Классный урок на «Радио России – Тамбов», эфир 28 апреля 2020 года
Автор ГТРК «ТАМБОВ» На чтение 16 мин. Просмотров 62 Опубликовано
Урок биологии для десятиклассников на канале «Радио России» в рамках совместного проекта ГТРК «Тамбов», Управления образования и науки Тамбовской области и ТГУ имени Г.Р. Державина ведут сегодня педагоги университета и 13-го Центра образования Тамбова. Татьяна Николаевна Киселева рассказывает о цитоплазматической наследственности и генотипическом определении пола. А Инна Вячеславовна Баженова — о взаимодействии неаллейных генов.
БИОЛОГИЯ ДЛЯ ДЕСЯТИКЛАССНИКОВ
Предмет: Биология (для 10 класса)
Педагог: Инна Вячеславовна Баженова – учитель биологии МАОУ «Центр образования №13 им. Н.А. Кузнецова»
Тема: Взаимодействие неаллейных генов.
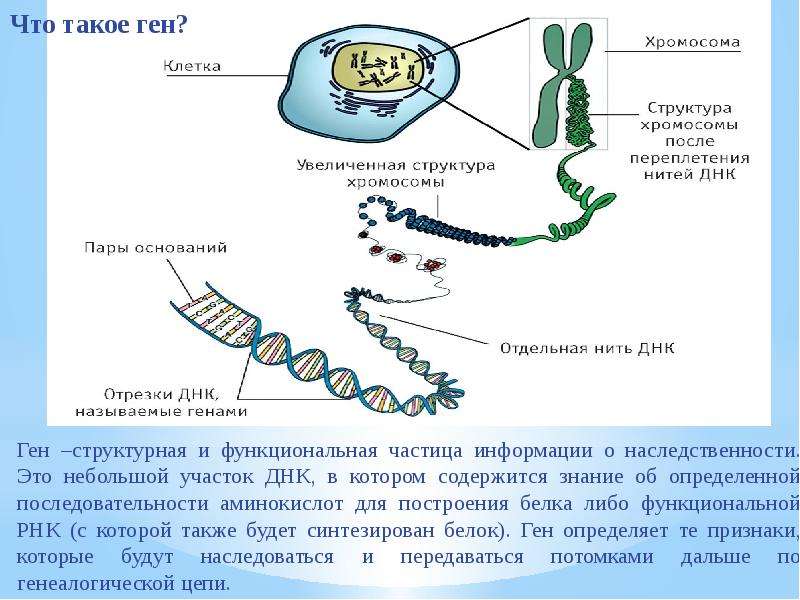
Генотип – это не простая совокупность генов, а система сложного взаимодействия между ними Взаимодействие генов — это совместное действие нескольких генов, в результате которого появляется признак, которого нет у родителей, или усиливается проявление уже имеющегося признака. Первый случай неаллельного взаимодействия был описан в качестве примера отклонения от законов Менделя английскими учеными У. Бетсоном и Р. Пеннетом в 1904 г. при изучении наследования формы гребня у кур.
Для объяснения результатов взаимодействия генов важно понимать механизм формирования признаков.
Ген — это участок ДНК, в котором закодирована информация об одном белке. В простейшем случае формирование признака может происходить в результате действия одного белка, синтез которого определяется одним геном:
— но обычно признак формируется в результате сложных биохимических процессов. В клетке происходит взаимодействие между белками-ферментами, синтез которых определяется генами, или между веществами, которые образуются под влиянием этих ферментов.
Возможны следующие типы проявления генов в фенотипе:
Взаимодействие генов — это совместное действие нескольких генов, в результате которого появляется признак, которого нет у родителей, или усиливается проявление уже имеющегося признака. Первый случай неаллельного взаимодействия был описан в качестве примера отклонения от законов Менделя английскими учеными У. Бетсоном и Р. Пеннетом в 1904 г. при изучении наследования формы гребня у кур.
Для объяснения результатов взаимодействия генов важно понимать механизм формирования признаков.
Ген — это участок ДНК, в котором закодирована информация об одном белке. В простейшем случае формирование признака может происходить в результате действия одного белка, синтез которого определяется одним геном:
— но обычно признак формируется в результате сложных биохимических процессов. В клетке происходит взаимодействие между белками-ферментами, синтез которых определяется генами, или между веществами, которые образуются под влиянием этих ферментов.
Возможны следующие типы проявления генов в фенотипе:
- один признак формируется в результате взаимодействия нескольких белков, синтез которых определяется несколькими генами:
- один ген определяет синтез белка, который влияет на формирование нескольких признаков:
 Возможно взаимодействие как между аллельными генами, так и между неаллельными.
Неаллельные гены — это гены, расположенные в различных участках (локусах) хромосом и кодирующие неодинаковые белки. Формы взаимодействия неаллельных генов:
Возможно взаимодействие как между аллельными генами, так и между неаллельными.
Неаллельные гены — это гены, расположенные в различных участках (локусах) хромосом и кодирующие неодинаковые белки. Формы взаимодействия неаллельных генов:
- комплементарность;
- эпистаз;
- полимерия.
 При скрещивании двух растений с белыми цветками у гибридов F1 цветки оказались пурпурными. При самоопылении растений из F1 в F2 наблюдалось расщепление растений по окраске цветков в отношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые у 7/16.
Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментом антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента — пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором — есть пропигмент, но нет фермента, переводящего пропигмент в пигмент.
Проведем скрещивание двух растений душистого горошка с белыми цветками. В одной из линий, АAbb, есть доминантный аллель А, а у другой, ааBB, есть доминантный аллель В. У растений, имеющих доминантные гены А и В одновременно, есть и пропигмент (обеспечиваемый А), и фермент (обеспечиваемый В), необходимые для образования пурпурного пигмента.
При скрещивании двух растений с белыми цветками у гибридов F1 цветки оказались пурпурными. При самоопылении растений из F1 в F2 наблюдалось расщепление растений по окраске цветков в отношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые у 7/16.
Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментом антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента — пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором — есть пропигмент, но нет фермента, переводящего пропигмент в пигмент.
Проведем скрещивание двух растений душистого горошка с белыми цветками. В одной из линий, АAbb, есть доминантный аллель А, а у другой, ааBB, есть доминантный аллель В. У растений, имеющих доминантные гены А и В одновременно, есть и пропигмент (обеспечиваемый А), и фермент (обеспечиваемый В), необходимые для образования пурпурного пигмента.
| гаметы | AB | Ab | aB | ab |
| AB | AABB | AABb | AaBB | AaBb |
| Ab | AABb | AAbb | AaBb | Aabb |
| aB | AaBB | AaBb | aaBB | aaBb |
| ab | AaBb | Aabb | aaBb | aabb |
 В этом случае расщепление будет 9:6:1. Например, это наблюдается при наследовании формы плодов у тыквы:
Возможно и возникновение четырёх фенотипов. Например, при скрещивании кур с различной формой гребня.
А_В_ — ореховидный
А_bb — гороховидный
aaB_ — розовидный
aabb — простой (листовидный)
Чтобы успешно решать задачи, надо помнить, что такое явление, как комплементарность, в принципе, возможно, и быть внимательным при написании генотипов особей и их гамет.
Пример решения задачи
Наследование слуха у человека определяется двумя доминантными генами из разных аллельных пар, один из которых детерминирует развитие слухового нерва, а другой – улитки. Определить вероятность рождения глухих детей, если оба родителя глухие, но по разным генетическим причинам (у одного отсутствует слуховой нерв, у другого улитка). По генотипу оба родителя являются дигомозиготными.
Здесь проявляется I закон Менделя — закон единообразия гибридов первого поколения. Возможен только один вариант генотипа ребенка от такого брака.
В этом случае расщепление будет 9:6:1. Например, это наблюдается при наследовании формы плодов у тыквы:
Возможно и возникновение четырёх фенотипов. Например, при скрещивании кур с различной формой гребня.
А_В_ — ореховидный
А_bb — гороховидный
aaB_ — розовидный
aabb — простой (листовидный)
Чтобы успешно решать задачи, надо помнить, что такое явление, как комплементарность, в принципе, возможно, и быть внимательным при написании генотипов особей и их гамет.
Пример решения задачи
Наследование слуха у человека определяется двумя доминантными генами из разных аллельных пар, один из которых детерминирует развитие слухового нерва, а другой – улитки. Определить вероятность рождения глухих детей, если оба родителя глухие, но по разным генетическим причинам (у одного отсутствует слуховой нерв, у другого улитка). По генотипу оба родителя являются дигомозиготными.
Здесь проявляется I закон Менделя — закон единообразия гибридов первого поколения. Возможен только один вариант генотипа ребенка от такого брака.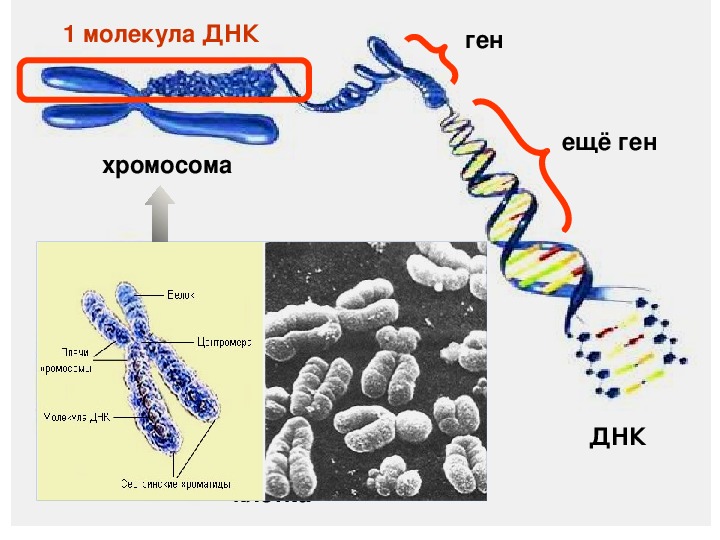
 Наследование окраски шерсти у собак (пример доминантного эпистаза):
A — черная окраска, а — коричневая, I — подавляет окраску, i — не подавляет.
Рецессивный эпистаз
Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена в гомозиготном состоянии аллелей гипостатичного гена (ii > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа ii (например, обе формы неокрашены).
Рецессивный эпистаз проявляется при наследовании окраски шерсти у домовых мышей.
Наследование окраски шерсти у собак (пример доминантного эпистаза):
A — черная окраска, а — коричневая, I — подавляет окраску, i — не подавляет.
Рецессивный эпистаз
Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена в гомозиготном состоянии аллелей гипостатичного гена (ii > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа ii (например, обе формы неокрашены).
Рецессивный эпистаз проявляется при наследовании окраски шерсти у домовых мышей.
 Доктор Бхенде обнаружил, что у людей рецессивных по гену h (hh) на поверхности эритроцитов не синтезируются агглютиногены — в результате этого они могут быть универсальными донорами.
Говоря проще о Бомбейском феномене: у людей с генотипом hh всегда обнаруживается первая группа крови при любом генотипе — I
AIA, IBIB, IAIB. Ген h подавляет гены IA и IB — на поверхности эритроцитов не образуются агглютиногены A и B.
Пример решения задачи
«Редкий рецессивный ген (h) в гомозиготном состоянии обладает эпистатическим действием по отношению к генам IA, IB и изменяет их действие до I группы крови (бомбейский феномен). Определите возможные группы крови у детей, если у мужа II гомозиготная, у жены IV и оба родителя гетерозиготны по эпистатическому гену».
Вероятность рождения детей с i(0) группой крови в данном случае равна 2/8, или 1/4 (25%). Генотипами, у которых будет i(0) группа крови являются: IAIAhh и IAIBhh.
Доктор Бхенде обнаружил, что у людей рецессивных по гену h (hh) на поверхности эритроцитов не синтезируются агглютиногены — в результате этого они могут быть универсальными донорами.
Говоря проще о Бомбейском феномене: у людей с генотипом hh всегда обнаруживается первая группа крови при любом генотипе — I
AIA, IBIB, IAIB. Ген h подавляет гены IA и IB — на поверхности эритроцитов не образуются агглютиногены A и B.
Пример решения задачи
«Редкий рецессивный ген (h) в гомозиготном состоянии обладает эпистатическим действием по отношению к генам IA, IB и изменяет их действие до I группы крови (бомбейский феномен). Определите возможные группы крови у детей, если у мужа II гомозиготная, у жены IV и оба родителя гетерозиготны по эпистатическому гену».
Вероятность рождения детей с i(0) группой крови в данном случае равна 2/8, или 1/4 (25%). Генотипами, у которых будет i(0) группа крови являются: IAIAhh и IAIBhh. Эпистатический рецессивный ген hh в гомозиготном состоянии всегда приводит к i(0) группе крови.
Эпистатический рецессивный ген hh в гомозиготном состоянии всегда приводит к i(0) группе крови.
 Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак.
У человека полимерное действие генов заложено в наследовании количественных признаков таких как рост, масса тела, интеллектуальные особенности, склонность к повышению артериального давления, устойчивость к инфекционным заболеваниям и других.
Пример решения задачи
«Цвет кожи у мулатов наследуется по типу полимерии. При этом данный признак контролируется 2 аутосомными несцепленными генами. Сын белой женщины и негра женился на белой женщине. Может ли этот ребенок быть темнее своего отца?»
В данном случае полимерия проявляется в том, что чем больше доминантных генов в генотипе (A и B), тем более темный цвет кожи имеет человек. Это правило мы и применим для решения.
В результате первого брака (вспоминаем закон единообразия Менделя) получается AaBb — средний мулат. По условиям задачи он берет в жены белую женщину aabb. Очевидно, что в этой семье ребенок не может быть темнее своего отца: дети могут быть или же средними мулатами (AaBb), как отец, либо белыми, как мать (aabb).
Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак.
У человека полимерное действие генов заложено в наследовании количественных признаков таких как рост, масса тела, интеллектуальные особенности, склонность к повышению артериального давления, устойчивость к инфекционным заболеваниям и других.
Пример решения задачи
«Цвет кожи у мулатов наследуется по типу полимерии. При этом данный признак контролируется 2 аутосомными несцепленными генами. Сын белой женщины и негра женился на белой женщине. Может ли этот ребенок быть темнее своего отца?»
В данном случае полимерия проявляется в том, что чем больше доминантных генов в генотипе (A и B), тем более темный цвет кожи имеет человек. Это правило мы и применим для решения.
В результате первого брака (вспоминаем закон единообразия Менделя) получается AaBb — средний мулат. По условиям задачи он берет в жены белую женщину aabb. Очевидно, что в этой семье ребенок не может быть темнее своего отца: дети могут быть или же средними мулатами (AaBb), как отец, либо белыми, как мать (aabb). При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1 для двух генов, 63:1 для трёх генов и т.д.
К взаимодействию неаллельных генов относят также явление плейотропии — множественного действия гена, влияния его на развитие нескольких признаков. Плейотропное действие генов является результатом серьезного нарушения обмена веществ, обусловленного мутантной структурой данного гена.
Летальный эффект при переходе в гомозиготное состояние характерен для многих плейотропных мутаций. Так, у лисиц доминантные гены, контролирующие платиновую и беломордую окраски меха, не оказывающие летального действия в гетерозиготе, вызывают гибель гомозиготных зародышей на ранней стадии развития. Аналогичная ситуация имеет место при наследовании серой окраски шерсти у овец породы ширази и недоразвития чешуи у зеркального карпа.
При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1 для двух генов, 63:1 для трёх генов и т.д.
К взаимодействию неаллельных генов относят также явление плейотропии — множественного действия гена, влияния его на развитие нескольких признаков. Плейотропное действие генов является результатом серьезного нарушения обмена веществ, обусловленного мутантной структурой данного гена.
Летальный эффект при переходе в гомозиготное состояние характерен для многих плейотропных мутаций. Так, у лисиц доминантные гены, контролирующие платиновую и беломордую окраски меха, не оказывающие летального действия в гетерозиготе, вызывают гибель гомозиготных зародышей на ранней стадии развития. Аналогичная ситуация имеет место при наследовании серой окраски шерсти у овец породы ширази и недоразвития чешуи у зеркального карпа. Летальный эффект мутаций приводит к тому, что животные этих пород могут быть только гетерозиготными и при внутрипородных скрещиваниях дают расщепление в соотношении 2 мутанта : 1 норма.
Схема наследования платиновой окраски у лис
F1
Летальный эффект мутаций приводит к тому, что животные этих пород могут быть только гетерозиготными и при внутрипородных скрещиваниях дают расщепление в соотношении 2 мутанта : 1 норма.
Схема наследования платиновой окраски у лис
F1| A | a | |
| A | AA погибают | Aa платин. |
| a | Aa платин. | aa черн. |
 Такое нарушение приводит к тому, что у человека формируются вывих хрусталика глаза, пороки клапана сердца, длинные и тонкие пальцы, пороки развития сосудов и частые вывихи суставов.
Синдром Марфана назван по имени педиатра, который наблюдал девочку с этим заболеванием на протяжении 20 лет. Имеется много интересных фактов о людях, имеющих характерные признаки патологии. Первая манекенщица (Лесли Хорнби – «Твигги»), которая была прототипом для всех чрезмерно худых моделей, болела синдромом Марфана. Наиболее известные личности, о которых есть подобные сведения: президент А. Линкольн, скрипач Н. Паганини, писатель Г. Х. Андерсен, композитор С. Рахманинов.
Такое нарушение приводит к тому, что у человека формируются вывих хрусталика глаза, пороки клапана сердца, длинные и тонкие пальцы, пороки развития сосудов и частые вывихи суставов.
Синдром Марфана назван по имени педиатра, который наблюдал девочку с этим заболеванием на протяжении 20 лет. Имеется много интересных фактов о людях, имеющих характерные признаки патологии. Первая манекенщица (Лесли Хорнби – «Твигги»), которая была прототипом для всех чрезмерно худых моделей, болела синдромом Марфана. Наиболее известные личности, о которых есть подобные сведения: президент А. Линкольн, скрипач Н. Паганини, писатель Г. Х. Андерсен, композитор С. Рахманинов.
Предмет: Биология (для 10 класса)
Педагог: Татьяна Николаевна Киселева – ассистент кафедры довузовской подготовки ТГУ им. Г.Р.Державина
Тема: Цитоплазматическая наследственность. Генотипитическое определение пола.
Цитоплазматическая наследственность.
Генетическое определение пола.
 Пластидная наследственность – выявлена при наследовании пестролистной окраски. Связана с наличием в клетках только окрашенных, только бесцветных или их смеси пластид.
Плазмидная наследственность – связана с генами, расположенными в плазмидах (коротких кольцевых молекул ДНК, находящихся вне нуклеотида бактериальной клетки), обеспечивает наследование устойчивости бактерий к действию лекарственных препаратов благодаря защитным белкам.
Характерной чертой цитоплазматической наследственности является наследование по линии матери. Это связано с тем, что в яйцеклетке и пластид, и митохондрий очень много, а в мужских гаметах этих органоидов практически нет (так как эти клетки практически лишены цитоплазмы). В сперматозоидах присутствуют митохондрии, но они все равно не проникают в яйцеклетку, так как при слиянии гамет в яйцеклетку попадает только ядро сперматозоида, содержащее генетический материал. Таким образом, все митохондрии и пластиды зиготы достаются ей в наследство только от материнского организма.
Пластидная наследственность – выявлена при наследовании пестролистной окраски. Связана с наличием в клетках только окрашенных, только бесцветных или их смеси пластид.
Плазмидная наследственность – связана с генами, расположенными в плазмидах (коротких кольцевых молекул ДНК, находящихся вне нуклеотида бактериальной клетки), обеспечивает наследование устойчивости бактерий к действию лекарственных препаратов благодаря защитным белкам.
Характерной чертой цитоплазматической наследственности является наследование по линии матери. Это связано с тем, что в яйцеклетке и пластид, и митохондрий очень много, а в мужских гаметах этих органоидов практически нет (так как эти клетки практически лишены цитоплазмы). В сперматозоидах присутствуют митохондрии, но они все равно не проникают в яйцеклетку, так как при слиянии гамет в яйцеклетку попадает только ядро сперматозоида, содержащее генетический материал. Таким образом, все митохондрии и пластиды зиготы достаются ей в наследство только от материнского организма. Показано, что хромосомная и нехромосомная наследственность могут взаимодействовать, приводя к сложным случаям наследования. Например, большинство белков митохондрий закодировано в ядерных генах и наследуется по правилам Менделя, а оставшиеся белки кодируются в ДНК самих митохондрий, которые передаются только по материнской линии.
Генетическое определение пола
Мы знаем, что большинство животных и двудомных растений являются раздельнополыми организмами, причем внутри вида количество особей мужского пола приблизительно равно количеству особей женского пола. Другими словами, каждому виду, имеющему четкое деление на мужские и женские особи, свойственно определенное соотношение полов, близкое 1:1.
Проблема происхождения половых отличий, механизм определения пола и поддержание определенного соотношения полов в группах животных организмов важна как для практической, так и для теоретической биологии.
Пол у животных чаще всего определяется в момент оплодотворения – сингамный тип определения пола.
Показано, что хромосомная и нехромосомная наследственность могут взаимодействовать, приводя к сложным случаям наследования. Например, большинство белков митохондрий закодировано в ядерных генах и наследуется по правилам Менделя, а оставшиеся белки кодируются в ДНК самих митохондрий, которые передаются только по материнской линии.
Генетическое определение пола
Мы знаем, что большинство животных и двудомных растений являются раздельнополыми организмами, причем внутри вида количество особей мужского пола приблизительно равно количеству особей женского пола. Другими словами, каждому виду, имеющему четкое деление на мужские и женские особи, свойственно определенное соотношение полов, близкое 1:1.
Проблема происхождения половых отличий, механизм определения пола и поддержание определенного соотношения полов в группах животных организмов важна как для практической, так и для теоретической биологии.
Пол у животных чаще всего определяется в момент оплодотворения – сингамный тип определения пола. Это – генетическое определение пола, так как зависит от баланса хромосом. Довольно часто его называют хромосомным определением пола.
Если определение пола происходит еще до оплодотворения в процессе созревания яйцеклеток, то такое определение пола называют прогамным (т.е. перед оплодотворением, до слияния гамет). Характерно для тли, коловраток…
Если определение пола происходит после оплодотворения, то подобный тип определения пола называют эпигамным (т.е. после слияния гамет).
У ряда многоклеточных организмов определение пола происходит вне связи с оплодотворением. Это партеногенез (пример: дафнии – в зависимости от условий существования образуются потомки мужского или женского пола).
Поскольку у большинства живых организмов пол чаще определяется в момент оплодотворения, то важнейшая роль в генетическом определении пола принадлежит хромосомному набору зиготы. Таким образом, определение пола обычно связывают с наличием одной пары хромосом, по которой отличается женский пол от мужского.
Это – генетическое определение пола, так как зависит от баланса хромосом. Довольно часто его называют хромосомным определением пола.
Если определение пола происходит еще до оплодотворения в процессе созревания яйцеклеток, то такое определение пола называют прогамным (т.е. перед оплодотворением, до слияния гамет). Характерно для тли, коловраток…
Если определение пола происходит после оплодотворения, то подобный тип определения пола называют эпигамным (т.е. после слияния гамет).
У ряда многоклеточных организмов определение пола происходит вне связи с оплодотворением. Это партеногенез (пример: дафнии – в зависимости от условий существования образуются потомки мужского или женского пола).
Поскольку у большинства живых организмов пол чаще определяется в момент оплодотворения, то важнейшая роль в генетическом определении пола принадлежит хромосомному набору зиготы. Таким образом, определение пола обычно связывают с наличием одной пары хромосом, по которой отличается женский пол от мужского. И одинаковые по внешнему виду хромосомы в клетках раздельнополых организмов называют аутосомами, а пару различающихся хромосом, неодинаковых у самца и самки называют половыми хромосомами.
Общее число, размер и форма хромосом – кариотип.
Вспомните например, хромосомный набор человека (кариотип). В наборе хромосом зиготы содержатся парные – гомологичные хромосомы. В женском кариотипе все хромосомы парные, в мужском имеется одна крупная равноплечая непарная хромосома, другая маленькая палочковидная, встречающаяся у мужчин. Таким образом, кариотип женщин содержит 22 пары хромосом — аутосом, одинаковые у мужчин и женщин. И одну пару хромосом, по которой различны оба пола — гетерохромосомы.
Половые хромосомы у женщин одинаковы – X-хромосомы.
Диплоидные (соматические) клетки женского организма содержат 2 X – хромосомы и в процессе овогенеза образуют яйцеклетки, имеющие по одной X – хромосоме.
Пол, образующий гаметы, одинаковые по половой хромосоме, называют гомогаметным.
И одинаковые по внешнему виду хромосомы в клетках раздельнополых организмов называют аутосомами, а пару различающихся хромосом, неодинаковых у самца и самки называют половыми хромосомами.
Общее число, размер и форма хромосом – кариотип.
Вспомните например, хромосомный набор человека (кариотип). В наборе хромосом зиготы содержатся парные – гомологичные хромосомы. В женском кариотипе все хромосомы парные, в мужском имеется одна крупная равноплечая непарная хромосома, другая маленькая палочковидная, встречающаяся у мужчин. Таким образом, кариотип женщин содержит 22 пары хромосом — аутосом, одинаковые у мужчин и женщин. И одну пару хромосом, по которой различны оба пола — гетерохромосомы.
Половые хромосомы у женщин одинаковы – X-хромосомы.
Диплоидные (соматические) клетки женского организма содержат 2 X – хромосомы и в процессе овогенеза образуют яйцеклетки, имеющие по одной X – хромосоме.
Пол, образующий гаметы, одинаковые по половой хромосоме, называют гомогаметным.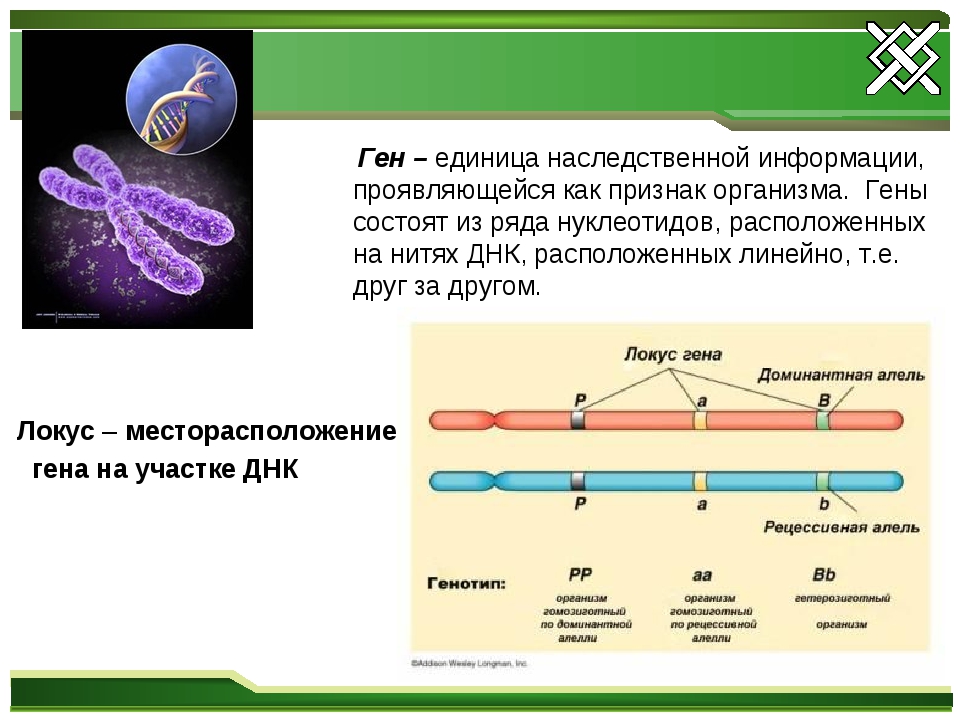 У мужчин в диплоидных клетках имеется 1 X- хромосома и 1 Y – хромосома. При сперматогенезе получаются гаметы 2-х сортов (X и Y).
Пол, который формируют гаметы, неодинаковые по половой хромосоме называют гетерогаметным.
Таким образом, у человека хромосомный набор выглядит так:
Женский: 2×2 + XX
Мужской: 2×2 + XY
Т.е. женский пол – гомогаметен, а мужской – гетерогаметен.
У бабочек, птиц гомогаметным полом является мужской пол, т.е. петух будет иметь XX набор хромосом, а курица XY. Y-хромосома определяет развитие семенников, а в дальнейшем развитие мужских половых признаков.
Подводя итог занятию, благодаря наличию ДНК не только в ядрах, но и в органеллах цитоплазмы живые организмы получают определенное преимущество в процессе эволюции. Дело в том, что ядро и хромосомы отличаются генетически обусловленной высокой устойчивостью к меняющимся условиям окружающей среды. В то же время хлоропласты и митохондрии развиваются до некоторой степени независимо от клеточного деления, непосредственно реагируя на воздействие окружающей среды.
У мужчин в диплоидных клетках имеется 1 X- хромосома и 1 Y – хромосома. При сперматогенезе получаются гаметы 2-х сортов (X и Y).
Пол, который формируют гаметы, неодинаковые по половой хромосоме называют гетерогаметным.
Таким образом, у человека хромосомный набор выглядит так:
Женский: 2×2 + XX
Мужской: 2×2 + XY
Т.е. женский пол – гомогаметен, а мужской – гетерогаметен.
У бабочек, птиц гомогаметным полом является мужской пол, т.е. петух будет иметь XX набор хромосом, а курица XY. Y-хромосома определяет развитие семенников, а в дальнейшем развитие мужских половых признаков.
Подводя итог занятию, благодаря наличию ДНК не только в ядрах, но и в органеллах цитоплазмы живые организмы получают определенное преимущество в процессе эволюции. Дело в том, что ядро и хромосомы отличаются генетически обусловленной высокой устойчивостью к меняющимся условиям окружающей среды. В то же время хлоропласты и митохондрии развиваются до некоторой степени независимо от клеточного деления, непосредственно реагируя на воздействие окружающей среды. Таким образом, они имеют потенциальную возможность обеспечить быстрые реакции организма на изменение внешних условий.
Домашнее задание: решить задачи на сцепленное с полом наследование.
Задача 1. Мужчина с нормальным зрение женился на женщине-дальтонике (рецессивный ген d сцеплен с Х-хромосомой). Определите генотипы родителей, соотношение фенотипов и генотипов в потомстве.
Задача 2. Мужчина, больной гемофилией вступает в брак с нормальной женщиной, отец которой страдал гемофилией. Определите вероятность рождения здоровых детей?
Задача 3. У кур встречается сцепленный с полом летальный ген (а), вызывающий гибель эмбрионов, гетерозиготы по этому гену жизнеспособны. Скрестили нормальную курицу с гетерозиготным по этому гену петухом (у птиц гетерогаметный пол — женский). Составьте схему решения задачи, определите генотипы родителей, пол и генотип возможного потомства и вероятность вылупления курочек от общего числа жизнеспособного потомства.
Задача 4. У человека наследование альбинизма не сцеплено с полом (А – наличие меланина в клетках кожи, а – отсутствие меланина в клетках кожи – альбинизм), а гемофилии – сцеплено с полом (XН – нормальная свёртываемость крови, Xh – гемофилия).
