровня, работающего с подмножеством исходного набора образцов. Анализы гликозильных связей и ЯМР составляют последний уровень, поскольку эти методы требуют много времени, а для ЯМР также довольно нечувствительны. Таким образом, эти методы не подходят для широкомасштабного отбора проб, но часто незаменимы для углубленного анализа отобранных проб. Какой бы метод анализа ни был выбран, отбор проб всегда будет создавать серьезные проблемы. Учитывая, что невозможно взять образцы стенок каждого сохранившегося растения, тогда какие растения и какие органы или ткани следует выбрать и имеется ли в наличии достаточное количество ткани? Кажется разумным выбрать виды, которые являются репрезентативными для таксономических групп или морфотипов, но такая расстановка приоритетов не всегда проста. После того, как растения выбраны, возникает дальнейшая сложность правильного отбора образцов индивидуальных клеточных стенок внутри этого растения.Один из вариантов — просто гомогенизировать все растение и извлечь как можно больше полисахаридов клеточной стенки. Это возможно, скажем, для небольших саженцев и микроводорослей, но не для древесных пород и деревьев. Очевидно, что необходимо выбрать определенные ткани, органы или стадии развития — но на каком основании? Эквивалентность также может быть проблематичной, потому что растения различаются по органам и тканям.
Какой бы метод анализа ни был выбран, отбор проб всегда будет создавать серьезные проблемы. Учитывая, что невозможно взять образцы стенок каждого сохранившегося растения, тогда какие растения и какие органы или ткани следует выбрать и имеется ли в наличии достаточное количество ткани? Кажется разумным выбрать виды, которые являются репрезентативными для таксономических групп или морфотипов, но такая расстановка приоритетов не всегда проста. После того, как растения выбраны, возникает дальнейшая сложность правильного отбора образцов индивидуальных клеточных стенок внутри этого растения.Один из вариантов — просто гомогенизировать все растение и извлечь как можно больше полисахаридов клеточной стенки. Это возможно, скажем, для небольших саженцев и микроводорослей, но не для древесных пород и деревьев. Очевидно, что необходимо выбрать определенные ткани, органы или стадии развития — но на каком основании? Эквивалентность также может быть проблематичной, потому что растения различаются по органам и тканям. У некоторых есть цветы и листья, у некоторых их нет. Кроме того, измененные условия роста могут влиять на экспрессию и структуру компонентов клеточной стенки в пределах одного и того же вида (Iraki et al., 1989а, б, в). Такие соображения важны, потому что мы знаем, что некоторые компоненты клеточной стенки могут быть очень избирательно распределены по растению (см. Раздел «Точное картирование разнообразия и гетерогенности клеточной стенки на клеточном и субклеточном уровнях») и их легко упустить. Еще одна важная задача — интерпретация различных анализов. Требуется значительная осторожность, учитывая практически невозможность действительно всеобъемлющего отбора проб. В случае положительной идентификации несколькими методами, конкретный полисахарид можно рассматривать как «присутствующий», но если пробы не были взяты из всех частей растения (включая все стадии развития), то неспособность идентифицировать конкретный полисахарид следует интерпретировать как присутствие этого полисахарида. быть «неизвестным», а не «отсутствующим».
У некоторых есть цветы и листья, у некоторых их нет. Кроме того, измененные условия роста могут влиять на экспрессию и структуру компонентов клеточной стенки в пределах одного и того же вида (Iraki et al., 1989а, б, в). Такие соображения важны, потому что мы знаем, что некоторые компоненты клеточной стенки могут быть очень избирательно распределены по растению (см. Раздел «Точное картирование разнообразия и гетерогенности клеточной стенки на клеточном и субклеточном уровнях») и их легко упустить. Еще одна важная задача — интерпретация различных анализов. Требуется значительная осторожность, учитывая практически невозможность действительно всеобъемлющего отбора проб. В случае положительной идентификации несколькими методами, конкретный полисахарид можно рассматривать как «присутствующий», но если пробы не были взяты из всех частей растения (включая все стадии развития), то неспособность идентифицировать конкретный полисахарид следует интерпретировать как присутствие этого полисахарида. быть «неизвестным», а не «отсутствующим». «При изучении эволюции клеточной стенки также важно учитывать полисахариды, которые могут встречаться на таких низких уровнях, что в других исследованиях они могут считаться функционально несущественными. Например, хотя данные о гликозильной связи и ICP-MS предполагают, что RG-II или RG-II-подобный олигосахарид встречается в очень низких количествах у бессосудистых мохообразных, менее 1% от такового у покрытосеменных (Matsunaga et al., 2004), его присутствие у этих растений, тем не менее, будет иметь значение с точки зрения эволюции лежащих в основе биосинтетических механизмов.
«При изучении эволюции клеточной стенки также важно учитывать полисахариды, которые могут встречаться на таких низких уровнях, что в других исследованиях они могут считаться функционально несущественными. Например, хотя данные о гликозильной связи и ICP-MS предполагают, что RG-II или RG-II-подобный олигосахарид встречается в очень низких количествах у бессосудистых мохообразных, менее 1% от такового у покрытосеменных (Matsunaga et al., 2004), его присутствие у этих растений, тем не менее, будет иметь значение с точки зрения эволюции лежащих в основе биосинтетических механизмов.
Многоуровневый подход к пониманию разнообразия и эволюции клеточной стенки
Авторы приняли многоуровневую стратегию картирования полисахаридов клеточной стенки и генетического разнообразия, чтобы получить представление о лежащих в основе эволюционных механизмах (рис. 1А). Первый уровень состоит из первичного скрининга полисахаридов клеточной стенки на основе углеводных микрочипов, зондированных с помощью mAb и CBM (Moller et al.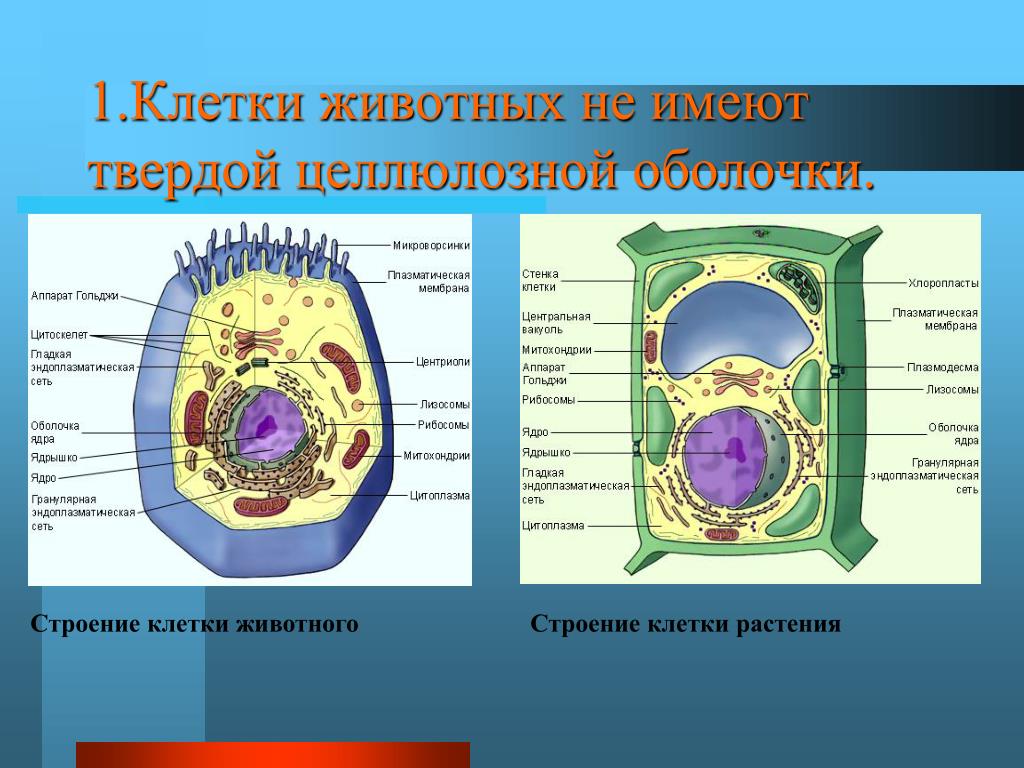 , 2007). Этот уровень ограничен доступностью охарактеризованных mAb и CBM и их способностью распознавать многочисленные эпитопы, которые встречаются в различных компонентах клеточной стенки растений.Очевидно, что в идеале было бы как можно больше охватывать все эпитопы, существующие в компонентах клеточной стенки. Однако это не текущая ситуация, и есть некоторые примечательные компоненты стенки, такие как RG-II, для которых еще предстоит получить эффективное mAb. Параллельно с углеводными микрочипами анализируются геномы и транскриптомы для идентификации GT, связанных с клеточной стенкой. Второй уровень анализа направлен на получение более подробной информации о некоторых полисахаридах и генах из подмножеств растений.Эти анализы выполняются с использованием установленных методов анализа полисахаридов, клонирования и секвенирования генов. Третий уровень направлен на получение окончательной информации и функций генов, белков и полисахаридов. В некоторых случаях экспрессируются гены и определяется биохимическая активность ГТ.
, 2007). Этот уровень ограничен доступностью охарактеризованных mAb и CBM и их способностью распознавать многочисленные эпитопы, которые встречаются в различных компонентах клеточной стенки растений.Очевидно, что в идеале было бы как можно больше охватывать все эпитопы, существующие в компонентах клеточной стенки. Однако это не текущая ситуация, и есть некоторые примечательные компоненты стенки, такие как RG-II, для которых еще предстоит получить эффективное mAb. Параллельно с углеводными микрочипами анализируются геномы и транскриптомы для идентификации GT, связанных с клеточной стенкой. Второй уровень анализа направлен на получение более подробной информации о некоторых полисахаридах и генах из подмножеств растений.Эти анализы выполняются с использованием установленных методов анализа полисахаридов, клонирования и секвенирования генов. Третий уровень направлен на получение окончательной информации и функций генов, белков и полисахаридов. В некоторых случаях экспрессируются гены и определяется биохимическая активность ГТ. На рисунке 1B показаны некоторые предварительные данные первичного скрининга полисахаридов и генов, связанных с клеточной стенкой. Комбинированный анализ наборов данных может дать представление о времени и механизме определенных эволюционных событий.Например, Xue and Fry (2012) предположили, что MLG ограничены хвощами, на основании результатов, полученных при обработке клеточных стенок различных монилофитов лихеназой, ферментом, который специфически фрагментирует эти (1 → 3) (1 → 4) -β — D -глюканы. Напротив, мы получили доказательства наличия MLG у Selaginella moellendorffii и отобранных зеленых водорослей Charophycean (CGA), используя микроматричный скрининг на полисахариды и лечение лихеназой (Harholt et al., 2012; Fangel and Willats, unpublished) . Поскольку последовательность генома доступна для S. moellendorffii , можно с уверенностью установить, что это растение не содержит ортологов генов CSLF и C SLH (Harholt et al., 2012), которые участвуют в синтез (1 → 3) (1 → 4) -β- D -глюкана у видов Poales (Burton et al.
