
Нуклеиновые кислоты.
Нуклеиновые кислоты – природные высокомолекулярные биополимеры, обеспечивающие хранение и передачу наследственной (генетической) информации в живых организмах.
Макромолекула нуклеиновых кислот, с молекулярной массой от 10000 Дальтон до нескольких миллионов, открыты в 1869 г. швейцарским химиком Ф. Мишером в ядрах лейкоцитов, входящих в состав гноя, отсюда и название (нуклеус – ядро).
Нуклеиновые кислоты представляют собой полимеры, мономерами которых являются нуклеотиды. Каждый нуклеотид состоит из азотистого основания, сахара пентозы и остатка фосфорной кислоты. Из нуклеотидов строятся длинные молекулы – полинуклеотиды.
Фосфат
Пентоза
Азотистое
основание
Связь между
фосфатом и сахаром
Рис. Строение нуклеотида.
Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т. е. представляет собой
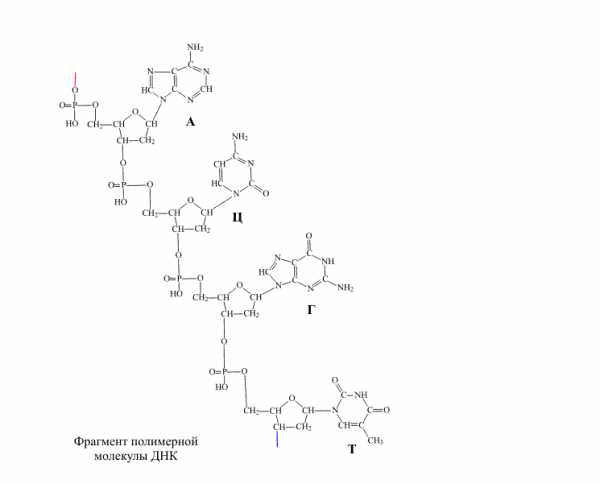
Основания, в обоих видах нуклеиновых кислот, содержатся четырех разных видов: два из них относятся к классу пуринов и два – к классу пиримидинов. К числу пуринов относятся аденин (А) и гуанин (Г), а к числу пиримидинов – цитизин (Ц) и тимин (Т) или урацил (У) (соответственно в ДНК или РНК).
Нуклеиновые кислоты являются кислотами потому, что в их молекуле содержится фосфорная кислота.
Роль нуклеотидов в организме не ограничивается тем, что они служат строительными блоками нуклеиновых кислот; некоторые важные коферменты также представляют совой нукоеотиды. Таковы, например, аденозинтрифосфат (АТФ), никотинамидадениндинуклеотид (НАД), никотинамидадениндинуклеотид-фосфат (НАДФ) и флавинадениндинуклеотид (ФАД).
Нуклеиновые кислоты
ДНКРНК
ядерная цитоплазматические иРНК тРНК рРНК
В настоящее время известно большое число разновидностей ДНК и РНК, отличных друг от друга по строению и значению в метаболизме.
Пример: в бактериях клеток кишечной палочки содержится около 1000 различных нуклеиновых кислот, а у животных и растений еще больше.
Каждый вид организмов содержит свой, характерный только для него, набор этих кислот. ДНК локализуется преимущественно в хромосомах клеточного ядра (99% всей ДНК клетки), а также в митохондриях и хлоропластах. РНК входит в состав ядрышек, рибосом митохондрий, пластид и цитоплазмы.
ДНК
Молекула ДНК является универсальным носителем генетической информации в клетках. Именно благодаря строению и функциям этой молекулы признаки передаются по наследству – от родителей потомкам, т.е. осуществляется всеобщее свойство живого – наследственность. Молекулы ДНК – самые крупные биополимеры.
Строение ДНК.
Структура молекул ДНК была расшифрована в 1953 г. Дж. Уотсоном и Ф. Криком. За это открытие они получили Нобелевскую премию.
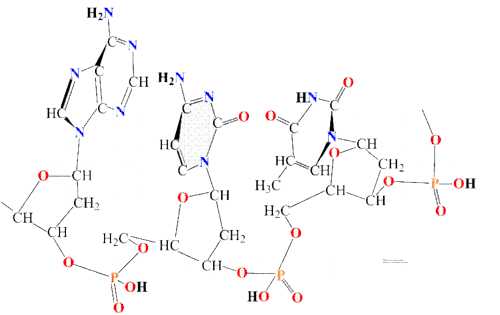
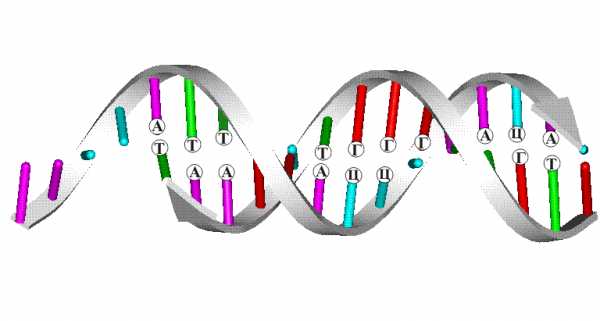
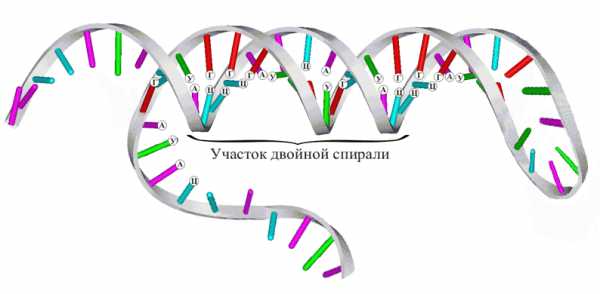
Согласно модели ДНК по Уотсону – Крику, молекула ДНК состоит из двух полинуклеотидных цепочек, закрученных вправо вокруг одной и той же оси, образуя двойную спираль. Цепи распложены антипараллельно, т.е. навстречу друг другу. Объединяются две полинуклеотидные цепи в единую молекулу ДНК при помощи водородных связей, возникающих между азотистым основанием нуклеотидов разных цепей. В полинуклеотидной цепочке соседние нуклеотиды связаны между собой ковалентными связями, которые образуются между дезоксирибозой, в молекуле ДНК (и рибозой в РНК), одного и остатком фосфорной кислоты другого нуклеотида.
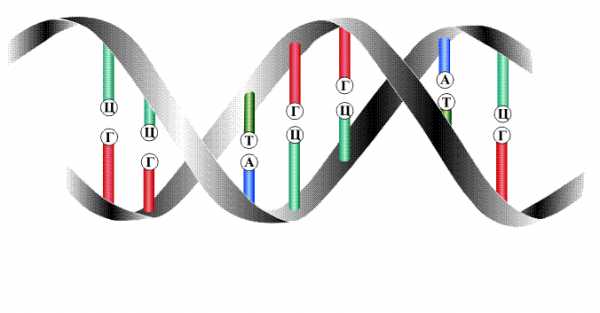
Цепи двойной спирали комплементарны друг другу, т. к. спаривание оснований происходит в строгом соответствии: аденин соединяется с тимином, а гуанин – с цитозином.
 В результате у всякого организма Рис. Спаривание нуклеотидов.
В результате у всякого организма Рис. Спаривание нуклеотидов.
число адениловых нуклеотидов равно числу тимидиловых, а число гуаниловых – числу
Строгое соответствие нуклеотидов, расположенных в парных антипараллельных нитях ДНК, называются комплементарностью. Это свойство лежит в основе образования новых молекул ДНК на базе исходной молекулы.
Таким образом, двойная спираль стабилизирована многочисленными водородными свойствами (между А и Т образуется две, а между Г и Ц – три) и гидрофобными взаимодействиями.
Вдоль оси молекулы соседние пары оснований располагаются на расстоянии 0,34 нм одна от другой. Полный оборот спирали приходится на 3,4 нм, т. е. на 10 пар оснований (один виток). Диаметр спирали – 2 нм. Расстояние между углеводными компонентами двух спаренных нуклеотидов 1,1 нм. Длина молекулы нуклеиновых кислот достигает сотен тысяч нанометров. Это значительно больше самой крупной макромолекулы белка, которая в развернутом виде достигает в длину не более 100-200 нм. Масса молекулы ДНК составляет 6*10 -12г.
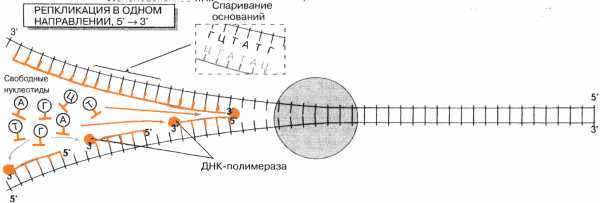 Процесс удвоения молекулы ДНК называется репликацией. Репликация происходит следующим образом. Под действием специальных ферментов (геликаза) разрываются водородные связи между нуклеотидами двух цепочек. Спираль раскручивается. К освободившимся связям, по принципу комплементарности, присоединяются соответствующие нуклеотиды ДНК, в присутствии фермента ДНК-полимеразы. Это наращивание может происходить только в направлении 5’→ 3′. Это означает непрерывного возможность копирования только одной цепи ДНК (на рисунке верхняя). Этот процесс называется непрерывнаярепликация. Копирование другой цепи должно всякий раз начинаться вновь, в результате в цепи возникают разрывы. Для их ликвидации необходим фермент – ДНК-лигаза. Такую репликацию называют прерывистой.
Процесс удвоения молекулы ДНК называется репликацией. Репликация происходит следующим образом. Под действием специальных ферментов (геликаза) разрываются водородные связи между нуклеотидами двух цепочек. Спираль раскручивается. К освободившимся связям, по принципу комплементарности, присоединяются соответствующие нуклеотиды ДНК, в присутствии фермента ДНК-полимеразы. Это наращивание может происходить только в направлении 5’→ 3′. Это означает непрерывного возможность копирования только одной цепи ДНК (на рисунке верхняя). Этот процесс называется непрерывнаярепликация. Копирование другой цепи должно всякий раз начинаться вновь, в результате в цепи возникают разрывы. Для их ликвидации необходим фермент – ДНК-лигаза. Такую репликацию называют прерывистой.
