
Блок 1.3. Молекулярные основы наследственности. Биосинтез белков и его регуляция
Репликация ДНК происходит
а) постмитотическомпериоде
б) синтетическомпериоде
в) премитотическомпериоде
г) анафазыпериоде
Свойство генетического материала клетки обеспечивающее передачу информации при размножении
а) репарации
б) стабильности
в) комплементарности
г) редупликации
Единица дискретности организма (отдельное свойство)
а) геном
б) признак
в) кодон
г) фенотип
Одна белковая молекулаопределяетразвитие
а) сложного признака
б) простого признака
в) независимого признака
г) неаллельных признаков
В результате совместного синтеза различных ферментов формируется
а) простой
б) специфический
в) сложный
г) элементарный
Нуклеотиды соединяются в полинуклеотидную цепь
а) пептидной связью
б) фосфодиэфирной связью
в) дисульфидной связью
г) водородной связью
С 5`-концом одной цепи ДНК соединён 3`-конец другой цепи
а) комплементарно
б) антипараллельно
в) стабильно
г) альтернативно
Последовательность аминокислот в пептиде зашифрована в ДНК при помощи
а) биохимического кода
б) специального кода
в) смыслового кода
г) генетического кода
Генетический код
а) последовательность аминокислот
б) последовательность фосфорных остатков
в) последовательность нуклеотидов
г) последовательность дезоксирибоз
Процессинг
а) раскручивание двух цепей молекулы ДНК и синтез комплементарных биоспиралей
б) «вырезание» с помощью фермента экзонуклеазы повреждённого участка ДНК и «сшивание» оставшихся участков
в) «вырезание» неинформативных участков из молекулы про-иРНК и «сшивание» оставшихся участков
г) процесс переноса и реализации информации в виде полипептида
Репарация ДНК
а) ошибочное включение в синтезируемую цепь ДНК нуклеотида, несущего химически изменённую форму
б) молекулярное восстановление исходной нуклеотидной последовательности ДНК
в) искажение последовательности нуклеотидов в одной из цепей ДНК
Полуконсервативный способ репликации ДНК
а) синтез двух молекул ДНК, одна из цепей собирается из старых материнских цепей, а другая из новых дочерних полинуклеотидных цепей
б) синтез двух молекул ДНК в которых одна цепь новая дочерняя, другая старая материнская
в) синтез второй цепи ДНК короткими фрагментами
г) синтез двух цепей ДНК, отличающихся друг от друга нуклеотидной последовательностью
Неперекрываемость генетического кода
а) кодирование одним нуклеотидом только одной аминокислоты
б) кодирование разных аминокислот несколькими триплетами
в) нахождение каждого отдельного нуклеотида в составе только одного триплета
г) единство кода для всех организмов
Трансляция
а) авторепродукция с помощью ДНК-полимеразы молекулы ДНК
б) «вырезание» неинформативных участков из молекулы про-иРНК и «сшивание» оставшихся участков
в) «переписывание» информации с молекулы ДНК на про-иРНК
г) репликация последовательности нуклеотидов в последовательность аминокислот в полипептиде
Матричная или информационная РНК
а) РНК имеющая нуклеотидную последовательность, несущею информацию о первичной структуре белка
б) структурный компонент рибосом, образующей пептидильный центр
в) РНК, транспортирующая аминокислоты на рибосомы
г) РНК, которая синтезируется в ядрышке ядра
Фермент, участвующий в вырезании повреждённого участка ДНК
а) экзонуклеаза
б) эндонуклеаза
в) ДНК-полимераза
г) лигаза
Фермент («редактор»), участвующий в узнавании повреждённого участка ДНК
а) экзонуклеаза
б) эндонуклеаза
в) ДНК-полимераза
г) лигаза
Фермент, участвующий в сшивании участка ДНК в процессе репарации
а) экзонуклеаза
б) эндонуклеаза
в) ДНК-полимераза
г) лигаза
Транскрипция
а) «переписывание» информации о синтезе белка с про-иРНК на иРНК
б) «переписывание» информации с молекулы ДНК на про-иРНК
в) «вырезание» неинформативных участков из молекулы про-иРНК
г) авторепродукция с помощью ДНК-полимеразы молекулы ДНК
Фаза инициации
а) начало синтеза пептида
б) сборка пептидной цепи
в) удлинение пептида
г) завершение синтеза полипептида
Неинформативные нуклеотидные последовательности генов эукариотических клеток
а) экзоны
б) интроны
в) кодоны
г) репликоны
Фрагмент молекулы ДНК, включающий промотор, транскрибируемую последовательность и терминатор
а) репликон
б) мРНК
в) транскриптон
г) кодон
Вырожденность генетического кода
а) каждый триплет кодирует только одну аминокислоту
б) многие аминокислоты шифруются несколькими триплетами
в) каждый отдельный нуклеотид входит в состав только одного триплета
г) соседние триплеты не перекрывают друг друга
Дестабилизирующие белки в ходе репликации ДНК
а) активируют нуклеотиды, участвующие в синтезе новой цепи
б) участвуют в разрыве одной из цепей ДНК, ослабляя напряжение в двойной спирали
в) растягивают остовы цепей молекулы ДНК, делая доступными их для связывания азотистых оснований
г) участвуют в расплетании двойной спирали ДНК в точках начала репликации
Фермент топоизомераза
а) активирует нуклеотиды, участвующие в синтезе новой цепи
б) участвует в разрыве фосфорнодиэфирной связи одной из цепей ДНК, ослабляя напряжение в двойной спирали
в) разрывает водородные связи двух цепей молекулы ДНК, делая доступными их для связывания комплементарных азотистых оснований
г) участвуют в расплетании двойной спирали ДНК в точках начала репликации