Таким образом, они имеют потенциальную возможность обеспечить быстрые реакции организма на изменение внешних условий.
Домашнее задание: решить задачи на сцепленное с полом наследование.
Задача 1. Мужчина с нормальным зрение женился на женщине-дальтонике (рецессивный ген d сцеплен с Х-хромосомой). Определите генотипы родителей, соотношение фенотипов и генотипов в потомстве.
Задача 2. Мужчина, больной гемофилией вступает в брак с нормальной женщиной, отец которой страдал гемофилией. Определите вероятность рождения здоровых детей?
Задача 3. У кур встречается сцепленный с полом летальный ген (а), вызывающий гибель эмбрионов, гетерозиготы по этому гену жизнеспособны. Скрестили нормальную курицу с гетерозиготным по этому гену петухом (у птиц гетерогаметный пол — женский). Составьте схему решения задачи, определите генотипы родителей, пол и генотип возможного потомства и вероятность вылупления курочек от общего числа жизнеспособного потомства.
Задача 4. У человека наследование альбинизма не сцеплено с полом (А – наличие меланина в клетках кожи, а – отсутствие меланина в клетках кожи – альбинизм), а гемофилии – сцеплено с полом (XН – нормальная свёртываемость крови, Xh – гемофилия). Определите генотипы родителей, а также возможные генотипы, пол и фенотипы детей от брака дигомозиготной нормальной по обеим аллелям женщины и мужчины альбиноса, больного гемофилией. Составьте схему решения задачи.
Задача 5. Глухота — аутосомный признак; дальтонизм – признак, сцепленный с полом. В браке здоровых родителей родился ребёнок глухой дальтоник. Составьте схему решения задачи. Определите генотипы родителей и ребёнка, его пол, генотипы и фенотипы возможного потомства, вероятность рождения детей с обеими аномалиями. Какие законы наследственности проявляются в данном случае? Ответ обоснуйте.
Определите генотипы родителей, а также возможные генотипы, пол и фенотипы детей от брака дигомозиготной нормальной по обеим аллелям женщины и мужчины альбиноса, больного гемофилией. Составьте схему решения задачи.
Задача 5. Глухота — аутосомный признак; дальтонизм – признак, сцепленный с полом. В браке здоровых родителей родился ребёнок глухой дальтоник. Составьте схему решения задачи. Определите генотипы родителей и ребёнка, его пол, генотипы и фенотипы возможного потомства, вероятность рождения детей с обеими аномалиями. Какие законы наследственности проявляются в данном случае? Ответ обоснуйте. Как управлять ДНК – Наука – Коммерсантъ
Наследственная информация любого организма, как известно, хранится в молекуле ДНК. У человека последовательность из четырех букв (оснований ДНК) общей длиной около 3 млрд — геном — несет полную информацию об организме. На основе этой информации он развивается и умеет приспосабливаться к изменяющимся условиям окружающей среды.
Казалось бы, если мы получим доступ к информации, хранящейся в геноме, то сможем узнать все о нашей биологической природе. Неудивительно, что генетики сфокусировали значительные усилия на задаче прочтения ДНК. Благодаря революции в технологиях работы с ДНК конца XX века и созданию новых технологий чтения ДНК в начале XXI века исследователи наконец получили доступ к наследственной информации: в настоящее время любой человек за относительно небольшую сумму может прочитать свою ДНК. Что же он узнает о себе? К сожалению, очень немногое. Проблема в том, что из всего массива генетического текста мы точно понимаем смысл только той части, которая кодирует белки (кодирующей части генов). А она составляет в геноме только несколько процентов.
Исследования последних 10–15 лет указывают на то, что значительная доля генома с пока неизвестной функцией, скорее всего, нужна для регуляции работы той самой небольшой доли генов. Таким образом, на десятки тысяч генов человека приходятся сотни тысяч (или, по другим оценкам, миллионы) «управленцев», которые указывают, где и когда нужно работать каждому отдельному гену. Это очень важное явление, которое называется дифференциальной экспрессией (активностью) генов. Дело в том, что все клетки организма содержат одинаковый — совершенно полный — набор генетической информации. Однако клетки очень сильно отличаются, и причина этого в том, что в разных клетках работают разные наборы генов с разной интенсивностью, что в результате и определяет, как устроена клетка и какие функции она выполняет.
Это очень важное явление, которое называется дифференциальной экспрессией (активностью) генов. Дело в том, что все клетки организма содержат одинаковый — совершенно полный — набор генетической информации. Однако клетки очень сильно отличаются, и причина этого в том, что в разных клетках работают разные наборы генов с разной интенсивностью, что в результате и определяет, как устроена клетка и какие функции она выполняет.
Для того чтобы читать информацию, регулирующую работу генов, в клетке имеется большой молекулярный аппарат транскрипции. Транскрипция — первый этап реализации наследственной информации: она переписывается с ДНК на небольшие копии — мРНК. Именно этот этап является ключевым с точки зрения регуляции активности (экспрессии) генов. Специфичные факторы транскрипции способны узнавать регуляторные элементы генома, связываться с ними и затем передавать посредством взаимодействия с другими белками сигнал на нужный ген.
Аппарат транскрипции — это одна из наиболее сложных молекулярных систем клетки: у человека имеется несколько тысяч таких факторов. Их слаженная работа обеспечивает работу всего генома. Факторы стоят на разных уровнях иерархии и выполняют очень разные молекулярные функции. Имеются факторы, которые специфически узнают определенную последовательность букв в ДНК. Другие факторы упаковывают или размечают молекулу ДНК и могут, например, переводить ее в нерабочее, «архивное», состояние. Существуют промежуточные факторы-посредники и т. д. Факторы транскрипции не работают поодиночке, а согласуют работу друг с другом: обычно для запуска одного гена требуется совместное присутствие нескольких десятков (до сотни) факторов.
Их слаженная работа обеспечивает работу всего генома. Факторы стоят на разных уровнях иерархии и выполняют очень разные молекулярные функции. Имеются факторы, которые специфически узнают определенную последовательность букв в ДНК. Другие факторы упаковывают или размечают молекулу ДНК и могут, например, переводить ее в нерабочее, «архивное», состояние. Существуют промежуточные факторы-посредники и т. д. Факторы транскрипции не работают поодиночке, а согласуют работу друг с другом: обычно для запуска одного гена требуется совместное присутствие нескольких десятков (до сотни) факторов.
Итак, над генами стоит большой, сложно устроенный руководящий орган. С одной стороны, он умеет читать ДНК клетки, с другой — аппарат транскрипции реагирует на сигналы, поступающие в клетку извне. В ответ на поступающие сигналы (например, гормональные) клетка должна реагировать изменением экспрессии определенных генов, а реализуется эта реакция клетки через аппарат транскрипции. Интересно, например, что самый большой аппарат транскрипции — у клеток на ранних стадиях развития организма, когда они еще только выбирают, в какой тип клеток они разовьются. А у клеток, уже достигших зрелости, таких как нейрон или клетка мышцы, обычно происходит «сокращение штатов»: остаются только те, которые обслуживают специфические для этих клеток гены.
А у клеток, уже достигших зрелости, таких как нейрон или клетка мышцы, обычно происходит «сокращение штатов»: остаются только те, которые обслуживают специфические для этих клеток гены.
Таким образом, значительная часть наследственной информации распознается факторами транскрипции. В последнее время активно изучаются схожие с ними архитектурные факторы, распознающие определенные последовательности в ДНК и формирующие ее пространственную структуру. Вероятно, существуют и другие классы белков, умеющих прочитывать регуляторную информацию генома, которая пока непонятна нам. Важная фундаментальная научная задача — понять, как работает весь этот аппарат.
Это важно также и с практической точки зрения. Прежде всего, нужно вспомнить, что множество болезней вызвано именно тем, что нарушается контроль за работой генов. Это может быть связано с тем, что факторы транскрипции не справляются с работой: они могут быть поломаны, их может быть недостаточно или, наоборот, слишком много. Яркий пример — раковая опухоль: в некоторых клетках происходит бесконтрольная активация генов, нужных для роста и размножения клеток, в результате начинается «бунт» этих клеток против своего организма. Виновные в таком сценарии чаще всего оказываются именно факторы транскрипции, причем имена многих участников уже известны ученым. Другой пример — из современной области стволовых клеток. Если мы будем знать, какие именно факторы транскрипции работают в той или иной ткани, то сможем вырастить в пробирке ткань заданного типа.
Виновные в таком сценарии чаще всего оказываются именно факторы транскрипции, причем имена многих участников уже известны ученым. Другой пример — из современной области стволовых клеток. Если мы будем знать, какие именно факторы транскрипции работают в той или иной ткани, то сможем вырастить в пробирке ткань заданного типа.
В настоящее время изучение регуляции активности одного-единственного гена и факторов, которые его контролируют,— это большая научная задача. Наверное, нет еще такого гена, про регуляцию которого мы бы знали все. У исследователей, занимающихся механизмами регуляции экспрессии генов, еще много нерешенных задач. В Институте биологии гена РАН в Москве также ведутся исследования в этой области. Мы изучаем факторы транскрипции, которые выполняют разнообразные функции. Например, был найден фактор, который способен объединять две активности: он готовит матрицу ДНК к транскрипции, высвобождая ее из «архивированного» состояния, и одновременно собирает другие белки, которые запускают саму транскрипцию. Так осуществляется эффективный запуск работы генов, которые находятся под контролем этого фактора. Мы показали, что такой сценарий реализуется при действии внешних сигналов (например, некоторых гормонов): изученный фактор привлекается на нужные гены-мишени и запускает их работу.
Так осуществляется эффективный запуск работы генов, которые находятся под контролем этого фактора. Мы показали, что такой сценарий реализуется при действии внешних сигналов (например, некоторых гормонов): изученный фактор привлекается на нужные гены-мишени и запускает их работу.
Эти результаты были получены на мушке дрозофиле. Оказалось, что у других организмов, в том числе у позвоночных, есть схожие с ним белки. У мыши такой фактор присутствует в стволовых клетках мозга и важен для того, чтобы они не потеряли этот статус. Таким образом, изучение даже одного фактора транскрипции проводилось в целой серии работ и открыло новые механизмы регуляции активности клеток на уровне молекул и целых клеток. Работы активно продолжаются и, несомненно, мы узнаем еще много нового о том, как функционирует геном.
Работа отдельного гена контролируется несколькими десятками регуляторных элементов, которые могут быть расположены далеко от самого гена по последовательности генетического текста. С каждым элементом и с самим геном связывается целый набор различных факторов транскрипции, которые взаимодействуют друг с другом и с геном и таким образом регулируют его работу.
С каждым элементом и с самим геном связывается целый набор различных факторов транскрипции, которые взаимодействуют друг с другом и с геном и таким образом регулируют его работу.
Юлий Шидловский, Институт биологии гена РАН
Молекула жизни: как ДНК стала «царицей биологии»
Впрочем, на этом «оптическом» приближении генетики и останавливались — методов, позволяющих заглянуть «внутрь» хромосомы, тогда не существовало. Поэтому генетика того времени существовала отдельно от наук «молекулярного уровня». Генетики скрещивали мух и кукурузу и наблюдали за результатом, не вдаваясь в микроскопические и тем более молекулярные детали происходящего. В этом смысле ранняя генетика была ближе к классическим зоологии и ботанике, чем к биохимии, от которой сегодня она неотделима.В «молекулярном» же мире науки сороковых годов безоговорочно царили белки. Даже их греческое название — протеины, «первичные», передаёт благоговение биохимиков перед этой обширнейшей группой молекул, выполнявших, как считалось, все основные роли в живых организмах. К тридцатым годам было понятно, что белки состоят из двадцати аминокислот — невероятные возможности для комбинаторики! Были открыты ферменты, даже предсказаны некоторые аспекты белковой структуры.
К тридцатым годам было понятно, что белки состоят из двадцати аминокислот — невероятные возможности для комбинаторики! Были открыты ферменты, даже предсказаны некоторые аспекты белковой структуры.
По сравнению с белками ДНК с её четырьмя «кирпичиками» — нуклеотидами — казалась безнадёжно «глупой» молекулой, не способной к каким-либо «осмысленным» функциям. Популярной теорией того времени было, что ДНК всегда состоит из регулярных повторов нуклеотидов, представляя собой что-то вроде слегка усложнённого крахмала.
Генетики «поместили» наследственность в хромосомы, а биохимики установили, что те состоят из белка и ДНК. Как нетрудно догадаться, подавляющее большинство учёных считало само собой разумеющимся фактом, что наследственную функцию выполняет именно хромосомный белок — ДНК считалась второстепенной. Ей в лучшем случае дозволялось помогать белку заниматься «умными» делами.
Мёртвые бактерии, мёртвые мыши
В такой атмосфере Эвери, Маклеод и Маккарти опубликовали результаты своего революционного эксперимента. В качестве объекта изучения были выбраны пневмококки — бактерии, вызывающие пневмонию. Эти бактерии существуют в двух вариантах: R-тип, не вызывающий заболевания, и S-тип, способный убить заражённую им мышь.
В качестве объекта изучения были выбраны пневмококки — бактерии, вызывающие пневмонию. Эти бактерии существуют в двух вариантах: R-тип, не вызывающий заболевания, и S-тип, способный убить заражённую им мышь.
Было известно, что бактерии S-типа могут «обращать в свою веру» неопасные R-бактерии. Более того, такое «обращение», или трансформация R-бактерий, возможна, даже если сами S-бактерии убиты нагреванием.
Содержание желтых пигментов в зерне твердой пшеницы биосинтез, генетический контроль, маркерная селекция
Abstract
Зерно с высоким содержанием каротиноидных пигментов ценится за ярко-желтый цвет пасты,
производимой из него, и провитаминную (витамин А) и антиоксидантную активность пигментов. Цель настоя-
щего обзора – обобщение современных знаний о биосинтезе и генетическом контроле накопления пигментов
в зерне твердой пшеницы и оценка основных результатов исследований и селекции за последние двадцать
лет за рубежом и в России. Признак «концентрация каротиноидных пигментов в зерне» (Ypc) относится к раз-
ряду количественных. Тем не менее превалирование сильных аддитивных эффектов генов и высокая насле-
дуемость способствовали значительному прогрессу в селекции по этому признаку. Методами молекулярного
маркирования локусов количественных признаков (QTL), контролирующих синтез каротиноидных пигментов
и значения индекса желтизны (IY), установлено их распределение по всем хромосомам генома твердой пшеницы.
Основные генетические локусы, определяющие более 60 % варьирования признака, были картированы
в хромосомах 7AL и 7BL. Вклад этих локусов связан с аллельными вариациями, влияющими на активность
фермента фитоенсинтетазы (PSY). В других хромосомах были локализованы минорные генетические факторы,
из которых наиболее значимы QTL, расположенные в хромосомах 3AS (ассоциирован с геном LCYE-ликопин-
ε-циклаза) и 4ВS (аллель Lpx-B1.1c). При этом показано, что аллель Lpx-B1.1c вносит вклад в снижение актив-
ности липоксигеназы, окисляющей каротиноиды в процессе изготовления конечных продуктов. Рассмотрены
и обсуждены проблемы использования молекулярных маркеров в селекционных программах, нацеленных на
увеличение концентрации пигментов в зерне и улучшение цветовых характеристик пасты.
Тем не менее превалирование сильных аддитивных эффектов генов и высокая насле-
дуемость способствовали значительному прогрессу в селекции по этому признаку. Методами молекулярного
маркирования локусов количественных признаков (QTL), контролирующих синтез каротиноидных пигментов
и значения индекса желтизны (IY), установлено их распределение по всем хромосомам генома твердой пшеницы.
Основные генетические локусы, определяющие более 60 % варьирования признака, были картированы
в хромосомах 7AL и 7BL. Вклад этих локусов связан с аллельными вариациями, влияющими на активность
фермента фитоенсинтетазы (PSY). В других хромосомах были локализованы минорные генетические факторы,
из которых наиболее значимы QTL, расположенные в хромосомах 3AS (ассоциирован с геном LCYE-ликопин-
ε-циклаза) и 4ВS (аллель Lpx-B1.1c). При этом показано, что аллель Lpx-B1.1c вносит вклад в снижение актив-
ности липоксигеназы, окисляющей каротиноиды в процессе изготовления конечных продуктов. Рассмотрены
и обсуждены проблемы использования молекулярных маркеров в селекционных программах, нацеленных на
увеличение концентрации пигментов в зерне и улучшение цветовых характеристик пасты.
Keywords: твердая пшеница, каротиноиды, концентрация пигментов, индекс желтизны, маркерная селекция
Abstract
Grain with high contents of yellow pigments will add the natural bright-yellow colour to the paste, which
unlike a paste with a high level of whiteness, are preferred by consumers. The provitamin activity (vitamin A) and
antioxidant activity of the carotenoid pigment increase the biological and nutritional value of the grain with high
contents of these pigments. The purpose of this review is to summarize modern knowledge about the biosynthesis
and genetic control of pigment accumulation in durum wheat and to assess the main results of research and
selection over the past 20 years abroad and in Russia. The trait “concentration carotenoid pigment in grain” (Ypc) is
quantitative. However, the prevalence of strong additive gene effects and high heritability have contributed to significant
progress in breeding for this trait. Molecular labeling of quantitative trait loci (QTL) that control the synthesis
of the carotenoid pigment and the yellowness index (YI) found that they are distributed across all chromosomes of
the durum wheat genome. The main QTLs, which determine 60 % of the variation of the trait, were mapped to 7AL
and 7BL chromosome. The contribution of these QTLs is associated with allelic variations that control the activity of
phytoene synthase (PSY). QTLs with minor effects found on the remaining chromosomes are also reliably mapped
using molecular markers. As confirmed in a number of experiments, most of them are QTLs located on 3AS (linked
to the LCYE (lycopene ε-cyclase) allele and on 4BS (the LpxB1.1c gene). It has been shown that the LpxB1.1c allele contributes to a decrease in the activity of lipoxygenase, which oxidases carotenoids during the production of end
products. This review considered and discusses the problems of molecular markers in breeding programs to increase
the concentration of pigments in the grain and improve the color characteristics of the paste.
The main QTLs, which determine 60 % of the variation of the trait, were mapped to 7AL
and 7BL chromosome. The contribution of these QTLs is associated with allelic variations that control the activity of
phytoene synthase (PSY). QTLs with minor effects found on the remaining chromosomes are also reliably mapped
using molecular markers. As confirmed in a number of experiments, most of them are QTLs located on 3AS (linked
to the LCYE (lycopene ε-cyclase) allele and on 4BS (the LpxB1.1c gene). It has been shown that the LpxB1.1c allele contributes to a decrease in the activity of lipoxygenase, which oxidases carotenoids during the production of end
products. This review considered and discusses the problems of molecular markers in breeding programs to increase
the concentration of pigments in the grain and improve the color characteristics of the paste.
Keywords: durum wheat, carotenoids, concentration pigment, yellow index, marker-assisted selection
Введение
Твердая пшеница (Triticum durum Desf. ) ежегодно вы-
ращивается во всем мире на площади около 17.0 млн га.
Производство зерна колеблется по годам от 32 до 42 млн
тонн. Продукты из твердой пшеницы используются почти
во всех странах, но основными регионами потребления
являются страны Средиземноморского бассейна, где зер-
но твердой пшеницы применяется в качестве сырья для
различных продуктов, в основном пасты и круп кус-кус и
булгур (Kabbaj et al., 2017). Содержание и структура белка
(качество клейковины), а также цвет изделий (макароны,
крупа) – наиболее ценные для пищевой, технологической
промышленности и на рынке конечных продуктов в этих
регионах (Sisson, 2008; Mazzeo et al., 2017). Желтый цвет
определяется накоплением каротиноидов в эндосперме,
обладающих провитаминными и антиоксидантными свойствами.
В процессе размола зерна, замеса теста и изго-
товления макарон происходит окисление кислородом воз-
духа полиненасыщенных жирных кислот, каротиноидов,
фенолов, что приводит к накоплению бурого (коричне-
вого) пигмента. Окисление катализируется ферментами:
липоксигеназой, полифенолоксидазой, пероксидазой, альдегидоксидазой
(N’Diaye et al.
) ежегодно вы-
ращивается во всем мире на площади около 17.0 млн га.
Производство зерна колеблется по годам от 32 до 42 млн
тонн. Продукты из твердой пшеницы используются почти
во всех странах, но основными регионами потребления
являются страны Средиземноморского бассейна, где зер-
но твердой пшеницы применяется в качестве сырья для
различных продуктов, в основном пасты и круп кус-кус и
булгур (Kabbaj et al., 2017). Содержание и структура белка
(качество клейковины), а также цвет изделий (макароны,
крупа) – наиболее ценные для пищевой, технологической
промышленности и на рынке конечных продуктов в этих
регионах (Sisson, 2008; Mazzeo et al., 2017). Желтый цвет
определяется накоплением каротиноидов в эндосперме,
обладающих провитаминными и антиоксидантными свойствами.
В процессе размола зерна, замеса теста и изго-
товления макарон происходит окисление кислородом воз-
духа полиненасыщенных жирных кислот, каротиноидов,
фенолов, что приводит к накоплению бурого (коричне-
вого) пигмента. Окисление катализируется ферментами:
липоксигеназой, полифенолоксидазой, пероксидазой, альдегидоксидазой
(N’Diaye et al. , 2017). Наличие спексов
(темных вкраплений) также может снижать уровень желтизны
семолины и макаронных изделий (Васильчук и
др., 2009).
, 2017). Наличие спексов
(темных вкраплений) также может снижать уровень желтизны
семолины и макаронных изделий (Васильчук и
др., 2009).
Результаты изучения наследования всего комплекса желтых пигментов в зерне твердой пшеницы показали, что оно носит количественный характер с высокими зна- чениями коэффициента наследуемости и преобладанием аддитивных эффектов генов (Clarke еt al.,1998; Borelli et al., 1999; Digesù et al., 2009; Blanco et al., 2011; Roncallo et al., 2012; Schulthess et al., 2013). В связи с этим право- мерно предположение о том, что этот признак удобен для молекулярного маркирования соответствующих QTL и их картирования на хромосомах.
Биосинтез, содержание и распределение каротиноидов в зерновке твердой пшеницы
Средняя концентрация каротиноидов в зерне твердой
пшеницы составляет 6.2 ± 0.13 мг/кг в сухом весе (Brandolini
et al., 2015) с варьированием, в зависимости от
сорта и условий среды (год, пункт), от 2.8 до 12.3 мг/кг
(Colasuonno et al., 2017a). По мнению Н. С. Васильчука
с коллегами (2009), в Поволжье при низкой активности
окислительных ферментов достаточно иметь зерно с кон-
центрацией 4.5 мг/кг для получения макарон золотисто-
желтого цвета. Каротиноиды – не единственные желтые
пигменты в зерне пшеницы и других злаков. Сравнивая
общую концентрацию каротиноидов, определенную ме-
тодом высокоэффективной жидкостной хромотографии (ВЭЖХ), с общим содержанием пигментов, A.M. Digesù
с коллегами (2009) показали, что доля каротиноидов у
культивируемых и диких тетраплоидных видов пше-
ницы составила 33.2 % от общего количества желтых
пигментов. По результатам своих исследований A. Blanco
с колегами (2011) также сообщили о доле каротиноидов,
составившей 37 % в общем объеме желтых пигментов, что
означает наличие в экстрактах твердой пшеницы неизвест-
ных желтых пигментов, поглощающих свет при 435 нм.
По данным (Fu et al., 2017), полученным при изучении
канадских сортов твердой пшеницы Navigator и Strongfield,
фенольные соединения могут вносить существенный
вклад в степень желтизны при экстракции пигментов.
С. Васильчука
с коллегами (2009), в Поволжье при низкой активности
окислительных ферментов достаточно иметь зерно с кон-
центрацией 4.5 мг/кг для получения макарон золотисто-
желтого цвета. Каротиноиды – не единственные желтые
пигменты в зерне пшеницы и других злаков. Сравнивая
общую концентрацию каротиноидов, определенную ме-
тодом высокоэффективной жидкостной хромотографии (ВЭЖХ), с общим содержанием пигментов, A.M. Digesù
с коллегами (2009) показали, что доля каротиноидов у
культивируемых и диких тетраплоидных видов пше-
ницы составила 33.2 % от общего количества желтых
пигментов. По результатам своих исследований A. Blanco
с колегами (2011) также сообщили о доле каротиноидов,
составившей 37 % в общем объеме желтых пигментов, что
означает наличие в экстрактах твердой пшеницы неизвест-
ных желтых пигментов, поглощающих свет при 435 нм.
По данным (Fu et al., 2017), полученным при изучении
канадских сортов твердой пшеницы Navigator и Strongfield,
фенольные соединения могут вносить существенный
вклад в степень желтизны при экстракции пигментов. Brandolini с коллегами (2008) пришли к выводу, что, хотя
каротиноиды являются наиболее важными пигментами
при определении желтого цвета пшеничной муки и мака-
рон, их точное измерение может быть достигнуто только
с помощью ВЭЖХ анализа. Тем не менее среди желтых
пигментов именно по каротиноидам имеется более под-
робная научная информация.
Brandolini с коллегами (2008) пришли к выводу, что, хотя
каротиноиды являются наиболее важными пигментами
при определении желтого цвета пшеничной муки и мака-
рон, их точное измерение может быть достигнуто только
с помощью ВЭЖХ анализа. Тем не менее среди желтых
пигментов именно по каротиноидам имеется более под-
робная научная информация.
К каротиноидам относятся пигменты – каротины и ксантофиллы. Химически они представляют собой изо- преноидные углеводороды, содержащие 40 углеродных атомов (Кретович, 1986). Биосинтез каротиноидов тща- тельно исследован на различных растениях – арабидоп- сисе, рисе, кукурузе, перце, томатах, апельсине и других культурах (Colasuonno et al., 2017a; Rodrigues-Concepcion et al., 2018; Sun et al., 2018).
В зерне твердой пшеницы представлен широкий набор
каротиноидных пигментов: лютеин, β-каротин, зеаксантин,
β-криптоксантин, β-апокаротенал, антраксантин,
тараксантин (лютеин-5,6-эпоксид), авоксантин и тритикоксантин.
Каротины α и β в основном находятся в за-
родыше, превалирующий в зерне среди каротиноидов
лютеин (86–94 %) одинаково распределен по слоям и
частям зерновки (Digesù et al. , 2009). Его доля в семолине
при размоле зерна твердой пшеницы составляет 83 %, во
фракциях отрубей – 75 % (Fu et al., 2017). Установлено, что
характер распределения желтых пигментов по фракциям,
выделяемым в процессе размола, варьирует, в зависимо-
сти от генотипа. Концентрация лютеина в эндосперме
сорта Navigator была выше, чем в отрубях, в то же время
у сорта Strongfield наблюдалась обратная закономерность
(Fu et al., 2017). В процессе помола и получения крупки
(семолины) концентрация большинства каротиноидных
пигментов уменьшается, что связано с увеличением кон-
такта измельченных частиц зерна с кислородом воздуха
и активностью ферментов. Лютеин и зеаксантин имеют
более высокую стабильность при помоле и изготовлении
конечных продуктов, по сравнению с другими каротиноидами
(Kean et al., 2011).
, 2009). Его доля в семолине
при размоле зерна твердой пшеницы составляет 83 %, во
фракциях отрубей – 75 % (Fu et al., 2017). Установлено, что
характер распределения желтых пигментов по фракциям,
выделяемым в процессе размола, варьирует, в зависимо-
сти от генотипа. Концентрация лютеина в эндосперме
сорта Navigator была выше, чем в отрубях, в то же время
у сорта Strongfield наблюдалась обратная закономерность
(Fu et al., 2017). В процессе помола и получения крупки
(семолины) концентрация большинства каротиноидных
пигментов уменьшается, что связано с увеличением кон-
такта измельченных частиц зерна с кислородом воздуха
и активностью ферментов. Лютеин и зеаксантин имеют
более высокую стабильность при помоле и изготовлении
конечных продуктов, по сравнению с другими каротиноидами
(Kean et al., 2011).
Накопление каротинов в зерне, особенно β-каротина,
обуславливает изменение интенсивности окраски семоли-
ны от желто-оранжевого до красноватого оттенка. Ксан-
тофиллы обеспечивают желто-оранжевую окраску крупки
и макарон. Схема биосинтеза каротиноидов показана на
рисунке. Исходным веществом для биохимического син-
теза каротинов служит 5-углеродный (С-5) изопреноид –
изопентилпирофосфат. Конденсация этого изопреноида
представляет собой основу для образования геранил-
геранилпирофосфата (С-20). В результате соединения
двух молекул геранилгеранилпирофосфата при участии
фермента фитоенсинтетазы образуется фитоен (С-40) –
первое промежуточное вещество в биосинтезе каротинов.
Этот этап является ключевым – скорость биосинтеза и на-
копления фитоена влияет на весь пул каротиноидов (Cazzonelli,
Pogson, 2010; Ke et al., 2019). Фитоен в результате
десатурации под действием ферментов фитоендесатуразы
(PDS), z-каротиндесатуразы (ZDS), каротинизомеразы
(CRTISO) и последовательного удаления четырех атомов
водорода превращается в ликопин. Ликопин – каротиноид,
определяющий красную и оранжевую окраску плодов, –
исходное вещество для синтеза α-каротина/лютеина (класс
ксантофиллов) – главного каротиноида зерна твердой
пшеницы и β-каротина/зеаксантина (класс ксантофил-
лов) – главного компонента каротиноидных пигментов
в зерне кукурузы (Zhang, Dubcovsky, 2008).
Схема биосинтеза каротиноидов показана на
рисунке. Исходным веществом для биохимического син-
теза каротинов служит 5-углеродный (С-5) изопреноид –
изопентилпирофосфат. Конденсация этого изопреноида
представляет собой основу для образования геранил-
геранилпирофосфата (С-20). В результате соединения
двух молекул геранилгеранилпирофосфата при участии
фермента фитоенсинтетазы образуется фитоен (С-40) –
первое промежуточное вещество в биосинтезе каротинов.
Этот этап является ключевым – скорость биосинтеза и на-
копления фитоена влияет на весь пул каротиноидов (Cazzonelli,
Pogson, 2010; Ke et al., 2019). Фитоен в результате
десатурации под действием ферментов фитоендесатуразы
(PDS), z-каротиндесатуразы (ZDS), каротинизомеразы
(CRTISO) и последовательного удаления четырех атомов
водорода превращается в ликопин. Ликопин – каротиноид,
определяющий красную и оранжевую окраску плодов, –
исходное вещество для синтеза α-каротина/лютеина (класс
ксантофиллов) – главного каротиноида зерна твердой
пшеницы и β-каротина/зеаксантина (класс ксантофил-
лов) – главного компонента каротиноидных пигментов
в зерне кукурузы (Zhang, Dubcovsky, 2008). Дальнейшее
гидроксилирование α-каротина приводит к образованию
желтого зейоксантина и лютеина. Трансформация β-каротина
продуцирует образование β-криптоксантина, зеа-
ксантина, антраксантина, виолаксантина и неоксантина.