Данный способ репликации ДНК, предложенный Уотсоном и Криком известен под названием
 Следовательно, порядок нуклеотидов в «старой» цепочке ДНК определяет порядок нуклеотидов в «новой», т.е. «старая» цепочка ДНК как бы является матрицей для синтеза «новой». Такие реакции называются реакции матричного синтеза; они характерны только для живого.
Следовательно, порядок нуклеотидов в «старой» цепочке ДНК определяет порядок нуклеотидов в «новой», т.е. «старая» цепочка ДНК как бы является матрицей для синтеза «новой». Такие реакции называются реакции матричного синтеза; они характерны только для живого.
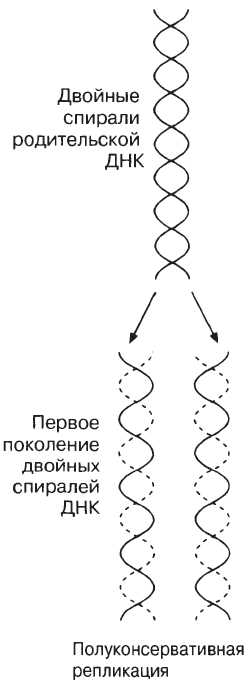
Репликация (редупликация) позволяет сохранить постоянство структуры ДНК. Синтезированная молекула ДНК абсолютно идентична исходной по последовательности нуклеотидов. Если под воздействием различных факторов в процессе репликации в молекуле ДНК происходят изменения в числе и порядке следования нуклеотидов, то возникают мутации. Способность молекул ДНК исправлять возникающие изменения и восстанавливать исходную называется репарацией.
Функции ДНК:
1) Хранение наследственной информации.
ДНК хранит информацию в виде последовательности нуклеотидов.
2) Воспроизведение и передача генетической информации.
Возможность передачи информации дочерним клеткам обеспечивается способностью хромосом к разделению на хроматиды с последующей редупликацией молекул ДНК. В ней закодирована генетическая информация о последовательности аминокислот в молекуле белка. Участок ДНК, несущий информацию об одной полипептидной цепи, называется геном.
3) Структурная.
ДНК присутствует в хромосомах в качестве структурного компонента, т.е. является химической основой хромосомного генетического материала (гена).
4) ДНК является матрицей для создания молекул РНК.
РНК.
РНК содержиться во всех живых клетках в виде одноцепочечных молекул. Она отличается от ДНК тем, что содержит в качестве пентозы рибозу (вместо дезоксирибозы), а в качестве одного из пиримидиновых оснований –
 В цепочке РНК нуклеотиды соединяются путем образования ковалентных связей (фосфодиэфирные связи) между рибозой одного нуклеотида и остатком фосфорной кислоты другого.
В цепочке РНК нуклеотиды соединяются путем образования ковалентных связей (фосфодиэфирные связи) между рибозой одного нуклеотида и остатком фосфорной кислоты другого.
В отличие от ДНК, молекулы РНК, представляют собой одноцепочечный линейный биополимер, состоящий из нуклеотидов.
Двухцепочечные РНК служат для хранения и воспроизведения наследственной информации у некоторых вирусов, т.е. выполняют у них функции хромосом – вирусная РНК.
Нуклеотиды одной молекулы РНК могут вступать в комплементарные взаимоотношения с другими нуклеотидами этой же цепочки, в результате образования вторичной и третичной структуры молекул РНК.
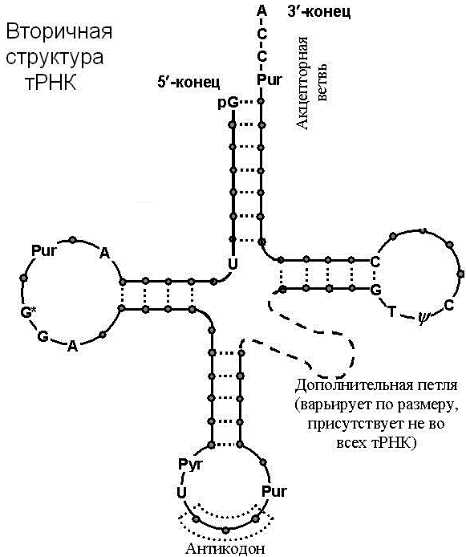
Рис. Строение транспортной РНК.
Рибисомальная РНК (рРНК) составляет 85% всей РНК клетки, она синтезируется в ядрышке, в соединение с белком входит в состав рибосом, митохондрий (митохондриальная РНК) и пластид (пластидная РНК). Содержит от 3 до 5 тыс. нуклеотидов. На рибосомах идет синтез белка.
Функции: рРНК выполняет структурную функцию (входит в состав рибосом) и участвует в формировании активного центра рибосом, где происходит образование пептидных связей между молекулами аминокислот в процессе биосинтеза белка.
Информационная РНК (иРНК) составляет 5% всей РНК в клетках. Она синтезируется в процессе транскрипции на определенном участке молекулы ДНК – гене. По строению иРНК комплементарна участку молекул ДНК, несущему информацию о синтезе определенного белка. Длина иРНК зависит от длины участка ДНК, с которого считывалась информация (может состоять из 300-30000 нуклеотидов)
Функции: иРНК переносит информацию о синтезе белка из ядра в цитоплазму на рибосомы и становится матрицей для синтеза молекул белка.
Транспортная РНК (тРНК) составляет около 10% всей РНК, синтезируется в ядрышке, имеет короткую цепь нуклеотидов и находится в цитоплазме. Она имеет функцию трилистника. У каждой аминокислоты имеется собственная семья молекул тРНК. Они доставляют содержащиеся в цитоплазме аминокислоты к рибосоме.
Функции: на одном конце находится триплет нуклеотидов (антикодон), кодирующий определенную аминокислоту. На другом конце триплет нуклеотидов, к которому присоединяется аминокислота. Для каждой аминокислоты – своя тРНК.
Похожие статьи:
poznayka.org
Нуклеиновые кислоты, их строение, свойства и функции
Подобно белкам, нуклеиновые кислоты — биополимеры, а их функция заключается в хранении, реализации и передаче генетической (наследственной) информации в живых организмах.
Существует два типа нуклеиновых кислот — дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Мономерами в нуклеиновых кислотах служат нуклеотиды. Каждый из них содержит азотистое основание, пятиуглеродный сахар (дезоксирибоза — в ДНК, рибоза — в РНК) и остаток фосфорной кислоты.
В ДНК входят четыре вида нуклеотидов, отличающихся по азотистому основанию в их составе, — аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т). В молекуле РНК также имеется 4 вида нуклеотидов с одним из азотистых оснований — аденином, гуанином, цитозином и урацилом (У). Таким образом, ДНК и РНК различаются как по содержанию сахара в нуклеотидах, так и по одному из азотистых оснований (табл. 1).
Таблица 1
Компоненты нуклеотидов ДНК и РНК
|
Нуклеиновая кислота |
Пятиуглеродный сахар |
Азотистые основания |
Остаток фосфорной кислоты |
||
|
ДНК |
Дезоксирибоза |
Аденин, гуанин, цитозин, тимин |
Остаток фосфорной кислоты |
||
|
РНК |
Рибоза |
Аденин, гуанин, цитозин, урацил |
Остаток фосфорной кислоты |
||
Молекулы ДНК и РНК существенно различаются по своему строению и выполняемым функциям.
Молекула ДНК может включать огромное количество нуклеотидов — от нескольких тысяч до сотен миллионов (поистине гигантские молекулы ДНК удается «увидеть» с помощью электронного микроскопа). В структурном отношении она представляет собой двойную спираль из полинуклеотидных цепей (рис. 1), соединенных с помощью водородных связей между азотистыми основаниями нуклеотидов. Благодаря этому полинуклеотидные цепи прочно удерживаются одна возле другой.
При исследовании различных ДНК (у разных видов организмов) было установлено, что аденин одной цепи может связываться лишь с тимином, а гуанин — только с цитозином другой. Следовательно, порядок расположения нуклеотидов в одной цепи строго соответствует порядку их расположения в другой. Этот феномен получил название комплементарности (т. е. дополнения), а противоположные полинуклеотидные цепи называются комплементарными. Именно этим обусловлено уникальное среди всех неорганических и органических веществ свойство ДНК — способность к самовоспроизведению или удвоению (рис. 2). При этом сначала комплементарные цепи молекул ДНК расходятся (под воздействием специального фермента происходит разрушение связей между комплементарными нуклеотидами двух цепей). Затем на каждой цепи начинается синтез новой («недостающей») комплементарной ей цепи за счет свободных нуклеотидов, всегда имеющихся в большом количестве в клетке. В результате вместо одной («материнской») молекулы ДНК образуются две («дочерние») новые, идентичные по структуре и составу друг другу, а также исходной молекуле ДНК. Этот процесс всегда предшествует клеточному делению и обеспечивает передачу наследственной информации от материнской клетки дочерним и всем последующим поколениям.
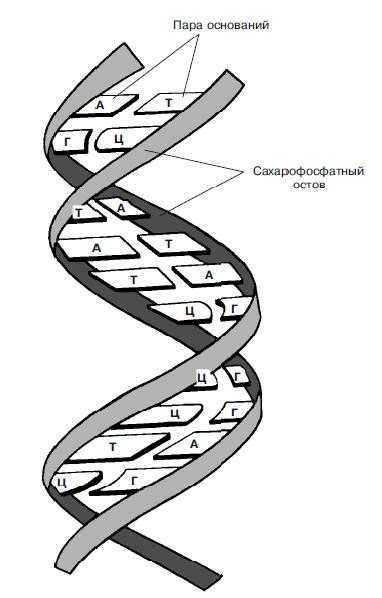
Рис. 1. Двойная спираль ДНК. Две цепи обвиты одна вокруг другой. Каждая цепь (изображенная в виде ленты) состоит из чередующихся остатков сахара и фосфатных групп. Водородные связи между азотистыми основаниями (А, Т, Г и Ц) удерживают две цепи вместе
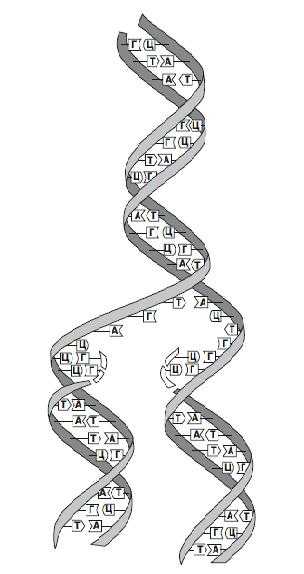
Рис. 2. Репликация ДНК. Двойная спираль «расстегивается» по слабым водородным связям, соединяющим комплементарные основания двух цепей. Каждая из старых цепей служит матрицей для образования новой: нуклеотиды с комплементарными основаниями выстраиваются против старой цепи и соединяются друг с другом
Молекулы РНК, как правило, одноцепочечные (в отличие от ДНК) и содержат значительно меньшее число нуклеотидов. Выделяют три вида РНК (табл. 2), различающиеся по величине молекул и выполняемым функциям, — информационную (иРНК), рибосомальную (рРНК) и транспортную (тРНК).