На рисунке 1B показаны некоторые предварительные данные первичного скрининга полисахаридов и генов, связанных с клеточной стенкой. Комбинированный анализ наборов данных может дать представление о времени и механизме определенных эволюционных событий.Например, Xue and Fry (2012) предположили, что MLG ограничены хвощами, на основании результатов, полученных при обработке клеточных стенок различных монилофитов лихеназой, ферментом, который специфически фрагментирует эти (1 → 3) (1 → 4) -β — D -глюканы. Напротив, мы получили доказательства наличия MLG у Selaginella moellendorffii и отобранных зеленых водорослей Charophycean (CGA), используя микроматричный скрининг на полисахариды и лечение лихеназой (Harholt et al., 2012; Fangel and Willats, unpublished) . Поскольку последовательность генома доступна для S. moellendorffii , можно с уверенностью установить, что это растение не содержит ортологов генов CSLF и C SLH (Harholt et al., 2012), которые участвуют в синтез (1 → 3) (1 → 4) -β- D -глюкана у видов Poales (Burton et al. , 2006; Doblin et al., 2009). Эти данные являются убедительным доказательством того, что (1 → 3) (1 → 4) -β- D -глюкан эволюционировал по крайней мере дважды в результате конвергентной эволюции.
, 2006; Doblin et al., 2009). Эти данные являются убедительным доказательством того, что (1 → 3) (1 → 4) -β- D -глюкан эволюционировал по крайней мере дважды в результате конвергентной эволюции.
РИСУНОК 1. (A) Многоуровневый подход необходим для оценки наличия полисахаридов клеточной стенки и родственных генов во всем царстве растений. (B) Отобранные данные первичного скрининга полисахаридов в различных видах растений и результаты поиска геномов и транскриптомов GT, связанных с клеточной стенкой. Точки внизу столбиков указывают на самую базовую группу растений, в которой на данный момент идентифицирован этот конкретный полисахарид или последовательность гена.Столбики показывают, что в большинстве случаев предполагается, что после эволюции ген или полисахарид сохраняется на протяжении всей эволюции и присутствует в более поздних расходящихся видах. Однако это отнюдь не универсально. Например, пунктирная полоса для (1 → 3) (1 → 4) -β- D -глюкана указывает на то, что появление этого полисахарида является прерывистым во всем царстве растений и возникло в результате конвергентной эволюции более одного раза. В некоторых случаях наличие компонентов клеточной стенки сомнительно. Например, лигнин был предварительно идентифицирован в некоторых зеленых водорослях Charophycean, но наиболее базальной группой, в которой он был определен окончательно, является Lycopodiophyta.Группа предполагаемых предковых CSL, которые являются общими предками CSLA s и CSLC s, была идентифицирована в некоторых хлорофитных водорослях Yin et al. (2009). Эти гены имеют общую гомологию как с CSLA s, так и с CSLC s, но, по-видимому, достаточно отличаются, чтобы гарантировать новое имя семейства, которому мы присвоили CSLK s, в соответствии с существующей алфавитной последовательностью существующих имен семейства CSL . (1) Цекос (1999); (2) Scherp et al.(2001); (3) Маки и Сто (1968); (4) Домозыч и др. (2009); (5) Соренсен и др. (2011); (6) Мацунага и др. (2004); (7) Эстевес и др. (2009); (8) Weng et al. (2008); (9) Робертс и др. (2002); (10) Аберкромби и др. (2011); (11) Инь и др.
В некоторых случаях наличие компонентов клеточной стенки сомнительно. Например, лигнин был предварительно идентифицирован в некоторых зеленых водорослях Charophycean, но наиболее базальной группой, в которой он был определен окончательно, является Lycopodiophyta.Группа предполагаемых предковых CSL, которые являются общими предками CSLA s и CSLC s, была идентифицирована в некоторых хлорофитных водорослях Yin et al. (2009). Эти гены имеют общую гомологию как с CSLA s, так и с CSLC s, но, по-видимому, достаточно отличаются, чтобы гарантировать новое имя семейства, которому мы присвоили CSLK s, в соответствии с существующей алфавитной последовательностью существующих имен семейства CSL . (1) Цекос (1999); (2) Scherp et al.(2001); (3) Маки и Сто (1968); (4) Домозыч и др. (2009); (5) Соренсен и др. (2011); (6) Мацунага и др. (2004); (7) Эстевес и др. (2009); (8) Weng et al. (2008); (9) Робертс и др. (2002); (10) Аберкромби и др. (2011); (11) Инь и др.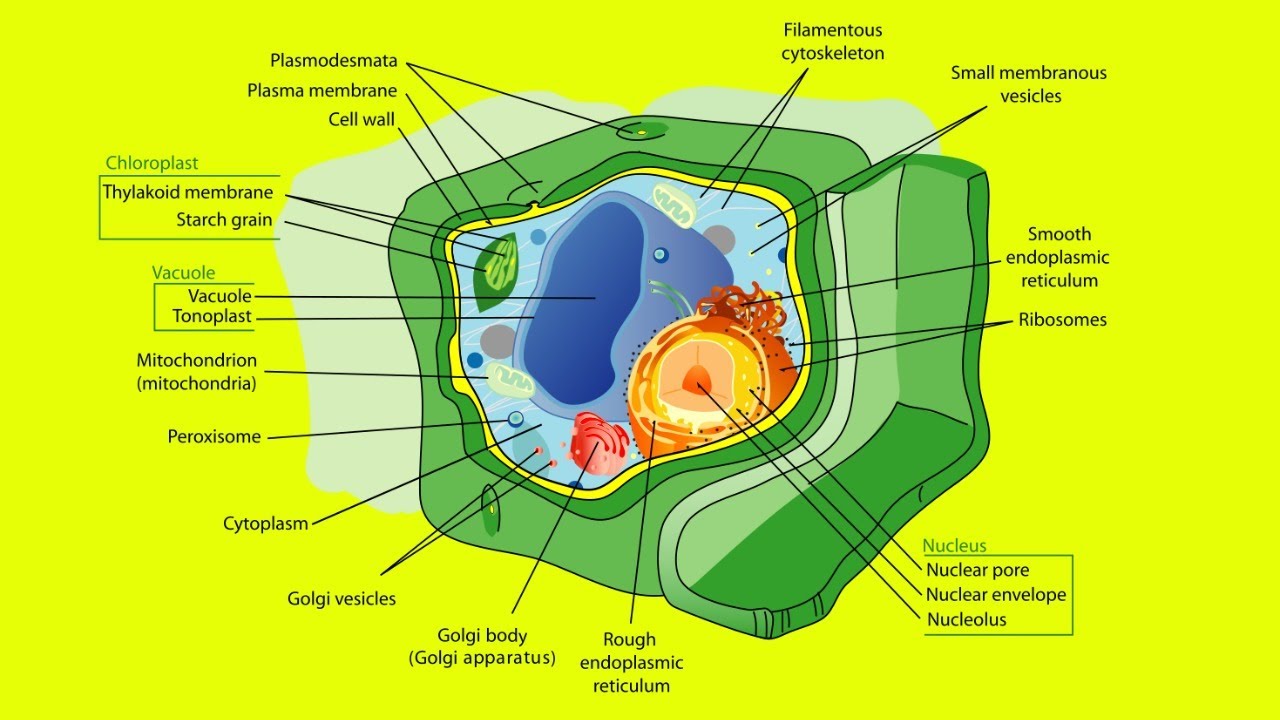 (2009); (12) Инь и др. (2010); (13) Egelund et al. (2007), Velasquez et al. (2011); (14) Qu et al. (2004), Harholt et al. (2012). ТК, терминальный комплекс.
(2009); (12) Инь и др. (2010); (13) Egelund et al. (2007), Velasquez et al. (2011); (14) Qu et al. (2004), Harholt et al. (2012). ТК, терминальный комплекс.
Анализ генома и транскриптома гликозилтрансфераз, связанных с клеточной стенкой
«По крайней мере, один GT для каждой гликозидной связи» является аксиомой нашего биоинформатического анализа биогенеза клеточной стенки.Редкие ГТ с двойной функцией известны у животных и растений, при этом CslA является примером ГТ, который может использовать два разных GDP-сахара в качестве субстрата. Принятие аксиомы одного GT на гликозидную связь должно позволить сделать вывод о минимальном количестве GT, участвующих в биосинтезе клеточной стенки. Однако это невозможно по двум причинам. Во-первых, эволюция цветковых растений сопровождалась очень значительной дупликацией генов. Вероятно, способствующим фактором является дифференциация сложных тканей покрытосеменных растений, требующая раздельного регулирования в пространстве и времени одной и той же каталитической активности. Mitchell et al. (2007) объединили эту линию мышления с анализом экспрессии и предположили, что определенные клады семейства GT61 должны включать гены, участвующие в синтезе полимера, имеющего особое значение для злаков, — предсказание, которое недавно оказалось верным (Anders et al., 2012). Обширный репертуар GT мха Physcomitrella patens , несмотря на то, что они не являются сосудистыми (то есть имеют только не одревесневшие ткани), а также разошлись до событий дупликации генов, связанных с эволюцией цветковых растений, предостерегает нас от обобщения. этот принцип чрезмерно.Вторая причина, по которой количество GT не может быть выведено из количества различных связей, состоит в том, что биосинтез некоторых полисахаридов оказался намного более сложным, чем ожидалось, исходя из структуры полисахарида; Биосинтез ксилана, обзор Scheller and Ulvskov (2010), является ярким примером. У двудольных растений GT из двух разных семейств (GT43 и GT47) участвуют в синтезе основной цепи ксилана, всего восемь GT для синтеза одной связи.
Mitchell et al. (2007) объединили эту линию мышления с анализом экспрессии и предположили, что определенные клады семейства GT61 должны включать гены, участвующие в синтезе полимера, имеющего особое значение для злаков, — предсказание, которое недавно оказалось верным (Anders et al., 2012). Обширный репертуар GT мха Physcomitrella patens , несмотря на то, что они не являются сосудистыми (то есть имеют только не одревесневшие ткани), а также разошлись до событий дупликации генов, связанных с эволюцией цветковых растений, предостерегает нас от обобщения. этот принцип чрезмерно.Вторая причина, по которой количество GT не может быть выведено из количества различных связей, состоит в том, что биосинтез некоторых полисахаридов оказался намного более сложным, чем ожидалось, исходя из структуры полисахарида; Биосинтез ксилана, обзор Scheller and Ulvskov (2010), является ярким примером. У двудольных растений GT из двух разных семейств (GT43 и GT47) участвуют в синтезе основной цепи ксилана, всего восемь GT для синтеза одной связи. Предполагается, что для биосинтеза пектина требуется не менее 67 ферментов, GT плюс метил- и ацетилтрансферазы (Mohnen, 2008).Слишком мало из них было идентифицировано до сих пор, чтобы судить, является ли подход Природы к биосинтезу пектина скудным или обширным, как в случае с ксиланом. Несмотря на эти ограничения, поиск геномов для их репертуара GT оказался плодотворным. Обычно это делается с использованием базы данных CAZy в качестве основы, см. Вставку 1. База данных CAZy является самой обширной базой данных GT и содержит GT из всех трех царств. Используя глобальный подход, включающий всю базу данных CAZy на экране, можно обнаружить более удаленные ортологии, примером чего может служить открытие маннозилглицератсинтазы GT78 в S.moellendorffii (Scheller et al., 2010). Но CAZy не является полным, и GT могут быть обнаружены за пределами CAZy, а некоторые виды деятельности трудно учесть в рамках существующей базы данных CAZy (Hansen et al., 2012). Подход, основанный на CAZy, применялся к тополю (Geisler-Lee et al.
