
biologia_2
Открытие генетической роли ДНК.
К 1944 г. О. Эйвери и его коллеги К. Маклеод и М. Маккарти открыли трансформирующую активность ДНК у пневмококков. Эти авторы продолжили работу Гриффита, описавшего феномен трансформации (передачи наследственных признаков) у бактерий. О. Эйвери, К. Маклеод, М. Маккарти показали, что при удалении белков, полисахаридов и РНК трансформация бактерий не нарушается, а при воздействии на индуцирующее вещество ферментом дезоксирибонуклеазой трансформирующая активность исчезает.
В этих экспериментах впервые была продемонстрирована генетическая роль молекулы ДНК. В 1952 г. А. Херши и М. Чейз подтвердили генетическую роль молекулы ДН К в опытах на бактериофаге Т2. Пометив его белок радиоактивной серой, а ДНК-радиоактивным фосфором,они инфицировали этим бактериальным вирусом кишечную палочку Е. coli. В потомстве фага было выявлено большое количество радиоактивного фосфора и лишь следы S. Отсюда следовало, что именно ДНК, а не белок фага проникает в бактерию, а затем после репликации передается фаговому потомству.
Строение нуклеотида ДНК. Типы нуклеотидов.
Нуклеотид ДНК состоит из
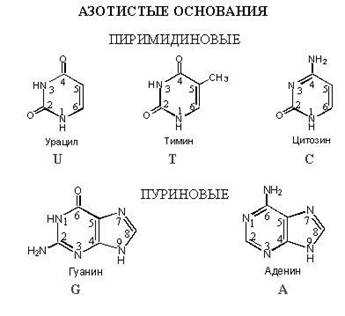
— азотистого основания (в ДНК 4 типа: аденин, тимин, цитозин, гуанин)
— моносахара дезоксирибозы
— фосфорной кислоты
Молекула нуклеотида состоит из трех частей — пятиуглеродного сахара, азотистого основания и фосфорной кислоты.
Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т. е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два типа нуклеиновых кислот — рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дезоксирибозу. В дезоксирибозе — ОН-группа при 2-м атоме углерода заменена на атом Н, т. е. в ней на один атом кислорода меньше, чем в рибозе.
В обоих типах нуклеиновых кислот содержатся основания четырех разных видов: два из них относятся к классу пуринов и два — к классу пиримидинов. Основной характер этим соединениям придает включенный в кольцо азот. К числу пуринов относятся аденин (А) и гуанин (Г), а к числу пиримидинов — цитозин (Ц) и тимин (Т) или урацил (У) (соответственно в ДНК или РНК). Тимин химически очень близок к урацилу (он представляет собой 5-метилурацил, т. е. урацил, в котором у 5-го углеродного атома стоит метильная группа). В молекуле пуринов имеется два кольца, а в молекуле пиримидинов — одно.
Нуклеотиды соединяются между собой прочной ковалентной связью через сахар одного нуклеотида и фосфорную кислоту другого. Получаетсяполинуклеотидная цепь. На одном ее конце – свободная фосфорная кислота (5’-конец), на другом – свободный сахар (3’-конец). (ДНК-полимераза может присоединять новые нуклеотиды только к 3’-концу.)
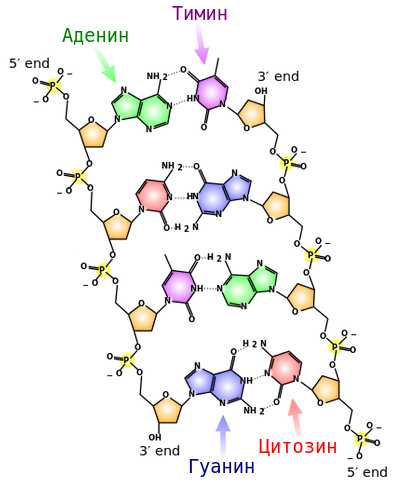
Две полинуклеотидные цепи соединяются друг с другом слабыми водородными связями между азотистыми основаниями. Соблюдаются 2 правила:
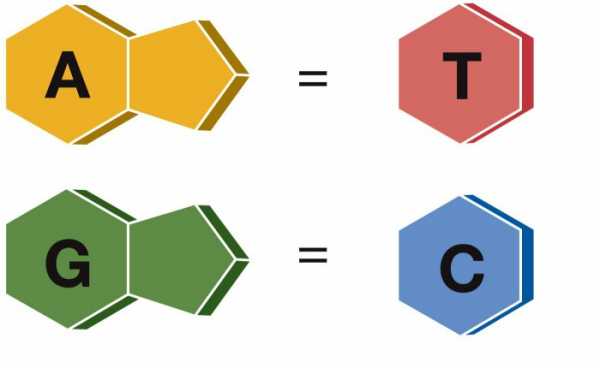
принцип комплементарности: напротив аденина всегда стоит тимин, напротив цитозина – гуанин (они подходят друг другу по форме и числу водородных связей – между А и Г две связи, между Ц и Г – 3).
принцип антипараллельности: там, где у одной полинуклеотидной цепи 5’-конец, у другой – 3’-конец, и наоборот.
Получается двойная цепь ДНК.
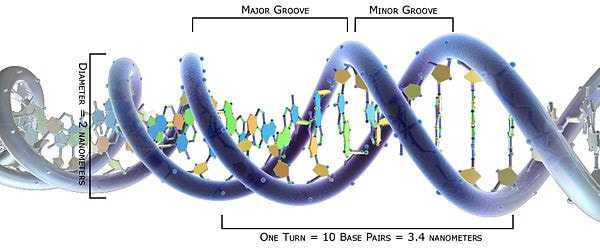
Она скручивается в двойную спираль, один виток спирали имеет длину 3,4 нм, содержит 10 пар нуклеотидов. Азотистые основания (хранители генетической информации) находятся внутри спирали, защищенные.
Структурная организация молекулы ДНК. Модель Дж.Уотсона и Ф.Крика
В 1950 г. английский физик М.Уилкинс получил рентгенограмму кристаллических волокон ДНК. Она показала, что молекула ДНК имеет определенную структуру, расшифровка которой помогла бы понять механизм функционирования ДНК. Рентгенограммы, полученные не на кристаллических волокнах ДНК, а на менее упорядоченных агрегатах, которые образуются при более высокой влажности, позволили Розалинд Франклин , коллеге М. Уилкинса, увидеть четкий крестообразный рисунок — опознавательный знак двойной спирали. Стало известно также, что нуклеотиды расположены друг от друга на расстоянии 0,34 нм, а на один виток спирали их приходится 10. Диаметр молекулы ДНК составляет около 2 нм. Из рентгеноструктурных данных, однако, было не ясно, каким образом цепи удерживаются вместе в молекулах ДНК.
Картина полностью прояснилась в 1953 г., когда американский биохимик Дж. Уотсон и английский физик Ф. Крик , исследуя структуру молекулы ДНК, пришли к выводу, что сахарофосфатный остов находится на периферии молекулы ДНК, а пуриновые и пиримидиновые основания — в середине. Причем последние ориентированы таким образом, что между основаниями из противоположных Цепей могут образоваться водородные связи. Из построенной ими модели выявилось, что какой-либо пурин в одной цепи всегда связан водородными связями с одним из пиримидинов в другой цепи. Такие пары имеют одинаковый размер по всей длине молекулы. Не менее важно то, что аденин может спариваться лишь с тимином, а гуанин только с с цитозином. При этом между аденином и тимином образуются две водородные связи, а между гуанином и цитозином – три.