Таблица 2
Три вида РНК
|
РНК |
Число нуклеотидов в молекуле |
|
Информационные |
До 30 000 |
|
Рибосомальные |
До 6000 |
|
Транспортные |
Около 100 |
Информационная РНК (и-РНК) располагается в ядре и цитоплазме клетки, имеет самую длинную полинуклеотидную цепь среди РНК и выполняет функцию переноса наследственной информации из ядра в цитоплазму клетки.
Транспортная РНК (т-РНК) также содержится в ядре и цитоплазме клет-ки, ее цепь имеет наиболее сложную структуру, а также является самой короткой (75 нуклеотидов). Т-РНК доставляет аминокислоты к рибосомам в процессе трансляции — биосинтеза белка.
Рибосомальная РНК (р-РНК) содержится в ядрышке и рибосомах клетки, имеет цепь средней длины. Все виды РНК образуются в процессе транскрипции соответствующих генов ДНК.
Источник: Краснодембский Е. Г.»Общая биология: Пособие для старшеклассников и поступающих в вузы»
Н. С. Курбатова, Е. А. Козлова «Конспект лекций по общей биологии»
xn--90aeobapscbe.xn--p1ai
Нуклеиновая кислота Википедия
Нуклеи́новая кислота (от лат. nucleus — ядро) — высокомолекулярное органическое соединение, биополимер (полинуклеотид), образованный остатками нуклеотидов. Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации.
История исследования
- В 1847 из экстракта мышц быка было выделено[1] вещество, которое получило название «инозиновая кислота». Это соединение стало первым изученным нуклеотидом. В течение последующих десятилетий были установлены детали его химического строения. В частности, было показано, что инозиновая кислота является рибозид-5′-фосфатом, и содержит N-гликозидную связь.
- В 1868 году швейцарским химиком Фридрихом Мишером при изучении некоторых биологических субстанций было открыто неизвестное ранее вещество. Вещество содержало фосфор и не разлагалось под действием протеолитических ферментов. Также оно обладало выраженными кислотными свойствами. Вещество было названо «нуклеином». Соединению была приписана брутто-формула C29H49N9O22P3.
- Уилсон обратил внимание на практическую идентичность химического состава «нуклеина» и открытого незадолго до этого «хроматина» — главного компонента хромосом[2]. Было выдвинуто предположение об особой роли «нуклеина» в передаче наследственной информации.
- В 1889 г Рихард Альтман ввел термин «нуклеиновая кислота», а также разработал удобный способ получения нуклеиновых кислот, не содержащих белковых примесей.
- Левин и Жакоб, изучая продукты щелочного гидролиза нуклеиновых кислот, выделили их основные составляющие — нуклеотиды и нуклеозиды, а также предложили адекватные структурные формулы, описывающие их свойства.
- В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры ДНК»[3], оказавшуюся впоследствии ошибочной[4].
- В 1935 году Клейн и Танхаузер с помощью фермента фосфатазы провели мягкое фрагментирование ДНК, в результате чего были получены в кристаллическом состоянии четыре ДНК-образующих нуклеотида[5]. Это открыло новые возможности для установления структуры этих соединений.
- В 1940-е годы научная группа в Кембридже под руководством Александера Тодда проводит широкие синтетические исследования в области химии нуклеотидов и нуклеозидов. В результате их работы были установлены все детали химического строения и стереохимии нуклеотидов. За цикл работ в этой области Александер Тодд был награждён Нобелевской премией в области химии в 1957 году.
- В 1951 году Чаргаффом была установлена закономерность содержания в нуклеиновых кислотах нуклеотидов разных типов, получившая впоследствии название Правило Чаргаффа.
- В 1953 году Уотсоном и Криком установлена вторичная структура ДНК, двойная спираль[6].
Способы выделения
Гелеобразный осадок нуклеиновой кислотыОписаны многочисленные методики выделения нуклеиновых кислот из природных источников. Основными требованиями, предъявляемыми к методу выделения, являются эффективное отделение нуклеиновых кислот от белков, а также минимальная степень фрагментации полученных препаратов. Классический метод выделения ДНК был описан в 1952 году и используется в настоящее время без значительных изменений[7]. Клеточные стенки исследуемого биологического материала разрушаются одним из стандартных методов, а затем обрабатываются анионным детергентом. При этом белки выпадают в осадок, а нуклеиновые кислоты остаются в водном растворе. ДНК может быть осаждена в виде геля осторожным добавлением этанола к её солевому раствору. Концентрацию полученной нуклеиновой кислоты, а также наличие примесей (белки, фенол) обычно определяют спектрофотометрически по поглощению на А260 нм.
Нуклеиновые кислоты легко деградируют под действием особого класса ферментов — нуклеаз. В связи с этим при их выделении важно обработать лабораторное оборудование и материалы соответствующими ингибиторами. Так, например, при выделении РНК широко используется такой ингибитор рибонуклеаз как DEPC.
Физические свойства
Нуклеиновые кислоты хорошо растворимы в воде, практически нерастворимы в органических растворителях. Очень чувствительны к действию температуры и критическим значениям уровня pH. Молекулы ДНК с высокой молекулярной массой, выделенные из природных источников, способны фрагментироваться под действием механических сил, например, при перемешивании раствора. Нуклеиновые кислоты фрагментируются ферментами — нуклеазами.
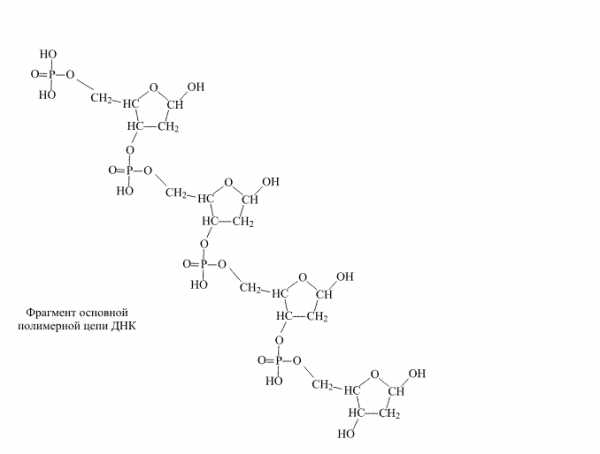
Строение
Фрагмент полимерной цепочки ДНКПолимерные формы нуклеиновых кислот называют полинуклеотидами.
Существуют 4 уровня структурной организации нуклеиновых кислот: первичная, вторичная, третичная и четвертичная. Первичная структура представляет собой цепочки из нуклеотидов, соединяющихся через остаток фосфорной кислоты (фосфодиэфирная связь). Вторичная структура — это две цепи нуклеиновых кислот соединённые водородными связями. Стоит отметить, что цепи соединяются по типу «голова-хвост» (3′ к 5′), по принципу комплементарности (азотистые основания находятся внутри этой структуры). Третичная структура, или же спираль, образуется за счет радикалов азотистых оснований (образуются водородные дополнительные связи, которые и сворачивают эту структуру, тем самым обуславливая её прочность). И наконец 4 структура — это комплексы гистонов и нитей хроматина.
Поскольку в нуклеотидах существует только два типа гетероциклических молекул, рибоза и дезоксирибоза, то и имеется лишь два вида нуклеиновых кислот — дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК).
Мономерные формы также встречаются в клетках и играют важную роль в процессах передачи сигналов или запасании энергии. Наиболее известный мономер РНК — АТФ, аденозинтрифосфорная кислота, важнейший аккумулятор энергии в клетке.
ДНК и РНК
Типы РНК
Ма́тричная рибонуклеи́новая кислота́ (мРНК, синоним — информацио́нная РНК, иРНК) — РНК, содержащая информацию о первичной структуре (аминокислотной последовательности) белков[8]. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.
Рибосо́мные рибонуклеи́новые кисло́ты (рРНК) — несколько молекул РНК, составляющих основу рибосомы. Основной функцией рРНК является осуществление процесса трансляции — считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами.
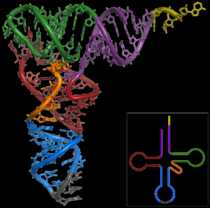 Структура транспортной РНК
Структура транспортной РНК
Транспортная РНК, тРНК — рибонуклеиновая кислота, функцией которой является транспортировка аминокислот к месту синтеза белка. Имеет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. тРНК также принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь — будучи в комплексе с аминокислотой — к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса.
Для каждой аминокислоты существует своя тРНК.
тРНК является одноцепочечной РНК, однако в функциональной форме имеет конформацию «клеверного листа». Аминокислота ковалентно присоединяется к 3′-концу молекулы с помощью специфичного для каждого типа тРНК фермента аминоацил-тРНК-синтетазы. На участке C находится антикодон, соответствующий аминокислоте.
Некодирующие РНК (non-coding RNA, ncRNA) — это молекулы РНК, которые не транслируются в белки. Ранее использовавшийся синоним, малые РНК (smRNA, small RNA), в настоящее время не используется, так как некоторые некодирующие РНК могут быть очень большими, например, Xist.
Последовательность ДНК, на которой транскрибируются некодирующие РНК, часто называют РНК-геном.
Некодирующие РНК включают в себя молекулы РНК, которые выполняют очень важные функции в клетке — транспортные РНК (тРНК), рибосомные РНК (рРНК), такие малые РНК, как малые ядрышковые РНК (snoRNA), микроРНК, siRNA, piRNA, а также длинные некодирующие РНК — Xist, Evf, Air, CTN, PINK, TUG1.
