
Вопрос №3. Мембранный принцип организации клетки
Цитоплазма представляет собой метаболический рабочий аппарат клетки. В ней сосредоточены общие и специальные органоиды, в цитоплазме протекают основные метаболические процессы.
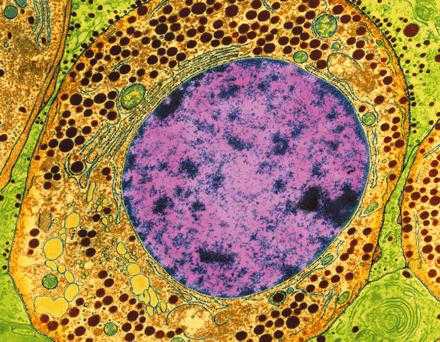
Общей чертой всех мембран клетки, внешней плазматической мембраны и всех внутриклеточных мембран и мембранных органоидов является то, что они представляют собой тонкие (6-10 нм) пласты липопротеидной природы (липиды в комплексе с белками), замкнутые сами на себя. В клетке нет открытых мембран со свободными концами. Мембраны клетки всегда ограничивают полости или участки, закрывая их со всех сторон и тем самым отделяя содержимое таких полостей от окружающей их среды. Так, плазматическая мембрана, покрывая всю поверхность клетки, имеющей сложную форму и многочисленные выросты, нигде не прерывается, она замкнута. Она отделяет содержимое цитоплазмы от окружающей клетку среды. Внутриклеточные замкнутые мембраны образуют пузырьки — вакуоли шаровидной или уплощенной формы. В последнем случае образуются плоские мембранные мешки, или цистерны. Часто полости, отграниченные мембранами, имеют сложную форму, напоминающую губку или сеть, но и в этом случае такие полости без перерывов отграничены мембраной. В подобных вариантах мембраны также разделяют две структурные фазы цитоплазмы: гиалоплазму от содержимого вакуолей и цистерн. Такое же свойство имеют мембраны митохондрий и пластид: они разделяют внутреннее содержимое от межмембранных полостей и от гиалоплазмы. Ядерная оболочка тоже может быть представлена в виде перфорированного полого двойного мембранного мешка шаровидной формы. Мембраны ядерной оболочки разграничивают, отделяют друг от друга кариоплазму и хромосомы от полости перинуклеарного пространства и от гиалоплазмы. Эти общие морфологические свойства клеточных мембран определяются их химическим составом, их липопротеидной природой.
Вопрос №4. Строение и функции органоидов и включений в клетке
Строение клетки.
Клетки находятся в межклеточном веществе, обеспечивающем их
механическую прочность, питание и дыхание. Основные части любой клетки –
цитоплазма и ядро.
Клетка покрыта мембраной, состоящей из нескольких слоёв молекул,
обеспечивающей избирательную проницаемость веществ. В цитоплазме
расположены мельчайшие структуры – органоиды. К органоидам клетки
относятся: эндоплазматическая сеть, рибосомы, митохондрии, лизосомы,
комплекс Гольджи, клеточный центр.
ВКЛЮЧЕНИЯ КЛЕТКИ, компоненты цитоплазмы, представляющие собой отложения веществ, временно выведенных из обмена или конечных его продуктов. Специфика включений клетки связана со специализацией соответствующих клеток, тканей и органов. Наиболее распространены трофические включения клетки — капли жира, глыбки гликогена, желток в яйцах. В растительных клетках включения представлены главным образом крахмальными и алейроновыми зёрнами и липидными каплями. К включениям клетки относят также секреторные гранулы в железистых клетках животных, кристаллы некоторых солей (главным образом оксалат кальция) в клетках растений. Особый вид включений клетки — остаточные тельца — продукты деятельности лизосом.
studfiles.net
ПРИНЦИПЫ СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ КЛЕТКИ — Мегаобучалка
В природе существует значительное разнообразие конкретных клеточных форм. Вместе с тем число основных типов клеточной организации ограниченно. Выделяют прокариотический и эукариотический типы с подразделением второго на подтип клеток простейших организмов и подтип клеток многоклеточных (рис. 1).
Клетки прокариотического типа имеют особенно малые размеры — не более 0,5—3,0 мкм в диаметре. У них нет морфологически обособленного ядра, так как ядерный материал в виде ДНК не отграничен от цитоплазмы оболочкой. В клетке отсутствует развитая система мембран. Генетический аппарат образован единственной кольцевой хромосомой, которая лишена основных белков — гистонов. Гистоны являются белками клеточных ядер. Благодари значительному количеству основных аминокислот аргинина и лизина они имеют щелочной характер. В хромосомах эукариотических клеток гистоны, соединяясь с ДНК, подавляют процесс считывания биологической информации. Различия прокариотических и эукариотических клеток но содержанию гистонов указывают на разные механизмы регуляции функции наследственного материала.
У прокариот отсутствует клеточный центр. Для них не типичны внутриклеточные перемещения цитоплазмы и амебоидное движение. Время, необходимое для образования двух дочерних клеток из материнской (время генерации), сравнительно мало и исчисляется десятками минут. Прокариотические клетки не делятся митозом. К этому типу клеток относятся бактерии и сине-зеленые водоросли.
Эукариотический тип клеточной организации представлен двумя подтипами. Особенностью организмов простейших является то, что они, исключая колониальные формы, в структурном отношении представляют собой клетку, в физиологическом — полноценную особь. В связи с этим в клетках некоторых простейших имеются миниатюрные образования, выполняющие на клеточном уровне функции органов, аппаратов и систем органов многоклеточного организма (цитостом, цитофаринкс и порошица, сократительные вакуоли, генеративное и вегетативное ядра инфузорий).
ЭВОЛЮЦИЯ КЛЕТКИ
Прокариотические клетки появились на Земле ранее эукариотических. По данным палеонтологии первые существовали 3,0— 3,5 млрд. лет тому назад, тогда как ископаемые останки вторых встречаются в породах, датируемых не ранее чем 1,0—1,4 млрд. лет. Это обстоятельство, а также сходство основных биохимических процессов заставляет думать о происхождении эукариотической клетки от прокариотической предковой формы.
Согласно симбиотической гипотезе, популярной в настоящее время, корпускулярные органеллы эукариотической клетки, имеющие собственный геном, характеризуются независимым происхождением и ведут начало от прокариотических клеток-симбионтов. Предположительно клеткой-хозяином служил анаэробный прокариот, способный лишь к амебоидному движению. Митохондрии и жгутики произошли путем изменений проникших в клетку-хозяина симбионтов — аэробных прокариотов и бактерий, напоминающих современных спирохет. Хлоропласты клеток зеленых растений возникли из симбионтов — прокариотических клеток сине-зеленых водорослей. Особое значение имело приобретение клеткой в процессе эволюции жгутиков с банальными тельцами, близкими к центриолям. Это позволило интенсифицировать двигательную активность и создать механизм митоза.
Трудным является вопрос о происхождении ядра. Первоначально объем информации и геномах клетки-хозяина, с одной стороны, и симбионтов — презумптивных митохондрий, центриолей и хлоропластов, с другой, был, по-видимому, сопоставим. В дальнейшем могла произойти утрата геномами симбионтов части генетических функций с перемещением блоков генов в геном клетки-хозяина. Это сопровождалось ростом объема ядерного генома, централизацией генетической регуляции функций клетки и потерей митохондриями и другими корпускулярными органеллами их автономности. Нельзя, однако, исключить, что ядро также представляет собой производное внутриклеточного симбионта. Происхождение внутриплазматических мембран шероховатой и гладкой цитоплазматической сети, пластинчатого комплекса и везикуло-вакуолярных структур объясняют эволюционными преобразованиями наружной мембраны ядерной оболочки. Симбиотическая гипотеза не объясняет ряд фактов. Так, белок бациллин, из которого состоят реснички и жгутики современных прокариот, отличен от белка тубулина эукариот. У бактерий не обнаружено структур с типичным для жгутиков, ресничек, базальных телец или центриолей эукариотической клетки составом микротрубочек «9 + 2» или «9 + 0».
Инвагинационная гипотеза происхождения эукариотической клетки исходит из того, что предковый формой был аэробный прокариот. Он содержал несколько геномов, каждый из которых прикреплялся к клеточной оболочке. Корпускулярные органеллы, имеющие ДНК, а также ядро, возникли в результате впячивания и отшнуровки фрагментов оболочки вместе с геномом с последующей функциональной специализацией в ядро, митохондрии, хлоропласты, усложнением ядерного генома, развитием цитоплазматических мембран. Эта гипотеза удовлетворительно объясняет наличие 2 мембран в оболочке ядра, митохондрий и хпоропластов. Она встречается с трудностями в объяснении различий в деталях процесса биосинтеза белка в корпускулярных органеллах и цитоплазме эукариотической клетки. В митохондриях и хлоропластах этот процесс в точности соответствует таковому в современных прокариотических клетках. Симбиотическая и инвагинационная гипотезы не исчерпывают все точки зрения на происхождение эукариотического типа клеточной организации.
Эволюционные возможности эукариотического типа клеточной организации превосходят таковые у прокариот. Указанную особенность объясняют, прежде всего, увеличением размеров ядерного генома. Количество генов в геноме бактерии и клетки человека соотносится как 1:100—1000. Известное значение могли иметь наличие в клетке двух аллелей каждого гена (диплоидность), многократное повторение некоторых генов. Все это расширяет масштабы мутационной изменчивости, совместимой с выживанием, и способствует образованию резерва наследственной изменчивости.
При переходе к эукариотическому типу усложнился также механизм регуляции жизнедеятельности клетки. На уровне генетического материала это проявилось в увеличении относительного числа регуляторных генов, замене единичных «голых» молекул ДНК множественными хромосомами, в которых ДНК комплексируется с белками. В итоге оказалось возможным считывать информацию по частям с разных блоков генов в разных клетках и в разное время. В бактериальной клетке, например, одновременно транскрибируется 80—100% информации генома. В клетках человека в зависимости от органа считывается от 8—10% (печень, почка, селезенка) до 44% (головной мозг). Использованию информации по частям принадлежит исключительная роль в образовании многоклеточных живых форм. Определенное значение для перехода к многоклеточности имело, по-видимому, и приобретение клетками эукариотического типа эластичной оболочки, что позволяет образовывать стабильные комплексы клеток. В условиях усложнения генетического аппарата эукариот важным эволюционным приобретением стал митоз как механизм воспроизведения в поколениях генетически сходных клеток. Возникший путем эволюции митоза мейотический процесс интенсифицировал комбинативную изменчивость. Благодаря отмеченным особенностям за 1 млрд. лет своего существования эукариотическая клетка обеспечила развитие разнообразных живых форм от простейших до млекопитающих и человека.
КЛЕТОЧНАЯ ТЕОРИЯ
Клеточная теория служит фундаментальным обобщением биологии. Она устанавливает связь всех проявлений жизни на Земле с клеткой, отмечает универсальность принципов клеточной организации, определяет клетку одновременно и как целостную самостоятельную единицу биологической активности, и как составную часть существ высшего порядка — многоклеточных растительных и животных организмов.
Клеточная теория сформулирована немецким исследователем — зоологом Т. Шванном (1839). В своих теоретических построениях Т. Шванн широко использовал работы ботаника М. Шлейдена, которого по праву считают соавтором клеточной теории. Исходя из предположения об общей природе или гомологичности растительных и животных клеток, доказываемой одинаковым механизмом их возникновения, Т. Шванн обобщил многочисленные данные в виде теории, согласно которой клетки являются структурной и функциональной основой живых существ. В конце прошлого столетия клеточная теория получила дальнейшее развитие в трудах патолога Р. Вирхова. Он сделал важный вывод о том, что клетки образуются лишь путем деления предшествующих клеток. Им также создана вызвавшая критику концепция «клеточного государства». Согласно последней многоклеточный организм состоит из относительно самостоятельных единиц (клеток), поставленных в своей жизнедеятельности в тесную зависимость друг от друга. Указанная концепция принижала значение интеграции клеток в единую систему организма. Она не воспринимала организм как целостность.
Клеточная теория в современном виде включает три главных положения. Первое положение соотносит клетку с живой, природой планеты в целом. Оно утверждает, что жизнь, какие бы сложные или простые формы она ни принимала, в ее структурном, функциональном и генетическом отношении обеспечивается в конечном итоге только клеткой. Выдающаяся роль клетки как первоисточника жизни обусловливается тем, что она является биологической единицей извлечения из внешней среды, превращения и использования организмами энергии и веществ. В клетке сохраняется и используется биологическая информация.
Согласно второму положению в настоящих условиях новые клетки возникают только путем деления предсуществующих клеток. В обосновании клеточной природы жизни на Земле тезису о единообразии путей возникновения клеток принадлежит особое место. Он был использован М. Шлейденом и Т. Шванном для доказательства гомологии разных типов клеток. Современная биология расширила круг доказательств этому положению. Независимо от индивидуальных морфофункциональных особенностей все клетки одинаковым образом: 1) хранят биологическую информацию; 2) редуплицируют наследственный материал с целью его передачи в ряду поколений; 3) используют информацию для осуществления своих функций на основе синтеза определенных белков-ферментов; 4) хранят и переносят энергию; 5) превращают энергию в работу; 6) регулируют обмен веществ.
Третье положение клеточной теории соотносит клетку с многоклеточными формами, для которых характерен принцип целостности и системной организации. Для системы свойственно наличие новых качеств, благодаря взаимному влиянию и взаимодействию единиц, составляющих эту систему. Структурно-функциональными единицами многоклеточных существ являются клетки. Вместе с тем многоклеточный организм характеризуется рядом специфических свойств, которые нельзя свести к свойствам и качествам отдельной клетки. В третьем положении клеточной теории мы встречаемся с уже знакомой проблемой соотношения части и целого, редукционизма и интегратизма. Системный подход как научное направление используется в биологических исследованиях с начала текущего столетия.