Свойства и функции ДНК.
Хранение наследственной информации (генетический код – способ записи ген.информации о последовательности аминокислот в белке с помощью нуклеотидов (Гамов)
Передача (репликация/удвоение)
Реализация (транскрипция)
Ауторепродукция ДНК. Репликон и его функционирование.
Процесс самовоспроизведения молекул нуклеиновых кислот, сопровождающийся передачей по наследству (от клетки к клетке) точных копий генетической информации; осуществляется с участием набора специфических ферментов (геликаза, контролирующая расплетание молекулы ДНК, ДНК-полимеразы, ДНК-лигаза), проходит по полуконсервативному типу с образованием репликативной вилки; на одной из цепей синтез комплементарной цепи непрерывен, а на другой происходит за счет образования фрагментов Дказаки. Высокоточный процесс, частота ошибок при котором не превышает 10-9; у эукариот может происходить сразу в нескольких точках одной молекулы ДНК; скорость у эукариот около 100, а у бактерий — около 1000 нуклеотидов в сек.
Репликон — единица процесса репликации участка генома, который находится под контролем одной точки инициации (начала) репликации. Термин предложен Ф. Жакобом и С. Бреннером в 1963 году. Геном прокариот представляет собой, как правило, один репликон. От точки инициации репликация идёт в обе стороны, в некоторых случаях с неравной скоростью. У эукариот геном состоит из многих (часто до неск. десятков тысяч) репликонов.
Генетический код, его свойства.
Генетический код – способ записи генетической инофрмации о последовательности аминокислот в белке с помощью нуклеотидов. Открытие ген. Кода принадлежит Георгию Гамову. 1954год.
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно.
Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин)[1]
Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов.
Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже).
Помехоустойчивость — мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
Понятие о гене. Свойства Гена.
Ген — структурная и функциональная единица наследственности живых организмов. Ген представляет собой последовательность ДНК, задающую последовательность определённого полипептида либо функциональной РНК. Гены определяют наследственные признаки организмов, передающиеся от родителейпотомству при размножении. При этом некоторые органеллы (митохондрии, пластиды) имеют собственную ДНК, не входящую в геном организма, которая определяет их признаки.
( Термин введен в 1909 году датским ботаником Вильгельмом Йогансеном)
дискретность — несмешиваемость генов;
стабильность — способность сохранять структуру;
лабильность — способность многократно мутировать;
множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
аллельность — в генотипе диплоидных организмов только две формы гена;
специфичность — каждый ген кодирует свой признак;
плейотропия — множественный эффект гена;
экспрессивность — степень выраженности гена в признаке;
пенетрантность — частота проявления гена в фенотипе;
амплификация — увеличение количества копий гена.
Особенности организации генома эукариот.
Геном эукариот:
большое число генов,
большее количество ДНК,
в хромосомах имеется очень сложная система контроля активности генов во времени и пространстве, связанная с дифференциацией клеток и тканей в онтогенезе организма.
Количество ДНК в хромосомах велико и возрастает по мере усложнения организмов. Для эукариот также характернаизбыточность генов. Так, у человека геном содержит число нуклеотидных пар, достаточное для образования более 2 млн. структурных генов, в то время как у человека имеется по данным 2000 года 31 тыс. всех генов.
Больше половины гаплоидного набора генома эукариотов составляют уникальные гены, представленные лишь по одному разу. У человека таких уникальных генов — 64%, у теленка — 55%, у дрозофилы — 70%.
Классы нуклеотидных последовательностей в ДНК эукариот, их характеристика, свойства и биологичесок значение.
Нуклеотидные последовательности в геноме эукариот
В конце 60-х годов работами американских ученых Р. Бриттена, Э. Дэвидсона и других была открыта фундаментальная особенность молекулярной структуры генома эукариот – нуклеотидные последовательности разной степени повторяемости. Это открытие было сделано с помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК. Различают следующие фракции в геноме эукариот.
1. Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки.
2. Низкочастотные повторы – последовательности, повторяющиеся десятки раз.
3. Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов.
4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие (~ 10 пн) некодирующие последовательности, которые входят в состав прицентромерного гетерохроматина.
Уровни организации генома эукариот.
Химический и структурный состав хромосом.
Молекулярно-биологические исследования позволили получить представление не только о химической структуре хромосом, но также и об их надмолекулярной организации и особенностях функционирования. В настоящее время известно, что хромосомы представляют собой нуклеопротеидные образования, состоящие из ДНК и белка. Кроме того, в хромосомах присутствует некоторое количество РНК, образующейся при транскрипции, и ионы Са+ и Mg+. Каждая хроматида, а в промежутке времени анафаза- S -период интерфазы и хромосома, содержит одну молекулу ДНК, которая определяет все функции хромосомы, связанные с хранением наследственной информации, её передачей и реализацией. Молекула ДНК в хромосомах тесно связана с двумя классами белков- гистонами (основные белки) и негистонами (кислые белки). Гистоны — это небольшие по величине белки с высоким содержанием заряженных аминокислот (лизина и аргинина). Суммарный положительный заряд позволяет гистонам связываться с ДНК независимо от нуклеотидного состава. Им принадлежит в основном структурная функция. Это очень стабильные белки, молекулы которых могут сохраняться в течение всей жизни клетки. В эукариотической клетке присутствуют 5 типов гистонов, которые распределяются на две основные группы: первая группа (их обозначают как Н2А, Н2В, НЗ, Н4), отвечает за формирование специфических дезоксирибонуклеопротеидных комплексов — нуклеосом. Вторая группа гистонов (HI) располагается между нуклеосомами и фиксирует укладку нуклеосомной цепи в более высокий уровень структурной организации (супернуклеосомную нить). Среди гистоновых белков, кроме структурных, встречаются такие, которые способны ограничивать доступность ДНК для ДНК — связывающих регуляторных белков и тем самым участвовать в регуляции активности генов. Негистоновые белки весьма разнообразны. Число их фракций превышает 100. Они присутствуют в меньших количествах в хромосомах в сравнении с гистонами и выполняют в основном регуляторную функцию. Участвуют в регуляции транскрипционной активности генов, в обеспечении редупликации и репарации ДНК. Большинство негистоновых белков хроматина присутствуют в клетках в небольшом количестве (минорные) — это регуляторные белки, узнающие специфические последовательности ДНК и связывающиеся с ними. Они вовлечены во многие генетические процессы, но известно о них пока что немного. Количественно преобладают негистоновые белки (мажорные), высокоподвижные, относительно малого размера, с большим электрическим зарядом — они всегда соединяются с нуклеосомами, содержащими активные гены. Кроме того, в группу негистоновых белков входит много ферментов.
Уровни упаковки наследственного материала у эукариот.
Таким образом, уровни упаковки ДНК следующие:
1) Нуклеосомный (2,5 оборота двуспиральной ДНК вокруг восьми молекул гистоновых белков).
2) Супернуклеосомный — хроматиновая спираль (хромонема).
3) Хроматидный — спирализованная хромонема.
4) Хромосома — четвертая степень сперализации ДНК.
