
Урок на тему «Брюхоногие и Двустворчатые моллюски»
Тема урока: Брюхоногие и Двустворчатые моллюски.
Цель образовательная: сформировать знания об особенностях строения и жизнедеятельности двустворчатых и брюхоногих моллюсков.
Воспитательная: продолжить формирование научного мировоззрения учащихся. Сформировать у учащихся ответственное и бережное отношение к природе. Реализовать задачи эстетического воспитания.
Развивающая: развитие памяти, логического мышления, научно-биологической речи учащихся. Формирование специальных умений и навыков. Формирование и развитие познавательного интереса учащихся к биологии
Тип урока: комбинированный.
Методы обучения: словесные ( рассказ, беседа), наглядные.
Средства обучения: учебник, таблица.
Ход урока
1.Организационный этап урока.
Вступительное
слово учителя.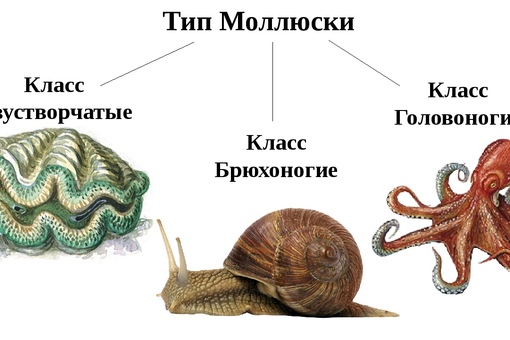
2.Проверка домашнего задания.
1. Назовите общие признаки моллюсков.
2. Каковы особенности внутреннего строения моллюсок?
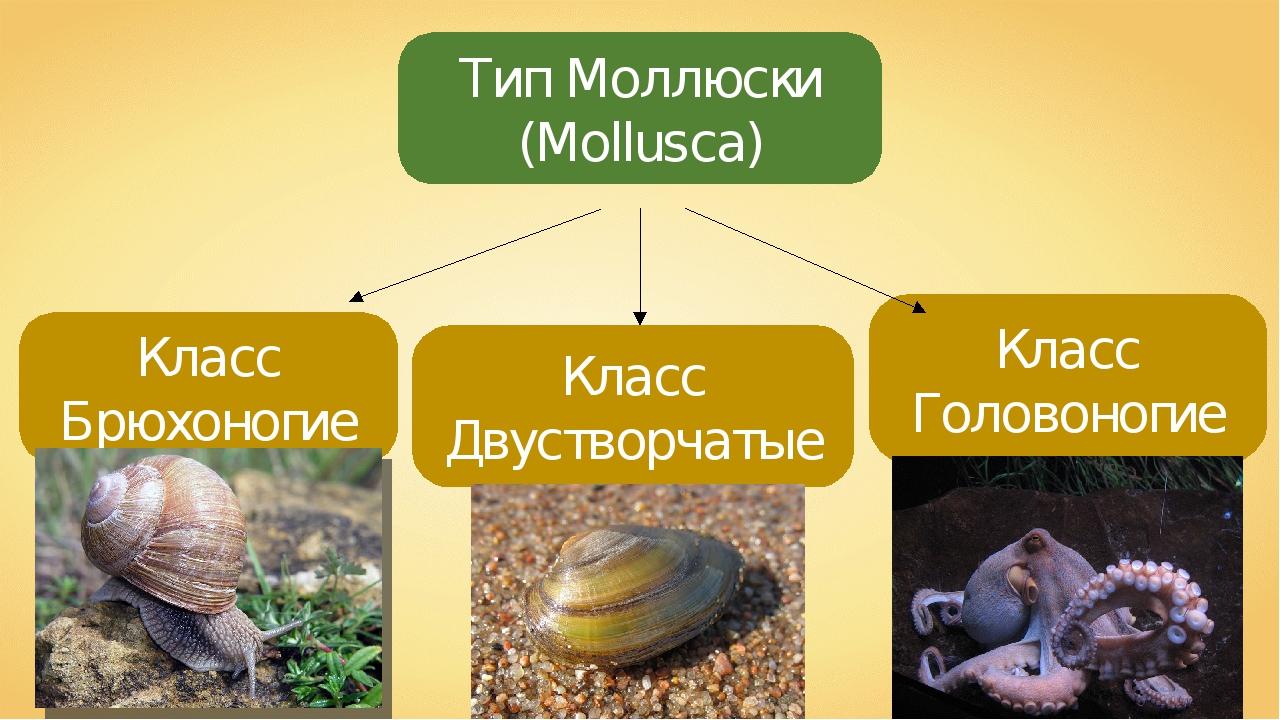
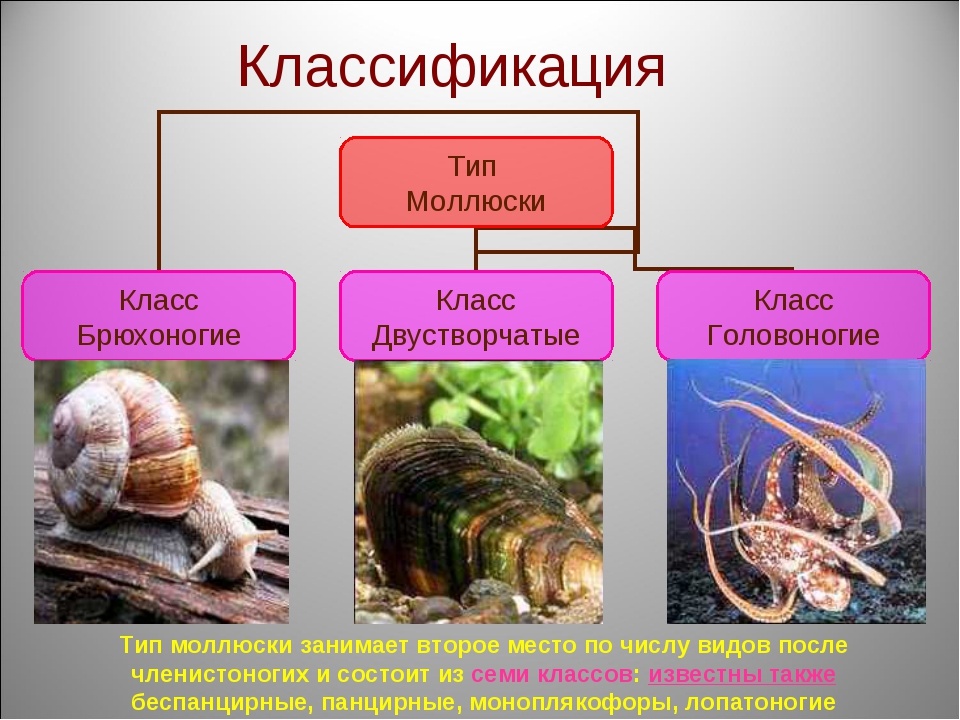
3. Какие выделяют классы моллюсок?
3.Мотивация учебной деятельности.
Моллюски, или мягкотелые, образуют обособленный тип беспозвоночных животных, характерной особенностью которых является наличие кожной складки – мантии, ведущих свое начало от древних неспециализированных многощетинковых червей.
Количество видов достигает 130 тысяч, выделяют семь классов, мы познакомимся только с тремя из них: Брюхоногие, Двустворчатые, Головоногие.
Формирование темы и целей урока.
4.Изучение нового материала.
Брюхоногие (слайд) обладают мощной
“ногой” с широкой подошвой для ползания; выделяемая при этом слизь облегчает
перемещение по жесткому субстрату. На хорошо выраженной голове имеются
подвижные выросты, способные вытягиваться, сокращаться и ощупывать
встречающиеся на пути предметы, а также глаза на одной из пар щупалец.
Внешнее строение.
Большинство Брюхоногих моллюсков имеет спирально закрученную раковину. У некоторых раковина недоразвита или полностью отсутствует (например, у голых слизней).
Тело состоит из трёх отделов: головы, туловища и ноги.
На голове расположены одна или две пары длинных мягких щупалец и пара глаз.
В туловище — внутренние органы.
Нога брюхоногих приспособлена к ползанию и представляет собой мускулистый вырост брюшной части тела (отсюда и название класса).
Среди брюхоногих есть фильтраторы, а также паразиты и хищники. Но в основном эти моллюски растительноядные или поедающие разлагающиеся донные отложения.
Прудовик
обыкновенный — обитает в пресных водоёмах и на мелководьях рек по всей
территории России. Питается растительной пищей, соскабливая тёркой мягкие ткани
растений.
Пищеварительная система.
В ротовой полости брюхоногих моллюсков находится мускулистый язык с хитиновыми зубами, образующими «тёрку» (или радулу). У растительноядных моллюсков тёрка (радула) служит для соскабливания растительной пищи, у хищных — помогает удерживать добычу.
В ротовую полость обычно открываются слюнные железы.
Ротовая полость переходит в глотку, а затем — в пищевод, который ведёт в желудок и кишечник. В него впадают протоки пищеварительной железы
. Непереваренные остатки пищи выбрасываются через анальное отверстие.Нервная система.
Нервная система (на рисунке показана жёлтым цветом) состоит из нескольких пар хорошо развитых нервных узлов, расположенных в разных частях тела, и отходящих от них нервов.
У
Брюхоногих моллюсков развиты органы чувств, они расположены в основном на
голове: глаза, щупальца — органы осязания, органы равновесия.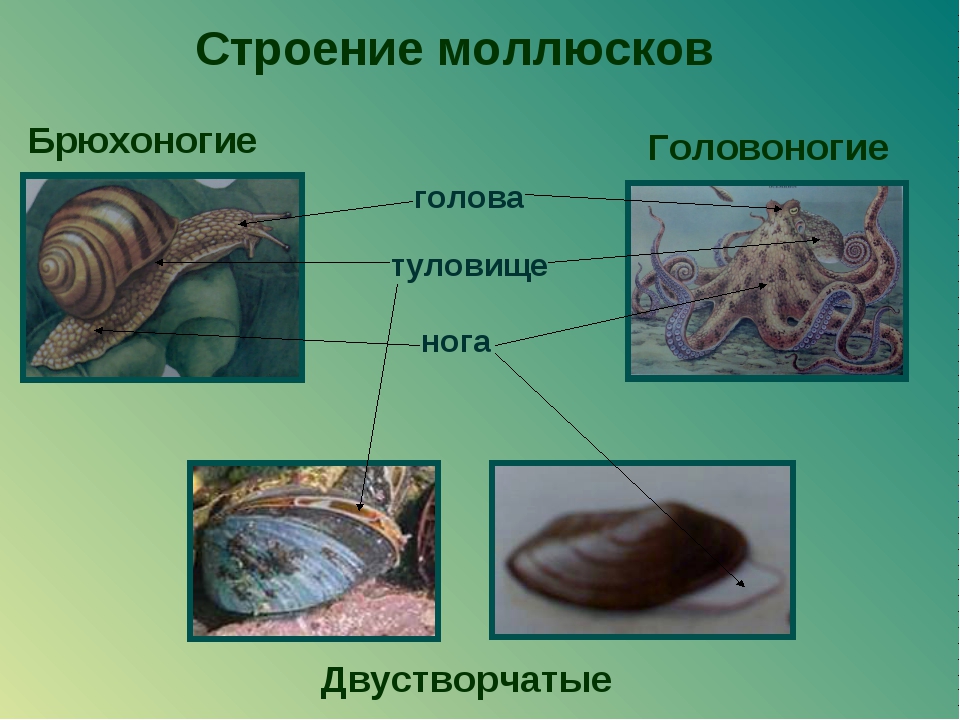 У
брюхоногих хорошо развиты органы обоняния — они могут
распознавать запахи.
У
брюхоногих хорошо развиты органы обоняния — они могут
распознавать запахи.
Кровеносная система.
Брюхоногие моллюски имеют незамкнутую кровеносную систему, состоящую из сердца и сосудов. Сердце состоит из двух камер: желудочка и предсердия.
Дыхание.
Дыхание у моллюсков, живущих в воде, осуществляется жабрами, а у наземных — с помощью
В мантийной полости у большинства водных брюхоногих моллюсков имеются одна или, реже, две жабры.
У прудовиков, катушек, виноградных улиток мантийная полость исполняет роль лёгкого. Кислород из атмосферного воздуха, заполняющего «лёгкое», проникает через стенку мантии в разветвлённые в ней кровеносные сосуды, а углекислый газ из кровеносных сосудов поступает в полость «лёгкого» и выходит наружу.
Выделительная система.
Органы выделения моллюсков — одна или две почки.
Ненужные
для организма продукты обмена веществ поступают из крови в почку, проток от
которой открывается в мантийную полость.
Освобождение крови от углекислого газа и обогащение кислородом происходит в органах дыхания (в жабрах или в лёгком).
Размножение.
Моллюски размножаются только половым путём.
Прудовики, катушки, слизни — гермафродиты.
Оплодотворённые яйца они обычно откладывают на листья растений и различные водные предметы или между комочками почвы. Из яиц выходят маленькие улитки.
Многие морские Брюхоногие — раздельнополые животные, они развиваются с личиночной стадией — парусником.
Двустворчатые (слайд ) моллюски надежно защищены прочными половинками раковин, удерживаемых мощным мускульным замком. “Нога” у большинства видов сплюснута с боков и хорошо приспособлена для рытья канавок на илистом или песчаном дне (полезно спросить учащихся – доводилось ли им видеть бороздки на поверхности прибрежного дна реки или озера?).
Класс
Двустворчатые моллюски насчитывает приблизительно 20 тысяч видов.
Представителей, которых мы сегодня с вами рассмотрим – это Беззубка и мидия.
Внешнее строение: туловище, нога. В отличии от брюхоногих, у двустворчатых отсутствует голова. Тело двусторонне симметричное. Раковина состоит из двух створок, которые скреплены биссусом.
Давайте запишем в тетрадях:
Биссус – это прочные белковые нити, с помощью которых моллюск прикрепляется к камням.
Тонкие листки мантии, свешиваясь с боков тела, прилегают к раковине, как подкладка одежды. Между ними расположена мантийная полость, в которой лежат пластинчатые жабры и нога. Нога двустворчатых имеет вид киля или клина (отсюда одно из названий класса — киленогие) и служит в основном для рытья песка или ила. Ползают они очень медленно, а иные стали сидячими. У таких нога редуцируется, как у мидий, или исчезает совсем, как у устриц.
Пищеварительная
система: Т. к. отсутствует голова, то
исчезают слюнные железы, радула, глотка (у Брюхоногих они имеются).
Давайте запишем: сифоны – это трубки на заднем конце тела моллюска, образованные мантией.
В это время она отдает кислород, насыщается углекислым газом, а пищевые частицы, предварительно склеенные слизью, поступают, захваченные ротовыми лопастями, в ротовое отверстие. Устрица средних размеров пропускает за 1 ч через свою мантийную полость до 10 л воды. Размножаясь во множестве, ракушки становятся мощным фактором очистки водоемов.
Дыхательная
система: пластинчатые жабры, снабжённые капиллярами.
Кровеносная система: также как и у брюхоногих незамкнутая. У нее любопытная особенность: сердце, состоящее из желудочка и двух предсердий, как бы наложено на кишечник.
Органы выделения: парные почки
Нервная система: три пары ганглиев (нервных узлов)
Органы чувств: химические органы чувств, органы равновесия, глаза, органы осязания.
Размножение: раздельнополые животные. Оплодотворение внутреннее – в мантийной полости.
|
Сравниваемые признаки |
Классы | |
|
Брюхоногие |
Двустворчатые |
|
|
Симметрия тела |
Ассиметричны |
Двусторонне-симметричны |
|
Части тела |
Голова, нога, туловище |
Туловище, нога |
|
Раковина |
Спирально закрученная |
Две створки |
|
Пищеварительная система |
Рот, глотка, радула,
пищевод, желудок, печень, слюнные железы, кишка, анальное отверстие. |
|
|
Органы дыхания |
Лёгкие |
Пластинчатые жабры |
|
Кровеносная система |
Незамкнутая, 1 предсердие, 1 желудочек. |
Незамкнутая, 2 предсердия, 1 желудочек. |
|
Выделительная система |
Левая почка |
Парные почки |
|
Нервная система |
Разбросанно-узлового типа. Перекрёст между нервными узлами. |
3 пары нервных узлов — ганглиев |
|
Размножение |
Гермафродиты. Личинка
парусник. |
Раздельнополые.Личинка парусник. Оплодотворение внутренне- в мантийной полости |

 Оплодотворение внутреннее, перекрёстное.
Оплодотворение внутреннее, перекрёстное.
Интересные факты: 1. Самым крупным беспозвоночным животным является гигантский кальмар из рода архитевтис. На берегу бухты Лайалл в Новой Зеландии штормом был выброшен кальмар длиной 17,37 м. Два самых длинных его щупальца были по15м. Вес таких кальмаров превышает 2т.
2. Самые большие глаза имеют гигантские кальмары, обитающие в Атлантике. Диаметр глаза может достигать 40см.
3. Самый крупный осьминог принадлежит к виду Октопус апполион. Обитает в Тихом океане. Вес – 53,8 кг, расстояние между щупальцами – 7 м.
4. Самой быстрой наземной улиткой считается обыкновенная садовая улитка Хеликс асперза. Расстояние в 31 см она преодолевает за 2 мин. 31 сек.
5.
Самые ценные раковины имеют фарфоровые улитки ципреи.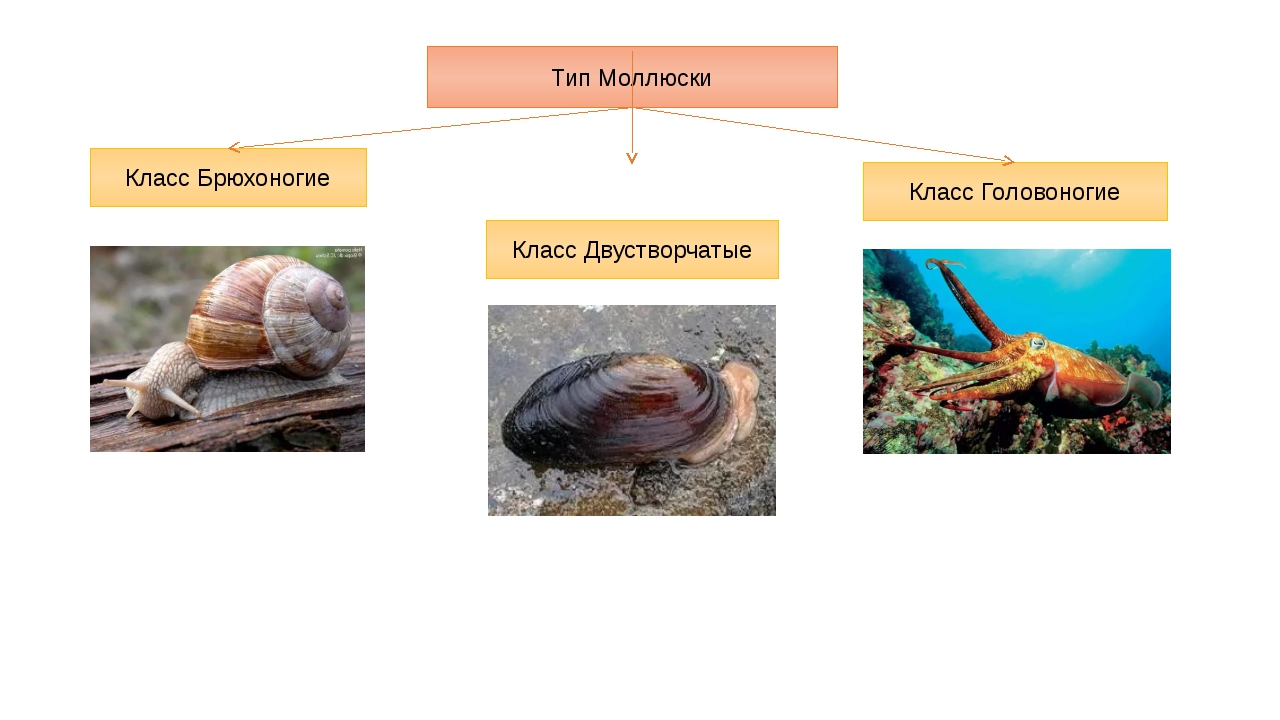 Так,
экземпляр вида ципреи был продан в Нью-Йорке одному
коллекционеру из Италии за 24 тыс. долларов.
Так,
экземпляр вида ципреи был продан в Нью-Йорке одному
коллекционеру из Италии за 24 тыс. долларов.
6. Самая большая жемчужина была размерами с голубиное яйцо и весила 6,8 г. Стоимость этой жемчужины – 350 кг золота.
7. Укус морских улиток может быть ядовитым. Яд конуса вызывает воспаление и иногда остановку дыхания.
8. Европейская устрица может менять свой пол несколько раз в год. Созревшая для размножения, она сначала функционирует как самка, а через некоторое время как самец и так по нескольку раз.
9. Присоски осьминогов обладают большой силой. На каждом щупальце их насчитывается до 100 штук. Одна присоска диаметром 6 мм удерживает вес в 170 г.
10. Ежегодно во всем мире добывается свыше 800 000 т мидий. По калорийности это соответствует более чем 6000 т. телятины.
5.Первичное закрепление.
ЛАБОРАТОРНАЯ РАБОТА №3
Изучение строения раковин моллюсков
Цель: изучить особенности строения раковин моллюсков, научиться выявлять черты сходства и различия
Оборудование и материалы: коллекция раковин моллюсков (беззубка,
катушка роговая, прудовик, садовая, виноградная улитки)
Теоретические сведения
Тип Моллюски
объединяет 230 тысяч видов. Это несегментированные вторичнополостные
беспозвоночные животные, тело которых подразделяется на голову, туловище, ногу.
Это несегментированные вторичнополостные
беспозвоночные животные, тело которых подразделяется на голову, туловище, ногу.
Тело моллюсков окружает кожная складка – мантия. Между мантией и туловищем образуется мантийная полость. Благодаря железам мантии у большинства моллюсков формируется защитная раковина, которая является скелетным образованием и служит опорой и защитой для моллюсков, надёжно защищая от большинства хищников и позволяя удобно перемещаться
Раковина образована комплексом органических и неорганических веществ и состоит их трех слоев.
Наружный слой образован органическим рогоподобным веществом, которое определяет окраску раковины.
Средний слой называют фарфоровым, он является наиболее мощным, состоит из кальция карбоната.
Внутренний слой
соприкасается с телом животного, имеет гладкую поверхность, защищая нежные
ткани мантии от механического повреждения. Основу его составляет тоже кальция
карбонат, пластинки которого располагаются параллельно поверхности раковины,
что создает эффект переливания световых волн.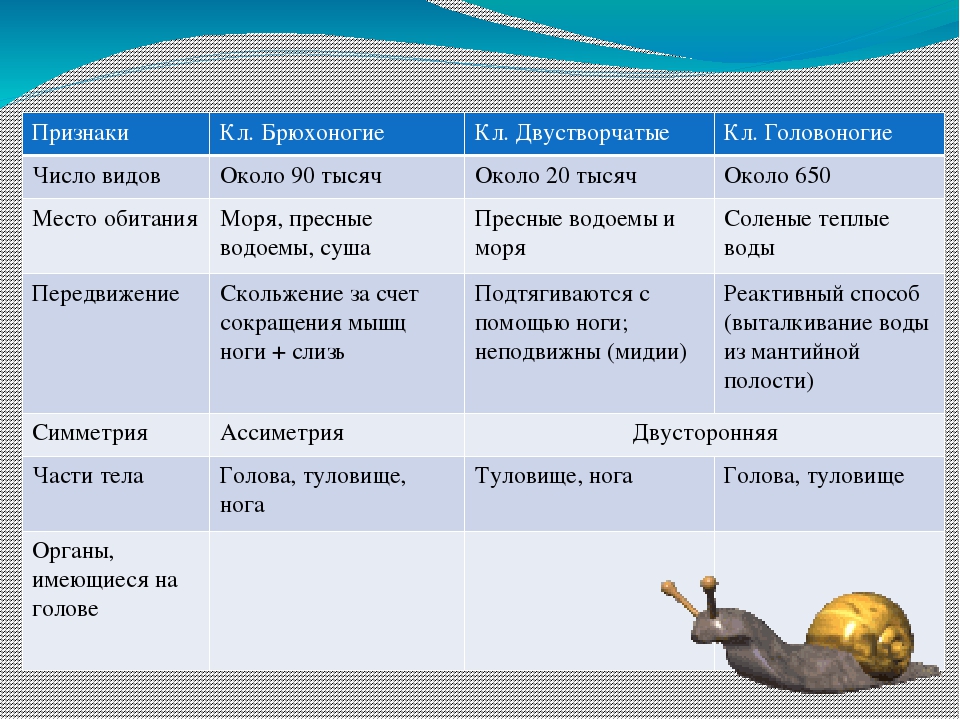 Поэтому этот слой называют перламутровым.
Поэтому этот слой называют перламутровым.
Прирост раковины происходит по краю мантии, из-за чего на створках видны концентрические линии. По ним, как и по годичным кольцам деревьев, можно судить о скорости роста и условиях жизни конкретных особей.
Брюхоногие, Двустворчатые, Головоногие. Особенности строения Моллюсков
Вопрос 1. Объясните, как моллюски приспо-соблены к своей среде обитания.
Среди моллюсков есть обитатели вод-ной и наземно-воздушной среды.
Многие как наземные, так и водные моллюски имеют раковину, которая и у тех, и у других выполняет роль пассивной защиты.
Важнейшим приспособлением к назем-ному образу жизни у сухопутных моллю-сков является дыхание легкими.
Водные моллюски приспособлены к по-лучению кислорода из воды — они дышат жабрами или поверхностью мантии.
Головоногие моллюски выработали осо-бый — реактивный — способ передвиже-ния, очень эффективный в водной среде.
Вопрос 2. Каковы черты сходства и различия брюхоногих и двустворчатых моллюсков?
Каковы черты сходства и различия брюхоногих и двустворчатых моллюсков?
Брюхоногие и двустворчатые моллюски являются многоклеточными животными, туловища которых не имеют сегментарно-го строения. Тело большинства видов дан-ных классов моллюсков полностью или частично покрыто раковиной. У брюхоно-гих раковина дельная, часто асимметрич-ная и закрученная, у двустворчатых — со-стоит из двух створок.
Голову, туловище и ногу можно разли-чить только у брюхоногих, у двустворча-тых голова отсутствует.
У брюхоногих хорошо развиты органы чувств — осязания, химического чувства, равновесия и зрения. У двустворчатых ор-ганы чувств развиты слабо.
Двустворчатые являются исключитель-но водными животными, в то время как среди брюхоногих есть обитатели и вод-ной, и наземно-воздушной среды.
Вопрос 3. Перечислите способы защиты от врагов известных вам моллюсков.
Некоторые брюхоногие и практически все двустворчатые моллюски способны полностью или частично прятаться в ра-ковину — это пассивный способ защиты.
Кальмары при спасении от преследова-ния могут несколько десятков метров про-летать над водой. Материал с сайта
Некоторые моллюски, например кара-катицы, осьминоги, в случае опасности способны менять окраску или выбрасы-вать чернильное вещество, вырабатывае-мое в особом органе — чернильном мешке. Перед применением этого средства за-щиты моллюск сначала темнеет, потом выбрасывает чернила в пленке, по форме напоминающей его тело. Преследователь хватает чернильную «бомбу» — пленка разрывается, чернила окрашивают боль-шой объем воды и парализуют обоняние врага. Это спасает жизнь моллюска: он после выброса чернил быстро бледнеет и почти невидимым уплывает.
Вопрос 4. Что позволяет головоногим быстро перемещаться в воде и спасаться от врагов?
Быстрое движение в воде головоногих моллюсков обеспечивается пульсирующим выбрасыванием воды из мантийной полос-ти (реактивное движение). Такой способ движения позволяет развить значитель-ную скорость плавания: кальмарам — до 40 км/ч, осьминогам — до 15 км/ч.
Не нашли то, что искали? Воспользуйтесь поиском
На этой странице материал по темам:
- передвижение у брюхоногих
- брюхоногие моллюски краткое содержание
- В чем различие брюхоногих, двустворчатых и головоногих моллюсков
- органы и способы передвижения двухтворчатых молюсков
- класс головоногие моллюски краткая биография
Тип моллюски — это мягкотелые животные, преимущественно с билатерально симметричным строением, населяющие как водоемы, так и сушу. Насчитывается более 120 тысяч видов.
Размеры зрелых моллюсков разных классов существенно отличаются — от пары миллиметров до 20м. Многие ведут сидячий или малоподвижный образ жизни, и лишь головоногие способны активно перемещаться в воде. Наука о моллюсках называется малакологией , она занимается изучением строения, развития мягкотелых, роли в окружающем мире.
Особенности строения Моллюсков
Внешнее строение
Туловище с двусторонней симметрией у двустворчатых и головоногих, или несимметричное — у брюхоногих.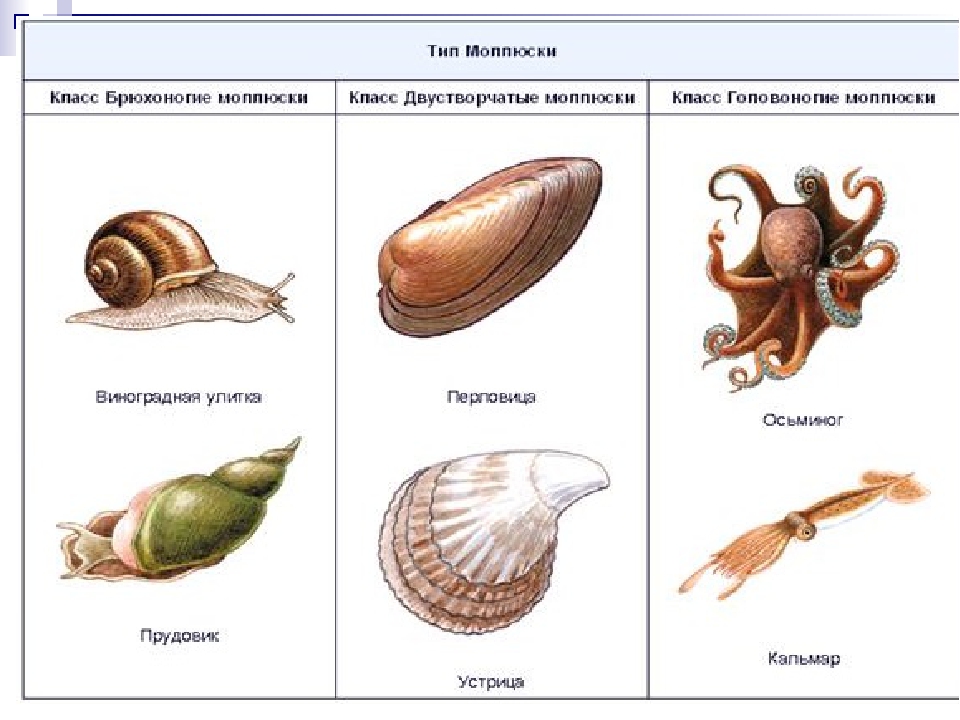 Выделяют такие его отделы: головная часть с органами зрения и щупальцами, собственно тело и нога — мышечное образование, служит для перемещения. Для всех двустворчатых характерно наличие ноги, у головоногих же она преобразовалась в щупальца и сифон.
Выделяют такие его отделы: головная часть с органами зрения и щупальцами, собственно тело и нога — мышечное образование, служит для перемещения. Для всех двустворчатых характерно наличие ноги, у головоногих же она преобразовалась в щупальца и сифон.
Тело моллюска окружено раковиной, служит местом крепления мышц. У брюхоногих моллюсков она имеет цельное строение в виде спирального завитка. У двустворчатых представлена двумя створками, которые связаны гибкими тяжами соединительной ткани. Большая часть головоногих лишены раковины.