Фермент геликаза
а) активирует нуклеотиды, участвующие в синтезе новой цепи
б) участвует в разрыве фосфорнодиэфирной связи одной из цепей ДНК, ослабляя напряжение в двойной спирали
в) разрывает водородные связи двух цепей молекулы ДНК, делая их доступными для связывания с комплементарными азотистыми основаниями
г) участвуют в расплетании двойной спирали ДНК в точках начала репликации
Цепь ДНК, имеющая 3/ конец, участвующая в репликации ДНК
а) лидирующая
б) кодогенная
в) консервативная
г) антисмысловая
Образуемые в ходе процессинга на 5`-концах мРНК колпачки (кэпы) обеспечивают
а) объединение 2-х субчастиц рибосом
б) «узнавание» молекул мРНК малыми субчастицами рибосом
в) образование комплекса аминоацил-тРНК
г) присоединение к стартовому кодону первой аминоацил-тРНК
В цитоплазме клеток содержится количество различных видов тРНК
а) 20
б) 40
в) 58
г) 61
Участвующий в негативном контроле транскрипции белок-регулятор
а) апоиндуктор
б) репрессор
в) ингибитор
г) супрессор
Участвующий в позитивном контроле транскрипции белок-регулятор
а) эффектор
б) интенсификатор
в) модификатор
г) апоиндуктор
Гены, ответственные за синтез белков общего назначения (белков мембран, рибосом)
а) модуляторы
б) конститутивные
в) регулируемые
г) функциональные
Негенетические факторы небелковой природы, регулирующие экспрессию генов
а) апоиндукторы
б) репрессоры
в) эффекторы
г) модификаторы
Эффекторы, запускающие транскрипцию
а) индукторы
б) апоиндукторы
в) активаторы
г) модуляторы
Эффекторы, запрещающие транскрипцию
а) репрессоры
б) корепрессоры
в) ингибиторы
г) индукторы
Свойство гена, обеспечивающее сохранность постоянства структуры при передаче из поколения в поколение
а) стабильность
б) специфичность действия
в) дискретность
г) «дозированность» действия
Кольцевая молекула ДНК прокариот упакована в виде
а) соленоида
б) нуклеосомы
в) доменов
г) линейной структуры
Однократная репликация ДНК в пределах одной хромосомы делает её структуру
а) однонитчатой
б) двухнитчатой
в) трёхнитчатой
г) четырёхнитчатой
Одна из особенностей строения молекулы ДНК препятствует одновременному синтезу
а) антипарпллельность
б) комплементарность
в) самоудвоение
г) стабильность
Единица транскрипции «транскриптон» представляет собой участок ДНК
а) промотор и структурная части гена (экзонов)
б) структурная часть гена (интроны,экзоны)и терминатор
в) промотор, структурная часть гена (интроны, экзоны) и терминатор
г) промотор,структурная часть гена (интроны,экзоны) и терминатор
Посттранскрипционные преобразования мРНК (процессинг) осуществляется в
а) цитоплазме клетки
б) ядре
в) рибосомах
г) ЭПС
Процессинг (созревание мРНК) в эукариотической клетке начинается с повтор
а) образования на переднем конце первичного транскрипта (5`-конце) колпачка(кэпа)
б) вырезания интронов и сшивания (сплайсинг)экзонов
в) метилирования азотистых оснований в транскрипте, стабилизирующих мРНК
г) формирования на 3`-конце транскрипта полиадениловой последовательностиА А А
Лактозный оперон E.coli включает в себя последовательности нуклеотидов
а) структурных генов Z,Y,A
б) промотора и структурных генов Z,Y,А
в) промотора,оператора,структурных генов Z,Y,А
г) оператора,структурных генов Z,Y,А
Гены «ответственные» за степень выраженности признака
а) гены-регуляторы
в) структурные гены
б) гены-модуляторы
г) гены-операторы
Гены «ответственные» за транскрибирование структурных генов
а) гены-регуляторы
в) структурные гены
б) гены-модуляторы
г) конститутивные гены
Свойство гена участвовать в разныхбиохимических процессах по формированию сложного признака
а) дозированность
б) дискретность
в) плейотропия
г) специфичность
Регуляторный белок связывается с нуклеотидными последовательностями
а) энхансера
б) промотора
в) блока Прибнова
г) энхансера и ТАТА-блока
Негативный контроль экспрессии генов осуществляется при участии белка-регулятора
а) апоиндуктора
б) индуктора
в) репрессора
г) корепрессора
Позитивный контроль экспрессии генов осуществляется при участии белка-регулятора
а) эффектора
б) апоиндуктора
в) активатора
г) индуктора
Гипотеза «Один ген-один фермент» была предложена
а) Бриджесом и Вольдейером
б) Уотсоном и Криком
в) Бидлом и Татумом
г) Бриджесом и Гальтоном
Однократная репликация ДНК в пределах одной хромосомы делает её структуру
а) однонитчатой
б) двухнитчатой
в) трёхнитчатой
г) четырёхнитчатой
Цепь ДНК, синтезируемая в ходе репликации отдельными фрагментами (Оказаки)
а) лидирующая
б) смысловая
в) антисмысловая
г) отстающая
Одна из особенностей строения молекулы ДНК препятствует одновременному синтезу двух её цепей при репликации
а) комплементарность
б) способность к образованию трёхмерной спирали
в) антипараллельность
г) сущесвование молекулы в двух вариантах:правозакрученной(В-ДНК) и левозакрученной(Z-ДНК)
После митоза хромосомы дочерней клетки содержат
а) одну молекулу ДНК
б) две молекулы ДНК
в) количество молекул соответствует содержанию их в профазе митоза
г) количество молекул соответствует содержанию их в анафазе митоза
Генетическая информация может считываться с участка ДНК в состоянии
а) компактизации(спирализации)
б) дезактивации
в) декомпактизации(деспирализации)