В интерфазном ядре хромосомы деконденсированы и представлены хроматином. Деспирализованный участок, содержащий гены, называется эухроматин (разрыхленный, волокнистый хроматин). Это необходимое условие для транскрипции. Во время покоя между делениями определенные участки хромосом и целые хромосомы остаются компактными.
Эти спирализованные, сильно окрашивающиеся участки, называются гетерохроматином. Они неактивны в отношении транскрипции. Различают факультативный и конститутивный гетерохроматин.
Факультативный гетерохроматин информативен, т.к. содержит гены и может переходить в эухроматин. Из двух гомологичных хромосом одна может гетерохроматической. Конститутивный гетерохроматин всегда гетерохроматичен, неиформативен (не содержит генов) и поэтому всегда неактивен в отношении транскрипции.
Хромосомная ДНК состоит из более 108 пар оснований, из которых образуется информативные блоки — гены, расположенные линейно. На их долю приходится до 25% ДНК. Ген — функциональная единица ДНК, содержащая информацию для синтеза полипептидов, или всех РНК. Между генами находятся спейсеры — неинформативные отрезки ДНК разной длины. Избыточные гены представлены большим числом — 104 идентичных копий. Примером являются гены для т-РНК, р-РНК, гистонов. В ДНК встречаются последовательности одних и тех же нуклеотидов. Они могут быть умеренно повторяющимися и высоко повторяющимися последовательностями. Умеренно повторяющиеся последовательности достигают 300 пар нуклеотидов с повторениями 102 — 104 и представляют чаще всего спейсеры, избыточные гены.
Высокоповторяющиеся последовательности (105 — 106) образуют конститутивный гетерохроматин. Около 75% всего хроматина не участвует в транскрипции, он приходится на высокоповторяющиеся последовательности и нетранскрибируемые спейсеры.
Морфологические особенности метафазной хромосомы.
Митотическая суперкомпактизация хроматина делает возможным изучение внешнего вида хромосом с помощью световой микроскопии. В первой половине митоза они состоят из двух хроматид, соединенных между собой в области первичной перетяжки (центромеры или кинетохора) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.
В зависимости от места положения центромеры и длины плеч, расположенных по обе стороны от нее, различают несколько форм хромосом: равноплечие, или метацентрические (с центромерой посередине), неравноплечие, или субметацентрические (с центромерой, сдвинутой к одному из концов), палочковидные, или акроцентрические (с центромерой, расположенной практически на конце хромосомы), и точковые —очень небольшие, форму которых трудно определить (рис. 3.52). При рутинных методах окраски хромосом они различаются по форме и соотносительным размерам. При использовании методик дифференциальной окраски выявляется неодинаковая флуоресценция или распределение красителя по длине хромосомы, строго специфические для каждой отдельной хромосомы и ее гомолога (рис. 3.53).
Таким образом, каждая хромосома индивидуальна не только по заключенному в ней набору генов, но и по морфологии и характеру дифференциального окрашивания.
Эу- и гетерохроматин, их биологическое значение.
Некоторые хромосомы во время клеточного деления выглядят конденсированными и интенсивно окрашенными. Такие различия были названы гетеропикнозом. Для обозначения районов хромосом, демонстрирующих положительный гетеропикноз на всех стадиях митотического цикла был предложен термин «гетерохроматин». Различают эухроматин — основную часть митотических хромосом, которая претерпевает обычный цикл компактизации декомпактизации во время митоза, и гетерохроматин — участки хромосом, постоянно находящиеся в компактном состоянии.
У большинства видов эукариот хромосомы содержат как эу-, так и гетерохроматиновые участки, причем последние составляют значительную часть генома. Гетерохроматин располагается в прицентромерных, иногда в прителомерных областях. Обнаружены гетерохроматиновые участки в эухроматиновых плечах хромосом. Они выглядят как вкрапления (интеркаляции) гетерохроматина в эухроматин. Такой гетерохроматин называют интеркалярным. Компактизация хроматина. Эухроматин и гетерохроматин различаются по циклам компактизации. Эухр. проходит полный цикл компактизации-декомпактизации от интерфазы до интерфазы, гетеро. сохраняет состояние относительной компактности. Дифференциальная окрашиваемость. Разные участки гетерохроматина окрашиваются разными красителями, некоторые районы — каким-то одним, другие — несколькими. Применяя различные красители и используя хромосомные перестройки, разрывающие гетерохроматиновые районы, у дрозофилы удалось охарактеризовать много небольших районов, где сродство к окраскам отлично от соседних участках.
Понятие о кариотипе (определение).Обща характеристика кариотипа человека.
Кариотип — диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида, являющийся видоспецифическим признаком и характеризующийся определенным числом, строением и генетическим составом хромосом.
Если число хромосом в гаплоидном наборе половых клеток обозначить п, то общая формула кариотипа будет выглядеть как 2п, где значение п различно у разных видов. Являясь видовой характеристикой организмов, кариотип может отличаться у отдельных особей некоторыми частными особенностями. Например, у представителей разного пола, имеются в основном одинаковые пары хромосом (аутосомы), но их кариотипы отличаются по одной паре хромосом (гетерохромосомы, или половые хромосомы). Иногда эти различия состоят в разном количестве гетерохромосом у самок и самцов (XX или ХО). Чаще различия касаются строения половых хромосом, обозначаемых разными буквами —X и Y (XX или XY).
Каждый вид хромосом в кариотипе, содержащий определенный комплекс генов, представлен двумя гомологами, унаследованными от родителей с их половыми клетками. Двойной набор генов, заключенный в кариотипе,— генотип — это уникальное сочетание парных аллелей генома. В генотипе содержится программа развития конкретной особи.
Денверская (1960) и Парижская (1971) классификация хромосом человека: основные принципы и сущность.
Денверская и Парижская классификация хромосом Хромосомы подразделяются на аутосомы (соматических клеток) и гетерохромосомы (половых клеток). По предложению Левитского (1924) диплоидный набор соматических хромосом клетки был назван кариотипом. Он характеризуется числом, формой, размерами хромосом. Для описания хромосом кариотипа по предложению С.Г. Навашина их располагают в виде идиограммы — систематизированного кариотипа. В 1960 году была предложена Денверская международная классификация хромосом, где хромосомы классифицированы по величине и расположению центромеры. В кариотипе соматической клетки человека различают 22 пары аутосом и пару половых хромосом. Набор хромосом в соматических клетках называют диплоидным, а в половых клетках —гаплоидным (он равен половине набора аутосом). В идиограмме кариотипа человека хромосомы делят на 7 групп, в зависимости от их размеров и формы. 1 — 1-3 крупные метацентрические. 2 — 4-5 крупные субметацентрические. 3 — 6-12 и Х-хромосома средние метацентрические. 4 — 13-15 средние акроцентрические. 5 — 16-18 относительно малые мета-субметацентрические. 6 — 19-20 малые метацентрические. 7 — 21-22 и Y-хромосома наиболее малые акроцентрические. Согласно Парижской классификации хромосомы разделены на группы по их размерам и форме, а также линейной дифференцировке.
studfiles.net
Нуклеотиды | Кинезиолог
Определение понятия
Нуклеотиды — это сложные мономеры, из которых собраны гетерополимерные молекулы. ДНК и РНК. Свободные нуклеотиды участвуют в сигнальных и энергетических процессах жизнедеятельности. ДНК-нуклеотиды и РНК-нуклеотиды имеют общий план строения, но различаются по строению сахара-пентозы. В ДНК-нуклеотидах используется сахар дезоксирибоза, а в РНК-нуклеотидах — рибоза.