Предполагается, что для биосинтеза пектина требуется не менее 67 ферментов, GT плюс метил- и ацетилтрансферазы (Mohnen, 2008).Слишком мало из них было идентифицировано до сих пор, чтобы судить, является ли подход Природы к биосинтезу пектина скудным или обширным, как в случае с ксиланом. Несмотря на эти ограничения, поиск геномов для их репертуара GT оказался плодотворным. Обычно это делается с использованием базы данных CAZy в качестве основы, см. Вставку 1. База данных CAZy является самой обширной базой данных GT и содержит GT из всех трех царств. Используя глобальный подход, включающий всю базу данных CAZy на экране, можно обнаружить более удаленные ортологии, примером чего может служить открытие маннозилглицератсинтазы GT78 в S.moellendorffii (Scheller et al., 2010). Но CAZy не является полным, и GT могут быть обнаружены за пределами CAZy, а некоторые виды деятельности трудно учесть в рамках существующей базы данных CAZy (Hansen et al., 2012). Подход, основанный на CAZy, применялся к тополю (Geisler-Lee et al. , 2006), а совсем недавно к Brachypodium distachyon (Vogel et al., 2010) и S. moellendorffii (Banks et al., 2011). что привело к соглашению об именах на основе CAZy для предполагаемых GT, которые могут быть отнесены к семейству генов, но не к функции.
, 2006), а совсем недавно к Brachypodium distachyon (Vogel et al., 2010) и S. moellendorffii (Banks et al., 2011). что привело к соглашению об именах на основе CAZy для предполагаемых GT, которые могут быть отнесены к семейству генов, но не к функции.
BOX 1 Текстовое поле под названием CAZy.
База данных CAZy, www.cazy.org, описывает семейства структурно связанных каталитических и углеводсвязывающих модулей (или функциональных доменов) ферментов, которые разрушают, модифицируют или создают гликозидные связи (Cantarel et al., 2009). GT делятся на 94 семейства (и продолжают расти). Классификация частично основана на сходстве последовательностей, частично на трехмерной информации о структуре белка. 94 семейства могут быть сгруппированы в небольшое количество GT-складок, создавая более высокий уровень в иерархии, формализованный как кланы для гликозидгидролаз, но еще не для GT.Большинство семейств включают несколько видов деятельности, поэтому отнесения GT к семейству редко бывает достаточно для определения точной каталитической активности GT. Однако можно с хорошей точностью спрогнозировать инвертирование или сохранение GT на основе их семейства CAZy, где сохранение относится к ситуации, когда перенесенный моносахарид оказывается в той же аномерной конформации, что и в донорном субстрате, и, наоборот, для инвертирующих GT.
Однако можно с хорошей точностью спрогнозировать инвертирование или сохранение GT на основе их семейства CAZy, где сохранение относится к ситуации, когда перенесенный моносахарид оказывается в той же аномерной конформации, что и в донорном субстрате, и, наоборот, для инвертирующих GT.
Применение основанной на CAZy классификации предполагаемых GT на больших филогенетических расстояниях может дать эволюционно релевантную информацию, как показано на примере конвергентной эволюции MLG.CAZy также может использоваться в качестве поддержки усилий по открытию генов, поскольку Egelund et al. (2007) использовали гомологию между генами Chlamydomonas и Arabidopsis в кладе A семейства GT77, чтобы сделать вывод о функции генов уменьшенной остаточной арабинозы (RRA) как кодирующих предполагаемые экстензин-арабинозилтрансферазы. Эта аннотация не доказана, но была подтверждена Velasquez et al. (2011). Экстенсин вместе с маннаном, насколько известно авторам, являются единственными известными компонентами общей клеточной стенки между зелеными водорослями Chlorophycean и Arabidopsis (рисунок 1; Estevez et al. , 2009).
, 2009).
Еще не было секвенировано CGA, что прискорбно, учитывая, что анализ клеточной стенки убедительно свидетельствует о том, что общим предком всех растений с ксилоглюканом и пектиновыми полимерами, типичными для сосудистых растений, был CGA (рис. 1). Доступен ряд наборов данных EST (Timme and Delwiche, 2010; Wodniok et al., 2011; Timme et al., 2012), которые могут быть подвергнуты тому же анализу на основе CAZy, что и полные геномы, хотя и менее безопасно. Наши неопубликованные наблюдения этой природы в сочетании с недавним филогенетическим анализом, основанным на геномных данных, приводят нас к предположению, что такие роды, как Penium и Coleochaete , представляют собой самые ранние версии клеточной стенки высших растений, в то время как такие виды, как Chara , которые внешне выглядит более продвинутым, существенно отклонился от основного эволюционного пути, ведущего к наземным растениям, и, следовательно, представляет собой менее привлекательную модель для отслеживания эволюционной истории клеточной стенки растений (Timme et al.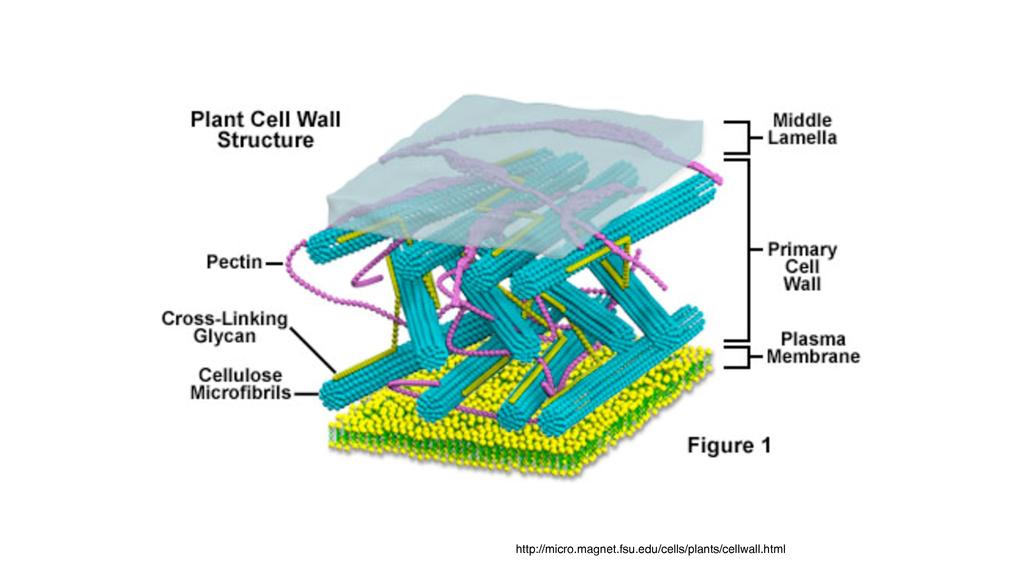 , 2012).
, 2012).
Точное картирование разнообразия и неоднородности клеточной стенки на клеточном и субклеточном уровнях
Целлюлоза широко распространена в большинстве клеточных стенок, но ясно, что известное структурное разнообразие полисахаридов гемицеллюлозы и пектиновых групп регулируется как таксономически, так и в отношении типов клеток и микроструктур клеточных стенок в тканях. Использование mAb и CBM для анализа in situ клеточных стенок остается ключевым подходом к определению молекулярной архитектуры стенок и их неоднородностей (Burton et al., 2010; Паттатил и др., 2010; Ли и др., 2011). Использование одних и тех же молекулярных зондов в гликомическом анализе представляет собой очень полезную и дополняющую деятельность, при которой специфические структурные особенности олигосахаридов могут быть широко изучены с точки зрения встречаемости и биохимии в дополнение к клеточным местоположениям.
Важно отметить, что, хотя нам известны некоторые широкие клеточные проявления специфических нецеллюлозных полисахаридов, такие как содержание ксилана во вторичных клеточных стенках двудольных растений, HG в первичных клеточных стенках и (пример на субклеточном уровне) Отсутствие пектинового галактана из ямок, у нас нет хорошего понимания клеточного распределения всех полимеров для большинства типов растительных клеток.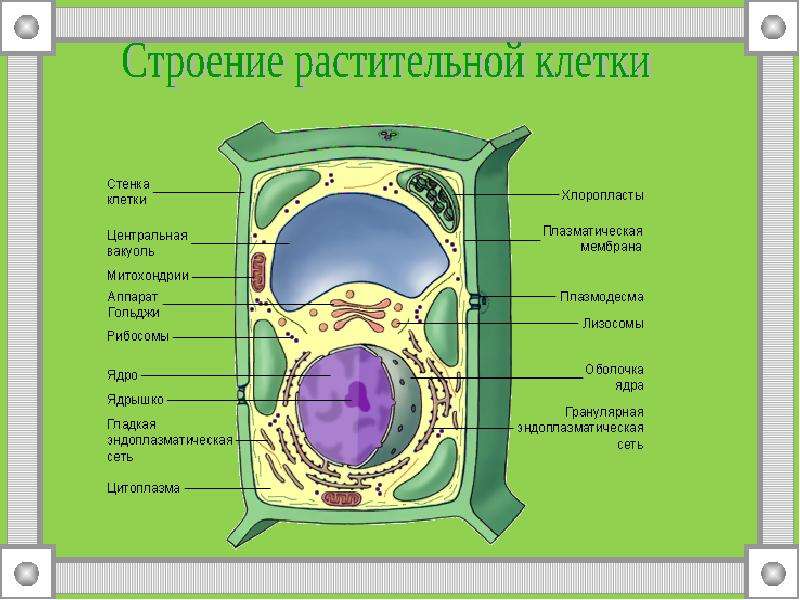 Даже для Arabidopsis систематическая оценка in situ основных полимеров в клеточных стенках основных органов еще не проведена. Некоторые из них можно предсказать — например, указанные выше, но распределение RG-I и других пектиновых эпитопов или гемицеллюлоз еще нельзя предсказать с уверенностью. Анализ структур клеточной стенки in situ стал более сложным, но также и более показательным в том смысле, что по мере расширения наборов зондов для конкретных полисахаридов может быть обнаружено большее разнообразие клеточных стенок и неоднородность.Например, это относится к пектиновому HG (Willats et al., 2001; Parre and Geitmann, 2005; Wolf et al., 2009), а также к пектиновому арабинану (Verhertbruggen et al., 2009), где mAb к линейному арабинану обнаруживает субструктуры арабинана в ограниченных межклеточных областях паренхимы, как показано на рисунке 2. Это также применимо к структурам ксилана в той же паренхиме (Hervé, 2009). Какова функциональная основа полисахаридной тонкой структуры как ксиланов, так и арабинанов, которые настолько тесно пространственно регулируются по отношению к таким факторам, как клеточная адгезия (рис.