Высокая упорядоченность внутреннего содержимого эукариотической клетки достигается путем компартментализации ее объема — подразделения на «ячейки», отличающиеся деталями химического (ферментного) состава.
megaobuchalka.ru
2.3. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ
РНК-предшественниц,из которых затем образуются более мелкие молекулы зрелых рРНК. В процессе созревания фибриллы преобразуются в рибонуклеопротеиновые зерна (гранулы), которыми представлен зернистый компонент.
Хроматиновые структуры в виде глыбок, рассеянных в нуклеоплазме, являются интерфазной формой существования хромосом клетки (см. 3.5.2.2).
В цитоплазме различают основное вещество (матрикс, гиалоплазма), включения и органеллы.Основное вещество цитоплазмы заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Обычный электронный микроскоп не выявляет в немкакой-либовнутренней организации. Белковый состав гиалоплазмы разнообразен. Важнейшие из белков представлены ферментами гаиколиза, обмена Сахаров, азотистых оснований, аминокислот и липидов. Ряд белков гиалоплазмы служит субъединицами, из которых происходит сборка таких структур, как микротрубочки.
Основное вещество цитоплазмы образует истинную внутреннюю среду клетки, которая объединяет все внутриклеточные структуры и обеспечивает взаимодействие их друг с другом. Выполнение матриксом объединяющей, а также каркасной функции может быть связано с выявляемой с помощью сверхмощного электронного микроскопа микротрабекулярной сети, образованной тонкими фибриллами толщиной 2—3нм и пронизывающей всю цитоплазму. Через гиалоплазму осуществляется значительный объем внутриклеточных перемещений веществ и структур. Основное вещество цитоплазмы следует рассматривать так же, как сложную коллоидную систему, способную переходить из золеобразного (жидкого) состояния в гелеобразное. В процессе таких переходов совершается работа. О функциональном значении таких переходов см. разд. 2.3.8.
Включениями (рис. 2.5) называют относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген), продуктами, подлежащими выведению из клетки (гранулы секрета), балластными веществами (некоторые пигменты).
Органеллы — это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции.
Выделяют органеллы общего значения испециальные. Последние в значительном количестве присутствуют в клетках, специализированных к выполнению определенной функции, но в незначительном количестве могут встречаться и в других типах клеток. К ним относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки, транспортирующие вещества — переносчики нервного возбуждения с одной нервной клетки на другую или клетку рабочего органа, миофибриллы, от которых зависит сокращение мышцы. Детальное рассмотрение специальных органелл входит в задачу курса гистологии.
К органеллам общего значения относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра. В растительных
studfiles.net
Андреев в.П. Принципы структурно-функциональной организации и биология эукариотической клетки
Для всякого живого существа
клетка является последним
морфологическим элементом,
из которого исходит всякая
жизнедеятельность – нормальная,
и патологическая»
Рудольф Вирхов
Содержание лекции
Введение
Организация плазматической мембраны
Функции плазматической мембраны
Управление ионными каналами
Цитозоль
Ядро
Эндоплазматический ретикулум
Цитоскелет
Аппарат Гольджи
Везикулы транспортные
Лизосомы
Пероксисомы
Митохондрии
Протеасомы
Межклеточные взаимодействия
Межклеточные сигнальные вещества
Органоиды клетки
Цитоскелет
Передача сигналов внутри клетки
Передача внешнего сигнала в клетку
Концепции современной цитологии
Основные хронологические вехи в изучении клетки
Введение
Эукариотическая клетка – это элементарная самовоспроизводящая живая система, обладающая способностью к обмену веществом, энергией и информацией с окружающей средой. Клетки существуют в форме самостоятельных одноклеточных организмов (амеба, малярийный плазмодий и др.) или входят в состав тканей многоклеточных организмов. В многоклеточных организмах клетки представляют собой элементы, подчиненные целостному организму. Интеграцию клеток осуществляют гуморальная, нервная и иммунная система. Для адекватной реакции на регуляторные сигналы «центра» каждая клетка обладает системой управления. Согласно взглядам В.Я. Александрова и М.Д. Голубовского клетка владеет разными видами целесообразного поведения, что у человека называется эрудиция и ум. Клеточная «эрудиция» – хранение большого набора генетических программ; клеточный «ум» – способность включать в определенных условиях соответствующую программу поведения. Например, в ответ на стресс клетки генерируют изменение своих компонентов и приступают к поиску адаптивных генетических программ поведения.
Организм человека построен приблизительно из десяти триллионов клеток, которые формируют четыре основных типа тканей: эпителиальную, соединительную, мышечную и нервную. Каждая ткань представлена многочисленными клеточными типами. К настоящему времени изучена структура и функция более 200 фенотипов клеток, приспособленных для выполнения многочисленных функций. (Открытие клеточных типов продолжается; так несколько лет назад в сетчатке глаза ученые обнаружили третий тип клеток, участвующих в формировании ритма сон-бодрствование).
В эукариотической клетке, как элементарной целостной живой системе, можно выделить четыре общебиологические эволюционно обусловленные подсистемы. К ним относятся: пограничная, представленная внешней клеточной мембраной-плазмалеммой, скелетно-двигательная (цитоскелет и сократительный аппарат), гомеостатическая (ядро и комплекс органелл) и подсистема, обеспечивающая реактивные свойства за счет рецепторно-транспортных механизмов.
Организация эукариотической клетки
Эукариотическая клетка организована системой мембран. Снаружи она ограничена плазматической мембраной. Внутренний объем клетки заполнен цитоплазмой, которая разделена на хорошо различимые, окруженные внутриклеточными мембранами отделы, называемыми мембранными органеллами. В них сосредоточено около 50% объема клетки. Остальное внутриклеточное пространство занимает компартмент, называемый цитозолем ( син. матрикс цитоплазмы, гиалоплазма).
Самой крупной органеллой является ядро клетки, его можно легко видеть в световой микроскоп. В нем находятся хромосомы, и осуществляется ДНК-направляемый синтез РНК (транскрипция). Перинуклеарное пространство (лат. nucleus – ядро), ограниченное наружной и внутренней мембранами, связано с эндоплазматической сетью, представляющей собой ограниченную мембранами систему связанных друг с другом канальцами полостей в виде уплощенных мешочков и цистерн. Другая, ограниченная мембранами органелла, также представляющая собой систему мембран, – комплекс Гольджи напоминает сложенные в стопку расширенные по краям уплощенные цистерны, в которых происходит модификация и сортировка белков и липидов. Митохондрии отвечают за производство энергии в виде аденозинтрифосфата (АТФ). Лизосомы и пероксисомы – маленькие глобулярные органеллы, предназначенные для выполнения специфических функций. Экзосомы и эндосомы – пузыреобразные органеллы (везикулы), участвующие в процессе обмена веществ между клеткой и ее окружением. Кроме «мембранных», в клетке находятся и немембранные органоиды – рибосомы, клеточный центр, реснички, жгутики и др. Регуляторные, а также состарившиеся, содержащие ошибки транскрипции белки разрушаются в протеасомах. Способность клетки изменять свою форму, перемещаться, передвигать органеллы в цитоплазме и разделять хромосомы во время митоза обеспечивает система белковых нитевидных структур – микрофиламентов, актиновых филаментов, микротрубочек, – напоминающая строительные леса (так называемый цитоскелет).
Состав биомембран
Все биомембраны, несмотря на огромное разнообразие, построены однотипно; они состоят из двух слоев полярных липидных молекул, в которые встроены белки. В клеточной мембране присутствуют липиды трех главных типов – фосфолипиды, холестерол и гликолипиды. Все они являются амфипатическими, то есть обладающими двумя противоположными свойствами. Каждая молекула мембранного липида имеет заряженную гидрофильную «головку» и два незаряженных гидрофобных «хвоста». Каждый «хвост» представляет собой длинную углеводородную цепь жирной кислоты, причем у фосфолипидов одна из этих цепей – предельная, т.е. не содержит двойных связей, а вторая – непредельная имеет одну или более двойных связей. Неполярные, гидрофобные участки фосфолипидных молекул, образуемые цепями жирных кислот, обращены внутрь липидного бислоя, формируя сплошную пленку – своеобразный барьер для растворенных в воде ионов и полярных молекул. Полярные гидрофильные группы фосфолипидных молекул обращены наружу и как бы смачиваются окружающим водным раствором. Структуру такого типа называют жидкокристаллической (рис. 1).
У разных клеток состав мембранных липидов может существенно различаться. Различный липидный состав характерен и для разных мембран одной и той же клетки. Они могут включать такие липиды как фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, кардиолипин, триглицериды, холестерол и др., от которых в основном зависят физико-химические свойства мембран. Наиболее распространенными мембранными липидами являются производные глицерин-3-фосфата, называемые глицерофосфолипидами, в которых две гидроксильные группы глицерола замещены остатками различных или одинаковых жирных кислот. Некоторые мембраны содержат, кроме того, сахарные остатки, связанные с липидами и белками ковалентными связями. Такими углеводными остатками могут быть глюкоза, галактоза, N-ацетилгалактозамин и др.
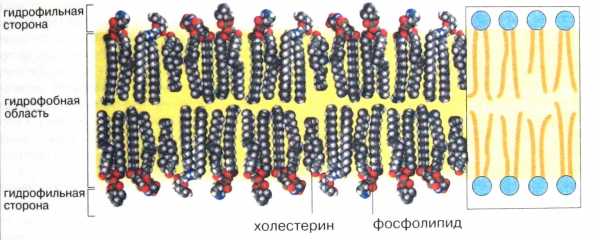
Рис.1. Мембранные липиды. (Из кн. Кольман Я., Рём К, – Г. «Наглядная биохимия»)
Жидкий кристалл – это как бы промежуточное состояние вещества между твердым кристаллом и аморфной жидкостью: в расположении образующих его молекул есть порядок, обусловленный правильной ориентацией молекул друг относительно друга. Для образования жидких кристаллов необходимо, чтобы молекулы были длинными и узкими. Такими молекулами в плазматической мембране являются жирные кислоты.
Соотношение этих компонентов является характерным для клетки или мембраны и существенно варьирует в зависимости от типа клеток или мембран. В мембранах эритроцитов имеется производное от многоатомного аминоспирта сфингозина – церамид (рис.2). К этому липиду ферменты присоединяют разветвленные цепочки различных сахаров. Такие гликолипиды называют ганглиозидами. В настоящее время идентифицирована структура более 60-ти молекул ганглиозидов. В нервных клетках ганглиев они составляют около 6 % общей массы липидов; в других типах клеток они встречаются в меньших количествах.
Все ганглиозиды выполняют важные функции в тех или иных клетках, являясь рецепторами для медиаторов, осуществляющих связь между клетками. Ганглиозиды в эритроцитах обладают антигенными свойствами и определяют группы крови людей 0, А, В-системы. Следует отметить, что молекулы ганглиозидов нередко являются «посадочными площадками» для вирусов или бактериальных токсинов, то есть выполняют несвойственные им в норме рецепторные функции. Так, холерный токсин проникает внутрь эпителиальных клеток кишечника, присоединяясь к ганглиозиду Gм1.
Плазматическая мембрана постоянно обновляется. Это происходит за счет процессов синтеза и разрушения отслуживших окисленных гликолипидов.
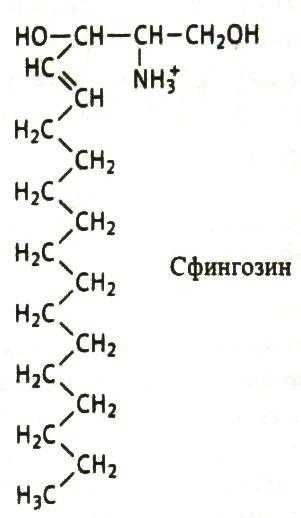
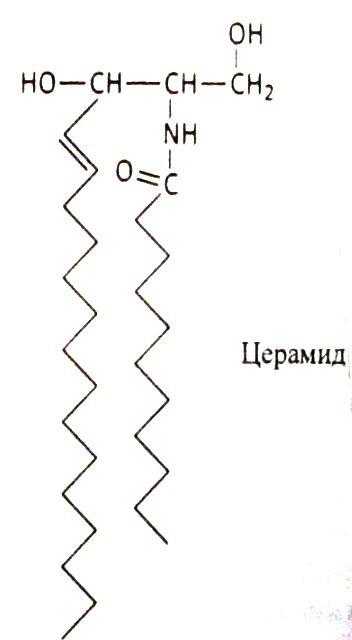
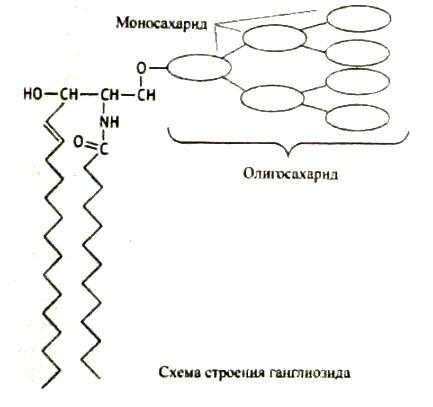
Рис. 2. Структура некоторых компонентов плазматической мембраны (из кн. В. Элиот, Д. Элиот «Биохимия и молекулярная биология».