Последние транскриптомные технологии (секвенирование РНК) и методы ДНК-микрочипов предполагают наличие более 30000 длинных некодирующих РНК (англ. long ncRNA). Примерно такое же количество малых регуляторных РНК содержится в геноме мыши.
Примечания
- ↑ J. Liebig (1847). «???». Annalen 62: 257.
- ↑ Edmund B. Wilson. An Atlas of the Fertilization and Karyokinesis of the Ovum. — N. Y.: Macmillan, 1895. — P. 4.
- ↑ P. A. Levene (1921). «???». J. Biol. Chem. 48: 119.
- ↑ Во время выдвижения «тетрануклеотидной структуры» химики критически относились к самой возможности существования макромолекул, вследствие чего ДНК была приписана структура с низкой молекулярной массой
- ↑ W. Klein, S. J. Thannhauser (1935). «???». Z. physiol. Chem. 231: 96.
- ↑ J. D. Watson, F. H. C. Crick (1953). «Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid». Nature 171: 737—738. DOI:10.1038/171737a0.
- ↑ Ernest R. M. Kay, Norman S. Simmons, Alexander L. (1952). «An Improved Preparation of Sodium Desoxyribonucleate». J. Am. Chem. Soc. 74 (7): 1724–1726. DOI:10.1021/ja01127a034.
- ↑ Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. Molecular Biology of the Cell. — 5. — Garland Science, 2008. — 1392 с. — ISBN 0815341059.
Литература
- Бартон Д., Оллис У. Д. Общая органическая химия. — М.: Химия, 1986. — Т. 10. — С. 32—215. — 704 с.
- Франк-Каменецкий М. Д. Самая главная молекула. — М.: Наука, 1983. — 160 с.
- Аппель Б., Бенеке И., Бенсон Я., под ред. С. Мюллер. Нуклеиновые кислоты от А до Я. — М.: Бином, 2012. — 352 с. — ISBN 978-5-9963-0376-2.
wikiredia.ru
Нуклеиновые кислоты — это… Что такое Нуклеиновые кислоты?
Нуклеи́новые кисло́ты (от лат. nucleus — ядро) — высокомолекулярные органические соединения, биополимеры (полинуклеотиды), образованные остатками нуклеотидов. Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации.
История исследования
- В 1847 из экстракта мышц быка было выделено[1] вещество, которое получило название «инозиновая кислота». Это соединение стало первым изученным нуклеотидом. В течение последующих десятилетий были установлены детали его химического строения. В частности, было показано, что инозиновая кислота является рибозид-5′-фосфатом, и содержит N-гликозидную связь.
- В 1868 году швейцарским химиком Фридрихом Мишером при изучении некоторых биологических субстанций было открыто неизвестное ранее вещество. Вещество содержало фосфор и не разлагалось под действием протеолитических ферментов. Также оно обладало сильновыраженными кислотными свойствами. Вещество было названо «нуклеином». Соединению была приписана брутто-формула C29H49N9O22P3.
- Уилсон обратил внимание на практическую идентичность химического состава «нуклеина» и открытого незадолго до этого «хроматина» — главного компонента хромосом[2]. Было выдвинуто предположение об особой роли «нуклеина» в передаче наследственной информации.
- В 1889 г Рихард Альтман ввел термин «нуклеиновая кислота», а также разработал удобный способ получения нуклеиновых кислот, не содержащих белковых примесей.
- Левин и Жакоб, изучая продукты щелочного гидролиза нуклеиновых кислот, выделили их основные составляющие — нуклеотиды и нуклеозиды, а также предложили адекватные структурные формулы, описывающие их свойства.
- В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры ДНК» [3], оказавшуюся впоследствии ошибочной[4].
- В 1935 году Клейн и Танхаузер с помощью фермента фосфатазы провели мягкое фрагментирование ДНК, в результате чего были получены в кристаллическом состоянии четыре ДНК-образующих нуклеотида[5]. Это открыло новые возможности для установления структуры этих соединений.
- В 1940-е годы научная группа в Кембридже под руководством Александера Тодда проводит широкие синтетические исследования в области химии нуклеотидов и нуклеозидов. В результате их работы были установлены все детали химического строения и стереохимии нуклеотидов. За цикл работ в этой области Александер Тодд был награждён Нобелевской премией в области химии в 1957 году.
- Чаргаффом было установлена закономерность содержания в нуклеиновых кислотах нуклеотидов разных типов, получившая впоследствии название Правило Чаргаффа.
- В 1953 году Уотсоном и Криком установлена вторичная структура ДНК, двойная спираль[6].
Способы выделения
Гелеобразный осадок нуклеиновой кислотыОписаны многочисленные методики выделения нуклеиновых кислот из природных источников. Основными требованиями, предъявляемыми к методу выделения, являются эффективное отделения нуклеиновых кислот от белков, а также минимальная степень фрагментации полученных препаратов. Типичная методика приводится в работе[7]. Клеточные стенки исследуемого биологического материала разрушаются одним из стандартных методов, а затем обрабатываются анионным детергентом. При этом белки выпадают в осадок, а нуклеиновые кислоты остаются в водном растворе. ДНК может быть осаждена в виде геля осторожным добавлением этанола к её солевому раствору.
Нуклеиновые кислоты легко деградируют под действием особого класса ферментов — нуклеаз. В связи с этим при их выделении важно обработать лабораторное оборудование и материалы соответствующими ингибиторами. Так, например, при выделении РНК широко используется такой ингибитор рибонуклеаз как DEPC.
Химические свойства
Нуклеиновые кислоты хорошо растворимы в воде, практически не растворимы в органических растворителях. Очень чувствительны к действию температуры и критических значений уровня pH. Молекулы ДНК с высокой молекулярной массой, выделенные из природных источников, способны фрагментироваться под действием механических сил, например при перемешивании раствора. Нуклеиновые кислоты фрагментируются ферментами — нуклеазами.
Строение
Фрагмент полимерной цепочки ДНКПолимерные формы нуклеиновых кислот называют полинуклеотидами. Цепочки из нуклеотидов соединяются через остаток фосфорной кислоты (фосфодиэфирная связь). Поскольку в нуклеотидах существует только два типа гетероциклических молекул, рибоза и дезоксирибоза, то и имеется лишь два вида нуклеиновых кислот — дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК).
Мономерные формы также встречаются в клетках и играют важную роль в процессах передачи сигналов или запасании энергии. Наиболее известный мономер РНК — АТФ, аденозинтрифосфорная кислота, важнейший аккумулятор энергии в клетке.
ДНК и РНК
Примечания
- ↑ J.Liebig (1847). «». Annalen 62: 257.
- ↑ Edmund B. Wilson An Atlas of the Fertilization and Karyokinesis of the Ovum — New York: Macmillan, 1895. — P. 4.
- ↑ P.A.Levene (1921). «». J.Biol.Chem. 48: 119.
- ↑ Во время выдвижения «тетрануклеотидной структуры» химики критически относились к самой возможности существования макромолекул, вследствие чего ДНК была приписана структура с низкой молекулярной массой
- ↑ W.Klein, S.J.Thannhauser (1935). «». Z. physiol. Chem. 231: 96.
- ↑ J. D. Watson, F. H. C. Crick (1953). «Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid». Nature 171: 737 — 738. DOI:doi:10.1038/171737a0.
- ↑ Ernest R. M. Kay, Norman S. Simmons, Alexander L. (1952). «An Improved Preparation of Sodium Desoxyribonucleate». J. Am. Chem. Soc. 74 (7): 1724–1726. DOI:10.1021/ja01127a034.
Литература
- Бартон Д., Оллис У.Д. Общая органическая химия — Москва: Химия, 1986. — Т. 10. — С. 32—215. — 704 с.
- Франк-Каменецкий М.Д. Самая главная молекула — Москва: Наука, 1983. — 160 с.
См. также
| Типы нуклеиновых кислот | |
|---|---|
| Азотистые основания | Пурины (Аденин, Гуанин) | Пиримидины (Урацил, Тимин, Цитозин) |
| Нуклеозиды | Аденозин | Гуанозин | Уридин | Тимидин | Цитидин |
| Нуклеотиды | монофосфаты (АМФ, ГМФ, UMP, ЦМФ) | дифосфаты (АДФ, ГДФ, УДФ, ЦДФ) | трифосфаты (АТФ, ГТФ, УТФ, ЦТФ) | циклические (цАМФ, цГМФ, cADPR) |
| Рибонуклеиновые кислоты | РНК | мРНК | тРНК | рРНК | антисмысловые РНК | gRNA | микроРНК | некодирующие РНК | piwi-interacting RNA | shRNA | малые интерферирующие РНК | малые ядерные РНК | малые ядрышковые РНК | тмРНК |
| Дезоксирибонуклеиновые кислоты | ДНК | кДНК | Геном | msDNA | Митохондриальная ДНК |
| Аналоги нуклеиновых кислот | en:glycerol nucleic acid | en:locked nucleic acid | ПНК | ТНК | Морфолино |
| Типы векторов | en:phagemid | Плазмиды | Фаг лямбда | en:cosmid | en:P1 phage | en:fosmid | en:Bacterial artificial chromosome | en:Yeast artificial chromosome | en:Human artificial chromosome |
3dic.academic.ru
Нуклеиновые кислоты – ДНК и РНК
Биологическая роль нуклеиновых кислот. Название «нуклеиновые кислоты» происходит от латинского слова «нуклеус», т. е. ядро: они впервые были обнаружены в клеточных ядрах. Биологическое значение нуклеиновых кислот очень велико. Они играют центральную роль в хранении и передаче наследственных свойств клетки, поэтому их часто называют веществами наследственности. Известно, что любая клетка возникает в результате деления материнской клетки. При этом дочерние клетки наследуют свойства материнской. Свойства же клетки определяются главным образом ее белками. Нуклеиновые кислоты обеспечивают в клетке синтез белков, точно таких же, как в материнской клетке.
Существуют два вида нуклеиновых кислот дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК).