г) активации
Хромосомы типа ламповых щёток можно обнаружить в
а) овоцитах первого порядка
б) овогониях
в) яйцеклетках
г) слюнных железах насекомых
Фаза инициации (начала синтеза пептида)включает в себя процессы
а) объединения 2-х субчастиц рибосом и присоединения к ней первой аминоацил-тРНК
б) «созревания» мРНК и присоединение её к меньшей субчастице рибосомы
в) формирования в матриксе цитоплазмы третичной структуры т-РНК и образования аминоацил-тРНК
г) перемещения тРНК из аминоацильного участка рибосомы в пептидильный
Стартовому кодону и РНК соответствует сочетание нуклеотидов
а) УАГ
б) УАА
в) АУГ
г) УГА
Посттрансляционные преобразования белков осуществляются в
а) ядре клетки
б) ядре и цитоплазме
в) цитоплазме
г) в комплексе Гольджи
Нуклеосома-как один из уровней упаковки хроматина представляет собой
а) гистоновый кор с участком ДНК из 146 нуклеотидов
б) 4 пары гистоновых коров, соединённых линкерами
в) гистоновый кор, состоящий из 8 молекул гистонов и линкера(60 п.н)
г) компактное образование, состоящее из 4-х гистоновых коров, соединённых Н1 фракцией гистонов
Комплекс ДНК с негистоновыми белками прокариот называется
а) нуклеопротеиновым
б) нуклеосомным
в) нуклеоидным
г) протеиноидным
Кольцевая молекула ДНК прокариот упакована в виде
а) соленоида
б) нуклеосомы
в) петель
г) плотной гладкой структуры
Если в образовавшейся после митоза клетке одна молекула ДНК «материнская»,а другая «дочерняя, способ репликации генетического материала называется
а) консервативным
б) матричным
в) полуконсервативным
г) дисперсионным
Рибонуклеопротеиновый комплекс является составной частью
а) ядрышка ядра
б) кариоплазмы
в) хроматина
г) порового комплекса
studfiles.net
Молекулярные основы наследственности
До 40-х годов нашего столетия большинство ученых считали, что гены имеют белковую природу. Выдающийся русский исследователь Н. К. Кольцов высказал мысль о том, что хромосома — это гигантская биологическая молекула, обладающая свойством самоудвоения, и что все признаки и свойства организма обусловлены строением белка и взаимодействием его молекул. В 1927 г. Н. К. Кольцов подготовил базу для понимания ауторепродукции хромосом, что составляет в настоящее время основу молекулярной генетики. Казалось вероятным, что именно в белках заключена наследственная информация о развитии всех признаков и свойств организма. Однако проведенные в последующем эксперименты на микроорганизмах с применением новейших методов исследований, рентгеноструктурного анализа, электронной микроскопии, меченых атомов и т. д. позволили установить, что генетическая информация сосредоточена в нуклеиновых кислотах.
ДОКАЗАТЕЛЬСТВА РОЛИ ДНК В НАСЛЕДСТВЕННОСТИ
В 1928 г. Ф. Гриффит впервые получил^ доказательства возможной передачи наследственных задатков от одной бактерии к другой. Ученый вводил мышам вирулентный капсульный и ави-рулентный бескапсульный штаммы пневмококков. При введении вирулентного штамма мыши заболевали пневмонией и погибали. При введении авирулентного штамма мыши оставались живыми. При введении вирулентного капсульного штамма, убитого нагреванием, мыши также не погибали. В следующем опыте он ввел смесь живой культуры авирулентного бескапсульного штамма со штаммом убитого нагреванием вирулентного капсульного и получил неожиданный результат — мыши заболели пневмонией и погибли. Из крови погибших животных были выделены бактерии, которые обладали вирулентностью и были способны образовать капсулу. Следовательно, живые бактерии авирулентного бескапсульного штамма трансформировались — приобрели свойства убитых болезнетворных бактерий. В дальнейшем другими учеными были подтверждены результаты опытов Ф. Гриффита в условиях пробирки. Основываясь на этих опытах, в 1944 г.
74
О. Эвери и его сотрудники Мак-Леод и Мак-Карти изучили роль разных веществ клетки в явлениях трансформации и получили убедительные доказательства того, что трансформирующим фактором является дезоксирибонуклеиновая кислота (ДНК). Было установлено, что под действием дезоксирибонуклеазы — фермента, специфически разрушающего ДНК, активность трансформирующего фактора исчезла. В то же время рибонуклеаза и протео-литические ферменты не изменяли биологической активности трансформирующего фактора.
Следующим доказательством генетической роли ДНК были эксперименты А. Херши и М. Чейза, проведенные с бактериофагом в 1952 г. Основные компоненты фага — ДНК и белок. ДНК фага была помечена радиоактивным фосфором (32Р), который включается только в ДНК. Белок фага пометили с помощью радиоактивной серы г%), которая включается только в белок. После заражения бактерий мечеными фагами было установлено, что в клетку бактерии проникает только молекула ДНК, а белковая оболочка фага остается снаружи. Тем не менее в клетках зараженных бактерий образовалось множество зрелых частиц фага. Это говорило о том, что в ДНК заключена наследственная информация о всех признаках и свойствах фага. Опыты А. Херши и М. Чейза еще раз подтвердили, что наследственная информация заключена в молекулах ДНК и передается ими по наследству. В последующем было установлено, что у некоторых прокариот наследственная информация зашифрована в молекулах РНК.