Эти реакции катализируются двумя негемовыми β-каротингидроксилазами
(ВСН1 и ВСН2) и двумя гемгидроксилазами
(CYP97A и CYP97C) соответственно (Sun et
al., 2018). Последняя фаза биосинтеза каротиноидов, катализируемая
неоксантиноксидазой (NXS), заключается
в превращении виолаксантина в неоксантин. Окисление
виолаксантина и неоксантина приводит к образованию
ксантоксина, превращаемого в растительный гормон –
абсцизовую кислоту (ABA), которая способствует регу-
лярному и сбалансированному накоплению пигментов в
растениях и формированию устойчивости к абиотическим
стрессам (Al-Babilli, Bowmeester, 2015; Nisar et al., 2015).
Еще одна ветвь трансформации β-каротина представляет
превращение его под действием ферментов диоксигеназ-
ной группы (CCD7, CCD8, CYP711A1) в стриголактоны –
ингредиенты гормональной природы, регулирующие
развитие и ростовые процессы растений (Colasuonno et
al.
Дальнейшее
гидроксилирование α-каротина приводит к образованию
желтого зейоксантина и лютеина. Трансформация β-каротина
продуцирует образование β-криптоксантина, зеа-
ксантина, антраксантина, виолаксантина и неоксантина.
Эти реакции катализируются двумя негемовыми β-каротингидроксилазами
(ВСН1 и ВСН2) и двумя гемгидроксилазами
(CYP97A и CYP97C) соответственно (Sun et
al., 2018). Последняя фаза биосинтеза каротиноидов, катализируемая
неоксантиноксидазой (NXS), заключается
в превращении виолаксантина в неоксантин. Окисление
виолаксантина и неоксантина приводит к образованию
ксантоксина, превращаемого в растительный гормон –
абсцизовую кислоту (ABA), которая способствует регу-
лярному и сбалансированному накоплению пигментов в
растениях и формированию устойчивости к абиотическим
стрессам (Al-Babilli, Bowmeester, 2015; Nisar et al., 2015).
Еще одна ветвь трансформации β-каротина представляет
превращение его под действием ферментов диоксигеназ-
ной группы (CCD7, CCD8, CYP711A1) в стриголактоны –
ингредиенты гормональной природы, регулирующие
развитие и ростовые процессы растений (Colasuonno et
al. , 2019).
, 2019).
The main components of the biosynthetic pathway are shown in black; all enzymes involved in carotenoid synthesis, in blue; enzymes of the dioxygenase group involved in carotenoid metabolism in growing plants under stress and accumulating hormone-like ingredients – abscisic acid and strigolactones – in red.
Методы определения общего и компонентного состава каротиноидных пигментов в зерне
Содержание пигментов в зерне, крупке и макаронах от-
носится к сложным признакам. Для общего и компонент-
ного определения их содержания существует несколько
мотодов. Эталонные методы для определения общего
содержания каротиноидов – стандартный метод 152 (ICC
Method 152, 1990) Международной ассоциации науки и
техники о зерне (ICC) и международный официальный
метод (ААСС 14-50.01 (AACC International, 2013). Эти
две процедуры основаны на экстракции всех пигментов
в водонасыщенном н-бутаноле с последующим спектрофотометрическим
количественным определением оптиче-
ской плотности спиртового экстракта при 435.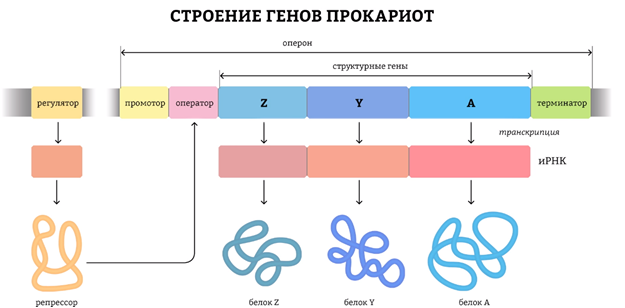 5 нм (длина
волны максимального поглощения лютеина, доминирующего
каротиноида в твердой пшенице). В качестве альтер-
нативы концентрации для характеристики насыщенно-
сти
пигментами зерна применяется индекс желтизны (YI)
на основе количественного определения коэффициента
отражения света. Хроматограф Minolta CR-300 (Konica Minolta Pty Ltd, Macquarie Park, NSW), оснащенный им-
пульсной ксеноновой дуговой лампой, – наиболее часто
используемый инструмент для анализа индекса желтизны.
Этот прибор позволяет определять коэффициенты индекса
желтизны и коричневатости (100-L) семолины и макарон.
В России в некоторых селекционных центрах для опреде-
ления индекса желтизны применяется прибор Specol 10
по методологии, предложенной Н.С. Васильчуком (2001).
Методы быстродействующей жидкостной хромотографии
высокого давления (HPLC) с применением инфракрасной
спектроскопии позволяют определить химический состав
каротиноидных пигментов и измерить количество каждого
компонента в крупке и пасте (Brandolini et al.
5 нм (длина
волны максимального поглощения лютеина, доминирующего
каротиноида в твердой пшенице). В качестве альтер-
нативы концентрации для характеристики насыщенно-
сти
пигментами зерна применяется индекс желтизны (YI)
на основе количественного определения коэффициента
отражения света. Хроматограф Minolta CR-300 (Konica Minolta Pty Ltd, Macquarie Park, NSW), оснащенный им-
пульсной ксеноновой дуговой лампой, – наиболее часто
используемый инструмент для анализа индекса желтизны.
Этот прибор позволяет определять коэффициенты индекса
желтизны и коричневатости (100-L) семолины и макарон.
В России в некоторых селекционных центрах для опреде-
ления индекса желтизны применяется прибор Specol 10
по методологии, предложенной Н.С. Васильчуком (2001).
Методы быстродействующей жидкостной хромотографии
высокого давления (HPLC) с применением инфракрасной
спектроскопии позволяют определить химический состав
каротиноидных пигментов и измерить количество каждого
компонента в крупке и пасте (Brandolini et al. , 2008; Fu
et al., 2017).
, 2008; Fu
et al., 2017).
Картирование локусов, контролирующих синтез пигментов
Концентрация пигментов в зерне твердой пшеницы кон-
тролируется различными генами с аддитивными эффекта-
ми и зависит от условий внешней среды (Васильчук и др.,
2009; Мальчиков, 2009; Schulthess, Schwember, 2013; Гапо-
нов и др., 2018; Мясникова и др., 2019). Систематическое
сортоизучение
в различных экологических условиях дает
необходимую информацию о свойствах сортов и наличии
у них соответствующих QTL. Достоверные, значительные
и стабильные различия между генотипами по величинам
YPC (Yellow Pigment Concentration) и YI (Yellowness Index)
свидетельствуют о функционировании локусов
количе-
ственного признака. Для маркирования и локализации
QTL используют рекомбинантно-инбредные линии (RIL),
созданные от двуродительских скрещиваний контрастных
по величине признака сортов и отобранные в поколениях
(от одного парного скрещивания) из беккроссных или
дигаплоидных популяций (Elouafi еt аl., 2001; Pozniak
et al. , 2007; Singh et al., 2009; Colasuonno et al., 2014).
В настоящее время для этих целей используется метод
полногеномного генотипирования для поиска корреляций
между генотипами и фенотипами в наборах селекционных
линий, образцов генетических коллекций и выявления
аллельных вариантов молекулярных маркеров и функцио-
нальных генов (Vargas et al., 2016; Colasuonno et al., 2017a;
Fiedler et al., 2017). В последнее десятилетие этот подход
получил распространение благодаря наличию большого
количества ДНК-маркеров, равномерно распределенных
в геноме, полученных на основе однонуклеотиного по-
лиморфизма (SNP) и совершенствования статистических
инструментов (Чесноков, Артемьева, 2011; Maccaferri et
al., 2011; Wang et al., 2014; Sehgal, Dreisigacker, 2019).
Картирование QTL предполагает хромосомную локали-
зацию локуса в геноме по результатам оценки признаков,
полученных в нескольких экспериментах.
, 2007; Singh et al., 2009; Colasuonno et al., 2014).
В настоящее время для этих целей используется метод
полногеномного генотипирования для поиска корреляций
между генотипами и фенотипами в наборах селекционных
линий, образцов генетических коллекций и выявления
аллельных вариантов молекулярных маркеров и функцио-
нальных генов (Vargas et al., 2016; Colasuonno et al., 2017a;
Fiedler et al., 2017). В последнее десятилетие этот подход
получил распространение благодаря наличию большого
количества ДНК-маркеров, равномерно распределенных
в геноме, полученных на основе однонуклеотиного по-
лиморфизма (SNP) и совершенствования статистических
инструментов (Чесноков, Артемьева, 2011; Maccaferri et
al., 2011; Wang et al., 2014; Sehgal, Dreisigacker, 2019).
Картирование QTL предполагает хромосомную локали-
зацию локуса в геноме по результатам оценки признаков,
полученных в нескольких экспериментах.
Воспроизводимость результатов может зависеть от
влияния
на признак в исследуемой картирующей попу-
ляции многочисленных генов с аддитивными эффектами,
родительских компонентов, взаимодействия генотип–сре-
да, числа используемых маркеров и способа измерения
каротиноидов. В настоящее время для YPC и YI иденти-
фицирован 81 QTL, включая синглетоны и кластеры QTL,
которые распределены по всем хромосомам (Colasuonno
et al., 2019). Локализация некоторых QTL была подтверждена с использованием нескольких картирующих популя-
ций, что указывает на присутствие стабильных аллелей,
влияющих на улучшение цвета и пищевой ценности зерна
твердой пшеницы. Стабильные QTL были обнаружены в
хромосомах 1А, 1В, 2А, 2В, 3В, 4А, 6А, 6В, 7А, 7В (Parker
et al., 1998; Hessler et al., 2002; Patil et al., 2008; Zhang et
al., 2009; Pozniak et al., 2012).
В настоящее время для YPC и YI иденти-
фицирован 81 QTL, включая синглетоны и кластеры QTL,
которые распределены по всем хромосомам (Colasuonno
et al., 2019). Локализация некоторых QTL была подтверждена с использованием нескольких картирующих популя-
ций, что указывает на присутствие стабильных аллелей,
влияющих на улучшение цвета и пищевой ценности зерна
твердой пшеницы. Стабильные QTL были обнаружены в
хромосомах 1А, 1В, 2А, 2В, 3В, 4А, 6А, 6В, 7А, 7В (Parker
et al., 1998; Hessler et al., 2002; Patil et al., 2008; Zhang et
al., 2009; Pozniak et al., 2012).
По эффектам на фенотипическую вариабельность
признаков
QTL распределяются на группы с сильным
(40 %), средним (10–40 %) и незначительным (< 10 %)
влиянием (Colasuonno et al., 2019). Главные локусы с
сильным влиянием на YPC и YI были картированы в
хромосоме 7АL и дистальной области 7BL (Elouafi et al.,
2001; Pozniak et al., 2007; Patil et al., 2008; Zhang, Dubcovsky,
2008). В частности, в длинном плече хромосомы 7A
были идентифицированы два QTL с противоположными
эффектами. Локус с негативным эффектом по отношению
к признакам YPC и YI был ассоциирован с аллельными
вариациями гена альдегидоксидазы (АО) – фермента,
катализирующего деградацию каротиноидов в результа-
те их окисления (Colasuonno et al., 2014, 2017b). Второй
(QTL-73), расположенный в области локализации гена
Psy-1, оказывал положительный эффект (до 60 % вариации)
на признак YPC. Аналогичные ассоциации QTL и
YPC установлены в хромосоме 7BL – негативный эффект
одного локуса на фенотипическое проявление YPC составлял
29 %, позитивное влияние второго локуса – 52 %
(He et al., 2008, 2009; Zhang, Dubcovsky, 2008). В публи-
кации (N’Diaye et al., 2017) сообщается о картировании
YPC и Yi методом конструирования гаплотипных блоков.
Гаплотипные блоки формируются с применением насы-
щенных молекулярных SNP карт. При этом SNP, распо-
ложенные в хромосоме в пределах 5.3 сМ, группируют в
один гаплотипный блок, определяющий один локус, ко-
торый обозначается префиксом “hap” с указанием номера
хромосомы и порядкового номера локуса на хромосоме.
Локус с негативным эффектом по отношению
к признакам YPC и YI был ассоциирован с аллельными
вариациями гена альдегидоксидазы (АО) – фермента,
катализирующего деградацию каротиноидов в результа-
те их окисления (Colasuonno et al., 2014, 2017b). Второй
(QTL-73), расположенный в области локализации гена
Psy-1, оказывал положительный эффект (до 60 % вариации)
на признак YPC. Аналогичные ассоциации QTL и
YPC установлены в хромосоме 7BL – негативный эффект
одного локуса на фенотипическое проявление YPC составлял
29 %, позитивное влияние второго локуса – 52 %
(He et al., 2008, 2009; Zhang, Dubcovsky, 2008). В публи-
кации (N’Diaye et al., 2017) сообщается о картировании
YPC и Yi методом конструирования гаплотипных блоков.
Гаплотипные блоки формируются с применением насы-
щенных молекулярных SNP карт. При этом SNP, распо-
ложенные в хромосоме в пределах 5.3 сМ, группируют в
один гаплотипный блок, определяющий один локус, ко-
торый обозначается префиксом “hap” с указанием номера
хромосомы и порядкового номера локуса на хромосоме. Этот подход подтвердил существование высокозначимого
QTL (hap_7A_32, маркер Tdurum_conting 54832_139) в
хромосоме 7АL, объясняющего 35.6 % фенотипической
дисперсии общего пигмента и индекса желтизны в семо-
лине твердой пшеницы и связанного с локусом Psy-A1.
Использование подобного метода подтвердило достовер-
ную значимость QTL в хромосомах 2A (hap_2A_18), 7B
(hap_7B_36) и 4B (hap_4B_6).
Этот подход подтвердил существование высокозначимого
QTL (hap_7A_32, маркер Tdurum_conting 54832_139) в
хромосоме 7АL, объясняющего 35.6 % фенотипической
дисперсии общего пигмента и индекса желтизны в семо-
лине твердой пшеницы и связанного с локусом Psy-A1.
Использование подобного метода подтвердило достовер-
ную значимость QTL в хромосомах 2A (hap_2A_18), 7B
(hap_7B_36) и 4B (hap_4B_6).
Незначительные по эффектам QTL обнаружены в хро- мосомах 3А (Parker et al., 1998), 4A и 5A (Hessler et al., 2002), 2A, 4B и 6В (Pozniak et al., 2007), 4B и 6B (Zhang, Dubcovsky, 2008), 1А, 3В и 5В (Patil et al., 2008), 3B и 5B (Howitt et al., 2009), 1A, 1B, 3B и 4А (Zhang et al., 2009).
Гены, участвующие в биосинтезе каротиноидов и их окислении в процессе изготовления конечных продуктов
В настоящее время получена информация о генах-канди-
датах, расположенных в регионах локализации локусов со
значимым влиянием на признаки YPC и YI. В данном слу-
чае к ним относятся гены, контролирующие активность
основных ферментов биосинтеза каротиноидов пшеницы,
и катаболические гены, вызывающие деградацию каро-
тиноидов. Количественные различия генотипов по на коплению пигментов связаны с аллельным разнообра-
зием генов PSY – фитоенсинтетазы (Pozniak et al., 2007;
He et al., 2008; Dibari et al., 2012; Colasuonno et al., 2014;
Campos et al., 2016), LCYE – ликопин-ε-циклазы (Howitt
et al., 2009; Crawford, Francki, 2013), LCYB – ликопин-β-
циклазы (Zeng et al., 2015), HYD – β-гидроксилазы (Qin
et al., 2012), PDS (фитоендесатуразы) и ZDS – каротин-
десатуразы (Cong et al., 2010). Катаболические гены контролируют
активность альдегидоксидазы – АО (Colasuonno
et al., 2017b), полифенолоксидазы – РРО (Watanabe et al.,
2004, 2006; Si et al., 2012), липоксигеназы – LOX или Lpx
(DeSimone еt al., 2010; Randhawa et al., 2013) и пероксида-
зы – PER (Ficco et al., 2014), снижающих концентрацию
пигментов и потребительские качества конечных продук-
тов. Наиболее значимые гены, их аллельные варианты и
маркеры обсуждаются в тексте и представлены в табл. 1.
Количественные различия генотипов по на коплению пигментов связаны с аллельным разнообра-
зием генов PSY – фитоенсинтетазы (Pozniak et al., 2007;
He et al., 2008; Dibari et al., 2012; Colasuonno et al., 2014;
Campos et al., 2016), LCYE – ликопин-ε-циклазы (Howitt
et al., 2009; Crawford, Francki, 2013), LCYB – ликопин-β-
циклазы (Zeng et al., 2015), HYD – β-гидроксилазы (Qin
et al., 2012), PDS (фитоендесатуразы) и ZDS – каротин-
десатуразы (Cong et al., 2010). Катаболические гены контролируют
активность альдегидоксидазы – АО (Colasuonno
et al., 2017b), полифенолоксидазы – РРО (Watanabe et al.,
2004, 2006; Si et al., 2012), липоксигеназы – LOX или Lpx
(DeSimone еt al., 2010; Randhawa et al., 2013) и пероксида-
зы – PER (Ficco et al., 2014), снижающих концентрацию
пигментов и потребительские качества конечных продук-
тов. Наиболее значимые гены, их аллельные варианты и
маркеры обсуждаются в тексте и представлены в табл. 1.
Genes and their markers associated with the concentration of yellow pigments in grains and processed products located in wheat genomes A and B
Как уже отмечено, ключевым ферментом в сложной це-
почке биосинтеза каротиноидов является PSY.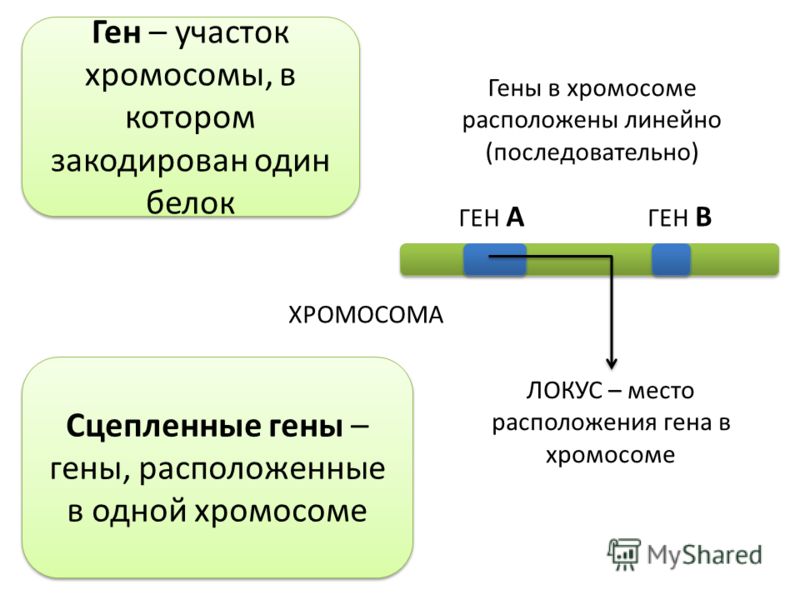 Известны
три различных гена, кодирующих активность этого фер-
мента: PSY 1, PSY 2, PSY 3, картированные в гомеологич-
ных хромосомах 7-, 5- и 3-й групп соответственно (Dibari
et al., 2012). Ген PSY-1 был локализован в хромосомах 7А
и 7В твердой пшеницы. Обнаружено, что ген PSY-B1,
расположенный в хромосоме 7В, сегрегирует совместно с
QTL, ассоциированным с каротиноидным пулом, с измен-
чивостью YI и YPC от низкого уровня (10 %) до среднего
(10–30 %). Ген PSY-A1, расположенный в хромосоме 7А,
ведет себя как кодоминантный маркер, объясняет фено-
типическую изменчивость, в зависимости от генофона, в
диапазоне от среднего (10–30 %) до высокого (30–50 %) и
очень высокого (> 50 %) уровня (Colasuonno et al., 2014).
В целом влияние альтернативных аллелей PSY-A1, по-
видимому, является наиболее важным в изменчивости
концентрации пигментов и индекса желтизны крупки.
Этот вывод основан на результатах изучения различных
популяций твердой пшеницы (Campos et al., 2016; Vargas
et al., 2016; Patil et al.
Известны
три различных гена, кодирующих активность этого фер-
мента: PSY 1, PSY 2, PSY 3, картированные в гомеологич-
ных хромосомах 7-, 5- и 3-й групп соответственно (Dibari
et al., 2012). Ген PSY-1 был локализован в хромосомах 7А
и 7В твердой пшеницы. Обнаружено, что ген PSY-B1,
расположенный в хромосоме 7В, сегрегирует совместно с
QTL, ассоциированным с каротиноидным пулом, с измен-
чивостью YI и YPC от низкого уровня (10 %) до среднего
(10–30 %). Ген PSY-A1, расположенный в хромосоме 7А,
ведет себя как кодоминантный маркер, объясняет фено-
типическую изменчивость, в зависимости от генофона, в
диапазоне от среднего (10–30 %) до высокого (30–50 %) и
очень высокого (> 50 %) уровня (Colasuonno et al., 2014).
В целом влияние альтернативных аллелей PSY-A1, по-
видимому, является наиболее важным в изменчивости
концентрации пигментов и индекса желтизны крупки.
Этот вывод основан на результатах изучения различных
популяций твердой пшеницы (Campos et al., 2016; Vargas
et al., 2016; Patil et al. , 2018). Ген PSY-B2, локализованный
в 5ВS, не оказывал существенного влияния на концен-
трацию каротиноидов в зерне и крупке (Colasuonno et
al., 2014). Локализованный в длинном плече хромосомы
5В ген PSY-B3 повышал экспрессию в листьях и корнях
в условиях абиотического стресса (засуха, засоление) и был связан с увеличением пула абсцизовой кислоты
(Dibari et al., 2012). Гены, контролирующие активность
других ферментов, также имеют значимое влияние на
концентрацию пигментов в зерне и конечных продуктах.
Так, ген ликопин-ε-циклазы (LCYE), ассоциированный
с QTL на хромосоме 3А, играет определяющую роль в
процессах накопления лютеина – главного каротиноида
зерна твердой пшеницы (Howitt et al., 2009). Ген z-каротиндесатуразы
(ZDS), маркируемый кодоминантным
функциональным маркером YP2A-1 на основе полимор-
физма двух аллелей, расположенный на хромосоме 2А,
объяснял 11.3 % фенотипической дисперсии YPC и IY в
популяции дигаплоидных линий (Dong et al., 2012).
, 2018). Ген PSY-B2, локализованный
в 5ВS, не оказывал существенного влияния на концен-
трацию каротиноидов в зерне и крупке (Colasuonno et
al., 2014). Локализованный в длинном плече хромосомы
5В ген PSY-B3 повышал экспрессию в листьях и корнях
в условиях абиотического стресса (засуха, засоление) и был связан с увеличением пула абсцизовой кислоты
(Dibari et al., 2012). Гены, контролирующие активность
других ферментов, также имеют значимое влияние на
концентрацию пигментов в зерне и конечных продуктах.
Так, ген ликопин-ε-циклазы (LCYE), ассоциированный
с QTL на хромосоме 3А, играет определяющую роль в
процессах накопления лютеина – главного каротиноида
зерна твердой пшеницы (Howitt et al., 2009). Ген z-каротиндесатуразы
(ZDS), маркируемый кодоминантным
функциональным маркером YP2A-1 на основе полимор-
физма двух аллелей, расположенный на хромосоме 2А,
объяснял 11.3 % фенотипической дисперсии YPC и IY в
популяции дигаплоидных линий (Dong et al., 2012).
Наиболее четко выражены гены-кандидаты, участвующие
в катаболическом пути окисления каротиноидов. Липоксигеназа (Lpx) у растений продуцирует активные
формы кислорода, приводящие к деградации каротинои-
дов и обесцвечиванию конечных продуктов, получаемых
из зерна твердой пшеницы (Borrelli et al., 2003). У твердой
пшеницы существуют разные гены Lpx (Borrelli, Trono,
2016). На стадии зрелого зерна у сортов с контрастной
активностью (окислительной способностью) была уста-
новлена различная степень транскрипции генов Lpx-1 и
Lpx-3, в то время как транскрипты Lpx-2 на стадии зрелого
зерна отсутствовали (De Simone et al., 2010). Локус Lpx-B1
расположен на коротком плече хромосомы 4B, в кото-
ром обнаружены три тесносцепленных гена: Lpx-B1.1,
Lpx-B1.2 и Lpx-B1.3. Ген Lpx-B1.1 представлен тремя
аллелями – Lpx-B1.1a, Lpx-B1.1b, Lpx-B1.1c (Hessler et
al., 2002; Carrera et al., 2007; Verlotta et al., 2010). Анализ
QTL у твердой пшеницы показал, что 35–54.0 % вариации
активности Lpx объясняется Lpx-B1. Аллель Lpx-B1.1c
отличается делецией в нуклеотидной последовательности
от второго интрона до последнего экзона (Carrera et al.
Липоксигеназа (Lpx) у растений продуцирует активные
формы кислорода, приводящие к деградации каротинои-
дов и обесцвечиванию конечных продуктов, получаемых
из зерна твердой пшеницы (Borrelli et al., 2003). У твердой
пшеницы существуют разные гены Lpx (Borrelli, Trono,
2016). На стадии зрелого зерна у сортов с контрастной
активностью (окислительной способностью) была уста-
новлена различная степень транскрипции генов Lpx-1 и
Lpx-3, в то время как транскрипты Lpx-2 на стадии зрелого
зерна отсутствовали (De Simone et al., 2010). Локус Lpx-B1
расположен на коротком плече хромосомы 4B, в кото-
ром обнаружены три тесносцепленных гена: Lpx-B1.1,
Lpx-B1.2 и Lpx-B1.3. Ген Lpx-B1.1 представлен тремя
аллелями – Lpx-B1.1a, Lpx-B1.1b, Lpx-B1.1c (Hessler et
al., 2002; Carrera et al., 2007; Verlotta et al., 2010). Анализ
QTL у твердой пшеницы показал, что 35–54.0 % вариации
активности Lpx объясняется Lpx-B1. Аллель Lpx-B1.1c
отличается делецией в нуклеотидной последовательности
от второго интрона до последнего экзона (Carrera et al. ,
2007). Этот аллель коррелирует с высоким уровнем жел-
тизны и относительно слабой деградацией пигментов в
макаронных изделиях (Carrera et al., 2007; Verlotta et al.,
2010). Изучение коллекции твердой пшеницы с включе-
нием в нее ландрасов и современных сортов позволило
идентифицировать три гаплотипа: первый включал гены и
аллели – Lpx-B1.3+Lpx-B1.b, второй – LpxB1.2+LpxB1.1a
и третий – LpxB1.2+LpxB1.1c. Эти гаплотипы демон-
стрировали, соответственно, высокий, средний и низкий
уровни функциональных транскриптов Lpx-B1 и фермен-
тативной активности в созревшем зерне.
,
2007). Этот аллель коррелирует с высоким уровнем жел-
тизны и относительно слабой деградацией пигментов в
макаронных изделиях (Carrera et al., 2007; Verlotta et al.,
2010). Изучение коллекции твердой пшеницы с включе-
нием в нее ландрасов и современных сортов позволило
идентифицировать три гаплотипа: первый включал гены и
аллели – Lpx-B1.3+Lpx-B1.b, второй – LpxB1.2+LpxB1.1a
и третий – LpxB1.2+LpxB1.1c. Эти гаплотипы демон-
стрировали, соответственно, высокий, средний и низкий
уровни функциональных транскриптов Lpx-B1 и фермен-
тативной активности в созревшем зерне.
Известны коммерческие сорта-носители разных ге-
нов, Lpx-B1.1, Lpx-B1.2, Lpx-B1.3, и аллелей в локусе
Lpx-B1.1. В частности, сорта Kofa и Aureo содержат
аллель Lpx-B1.1c. Описаны возможности накопления
каротиноидов и генетика липоксигеназы у сортов: Primadur (имеет высокое содержание каротиноидов и высокую
активность липоксигеназы), Cosmodur (высокая концентрация
каротиноидов, низкая активность липоксигеназы –
Lpx-B1. 1с), Trinakria (низкая концентрация каротинои-
дов, высокая активность липоксигеназы), Creso (низкое
содержание каротиноидов и низкая активность липокси-
геназы – Lpx-B1.1с) (De Simone et al., 2010). По данным
A. Verlotta с коллегами (2010) в табл. 2 показано рас-
пределение сортов разных периодов селекции (до 1971 г.
и в 1971–2005 гг.) по их принадлежности к различным
гаплотипам. Вызывает внимание увеличение частоты
встречаемости второго и третьего гаплотипов в сортах
последних периодов селекции. Необходимо подчеркнуть,
что этот результат достигнут селекционерами без по-
нимания и учета в селекционных процедурах генетики
окислительных процессов в зерне.
1с), Trinakria (низкая концентрация каротинои-
дов, высокая активность липоксигеназы), Creso (низкое
содержание каротиноидов и низкая активность липокси-
геназы – Lpx-B1.1с) (De Simone et al., 2010). По данным
A. Verlotta с коллегами (2010) в табл. 2 показано рас-
пределение сортов разных периодов селекции (до 1971 г.
и в 1971–2005 гг.) по их принадлежности к различным
гаплотипам. Вызывает внимание увеличение частоты
встречаемости второго и третьего гаплотипов в сортах
последних периодов селекции. Необходимо подчеркнуть,
что этот результат достигнут селекционерами без по-
нимания и учета в селекционных процедурах генетики
окислительных процессов в зерне.
Note. The “+” sign indicates the belonging to the haplotype.
Второй по значимости воздействия на цвет конечных
продуктов катаболический фермент – полифенолоксида-
за (РРО). Активность фермента контролируют два гена,
идентифицированных на гомеологичных хромосомах
второй группы, 2А и 2В (Jimenez, Dubcovsky, 1999; Simeone
et al. , 2002; Watanabe et al., 2004, 2006). В частности,
R. Simeone с коллегами (2002) сообщили о значительном
генетическом эффекте на активность РРО локуса на длин-
ном плече 2А хромосомы. Гены Тс1 и Тс2 в дистальных
частях 2AL и 2BL на расстоянии 46.08 и 40.7 сМ от цен-
тромеры картированы N. Watanabe с коллегами (2004).
Изучение RIL, полученной от скрещивания сортов Jennah
Khetifa и Cham 1, показало, что локус в хромосоме 2А
обеспечивал 49.1 % активности РРО, низкая активность
сегрегировала с молекулярным маркером Xgwm312@2A
(Watanabe et al., 2006).
, 2002; Watanabe et al., 2004, 2006). В частности,
R. Simeone с коллегами (2002) сообщили о значительном
генетическом эффекте на активность РРО локуса на длин-
ном плече 2А хромосомы. Гены Тс1 и Тс2 в дистальных
частях 2AL и 2BL на расстоянии 46.08 и 40.7 сМ от цен-
тромеры картированы N. Watanabe с коллегами (2004).
Изучение RIL, полученной от скрещивания сортов Jennah
Khetifa и Cham 1, показало, что локус в хромосоме 2А
обеспечивал 49.1 % активности РРО, низкая активность
сегрегировала с молекулярным маркером Xgwm312@2A
(Watanabe et al., 2006).