От боковых частей туловища отходит мантия, высланная эпителиальными клетками. Она вместе с телом формирует полость, где находятся жаберные дуги, органы чувств, выводные протоки желез пищеварительного тракта, мочеполовой системы, анус.
Моллюски это целомические организмы, но вторичная полость у них сохранилась только возле сердца и половых органов. Основная же часть внутреннего пространства представлена гемоцелем.
Внутреннее строение
Система пищеварения моллюсков
разделена на три части: переднюю, среднюю и заднюю кишки. У многих представителей в глотке находится радула — язычок, предназначен для измельчения пищи. На нем есть хитиновые пластинки с зубцами. С помощью радулы они поглощают бактерий или еду растительной природы. В полость глотки выделяется слюна и склеивает пищевые частицы. Затем еда попадает в желудок, куда открывается пищеварительная железа (печень). После переваривания остатки выводятся наружу через анальное отверстие.
У многих представителей в глотке находится радула — язычок, предназначен для измельчения пищи. На нем есть хитиновые пластинки с зубцами. С помощью радулы они поглощают бактерий или еду растительной природы. В полость глотки выделяется слюна и склеивает пищевые частицы. Затем еда попадает в желудок, куда открывается пищеварительная железа (печень). После переваривания остатки выводятся наружу через анальное отверстие.
Система кровообращения незамкнутая, в сердце различают желудочек и обычно два (редко четыре) предсердия. Из кровеносного русла кровь поступает в синусы и лакуны, расположенные между органами, дальше опять переходит в сосуды и идет к органам дыхания.
Дыхание у водных видов осуществляется жабрами у обитателей суши — легкими. Лёгочная ткань оснащена густой сосудистой сетью, где идет обмен кислорода и СО 2 . Легкое сообщается с наружной средой дыхальцем.
Нервная система моллюсков
состоит из пяти пар нервных узлов, объединенных волокнистыми тяжами. Неодинаковое развитие органов чувств у моллюсков свидетельствует о разном образе жизни представителей типа.
Неодинаковое развитие органов чувств у моллюсков свидетельствует о разном образе жизни представителей типа.
К примеру, головоногие имеют достаточно развитое зрение, строение глаза схожее со строением глаза позвоночных. Хищная природа вынудила их приспосабливаться к переменчивым условиям окружающей среды через усложнение зрительного аппарата. У них образовался своеобразный тип аккомодации, который осуществлялся посредством смены расстояния между сетчаткой и хрусталиком.
Размножаются моллюски половым путем . Встречаются как раздельнополые (с наружным оплодотворением), так и гермафродиты (с внутренним). У морских двустворчатых и брюхоногих моллюсков — развитие непрямое, есть стадия личинки, у остальных – прямое.
Особенности строения моллюсков по сравнению с кольчатыми червями
Какие новые органы появились у моллюсков по сравнению с червями?
Моллюски обладают специализированными органами. Это выделительная, пищеварительная система, которая включает ряд отделов, есть сердце, печень. Дыхательные органы — жабры или легочная ткань.
Дыхательные органы — жабры или легочная ткань.
Кровеносная система незамкнутая, у кольчатых червей – замкнутая.
Нервная система моллюсков имеет вид нервных ганглиев, объединённых друг с другом нервными волокнами. У кольчатых червей есть только в брюшной области нервная цепь, которая разветвляется по сегментам.
Как моллюски приспособлены к своей среде обитания?
Представители типа населяют водные просторы и поверхность суши. Для существования вне водоема и дыхания атмосферным воздухом у мягкотелых появились легочная ткань. Жители водоемов получают О 2 с помощью жаберных дуг.
Как моллюски защищаются от врагов?
Для перемещения в воде головоногие приспособились к реактивному передвижению, так они могут быстро убегать от врагов.
Защитой от хищников служат ядовитые и химические вещества (чернила). Некоторые способны за секунды закапываться в песчаное дно при наличии угрозы или скрыться, используя пружинящую ногу.
Какую функцию выполняет раковина моллюсков?
В первую очередь — это опорная функция, служит наружным скелетом. Также прочная раковина двустворчатых и брюхоногих моллюсков нужна для защиты от неблагоприятных факторов. Так, при приближении опасности они прячутся в них и становятся недоступными для большинства рыб.
Также прочная раковина двустворчатых и брюхоногих моллюсков нужна для защиты от неблагоприятных факторов. Так, при приближении опасности они прячутся в них и становятся недоступными для большинства рыб.
Cходство и различие брюхоногих и двустворчатых моллюсков
| Свойства | Брюхоногие | Двустворчатые |
|---|---|---|
| Внесистематическая категория | Многоклеточные организмы | |
| Внешние покровы | Тело окружено раковиной (целиком или отчасти) | |
| Раковина | Сдельная, ассиметричная и закрученная | Имеет две створки |
| Строение тела | Голова, туловище и нога | Туловище, нога |
| Анализаторы | Тактильная, химическая рецепция, равновесие и зрение. | Недостаточно развиты |
| Среда обитания | Вода и суша | Водоемы |
Значение моллюсков в природе и жизни человека
Они неотъемлемая составляющая пищевой цепочки. Мягкотелых употребляют лягушки, рыбы, птицы. Тюлени употребляют головоногих моллюсков, морские звезды – двустворчатых.
Мягкотелых употребляют лягушки, рыбы, птицы. Тюлени употребляют головоногих моллюсков, морские звезды – двустворчатых.
Через тело моллюска проходит вода и очищается от загрязняющих факторов. А моллюски в свою очередь получают частицы еды из отфильтрованной воды.
Створки мягкотелых принимают участие в формировании осадочных пород.
Широко применяются в кулинарии, считаются деликатесом во многих странах. Это мясо мидий, морские гребешки, устрицы, каракатицы и осьминоги. Из-за популярности блюд из экзотических животных, их стали выращивать на специально оборудованных фермах.
Между створками раковины образуется ценное ювелирное сырье — жемчуг. Жемчужина формируется после попадания внутрь инородного тела. Поскольку мышцы моллюсков недостаточно развиты, они не могут выбросить его наружу. Чтобы нейтрализовать чужеродный предмет, вокруг него формируется капсула и с новообразованной жемчужиной моллюск живет всю жизнь.
Сейчас жемчуг добывают в искусственно созданных условиях. Приоткрыв створки, под мантию кладут инородные объекты, а моллюска перемещают в водоем с благоприятными для жизни условиями и через три года получают жемчуг.
Приоткрыв створки, под мантию кладут инородные объекты, а моллюска перемещают в водоем с благоприятными для жизни условиями и через три года получают жемчуг.
Каракатиц и осьминогов используют для добычи чернильного вещества, из которого изготавливают тушь.
Вредители сельского хозяйства — слизни, уничтожают зерновые культуры, огородные растения (картофель, капусту, томаты).
Плоские черви, вызывающие заболевания человека и животных, используют моллюсков как промежуточных хозяев.
Моллюски относятся к одним из наиболее древних беспозвоночных животных. Отличаются наличием вторичной полости тела и довольно сложно устроенными внутренними органами. Многие из них имеют известковую раковину, которая довольно хорошо защищает их тело от посягательств многочисленных врагов.
Не так уж часто об этом вспоминают, но немало видов этого типа ведут хищнический образ жизни. В этом им помогает развитая слюнная железа. Кстати, а что такое слюнная железа у моллюсков? Под этим обобщающим понятием подразумевается довольно обширный спектр специфических органов, располагающихся в глотке и ротовой полости. Предназначаются они для секреции различных веществ, характеристики которых могут очень сильно отличаться от нашего понимания слова «слюна».
Предназначаются они для секреции различных веществ, характеристики которых могут очень сильно отличаться от нашего понимания слова «слюна».
Как правило, у моллюсков имеется одна или две пары таких желез, которые у некоторых видов достигают весьма впечатляющих размеров. У большинства хищных видов в секрете, который они выделяют, содержится от 2,18 до 4,25% химически чистой серной кислоты. Она помогает как отбиваться от хищников, так и охотиться на своих сородичей (серная кислота отлично растворяет их известковые раковины). Вот что такое слюнная железа у моллюсков.
Прочее природное значение
Многие из видов слизней, а также виноградная улитка, наносят огромный вред сельскому хозяйству по всему миру. В то же время именно моллюски играют важнейшую роль в общемировой очистке воды, так как для питания ими используется отфильтрованная из нее органика. Во многих странах крупных разводят на морских фермах, так как они являются ценным пищевым продуктом, в котором содержится много белка. Эти представители и устрицы) используются даже в диетическом питании.
Эти представители и устрицы) используются даже в диетическом питании.
В бывшем СССР редкими и исчезающими считались сразу 19 представителей этого древнейшего типа. Несмотря на многообразие моллюсков, к ним следует относиться бережно, так как они крайне важны для правильного функционирования многих природных биотопов.
Вообще, моллюски нередко отличаются важнейшим практическим значением и для человека. К примеру, жемчужница массово разводится во многих приморских странах, так как этот вид является поставщиком природного жемчуга. Некоторые моллюски представляют собой большую ценность для медицины, химической и перерабатывающей промышленности.
Хотите знать интересные факты о моллюсках? В Античный период и Средневековье неприметные головоногие порой были основой благосостояния целых государств, так как из них добывался ценнейший пурпур, которым окрашивались королевские мантии и одеяния знати!
Тип моллюсков
Всего он насчитывает более 130 000 видов (да, многообразие моллюсков невероятно). Моллюски по общей численности уступают только членистоногим, являются вторыми по распространенности живыми организмами на планете. Большинство из них обитает в воде, и лишь сравнительно небольшое количество видов избрали местом своего проживания сушу.
Моллюски по общей численности уступают только членистоногим, являются вторыми по распространенности живыми организмами на планете. Большинство из них обитает в воде, и лишь сравнительно небольшое количество видов избрали местом своего проживания сушу.
Общая характеристика
Практически все животные, которые входят в состав этого типа, отличаются сразу несколькими специфическими чертами. Вот принятая на сегодняшний день общая характеристика моллюсков:
- Во-первых, трехслойность. Их система органов формируется из эктодермы, энтодермы и мезодермы.
- Симметрия билатерального типа, вызванная значительным смещением большей части их органов.
- Тело несегментированное, в большинстве случаев защищенное сравнительно прочной известковой раковиной.
- Имеется кожная складка (мантия), которая обволакивает все их тело.
- Для передвижения служит хорошо выраженный мускульный вырост (нога).
- Целомическая полость очень плохо выражена.
- Имеются практически все те же системы органов (в упрощенном варианте, конечно же), как и у высших животных.

Таким образом, общая характеристика моллюсков указывает на то, что перед нами — достаточно развитые, но все же примитивные животные. Неудивительно, что именно моллюсков многие ученые считают основными предками большого количества живых организмов на нашей планете. Для наглядности приведем таблицу, в которой более подробно расписываются характеристики двух наиболее распространенных классов.
Рассматриваемый признак | Классы моллюсков |
|
Двустворчатые | Брюхоногие |
|
Тип симметрии | Двусторонний. | Симметрия отсутствует, некоторые органы полностью редуцированы. |
Наличие или отсутствие головы | Полностью атрофирована, как и все системы органов, которые к ней исторически относились. | Имеется, как и весь комплект органов (ротовая полость, глаза). |
Органы дыхания | Жабры или легкое (прудовик, к примеру). |
|
Тип раковины | Двустворчатая. | Цельная, может быть закручена в разные стороны (прудовики, ампулярия) или же в спираль (Катушка озерная). |
Половой диморфизм, половая система | Раздельнополые, самцы зачастую мельче. | Гермафродиты, иногда раздельнополые. Диморфизм выражен слабо. |
Тип питания | Пассивное (фильтрация воды). Вообще, эти моллюски в природе способствуют отличному очищению воды, так как отфильтровывают из нее тонны органических примесей. | Активное, встречаются хищные виды (Конусы (лат. |
Ареал обитания | Моря и пресные водоемы. | Все типы водоемов. Встречаются также наземные моллюски (Виноградная улитка). |

 Conidae)).
Conidae)).Детальная характеристика
У тело все же симметрично, хотя у двустворчатых видов этого не наблюдается. Разделение тела на сегменты сохранилось только у очень примитивных видов. Вторичная полость тела представлена сумкой, окружающей сердечную мышцу и половые органы. Все пространство между органами полностью заполнено паренхимой.
Туловище у большинства можно поделить на следующие отделы:
- Голова.
- Туловище.
- Мускульная нога, посредством которой осуществляется передвижение.
У всех двустворчатых видов голова полностью редуцирована. Под ногой подразумевается массивный мускульный отросток, который развивается из основания брюшной стенки. У самого основания тела кожный покров образует большую складку, мантию. Между ней и телом имеется довольно большая полость, в которой размещены следующие органы: жабры, а также выводы половой и выделительной систем. Именно мантия секретирует те вещества, которые, вступив в реакцию с водой, образуют прочную раковину.
Между ней и телом имеется довольно большая полость, в которой размещены следующие органы: жабры, а также выводы половой и выделительной систем. Именно мантия секретирует те вещества, которые, вступив в реакцию с водой, образуют прочную раковину.
Раковина может быть как полностью сплошной, так и состоять из двух створок или нескольких пластинок. В состав этой оболочки входит много углекислого газа (разумеется, в связанном состоянии — СаСО 3), а также конхиолина, особого органического вещества, которое синтезируется телом моллюска. Впрочем, у многих видов моллюсков раковина полностью или частично редуцирована. У слизней от нее осталась только микроскопических размеров пластинка.
Характеристика пищеварительной системы
Брюхоногие моллюскиНа переднем конце головы имеется рот. Главным органом в нем является мощный мускулистый язык, который покрыт особо прочной хитиновой теркой (радулой). С ее помощью улитки соскребают налет водорослей или другой органики со всех доступных поверхностей. У хищных видов (о них мы поговорим ниже) язык переродился в гибкий и жесткий хоботок, который предназначается для вскрытия раковин других моллюсков.
У хищных видов (о них мы поговорим ниже) язык переродился в гибкий и жесткий хоботок, который предназначается для вскрытия раковин других моллюсков.
У Конусов (о них также будет сказано отдельно) отдельные сегменты радулы выступают за пределы ротовой полости и образуют своего рода гарпуны. С их помощью эти представители моллюсков буквально метают свой яд в жертву. У некоторых хищных брюхоногих язык превратился в особое «сверло», которым они буквально высверливают в раковине своей добычи отверстия для впрыскивания яда.
ДвустворчатыеВ их случае все намного проще. Они просто неподвижно лежат на дне (или висят, намертво прикрепленные к субстрату), профильтровывая сквозь свое тело сотни литров воды с растворенной в ней органикой. Отфильтрованные частицы поступают прямо в объемистый желудок.
Органы дыхания
Большая часть видов дышит жабрами. Есть «передние» и «задние» виды. У первых жабры располагаются в передней части тела и их верхушка направлена вперед. Соответственно, во втором случае верхушка смотрит назад. Некоторые утратили жабры в прямом понимании этого слова. Эти большие моллюски дышат прямо через кожу.
Соответственно, во втором случае верхушка смотрит назад. Некоторые утратили жабры в прямом понимании этого слова. Эти большие моллюски дышат прямо через кожу.
Для этого у них развился специальный кожный орган адаптивного типа. У сухопутных видов и вторичных водных моллюсков (их предки снова вернулись в воду) часть мантии заворачивается, образуя своеобразное легкое, стенки которого густо пронизаны кровеносными сосудами. Чтобы подышать, такие улитки поднимаются на поверхность воды и набирают запас воздуха при помощи специального дыхальца. Сердце, расположенное неподалеку от простейшей «конструкции», состоит из одного предсердия и желудочка.
Основные классы, входящие в состав типа
Как же подразделяется тип моллюски? Классы моллюсков (всего их восемь штук) «увенчаны» тремя наиболее многочисленными:
- Брюхоногие (Gastropoda). Сюда относятся тысячи видов улиток всех размеров, главным отличительным признаком которых является низкая скорость передвижения и хорошо развитая мускулистая нога.

- Двустворчатые (Bivalvia). Раковина из двух створок. Как правило, все входящие в класс виды являются оседлыми, малоподвижными. Передвигаться могут как с помощью мускулистой ноги, так и посредством реактивной тяги, выбрасывая воду под давлением.
- Головоногие (Cephalopoda). Подвижные моллюски, раковины или лишены вовсе, или она находится в зачаточном состоянии.
Кто еще входит в тип моллюски? Классы моллюсков довольно разнообразны: кроме всех вышеперечисленных, имеются еще Лопатоногие, Панцирные и Ямкохвостые, Бороздчатобрюхие и Моноплакофоры. Все они относятся к ныне живущим и здравствующим.
Какие ископаемые содержит тип моллюски? Классы моллюсков, которые уже вымерли:
- Rostroconchia.
- Тентакулиты.
К слову говоря, те же Моноплакофоры до 1952 года считались полностью вымершими, но в то время судно «Галатея» с научно-исследовательской экспедицией на борту выловило несколько новых организмов, которые были отнесены к новому виду Neopilina galatheae. Как можно заметить, название моллюсков этого вида было дано по имени исследовательского судна, которое их обнаружило. Впрочем, в научной практике это не редкость: виды куда чаще обозначаются в честь открывшего их исследователя.
Как можно заметить, название моллюсков этого вида было дано по имени исследовательского судна, которое их обнаружило. Впрочем, в научной практике это не редкость: виды куда чаще обозначаются в честь открывшего их исследователя.
Так что не исключено, что все последующие годы и новые исследовательские миссии смогут обогатить тип моллюски: классы моллюсков, которые ныне считаются вымершими, вполне могут сохраняться где-то в бездонных глубинах мирового океана.
Как бы странно это ни звучало, но одними из самых опасных и невероятных хищников на нашей планете считаются… внешне безобидные брюхоногие моллюски. К примеру, улитки Конусы (лат. Conidae), яд которых настолько необычен, что его используют современные фармацевты при изготовлении некоторых видов редких лекарств. Кстати, название моллюсков этого семейства полностью оправдано. Их форма и в самом деле более всего похожа на усеченный конус.
Они могут быть настойчивыми охотниками, исключительно безжалостно расправляясь с пойменной добычей. Конечно же, в роли последней чаще выступают колониальные, малоподвижные виды животных, так как за другими улиткам попросту не угнаться. Сама жертва своими размерами может превосходить охотника в десятки раз. Хотите знать еще интересные факты о моллюсках? Да пожалуйста!
Конечно же, в роли последней чаще выступают колониальные, малоподвижные виды животных, так как за другими улиткам попросту не угнаться. Сама жертва своими размерами может превосходить охотника в десятки раз. Хотите знать еще интересные факты о моллюсках? Да пожалуйста!
О методах охоты улиток
Чаще всего коварный моллюск пользуется своим самым мощным органом, сильной мускулистой ногой. Она может прикрепляться к добыче с силой, эквивалентной приложению усилия в 20 кг! Хищной улитке этого вполне достаточно. К примеру, «пойманная» устрица раскрывается менее чем за час при усилии только в десяток килограммов! Словом, жизнь моллюсков куда опаснее, чем об этом принято думать…
Другие виды брюхоногих и вовсе предпочитают ничего не давить, аккуратно просверливая раковину добычи при помощи специального хоботка. Вот только процесс этот простым и быстрым не назвать при всем желании. Так, при толщине раковины всего в 0,1 мм сверление может отнимать вплоть до 13 часов! Да уж, такой способ «охоты» подходит исключительно для улиток. ..
..
Растворение!
Чтобы растворить чужую раковину и самого его владельца, моллюск использует серную кислоту (вы ведь уже знаете, что такое слюнная железа у моллюсков). Так разрушение идет намного проще и быстрее. После того как дырка проделана, хищник начинает не торопясь выедать свою добычу из «упаковки», пользуясь для этого своим хоботком. В какой-то мере этот орган можно смело считать аналогом нашей руки, так как он принимает непосредственное участие в захвате и удержании добычи. Кроме того, этот манипулятор зачастую может вытягиваться так, что превышает длину тела самого охотника.
Именно так улитки могут доставать свою добычу даже из глубоких щелей и больших раковин. Еще раз напоминаем, что именно из хоботка в теле жертвы впрыскивается сильный яд, основу которого составляет химически чистая серная кислота (выделяемая из «безобидных» слюнных желез). Словом, отныне вы точно знаете, что такое слюнная железа у моллюсков и для чего она им нужна.
К Двустворчатым моллюскам относится около 20 тыс. видов. Это донные малоподвижные животные. В реках и озерах живут беззубка, перловица. Хорошо известен морской моллюск мидия. Питаются двустворчатые моллюски мелким планктоном и взвешенными в воде частицами, играя важнейшую роль в очистке воды.
видов. Это донные малоподвижные животные. В реках и озерах живут беззубка, перловица. Хорошо известен морской моллюск мидия. Питаются двустворчатые моллюски мелким планктоном и взвешенными в воде частицами, играя важнейшую роль в очистке воды.
Внешнее строение. Тело двустворчатых моллюсков продолговатое, двусторонне-симметричное, сплюогутое с боков. Головы нет (рис. 76). В теле различают туловище и у многих — ногу.
Рис. 76. Разнообразие двустворчатых моллюсков: 1 — перловица; 2 — мидия; 3 — устрица; 4 — гребешок
У беззубки нога имеет клиновидную форму и служит для передвижения в песке и иле. При этом моллюск выдвигает ногу вперед, затем расширяет ее, закрепляя в грунте, и подтягивает тело (рис. 77).
Рис. 77. Схема передвижения беззубки
У мидии, ведущей неподвижный образ жизни, нога утратила двигательную функцию. Специальными железами мидия выделяет прочные белковые нити — биссус (от греч. биссос — «тонкая пряжа»), с помощью которых она прикрепляется к камням.
Тело двустворчатых покрыто мантией, которая свободно свешивается по бокам тела в виде двух больших складок. На заднем конце тела мантия часто срастается и образует две трубки — сифоны.
Наружная сторона мантийных складок формирует известковую раковину. У беззубки длина ее может достигать 10 см, у мидии — 20 см. Раковина состоит из двух симметричных створок, охватывающих тело с боков. Короткая поперечная лента эластичного вещества соединяет створки на спинной стороне. Створки замыкаются особыми мускулами-замыкателями. У беззубки имеется два таких мускула, а у мидии — один. Когда моллюск расслабляет мускулы, створки расходятся и остаются полуоткрытыми.
У некоторых моллюсков края створок на спинной стороне образуют выросты — зубы. Это замок, усиливающий скрепление створок. У беззубки таких выростов нет, за что она и получила свое название. У беззубки и мидии внутренняя поверхность раковины выстлана прочным блестящим перламутровым слоем. Инородные частицы (например, песчинки), попадающие между мантией и створкой раковины, обволакиваются слоями перламутра и превращаются в жемчуг (рис. 78).
78).
Рис. 78. Схема образования жемчужины: 1 — раковина; 2 — мантия (наружный слой) 3 — песчинка: 4 — жемчужина
Пищеварительная система. Редукция головы у двустворчатых моллюсков привела к исчезновению многих органов пищеварения, которые есть у брюхоногих: глотки, тёрки, челюстей, слюнных желез (рис. 79).
Рис. 79. Внутреннее строение беззубки при продольном (А) и поперечном (Б) разрезе: 1 — нога; 2 — ротовое отверстие; 3 — пищевод; 4 — печень; 5 — желудок; 6 — кишка; 7 — сердце; 8 — почка; 9 — анальное отверстие; 10 — жабры; 11 — мантия; 12 — раковина; 13 — яичник
Рот, окруженный двумя парами лопастей, расположен на переднем конце тела, у основания ноги. Он ведет в короткий пищевод, который открывается в мешковидный желудок. Кишка спускается от желудка в основание ноги, делает несколько изгибов и заканчивается на заднем конце тела анальным отверстием.
Двустворчатые моллюски относятся к животным-фильтраторам. Они питаются планктоном и взвешенными в воде мелкими органическими частичками. На жабрах этих моллюсков находятся многочисленные очень маленькие, постоянно колеблющиеся реснички. Их движение создает ток воды в мантийной полости: через вводной сифон вода засасывается в мантийную полость. С током воды приносятся мелкие пищевые частицы. Они осаждаются выделяемой слизью и направляются к ротовым лопастям. Ротовые лопасти освобождают пищу от несъедобных частиц. Съедобные частицы отправляются в рот, несъедобные — через выводной сифон наружу. Через него из организма выносятся и экскременты. Двустворчатые моллюски могут за небольшое время профильтровывать большой объем воды. Например, мидия фильтрует за час до 5 л воды.
На жабрах этих моллюсков находятся многочисленные очень маленькие, постоянно колеблющиеся реснички. Их движение создает ток воды в мантийной полости: через вводной сифон вода засасывается в мантийную полость. С током воды приносятся мелкие пищевые частицы. Они осаждаются выделяемой слизью и направляются к ротовым лопастям. Ротовые лопасти освобождают пищу от несъедобных частиц. Съедобные частицы отправляются в рот, несъедобные — через выводной сифон наружу. Через него из организма выносятся и экскременты. Двустворчатые моллюски могут за небольшое время профильтровывать большой объем воды. Например, мидия фильтрует за час до 5 л воды.
Органы дыхания. У беззубки и мидии жабры пластинчатые. Они расположены под мантией с обеих сторон тела животного. Ток воды приносит (за счет работы ресничек) к жабрам обогащенную кислородом воду и выносит воду, богатую углекислым газом.
Кровеносная система
у двустворчатых моллюсков незамкнутая. В сердце у беззубки два предсердия и один желудочек. От желудочка берут начало два крупных сосуда — передняя и задняя аорты, которые распадаются на ряд артерий. Из артерий кровь попадает в систему полостей, лежащих в соединительной ткани. Из них по венам направляется к жабрам. В жабрах расположена густая сеть тончайших кровеносных сосудов (капилляров). Здесь кровь обогащается кислородом и по сосудам направляется к предсердиям. Сердце сокращается 3-20 раз в минуту.
От желудочка берут начало два крупных сосуда — передняя и задняя аорты, которые распадаются на ряд артерий. Из артерий кровь попадает в систему полостей, лежащих в соединительной ткани. Из них по венам направляется к жабрам. В жабрах расположена густая сеть тончайших кровеносных сосудов (капилляров). Здесь кровь обогащается кислородом и по сосудам направляется к предсердиям. Сердце сокращается 3-20 раз в минуту.
Выделительная система состоит из двух почек. Почки имеют вид двух обширных трубчатых, сложенных вдвое мешочков, одна сторона которых сообщается с околосердечной сумкой (остаток ценома), а другая — с мантийной полостью. В нее выходят вредные продукты жизнедеятельности и через выводной сифон удаляются из организма.
Нервная система. Состоит из трех пар нервных узлов (нервных ганглиев) и многочисленных отходящих от них нервов. Ганглии соединены между собой нервными стволами. С периферии сигналы по нервам передаются в ганглии, а из них — к мышцам.
Органы чувств
развиты слабо в результате малоподвижного образа жизни двустворчатых моллюсков и редукции головы.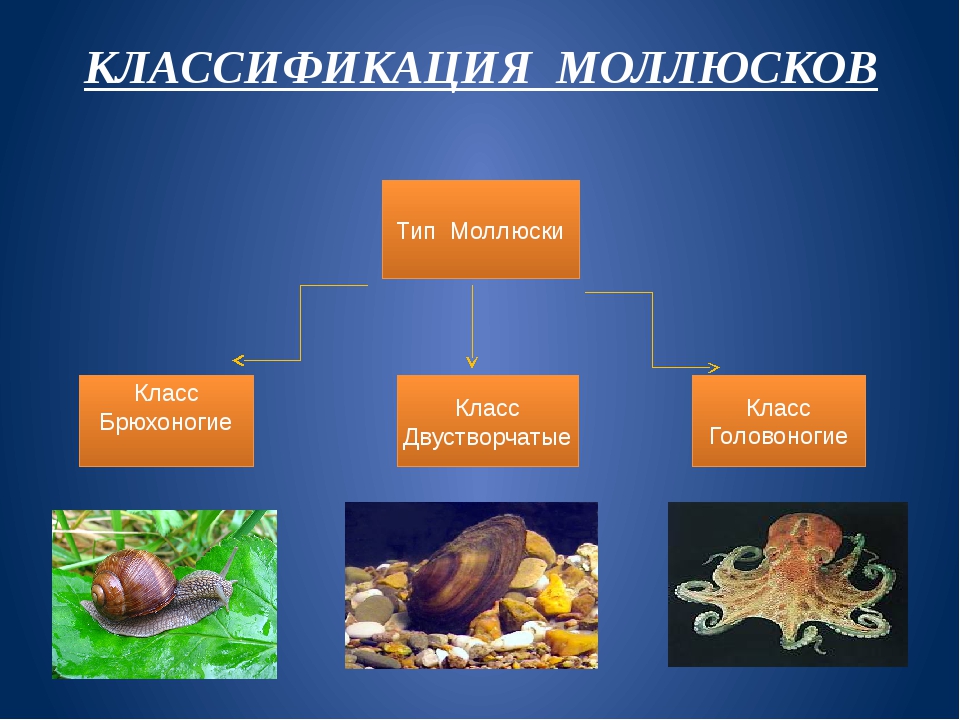 Имеются органы равновесия. Органами осязания служат ротовые лопасти. Осязательные клетки находятся также в ноге, по краю мантии и в жабрах. У некоторых моллюсков органами осязания служат различные щупальцевидные придатки, развивающиеся но краю мантии. У основания жаберных пластинок находятся органы химического чувства. У некоторых моллюсков по краю мантии располагаются глаза. У очень подвижных гребешков их свыше 100.
Имеются органы равновесия. Органами осязания служат ротовые лопасти. Осязательные клетки находятся также в ноге, по краю мантии и в жабрах. У некоторых моллюсков органами осязания служат различные щупальцевидные придатки, развивающиеся но краю мантии. У основания жаберных пластинок находятся органы химического чувства. У некоторых моллюсков по краю мантии располагаются глаза. У очень подвижных гребешков их свыше 100.
Размножение. Беззубка и мидия — раздельнополые животные. Сперматозоиды, образующиеся в семенниках самцов, через сифон попадают в воду и проникают в мантийную полость самок, где и происходит оплодотворение яйцеклеток. Успешное оплодотворение возможно только при большом скоплении моллюсков.
У мидии из яйца выходит маленькая личинка (рис. 80). Через некоторое время она превращается в другую личинку, называемую парусником. Парусник некоторое время плавает в толще воды, затем оседает на камень, скалу, другие твердые предметы и постепенно превращается в молодого моллюска.
Рис. 80. Личинки: 1 — мидии: 2 — беззубки
Личинки беззубки имеют на раковинке зубчики и клейкие нити, с помощью которых они прикрепляются к жабрам и коже проплывающих мимо рыб. В месте прикрепления личинки на теле рыбы образуется опухоль, внугри которой развивается моллюск. Через некоторое время он выходит наружу и падает на дно. Так с помощью рыб происходит развитие и расселение беззубок.
Двустворчатые моллюски играют огромную роль в водных биоценозах, отфильтровывая воду. Беззубками питаются некоторые водные животные.
К двустворчатым моллюскам принадлежат животные различных размеров, длиной от нескольких миллиметров до 1,5 м. А масса самого крупного двустворчатого моллюска — тридакны — может превышать 250 кг. Двустворчатые моллюски широко распространены в Мировом океане. Особенно много их в прибрежных мелководных участках теплых морей. Около 20% всех известных видов двустворчатых моллюсков населяют пресные воды, на суше они не встречаются. Двустворчатых моллюсков, например устриц, мидий, морских гребешков, сердцевидок, люди издавна употребляются в пищу. Некоторые из этих моллюсков, а также жемчужницы образуют перламутр и жемчуг. Их не только добывают с морского дна, но и специально выращивают на морских фермах, помещая между створкой раковины и мантией песчинку.