Дезоксирибонуклеиновая кислота (ДНК). Роль хранителя наследственной информации у всех клеток – животных и растительных – принадлежит ДНК. Схема строения ДНК изображена на рисунке [74]. Молекула ДНК представляет собой две спирально закрученные одна вокруг другой нити. Ширина такой двойной спирали ДНК невелика, около 2 нм. Длина же ее в десятки тысяч раз больше – она достигает сотен тысяч нанометров. Между тем самые крупные белковые молекулы в развернутом виде достигают в длину не более 100-200 нм. Таким образом, вдоль молекулы ДНК могут быть уложены одна за другой тысячи белковых молекул. Молекулярная масса ДНК соответственно исключительно велика – она достигает десятков и даже сотен миллионов.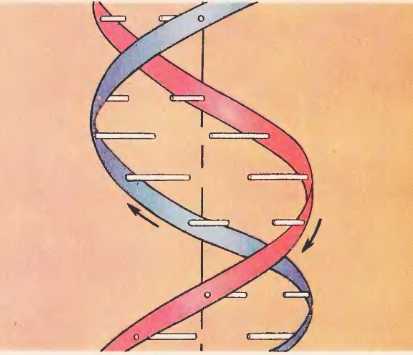 Рис. 74. Схема строения ДНК (двойная спираль)
Рис. 74. Схема строения ДНК (двойная спираль)
Обратимся к структуре ДНК. Каждая нить ДНК представляет собой полимер, мономерами которого являются нуклеотиды. Нуклеотид это химическое соединение остатков трех веществ: азотистого основания, углевода (моносахарида – дезоксирибозы) и фосфорной кислоты. ДНК всего органического мира образованы соединением четырех видов нуклеотидов. Их структуры приведены на рисунке [75]. Как видно, у всех четырех нуклеозидов углевод и фосфорная кислота одинаковы.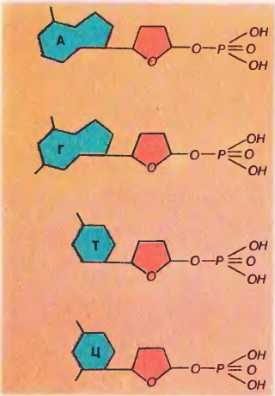 Рис. 75. Четыре нуклеотида, из которых построены все ДНК живой природы
Рис. 75. Четыре нуклеотида, из которых построены все ДНК живой природы
Нуклеотиды отличаются только по азотистым основаниям, в соответствии с которыми их называют: нуклеотид с азотистым основанием аденин (сокращенно А), нуклеотид с гуанином (Г), нуклеотид с тимином (Т) и нуклеотид с цитозином (Ц). По размерам А ранен Г, а Т равен Ц; размеры А и Г несколько больше, чем Т и Ц.
Соединение нуклеотидов в нити ДНК происходит через углевод одного нуклеотида и фосфорную кислоту соседнего. Они соединяются прочной ковалентной связью [76].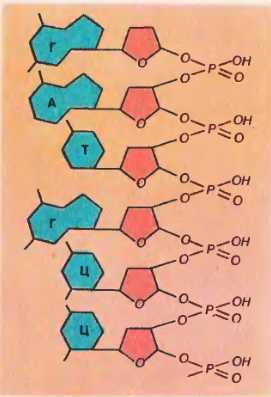 Рис. 76. Соединение нуклеотидов в полинуклеотидную цепь
Рис. 76. Соединение нуклеотидов в полинуклеотидную цепь
Итак, каждая нить ДНК представляет собой полинуклеотид. Это длинная цепь, в которой в строго определенном порядке расположены нуклеотиды.
Рассмотрим теперь, как располагаются относительно друг друга нити ДНК, когда образуется двойная спираль, и какие силы сдерживают их рядом. Представление об этом дает рисунок 77, на котором изображен небольшой участок двойной спирали.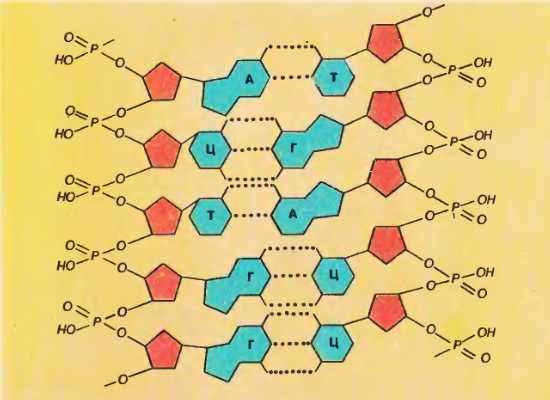 Рис. 77. Участок двойной спирали ДНК
Рис. 77. Участок двойной спирали ДНК
Как видно, азотистые основания одной цепи «стыкуются» с азотистыми основаниями другой. Основания подходят друг к другу настолько близко, что между ними возникают водородные связи.
В расположении стыкующихся нуклеотидов имеется важная закономерность, а именно: против А одной цепи всегда оказывается Т на другой цепи, а против Г одной цепи – всегда Ц. Оказывается, что только при таком сочетании нуклеотидов обеспечивается, во-первых, одинаковое но всей длине двойной спирали расстояние между цепями и, во-вторых, образование между противолежащими основаниями максимального числа водородных связей (три водородные связи между Г и Ц и две водородные связи между А и Т) . В каждом из этих сочетаний оба нуклеотида как бы дополняют друг друга. Слово «дополнение» на латинском языке «комплемент». Принято поэтому говорить, что Г является комплементарным Ц, а Т комплементарен А. Если на каком-нибудь участке одной цени ДНК один за другим следуют нуклеотиды А, Г, Ц, Т, А, Ц. Ц, то на противолежащем участке другой цепи окажутся комплементарные им Т, Ц, Г, А, Т, Г, Г. Таким образом, если известен порядок следования нуклеотидов в одной цени, то по принципу комплементарности сразу же выясняется порядок нуклеотидов в другой цени.
Большое число водородных связей обеспечивает прочное соединение нитей ДНК, что придает молекуле устойчивость и в то же время сохраняет ее подвижность: под влиянием фермента дезоксирибонуклеазы она легко раскручивается.
ДНК содержится в ядре клетки, а также в митохондриях и хлоропластах. В ядре ДНК входит в состав хромосом, где она находится в соединении с белками.
Удвоение ДНК. Принцип комплементарности, лежащий а основе структуры ДНК, позволяет понять, как синтезируются новые молекулы ДНК незадолго перед делением клетки. Этот синтез обусловлен замечательной способностью молекулы ДНК к удвоению и определяет передачу наследственных свойств от материнской клетки к дочерним.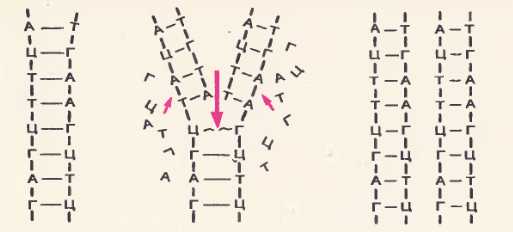 Рис. 78. Удвоение ДНК
Рис. 78. Удвоение ДНК
Как происходит удвоение ДНК, показано на рисунке 78. Двойная спираль ДНК под влиянием фермента начинает с одного конца раскручиваться, и на каждой цепи из находящихся в окружающей среде свободных нуклеотидов собирается новая цепь. Сборка новой цепи идет в точном соответствии с принципом комплементарности. Против каждого А встает Т, против Г Ц и т. д. В результате вместо одной молекулы ДНК возникают две молекулы такого же точно нуклеотидного состава, как и первоначальная. Одна цепь в каждой вновь образовавшейся молекуле ДНК происходит из первоначальной молекулы, а другая синтезируется вновь.
Рибонуклеиновые кислоты (РНК). Структуры РНК сходны со структурами ДНК. РНК, как и ДНК, полинуклеотиды, но, в отличие от ДНК, молекула РНК одноцепочечная. Как и в ДНК, структура РНК создается чередованием четырех типов нуклеотидов, но состав нуклеотидов РНК несколько отличается от нуклеотидов ДНК, т. е. углевод в РНК не дезоксирибоза, а рибоза, отсюда и название РНК – рибонуклеиновая кислота. Кроме того, в РНК вместо азотистого основания тимина входит другое, близкое по строению основание, называемое урацилом (У).
В клетке имеется несколько видов РНК. Все они участвуют в синтезе белка. Первый вид – транспортные РНК (т-РНК). Это самые маленькие по размерам РНК. Они связывают аминокислоты и транспортируют их к месту синтеза белка. Второй вид – информационные РНК (и-РНК) . По размерам они раз в 10 больше т-РНК. Их функция состоит в переносе информации о структуре белка от ДНК к месту синтеза белка. Третий вид – рибосомные РНК (р-РНК). Они имеют наибольшие размеры молекулы и входят в состав рибосом.
1. Какие виды нуклеиновых кислот обнаружены в клетке? 2. Какова биологическая роль ДНК? 3. Охарактеризуйте структуру ДНК. 4. В чем сущность принципа комплиментарности? 5. Как осуществляется удвоение ДНК? 6. Какие виды РНК имеются в клетке? 7. Сравните структуры ДНК и РНК.
blgy.ru
НУКЛЕИНОВЫЕ КИСЛОТЫ | Энциклопедия Кругосвет
Содержание статьиНУКЛЕИНОВЫЕ КИСЛОТЫ – биологические полимерные молекулы, хранящие всю информацию об отдельном живом организме, определяющие его рост и развитие, а также наследственные признаки, передаваемые следующему поколению. Нуклеиновые кислоты есть ядрах клеток всех растительных и животных организмов, что определило их название (лат. nucleus – ядро).
Состав полимерной цепи нуклеиновых кислот.
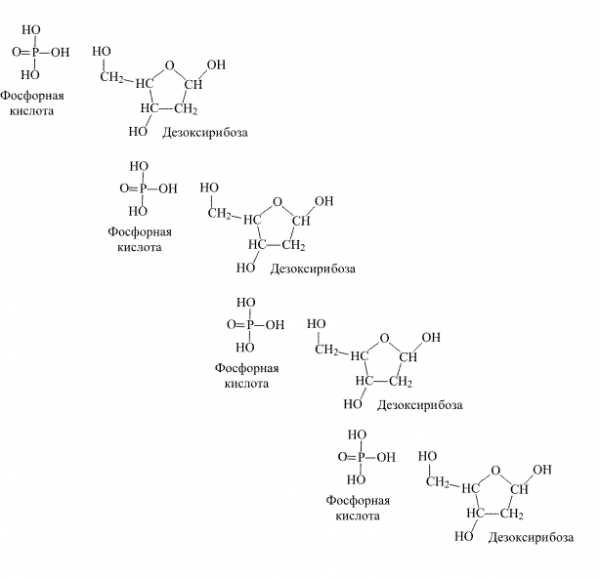
Полимерная цепь нуклеиновых кислот собрана из фрагментов фосфорной кислоты Н3РО3 и фрагментов гетероциклических молекул, представляющих собой производные фурана. Есть лишь два вида нуклеиновых кислот, каждая построена на основе одного из двух типов таких гетероциклов – рибозы или дезоксирибозы (рис. 1).