Даже для Arabidopsis систематическая оценка in situ основных полимеров в клеточных стенках основных органов еще не проведена. Некоторые из них можно предсказать — например, указанные выше, но распределение RG-I и других пектиновых эпитопов или гемицеллюлоз еще нельзя предсказать с уверенностью. Анализ структур клеточной стенки in situ стал более сложным, но также и более показательным в том смысле, что по мере расширения наборов зондов для конкретных полисахаридов может быть обнаружено большее разнообразие клеточных стенок и неоднородность.Например, это относится к пектиновому HG (Willats et al., 2001; Parre and Geitmann, 2005; Wolf et al., 2009), а также к пектиновому арабинану (Verhertbruggen et al., 2009), где mAb к линейному арабинану обнаруживает субструктуры арабинана в ограниченных межклеточных областях паренхимы, как показано на рисунке 2. Это также применимо к структурам ксилана в той же паренхиме (Hervé, 2009). Какова функциональная основа полисахаридной тонкой структуры как ксиланов, так и арабинанов, которые настолько тесно пространственно регулируются по отношению к таким факторам, как клеточная адгезия (рис.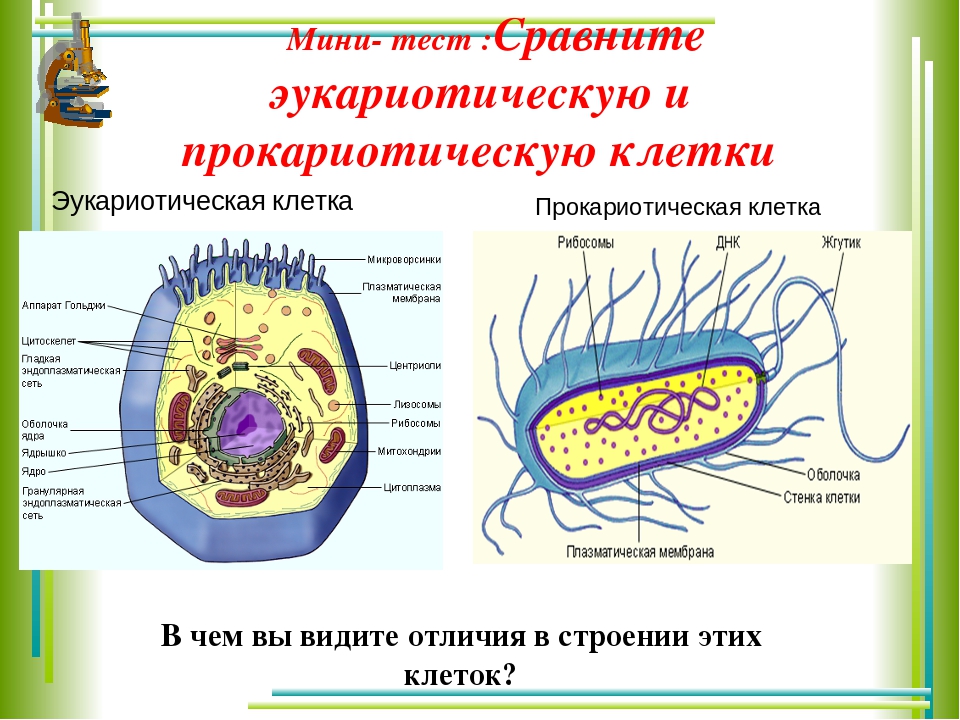 2)? Будет ли это парадигмой для нецеллюлозных полимеров стенок, для которых в настоящее время у нас есть один или ограниченное количество зондов, таких как пектиновый галактан и ксилоглюкан? В ближайшие несколько лет мы увидим более подробные систематические оценки молекулярных архитектур.Это будет в сочетании с ферментативной и / или химической предварительной обработкой, которая в некоторых случаях требуется для оптимизации обнаружения полисахаридов, а в случае маскировки полисахаридов (когда один класс полимеров блокирует доступ к другому классу полимеров) указывает на важные особенности клеточной стенки. архитектуры, отражающие доступ к белкам (Vreeland et al., 1984; Marcus et al., 2010; Davies et al., 2012).
2)? Будет ли это парадигмой для нецеллюлозных полимеров стенок, для которых в настоящее время у нас есть один или ограниченное количество зондов, таких как пектиновый галактан и ксилоглюкан? В ближайшие несколько лет мы увидим более подробные систематические оценки молекулярных архитектур.Это будет в сочетании с ферментативной и / или химической предварительной обработкой, которая в некоторых случаях требуется для оптимизации обнаружения полисахаридов, а в случае маскировки полисахаридов (когда один класс полимеров блокирует доступ к другому классу полимеров) указывает на важные особенности клеточной стенки. архитектуры, отражающие доступ к белкам (Vreeland et al., 1984; Marcus et al., 2010; Davies et al., 2012).
РИСУНОК 2. Архитектурная неоднородность в первичных клеточных стенках .Эквивалентные поперечные срезы кортикальной области стеблей табака с иммунофлуоресцентной визуализацией двух эпитопов арабинана (mAb LM6 и LM13), двух эпитопов ксилана (mAb LM10 и CBM15) и одного эпитопа ксилоглюкана (mAb LM15). Стрелки указывают на межклеточный матрикс, разделяемый соседними клетками. Эпитопы ксилана и ксилоглюкана показаны после удаления пектинового гомогалактуронана. Структурные особенности пектинового арабинана, ксилана и ксилоглюкана демонстрируют пространственную неоднородность в отношении утолщения клеточной стенки и межклеточных областей.mAb, моноклональное антитело; CBM, модуль связывания углеводов. Масштабная линейка = 100 мкм.
Стрелки указывают на межклеточный матрикс, разделяемый соседними клетками. Эпитопы ксилана и ксилоглюкана показаны после удаления пектинового гомогалактуронана. Структурные особенности пектинового арабинана, ксилана и ксилоглюкана демонстрируют пространственную неоднородность в отношении утолщения клеточной стенки и межклеточных областей.mAb, моноклональное антитело; CBM, модуль связывания углеводов. Масштабная линейка = 100 мкм.
Очевидно, что мы сталкиваемся со многими проблемами в понимании эволюции клеточной стенки, не в последнюю очередь из которых входит достаточный отбор образцов с последующим соответствующим синтезом и интерпретацией больших наборов данных, включая разнообразную информацию, такую как данные о последовательностях генов и белков, а также о связях сахаров и распределении эпитопов. Однако, поскольку молекулярная архитектура клеток и тканей документирована, другая важная проблема, которая привлекает внимание, — это функция отдельных компонентов стенок и целых клеточных стенок, которые могут сильно различаться по количественному и качественному составу. Почему некоторые клеточные стенки имеют гемицеллюлозы ксилоглюкана, ксилана и маннана в различных пространственных распределениях, как, например, в широко изученной стволовой системе табака, как показано на рисунке 2? Как эти неоднородности объединены в функциональное целое с точки зрения свойств и функций стен? In vitro анализ композитов, образованных из целлюлозы, пектинов и ксилоглюканов, дал бесценную информацию о свойствах некоторых компонентов стенок (Chanliaud et al., 2002). Однако структурная и композиционная сложность встречающихся в природе клеточных стенок означает, что подход in vitro не может разумно применяться для исследования функциональных свойств всего разнообразия существующих стенок. In vivo. Были разработаны методы исследования биомеханики стенок на тканевом и нижних уровнях (Spatz et al., 1998; Burget, 2006), способствующие лучшему пониманию того, как собираются стенки, и мельчайших деталей доменов стенки и их наномеханические свойства, вероятно, проявятся в течение следующих нескольких лет, поскольку подробный анализ in situ будет совмещен с генетическим и ферментативным вмешательством.
Почему некоторые клеточные стенки имеют гемицеллюлозы ксилоглюкана, ксилана и маннана в различных пространственных распределениях, как, например, в широко изученной стволовой системе табака, как показано на рисунке 2? Как эти неоднородности объединены в функциональное целое с точки зрения свойств и функций стен? In vitro анализ композитов, образованных из целлюлозы, пектинов и ксилоглюканов, дал бесценную информацию о свойствах некоторых компонентов стенок (Chanliaud et al., 2002). Однако структурная и композиционная сложность встречающихся в природе клеточных стенок означает, что подход in vitro не может разумно применяться для исследования функциональных свойств всего разнообразия существующих стенок. In vivo. Были разработаны методы исследования биомеханики стенок на тканевом и нижних уровнях (Spatz et al., 1998; Burget, 2006), способствующие лучшему пониманию того, как собираются стенки, и мельчайших деталей доменов стенки и их наномеханические свойства, вероятно, проявятся в течение следующих нескольких лет, поскольку подробный анализ in situ будет совмещен с генетическим и ферментативным вмешательством.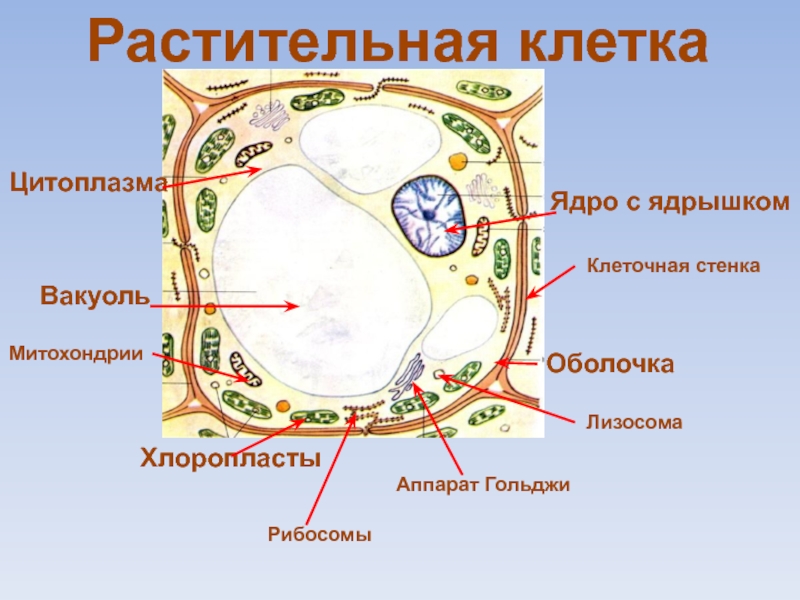 Интеграция этих знаний станет серьезной проблемой и представляет собой захватывающий рубеж для биологии клеточной стенки.
Интеграция этих знаний станет серьезной проблемой и представляет собой захватывающий рубеж для биологии клеточной стенки.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Список литературы
Аберкромби, Дж. М., О’Мира, Б. К., Моффат, А. Р., и Уильямс, Дж. Х. (2011). Эволюция развития клеточных стенок пыльцевых трубок цветковых растений: паттерны экспрессии генов каллозосинтазы (CalS). EvoDevo 2, 14.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Андерс, Н., Уилкинсон, Мэриленд, Лавгроув, А., Фриман, Дж., Трифона, Т., Пелльни, Т.К., Веймар, Т., Мортимер, Дж. К., Стотт, К., Бейкер, Дж. М., Дефоин-Платель, М., Шури , PR, Дюпри, П., и Митчелл, RAC (2012). Гликозилтрансферазы семейства 61 опосредуют перенос арабинофуранозила на ксилан в травах. Proc. Natl. Акад. Sci. США 17, 989–993.
Proc. Natl. Акад. Sci. США 17, 989–993.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Бачич А., Харрис А. Дж. И Стоун Б. А. (1988). «Структура и функция стенок растительных клеток», в The Biochemistry of Plants , ed J.Прейсс (Нью-Йорк: Academic Press), 297–371.
Бэнкс, Дж. А., Нишияма, Т., Хасебе, М., Боуман, Дж. Л., Грибсков, М., деПамфилис, К., Альберт, В. А., Аоно, Н., Аояма, Т., Эмброуз, Б. А., Эштон, Н. В. , Axtell, MJ, Barker, E., Barker, MS, Bennetzen, JL, Bonawitz, ND, Chapple, C., Cheng, C., Correa, LG, Dacre, M., DeBarry, J., Dreyer, I. , Элиас, М., Энгстрем, Э.М., Эстель, М., Фенг, Л., Финет, К., Флойд, С.К., Фроммер, В.Б., Фуджита, Т., Грамцов, Л., Гутенсон, М., Хархольт, Дж., Хаттори, М., Хейл, А., Хираи, Т., Хиваташи, Ю., Исикава, М., Ивата, М., Кароль, К.Г., Келер, Б., Колукисаоглу, У., Кубо, М., Курата, Т., Лалонд, С., Ли, К., Ли, Ю., Литт, А., Лайонс, Э., Мэннинг, Г., Маруяма, Т., Майкл, Т. П., Миками, К., Миядзаки , С. , Моринага, С., Мурата, Т., Мюллер-Робер, Б., Нельсон, Д.Р., Обара, М., Огури, Ю., Олмстед, Р.Г., Онодера, Н., Петерсен, Б.Л., Пилс, Б., Пригге, М., Ренсинг, С.А., Рианьо-Пачон, Д.М., Робертс, А.В., Сато, Ю., Шеллер, Х.В., Шульц, Б., Шульц, К., Шакиров, Е.В., Шибагаки, Н., Шинохара, Н., Шиппен, Д.Э., Соренсен, И., Сотоока, Р., Сугимото, Н., Сугита, М., Сумикава, Н., Танурджич , М., Тайссен, Г., Ульвсков, П., Вакадзуки, С., Вен, Дж. К., Уиллатс, В. В., Випф, Д., Вольф, П. Г., Янг, Л., Циммер, А. Д., Чжу, К., Митрос, Т., Хеллстен, У., Локе, Д., Отиллар, Р., Саламов, А., Шмутц, Дж., Шапиро, Х., Линдквист, Э., Лукас, С., Рохсар, Д., и Григорьев И.В. (2011). Геном Selaginella определяет генетические изменения, связанные с эволюцией сосудистых растений. Наука 332, 960–963.