Некоторые из этих моллюсков, а также жемчужницы образуют перламутр и жемчуг. Их не только добывают с морского дна, но и специально выращивают на морских фермах, помещая между створкой раковины и мантией песчинку.
Лабораторная работа № 4
- Тема. Внешнее строение раковин пресноводных и морских моллюсков (по выбору — пункт 2 или 3).
- Цель. Установить сходство и различия в строении раковин моллюсков.
- Оборудование: пинцет, раковины моллюсков: морской гребешок, мидия, перловица, беззубка, роговая катушка, большой прудовик и др.
Ход работы
- Рассмотрите раковины морского гребешка и мидии. Выясните их сходство и различия. Объясните наличие выступов и углублений на спинной стороне раковин. Обратите внимание на форму и цвет наружного и внутреннего перламутрового слоя раковин.
- Рассмотрите раковины перловицы (или беззубки), определите переднюю и заднюю части. Отметьте сходство и различия во внешнем строении. Определите возраст моллюсков по годичным кольцам, расположенным на раковине.
 Соскребите скальпелем часть рогового слоя до известкового. Рассмотрите внутренний перламутровый слой.
Соскребите скальпелем часть рогового слоя до известкового. Рассмотрите внутренний перламутровый слой. - Рассмотрите раковины большого прудовика и роговой катушки. Отметьте сходство и различия внешнего строения раковин. Подсчитайте число оборотов в завитке каждой раковины.
- Зарисуйте по одной раковине из каждой пары. Обозначьте на рисунке основные части внешнего и внутреннего строения раковин. Надпишите названия этих частей.
- Напишите основные отличительные признаки раковины каждого моллюска. Объясните, по каким из них можно определить среду обитания, возраст и образ жизни моллюска.
Двустворчатые моллюски широко распространены в морях. Они являются очистителями-фильтраторами воды. Их тело заключено в двустворчатую раковину. Головы нет. Человек употребляет этих моллюсков в пищу, добывает из них жемчуг и перламутр.
Упражнения по пройденному материалу
- Назовите представителей двустворчатых моллюсков, используя рисунок 76 (с. 107). Каковы отличительные признаки их внешнего строения?
- Из каких слоев состоит раковина моллюсков? Какими веществами они образованы?
- Каковы особенности внутреннего строения и процессов жизнедеятельности двустворчатых моллюсков? Поясните на примере беззубки и мидии.
- Охарактеризуйте значение двустворчатых моллюсков в природе и жизни человека.
Какие признаки моллюсков послужили основой для выделения в типе Моллюски трех основных классов?
Три основных класса в типе Моллюски выделены на основании особенностей внешнего строения организма.
Каково значение моллюсков в природе и жизни человека?
Моллюски являются важным звеном в цепях питания. Многие из них употребляются в пищу. Среди моллюсков есть фильтраторы и трупоеды, поэтому они являются санитарами водоемов. Двустворчатые моллюски являются производителями жемчуга.
Среди моллюсков есть вредители садов и огородов.
Вопросы
1. Объясните, как моллюски приспособлены к своей среде обитания?
Большинство моллюсков – водные обитатели, поэтому их дыхательная система приспособлена к дыханию в воде. У многих есть жабры. Брюхоногие и двустворчатые моллюски, которые не способны к быстрому передвижению, имеют защитные раковины. Наземные моллюски выделяют большое количество слизи, которая защищает их от высыхания.
Наземные моллюски выделяют большое количество слизи, которая защищает их от высыхания.
2. Каковы черты сходства и различия брюхоногих и двустворчатых моллюсков?
Оба класса моллюсков являются исключительно водными животными. Головоногие моллюски, в отличие от двустворчатых, не имеют раковины, обитают только в соленой воде. Двустворчатые моллюски не способны к быстрому передвижению, не имеют головного отдела и характеризуются менее развитой нервной системой.
3. Как моллюски защищаются от врагов?
Двустворчатые и брюхоногие моллюски защищаются от врагов, прячась в своих раковинах. Головоногие моллюски способны быстро передвигаться, менять окраску и выбрасывать чернильные пятна.
4. Что позволяет головоногим моллюскам быстро перемещаться в воде и спасаться от врагов?
Быстро передвигаться головоногим моллюскам помогает пульсирующее выбрасывание воды из мантийной полости.
5. Как люди используют раковины моллюсков?
Раковины моллюсков использовались как материал для изготовления различных инструментов: рыболовных крючков, резцов, скребков, насадок для мотыги.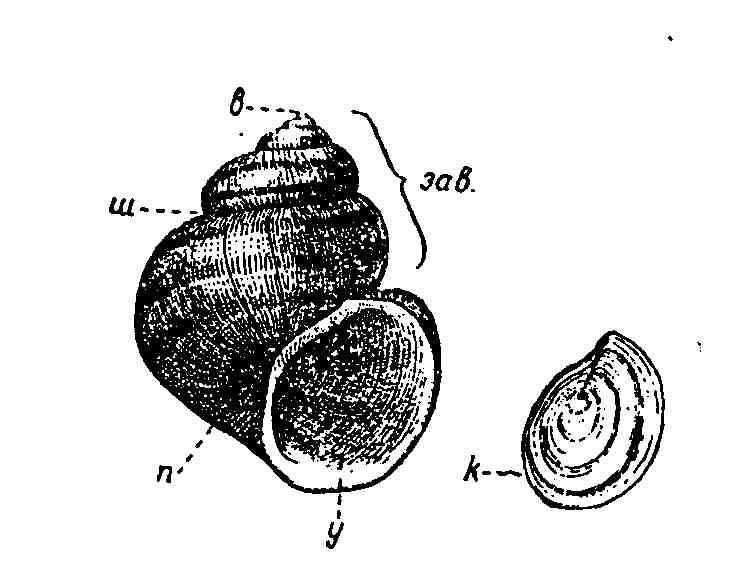 Сами раковины употреблялись в качестве сосудов, а также музыкальных инструментов (конх) и украшений. Добываемый из раковин перламутр используется для изготовления различных изделий, например, пуговиц, а также для инкрустаций. В некоторых местностях раковины служили деньгами — к примеру, раковины каури на островах Океании.
Сами раковины употреблялись в качестве сосудов, а также музыкальных инструментов (конх) и украшений. Добываемый из раковин перламутр используется для изготовления различных изделий, например, пуговиц, а также для инкрустаций. В некоторых местностях раковины служили деньгами — к примеру, раковины каури на островах Океании.
Задания
Предложите варианты борьбы с брюхоногими моллюсками – вредителями содовых и огородных культур, опираясь на особенности строения и жизнедеятельности этих животных.
Механические средства борьбы предполагают сбор вредителей вручную, а также устройство ловушек для них. Удобнее всего собирать улиток и слизней при помощи пинцета. Поскольку вредители днем предпочитают темные и влажные места, для них готовят соответствующие ловушки. Например, укладывают между грядками и на дорожках смоченные фруктовым соком или пивом капустные листы, мешковину, ветошь или доски. Днем вредители заползут в ловушки, а вечером их останется только собрать. Помимо этого, можно организовать и такие ловушки – неглубокие емкости вкапывают на уровне земли, наполняя их крепким солевым или мыльным раствором, и накрывают мешковиной. При контакте с мыльной или соленой жидкостью слизни погибают. Тот факт, что у садовых улиток и слизней очень мягкое тельце также можно использовать против них. Для этого достаточно рассыпать вблизи растений сухой пористый материал – измельченную яичную скорлупу, ракушки или мелкий гравий. Поскольку такая поверхность неприятна для моллюсков, к растениям они вряд ли подберутся. Кстати, в этом плане вредителям очень не нравится известь и суперфосфат, поскольку эти вещества впитывают слизь и влагу с их тел, усложняя передвижение. Однако следует отметить, что в дождливую погоду эффективность такого средства снижается. Еще одним непреодолимым барьером для слизней и улиток является вода. Создать для них такую преграду можно при помощи пластиковых желобов, наполненных водой. Опять же, их можно вкопать в землю. Брюхоногие моллюски вряд ли пожелают преодолевать эти препятствия, а если и попытаются, то просто попадут в воду и не смогут из нее выбраться.
При контакте с мыльной или соленой жидкостью слизни погибают. Тот факт, что у садовых улиток и слизней очень мягкое тельце также можно использовать против них. Для этого достаточно рассыпать вблизи растений сухой пористый материал – измельченную яичную скорлупу, ракушки или мелкий гравий. Поскольку такая поверхность неприятна для моллюсков, к растениям они вряд ли подберутся. Кстати, в этом плане вредителям очень не нравится известь и суперфосфат, поскольку эти вещества впитывают слизь и влагу с их тел, усложняя передвижение. Однако следует отметить, что в дождливую погоду эффективность такого средства снижается. Еще одним непреодолимым барьером для слизней и улиток является вода. Создать для них такую преграду можно при помощи пластиковых желобов, наполненных водой. Опять же, их можно вкопать в землю. Брюхоногие моллюски вряд ли пожелают преодолевать эти препятствия, а если и попытаются, то просто попадут в воду и не смогут из нее выбраться.
Улиток и слизней отпугивает запах пряных трав – петрушки, лавра, лаванды, розмарина, тимьяна, сантолина и шалфея. Посадив их по периметру грядок, вы тем самым защитите урожай от многих вредителей. Кроме того из чеснока, горчицы и горького перца можно приготовить специальные фито-настои, которые также отпугнут желающих полакомиться вашим урожаем.
Посадив их по периметру грядок, вы тем самым защитите урожай от многих вредителей. Кроме того из чеснока, горчицы и горького перца можно приготовить специальные фито-настои, которые также отпугнут желающих полакомиться вашим урожаем.
Классы Брюхоногие и двустворчатые моллюски
Кто такие брюхоногие моллюски?
Многочисленный класс моллюсков. Наземные, пресноводные и морские организмы. Хищники или растительноядные, встречаются случаи паразитизма. Некоторые морские виды ядовиты. Ряд видов являются
промежуточными хозяевами гельминтов. Наземные формы могут повреждать сельскохозяйственные культуры. Некоторые брюхоногие моллюски — объект промысла.
Особенности внешнего строения брюхоногих моллюсков
Форма тела и размеры
Тело брюхоногих моллюсков несегментоване, асимметричное. Оно состоит из трех основных отделов — головы, туловища и ноги. Размеры варьируют в широких пределах — от нескольких миллиметров до 60 см.
Покровы тела
Туловище моллюсков окружен кожной складкой — мантией. Пространство между туловищем и мантией называется мантийной полости. На спинной стороне тела, как правило, есть защитная внешняя черепашка (может редуцироваться) из белка (конхиолин), укрепленного известью. У большинства видов черепашка спирально закрученная, а в некоторых редуцирована. Отверстие раковины называется устьем.
Особенности внутреннего строения брюхоногих моллюсков
Опорно-двигательная система
В ноге брюхоногих моллюсков расположена сложная система полостей, заполненных кровью, образующих гидростатическое скелет. Движение обеспечивается благодаря волнам мышечных сокращений, возникающих на поверхности ноги, прилегающей к субстрату, которым передвигается моллюск.
Пищеварительная система
В состав пищеварительной системы брюхоногих моллюсков входят ротовая полость, глотка, пищевод, желудок, средняя кишка и задняя кишка. Кроме того, есть слюнные железы и печень. Для брюхоногих моллюсков характерно наличие в глотке языковидные терки (Радуль), покрытой хитиновыми зубцами. У хищных видов кроме терки также роговые или известковые челюсти для отрывания и перетирания кусков добычи.
Кроме того, есть слюнные железы и печень. Для брюхоногих моллюсков характерно наличие в глотке языковидные терки (Радуль), покрытой хитиновыми зубцами. У хищных видов кроме терки также роговые или известковые челюсти для отрывания и перетирания кусков добычи.
Дыхательная система
Типичная для типа Моллюски.
Вопрос. Как устроена дыхательная система моллюсков?
Кровеносная система
Типичная для типа Моллюски.
Вопрос. Как устроена кровеносная система моллюсков?
Выделительная система
Типичная для типа Моллюски.
Вопрос. Как устроена выделительная система моллюсков?
Нервная система
Разбросано-узлового типа.
Органы чувств представлены глазами, органами химического чувства, органами равновесия, главными щупальцами (органы осязания).
Размножение
Среди брюхоногих моллюсков встречаются как раздельнополые формы, так и гермафродиты. У некоторых видов развитие прямое, а в большинстве — косвенный, с метаморфозом.
У некоторых видов развитие прямое, а в большинстве — косвенный, с метаморфозом.
Кто такие двустворчатые моллюски?
Морские или пресноводные моллюски, имеющие двустороннюю симметрию. Донни, малоподвижные животные. Большинство двустворчатых моллюсков является фильтраторы, детрит-й планктонофагамы, некоторые — хищниками. Многие виды являются объектом промысла или разводятся искусственно. Некоторые морские виды способны повреждать гидротехнические сооружения.
Особенности внешнего строения двустворчатых моллюсков
Форма тела и размеры
Тело двустворчатых моллюсков несегментоване, имеет двустороннюю симметрию. Оно состоит из двух основных отделов — туловища и ноги. У некоторых видов нога редуцируется. Размеры варьируют в широких пределах — от нескольких миллиметров до 1,5 м.
Покровы тела
Туловище двустворчатых моллюсков окружен кожной складкой — мантией, образует сифоны (сквозь них моллюск втягивает и удаляет воду из мантийной полости). Черепашка состоит из двух створок, охватывающих тело с боков. В некоторых моллюсков раковина изнутри выстлана перламутром. Для большинства двустворчатых характерно наличие на ноге бисуснои железы (выделяет прочные нити — бисус, с помощью которых моллюск прикрепляется к субстрату).
Черепашка состоит из двух створок, охватывающих тело с боков. В некоторых моллюсков раковина изнутри выстлана перламутром. Для большинства двустворчатых характерно наличие на ноге бисуснои железы (выделяет прочные нити — бисус, с помощью которых моллюск прикрепляется к субстрату).
Особенности внутреннего строения моллюсков
Опорно-двигательная система
Типичная для типа Моллюски.
Вопрос. Как устроена опорно-двигательная система моллюсков?
Пищеварительная система
В состав пищеварительной системы двустворчатых моллюсков входят рот, пищевод, желудок, средняя кишка и задняя кишка. Рот имеет две пары лопастей направляют в него пищевые частицы. Слюнные железы, язык с теркой и глотка отсутствуют. Есть печень. Дыхательная система
Органом дыхания двустворчатых моллюсков являются жабры. У большинства видов они превращены в жаберные пластинки, но в наиболее примитивных форм жабры имеют вид перистых выростов. Кровеносная система Типичная для типа Моллюски.
Кровеносная система Типичная для типа Моллюски.
Вопрос. как устроена кровеносная система моллюсков?
Выделительная система Типичная для типа Моллюски.
Вопрос. как устроена выделительная система моллюсков? Нервная система разбросанных-узлового типа.
Органы чувств развиты слабо. У некоторых видов они представлены глазами, органами химического чувства и равновесия.
Размножение
Среди двустворчатых моллюсков чаще встречаются раздельнополые формы, но есть и гермафродиты. Развитие косвенный, с метаморфозом.
Классы моллюсков: Брюхоногие, Двустворчатые, Головоногие | Биология. Реферат, доклад, сообщение, краткое содержание, конспект, сочинение, ГДЗ, тест, книга
Вопрос 1. Объясните, как моллюски приспособлены к своей среде обитания.
Среди моллюсков есть обитатели водной и наземно-воздушной среды.
Многие как наземные, так и водные моллюски имеют раковину, которая и у тех, и у других выполняет роль пассивной защиты.
Важнейшим приспособлением к наземному образу жизни у сухопутных моллюсков является дыхание легкими.
Водные моллюски приспособлены к получению кислорода из воды — они дышат жабрами или поверхностью мантии.
Головоногие моллюски выработали особый — реактивный — способ передвижения, очень эффективный в водной среде.
Вопрос 2. Каковы черты сходства и различия брюхоногих и двустворчатых моллюсков?
Брюхоногие и двустворчатые моллюски являются многоклеточными животными, туловища которых не имеют сегментарного строения. Тело большинства видов данных классов моллюсков полностью или частично покрыто раковиной. У брюхоногих раковина дельная, часто асимметричная и закрученная, у двустворчатых — состоит из двух створок.
Голову, туловище и ногу можно различить только у брюхоногих, у двустворчатых голова отсутствует.
У брюхоногих хорошо развиты органы чувств — осязания, химического чувства, равновесия и зрения. У двустворчатых органы чувств развиты слабо.
У двустворчатых органы чувств развиты слабо.
Двустворчатые являются исключительно водными животными, в то время как среди брюхоногих есть обитатели и водной, и наземно-воздушной среды.
Вопрос 3. Перечислите способы защиты от врагов известных вам моллюсков.
Некоторые брюхоногие и практически все двустворчатые моллюски способны полностью или частично прятаться в раковину — это пассивный способ защиты.
Кальмары при спасении от преследования могут несколько десятков метров пролетать над водой. Материал с сайта //iEssay.ru
Некоторые моллюски, например каракатицы, осьминоги, в случае опасности способны менять окраску или выбрасывать чернильное вещество, вырабатываемое в особом органе — чернильном мешке. Перед применением этого средства защиты моллюск сначала темнеет, потом выбрасывает чернила в пленке, по форме напоминающей его тело. Преследователь хватает чернильную «бомбу» — пленка разрывается, чернила окрашивают большой объем воды и парализуют обоняние врага. Это спасает жизнь моллюска: он после выброса чернил быстро бледнеет и почти невидимым уплывает.
Это спасает жизнь моллюска: он после выброса чернил быстро бледнеет и почти невидимым уплывает.
Вопрос 4. Что позволяет головоногим быстро перемещаться в воде и спасаться от врагов?
Быстрое движение в воде головоногих моллюсков обеспечивается пульсирующим выбрасыванием воды из мантийной полости (реактивное движение). Такой способ движения позволяет развить значительную скорость плавания: кальмарам — до 40 км/ч, осьминогам — до 15 км/ч.
На этой странице материал по темам:- класс двустворчатые моллюски
- сочинение двустворчатые молюски
- брюхоногие моллюски краткое содержание
- способы защиты у головоногих моллюсков
- способ передвижения двухстворчатых моллюсок
Роботы копируют моллюска-чемпиона | Научные открытия и технические новинки из Германии | DW
Моллюски в большинстве своем неспособны перемещаться сколько-нибудь быстро, а брюхоногие моллюски даже являются символом медлительности: выражение «ползти как улитка» в той или иной форме имеется во многих языках мира. Исключение составляют лишь головоногие: кальмары, каракатицы, осьминоги. Что же касается двух других основных классов — двустворчатых и уже упомянутых брюхоногих, — то они ведут либо малоподвижный, либо вовсе неподвижный образ жизни. Однако и среди неторопливых моллюсков есть, конечно, свои чемпионы по скоростному передвижению. К ним относится двустворчатый моллюск, известный под названием «морской черенок» (Ensis ensis).
Исключение составляют лишь головоногие: кальмары, каракатицы, осьминоги. Что же касается двух других основных классов — двустворчатых и уже упомянутых брюхоногих, — то они ведут либо малоподвижный, либо вовсе неподвижный образ жизни. Однако и среди неторопливых моллюсков есть, конечно, свои чемпионы по скоростному передвижению. К ним относится двустворчатый моллюск, известный под названием «морской черенок» (Ensis ensis).
Моллюск-чемпион
Для этого вида, обитающего в песчаных грунтах и широко распространенного по всему миру, включая Северное и Средиземное моря, характерны сильно удлиненная раковина, формой напоминающая ножны, и мускулистая нога, позволяющая быстро закапываться в грунт. Конечно, быстро — понятие относительное, и та скорость, с которой морской черенок умудряется вертикально погружаться в песок, кому-то может показаться скромной, но для моллюсков это рекорд: сантиметр в минуту.
Умение столь стремительно зарываться в морское дно привлекло к черенку пристальное внимание ученых. Оказалось, что моллюск добивается такого эффекта, используя хитроумный прием, говорит Керстин Нордстром (Kerstin Nordstrom), научная сотрудница Института электроники и прикладной физики Мэрилендского университета в Колледж-Парке: «Моллюск рывком сдвигает створки своей раковины и тем самым взвихривает под собой песок. В результате плотный грунт на короткое время становится податливым, как бы разжижается. Образуется суспензия, в нее моллюск легко вдвигает свою ногу и может втянуться вглубь сразу на значительное расстояние. Затем черенок снова раздвигает створки раковины, и весь процесс повторяется».
Оказалось, что моллюск добивается такого эффекта, используя хитроумный прием, говорит Керстин Нордстром (Kerstin Nordstrom), научная сотрудница Института электроники и прикладной физики Мэрилендского университета в Колледж-Парке: «Моллюск рывком сдвигает створки своей раковины и тем самым взвихривает под собой песок. В результате плотный грунт на короткое время становится податливым, как бы разжижается. Образуется суспензия, в нее моллюск легко вдвигает свою ногу и может втянуться вглубь сразу на значительное расстояние. Затем черенок снова раздвигает створки раковины, и весь процесс повторяется».
Десятикратная эффективность
По сути дела, моллюск сам себе создает зыбучий песок, в котором быстро тонет. Больше всего исследователей поразила эффективность этого трюка: ни один из разработанных инженерами механизмов для земляных работ не в состоянии конкурировать с морским черенком, говорит Керстин Нордстром: «Существуют технические устройства с приводом типа гребного винта, есть шнековые буры, ввинчивающиеся в грунт, есть и машины, погружающиеся в песок за счет вибрации. Но морской черенок делает это намного эффективнее. Конкретно — в 10 раз эффективнее, чем любая из имеющихся сегодня технологий».
Но морской черенок делает это намного эффективнее. Конкретно — в 10 раз эффективнее, чем любая из имеющихся сегодня технологий».
Специалисты Мэрилендского университета решили в лаборатории искусственно воспроизвести прием, применяемый морским черенком, и сконструировали механизм, в котором гидравлический привод мог рывком сводить и разводить две продолговатые металлические створки.
Прототип RoboClam
Прототип получил название RoboClam, то есть «робот-двустворчатый моллюск». А чтобы убедиться в том, что он действительно способен разжижать песок, исследователи производили испытания в специальном контейнере с лазерной подсветкой. «Мы разработали такую опытную схему, которая позволяет нам как бы заглянуть внутрь песка, — поясняет Керстин Нордстром. — Мы взяли небольшой аквариум, налили в него подкрашенной воды и засыпали доверху мелкими крупинками, имитирующими песок. Этот аквариум освещается с разных сторон лучами лазера. В результате частицы красящего пигмента в воде начинают светиться, крупинки же выглядят темными точками на светлом фоне.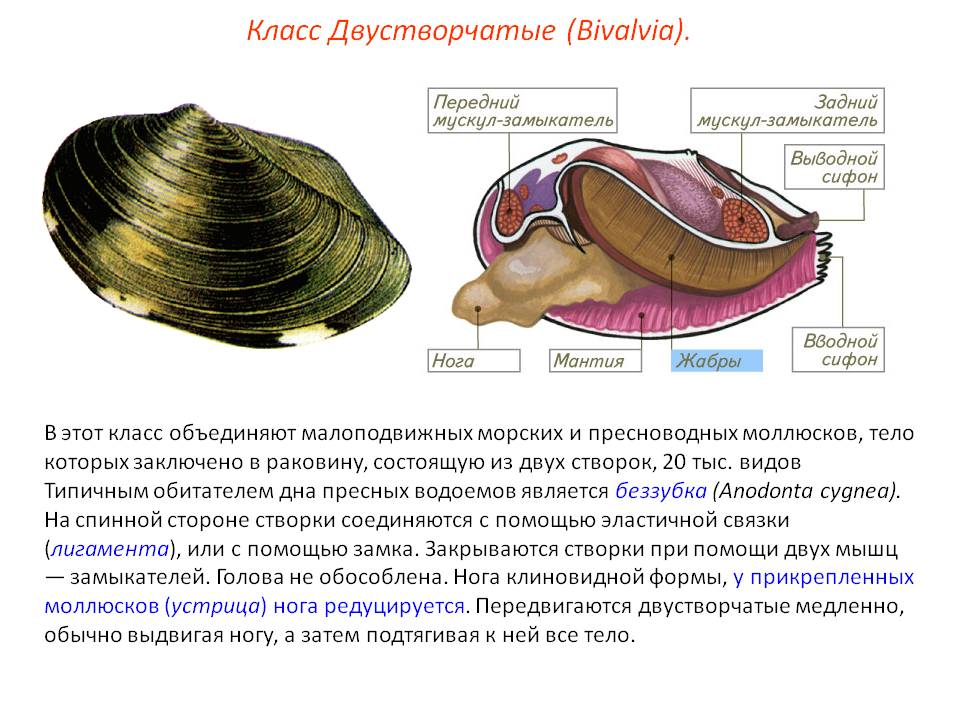 Запустив в аквариум нашего робота, мы можем наблюдать, что происходит с крупинками».
Запустив в аквариум нашего робота, мы можем наблюдать, что происходит с крупинками».
Оказалось, что если робот машет створками в определенном ритме, крупинки в лазерном свете действительно начинают вести себя как зыбучий песок: легко теряют сцепление друг с другом, в результате чего вязкость среды резко падает.
Проблема — в размерах
Своего робота разработчики представили на прошедшей недавно в Денвере, штат Колорадо, весенней сессии Американского физического общества. Но говорить о каком-либо практическом применении этого устройства пока не приходится. Сегодняшний прототип для этого просто слишком мал — размером с одноразовую газовую зажигалку, то есть примерно с того самого моллюска, что послужил образцом при его создании. Мэрилендские специалисты работают теперь над устройствами более внушительных размеров, полагая, что они найдут применение в роботах, обследующих в автономном режиме морское дно или подводные трубопроводы. «Большинство этих подводных роботов невелики, а потому легко уносятся морскими течениями, — поясняет Керстин Нордстром. — Чтобы удержаться на одном месте, они должны как-то закрепиться на дне. Но это требует довольно значительного расхода энергии, а ее мало, ведь все эти роботы питаются от аккумуляторов».
— Чтобы удержаться на одном месте, они должны как-то закрепиться на дне. Но это требует довольно значительного расхода энергии, а ее мало, ведь все эти роботы питаются от аккумуляторов».
Вот здесь-то и пригодится позаимствованный у моллюска высокоэффективный механизм погружения в грунт. Впрочем, интерес к разработке проявляют уже и военные: они намерены использовать эту новую технологию для разрыхления морского дна с целью обезвреживания донных мин.
Коллекция моллюсков Государственный Дарвиновский музей
Хранитель А.Р. Алякринский.
Насчитывает 20325 экземпляров. Коллекция начала формироваться А.Ф. Котсом в начале ХХ века. Среди ранних поступлений – коллекция морских тропических моллюсков Е.К. Попова и коллекция наземных брюхоногих моллюсков с Гавайских островов. В 70-х годах в музей поступило более 1000 экземпляров тропических моллюсков, собранных во время Тихоокеанских экспедиций Института океанологии Академии Наук (1971 и 1977 гг.) на литорали островов Новая Гвинея, Фиджи, Западное Самоа, Новая Каледония, Науру, Тувалу, Лорд-Хау, архипелага Новые Гебриды.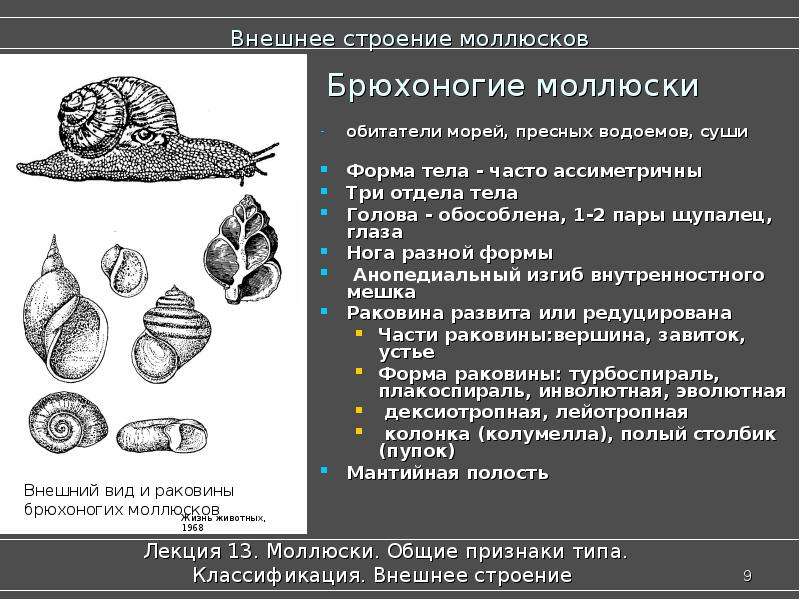 В период с 1981 по 1986 гг. московским коллекционером К.Н. Гайденко было подарено музею 7479 раковин морских брюхоногих и двустворчатых моллюсков. В 1992 г. музеем была приобретена ценная коллекция наземных моллюсков России и сопредельных стран, собранная А.Г. Кузнецовым (около 1100 экз.). В последние годы коллекция интенсивно пополняется силами сотрудника отдела фондов ГДМ А.Р. Алякринского (около 4000 экз. морских, пресноводных и наземных моллюсков из Средней Азии, Сибири, Дальнего Востока, различных регионов Юго-Восточной Азии).
В период с 1981 по 1986 гг. московским коллекционером К.Н. Гайденко было подарено музею 7479 раковин морских брюхоногих и двустворчатых моллюсков. В 1992 г. музеем была приобретена ценная коллекция наземных моллюсков России и сопредельных стран, собранная А.Г. Кузнецовым (около 1100 экз.). В последние годы коллекция интенсивно пополняется силами сотрудника отдела фондов ГДМ А.Р. Алякринского (около 4000 экз. морских, пресноводных и наземных моллюсков из Средней Азии, Сибири, Дальнего Востока, различных регионов Юго-Восточной Азии).
Коллекция конусов Дарвиновского музея содержит 502 экземпляра 136 видов. Ее иллюстрированное описание издано в виде каталога в 2005 г.
Моллюски мурексы (семейство Muricidae), как и конусы, хищники. Основной объект их питания – двустворчатые моллюски. Чтобы съесть такого моллюска мурексы просверливают в раковине жертвы маленькое отверстие, или же раздавливают последнюю, зажав ее между роговой крышечкой и устьем собственной раковины. Многие мурексы имеют очень красивую раковину. У некоторых видов она украшена причудливым выростами и шипами. В коллекции музея хранится 490 экземпляров мурексов.
У некоторых видов она украшена причудливым выростами и шипами. В коллекции музея хранится 490 экземпляров мурексов.
Ципреи, или фарфоровые улитки, также очень красивы. Поверхность их раковин абсолютно гладкая, словно отполированная. В течение столетий раковины некоторых видов ципрей использовались в качестве платежного средства, выполняя роль своего рода разменной монеты, причем не только в тропических странах, в акватории которых они обитают, но и далеко за их пределами. В музее имеется 506 экземпляров ципрей, часть из которых выставлена в одной из витрин зала «Зоогеография».
В отличии от предыдущих семейств морских моллюсков, представители семейства ахатинеллид (Achatenillidae) ведут наземный образ жизни. Все они обитают на тропических островах Тихого океана, в основном на Гавайях. Их раковины имеют очень яркую окраску и отличаются сильной индивидуальной изменчивостью. В музее хранится около 1400 экземпляров представителей данного семейства.
В большинстве своем коллекция моллюсков Дарвиновского музея тщательно этикетирована, хорошей сохранности и представляет определенную научную ценность для специалистов-малакологов.
Раковины морских гребешков Chlamys rubida Hinds (вверху слева), Pecten irradians Lam. (вверху справа), Lyropecten nodosa L. (внизу).
Раковины конусов. Слева направо: Conus tessulatus Born., Conus bengalensis Ocut., Conus amadis Gmelin, Conus vitulinus Hwass.