Липиды мембран синтезируются на эндоплазматической сети и переносятся в плазматическую мембрану. Процесс переноса осуществляется транспортными пузырьками. При некоторых наследственных заболевания человека ферменты, необходимые для разрушения этих гликолипидов, оказываются дефектными, и клетка не может расщеплять гликолипиды. Это приводит к их накоплению в клетке, и, в конце концов, к гибели клетки. Заболевания, вызванные неспособностью клетки к расщеплению сложных гликолипидов, называются мукополисахаридозами.
Текучесть мембран зависит от липидного состава и температуры окружающей среды. Существенное значение для структуры клеточных мембран имеют полиненасыщенные (с двойными связями между углеродными атомами) жирные кислоты (ПНЖК). В промежутках между изгибами ПНЖК располагается спирт холестерол (холестерин). Он предотвращает кристаллизацию мембраны или переход в гель, придавая им текучесть. Однако полиненасыщенные жирные кислоты не синтезируются в организме человека и высших животных, поэтому их поступление с пищей необходимо для правильной деятельности клеточных мембран. Отсутствие ПНЖК приводит к тяжелым общим расстройствам, провалам памяти, выпадению волос, шелушению кожи. (Источниками ПНЖК являются растительные масла, не подвегнувшиеся термической обработке и рыба холодных морей).
Специфические функции биологических мембран осуществляются главным образом белками. Типы белков и их количества в мембране отражают ее функцию. Так в плазматической мембране белки составляют приблизительно 50% от ее массы, а в мембране митохондрии – 75%. По своей функциональной роли мембранные белки разделяются на ферментативные, транспортные и регуляторные. Но такое разделение носит только условный характер, поскольку, например, ионный канал может одновременно участвовать во всех этих функциях.
Большинство мембранных белков являются интегральными. Эти протеины или погружены в толщу липидного слоя, или пронизывают мембрану насквозь (трансмембранные белки). Они удерживаются в билипидном слое за счет нековалентных связей. Их гидрофильные аминокислоты взаимодействуют с фосфатными группами фосфолипидов, а гидрофобные – с цепями жирных кислот. Между белками и жирными кислотами могут возникать ковалентные связи. Для интегральных белков некоторых клеток характерна латеральная подвижность; они могут перераспределяться в мембранах в результате взаимодействия с периферическими белками, элементами цитоскелета, молекулами в мембране соседней клетки и компонентами внеклеточного матрикса. Кроме того, выделяют так называемые периферические белки, молекулы которых не встроены в мембрану, а за счет слабых взаимодействий удерживаются на ее поверхности. Периферические мембранные белки (фибриллярные, глобулярные) находятся как на наружной, так и внутренней поверхности мембраны и нековалентно – за счет водородных, гидрофобных, электростатических взаимодействий – связаны с интегральными мембранными белками. К периферическим белкам внутренней поверхности мембраны относятся белки цитоскелета (например, анкирин от лат. аnkir – якорь), которые обеспечивают механическую связь мембран с цитоскелетом (микрофиламентами и микротрубочками), семейство G-белков (гуанозинтрифосфатаз), посредством которых активируется фермент аденилатциклаза и мн. др.
Примером периферического белка наружной поверхности мембраны является фибронектин. Этот гликопротеин локализован на наружной поверхности мембраны почти всех клеток. Фибронектин обеспечивает прикрепление клеток к другим клеткам, а также способствует соединению клетки с внеклеточным матриксом. Отмечено резкое снижение содержания фибронектина в мембранах опухолевых клеток; это придает им большую подвижность и способствует метастазированию. Белки и некоторые липидные молекулы, расположенные на наружной стороне плазматической мембраны, несут ковалентно связанные углеводные компоненты – олигосахариды (рис3).
Эти гликопротеины и гликолипиды вместе с дополнительными несвязанными с мембраной гликопротеинами и полисахаридами образуют клеточную оболочку – гликокаликс. Гликокаликс, покрывающий микроворсинки каемчатых клеток эпителия кишечника, содержит ферменты, завершающие расщепление белков, углеводов (пристеночное пищеварение). Однако основные функции гликокаликса – межклеточное узнавание и межклеточные взаимодействия.
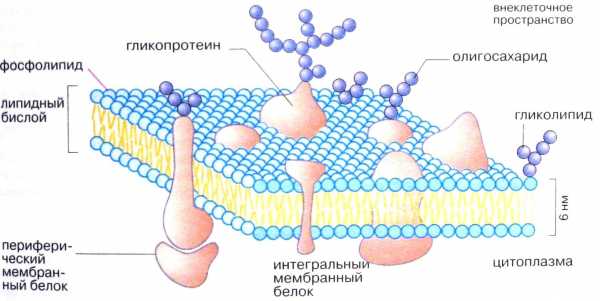
Рис.3. Структура плазматической мемраны. (Из кн. Кольман Я., Рём К, – Г. «Наглядная биохимия»)
Углеводные остатки гликопротеинов и гликолипидов наружной мембраны клетки обладают специфическими антигенными свойствами. Все клетки одного организма несут сходные поверхностные антигены, которые отличаются от поверхностных антигенов клеток любого другого организма. Это свойство используется иммунной системой для разделения всех клеток «на свои» и «чужие». Успех переливания крови или трансплантации другой ткани во многом определяют антигены групп крови 0, А, В-системы. Их антигенные свойства обусловлены структурой углеводных остатков гликолипидов на поверхности эритроцитов. (Схема строения ганглиозидов, определяющих IV, III, II и I-ю группы крови 0, А, В-системы представлена на рис. 4-7.)
Кроме этих антигенов индивидуальную поверхность клеток определяют так называемые трансплантационные антигены или антигены гистосовместимости. В этом случае антигенами служат полипептидные цепи группы трансмембранных белков. Эти белки-антигены кодируются в геноме млекопитающих многими генами, так называемого главного комплекса гистосовместимости. Бóльшая часть Т-лимфоцитов узнает чужеродные антигены только в том случае, если эти антигены ассоциированы на клеточных поверхностях с антигенами главного комплекса гистосовместимости.
Антигéны главного комплекса гистосовместимости (ГКГ) или антигены МНС (англ.majorhistocompatibilitycomplex, произносят как эм, эйч, си) – семейство антигенов, определяющих отторжение чужеродного трансплантата, т.е. тканевую несовместимость. У человека они впервые были выявлены в лейкоцитах и поэтому получили названиеHLA(англ.humanleucocyteantigen, произносят как эйч, эль, эй). Существует два основных класса антигенов (молекул) ГКГ: антигены ГКГ классаIи антигены ГКГ классаII. Все они являются мембранными гликопротеинами (рис. 8).
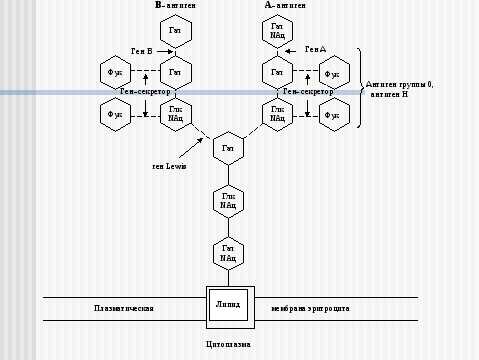
Рис. 4. Схема связи остатков сахаров с наружной мембраной эритроцита у людей с IV (АВ) группой крови 0, А, В-системы.
Глк – D-глюкоза, Гал – D-галактоза, Фук – L-фукоза, Глк NАц – N-ацетил- D-глюкозамин, Гал NАц – N—aцетил- D-галактозамин.
Прерывистые связи между остатками сахаров означают, что существует много типов соединений.
Ген Lewis, доминантный ген-секретор, ген-А, ген-В кодируют ферменты-трансферазы, переносящие и присоединяющие остатки сахаров. При наличии гена А или В, и рецессивного гена-секретора в гомозиготном состоянии антигены А и В не образуются (бомбейский феномен).
Примечание: Отличительной особенностью антигена Н является наличие его в биологических жидкостях секреторов групповых веществ и отсутствие – у несекреторов. Антиген 0, в отличие от антигена Н, А и В, с секретами (слюна, сперма) не выделяется.
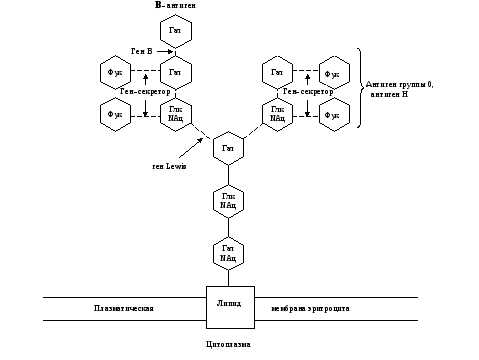
Рис. 5. Схема связи остатков сахаров с наружной мембраной эритроцита у людей с III (В) группой крови 0, А, В-системы.
Глк – D-глюкоза, Гал – D-галактоза, Фук – L-фукоза, Глк NАц – N-ацетил— D-глюкозамин, Гал NАц – N-aцетил— D-галактозамин.
Прерывистые связи между остатками сахаров означают, что существует много типов соединений.
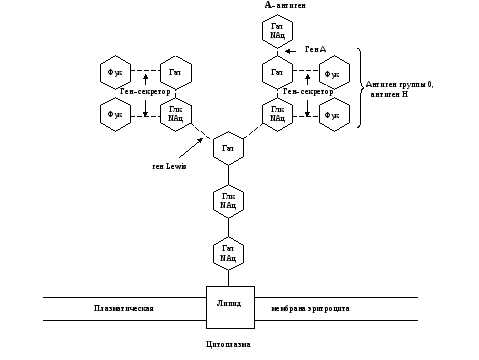
Рис. 6. Схема связи остатков сахаров с наружной мембраной эритроцита у людей с II (А) группой крови 0, А, В-системы.
Глк – D-глюкоза, Гал – D-галактоза, Фук – L-фукоза, Глк NАц – N-ацетил- D-глюкозамин, Гал NАц – N—aцетил- D-галактозамин.
Прерывистые связи между остатками сахаров означают, что существует много типов соединений.
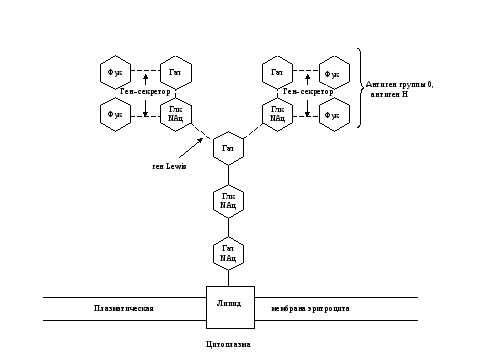
Рис. 7. Схема связи остатков сахаров с наружной мембраной эритроцита у людей с I (0) группой крови 0, А, В-системы.
Глк – D-глюкоза, Гал – D-галактоза, Фук – L-фукоза, Глк NАц – N-ацетил- D-глюкозамин, Гал NАц – N—aцетил- D-галактозамин.
Прерывистые связи между остатками сахаров означают, что существует много типов соединений.
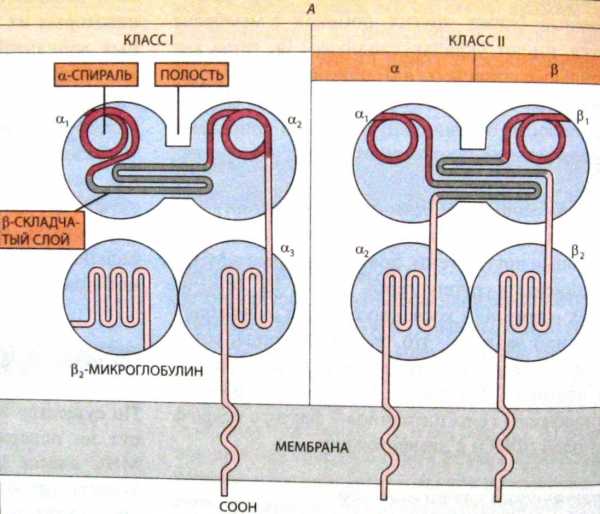
Рис. 8. Схема структуры трансмембранных гликопротеинов МНС класс 1 (слева) и МНС класса 2 (справа). (Из кн. Б. Албертс и др. «Молекулярная биология клетки», том 5.)
Класс IГКГ содержится на оболочках всех соматических клеток, обладающих ядром. Они презентируют вирусные антигены цитотоксическим Т-лимфоцитам. ГКГ классаIIсвязаны с плазматической мембраной В-лимфоцитов, макрофагов, дендритных и некоторых других антигенпредставляющих (презентирующих) клеток, которые представляют в иммунных ответах для распознавания Т-лимфоцитам переработанные ими до пептидов чужеродные антигены, вслед, за чем запускается развитие иммунных защитных реакций.
Функции мембран
Ограничение и обособление клеток и органелл. Обособление клеток от межклеточной среды обеспечивается плазматической мембраной, защищающей клетки от механического и химического воздействий.
Контролируемый транспорт метаболитов и ионов через поры и посредством переносчиков определяет внутреннюю среду, что существенно для гомеостаза, т.е. поддержания постоянной концентрации метаболитов и неорганических ионов, и других физиологических параметров.
Восприятие внеклеточных сигналов и их передача внутрь клетки, а также инициация сигналов.
Ферментативный катализ. В мембранах локализованы наиболее важные реакции энергетического обмена, такие, как окислительное фосфорилирование.
Контактное взаимодействие с межклеточным матриксом и взаимодействие с другими клетками при образовании тканей.
Заякоривание цитоскелета, обеспечивающее поддержание формы клеток и органелл и клеточной подвижности.