Два гомологичных семейства РРО были картированы
по второй гомеологичной группе хромосом и названы
РРО-1 (РРО-A1 и РРО-В1) и РРО-2 (РРО-А2 и РРО-В2)
(Beecher et al., 2012). Использование маркера, специфич-
ного для мягкой пшеницы (РРО18), для анализа твердой
пшеницы позволило обнаружить четыре аллеля РРО-А1:
РРО-А1b, PPO-A1f, PPO-A1e, PPO-A1g (He et al., 2009).
Используя 111 образцов твердой пшеницы, F. Taranto с
коллегами (2012) определили связь различных аллелей
РРО-А1 с уровнем активности фермента. Аллель РРО-А1f
был связан с высокой, тогда как РРО-А1b и РРО-А1g –
с низкой активностью фермента. Эти ученые разработали
также новый маркер (MG18), способный обнаружить те
же аллели, что и при помощи маркера РРО18, но более
эффективно и с более низкой вариабельностью актив-
ности РРО внутри каждой группы сортов, несущих один
и тот же аллель (Taranto et al., 2012). В маркер-ассоции-
рованной селекции с целью снижения активности РРО
целесообразно использовать и паралогичные гены РРО-В1
и РРО-В2 с применением соответствующих маркеров
MG08 и MG33, предложенных F. Taranto с коллегами
(2015). Гены PPO-B1 и PPO-B2 были расположены на
расстоянии 11.4 сМ от центромеры на хромосоме 2BL.
Скрининг коллекции твердой пшеницы с помощью мар-
керов MG08 и MG33 позволил идентифицировать четыре
и два аллеля соответственно, включая три новых аллеля
гена PPO-B1: PPO- B1b, PPO-B1c и PPO-B1d, и один но-
вый аллель PPO-B2 – PPO-B2d. Маркер MG33 способен распознавать два аллеля РРО-В2, связанных с высокой
(РРО-В2d) и низкой (РРО-В2а) активностью фермента
(Taranto et al.
Аллель РРО-А1f
был связан с высокой, тогда как РРО-А1b и РРО-А1g –
с низкой активностью фермента. Эти ученые разработали
также новый маркер (MG18), способный обнаружить те
же аллели, что и при помощи маркера РРО18, но более
эффективно и с более низкой вариабельностью актив-
ности РРО внутри каждой группы сортов, несущих один
и тот же аллель (Taranto et al., 2012). В маркер-ассоции-
рованной селекции с целью снижения активности РРО
целесообразно использовать и паралогичные гены РРО-В1
и РРО-В2 с применением соответствующих маркеров
MG08 и MG33, предложенных F. Taranto с коллегами
(2015). Гены PPO-B1 и PPO-B2 были расположены на
расстоянии 11.4 сМ от центромеры на хромосоме 2BL.
Скрининг коллекции твердой пшеницы с помощью мар-
керов MG08 и MG33 позволил идентифицировать четыре
и два аллеля соответственно, включая три новых аллеля
гена PPO-B1: PPO- B1b, PPO-B1c и PPO-B1d, и один но-
вый аллель PPO-B2 – PPO-B2d. Маркер MG33 способен распознавать два аллеля РРО-В2, связанных с высокой
(РРО-В2d) и низкой (РРО-В2а) активностью фермента
(Taranto et al. , 2015).
, 2015).
Значительное влияние на признаки YI, YPC и цвет ма- каронных изделий оказывает альдегидоксидаза (АО; ЕС 1.2.3.1) (Colasuonno et al., 2017b). Три изоформы фермента пшеницы, АО1, АО2, АО3, были локализованы на 2-, 5-, 7-хромосомных группах соответственно. Третий ген альдегидоксидазы, АО-А3, расположенный на хромосо- ме 7AL, связан с QTL, влияющим на признаки YI и YPC (Colasuonno et al., 2017b). Эксперименты с двумя сортами, Cicco (низкое содержание каротиноидов в зерне) и Svevo (высокое содержание каротиноидов), с применением метода qRT-PCR выявили высокий уровень экспрессии гена АО-АЗ у первого сорта и низкий у второго, что подтверждает отрицательный эффект продуктов гена в период накопления каротиноидов. На основе SNP данных, со- ответствующих изоформе фермента, на хромосоме 7AL картирован маркер IWB59875, который предложен для процедур маркерной селекции для повышения содержа- ния каротиноидов в зерне и цвета макаронных изделий (Colasuonno et al., 2017b).
Пероксидазы – ферменты, катализирующие общую
реакцию: ROOH + H2O2 = ROH + H2O + 1/2O2 (Feillet et al.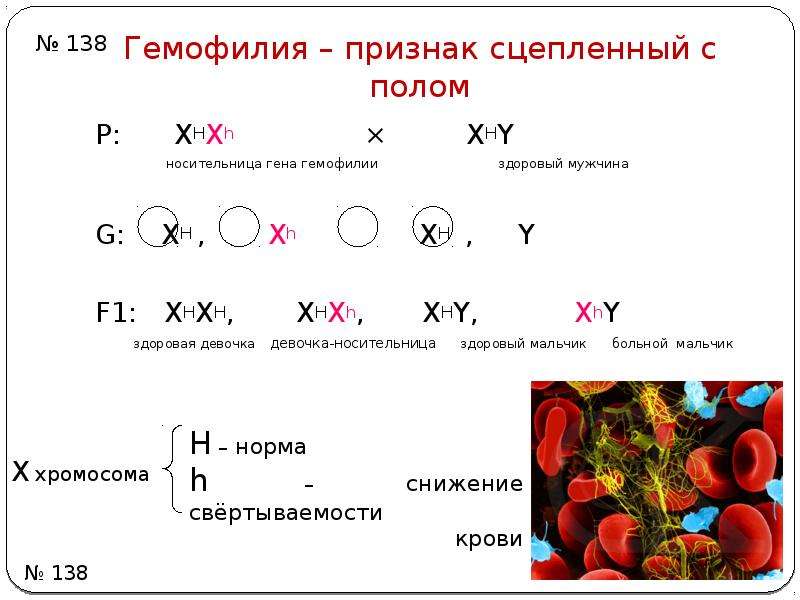 ,
2000). Паста, произведенная из зерна сортов с высоким
уровнем активности пероксидазы, имеет буро-корич-
невый цвет и низкие потребительские качества (Sisson,
2008). В то же время в процессе изготовления макарон
пероксидаза не проявляется в связи с недоступностью
перекиси водорода (Ficco et al., 2014). Активность фер-
ментов этой группы в зерне твердой пшеницы значительно
меньше, чем в зерне мягкой пшеницы. В связи с этим
большинство исследований по их изучению проведено
на мягкой пшенице. В литературе отсутствуют сведения
о специфических маркерах в геноме твердой пшеницы,
связанных с QTL или генами низкой активности. В ряде
публикаций отмечено, что гены пероксидазы расположе-
ны в гомеологичных хромосомах групп 1, 2, 3, 4 и 7 (Liu
et al., 1990; Wei et al., 2015). В зерне твердой пшеницы
установлено функционирование 12 изоформ перокси-
дазы, различающихся по активности в период налива,
созревания и прорастания зерна. Некоторые изоформы
имеют специфическую локализацию в зерне – перикарп,
эндосперм, зародыш.
,
2000). Паста, произведенная из зерна сортов с высоким
уровнем активности пероксидазы, имеет буро-корич-
невый цвет и низкие потребительские качества (Sisson,
2008). В то же время в процессе изготовления макарон
пероксидаза не проявляется в связи с недоступностью
перекиси водорода (Ficco et al., 2014). Активность фер-
ментов этой группы в зерне твердой пшеницы значительно
меньше, чем в зерне мягкой пшеницы. В связи с этим
большинство исследований по их изучению проведено
на мягкой пшенице. В литературе отсутствуют сведения
о специфических маркерах в геноме твердой пшеницы,
связанных с QTL или генами низкой активности. В ряде
публикаций отмечено, что гены пероксидазы расположе-
ны в гомеологичных хромосомах групп 1, 2, 3, 4 и 7 (Liu
et al., 1990; Wei et al., 2015). В зерне твердой пшеницы
установлено функционирование 12 изоформ перокси-
дазы, различающихся по активности в период налива,
созревания и прорастания зерна. Некоторые изоформы
имеют специфическую локализацию в зерне – перикарп,
эндосперм, зародыш. Наиболее важная изоформа, Р-5,
расположена в эндосперме и оказывает значимое влияние
на потемнение (коричневатость) макаронных изделий
(Feillet et al., 2000). При использовании полногеномного
секвенирования и нулли-тетрасомных линий сорта Чайниз
Спринг обнаружены и локализованы два гена, TaPod-A2
и TaPod-D1, на хромосомах 7AS и 7DS. Анализ SNP выявил
для двух аллелей локуса TaPod-D1 два функциональ-
ных маркера, POD-7D1 и POD-7D6, с высокой и низкой
активностью пероксидазы соответственно (Geng et al.,
2019). Эти данные, полученные на мягкой пшенице, с
учетом ортологичности геномов А и В в перспективе
можно адаптировать к проблемам маркерной селекции
твердой пшеницы. В отечественной литературе известна
публикация А.А. Вьюшкова (2004), в которой приведены
значительные сортовые различия твердой пшеницы по
активности пероксидазы в крупке (эндосперме).
Наиболее важная изоформа, Р-5,
расположена в эндосперме и оказывает значимое влияние
на потемнение (коричневатость) макаронных изделий
(Feillet et al., 2000). При использовании полногеномного
секвенирования и нулли-тетрасомных линий сорта Чайниз
Спринг обнаружены и локализованы два гена, TaPod-A2
и TaPod-D1, на хромосомах 7AS и 7DS. Анализ SNP выявил
для двух аллелей локуса TaPod-D1 два функциональ-
ных маркера, POD-7D1 и POD-7D6, с высокой и низкой
активностью пероксидазы соответственно (Geng et al.,
2019). Эти данные, полученные на мягкой пшенице, с
учетом ортологичности геномов А и В в перспективе
можно адаптировать к проблемам маркерной селекции
твердой пшеницы. В отечественной литературе известна
публикация А.А. Вьюшкова (2004), в которой приведены
значительные сортовые различия твердой пшеницы по
активности пероксидазы в крупке (эндосперме).
Результаты и перспективы применения MAS в селекции твердой пшеницы
Несмотря на большое количество работ по локализации и
маркированию QTL, связанных с высокой концентрацией
каротиноидов в зерне и крупке, результаты прямой про-
верки применения MAS (селекция с помощью маркеров)
представлены ограниченно. Patil с коллегами (2018) со-
общили о высокой эффективности
использования маркера
Psy-A1SSRe, сцепленного с QTL и локусом Psy-A1, на
хромосоме 7AL. Маркер
Psy-1SSR разработан на основе
вариаций в промоторной
области PSY-1, он позволяет
идентифицировать восемь аллелей
Psy-A1 и семь аллелей
Psy-B1 одновременно. Маркер Psy-A1SSRe, расположен-
ный в 7AL в большом QTL для YPC, вместе с ранее уста-
новленным маркером Qyp.macs-7A был идентифицирован
в популяции RIL-PDW233/Bhalegaon 4. Родительский
сорт PDW 233 в этой популяции является носителем QTL
высокой концентрации пигментов. Эти маркеры были
применены для улучшения индийских сортов MACS 3125
и HI8498 с низкой концентрацией пигментов (3.57 и
3.26 ppm соответственно), которые были взяты в качестве
рекуррентных родителей в скрещиваниях с PDW 233 –
донором QTL для YPC с высокой концентрацией пиг-
ментов (8.36 ppm). Селекционные линии, полученные
с применением методов MAS на основе MACS 3125 и
HI 8498, показали значительное увеличение YPC: 6.
Patil с коллегами (2018) со-
общили о высокой эффективности
использования маркера
Psy-A1SSRe, сцепленного с QTL и локусом Psy-A1, на
хромосоме 7AL. Маркер
Psy-1SSR разработан на основе
вариаций в промоторной
области PSY-1, он позволяет
идентифицировать восемь аллелей
Psy-A1 и семь аллелей
Psy-B1 одновременно. Маркер Psy-A1SSRe, расположен-
ный в 7AL в большом QTL для YPC, вместе с ранее уста-
новленным маркером Qyp.macs-7A был идентифицирован
в популяции RIL-PDW233/Bhalegaon 4. Родительский
сорт PDW 233 в этой популяции является носителем QTL
высокой концентрации пигментов. Эти маркеры были
применены для улучшения индийских сортов MACS 3125
и HI8498 с низкой концентрацией пигментов (3.57 и
3.26 ppm соответственно), которые были взяты в качестве
рекуррентных родителей в скрещиваниях с PDW 233 –
донором QTL для YPC с высокой концентрацией пиг-
ментов (8.36 ppm). Селекционные линии, полученные
с применением методов MAS на основе MACS 3125 и
HI 8498, показали значительное увеличение YPC: 6. 16–7.7
и 5.0–7.46 ppm соответственно. В настоящее время MAS
используется в CIMMYT и в Канаде для отбора селекци-
онных линий с низкой активностью липоксигеназы при
помощи маркера LOXA, нацеленного на аллель Lpx-B1.1c
(Randhawa et al., 2013; Dreisigacker et al., 2016; N’Diaye
et al., 2017, 2018).
16–7.7
и 5.0–7.46 ppm соответственно. В настоящее время MAS
используется в CIMMYT и в Канаде для отбора селекци-
онных линий с низкой активностью липоксигеназы при
помощи маркера LOXA, нацеленного на аллель Lpx-B1.1c
(Randhawa et al., 2013; Dreisigacker et al., 2016; N’Diaye
et al., 2017, 2018).
В России во всех лабораториях, осуществляющих селекцию
на увеличение концентрации каротиноидов в зерне,
индекса желтизны семолины и конечных продуктов,
применяются методы традиционной селекции. За период
научной селекции и особенно за последние 30 лет эти
признаки были улучшены. Сорта, созданные на первых
этапах, – Мелянопус 69, Гордеиформе 432, Мелянопус
26,
Гордеиформе 179, Гордеиформе 675, накапливают в зерне
3.6–5.0 ppm каротиноидных пигментов. Сорта, широко
возделывавшиеся в 60-80-х годах ХХ в., – Безенчук-
ская 105, Харьковская 46, Безенчукская 139, – превышают
этот уровень незначительно (~5 %) (Мясникова и др.,
2019). Положительные изменения наблюдались у сортов
Светлана (1987 – год включения в реестр) и Саратовская
золотистая (1993), которые накапливали 6.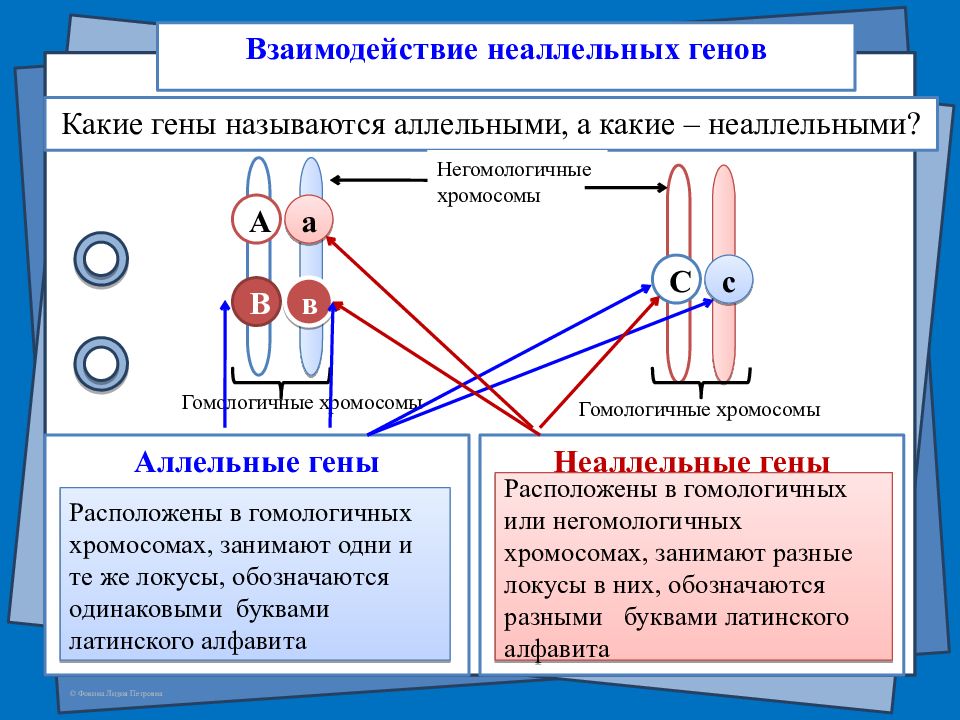 0–7.0 ppm
пигментов в зерне (Васильчук,
2001). Среди современных
сортов яровой твердой пшеницы заметно выделяются по
содержанию каротиноидов в зерне Безенчукская золоти-
стая (8.5–9.0 ppm) и Безенчукская крепость (7.5–8.5 ppm).
Весь набор современных сортов образует непрерывный
ряд изменчивости с шагом в 10–15 % и разницей между
крайними вариантами в 200 % (Безенчукская золотистая –
Алтайская нива). Изучение отечественных сортов разных
этапов селекции в одиннадцати экологических средах
позволило установить, что фенотипическое варьирование
признака определяется генотипической и средовой вариансами
с незначительными эффектами генотип-средовых
взаимодействий. По результатам кластерного анализа параметров адаптивности, стабильности и отзывчивости,
сорта отчетливо распределялись на кластеры. В группу
с оптимальным сочетанием величины, стабильности и
отзывчивости признака вошли Безенчукская золотистая,
Безенчукская крепость, Безенчукская 210, Саратовская зо-
лотистая. Эти генотипы рекомендуется использовать для
создания рекомбинантных инбредных линий с целью мар-
кирования QTL, контролирующих синтез каротиноидов
в зерне твердой пшеницы, и организации на этой основе
маркер-опосредованной технологии селекции (Мясникова
и др.
0–7.0 ppm
пигментов в зерне (Васильчук,
2001). Среди современных
сортов яровой твердой пшеницы заметно выделяются по
содержанию каротиноидов в зерне Безенчукская золоти-
стая (8.5–9.0 ppm) и Безенчукская крепость (7.5–8.5 ppm).
Весь набор современных сортов образует непрерывный
ряд изменчивости с шагом в 10–15 % и разницей между
крайними вариантами в 200 % (Безенчукская золотистая –
Алтайская нива). Изучение отечественных сортов разных
этапов селекции в одиннадцати экологических средах
позволило установить, что фенотипическое варьирование
признака определяется генотипической и средовой вариансами
с незначительными эффектами генотип-средовых
взаимодействий. По результатам кластерного анализа параметров адаптивности, стабильности и отзывчивости,
сорта отчетливо распределялись на кластеры. В группу
с оптимальным сочетанием величины, стабильности и
отзывчивости признака вошли Безенчукская золотистая,
Безенчукская крепость, Безенчукская 210, Саратовская зо-
лотистая. Эти генотипы рекомендуется использовать для
создания рекомбинантных инбредных линий с целью мар-
кирования QTL, контролирующих синтез каротиноидов
в зерне твердой пшеницы, и организации на этой основе
маркер-опосредованной технологии селекции (Мясникова
и др. , 2019).
, 2019).
Заключение
Результаты исследований, рассмотренные в этой статье,
показывают реализованные за последние двадцать лет
цели в понимании биохимических, генетических ме-
ханизмов регулирования метаболизма каротиноидных
пигментов в твердой пшенице. Данные, полученные на
разнообразном растительном материале с применением
современных классов ДНК-маркеров, и согласованные в
разнообразных экспериментах генетические карты по-
зволили выявить наиболее важные гены, участвующие в
контроле биосинтеза, накопления и катаболизма кароти-
ноидов. Наиболее изученными и подтвержденными в ряде
экспериментов QTL являются те, которые расположены
на хромосомах 3AS (связаны с геном LCYE), 7AL и 7BL
(оба тесно связаны с аллелями PSY 1). Перспективно
для MAS применение маркера LOXA, нацеленного на
аллель Lpx-B1.1c для снижения активности липоксиге-
назы. Технология MAS с использованием известных для
этих QTL маркеров отличается от методов традиционной
фенотипической селекции высокой производительностью
и эффективностью. В то же время в ряде исследований
были представлены данные о том, что большинство QTL
не могут быть широко использованы в процедурах MAS,
что связано с уровнем их валидации в фенотипических
соотношениях в локально адаптированных группах се-
лекционного материала (Pozniak et al., 2012).
В то же время в ряде исследований
были представлены данные о том, что большинство QTL
не могут быть широко использованы в процедурах MAS,
что связано с уровнем их валидации в фенотипических
соотношениях в локально адаптированных группах се-
лекционного материала (Pozniak et al., 2012).
Другие QTL, представленные и охарактеризованные
в обзоре, требуют дополнительных исследований для
идентификации генов-кандидатов, участвующих в накоплении/
деградации каротиноидов. В перспективе можно
ожидать широкое внедрение методов геномной селекции
в программы улучшения цвета конечных продуктов из
твердой пшеницы. С учетом аддитивных эффектов генов,
контролирующих уровень пигментов и превалирование
генотипа признака над средой, применение методов геномной
селекции позволит ускорить селекционный про-
цесс на основе не только молекулярной идентификации
функционирующих в популяции необходимых QTL, но и
целенаправленного получения трансгрессий в результате
пирамидирования различных генов. В связи с этим даль-
нейший акцент в селекционно-генетических мероприя-
тиях будет сделан на анализе генетической изменчивости
в коллекциях зародышевой плазмы твердой пшеницы и
мутантных популяциях. Перспективны методы редакти-
рования генома с использованием CRIS-Cas9, если их
применять, исходя из понимания функций гомеологичных
генов с аддитивным эффектом (Patil et al., 2018). Характе-
ристика каждого гена позволит вырабатывать стратегии диверсификации генетической системы каротиноидных
пигментов и расширить существующие вариации, до-
ступные селекционерам.
Перспективны методы редакти-
рования генома с использованием CRIS-Cas9, если их
применять, исходя из понимания функций гомеологичных
генов с аддитивным эффектом (Patil et al., 2018). Характе-
ристика каждого гена позволит вырабатывать стратегии диверсификации генетической системы каротиноидных
пигментов и расширить существующие вариации, до-
ступные селекционерам.
Непосредственное применение маркеров QTL, апробированных
на растительном материале зарубежных стран
в России в процедурах MAS и геномной селекции, воз-
можно, если в качестве доноров использовать сорта за-
рубежной селекции, несущие соответствующие QTL и
маркеры.
При этом маркеры должны быть эффектив-
ными в экологических зонах российских селекционных
центров. Однако и в этом случае могут быть проблемы
преодоления недостаточной адаптивности привлекаемых
в качестве исходного материала генотипов. Селекция на
увеличение концентрации пигментов в зерне и продуктах
его переработки в России, безусловно, может использо-
вать генетический материал и маркеры, разработанные
в иностранных центрах, но базироваться она должна на
отечественном исходном материале и адаптированных
к нему технологиях маркер-опосредованной селекции.
Conflict of interest
The authors declare no conflict of interest.
References
Васильчук Н.С. Селекция яровой твердой пшеницы. Саратов, 2001. [Vasil’chuk N.S. Spring Durum Wheat Breeding. Saratov, 2001. (in Russian)]
Васильчук Н.С., Гапонов С.Н., Еременко Л.В., Паршикова Т.М., Попова В.М., Шутарева Г.М., Куликова В.А. Селекция твердой яровой пшеницы на высокое содержание каротиноидов в зерне. В: Сборник научных трудов ГНУ НИИСХ Юго-Востока Рос- сельхозакадемии. Саратов: ООО Ракурс, 2009;89-90. [Vasil’chuk N.S., Gaponov S.N., Eremenko L.V., Parshikova T.M., Popova V.M., Shutareva G.M., Kulikova V.A. Breeding of spring durum wheat for carotenoid-rich grain. In: Collection of Scientific Papers of the South-East Agricultural Research Institute of the Russian Agricultural Academy. Saratov: Rakurs Company, 2009;89-90. (in Russian)]
Вьюшков А.А. Селекция яровой пшеницы в Среднем Поволжье.
Самара: ООО «СамЛюкс», 2004.
[Vyushkov A.A. Spring Wheat Breeding in the Middle Volga region.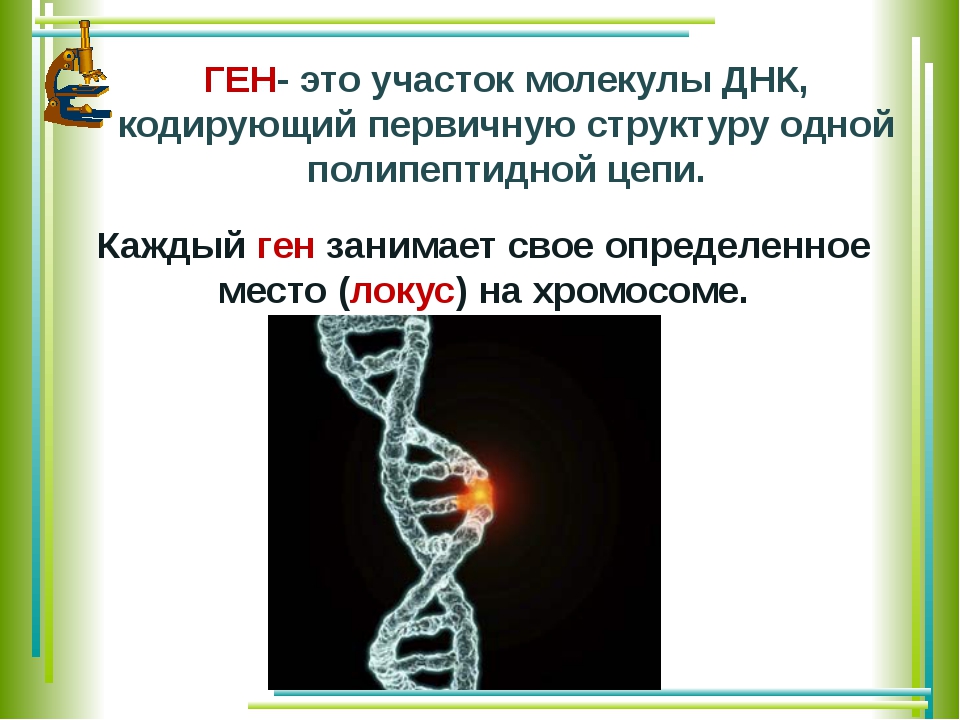 Samara: SamLyuks Company, 2004. (in Russian)]
Samara: SamLyuks Company, 2004. (in Russian)]
Гапонов С.Н., Попова В.М., Шутарева Г.И., Цетва Н.М., Цет- ва И.С., Паршикова Т.М. Получение новых источников для се- лекции яровой твердой пшеницы – гарантия создания стабиль- ных стрессоустойчивых сортов. Аграр. вестн. Юго-Востока. 2018;3:30-31. [Gaponov S.N., Popova V.M., Shutareva G.I., Tsetva N.M., Tsetva I.S., Parshikova T.M. Obtaining new sources for breeding spring durum wheat warrants the raise of stable stress-resistant varieties. Agrarnyy Vestnik Yugo-Vostoka = Agrarian Bulletin of the South- East. 2018;3:30-31. (in Russian)]
Кретович В.Л. Биохимия растений. М.: Высш. шк., 1986. [Kretovich V.L. Plant Biochemistry. Moscow: Vysshaya shkola, 1986. (in Russian)]
Мальчиков П.Н. Селекция яровой твердой пшеницы в Среднем Поволжье. Кинель, 2009. [Malchikov P.N. Spring Durum Wheat Breeding in the Middle Volga Region. Kinel, 2009. (in Russian)]
Мясникова М.Г., Мальчиков П.Н., Шаболкина Е.Н., Анисимки-
на Н.В., Розова М. А., Чахеева Т.В. Результаты селекции твер-
дой пшеницы в России на содержание каротиноидных пигмен-
тов в зерне. Зерновое хозяйство России. 2019;6(66):37-40. DOI
10.31367/2079-8725-2019-66-6-37-40.
[Myasnikova M.G., Malchikov P.N., Shabolkina E.N., Anisimkina
N.V., Rozova M.A., Chakheeva T.V. The results of durum wheat
breeding in Russia for carotenoid pigments content in kernels. Zernovoe
Khozjaistvo Rossii = Grain Economy of Russia. 2019;6(66):
37-40. DOI 10.31367/2079-8725-2019-66-6-37-40. (in Russian)]
А., Чахеева Т.В. Результаты селекции твер-
дой пшеницы в России на содержание каротиноидных пигмен-
тов в зерне. Зерновое хозяйство России. 2019;6(66):37-40. DOI
10.31367/2079-8725-2019-66-6-37-40.
[Myasnikova M.G., Malchikov P.N., Shabolkina E.N., Anisimkina
N.V., Rozova M.A., Chakheeva T.V. The results of durum wheat
breeding in Russia for carotenoid pigments content in kernels. Zernovoe
Khozjaistvo Rossii = Grain Economy of Russia. 2019;6(66):
37-40. DOI 10.31367/2079-8725-2019-66-6-37-40. (in Russian)]
Чесноков Ю.В., Артемьева А.М. Ассоциативное картирование у растений. С.-х. биология. 2011;46(5):3-16. [Chesnokov Yu.V., Artemyeva A.M. Associative mapping in plants. Selskokhozyaystvennaya Biologiya = Agricultural Biology. 2011; 46(5):3-16. (in Russian)]
AACC International. AACC International Official Method 14-50.01. In: Approved Methods of the American Association of Cereal Chemists. Tenth ed. (MN, USA: St. Paul), 2013.
Al-Babili S., Bouwmeester H.J. Strigolactones, a novel carotenoidderived
plant hormone. Annu. Rev. Plant Biol. 2015;66(1):161-186.
DOI 10.1146/annurev-arplant-043014-14759.
Annu. Rev. Plant Biol. 2015;66(1):161-186.
DOI 10.1146/annurev-arplant-043014-14759.
Beecher B.S., Carter A.H., See D.R. Genetic mapping of new seed-expressed polyphenol oxidase genes in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2012;124:1463-1473. DOI 10.1007/s00122- 012-1801-2.
Blanco A., Colasuonno P., Gadaleta A., Mangini G., Schiavulli A., Simeone R., Digesu A.M., De Vita P., Mastrangelo A.M., Cativelly L. Quantitative trait loci for yellow pigment concentration and individual carotenoid compounds in durum wheat. J. Cereal Sci. 2011; 54(2):255-264. DOI 10.1016/j.jcs.2011.07.002.
Borrelli G.M., De Leonardis A.M., Fares C., Platani C., Di Fonzo N. Effects of modifyed processing conditions on oxidative properties of semolina dough and pasta. Cereal Chem. 2003;80:225-231. DOI 10.1094/cchem.2003.80.2.225.
Borelli G.M., Troccoli А., Di Fonzo N., Fares C. Durum wheat lipoxygenase
activity and other parameters that affect pasta color. Cereal
Chem. 1999;76:335-340. DOI 10.1094/CCHEM.1999.76.3.335.
DOI 10.1094/CCHEM.1999.76.3.335.
Borrelli G.M., Trono D. Molecular approaches to genetically improve the accumulation of health-promoting secondary metabolites in staple crops-A case study: the Lipoxygenase-B1 genes and regulation of the carotenoid content in pasta products. Int. J. Mol. Sci. 2016;17:1177. DOI 10.3390/ijms17071177.