Урок 18. Многообразие моллюсков. Брюхоногие и Двустворчатые | Поурочные планы по биологии 7-8 класс
Урок 18. Многообразие моллюсков. Брюхоногие и Двустворчатые
09.01.2015 3138 0Тема урока: Многообразие моллюсков, значение их в природе и в жизни человека. Двустворчатые и брюхоногие моллюски.
Цель урока: Расширить знания о разнообразии моллюсков, их приспособленности к факторам среды, показать их роль в природе и практическое значение для человека.
Оборудование: Таблицы, рисунки моллюсков, раковины морских моллюсков,
диафильм «Многообразие моллюсков».
Ход урока
I. Организационный момент
II. Актуализация опорных знаний.Самостоятельная работа по вариантам
Вариант I
Выпишите цифры, которые соответствуют сведениям, относящимся к большому прудовику.
1. Тело животного мягкое, есть мантия.
2. Снаружи тело покрыто двустворчатой раковиной.
3. Раковина единая, имеет вид башенки.
4. Хищник: поедает рыб, крабов, моллюсков.
5. Питается водными растениями.
6. Дышит атмосферным воздухом.
7. Дышит кислородом, растворенным в воде, проникающим через жабры.
8. Кровеносная система незамкнутая.
9. Кровеносная система замкнутая.
10. Есть
глаза, расположенные у основания щупалец.
Есть
глаза, расположенные у основания щупалец.
11. Органы зрения отсутствуют.
12. Нервная система состоит из нескольких пар узлов.
13. Нервные клетки не образуют узлов.
14. Обитает в пресных водоемах.
15. Обитает в морях.
16. Голова хорошо развита.
17. Личинки паразитируют на теле рыб.
18. Гермафродит.
19. Имеет терку (радулу).
Ответы: I, 3, 5, 6, 8, 10, 12, 14, 16, 18, 19.
Вариант II
Из перечисленных признаков выберите те, которые характерны для двустворчатых моллюсков.
1. Тело разделено на туловище и мускулистую ногу.
2.
Передвигаются
за счет воды, выталкиваемой из мантийной полости.
3. Органами пищеварения служат рот с роговыми челюстями, глотка, пищевод, желудок, кишечник, заканчивающийся анальным отверстием.
4. Органами дыхания служат парные жабры, имеющие вид листочков.
5. Развитие протекает без превращений, стадии личинки отсутствуют.
6. Головы нет.
7. Раздельнополы.
8. Раковина состоит из трех слоев: рогового, фарфоровидного и перламутрового.
9. Створки раковины закрываются за счет сокращения мышц-замыкателей.
10. Живут в пресных водоемах и очищают их от зарастания водными растениями.
11. Органы чувств развиты слабо.
12. Фильтраторы воды.
13.
Ведут
малоподвижный образ жизни.
14. Кровеносная система замкнутая. Она состоит из сердца и сосудов.
15. Хищники: поедают различных беспозвоночных животных.
16. Дышат кислородом, растворенным в воде.
17. Тело животного мягкое, есть мантия.
18. Имеют терку (радулу).
19. Среди них присутствуют наземные виды.
Ответы: 1, 4, 6, 7, 8, 9, И, 12, 13, 16, 17.
III. Изучение нового материала’
500-600 млн. лет назад в древнем океане жили
разнообразные кольчатые черви. Одни из них были хищниками, другие питались
водорослями, третьи — органическими остатками, четвертые были фильтраторами. И
вот среди этих червей выделилась небольшая группа, у которой на покровах тела
стали появляться защитные образования в виде простых известковых игл. Со
временем эти образования усложнялись, и в конце концов получилось то, что
сегодня мы называем моллюсками. Наиболее примитивный представитель типа —
хитон, обитатель прибрежных подводных скал, — сохранил членистость раковины и
туловища (кадр 3).
Наиболее примитивный представитель типа —
хитон, обитатель прибрежных подводных скал, — сохранил членистость раковины и
туловища (кадр 3).
За прошедшие миллионы лет моллюски заселили всю Землю. По количеству видов они занимают второе место после насекомых. Известно около 130 тысяч видов ныне живущих моллюсков и примерно 50 тысяч видов вымерших.
Когда впервые пересеклись пути человека и моллюска, сейчас установить почти невозможно. Однако раскопки стоянок и захоронений древних людей свидетельствуют о том, что это было очень давно. Так, в Северной Америке, в штате Миннесота, при строительстве железной дороги был найден скелет девочки ледникового периода, на шее которой висела раковина из далеких южных морей. В 1876 г. Во Франции тоже при постройке железной дороги было найдено древнее захоронение. Вместе с людьми лежало оружие, инструменты, предметы обихода и… ожерелье из морских раковин.
Вообще все раковины, найденные при раскопках, можно
отнести к двум группам: одни представляли для человека эстетическую ценность,
содержимое других употребляли в пищу (например, найденные скопления на о. Крит
и известные под названием кухонные кучи).
Крит
и известные под названием кухонные кучи).
Можно предположить, что моллюски и их раковины сыграли не последнюю роль в судьбе человечества. С моллюсками связаны многочисленные истории и легенды.
Познакомимся с некоторыми представителями классов Двустворчатые и Брюхоногие.
О По ходу изучения темы составьте в тетради таблицу «Значение моллюсков».
Среди брюхоногих моллюсков наиболее известны различные
виды ципрей (кадр 8). Раковины ципрей, так называемые каури, использовались
когда-то в качестве денег в Японии, Китае и других странах бассейна Тихого
океана (кадр 9). Для этого использовались раковины двух видов брюхоногих моллюсков
— ципреи моны и ципреи апнулюс. Эти раковины как бы самой природой
предназначены выполнять функции денег — имеют гладкую поверхность, округлую
форму и большую прочность. Предполагается, что впервые раковины каури стали
использовать в качестве денег в Китае примерно полторы тысячи лет до нашей
эры. Позднее они проникли в Корею и Японии. Известно также, что деньги каури
были распространены в Индии, Сиаме, на Филиппинах, в странах Африки.
Использовались каури и на Руси под названием ужо- вок, зерновок, змеиных
головок. Было это в так называемый безмонетный период (XII—XIV вв.). Их и
сейчас находят при раскопках в Новгородской, Псковской областях.
Позднее они проникли в Корею и Японии. Известно также, что деньги каури
были распространены в Индии, Сиаме, на Филиппинах, в странах Африки.
Использовались каури и на Руси под названием ужо- вок, зерновок, змеиных
головок. Было это в так называемый безмонетный период (XII—XIV вв.). Их и
сейчас находят при раскопках в Новгородской, Псковской областях.
Следующий моллюск, с которым мы познакомимся — рапана.
Его раковины в качестве сувениров продаются во всех черноморских городах (кадр
10; демонстрация раковины) и вам они всем хорошо знакомы. Но мало кто знает,
что появилась рапана в Черном море всего пятьдесят лет назад. До 1974 года она
была известна только в Японском.и Желтом морях и на японском побережье Тихого
океана. Как рапана могла перебраться из Японского моря в Черное? Об этом можно
только догадываться. Откладываемые рапаной яйца снабжены своеобразным
скафандром — яйцевой капсулой, которая имеет вид цилиндрического стручка,
прикрепляющегося подошвой к субстрату. В каждой такой капсуле содержится от
200 до 1000 яиц. Через месяц из этих яиц развиваются личинки-велигеры
(парусники), снабженные двулопастным парусом. Вероятно, именно в виде личинок и
были перенесены рапаны ка- ким-то кораблем из Японского в Черное море. Там они
хорошо прижились, только измельчали, т.к. вода в Черном море менее соленая, чем
в Японском. Рапаны — хищники.
В каждой такой капсуле содержится от
200 до 1000 яиц. Через месяц из этих яиц развиваются личинки-велигеры
(парусники), снабженные двулопастным парусом. Вероятно, именно в виде личинок и
были перенесены рапаны ка- ким-то кораблем из Японского в Черное море. Там они
хорошо прижились, только измельчали, т.к. вода в Черном море менее соленая, чем
в Японском. Рапаны — хищники.
Нападают они на двустворчатых моллюсков. Гипобронхиальная железа рапаны содержит в составе слизи ядовитые вещества, оказывающие на двустворчатых моллюсков парализующее действие. В Черном море, например, они полностью уничтожили устриц на Гудаутской промысловой устричной ферме. Вот почему в настоящее время устричные парки обязательно отгораживают от моря заборами.
Очень интересны моллюски, относящиеся к семейству
Мурицид. Их гипобронхиальная железа состоит из трех отделений. Клетки двух
выделяют слизь, а центральное содержит мелкие гранулы, придающие клеткам молочно-белый
цвет.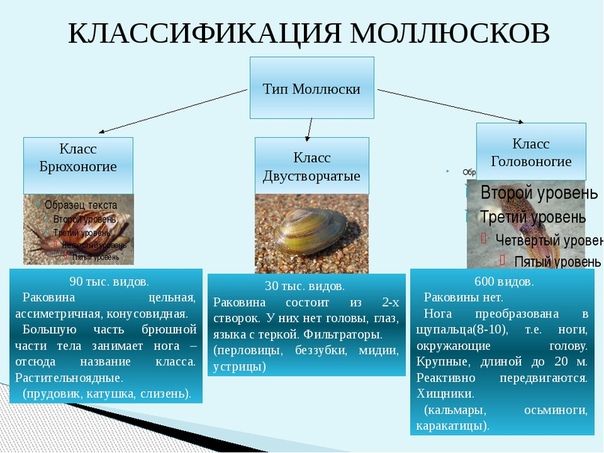 Это пурпур. Соприкасаясь с воздухом, они начинают менять цвет. Существует
легенда, рассказывающая о том, как был открыт пурпур моллюсков. Как-то царь
финикийского города Тира по имени Феникс прогуливался по берегу моря со своей
собакой. Собака рылась в песке, трепала водоросли, и когда подбежала к царю, он
заметил, что морда ее покрыта пятнами кроваво-красного цвета. Царь догадался,
что на морде собаки была не кровь, а что-то другое. Так три с половиной тысячи
лет назад был открыт пурпур моллюсков.
Это пурпур. Соприкасаясь с воздухом, они начинают менять цвет. Существует
легенда, рассказывающая о том, как был открыт пурпур моллюсков. Как-то царь
финикийского города Тира по имени Феникс прогуливался по берегу моря со своей
собакой. Собака рылась в песке, трепала водоросли, и когда подбежала к царю, он
заметил, что морда ее покрыта пятнами кроваво-красного цвета. Царь догадался,
что на морде собаки была не кровь, а что-то другое. Так три с половиной тысячи
лет назад был открыт пурпур моллюсков.
Пурпур Мурицид разного цвета. Пурпур Мурекса
брандариса — красно-фиолетового цвета, мурексатрункул юса —
голубовато-фиолетового, у рапаны (относящейся к тому же семейству) пурпур
чисто фиолетового цвета. (Кадры 11, 12). Процесс получения пурпура из моллюсков
очень трудоемкий. Тела моллюсков вынимают из раковин, вымачивают в соленой
воде, вялят на солнце, варят, а в результате из десяти тысяч моллюсков получают
всего 1 грамм краски. Шерстяные и льняные ткани погружают в раствор, и л ишь
при высыхании на солнце они приобретают свой окончательный цвет. Окрашенные
таким образом ткани очень дорогие. Например, в Древнем Риме платье, окрашенное
пурпуром, в перерасчете на современные деньги стоило 112 000 долларов.
Окрашенные
таким образом ткани очень дорогие. Например, в Древнем Риме платье, окрашенное
пурпуром, в перерасчете на современные деньги стоило 112 000 долларов.
Существуют ли моллюски, опасные для человека? Да. В семействе Конусов есть моллюски с очень красивыми конусовидными раковинами (кадр 15). Они — хищники. Обитают на мелководьях, среди кораллов и водорослей. Пищей им служат различные черви, моллюски, рыбы. Охотятся они с помощью секрета ядовитых желез, который стекает по каналам, идущим внутри зубов радулы. Считается, что яд этот оказывает паралитическое действие. Конусы очень опасны и для человека, нападения их нередко смертельны. Существует мнение, что конусы опаснее акул, так как, например, в бассейне Тихого океана от акул за год погибает в среднем один человек, а от конусов — двое-трое.
Громадные раковины улиток-тритонов использовались
народами Океании для подачи звуковых сигналов. Теперь это просто желанный
сувенир для туристов (кадр 16). Пустые раковины морских улиток используются
различными видами рака-отшельника в качестве временного жилища. Подрастая, рак
меняет раковину на более вместительную (кадр 17).
Пустые раковины морских улиток используются
различными видами рака-отшельника в качестве временного жилища. Подрастая, рак
меняет раковину на более вместительную (кадр 17).
Среди морских брюхоногих есть виды, лишенные раковины, например, клионы. Эти свободноплавающие моллюски — излюбленная пища некоторых китов (кадр 18).
Виноградная улитка имеет ценность как продукт питания и в ряде стран употребляется в пищу в вареном виде. Особенный спрос она находит в Испании, во Франции, в Италии; здесь уже во времена древнего Рима ее весьма ценили как пищевой продукт. При массовом размножении может приносить вред плодоводческим хозяйствам (кадр 20).
Однако наиболее значительный вред сельскому хозяйству
приносят наземные легочные улитки, лишенные раковины, — так называемые голые
слизни. Массовое появление слизней сопровождается большими повреждениями
посевов, иногда на десятках тысяч гектаров. Причем часть посевов гибнет
полностью. Кроме того, слизни уничтожают огромное количество огородных культур
(кадр 21).
Кроме того, слизни уничтожают огромное количество огородных культур
(кадр 21).
Учащиеся завершают заполнение таблицы по брюхоногим. Подводятся итоги.
Домашнее задание
§37, вопросы после параграфа, закончить заполнение таблицы, используя сведения о корабельном черве (стр. 110) и другие данные параграфа.
Брюхоногие и двустворчатые моллюски
Брюхоногие и двустворчатые моллюскиПо окончании этой лабораторной работы вы должны уметь:
- Идентифицировать окаменелость как брюхоногого или двустворчатого моллюска и быть в состоянии идентифицировать оба группы в порядке, используя предоставленные диаграммы .
- Уметь определить вероятный образ жизни брюхоногого моллюска или двустворчатого моллюска.
- Знать строение скелета и материал каждого из эти животные.
- Знать экологические характеристики каждого из этих животные.
- Знать геологический диапазон каждой из этих групп.

- Знать несколько важных родов (упомянутых по имени в этот раздаточный материал) для каждой группы.
Дисплей: брюхоногие моллюски и двустворчатые моллюски
Брюхоногие моллюски
I. Морфология мягких частей: В полимерном блоке определите головку и остальную часть стопы.
II. Морфология твердой части: Найдите следующие признаки: оборот, апертура, костная мозоль, сифональная вырезка, сифональный канал
III.Классификация: Разработана новая кладистическая классификация моллюсков.
опубликовано в 2005 г. Оно сложное и требует знания морфологии мягких тканей.
животных. По этой причине мы немного окунемся в традиционную
отряды переднежаберных: археогастроподы, мезогастроподы и неогастроподы.
Есть несколько функций оболочки, которые являются универсальными для каждого из этих
группы. Ископаемые брюхоногие помещаются в отряды путем идентификации семейства,
затем посмотрите, в каком порядке находится эта семья. В кладистическом анализе
эти заказы просто разваливаются. ВАМ НЕ НУЖНО ЗАПОМИНАТЬ КЛАССИФИКАЦИЮ.
Я хочу, чтобы вы были достаточно знакомы с морфологией улиток, чтобы определить их местонахождение.
в заказ с учетом описания заказов.
В кладистическом анализе
эти заказы просто разваливаются. ВАМ НЕ НУЖНО ЗАПОМИНАТЬ КЛАССИФИКАЦИЮ.
Я хочу, чтобы вы были достаточно знакомы с морфологией улиток, чтобы определить их местонахождение.
в заказ с учетом описания заказов.
Археогастроподы (№ 579, 1305, 578): от палеозоя до современности, травоядные
- брюхоногие моллюски с высокой скоростью расширения и небольшим скручиванием или без него: блюдечки и морское ушко
- несколько групп с низкими скоростями перевода и простыми отверстиями
- верхняя часть и тюрбан-чашечки
Мезогастроподы (№ 1604, 474): от позднего палеозоя до недавнего времени, травоядные и хищники
- от закругленных до остроконечных
- от простой до сложной апертуры
- иногда сифонная выемка
Необрюхоногие (№ 459, 1303, 473): от юры до современности
- отверстия обычно удлиненные
- сифонные выемки и каналы
Легочные (№): наземные улитки
- тонкие гладкие оболочки
- простые отверстия
IV. Образ жизни и морфология
Образ жизни и морфология
- несвернутые или плоскодонные раковины (№ 1189, морское ушко) – часто живые прикрепленные качаться в приливной или сублиторальной
- гладкие округлые формы (№ 458) — глубоко закапываются в песок и грязь
- высокие неорнаментированные шпили (коричневые без номеров) — неглубоко зарытые в песок
- толстое украшение (#20, 1555)-защита от раздавливающих хищников (как крабы), поэтому живут на открытом воздухе на каменистом или песчаном субстрате
Двустворчатые моллюски
И.Морфология мягких частей . В смоляном блоке смотрите специализированный морфология мидии. Сравните с более общей морфологией двустворчатых моллюсков. на схеме в нашей книге.
II. Морфология твердой части. Найдите следующие признаки: зубной ряд, шарнир, паллиальная линия и синус, мышечные рубцы.
III. Классификация: Традиционная классификация двустворчатых моллюсков. находится в еще большем беспокойстве, чем брюхоногие моллюски. Признать несколько групп
их общими именами (ниже).
находится в еще большем беспокойстве, чем брюхоногие моллюски. Признать несколько групп
их общими именами (ниже).
- Мидии (987): обитают в каменистой литорали, прикрепляясь к камням с нити силы, непревзойденной конструкциями любых других существ, включая людей. Без сифонов, уменьшенная ножка.
- Пектины (№): свободно плавающие. Одна крупная мышца закрывает скорлупу (цилиндрический «гребешок», который вы едите. Ищите мышечный шрам.
- Устрицы (# 1704, № #): некоторые прикреплены, другие покоятся в мягком субстрате. Широкие экологические допуски.Раковины асимметричные, часто неправильной формы.
- Раковины ковчега (#21): длинный прямой шарнир, таксодонтный зубной ряд, массивные ребра. Живи привязанным к камням
- Венероиды: венериные моллюски (поедание моллюсков) (№ 1608, 1196), моллюски-бритвы (№ 496),
гигантские моллюски (без #) и другие группы. Гетеродонтный зубной ряд. «Моллюск»-
форменные норники.

- Рудисты (Hippuritoida) (№№): вымершие моллюски в форме кораллов с один клапан в виде «крышки» на конусообразном нижнем клапане. Строители рифов.
IV. Образ жизни: раковины соответствуют требованиям окружающей среды
- роющие двустворчатые моллюски (№ 1236, 25): симметричные створки, два одинаковых аддуктора
мышцы, отчетливая мантивная линия
- глубокие норы — глубокий мантийный синус, тонкая, гладкая оболочка
- неглубокие норники — неглубокий мундштук, толстый или сильно орнаментированный ракушки
- быстро роющий (движущиеся отложения, много хищников) — маленькая раковина, сплюснутая оболочка
- медлительный роющий — большие раковины, выпуклые раковины
- двустворчатые моллюски (пектиновая коробка, № 596): тонкая раковина, симметричные створки
с «крыльями» (ушными раковинами), одиночной большой приводящей мышцей
- плоские поплавки: один клапан более плоский, чем другой
- вертикальные поплавки: створки одинаково закругленные
- мелководье: ребра Глубоководные
- : гладкие
- двустворчатые эпифауны:
- биссус (скреплен нитями) (без #): приводящие мышцы разного размера,
зияние биссуса, либо очень заостренный клюв, либо длинный прямой шарнир с «крыльями».

- цементированные (#382): сильно асимметричные створки, шипы, могут иметь одиночные мышца
- мягкая подложка (#1738): очень асимметричные клапаны, меньшего размера в виде крышки створка на глубоко вогнутой нижней створке.
- биссус (скреплен нитями) (без #): приводящие мышцы разного размера,
зияние биссуса, либо очень заостренный клюв, либо длинный прямой шарнир с «крыльями».
Дисплей: другие моллюски
Скафоподы (#843) называются бивневыми панцирями. Организм живет головой вниз закрепился в осадке. Они очень мало изменились за геологическое время, и никогда не отличались большим разнообразием.
Tentaculites : (#155) этот организм является одним из нескольких малоизвестных и загадочные ранние существа, которые интерпретируются как моллюски. Крошечный конический оболочка — кальцит. Tentaculites жили в раннем палеозое.
Вопросы
1. #1739 — Этот мезогастропод принадлежит к семейству Natacidae, лунных улиток.
Эти улитки — хищники, они ловят двустворчатых моллюсков и просверливают отверстия в
оболочки, затем переваривая содержимое. Посмотрите внимательно на натацид и его добычу,
и сделайте вывод, является ли эта улитка эпифауной или инфауной.
Посмотрите внимательно на натацид и его добычу,
и сделайте вывод, является ли эта улитка эпифауной или инфауной.
2. #123 -Красивый перламутровый перламутровый слой внутри отметин ракушки место прикрепления мягких тканей улитки к раковине.Что это было жизненная привычка улитки? Каковы ваши доказательства?
3. #1329 — Как сохраняется этот брюхоногий моллюск?
4. Этот легочный имеет типично тонкую гладкую оболочку. Зачем пульмонировать улиток эволюция таких раковин, в то время как морские улитки неуклонно становились толще, более сильно скульптурные снаряды?
5. Расположите каждого из этих брюхоногих в порядке Archeogastropoda, Mesogastropoda, или Neogastropoda: № 1242, 1307, 1667, 1017, 1877.
6. #1633 — обратите внимание на асимметричную форму этого пектина. Как вы это объясните?
7. #26 — Обратите внимание на следы на панцире какого-то скучного животного. Какие защита этого двустворчатого моллюска развилась против таких нападений? Каковы ваши доказательства?
8. Нет # — Назовите возможные функции шипов у этого двустворчатого моллюска.
Нет # — Назовите возможные функции шипов у этого двустворчатого моллюска.
9. Определите вероятный образ жизни каждого двустворчатого моллюска и морфологические признаки. подтверждающие вашу гипотезу: № 1074, 22, 25, 592.
Брюхоногие и двустворчатые моллюски
Брюхоногие и двустворчатые моллюскиПо окончании этой лабораторной работы вы должны уметь:
- Идентифицировать окаменелость как брюхоногого или двустворчатого моллюска и уметь идентифицировать двустворчатых моллюсков до уровня отряда, используя предоставленную таблицу.
- Уметь определить вероятный образ жизни брюхоногого моллюска или двустворчатого моллюска.
- Знать строение скелета и материал каждого из эти животные.
- Знать экологические характеристики каждого из этих животные.
- Знать геологический диапазон каждой из этих групп.
- Знать несколько важных родов (упомянутых по имени в
этот раздаточный материал) для каждой группы.

Дисплей: брюхоногие моллюски и двустворчатые моллюски
Брюхоногие моллюски
I. Морфология мягких частей:
- A. В полимерном блоке найдите головку и остальную часть стопы. Не забудьте перевернуть его, чтобы посмотреть на дно улитки.
- B. #1880 Littorina & #1326 : Жаберная крышка — это маленький люк, который делают некоторые улитки.
Он может быть как белковым, так и кальцинированным. Оперкулюм прилагается
к ноге улитки, и когда она втягивается в свою раковину,
operculum закрывает отверстие.
II. Морфология твердой части: № 486 и без номера Fasciolaria.
Найдите эти признаки: оборот, апертура,
костная мозоль, сифональная вырезка, сифональный канал. Обратите внимание, что внутренности
современные улитки на выставке гладкие и блестящие. Брюхоногие моллюски
обычно кладут гладкий слой перламутра (смесь арагонита и
белок) на той части оболочки, с которой регулярно контактирует их тело.
- A. Ненумерованные фасциолярии
-
Б. № 486 — Фасциолярия
III. Классификация: Разработана новая кладистическая классификация моллюсков.
опубликовано в 2005 г. Оно сложное и требует знания морфологии мягких тканей.
животных. По этой причине вам не потребуется изучать таксономию брюхоногих моллюсков.
IV. Образ жизни и морфология: брюхоногих моллюсков живут в широком
диапазон сред.Как группа, они имеют одни из самых широких
экологическая толерантность любых беспозвоночных. Большая часть широкого
морфология брюхоногих моллюсков – это приспособления к различным
среды:
- A. скалистые поверхности в воде с высокой энергией:
- 2.
 несвернутые или плоскодонные раковины ( # 1879 Colisella , морское ушко ) — часто живые прикрепленные
качаться в приливной или сублиторальной
несвернутые или плоскодонные раковины ( # 1879 Colisella , морское ушко ) — часто живые прикрепленные
качаться в приливной или сублиторальной
- B. рытье глубоких нор в песке и иле:
- гладкие закругленные формы ( #458 Ampullonatica , #474 Natica ),
обтекаемой формы ( ненумерованные оливковые скорлупы ). Многие из этих
роющие улитки — хищники моллюсков, преследующие моллюсков через
отложений и бурения в них.Оливки — самые быстрые роющие норы среди брюхоногих моллюсков.
- C. неглубокое рытье нор в песке
— эти брюхоногие передвигаются ногой прямо под поверхностью и частично заглубляют раковину.
- высокие гладкие шпили или высокие шпили с гофрами, помогающими
удерживать раковину на месте, пока лапа брюхоногого моллюска проталкивается вперед
песок ( коричневый ненумерованный, четыре белых ненумерованных экземпляра )
- D.
 толстый или сложный орнамент
толстый или сложный орнамент
- 1.утолщение панциря с выступами и выпуклостями может быть защитой от раздавливающих хищников (например, крабы) для улиток, живущих на открытом воздухе на каменистом или песчаном субстрате ( #1555 Busycon , #20 Salterella , #1645 Ecphora , четыре ненумерованных экземпляра )
- 2. Для улиток, живущих в зыбучем песке или иле, выступы снегоступов могут стабилизировать их (три ненумерованных экземпляра)
- 1. широкие раковины с куполообразными шпилями или широкие треугольные
поперечные сечения и плоское днище, которые эффективно прижимаются к скале:
тюрбаны ( # 1305 и # 1189 Tegula ) и топы ( ненумерованная розовая ракушка )
- A.Палеозойские формы: (#578 Straparolus , #579 Worthenia, #1328, #1668 Trepospira, #1771-75 Bellerophon 6 , #981 52 0 Busanopsi). Немного ранние брюхоногие моллюски не образовывали спирали, а скручивались почти в один самолет (№578, №981, №1771-75).
- B.
 Cowries : Перламутровая внешняя поверхность раковины говорит вам
что мягкие ткани улитки обернуты снаружи
оболочка в жизни, маскирующая оболочку.Каури травоядны, многие
из них днем прячутся в расщелинах рифов и выныривают только
ночь. У взрослого каури сложно разглядеть спираль улитки.
потому что внешний край раковины утолщается и загибается внутрь, когда
животное достигает половой зрелости.
Cowries : Перламутровая внешняя поверхность раковины говорит вам
что мягкие ткани улитки обернуты снаружи
оболочка в жизни, маскирующая оболочку.Каури травоядны, многие
из них днем прячутся в расщелинах рифов и выныривают только
ночь. У взрослого каури сложно разглядеть спираль улитки.
потому что внешний край раковины утолщается и загибается внутрь, когда
животное достигает половой зрелости. - C. Шишки : Раковины шишек являются хищниками. Радула — это твердая ротовая часть в форме рашпиля, которую многие другие улитки используют для соскребания водорослей с камней — в Conus специализируется на гарпуне, который впрыскивает токсины в добычу.Многие Conus опасны для человека.
- D. Crepidula : Эти брюхоногие прикрепляются к раковинам
живые или мертвые существа и фильтрующие корма. Они могут образовывать большие
брачные стопки, где каждая улитка становится противоположным полом улитки.
укладывается поверх.

- E. Верметиды: Эти брюхоногие моллюски очень странные. Они ни травоядные и не хищники. Вместо этого они строят длинные трубы вместо спиральные раковины и фильтрующая подача на одном конце, как у многих морских черви.Их раковины выглядят как большие трубки серпулидных червей, но внутренности имеют блестящий перламутр, как и у других брюхоногих моллюсков.
- Некоторые брюхоногие моллюски не строят минерализованные скелеты, в том числе слизни.
на суше и голожаберные и морские зайцы в океане. У них нет
летопись окаменелостей, но мы, вероятно, увидим некоторые из них во время нашей экскурсии.
Двустворчатые моллюски
I. Морфология мягких частей . В блоке смолы посмотрите специализированный морфология мидии. Сравните с более общей морфологией двустворчатых моллюсков. на схеме в нашей книге.
II. Морфология твердой части.
- Найдите следующие признаки: зубной ряд (E), шарнир (F), мышечные шрамы (C).
 То
Блестящая линия, идущая от одного мышечного рубца к другому, называется мантийным
линия (В), место, где находится мантия (ткань, выделяющая оболочку)
прикреплен к оболочке.
То
Блестящая линия, идущая от одного мышечного рубца к другому, называется мантийным
линия (В), место, где находится мантия (ткань, выделяющая оболочку)
прикреплен к оболочке.
- Один из этих моллюсков роет норы, и моллюску нужно место, чтобы вытащить свои сифоны. Это место — паллиальный синус (А)
где было большое углубление для моллюска, чтобы втянуть
сифоны.У моллюсков, которые не зарываются, как и у другого нашего образца, нет паллиального синуса,
а мантийная линия проходит по краю раковины от мышечного рубца
к мышечному рубцу.
III. Классификация: Традиционная классификация двустворчатых моллюсков. находится в еще большем беспокойстве, чем брюхоногие моллюски. Признать несколько групп их общими именами (ниже).
- A. Мидии ( #987 Myalina , без номера Mytilus ): обитают в каменистой литорали, прикреплены к скалам с
нити силы, непревзойденной конструкциями любых других существ,
включая людей.Без сифонов, уменьшенная ножка.

- B. Pectens ( # 1676 Pecten , без номера ): вольные пловцы. Одна крупная мышца закрывает скорлупу (цилиндрический «гребешок», который вы едите. Ищите мышечный шрам.
- C. Oysters ( #1470, #1704, #1444 Exogyra , без номера Gryphea ): некоторые прикреплены, другие покоятся в мягком субстрате. Широкие экологические допуски. Раковины асимметричные, часто неправильной формы.
- D. Раковины ковчега ( # 21 ): длинный прямой шарнир, таксодонтный зубной ряд, тяжелые ребра.Живи привязанным к камням
- E. Венероиды: моллюски Венеры (поедающие моллюсков) ( #1608, 1196 ), моллюски-бритвы ( #496 ), гигантские моллюски ( без номера ) и другие группы. Гетеродонтный зубной ряд. «Моллюск»- форменные норники.
- F. Rudists (Hippuritoida) ( ненумерованный маленький экземпляр, ненумерованный крупный экземпляр ): вымершие моллюски в форме кораллов, с
один клапан в виде «крышки» на конусообразном нижнем клапане.
 Строители рифов.
Строители рифов.
IV.Образ жизни: раковины соответствуют требованиям окружающей среды
- А. роющие двустворчатые моллюски ( № 1236, 25 ): симметричные створки, два одинаковых аддуктора
мышцы, отчетливая мантивная линия
- 1. глубокие норы ( без номера ) — глубокий мантийный синус, тонкий гладкая раковина (недоступная для раздавливающих хищников, или раковины с гребнями параллельно краю раковины для копания
- 2. неглубокие норы ( # 25 , # 1236 Glycymeris ) — мантийный синус маленький или отсутствует, толстый или сильно орнаментированный ракушки
- 3.быстро роющийся в зыбучих отложениях со многими хищниками ( без номера Насута ) — маленькая раковина, сплюснутая оболочка
- 4. медлительный роющий ( без номера Clinocardium ) — выпуклая раковина, толстый орнамент (часто это неглубокие роющие)
- B.