, Моринага, С., Мурата, Т., Мюллер-Робер, Б., Нельсон, Д.Р., Обара, М., Огури, Ю., Олмстед, Р.Г., Онодера, Н., Петерсен, Б.Л., Пилс, Б., Пригге, М., Ренсинг, С.А., Рианьо-Пачон, Д.М., Робертс, А.В., Сато, Ю., Шеллер, Х.В., Шульц, Б., Шульц, К., Шакиров, Е.В., Шибагаки, Н., Шинохара, Н., Шиппен, Д.Э., Соренсен, И., Сотоока, Р., Сугимото, Н., Сугита, М., Сумикава, Н., Танурджич , М., Тайссен, Г., Ульвсков, П., Вакадзуки, С., Вен, Дж. К., Уиллатс, В. В., Випф, Д., Вольф, П. Г., Янг, Л., Циммер, А. Д., Чжу, К., Митрос, Т., Хеллстен, У., Локе, Д., Отиллар, Р., Саламов, А., Шмутц, Дж., Шапиро, Х., Линдквист, Э., Лукас, С., Рохсар, Д., и Григорьев И.В. (2011). Геном Selaginella определяет генетические изменения, связанные с эволюцией сосудистых растений. Наука 332, 960–963.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Бертон, Р. А., Уилсон, С. М., Хрмова, М., Харви, А. Дж., Ширли, Н. Дж., Медхерст, А., Стоун, Б. А., Ньюбигин, Э. Дж., Бэчич, А. , и Финчер, Г. Б. (2006). Целлюлозосинтазоподобные гены CslF опосредуют синтез (1,3; 1,4) -β- D -глюканов клеточной стенки. Наука 311, 1940–1942.
, и Финчер, Г. Б. (2006). Целлюлозосинтазоподобные гены CslF опосредуют синтез (1,3; 1,4) -β- D -глюканов клеточной стенки. Наука 311, 1940–1942.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Кантарель, Б.Л., Коутиньо, П. М., Ранкурель, К., Бернар, Т., Ломбард, В., и, Хенриссат, Б. (2009). База данных углеводно-активных ферментов (CAZy): экспертный ресурс по гликогеномике. Nucleic Acids Res. 37, 233–238.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Карпита, Н. К., и Жибо, Д. М. (1993). Структурные модели первичных клеточных стенок цветковых растений: соответствие молекулярной структуры физическим свойствам стенок во время роста. Завод J. 3, 1–30.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Дэвис, Л. Дж., Лилли, К. Дж., Нокс, Дж. П., и Урвин, П. Э. (2012). Синцитии, образованные взрослой самкой Heterodera schachtii на корнях Arabidopsis thaliana , имеют отчетливую молекулярную архитектуру клеточной стенки. Новый Фитол . (в прессе).
Новый Фитол . (в прессе).
Доблин, М.С., Петтолино, Ф.А., Уилсон, С.М., Кэмпбелл, Р., Бертон, Р.А., Финчер, Г.Б., Ньюбигин, Э., и Бачич, А. (2009). Ген CSLH, подобный синтазе целлюлозы ячменя, опосредует синтез (1,3; 1,4) -β- D -глюкана в трансгенном Arabidopsis . Proc. Natl. Акад. Sci. США 106, 5996–6001.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Домозич, Д. С., Соренсен, И., Уиллатс, В. Г. Т. (2009). Распределение полимеров клеточной стенки во время развития антеридиев и сперматогенеза у зеленой водоросли Charophycean, Chara corallina . Ann. Бот. 104, 1045–1056.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Эгелунд, Дж., Обель, Н., Ульвсков, П., Геши, Н., Паули, М., Бачич, А., и Петерсен, Б. Л. (2007). Молекулярная характеристика двух мутантов гликозилтрансферазы Arabidopsis thaliana , rra1 и rra2, которые имеют пониженное остаточное содержание арабинозы в полимере, прочно связанном с остатком целлюлозной стенки. Завод Мол. Биол. 64, 439–451.
Завод Мол. Биол. 64, 439–451.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Эстевес, Дж.М., Фернандес, П. В., Касулин, Л., Дюпри, П., и Чиансия, М. (2009). Химическая и in situ характеристика макромолекулярных компонентов клеточных стенок зеленых морских водорослей Codium fragile . Гликобиология 19, 212–228.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Фейнгольд, Д. С., Авигад, Г. (1980). «Трансформации сахарных нуклеотидов в растениях», в Биохимия растений: всеобъемлющий трактат , ред.К. Штумпф и Э. Э. Конн (Нью-Йорк: Academic Press), 101–170.
Фрай, С. К. (2001). Клеточная стенка растущих растений: химический и метаболический анализ , Репринтное издание. Нью-Джерси: The Blackburn Press.
Фрай, С. К. (2004). Первичный метаболизм клеточной стенки: отслеживание карьеры полимеров стенки в живых растительных клетках. New Phytol. 161, 641–675.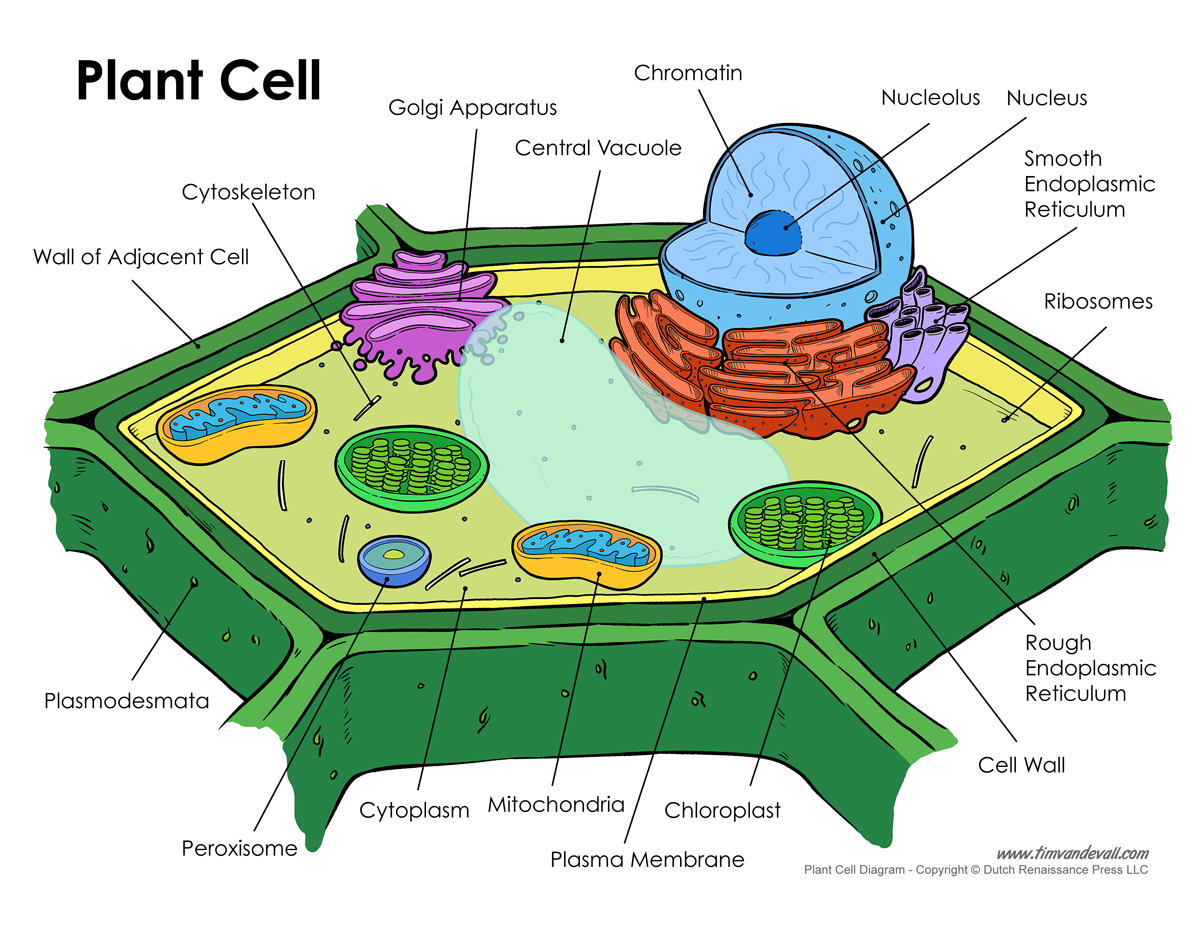
CrossRef Полный текст
Фрай, С. К., Молер, К. Э., Нессельроде, Б. Х. У. А., и Франкова, Л.(2008). Β-глюкан со смешанными связями: эндотрансглюкозилаза ксилоглюкана, новый фермент ремоделирования стенок из хвоща (хвоща) и харофитных водорослей. Завод J. 55, 240–252.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Гейслер-Ли, Дж., Гейслер, М., Коутиньо, П.М., Сегерман, Б., Нишикубо, Н., Такахаши, Дж., Аспеборг, Х., Джерби, С., Мастер, Э., Андерссон-Гуннерас , С., Сундберг, Б., Карпински, С., Тери, Т.Т., Клецковски, Л.А., Хенриссат, Б., и Меллерович, Э.Дж. (2006). Углеводноактивные ферменты тополя. Идентификация генов и анализ экспрессии. Plant Physiol. 140, 946–962.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Губе, Ф., Джексон, П., Дери, М. Дж., И Дюпри, П. (2002). Анализ полисахаридов с использованием электрофореза в углеводном геле: метод изучения полисахаридов и полисахаридгидролаз стенок растительных клеток. Анал. Биохим. 300, 53–68.
Анал. Биохим. 300, 53–68.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Хансен, С.Ф., Хархольт Дж., Оикава А. и Шеллер Х. В. (2012). Растительные гликозилтрансферазы за пределами CAZy: взгляд на семейства DUF. Фронт. Завод Sci . 3:59. DOI: 10.3389 / fpls.2012.00059
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Хархольт, Дж., Соренсен, И., Фангель, Дж. У., Робертс, А., Уиллатс, В. Г. Т., Шеллер, Х. В., Бент Ларсен Петерсен, Б. Л., Бэнкс, Дж. А., и Ульвсков, П. (2012). Репертуар гликозилтрансфераз шиповника Selaginella moellendorffii и сравнительное исследование структуры клеточной стенки. PLoS ONE 7, e35846. DOI: 10.1371 / journal.pone.0035846
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Хеплер, П. К., Фоскет, Д. Э., и Ньюкомб, Э. Х. (1970). Лигнификация во время формирования вторичной стенки в Coleus : исследование с помощью электронной микроскопии. Am. J. Bot. 57, 85–96.