Рис. 1. СТРОЕНИЕ РИБОЗЫ И ДЕЗОКСИРИБОЗЫ.
Название рибоза (от лат. Rib – ребро, скрепка) имеет окончание – оза, что указывает на принадлежность к классу сахаров (например, глюкоза, фруктоза). У второго соединения нет группы ОН (окси-группа), которая в рибозе отмечена красным цветом. В связи с этим втрое соединение называют дезоксирибозой, т.е., рибоза, лишенная окси-группы.
Полимерная цепь, построенная из фрагментов рибозы и фосфорной кислоты, представляет собой основу одной из нуклеиновых кислот – рибонуклеиновой кислоты (РНК). Термин «кислота» в названии этого соединения употреблен потому, что одна из кислотных групп ОН фосфорной кислоты остается незамещенной, что придает всему соединению слабокислый характер. Если вместо рибозы в образовании полимерной цепи участвует дезоксирибоза, то образуется дезоксирибонуклеиновая кислота, для которой повсеместно принято широко известное сокращение ДНК.
Структура ДНК.
Молекула ДНК служит отправной точкой в процессе роста и развития организма. На рис. 2 показано, как объединяются в полимерную цепь два типа чередующихся исходных соединений, показан не способ синтеза, а принципиальная схема сборки молекулы ДНК.
В окончательном варианте полимерная молекула ДНК содержит в боковом обрамлении азотсодержащие гетероциклы. В образовании ДНК участвуют четыре типа таких соединений, два из них представляют собой шестичленные циклы, а два – конденсированные циклы, где шестичленное кольцо спаяно с пятичленным (рис. 3).

Рис. 3. СТРОЕНИЕ АЗОТСОДЕРЖАЩИХ ГЕТЕРОЦИКЛОВ, входящих в состав ДНК
На втором этапе сборки к свободным группам ОН дезоксирибозы присоединяются показанные выше азотсодержащие гетероциклические соединения, образуя у полимерной цепи боковые подвески (рис. 4).
Присоединенные к полимерной цепи молекулы аденина, тимина, гуанина и цитозина обозначают первыми буквами названий исходных соединений, то есть, А, Т, Г и Ц.
Сама полимерная цепь ДНК имеет определенную направленность – при мысленном продвижении вдоль молекулы в прямом и обратном направлении одни и те же группировки, входящие в состав цепи, встречаются на пути в разной последовательности. При движении в одном направлении от одного атома фосфора к другому вначале на пути следования идет группа СН2, а затем две группы СН (атомы кислорода можно не принимать во внимание), при движении в противоположном направлении последовательность этих групп будет обратной (рис. 5).
Рис. 5. НАПРАВЛЕННОСТЬ ПОЛИМЕРНОЙ ЦЕПИ ДНК. При описании того, в каком порядке чередуются присоединенные гетероциклы, принято использовать прямое направление, то есть от группы СН2 к группам СН.
Само понятие «направление цепи» помогает понять то, как располагаются две цепи ДНК при их объединении, а также имеет прямое отношение к синтезу белка.
На следующей стадии две молекулы ДНК объединяются, располагаясь таким образом, чтобы начало и концы цепей были направлены в противоположные стороны. В этом случае гетероциклы двух цепей обращены навстречу друг другу и оказываются расположенными неким оптимальным образом, имеется в виду, что между парами группировок С=О и NH2 , а также между єN и NH=, входящими в состав гетероциклов, возникают водородные связи (см. ВОДОРОДНАЯ СВЯЗЬ). На рис. 6 показано, как располагаются две цепи относительно друг друга и как при этом возникают водородные связи между гетероциклами. Самая важная деталь – состоит в том, что пары, связанные водородными связями, жестко определены: фрагмент А всегда взаимодействует с Т, а фрагмент Г – всегда с Ц. Строго определенная геометрия этих групп приводит к тому, что эти пары исключительно точно подходят друг другу (как ключ к замку), пара А-Т связана двумя водородными связями, а пара Г-Ц – тремя связями.
Водородные связи заметно слабее обычных валентных связей, но из-за большого их количества вдоль всей полимерной молекулы соединение двух цепей становится достаточно прочным. В молекуле ДНК содержится десятки тысяч групп А, Т, Г и Ц и порядок их чередования в пределах одной полимерной молекулы может быть различным, например, на определенном участке цепи последовательность может иметь вид: —А—А—Т—Г—Ц—Г—А—Т-. Поскольку взаимодействующие группы строго определены, то на противолежащем участке второй полимерной молекулы обязательно будет последовательность –Т—Т—А—Ц—Г—Ц—Т—А-. Таким образом, зная порядок расположения гетероциклов в одной цепи, можно указать их размещение в другой цепи. Из этого соответствия следует, что суммарно в сдвоенной молекуле ДНК количество групп А равно количеству групп Т, а количество групп Г – количеству Ц (правило Э.Чаргаффа).
Две молекулы ДНК, связанные водородными связями, показаны на рис. 5 в виде двух плоско лежащих цепей, однако в действительности они располагаются иным образом. Истинное направление в пространстве всех связей, определяемое валентными углами и стягивающими водородными взаимодействиями, приводит к определенном изгибам полимерных цепей и повороту плоскости гетероциклов, что приблизительно показано в первом видеофрагменте рис. 7 с помощью структурной формулы. Гораздо точнее всю пространственную конструкцию можно передать только с помощью объемных моделей (рис. 7, второй видеофрагмент). При этом возникает сложная картина, поэтому принято использовать упрощенные изображения, которые особенно широко применяют при изображении структуры нуклеиновых кислот или белков. В случае нуклеиновых кислот полимерные цепи изображают в форме плоских лент, а гетероциклические группировки А, Т, Г и Ц – в виде боковых стержней или простых валентных штрихов, имеющих различные цвета, либо содержащих на конце буквенные обозначения соответствующих гетероциклов (рис. 7, третий видеофрагмент).
Во время поворота всей конструкции вокруг вертикальной оси (рис. 8) отчетливо видна спиральная форма двух полимерных молекул, которые как бы навиты на поверхность цилиндра, это широко известная двойная спираль ДНК.
При таком упрощенном изображении не исчезает основная информация – порядок чередования группировки А, Т, Г и Ц, определяющий индивидуальность каждого живого организма, вся информация записана четырехбуквенным кодом.
Строение полимерной цепи и обязательное присутствие четырех типов гетероциклов однотипно для всех представителей живого мира. У всех животных и высших растений количество пар А – Т всегда несколько больше, чем пар Г – Ц. Отличие ДНК млекопитающих от ДНК растений в том, что у млекопитающих пара А – Т на всем протяжении цепи встречается ненамного чаще (приблизительно в 1,2 раза), чем пара Г – Ц. В случае растений предпочтительность первой пары гораздо более заметна (приблизительно в 1,6 раза).
ДНК – одна из самых больших известных на сегодня полимерных молекул, у некоторых организмов ее полимерная цепь состоит из сотен миллионов звеньев. Длина такой молекулы достигает нескольких сантиметров, это очень большая величина для молекулярных объектов. Т.к. поперечное сечение молекулы всего 2 нм (1нм = 10–9 м), то ее пропорции можно сопоставить с железнодорожным рельсом длиной в десятки километров.
Химические свойства ДНК.
В воде ДНК образует вязкие растворы, при нагревании таких растворов до 60° С или при действии щелочей двойная спираль распадается на две составляющие цепи, которые вновь могут объединиться, если вернуться к исходным условиям. В слабокислых условиях происходит гидролиз, в результате частично расщепляются фрагменты –Р-О-СН2— с образованием фрагментов –Р-ОН и НО-СН2 , соответственно результате образуются мономерные, димерные (сдвоенные) или тримерные (утроенные) кислоты, представляющие собой звенья, из которых была собрана цепь ДНК (рис. 9).

Рис. 9. ФРАГМЕНТЫ, ПОЛУЧАЕМЫЕ ПРИ РАСЩЕПЛЕНИИ ДНК.
Более глубокий гидролиз позволяет отделить участки дезоксирибозы от фосфорной кислоты, а также группировку Г от дезоксирибозы, т.е., более детально разобрать молекулу ДНК на составляющие компоненты. При действии сильных кислот (помимо распада фрагментов –Р(О)-О-СН2-) отщепляются и группировки А и Г. Действие иных реагентов (например, гидразина) позволяет отделить группировки Т и Ц. Более деликатное расщепление ДНК на компоненты проводят с помощью биологического препарата – дезоксирибонуклеазы, выделяемой из поджелудочной железы (окончание —аза всегда указывает на то, что данное вещество представляет собой катализатор биологического происхождения – фермент). Начальная часть названия – дезоксирибонуклеаза – указывает, какое именно соединение расщепляет этот фермент. Все указанные способы расщепления ДНК ориентированы, в первую очередь, на детальный анализ ее состава.
Самая важная информация, содержащаяся в молекуле ДНК, – порядок чередования групп А, Т, Г и Ц , ее получают с помощью специально разработанных методик. Для этого создан широкий набор ферментов, которые находят в молекуле ДНК строго определенную последовательность, например, Ц—T—Г—Ц—A—Г (а также соответствующую ей последовательность на противоположной цепи Г—А—Ц—Г—Т—Ц) и вычленяют ее из состава цепи. Таким свойством обладает фермент Pst I (торговое наименование, оно образуется из названия того микроорганизма Providencia stuartii, из которого получают этот фермент). При использовании другого фермента Pal I удается найти последовательность Г—Г—Ц—Ц. Далее сопоставляются результаты, полученные при действии широкого набора различных ферментов по заранее разработанной схеме, в результате удается определить последовательность таких групп на определенном участке ДНК. Сейчас подобные методики доведены до стадии широкого применения, они используются в самых разнообразных областях, далеких от научных биохимических исследований, например, при идентификации останков живых организмов или установлении степени родства.