 плавающие двустворчатые моллюски ( пектиновый мешок , симметричная пектиновая сумка, № 256): клапаны
с «крыльями» (ушными раковинами), одиночной большой приводящей мышцей
плавающие двустворчатые моллюски ( пектиновый мешок , симметричная пектиновая сумка, № 256): клапаны
с «крыльями» (ушными раковинами), одиночной большой приводящей мышцей
- плоские поплавки: один клапан более плоский, чем другой
- вертикальные поплавки: створки одинаково закругленные
- мелководье: ребра Глубоководные
- : гладкие
- С.эпифауны двустворчатых моллюсков:
- 1. биссальные (прикреплены нитями) ( #1074 Mytilus ): приводящие мышцы разного размера, зияние биссуса, либо очень заостренный клюв, либо длинный прямой шарнир с «крыльями».
- 2. цементированные ( #590 Ostrea , пектин скальный без номера ): сильно асимметричные створки, шипы,
может иметь один
мышца. Обратите внимание на гребень скалы, который выглядит очень нормально.
pecten, похороненный в сумасшествии новой оболочки.Этот вид
плавает, как молодой гребешок, затем оседает, прикрепляется к скале,
и всю оставшуюся жизнь отращивает раковину, похожую на устрицу (можно также
увидеть, где какой-то скучный организм врезался в скорлупу, но гребень сохранился
раскалывать скорлупу внутри, чтобы избежать нападения хищников).

- 3. мягкий субстрат ( # 589 Gryphea , без номера Exogyra ): сильно асимметричные створки, крышка меньшего размера створка на глубоко вогнутой нижней створке.
В.Доказательства хищников или паразитов на панцирях
- A. Двустворчатые моллюски, которые не зарываются, часто подвергаются нападениям со стороны хищники. Из-за этого эти двустворчатые моллюски обычно строят раковины. более толстые, с тяжелым орнаментом или гофрированием, чтобы противостоять обоим дробление и сверление хищников. У них могут быть шипы для отпугивать хищников, таких как морские звезды или улитки.
- B. Некоторые двустворчатые моллюски становятся жертвами улиток, которые просверливают отверстий в скорлупу, впрыснуть пищеварительные соки, а затем высосать полупереваренный моллюск
- C. Другие двустворчатые моллюски поражаются паразитами , использующими раковину
моллюск К ним относятся клиониевые губки, растворяющие отверстия в
панцирь, а затем двигаться в открытые туннели, и скучные моллюски, которые
просверлить отверстия в оболочке, в которой они живут.

Дисплей: другие моллюски
Хитоны (без номера): Несмотря на название и множество сегментов,
эти организмы являются моллюсками, а не членистоногими. Их скелет
карбонат кальция, как и у других моллюсков, а не хитин, как
членистоногие.Тело организма очень похоже на
морское ушко. Обычно они любят каменистую среду и цепляются за
камни. Этот покрыт инкрустация spnge (красный) и
известковые водоросли (розовые)
Скафоподы ( без номера ) называются бивневыми панцирями. Организм
живет вниз головой
закрепился в осадке. Они очень мало изменились за геологические
время,
и никогда не отличались большим разнообразием. Обратите внимание на толщину
панцирь, особенно в ископаемом образце.Поздно вы можете захотеть
спутать их с некоторыми видами головоногих, но панцирь на
скафаподы обычно намного толще.
Tentaculites : ( # 155 ) этот организм является одним из нескольких малоизвестных
и загадочные ранние существа, которые интерпретируются как моллюски.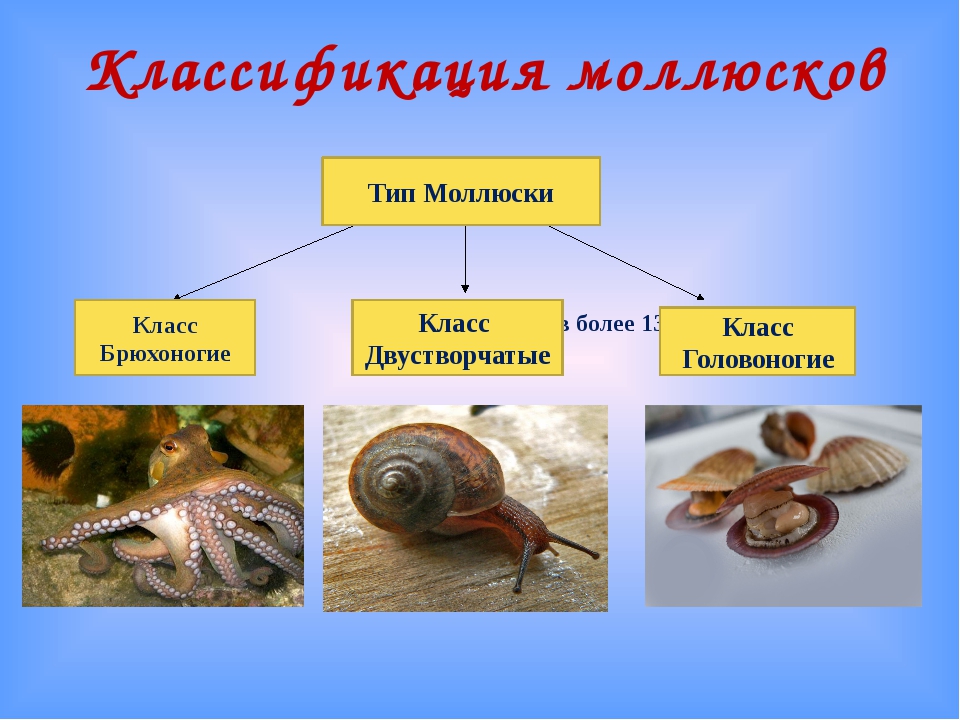 Крошечный конический
оболочка — кальцит. Tentaculites жили в раннем палеозое.
Крошечный конический
оболочка — кальцит. Tentaculites жили в раннем палеозое.
Вопросы
1. #1739 — Этот мезогастропод принадлежит к семейству Natacidae, лунных улиток.Эти улитки — хищники, они ловят двустворчатых моллюсков и просверливают отверстия в оболочки, затем переваривая содержимое. Посмотрите внимательно на натацид и его добычу, и сделайте вывод, является ли эта улитка эпифауной или инфауной.
2. #123 -Красивый перламутровый перламутровый слой внутри отметин ракушки место прикрепления мягких тканей улитки к раковине. Что это было жизненная привычка улитки? Каковы ваши доказательства?
3. #1329 — Как сохраняется этот брюхоногий моллюск?
4.Эта легочная (сухопутная улитка) имеет типично тонкую гладкую раковину. Зачем пульмонировать улиток эволюция таких раковин, в то время как морские улитки неуклонно становились толще, более сильно скульптурные снаряды?
5. Что вы можете сказать об образе жизни каждого из этих брюхоногих моллюсков?
#1667:
#1017:
#2010:
#1307:
#1311:
#1879:
#1242:
#1877:
без номера коричневый и белый:
6. #1633 — обратите внимание на асимметричную форму этого пектина.Как вы это объясните?
#1633 — обратите внимание на асимметричную форму этого пектина.Как вы это объясните?
7. #26 — Обратите внимание на следы на панцире какого-то скучного животного. Какие защита этого двустворчатого моллюска развилась против таких нападений? Каковы ваши доказательства?
8. Нет # — Назовите возможные функции шипов у этого двустворчатого моллюска.
9. Определите вероятный образ жизни (норка глубокая, нора неглубокая,
быстрая нора, медленная нора, пловец, эпифауна на мягком субстрате, эпифауна
зацементированы) каждого двустворчатого моллюска и морфологические признаки
подтверждение вашей гипотезы:
#1472
#1504
#14
#20
#592
без номера
крошечный белый
границ | Связь глубины и разнообразия двустворчатых моллюсков и брюхоногих моллюсков в двух контрастных субарктических морских регионах
Введение
В бентической области сосредоточена значительная часть биоразнообразия океана (Snelgrove, 1999). Морские беспозвоночные, которые обитают на (эпифауне) или в (инфауне) бентических отложениях, составляют значительную часть этого разнообразия и обеспечивают ключевые экосистемные услуги, например, за счет структурной среды обитания и биотурбации (Snelgrove, 1999; Queirós et al., 2013). ). Подавляющее большинство (91%) морской бентической среды обитания находится на глубине ниже 200 м, где преобладает темнота, а автотрофная деятельность незначительна (Kennish, 2000). Доступ к глубокому океану — очень дорогостоящее и сложное с точки зрения логистики мероприятие, объясняющее, почему экосистемы в глубоком море остаются недостаточно изученными по сравнению с мелководными или наземными системами (May and Godfrey, 1994; Higgs and Attrill, 2015).Как никогда важно задокументировать и понять модели биоразнообразия и экологические процессы в глубоководных бентических экосистемах, поскольку эти системы характеризуются экологической стабильностью (Seibel and Walsh, 2003; Hofmann et al., 2011), ограниченной доступностью пищи (Lutz et al.
Морские беспозвоночные, которые обитают на (эпифауне) или в (инфауне) бентических отложениях, составляют значительную часть этого разнообразия и обеспечивают ключевые экосистемные услуги, например, за счет структурной среды обитания и биотурбации (Snelgrove, 1999; Queirós et al., 2013). ). Подавляющее большинство (91%) морской бентической среды обитания находится на глубине ниже 200 м, где преобладает темнота, а автотрофная деятельность незначительна (Kennish, 2000). Доступ к глубокому океану — очень дорогостоящее и сложное с точки зрения логистики мероприятие, объясняющее, почему экосистемы в глубоком море остаются недостаточно изученными по сравнению с мелководными или наземными системами (May and Godfrey, 1994; Higgs and Attrill, 2015).Как никогда важно задокументировать и понять модели биоразнообразия и экологические процессы в глубоководных бентических экосистемах, поскольку эти системы характеризуются экологической стабильностью (Seibel and Walsh, 2003; Hofmann et al., 2011), ограниченной доступностью пищи (Lutz et al. , 2007) и низкие температуры, ограничивающие скорость процессов (Childress, 1995), могут быть особенно чувствительны к естественным и антропогенным изменениям окружающей среды, происходящим с беспрецедентной скоростью (Crain et al., 2008; Gehlen et al., 2014; Роджерс, 2015).
, 2007) и низкие температуры, ограничивающие скорость процессов (Childress, 1995), могут быть особенно чувствительны к естественным и антропогенным изменениям окружающей среды, происходящим с беспрецедентной скоростью (Crain et al., 2008; Gehlen et al., 2014; Роджерс, 2015).
Со времени первых работ Хесслера и Сандерса (1967 г.) многое стало известно об общих закономерностях биоразнообразия бентических видов в глубинах океана и о том, как оно меняется в зависимости от батиметрических градиентов (Рекс и Эттер, 2010 г.). Альфа-разнообразие (α) описывает локальное разнообразие, например, в пределах одного образца или участка. Региональные и связанные с глубиной изменения альфа-разнообразия были в центре внимания большинства ранних исследований, так что в настоящее время пространственные тренды считаются достаточно хорошо изученными для некоторых частей Северной Атлантики (Rex and Etter, 2010).В целом, α-разнообразие донной макрофауны в Северной Атлантике демонстрирует одномодальный характер разнообразия: оно увеличивается по направлению к нижней батиали или верхней абиссальной глубине и уменьшается по направлению к более глубокой бездне (Rex, 1973; Andrew and Scott, 2000). Характер бентического разнообразия к северу от Гренландско-Исландско-Фарерского хребта (ГИФ) в Северных морях и Северном Ледовитом океане заметно отличается от тех, которые обычно описываются для Северной Атлантики. В высокоширотных регионах взаимосвязь между α-разнообразием и глубиной является унимодальной, что обычно наблюдается к югу от хребта GIF, но максимум α-разнообразия приходится на гораздо меньшую глубину (на большей батиальной или меньшей глубине) с заметно низким разнообразием на более низких батиальных и абиссальных глубинах ( Сваварссон, 1997; Бетт, 2001).Однако разнообразие на шельфе не считается особенно низким (Piepenburg et al., 2011).
Характер бентического разнообразия к северу от Гренландско-Исландско-Фарерского хребта (ГИФ) в Северных морях и Северном Ледовитом океане заметно отличается от тех, которые обычно описываются для Северной Атлантики. В высокоширотных регионах взаимосвязь между α-разнообразием и глубиной является унимодальной, что обычно наблюдается к югу от хребта GIF, но максимум α-разнообразия приходится на гораздо меньшую глубину (на большей батиальной или меньшей глубине) с заметно низким разнообразием на более низких батиальных и абиссальных глубинах ( Сваварссон, 1997; Бетт, 2001).Однако разнообразие на шельфе не считается особенно низким (Piepenburg et al., 2011).
Бета (β) разнообразие описывает изменения в составе сообщества в пределах региона или по градиенту (Whittaker, 1960, 1972). Увеличение усилий по отбору проб и доступность данных облегчили исследования β-разнообразия в океане, при этом результаты интерпретируются почти исключительно как пространственное замещение видов по глубине или по горизонтали (McClain and Rex, 2015). Однако на β-разнообразие также может влиять потеря (или увеличение) видов, в результате чего более мелкие сообщества образуют упорядоченные подмножества видового состава более крупных сообществ, что также называют «гнездовостью» (Ulrich and Gotelli, 2007; Almeida-Neto). и другие., 2008). Особая экологическая значимость замены или гнездования видов привела к многочисленным усилиям, направленным на математическую количественную оценку вклада этих компонентов в β-разнообразие (McClain and Rex, 2015). В последние годы в исследованиях глубоководного разнообразия использовался метод, представленный Baselga (2010, 2012), для разделения β-разнообразия на два аддитивных компонента: несходство, возникающее в результате гнездования, и несходство, возникающее в результате оборота видов. Это разделение позволило по-новому взглянуть на модели глубоководного разнообразия (Wagstaff et al., 2014; Стюарт и др., 2017). Например, Браулт и др. (2013b) пришли к выводу, что скопления абиссальных неогастропод, вероятно, поддерживаются динамикой истока и стока, частично основанной на значительном увеличении гнездовья с глубиной наряду с уменьшением α-разнообразия.
Однако на β-разнообразие также может влиять потеря (или увеличение) видов, в результате чего более мелкие сообщества образуют упорядоченные подмножества видового состава более крупных сообществ, что также называют «гнездовостью» (Ulrich and Gotelli, 2007; Almeida-Neto). и другие., 2008). Особая экологическая значимость замены или гнездования видов привела к многочисленным усилиям, направленным на математическую количественную оценку вклада этих компонентов в β-разнообразие (McClain and Rex, 2015). В последние годы в исследованиях глубоководного разнообразия использовался метод, представленный Baselga (2010, 2012), для разделения β-разнообразия на два аддитивных компонента: несходство, возникающее в результате гнездования, и несходство, возникающее в результате оборота видов. Это разделение позволило по-новому взглянуть на модели глубоководного разнообразия (Wagstaff et al., 2014; Стюарт и др., 2017). Например, Браулт и др. (2013b) пришли к выводу, что скопления абиссальных неогастропод, вероятно, поддерживаются динамикой истока и стока, частично основанной на значительном увеличении гнездовья с глубиной наряду с уменьшением α-разнообразия. Хотя этот метод не является широко используемым в исследованиях глубоководного разнообразия, из-за отсутствия сопоставимых стратегий отбора проб, метод регионального сравнения моделей α- и β-разнообразия также может помочь определить процессы, которые определяют модели глубоководного разнообразия (e .г., Браулт и др., 2013а).
Хотя этот метод не является широко используемым в исследованиях глубоководного разнообразия, из-за отсутствия сопоставимых стратегий отбора проб, метод регионального сравнения моделей α- и β-разнообразия также может помочь определить процессы, которые определяют модели глубоководного разнообразия (e .г., Браулт и др., 2013а).
Моллюски представляют собой богатый видами и разнообразный тип, виды которого входят в число наиболее заметных и биоразнообразных макрофауны беспозвоночных в морской среде (Alongi, 1990; Linse et al., 2006). Внутри типа Mollusca брюхоногие моллюски и двустворчатые моллюски являются наиболее богатыми видами классами (Horton et al., 2019). Виды, принадлежащие к обеим группам, могут играть решающую роль в качестве ключевых видов (Paine, 1969) или инженеров экосистемы в результате их механизмов питания или признаков кальцификации (например,г., Холл-Спенсер и Мур, 2000; Гутьеррес и др., 2003). Считается, что кальцифицирующие моллюски особенно подвержены закислению океана и одновременному снижению состояния насыщения карбонатом кальция (Ω) (Gazeau et al. , 2013; Gattuso et al., 2015). В результате повышенного давления Ω уменьшается с глубиной (Millero, 2007), но определение относительной экологической важности Ω и других параметров окружающей среды, связанных с глубиной, таких как наличие пищи и характеристики отложений, представляет собой сложную задачу.Сбор информации о встречаемости видов и биоразнообразии кальцифицирующих моллюсков является важным первым шагом к пониманию последствий изменений Ω, температуры и других крупномасштабных изменений окружающей среды в глубоководных бентических местообитаниях (Urban et al., 2016). Потребность в фоновых данных особенно актуальна для Северных морей из-за естественно низкого значения Ω морской воды и того факта, что Ω уменьшается с угрожающей скоростью как в поверхностных, так и в глубоких водах (Olafsson et al., 2009; Skogen et al. , 2014).
, 2013; Gattuso et al., 2015). В результате повышенного давления Ω уменьшается с глубиной (Millero, 2007), но определение относительной экологической важности Ω и других параметров окружающей среды, связанных с глубиной, таких как наличие пищи и характеристики отложений, представляет собой сложную задачу.Сбор информации о встречаемости видов и биоразнообразии кальцифицирующих моллюсков является важным первым шагом к пониманию последствий изменений Ω, температуры и других крупномасштабных изменений окружающей среды в глубоководных бентических местообитаниях (Urban et al., 2016). Потребность в фоновых данных особенно актуальна для Северных морей из-за естественно низкого значения Ω морской воды и того факта, что Ω уменьшается с угрожающей скоростью как в поверхностных, так и в глубоких водах (Olafsson et al., 2009; Skogen et al. , 2014).
Хребет GIF действует как топографический барьер, разделяющий два океанических бассейна: бассейн Северного моря на севере и бассейн Северной Атлантики на юге. В то время как максимальная глубина порога Гренландско-Исландского хребта и хребта Исландия-Фарерские острова составляет 620 и 480 м соответственно, морское дно достигает глубины более 3000 м к югу и северу от хребта GIF (Jakobsson, 2002). Биогеографические границы на гребне GIF совпадают с переходом между более холодными и более теплыми водными массами, при этом Исландия совпадает с Арктическим фронтом (Hansen, Meincke, 1979).Таким образом, считается, что остров занимает ключевое положение в Северной Атлантике по биогеографическим ареалам видов (Briggs, 1970; Dahlgren et al., 2000; Jöst et al., 2018). Сравнение биоразнообразия ранее проводилось между бентосными местообитаниями непосредственно к северу и югу от хребта GIF для гипербентических амфипод (Weisshappel and Svavarsson, 1998; Weisshappel, 2000), бентических изопод (Svavarsson et al., 1993; Svavarsson, 1997; Brix et al., 2018; Schnurr et al., 2018), остракоды (Jöst et al., 2018) и фораминифер (Gudmundsson, 1998). Однако сравнение видового разнообразия моллюсков между регионами до настоящего времени не проводилось.
В то время как максимальная глубина порога Гренландско-Исландского хребта и хребта Исландия-Фарерские острова составляет 620 и 480 м соответственно, морское дно достигает глубины более 3000 м к югу и северу от хребта GIF (Jakobsson, 2002). Биогеографические границы на гребне GIF совпадают с переходом между более холодными и более теплыми водными массами, при этом Исландия совпадает с Арктическим фронтом (Hansen, Meincke, 1979).Таким образом, считается, что остров занимает ключевое положение в Северной Атлантике по биогеографическим ареалам видов (Briggs, 1970; Dahlgren et al., 2000; Jöst et al., 2018). Сравнение биоразнообразия ранее проводилось между бентосными местообитаниями непосредственно к северу и югу от хребта GIF для гипербентических амфипод (Weisshappel and Svavarsson, 1998; Weisshappel, 2000), бентических изопод (Svavarsson et al., 1993; Svavarsson, 1997; Brix et al., 2018; Schnurr et al., 2018), остракоды (Jöst et al., 2018) и фораминифер (Gudmundsson, 1998). Однако сравнение видового разнообразия моллюсков между регионами до настоящего времени не проводилось.
Основная цель этого исследования заключалась в том, чтобы лучше понять модели биоразнообразия в высоких широтах Северной Атлантики путем изучения моделей α- и β-разнообразия моллюсков вдоль батиметрического градиента к северу и югу от хребта GIF. Второстепенной целью этого исследования было выявление видов двустворчатых моллюсков и брюхоногих моллюсков, которые можно было бы использовать в качестве видов-индикаторов для мониторинга экологической целостности к северу от хребта МФП (Carignan and Villard, 2002).Будущие усилия по мониторингу предпочтительно должны включать различные таксоны, но поскольку двустворчатые моллюски и брюхоногие моллюски, как известно, чувствительны к Ω, они могут обеспечить раннее предупреждение о естественных ответных реакциях в районах, подвергающихся быстрому закислению океана, как это наблюдается к северу от хребта GIF (Gattuso et al. др., 2015). Для достижения этих целей мы используем данные о двустворчатых моллюсках и брюхоногих моллюсках, собранные в ходе программы отбора проб BIOICE (Benthic Invertebrates of Icelandic waters).
Материалы и методы
Районы исследования
Районы исследования охватывают исландский шельф и склон в пределах Исландского моря к северу от хребта GIF, а также исландский шельф и склон к югу от хребта GIF (рис. 1).Эти регионы имеют топографическое сходство, поскольку они оба включают исландский шельф и склон, а также части Срединно-Атлантического хребта, но широкое разделение хребтом GIF приводит к региональным гидрографическим различиям (Stefánsson, 1962). Исландское море является частью северных морей наряду с Норвежским и Гренландским морями (Jakobsson, 2002). Гидрографические свойства Исландского моря обычно описываются как арктические промежуточные воды, лежащие над арктическими глубокими водами (Stefánsson, 1962; Swift et al., 1980). Для этих вод характерны отрицательные температуры по всей толще воды зимой и неглубокий (∼200 м) смешанный слой летом, в котором температуры превышают 0°С (Olafsson, 2003; Olafsson et al., 2009). Океанографическая среда к югу от хребта отражает динамическое взаимодействие водных масс с различными физическими свойствами, с температурой от 2 до >10°C (Hansen, Østerhus, 2000; Malmberg, Valdimarsson, 2003). Потепление и закисление океана наблюдались на поверхности как в Исландии, так и в море Ирмингера (к юго-западу от Исландии), при этом потепление и закисление наблюдались на глубине не менее 1800 м в Исландском море за последние два десятилетия (Olafsson et al., 2009, 2010).
Потепление и закисление океана наблюдались на поверхности как в Исландии, так и в море Ирмингера (к юго-западу от Исландии), при этом потепление и закисление наблюдались на глубине не менее 1800 м в Исландском море за последние два десятилетия (Olafsson et al., 2009, 2010).
Рисунок 1. Районы исследования к северу и югу от хребта Гренландия-Исландия-Фареры и географическое положение образцов BIOICE, включенных в исследование. Красные треугольники и синие кружки — места проб бентоса, отобранных при буксировке модифицированных саней RP и саней Sneli соответственно. Горизонтали нанесены с интервалом в 1000 м по глубине.
Биологические данные
пробы бентоса были собраны на глубине от 20 до 3020 м в период с 1991 по 2004 год в рамках программы BIOICE (Gudmundsson, 1998).В программе использовались четыре типа буксируемых орудий для отбора проб, в зависимости от типа дна, но это исследование включает только данные, собранные с использованием модифицированных саней RP (Rothlisberg and Pearcy, 1977; Brattegard and Fosså, 1991) и саней Sneli (Sneli, 1998). ), поскольку эти снасти чаще всего использовались и собирали образцы скоплений сходных видов (дополнительный материал 1). Записи о видах и их таксономическая классификация были проверены вручную и при необходимости отредактированы, чтобы соответствовать записям в базе данных «Всемирный регистр морских видов» и синонимам, стандартизированным до единого названия.Предоставляется информация о данных образцов, рассмотренных для исследования, список всех зарегистрированных видов, количество образцов, в которых они были зарегистрированы, и их наблюдаемый диапазон глубин в каждом районе исследования (Дополнительный материал 2).
), поскольку эти снасти чаще всего использовались и собирали образцы скоплений сходных видов (дополнительный материал 1). Записи о видах и их таксономическая классификация были проверены вручную и при необходимости отредактированы, чтобы соответствовать записям в базе данных «Всемирный регистр морских видов» и синонимам, стандартизированным до единого названия.Предоставляется информация о данных образцов, рассмотренных для исследования, список всех зарегистрированных видов, количество образцов, в которых они были зарегистрированы, и их наблюдаемый диапазон глубин в каждом районе исследования (Дополнительный материал 2).
Аналитический подход
Данные проб, собранные с использованием модифицированных салазок RP и салазок Sneli, были объединены для анализа биоразнообразия, поскольку наборы видов, отобранные для разных типов снастей, были схожими. Общее количество проб, отобранных в пределах исследуемых районов, составило 366 и 383 пробы к северу и югу от хребта ГИФ соответственно. Образцы, собранные с помощью салазок RP и салазок Снели, внесли почти равный вклад в общее количество образцов в обоих регионах.
Образцы, собранные с помощью салазок RP и салазок Снели, внесли почти равный вклад в общее количество образцов в обоих регионах.
Количество ненаблюдаемых видов, в дополнение к наблюдаемому видовому богатству, оценивалось на основе одиночных и двойных видов в матрицах видов (Chao, 1987) вместе со стандартной ошибкой (Chiu et al., 2014). Оценки проводились для каждого района и отдельно для четырех 500-метровых диапазонов глубин, между поверхностью и глубиной 2000 м, ниже которых данные проб считались недостаточными для оценки видового богатства.
Альфа-разнообразие оценивалось с использованием ожидаемого числа видов Сандерса-Херлберта (Sanders, 1968; Hurlbert, 1971) в выборке, нормализованной к 20 особям [ E ( S 20 )], исключая из анализа образцы, где было собрано менее 20 особей брюхоногих или двустворчатых моллюсков. Соответственно, количество образцов, доступных для анализа, было сокращено до 252 и 250 для двустворчатых моллюсков к северу и югу от хребта GIF соответственно, и до 146 и 197 образцов для брюхоногих моллюсков в тех же регионах соответственно. Чтобы исследовать, как E ( S 20 ) менялся с глубиной в каждом регионе, образцы были усреднены по 100-метровым бинам, и для определения тенденций в E ( S 20 ) использовалась линейная регрессия наименьших квадратов. в зависимости от глубины в пределах батиальной зоны, или от 300 до 2000 м.
Чтобы исследовать, как E ( S 20 ) менялся с глубиной в каждом регионе, образцы были усреднены по 100-метровым бинам, и для определения тенденций в E ( S 20 ) использовалась линейная регрессия наименьших квадратов. в зависимости от глубины в пределах батиальной зоны, или от 300 до 2000 м.
Чтобы изучить взаимосвязь глубины и региона с видовыми сообществами двустворчатых моллюсков и брюхоногих моллюсков, был проведен двумерный анализ NMDS по матрицам присутствия-отсутствия (бинарных) видов, исключая виды, встречающиеся менее чем в 5% образцов в каждом регионе.Анализ NMDS был основан на различии образцов Серенсена (Брея-Кертиса) (Sørensen, 1948). Вклад глубины и области в MDS1 (первичная ось NMDS) и MDS2 (вторичная ось NMDS) анализировали с использованием линейной регрессии наименьших квадратов и критерия Стьюдента t соответственно.
Оптимальные методы анализа β-разнообразия вдоль градиентов окружающей среды требуют данных, собранных с использованием стандартизированных методов выборки, но, как и большинство исследований моделей глубоководного разнообразия макро- и/или мейофауны (Rex and Etter, 2010), это исследование опирается на методы, которые не стандартизированы. Салазки для отбора проб в BIOICE обычно буксировались в течение 20 минут, но среднее время буксировки составляло 19 ± 6 минут (дополнительный материал 2), а буксируемый нижний субстрат был неизвестен. Мы признаем проблемы, связанные с отсутствием стандартизированного отбора проб, а также проблемы, связанные с неслучайным и бессистемным пространственным распределением мест отбора проб в программе BIOICE, особенно с ограниченными усилиями по отбору проб на глубинах ниже 1500 м. Чтобы решить эти проблемы, данные выборки были объединены в ячейки глубиной 100 м, при этом ячейки, содержащие менее 100 и 50 отдельных двустворчатых моллюсков и брюхоногих моллюсков, соответственно, были исключены из анализа β-разнообразия.Данные были преобразованы в матрицы присутствия-отсутствия (бинарные) видов. Анализ был ограничен верхним пределом глубины 300 м, чтобы исключить мелководные сообщества на шельфе Исландии при анализе β-разнообразия. В результате анализ был применен к диапазону глубин от 300 до 1600 м, с 14 интервалами по глубине для двустворчатых моллюсков и 13 интервалами по глубине для брюхоногих моллюсков из-за исключения интервала по глубине 1200 м.
Салазки для отбора проб в BIOICE обычно буксировались в течение 20 минут, но среднее время буксировки составляло 19 ± 6 минут (дополнительный материал 2), а буксируемый нижний субстрат был неизвестен. Мы признаем проблемы, связанные с отсутствием стандартизированного отбора проб, а также проблемы, связанные с неслучайным и бессистемным пространственным распределением мест отбора проб в программе BIOICE, особенно с ограниченными усилиями по отбору проб на глубинах ниже 1500 м. Чтобы решить эти проблемы, данные выборки были объединены в ячейки глубиной 100 м, при этом ячейки, содержащие менее 100 и 50 отдельных двустворчатых моллюсков и брюхоногих моллюсков, соответственно, были исключены из анализа β-разнообразия.Данные были преобразованы в матрицы присутствия-отсутствия (бинарные) видов. Анализ был ограничен верхним пределом глубины 300 м, чтобы исключить мелководные сообщества на шельфе Исландии при анализе β-разнообразия. В результате анализ был применен к диапазону глубин от 300 до 1600 м, с 14 интервалами по глубине для двустворчатых моллюсков и 13 интервалами по глубине для брюхоногих моллюсков из-за исключения интервала по глубине 1200 м. Для исследования динамики β-разнообразия, включая вклад вложенности в β-разнообразие, мы используем два независимых метода.
Для исследования динамики β-разнообразия, включая вклад вложенности в β-разнообразие, мы используем два независимых метода.