Структура нуклеотида
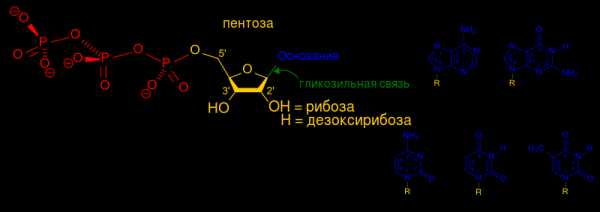
В каждом нуклеотиде можно выделить 3 части:
1. Углевод — это пятичленный сахар-пентоза (рибоза или дезоксирибоза).
2. Фосфорный остаток (фосфат) — это остаток фосфорной кислоты.
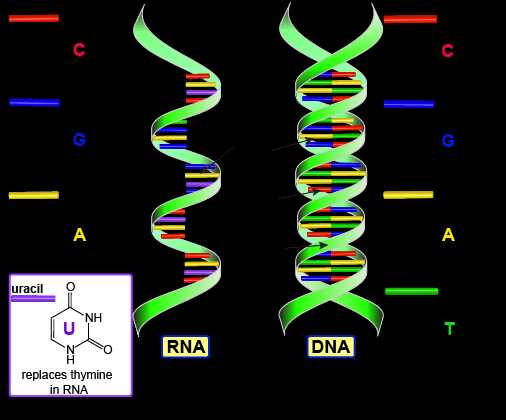
3. Азотистое основание — это соединение, в котором много атомов азота. В нуклеиновых кислотах используется всего 5 видов азотистых оснований : Аденин, Тимин, Гуанин, Цитозин, Урацил. В ДНК — 4 вида: Аденин, Тимин, Гуанин, Цитозин. В РНК — тоже 4 вида: Аденин, Урацил, Гуанин, Цитозин, Легко заметить, что в РНК происходит замещение Тимина на Урацил по сравнению с ДНК.
Общая структурная формула пентозы (рибозы или дезоксирибозы), молекулы которой образуют «скелет» нуклеиновых кислот:
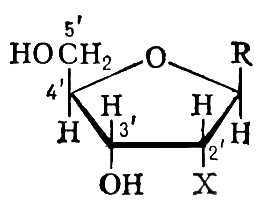
Если Х заменить на Н (Х = Н) — то получаются дезоксирибонуклеозиды; если Х заменить на ОН (Х = ОН) — то получаются рибонуклеозиды. Если вместо R подставить азотистое основание (пуриновое или пиримидиновое) — то получится конкретный нуклеотид.
Важно обратить внимание на те положения атомов углерода в пентозе, которые обозначены как 3′ и 5′. Нумерация атомов углерода начинается от атома кислорода вверху и идёт по часовой стрелке. Последним получается атом углерода (5′), который располагается за пределами пентозного кольца и образует, можно сказать, «хвостик» у пентозы. Так вот, при наращивании цепочки из нуклеотидов фермент может присоединить новый нуклеотид только к углероду 3′ и ни к какому другому. Поэтому 5′-конец нуклеотидной цепочки никогда не сможет иметь продолжения, удлинняться может только 3′-конец.
Азотистые основания
Нуклеотиды
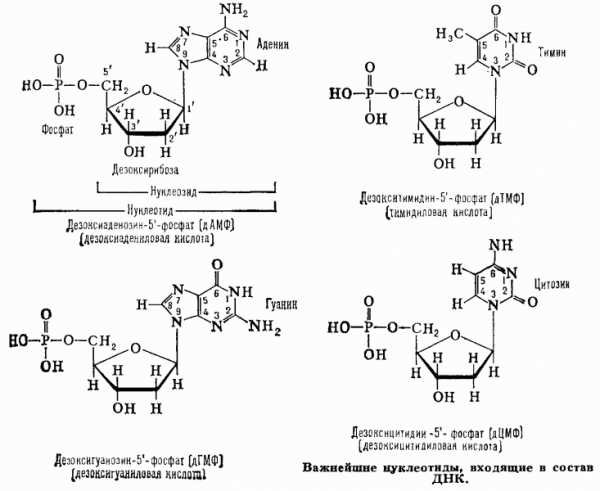
Сравните нуклеотид для РНК с нуклеотидом для ДНК.
Попробуйте узнать, какой это нуклеотид, в таком представлении:
АТФ — свободный нуклеотид
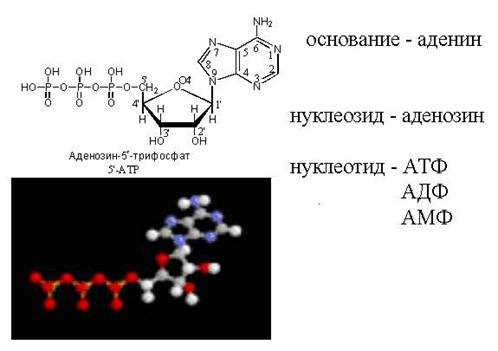
цАМФ — «закольцованная» молекула АТФ
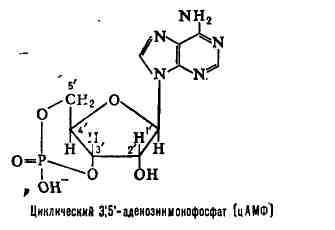
Схема строения нуклеотида
Обратите внимание на то, что активированный нуклеотид, способный наращивать цепочку ДНК или РНК, имеет «трифосфатный хвостик». Именно этим «энергонасыщенным» хвостиком он может присоединиться к уже имеющейся цепочке растущей нуклеиновой кислоты. Фосфатный хвостик сидит на 5-м атоме углерода, так что это положение углерода уже занято фосфатами и предназнено для прикрепления. К чему же его прикрепить? Только к углероду в положении 3′. После прикрепления данный нуклеотид сам станет мишенью дла прикрепления следующего нуклеотида. «Принимающая сторона» предоставляет углерод в положении 3′, а «прибывающая сторона» цепляется к нему фосфатным хвостиком, находящимся в положении 5′. В целом цепочка растёт со стороны 3′.
Наращивание нуклеотидной цепочки ДНК
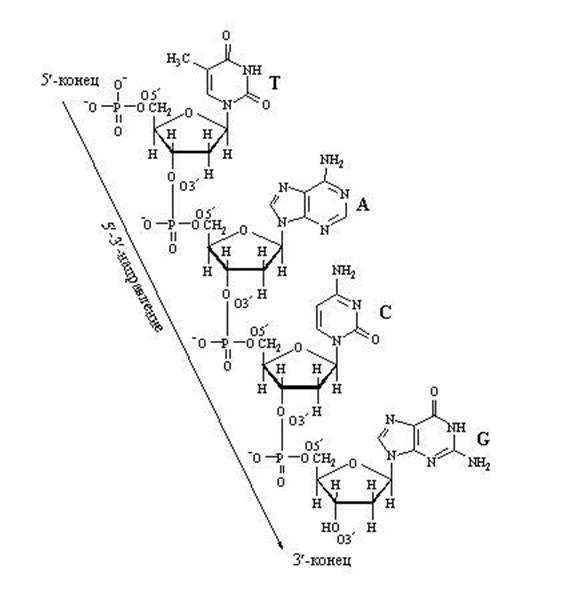
Наращивание цепочки за счёт «продольных» связей между нуклеотидами может идти только в одном направлении: от 5′ ⇒ к 3′, т.к. новый нуклеотид можно присоединить только к 3′-концу цепочки, но не к 5′-концу.