Brandolini A., Hidalgo A., Gabriele S., Heun M. Chemical composition of wild and feral diploid wheats and their bearing on domesticated wheats. J. Cereal Sci. 2015;63:122-127. DOI 10.1016/j.jcs.2015. 03.005.
Brandolini A., Hidalgo A., Moscaritolo S. Chemical composition and pasting properties of einkorn (Triticum monococcum L. subsp. monococcum) whole meal flour. J. Cereal Sci. 2008;47:599-609. DOI 10.1016/j.jcs.2007.07.005.
Campos K.M., Royo C., Schulthess A., Villegas D., Matus I., Ammar
K., Schwember A.R. Association of phytoene synthase Psy1- A1
and Psy1-B1 allelic variants with semolina yellowness in durum
wheat (Triticum turgidum L. var. durum). Euphytica. 2016;207:109-
117. DOI 10.1007/s10681-015-1541-x.
var. durum). Euphytica. 2016;207:109-
117. DOI 10.1007/s10681-015-1541-x.
Carrera A., Echenique V., Zhang W., Helguera M., Manthey F., Schrager A., Picca A., Cervigna G., Dubcovsky J. A deletion at the Lpx- B1 locus is associated with low lipoxygenase activity and improved pasta color in durum wheat (Triticum turgidum ssp. durum). J. Cereal Sci. 2007;45:67-77. DOI 10.1016/j.jcs.2006.07.001.
Cazzonelli C.I., Pogson B.J. Source to sink: regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010;15:266-274. DOI 10.1016/j.tplants.2010.02.003.
Clarke J.M., Clarke F.R., McCaig T.N. Heritability of content in three durum wheat crosses. In: Proc. Ninth Int. Wheat Genetics Symposium. Saskatoon, 1998;2:182-184.
Colasuonno P., Gadaleta A., Giancaspro A., Nigro D., Giove S., Incerti
O., Mangini G., Signorile A., Simeone R., Blanco A. Development
of a high-density SNP-based linkage map and detection of yellow
pigment content QTLs in durum wheat. Mol. Breed. 2014;34:1563-
1578.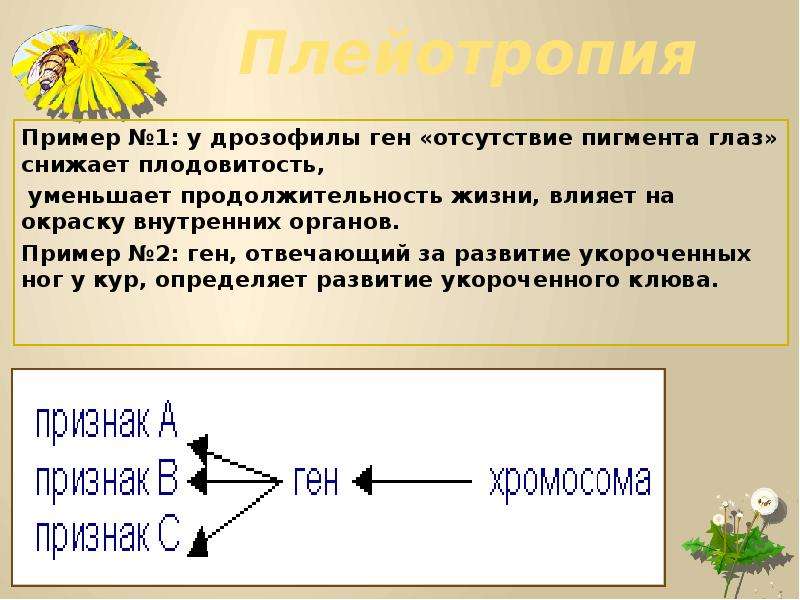 DOI 10.1007/s11032-014-0183-3.
DOI 10.1007/s11032-014-0183-3.
Colasuonno P., Lozito M.L., Marcotuli I., Nigro D., Giancaspro A., Mangini G., De Vita P., Mastrangelo A.M., Pecchioni N., Houston K., Simeone R., Gadaleta A., Blanco A. The carotenoid biosynthetic and catabolic genes in wheat and their association with yellow pigments. BMC Genomics. 2017a;18:122. DOI 10.1186/s12864- 016-3395-6.
Colasuonno P., Marcotuli I., Blanco A., Maccaferri M., Condorelli G.E., Tuberosa R., Parada R., de Camargo A.C., Schwember A.R., Gadaleta A. Carotenoid pigment content in durum wheat (Triticum turgidum L. var durum): an overview of quantitative trait loci and candidate genes. Front. Plant Sci. 2019;10:1347. DOI 10.3389/fpls. 2019.01347.
Colasuonno P., Marcotuli I., Lozito M. L., Simeone R., Blanco A., Gadaleta A. Characterization of aldehyde oxidase (AO) genes involved in the accumulation of carotenoid pigments in wheat grain. Front. Plant Sci. 2017b;8:863. DOI 10.3389/fpls.2017.00863.
Cong L., Wang C., Li Z. , Chen L., Yang G., Wang Y., He G. cDNA
cloning and expression analysis of wheat (Triticum aestivum L.)
phytoene and ζ-carotene desaturase genes. Mol. Biol. Rep. 2010;37:
3351-3361. DOI 10.1007/s11033-009-9922-7.
, Chen L., Yang G., Wang Y., He G. cDNA
cloning and expression analysis of wheat (Triticum aestivum L.)
phytoene and ζ-carotene desaturase genes. Mol. Biol. Rep. 2010;37:
3351-3361. DOI 10.1007/s11033-009-9922-7.
Crawford A.C., Francki M.G. Chromosomal location of wheat genes of the carotenoid biosynthetic pathway and evidence for a catalase gene on chromosome 7A functionally associated with flour b* colour variation. Mol. Genet. Genomics. 2013;288:483-493. DOI 10.1007/ s00438-013-0767-3.
De Simone V., Menzo V., De Leonardis A.M., Ficco D.B.M., Trono D., Cattivelli L., De Vita P. Different mechanisms control lipoxygenase activity in durum wheat kernels. J. Cereal Sci. 2010;52:121-128. DOI 10.1016/j.jcs.2010.04.003.
Dibari B., Murat F., Chosson A., Gautier V., Poncet C., Lecomte P.,
Mercier I., Berges H., Pont C., Blanco A., Salse J. Deciphering the
genomic structure, function and evolution of carotenogenesis related
phytoene synthases in grasses. BMC Genomics. 2012;13:221. DOI
10.1186/1471-2164-13-221.
DOI
10.1186/1471-2164-13-221.
Digesù A.M., Platani C., Cattivelli L., Mangini G., Blanco A. Genetic variability in yellow pigment components in cultivated and wild tetraploid wheats. J. Cereal Sci. 2009;50:210-218. DOI 10.1016/ j.jcs.2009.05.002.
Dong C.H., Ma Z.Y., Xia X.C., Zhang L.P., He Z.H. Allelic variation at the TaZds-A1 locus on wheat chromosome 2A and development of a functional marker in common wheat. J. Integr. Agric. 2012;11: 1067-1074. DOI 10.1016/S2095-3119(12)60099-9.
Dreisigacker S., Sehgal D., Reyes Jaimez A., Luna Garrido B., Muñoz Zavala S., Núñez Ríos C. CIMMYT Wheat Molecular Genetics: Laboratory Protocols and Applications to Wheat Breeding. Mexico: CIMMYT, 2016.
Elouafi I., Nachit M.M., Martin L.M. Identification of a microsatellite on chromosome 7B showing a strong linkage with yellow pigment in durum wheat (Triticum turgidum L. var. durum). Hereditas. 2001;135:255-261. DOI 10.1111/j.1601-5223.2001.t01-1-00255.x.
Feillet P., Autran J. -C., Icard-Vernière C. Mini review pasta brownness:
An assessment. J. Cereal Sci. 2000;32:215-233. DOI 10.1006/jcrs.
2000.0326.
-C., Icard-Vernière C. Mini review pasta brownness:
An assessment. J. Cereal Sci. 2000;32:215-233. DOI 10.1006/jcrs.
2000.0326.
Ficco D.B.M., Mastrangelo A.M., Trono D., Borrelli G.M., De Vita P., Fares C., Beleggia R., Platani C., Papa R. The colours of durum wheat: a review. Crop Pasture Sci. 2014;65(1):1-15. DOI 10.1071/ cp13293.
Fiedler J.D., Salsman E., Liu Y., Michalak de Jiménez M., Hegstad J.B., Chen B., Manthey F.A., Chao S., Xu S., Elias M.E., Li X. Genomewide association and prediction of grain and semolina quality traits in durum wheat breeding populations. Plant Genome. 2017;10(3): 1-12. DOI 10.3835/plantgenome2017.05.0038.
Fu B.X., Chiremba C., Pozniak C.J., Wang K., Nam S. Total phenolic and yellow pigment contents and antioxidant activities of durum wheat milling fractions. Antioxidants. 2017;6:78. DOI 10.3390/ antiox6040078.
Geng H., Shi J., Fuerst E.P., Wei J., Morris C.F. Phisical mapping of peroxidase
genes and development of functional markers for TaPod-
D1
on bread wheat chromosome 7D. Front. Plant Sci. 2019;10:523.
DOI 10.3389/fpls.2019.00523.
Front. Plant Sci. 2019;10:523.
DOI 10.3389/fpls.2019.00523.
He X.Y., He Z.H., Morris C.F., Xia X.C. Cloning and phylogenetic analysis of polyphenol oxidase genes in common wheat and related species. Genet. Resour. Crop Evol. 2009;56:311. DOI 10.1007/ s10722-008-9365-3.
He X.Y., Zhang Y.L., He Z.H., Wu Y.P., Xiao Y.G., Ma C.X., Xia X.C. Characterization of phytoene synthase 1 gene (Psy1) located on common wheat chromosome 7A and development of a functional marker. Theor. Appl. Genet. 2008;116:213-221. DOI 10.1007/ s00122-007-0660-8.
Hessler T.G., Thomson M.J., Benscher D., Nachit M.M., Sorrells M.E. Association of a lipoxygenase locus, Lpx-B1, with variation in lipoxygenase activity in durum wheat seeds. Crop Sci. 2002;42(5): 1695-1700. DOI 10.2135/cropsci2002.1695.
Howitt C.A., Cavanagh C.R., Bowerman A.F., Cazzonelli C., Rampling
L., Mimica J.L., Pogson B.J. Alternative splicing, activation
of cryptic exons and amino acid substitutions in carotenoid biosynthetic
genes are associated with lutein accumulation in wheat
endosperm. Funct. Integr. Genomics. 2009;9(3):363-376. DOI
10.1007/s10142-009-0121-3ICC.
Funct. Integr. Genomics. 2009;9(3):363-376. DOI
10.1007/s10142-009-0121-3ICC.
ICC Method 152, in Standard Methods of the International Association for Cereal Science and Technology. Detmold, Germany: Verlag Moritz Schäfer ICC, 1990.
Jimenez M., Dubcovsky J. Chromosome location of genes affecting polyphenol oxidase activity in seeds of common and durum wheat. Plant Breed. 1999;118:395-398. DOI 10.1046/j.1439-0523.1999. 00393.x.
Kabbaj H., Sall A.T., Al-Abdallat A., Geleta M., Amri A., Filali-Maltouf A., Belkadi B., Ortiz R., Bassi F.M. Genetic diversity within a Global panel of durum wheat (Triticum durum) landraces and modern germplasm reveals the history of alleles exchange. Front. Plant Sci. 2017;8:1277. DOI 10.3389/fpls.2017.01277.
Ke Q., Kang L., Kim H.S., Xie T., Liu C., Ji C.Y., Kim H.S., Park W.S.,
Ahn M.J., Wang S., Li H., Deng X., Kwak S.S. Down-regulation
of lycopene ε-cyclase expression in transgenic sweetpotato plants
increases the carotenoid content and tolerance to abiotic stress.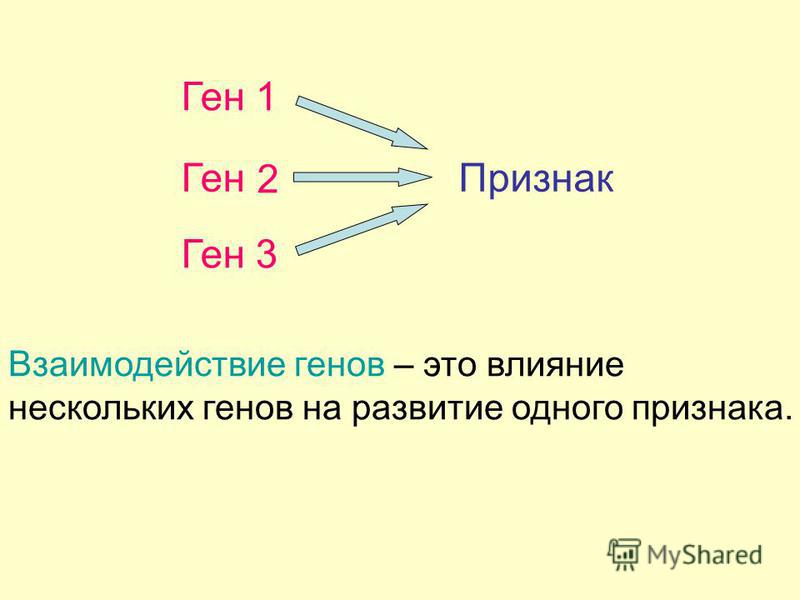 Plant
Sci. 2019;281:52-60. DOI 10.1016/j.plantsci.2019.01.002.
Plant
Sci. 2019;281:52-60. DOI 10.1016/j.plantsci.2019.01.002.
Kean E.G., Bordenave N., Ejeta G., Hamaker B.R., Ferruzzi M.G. Carotenoid bioaccessibility from whole grain and decorticated yellow endosperm sorghum porridge. J. Cereal Sci. 2011;54:450-459. DOI 10.1016/j.jcs.2011.08.010.
Liu C.J., Chao S., Gale M.D. The genetical control of tissue-specific peroxidases, Per-1, Per-2, Per-3, Per-4, and Per-5 in wheat. Theor. Appl. Genet. 1990;79(3):305-313. DOI 10.1007/bf01186072.
Maccaferri M., Sanguineti M.C., Demontis A., El-Ahmed A., Garcia del Moral L., Maalouf F., Nachit M., Nserallah N., Ouabbou H., Rhouma S. Association mapping in durum wheat grown across a broad range of water regimes. J. Exp. Bot. 2011;62(2):409-438. DOI 10.1093/jxb/erq287.
Mazzeo M.F., Di Stasio L., D’Ambrosio C., Arena S., Scaloni A.,
Corneti S., Ceriotti A., Tuberosa R., Siciliano R.A., Picariello G.,
Mamone G. Identification of early represented gluten proteins during
durum wheat grain development. J. Agric. Food Chem. 2017;
65(15):3242-3250. DOI 10.1021/acs.jafc.7b00571.
J. Agric. Food Chem. 2017;
65(15):3242-3250. DOI 10.1021/acs.jafc.7b00571.
N’Diaye A., Haile J.K., Cory A.T., Clarke F.R., Clarke J.M., Knox R.E., Pozniak C.J. Single marker and haplotype-based association analysis of semolina and pasta colour in elite durum wheat breeding lines using a high-density consensus map. PloS One. 2017;12(1): e0170941. DOI 10.1371/journal.pone.0170941.
N’Diaye A., Haile J.K., Nilsen K.T., Walkowiak S., Ruan Y., Singh A.K., Clarke F.R., Clarke J.M., Pozniak C.J. Haplotype loci under selection in Canadian durum wheat germplasm over 60 years of breeding: Association with grain yield, quality traits, protein loss, and plant height. Front. Plant Sci. 2018;9(1589):1-19. DOI 10.3389/ fpls.2018.01589.
Nisar N., Li L., Lu S., Khin N.C., Pogson B.J. Carotenoid metabolism in
plants. Mol. Plant. 2015;8:68-82. DOI 10.1016/j.molp.2014.12.007.
Parker G.D., Chalmers K.J., Rathjen A.J., Langridge P. Mapping
loci associated with flour colour in wheat (Triticum aestivum L.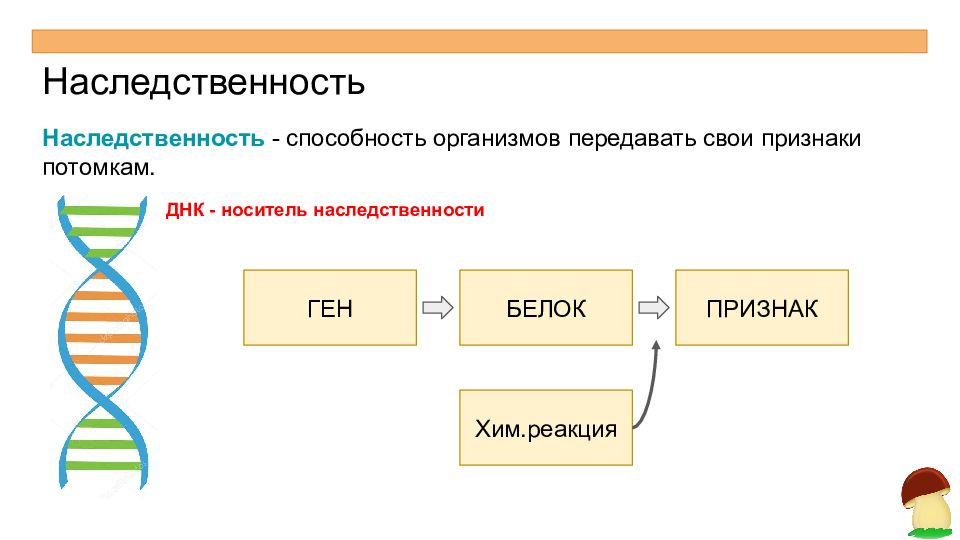 ).
Theor.
Appl. Genet. 1998;97(1-2):238-245. DOI 10.1007/s0012200
50891.
).
Theor.
Appl. Genet. 1998;97(1-2):238-245. DOI 10.1007/s0012200
50891.
Patil R.M., Oak M., Deshpande A., Tamhankar S. Development of a robust marker for Psy-1 homoeologs and its application in improvement of yellow pigment content in durum wheat. Mol. Breed. 2018; 38;136. DOI 10.1007/s11032-018-0895-x.
Patil R., Oak M., Tamhankar S., Sourdille P., Rao V. Mapping and validation of a major QTL for yellow pigment content on 7AL in durum wheat (Triticum turgidum L. ssp. durum). Mol. Breed. 2008; 21(4):485-496. DOI 10.1007/s11032-007-9147-1.
Pozniak C.J., Clarke J.M., Clarke F.R. Potential for detection of marker- trait associations in durum wheat using unbalanced, historical phenotypic datasets. Mol. Breed. 2012;30:1537-1550. DOI 10.1007/ s11032-012-9737-4.
Pozniak C.J., Knox R.E., Clarke F.R., Clark J.M. Identification of QTL
and association of a phytoene synthase gene with endosperm colour
in durum wheat. Theor. Appl. Genet. 2007;114:525-537. DOI
10.1007/s00122-006-0453-5.
Qin X., Zhang W., Dubcovsky J., Tian L. Cloning and comparative analysis of carotenoid beta-hydroxylase genes provides new insights into carotenoid metabolism in tetraploid (Triticum turgidum ssp. durum) and hexaploid (Triticum aestivum) wheat grains. Plant Mol. Biol. 2012;80:631-646. DOI 10.1007/s11103-012-9972-4.
Randhawa H.S., Asif M., Pozniak C., Clarke J.M., Graf R.J., Fox S.L., Humphaeys D.G., Knox R.E., DePaw R.M., Singh A.K., Cuthbert R.D., Hucl P., Spaner D. Application of molecular markers to wheat breeding in Canada. Plant Breed. 2013;132:458-471. DOI 10.1111/pbr.12057.
Ravel C., Dardevet M., Leenhardt F., Bordes J., Joseph J.L., Perretant M.R., Exbrayat F., Poncet C., Balfourier F., Chanliaud E., Charmet G. Improving the yellow pigment content of bred wheat flour by selecting the three homeologous copies of Psy1. Mol. Breed. 2013;31:87-89. DOI 10.1007/S11032-012-9772-1.
Rodriguez-Concepcion M., Avalos J., Bonet M.L., Boronat A., Gomez-
Gomez L., Hornero-Mendez D. , Limon M.C., Melendez-Martinez
A.J., Olmedilla-Alonso B., Palou A., Ribot J., Rodrigo M.J.,
Zacarias L., Zhu C. A global perspective on carotenoids: Metabolism,
biotechnology, and benefits for nutrition and health. Prog. Lipid
Res. 2018;70:62-93. DOI 10.1016/j.plipres.2018.04.004.
, Limon M.C., Melendez-Martinez
A.J., Olmedilla-Alonso B., Palou A., Ribot J., Rodrigo M.J.,
Zacarias L., Zhu C. A global perspective on carotenoids: Metabolism,
biotechnology, and benefits for nutrition and health. Prog. Lipid
Res. 2018;70:62-93. DOI 10.1016/j.plipres.2018.04.004.
Roncallo P.F., Cervigni G.L., Jensen C., Miranda R., Carrera A.D., Helguera M., Echenique V. QTL analysis of main and epistatic effects for flour color traits in durum wheat. Euphytica. 2012;185:77-92. DOI 10.1007/s10681-012-0628-x.
Roncallo P., Echenique V. Identification of molecular markers associated with yield and quality traits for Argentinean durum wheat breeding programs. In: Options Méditerranéennes. Proceedings of the International Symposium on Genetics and Breeding of Durum Wheat. Rome, Italy: CHEAM, 2014;577-582.
Schulthess A., Schwember A.R. Improving durum wheat (Triticum
turgidum L. var. durum) grain yellow pigment content through
plant breeding. Сienc. Invest. Agr. 2013;40:475-490. http://dx.doi.
org/10.7764/rcia.v40i3.1157.
http://dx.doi.
org/10.7764/rcia.v40i3.1157.
Sehgal D., Dreisigacker S. Haplotypes-based genetic analysis: benefits and challenges. Vavilovskii Zhurnal Genetiki i Selektsii = Vavilov Journal of Genetics and Breeding. 2019;23(7):803-808. DOI 10.18699/VJ19.37.
Simeone R., Pasqualone A., Clodoveo M., Blanco A. Genetic mapping of polyphenol oxidase in tetraploid wheat. Cell. Mol. Biol. Lett. 2002;7:763-769.
Si H., Ma C., Wang X., He X. Variability of polyphenol oxidase (PPO) alleles located on chromosomes 2A and 2D can change the wheat kernel PPO activity. Aust. J. Crop Sci. 2012;6(3):444-449.
Singh A., Reimer S., Pozniak C.J., Clarke F.R., Clarke J.M., Knox R.E., Singh A.K. Allelic variation at Psy1-A1 and association with yellow pigment in durum wheat grain. Theor. Appl. Genet. 2009;118:1539- 1548. DOI 10.1007/s00122-009-1001-x.
Sissons M. Role of durum wheat composition on the quality of pasta and bread. Food Global Sci. 2008;2:75-90.
Sun T., Yuan H., Cao H. , Yazdani M., Tadmor Y., Li L. Carotenoid metabolism
in plants: The role of plastids. Mol. Plant. 2018;11:58-74.
DOI 10.1016/j.molp.2017.09.010.
, Yazdani M., Tadmor Y., Li L. Carotenoid metabolism
in plants: The role of plastids. Mol. Plant. 2018;11:58-74.
DOI 10.1016/j.molp.2017.09.010.
Taranto F., Delvecchio L.N., Mangini G., Del Faro L., Blanco A., Pasqualone A. Molecular and physico-chemical evaluation of enzymatic browning of whole meal and dough in a collection of tetraploid wheats. J. Cereal Sci. 2012;55:405-414. DOI 10.1016/j.jcs. 2012.02.005.
Taranto F., Mangini G., Pasqualone A., Gadaleta A., Blanco A. Mapping and allelic variations of Ppo-B1 and Ppo-B2 gene-related polyphenol oxidase activity in durum wheat. Mol. Breed. 2015;35:80. DOI 10.1007/s11032-015-0272-y.
Vargas V.H., Schulthess A., Royo C., Matus I., Schwember A.R. Transcripts levels of Phytoene synthase 1 (Psy-1) are associated to semolina yellowness variation in durum wheat (Triticum turgidum L. ssp. durum). J. Cereal Sci. 2016;68:155-163. DOI 10.1016/j.jcs.2016. 01.011.
Verlotta A., De Simone V., Mastrangelo A.M., Cattivelli L., Papa R. ,
Trono D. Insight into durum wheat Lpx-B1: a small gene family
coding for the lipoxygenase responsible for carotenoid bleaching in
mature grains. BMC Plant Biol. 2010;10:263. DOI 10.1186/1471-
2229-10-263.
,
Trono D. Insight into durum wheat Lpx-B1: a small gene family
coding for the lipoxygenase responsible for carotenoid bleaching in
mature grains. BMC Plant Biol. 2010;10:263. DOI 10.1186/1471-
2229-10-263.
Wang S., Wong D., Forrest K., Allen A., Chao S., Huang B.E., Maccaferri M., Salvi S., Milner S.S., Cattivelli L., Mastrangello A.M., Whan A., Stephan S., Sarker G., Wieseke R., Plieske S. Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014;12:787-796. DOI 10.1111/pbi.12183.
Watanabe N., Akond A.S.M.G.M., Nachit M.M. Genetic mapping of the gene affecting polyphenol oxidase activity in tetraploid durum wheat. J. Appl. Genet. 2006;47:201-205. DOI 10.1007/BF031 94624.
Watanabe N., Takeuchi A., Nakayama A. Inheritance and chromosomal
location of the homoeologous genes affecting phenol colour reaction
of kernels in durum wheat. Euphytica. 2004;139:87-93. DOI
10.1007/s10681-004-2255-7.
Wei J., Geng H., Zhang Y., Liu J., Wen W., Zhang Y., Xia X., Chen X., He Z. Mapping quantitative trait loci for peroxidase activity and developing gene-specific markers for TaPod-A1 on wheat chromosome 3AL. Theor. Appl. Genet. 2015;128:2067-2076. DOI 10.1007/ s00122-015-2567-0.
Zeng J., Wang X., Miao Y., Wang C., Zang M., Chen X., Li M., Li X., Wang O., Li K., Chang J., Wang G., Yang G., He G. Metabolic engineering of wheat provitamin A by simultaneously overexpressing CrtB and silencing carotenoid hydroxylase (TaHYD). J. Agric. Food Chem. 2015;63:9083-9092. DOI 10.1021/acs.jafc.5b04279.
Zhang W., Dubcovsky J. Association between allelic variation at the Phytoene synthase 1 gene and yellow pigment content in the wheat grain. Theor. Appl. Genet. 2008;116:635-645. DOI 10.1007/s00122- 007-0697-8.
Zhang Y., Wu Y., Xiao Y., He Z., Zhang Y., Yan J., Zhang Y., Xia X.,
Ma C. QTL mapping for flour and noodle colour components and
yellow pigment content in common wheat. Euphytica. 2009;165(3):
435-444. DOI 10.1007/s10681-008-9744-z.
2009;165(3):
435-444. DOI 10.1007/s10681-008-9744-z.
Что такое ген? (для детей)
Гены (скажем: jeenz ) играют важную роль в определении физических качеств — того, как мы выглядим — и многих других вещей о нас. Они несут информацию, которая делает вас тем, кто вы есть и как вы выглядите: вьющиеся или прямые волосы, длинные или короткие ноги, даже то, как вы можете улыбаться или смеяться. Многие из этих вещей передаются от одного поколения к другому в семье по генам.
Что такое ген?
Гены несут информацию, которая определяет ваши черты (скажем: trates ), то есть черты или характеристики, которые передаются вам — или унаследованы — от ваших родителей.Каждая клетка человеческого тела содержит от 25 000 до 35 000 генов.
Например, если у обоих ваших родителей зеленые глаза, вы можете унаследовать от них черту зеленых глаз. Или, если у вашей мамы есть веснушки, у вас тоже могут быть веснушки, потому что вы унаследовали черту веснушек. Гены есть не только у людей — они есть у всех животных и растений.
Гены есть не только у людей — они есть у всех животных и растений.
Где эти важные гены? Они такие маленькие, что их не видно. Гены находятся на крошечных структурах, похожих на спагетти, называемых хромосомами (скажем: KRO-moh-somes).А хромосомы находятся внутри клеток. Ваше тело состоит из миллиардов клеток. Клетки — это очень маленькие единицы, из которых состоит все живое. Клетка настолько крошечная, что увидеть ее можно только в сильный микроскоп.
Хромосомы входят в соответствующие наборы из двух (или пар), и всего в одной хромосоме находятся сотни, а иногда и тысячи генов. Хромосомы и гены состоят из ДНК, что является сокращением от дезоксирибонуклеиновой (скажем: ди-окс-си-ри-бо-нью-глиняной) кислоты.
Большинство клеток имеют одно ядро (скажем: NOO-clee-us).Ядро — это небольшая яйцевидная структура внутри клетки, которая действует как мозг клетки. Он сообщает каждой части клетки, что делать. Но откуда ядро так много знает? Он содержит наши хромосомы и гены. Каким бы крошечным оно ни было, ядро содержит больше информации, чем самый большой словарь, который вы когда-либо видели.
Каким бы крошечным оно ни было, ядро содержит больше информации, чем самый большой словарь, который вы когда-либо видели.
У человека ядро клетки содержит 46 отдельных хромосом или 23 пары хромосом (хромосомы бывают парами, помните? 23 x 2 = 46). Половина этих хромосом принадлежит одному из родителей, а половина — другому.
Под микроскопом мы видим, что хромосомы бывают разной длины и разной формы. Когда они выстраиваются в линию по размеру и схожему рисунку полос, первые двадцать две пары называются аутосомами; последняя пара хромосом называется половыми хромосомами, X и Y. Половые хромосомы определяют, мальчик вы или девочка: у женщин две хромосомы X, а у мужчин — одна X и одна Y.
Но не у каждого живого существа внутри клетки есть 46 хромосом.Например, у клетки плодовой мушки всего четыре хромосомы!
Как работают гены?