Am. J. Bot. 57, 85–96.
CrossRef Полный текст
Эрве К., Роговски А., Гилберт Х. Дж. И Нокс Дж. П. (2009). Ферментативная обработка показывает разные способности к распознаванию и разложению ксилана в первичных и вторичных стенках растительных клеток. Plant J. 58, 413–422.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Ираки, Н. М., Брессан, Р. А., Хасегава, П. М., и Карпита, Н. К. (1989a). Изменение физической и химической структуры первичной клеточной стенки ограниченных ростом растительных клеток, адаптированных к осмотическому стрессу. Plant Physiol. 91, 39–47.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Ираки, Н. М., Сингх, Н., Брессан, Р. А., и Карпита, Н.С. (1989b). Клеточные стенки клеток табака и изменения в составе, связанные со снижением роста при адаптации к водному и физиологическому стрессу. Plant Physiol. 91, 48–53.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Ираки, Н.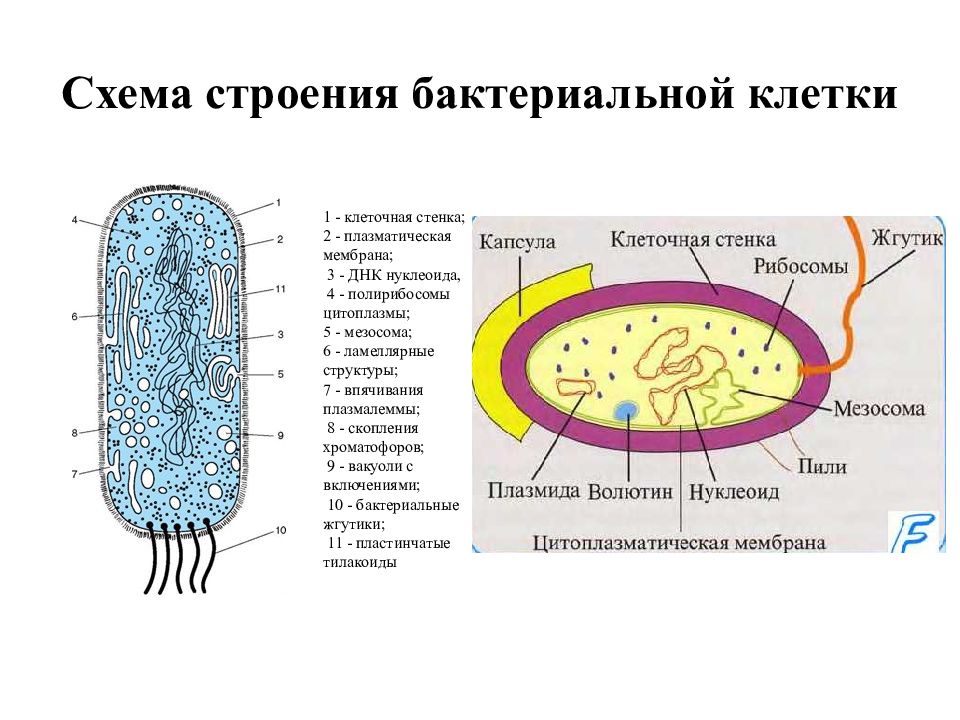 М., Брессан, Р. А., и Карпита, Н. К. (1989c). Внеклеточные полисахариды и белки культур клеток табака и изменения в составе, связанные с ограничивающей рост адаптацией к водному и физиологическому стрессу. Plant Physiol. 91, 54–61.
М., Брессан, Р. А., и Карпита, Н. К. (1989c). Внеклеточные полисахариды и белки культур клеток табака и изменения в составе, связанные с ограничивающей рост адаптацией к водному и физиологическому стрессу. Plant Physiol. 91, 54–61.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Кенрик П. и Крейн П. Р. (1997). Происхождение и ранняя эволюция растений на суше. Природа 389, 33–39.
CrossRef Полный текст
Леруксель, О., Чу, Т. С., Севено, М., Усадель, Б., Фэй, Л., Леруж, П., и Поли, М. (2002). Быстрое структурное фенотипирование мутантов клеточной стенки растений с помощью ферментативного фингерпринтинга олигосахаридов. Plant Physiol. 130, 1754–1763.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Маки У. и Сто, Р. Д. П. Р. (1968). Наличие микрофибрилл маннана в зеленых водорослях Codium fragile и Acetabularia crenulata . Planta 253, 249–253.
CrossRef Полный текст
Маркус, С.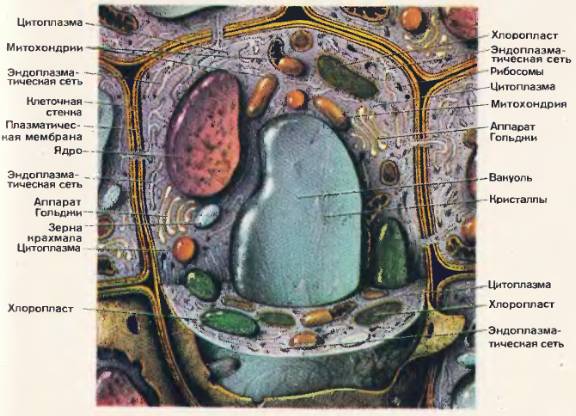 Е., Блейк, А.В., Бенианс, ТАС, Ли, KJD, Пойзер, К., Дональдсон, Л., Леру, О., Роговски, А., Петерсен, Х.Л., Борастон, А., Гилберт, Х.Дж., Уиллатс, В.Г. Т., и Нокс, Дж. П. (2010). Ограниченный доступ белков к полисахаридам маннана в стенках интактных растительных клеток. Завод J. 64, 191–203.
Е., Блейк, А.В., Бенианс, ТАС, Ли, KJD, Пойзер, К., Дональдсон, Л., Леру, О., Роговски, А., Петерсен, Х.Л., Борастон, А., Гилберт, Х.Дж., Уиллатс, В.Г. Т., и Нокс, Дж. П. (2010). Ограниченный доступ белков к полисахаридам маннана в стенках интактных растительных клеток. Завод J. 64, 191–203.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Мацунага, Т., Исии, Т., Мацумото, С., Хигучи, М., Дарвилл, А., Альберсхайм, П., и О’Нил, М.А. (2004). Возникновение полисахарида первичной клеточной стенки рамногалактуронана II у птеридофитов, ликофитов и мохообразных: последствия для эволюции сосудистых растений. Plant Physiol. 134, 339–351.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Митчелл Р. А., Дюпри П. и Шьюри П. Р. (2007). Новый подход к биоинформатике определяет гены-кандидаты для синтеза и ферулоилирования арабиноксилана. Plant Physiol. 144, 43–53.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Моллер И. , Соренсен И., Бернал А. Дж., Блаукопф К., Ли К., Обро Дж., Петтолино Ф., Робертс, А., Миккельсен, Дж. Д., Нокс, Дж. П., Бачич, А., и Уиллатс, В. Г. Т. (2007). Высокопроизводительное картирование полимеров клеточной стенки внутри и между растениями с использованием новых микрочипов. Plant J. 50, 1118–1128.
, Соренсен И., Бернал А. Дж., Блаукопф К., Ли К., Обро Дж., Петтолино Ф., Робертс, А., Миккельсен, Дж. Д., Нокс, Дж. П., Бачич, А., и Уиллатс, В. Г. Т. (2007). Высокопроизводительное картирование полимеров клеточной стенки внутри и между растениями с использованием новых микрочипов. Plant J. 50, 1118–1128.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Mouille, G., Robin, S., Lecomte, M., Pagant, S., and Höfte, H. (2003). Классификация и идентификация мутантов клеточной стенки арабидопсиса с использованием инфракрасной микроскопии с преобразованием Фурье (FT-IR). Plant J. 35, 393–404.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Никлас, К. Дж. (2004). Стенки клеток, скрепляющие древо жизни. BioScience 54, 831–841.
CrossRef Полный текст
Парк С., Шумлански А. Л., Гу Ф., Го Ф. и Нильсен Э. (2011). Роль CSLD3 во время синтеза клеточной стенки в апикальных плазматических мембранах растущих на кончике корневых волосковых клеток.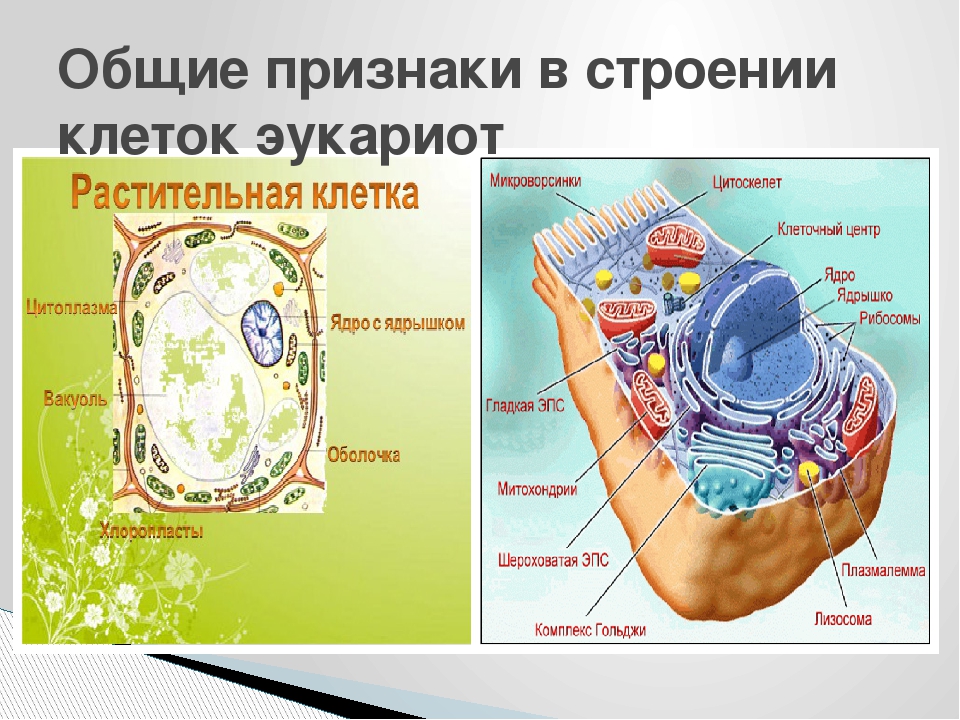 Nat. Клетка. Биол. 13, 973–980.