БИОЛОГИЧЕСКАЯ РОЛЬ НУКЛЕИНОВЫХ КИСЛОТ
Генетическая информация реализуется в процессе биосинтеза белков. Все основные свойства живых существ определяются структурой и функцией белковых молекул. В последние 40 лет в ряде лабораторий разных стран мира было выяснено, что синтез специфических белков предопределен генетически. Материальным субстратом наследственности является ДНК. В молекулах ДНК зашифрована наследственная информация о строении каждого белка. ДНК обеспечивает хранение и передачу генетической информации из поколения в поколение. Участок молекулы ДНК, кодирующий первичную структуру полипептида, молекулы транспортной или рибосомной РНК, называется геном. Реализация наследственной информации осуществляется с участием рибонуклеиновых кислот (РНК).
Белки — структурная основа всех клеток, органов и тканей организма. В сочетании с другими веществами они участвуют в формировании различного рода клеточных структур.
Многочисленными исследованиями установлено, что белки различаются как у отдельных видов микроорганизмов, растений и животных, так и в пределах одного вида. Главные структурные
75
элементы белковых молекул — 20 аминокислот. Специфика строения белковой молекулы определяется наличием определенных аминокислот и порядком их расположения в полипептидных цепях. К настоящему времени достигнуты значительные успехи в раскрытии химической структуры различных белков и полипептидов. Рассмотрим, как влияет содержание аминокислот и их чередование в полипептидных цепях гормонов гипофиза оксито-цина и вазопрессина на их биологическую роль. Эти гормоны включают по 9 аминокислотных остатков:
окситоцин: цистеил — тирозил — изолейцил — глутамил — ас-парагил — цистеил — пролил — лейцил — глицин;
вазопрессин: цистеил — тирозил — фенилаланил — глутамил — аспарагил — цистеил — пролил — аргинил — глицин. Как видим, разница состоит только в том, что в окситоцине на третьем месте стоит аминокислота изолейцин, на восьмом лейцин, а у вазопрессина соответственно фенилаланин и аргинин. Эти небольшие изменения обусловили разную биологическую роль гормонов: окситоцин вызывает сокращение матки во время родов, а вазопрессин увеличивает кровяное давление.
Обнаружено, что виды, породы и отдельные индивидуумы имеют незначительные специфические отличия в строении ферментов и других белковых молекул, выполняющих одинаковые функции.
Однако имеется много случаев, когда незначительные изменения в структуре белка приводят к серьезным последствиям. Так, известно, что от 5 до 20 % коренного населения Африки, Индии и Средиземноморских стран имеют аномальный гемоглобин S, который отличается от нормального гемоглобина А только по одной аминокислоте. Ниже приведено чередование аминокислот на определенном участке гемоглобина А и S:
гемоглобин А —лей-тре-про-глу-глу-лиз;
гемоглобин S —лей-тре-про-вал-глу-лиз.
Незначительные изменения в строении гемоглобина являются причиной тяжелого наследственного заболевания — серповидно-клеточной анемии. Эритроциты больных серповидноклеточной анемией имеют форму серпа, а не округлую, как в норме.
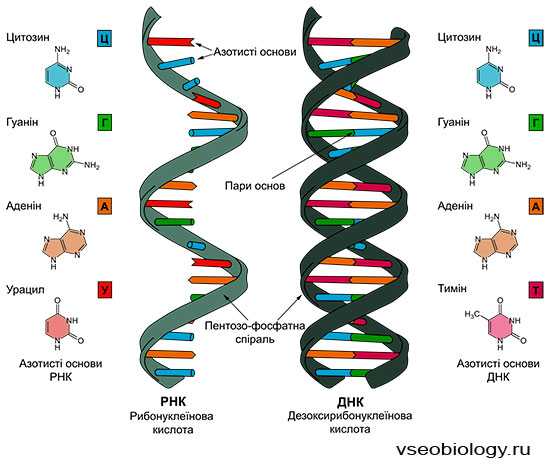
ХИМИЧЕСКИЙ СОСТАВ И СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Нуклеиновые кислоты впервые открыл И. Ф. Мишер в 1868 г. Он выделил из ядер клеток особое вещество кислотной природы и назвал его нуклеином. Впоследствии ему дали название «нуклеиновая кислота». Было обнаружено два типа нуклеиновых кислот. Их назвали в зависимости от углеводного компонента, входящего в состав. Нуклеиновую кислоту, в состав которой входит углевод дезоксирибоза, назвали дезоксирибонуклеиновой кислотой (ДНК), а в состав которой входит углевод рибоза, — рибонуклеино-
76
вой кислотой (РНК). В период с 1900 по 1932 г. был определен химический состав нуклеиновых кислот. Они включают следующие компоненты:
РНК
Аденин, гуанин Цитозин, урацил Рибоза
ДНК
Аденин, гуанин Цитозин, тимин Дезоксирибоза
Пуриновые основания Пиримидиновые основания Углеводный компонент
Обе нуклеиновые кислоты включают остатки фосфорной кислоты. Различие заключается в том, что в состав РНК входит азотистое основание урацил вместо тимина и рибоза вместо дезоксирибозы.
В 1936 г. на кафедре биохимии растений Московского университета А. Н. Белозерский с И. И. Дубровской впервые выделили ДНК в чистом виде из растительного материала. К середине 40-х годов было выяснено, что ДНК и РНК одновременно присутствуют в каждом живом организме.
В конце 40-х — начале 50-х годов при изучении нуклеиновых кислот стали использовать новые физические и химические методы исследования. В 1950 г. Э. Чаргафф установил правила нукле-отидных отношений, лежащие в основе строения всех ДНК.
Правила Чаргаффа заключаются в том, что в ДНК содержание аденина равно содержанию тимина (А = Т), а содержание гуанина равно содержанию цитозина (Г = И), отсюда А + Г/Т + Ц = 1; сумма пуриновых нуклеотидов равна сумме пиримидиновых нуклео-тидов. В соответствии с этим правилом нуклеотидный состав разных организмов может варьировать только по величине А+Т/Г+Ц.