У каждого гена своя собственная работа. ДНК в гене дает конкретные инструкции — как в рецепте поваренной книги — для производства белков (скажем, PRO-подростков) в клетке.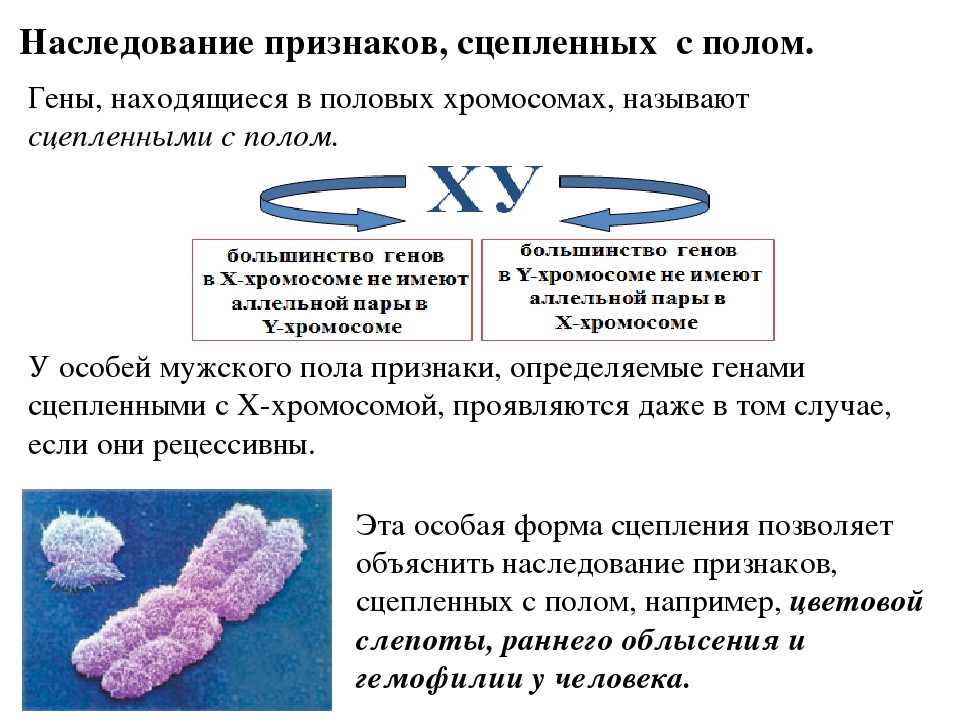 Белки — это строительные блоки всего в вашем теле. Кости и зубы, волосы и мочки ушей, мышцы и кровь состоят из белков. Эти белки помогают нашему телу расти, правильно работать и оставаться здоровым. По сегодняшним оценкам ученых, каждый ген в организме может вырабатывать до 10 различных белков.Это более 300 000 белков!
Белки — это строительные блоки всего в вашем теле. Кости и зубы, волосы и мочки ушей, мышцы и кровь состоят из белков. Эти белки помогают нашему телу расти, правильно работать и оставаться здоровым. По сегодняшним оценкам ученых, каждый ген в организме может вырабатывать до 10 различных белков.Это более 300 000 белков!
Гены, как и хромосомы, тоже попадают в пары. У каждого из ваших родителей есть по две копии каждого из своих генов, и каждый родитель передает только одну копию, чтобы составить гены, которые у вас есть. Гены, которые передаются вам, определяют многие ваши черты, такие как цвет волос и цвет кожи.
Может быть, у матери Эммы один ген каштановых волос и один — рыжих, и она передала ген рыжих волос Эмме. Если у ее отца два гена рыжих волос, это может объяснить ее рыжие волосы.Эмма получила два гена рыжих волос, по одному от каждого из ее родителей.
Вы также можете увидеть, как работают гены, если подумать обо всех породах собак. У всех них есть гены, которые делают их собаками, а не кошками, рыбами или людьми. Но те же гены, которые делают собаку собакой, также определяют разные собачьи черты. Итак, одни породы маленькие, а другие большие. У одних длинный мех, у других — короткий. У далматинов есть гены белого меха и черных пятен, а у игрушечных пуделей есть гены, которые делают их маленькими с вьющейся шерстью.Вы уловили идею!
Но те же гены, которые делают собаку собакой, также определяют разные собачьи черты. Итак, одни породы маленькие, а другие большие. У одних длинный мех, у других — короткий. У далматинов есть гены белого меха и черных пятен, а у игрушечных пуделей есть гены, которые делают их маленькими с вьющейся шерстью.Вы уловили идею!
Когда есть проблемы с генами
Ученые очень заняты изучением генов. Они хотят знать, какие белки вырабатывает каждый ген и что эти белки делают. Они также хотят знать, какие болезни вызваны неправильными генами. Измененные гены называются мутациями. Исследователи считают, что мутации могут быть частично виноваты в проблемах с легкими, раке и многих других заболеваниях. Другие болезни и проблемы со здоровьем возникают, когда отсутствуют гены или лишние части генов или хромосом.
Некоторые из этих генных проблем могут быть унаследованы от родителей. Например, возьмем ген, который помогает организму вырабатывать гемоглобин (скажем: HEE-muh-glow-bin). Гемоглобин — важный белок, необходимый эритроцитам для переноса кислорода по всему телу. Если родители передают своему ребенку измененные гены гемоглобина, ребенок может вырабатывать только гемоглобин того типа, который не работает должным образом. Это может вызвать состояние, известное как анемия (скажем: э-э-э-э-э-э), состояние, при котором у человека меньше здоровых эритроцитов.Серповидно-клеточная анемия — это один из видов анемии, который передается по генам от родителей к детям.
Гемоглобин — важный белок, необходимый эритроцитам для переноса кислорода по всему телу. Если родители передают своему ребенку измененные гены гемоглобина, ребенок может вырабатывать только гемоглобин того типа, который не работает должным образом. Это может вызвать состояние, известное как анемия (скажем: э-э-э-э-э-э), состояние, при котором у человека меньше здоровых эритроцитов.Серповидно-клеточная анемия — это один из видов анемии, который передается по генам от родителей к детям.
Муковисцидоз (скажем: SIS-tick fi-BRO-sus) или CF — еще одно заболевание, которое наследуют некоторые дети. Родители с измененным геном CF могут передать его своим детям. Людям с муковисцидозом часто трудно дышать, потому что в их организме вырабатывается много слизи (скажем: MYOO-kus) — слизи, которая выходит из носа, когда вы больны, и которая застревает в легких. Людям с МВ необходимо лечение на протяжении всей жизни, чтобы их легкие были как можно более здоровыми.
Что такое генная терапия?
Генная терапия — это новый вид медицины, настолько новый, что ученые все еще проводят эксперименты, чтобы проверить, работает ли оно. Он использует технологию генной инженерии для лечения болезни, вызванной каким-либо образом изменившимся геном. Один из тестируемых методов — замена больных генов здоровыми. Испытания генной терапии — где исследования проводятся на людях — и другие исследования могут привести к новым способам лечения или даже предотвращения многих заболеваний.
Он использует технологию генной инженерии для лечения болезни, вызванной каким-либо образом изменившимся геном. Один из тестируемых методов — замена больных генов здоровыми. Испытания генной терапии — где исследования проводятся на людях — и другие исследования могут привести к новым способам лечения или даже предотвращения многих заболеваний.
Гены и хромосомы — Основы
Чтобы предотвратить ошибки во время репликации, в клетках есть функция «корректуры», которая помогает гарантировать правильное спаривание оснований.Существуют также химические механизмы восстановления ДНК, которая не была скопирована должным образом. Однако из-за миллиардов пар оснований, вовлеченных в процесс синтеза белка, и сложности процесса, могут возникать ошибки. Такие ошибки могут возникать по многим причинам (включая воздействие радиации, лекарств или вирусов) или без видимых причин. Незначительные вариации ДНК очень распространены и встречаются у большинства людей. Большинство вариаций не влияет на последующие копии гена.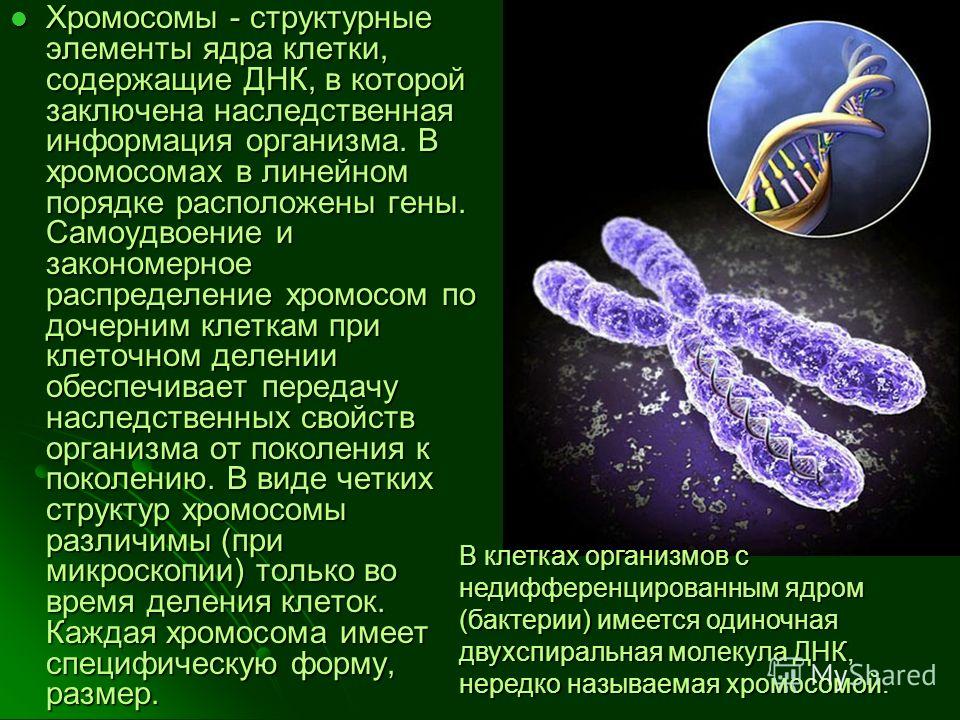 Ошибки, которые повторяются в последующих копиях, называются мутациями.
Ошибки, которые повторяются в последующих копиях, называются мутациями.
Унаследованные мутации — это мутации, которые могут передаваться потомству. Мутации могут передаваться по наследству только тогда, когда они затрагивают репродуктивные клетки (сперматозоиды или яйцеклетки). Мутации, не влияющие на репродуктивные клетки, влияют на потомков мутировавшей клетки (например, становятся раком), но не передаются потомству.
Мутации могут быть уникальными для отдельного человека или семьи, и наиболее опасные мутации встречаются редко. Мутации, которые становятся настолько распространенными, что затрагивают более 1% населения, называются полиморфизмами (например, группы крови человека A, B, AB и O).Большинство полиморфизмов практически не влияют на фенотип (фактическая структура и функция тела человека).
Мутации могут включать малые или большие сегменты ДНК. В зависимости от размера и местоположения мутация может не иметь видимого эффекта или может изменять аминокислотную последовательность в белке или уменьшать количество продуцируемого белка. Если белок имеет другую аминокислотную последовательность, он может функционировать по-другому или не функционировать совсем. Отсутствующий или нефункционирующий белок часто опасен или фатален.Например, при фенилкетонурии Фенилкетонурия (ФКУ) Фенилкетонурия — это нарушение метаболизма аминокислот, которое возникает у младенцев, рожденных без способности нормально расщеплять аминокислоту, называемую фенилаланином. Фенилаланин, который токсичен … читать дальше, мутация приводит к дефициту или отсутствию фермента фенилаланингидроксилазы. Этот дефицит позволяет аминокислоте фенилаланину (всасываемой с пищей) накапливаться в организме, что в конечном итоге вызывает серьезную умственную отсталость.В редких случаях мутация вносит выгодное изменение. Например, в случае серповидноклеточного гена, когда человек наследует две копии аномального гена, у человека разовьется серповидноклеточная болезнь Серповидноклеточная болезнь Серповидноклеточная болезнь — это наследственная генетическая аномалия гемоглобина (обнаруженный белок, переносящий кислород.
Если белок имеет другую аминокислотную последовательность, он может функционировать по-другому или не функционировать совсем. Отсутствующий или нефункционирующий белок часто опасен или фатален.Например, при фенилкетонурии Фенилкетонурия (ФКУ) Фенилкетонурия — это нарушение метаболизма аминокислот, которое возникает у младенцев, рожденных без способности нормально расщеплять аминокислоту, называемую фенилаланином. Фенилаланин, который токсичен … читать дальше, мутация приводит к дефициту или отсутствию фермента фенилаланингидроксилазы. Этот дефицит позволяет аминокислоте фенилаланину (всасываемой с пищей) накапливаться в организме, что в конечном итоге вызывает серьезную умственную отсталость.В редких случаях мутация вносит выгодное изменение. Например, в случае серповидноклеточного гена, когда человек наследует две копии аномального гена, у человека разовьется серповидноклеточная болезнь Серповидноклеточная болезнь Серповидноклеточная болезнь — это наследственная генетическая аномалия гемоглобина (обнаруженный белок, переносящий кислород. в красных кровяных тельцах) характеризуются серповидными (серповидными) эритроцитами и хроническими … подробнее. Однако, когда человек наследует только одну копию гена серповидных клеток (называемого носителем), у человека появляется некоторая защита от малярии. Малярия Малярия — это инфицирование красных кровяных телец одним из пяти видов Plasmodium, простейшими.Малярия вызывает жар, озноб, потливость, общее недомогание (недомогание), а иногда и диарею … подробнее (заражение крови). Хотя защита от малярии может помочь носителю выжить, серповидно-клеточная анемия (у человека, имеющего две копии гена) вызывает симптомы и осложнения, которые могут сократить продолжительность жизни.
в красных кровяных тельцах) характеризуются серповидными (серповидными) эритроцитами и хроническими … подробнее. Однако, когда человек наследует только одну копию гена серповидных клеток (называемого носителем), у человека появляется некоторая защита от малярии. Малярия Малярия — это инфицирование красных кровяных телец одним из пяти видов Plasmodium, простейшими.Малярия вызывает жар, озноб, потливость, общее недомогание (недомогание), а иногда и диарею … подробнее (заражение крови). Хотя защита от малярии может помочь носителю выжить, серповидно-клеточная анемия (у человека, имеющего две копии гена) вызывает симптомы и осложнения, которые могут сократить продолжительность жизни.
Естественный отбор относится к концепции, согласно которой мутации, ухудшающие выживаемость в данной среде, с меньшей вероятностью передаются потомству (и, таким образом, становятся менее распространенными в популяции), тогда как мутации, улучшающие выживаемость, становятся все более распространенными. Таким образом, полезные мутации, хотя изначально редки, в конечном итоге становятся обычным явлением. Медленные изменения, происходящие с течением времени, вызванные мутациями и естественным отбором в скрещивающейся популяции, в совокупности называются эволюцией .
Таким образом, полезные мутации, хотя изначально редки, в конечном итоге становятся обычным явлением. Медленные изменения, происходящие с течением времени, вызванные мутациями и естественным отбором в скрещивающейся популяции, в совокупности называются эволюцией .
Признаки — Определение и примеры
Признаки
сущ., Единственное число: признак
[tɹeɪts]
Определение: характеристики или атрибуты, выраженные генами и / или находящиеся под влиянием окружающей среды
Определение признаков
«Признак» — довольно известное слово в английском языке. Что означает черта?
Черта определяется как качество кого-то или чего-то примечательного. Как правило, определение на английском языке совпадает с определением биологии. Тогда возникает вопрос: , что такое черта в биологии? Как биология определяет «черты»? Определение черты в биологии — это особая характеристика, которую организм может удерживать или отображать.
Признаки — это характеристики или атрибуты организма, которые выражаются генами и / или находятся под влиянием окружающей среды.Черты включают физические атрибуты организма, такие как цвет волос, форма, размер листьев и т. Д., А также поведенческие характеристики, такие как гнездование птиц. Этимология: от латинского tractus. См. Также: фенотип, наследование, ген.
Типы черт
Организм может отображать множество типов черт. Все признаки представляют собой либо генетических признаков , которые основаны на генах организма, либо поведенческих признаков , которые определяются взаимодействием окружающей среды с генами.
1. Генетические признаки
Как гены определяют особенности организма? Генетические признаки контролируются генами, которые наследуются от обоих родителей. Гены определяют наши индивидуальные и независимые характеристики как кодированные определенными сегментами нашей ДНК.
Эти гены кодируют наш генотип, который является генетической структурой организма. Поскольку генотип определяет фенотип, фенотипические признаки — это физических признаков , которые проявляются организмом, определяемым генетическими признаками, которые они унаследовали.
2. Поведенческие черты
Поведенческие черты — это те действия, которые наблюдаются у организмов всех их видов. Это наблюдаемая или присущая окружающей среде черта и передается по наследству через обучение или инстинкт. Однако следует отметить, что поведенческие черты представляют собой смесь унаследованного поведения, основанного на генетике, и поведения, приобретенного на основе социальных сигналов и ближайшего окружения.
Рис. 1. Змея со своим экологическим признаком, маскировкой. Предоставлено: Зденек Розенталер. Окружение играет большую роль в том, как черты будут отображаться в конце. Определенные черты могут нормально проявляться одним образом — поведенческими или генетическими — но могут различаться в зависимости от среды, в которой находится организм. Для некоторых подвижных организмов, таких как некоторые животные и люди, этими факторами окружающей среды можно управлять, если они могут отойти от них. Для других, с другой стороны, это не вариант, и они должны соответствовать и адаптироваться к этим условиям. Это часто может изменить способ изображения физических черт в будущих поколениях.
Для некоторых подвижных организмов, таких как некоторые животные и люди, этими факторами окружающей среды можно управлять, если они могут отойти от них. Для других, с другой стороны, это не вариант, и они должны соответствовать и адаптироваться к этим условиям. Это часто может изменить способ изображения физических черт в будущих поколениях.
Иногда признаки, основанные на окружающей среде, могут быть очень полезны диким и даже домашним животным, а также людям. Маленькие змеи, рожденные с чешуйчатыми узорами, которые сочетаются с окружающей средой, с меньшей вероятностью станут жертвами хищников. Эта черта, камуфляж, — отличная экологическая черта, которая может помочь организмам выжить. Другие физические особенности, такие как красота и узор перьев павлина, могут облегчить поиск партнера.
Примеры черт
Достаточно легко увидеть примеры физических черт в среднестатистическом организме.Многие живые существа проявляют свои черты и передают их будущим поколениям. Например, у растений есть множество черт, которые может заметить средний человек.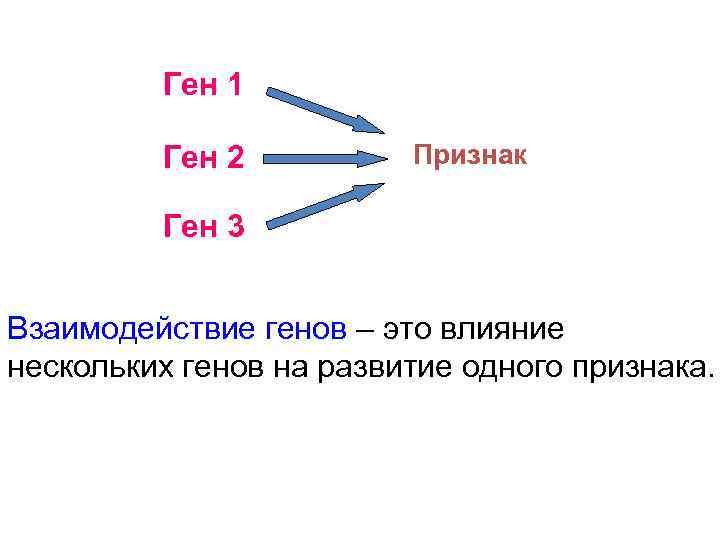 Форма листьев, цвет цветов и даже узоры ветвления некоторых деревьев — все это разные типы черт, которые передаются генетически и проявляются физически.
Форма листьев, цвет цветов и даже узоры ветвления некоторых деревьев — все это разные типы черт, которые передаются генетически и проявляются физически.
-
Признаки растения гороха Менделя
На Рисунке 2 ниже мы видим один из наиболее распространенных примеров физических признаков растений в биологии.Признаки растения гороха Менделя показывают различные фенотипы или физические признаки, которые приобретает растение гороха, в зависимости от того, является ли его генотип доминантным или рецессивным. Растения могут быть высокими или короткими, желтыми или зелеными и даже осевыми или конечными — все в зависимости от унаследованных ими генов.
Рис. 2. Признаки растения гороха Менделя.ЧИТАТЬ: Закон Менделя и Менделевская генетика
Многие животные также обладают определенными физическими чертами. Эти черты наследуются от родительских поколений и передаются потомкам и другим будущим поколениям.Родительские организмы могут производить потомство, которое имеет физические черты, сильно отличающиеся от других потомков того же родителя.
Рисунок 3 ниже показывает нам, что, несмотря на то, что у одной из собак-родителей шерсть более темно-коричневая, чем у другого родителя, все щенки помета физически выглядели как другой родитель, имея более светлый цвет шерсти.
Рис. 3. Судя по фотографии, «доминирующей» чертой является кремовый окрас шерсти. Фото: Sweethome.Другие хорошо известные физические генетические особенности, которые люди передают своему потомству, показаны на Рисунке 4.Они являются примерами моногенного признака, что означает признак, определяемый одним геном.
Рис. 4: Примеры моногенных признаков у человека. Кредит: bio.libretexts.org, CC BY-NC-SA 3.0.Полигенные признаки — это признаки, которые контролируются полигенами (множественными неаллельными генами). Примерами являются рост, цвет кожи, цвет волос и цвет глаз людей. Признаки определяются не одним геном, а множеством генов.
У животных поведенческие черты могут варьироваться от того, как животные двигаются, до того, как они защищаются от различных хищников и раздражителей.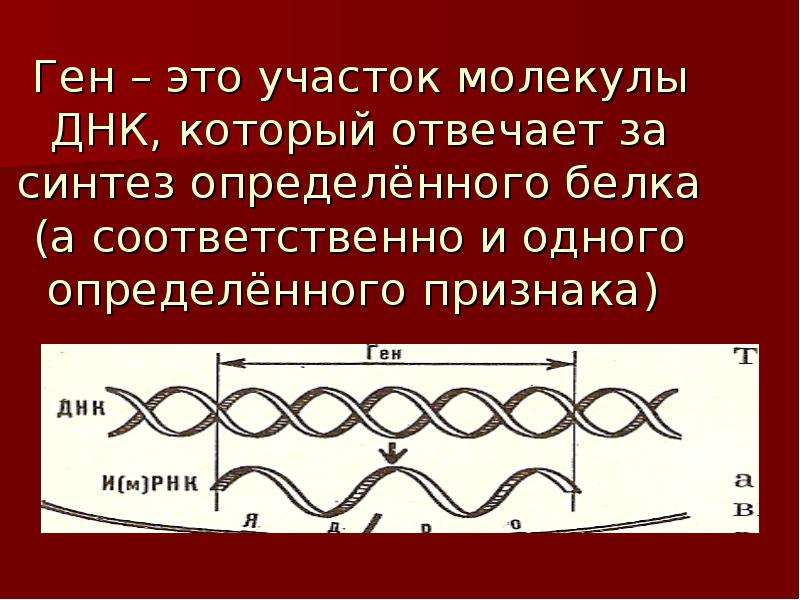 Опоссумы печально известны тем, что притворяются мертвыми, чтобы отогнать хищников и избежать быть съеденными. Это поведение наблюдается и имитируется их потомками и передается из поколения в поколение.
Опоссумы печально известны тем, что притворяются мертвыми, чтобы отогнать хищников и избежать быть съеденными. Это поведение наблюдается и имитируется их потомками и передается из поколения в поколение.
Другие животные, такие как шипящие тараканы, используют звуки, чтобы обмануть хищников и заставить их думать, что они более опасны, чем кажутся. Эти звуки снова узнаются их потомками и используются в будущих поколениях.
Некоторые животные также учатся и развивают черты характера в зависимости от среды обитания.Эти экологические характеристики могут варьироваться от внутренних изменений, таких как гормоны и температура тела, до внешних изменений, таких как физические характеристики. Например, животные, которые живут в районах, где они живут в разные времена года, будут претерпевать физические изменения с приближением зимы. Такие животные, как заяц-снегоступы, полагаются на камуфляж, чтобы избежать хищников. Следовательно, по мере смены сезонов они меняют цвет своей шубы — на белый зимой и коричневый осенью и весной, чтобы они могли более эффективно сочетаться с окружающей средой.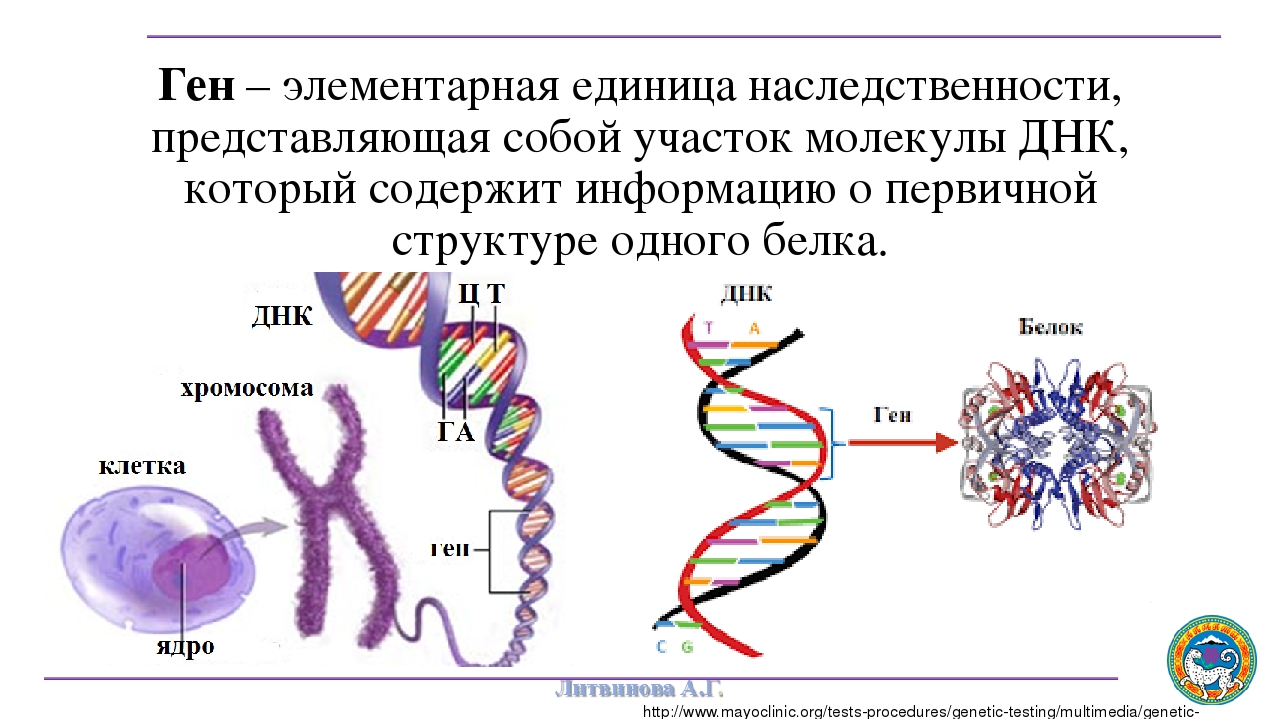
Посмотрите это видео, чтобы узнать больше о чертах характера.
Ссылки
- Accelerate Learning.
 2021. Экологические особенности. Ускоренное обучение, http://davis314.weebly.com/uploads/1/0/6/3/106359787/environmental_traits.pdf
2021. Экологические особенности. Ускоренное обучение, http://davis314.weebly.com/uploads/1/0/6/3/106359787/environmental_traits.pdf - Команда Сети животных. 2021. Девять удивительных животных-защитников, которые используют, чтобы избежать хищников. Сеть животных. https://animals.net/nine-awesome-defenses-animals-use-to-avoid-predators/
- Generation Genius. 2021 г.Наследственные черты и генетика для детей. Поколение Гений. https://www.generationgenius.com/inherited-traits-and-genetics-for-kids/
- Lobo, I. (2008) Влияние окружающей среды на экспрессию генов. Nature Education 1 (1): 39
https://www.nature.com/scitable/topicpage/environmental-influences-on-gene-expression-536/ - Национальный институт исследования генома человека. 2021. Черты характера. О геномике. https://www.genome.gov/genetics-glossary/Trait
- Природное образование. 2014 г.Генетика. Scitable by Nature Education. https://www.nature.com/scitable/topicpage/each-organism-s-traits-are-inherited-from-6524917/
- PBS.
 2021. Воздействие окружающей среды на черты характера. Наука о жизни. https://www.pbslearningmedia.org/subjects/science/life-science/genetics-and-heredity/environmental-effects-on-traits/
2021. Воздействие окружающей среды на черты характера. Наука о жизни. https://www.pbslearningmedia.org/subjects/science/life-science/genetics-and-heredity/environmental-effects-on-traits/ - Writer, S. 2020. Что такое поведенческая черта? Ссылка. https://www.reference.com/world-view/behavioral-trait-9d7ca58594464c10
© BiologyOnline.com. Контент предоставлен и модерируется редакторами Biology Online.
Близнецы могут помочь нам понять, как гены и окружающая среда формируют нас · Границы для молодых умов
Абстрактные
Вы когда-нибудь задумывались, почему вы отличаетесь от других людей? Различия, такие как рост и цвет волос, могут возникать из-за различий в ваших генах, называемых мутациями. Понимание того, как мутации формируют нас, может быть немного сложным, потому что на нас также влияет окружающий мир.Близнецы, братья и сестры могут помочь нам понять, разные ли люди из-за генетических различий или из-за того, что они живут в разных средах. Однояйцевые близнецы имеют одни и те же гены, в то время как неидентичные близнецы в среднем разделяют половину генов с другим близнецом. Мы можем измерить и сравнить степень сходства черт между парами однояйцевых и неидентичных близнецов, чтобы понять, какая разница в чертах между людьми вызвана генами, а какая — окружающей средой.
Однояйцевые близнецы имеют одни и те же гены, в то время как неидентичные близнецы в среднем разделяют половину генов с другим близнецом. Мы можем измерить и сравнить степень сходства черт между парами однояйцевых и неидентичных близнецов, чтобы понять, какая разница в чертах между людьми вызвана генами, а какая — окружающей средой.
Каждый человек в мире отличается от всех остальных. У вас могут быть разные цвета волос, кожи или глаз или другой рост, а также целый ряд других черт , которые делают вас уникальным. С другой стороны, у вас очень схожие черты с членами вашей семьи. У вас может быть тот же цвет волос, что и у вашего отца, и такой же цвет глаз, как у вашей мамы. Вы можете немного походить на своего брата или сестру. Это связано с тем, что семьи имеют общие вариации (также называемые мутациями ) в своих генах .
Можно сказать, что гены — это небольшие кусочки биологической информации, передаваемые от родителей к их детям, закодированные в ДНК . Думайте о генах как о рецепте с инструкциями, в которых перечислены ингредиенты, количество и шаги по приготовлению пищи. В этом примере , вы — еда, а ваша ДНК — это и бумага, и буквы, использованные для написания рецепта. Хотя наши гены подобны рецептам, имейте в виду, что это очень сложный рецепт с множеством (множеством!) Ингредиентов и шагов, которые мы все еще пытаемся открыть и понять.В мире есть много видов еды, как и много разных людей. Кексы отличаются от печенья тем, что у них разные рецепты. Люди отличаются друг от друга из-за разницы в генах (рецептах). Некоторые люди более похожи, чем другие (например, вы и ваш брат или сестра больше похожи, чем вы и ваши друзья). Это как кексы и кексы — они не одно и то же, но очень похожи. Это потому, что их рецепты похожи, но не совсем одинаковы.Иногда определенные мутации в ваших генах могут быть плохими для вас и вызвать болезнь, в то время как другие мутации могут защитить вас от болезней. Некоторые мутации могут быть даже хорошими или плохими в зависимости от окружающей среды! По нашей аналогии, мутация, вызывающая болезнь, будет похожа на использование неправильного рецепта.