Nat. Клетка. Биол. 13, 973–980.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Паттатил, С., Авчи, У., Болдуин, Д., Свеннес, А.Г., МакГилл, Дж. А., Поппер, З., Буттен, Т., Альберт, А., Дэвис, Р., Ченнаредди, К., Донг, Р., О ‘ Ши, Б., Росси, Р., Лефф, К., Фрешур, Г., Нарра, Р., О’Нил, М., Йорк, WS, и Хан, М.Г. (2010). Комплексный набор моноклональных антител, направленных на гликаны клеточной стенки растений. Plant Physiol. 153, 514–525.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Поппер, З.А., Мишель, Г., Эрве, К., Домозич, Д.С., Уиллатс, В. Г., Туохи, М. Г., Клоарег, Б., и Стенгель, Д. Б. (2011). Эволюция и разнообразие клеточных стенок растений: от водорослей до цветковых растений. Annu. Преподобный завод. Биол. 62, 567–590.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Qu, Y., Egelund, J., Gilson, P.R., Houghton, F., Gleeson, P.A., Schultz, C.J., и Bacic, A.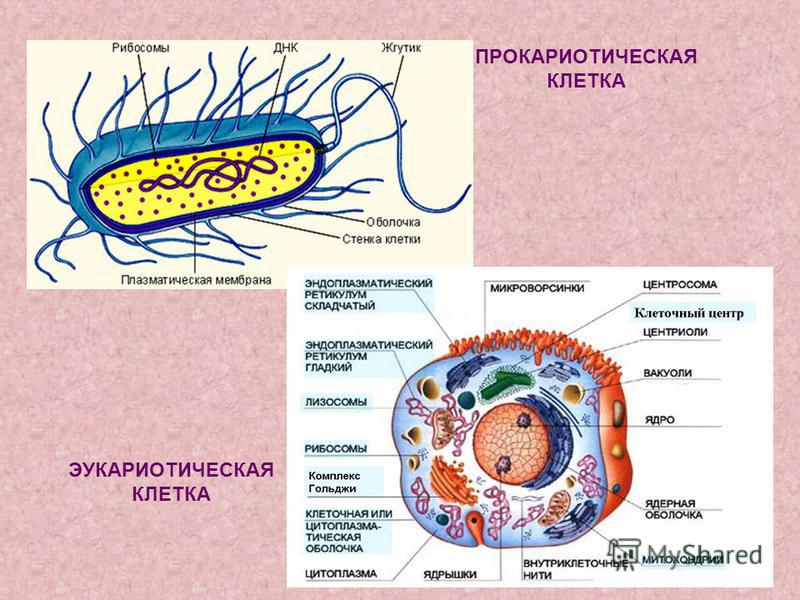 (2004). Идентификация новой группы предполагаемых Arabidopsis thaliana β- (1,3) -галактозилтрансфераз. Завод Мол. Биол. 68, 43–59.
(2004). Идентификация новой группы предполагаемых Arabidopsis thaliana β- (1,3) -галактозилтрансфераз. Завод Мол. Биол. 68, 43–59.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Scherp, P., Grotha, R., and Kutschera, U. (2001). Встречаемость и филогенетическое значение каллозы, связанной с цитокинезом, у зеленых водорослей, мохообразных, папоротников и семенных растений. Отчеты о растительных клетках 20, 143–149.
CrossRef Полный текст
Соренсен, И., Петтолино, Ф. А., Бачич, А., Ральф, Дж., Лу, Ф., О’Нил, М. А., Фей, З., Роуз, Дж. К. К., Домозыч, Д. С., Уиллатс, В. Г. Т. (2011). Зеленые водоросли charophycean дают представление о раннем происхождении стенок растительных клеток. Plant J. 68, 201–211.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Spatz, H.-C., Rowe, N., Speck, T., and Daviero, V. (1998). Биомеханика полостебельных сфенопсид: II. Каламиты — иметь или не иметь вторичную ксилему. Rev. Palaeobot. Палинол. 102, 63–77.
Rev. Palaeobot. Палинол. 102, 63–77.
CrossRef Полный текст
Тимме Р.Э., и Делвиче, К. Ф. (2010). Выявление эволюционного происхождения молекулярных процессов растений: сравнение транскриптомов Coleochaete (Coleochaetales) и Spirogyra (Zygnematales). BMC Plant Biol. 10, 96. DOI: 10.1186 / 1471-2229-10-96
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Цекос И. (1999). Сайты синтеза целлюлозы у водорослей: разнообразие и эволюция целлюлозосинтезирующих ферментных комплексов. Дж.Phycol. 35, 635–655.
CrossRef Полный текст
Веласкес, С.М., Рикарди, М.М., Дорош, Дж.Г., Фернандес, П.В., Надра, А.Д., Поль-Фачин, Л., Эгелунд, Дж., Гилле, С., Хархольт, Дж., Чианция, М., Верли, Х., Паули, М., Бачич, А., Олсен, К.Е., Ульвсков, П., Петерсен, Б.Л., Сомервилл, К., Юсем, Н.Д., и Эстевес, Дж. М. . (2011). O -Гликозилированные белки клеточной стенки необходимы для роста корневых волосков. Наука 332, 1401–1403.
Наука 332, 1401–1403.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Verhertbruggen, Y., Маркус, С. Е., Хэгер, А., Верхоф, Р., Шолс, Х. А., Макклири, Б. В., Макки, Л., Гилберт, Х. Дж., И Нокс, Дж. П. (2009). Сложность развития полисахаридов арабинана и их переработка в стенках клеток растений. Плант Дж. 59, 413–425.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Фогель, Дж. П., Гарвин, Д. Ф., Моклер, Т. К., Шмутц, Дж., Рохсар, Д., Беван, М. В., Барри, К., Лукас, С., Хармон-Смит, М., Лейл, К., Тайс , Х., Шмутц, Дж., Гримвуд, Дж., Маккензи, Н., Беван, М.В., Хо, Н., Гу, YQ, Лазо, Г.Р., Андерсон, О.Д., Фогель, Дж. П., Ю, Ф.М., Луо, М.С., Дворжак, Дж., Райт, Дж., Фебрер , М., Беван, М.В., Идзяк, Д., Хастерок, Р., Гарвин, Д.Ф., Линдквист, Э., Ван, М., Фокс, С.Е., Прист, HD, Филичкин, С.А., Гиван, С.А., Брайант, Д.У., Чанг, Д.Х., Моклер, ТК, Ву, Х., Ву, В., Ся, А.П., Шнабл, П.С., Кальянараман, А., Барбазук, Б. , Майкл, Т.П., Хазен, С.П., Брэгг, Дж. Лауденсия-Чингкуанко, Д., Фогель, JP, Гарвин, Д.Ф., Вен, Ю., Маккензи, Н., Беван, М. В., Хаберер, Г., Спаннагл, М., Майер, К., Раттей, Т., Митрос, Т., Рохсар, Д., Ли, С. Дж. , Роуз, Дж. К., Мюллер, Л. А., Йорк, Т. Л., Уикер, Т., Бухманн, Дж. П., Тансканен, Дж., Шульман, А. Х., Гундлах, Х., Райт, Дж., Беван, М., де Оливейра, А. К. , Maia Lda, C., Belknap, W., Gu, YQ, Jiang, N., Lai. J., Zhu, L., Ma, J., Sun, C., Pritham, E., Salse, J., Murat, F., Abrouk, M., Haberer. Г., Spannagl. М., Майер, К., Бруггманн, Р., Мессинг, Дж., Ю, Ф. М., Луо, М.К., Дворжак, Дж., Фальгрен, Н., Фокс, С. Е., Салливан, К. М., Моклер, Т. К., Кэррингтон, Дж. К., Чепмен, Э. Дж., Мэй, Г. Д., Чжай, Дж., Гансманн, М., Гуразада, С. Г. , Герман, М., Мейерс, BC, Грин, П.Дж., Брэгг, Дж. Н., Тайлер, Л., Ву, Дж., Гу, YQ, Лазо, Г.Р., Лауденсия-Чингкуанко, Д., Томсон, Дж., Фогель, JP, Hazen, SP, Chen, S., Scheller, HV, Harholt, J., Ulvskov, P., Fox, SE, Filichkin, SA, Fahlgren, N., Kimbrel, JA, Chang, JH, Sullivan, CM, Чепмен, Э.
, Майкл, Т.П., Хазен, С.П., Брэгг, Дж. Лауденсия-Чингкуанко, Д., Фогель, JP, Гарвин, Д.Ф., Вен, Ю., Маккензи, Н., Беван, М. В., Хаберер, Г., Спаннагл, М., Майер, К., Раттей, Т., Митрос, Т., Рохсар, Д., Ли, С. Дж. , Роуз, Дж. К., Мюллер, Л. А., Йорк, Т. Л., Уикер, Т., Бухманн, Дж. П., Тансканен, Дж., Шульман, А. Х., Гундлах, Х., Райт, Дж., Беван, М., де Оливейра, А. К. , Maia Lda, C., Belknap, W., Gu, YQ, Jiang, N., Lai. J., Zhu, L., Ma, J., Sun, C., Pritham, E., Salse, J., Murat, F., Abrouk, M., Haberer. Г., Spannagl. М., Майер, К., Бруггманн, Р., Мессинг, Дж., Ю, Ф. М., Луо, М.К., Дворжак, Дж., Фальгрен, Н., Фокс, С. Е., Салливан, К. М., Моклер, Т. К., Кэррингтон, Дж. К., Чепмен, Э. Дж., Мэй, Г. Д., Чжай, Дж., Гансманн, М., Гуразада, С. Г. , Герман, М., Мейерс, BC, Грин, П.Дж., Брэгг, Дж. Н., Тайлер, Л., Ву, Дж., Гу, YQ, Лазо, Г.Р., Лауденсия-Чингкуанко, Д., Томсон, Дж., Фогель, JP, Hazen, SP, Chen, S., Scheller, HV, Harholt, J., Ulvskov, P., Fox, SE, Filichkin, SA, Fahlgren, N., Kimbrel, JA, Chang, JH, Sullivan, CM, Чепмен, Э. Дж., Кэррингтон, Дж.К., Моклер, Т.С., Бартли, Л.Е., Цао, П., Юнг, К.Х., Шарма, М.К., Вега-Санчес, М., Рональд, П., Дардик, С.Д., Де Бодт, С., Верелст, В. , Inzé, D., Heese, M., Schnittger, A., Yang, X., Kalluri, UC, Tuskan, GA, Hua, Z., Vierstra, RD, Garvin, DF, Cui, Y., Ouyang, S ., Sun, Q., Liu, Z., Yilmaz, A., Grotewold, E., Sibout, R., Hematy, K., Mouille, G., Höfte, H., Michael, T., Pelloux, J ., О’Коннор, Д., Шнабл, Дж., Роу, С., Хармон, Ф., Касс, К.Л., Седбрук, Дж. К., Бирн, М., Уолш, С., Хиггинс, Дж., Беван, М., Ли, П., Брутнелл, Т., Унвер, Т., Будак, Х., Белкрэм, Х., Чарльз, М., Чалхуб, Б., и Бакстер, I. . (2010). Секвенирование и анализ генома модельной травы Brachypodium distachyon . Природа 463, 763–768.
Дж., Кэррингтон, Дж.К., Моклер, Т.С., Бартли, Л.Е., Цао, П., Юнг, К.Х., Шарма, М.К., Вега-Санчес, М., Рональд, П., Дардик, С.Д., Де Бодт, С., Верелст, В. , Inzé, D., Heese, M., Schnittger, A., Yang, X., Kalluri, UC, Tuskan, GA, Hua, Z., Vierstra, RD, Garvin, DF, Cui, Y., Ouyang, S ., Sun, Q., Liu, Z., Yilmaz, A., Grotewold, E., Sibout, R., Hematy, K., Mouille, G., Höfte, H., Michael, T., Pelloux, J ., О’Коннор, Д., Шнабл, Дж., Роу, С., Хармон, Ф., Касс, К.Л., Седбрук, Дж. К., Бирн, М., Уолш, С., Хиггинс, Дж., Беван, М., Ли, П., Брутнелл, Т., Унвер, Т., Будак, Х., Белкрэм, Х., Чарльз, М., Чалхуб, Б., и Бакстер, I. . (2010). Секвенирование и анализ генома модельной травы Brachypodium distachyon . Природа 463, 763–768.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Вриланд В., Сломич М. и Летч В. М. (1984). Моноклональные антитела как молекулярные зонды для антигенов клеточной стенки бурой водоросли Fucus . Planta 163, 506–517.