К 1952 г. Р. Франклин и М. Уилкинс добились получения высококачественных рентгенограмм ДНК, показавших, что она имеет форму спирали и двойственную структуру.
В 1953 г. Дж. Уотсон и Ф. Крик, опираясь на данные рентге-ноструктурного анализа и правила Чаргаффа, установили структуру ДНК. Согласно их модели молекула ДНК имеет двойную спираль, состоящую из двух полинуклеотидных цепей с общей осью (рис. 17). Диаметр двойной спирали ДНК равен 2 нм, а расстояние между витками 3,4 нм. На каждый виток спирали приходится 10 пар нуклеотидов, отсюда расстояние между азотистыми основаниями равно 0,34 нм.
Структурными единицами полинуклеотидных цепей являются нуклеотиды. В состав нуклеотида входят: одно из азотистых оснований — пуриновое (аденин или гуанин) или пиримидиновое (тимин или цитозин), дезоксирибоза, фосфатный остаток. Эти компоненты соединены друг с другом в следующем порядке: азотистое основание — дезоксирибоза — фосфатный остаток. Соединение одного из оснований с дезоксирибозой приводит к образованию нуклеозида. В случае присоединения фосфатной группы к углеводной части нуклеозида образуется нуклеотид.
77
Дезоксирибоза в нуклеотидах соединяется с основаниями гли-козидной связью, а с фосфорной кислотой — эфирными связями. Следовательно, по химическому составу любой нуклеотид — это фосфорный эфир нуклеозидов. В соответствии с этим нукле-отиды называются дезоксиадениловой, дезоксигуаниловой, дезокси-цитидиловой и тимидиловой кислотами.
Наряду с главными азотистыми основаниями ДНК содержит также метилированные основания, такие, как 5-метилцитозин, 5-оксиметилцитозин и др. У животных количество 5-метилцито-зина в ДНК обычно не превышает 1,5—2 %.
В каждой из цепей ДНК нуклеотиды последовательно соединены друг с другом с помощью остатка фосфорной кислоты и молекулы дезоксирибозы. Дезоксирибоза связывается с одной молекулой фосфорной кислоты через углерод в положении 3′, а с другой — через углерод 5′, образуя углеводно-фосфатный остов (рис. 18).
Обе цепи в молекуле ДНК имеют противоположную полярность. Это означает, что межнуклеотидная связь в одной цепи имеет направление 5′-»3′, а в другой

комплементарностью, и, следовательно, цепи в ДНК комплементарны, они взаимно дополняют друг друга.
Углеводно-фосфатный остов по всей длине во всех молекулах ДНК имеет однотипную структуру и не может нести генетической информации. В противоположность этому рас-
Рнс. 17. Схема строения молекулы ДНК из двух спирально закрученных цепей (по Д. Уотсо-ну и Ф. Крику) (Цифры указывают на расстоя-
ния ■ А между разными точками молекулы)
78
Рис. 18. Схема отрезка двухцепочной молекулы ДНК (по С. М. Гершензону)
положение пуриновых и пиримидиновых оснований нуклеотидов вдоль цепи ДНК очень изменчиво и характерно для каждого данного типа молекул ДНК. Значит, наследственная информация зашифрована различной последовательностью оснований.
Нуклеотидный состав ДНК значительно варьирует в зависимости от принадлежности организма к той или иной систематической группе (табл. 7). Специфичность ДНК выражается соотношением А + Т/Г + Ц, получившим название коэффициента видовой специфичности.
|
7. Нуклеотидный состав ДНК из различных (по А. Ленинджеру, 1976) |
ИСТОЧНИКОВ | ||||||
|
Нуклеотидный состав, |
мол. % |
Коэффициент | |||||
|
Объект |
А |
Г |
Ц |
т |
специфичности А + Т/Г + Ц | ||
30,9 19,9 19,8 29,4 1,52
Человек Животные:
28,3 1,36
29,2 1,38
27.1 1,19 32,9 1,79 24,9 1,00
23,6 0,93
1,50
2,70 21,1 0,72
0,35
|
29,3 |
21,4 |
21,0 |
|
28,8 |
20,5 |
21,5 |
|
27,3 |
22,7 |
22,8 |
|
31,3 |
18,7 |
17,1 |
|
25,0 |
25,1 |
25,0 |
|
24,7 |
26,0 |
25,7 |
|
30,8 |
21,0 |
19,0 |
|
36,9 |
14,0 |
12,8 |
|
21,0 |
29,0 |
28,9 |
|
13,4 |
37,1 |
37,1 |
овиа
курица
Растения, грибы, зерна пшеницы Дрожжи
Aspergillus niger Бактерии:
Е. coli
Staphylococcus ayreus
Clostridium perfinngens
Bnicella abortus
Sarcina lutea
79
топоизомераза
В ДНК животных наблюдается избыток А + Т по отношению к Г + Ц. У грибов и бактерий встречаются формы как богатые А + Т, так и с преобладанием Г + Ц, в то же время есть близкие по коэффициенту специфичности к животным. Это говорит о том, что изменчивость в расположении оснований уже достаточна для того, чтобы обеспечить различия между генами этих организмов.Молекулы ДНК состоят примерно из 2-Ю3— МО8 и более нуклеотидов и имеют большую относительную молекулярную массу.
Репликация (удвоение) ДНК. ДНК находится в хромосомах, и репликация ее происходит перед каждым удвоением хромосом и делением клетки. Дж. Уотсон и Ф. Крик предложили схему удвоения ДНК, согласно которой спиралевидная двухцепочная ДНК сначала раскручивается (расплетается) вдоль оси. При этом водородные связи между азотистыми основаниями рвутся и цепи расходятся. Одновременно к нуклеотидам каждой цепи пристраиваются комплементарные азотистые основания нуклеотидов второй цепи, где против аденина встает тимин, против тимина — аденин, против гуанина — цитозин и т. д., которые с помощью ферментов ДНК-полимераз связываются в новые полинуклеотидные цепи. В результате из одной образуются две новые дочерние молекулы ДНК. Каждая дочерняя молекула, наследуя структуру одной цепи материнской молекулы, строго сохраняет специфичность заключенной в ней информации. Поскольку матрицей для репликации служит одна из двух цепей молекулы, такой тип синтеза ДНК носит название полуконсервативной ауторепродукции.