Думайте о генах как о рецепте с инструкциями, в которых перечислены ингредиенты, количество и шаги по приготовлению пищи. В этом примере , вы — еда, а ваша ДНК — это и бумага, и буквы, использованные для написания рецепта. Хотя наши гены подобны рецептам, имейте в виду, что это очень сложный рецепт с множеством (множеством!) Ингредиентов и шагов, которые мы все еще пытаемся открыть и понять.В мире есть много видов еды, как и много разных людей. Кексы отличаются от печенья тем, что у них разные рецепты. Люди отличаются друг от друга из-за разницы в генах (рецептах). Некоторые люди более похожи, чем другие (например, вы и ваш брат или сестра больше похожи, чем вы и ваши друзья). Это как кексы и кексы — они не одно и то же, но очень похожи. Это потому, что их рецепты похожи, но не совсем одинаковы.Иногда определенные мутации в ваших генах могут быть плохими для вас и вызвать болезнь, в то время как другие мутации могут защитить вас от болезней. Некоторые мутации могут быть даже хорошими или плохими в зависимости от окружающей среды! По нашей аналогии, мутация, вызывающая болезнь, будет похожа на использование неправильного рецепта. Если вы добавите соль в смесь для торта вместо сахара, пирог не получится хорошо. Хотя вы можете подумать, что мутация — это что-то плохое (или, может быть, что-то классное, что есть у некоторых супергероев), правда в том, что большинство мутаций не являются ни хорошими, ни плохими — их называют нейтральными.
Если вы добавите соль в смесь для торта вместо сахара, пирог не получится хорошо. Хотя вы можете подумать, что мутация — это что-то плохое (или, может быть, что-то классное, что есть у некоторых супергероев), правда в том, что большинство мутаций не являются ни хорошими, ни плохими — их называют нейтральными.
Рецепт — не единственное, что влияет на выход торта. Среда также играет важную роль. Если вы сделаете духовку слишком горячей, пирог подгорит, но если он недостаточно горячий, он не будет готовиться и плюхнется. Но ждать! Использование неправильных ингредиентов или правильных в неправильном количестве также может сделать торт провальным! [1]. Если вы найдете выпавший торт, вы не всегда будете знать, что его вызвало. Это была ошибка в рецепте? Или это было неправильно приготовлено? Это важное различие между генами и рецептами.Хотя вы можете видеть симптомы болезни так же, как вы видите провалившийся торт, рецепт в ДНК написан на другом языке, и мы не можем легко вернуться и прочитать его, чтобы найти ошибку.
Окружающая среда сильно влияет на человеческие качества. Подумайте, например, о цвете кожи. Вы можете родиться с тем же цветом кожи, что и ваш друг. Если вашему другу нравится заниматься спортом на улице, а вам нравится читать в помещении, солнце сделает кожу вашего друга темнее, а ваша кожа останется незагорелой.Вы будете выглядеть по-другому, даже если у вас одинаковые гены цвета кожи. Опять же, гены также могут придавать людям разные цвета кожи. Когда вы видите людей с разным цветом кожи, трудно сказать, вызваны ли эти различия генами (генетическими), вызванными воздействием солнца (окружающая среда) или даже комбинацией обоих генов и окружающей среды! На самом деле, все различия между вами и всеми людьми вокруг вас происходят из-за различий в ваших генах, в вашем окружении или их комбинации.Например, вы можете быть очень умным, потому что родители заставляют вас много читать (это зависит от вашего окружения). Или вы можете быть очень умным, потому что у вас есть гены, которые влияют на ваш мозг, позволяя вам быстрее читать или легче понимать вещи [2].
Понимание того, как мутации влияют на определенную черту в группе людей, очень важно, потому что это помогает нам понять, почему мы все разные. Кроме того, понимание мутаций может помочь нам изучить способы, которыми гены вызывают некоторые заболевания.С другой стороны, выяснение того, как окружение влияет на черту, также полезно, потому что окружение иногда можно изменить. Например, ученые выяснили, что болезнь Диабет лишь немного зависит от генов (наследуемость составляет 26%; [3]). Зная это, врачи могут помочь людям избежать заражения из-за изменений окружающей среды (например, придерживаться здоровой диеты или больше заниматься спортом), даже если у них есть генов диабета!
Чтобы понять влияние генов vs.окружающей среды по определенному признаку, мы могли бы изучить группу людей и сравнить различия в их генах и различия в интересующих признаках. Это похоже на сравнение всех рецептов, используемых для разных тортов, чтобы увидеть, является ли какой-либо конкретный ингредиент общим для всех гибких тортов. Если есть общий ингредиент в мягких тортах, и этот ингредиент отсутствует в не мягких тортах, то этот конкретный ингредиент, вероятно, является причиной хлопья торта. Таким же образом мы можем увидеть, есть ли у всех людей с различиями в чертах определенные мутации.Затем мы можем использовать математику, чтобы понять, какая часть различий в признаке объясняется мутациями. Проблема этого метода в том, что чтение ДНК многих людей может быть очень дорогим и сложным. К счастью, почти 100 лет назад ученые нашли мощный способ изучить влияние генов на окружающую среду без необходимости считывать ДНК. В этом методе используются близнецы [4].
Если есть общий ингредиент в мягких тортах, и этот ингредиент отсутствует в не мягких тортах, то этот конкретный ингредиент, вероятно, является причиной хлопья торта. Таким же образом мы можем увидеть, есть ли у всех людей с различиями в чертах определенные мутации.Затем мы можем использовать математику, чтобы понять, какая часть различий в признаке объясняется мутациями. Проблема этого метода в том, что чтение ДНК многих людей может быть очень дорогим и сложным. К счастью, почти 100 лет назад ученые нашли мощный способ изучить влияние генов на окружающую среду без необходимости считывать ДНК. В этом методе используются близнецы [4].
Близнецы — это особый тип братьев и сестер, потому что они рождаются в одно время. Есть два типа близнецов.Однояйцевые близнецы очень похожи друг на друга и всегда либо мальчики, либо девочки. Неидентичные близнецы могут выглядеть по-разному, и даже могут быть мальчиком и девочкой. Однояйцевые близнецы имеют все свои гены, в то время как неидентичные близнецы, как и братья и сестры, не являющиеся близнецами, имеют половину общих генов.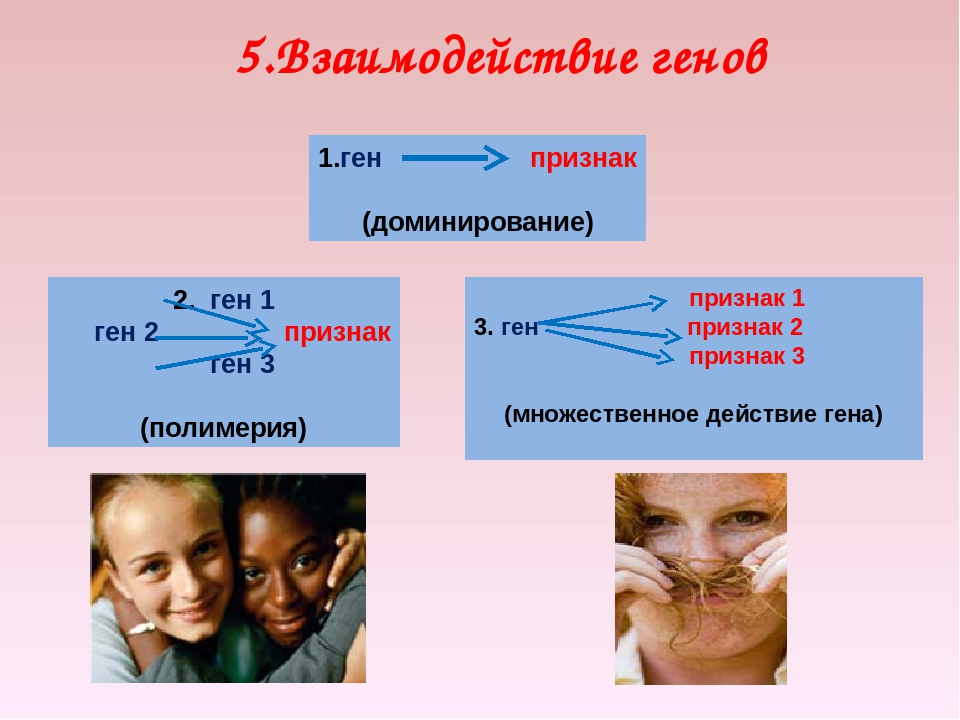 Итак, мы можем предположить, что любые различия в чертах между однояйцевыми близнецами происходят из окружающей среды, а не из-за различий в их генах. Измеряя признак (например, цвет кожи) в больших группах пар близнецов и братьев и сестер, мы можем понять влияние генов противокружающая среда по признаку, не глядя непосредственно на гены людей, чтобы увидеть их мутации.
Итак, мы можем предположить, что любые различия в чертах между однояйцевыми близнецами происходят из окружающей среды, а не из-за различий в их генах. Измеряя признак (например, цвет кожи) в больших группах пар близнецов и братьев и сестер, мы можем понять влияние генов противокружающая среда по признаку, не глядя непосредственно на гены людей, чтобы увидеть их мутации.
Пример результатов двух исследований близнецов показан на рисунке 1. В одном исследовании у австралийских близнецов (как идентичных, так и неидентичных) был измерен интеллект с помощью теста IQ (интеллектуального коэффициента). В другом исследовании близнецов спрашивали, сколько часов в сутки они обычно спят. Оба исследования были направлены на то, чтобы выяснить, насколько гены влияют на интересующие нас черты (IQ и время сна).Чтобы понять, насколько генетическим является признак, они сравнили, насколько похожи пары однояйцевых близнецов и насколько похожи пары неидентичных близнецов. Поскольку у однояйцевых близнецов общие гены, их измерения IQ и времени сна будут тем более похожими, чем большую роль в этом играют гены (то есть, различия между двумя однояйцевыми близнецами должны быть вызваны окружающей средой, потому что их гены одинаковы). С другой стороны, мы ожидаем, что неидентичные близнецы будут менее похожи, чем однояйцевые близнецы, но мы все же ожидаем, что они будут в чем-то похожими, потому что некоторые из их генов являются общими.Имейте в виду, что сравниваются не показатели черт, а то, насколько близнецы похожи (оба с высоким IQ или оба спят одинаковое количество времени). На Рисунке 1 сходство можно наблюдать по тому, насколько близко точки (пары близнецов) находятся к зеленой линии посередине (линия идеального сходства). Эти исследования показали, что IQ очень передается по наследству (хотя окружающая среда по-прежнему играет роль в определении вашего IQ). Мы можем сказать это, потому что IQ однояйцевых близнецов почти всегда одинаков, а неидентичные близнецы — лишь иногда.С другой стороны, график сна однояйцевых близнецов показывает большой разброс от средней зеленой линии. Это означает, что между однояйцевыми близнецами наблюдаются большие различия, а поскольку у них одни и те же гены, эти большие различия вызваны окружающей средой.
С другой стороны, мы ожидаем, что неидентичные близнецы будут менее похожи, чем однояйцевые близнецы, но мы все же ожидаем, что они будут в чем-то похожими, потому что некоторые из их генов являются общими.Имейте в виду, что сравниваются не показатели черт, а то, насколько близнецы похожи (оба с высоким IQ или оба спят одинаковое количество времени). На Рисунке 1 сходство можно наблюдать по тому, насколько близко точки (пары близнецов) находятся к зеленой линии посередине (линия идеального сходства). Эти исследования показали, что IQ очень передается по наследству (хотя окружающая среда по-прежнему играет роль в определении вашего IQ). Мы можем сказать это, потому что IQ однояйцевых близнецов почти всегда одинаков, а неидентичные близнецы — лишь иногда.С другой стороны, график сна однояйцевых близнецов показывает большой разброс от средней зеленой линии. Это означает, что между однояйцевыми близнецами наблюдаются большие различия, а поскольку у них одни и те же гены, эти большие различия вызваны окружающей средой. Это означает, что продолжительность сна имеет низкую наследуемость.
Это означает, что продолжительность сна имеет низкую наследуемость.
- Рис. 1. Данные о близнецах помогают нам понять генетические эффекты и влияние окружающей среды.
- Результаты двух реальных исследований близнецов. Два вверху — это результаты IQ (показатель интеллекта), а те, что внизу, показывают результаты времени сна.На графиках каждая точка — это пара близнецов. Ось X представляет оценку (или количество часов сна), полученное первым близнецом, а ось Y представляет собой оценку, полученную другим близнецом (братом). Если оба близнеца очень умны, точка, представляющая их, будет в верхнем правом углу (они оба получили высокие баллы). Если у пары близнецов одни и те же измерения (IQ или часы сна), они будут лежать на зеленой линии. Если их размеры сильно различаются, их точка будет далеко от зеленой линии.
Подобные исследования выявили, что личность, интеллект, плохое зрение и даже психические заболевания, такие как биполярное расстройство и шизофрения, имеют наследственность от средней до высокой (т. е., на них сильно влияют гены). Практически нет предела тому, какие признаки или болезни можно изучать. Если его можно измерить или классифицировать, мы сможем оценить его наследуемость! Мы изучаем близнецов, чтобы понять, какая разница в чертах характера между людьми вызвана генами, а какая — окружающей средой. Эти исследования важны, потому что они помогают ученым количественно оценить генетических и изменяемых факторов окружающей среды , которые увеличивают риск определенных заболеваний.Ученые провели много подобных исследований. К настоящему времени изучено около 18 000 человеческих черт, включая рост, массу тела и несколько заболеваний [5]. Итак, когда вы видите пару близнецов, вспомните, насколько они генетически особенные и ценные для науки и медицинских исследований.
е., на них сильно влияют гены). Практически нет предела тому, какие признаки или болезни можно изучать. Если его можно измерить или классифицировать, мы сможем оценить его наследуемость! Мы изучаем близнецов, чтобы понять, какая разница в чертах характера между людьми вызвана генами, а какая — окружающей средой. Эти исследования важны, потому что они помогают ученым количественно оценить генетических и изменяемых факторов окружающей среды , которые увеличивают риск определенных заболеваний.Ученые провели много подобных исследований. К настоящему времени изучено около 18 000 человеческих черт, включая рост, массу тела и несколько заболеваний [5]. Итак, когда вы видите пару близнецов, вспомните, насколько они генетически особенные и ценные для науки и медицинских исследований.
Глоссарий
Черта : ↑ Определенная характеристика или характеристика человека. Например, чей-то цвет волос или рост.
Мутация : ↑ Форма или версия гена. Мутации могут возникать внезапно в любое время и обычно не являются ни хорошими, ни плохими для вас.
Мутации могут возникать внезапно в любое время и обычно не являются ни хорошими, ни плохими для вас.
Ген : ↑ Часть ДНК, которая передается от родителей к детям. Гены содержат информацию о ваших качествах.
ДНК : ↑ Молекула внутри ваших клеток, в которой хранятся все ваши гены. Ваша ДНК содержит по одной копии ДНК ваших родителей, что делает вас похожими на них обоих.
Диабет : ↑ Заболевание, которое возникает, когда организм не может нормально использовать глюкозу (разновидность сахара).
Наследственность : ↑ Степень изменения признака, вызванного генетическими различиями.
Сходство : ↑ Оценка, которая измеряет сходство вещей. В этом случае мы измеряем, насколько близнецы похожи на своих близнецов.
Возможность изменения : ↑ Что-то, что можно изменить. Например, количество упражнений, которые вы делаете.
Например, количество упражнений, которые вы делаете.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
[1] ↑ Мур, Дж. 2018. Подход к случаю: как уберечь торт от падения . Получено с: https://www.craftsy.com/cake-decorating/article/how-to-keep-a-cake-from-falling/
.[2] ↑ Джоши, А. А., Лепоре, Н., Джоши, С. Х., Ли, А. Д., Барышева, М., Стейн, Дж. Л. и др. 2011. Вклад генов в толщину и объем коры. Нейроотчет . 22: 101–5. DOI: 10.1097 / WNR.0b013e3283424c84
[3] ↑ Поульсен, П., Кивик, К. О., Вааг, А., и Бек-Нильсен, Х. 1999. Наследование сахарного диабета типа II (инсулино-зависимый) и аномальной толерантности к глюкозе — популяционное исследование близнецов. Диабетология . 42: 139–45.
42: 139–45.
[4] ↑ Лью, С. Х. М., Элснер, Х., Спектор, Т. Д., и Хаммонд, К. Дж. 2005. Первое «классическое» исследование близнецов? Анализ аномалии рефракции у монозиготных и дизиготных близнецов, опубликованный в 1922 году. Twin Res. Гм. Genet. 8: 198–200. DOI: 10.1375 / 1832427054253158
[5] ↑ Полдерман, Т.J., Benyamin, B., De Leeuw, C.A., Sullivan, P.F., Van Bochoven, A., Visscher, P.M, et al. 2015. Мета-анализ наследуемости человеческих черт на основе пятидесятилетних исследований близнецов. Nat. Genet. 47: 702. DOI: 10,1038 / нг. 3285
| Множественный выбор Определите буква выбора, которая лучше всего завершает утверждение или отвечает на вопрос.
|
||||||||||||
| А B C D |
1. |
Факторы, управляющие признаками, называются
|
||||||||||
| А B C D |
2. |
Ученые называют организм, имеющий два разных аллеля признака а
|
||||||||||
| А B C D |
3. |
Что Обозначение TT означает для генетиков?
|
||||||||||
| А B C D |
4. |
Что Обозначение Tt означает для генетиков?
|
||||||||||
| А B C D |
5. |
Что это вероятность?
|
||||||||||
| А B C D |
6. |
Что показывает квадрат Пеннета?
|
||||||||||
| А B C D |
7. |
Если гомозиготная черная морская свинка ( BB ) скрещена с гомозиготной белой морской свинкой ( bb ), какова вероятность того, что у потомства будет черный мех?
|
||||||||||
| А B C D |
8. |
An внешний вид организма — его
|
||||||||||
| А B C D |
9. |
Что кодоминант в генетике?
|
||||||||||
| А B C D |
10. |
Что бывает во время мейоза?
|
||||||||||
| А B C D |
11. |
Когда половые клетки объединяются, чтобы произвести потомство, каждая половая клетка будет способствовать
|
||||||||||
| А B C D |
12. |
Что это мутация?
|
||||||||||
| А B C D |
13. |
Какой термин относится к физическим характеристикам, которые изучаются в генетике?
|
||||||||||
| А B C D |
14. |
различные формы гена называются
|
||||||||||
| А B C D |
15. |
Где происходит ли синтез белка?
|
||||||||||
| А B C D |
16. |
Что Обозначение tt означает для генетиков?
|
||||||||||
| А B C D |
17. |
An генотип организма — его
|
||||||||||
| А B C D |
18. |
Какой азотистое основание в РНК НЕ является частью ДНК?
|
||||||||||
| А B C D |
19. |
An Организм, имеющий два идентичных аллеля для признака, —
|
||||||||||
| А B C D |
20. |
А гетерозиготный организм имеет
|
||||||||||
| А B C D |
21. |
Какой из этих признаков контролируется геном с множественными аллелями?
|
||||||||||
| А B C D |
22. |
Какой сочетание половых хромосом приводит к появлению человека мужского пола?
|
||||||||||
| А B C D |
23. |
Почему Связанные с полом признаки чаще встречаются у мужчин, чем у женщин?
|
||||||||||
| А B C D |
24. |
Как генетик использует родословные?
|
||||||||||
| А B C D |
25. |
Что это родословная?
|
||||||||||
| А B C D |
26. |
Генетические нарушения вызваны
|
||||||||||
| А B C D |
27. |
Какой генетическое заболевание заставляет организм производить необычно густую слизь в легких и кишечник?
|
||||||||||
| А B C D |
28. |
В результате клонирования образуются два организма:
|
||||||||||
| А B C D |
29. |
Какой из этих примеров преимуществ генной инженерии?
|
||||||||||
| А B C D |
30. |
Нет два человека имеют одинаковую ДНК, за исключением
|
||||||||||
| А B C D |
31. |
Сцепленные с полом гены — это гены на
|
||||||||||
| А B C D |
32. |
А Перевозчик — это лицо, имеющее
|
||||||||||
| Короткий ответ
|
||||||||||||
|
Используйте схему, чтобы ответить на каждый вопрос .
|
||||||||||||
|
33. |
Какой черта — белые цветы или пурпурные цветы — контролируется доминантным аллелем? Что контролируется рецессивный аллель? Откуда вы знаете?
|
|||||||||||
|
34. |
В у какого поколения родители чистокровные? В каком поколении они гибриды?
|
|||||||||||
|
35. |
В поколение F 1 , каков генотип потомства? Какая у них фенотип?
|
|||||||||||
|
Используйте схему, чтобы ответить на каждый вопрос .
|
||||||||||||
|
36. |
Что обозначают круги в родословной? Что представляют собой квадраты?
|
|||||||||||
|
37. |
Какой пары особей в родословной имеют детей?
|
|||||||||||
|
38. |
Какой особи имеют черту, которая прослеживается по родословной?
|
|||||||||||
|
39. |
Какой особи являются носителями признака, прослеженного по родословной?
|
|||||||||||
| Очерк
|
||||||||||||
|
40. |
В гороха, аллель гладких стручков (S) доминирует над аллелем защемленных стручков (ов). Постройте квадрат Пеннета, который показывает нечто среднее между заводом SS и заводом Ss. Предсказать, что процент потомства, вероятно, будет иметь гладкие стручки.
|
|||||||||||
|
41. |
Это Может ли сын унаследовать аллель Х-хромосомы от своего отца? Объясните почему или почему нет.
|
|||||||||||







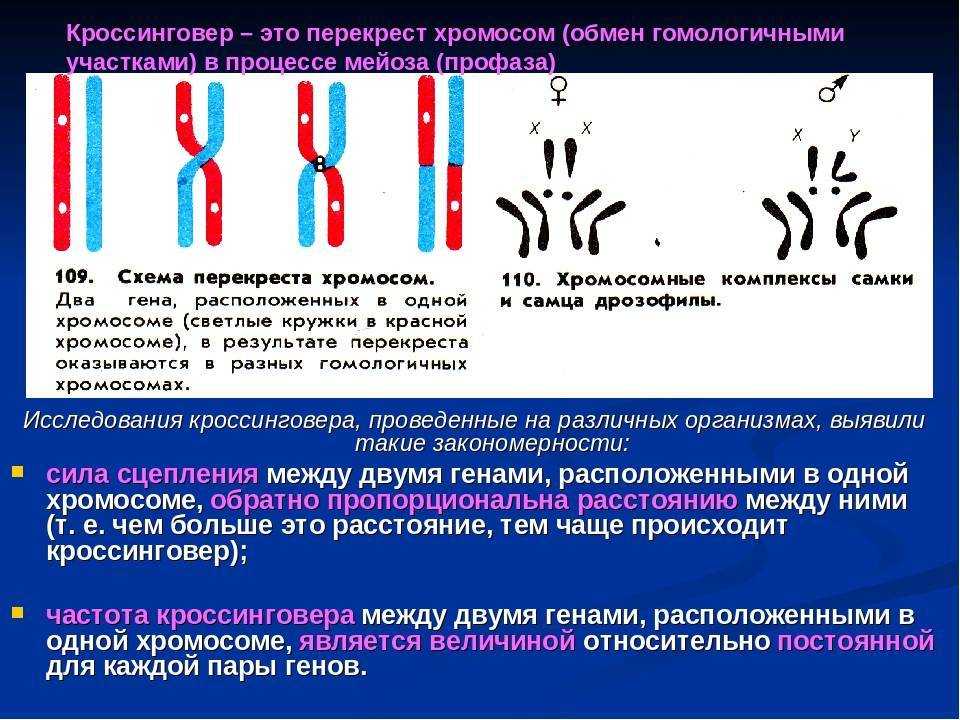













8.4: Простое наследование — биология LibreTexts
Ямочки
У этого человека есть генетическая особенность — ямочки на щеках, когда он улыбается. Генетические признаки — это характеристики, закодированные в ДНК. Некоторые генетические признаки, такие как ямочки на щеках, имеют простой образец наследования, как и черты, которые Грегор Мендель изучал на растениях гороха. То, как эти черты наследуются потомками от родителей, называется простым наследованием.
Генетические признаки — это характеристики, закодированные в ДНК. Некоторые генетические признаки, такие как ямочки на щеках, имеют простой образец наследования, как и черты, которые Грегор Мендель изучал на растениях гороха. То, как эти черты наследуются потомками от родителей, называется простым наследованием.
Что такое простое наследование?
Простое (или менделевское) наследование относится к наследованию признаков, контролируемых одним геном с двумя аллелями, один из которых может быть полностью доминантным по отношению к другому. Тип наследования простых признаков зависит от того, контролируются ли признаки генами на аутосомах или генами на половых хромосомах.
- Аутосомные признаки контролируются генами одной из 22 пар аутосом человека.Аутосомы — это все хромосомы, кроме хромосомы X или Y, и они не различаются между мужчинами и женщинами, поэтому аутосомные черты наследуются одинаково, независимо от пола родителей или потомства.

- Признаки, контролируемые генами половых хромосом, называются признаками, сцепленными с полом. Из-за небольшого размера Y-хромосомы большинство связанных с полом признаков контролируются генами на X-хромосоме. Эти признаки называются X-сцепленными признаками . Х-сцепленные признаки с одним геном имеют другой образец наследования, чем аутосомные признаки с одним геном, потому что у мужчин есть только одна Х-хромосома.Мужчины всегда наследуют свою Х-хромосому от своей матери, и они передают свою Х-хромосому всем своим дочерям, но никому из своих сыновей.
Изучение шаблонов наследования
Есть два очень полезных инструмента для изучения того, как черты передаются от одного поколения к другому. Один инструмент — родословная, другой — квадрат Пеннета.
Родословная
В таблицах ниже указаны родословных . Родословная показывает, как особенность передается из поколения в поколение в семье. Родословная может показать, например, является ли признак аутосомно-доминантным, аутосомно-рецессивным или X-сцепленным признаком. Родословные показывают отношения и идентифицируют людей с заданной чертой. В родословных ниже:
Родословная может показать, например, является ли признак аутосомно-доминантным, аутосомно-рецессивным или X-сцепленным признаком. Родословные показывают отношения и идентифицируют людей с заданной чертой. В родословных ниже:
- пораженных человека показаны красным, здоровые лица показаны синим
- мужчины показаны квадратами, женщины показаны кружками
- Верхний ряд родословной — исходная пара. Две особи, соединенные горизонтальной линией, являются размножающимися парами.Дети пары связаны с ними вертикальными линиями.
- В следующем ряду родословной показаны дети пары, а также партнеры детей. И третий ряд родословной показывает следующее поколение (внуки пары в верхней части родословной). В более крупных родословных может быть больше строк с указанием дополнительных поколений.
Рисунок \ (\ PageIndex {2} \) — это пример родословной аутосомно-доминантного признака. Эта родословная начинается с пораженного самца и незатронутой самки. Первый ребенок пары (слева) — больной мужчина, партнером которой является здоровая женщина. У них рождается двое детей: пораженная женщина и здоровый мужчина. Следующий ребенок — незатронутый самец, находящийся в паре с незатронутой самкой. Их дети — две здоровые женщины и здоровый мужчина. Следующий ребенок — больная девочка, не имеющая партнеров. Четвертый ребенок — незатронутая женщина без партнера. Самый младший ребенок — пораженная женщина, у которой партнерша с здоровым мужчиной. Их дети — пораженный самец, за которым следует здоровый самец, за которым следует больная самка, а за ней — здоровая самка.
Первый ребенок пары (слева) — больной мужчина, партнером которой является здоровая женщина. У них рождается двое детей: пораженная женщина и здоровый мужчина. Следующий ребенок — незатронутый самец, находящийся в паре с незатронутой самкой. Их дети — две здоровые женщины и здоровый мужчина. Следующий ребенок — больная девочка, не имеющая партнеров. Четвертый ребенок — незатронутая женщина без партнера. Самый младший ребенок — пораженная женщина, у которой партнерша с здоровым мужчиной. Их дети — пораженный самец, за которым следует здоровый самец, за которым следует больная самка, а за ней — здоровая самка.
При аутосомно-доминантном признаке у ребенка с этим признаком всегда будет хотя бы один родитель с этим признаком. При аутосомно-рецессивном признаке у двух человек без признака может родиться ребенок с этим признаком.
Рисунок \ (\ PageIndex {3} \): Родословная аутосомно-рецессивного признака.
Рисунок \ (\ PageIndex {3} \) является родословной аутосомно-рецессивного признака. В этой родословной полузатененные особи являются гетерозиготными (и, следовательно, не проявляют признака).В этом примере у двух гетерозиготных особей четверо детей. Их младший ребенок — больной самец. Этот особь спаривается с незатронутой самкой, чтобы произвести на свет двух гетерозиготных детей и двух здоровых детей. Следующая младшая из исходной пары — гетерозиготная женщина. Второй ребенок исходной пары — незатронутый мужчина, который в партнерстве с незатронутой женщиной произвел на свет двух незатронутых детей. Старший ребенок исходной пары — здоровая женщина, партнером которой является гетерозиготный мужчина.Их дети — здоровая самка, здоровый самец и гетерозиготный самец.
Punnett Square
Квадрат Пеннета — это диаграмма, которая позволяет легко определить ожидаемые соотношения возможных генотипов у потомков от двух родителей. Спаривание между двумя родителями называется крестом. Площадь Пеннета названа в честь ее разработчика, британского генетика Реджинальда К. Пеннета. Вы можете увидеть гипотетический пример на рисунке \ (\ PageIndex {4} \). В этом случае ген является аутосомным, и оба родителя являются гетерозиготами (Aa) для этого гена.Половина гамет, производимых каждым родителем, будет иметь аллель A , а половина — аллель и . Это потому, что эти два аллеля находятся на гомологичных хромосомах, которые всегда разделяются и переходят в отдельные гаметы во время мейоза. Согласно закону сегрегации Менделя, аллели в гаметах от каждого родителя записаны на стороне и поперек квадрата Пеннета. Заполнение ячеек квадрата Пеннета дает возможные генотипы их детей.Он также показывает наиболее вероятные соотношения генотипов, которые в данном случае составляют 25 процентов AA, 50 процентов Aa, и 25 процентов AA.
Площадь Пеннета названа в честь ее разработчика, британского генетика Реджинальда К. Пеннета. Вы можете увидеть гипотетический пример на рисунке \ (\ PageIndex {4} \). В этом случае ген является аутосомным, и оба родителя являются гетерозиготами (Aa) для этого гена.Половина гамет, производимых каждым родителем, будет иметь аллель A , а половина — аллель и . Это потому, что эти два аллеля находятся на гомологичных хромосомах, которые всегда разделяются и переходят в отдельные гаметы во время мейоза. Согласно закону сегрегации Менделя, аллели в гаметах от каждого родителя записаны на стороне и поперек квадрата Пеннета. Заполнение ячеек квадрата Пеннета дает возможные генотипы их детей.Он также показывает наиболее вероятные соотношения генотипов, которые в данном случае составляют 25 процентов AA, 50 процентов Aa, и 25 процентов AA.