Структура РНК
во многом напоминает ДНК, отличие в том, что в основной цепи фрагменты фосфорной кислоты чередуются с рибозой, а не с дезоксирибозой (рис.). Второе отличие – к боковому обрамлению присоединяется гетероцикл урацил (У) вместо тимина (Т), остальные гетероциклы А, Г и Ц те же, что у ДНК. Урацил отличается от тимина отсутствием метильной группы, присоединенной к циклу, на рис. 10 эта метильная группа выделена красным цветом.
Рис. 10. ОТЛИЧИЕ ТИМИНА ОТ УРАЦИЛА – отсутствие у второго соединения метильной группы, выделенной в тимине красным цветом.
Фрагмент молекулы РНК показан на рис. 11, порядок следования группировок А, У, Г и Ц, а также их количественное соотношение может быть различным.

Рис.11. ФРАГМЕНТ МОЛЕКУЛЫ РНК. Основное отличие от ДНК – наличие группировок ОН в рибозе (красный цвет) и фрагмента урацила (синий цвет).
Полимерная цепь РНК приблизительно в десять раз короче, чем у ДНК. Дополнительное отличие в том, что молекулы РНК не объединяются в двойные спирали, состоящие из двух молекул, а обычно существуют в виде одиночной молекулы, которая на некоторых участках может образовывать сама с собой двухцепные спиральные фрагменты, чередующиеся с линейными участками. На спиральных участках взаимодействие пар соблюдается также строго, как в ДНК. Пары, связанные водородными связями и формирующие спираль (А—У и Г—Ц), возникают на тех участках, где расположение групп оказывается благоприятным для такого взаимодействия (рис. 12).
Для подавляющего большинства живых организмов количественное содержание пар А—У больше чем Г—Ц, у млекопитающих в 1,5–1,6 раза, у растений – в 1,2 раза. Существует несколько типов РНК, роли, которых в живом организме различны.
Химические свойства РНК
напоминают свойства ДНК, однако наличие дополнительных групп ОН в рибозе и меньшее (в сравнении с ДНК) содержание стабилизированных спиральных участков делает молекулы РНК химически более уязвимыми. При действии кислот или щелочей основные фрагменты полимерной цепи Р(О)-О-СН2 легко гидролизуются, группировки А, У, Г и Ц отщепляются легче. Если нужно получить мономерные фрагменты (подобные тем, что на рис. 9), сохранив при этом химически связанные гетероциклы, используют деликатно действующие ферменты, называемые рибонкулеазами.
Участие ДНК и РНК в синтезе белков
– одна из основных функций нуклеиновых кислот. Белки – важнейшие компоненты каждого живого организма. Мышцы, внутренние органы, костная ткань, кожный и волосяной покров млекопитающих состоят из белков. Это полимерные соединения, которые собираются в живом организме из различных аминокислот. В такой сборке управляющую роль играют нуклеиновые кислоты, процесс проходит в две стадии, причем на каждой из них определяющий фактор – взаимоориентация азотсодержащих гетероциклов ДНК и РНК.
Основная задача ДНК – хранить записанную информацию и предоставлять в тот момент, когда начинается синтез белков. В связи с этим понятна повышенная химическая устойчивость ДНК в сравнении с РНК. Природа позаботилась о том, чтобы сохранить по возможности основную информацию неприкосновенной.
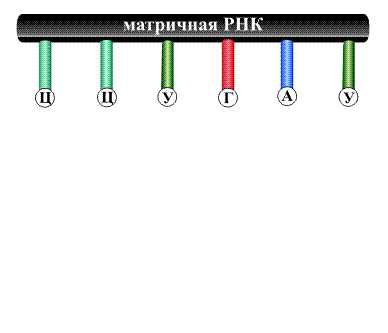
На первой стадии часть двойной спирали раскрывается, освободившиеся ветви расходятся, и на группах А, Т, Г и Ц, оказавшихся доступными, начинается синтез РНК, называемой матричной РНК, поскольку она как копия с матрицы точно воспроизводит информацию, записанную на раскрывшемся участке ДНК. Напротив группы А, принадлежащей молекуле ДНК, располагается фрагмент будущей матричной РНК, содержащий группу У, все остальные группы располагаются друг напротив друга в точном соответствии с тем, как это происходит при образовании двойной спирали ДНК (рис. 13).
По указанной схеме образуются полимерная молекула матричной РНК, содержащая несколько тысяч мономерных звеньев.
На втором этапе матричная ДНК перемещается из ядра клетки в околоядерное пространство – цитоплазму. К полученной матричной РНК подходят так называемые транспортные РНК, которые несут с собой (транспортируют) различные аминокислоты. Каждая транспортная РНК, нагруженная определенной аминокислотой, приближается к строго обусловленному участку матричной РНК, нужное место обнаруживается с помощью все того же принципа взаимосоответствия групп А—У, и Г—Ц. В конечном итоге две аминокислоты, оказавшиеся рядом, взаимодействуют между собой, так начинается сборка будущей белковой молекулы (рис. 14).
Важная деталь состоит в том, что временное взаимодействие матричной и транспортной РНК проходит всего по трем группам, например, к триаде Ц—Ц—У матричной кислоты может подойти только соответствующая ей тройка Г—Г—А транспортной РНК, которая непременно несет с собой аминокислоту глицин (рис. 14). Точно также к триаде Г—А—У может приблизиться лишь набор Ц—У—А, транспортирующий только аминокислоту лейцин. Таким образом, последовательность групп в матричной РНК указывает, в каком порядке должны соединяться аминокислоты. Кроме того, система содержит в закодированном виде дополнительные регулирующие правила, некоторые последовательности из трех групп матричной РНК указывает на то, что в этом месте синтез белка должен остановиться, т.е. молекула достигла необходимой длины.
Показанный на рис. 14 синтез белка проходит с участием еще одного – третьего вида РНКислот, они входят в состав рибосом и потому их называют рибосомными. Рибосома, представляющая собой ансамбль определенных белков рибосомных РНК, обеспечивает взаимодействие матричной и транспортной РНК, играя роль конвейерной ленты, которая передвигает матричную РНК на один шаг после того, как произошло соединение двух аминокислот.
Основной смысл двухстадийной схемы, показанной на рис. 13 и 14, состоит в том, что полимерная цепь белковой молекулы собирается из различных аминокислот в намеченном порядке и строго по тому плану, который был записан в закодированном виде на определенном участке ДНК. Таким образом, ДНК представляет собой отправную точку всего этого запрограммированного процесса.
В процессе жизнедеятельности белки постоянно расходуются, и потому они регулярно воспроизводятся по описанной схеме, весь синтез белковой молекулы, состоящей из сотен аминокислот, проходит в живом организме приблизительно в течение одной минуты.
Первые исследования нуклеиновых кислот были проведены во второй половине 19 в., понимание того, что в ДНК зашифрована вся информация о живом организме, пришло в середине 20 в., структуру двойной спирали ДНК установили в 1953 Дж.Уотсон и Ф.Крик на основании данных рентгеноструктурного анализа, что признано крупнейшим научным достижением 20 столетия. В середине 70-х годов 20 в. появились методики расшифровки детальной структуры нуклеиновых кислот, а вслед за тем были разработаны способы их направленного синтеза. Сегодня ясны далеко не все процессы, происходящие в живых организмах с участием нуклеиновых кислот, и сегодня это одна из самых интенсивно развивающихся областей науки.
Михаил Левицкий
www.krugosvet.ru
НУКЛЕИНОВЫЕ КИСЛОТЫ — это… Что такое НУКЛЕИНОВЫЕ КИСЛОТЫ?
Вирусная РНК может транскрибироваться ферментом — обратной транскриптазой — в ДНК, которая иногда включается в хромосомную ДНК клетки-хозяина. Теперь эта ДНК несет вирусные гены, и после транскрипции в клетке может появиться вирусная РНК. Таким образом, спустя длительное время, в течение которого никакого вируса в клетке не обнаруживается, он снова в ней появится без повторного заражения. Вирусы, генетический материал которых включается в хромосому клетки-хозяина, часто являются причиной рака.
Генетическая информация, закодированная в нуклеотидной последовательности ДНК, переводится не только на язык нуклеотидной последовательности РНК, но и на язык аминокислот — мономерных единиц белков. Белковая молекула — это цепочка из аминокислот. Каждая аминокислота содержит кислую карбоксильную группу -COOH и оснвную аминогруппу -Nh3. Карбоксильная группа одной аминокислоты связывается с аминогруппой другой, образуя амидную связь, и этот процесс продолжается, пока не образуется цепь, содержащая до 1000 аминокислот (см. также
). В белках присутствует 20 разных аминокислот, от последовательности которых зависят их природа и функции. Эта последовательность определяется нуклеотидной последовательностью соответствующего гена — участка ДНК, кодирующего данный белок. Однако сама ДНК не является матрицей при синтезе белка. Сначала она транскрибируется в ядре с образованием матричной РНК (мРНК), которая диффундирует в цитоплазму, и на ней как на матрице синтезируется белок. Процесс ускоряется благодаря тому, что на каждой молекуле мРНК может одновременно синтезироваться множество белковых молекул. Репликация нуклеиновых кислот осуществляется благодаря образованию водородных связей между комплементарными основаниями исходной и дочерней цепей. Аминокислоты не образуют водородных связей с основаниями, так что прямое копирование матрицы невозможно. Они взаимодействуют с матрицей опосредованно, через «адапторные» нуклеиновые кислоты — небольшие молекулы транспортных РНК (тРНК), состоящие примерно из 80 оснований и способные связываться с мРНК. Каждая тРНК содержит специфическую последовательность из трех оснований, антикодон, который комплементарен группе из трех оснований, кодону, в мРНК. Антикодоны взаимодействуют с кодонами по правилу комплементарности, примерно так же, как взаимодействуют две цепи ДНК. Таким образом, последовательность оснований в мРНК определяет порядок присоединения тРНК, несущих аминокислоты. Схематически перенос информации от ДНК к белку можно представить следующим образом:

Последовательность оснований в ДНК задает порядок следования аминокислот в белке, поскольку каждая аминокислота присоединяется специфическим ферментом только к определенным тРНК, а те, в свою очередь, — только к определенным кодонам в мРНК. Комплексы тРНК-аминокислота связываются с матрицей по одному в каждый данный момент времени. Ниже перечислены основные этапы белкового синтеза (см. также рисунок).