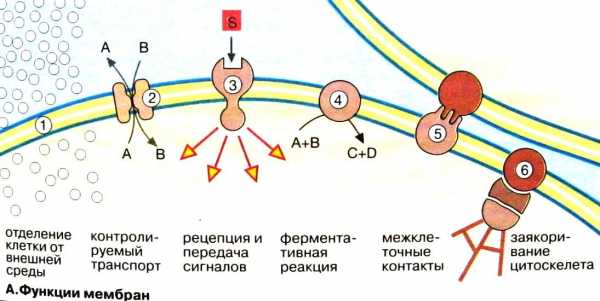
Основная функция наружной клеточной мембраны – сохранение внутренней среды клетки. При участии плазматической мембраны происходит узнавание и агрегация, как соседних клеток, так и клеток с компонентами внеклеточного матрикса. Формирование клеточной поверхности, которая способна к узнаванию других клеток, контакту с ними и восприятию разнообразных раздражителей, воздействующих на клетку, обеспечивается, прежде всего, углеводными группами, входящими в состав гликопротеидов клеточной мембраны.
studfiles.net
Основные положения клеточной теории
Клетки открыты в 1665 г. Р. Гуком. Клеточная теория, одно из величайших открытий 19-го века, была сформулирована в 1838 г. немецкими учёными М. Шлейденом и Т. Шванном, а в дальнейшем развита и дополнена Р. Вирховым. Клеточная теория включает в себя следующие положения:
1.Клетка является наименьшей единицей живого.
2.Клетки разных организмов имеют сходное строение, что свидетельствует о единстве живой природы.
3.Размножение клеток происходит путём деления исходной, материнской клетки (постулат: каждая клетка — из клетки).
4.Многоклеточные организмы состоят из сложных ансамблей клеток и их производных, объединённых в системы тканей и органов, а последние — в целостный организм с помощью нервных, гуморальных и иммунных механизмов регуляции.
Клеточная теория объединила представления о клетке как наименьшей структурной, генетической и функциональной единице животных и растительных организмов. Она вооружила биологию и медицину пониманием общих закономерностей строения живого.
Меры длины, применяемые в цитологии
1 мкм (микрометр) – 10–3 мм (10–6 м)
1 нм (нанометр) – 10–3 η (10–9 м)
1 A (амстрем) – 0,1 нм (10–10 м)
Общая организация животных клеток
Все клетки организма человека и животных имеют общий план строения. Они состоят из цитоплазмы и ядра и отделены от окружающей среды клеточной оболочкой.
Организм человека состоит примерно из 1013 клеток, подразделяющихся более чем на 200 типов. В зависимости от своей функциональной специализации, различные клетки организма могут значительно отличаться по своей форме, величине и внутреннему устройству. В организме человека встречаются круглые (клетки крови), плоские, кубические, призматические (эпителиальные), веретеновидные (мышечные), отростчатые (нервные) клетки. Их размеры колеблются от 4-5 мкм (клетки-зёрна мозжечка и малые лимфоциты) до 250 мкм (яйцеклетка). Отростки некоторых нервных клеток имеют длину более 1 метра (у нейронов спинного мозга, отростки которых идут до кончиков пальцев конечностей). При этом форма, величина и внутреннее строение клеток всегда наилучшим образом соответствуют выполняемым ими функциям.
Структурные компоненты клетки
Цитоплазма – часть клетки, отделённая от окружающей среды клеточной оболочкой и включающая в себя гиалоплазму, органеллы и включения.
Все мембраны в клетках имеют общий план строения, который обобщён в понятии универсальная биологическая мембрана (рис. 2- 1А).
У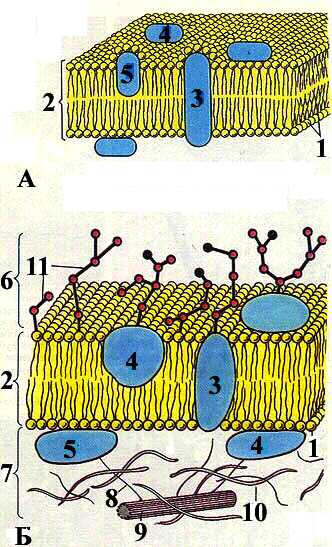 ниверсальная
биологическая мембрана
образована двойным слоем молекул
фосфолипидов общей толщиной 6 мкм. При
этом гидрофобные хвосты молекул
фосфолипидов обращены внутрь, навстречу
друг другу, а полярные гидрофильные
головки обращены наружу мембраны,
навстречу воде. Липиды обеспечивают
основные физико-химические свойства
мембран, в частности, их текучесть
при температуре тела. В этот двойной
слой липидов встроены белки. Их
подразделяют на интегральные
(пронизывают весь бислой липидов),
полуинтегральные
(проникают до половины липидного
бислоя), или поверностные (располагаются
на внутренней или наружной поверхности
липидного бислоя).
ниверсальная
биологическая мембрана
образована двойным слоем молекул
фосфолипидов общей толщиной 6 мкм. При
этом гидрофобные хвосты молекул
фосфолипидов обращены внутрь, навстречу
друг другу, а полярные гидрофильные
головки обращены наружу мембраны,
навстречу воде. Липиды обеспечивают
основные физико-химические свойства
мембран, в частности, их текучесть
при температуре тела. В этот двойной
слой липидов встроены белки. Их
подразделяют на интегральные
(пронизывают весь бислой липидов),
полуинтегральные
(проникают до половины липидного
бислоя), или поверностные (располагаются
на внутренней или наружной поверхности
липидного бислоя).
Рис. 2-1. Строение биологической мембраны (А) и клеточной оболочки (Б).
1. Молекула липида.
2. Бислой липидов.
3. Интегральные белки.
4. Полуинтегральные белки.
5. Периферические белки.
6. Гликокаликс.
7. Подмембранный слой.
8. Микрофиламенты.
9. Микротрубочки.
10. Микрофибриллы.
11. Молекулы гликопротеинов и гликолипидов.
(По О. В. Волковой, Ю. К. Елецкому).
При этом белковые молекулы располагаются в липидном бислое мозаично и могут «плавать» в «липидном море» наподобие айсбергов, благодаря текучести мембран. По своей функции эти белки могут быть структурными (поддерживать определённую структуру мембраны), рецепторными (образовывать рецепторы биологически активных веществ), транспортными (осуществляют транспорт веществ через мембрану) и ферментными (катализируют определённые химические реакции). Эта наиболее признанная в настоящее время жидкостно-мозаичная модель биологической мембраны была предложена в 1972 г. Singer и Nikolson.
Мембраны выполняют в клетке разграничительную функцию. Они разделяют клетку на отсеки, компартменты, в которых процессы и химические реакции могут идти независимо друг от друга. Например, агрессивные гидролитические ферменты лизосом, способные расщеплять большинство органических молекул, отделены от остальной цитоплазмы с помощью мемраны. В случае её разрушения происходит самопереваривание и гибель клетки.
Имея общий план строения, разные биологические мембраны клетки различаются по своему химическому составу, организации и свойствам, в зависимости от функций структур, которые они образуют.
studfiles.net
Что такое цитоплазма? Структура, состав и свойства цитоплазмы
Что такое цитоплазма? Каково ее строение и состав? Какие функции она выполняет? В этой статье мы подробно ответим на все эти вопросы. Кроме того, мы рассмотрим структурные особенности цитоплазмы и ее свойства, а также поговорим о делении коллоидного раствора, строении клеточных мембран и важнейших клеточных органоидах.
Структурные единицы всех тканей и органов клетки. Два типа их структурной организации
Известно, что клетки образуют ткани всех растений и животных. Эти структурные единицы всего живого могут различаться по форме, размерам и даже по внутреннему строению. Но в то же время они имеют схожие принципы в процессах жизнедеятельности, в том числе в обмене веществ, росте и развитии, раздражимости и изменчивости. Самые простейшие формы жизни состоят из единственной клетки и размножаются делением.
Учеными было выделено два типа организации клеточной структуры:
- прокариотический;
- эукариотический.
Они имеют множество различий в своем строении. В прокариотической клетке структурно оформленное ядро отсутствует. Ее единственная хромосома находится непосредственно в цитоплазме, то есть никак не отделяется от других элементов. Такое строение свойственно бактериям. Их цитоплазма бедна по составу структур, но в ней имеются мелкие рибосомы. Эукариотическая устроена гораздо сложнее прокариотической клетки. Ее ДНК, связанная с белком, находится в хромосомах, располагающихся в обособленном клеточном органоиде — ядре. Оно отделяется от других органоидов клетки пористой мембраной и состоит из таких элементов как: хроматин, ядерный сок и ядрышко. Тем не менее есть и нечто общее у двух типов клеточной организации. И прокариоты, и эукариоты имеют оболочку. А их внутреннее содержимое представлено особым коллоидным раствором, в котором находятся различные органоиды и временные включения.
Эукариотическая клетка: цитоплазма. Ее состав и функции
Итак, переходим к сути нашего исследования. Что такое цитоплазма? Давайте рассмотрим более подробно это клеточное образование. Цитоплазма представляет собой архиважную составляющую клетки, располагающуюся между ядром и плазматической мембраной. Полужидкая, она пронизана канальцами, микротрубочками, микрофиламентами и филаментами. Также под цитоплазмой можно понимать коллоидный раствор, который характеризуется движением коллоидных частиц и прочих компонентов. В этой полужидкой среде, состоящей из воды, различных органических и неорганических соединений, располагаются клеточные структуры-органоиды, а также временные включения. Важнейшие функции цитоплазмы таковы. Она осуществляет оформление всех клеточных компонентов в единую систему. Благодаря наличию канальцев и микротрубочек цитоплазма выполняет функцию клеточного скелета и предоставляет среду для осуществления физиологических и биохимических процессов. Кроме этого, она дает возможность для функционирования всех клеточных органоидов и обеспечивает передвижение. Эти функции клетки цитоплазмы чрезвычайно важны, так как позволяют структурной единице всего живого осуществлять свою нормальную жизнедеятельность. Теперь вы знаете, что такое цитоплазма. А также осведомлены о том, какое положение в клетке она занимает и какую «работу» выполняет. Далее мы рассмотрим состав и структуру коллоидного раствора более подробно.
Есть ли отличия в цитоплазме растительной и животной клеток?
Мембранными органоидами, находящимися в коллоидном растворе, считаются аппарат Гольджи, эндоплазматическая сеть, митохондрии, лизосомы, пластиды и наружная цитоплазматическая мембрана. В клетках животных и растений состав полужидкой среды отличается. Цитоплазма в растительной клетке имеет специальные органоиды – пластиды. Они представляют собой специфичные белковые тельца, которые различаются по функциям, форме и окрашиваются пигментами в разные цвета. Пластиды располагаются в цитоплазме и способны передвигаться вместе с ней. Они растут, размножаются и вырабатывают органические соединения, содержащие ферменты. Цитоплазма в растительной клетке имеет три вида пластид. Желтоватые или оранжевые называются хромопластами, зеленые – хлоропластами, а бесцветные — лейкопластами. Есть и еще одна характерная особенность — комплекс Гольджи представлен диктиосомами, рассеянными по цитоплазме. В клетках животных, в отличие от растительных, имеется два слоя цитоплазмы. Наружный называется эктоплазма, а внутренний — эндоплазма. Первый слой прилегает к клеточной мембране, а второй — находится между ним и пористой ядерной мембраной. Эктоплазма имеет в своем составе большое количество микрофиламента — нитей из молекул глобулярного белка актина. Эндоплазма содержит различные органоиды, гранулы и характеризуется меньшей вязкостью.

Гиалоплазма в эукариотической клетке
Основу цитоплазмы эукариотов составляет так называемая гиалоплазма. Она представляет собой слизистый, бесцветный, неоднородный раствор, в котором постоянно протекают процессы обмена веществ. Гиалоплазма (иными словами матрикс) это коллоидная система со сложным строением. В ее состав включаются растворимые РНК и белки, липиды и полисахариды. Еще в гиалоплазме содержится значительное количество нуклеотидов, аминокислот, а также ионов неорганических соединений типа Na— или Са2+.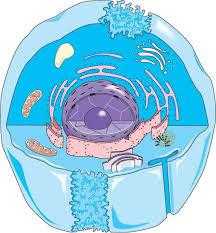 Матрикс не имеет гомогенной структуры. Он представлен в двух формах, которые называются гель (твердая) и золь (жидкая). Между ними происходят взаимопереходы. В жидкой фазе имеется система тончайших белковых нитей, которые называются микротрабекулами. Они связывают все структуры внутри клетки. А в местах их пересечения находятся группы рибосом. Микротрабекулы вместе с микротрубочками и микрофиламентами формируют цитоплазматический скелет. Он определяет и упорядочивает местоположение всех клеточных органелл.
Матрикс не имеет гомогенной структуры. Он представлен в двух формах, которые называются гель (твердая) и золь (жидкая). Между ними происходят взаимопереходы. В жидкой фазе имеется система тончайших белковых нитей, которые называются микротрабекулами. Они связывают все структуры внутри клетки. А в местах их пересечения находятся группы рибосом. Микротрабекулы вместе с микротрубочками и микрофиламентами формируют цитоплазматический скелет. Он определяет и упорядочивает местоположение всех клеточных органелл.