Пары нуклеотидов, связанные «поперечными» комплементарными связями своих азотистых оснований
Участок двойной спирали ДНК
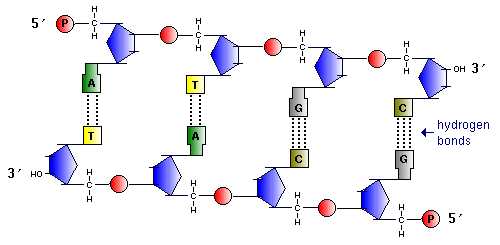
Найдите признаки антипараллельности двух цепей ДНК.
Найдите пары нуклеотидов с двойными и тройными комплементарными связями.
kineziolog.su
Типы нуклеиновых кислот — строение нуклеотида ДНК и РНК. Двойная спираль ДНК
Нуклеиновые кислоты — ДНК и РНК — являются важнейшими макромолекулами обеспечивающие непрерывность жизни. ДНК и РНК — несут генетический план строения и функционирования клетки.
Строение ДНК и РНК
Двумя основными типами нуклеиновых кислот являются дезоксирибонуклеиновая кислота (ДНК, DNA) и рибонуклеиновая кислота (РНК, RNA).
Мономер нуклеиновых кислот — нуклеотид
ДНК и РНК состоят из мономеров. Мономерами нуклеиновых кислот являются нуклеотиды. В результате синтеза ДНК или РНК нуклеотиды объединяются друг с другом с образованием полинуклеотида.
Строение нуклеотида
Каждый нуклеотид состоит из трех компонентов:
- азотистого основания
- пентозного (пятиуглеродного) сахара
- фосфатной группы
Азотистое основание присоединено к молекуле сахара, которая присоединена к одной или нескольким фосфатным группам.
Каждый нуклеотид в дезоксирибонуклеиновой кислоте (ДНК) содержит один из четырех возможных азотистых оснований:
- аденин (A),
- гуанин (G),
- цитозин (C),
- тимин (T).
Аденин и гуанин классифицируются как пурины. Первичная структура пуринового основания – два гетероциклических кольца содержащих углерод и азот.
Цитозин, тимин и урацил классифицируются как пиримидины, у которых в качестве первичной структуры одно гетероциклическое кольцо.
Сахар-пентоза в ДНК — дезоксирибоза, а в РНК — рибоза. Различием между сахарами в ДНК и РНК является присутствие гидроксильной группы на втором углероде рибозы или водорода на втором углероде дезоксирибозы.
Углеродные атомы молекулы сахара пронумерованы как 1′, 2′, 3 ‘, 4′ и 5’.
Фосфатный остаток присоединяют к гидроксильной группе 5′-углерода одного сахара и гидроксильной группе 3′-углерода сахара следующего нуклеотида, который образует 5′-3′-фосфодиэфирную связь.
Структура двойной спирали ДНК
ДНК имеет двухспиральную структуру.
Сахар и фосфат лежат снаружи спирали, образуя основу ДНК.
Азотистые основания уложены во внутреннюю часть.
Пары азотистых оснований связаны друг с другом водородными связями.
Каждая пара оснований в двойной спирали отделена от следующей пары оснований на 0,34 нм.
Две нити спирали движутся в противоположных направлениях — одна нить спирали имеет направление от 5′ к 3′, а другая – направление от 3′ к 5′.
Разрешены только определенные типы спариваний оснований. Например, определенный пурин может спариваться только с определенным пиримидином, т.е. аденин (A) может спариваться (комплементарен) с тимином (T), а гуанин (G) комплементарен цитозину (C).
РНК
Состав РНК
РНК обычно одноцепочечна и состоит из рибонуклеотидов, которые связаны фосфодиэфирными связями. Рибонуклеотид в цепи РНК содержит рибозу (пентозный сахар), один из четырех азотистых оснований (A, U (урацил), G и C) и фосфатную группу.
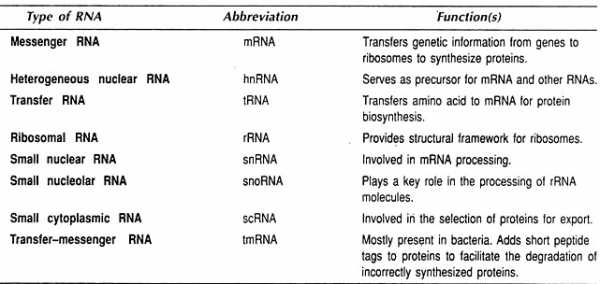
Виды и функции РНК
Четыре основных типа РНК — информационная РНК (мРНК), рибосомная РНК (рРНК), транспортная РНК (тРНК) и микро РНК (miRNA).
Статьи в категории
biology.reachingfordreams.com
Нуклеотиды. Состав. Строение.
Доклад
Студенток 1 курса 13 группы Института фзической культуры и спорта
Факультета физической культуры для лиц с отклонениями в состоянии здоровья (адаптивная физическая культура)
Размус Алены
Семеновой Екатерины
по теме: «Нуклеиновые кисллоты».
Содержание:
Нуклеиновые кислоты. Определение. Открытие. Виды нуклеиновых кислот.
Нуклеотид. Состав. Строение.
Правило Чааргафа
ДНК. Модель Уотсона и Крика. Структура. Состав. Функции.
РНК. Состав и ее разнообразие.
ДНК – носитель наследственной информации.
Краткие итоги.
Нуклеиновые кислоты.
Нуклеиновые кислоты (Нк) – биополимеры, обеспечивающие хранение и передачу наследственной (генетической) информации в живых организмах.
Впервые Нк были описаны в 1868 году швейцарским биохимиком Фридрихом Мишером (1844-1895).
Из
остатков клеток, содержащихся в гное,
он выделил вещество, в состав которого
входили N2
и P.
Ученый назвал это вещество нуклеином
(лат.
nucleus
– ядро), полагая, что оно содержится
лишь в ядрах клеток. Позднее небелковая
часть этого вещства была названа
нуклеиновой
кислотой.

Нуклеиновые кислоты в природе существут двух типов, различающиеся по составу, строению и функциям. Одна названа ДНК (дизоксирибонуклиновая кислота), а вторая РНК (рибонуклиновая кислота).
Нуклеиновые кислоты – это важнейшие биополимеры, определяющие основные свойства живого.
ДНК – это полимерная молекула, состоящая из десятков тысяч или миллионов мономеров – дезоксирибонуклеотидов.
Определение размеров молекул ДНК стало возможным только после разработки специальных методов: электронной микроскопии, ультрацентрифугирования, электрофореза. При полном гидролизе эти молекулы расщепляются до пуриновых и пеиримидиновых оснований, пятиугольного моносахарида дезоксирибозы и фосфорной кислоты.