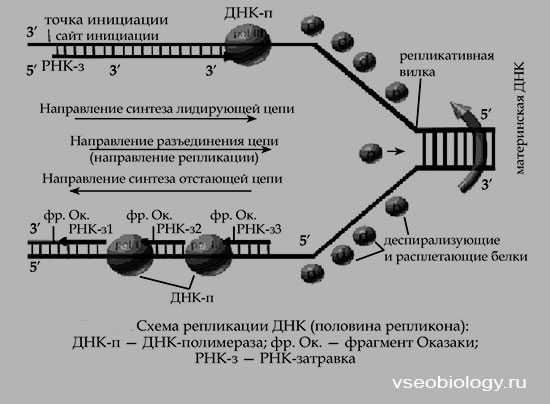
Дальнейшие исследования показали, что репликация бактериальных и других молекул ДНК начинается в определенной точке старта. В хромосомах эукариот обнаружено по нескольку таких начальных точек. Цепи ДНК в точке инициации репликации разъединяются под влиянием особого белка геликазы (рис. 19). Возникают одноцепочные участки ДНК, которые становятся матрицами для репликации-притяжения комплементарных нуклеотидов. Эти одноцепочные участки связываются с особыми белками, которые их стабилизируют (препятствуют их комплементарному взаимодействию). Особый фермент топоизомераза (у прокариот назьгаается ДНК-гиразой) способствует расщеплению спирали ДНК в области репликационной вилки.
Репликация на материнской цепи, идущей от точки старта в направлении 5′-*3′, идет в виде сплошной линии. Эта цепь получила название лидирующей. Синтез на второй цепи 3′->5′ идет отдельными фрагментами в противоположном направлении (тоже 5′-»3′)- Эта цепь получила название запаздывающей. Фрагментами являются небольшие участки ДНК (у кишечной палочки около 2000 нуклеотидов, у эукариот около 200). Они называются по
80
фрагменты Окозаки
Рис. 19. Синтез лидирующей (вверху) и запаздывающей (внизу) цепей ДНК в вилке репликации (по В. Alberts, R. Sternglanz)
имени открывшего их японского ученого Р. Оказаки. После завершения синтеза фрагменты Оказаки соединяются при помощи фермента лигазы в общую полинуклеотидную цепочку. У эукариот репликация ДНК и соединение различных ее репликационных участков происходят в фазе S-периода интерфазы. После завершения этой фазы в каждой хромосоме имеется две молекулы ДНК, которые становятся двумя идентичными хроматидами.
Структура, способная к репликации (хромосома, плазмида, вирусный геном), называется репликоном.
Самоудвоение молекул ДНК — основа устойчивости генетической информации данного вида и обеспечивает материальную непрерывность наследственного вещества клетки.
studfiles.net
Лекция Молекулярные основы наследственности Биосинтез белка Транскрипция 1
 Лекция Молекулярные основы наследственности. Биосинтез белка. Транскрипция. 1
Лекция Молекулярные основы наследственности. Биосинтез белка. Транскрипция. 1
 План 1. Организация наследственного материала. а) Историческое развитие концепции гена. б) Фукционально-генетическая классификация генов. 2. Особенности транскрипции. 3. Транскрипция у прокариот. Регуляция транскрипции у прокариот. Схема работы лактозного оперона. 4. Транскрипция у эукариот. Регуляция транскрипции у эукариот. 2
План 1. Организация наследственного материала. а) Историческое развитие концепции гена. б) Фукционально-генетическая классификация генов. 2. Особенности транскрипции. 3. Транскрипция у прокариот. Регуляция транскрипции у прокариот. Схема работы лактозного оперона. 4. Транскрипция у эукариот. Регуляция транскрипции у эукариот. 2
 1. Организация наследственного материала. а) Историческое развитие концепции гена. б) Фукционально-генетическая классификация генов.
1. Организация наследственного материала. а) Историческое развитие концепции гена. б) Фукционально-генетическая классификация генов.
 Организация наследственного материала Современные представления о природе генетического аппарата позволяют выделить три уровня его организации: • Генный • Хромосомный • Геномный
Организация наследственного материала Современные представления о природе генетического аппарата позволяют выделить три уровня его организации: • Генный • Хромосомный • Геномный
 Геномный уровень организации наследственного материала • Геном это совокупность наследственного материала в гаплоидном наборе хромосом клеток организмов соответствующего биологического вида. • Геном видоспецифичен. • Геном сбалансированный, эволюционно «проработанной» и отобранный объем генетической информации. • С геномным уровнем генетического аппарата эукариот связывают переход к избирательной транскрипции генов по времени (период онтогенеза), месту (тип клеток) и объему.
Геномный уровень организации наследственного материала • Геном это совокупность наследственного материала в гаплоидном наборе хромосом клеток организмов соответствующего биологического вида. • Геном видоспецифичен. • Геном сбалансированный, эволюционно «проработанной» и отобранный объем генетической информации. • С геномным уровнем генетического аппарата эукариот связывают переход к избирательной транскрипции генов по времени (период онтогенеза), месту (тип клеток) и объему.