Органические и неорганические вещества в коллоидном растворе клетки
Давайте рассмотрим, каков же химический состав цитоплазмы? Вещества, содержащиеся в клетке, можно классифицировать на две группы — органические и неорганические. Первая представлена белками, углеводами, жирами и нуклеиновыми кислотами. Углеводы в цитоплазме представлены моно-, ди- и полисахаридами. К моносахаридам, бесцветным кристаллическим веществам, обычно сладковатым на вкус, относят фруктозу, глюкозу, рибозу и т. д. Крупные молекулы полисахаридов состоят из моносахаридов. В клетке они представлены крахмалом, гликогеном и целлюлозой. Липиды, то есть молекулы жиров, образуются остатками глицерина и жирных кислот. Структура цитоплазмы: неорганические вещества представлены в первую очередь водой, которая, как правило, составляет до 90% массы. Она выполняет в цитоплазме важные функции.  Вода является универсальным растворителем, придает упругость, принимает непосредственное участие в перемещении веществ как внутри, так и между клетками. Что касается макроэлементов, формирующих основу биополимеров, то более 98% всего состава цитоплазмы занимают кислород, водород, углерод и азот. Кроме них в клетке содержатся натрий, кальций, сера, магний, хлор и др. Минеральные соли находятся в виде анионов и катионов, при этом их соотношение определяет кислотность среды.
Вода является универсальным растворителем, придает упругость, принимает непосредственное участие в перемещении веществ как внутри, так и между клетками. Что касается макроэлементов, формирующих основу биополимеров, то более 98% всего состава цитоплазмы занимают кислород, водород, углерод и азот. Кроме них в клетке содержатся натрий, кальций, сера, магний, хлор и др. Минеральные соли находятся в виде анионов и катионов, при этом их соотношение определяет кислотность среды.
Свойства коллоидного раствора в клетке
Рассмотрим далее, каковы основные свойства цитоплазмы. Во-первых, это постоянный циклоз. Он представляет собой внутриклеточное движение цитоплазмы. Впервые оно было зафиксировано и описано в 18-м столетии итальянским ученым Корти. Циклоз осуществляется во всей протоплазме, в том числе и в тяжах, связывающих цитоплазму с ядром. Если движение по каким-либо причинам прекращается — погибает эукариотическая клетка. Цитоплазма обязательно находится в постоянном циклозе, который обнаруживается по перемещению органоидов. Скорость движения матрикса зависит от различных факторов, в том числе от света и температуры. К примеру, в эпидермисе чешуи лука скорость циклоза составляет около 6 м/с. Движение цитоплазмы в растительном организме оказывает огромное влияние на его рост и развитие, способствуя транспорту веществ между клетками. Вторым важным свойством является вязкость коллоидного раствора. Она сильно варьируется в зависимости от вида организма. У некоторых живых существ вязкость цитоплазмы может совсем незначительно превышать вязкость воды, у других, наоборот, достигать вязкости глицерина. Считается, что она зависит от обмена веществ. Чем интенсивнее происходит обмен, тем ниже становится вязкость коллоидного раствора. 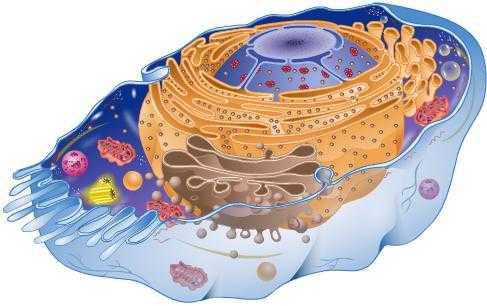 Еще одним немаловажным свойством является полупроницаемость. Цитоплазма в своем составе имеет пограничные мембраны. Они, благодаря особому своему строению, имеют возможность избирательно пропускать молекулы одних веществ и не пропускать других. Избирательная проницаемость цитоплазмы играет важнейшую роль в процессе жизнедеятельности. Она не постоянна в течение жизни, меняется с возрастом и увеличивается у растительных организмов при повышении интенсивности освещения и температуры. Сложно переоценить значение цитоплазмы. Она участвует в энергетическом обмене, транспорте питательных веществ, выведении экзотоксинов. Также матрикс считается осмотическим барьером и участвует в регуляции процессов развития, роста и клеточного деления. В том числе цитоплазма играет большую роль при репликации ДНК.
Еще одним немаловажным свойством является полупроницаемость. Цитоплазма в своем составе имеет пограничные мембраны. Они, благодаря особому своему строению, имеют возможность избирательно пропускать молекулы одних веществ и не пропускать других. Избирательная проницаемость цитоплазмы играет важнейшую роль в процессе жизнедеятельности. Она не постоянна в течение жизни, меняется с возрастом и увеличивается у растительных организмов при повышении интенсивности освещения и температуры. Сложно переоценить значение цитоплазмы. Она участвует в энергетическом обмене, транспорте питательных веществ, выведении экзотоксинов. Также матрикс считается осмотическим барьером и участвует в регуляции процессов развития, роста и клеточного деления. В том числе цитоплазма играет большую роль при репликации ДНК.
Особенности клеточного размножения
Все растительные и животные клетки размножаются делением. Известно три вида — непрямое, прямое и редукционное. Первый иначе называется амитоз. Непрямое размножение происходит следующим образом. Первоначально «перешнуровывается» ядро, а затем происходит деление цитоплазмы. В итоге формируются две клетки, которые постепенно вырастают до размеров материнской. Такой вид деления у животных встречается крайне редко. Как правило, у них происходит непрямое деление, то есть митоз. Оно значительно сложнее амитоза и характеризуется тем, что происходит усиление синтеза в ядре и удвоение количества ДНК. Митоз имеет четыре фазы, которые называются — профаза, метафаза, анафаза и телофаза.
- Первая фаза характеризуется формированием клубка хроматиновых нитей на месте ядра, а впоследствии хромосом в виде «шпилек». В этот период происходит расхождение центриолей к полюсам и формирование ахроматинового веретена деления.
- Второй этап митоза отличается тем, что хромосомы, достигая максимальной спирализации, начинают располагаться на экваторе клетки упорядоченно.
- В третьей фазе происходит расщепление хромосомы на две хроматиды. При этом нити веретена сокращаются и оттягивают дочерние хромосомы к противоположным полюсам.
- В четвертой фазе митоза происходит диспирализация хромосом, а также формирование вокруг них ядерной оболочки. Одновременно происходит деление цитоплазмы. У дочерних клеток имеется диплоидный набор хромосом.
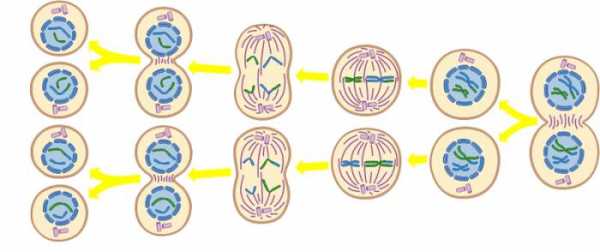
Редукционное деление свойственно исключительно половым клеткам. При таком типе клеточного размножения происходит формирование из хромосом парных образований. Исключение составляет одна непарная хромосома. В результате редукционного деления в двух дочерних клетках получается половинный хромосомный набор. Непарная находится лишь в одной дочерней клетке. Половые клетки, имеющие половинный набор хромосом, созревшие и способные к оплодотворению, называются женской и мужской гаметами.
Понятие цитоплазматической мембраны
У всех клеток животных, растений и даже у простейших бактерий есть особый поверхностный аппарат, который ограничивает и защищает матрикс от внешней среды. Цитоплазматическая мембрана (плазмалемма, клеточная мембрана, плазматическая мембрана) представляет собой избирательно проницаемый слой молекул (протеины, фосфолипиды), который охватывает цитоплазму. Он включает три подсистемы:
- плазматическую мембрану;
- надмембранный комплекс;
- субмембранный опорно-сократительный аппарат гиалоплазмы.
Строение мембраны цитоплазмы таково: она содержит два слоя молекул липидов (бислой), при этом каждая такая молекула имеет хвост и головку. Хвосты обращены друг к другу. Они гидрофобны. Головки гидрофильны и обращены внутрь и наружу клетки. В бислой включены молекулы белка. Причем он асимметричен, а в монослоях располагаются разные липиды. Например, в эукариотической клетке молекулы холестерина находятся во внутренней, прилегающей к цитоплазме, половине мембраны. Гликолипиды располагаются исключительно в наружном слое, причем их углеводные цепи всегда направлены наружу. Цитоплазматическая мембрана выполняет важнейшие функции, в том числе ограничивает внутреннее содержимое клетки от внешней среды, позволяет проникать определенным веществам (глюкозе, аминокислотам) внутрь клетки. Плазмалемма осуществляет перенос веществ внутрь клетки, а также их вывод наружу, то есть выделение. Через поры проникают вода, ионы и мелкие молекулы веществ, а крупные твердые частицы транспортируются в клетку при помощи фагоцитоза. На поверхности мембрана образует микроворсинки, впячивания и выпячивания, что позволяет не только эффективно всасывать и выделять вещества, но и соединяться с другими клетками. Мембрана предоставляет возможность прикрепления «единицы всего живого» к различным поверхностям и способствует движению.
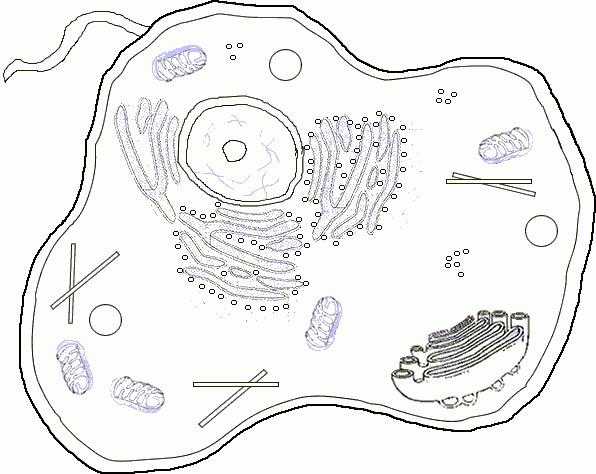
Органоиды в составе цитоплазмы. Эндоплазматическая сеть и рибосомы
Помимо гиалоплазмы, цитоплазма содержит в себе и множество микроскопических органоидов, которые различаются по строению. Их присутствие в растительных и животных клетках свидетельствует о том, что все они выполняют важнейшие функции и жизненно необходимы. В какой-то степени эти морфологические образования сравнимы с органами тела человека или животных, что и дало возможность называть их органоидами. В цитоплазме различают видимые в световой микроскоп органеллы -пластинчатый комплекс, митохондрии и центросому. При помощи электронного микроскопа в матриксе обнаруживаются микротрубочки, лизосомы, рибосомы и плазматическая сеть. Цитоплазма клеточная пронизана различными каналами, которые и получили название «эндполазматическая сеть». Их мембранные стенки контактируют со всеми другими органеллами и составляют единую систему, осуществляющую энергетический обмен, а также перемещение внутри клетки веществ. В стенках этих каналов находятся рибосомы, которые выглядят как мельчайшие гранулы. Они могут располагаться одиночно или группами. Рибосомы состоят из практически равного количества рибонуклеиновой кислоты и белков. Также в их состав включен магний. Рибосомы могут не только находиться в каналах ЭПС, но и свободно лежать в цитоплазме, а также встречаться в ядре, где они и образуются. Совокупность каналов, имеющих рибосомы, называются гранулярной эндоплазматической сетью. На них, кроме рибосом, располагаются ферменты, способствующие синтезу углеводов и жиров. Во внутренних полостях каналов находятся продукты жизнедеятельности клетки. Иногда в расширениях ЭПС формируются вакуоли — полости, заполненные клеточным соком и ограниченные мембраной. Эти органоиды поддерживают тургорное давление. Лизосомы представляют собой мелкие образования овальной формы. Они рассеяны по цитоплазме. Формируются лизосомы в ЭПС или комплексе Гольджи, где наполняются гидролитическими ферментами. Лизосомы предназначены для переваривания частиц, попавших внутрь клетки вследствие фагоцитоза.

Цитоплазма: строение и функции ее органоидов. Пластинчатый комплекс Гольджи, митохондрии и центросома
Комплекс Гольджи представлен в растительных клетках отдельными тельцами, оформленными мембранами, а в животных — канальцами, пузырьками и цистернами. Этот органоид предназначен для химического изменения, уплотнения и последующего вывода в цитоплазму продуктов клеточной секреции. Также в нем осуществляется синтез полисахаридов и образование гликопротеидов. Митохондрии — это тельца палочковидной, нитевидной или зернистой формы. Они ограничиваются двумя мембранами, которые состоят из двойных слоев фосфолипидов и белков. От внутренних мембран этих органелл отходят кристы, на стенках которых находятся ферменты. С их помощью происходит синтез аденозинтрифосфорной кислоты (АТФ). Митохондрии иногда называют «клеточными электростанциями», так как они поставляют значительную часть аденозинового трифосфата. Он используется клеткой как источник химической энергии. Кроме того, митохондрии выполняют и другие функции, в том числе: передачу сигналов, некроз клеток, клеточное дифференцирование. Центросома (клеточный центр) состоит из двух центриолей, которые располагаются под углом друг к другу. Этот органоид имеется у всех животных и растений (кроме простейших и низших грибов) и отвечает за определение полюсов при митозе. В делящейся клетке сначала разделяется центросома. При этом образуется ахроматиновое веретено, которое задает ориентиры хромосомам, расходящимся к полюсам. Кроме обозначенных органоидов в клетке могут находиться и органеллы специального назначения, например, реснички и жгутики. Также на определенных этапах жизнедеятельности в ней могут иметься и включения, то есть временные элементы. Например, такие питательные вещества как: капельки жира, белки, крахмал, гликоген и т. д.