Прогнозирование генотипов и фенотипа с помощью квадратов Пеннета
Мендель разработал закон сегрегации, следуя только одной характеристике, такой как цвет стручка, у своих растений гороха.В моногибридном скрещивании, таком как тот, что на рисунке \ (\ PageIndex {5} \), квадрат Паннета показывает все возможные комбинации при объединении одного материнского (биологического материнского) аллеля с одним отцовским (биологическим отцовским) аллелем. В этом примере оба организма гетерозиготны по цвету цветов Bb (пурпурный). Оба растения производят гаметы, содержащие аллели B и b. Если гаметы от обоих родителей содержат доминантные аллели, полученное растение будет гомозиготным доминантным и с пурпурными цветками.Если гаметы от обоих родителей содержат рецессивные аллели, полученное растение будет гомозиготным рецессивным и с белыми цветками. Если гамета от одного родителя содержит доминантный аллель, а гамета от другого родителя содержит рецессивный аллель, полученное растение будет гетерозиготным с фиолетовыми цветками. Вероятность того, что у любого единственного потомства появится доминантный признак, составляет 3: 1, или 75%.
Вероятность того, что у любого единственного потомства появится доминантный признак, составляет 3: 1, или 75%.
Дигибридный кросс
Для моногибридного скрещивания мы рассматриваем только один ген. Следовательно, за пределами квадрата Пеннета будут только отдельные буквы (отдельные аллели). Для дигибридного скрещивания используются пары аллелей. Это означает, что снаружи квадрата будут пары букв. Квадрат Пеннета для моногибридного скрещивания делится на четыре квадрата, тогда как квадрат Пеннета для дигибридного скрещивания делится на 16 квадратов.Сколько ящиков потребуется квадрату Пеннета, если исследовать три признака? Квадраты заполнены возможными комбинациями аллелей, образующихся при объединении гамет, например, в зиготе.
Эти типы крестов могут быть сложными в установке, а квадрат, который вы создадите, будет иметь размер 4×4. Это простое руководство проведет вас через этапы решения типичного дигибридного скрещивания, распространенного в генетике. Этот метод также может работать для любого скрещивания, включающего две черты.
Это простое руководство проведет вас через этапы решения типичного дигибридного скрещивания, распространенного в генетике. Этот метод также может работать для любого скрещивания, включающего две черты.
Рассмотрим этот крест
Этот крест фокусируется на двух признаках гороха.
- Признак желтого горошка (Y) является доминирующим по отношению к признаку зеленого горошка (y).
- Признак круглого горошка (R) является доминирующим по отношению к признаку морщинистого гороха (r).
На рисунке \ (\ PageIndex {6} \) показаны кресты двух поколений. В родительском (P) поколении скрещиваются два гомозиготных растения: растение, производящее желтый круглый горошек (YYRR), скрещивается с растением, которое дает зеленый морщинистый горох (yyrr). Квадрат Паннета для этого скрещивания не показан, но все потомки будут гетерозиготными (иметь генотип YyRr) и давать желтый круглый горошек.Чтобы получить это:
- Родитель с генотипом YYRR производит гаметы, все из которых являются YR
- Родитель с генотипом yr производит гаметы, которые имеют возраст
- Гаметы YR и yr дают потомство YyRr

Скрещиваются два гетерозиготных растения (YyRr). Какие гаметы производит каждое из этих растений? Когда образуются гаметы, они могут иметь либо доминантный R, либо рецессивный r.И у них может быть либо доминантный Y, либо рецессивный y. Объедините R и Y каждого родителя, чтобы представить сперму и яйцеклетку.
- Гамета, которая получает аллель Y:
- может либо иметь аллель R, либо быть YR
- или он может получить аллель r и быть Yr
- Гамета, которая получает аллель y:
- может иметь аллель R и быть yR
- или он может получить аллель r и быть годом
Завод YyRr будет производить 4 разных гаметы: YR, Yr, yR, yr.Эти гаметы написаны снаружи квадрата Пеннета на рисунке \ (\ PageIndex {6} \). Затем гаметы объединяются, чтобы сформировать генотипы потомства (написанные в центре квадрата Паннета). Полученное потомство будет иметь следующие соотношения:
- 9/16 круглый, горошек желтый; имея генотипы
- ГГР (1)
- ГГРР (2)
- ГГРр (2)
- ГгРр (4)
- 3/16 круглая, зеленый горошек; с генотипами yyRR (1) и yyRr (2)
- 3/16 горошек желтый, морщинистый; с генотипами YYrr (3)
- 1/16 зеленого горошка морщинистого; имеющий генотип yyrr
Аутосомные моногенные черты человека
Аутосомные признаки с одним геном включают вдовий пик и веснушки, оба из которых проиллюстрированы ниже. Пик вдовы означает точку на линии роста волос в центре лба. Предположим, что доминантный и рецессивный аллели гена пика вдовы представлены W и w соответственно. Поскольку это доминантный признак, люди с генотипом WW и генотипом Ww будут иметь пик вдовы, и только люди с генотипом ww не будут иметь этого признака.
Пик вдовы означает точку на линии роста волос в центре лба. Предположим, что доминантный и рецессивный аллели гена пика вдовы представлены W и w соответственно. Поскольку это доминантный признак, люди с генотипом WW и генотипом Ww будут иметь пик вдовы, и только люди с генотипом ww не будут иметь этого признака.
Предположим, что доминантный и рецессивный аллели веснушек представлены F и f соответственно. Поскольку это доминантный признак, у людей с генотипом FF и генотипом Ff будут веснушки, и только люди с генотипом ff не будут иметь этого признака.
Рисунок \ (\ PageIndex {8} \): Веснушки — аутосомно-доминантный признак с одним геном.Половое наследование
Что определяет, мальчик или девочка ребенок? Вспомните, что у вас 23 пары хромосом, и одна из этих пар — половые хромосомы. Обычно у каждого человека две половые хромосомы. Позже вы узнаете, что из-за нерасхождения у мужчин и женщин может быть на одну меньше или одну дополнительную Х-хромосому. Ваши половые хромосомы могут быть X или Y. У женщин две X-хромосомы (XX), а у мужчин — одна X-хромосома и одна Y-хромосома (XY). Если ребенок унаследует Х-хромосому от отца и Х-хромосому от матери, каков будет пол ребенка? У ребенка будет две Х-хромосомы, поэтому он будет женским. Если сперма отца несет Y-хромосому, ребенок будет мужчиной.Обратите внимание, что мать может передавать только Х-хромосому, поэтому пол ребенка определяется отцом. У отца есть 50-процентная вероятность передачи хромосомы Y или X, поэтому есть 50-процентная вероятность, что ребенок будет мужчиной, и есть 50-процентная вероятность, что ребенок будет женским. Этот шанс 50:50 встречается у каждого ребенка. Первые пять детей пары могут быть мальчиками. Шестой ребенок по-прежнему имеет шанс быть девочкой 50:50. Квадрат Пеннета также можно использовать, чтобы показать, как хромосомы X и Y передаются от родителей к их детям.
Обычно у каждого человека две половые хромосомы. Позже вы узнаете, что из-за нерасхождения у мужчин и женщин может быть на одну меньше или одну дополнительную Х-хромосому. Ваши половые хромосомы могут быть X или Y. У женщин две X-хромосомы (XX), а у мужчин — одна X-хромосома и одна Y-хромосома (XY). Если ребенок унаследует Х-хромосому от отца и Х-хромосому от матери, каков будет пол ребенка? У ребенка будет две Х-хромосомы, поэтому он будет женским. Если сперма отца несет Y-хромосому, ребенок будет мужчиной.Обратите внимание, что мать может передавать только Х-хромосому, поэтому пол ребенка определяется отцом. У отца есть 50-процентная вероятность передачи хромосомы Y или X, поэтому есть 50-процентная вероятность, что ребенок будет мужчиной, и есть 50-процентная вероятность, что ребенок будет женским. Этот шанс 50:50 встречается у каждого ребенка. Первые пять детей пары могут быть мальчиками. Шестой ребенок по-прежнему имеет шанс быть девочкой 50:50. Квадрат Пеннета также можно использовать, чтобы показать, как хромосомы X и Y передаются от родителей к их детям. Это проиллюстрировано на квадрате Пеннета ниже. Это может помочь вам понять схему наследования признаков, связанных с полом.
Это проиллюстрировано на квадрате Пеннета ниже. Это может помочь вам понять схему наследования признаков, связанных с полом.
Х-сцепленные менделевские черты у людей
Одним из примеров признака, связанного с полом, является красно-зеленая дальтонизм.Люди с этим типом дальтонизма не могут отличить красный от зеленого. Они часто видят эти цвета как оттенки коричневого (рисунок \ (\ PageIndex {11} \) ).
Мужчины гораздо чаще страдают дальтонизмом, чем женщины, потому что дальтонизм — это рецессивная характеристика, сцепленная с полом. Поскольку у мужчин только одна Х-хромосома, у них есть только один аллель для любого Х-сцепленного признака. Следовательно, рецессивный X-сцепленный аллель всегда выражен у мужчин. Поскольку у женщин две X-хромосомы, у них есть два аллеля для любого X-сцепленного признака.У женщин может быть одна Х-хромосома с геном дальтонизма и одна Х-хромосома с нормальным геном цветового зрения. Поскольку дальтонизм рецессивен, доминантный нормальный ген будет маскировать рецессивный ген дальтонизма. Самки с одним аллелем дальтонизма и одним нормальным аллелем называются носителями. Они несут аллель, но не выражают его. Самки должны унаследовать две копии рецессивного аллеля, чтобы выразить Х-сцепленный рецессивный признак. Это объясняет, почему Х-сцепленные рецессивные признаки реже встречаются у женщин, чем у мужчин, и почему они демонстрируют иной образец наследования, чем аутосомные признаки.
Поскольку у мужчин только одна Х-хромосома, у них есть только один аллель для любого Х-сцепленного признака. Следовательно, рецессивный X-сцепленный аллель всегда выражен у мужчин. Поскольку у женщин две X-хромосомы, у них есть два аллеля для любого X-сцепленного признака.У женщин может быть одна Х-хромосома с геном дальтонизма и одна Х-хромосома с нормальным геном цветового зрения. Поскольку дальтонизм рецессивен, доминантный нормальный ген будет маскировать рецессивный ген дальтонизма. Самки с одним аллелем дальтонизма и одним нормальным аллелем называются носителями. Они несут аллель, но не выражают его. Самки должны унаследовать две копии рецессивного аллеля, чтобы выразить Х-сцепленный рецессивный признак. Это объясняет, почему Х-сцепленные рецессивные признаки реже встречаются у женщин, чем у мужчин, и почему они демонстрируют иной образец наследования, чем аутосомные признаки.
Согласно этому квадрату Пеннета (Таблица \ (\ PageIndex {1} \)), сын женщины с чертой дальтонизма и мужчины с нормальным зрением с вероятностью 50% страдают дальтонизмом. На рисунке \ (\ PageIndex {10} \) показана простая родословная этого признака.
На рисунке \ (\ PageIndex {10} \) показана простая родословная этого признака.
| х б | X B | |
|---|---|---|
| X B |
х б х ( несущая розетка) |
X B X B (здоровая самка) |
| Y |
X b Y (мужчина дальтоник) |
X B Y (здоровый самец) |
 Дальтонизм — это рецессивный признак, связанный с Х-хромосомой. Мать-носитель и здоровый отец могут произвести на свет здоровых сыновей и дочерей, дочерей-носителей и сыновей с дальтонизмом.
Дальтонизм — это рецессивный признак, связанный с Х-хромосомой. Мать-носитель и здоровый отец могут произвести на свет здоровых сыновей и дочерей, дочерей-носителей и сыновей с дальтонизмом. Другой пример рецессивного Х-сцепленного менделевского признака — гемофилия. Это заболевание, характеризующееся неспособностью крови к нормальному свертыванию. Королева Англии Виктория была разносчиком беспорядка. Две из пяти дочерей королевы Виктории унаследовали аллель гемофилии от своей матери и были носителями. Когда они вступили в брак с членами королевской семьи в других европейских странах, они распространили аллель по всей Европе, включая королевские семьи Испании, Германии и России.Сын Виктории принц Леопольд также унаследовал аллель гемофилии от своей матери и действительно страдал от этого заболевания. По этим причинам гемофилию когда-то в народе называли «королевской болезнью».
Особенность: Мое человеческое тело
Вы дальтоник или думаете, что можете быть? Если вы унаследовали это Х-сцепленное рецессивное заболевание, мир без явных различий между определенными цветами кажется вам нормальным. Это все, что вы когда-либо знали. Вот почему некоторые люди с дальтонизмом даже не подозревают об этом.Были разработаны простые тесты, чтобы определить, страдает ли человек дальтонизмом, и определить степень этого дефицита зрения. Пример такого теста изображен ниже. Что вы видите, когда смотрите на этот круг? Вы отчетливо видите число 74? Если это так, у вас, вероятно, нормальное зрение в красно-зеленом цвете. Если вы не видите номер, у вас может быть красно-зеленая дальтонизм.
Это все, что вы когда-либо знали. Вот почему некоторые люди с дальтонизмом даже не подозревают об этом.Были разработаны простые тесты, чтобы определить, страдает ли человек дальтонизмом, и определить степень этого дефицита зрения. Пример такого теста изображен ниже. Что вы видите, когда смотрите на этот круг? Вы отчетливо видите число 74? Если это так, у вас, вероятно, нормальное зрение в красно-зеленом цвете. Если вы не видите номер, у вас может быть красно-зеленая дальтонизм.
Дальтонизм может вызвать ряд проблем.Они могут варьироваться от незначительных разочарований до явных опасностей. Например:
- Если вы дальтоник, вам может быть сложно согласовать цвета одежды и мебели. Вы можете использовать комбинации цветов, которые люди с нормальным цветовым зрением считают странными или противоречивыми.
- Многие светодиодные индикаторы имеют красный или зеленый цвет.
 Например, удлинители и электронные устройства могут иметь световые индикаторы, показывающие, включены они (зеленый) или выключены (красный).
Например, удлинители и электронные устройства могут иметь световые индикаторы, показывающие, включены они (зеленый) или выключены (красный). - Тест-полоски для определения pH, жесткой воды, химикатов для бассейнов и других стандартных тестов также часто имеют цветовую маркировку.Например, лакмусовая бумага для проверки pH становится красной в присутствии кислоты, но если вы дальтоник, возможно, вы не сможете прочитать результат теста.
- Любите хорошо прожаренный стейк? Если вы страдаете дальтонизмом, возможно, вы не сможете определить, осталось ли мясо недоваренным (красным) или правильно приготовленным на гриле. Вы также не сможете отличить спелые (красные) от незрелых (зеленых) фруктов и овощей, таких как помидоры. А некоторые продукты, такие как темно-зеленый шпинат, могут больше походить на грязь, чем на пищу, и быть совершенно неаппетитными.
- Карты погоды часто имеют цветовую кодировку. Это дождь (зеленый) в вашем прогнозе или зимняя смесь мокрого снега и ледяного дождя (розового или красного)? Если вы не заметите разницы, вы можете выйти на дорогу, когда этого не следует делать, и подвергнуть себя опасности.
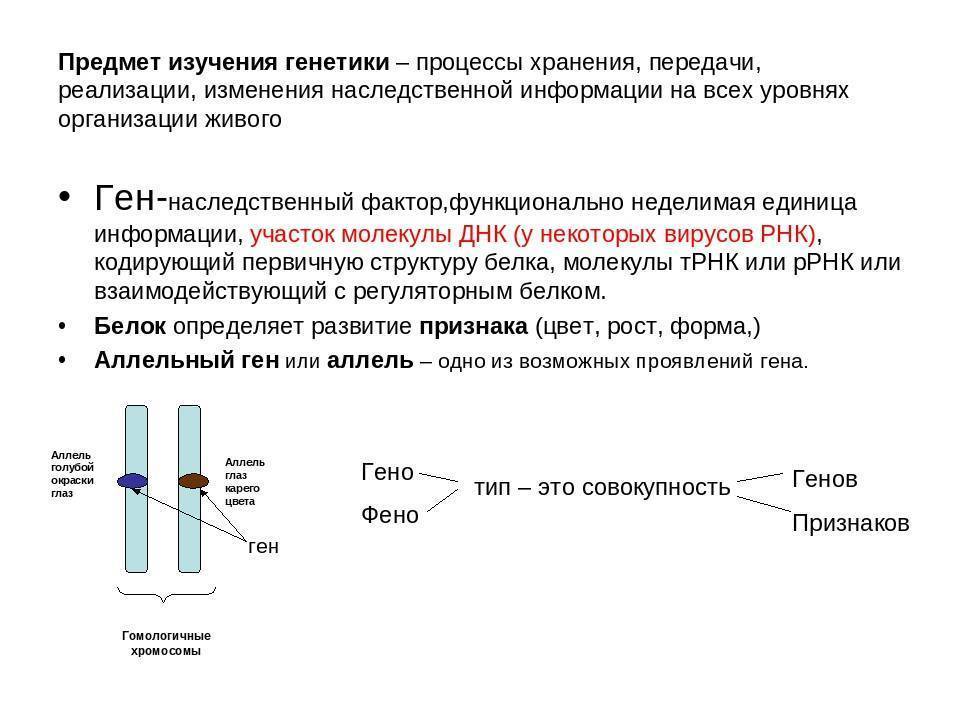
- Умение отличить красный светофор от зеленого может быть вопросом жизни или смерти. Это может быть очень сложно для человека с красно-зеленой дальтонизмом. Вот почему в некоторых странах людям с этим дефектом зрения не разрешается водить машину.
Знание о таких условиях, как дальтонизм, также важно для всех, кто создает контент в Интернете. Создание веб-страниц, удобочитаемых для всех пользователей, является важным навыком для множества рабочих мест. Вы можете использовать онлайн-инструменты (такие как Toptal Color Blind Filter), чтобы убедиться, что созданный вами контент доступен для всех ваших клиентов.
Обзор
- Определите генетические признаки и простое наследование.
- Объясните, почему аутосомные и Х-сцепленные признаки имеют разные модели наследования.
- Что такое родословная и почему она полезна для изучения того, как черты передаются от одного поколения к другому?
- Что такое квадрат Пеннета и что он показывает?
- Укажите примеры человеческих аутосомных и X-сцепленных черт.
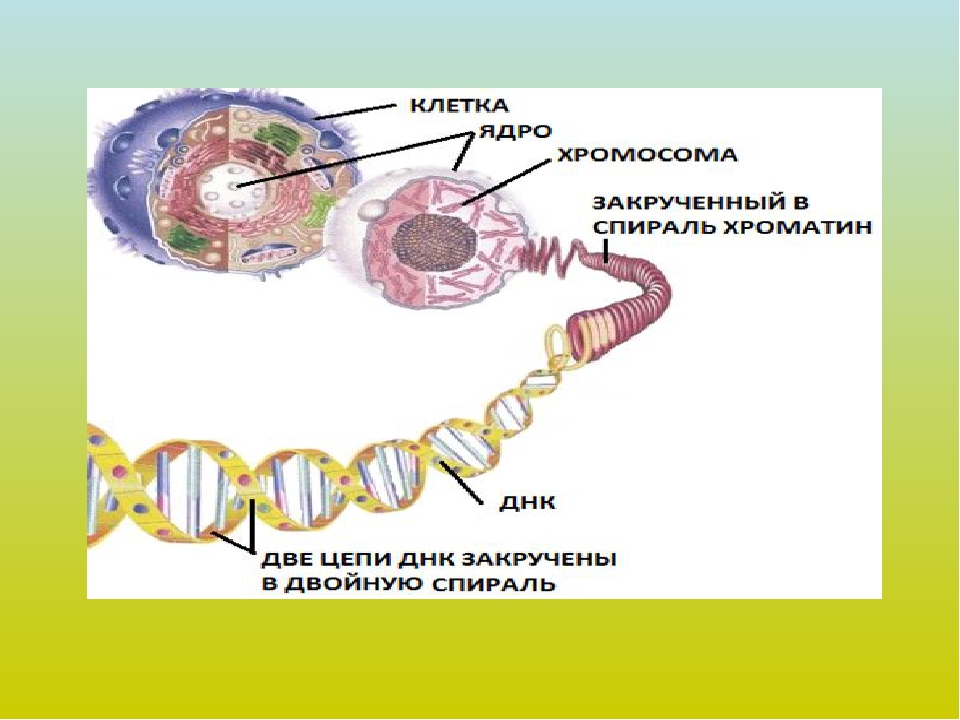
- Представьте себе гипотетический ген человека, который имеет два аллеля: Q и q . Q является доминантным по отношению к q , и наследование этого гена простое.Ответьте на следующие вопросы об этом гене.
- Если женщина имеет генотип Qq , а ее партнер — генотип QQ , перечислите каждую из их возможных гамет и укажите, какая доля их гамет будет иметь каждый аллель.
- Каковы вероятные пропорции их потомков: QQ , Qq или qq ?
- Это аутосомный признак или X-сцепленный признак? Откуда вы знаете?
- Каковы шансы, что их потомство проявит доминантный признак Q ? Поясните свой ответ.
- Объясните, почему отцы всегда передают свою Х-хромосому дочерям.
- Верно или неверно .
 Женщины чаще страдают Х-сцепленными заболеваниями, чем мужчины.
Женщины чаще страдают Х-сцепленными заболеваниями, чем мужчины. - Верно или неверно . Большинство аутосомных черт человека контролируются одним геном с двумя аллелями, как у гороха Менделя.
- Для каждого из приведенных ниже сценариев выберите, будете ли вы использовать квадрат Пеннета или родословную.Выбирайте только тот, который лучше всего подходит для сценария.
- Мужчина и женщина знают генотипы, и вы хотите предсказать возможные генотипы их потомков.
- Вы хотите документально подтвердить, какие члены вашей семьи болели или болеют раком груди.
Атрибуции
- Молодая женщина с ямочками от «Мне так скучно», CC BY-SA 3.0 через Wikimedia Commons
- Аутосомно-доминантная родословная Джерома Уокера, CC BY-SA 3.0 через Wikimedia Commons
- Аутосомно-рецессивная родословная Джерома Уокера, CC BY-SA 3.0 через Wikimedia Commons
- Punnett, miguelferig, выделенный CC0 через Wikimedia Commons
- Площадь Пеннета, автор — Мариана Руис Вильярреал (LadyofHats), CC BY-NC 3.
 0 через CK-12
0 через CK-12 - Дигибридный крест от CNX OpenStax, под лицензией CC BY 4.0 через Wikimedia Commons
- Пик вдовы, созданный Мандипом Гревалом по лицензии CC BY-SA 4.0, из фотографий
- Male Widows Peak, автор Jmblock2, лицензия CC BY-SA 4.0 через Wikimedia Commons
- Omer Mor от Omer Mor по лицензии CC BY-SA 4.0 через Wikimedia Commons
- Freckles от Ayo Ogunseinde, по лицензии Unsplash
- X- и Y-хромосомы Марии Джексон, Лии Маркс, Герхарда Мэй и Джоанны Уилсон, лицензированных CC BY-NC-ND 4.0 через Research Gate
- Родословная дальтонизма от Джоди Со, CC BY-NC 3.0 через CK-12
- Ишара общественное достояние через Wikimedia Commons
- Текст адаптирован из Human Biology по лицензии CK-12 CC BY-NC 3.0
Программа «Происхождение человека» Смитсоновского института
ДНК
Благодаря новостным статьям и криминальным историям мы все знаем, что ДНК в наших клетках отражает уникальную идентичность каждого человека и то, насколько тесно мы связаны друг с другом. То же верно и для взаимоотношений между организмами. ДНК или дезоксирибонуклеиновая кислота — это молекула, которая составляет геном организма в ядре каждой клетки. Он состоит из генов, которые представляют собой молекулярные коды белков — строительных блоков наших тканей и их функций. Он также состоит из молекулярных кодов, которые регулируют выработку генов, то есть время и степень образования белка. ДНК определяет рост организма и физиологию его крови, костей и мозга.
То же верно и для взаимоотношений между организмами. ДНК или дезоксирибонуклеиновая кислота — это молекула, которая составляет геном организма в ядре каждой клетки. Он состоит из генов, которые представляют собой молекулярные коды белков — строительных блоков наших тканей и их функций. Он также состоит из молекулярных кодов, которые регулируют выработку генов, то есть время и степень образования белка. ДНК определяет рост организма и физиологию его крови, костей и мозга.
ДНК, таким образом, особенно важна в изучении эволюции. Количество различий в ДНК — это критерий различия между одним видом и другим — и, следовательно, насколько они близки или отдалены друг от друга.
В то время как генетическая разница между отдельными людьми сегодня ничтожна — в среднем около 0,1%, — исследование одних и тех же аспектов генома шимпанзе показывает разницу примерно в 1,2%. Бонобо ( Pan paniscus ), который является близким родственником шимпанзе ( Pan troglodytes ), отличается от человека в той же степени. Разница в ДНК горилл, другой африканской обезьяны, составляет около 1,6%. Что наиболее важно, шимпанзе, бонобо, и человек демонстрируют одинаковое отличие от горилл. Разница в 3,1% отличает нас и африканских обезьян от азиатской большой обезьяны, орангутана. Как складываются обезьяны? Все человекообразные обезьяны и люди отличаются от макак-резусов, например, примерно на 7% по своей ДНК.
Разница в ДНК горилл, другой африканской обезьяны, составляет около 1,6%. Что наиболее важно, шимпанзе, бонобо, и человек демонстрируют одинаковое отличие от горилл. Разница в 3,1% отличает нас и африканских обезьян от азиатской большой обезьяны, орангутана. Как складываются обезьяны? Все человекообразные обезьяны и люди отличаются от макак-резусов, например, примерно на 7% по своей ДНК.
Генетики придумали множество способов вычисления процентов, которые дают разные представления о том, насколько похожи шимпанзе и люди.Например, различие между шимпанзе и человеком на 1,2% включает измерение только замен в базовых строительных блоках тех генов, которые являются общими для шимпанзе и человека. Однако сравнение всего генома показывает, что сегменты ДНК также были удалены, дублированы снова и снова или вставлены из одной части генома в другую. Если подсчитать эти различия, можно обнаружить еще 4-5% различий между геномами человека и шимпанзе.
Независимо от того, как производятся вычисления, главный момент все же остается в силе: люди, шимпанзе и бонобо более тесно связаны друг с другом, чем с гориллами или другими приматами. С точки зрения этого мощного теста на биологическое родство, люди не только родственны человекообразным обезьянам — мы едины. Доказательства ДНК оставляют нас с одним из величайших сюрпризов в биологии: стена между человеком, с одной стороны, и обезьяной или животным, с другой, была прорвана. Древо эволюции человека встроено в человекообразных обезьян.
С точки зрения этого мощного теста на биологическое родство, люди не только родственны человекообразным обезьянам — мы едины. Доказательства ДНК оставляют нас с одним из величайших сюрпризов в биологии: стена между человеком, с одной стороны, и обезьяной или животным, с другой, была прорвана. Древо эволюции человека встроено в человекообразных обезьян.
Сильное сходство между людьми и африканскими приматами привело к тому, что Чарльз Дарвин в 1871 году предсказал, что Африка была вероятным местом, где человеческое происхождение ответвилось от других животных, то есть местом, где был общий предок шимпанзе, людей и горилл. когда-то жил.Доказательства ДНК показывают удивительное подтверждение этого смелого предсказания. Африканские человекообразные обезьяны, включая людей, имеют более тесные родственные связи друг с другом, чем африканские обезьяны с орангутанами или другими приматами. Едва ли когда-либо существовало такое смелое, столь «доступное» для своего времени научное предсказание, как сделанное в 1871 году, когда человеческая эволюция началась в Африке.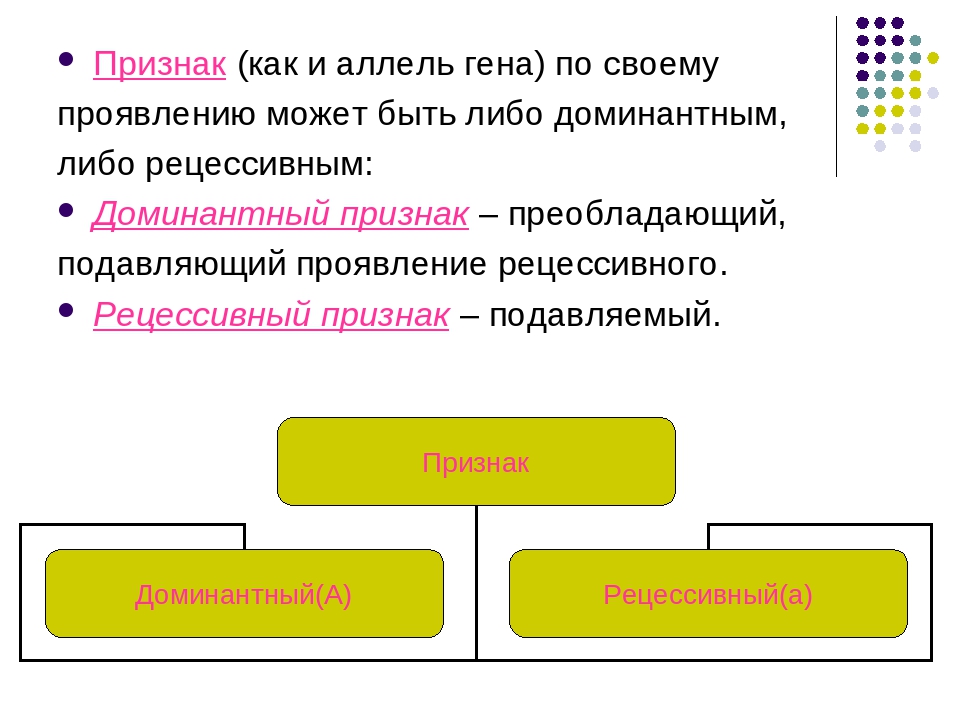
Доказательства ДНК подтверждают этот вывод, и окаменелости тоже. Несмотря на то, что Европа и Азия искали в поисках ранних человеческих окаменелостей задолго до того, как даже задумывались об Африке, продолжающиеся открытия окаменелостей подтверждают, что первые 4 миллиона лет или около того в истории эволюции человека имели место исключительно на африканском континенте.Именно там продолжаются поиски окаменелостей в точке ветвления шимпанзе и человеческих родословных от нашего последнего общего предка или около нее.
Генеалогическое древо приматов
В результате миллиардов лет эволюции люди имеют общие гены со всеми живыми организмами. Процент генов или ДНК, которые разделяют организмы, фиксирует их сходство. Мы разделяем больше генов с организмами, которые более тесно связаны с нами.
Люди принадлежат к биологической группе, известной как приматы, и относятся к человекообразным обезьянам, одной из основных групп эволюционного древа приматов. Помимо сходства в анатомии и поведении, на наше близкое биологическое родство с другими видами приматов указывают данные ДНК. Это подтверждает, что наши ближайшие живые биологические родственники — шимпанзе и бонобо, с которыми мы имеем много общих черт. Но мы не произошли напрямую от приматов, живущих сегодня.
Помимо сходства в анатомии и поведении, на наше близкое биологическое родство с другими видами приматов указывают данные ДНК. Это подтверждает, что наши ближайшие живые биологические родственники — шимпанзе и бонобо, с которыми мы имеем много общих черт. Но мы не произошли напрямую от приматов, живущих сегодня.
также показывает, что наш вид и шимпанзе произошли от общего вида-предка, который жил между 8 и 6 миллионами лет назад. Последний общий предок обезьян и обезьян жил около 25 миллионов лет назад.
(© Авторское право Смитсоновского института) .