CrossRef Полный текст
Willats, W. G. T., Orfila, C., Limberg, G., Buchholt, H. C., van Alebeek, G.-J. В. М., Вораген, А. Г. Дж., Маркус, С. Е., Кристенсен, Т. М. И., Миккельсен, Дж. Д., Мюррей, Б. С. и Нокс, Дж. П. (2001). Модуляция степени и характера метилэстерификации пектинового гомогалактуронана в стенках растительных клеток: последствия для действия пектинметилэстеразы, свойств матрикса и клеточной адгезии. J. Biol. Chem. 276, 19404–19413.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Водниок, С., Бринкманн, Х., Глёкнер, Г., Хайдель, А. Дж., Филипп, Х., Мелконян, М., и Беккер, Б. (2011). Происхождение наземных растений: решает ли спряжение зеленых водорослей? BMC Evol. Биол. 11, 104. DOI: 10.1186 / 1471-2148-11-104
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Инь, Ю., Чен, Х., Хан, М. Г., Монен, Д., и Сюй, Ю. (2010). Эволюция и функция семейства гликозилтрансфераз, связанных с синтезом клеточной стенки растений 8. Plant Physiol .153, 1729–1746.
Plant Physiol .153, 1729–1746.
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Есть ли у клеток животных клеточная мембрана? — Видео и стенограмма урока
Функция мембраны клеток животных
Итак, что же на самом деле делает мембрана клеток животных? Итак, в нашем первоначальном определении говорилось, что клеточная мембрана полупроницаема и окружает клетку. Его полупроницаемая природа означает, что мембрана может выполнять две основные функции:
- Формирование внешней границы клетки и, следовательно, удерживание клетки вместе.
- Позволяет одним веществам проходить через него, запрещая это другим.
Первая из этих функций довольно проста. Точно так же, как водный баллон удерживает воду внутри, клеточная мембрана удерживает внутри все клеточные компоненты (в этом нет ничего удивительного). Итак, вместо того, чтобы сосредотачиваться на очевидной функции, давайте посмотрим на вторую задачу: контроль входа и выхода из ячейки. Контролируя то, что попадает в клетку и выходит из нее, мембрана действительно действует как регулирующая структура.Примеры веществ, которые он регулирует, включают (но не ограничиваются ими) натрий, калий, сахар и аминокислоты. Каждый из них аналогичным образом взаимодействует с мембраной клетки животных.
Контроль входа и выхода
Как клеточная мембрана контролирует то, что входит и выходит из клетки? Чтобы это выяснить, мы должны сначала понять, из чего сделана мембрана.
T
На его рисунке изображено поперечное сечение мембраны клетки животных.Обратите особое внимание на зеленый канал и белки-переносчики, проходящие через мембрану. Белки — это структуры внутри клеточной мембраны, которые позволяют веществам входить или выходить из клетки. Думайте о них как о туннелях, соединяющих внутреннюю часть клетки с внешней средой. Разные молекулы будут использовать разные туннели (белки) для пересечения мембраны. Другим молекулам будет отказано в доступе, потому что для них нет белка. Например, бактерии не могут просто проникнуть внутрь клетки, потому что для нее нет отверстия.Именно благодаря этим белкам мембрана клетки животных способна регулировать вход и выход из клетки.
Краткое содержание урока
Хорошо, давайте сделаем пару минут, чтобы повторить то, что мы узнали. Клетки животных, например клетки вашего тела, содержат клеточную мембрану , которая образует внешнюю часть клетки. Клеточная мембрана полупроницаемая , что означает, что она пропускает только определенные предметы. Эта мембрана выполняет две жизненно важные функции: она создает внешнюю границу клетки и удерживает клетку вместе, а также контролирует вход и выход материала из клетки.Мембрана животной клетки способна выполнять эту вторую задачу благодаря встроенным в нее белкам. Важно помнить, что белков — это структуры внутри клеточной мембраны, которые позволяют веществам входить или выходить из клетки.
Biology4Kids.com: Структура клетки: клеточные стенки
Клеточные мембраны окружают каждую клетку, которую вы будете изучать. Клеточные стенки из целлюлозы встречаются только вокруг растительных клеток и некоторых других организмов. Целлюлоза — это специальный сахар, который классифицируется как структурный углевод и не используется для получения энергии. Если растительная клетка похожа на воздушный шар, клеточная стенка похожа на картонную коробку, которая защищает воздушный шар. Воздушный шар защищен от внешнего мира структурой, обеспечивающей защиту и поддержку.
Хотя многие сахара, такие как глюкоза, могут растворяться в воде (H 2 0), целлюлоза не растворяется в воде и может образовывать длинные цепочки, поддерживающие растения. Когда вы едите растительный материал, вы даже не можете переваривать и расщеплять целлюлозу для получения энергии.У коров и других травоядных есть особые бактерии в желудках, которые переваривают полимеры целлюлозы .
Хотя клеточные стенки защищают клетки, они также позволяют растениям расти до больших высот. У вас есть скелет, который вас поддерживает. Красное дерево высотой 100 футов — нет. Он использует прочные клеточные стенки для поддержания своей формы. Для общей поддержки, плотные клетки в сердцевине ствола могут позволить дереву вырасти очень высоко. Стенки клеток слегка эластичны для более мелких растений, листьев и тонких веток.Ветер может толкать их из стороны в сторону, и они отскакивают назад. Большие секвойи нуждаются в прочности при сильном ветре и очень мало раскачиваются (кроме вершины).
Клеточная стенка — это не непроницаемая крепость вокруг нежной растительной клетки. В клеточных стенках между растительными клетками есть небольшие отверстия, называемые Plasmodesmata . Клеточные мембраны соседних клеток могут соединяться через эти отверстия. Связи позволяют переносить питательные вещества, отходы и ионы (симпластические пути).Молекулы также могут проходить через пространства внутри клеточных стенок, полностью избегая клеток (апопластические пути).Здорово, что питательные вещества могут переходить от клетки к клетке, но есть проблема и со всеми отверстиями. Клетки могут терять воду. Растения теряют большое количество воды в середине дня или в очень жаркие дни. Когда воздух нагревается и давление водяного пара уменьшается, растения теряют воду в процессе транспирации. Вода выходит через поры на поверхности растения, называемые устьицами.Даже когда клетки растений теряют воду, их основная форма сохраняется за счет клеточных стенок. Растение может поникнуть или увядать, но оно может восстановиться, когда вода вернется в систему. Он будет выглядеть так же, как и при запуске.
Вы можете слышать о клеточных стенках у других видов. Бактерии имеют структуру, называемую клеточной стенкой. У грибов и некоторых простейших также есть клеточные стенки. Они не такие, как стенки растительных клеток из целлюлозы. Другие стенки могут быть сделаны из белков или вещества, называемого хитином. Хитин — еще один структурный углевод. Все они служат одной цели — защите и поддержанию структуры, но это очень разные молекулы.Клеточные стенки (видео Общества Макса Планка)
Полезные справочные ссылки
Encyclopedia.com:http://www.encyclopedia.com/topic/Cell_Wall.aspx
Википедия:
http: // en.wikipedia.org/wiki/Cell_wall
Британская энциклопедия:
http://www.britannica.com/EBchecked/topic/101396/cell/37446/The-plant-cell-wall?anchor=ref313833
Понимание клеточной мембраны и клеточной стенки
Если вы считаете, что контент, доступный через Веб-сайт (как определено в наших Условиях обслуживания), нарушает одно или больше ваших авторских прав, сообщите нам, отправив письменное уведомление («Уведомление о нарушении»), содержащее в информацию, описанную ниже, назначенному ниже агенту.Если репетиторы университета предпримут действия в ответ на ан Уведомление о нарушении, оно предпримет добросовестную попытку связаться со стороной, которая предоставила такой контент средствами самого последнего адреса электронной почты, если таковой имеется, предоставленного такой стороной Varsity Tutors.
Ваше Уведомление о нарушении прав может быть отправлено стороне, предоставившей доступ к контенту, или третьим лицам, таким как в виде ChillingEffects.org.
Обратите внимание, что вы будете нести ответственность за ущерб (включая расходы и гонорары адвокатам), если вы существенно искажать информацию о том, что продукт или действие нарушает ваши авторские права.Таким образом, если вы не уверены, что контент находится на Веб-сайте или по ссылке с него нарушает ваши авторские права, вам следует сначала обратиться к юристу.
Чтобы отправить уведомление, выполните следующие действия:
Вы должны включить следующее:
Физическая или электронная подпись правообладателя или лица, уполномоченного действовать от их имени; Идентификация авторских прав, которые, как утверждается, были нарушены; Описание характера и точного местонахождения контента, который, по вашему мнению, нарушает ваши авторские права, в \ достаточно подробностей, чтобы позволить репетиторам Varsity найти и точно идентифицировать этот контент; например нам требуется а ссылка на конкретный вопрос (а не только на название вопроса), который содержит содержание и описание к какой конкретной части вопроса — изображению, ссылке, тексту и т. д. — относится ваша жалоба; Ваше имя, адрес, номер телефона и адрес электронной почты; и Ваше заявление: (а) вы добросовестно считаете, что использование контента, который, по вашему утверждению, нарушает ваши авторские права не разрешены законом, владельцем авторских прав или его агентом; (б) что все информация, содержащаяся в вашем Уведомлении о нарушении, является точной, и (c) под страхом наказания за лжесвидетельство вы либо владелец авторских прав, либо лицо, уполномоченное действовать от их имени.
Отправьте жалобу нашему уполномоченному агенту по адресу:
Чарльз Кон
Varsity Tutors LLC
101 S. Hanley Rd, Suite 300
St. Louis, MO 63105
Или заполните форму ниже:
Из чего состоит клеточная стенка и как ее разрушить?
Основное различие между растительными и животными клетками состоит в том, что у растительных клеток есть клеточная стенка и клеточная мембрана, а у животных клеток есть только клеточная мембрана.Щелкните здесь для быстрого визуального сравнения клеток растений и животных.
Клеточная стенка расположена вне клеточной мембраны у растений, бактерий, грибов и водорослей. Это прочный жесткий слой, обеспечивающий структурную поддержку и защиту ячейки. Многие клетки растений имеют как первичную клеточную стенку, которая вмещает клетку по мере ее роста, так и вторичную клеточную стенку, которая развивается внутри первичной стенки после того, как клетка перестает расти. Вторичные клеточные стенки намного толще и прочнее, чем первичные клеточные стенки, и являются основным компонентом древесины.
Стенка растительной клетки состоит из целлюлозы, сложного углевода, состоящего из нескольких тысяч молекул глюкозы, соединенных встык. Кроме того, клеточная стенка содержит две группы разветвленных полисахаридов, пектины и сшивающие гликаны.
Разрушение клеточных стенок
Не все методы лизиса клеток подходят для лизиса растительных клеток. Ультразвуковые гомогенизаторы или ультразвуковые аппараты, например, непригодны, поскольку клеточные стенки гораздо более устойчивы к ультразвуковой обработке, чем клеточные мембраны.Органические растворители, такие как спирты, эфир или хлороформ, могут разрушать клетки, проникая через клеточные стенки и мембраны. Эти растворители часто используются в сочетании со сдвигающими силами для лизиса растительных клеток.
Гомогенизаторы высокого давления для лизиса растительных клеток Гомогенизаторы высокого давления
DeBEE используют такие силы, как сдвиг, кавитация и удар, для разрушения клеток растений. Эти силы регулируются, что позволяет пользователю оптимизировать разрыв ячейки в соответствии с конкретным типом ячейки.Оператор может лизировать даже самые сложные клетки, такие как дрожжи или грибы. Другие клетки, такие как кишечная палочка, бактерии, ткани млекопитающих и клетки насекомых, могут быть разрушены с помощью наиболее эффективного процесса для наивысшей скорости восстановления.