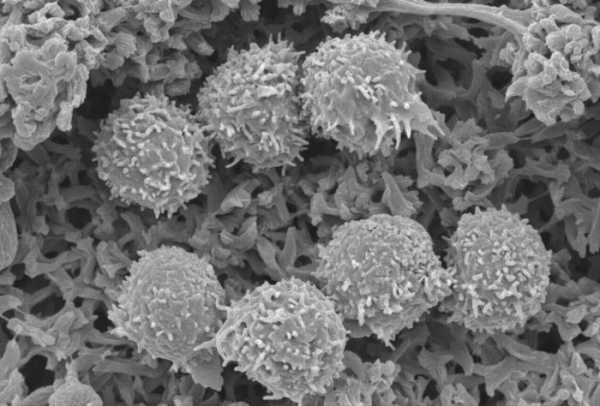
Лимфоциты — важнейшие клетки иммунной системы
Лимфоциты — это важные клетки, относящиеся к группе лейкоцитов крови человека и животных и участвующие в иммунологических реакциях. Они подразделяются по размеру и структурным особенностям на три подгруппы:
- малые — диаметром менее 8 мкм;
- средние — диаметром от 8 до 11 мкм;
- большие — диаметром свыше 11 мкм.
Малые лимфоциты преобладают в крови животных. Они имеют крупное ядро округлой формы, преобладающее над объемом цитоплазмы. Цитоплазма лимфоцитов этой подгруппы выглядит как ядерный ободок или серп, прилежащий к какой-либо стороне ядра. Часто в матриксе содержится некоторое количество азурофильных гранул мелкого размера. Митохондрии, элементы пластинчатого комплекса и канальцы ЭПС немногочисленны и находятся около ядерного углубления. Средние и большие лимфоциты устроены несколько иначе. Их ядра имеют бобовидную форму, содержат меньшее количество хроматина конденсированного. В них легко различить ядрышко. Цитоплазма лимфоцитов второй и третьей групп имеет более широкий ободок. Известно два класса лимфоцитов, так называемые В- и T-лимфоциты. Первые образуются у животных в миеловидной ткани костного мозга. Эти клетки имеют способность образовывать иммуноглобулины. С их помощью В-лимфоциты взаимодействуют с антигенами, распознавая последних. Т-лимфоциты образуются из костномозговых клеток в тимусе (в его корковой части долек). В их цитоплазматической мембране находятся поверхностные антигены гистосовместимости, а также многочисленные рецепторы, при помощи которых осуществляется распознавание чужеродных частиц. Малые лимфоциты, в основном, представлены T-лимфоцитами (более 70%), среди которых имеется большое количество долгоживущих клеток. Подавляющее большинство B-лимфоцитов живут недолго — от одной недели до месяца.
Надеемся, наша статья оказалась полезной, и теперь вы знаете, что такое цитоплазма, гиалоплазма и плазмелемма. А также осведомлены, каковы функции, строение и значение для жизнедеятельности организма этих клеточных образований.
fb.ru
Реферат — Цитоплазма эукариотической клетки
Скачать реферат: Цитоплазма эукариотической клетки |
|||
Содержание реферата
1. Роль ученых в открытии клеточных мембран
2.
Цитоплазма эукариотической клетки
3. Гиалоплазма — внутренняя среда клетки
4. Мембраны цитоплазмы
4.1. Химия и свойства клеточных мембран
4.2. Структура клеточных мембран
5. Плазматическая мембрана
5.1. Рост плазматической мембраны
6. Функции плазматической мембраны
6.1. Транспортные функции
6.2. Рецепторные функций плазматической мембраны
6.3. Межклеточные контакты
6.4. Простой контакт. Соединение типа «замка»
6.5. Плотный замыкающий контакт
6.6. Промежуточный контакт
6.7. Щелевидный контакт
Список литературы
1. Роль ученых в открытии клеточных мембран
До конца XVIII века предполагали, что клетки разделены общей стенкой и не могут
быть изолированными друг от друга. В дальнейшем развитии данной науки большую роль
сыграли исследования французского ботаника Бриссо де Мирбеля (1776-1854), в которых
впервые высказывалась идея сравнительного изучения растительных тканей. Мирбель
впервые предположил, что соприкасающиеся клетки разделены общей перегородкой — мембраной.
Немецкий естествоиспытатель Рудольфи пришел к заключению об обособленности клеток,
о наличии у них собственных мембран, окружающих их со всех сторон. Всякие сомнения
о том, что клетки могут быть выделены и изолированы, окончательно исчезли после
работы ботаника И. Молденгауера (1766-1827), из которой следовало, что клетка является
самостоятельной структурой, покрытой оболочкой. Итак, сформировалось новое представление
о клетке.
Наиболее четко его сформулировал несколько позже Ф. Мейен. Его определение таково:
«Клетка растительного организма представляет собой пространство, вполне замкнутое
вегетативной мембраной». Ф. Мейен ясно изложил этот взгляд, в начале ХIХ в. сменивший
первоначальное воззрение на клетку как на пору. Но конкретное содержание анатомии
растений по-прежнему принципиально мало чем отличалось от старого. Исследователи
продолжали системно описывать строение различных органов растений, а также детализировать
и уточнять изученное. Этот период справедливо считается периодом собирания материала,
периодом накопления многочисленных сведений о тончайшей структуре растений.
Итак, сам факт наличия клеток в различных тканях сомнения не вызывал. Но исчерпывается
ли все разнообразие структур растительных организмов только клетками? Вопрос оставался
еще не до конца ясным. Более того, большинство ученых отвечало на этот вопрос отрицательно,
указывая, наряду с клетками, на сосуды и волокна растений как на вполне своеобразные
структуры. Обобщая все знания в этой области, Мейен прямо писал, что растительные
ткани состоят из клеток, волокон и сосудов. Возникновение этих последних из метаморфизирующихся
клеток было окончательно установлено лишь несколько позднее — в 30-х гг. прошлого
века.
2. Цитоплазма эукариотической клетки
Цитоплазма представляет собой метаболический рабочий аппарат клетки. В ней сосредоточены
общие и специальные органоиды, в цитоплазме протекают основные метаболические процессы.
В последние десятилетия достигнуты особенно большие успехи в дискретном морфофункциональном
анализе отдельных органоидов и мембранных систем метаболического аппарата цитоплазмы.
Это делает возможным представить себе работу метаболического аппарата как целостной
системы.
В современных исследованиях метаболического аппарата клетки особенно ярко проявляется
тенденция к изучению функционального значения морфологических структур — анализу
функции через структурно-биохимическую организацию этих структур. Важнейшие успехи
в этом плане достигнуты в изучении основных органоидов белкового синтеза (рибосом)
и систем, обеспечивающих энергетический обмен (митохондрий, хлоропластов и сопрягающих
мембран прокариотных клеток). Крупным обобщением в изучении метаболического аппарата
цитоплазмы являются представления о нем как о пластичной дифференцированной трехфазной
системе, состоящей из основной цитоплазмы, внутриклеточных мембран и содержимого
мембранной системы. Таким образом метаболический аппарат цитоплазмы, с одной стороны,
сложен специализированными структурами, выполняющими частные функции, а с другой
стороны, является единой целостной системой, находящейся в тесной взаимосвязи и
с поверхностным, и с ядерным аппаратами клетки.
Начало такому подходу положил еще в 60-х гг. Де Дюв в учении об экзоплазматических
мембранных системах. В дальнейшем этот подход развивался и в учении о лизосомах,
и в современных представлениях об аппарате Гольджи и эндоплазматической сети, а
также в представлениях о динамичном взаимодействии внутриклеточных мембранных систем.
Весьма перспективным, на наш взгляд, является также оформившееся в последнее время
представление об основной цитоплазме — гиалоплазме. В противовес бытовавшему раньше
взгляду на гиалоплазму как на однородную живую коллоидную систему оно трактует основную
цитоплазму как дифференцированную, гетерогенную фазу цитоплазмы, способную к формированию
сложных структур. С этим свойством гиалоплазмы мы уже частично познакомились на
примере субмембранной системы поверхностного аппарата с ее динамичными механо-химическими
и опорными структурами.
В настоящее время становится ясно, что такие примембранные системы гиалоплазмы формируются
не только в области плазматических мембран, но и в области всех мембранных, а иногда
даже и немембранных (клеточный центр) структур цитоплазмы. Эту часть цитоплазмы
предлагается выделить в особую субсистему гиалоплазмы, обозначив ее термином «цитозоль».
Естественно, что функции и морфобиохимическая организация такого цитозоля в области
шероховатой и гладкой эндоплазматической сети (ЭПС), мембран аппарата Гольджи, митохондрий,
клеточного центра, ядерной оболочки и т. д. будут различны и специфичны для тех
структур, вокруг которых формируются соответствующие участки цитозоля.
С учетом структурного и функционального разнообразия органоидов, обеспечивающих
внутриклеточный метаболизм в цитоплазме клеток, функция соответствующих каждому
конкретному органоиду и практически входящих в его состав участков цитозоля не будет
ограничиваться только опорно-сократительной. Структуры и ферменты, входящие в состав
цитозоля, специфичны для каждого органоида и принимают непосредственное участие
в обеспечении протекающих в данном органоиде биохимических процессов.
Рассмотрение цитоплазмы целесообразно начать с характеристики проблемы структурно-биохимической
организации рибосом — молекулярных машин белкового синтеза. В этом случае особенно
ярко проявляется значение структурной организации биохимических процессов в клеточных
системах и достигнуты большие успехи в ее изучении.
Второй областью биологии клетки, где структурная организация биохимических процессов
также изучена весьма детально, является анализ так называемых сопрягающих мембран
— аппарата энергетического обмена клеток. Их мы рассмотрим при характеристике митохондрий,
пластид и других структур цитоплазмы, отвечающих за энергетический обмен в клетке.
Далее мы обсудим результаты дискретного анализа основных мембран.
Собственно тело клетки и ее содержимое отделено от внешней среды или от соседних
элементов у многоклеточных организмов плазматической мембраной. Кнаружи от плазматической
мембраны, экстрацеллюлярно, расположена клеточная оболочка, или стенка, особенно
хорошо выраженная у растений и прокариотических организмов; у клеток животных она
отсутствует или выражена очень слабо. Все внутреннее содержимое клетки, за исключением
ядра, носит название цитоплазмы. Это общий термин, который подчеркивает разделение
клетки на два главных компонента: цитоплазму и ядро.
Цитоплазма эукариотических клеток неоднородна по своему строению и составу и включает
в себя гиалоплазму, мембранные и немембранные компоненты. К мембранным компонентам
относится вакуолярная система (эндоплазматическая сеть, аппарат Гольджи, лизосомы,
вакуоли растений), мембранные органеллы (митохондрии и пластиды). К немембранным
компонентам относятся центриоли, характерные для животных клеток, и немембранные
макромолекулярные комплексы и структуры, такие как рибосомы, микротрубочки и микрофиламенты.
Такое разделение цитоплазмы на отдельные компоненты не означает их структурной и
функциональной обособленности друг от друга.
3. Гиалоплазма — внутренняя среда клетки
Термины «гиалоплазма» (от hyaline — просвечивающийся, прозрачный), «основная плазма»,
или «матрикс цитоплазмы», обозначают очень важную часть клетки, ее истинную, внутреннюю
среду. В электронном микроскопе матрикс цитоплазмы имеет вид гомогенного или тонкозернистого
вещества с низкой электронной плотностью. Из старых наблюдений за физико-химическими
свойствами цитоплазмы было получено представление о том, что гиалоплазма является
сложной коллоидной системой, включающей в себя различные биополимеры: белки, нуклеиновые
кислоты, полисахариды и др. Эта система способна переходить из золеобразного (жидкого)
состояния в гель и обратно. Так, например, при высоких гидростатических давлениях
цитоплазма не уплотняется, а обратимо разжижается. Это явление объясняется нарушением
связей между молекулами в составе гиалоплазмы.
Благодаря электронно-микроскопическим и физико-химическим методам исследования возникли
представления об основной плазме как об организованной, упорядоченной многокомпонентной
системе. Отдельные зоны гиалоплазмы могут менять свое агрегатное состояние — в зависимости
от условий или от функциональной задачи. Так, известно, что отдельные молекулы белков-тубулинов
могут быть диспергированы в гиалоплазме, но в определенные моменты они начинают
собираться и строить длинные трубчатые структуры — микротрубочки.
Этот процесс самосборки микротрубочек обратим: при изменении условий жизни клетки
(повышение давления или изменение проницаемости мембран клетки) микротрубочки распадаются
до мономерных молекул тубулинов. Таким же образом в бесструктурной, на взгляд, гиалоплазме
могут возникать и распадаться различные фибриллярные, нитчатые комплексы белковых
молекул. Основная плазма может принимать участие в образовании клеточных мембран,
нитей и микрофиламентов. У амеб внешние слои цитоплазмы (эктоплазма) или зона цитоплазмы
в области псевдоподии практически состоят из одной гиалоплазмы, там редко обнаруживаются
мембраны, митохондрии и другие структуры. Однако после воздействия ионов, способствующих
набуханию клетки происходят быстрые и резкие перестройки этих участков цитоплазмы
— в них появляются мембранные мелкие вакуоли. Следовательно, основная плазма должна
содержать все строительные компоненты (липиды и белки), которые необходимы для образования
мембран.
Фибриллярные цитоплазматические структуры также могут возникать в результате специфических
агрегационных процессов в гиалоплазме. Если из гомогенатов клеток осадить крупные
ядра, мембранные структуры, а затем рибосомы, то оставшаяся надосадочная жидкость
будет заключать в себе основные химические компоненты гиалоплазмы. В состав гиалоплазмы
из микромолекул входят главным образом различные глобулярные белки и ферменты цитоплазматического
матрикса. Они составляют 20-25 % общего содержания белков в эукариотической клетке.