Пуриновые основания – производные пурин. Из них в сосатв нуклеиновых кислот входят аденин и гуанин:
Пиримидиновые основания, содержащиеся в нуклеиновых кислотах, — цитозин и тимин в ДНК, цитозин и урацил в РНК – это производные пиримидина:
Тимин отличается от урацила наличием метильной группы (-СН3). Пуриновые и пиримидиновые основания называются азотистыми основаниями.
При мягком гидролизе нуклеиновых кислот получали соединения, дезоксирибоза которых была связана с пуриновым или пиримидиновым основанием посредством атома N2. Подобные соединения получили название нуклеозидов. Нуклеозиды,соединяясь с одной молекулой фосфорной кислоты, образуют более сложные вещества – нуклеотиды. Именно они являются мономерами нуклеиновых кислот ДНК и РНК.
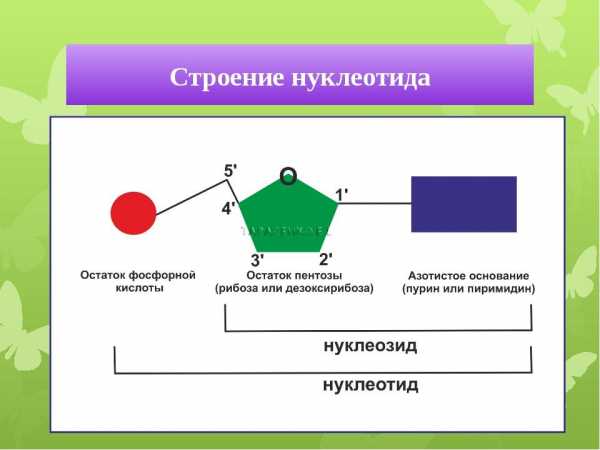
Итак, нуклеотид состояит из остатков азотистого основания, сахара – пентозы и фофорной кислоты.
Правило Эрвина Чааргафа.
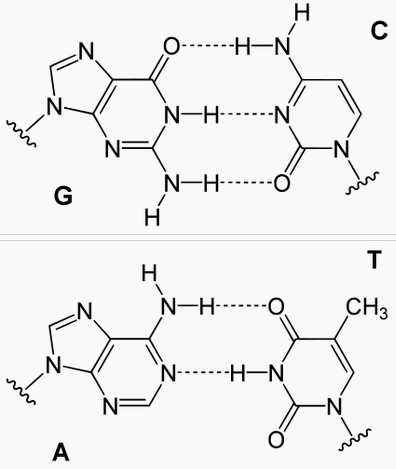
Нуклеотидный состав ДНК впервые количественно проанализировал американский биохимик Эрвин Чааргаф, который в 1951 году доказал, что в составе ДНК имеются четыре основания. Э. Чааргаф обнаружил, что у всех изученных им видов количество пуринового основания аденина (А) равно количеству пиримидинового основания тимина (Т), т.е. А=Т.
Сходным образом количество пуринового основания гуанина (Г) всегда равно количеству пиримидинового основания цитозина (Ц), т.е. Г=Ц. Таким образом, число пуриновых ДНК всегда равно числу пиримидиновых, т.е. количеству аденина равно количеству имина, а количество гуанина – количеству цитозина. Эта закономерность получило название правила Чааргафа.
Днк. Модель Уотсона и Крика. Структура. Состав. Функции.
В 1950 году английский физик Морис Хью Уилкинс получил рентгенограмму ДНК. Она показала, что молекула ДНК имеет определенную вторичную структуру, расшифровка которой помогла бы понять механизм функционирования ДНК. Рентегонграммы, полученные на высокоочищенной ДНК, позволили Розалинде Франклин, коллеге Уилкинса, увидеть четкий крестообразный рисунок – опознавательный знак двоной спирали. Стало извесно также, что нуклеотиды расположены друг от лруга на растоянии 0, 34 нм, а на один виток спирали их приходится 10. Диаметр молекулы ДНК составляет около 2 нм. Из рентгеноструктурных данных, однако, было неясно, каким образом цепи удерживаются вместе в молекулах ДНК.
Картна полностью прояснилась в 1953 году, когда американский биохимик Джеймс Уотсон и английский физик Фрэнсис Крик, рассмотрев совокупность известных данных о строении ДНК, пришли к выводу, что сахарофосфатный остов находится на периферии молекулы ДНК, а пуриновые и пиримидиновые основания – в середине. Причем последние ориентированы таким образом, что между основаниями противоположных цепей могут образовываться водородные связи. Из построенной ими модели выявилось, что пурин в одной цепи всегда связан водородными связями с противолежащим пиримидином в другой цепи.
Такие пары имеют одинаковый размер по всей длинне молекулы. Не менее важно то,что аденин может спариваться лишь с тимином, а гуанин только с цитозином. При этом между аденином и тимином образуются две водородные связи, а между гуанином и цитозином – три.
В
каждой из цепей ДНК основания могут
чередоваться всеми возможными способами.
Если известна последовательность
оснований в одной цепи (например, Т
– Ц – Г – Ц – А – Т),
то благодаря специфичности спаривания
(принцип
дополнения, т.е.
комплементарности)
становится извсетной и последовательность
оснований ее партнера – второй цепи (А
– Г – Ц – Г – Т – А).
Противолежащие последовательности и
соответствующие полинуклеотидные цепи
называют комплементарными.
Хотя водородные связи, стабилизирующие
пары оснований, относительно слабы,
каждая молекула ДНК содержит так много
пар, что в физиологических условиях
(темпиратура, pH)
комплименарные цепи никогда самостоятельно
не разделяются.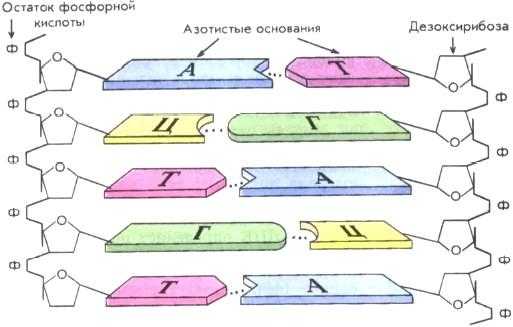
В начале 50-х годов большая группа ученых под руководством английского ученого А. Тодда установила точную структуру связей, соединяющих нуклеотиды одной цепи. Все эти связи оказались одинаковыми: углеродный атом в 5′-положении остатка дезоксирибозы (цифры со штрихами обозначают углеродные атомы в пятичленном сахаре – рибозе или дизоксирибозе) одного нуклеотида соединяется через фосфатную группу с углеродным атомом в 3’-положени соседнего нуклеотида. Никаких признаков необычных связей обнаружено не было. А. Тодд с сотрудниками пришли к выводу, что полинуклеотидные цепи ДНК, так же как и полипиптидные цепи белка, строго линейные. Регулярно расположенные связи между сахарами и фосфатными группами образуют скелет полинуклеотидной цепи.
Напротив
5′-конца одной цеп находится 3’-конец
комплементарной цепи. Такая ориентация
цепей названа антипараллельной.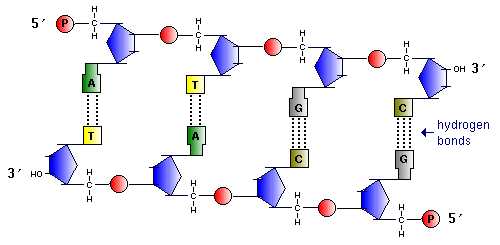
У всех живущих на Земле организмов ДНК представлена двухцепоными спиральными молекулами. Исключение составляют одноцепочные молекулы ДНК некоторых фагов – вирусов, поражающих бактериальные клетки. Эти одноцепочные ДНК всегда кольцевые. Двухцепочные ДНК бывают и кольцевые и линейные. Бактерии содержат только кольцевые формы ДНК. У растений, грибов и животных имеются и линейные (в ядре клетки), и кольцевые (в хлоропластахи митохондриях) молекулы.