Baselga (2010, 2012) описал, как β-разнообразие можно математически разделить на две составляющие: (A) β-разнообразие, возникающее в результате замены видов между участками, что обычно называют оборотом видов, и (B) часть несходства, которая можно отнести к эффекту потери (или приобретения) вида или вложенности видов. Расчеты β-разнообразия и относительного вклада оборота и гнездовости основаны на несходстве Серенсена (Брея-Кертиса) (Sørensen, 1948).Были сгенерированы множественные показатели различия мест (обозначены заглавными буквами), которые дали единый балл для β-разнообразия (β SØR ) и относительного вклада оборота видов (β SIM ) и гнездовости (β SNE ) в несходство в каждом регионе. Сравнение нескольких показателей несхожести сайтов можно проводить только между регионами с одинаковыми размерами выборки (Baselga, 2012). Однако повторная выборка оценок несходства для подмножества исходной базы данных дает средние показатели со стандартными ошибками, которые можно сравнивать между регионами с разными размерами выборки (Baselga, 2012).Хотя бины глубины были стандартизированы между регионами, мы сообщаем множественные оценки различий сайтов, основанные на 100 повторениях выборки, чтобы повысить сопоставимость результатов. Путем попарного сравнения образцов (обозначенных строчными буквами) мы также оценили, изменялись ли показатели β-разнообразия, обозначаемые как β sør , β sim, и β sne , в зависимости от разделения по глубине образца. Взаимосвязь между показателями парного несходства и разделением выборки по глубине была исследована с помощью линейной регрессии наименьших квадратов, но, признавая, что парные сравнения завышают степени свободы, регрессия считалась значимой только тогда, когда результирующее значение p для наклона было равно, или ниже 0.001.
Однако повторная выборка оценок несходства для подмножества исходной базы данных дает средние показатели со стандартными ошибками, которые можно сравнивать между регионами с разными размерами выборки (Baselga, 2012).Хотя бины глубины были стандартизированы между регионами, мы сообщаем множественные оценки различий сайтов, основанные на 100 повторениях выборки, чтобы повысить сопоставимость результатов. Путем попарного сравнения образцов (обозначенных строчными буквами) мы также оценили, изменялись ли показатели β-разнообразия, обозначаемые как β sør , β sim, и β sne , в зависимости от разделения по глубине образца. Взаимосвязь между показателями парного несходства и разделением выборки по глубине была исследована с помощью линейной регрессии наименьших квадратов, но, признавая, что парные сравнения завышают степени свободы, регрессия считалась значимой только тогда, когда результирующее значение p для наклона было равно, или ниже 0.001.
Направление вложенности вдоль батиметрического градиента исследовано с помощью программы BINMATNEST (Rodríguez-Girones and Santamaría, 2006, 2010).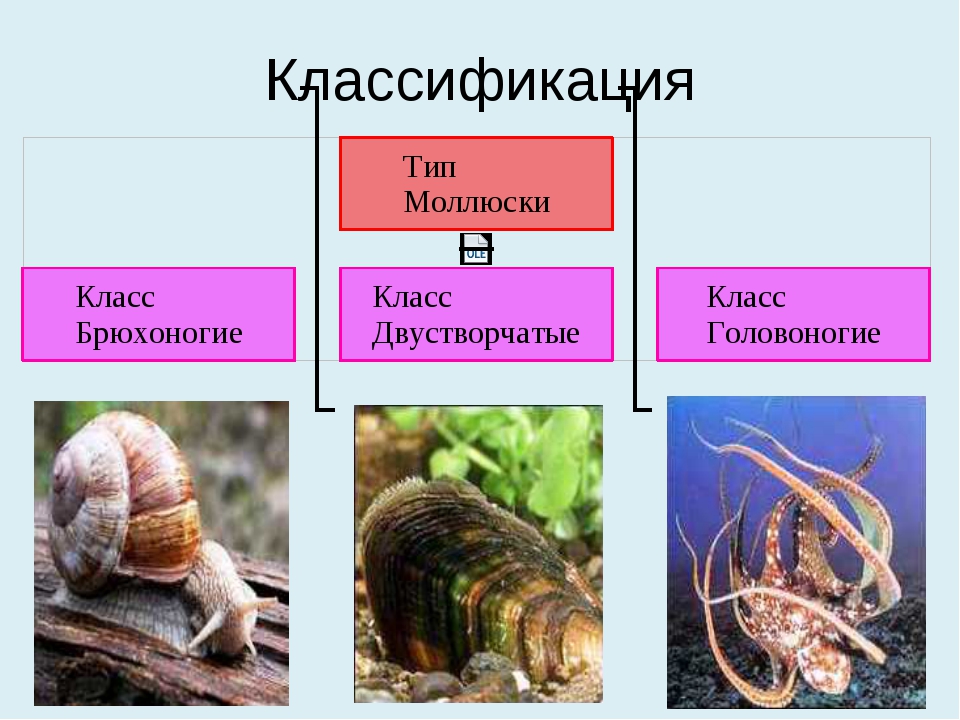 «Температура» матрицы присутствия-отсутствия видов, которая определяется как сумма «сюрпризов» в упорядоченной матрице (в диапазоне от 0 до 100°), была определена, отражая полностью вложенную матрицу во вложенный шаблон соответственно. Анализ также ранжирует бины глубины на основе вложенности, которую можно использовать для информирования о направлении вложенности вдоль градиента окружающей среды (Stuart et al., 2017). Чтобы проверить наличие существенной вложенности в матрицах видов и ранжировать бины по глубине на основе вложенности, было выполнено вычисление 1000 нулевых матриц, и матрицы видов были протестированы на случайность с использованием рекомендуемого выбора нулевой модели в BINMATNEST (модель 3) ( Родригес-Хиронес и Сантамария, 2006 г.).
«Температура» матрицы присутствия-отсутствия видов, которая определяется как сумма «сюрпризов» в упорядоченной матрице (в диапазоне от 0 до 100°), была определена, отражая полностью вложенную матрицу во вложенный шаблон соответственно. Анализ также ранжирует бины глубины на основе вложенности, которую можно использовать для информирования о направлении вложенности вдоль градиента окружающей среды (Stuart et al., 2017). Чтобы проверить наличие существенной вложенности в матрицах видов и ранжировать бины по глубине на основе вложенности, было выполнено вычисление 1000 нулевых матриц, и матрицы видов были протестированы на случайность с использованием рекомендуемого выбора нулевой модели в BINMATNEST (модель 3) ( Родригес-Хиронес и Сантамария, 2006 г.).
Выбор вида индикатора
Было идентифицировано вида двустворчатых моллюсков и брюхоногих моллюсков, которые можно считать подходящими для мониторинга с целью выявления ранних изменений в бентических сообществах, вызванных изменениями окружающей среды к северу от хребта GIF, с особым вниманием к закислению океана и уменьшению Ω. С целью исключения редких или вездесущих видов виды были перечислены на основе их частоты отбора проб и численности в пробах саней в наборе данных BIOICE, который включал 366 проб к северу от хребта GIF. В программе BIOICE виды учитывались в пробах, состоящих не менее чем из 100 особей, поэтому, хотя набор данных не включает полные измерения численности очень распространенных видов, можно определить виды двустворчатых моллюсков и брюхоногих моллюсков как многочисленные или нет на основе заранее определенный критерий.Виды были включены в список, если они соответствовали следующим критериям: (A) обнаружены в 40 или более образцах; (B) не менее 25% выборок включали пять или более человек; (C) Обнаружен в 25% или более проб, собранных в пределах наблюдаемого диапазона глубин; (D) не наблюдается на глубине более 1500 м в наборе данных BIOICE.
С целью исключения редких или вездесущих видов виды были перечислены на основе их частоты отбора проб и численности в пробах саней в наборе данных BIOICE, который включал 366 проб к северу от хребта GIF. В программе BIOICE виды учитывались в пробах, состоящих не менее чем из 100 особей, поэтому, хотя набор данных не включает полные измерения численности очень распространенных видов, можно определить виды двустворчатых моллюсков и брюхоногих моллюсков как многочисленные или нет на основе заранее определенный критерий.Виды были включены в список, если они соответствовали следующим критериям: (A) обнаружены в 40 или более образцах; (B) не менее 25% выборок включали пять или более человек; (C) Обнаружен в 25% или более проб, собранных в пределах наблюдаемого диапазона глубин; (D) не наблюдается на глубине более 1500 м в наборе данных BIOICE.
За исключением программного обеспечения BINMATNEST, весь анализ данных был выполнен с использованием R (R-Core-Team, 2018), а также пакетов vegan (Oksanen et al. , 2015) и betapart для R (Baselga и Orme). , 2012).
, 2015) и betapart для R (Baselga и Orme). , 2012).
Результаты
Видовое богатство
Двустворчатые моллюски, как правило, были более многочисленны в образцах, чем брюхоногие моллюски, но брюхоногие моллюски были примерно в два раза богаче видами двустворчатых моллюсков в обоих исследуемых регионах. Общее наблюдаемое видовое богатство двустворчатых моллюсков составляло 85 видов к северу от хребта GIF и 114 видов к югу от хребта GIF, при этом 71 вид был общим для обоих регионов. На основе анализа необнаруженного видового богатства оценочное видовое богатство двустворчатых моллюсков (± стандартная ошибка) составило 95 ± 8 и 137 ± 15 к северу и югу от хребта GIF соответственно.Видовое богатство двустворчатых моллюсков оценивается в 7 ± 1 в диапазоне глубин 1501–2000 м к северу от хребта ГИП по сравнению с 34 ± 6 к югу от хребта ГИП (рис. 2). Наблюдаемое видовое богатство брюхоногих моллюсков составляло 173 и 189 видов к северу и югу от хребта GIF соответственно, при этом 133 вида были разделены между регионами. Расчетное общее видовое богатство брюхоногих моллюсков (± стандартная ошибка) составило 201 ± 12 и 266 ± 33 к северу и югу от хребта GIF. Видовое богатство брюхоногих моллюсков, оцененное в диапазоне глубин 1501–2000 м, составило 19 ± 11 и 73 ± 9 севернее и южнее хребта ГФ соответственно (рис. 2).
Расчетное общее видовое богатство брюхоногих моллюсков (± стандартная ошибка) составило 201 ± 12 и 266 ± 33 к северу и югу от хребта GIF. Видовое богатство брюхоногих моллюсков, оцененное в диапазоне глубин 1501–2000 м, составило 19 ± 11 и 73 ± 9 севернее и южнее хребта ГФ соответственно (рис. 2).
Рис. 2. Расчетное видовое богатство в интервалах глубин 500 м на основе (Chao, 1987). Столбики погрешностей представляют собой стандартные ошибки оценок, основанных на Chiu et al. (2014) для (A) двустворчатых моллюсков к северу от хребта GIF и (B) двустворчатых моллюсков к югу от хребта GIF, (C) брюхоногих к северу от хребта GIF и (D) брюхоногих к югу от гребня GIF хребет.
Альфа-разнообразие
Анализ E ( S 20 ) для двустворчатых моллюсков и брюхоногих моллюсков к северу от хребта GIF указывает на унимодальную связь с глубиной, достигающую максимум ~ 5 видов между 100 и 600 м (рис. 3).Модель линейной регрессии E ( S 20 ), установленная в диапазоне глубин от 300 до 2000 м к северу от хребта GIF, показывает резкое снижение α-разнообразия как двустворчатых моллюсков, так и брюхоногих моллюсков вдоль батиметрического градиента с высоким R Значения 2 найдены для двустворчатых ( R 2 = 0,92) и брюхоногих моллюсков ( R 2 = 0,77) соответственно (табл. 1). К югу от хребта GIF аналогичная одномодальная связь между α-разнообразием и глубиной наблюдалась у двустворчатых моллюсков с небольшим уменьшением глубины от 300 до 2000 м.В отличие от севера, значения R 2 у двустворчатых моллюсков на юге значительно ниже ( R 2 = 0,22), а у брюхоногих моллюсков E ( S 20 90 20 ) четкой тенденции не выявлено. к югу от хребта GIF (рис. 3).
3).Модель линейной регрессии E ( S 20 ), установленная в диапазоне глубин от 300 до 2000 м к северу от хребта GIF, показывает резкое снижение α-разнообразия как двустворчатых моллюсков, так и брюхоногих моллюсков вдоль батиметрического градиента с высоким R Значения 2 найдены для двустворчатых ( R 2 = 0,92) и брюхоногих моллюсков ( R 2 = 0,77) соответственно (табл. 1). К югу от хребта GIF аналогичная одномодальная связь между α-разнообразием и глубиной наблюдалась у двустворчатых моллюсков с небольшим уменьшением глубины от 300 до 2000 м.В отличие от севера, значения R 2 у двустворчатых моллюсков на юге значительно ниже ( R 2 = 0,22), а у брюхоногих моллюсков E ( S 20 90 20 ) четкой тенденции не выявлено. к югу от хребта GIF (рис. 3).
Рисунок 3. Hurlbert-Sanders ожидаемое количество видов в выборке из 20 особей, E ( S 20 ), для (A) двустворчатых моллюсков к северу от хребта GIF, (B) 9002 двустворчатых моллюсков к югу от хребта ГИП, (В) брюхоногих к северу от хребта ГИП и (Г) брюхоногих к югу от хребта ГИП. Разреженные пробы усреднялись по 100-метровым бинам с планками погрешностей, представляющими стандартные отклонения. Там, где для разрежения был доступен только один образец, полоса ошибок не отображается. Черные линии и заштрихованная область соответствуют моделям линейной регрессии и стандартной ошибке соответственно.
Разреженные пробы усреднялись по 100-метровым бинам с планками погрешностей, представляющими стандартные отклонения. Там, где для разрежения был доступен только один образец, полоса ошибок не отображается. Черные линии и заштрихованная область соответствуют моделям линейной регрессии и стандартной ошибке соответственно.
Таблица 1. Результаты моделей линейной регрессии методом наименьших квадратов E ( S 20 ) в зависимости от глубины от 300 до 2000 м.
Сообщества видов
Глубина влияла на состав сообщества сильнее, чем регион, как у двустворчатых моллюсков, так и у брюхоногих моллюсков, но взаимосвязь между этими факторами и видовым составом была сильнее для двустворчатых моллюсков по сравнению с брюхоногими (рис. 4).Оценка MDS1 для сообществ двустворчатых моллюсков сильно коррелировала с логарифмом глубины (корреляция Пирсона продукт-момент: r = 0,84, t = 33, df = 469, p <0,001), а оценки MDS2 различались. между исследуемыми регионами ( t -тест: t = -26, df = 458, p < 0,001). Показатель MDS1 для сообществ брюхоногих моллюсков также значительно коррелировал с логарифмом глубины, хотя и не так сильно, как у двустворчатых моллюсков (корреляция Пирсона продукт-момент: r = 0.50, t = -12, df = 406, p < 0,001) и MDS2 также различались по регионам ( t -test: t = 11, df < 0,001).
между исследуемыми регионами ( t -тест: t = -26, df = 458, p < 0,001). Показатель MDS1 для сообществ брюхоногих моллюсков также значительно коррелировал с логарифмом глубины, хотя и не так сильно, как у двустворчатых моллюсков (корреляция Пирсона продукт-момент: r = 0.50, t = -12, df = 406, p < 0,001) и MDS2 также различались по регионам ( t -test: t = 11, df < 0,001).
Рисунок 4. Графики двумерного неметрического многомерного масштабирования (NMDS) для (A) сообществ двустворчатых моллюсков и (B) брюхоногих моллюсков. Для каждого класса на центральном графике показаны результаты анализа с выборочными данными, представляющими бассейн Исландского моря к северу от хребта GIF (синие квадраты) и регион к югу от хребта GIF (красные кружки).Размер точки относится к логарифму глубины выборки. Сверху показана корреляция оценок MDS1 и логарифма глубины, которая была значимой для двустворчатых моллюсков и брюхоногих моллюсков, а справа на диаграмме показаны значительные региональные различия для оценок MDS2 в обеих группах.
Бета-разнообразие
Анализ динамики β-разнообразия в сообществах двустворчатых моллюсков и брюхоногих моллюсков на высоте 300–1600 м показывает, что, хотя общее β-разнообразие сопоставимо между исследуемыми районами, существуют различия в относительном вкладе гнездования и оборота в β-разнообразие.
У двустворчатых моллюсков β-разнообразие было выше к северу от хребта GIF по сравнению с таковым к югу от хребта GIF р < 0,001; рис. 5). Множественные метрики сайта (± стандартное отклонение) для региона к северу от хребта GIF указывают на аналогичный вклад вложенности (β SNE ) и оборота (β SIM ) в общее β-разнообразие (β SØR ) в область с β SØR = 0.76 ± 0,02, β SIM = 0,39 ± 0,02 и β SNE = 0,37 ± 0,03. Гнездовость в меньшей степени способствовала региональному различию к югу от хребта GIF, где β SØR = 0,74 ± 0,01, β SIM = 0,58 ± 0,03 и β SNE = 0,17 ± 0,03. Попарные оценки предполагают, что различия видовых сообществ увеличиваются с увеличением глубины разделения в обоих регионах, т. Е. β sør положительно связан с глубиной разделения (таблица 2). Увеличение β sør отражает более высокие баллы β sne с увеличением глубины разделения к северу от хребта GIF, тогда как оборот видов в большей степени способствует изменению видовых сообществ к югу от хребта GIF (рис. 5).
Попарные оценки предполагают, что различия видовых сообществ увеличиваются с увеличением глубины разделения в обоих регионах, т. Е. β sør положительно связан с глубиной разделения (таблица 2). Увеличение β sør отражает более высокие баллы β sne с увеличением глубины разделения к северу от хребта GIF, тогда как оборот видов в большей степени способствует изменению видовых сообществ к югу от хребта GIF (рис. 5).
Рис. 5. Попарное несходство комплексов двустворчатых моллюсков (слева) и брюхоногих моллюсков (справа) в зависимости от разделения глубин между глубинными ячейками в Исландском море к северу от хребта GIF (синий квадрат) и к югу от хребта GIF (красные точки) ). Попарное общее β-разнообразие, или β sør , делится на β sim или несходство в результате замены видов и β sne или несходство в результате вложенности или потери (или увеличения) видов.Линии регрессии показывают, где линейная регрессия наименьших квадратов привела к p < 0,001 для наклона.
Таблица 2. Результаты моделей линейной регрессии наименьших квадратов Baselga (2010, 2012) попарных метрик β-разнообразия как функции разделения по глубине между 300 и 1600 м.
У брюхоногих β-разнообразие было выше к северу от хребта GIF по сравнению с таковым к югу от хребта GIF ( t -критерий парных оценок для β sør : t = 47.8, df = 163, p < 0,001; Рисунок 5). Метрики нескольких участков (± стандартное отклонение) для региона к северу от хребта GIF, β SØR = 0,81 ± 0,01, β SIM = 0,60 ± 0,02 и β SNE = 0,21 ± 0,02, предполагают меньший вклад гнездовья к β-разнообразию у брюхоногих моллюсков по сравнению с двустворчатыми моллюсками. Множественные показатели β-разнообразия брюхоногих моллюсков к югу от хребта GIF: β SØR = 0,75 ± 0,01, β SIM = 0,64 ± 0,02 и β SNE = 0.11 ± 0,02. Результаты попарных сравнений для брюхоногих говорят об увеличении β-разнообразия (β sør ) с увеличением глубины разделения в обоих регионах (табл. 2). Увеличение β sne с увеличением глубины разделения наблюдалось к северу от гребня GIF у брюхоногих моллюсков, аналогично тенденции, наблюдаемой у двустворчатых моллюсков (рис. 5). Регрессионные модели для брюхоногих моллюсков не выявили тенденции в β sim с увеличением глубины разделения для любого региона.
2). Увеличение β sne с увеличением глубины разделения наблюдалось к северу от гребня GIF у брюхоногих моллюсков, аналогично тенденции, наблюдаемой у двустворчатых моллюсков (рис. 5). Регрессионные модели для брюхоногих моллюсков не выявили тенденции в β sim с увеличением глубины разделения для любого региона.
Тест БИНМАТНЕСТ отклонил нулевую гипотезу об отсутствии гнездования в матрицах видов двустворчатых моллюсков и брюхоногих моллюсков в обоих регионах.Корреляция рангового порядка вложенности для каждого интервала глубины также указывает на значительное линейное увеличение вложенности с увеличением глубины (рис. 6). Связь была относительно сильной к северу от хребта GIF, но относительно слабой к югу от хребта GIF как у двустворчатых моллюсков, так и у брюхоногих моллюсков.
Рис. 6. Ранговый порядок гнездования, рассчитанный с помощью программы BINMATNEST для (A) двустворчатых моллюсков и (B) брюхоногих моллюсков, север (синие квадраты и линия) и юг (красные точки и линия) хребта GIF .
Идентификация видов-индикаторов
Десять видов двустворчатых моллюсков и пять видов брюхоногих моллюсков соответствовали критериям мониторинга экологической целостности донной экосистемы к северу от хребта МФП (табл. 3).
Таблица 3. Виды двустворчатых моллюсков и брюхоногих моллюсков, пригодные для мониторинга экологической целостности донной экосистемы к северу от хребта МФП.
Обсуждение
Видовое богатство исландского шельфа к северу и югу от хребта GIF было одинаковым.Этот вывод согласуется с выводами Piepenburg et al. (2011), которые показали, что видовое богатство бентоса на шельфе (выше ∼200 м) в арктических морях не особенно низкое по сравнению с видовым богатством в более низких широтах, как предполагалось ранее на основе ограниченных выборочных данных.
Альфа-разнообразие
Мы наблюдали заметное снижение α-разнообразия с увеличением глубины между 300 и 2000 м как у двустворчатых моллюсков, так и у брюхоногих моллюсков к северу от хребта GIF. Примечательно, что оценки видового богатства указывают на уменьшение на порядок величины от диапазона глубин 501–1000 м до диапазона глубин 1501–2000 м у двустворчатых моллюсков. Эти результаты согласуются с ранее описанным уменьшением α-разнообразия с увеличением глубины у других бентосных таксонов в Северных морях, включая изопод (Svavarsson, 1997), амфипод (Stephensen, 1940; Weisshappel and Svavarsson, 1998), полихет (Narayanaswamy et al., 2005; Oug et al., 2017), фораминифер (Mackensen et al., 1985; Gudmundsson, 2002) и макрофауну в целом (Bett, 2001).На самом деле исследования бентического разнообразия в глубоководном бассейне Северного моря последовательно описывают низкое видовое богатство по сравнению с глубоководными зонами других морей и океанических бассейнов, включая северную часть Атлантического океана к югу от хребта GIF (Bouchet and Warén, 1979; Dahl, 1979; Рекс и др., 2000). Причина низкого разнообразия в глубоководном бассейне Северного моря, вероятно, связана с низкими температурами и ограниченной доступностью энергии, как обсуждается далее в следующей главе о бета-разнообразии.
Примечательно, что оценки видового богатства указывают на уменьшение на порядок величины от диапазона глубин 501–1000 м до диапазона глубин 1501–2000 м у двустворчатых моллюсков. Эти результаты согласуются с ранее описанным уменьшением α-разнообразия с увеличением глубины у других бентосных таксонов в Северных морях, включая изопод (Svavarsson, 1997), амфипод (Stephensen, 1940; Weisshappel and Svavarsson, 1998), полихет (Narayanaswamy et al., 2005; Oug et al., 2017), фораминифер (Mackensen et al., 1985; Gudmundsson, 2002) и макрофауну в целом (Bett, 2001).На самом деле исследования бентического разнообразия в глубоководном бассейне Северного моря последовательно описывают низкое видовое богатство по сравнению с глубоководными зонами других морей и океанических бассейнов, включая северную часть Атлантического океана к югу от хребта GIF (Bouchet and Warén, 1979; Dahl, 1979; Рекс и др., 2000). Причина низкого разнообразия в глубоководном бассейне Северного моря, вероятно, связана с низкими температурами и ограниченной доступностью энергии, как обсуждается далее в следующей главе о бета-разнообразии. Представленные здесь тренды α-разнообразия также аналогичны описанным для Северного Ледовитого океана в целом (Bodil et al., 2011).
Представленные здесь тренды α-разнообразия также аналогичны описанным для Северного Ледовитого океана в целом (Bodil et al., 2011).
Паттерны α-разнообразия, наблюдаемые к югу от хребта GIF, т. е. уменьшение E ( S 20 ) у двустворчатых моллюсков вдоль батиметрического градиента и отсутствие тренда E ( S 20 ) у брюхоногих, контрастирует с ранее описанными моделями разнообразия непосредственно к югу от хребта GIF и в более широкой Северной Атлантике (Rex and Etter, 2010). Например, Svavarsson (1997) сообщает о значительном увеличении количества изопод E ( S 200 ) непосредственно к югу от хребта GIF с 200 до 1500 м, но известно, что тенденции разнообразия изопод отклоняются от общих тенденций в другой макрофауне. поскольку отряд включает в себя множество видов, специально адаптированных к глубоководным условиям (Hessler et al., 1979; Рекс и Эттер, 2010). В более широкой Северной Атлантике Bett (2001) и Narayanaswamy et al. (2005) также сообщили об увеличении общей макрофауны и полихет E ( S n ) на глубине от 200 до 2000 м в желобе Роколл, к югу от хребта Уивилл-Томсон. Кроме того, Браулт и соавт. (2013a) описал увеличение E ( S 20 ) от шельфа к глубинам более 2000 м у двустворчатых моллюсков в западной и восточной части Северной Атлантики.Объяснение наблюдаемого отклонения от общего тренда непосредственно к югу от хребта GIF неизвестно и требует исчерпывающих данных об экологических условиях региона. Вероятным объяснением является то, что океанографические условия непосредственно к югу от хребта GIF могут повлиять на модели биоразнообразия (Gudmundsson, 1998), в частности, на относительно быстрое течение ниже глубины ∼1500 м, несущее Североатлантические глубоководные воды (NADW), смешанные с прохладными поверхностными водами. от северных морей на запад вдоль континентального склона Исландии (Dickson et al., 1990; Логеманн и др., 2013). Текущие режимы могут также воздействовать на свойства отложений за счет ресуспендирования мелких отложений, при этом размер частиц отложений признан важным фактором разнообразия бентоса (Etter and Grassle, 1992; Gray, 2002).
(2005) также сообщили об увеличении общей макрофауны и полихет E ( S n ) на глубине от 200 до 2000 м в желобе Роколл, к югу от хребта Уивилл-Томсон. Кроме того, Браулт и соавт. (2013a) описал увеличение E ( S 20 ) от шельфа к глубинам более 2000 м у двустворчатых моллюсков в западной и восточной части Северной Атлантики.Объяснение наблюдаемого отклонения от общего тренда непосредственно к югу от хребта GIF неизвестно и требует исчерпывающих данных об экологических условиях региона. Вероятным объяснением является то, что океанографические условия непосредственно к югу от хребта GIF могут повлиять на модели биоразнообразия (Gudmundsson, 1998), в частности, на относительно быстрое течение ниже глубины ∼1500 м, несущее Североатлантические глубоководные воды (NADW), смешанные с прохладными поверхностными водами. от северных морей на запад вдоль континентального склона Исландии (Dickson et al., 1990; Логеманн и др., 2013). Текущие режимы могут также воздействовать на свойства отложений за счет ресуспендирования мелких отложений, при этом размер частиц отложений признан важным фактором разнообразия бентоса (Etter and Grassle, 1992; Gray, 2002). Численное моделирование отложений показало, что мелкозернистые отложения или глина не увеличивались с увеличением глубины к югу от хребта GIF, как это наблюдалось к северу от хребта GIF (Ostmann et al., 2014). Тем не менее, из-за вероятного неравномерного расположения отложений вдоль батиметрического градиента (Gray, 1981) и нескольких мест отбора проб бентоса (11), включенных в исследование (Ostmann et al., 2014), важность состава отложений в формировании тенденций разнообразия к югу от хребта GIF остается неопределенной.
Численное моделирование отложений показало, что мелкозернистые отложения или глина не увеличивались с увеличением глубины к югу от хребта GIF, как это наблюдалось к северу от хребта GIF (Ostmann et al., 2014). Тем не менее, из-за вероятного неравномерного расположения отложений вдоль батиметрического градиента (Gray, 1981) и нескольких мест отбора проб бентоса (11), включенных в исследование (Ostmann et al., 2014), важность состава отложений в формировании тенденций разнообразия к югу от хребта GIF остается неопределенной.
Сравнение двустворчатых моллюсков и брюхоногих моллюсков
Общие тенденции α- и β-разнообразия двустворчатых моллюсков и брюхоногих моллюсков к северу и югу от хребта GIF были одинаковыми, но между группами были и заметные различия. Различие не является неожиданным, поскольку двустворчатые моллюски и брюхоногие моллюски развивались независимо на протяжении почти 500 миллионов лет и имеют разные требования к среде обитания и механизмы питания, которые следует учитывать (Ponder and Lindberg, 2008). Большинство двустворчатых моллюсков имеют низкую подвижность, включают многие виды инфауны и, как правило, питаются взвесью или отложениями (Roy et al., 2000). С другой стороны, брюхоногие моллюски более богаты видами, чем двустворчатые моллюски, функционально более разнообразны, действуют как хищники, травоядные, всеядные, питающиеся отложениями или плотоядные, и в основном относятся к эпифауне (Hughes, 1986).
Большинство двустворчатых моллюсков имеют низкую подвижность, включают многие виды инфауны и, как правило, питаются взвесью или отложениями (Roy et al., 2000). С другой стороны, брюхоногие моллюски более богаты видами, чем двустворчатые моллюски, функционально более разнообразны, действуют как хищники, травоядные, всеядные, питающиеся отложениями или плотоядные, и в основном относятся к эпифауне (Hughes, 1986).
Учитывая это, вполне возможно, что большее филогенетическое и функциональное разнообразие брюхоногих может объяснить большую необъяснимую изменчивость в общих сообществах брюхоногих моллюсков по сравнению с сообществами двустворчатых моллюсков.Несмотря на эти различия, наборы видов обеих таксономических групп различались больше в зависимости от глубины воды, чем от региона, что указывает на то, что глубина воды и, в частности, изменения окружающей среды, связанные с глубиной воды, являются важным фактором β-разнообразия в обоих регионах. Это согласуется с McClain et al. (2012), которые описали глубину как лучший показатель β-разнообразия глубоководных двустворчатых моллюсков, чем географическое разделение. Наше исследование данных BIOICE также показало, что значительное количество видов было общим для регионов к северу и югу от хребта GIF, несмотря на различия в окружающей среде.
(2012), которые описали глубину как лучший показатель β-разнообразия глубоководных двустворчатых моллюсков, чем географическое разделение. Наше исследование данных BIOICE также показало, что значительное количество видов было общим для регионов к северу и югу от хребта GIF, несмотря на различия в окружающей среде.
Бета-разнообразие
Возможно, что наблюдаемые закономерности β-разнообразия в сочетании с быстрым уменьшением α-разнообразия вдоль батиметрического градиента могут в некоторой степени отражать геологическую историю Северной Атлантики (Barry et al., 2013). Во время последнего ледникового максимума ледяные щиты покрывали северные моря, ограничивая первичную продукцию на поверхности и последующий нисходящий поток органического вещества на морское дно. Бентическая среда резко изменилась, когда лед отступил примерно 14 тысяч лет назад, что привело к значительному увеличению первичной продукции на поверхности и изменению циркуляции воды (Смит и др., 1997; Мюллер и др. , 2009). В то время как плейстоценовые оледенения, вероятно, уничтожили большую часть шельфовой и глубоководной фауны в Арктике, другая шельфовая фауна нашла убежище в глубоководных районах и считается предковой фауной некоторых представителей современной арктической глубоководной фауны (Nesis, 1984). Таким образом, большая часть фауны шельфов северных морей, вероятно, была завезена из Северной Атлантики в голоценовый период времени.
, 2009). В то время как плейстоценовые оледенения, вероятно, уничтожили большую часть шельфовой и глубоководной фауны в Арктике, другая шельфовая фауна нашла убежище в глубоководных районах и считается предковой фауной некоторых представителей современной арктической глубоководной фауны (Nesis, 1984). Таким образом, большая часть фауны шельфов северных морей, вероятно, была завезена из Северной Атлантики в голоценовый период времени.