В бактериальных клетках, бедных мембранными элементами, на долю белков гиалоплазмы
может приходиться около 50 % всех белков. К важнейшим ферментам матрикса относятся
ферменты гликолиза, ферменты метаболизма сахаров, азотистых оснований, аминокислот,
липидов и других важных соединений. В матриксе располагаются ферменты активации
аминокислот при синтезе белка, трансферные РНК. Осмотические и буферные свойства
клетки в значительной степени определяются составом и структурой гиалоплазмы.
Важнейшая роль гиалоплазмы заключается в том, что эта полужидкая среда объединяет
все клеточные структуры и обеспечивает химическое взаимодействие их друг с другом.
Через гиалоплазму осуществляется большая часть внутриклеточных транспортных процессов:
перенос аминокислот, жирных кислот, нуклеотидов, сахаров. В гиалоплазме идет постоянный
поток ионов к плазматической мембране и от нее, к митохондриям, ядру и вакуолям.
Гиалоплазма является основным вместилищем и зоной перемещения массы молекул АТФ.
В ней же происходит отложение запасных продуктов: гликогена, жировых капель. В структурном
и морфологическом отношении гиалоплазма еще изучена плохо.
4. Мембраны цитоплазмы
Общей чертой всех мембран клетки, внешней плазматической мембраны и всех внутриклеточных
мембран и мембранных органоидов является то, что они представляют собой тонкие
(6-10 нм) пласты липопротеидной природы (липиды в комплексе с белками), замкнутые
сами на себя. В клетке нет открытых мембран со свободными концами. Мембраны клетки
всегда ограничивают полости или участки, закрывая их со всех сторон и тем самым
отделяя содержимое таких полостей от окружающей их среды. Так, плазматическая мембрана,
покрывая всю поверхность клетки, имеющей сложную форму и многочисленные выросты,
нигде не прерывается, она замкнута. Она отделяет содержимое цитоплазмы от окружающей
клетку среды.
Внутриклеточные замкнутые мембраны образуют пузырьки — вакуоли шаровидной или уплощенной
формы. В последнем случае образуются плоские мембранные мешки, или цистерны. Часто
полости, отграниченные мембранами, имеют сложную форму, напоминающую губку или сеть,
но и в этом случае такие полости без перерывов отграничены мембраной. В подобных
вариантах мембраны также разделяют две структурные фазы цитоплазмы: гиалоплазму
от содержимого вакуолей и цистерн. Такое же свойство имеют мембраны митохондрий
и пластид: они разделяют внутреннее содержимое от межмембранных полостей и от гиалоплазмы.
Ядерная оболочка тоже может быть представлена в виде перфорированного полого двойного
мембранного мешка шаровидной формы. Мембраны ядерной оболочки разграничивают, отделяют
друг от друга кариоплазму и хромосомы от полости перинуклеарного пространства и
от гиалоплазмы. Эти общие морфологические свойства клеточных мембран определяются
их химическим составом, их липопротеидной природой.
4.1. Химия и свойства клеточных мембран
Основными химическими компонентами клеточных мембран служат липиды (40 %) и белки
(60 %), кроме того, во многих мембранах обнаружены углеводы. К липидам относится
большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность)
и растворимостью в органических растворителях и жирах (липофильность).
Состав липидов, входящих в мембраны клетки, очень разнообразен. Характерными представителями
липидов, встречающихся в клеточных мембранах, являются фосфолипиды, сфингомиелины
и из стероидных липидов — холестерин. Глицеролипиды, представляют собой сложные
эфиры трехатомного спирта, глицерина с двумя жирными кислотами и с фосфорной кислотой,
которая, в свою очередь, может быть связана с различными химическими группами (холин,
серин, инозит, этаноламин и др.).
Другой группой мембранных липидов являются сфингомиелины, где глицерин замещен аминоспиртом
сфингозином. Из липидов, относящихся к стероидам, больше всего в мембранах холестерина.
В растительных клетках холестерин не обнаружен, его там заменяют фитостерины. У
бактерий стерины отсутствуют. Характерной особенностью мембранных липидов является
разделение их молекулы на две функционально различные части: неполярные хвосты,
не несущие зарядов и состоящие из жирных кислот, и заряженные полярные головки.
Полярные головки несут на себе отрицательные заряды или могут быть нейтральными
(в случае, если они имеют одновременно положительные и отрицательные заряды).
Было обнаружено, что клеточные мембраны сильно отличаются друг от друга по составу
липидов. Так, плазматические мембраны клеток животных богаты холестерином (до
30 %), в них мало лецитина, в то время как мембраны митохондрий, наоборот, богаты
фосфолипидами и бедны холестерином. Из общего количества липидов содержание лецитина
во фракциях эндоплазматической сети составляет 60-70 % от всех фосфолипидов, в то
время как в плазматической мембране его может быть 25-35 %.
В целом для плазматической мембраны характерно высокое содержание холестерина и
сфинголипидов, а также преобладание насыщенных и мононенасыщенных жирных кислот
в составе фосфолипидов, тогда как в митохондриях, эндоплазматической сети и во многих
других цитоплазматических мембранах содержится мало холестерина и сфинголипидов
и сравнительно много полиненасыщенных жирных кислот. Видимо, в связи с этим мембраны
цитоплазмы менее жесткие, чем плазматическая мембрана, они более легкоплавки.
Количество белков в клеточных мембранах также различно. Так, в митохондриях, по
сравнению с другими мембранами, белков больше всего. Очень разнятся мембраны набором
белковых молекул. Часть из них связана с липидными головками с помощью ионных связей
и поэтому легко экстрагируются из мембран растворами солей. Другие образуют солевые
связи с полярными участками липидов через взаимодействие с ионами Mg2+ или Са2+.
Белки также экстрагируются с помощью хелатных соединений, таких как версен (ЭДТА).
Большая же часть белков взаимодействует с липидами в составе мембран на основе гидрофобных
связей.
Стало известно, что многие мембранные белки состоят как бы из двух частей: из участков,
богатых полярными (несущими заряд) аминокислотами, и участков, обогащенных неполярными
аминокислотами (глицином, аланином, валионом, лейцином). Такие белки в липидных
слоях мембран располагаются так, что их неполярные участки как бы погружаются в
«жирную» часть мембраны, где находятся гидрофобные участки липидов. Полярная (гидрофильная)
же часть таких белков взаимодействует с головками липидов и обращена в сторону водной
фазы. Поэтому такие белки, связанные с липидами путем гидрофобных взаимодействий,
практически не экстрагируются в водных фазах. Их можно выделить, лишь разрушая мембрану,
экстрагируя из нее липиды органическими растворителями или детергентами.
По биологической роли мембранные белки можно разделить на три группы: ферменты,
рецепторные белки и структурные белки. Набор ферментов в составе мембран может быть
очень велик и разнообразен (например, в плазматической мембране клеток печени обнаружено
не менее 24 различных ферментов).
В разных мембранах существует характерный набор ферментов. Иными словами, химическая
разнокачественность мембран определяется не только липидами, но и белками. Общим
для всех клеточных мембран является их липопротеидное строение; несходство же заключается
в количественных и качественных различиях по липидному и белковому составу. Например,
состав и количество липидов и белков будут совсем разными в мембранах эндоплазматической
сети и внутренней мембране митохондрий.
Углеводный компонент мембран представлен главным образом гликопротеинами — молекулами
белков, ковалентно (в отличие от нуклеопротеидов и липопротеидов) связанных с цепочками
углеводов.
4.2. Структура клеточных мембран
Представление о том, что в основе клеточных мембран лежит двойной липидный слой,
получено еще в 20-х гг. В 1925 г. была опубликована работа, которая указывала на
образование сплошного билипидного слоя плазматической мембраны. Несмотря на ряд
ошибок, общая идея о существовании билипидного слоя, оказалась справедливой. Выяснилось,
что если экстрагировать липиды из оболочки эритроцитов, а затем поместить их на
поверхность водного мениска, то можно рассчитать площадь, занимаемую образовавшимся
монослоем липидов. Оказалось, что эта площадь вдвое больше площади, занимаемой поверхностью
эритроцитов, из которых были экстрагированы липиды.
Ученые предположили, что в мембранах эритроцитов липиды располагаются в два слоя.
К тому же оказалось, что поверхностное натяжение клетки значительно ниже, чем поверхностное
натяжение искусственного липидного слоя. Далее исследователи обнаружили, что при
добавлении белка к маслу поверхностное натяжение снижается до величины, характерной
для поверхностного натяжения клеток. Исходя из этого заключили, что в состав мембраны
входит белок.
Так возникла гипотеза о структуре мембраны, согласно которой она представляет собой
трехслойный «сэндвич» (белок-липид-белок). Липидный слой в центре — это бимолекулярный
слой, в котором гидрофобные концы молекул липидов направлены друг к другу, а гидрофильные
— к белковым слоям, лежащим по обе стороны от липидного слоя. Эта гипотеза получила
прямые подтверждения, когда стало возможным использовать для изучения мембран клетки
электронную микроскопию. Благодаря ей удалось увидеть особенно хорошо на мембранах
миелиновой оболочки нервов, что мембрана действительно трехслойная: два темных слоя
по 2,5 нм каждый и более широкий светлый слой между ними.
Трехслойность клеточных мембран удается показать в электронном микроскопе с помощью
метода замораживания-скалывания, что особенно важно, т. к. в этом случае есть возможность
наблюдать специфические ферменты или даже ферментные системы. В начале 60-х гг.
Робертсоном была сформулирована гипотеза о единой, унитарной «элементарной» биологической
мембране. Вопрос о возникновении, образовании мембран в живой клетке еще недостаточно
изучен. Известно, что количество и величина мембранных элементов в клетках может
возрастать и уменьшаться, т. е. можно говорить о мембраногенезе и о разрушении мембран.
5. Плазматическая мембрана
Плазматическая мембрана, или плазмалемма, среди различных клеточных мембран занимает
особое место. Это поверхностная периферическая структура, ограничивающая клетку
снаружи, что обусловливает ее непосредственную связь с внеклеточной средой, а следовательно,
со всеми веществами и стимулами, воздействующими на клетку. В химическом отношении
плазматическая мембрана представляет собой липопротеиновый комплекс.
Главные принципы организации мембран были уже обсуждены в предыдущем разделе. Она
имеет толщину около 10 нм и представляет собой самую толстую из клеточных мембран.
Основными компонентами плазматических мембран являются липиды (около 40 %), белки
(более 60 %) и углеводы (около 1 %). Как уже указывалось, плазмалемма по сравнению
с другими мембранами более богата холестерином, в ее фосфолипидах преобладают насыщенные
жирные кислоты.
Состав белков, входящих в плазматическую мембрану, очень разнообразен, что определяется
многообразием ее функциональных нагрузок. Среди ферментов обнаружены 5′- нуклеотидаза,
Mg-зависимая АТФаза, активируемая ионами Na и К, щелочная и кислая фосфатаза, аденилатциклаза,
РНКаза, обнаружена протеолитическая активность. Ферменты дыхательной цепи и гликолитические
ферменты в плазматической мембране эукариотических клеток полностью отсутствуют.
Зато у прокариотических клеток именно в составе плазматической мембраны локализованы
элементы цепи переноса электронов и окислительного фосфорилирования.
Необходимо подчеркнуть, что часть белков, обнаруживаемых во фракциях плазматической
мембраны, может быть и не связана непосредственно с билипидным слоем. Многие ферменты,
обнаруживающие свою активность во фракциях плазматической мембраны, могут быть локализованы
в гликокаликсе, гликопротеиновом комплексе, ассоциированном с плазматической мембраной.
В состав именно гликокаликса входят различные углеводы, которые могут составлять
около 1 % от сухого веса мембран.
Углеводы образуют длинные ветвящиеся цепочки полисахаридов, связанных с белками
слоя, часто определяющего форму клетки. К плазматической мембране здесь изнутри
могут примыкать мембранные мешочки; в этом случае у поверхности клеток имеются три
мембранных слоя: собственно плазматическая мембрана и две мембраны пелликулярных
альвеол.
У инфузории туфельки пелликула образует утолщения, располагающиеся в виде шестиугольников,
в центре которых выходят реснички. Жесткость пелликулярных образований может быть
связана также с элементами цитоплазмы, подстилающими плазматическую мембрану, с
кортикальным слоем. Так, в гребнях пелликулы эвглены вблизи мембраны обнаруживаются,
кроме мембранных вакуолей, параллельные пучки микротрубочек и микрофиламентов. Такая
фибриллярная периферическая арматура вместе со складчатой многослойной мембранной
периферией создает жесткую структуру пелликулы.
5.1. Рост плазматической мембраны
После деления клеток происходит увеличение объемов растущих дочерних клеток, и в
связи с этим происходит рост клеточной поверхности, увеличение площади плазматической
мембраны. Но это не единственный пример быстрого роста объема и поверхности. Поверхность
быстро растущих клеток в тычиночных нитях злаков может за 1 ч увеличиться в 65 раз,
т. е. каждую минуту плазмалемма нарастает на ее первоначальную величину. Такую большую
скорость роста плазматической мембраны можно объяснить только тем, что происходит
быстрое встраивание готовых мембранных предшественников, липидов и белков, в старый
липопротеидный слой. Подобный способ роста вполне возможен, т. к. известно, что
в клетке происходит постоянная перестройка мембран — замена одних липидов на другие,
изменение их количества, изменение состава белков и т. д. Следовательно, мембраны
представляют собой очень лабильные соединения.