Функции ДНК:
Хранение информации
Передача и воспроизведение в ряду поколений генетической информации
ДНК определяет, какие белки и в каких количествах необходимо синтезировать
studfiles.net
строение нуклеотидов днк и строение нуклеотидов рнк
ДНК — полимер, мономерами которой являются дезоксирибонуклеотиды. Модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. Дж. Уотсоном и Ф. Криком (для построения этой модели они использовали работы М. Уилкинса, Р. Франклин, Э. Чаргаффа) .Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т. е. представляет собой двойную спираль (исключение — некоторые ДНК-содержащие вирусы имеют одноцепочечную ДНК) . Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0,34 нм, на один оборот спирали приходится 10 пар нуклеотидов. Длина молекулы может достигать нескольких сантиметров. Молекулярный вес — десятки и сотни миллионов. Суммарная длина ДНК ядра клетки человека — около 2 м. В эукариотических клетках ДНК образует комплексы с белками и имеет специфическую пространственную конформацию.
Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов. Пиримидиновые основания ДНК (имеют в составе своей молекулы одно кольцо) — тимин, цитозин. Пуриновые основания (имеют два кольца) — аденин и гуанин.
Строение и функции РНК
РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК) . Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК.
Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов.
Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой.
Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК) , 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
otvet.mail.ru
Строение ДНК — Науколандия
По своему химическому строению ДНК (дезоксирибонуклеиновая кислота) является биополимером, мономерами которого являются нуклеотиды. То есть ДНК — это полинуклеотид. Причем молекула ДНК обычно состоит из двух цепей, закрученных друг относительно друга по винтовой линии (часто говорят «спирально закрученных») и соединенных между собой водородными связями.
Цепочки могут быть закручены как в левую, так и в правую (чаще всего) сторону.
У некоторых вирусов ДНК состоит из одной цепи.
Каждый нуклеотид ДНК состоит из 1) азотистого основания, 2) дезоксирибозы, 3) остатка фосфорной кислоты.
Двойная правозакрученная спираль ДНК
В состав ДНК входят следующие азотистые основания: аденин, гуанин, тимин и цитозин. Аденин и гуанин относятся к пуринам, а тимин и цитозин — к пиримидинам. Иногда в состав ДНК входит урацил, который обычно характерен для РНК, где замещает тимин.
Азотистые основания одной цепи молекулы ДНК соединяются с азотистыми основаниями другой строго по принципу комплементарности: аденин только с тимином (образуют между собой две водородные связи), а гуанин только с цитозином (три связи).
Азотистое основание в самом нуклеотиде соединено с первым атомом углерода циклической формы дезоксирибозы, которая является пентозой (углеводом с пятью атомами углерода). Связь является ковалентной, гликозидной (C-N). В отличие от рибозы у дезоксирибозы отсутствует одна из гидроксильных групп. Кольцо дезоксирибозы формируют четыре атома углерода и один атом кислорода. Пятый атом углерода находится вне кольца и соединен через атом кислорода с остатком фосфорной кислоты. Также через атом кислорода у третьего атома углерода присоединяется остаток фосфорной кислоты соседнего нуклеотида.
Таким образом, в одной цепи ДНК соседние нуклеотиды связаны между собой ковалентными связями между дезоксирибозой и фосфорной кислотой (фосфодиэфирная связь). Образуется фосфат-дезоксирибозный остов. Перпендикулярно ему, навстречу другой цепочке ДНК, направлены азотистые основания, которые соединяются с основаниями второй цепочки водородными связями.
Строение ДНК таково, что остовы соединенных водородными связями цепочек направлены в разные стороны (говорят «разнонаправлены», «антипараллельны»). С той стороны, где одна заканчивается фосфорной кислотой, соединенной с пятым атомом углерода дезоксирибозы, другая заканчивается «свободным» третьим атомом углерода. То есть остов одной цепочки перевернут как бы с ног на голову относительно другой. Таким образом, в строении цепочек ДНК различают 5′-концы и 3′-концы.
При репликации (удвоении) ДНК синтез новых цепочек всегда идет от их 5-го конца к третьему, так как новые нуклеотиды могут присоединяться только к свободному третьему концу.
В конечном итоге (опосредованно через РНК) каждые идущие подряд три нуклеотида в цепи ДНК кодируют одну аминокислоту белка.
Открытие строения молекулы ДНК произошло в 1953 году благодаря работам Ф. Крика и Д. Уотсона (чему также способствовали ранние работы других ученых). Хотя как химическое вещество ДНК было известно еще в XIX веке. В 40-х годах XX века стало ясно, что именно ДНК является носителем генетической информации.
Двойная спираль считается вторичной структурой молекулы ДНК. У клетках эукариот подавляющее количество ДНК находится в хромосомах, где связана с белками и другими веществами, а также подвергается более плотной упаковке.
scienceland.info
особенности, схема. Какое строение имеет молекула ДНК? :: SYL.ru
ДНК является универсальным источником и хранителем наследственной информации, которая записана с помощью специальной последовательности нуклеотидов, она определяет свойства всех живых организмов.

Средняя молекулярная масса нуклеотида принимается равной 345, а количество нуклеотидных остатков может достигать нескольких сот, тысяч и даже миллионов. ДНК в основной своей массе находится в ядрах клеток. Немного содержится в хлоропластах и митохондриях. Однако ДНК ядра клетки – это не одна молекула. Она состоит из множества молекул, которые распределены по разным хромосомам, их количество меняется в зависимости от организма. Это и есть особенности строения ДНК.
История открытия ДНК
Строение и функции ДНК были открыты Джеймсом Уотсоном и Френсисом Криком, им даже была вручена Нобелевская премия в 1962 году.

Но впервые обнаружил нуклеиновые кислоты швейцарский ученый Фридрих Иоганн Мишер, работавший в Германии. В 1869 году он изучал животные клетки – лейкоциты. Для их получения использовал повязки с гноем, достававшиеся ему из больниц. Из гноя Мишер вымывал лейкоциты, а из них выделял белок. В ходе этих исследований ученому удалось установить, что в лейкоцитах кроме белков имеется еще что-то, какое-то неизвестное на тот момент вещество. Оно представляло собой нитевидный или хлопьевидный осадок, который выделялся, если создать кислую среду. Осадок сразу растворялся при добавлении щелочи.
Ученый с помощью микроскопа обнаружил, что при отмывании лейкоцитов с помощью соляной кислоты от клеток остаются ядра. Тогда он сделал заключение, что в ядре есть неизвестное вещество, названное им нуклеином (слово nucleus в переводе означает ядро).
Проведя химический анализ, Мишер выяснил, что новое вещество в своем составе имеет углерод, водород, кислород и фосфор. В то время фосфорорганических соединений было известно немного, поэтому Фридрих решил, что обнаружил новый класс соединений, находящихся в ядре клетки.