Хотя геологическая история Северной Атлантики, вероятно, сыграла значительную роль в формировании видового состава северных морей, недавние исследования предоставили доказательства, подтверждающие предыдущую гипотезу, указывающую на то, что динамика энергии, включая химическую энергию (т.например, доступность пищи) и кинетическая энергия (т. е. температура) являются наиболее важными движущими силами моделей как α-, так и β-разнообразия в бентической морской среде (Tittensor et al., 2010; Brault et al., 2013a; Woolley et al. и др., 2016; Ясухара и Дановаро, 2016). Температура является особенно важным фактором при температурах ниже 5°C (Yasuhara and Danovaro, 2016), и, таким образом, переход от ∼2–4°C к югу от хребта GIF к ∼–1°C к северу от хребта, несомненно, представляет присутствие многих видов. с физиологическим барьером.Физиологический эффект гидростатического давления в сочетании с низкими температурами также может объяснить особенно низкое разнообразие, наблюдаемое в глубоководном бассейне Северного моря (Brown and Thatje, 2014). Доступность пищи, температура и гидростатическое давление не являются взаимоисключающими факторами, но могут синергически влиять на продуктивность видов (например, Brockington and Clarke, 2001; Sebert, 2002). Поверхность океана к северу и югу от хребта GIF демонстрирует значительную сезонность в первичной продукции на поверхности, но в Исландском море, к северу от хребта GIF, периоды цветения фитопланктона заметно короче (Pálsson et al., 2012; МакГинти и др., 2016). Следовательно, бентосное сообщество в глубоководных северных морях может состоять в основном из видов, которые успешно адаптировались к экстремальным энергетическим ограничениям в регионе, то есть к отрицательным температурам и высокой пульсации химической энергии.
На основании разделения Baselga (2010) и рангового порядка BINMATNEST для сообществ двустворчатых моллюсков и брюхоногих моллюсков к северу от хребта GIF увеличение несходства сообществ в значительной степени связано с увеличением гнездовья или исчезновением видов с глубиной и связано со снижением α- разнообразие.Браулт и др. (2013b) также сообщили о снижении α-разнообразия в сочетании с увеличением гнездовья от батиальных до абиссальных глубин у необрюхоногих моллюсков. Они утверждали, что эта закономерность свидетельствует о динамике стока-источника, т. е. о том, что абиссальные популяции не являются самоподдерживающимися из-за плохого качества среды обитания на больших глубинах и требуют интродукции особей из более мелких местообитаний (Rex et al., 2005; Brault). и др., 2013b). Точно так же увеличение количества гнезд к северу от хребта GIF в сочетании с уменьшением α-разнообразия может указывать на то, что по крайней мере некоторые популяции двустворчатых моллюсков и брюхоногих моллюсков не являются самоподдерживающимися в более глубокой части их наблюдаемых диапазонов глубин, а вместо этого требуют интродукции из «исходной» популяции на меньших глубинах, где пищи больше, а состояние насыщения карбонатом кальция (Ω) выше.Например, было показано, что двустворчатым моллюскам для успешного личиночного развития требуется видовая специфичность Ω (Waldbusser et al., 2015). Однако, как отмечают Brault et al. (2013b) такие модели разнообразия не дают достаточных доказательств для установления существования динамики сток-источник в метапопуляции. Также необходима информация о генетической структуре популяций «источника» и «приемника» (например, Manier and Arnold, 2005).
Вклад оборота видов в β-разнообразие был значительно выше к югу от хребта GIF, где результирующая гнездовья несходство было менее значительным.Тем не менее, анализ вложенности BINMATNEST показал увеличение вложенности с глубиной к югу от хребта GIF, хотя корреляция была относительно слабой по сравнению с таковой к северу от хребта GIF. Это было неожиданным открытием, поскольку ожидалось, что смена видов будет в большей степени доминировать в β-разнообразии в этом регионе. Однако этот вывод, вероятно, связан с неожиданным уменьшением α-разнообразия с увеличением глубины, наблюдаемым к югу от хребта ГИП, что опять-таки, возможно, является результатом особого океанографического режима непосредственно к югу от хребта ГИП.
Совершенно очевидно, что важно углубить понимание относительного вклада различных факторов окружающей среды, связанных с глубиной (например, гидростатического давления, доступности пищи, температуры, Ω и pH) в модели биоразнообразия в глубоководных районах. Исследования, направленные на углубление этого понимания, остаются сложной задачей из-за значительной корреляции множества параметров окружающей среды вдоль градиента глубины. Недавние исследования, однако, признали доступность энергии в качестве основной движущей силы моделей разнообразия в глубоководных районах (Brault et al., 2013а; Вулли и др., 2016). Тем не менее вполне вероятно, что другие параметры окружающей среды также играют роль в формировании моделей разнообразия в отдельных группах. Например, уменьшение Ω, связанное с глубиной, может быть важным фактором биоразнообразия в таксономических группах, исследованных в этом исследовании, и других кальцифицирующих таксонах.
Идентификация видов-индикаторов
Большинство видов двустворчатых моллюсков и брюхоногих моллюсков, отобранных к северу от хребта GIF, были зарегистрированы на глубинах менее 500 м, при этом несколько видов демонстрировали широкое распространение по всему батиальному ареалу.Интересно, что эпибентосный двустворчатый моллюск Hyalopecten frigidus был обычным явлением в пробах, собранных на глубине ниже 2000 м, что позволяет предположить, что он хорошо приспособлен к энергетическим ограничениям глубоководного бассейна Северного моря и низкого Ω. В немногочисленных пробах, отобранных ниже 2000 м к северу от хребта ГИП, видов брюхоногих обнаружено не было.
Всего было 15 видов, считающихся подходящими для мониторинга ранних признаков изменений в бентических сообществах в условиях закисления океана к северу от хребта GIF.Образцы, отобранные для программы BIOICE, были собраны за 10–27 лет до этого исследования, поэтому важно понимать, что изменения в морской среде к северу от хребта GIF продолжаются и что изменения окружающей среды уже могли вызвать изменения в бентических сообществах в этот регион. Желательно, чтобы организация проекта мониторинга потребовала сбора предварительных данных о бентических сообществах и информации о естественной (например, годовой и сезонной) изменчивости бентических сообществ в регионе.
Резюме и заключение
Биоразнообразие двустворчатых моллюсков и брюхоногих моллюсков существенно различается между батиметрическими градиентами к северу и югу от хребта МФП. Явное снижение α-разнообразия и увеличение гнездовья с глубиной наблюдались к северу от хребта ГИП, тогда как умеренное снижение и отсутствие снижения α-разнообразия наблюдалось у двустворчатых моллюсков и брюхоногих соответственно к югу от хребта ГИП, где оборот видов является доминирующим компонентом β-разнообразия.Методы, примененные к набору данных BIOICE в этом исследовании, были разработаны для решения проблем, связанных со стратегией нерандомизированной выборки и отсутствием стандартизированных методов выборки. Тем не менее, важно признать эти проблемы и потенциальное влияние на результаты этого исследования. Поскольку ограниченное количество выборочных данных с глубин ниже 1600 м было в значительной степени исключено из анализа, это исследование не рассматривает модели биоразнообразия на этих больших глубинах. Для будущих исследований рекомендуется применять стратегии выборки, которые лучше разработаны для ответов на вопросы о моделях биоразнообразия, чтобы подтвердить результаты этого исследования.Представленные здесь данные вносят значительный вклад в изучение биоразнообразия и видового состава моллюсков вдоль батиметрических градиентов в высоких широтах Северной Атлантики. Эта информация важна для будущих попыток оценить степень уязвимости сообществ двустворчатых моллюсков и брюхоногих моллюсков в Северных морях, где наблюдаются и прогнозируются экологические изменения, связанные с потеплением и закислением океана (Olafsson et al., 2009; Bopp et al., 2013). ) и за усилия по прогнозированию последствий этих изменений для глубоководных бентических сообществ в целом (Widdicombe and Spicer, 2008; Urban et al., 2016).
Добавляя к общему пониманию модели биоразнообразия в Северной Атлантике, включая Северные моря, наши результаты обеспечивают важную основу для будущей работы, направленной на количественную оценку реакции экосистем на быстрые изменения окружающей среды, происходящие в регионе, — информация, необходимая для формирования стратегий сохранения которые минимизируют пагубные последствия таких изменений в будущем (Cavanagh et al., 2016).
Доступность данных
Необработанные данные, подтверждающие выводы этой рукописи, будут предоставлены авторами без неоправданных оговорок любому квалифицированному исследователю.
Вклад авторов
HE написал рукопись и проанализировал данные. Н. М. принимал участие в написании рукописи и анализе данных. GG отредактировал рукопись и дал рекомендации по поводу данных о видах.
Финансирование
Эта работа была поддержана «Европейским проектом по закислению океана» EPOCA, который получил финансирование от Седьмой рамочной программы Европейского сообщества (FP7/2007-2013) в соответствии с соглашением о гранте №. 211384. HE также получил грант от Fisheries Iceland.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы хотели бы отметить ценную работу всех тех, кто принимал участие в отборе проб и идентификации двустворчатых моллюсков и брюхоногих моллюсков, собранных в рамках программы BIOICE, особенно Хенка Х. Джикстры и Джона-Арне Снели. Мы также хотим поблагодарить Jörundur Svavarsson и Jed McDonald за подробные комментарии к рукописи.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fmars.2019.00129/full#supplementary-material
.Каталожные номера
Алмейда-Нето М., Гимарайнш П., Гимарайнш П. Р., Лойола Р. Д. и Ульрих В. (2008). Последовательная метрика для анализа вложенности в экологических системах: согласование концепции и измерения. Ойкос 117, 1227–1239. doi: 10.1111/j.0030-1299.2008.16644.x
Полнотекстовая перекрестная ссылка | Академия Google
Аонги, Д.М. (1990). Экология тропических мягкодонных донных экосистем. Океаногр. Мар биол. Анну. Ред. 28, 381–496.
Академия Google
Эндрю К. и Скотт Л. (2000). Пространственные закономерности разнообразия в море: видовое богатство мшанок в Северной Атлантике. Дж. Аним. Экол. 69, 799–814. doi: 10.1046/j.1365-2656.2000.00440.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Барри, Т., Berteaux, D., Bültmann, H., Christiansen, J.S., Cook, J.A., Dahlberg, A., et al. (2013). Оценка биоразнообразия Арктики, 2013 г. Доступно на: https://www.caff.is/assessment-series/233-arctic-biodiversity-assessment-2013
Академия Google
Басельга, А. (2010). Разделение компонентов оборота и вложенности бета-разнообразия. Глоб. Экол. Биогеогр. 19, 134–143. doi: 10.1111/j.1466-8238.2009.00490.x
Полнотекстовая перекрестная ссылка | Академия Google
Басельга, А.(2012). Взаимосвязь между заменой видов, несходством, обусловленным вложенностью, и вложенностью. Глоб. Экол. Биогеогр. 21, 1223–1232. doi: 10.1111/j.1466-8238.2011.00756.x
Полнотекстовая перекрестная ссылка | Академия Google
Baselga, A., and Orme, C.D.L. (2012). betapart: пакет R для изучения бета-разнообразия. Методы Экол. Эвол. 3, 808–812. doi: 10.1111/j.2041-210X.2012.00224.x
Полнотекстовая перекрестная ссылка | Академия Google
Бетт, Б.Дж. (2001). Исследование окружающей среды на атлантической окраине Великобритании: введение и обзор экологии батиального бентоса. Прод. Полка Рез. 21, 917–956. doi: 10.1016/S0278-4343(00)00119-9
Полнотекстовая перекрестная ссылка | Академия Google
Бодил, Б.А., Амброуз, В.Г. мл., Бергманн, М., Клаф, Л.М., Гебрук, А.В., Хасеманн, С., и соавт. (2011). Разнообразие арктического глубоководного бентоса. Мар. Биодайверы. 41, 87–107. doi: 10.1007/s12526-010-0078-4
Полнотекстовая перекрестная ссылка | Академия Google
Бопп, Л., Resplandy, L., Orr, J.C., Doney, S.C., Dunne, J.P., Gehlen, M., et al. (2013). Множественные факторы стресса для океанских экосистем в 21 веке: прогнозы с использованием моделей CMIP5. Биогеонауки 10, 6225–6245. doi: 10.5194/bg-10-6225-2013
Полнотекстовая перекрестная ссылка | Академия Google
Буше П. и Варен А. (1979). Абиссальная фауна моллюсков Норвежского моря и ее связь с другими фаунами. Сарсия 64, 211–243. дои: 10.1080/00364827.1979.10411383
Полнотекстовая перекрестная ссылка | Академия Google
Браттегард, Т.и Фосса, Дж. Х. (1991). Воспроизводимость эпибентического пробоотборника. J. Mar. Biol. доц. Великобритания 71, 153–166. дои: 10.1017/S0025315400037462
Полнотекстовая перекрестная ссылка | Академия Google
Браулт, С., Стюарт, К.Т., Вагстафф, М.К., Макклейн, К.Р., Аллен, Дж.А., и Рекс, М.А. (2013a). Контрастные модели α- и β-разнообразия у глубоководных двустворчатых моллюсков восточной и западной части Северной Атлантики. Deep Sea Res. Часть II Верх. Стад. океаногр. 92, 157–164. doi: 10.1016/j.дср2.2013.01.018
Полнотекстовая перекрестная ссылка | Академия Google
Браулт, С., Стюарт, К.Т., Вагстафф, М.К., и Рекс, Массачусетс (2013b). Географические свидетельства динамики стока-источника у глубоководных необрюхоногих в восточной части Северной Атлантики: подход с использованием гнездового анализа. Глоб. Экол. Биогеогр. 22, 433–439. doi: 10.1111/geb.12005
Полнотекстовая перекрестная ссылка | Академия Google
Брикс С., Странски Б., Малютина М., Пабис К., Сваварссон Дж. и Риль Т.(2018). Распространение изопод (Crustacea) в исландских и прилегающих водах. Мар. Биодайверы. 48, 783–811. doi: 10.1007/s12526-018-0871-z
Полнотекстовая перекрестная ссылка | Академия Google
Брокингтон, С., и Кларк, А. (2001). Относительное влияние температуры и пищи на метаболизм морского беспозвоночного. Дж. Экспл. Мар биол. Экол. 258, 87–99. doi: 10.1016/S0022-0981(00)00347-6
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Браун, А.и Татье, С. (2014). Объяснение моделей батиметрического разнообразия морских донных беспозвоночных и донных рыб: физиологический вклад в адаптацию жизни на глубине. биол. Ред. 89, 406–426. doi: 10.1111/brv.12061
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кавана, Р. Д., Бросайт, С., Пиллинг, Г. М., Грант, С. М., Мерфи, Э. Дж., и Остин, М. К. (2016). Оценка биоразнообразия и экосистемных услуг: полезный способ управления и сохранения морских ресурсов? Проц.Р. Соц. Б биол. науч. 283:20161635. doi: 10.1098/rspb.2016.1635
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чайлдресс, Дж. Дж. (1995). Существуют ли физиологические и биохимические адаптации обмена веществ у глубоководных животных? Тренды Экол. Эвол. 10, 30–36. дои: 10.1016/S0169-5347(00)88957-0
Полнотекстовая перекрестная ссылка | Академия Google
Чиу, К.-Х., Ван, Ю.-Т., Вальтер, Б.А., и Чао, А. (2014). Улучшенная непараметрическая нижняя граница видового богатства с помощью модифицированной формулы частоты Гуд-Тьюринга. Биометрия 70, 671–682. doi: 10.1111/biom.12200
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Crain, C.M., Kroeker, K., и Halpern, B.S. (2008). Интерактивное и кумулятивное воздействие нескольких факторов стресса человека в морских системах. Экол. лат. 11, 1304–1315. doi: 10.1111/j.1461-0248.2008.01253.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Даль, Э. (1979). Amphipoda Gammaridea из глубин Норвежского моря.Предварительный отчет. Сарсия 64, 57–59. дои: 10.1080/00364827.1979.10411363
Полнотекстовая перекрестная ссылка | Академия Google
Дальгрен, Т. Г., Вайнберг, Дж. Р., и Галаныч, К. М. (2000). Филогеография океанического квахога ( Arctica islandica ): влияние палеоклимата на генетическое разнообразие и ареал видов. Мар. Биол. 137, 487–495. doi: 10.1007/s002270000342
Полнотекстовая перекрестная ссылка | Академия Google
Диксон, Р. Р., Gmitrowicz, E.M., and Watson, A.J. (1990). Глубоководное обновление в северной части Северной Атлантики. Природа 344, 848–850. дои: 10.1038/344848a0
Полнотекстовая перекрестная ссылка | Академия Google
Эттер, Р. Дж., и Грассл, Дж. Ф. (1992). Закономерности разнообразия видов в глубоком море в зависимости от разнообразия размеров частиц отложений. Природа 360, 576–578. дои: 10.1038/360576a0
Полнотекстовая перекрестная ссылка | Академия Google
Гаттузо, Дж. П., Маньян, А., Billé, R., Cheung, W.W.L., Howes, E.L., Joos, F., et al. (2015). Контрастные варианты будущего для океана и общества в зависимости от различных сценариев антропогенных выбросов CO2. Наука 349:aac4722. doi: 10.1126/science.aac4722
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Gazeau, F., Parker, L.M., Comeau, S., Gattuso, J.-P., O’Connor, W.A., Martin, S., et al. (2013). Воздействие закисления океана на морских моллюсков. Мар. Биол. 160, 2207–2245.doi: 10.1007/s00227-013-2219-3
Полнотекстовая перекрестная ссылка | Академия Google
Гелен, М., Сеферян, Р., Джонс, Д. О. Б., Рой, Т., Рот, Р., Барри, Дж., и др. (2014). Прогнозируемое снижение pH к 2100 году может поставить под угрозу глубоководное биоразнообразие Северной Атлантики. Биогеонауки 11, 6955–6967. doi: 10.5194/bg-11-6955-2014
Полнотекстовая перекрестная ссылка | Академия Google
Грей, Дж. С. (1981). Экология морских отложений. Кембридж: Издательство Кембриджского университета.
Академия Google
Грей, Дж. С. (2002). Видовое богатство морских мягких отложений. Мар. Экол. прог. сер. 244, 285–297. doi: 10.3354/meps244285
Полнотекстовая перекрестная ссылка | Академия Google
Гудмундссон, Г. (1998). Пределы распространения видов Pyrgo на биогеографических рубежах Арктики и Северо-Атлантического бореала. Дж. Форам. Рез. 28, 240–256.
Академия Google
Гудмундссон, Г. (2002).Эволюция и биполярное распространение Cornuspiroides (foraminifera): филогенетический вывод с использованием морфологических признаков и экономии. Дж. Форам. Рез. 32, 308–318. дои: 10.2113/32.3.308
Полнотекстовая перекрестная ссылка | Академия Google
Гутьеррес, Дж. Л., Джонс, К. Г., Страйер, Д. Л., и Ирибарн, О. О. (2003). Моллюски как инженеры экосистем: роль производства раковин в водной среде обитания. Ойкос 101, 79–90. doi: 10.1034/j.1600-0706.2003.12322.x
Полнотекстовая перекрестная ссылка | Академия Google
Холл-Спенсер, Дж., и Мур, П. (2000). Limaria hians (Mollusca: Limacea): заброшенный рифообразующий краеугольный камень. Аква. Консерв. 10, 267–277. doi: 10.1002/1099-0755(200007/08)10:4<267::AID-AQC407>3.0.CO;2-B
Полнотекстовая перекрестная ссылка | Академия Google
Хансен, Б., и Мейнке, Дж. (1979). Водовороты и меандры в районе Исландско-Фарерского хребта. Deep Sea Res. Часть А Океаногр.Рез. Пап. 26, 1067–1082. дои: 10.1016/0198-0149(79)
Полнотекстовая перекрестная ссылка | Академия Google
Хансен, Б., и Остерхус, С. (2000). Обмены между Северной Атлантикой и Северными морями. Прог. океаногр. 45, 109–208. doi: 10.1016/S0079-6611(99)00052-X
Полнотекстовая перекрестная ссылка | Академия Google
Хесслер, Р. Р., и Сандерс, Х. Л. (1967). Разнообразие фауны глубоководных районов. Deep Sea Res. океаногр. Абстр. 14, 65–70. дои: 10.1016/0011-7471(67)-0
Полнотекстовая перекрестная ссылка | Академия Google
Хесслер, Р.Р., Уилсон Г.Д. и Тистл Д. (1979). Глубоководные изоподы: биогеографический и филогенетический обзор. Сарсия 64, 67–75. дои: 10.1080/00364827.1979.10411365
Полнотекстовая перекрестная ссылка | Академия Google
Хиггс, Н., и Аттрилл, М. (2015). Предубеждения в биоразнообразии: широко распространенные виды сначала обнаруживаются в глубоком море. Перед. мар. 2:61. doi: 10.3389/fmars.2015.00061
Полнотекстовая перекрестная ссылка | Академия Google
Хофманн, Г.E., Smith, J.E., Johnson, K.S., Send, U., Levin, L.A., Micheli, F., et al. (2011). Высокочастотная динамика рН океана: сравнение нескольких экосистем. PLoS One 6:e28983. doi: 10.1371/journal.pone.0028983
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Horton, T., Kroh, A., Ahyong, S., Bailly, N., Boyko, C.B., Brandão, S.N., et al. (2019). Всемирный регистр морских видов WoRMS. Доступно на: http://www.marinespecies.org на VLIZ
Академия Google
Хьюз, Р.Н. (1986). Функциональная биология морских брюхоногих моллюсков. Берлин: Springer.
Академия Google
Якобссон, М. (2002). Гипсометрия и объем Северного Ледовитого океана и составляющих его морей. Геохим. Геофиз. Геосист. 3, 1–18. дои: 10.1029/2001gc000302
Полнотекстовая перекрестная ссылка | Академия Google
Йост, А. Б., Ясухара, М., Окахаши, Х., Брикс, С., Мартинес Арбису, П., и Остманн, А. (2018). Биогеографическое распространение вида Cytheropteron (Ostracoda) в исландских водах (субполярная часть Северной Атлантики). Мар. Биодайверы. 48, 763–782. doi: 10.1007/s12526-018-0867-8
Полнотекстовая перекрестная ссылка | Академия Google
Кенниш, MJ (2000). Практическое пособие по морским наукам. Бока-Ратон, Флорида: CRC Press.
Академия Google
Линс, К., Гриффитс, Х.Дж., Барнс, Д.К.А., и Кларк, А. (2006). Биоразнообразие и биогеография антарктических и субантарктических моллюсков. Deep Sea Res. Часть II Верх. Стад. океаногр. 53, 985–1008. дои: 10.1016/j.dsr2.2006.05.003
Полнотекстовая перекрестная ссылка | Академия Google
Логеманн, К., Олафссон, Дж., Сноррасон, А., Вальдимарссон, Х., и Мартейнсдоттир, Г. (2013). Циркуляция исландских вод – модельное исследование. Науки об океане. 9, 931–955. doi: 10.5194/osd-10-763-2013
Полнотекстовая перекрестная ссылка | Академия Google
Лутц, М. Дж., Калдейра, К., Данбар, Р. Б., и Беренфельд, М. Дж. (2007). Сезонные ритмы чистой первичной продукции и потока органического углерода в виде твердых частиц на глубину описывают эффективность биологического насоса в мировом океане. Ж. Геофиз. Рез. Океаны 112, 1–26. дои: 10.1029/2006JC003706
Полнотекстовая перекрестная ссылка | Академия Google
Макенсен, А., Сейруп, Х., и Янсен, Э. (1985). Распределение живых бентосных фораминифер на континентальном склоне и возвышенности у юго-западной части Норвегии. Мар. Микропалеонтол. 9, 275–306. дои: 10.1016/0377-8398(85)-5
Полнотекстовая перекрестная ссылка | Академия Google
Мальмберг, С.-А., и Вальдимарссон, Х. (2003). Гидрографические условия в водах Исландии, 1990–1999 гг. ICES Mar. Sci. Симп. 219, 50–60.
Академия Google
Манье, М.К., и Арнольд, С.Дж. (2005). Популяционный генетический анализ определяет динамику «источник-приемник» для двух видов симпатрических подвязочных змей ( Thamnophis elegans и Thamnophis sirtalis ). Мол. Экол. 14, 3965–3976. doi: 10.1111/j.1365-294X.2005.02734.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
May, R.M., and Godfrey, J. (1994).Биологическое разнообразие: различия между сушей и морем. Филос. Транс. Р. Соц. Лонд. Б биол. науч. 343, 105–111. doi: 10.1098/rstb.1994.0014
Полнотекстовая перекрестная ссылка | Академия Google
Макклейн, Ч.Р., и Рекс, Массачусетс (2015). К концептуальному пониманию β-разнообразия глубоководного бентоса. Год. Преподобный Экол. Эвол. Сист. 46, 623–642. doi: 10.1146/annurev-ecolsys-120213-0
Полнотекстовая перекрестная ссылка | Академия Google
Макклейн, К.Р., Стеген, Дж. К., и Херлберт, А. Х. (2012). Расселение, экологические ниши и оборот глубоководных двустворчатых моллюсков в океаническом масштабе. Проц. Р. Соц. Лонд. Б биол. науч. 279, 1993–2002 гг. doi: 10.1098/rspb.2011.2166
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
МакГинти, Н., Гудмундссон, К., Агустсдоттир, К., и Мартейнсдоттир, Г. (2016). Экологические и климатические эффекты изменчивости хлорофилла- a вокруг Исландии с использованием реконструированных полей спутниковых данных. Дж. Мар. Сист. 163, 31–42. doi: 10.1016/j.jmarsys.2016.06.005
Полнотекстовая перекрестная ссылка | Академия Google
Мюллер, Дж., Массе, Г., Штейн, Р., и Белт, С. Т. (2009). Изменчивость состояния морского льда в проливе Фрама за последние 30 000 лет. Нац. Geosci. 2, 772–776. дои: 10.1038/ngeo665
Полнотекстовая перекрестная ссылка | Академия Google
Нараянасвами, Б. Э., Бетт, Б. Дж., и Гейдж, Дж. Д. (2005). Экология фауны батиальных полихет на границе Арктики и Атлантики (Фареро-Шетландский пролив, Северо-Восточная Атлантика). Мар. Биол. Рез. 1, 20–32. дои: 10.1080/17451000510018971
Полнотекстовая перекрестная ссылка | Академия Google
Несис, К. (1984). Гипотеза о происхождении западных и восточных арктических ареалов морских донных животных. Сов. Дж. Мар. Биол. 9, 235–243.
Академия Google
Оксанен Дж., Бланше Ф.Г., Киндт Р., Лежандр П., Минчин П.Р., О’Хара Р.Б. и соавт. (2015). веган: Экологический пакет сообщества. Пакет R версии 2.3-0. Доступно по адресу: http://CRAN.R-project.org/package=(vegan
Академия Google
Олафссон, Дж. (2003). Питательные вещества зимнего смешанного слоя в морях Ирмингера и Исландии, 1990-2000 гг. ICES Mar. Sci. Симп. 219, 329–332.
Академия Google
Олафссон, Дж., Олафсдоттир, С.Р., Бенуа-Каттин, А., Даниэльсен, М., Арнарсон, Т.С., и Такахаши, Т. (2009). Скорость закисления Исландского моря по измерениям временных рядов. Биогеонауки 6, 2661–2668.doi: 10.5194/bg-6-2661-2009
Полнотекстовая перекрестная ссылка | Академия Google
Остманн, А., Шнурр, С., и Мартинес Арбису, П. (2014). Морская среда вокруг Исландии: гидрография, отложения и первые прогностические модели характеристик глубоководных отложений Исландии. Пол. Полярный рез. 35, 151–176. doi: 10.2478/popore-2014-0021
Полнотекстовая перекрестная ссылка | Академия Google
Оуг Э., Баккен Т., Конгсруд Дж. А. и Алвестад Т. (2017). Многощетинковые кольчатые черви в глубоководных северных морях: сильные батиметрические градиенты, низкое разнообразие и неразвитая таксономия. Deep Sea Res. Часть II Верх. Стад. океаногр. 137, 102–112. doi: 10.1016/j.dsr2.2016.06.016
Полнотекстовая перекрестная ссылка | Академия Google
Палссон, О.К., Гисласон, А., Гудфиннссон, Х.Г., Гуннарссон, Б., Олафсдоттир, С.Р., Петурсдоттир, Х., и др. (2012). Структура экосистемы Исландского моря и недавние изменения в популяции мойвы ( Mallotus villosus ). ICES J. Mar. Sci. 69, 1242–1254. дои: 10.1093/icesjms/fss071
Полнотекстовая перекрестная ссылка | Академия Google
Пипенбург, Д., Archambault, P., Ambrose, W.G., Blanchard, A.L., Bluhm, B.A., Carroll, M.L., et al. (2011). К панарктической инвентаризации видового разнообразия макро- и мегабентической фауны арктических шельфовых морей. Мар. Биодайверы. 41, 51–70. doi: 10.1007/s12526-010-0059-7
Полнотекстовая перекрестная ссылка | Академия Google
Пондер, В.Ф., и Линдберг, Д.Р. (2008). Филогения и эволюция моллюсков. Беркли, Калифорния: Издательство Калифорнийского университета. дои: 10.1525/Калифорния/9780520250925.001.0001
Полнотекстовая перекрестная ссылка | Академия Google
Queirós, A.M., Birchenough, S.N.R., Bremner, J., Godbold, J.A., Parker, R.E., Romero-Ramirez, A., et al. (2013). Классификация биотурбации европейских морских млекопитающих. Экол. Эвол. 3, 3958–3985. doi: 10.1002/ece3.769
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
R-Core-Team (2018). R: Язык и среда для статистических вычислений. Вена: Фонд статистических вычислений R.
Академия Google
Рекс, Массачусетс, и Эттер, Р. Дж. (2010). Глубоководное биоразнообразие: модель и масштаб. Кембридж, Массачусетс: Издательство Гарвардского университета.
Академия Google
Rex, M.A., McClain, C.R., Johnson, N.A., Etter, R.J., Allen, J.A., Bouchet, P., et al. (2005). Гипотеза источника-поглотителя абиссального биоразнообразия. утра. Нац. 165, 163–178. дои: 10.1086/427226
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рекс, М.А., Стюарт, К.Т., и Койн, Г. (2000). Широтные градиенты видового богатства глубоководного бентоса Северной Атлантики. Проц. Натл. акад. науч. США 97, 4082–4085. doi: 10.1073/pnas.050589497
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Родригес-Хиронес, А.М., и Сантамария, Л. (2010). Как поведение при поиске пищи и разделение ресурсов могут влиять на эволюцию цветов и структуру сетей опыления. Открытый экол. Дж. 3, 1–11. дои: 10.2174/1874213001003040001
Полнотекстовая перекрестная ссылка | Академия Google
Родригес-Хиронес, Массачусетс, и Сантамария, Л. (2006). Новый алгоритм расчета температуры вложенности матриц присутствия-отсутствия. Ж. Биогеогр. 33, 924–935. doi: 10.1111/j.1365-2699.2006.01444.x
Полнотекстовая перекрестная ссылка | Академия Google
Роджерс, AD (2015). Изменение окружающей среды в глубоком океане. Год. Преподобный Окружающая среда. Ресурс. 40, 1–38.doi: 10.1146/annurev-environ-102014-021415
Полнотекстовая перекрестная ссылка | Академия Google
Ротлисберг П. и Пирси В. (1977). Эпибентический пробоотборник, использованный для изучения онтогенеза вертикальной миграции Pandalus jordani (Decapoda, Caridea). Рыба. Бык. 74, 994–997.