Но описан и другой случай образования новой плазматической мембраны. Плазмодий миксомицета
Physarum pоlycephalum протыкали стеклянным капилляром и полученный цилиндр плазмы,
лишенный плазматической мембраны, фиксировали сразу же и далее через 1, 2, 3 и
6 с после начала эксперимента, затем изучали в электронном микроскопе. Для цитоплазмы
этого гриба характерно наличие большого числа мелких пузырьков, содержащих слизь,
которая покрывает весь плазмодий снаружи. Можно было видеть, что такие пузырьки,
рассеянные в основной плазме, собирались у незащищенного мембраной края плазмы,
образуя уплощенные вакуоли, располагающиеся параллельно наружной поверхности обнаженного
кусочка. Такие плоские вакуоли сливались и образовывали сплошную мембрану — новую
плазматическую мембрану, изолирующую компоненты плазмы от внешней среды.
По-видимому, такой процесс обновления плазматической мембраны происходит не только
при больших ее повреждениях, но и постоянно, в процессе жизнедеятельности клетки.
Так, в результате экзоцитоза мембраны вакуолей, сливаясь с плазматической мембраной,
как бы встраиваются в нее, тем самым увеличивая общую площадь поверхности клетки.
Наблюдения за процессом секреции (одна из форм экзоцитоза) привели многих исследователей
к представлению о том, что в клетке постоянно имеются потоки мембранных элементов:
от плазматической мембраны внутрь цитоплазмы (эндоцитоз) и, наоборот, ток мембранных
структур из цитоплазмы к поверхности клетки (экзоцитоз). В этом круговороте мембран
ведущая роль отводится системе мембранных вакуолей аппарата Гольджи.
6. Функции плазматической мембраны
Плазматическая мембрана выполняет целый ряд важнейших клеточных функций, ведущими
из которых являются функция разграничения веществ цитоплазмы от внешней среды и
функции транспорта различных веществ как внутрь клетки, так и из нее. K транспортным
функциям относятся пассивный транспорт воды, ионов, низкомолекулярных веществ и
активный перенос этих веществ против градиентов концентрации, а также разные формы
транспорта высокомолекулярных соединений и комплексов (эндоцитоз).
Плазматическая мембрана, кроме того, участвует в выведении из клеток продуктов,
образованных в ней. Она же участвует в процессах внеклеточного расщепления биополимеров.
На поверхности плазмалеммы располагаются различные рецепторные структуры, специфически
взаимодействующие с внеклеточными факторами и с соседними клетками. Тем самым клеточная
мембрана участвует в передаче сигналов внутрь клетки. Плазматическая мембрана принимает
участие в межклеточных взаимодействиях в многоклеточных организмах. Отдельные участки
плазматической мембраны в специализированных клетках животных принимают участие
в построении специальных отростков клетки, таких как микроворсинки, реснички, рецепторные
выросты и др. Наконец, плазматическая мембрана играет важную роль при делении клетки.
Это краткое перечисление функций плазматической мембраны далеко от полноты, однако
из него видна чрезвычайная важность этой периферической структуры любой клетки,
будь то клетка бактерий или эукариотических организмов.
6.1. Транспортные функции
Плазматическая мембрана, как и другие клеточные липопротеидные мембраны, полупроницаема.
Это значит, что через нее с различной скоростью проходят разные молекулы, и чем
больше размер молекул, тем меньше скорость их прохождения через мембрану. Это свойство
определяет плазматическую мембрану как осмотический барьер.
Максимальной проникающей способностью обладает вода и растворенные в ней газы, значительно
медленнее проникают сквозь мембрану ионы (примерно в 104 раза медленнее). Поэтому
если клетку, например эритроцит, поместить в среду, где концентрация солей будет
ниже, чем в клетке (гипотония), то вода снаружи устремится внутрь клетки, что приведет
к увеличению ее объема и к разрыву плазматической мембраны (гипотонический «шок»).
Наоборот, при помещении эритроцита в растворы солей более высокой концентрации,
чем в клетке, произойдет выход воды из нее во внешнюю среду. Клетка при этом сморщится,
уменьшится в объеме. Такой пассивный транспорт воды из клетки и в клетку все же
идет с низкой скоростью. Следовательно, в клеточной мембране, в ее липопротеидном
слое существуют специальные поры для проникновения воды и ионов.
6.2. Рецепторные функции плазматической мембраны
Эти функции связаны с локализацией на плазматической мембране специальных структур,
нацеленных на специфическое узнавание химических или физических факторов. Клеточная
поверхность обладает большим набором компонентов — рецепторов, определяющих возможность
специфических реакций с различными агентами. В качестве таких рецепторов на поверхности
клетки могут выступать белки мембраны или элементы гликокаликса (полисахариды, гликопротеиды).
Считается, что такие чувствительные к отдельным веществам участки могут быть разбросаны
по поверхности клетки или собраны в небольшие зоны. Так, на поверхности бактериальных
клеток или клеток животных существует ограниченное число мест, с которыми могут
связываться вирусные частицы. Интересно, что разные вирусы связываются обычно с
разными участками клеточной периферии, причем одна и та же бактериальная клетка
может иметь на поверхности несколько разных типов рецепторов.
Применение различных иммуннологических методов показало, что на поверхности клетки
локализуются антигенно активные компоненты, специфически реагирующие с антителами
или иммунными клетками. Большая часть таких поверхностных антигенов содержит углеводные
группировки (гликолипиды, гликопротеиды). Антигенными компонентами бактериальной
стенки также чаще всего являются липополисахариды и гликолипиды. На поверхности
клетки, часто прямо в ее липопротеидной мембране, располагаются различные рецепторы,
связывающие физиологически активные вещества, такие как разные гормоны, медиаторы
и пр.
Разные клетки животных организмов могут обладать разными наборами рецепторов или
же разной чувствительностью одного и того же рецептора. Роль многих клеточных рецепторов
заключается в передаче сигналов с поверхности внутрь клетки. В настоящее время хорошо
изучена система передачи стимула клеткам с помощью некоторых гормонов, в состав
которых входят пептидные цепочки. Обнаружено, что эти гормоны связываются со специфическими
рецепторами на поверхности плазматической мембраны клетки.
Разнообразие и специфичность наборов рецепторов на поверхности клеток приводит к
созданию очень сложной системы маркеров, позволяющих отличать свои клетки (той же
особи или того же вида) от чужих. Сходные клетки вступают друг с другом во взаимодействия,
приводящие к слипанию поверхностей (конъюгация у простейших и бактерий, образование
тканевых клеточных комплексов). При этом клетки, отличающиеся набором детерминантных
маркеров или не воспринимающие их, либо исключаются из такого взаимодействия, либо
у высших животных уничтожаются в результате иммуннологических реакций.
С плазматической мембраной связана локализация специфических рецепторов, реагирующих
на физические факторы. Так, в плазматической мембране или в ее производных у фотосинтетических
бактерий и сине-зеленых водорослей локализованы белки — рецепторы (хлорофиллы),
взаимодействующие с квантами света. В плазматической мембране светочувствительных
клеток животных расположена специальная система фоторецепторных белков (родопсин),
с помощью которых световой сигнал превращается в химический, что, в свою очередь,
приводит к генерации электрического импульса.
6.3. Межклеточные контакты
Плазматическая мембрана, как уже говорилось, принимает активное участие в межклеточных
контактах, связанных с конъюгацией одноклеточных организмов. У многоклеточных организмов
за счет межклеточных взаимодействий образуются сложные клеточные ансамбли, поддержание
которых может осуществляться разными путями. В зародышевых, эмбриональных тканях,
особенно на ранних стадиях развития, клетки остаются в связи друг с другом за счет
способности их поверхностей слипаться. Это свойство соединения клеток может определяться
свойствами их поверхности, которые специфически взаимодействуют друг с другом.
Механизм этих связей еще недостаточно изучен, но, вероятнее всего, он обеспечивается
взаимодействием между липопротеидами и гликокаликсом плазматических мембран. При
таком межклеточном взаимодействии эмбриональных клеток между плазматическими мембранами
всегда остается щель шириной около 20 нм, заполненная гликокаликсом. Обработка ткани
ферментами, нарушающими целостность гликокаликса (муказы, действующие гидролитически
на муцины, мукополисахариды) или повреждающими плазматическую мембрану (протеазы),
приводит к обособлению клеток друг от друга, к их диссоциации. Однако если удалить
фактор диссоциации, то клетки могут снова собираться, реагрегировать.
Так, можно диссоциировать клетки разных по окраске губок — оранжевых и желтых. Оказалось,
что в смеси этих клеток образуются два типа агрегатов: состоящие только из желтых
и только из оранжевых клеток. При этом смешанные клеточные суспензии самоорганизуются,
восстанавливая исходную многоклеточную структуру. Сходные результаты были получены
с суспензиями разделенных клеток эмбрионов амфибий; в этом случае происходит избирательное
пространственное обособление клеток эктодермы от энтодермы и от мезенхимы.
Более того, если для реагрегации используются ткани поздних стадий развития зародышей,
то в пробирке самостоятельно собираются различные клеточные ансамбли, обладающие
тканевой и органной специфичностью, образуются эпителиальные агрегаты, сходные с
почечными канальцами, и т. д. Соединения между клетками в составе тканей и органов
многоклеточных животных организмов могут образовываться сложными специальными структурами,
которые называют собственно межклеточными контактами. Эти структурированные межклеточные
контакты особенно выражены в покровных пограничных тканях, в эпителиях.
Возможно, что первичное обособление пласта клеток, связанных друг с другом с помощью
специальных структурированных межклеточных контактов, в филогенезе животных обеспечило
образование и развитие тканей и органов. Благодаря электронной микроскопии накопилось
множество данных об ультраструктуре этих соединительных образований. К сожалению,
их биохимический состав и молекулярная структура еще недостаточно точно изучены.
Изучая соединения клеток в эпителиальных пластах, можно обнаружить следующие структуры,
связывающие клетки друг с другом: простой контакт, соединение типа «замка», плотный
контакт, промежуточный контакт, или зона слипания, десмосомный контакт, щелевидный
контакт.
6.4. Простой контакт. соединение типа «замка»
Простой контакт встречается среди большинства прилежащих друг к другу клеток различного
происхождения. Большая часть поверхности контактирующих клеток эпителия также связана
с помощью простого контакта, где плазматические мембраны соприкасающихся клеток
разделены пространством 15-20 нм. Как уже говорилось, это пространство представляет
собой надмембранные компоненты клеточных поверхностей. Ширина щели между мембранами
клеток может быть и больше 20 нм, образуя расширения, полости, но не меньше 10 нм.
Со стороны цитоплазмы к этой зоне плазматической мембраны не примыкают никакие специальные
дополнительные структуры.
Соединение типа «замка» являет собой выпячивание плазматической мембраны одной клетки
в инвагинат (впячивание) другой. На срезе такой тип соединения напоминает плотничий
шов. Межмембранное пространство и цитоплазма в зоне «замков» имеют те же характеристики,
что и в областях простого контакта.
6.5. Плотный замыкающий контакт
Он представляет собой зону, где внешние слои двух плазматических мембран максимально
сближены. Часто видна трехслойность мембраны в этом контакте: два внешних осмофильных
слоя обеих мембран сливаются в общий слой толщиной в 2-3 нм. Слияние мембран происходит
не по всей площади плотного контакта, а представляет собой ряд точечных соединений.
Со стороны цитоплазмы в этой зоне часто встречаются многочисленные фибриллы около
8 нм в диаметре, располагающиеся параллельно поверхности плазмалеммы.
Такого типа контакты были обнаружены между фибробластами в культуре ткани, между
эмбриональным эпителием и клетками мезенхимы.
6.6. Промежуточный контакт (или зона слипания)
В этом месте межмембранное расстояние несколько расширено (до 25-30 нм) и, в отличие от простого контакта, заполнено плотным содержимым, вероятнее всего, белковой природы.
6.7. Щелевидный контакт
Он представляет собой область протяженностью 0,5-3 мкм, где плазматические мембраны разделены промежутком в 2-3 нм, что после осмирования придает всей этой структуре семислойный вид. Со стороны цитоплазмы никаких специальных примембранных структур не обнаруживается. Этот тип соединения встречается во всех типах тканей. Функциональная роль щелевидного контакта заключается, видимо, в передаче ионов и молекул от клетки к клетке. Например, в сердечной мышце передача потенциала действия от клетки к клетке происходит через тип контакта, где ионы могут свободно переходить по межклеточным соединениям. Поддержание такой ионной связи между клетками зависит от энергии, получаемой благодаря окислительному фосфорилированию.
Список литературы
1. Вермель Е. М. История учения о клетке. М.: Наука, 1970. 259 с.
2. Гуляев Г. В., Мальченко В. В. Словарь терминов по генетике, цитологии, селекции,
семеноводству и семеноведению. М.: Россельхозиздат, 1983. 240 с.
3. Де Дюв К. Путешествие в мир живой клетки. М.: Мир, 1987. 253 с.
4. Заварзин А. А., Харазова А. Д. Основы общей цитологии: Учебное пособие. Л.: Изд-во
Ленингр. ун-та, 1982. 240 с.
5. От молекул до человека./ Под. ред. Наумова Н. П. М.: Просвещение, 1973. 480 с.
6. Паушева З. П. Практикум по цитологии растений. М.: Агропромиздат, 1988. 271 с.
7. Трошин А. С., Браун А. Д., Вахтин Ю. Б., Жилкин Л. Н., Суханова К. М. Цитология.
М.: Просвещение, 1970. 304 с.
8. Ченцов Ю. С. Общая цитология. М.: МГУ, 1978. 344 с.
© Реферат плюс
referatplus.ru