ЭТАПЫ БЕЛКОВОГО СИНТЕЗА
1. Ферменты, называемые аминоацил-тРНК-синтетазами, присоединяют аминокислоты к соответствующим тРНК. Таких ферментов 20, по одному для каждой аминокислоты. 2. Молекула мРНК присоединяется своим первым кодоном к небольшой частице, называемой рибосомой. Рибосомы состоят из примерно равных количеств рРНК и белка. Структура и функция рибосом весьма сложны, но главная их задача — облегчение взаимодействия мРНК и тРНК и ускорение полимеризации аминокислот, связанных с разными тРНК. 3. тРНК, нагруженная аминокислотой, связывается с соответствующим кодоном мРНК, которая, в свою очередь, контактирует с рибосомой. Образуется комплекс рибосома-мРНК-тРНК-аминокислота. 4. мРНК, подобно ленте на конвейере, продвигается по рибосоме на один кодон вперед. 5. Следующая тРНК, нагруженная аминокислотой, присоединяется ко второму кодону. 6. Первая и вторая аминокислоты связываются между собой. 7. Первая тРНК отсоединяется от комплекса, и теперь вторая тРНК несет две аминокислоты, связанные между собой. 8. мРНК снова продвигается на один кодон вперед, и все события повторяются, а растущая аминокислотная цепь удлиняется на одну аминокислоту. Процесс продолжается, пока не будет достигнут последний, «стоп»-кодон и последняя тРНК не отделится от готовой белковой цепи. В бактериальных клетках цепь из 100-200 аминокислот собирается за несколько секунд. В животных клетках этот процесс занимает около минуты.
Генетический код. Итак, каждая аминокислота в белке опосредованно детерминируется определенным кодоном (группой из 3 оснований) в мРНК и в конечном счете в ДНК. Поскольку в нуклеиновых кислотах имеется четыре вида оснований, число возможных кодонов составляет 4ґ4ґ4 = 64. Соответствие между кодонами и аминокислотами, которые они кодируют, называется генетическим или биологическим кодом. Это соответствие было установлено опытным путем: к разрушенным клеткам добавляли синтетические полинуклеотиды известного состава и смотрели, какие аминокислоты включаются в белки. Позднее появилась возможность прямо сравнить последовательности аминокислот в вирусных белках и оснований в вирусных нуклеиновых кислотах. Чрезвычайно интересно, что генетический код, за редкими исключениями, одинаков для всех организмов — от вирусов до человека. Одно из таких исключений составляют изменения в генетическом коде, используемом митохондриями. Митохондрии — это небольшие автономные субклеточные частицы (органеллы), присутствующие во всех клетках, кроме бактерий и зрелых эритроцитов. Предполагают, что когда-то митохондрии были самостоятельными организмами; проникнув в клетки, они со временем стали их неотъемлемой частью, но сохранили некоторое количество собственной ДНК и синтезируют несколько митохондриальных белков.
Вообще говоря, каждой аминокислоте соответствует более одного кодона. Большинство кодонов, кодирующих одну и ту же аминокислоту, имеют два одинаковых первых основания, но в трех случаях (для лейцина, серина и аргинина) имеются два альтернативных набора первых дублетов в кодонах, соответствующих одной и той же аминокислоте. Природа основания в третьем положении не столь важна; одна и та же аминокислота — глицин — может кодироваться по-разному: ГГУ, ГГЦ, ГГА и ГГГ. Однако кодоны для двух разных аминокислот могут иметь два одинаковых первых основания, и тогда различие между ними будет определяться природой третьего основания — пурином или пиримидином. Так, гистидин кодируется триплетами ЦАУ и ЦАЦ, а глутамин — ЦАА и ЦАГ. Три кодона, УАА, УАГ и УГА, не кодируют никаких аминокислот и называются «бессмысленными». Одна молекула ДНК кодирует много белковых цепей. Каждый отрезок, кодирующий одну цепь, называют цистроном. Начало и конец цистрона, а также граница раздела между ними помечаются с помощью своего рода знаков химической пунктуации. По крайней мере у бактерий в начале цистрона находится метиониновый кодон АУГ. Логично предположить, что первой аминокислотой в белке всегда должен быть метионин, но часто несколько первых аминокислот отщепляются ферментативно после окончания синтеза белка. Конец белковой цепи помечается одним или несколькими «бессмысленными» кодонами. У бактерий (прокариот) практически вся ДНК кодирует какие-либо белки или тРНК. Однако у высших форм (эукариот) значительная часть ДНК состоит из простых повторяющихся последовательностей и «молчащих» генов, которые не транскрибируются в РНК и поэтому не транслируются в белки. Кроме того, исходно синтезированная мРНК содержит участки, не детерминирующие никаких белковых последовательностей. Такие участки (интроны), расположенные между кодирующими участками (экзонами), перед началом синтеза белка удаляются специальными ферментами. Почему в ДНК существуют эти казалось бы бесполезные сегменты — неясно; возможно, они выполняют регуляторные функции. У простейшей Tetrahymena РНК сама удаляет свои интроны и соединяет свободные концы цепей, действуя как фермент по отношению к себе самой. Это единственное известное исключение из правила, согласно которому нуклеиновые кислоты не обладают ферментативной активностью.
Транспортные РНК и супрессия. Смысл информации, содержащейся в ДНК, если переводить ее на язык аминокислот, определяется как самой ДНК, так и считывающим механизмом, т.е. зависит не только от того, какие кодоны есть в ДНК и в какой последовательности они расположены, но также и от того, какие именно аминокислоты (и к каким тРНК) присоединяют аминоацил-тРНК-синтетазы. Конечно, природа синтетаз и тРНК тоже определяется ДНК, и в этом смысле ДНК является первичным детерминантом белковой последовательности. Тем не менее суммарная детерминация представляет собой функцию всей системы, поскольку результат зависит от исходных компонентов. Если бы соответствие между тРНК и аминокислотами было другим, смысл кодонов тоже изменился бы. Известно, что мутации в ДНК изменяют считывающий механизм и в результате меняют — пусть и незначительно — смысл кодонов. Так, в бактерии Escherichia coli глициновая тРНК обычно узнает в мРНК кодон ГГА; мутация в ДНК, с которой транскрибируется эта тРНК, изменяет антикодон глициновой тРНК таким образом, что теперь он узнает кодон АГА, соответствующий аргинину, и в белковой молекуле вместо аргинина появляется глицин. Это не обязательно имеет фатальные последствия, поскольку не все аргинины кодируются триплетом АГА и есть аргининовые тРНК, по-прежнему узнающие «свои» АГА. В результате измененными оказываются не все белковые молекулы. Иногда такие мутации, изменяющие антикодон, подавляют (супрессируют) мутации в кодоне. Например, если в результате мутации глициновый кодон ГГА превращается в АГА, он все же может прочитываться как глицин, если антикодон глициновой тРНК, в свою очередь, изменился так, что эта тРНК стала узнавать АГА. В этом случае вторая «ошибка» устраняет первую. Мутации, приводящие к изменению антикодонов, могут иметь разные последствия, поскольку один и тот же кодон может узнаваться несколькими тРНК. Вообще говоря, узнавание осуществляется благодаря комплементарности оснований кодона и антикодона, однако одно из оснований кодона может модифицироваться таким образом, что антикодон будет узнавать даже неполностью комплементарный кодон. В результате одна и та же тРНК может взаимодействовать с несколькими разными кодонами, кодирующими одну и ту же аминокислоту. Этот феномен неполного соответствия кодона и антикодона был назван Ф. Криком «шатанием».
Регуляция активности генов. Для организма было бы катастрофой, если бы во всех его клетках одновременно работали все гены и синтезировались все закодированные ими белки. Бактерии, например, должны все время приспосабливаться к условиям среды, синтезируя нужные ферменты. Все клетки высших организмов имеют один и тот же набор генов, но, к счастью, клетки мозга не продуцируют пищеварительные ферменты, а в хрусталике глаза не синтезируются мышечные белки. Активность гена характеризуется тем, транскрибируется ли он с образованием соответствующей мРНК. ДНК — длинная молекула, и в определенных ее участках имеются последовательности, называемые промоторами, которые распознаются специфическим транскрибирующим ферментом — полимеразой. В этих участках и только в них начинается транскрипция, продолжаясь до тех пор, пока не достигнет последовательности оснований, означающей конец считывания. Существуют особые репрессорные белки, которые связываются с ДНК поблизости от промотора в участке, называемом оператором. Образовавшийся комплекс блокирует транскрипцию, и мРНК не синтезируется. Таким образом, репрессорные белки являются ингибиторами транскрипции. С другой стороны, существуют небольшие молекулы, которые образуют комплекс с репрессорами и снимают их блокирующее действие на транскрипцию. Иными словами, они ингибируют ингибиторы. Так, у бактерий в норме отсутствуют ферменты, катализирующие расщепление некоторых сахаров; однако если один из этих сахаров появляется в среде, он образует комплекс с репрессором, ингибирование снимается и запускается синтез соответствующего фермента. Ферменты, синтез которых индуцируется собственными субстратами, называются индуцибельными. В ряде случаев, наоборот, репрессорный белок не блокирует транскрипцию мРНК, если он не связан с определенной молекулой. У бактерий некоторые ферменты, участвующие в синтезе определенных аминокислот, образуются только в отсутствие этих аминокислот, т.е. бактерии производят данные ферменты лишь по мере надобности. Если добавить в среду соответствующую аминокислоту, она образует комплекс с репрессором и активирует его, а тем самым ингибирует транскрипцию соответствующих генов. Уже образовавшаяся мРНК вскоре расщепляется, и синтез ферментов останавливается. Такие ферменты являются отрицально индуцибельными. Поскольку репрессорные белки сами кодируются генами, работа которых, в свою очередь, может регулироваться другими генами, а синтез малых молекул-индукторов и гормонов также в конечном счете регулируется генами, механизмы регуляции генной активности могут быть очень сложными.
ЛИТЕРАТУРА
Ичас М. Биологический код. М., 1971 Шабарова З.А., Богданов А.А. Химия нуклеиновых кислот и их компонентов, М., 1978 Зенгер В. Принципы структурной организации нуклеиновых кислот. М., 1987
Энциклопедия Кольера. — Открытое общество. 2000.
dic.academic.ru