 Хромосомный уровень организации наследственного материала Связан с его большим объемом по сравнению с прокариотической клеткой. • Распределение основной массы генетического материала в ограниченном числе ядерных структур – хромосом – обеспечивает упорядоченность его пространственной организации по группам сцепления (хромосомам – хромосом и закономерное пространственное взаиморасположение хромосом в клеточном ядре. •
Хромосомный уровень организации наследственного материала Связан с его большим объемом по сравнению с прокариотической клеткой. • Распределение основной массы генетического материала в ограниченном числе ядерных структур – хромосом – обеспечивает упорядоченность его пространственной организации по группам сцепления (хромосомам – хромосом и закономерное пространственное взаиморасположение хромосом в клеточном ядре. •
 Генный уровень организации наследственного материала. • Ген – это единица наследственности и изменчивости. • По современным представлениям ген – это участок молекулы ДНК, кодирующий информацию о синтезе определенного полипептида или нуклеиновой кислоты.
Генный уровень организации наследственного материала. • Ген – это единица наследственности и изменчивости. • По современным представлениям ген – это участок молекулы ДНК, кодирующий информацию о синтезе определенного полипептида или нуклеиновой кислоты.
 а) Историческое развитие концепции гена
а) Историческое развитие концепции гена
 б) Фукционально-генетическая классификация генов.
б) Фукционально-генетическая классификация генов.
 2. Особенности транскрипции.
2. Особенности транскрипции.
 Так выглядит транскрипция – синтез РНК для последующего производства белка http: //www. vechnayamolodost. ru/pages/drugienaukiozhizni/rodkoodnrntrimndr 80. html
Так выглядит транскрипция – синтез РНК для последующего производства белка http: //www. vechnayamolodost. ru/pages/drugienaukiozhizni/rodkoodnrntrimndr 80. html
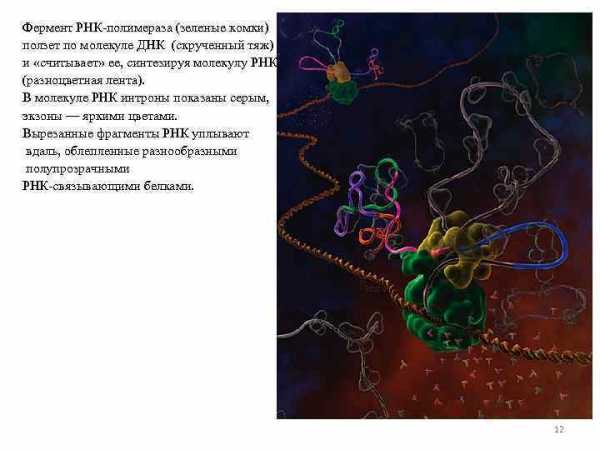 Фермент РНК-полимераза (зеленые комки) ползет по молекуле ДНК (скрученный тяж) и «считывает» ее, синтезируя молекулу РНК (разноцветная лента). В молекуле РНК интроны показаны серым, экзоны — яркими цветами. Вырезанные фрагменты РНК уплывают вдаль, облепленные разнообразными полупрозрачными РНК-связывающими белками. 12
Фермент РНК-полимераза (зеленые комки) ползет по молекуле ДНК (скрученный тяж) и «считывает» ее, синтезируя молекулу РНК (разноцветная лента). В молекуле РНК интроны показаны серым, экзоны — яркими цветами. Вырезанные фрагменты РНК уплывают вдаль, облепленные разнообразными полупрозрачными РНК-связывающими белками. 12
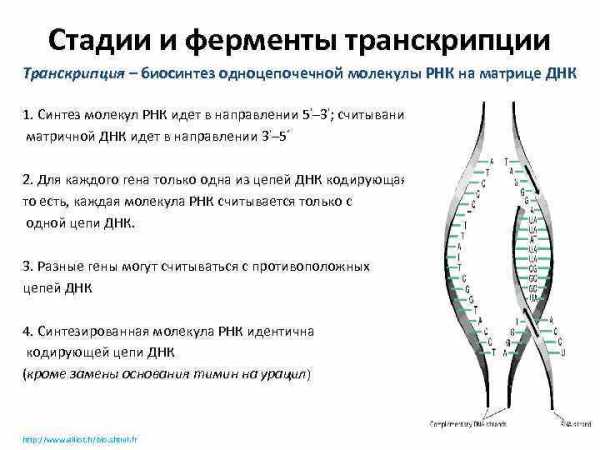 Стадии и ферменты транскрипции Транскрипция – биосинтез одноцепочечной молекулы РНК на матрице ДНК 1. Синтез молекул РНК идет в направлении 5’– 3′; считывание матричной ДНК идет в направлении 3’– 5‘ 2. Для каждого гена только одна из цепей ДНК кодирующая, то есть, каждая молекула РНК считывается только с одной цепи ДНК. 3. Разные гены могут считываться с противоположных цепей ДНК 4. Синтезированная молекула РНК идентична кодирующей цепи ДНК (кроме замены основания тимин на урацил) http: //www. alliot. fr/bio. shtml. fr 13