Академия Google
Рой К., Яблонски Д. и Валентайн Дж. В. (2000). Анализ градиентов широтного разнообразия: функциональные группы и клады морских двустворчатых моллюсков. Проц. Р. Соц. Лонд. Б биол. науч. 267, 293–299. doi: 10.1098/rspb.2000.0999
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шнурр С., Осборн К.Дж., Малютина М., Дженнингс Р., Брикс С., Дрискелл А. и соавт. (2018). Скрытое разнообразие в двухвидовых комплексах изопод муннопсид (Crustacea) на переходе между самой северной частью Северной Атлантики и северными морями. Мар. Биодайверы. 48, 813–843. doi: 10.1007/s12526-018-0877-6
Полнотекстовая перекрестная ссылка | Академия Google
Себерт, П.(2002). Рыба под высоким давлением: столетняя история. Комп. Биохим. Физиол. Мол. интегр. Физиол. 131, 575–585. doi: 10.1016/S1095-6433(01)00509-8
Полнотекстовая перекрестная ссылка | Академия Google
Сейбел, Б.А., и Уолш, П.Дж. (2003). Биологическое воздействие глубоководной закачки углекислого газа, выведенное из показателей физиологической работоспособности. Дж. Экспл. биол. 206, 641–650. doi: 10.1242/jeb.00141
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Скоген, М.Д., Олсен А., Боршейм К.Ю., Сандо А.Б. и Скьелван И. (2014). Моделирование закисления океана в Северном и Баренцевом морях в нынешнем и будущем климате. Дж. Мар. Сист. 131, 10–20. doi: 10.1016/j.jmarsys.2013.10.005
Полнотекстовая перекрестная ссылка | Академия Google
Smith, J.E., Risk, M.J., Schwarcz, H.P., and McConnaughey, T.A. (1997). Быстрое изменение климата в Северной Атлантике во время позднего дриаса, зафиксированное глубоководными кораллами. Природа 386, 818–820.дои: 10.1038/386818a0
Полнотекстовая перекрестная ссылка | Академия Google
Снелгроув, ПВР (1999). Достигая дна морского биоразнообразия: осадочные среды обитания: дно океана является наиболее распространенной средой обитания на Земле и поддерживает высокое биоразнообразие и ключевые экосистемные услуги. Биологические науки 49, 129–138. дои: 10.2307/1313538
Полнотекстовая перекрестная ссылка | Академия Google
Снели, Ж.-А. (1998). Простые бентические сани для мелководного и глубоководного отбора проб. Сарсия 83, 69–72. дои: 10.1080/00364827.1998.10413670
Полнотекстовая перекрестная ссылка | Академия Google
Соренсен, Т. (1948). Метод установления групп равной амплитуды в социологии растений, основанный на сходстве видов, и его применение к анализу растительности датских общин. биол. скр. 5, 1–34.
Академия Google
Стефанссон, У. (1962). Воды Северной Исландии. Рит Фискидейлдар 3:269.
Академия Google
Стефенсен, К.(1940). Морская амфипода. Зоол. Исландия 3, 1–11.
Академия Google
Стюарт, К.Т., Браулт, С., Роу, Г.Т., Вей, К.-Л., Вагстафф, М., Макклейн, К.Р., и соавт. (2017). Гнездование и замена видов вдоль батиметрических градиентов в глубоководных районах отражают продуктивность: тест с сообществами полихет в олиготрофной северо-западной части Мексиканского залива. Ж. Биогеогр. 44, 548–555. дои: 10.1111/jbi.12810
Полнотекстовая перекрестная ссылка | Академия Google
Сваварссон, Дж., Стромберг, Дж.-О., и Браттегард, Т. (1993). Фауна глубоководных аселлот (Isopoda, Crustacea) Северных морей: видовой состав, закономерности распространения и происхождение. Ж. Биогеогр. 20, 537–555. дои: 10.2307/2845725
Полнотекстовая перекрестная ссылка | Академия Google
Свифт, Дж. Х., Аагаард, К., и Мальмберг, С.-А. (1980). Вклад разлива Датского пролива в глубинную часть Северной Атлантики. Deep Sea Res. Часть А Океаногр. Рез. Пап. 27, 29–42. дои: 10.1016/0198-0149(80)-9
Полнотекстовая перекрестная ссылка | Академия Google
Tittensor, D.P., Mora, C., Jetz, W., Lotze, H.K., Ricard, D., Berghe, E.V., et al. (2010). Глобальные закономерности и предикторы морского биоразнообразия по таксонам. Природа 466, 1098–1101. doi: 10.1038/nature09329
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ульрих, В., и Готелли, Нью-Джерси (2007). Нулевой модельный анализ паттернов вложенности видов. Экология 88, 1824–1831.дои: 10.1890/06-1208.1
Полнотекстовая перекрестная ссылка | Академия Google
Урбан, М. К., Бочеди, Г., Хендри, А. П., Михуб, Ж.-Б., Пеер, Г., Сингер, А., и др. (2016). Улучшение прогноза биоразнообразия в условиях изменения климата. Наука 353:aad8466. doi: 10.1126/science.aad8466
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Wagstaff, M.C., Howell, K.L., Bett, B.J., Billett, D.S., Brault, S., Stuart, C.T., et al. (2014). β-разнообразие глубоководных голотурий и астероидов вдоль батиметрического градиента (СВ Атлантика). Мар. Экол. прог. сер. 508, 177–185. doi: 10.3354/meps10877
Полнотекстовая перекрестная ссылка | Академия Google
Waldbusser, G.G., Hales, B., Langdon, C.J., Haley, B.A., Schrader, P., Brunner, E.L., et al. (2015). Чувствительность состояния насыщения личинок морских двустворчатых моллюсков к закислению океана. Нац. Клим. Изменить 5, 273–280. doi: 10.1038/nclimate2479
Полнотекстовая перекрестная ссылка | Академия Google
Вайссаппель, Дж., и Сваварссон, Дж. (1998).Бентосные амфиподы (Crustacea: Malacostraca) в исландских водах: разнообразие фауны от мелких до средних глубин Северного Ледовитого и Северной Атлантики. Мар. Биол. 131, 133–143. дои: 10.1007/s002270050304
Полнотекстовая перекрестная ссылка | Академия Google
Вайссаппель, Дж. Б. (2000). Распространение и разнообразие гипербентосных амфипод семейства Eusiridae в различных морях Гренландско-Исландско-Фарерского хребта. Сарсия 85, 227–236.дои: 10.1080/00364827.2000.10414575
Полнотекстовая перекрестная ссылка | Академия Google
Уиттакер, Р. Х. (1960). Растительность гор Сискию, Орегон и Калифорния. Экол. моногр. 30, 279–338. дои: 10.2307/1943563
Полнотекстовая перекрестная ссылка | Академия Google
Виддикомб, С., и Спайсер, Дж. И. (2008). Прогнозирование воздействия подкисления океана на биоразнообразие бентоса: что может сказать нам физиология животных? Дж. Экспл. Мар биол. Экол. 366, 187–197.doi: 10.1016/j.jembe.2008.07.024
Полнотекстовая перекрестная ссылка | Академия Google
Woolley, S.N.C., Tittensor, D.P., Dunstan, P.K., Guillera-Arroita, G., Lahoz-Monfort, J.J., Wintle, B.A., et al. (2016). Модели глубоководного разнообразия определяются наличием энергии. Природа 533, 393–396. doi: 10.1038/nature17937
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
двустворчатых моллюсков, ракообразных и брюхоногих моллюсков, боже мой!
Свежие морепродукты – настоящее кулинарное удовольствие.В то время как посетители наслаждаются двустворчатыми моллюсками, ракообразными и брюхоногими в любое время года, лето традиционно ассоциируется с поеданием многих из этих существ, в основном с твердым панцирем. Для студентов кулинарной школы Боулдера небольшая предыстория и несколько советов по использованию этих трех типов наземных и морских существ в блюдах могут стать полезным кулинарным вдохновением.
Двустворчатые моллюски
Во всем мире регулярно едят много двустворчатых моллюсков, от моллюсков и морских гребешков до мидий и устриц.Также существует множество менее распространенных двустворчатых моллюсков, и некоторых из них едят в меньшем количестве, где их достаточно, чтобы их можно было ловить, готовить и употреблять в пищу.
Что делает двустворчатого моллюска двустворчатым? Технический ответ предполагает их научную классификацию. С практической точки зрения, эти существа отличаются отсутствием внутренних костей — это означает, что их легко съесть целиком — относительно плоским телом и шарнирным панцирем.
Двустворчатых моллюсков можно есть самыми разными способами, от потребления в сыром виде до жарки во фритюре и приготовления супов.Ищете вдохновение? Рассмотрим эти популярные способы приготовления двустворчатых моллюсков:
- Классический моллюск из Новой Англии, который может включать в себя омаров и креветок, оба из ракообразных, смешивает морепродукты и другие ингредиенты, характерные для этого региона, в вкусную и сытную еду. The Food Network предложила восхитительную версию рецепта от уважаемого шеф-повара Ины Гартен.
- Морские гребешки — известное и популярное ресторанное блюдо. Рассмотрите этот рецепт и дополнительные рекомендации по приготовлению гребешков от Serious Eats.
- Жареные моллюски популярны в прибрежных лачугах с морепродуктами, где их много. В Leite’s Culinaria есть рецепт воспроизведения этого вкуса.
Ракообразные
Крабы, омары, раки и креветки относятся к наиболее часто потребляемым ракообразным. Все эти животные живут рядом с водой или в воде и характеризуются твердыми панцирями, в основном мягкими внутренностями и ветвящимися конечностями, такими как клешни омара или краба.
Ракообразные также выигрывают от ряда проверенных препаратов и презентаций.Вот лишь некоторые из них:
Брюхоногие моллюски
Улитки и слизни — общие названия представителей класса брюхоногих. Эти мягкие, обычно маленькие существа могут иметь или не иметь панцирь и могут жить на суше или в воде в зависимости от вида. В то время как большинство видов слизней только появляются в качестве возможного регулярного источника пищи, улитки уже давно служат деликатесом. Вот несколько рецептов, позволяющих максимально использовать брюхоногих моллюсков:
- Этот рецепт жареных морских улиток от Cooking Channel прост в приготовлении и понравится многим.
- Улитки в скорлупе с маслом с травами — проверенное временем блюдо, которое требует более тщательного приготовления, но в итоге окупается. Рассмотрим этот рецепт от Food Network и частного шеф-повара Джесси Брюна.
Богатство, численность и пространственная неоднородность брюхоногих моллюсков и двустворчатых моллюсков в коралловых экосистемах мексиканской тропической части Тихого океана | Журнал изучения моллюсков
Получить помощь с доступом
Институциональный доступ
Доступ к контенту с ограниченным доступом в Oxford Academic часто предоставляется посредством институциональных подписок и покупок.Если вы являетесь членом учреждения с активной учетной записью, вы можете получить доступ к контенту следующими способами:
Доступ на основе IP
Как правило, доступ предоставляется через институциональную сеть к диапазону IP-адресов. Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с проверкой подлинности IP.
Войдите через свое учреждение
Выберите этот вариант, чтобы получить удаленный доступ за пределами вашего учреждения.
Технология Shibboleth/Open Athens используется для обеспечения единого входа между веб-сайтом вашего учебного заведения и Oxford Academic.
- Щелкните Войти через свое учреждение.
- Выберите свое учреждение из предоставленного списка, после чего вы перейдете на веб-сайт вашего учреждения для входа.
- При посещении сайта учреждения используйте учетные данные, предоставленные вашим учреждением.Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если вашего учреждения нет в списке или вы не можете войти на веб-сайт своего учреждения, обратитесь к своему библиотекарю или администратору.
Вход с помощью читательского билета
Введите номер своего читательского билета, чтобы войти в систему. Если вы не можете войти в систему, обратитесь к своему библиотекарю.
Члены общества
Многие общества предлагают своим членам доступ к своим журналам с помощью единого входа между веб-сайтом общества и Oxford Academic. Из журнала Oxford Academic:
- Щелкните Войти через сайт сообщества.
- При посещении сайта общества используйте учетные данные, предоставленные этим обществом. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если у вас нет учетной записи сообщества или вы забыли свое имя пользователя или пароль, обратитесь в свое общество.
Некоторые общества используют личные аккаунты Oxford Academic для своих членов.
Личный кабинет
Личную учетную запись можно использовать для получения оповещений по электронной почте, сохранения результатов поиска, покупки контента и активации подписок.
Некоторые общества используют личные учетные записи Oxford Academic для предоставления доступа своим членам.
Институциональная администрация
Для библиотекарей и администраторов ваша личная учетная запись также предоставляет доступ к управлению институциональной учетной записью. Здесь вы найдете параметры для просмотра и активации подписок, управления институциональными настройками и параметрами доступа, доступа к статистике использования и т. д.
Просмотр ваших зарегистрированных учетных записей
Вы можете одновременно войти в свою личную учетную запись и учетную запись своего учреждения.Щелкните значок учетной записи в левом верхнем углу, чтобы просмотреть учетные записи, в которые вы вошли, и получить доступ к функциям управления учетной записью.
Выполнен вход, но нет доступа к содержимому
Oxford Academic предлагает широкий ассортимент продукции. Подписка учреждения может не распространяться на контент, к которому вы пытаетесь получить доступ. Если вы считаете, что у вас должен быть доступ к этому контенту, обратитесь к своему библиотекарю.
Двустворчатые моллюски и брюхоногие моллюски природного парка Tubbataha Reefs, Филиппины
Эбботт, Р.Т. и С.П. Танец. 2000. Сборник ракушек. Издательство Одиссей, США. 411 стр.
Обри, У. 1999. Nuove Terebre e Antichi Versi (Новые теребры и древние стихи) . L’informatore Piceno Ed., Ancona, 47 стр.
Batomalaque, G.A., B.G.P. Арсе, М.Б.М. Эрнандес и I.K.C. Фонтанилья. 2010. Исследование и пространственное распределение береговой малакофауны на острове Гранде , залив Субик. Philippine Journal of Science 139 (2): 149-159 (http://philjournalsci.dost.gov.ph/vol139no2/volume_139_No2.html).
СИТЕС. 2014. Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения. Приложения I, II и III . Доступно на http://www.cites.org. Захвачено 6 августа 2014 г.
Долороза, Р. Г., С. Ф. Коналес и Н. А. Бандал. 2011. Первоначальная оценка запаса Terebra maculata (Gastropoda: Terebridae) в природном парке рифов Туббатаха , Палаван, Филиппины. The Philippine Scientist 48: 85–99 (http://www.philjol.info/philjol/index.php/PSCI/article/view/2482/2125).
Долороса, Р.Г., С.Ф. Коналес и Н.А. Бандал. 2013. Статус рогатого шлема Cassis cornuta в природном парке рифов Туббатаха и его торговля в городе Пуэрто-Принсеса, Филиппины. Atoll Research Bulletin 595:1−17 (doi: 10.5479/si.00775630.595).
Долороса, Р.Г. и J.B.S. Джонтила. 2012. Заметки об распространенных макробентосных рифовых беспозвоночных в природном парке Туббатаха Рифс, Филиппины. Science Diliman 24: 1–11 (http://journals.upd.edu.ph/index.php/sciencediliman/article/viewFile/3733/3434).
Долороса, Р.Г. и С. Шоппе. 2005. Фокусные бентические моллюски (Mollusca: Bivalvia и Gastropoda) на выбранных участках в Национальном морском парке рифов Туббатаха, Палаван, Филиппины. Science Diliman 17: 1–10 (http://journals.upd.edu.ph/index.php/sciencediliman/article/viewFile/1/11).
Дыгико, М.П. 2006. Рифы Туббатаха, морская охраняемая территория, которая работает, тематическое исследование на Филиппинах .WWF-Филиппины, Манила. 22 стр. (http://wwf.org.ph/wwf3/downloads/publications/TubbatahaCaseStudy.pdf).
Эстасьон, Дж., В. Палаганас, Р. Перес и М. Алава. 1993. Бентические характеристики островов и рифов в море Сулу, Филиппины. Журнал Силлимана 36: 15–44.
Лаурета Л.В. 2008. Сборник экономически важных морских ракушек в Панай, Филиппины . Издательство Филиппинского университета, Кесон-Сити. 147 стр.
Поппе, Т.Г. 2008а. Филиппинские морские моллюски. Том. 1 (Gastropoda — Часть 1), Hackenheim: ConchBooks. 759 стр.
Поппе, Т.Г. 2008б. Филиппинские морские моллюски. Том. 2 (Gastropoda — Часть 2), Hackenheim: ConchBooks. 848 стр.
Поппе, Т.Г. 2010а. Филиппинские морские моллюски. Том. 3 (брюхоногие — часть 3 и двустворчатые моллюски, часть 1), Hackenheim: ConchBooks. 665 стр.
Поппе, Т.Г. 2010а. Филиппинские морские моллюски. Том. 3 (Bivalvia — Part 2 и Scapphopoda, Polyplacophora, Cephalopdoa и Addenda), Hackenheim: ConchBooks.675 стр.
Richer de Forges, B., S.H. Тан, П. Буше, Нг, П.к.Л. и Н. Сагуил. 2009 г. Панглао 2005 г. — Исследование глубоководной бентической фауны моря Бохоль и прилегающих вод. The Raffles Bulletin of Zoology, Supplement 20: 21–38 (http://rmbr.nus.edu.sg/rbz/biblio/s20/s20rbz021-038.pdf).
Сонгко, А.М. 2012. Полевое руководство по основным хищникам природного парка рифов Туббатаха, Кагаянсилло, Палаван, Филиппины . Пуэрто-Принсеса: Фонд Pilipinas Shell.42 стр.
Спрингстин Ф.Дж. и Ф.М. Леобрера. 1986. Панцири Филиппин . Манила: Музей ракушек Карфель. 377 стр.
Террин, Ю. 2007. Путеводитель по современным теребридам (Mollusca: Neogastropoda) для коллекционеров. Хакенхайм: ConchBooks. 57 стр.
ТМО. 2013. Природный парк рифов Туббатаха: Годовой отчет по исследованиям и мониторингу. Офис управления Туббатаха, Пуэрто-Принсеса. 71 стр.
WORMS. 2014. Всемирный регистр морских видов. Доступен на http://www.marinespecies.org. Снято 8 августа 2014 г.
Yamaguchi, M. 1996. Сообщества моллюсков на мелководье рифа Туббатаха, Республика Филиппины; стр. 61–71, в: Департамент окружающей среды и природных ресурсов (DENR) и Центр морских парков Японии (MPCJ). Отчет о проекте исследования ресурсов и сохранения национального морского парка рифов Туббатаха. 200 стр.
Мэриленд Моллюски
МОЛЛЮСКИ МЭРИЛЕНД
В отряде Mollusca зарегистрировано девять классов: семь существующих в настоящее время (два вымерших).В Мэриленде встречаются три класса: двустворчатые, головоногие и брюхоногие.
ДВУСТВОРЧАТЫЕ (класс Bivalvia)
Несмотря на свое название, состоящее из двух раковин, соединенных шарниром, двустворчатые моллюски не единственные существа с этой чертой. Например, семейство Juliidae внутри класса Gastropoda также имеет двойную раковину. Однако двустворчатые моллюски — единственный класс с этой чертой, обнаруженный в Мэриленде. Как и большинство моллюсков, двустворчатые моллюски приносят большую пользу окружающей среде и обычно встречаются в ручьях или реках с песчаным (или каменистым) и илистым дном на участках с медленным течением.В Мэриленде двустворчатые моллюски также исторически предпочитали Чесапикский залив, что привело к большому рынку морепродуктов и аквакультуры.
Улитки на моллюске, Национальный прибрежный туристический центр острова Ассатиг, 11800 Marsh View Lane, Берлин, Мэриленд, апрель 2019 г. Фото Сары А. Хэнкс.
Ковчег
Ковчег — это моллюски. У них белые, иногда коричневые, ребристые раковины с отверстием для еды, ножка, которая закрепляет их на месте, и прямая линия шарнира.Живя в соленой воде, они зарываются в осадок и питаются детритом и планктоном. Их название происходит от мимолетного сходства с кораблем.
- Кровавые ковчеги (Anadara ovalis) имеют коричневый войлочный слой на панцире, который может действовать как камуфляж. У них красная кровь, отсюда и их название. Они живут на мелководье.
Оболочки тяжеловесных ковчегов (Noetia ponderosa) имеют сердцевидную форму сбоку и, как и кровавые ковчеги, покрыты коричневым войлочным слоем.Они могут вырасти до 2,5 дюймов в длину.
Поперечные ковчеги (Anadara transversa) вырастают примерно до 1 дюйма в длину. Вместо того, чтобы рыть норы, они могут жить среди трав.
Моллюск азиатский (Corbicula fluminea)
Азиатский моллюск имеет внешнюю оболочку желто-зеленого цвета с белыми пятнами, а внутреннюю оболочку пурпурную. Снаружи имеются концентрические кольца. Они вырастают до 1,75 дюйма в длину. Они питаются водорослями, которые фильтруют из воды.Азиатские моллюски живут на илистом или песчаном дне пресноводных озер и ручьев. Азиатский моллюск является инвазивным видом. Также известен как Золотой моллюск .
Моллюск с твердой оболочкой (Mercenaria mercenaria)
У моллюсков с твердой оболочкой раковина белая или серая, иногда с коричневыми пятнами. Темные годичные кольца отходят от середины. Мускулистая стопа позволяет им передвигаться по грязи. Внутренняя часть раковины белая и обычно имеет пурпурное пятно. Они могут вырасти до 4 дюймов в ширину.Моллюски с твердым панцирем являются фильтраторами, поглощающими водоросли и планктон вместе с водой. Живя в илистых отмелях, бухтах и эстуариях, они зарываются в песок. Также известен как Quahog или Round Clam .
Раковина твердого моллюска (Mercenaria mercenaria), побережье национального парка острова Ассатиг, Берлин (округ Вустер), штат Мэриленд, октябрь 2016 г. Фото Сары А. Хэнкс.
Моллюск Macoma
Macoma Моллюски очень маленькие, с тонкой хрупкой раковиной.У них есть одна ножка, которая закрепляет их на месте, и два сифона: один втягивает пищу и воду, а другой выбрасывает отходы. Они торчат из песка или грязи. Моллюски Macoma обитают на мелководье, где они зарываются в песок или ил, перемещаясь при необходимости.
- Балтийские моллюски Macoma (Macoma balthica) имеют раковины овальной или треугольной формы, которые могут быть белыми, розовыми или оранжевыми и могут иметь розовый оттенок. Кольца роста отходят от середины раковины. Они могут вырасти до 1,5 дюймов в длину.
Суженные моллюски Macoma (Macoma tenta) имеют удлиненные раковины жемчужного цвета с изгибом на заднем конце. На гладкой оболочке есть несколько линий роста. Суженные моллюски Macoma вырастают до 0,75 дюймов в длину.
Моллюск с мягкой оболочкой (Mya arenaria)
Моллюск с мягкой оболочкой У моллюсков овальная форма, тонкая и очень хрупкая раковина, за что они и получили свое название. Они белые, но имеют коричневый или серый налет. Одна их нога служит якорем, а два сифона, торчащие из грязи, втягивают и выталкивают морскую воду.Моллюски с мягкой оболочкой могут вырасти до 4 дюймов в длину. Они живут в илистых отмелях, зарываясь на глубину до фута под поверхность.
Мидии, атлантические ребристые (Geukensia demissa)
Атлантические ребристые мидии имеют темные ребристые раковины, которые могут быть коричневыми, черными или зелеными, с внутренней частью белой или синей. Они могут вырасти до 4 дюймов в длину. Они живут в болотах и илистых отмелях, где прикрепляются к твердым поверхностям. Используя свои биссусные нити, они также могут внедряться в осадок.Во время прилива они открывают свои раковины, чтобы набирать воду и выбрасывать отходы.
Мидия, ланцет желтый (Elliptio lanceolata)
ланцет желтый Мидии имеют ярко-желтые раковины с коричневыми участками, а внутренний слой обычно синий. Они могут вырасти более чем на три дюйма. Желтые ланцетные мидии обычно живут в песке пресноводных ручьев и рек. Фильтруя воду, они поедают водоросли. Когда-то использовавшаяся для изготовления пуговиц, плохое качество воды из-за загрязнения и слишком большого количества отложений уменьшило популяцию желтой ланцетной мидии.Желтая ланцетная мидия — вид, находящийся под угрозой исчезновения.
Мидия, зебра (Dreissena polymorpha)
Мидии-зебры имеют D-образную раковину с темными и светлыми полосами, отсюда и их название. Они могут вырасти до 2 дюймов в длину. Они всасывают и выделяют воду, при этом каждая мидия фильтрует до одного галлона воды в день. Как правило, они живут в пресной воде, но могут быть найдены и в солоноватых районах, цепляясь за отложения или твердые поверхности своими биссусными нитями.Мидия-зебра прикрепляется к местным мидиям и моллюскам и строит на них колонии, в конечном итоге убивая их. Мидия-зебра — инвазивный вид.
Устрица восточная (Crassostrea virginica)
Устрицы восточные имеют серо-коричневые раковины и белые внутренности. Они могут вырасти до 8 дюймов в длину. Они живут на морском дне в районах, известных как устричные заросли. Восточные устрицы втягивают воду, питаются водорослями и выделяют отходы. Одна устрица может фильтровать до 50 галлонов воды каждый день.Восточные устрицы могут производить жемчуг, но на сегодняшнем рынке он не имеет ценности. Они важны тем, что их отложения ракушек создают ложа и рифы, образуя среду обитания для других существ. Некоторые отложения из-за непрерывной слоистости могут иметь толщину до 50 футов. На самом деле город Крисфилд в графстве Сомерсет построен на фундаменте из устричных раковин. Также известен как American Oyster .
Гребешок гнедой (Argopecten irradians)
Гнедой гребешок имеет тонкую веерообразную раковину с 13 и 22 ребрами, расходящимися от середины.Их верхняя или левая створки могут быть коричневыми, черными, серыми, желтыми или красными, а нижняя или правая створки более светлого цвета. Внутренняя часть раковины белая и может иметь фиолетовый оттенок. С восемнадцатью парами голубых глаз по краю их панциря они видят движения и хищников. Они могут вырасти до 3,5 дюймов. Из воды они фильтруют планктон. Они живут на дне морских заливов, гаваней и эстуариев, особенно среди взморника, и могут плавать на короткие расстояния.
Корабельный червь Гулда (Bankia gouldi)
Корабельный червь Гулда имеет длинное червеобразное тело с передним концом, покрытым ребристым панцирем, который используется для проникновения в необработанную древесину.У них есть два сифона, которые используются для приема кислорода и пищи, включая древесину и планктон. Известью, которую они выделяют, они выстилают свою нору. Они живут в сваях, доках, корягах и корпусах кораблей. Туннели корабельных червей никогда не пересекаются.
Wedgemussel, Карликовый (Alasmidonta heterodon)
Карликовый Wedgemussels имеют коричневые или оливковые раковины трапециевидной формы. Внутренняя часть синего или белого цвета, а задняя часть радужная. Они вырастают до 1,78 дюйма в длину и живут в чистых пресноводных ручьях и реках.Их личинки прикрепляются к рыбе-хозяину, чтобы развиваться, и отпадают, когда становятся молодыми мидиями, оставляя рыбу невредимой. Их раковины, которые раньше использовались для изготовления пуговиц, были жизненно важны в японской индустрии культивированного жемчуга. Из-за деградации среды обитания и отсутствия рыбы-хозяина численность карликовой мидии сократилась, и она занесена в список находящихся под угрозой исчезновения.
ГОЛОВОЛОДЫ (класс Головоногие)
Головоногие моллюски — водные моллюски с выдающейся головой и набором щупалец.Два вида головоногих, которых часто называют осьминогами или кальмарами, обитают в водах Мэриленда.
Кальмар, атлантический короткий (Lolliguncula brevis)
Краткий Кальмары имеют большую мантию, два больших глаза, клюв и длинные мягкие тела с пятью парами придатков или щупалец, отходящих от головы. Они красно-коричневые или желто-коричневые, но могут менять цвет. Для движения вода проталкивается через воронку, расположенную под головками. Если приближается хищник, воронка сдвинется вперед и очень быстро оттолкнет кратких кальмаров назад, или кальмар испустит чернильное облако.Они могут вырасти до 5 дюймов в длину. Питаются рыбой и ракообразными. Краткие кальмары живут стаями в теплой мелководье.
Кальмар, длинноперый прибрежный (Loligo pealeii)
Прибрежный длинноперый кальмар красного, оранжевого или розового цвета с коричневыми пятнами, но может менять свой цвет. У них большие мантии, клювы, два больших глаза и десять придатков, включая восемь рук и два щупальца. Они могут вырасти до 18 дюймов в длину. В зависимости от своего размера они едят планктон, ракообразных, рыбу и других кальмаров.У континентального шельфа они живут стаями, а в зимние месяцы мигрируют в более теплые воды. Чтобы отвлечь хищников, они брызгают чернилами в воду. Также известен как Longfin Squid и North Atlantic Squid .
брюхоногие (класс брюхоногих)
класс брюхоногих состоит из 611 зарегистрированных семейств улиток и слизней (примерно 202 вымерших семейства). Из классов в царстве Animalia Gastropoda уступает только Insecta по общему количеству зарегистрированных видов.Хотя многие брюхоногие живут в воде, большинство людей сталкиваются с ними на суше, поскольку многие брюхоногие помогают в переработке лесных питательных веществ. Предпочитая прохладную влажную среду, улитки и слизни питаются древесным детритом, листьями и другими биологическими отходами. Брюхоногие помогают процессу разложения и ускоряют повторное введение минералов в почву. Вымерший вид морской улитки, Ecphora gardnerae gardnerae , является окаменелостью штата Мэриленд.
Слизень (класс Gastropoda), Балтимор, Мэриленд, август 2015 г.Фото Сары А. Хэнкс.
Дрель, атлантическая устрица (Urosalpinx cinerea)
Атлантическая устрица Дрель – это улитки. Их заостренные раковины овальной формы могут быть коричневыми, серыми, желтыми или фиолетовыми с вертикальными ребрами и множеством завитков. Они вырастают до 1 дюйма в длину. Они просверливают раковины устриц и поедают их, отсюда и их название, но они также едят и других моллюсков. Атлантические устрицы обитают на устричных рифах и среди трав на мелководье.
Грязевые улитки, Новая Зеландия (Potamopyrgus antipodarum)
Новая Зеландия Грязевые улитки вырастают до 12 миллиметров в длину в своих родных водах, но обычно имеют длину от 4 до 6 миллиметров в других местах. Их длинные желобчатые раковины правые (отверстия справа), серые или коричневые с 7 или 8 оборотами. Они живут как в пресных, так и в солоноватых водах, питаются водорослями, детритом и листьями. Обладая способностью к бесполому размножению, они имеют высокую скорость размножения и очень быстро колонизируют территорию.
Инвазивный вид, новозеландская грязевая улитка была впервые обнаружена в штате Мэриленд в реке Порох в сентябре 2017 года.
Барвинок болотный (Littorina irrorata)
Барвинок болотный — улитки. У них желобчатая раковина серо-белого цвета с коричнево-красными пятнами и от восьми до десяти оборотов на раковине. Они вырастают до 1 дюйма в длину. Обитая в солончаках и заболоченных местах, они поедают водоросли и грибы, растущие на болотной траве.
Морской слизень
Морской слизень напоминает садового слизняка.Они лишены панциря, имеют щупальца на голове и различные наросты на мягком теле. Они могут вырасти до 1,5 дюймов в длину. Морские слизни живут на мелководье, включая болота и реки, а также в твердых сооружениях, таких как рифы и буи.
-
Сакоглоссаны зеленые. Они едят водоросли. В Чесапикском заливе обитают четыре вида: морской слизень-крестоносец (Hermaea Crusata) , сумеречный морской слизень (Stiliger fuscatus) , изумрудный морской слизень (Elysia chlorotica) и морской слизень-кошечка (Elysia catula) .
Голожаберники ярко окрашены и сливаются с окружающей средой. Они едят гидроидов и других мелких существ. В Чесапикском заливе обитает четыре вида: моллюск голожаберный (Doridella obscura) , гребенчатый голожаберный (Polycerella conyma) , жесткоспинный голожаберный (Dorisverrucosa) и полосатый голожаберный (Cratena 162) 90.