Таким образом, в XIX веке было открыто существование нуклеиновых кислот. Однако в то время никто не мог даже подумать о том, какая важная роль им принадлежит.
Вещество наследственности
Строение ДНК продолжали исследовать, и в 1944 году группа бактериологов под руководством Освальда Эвери получила доказательства того, что эта молекула заслуживает серьезного внимания. Ученый на протяжении многих лет занимался изучением пневмококков, организмов, которые вызывали пневмонию или заболевание легких. Эвери проводил опыты, смешивая пневмококки, вызывающие заболевание, с теми, которые безопасны для живых организмов. Сначала болезнетворные клетки убивали, а после добавляли к ним те, которые заболеваний не вызывают.
Результаты исследований поразили всех. Были такие живые клетки, которые после взаимодействия с мертвыми научались вызывать болезнь. Ученый выяснил природу вещества, которое участвует в процессе передачи информации живым клеткам от мертвых. Молекула ДНК и оказалась этим веществом.

Строение
Итак, необходимо разобраться с тем, какое строение имеет молекула ДНК. Открытие ее структуры стало значимым событием, это привело к образованию молекулярной биологии – новой отрасли биохимии. ДНК в больших количествах находится в ядрах клеток, однако размеры и количество молекул зависят от вида организма. Установлено, что ядра клеток млекопитающих содержат много этих клеток, они распределены по хромосомам, их насчитывается 46.
Изучая строение ДНК, в 1924 году Фельген впервые установил ее локализацию. Доказательства, полученные в ходе экспериментов, показали, что ДНК находится в митохондриях (1-2%). В других местах эти молекулы могут находиться при вирусной инфекции, в базальных тельцах, а также в яйцеклетках некоторых животных. Известно, что чем сложнее организм, тем масса ДНК больше. Количество молекул, находящихся в клетке, зависит от функции и составляет обычно 1-10%. Меньше всего их находится в миоцитах (0,2%), больше – в половых клетках (60%).
Строение ДНК показало, что в хромосомах высших организмов они связаны с простыми белками – альбуминами, гистонами и прочими, которые все вместе образуют ДНП (дезоксирибонуклеопротеид). Обычно большая молекула нестойкая, и для того чтобы она оставалась целой и неизменной в ходе эволюции, создана так называемая репарирующая система, которая состоит из ферментов – лигаз и нуклеаз, отвечающих за «ремонт» молекулы.
Химическое строение ДНК
ДНК является полимером, полинуклеотидом, состоящим из огромного числа (до десятков тысяч миллионов) мононуклеотидов. Строение ДНК имеет следующий вид: мононуклеотиды содержат азотистые основания – цитозин (Ц) и тимин (Т) – из производных пиримидинов, аденин (А) и гуанин (Г) – из производных пурина. Кроме азотистых оснований, в составе молекулы человека и животных имеется 5-метилцитозин — минорное пиримидиновое основание. С фосфорной кислотой и дезоксирибозой связываются азотистые основания. Схема строения ДНК продемонстрирована ниже.
Правила Чаргаффа
Строение и биологическая роль ДНК изучались Э. Чаргаффом в 1949 году. В ходе исследований он выявил закономерности, которые наблюдаются в количественном распределении азотистых оснований:
- ∑Т + Ц = ∑А + Г (то есть число пиримидиновых оснований равно числу пуриновых).
- Всегда количество остатков аденина равно количеству остатков тимина, а количество гуанина равно цитозину.
- Коэффициент специфичности имеет формулу: Г+Ц/А+Т. Например, у человека он равен 1,5, у быка – 1,3.
- Сумма «А + Ц» равна сумме «Г + Т», то есть аденина и цитозина имеется столько же, сколько гуанина и тимина.
Модель строения ДНК
Ее создали Уотсон и Крик. Остатки фосфатов и дезоксирибоз располагаются по хребту двух закрученных спиралеобразным образом полинуклеотидных цепей. Определено, что плоскостные структуры пиримидиновых и пуриновых оснований располагаются перпендикулярно оси цепи и образуют как бы ступени лестницы в виде спирали. Установлено также, что А всегда соединяется с Т при помощи двух водородных связей, а Г прикреплено к Ц уже тремя такими же связями. Этому явлению дали название «принцип избирательности и комплементарности».
Уровни структурной организации
Изогнутая как спираль полинуклеотидная цепь – это первичная структура, которая имеет определенный качественный и количественный набор мононуклеотидов, связанных 3’,5’-фосфодиэфирной связью. Таким образом, каждая из цепей имеет 3’-конец (дезоксирибоза) и 5’-конец (фосфатный). Участки, которые содержат в себе генетическую информацию, названы структурными генами.
Двухспиральная молекула – это вторичная структура. Причем ее полинуклеотидные цепи антипараллельны и связываются водородными связями между комплементарными основаниями цепей. Установлено, что в каждом витке этой спирали содержится 10 нуклеотидных остатков, длина ее равняется 3,4 нм. Эту структуру поддерживают также Ван-дер-Ваальсовы силы взаимодействия, которые наблюдаются между основаниями одной цепи, включающие отталкивающие и притягивающие компоненты. Эти силы объясняются взаимодействием электронов в соседних атомах. Электростатическое взаимодействие также стабилизирует вторичную структуру. Оно возникает между заряженными положительно молекулами гистонов и заряженной отрицательно нитью ДНК.
Третичная структура – это намотка цепей ДНК на гистоны или суперспирализация. Описано пять видов гистонов: Н1, Н2А, Н2В, Н3, Н4.
Укладка нуклеосом в хроматин – это четвертичная структура, поэтому молекула ДНК, имеющая длину несколько сантиметров, может складываться до 5 нм.
Функции ДНК
Основными функциями ДНК являются:
- Хранение наследственной информации. Последовательность аминокислот, находящихся в молекуле белка, определяется порядком, в котором расположены нуклеотидные остатки в молекуле ДНК. Также в ней зашифрована вся информация о свойствах и признаках организма.
- ДНК способна передавать наследственную информацию следующему поколению. Это возможно из-за способности к репликации – самоудвоению. ДНК способна распадаться на две комплементарные цепочки, и на каждой из них (в соответствии с принципом комплементарности) восстанавливается исходная последовательность нуклеотидов.
- При помощи ДНК происходит биосинтез белков, ферментов и гормонов.
Заключение
Строение ДНК позволяет ей являться хранителем генетической информации, а также передавать ее следующим поколениям. Какие есть особенности у этой молекулы?
- Стабильность. Это возможно благодаря гликозидным, водородным и фосфодиэфирным связям, а также механизму репарации индуцированных и спонтанных повреждений.
- Возможность репликации. Этот механизм позволяет в соматических клетках сохранять диплоидное число хромосом.
- Существование генетического кода. При помощи процессов трансляции и транскрипции последовательность оснований, находящихся в ДНК, преобразуется в последовательность аминокислот, находящихся в полипептидной цепи.
- Способность к генетической рекомбинации. При этом образуются новые сочетания генов, которые сцеплены между собой.
Таким образом, строение и функции ДНК позволяют ей играть неоценимую роль в организмах живых существ. Известно, что длина 46-ти молекул ДНК, находящихся в каждой клетке человека, равна почти 2 м, а число нуклеотидных пар составляет 3,2 млрд.
www.syl.ru