Стадии и ферменты транскрипции Транскрипция – биосинтез одноцепочечной молекулы РНК на матрице ДНК 1. Синтез молекул РНК идет в направлении 5’– 3′; считывание матричной ДНК идет в направлении 3’– 5‘ 2. Для каждого гена только одна из цепей ДНК кодирующая, то есть, каждая молекула РНК считывается только с одной цепи ДНК. 3. Разные гены могут считываться с противоположных цепей ДНК 4. Синтезированная молекула РНК идентична кодирующей цепи ДНК (кроме замены основания тимин на урацил) http: //www. alliot. fr/bio. shtml. fr 13
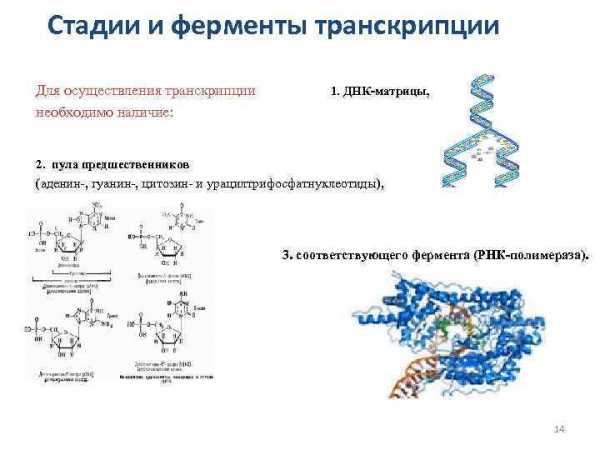 Стадии и ферменты транскрипции Для осуществления транскрипции необходимо наличие: 1. ДНК-матрицы, 2. пула предшественников (аденин-, гуанин-, цитозин- и урацилтрифосфатнуклеотиды), 3. соответствующего фермента (РНК-полимераза). 14
Стадии и ферменты транскрипции Для осуществления транскрипции необходимо наличие: 1. ДНК-матрицы, 2. пула предшественников (аденин-, гуанин-, цитозин- и урацилтрифосфатнуклеотиды), 3. соответствующего фермента (РНК-полимераза). 14
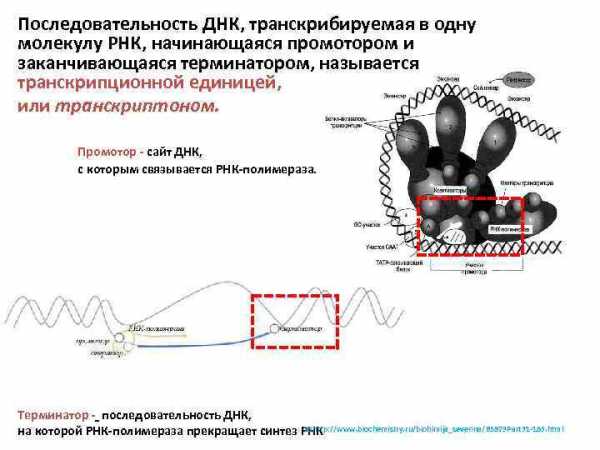 Последовательность ДНК, транскрибируемая в одну молекулу РНК, начинающаяся промотором и заканчивающаяся терминатором, называется транскрипционной единицей, или транскриптоном. Промотор — сайт ДНК, с которым связывается РНК-полимераза. Терминатор — последовательность ДНК, 15 http: //www. biochemistry. ru/biohimija_severina/B 5873 Part 31 -185. html на которой РНК-полимераза прекращает синтез РНК
Последовательность ДНК, транскрибируемая в одну молекулу РНК, начинающаяся промотором и заканчивающаяся терминатором, называется транскрипционной единицей, или транскриптоном. Промотор — сайт ДНК, с которым связывается РНК-полимераза. Терминатор — последовательность ДНК, 15 http: //www. biochemistry. ru/biohimija_severina/B 5873 Part 31 -185. html на которой РНК-полимераза прекращает синтез РНК
 Транскрипция – матричный процесс, в котором выделяют три стадии Стадии транскрипции ИНИЦИАЦИЯ (самая медленная стадия) промотор ЭЛОНГАЦИЯ Процессы, происходящие на стадиях транскрипции 1. 2. 3. Связывание РНК-полимеразы с ДНК Расплетание ДНК на участке 10 -20 нуклеотидов Формирование первых фосфодиэфирных связей Удлинение цепи РНК (самая длительная стадия) ТЕРМИНАЦИЯ (самая короткая стадия) терминатор 1. Остановка синтеза РНК 2. Распад тройного комплекса ДНК—-РНК-полимераза——РНК 16
Транскрипция – матричный процесс, в котором выделяют три стадии Стадии транскрипции ИНИЦИАЦИЯ (самая медленная стадия) промотор ЭЛОНГАЦИЯ Процессы, происходящие на стадиях транскрипции 1. 2. 3. Связывание РНК-полимеразы с ДНК Расплетание ДНК на участке 10 -20 нуклеотидов Формирование первых фосфодиэфирных связей Удлинение цепи РНК (самая длительная стадия) ТЕРМИНАЦИЯ (самая короткая стадия) терминатор 1. Остановка синтеза РНК 2. Распад тройного комплекса ДНК—-РНК-полимераза——РНК 16
 3. Транскрипция у прокариот. Регуляция транскрипции у прокариот. Схема работы лактозного оперона.
3. Транскрипция у прокариот. Регуляция транскрипции у прокариот. Схема работы лактозного оперона.
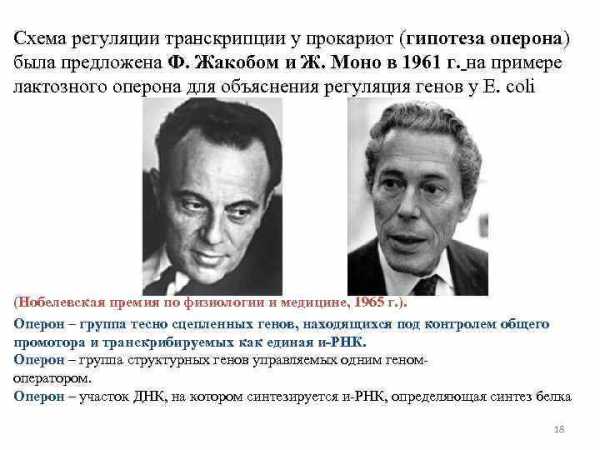 Схема регуляции транскрипции у прокариот (гипотеза оперона) была предложена Ф. Жакобом и Ж. Моно в 1961 г. на примере лактозного оперона для объяснения регуляция генов у E. coli (Нобелевская премия по физиологии и медицине, 1965 г. ). Оперон – группа тесно сцепленных генов, находящихся под контролем общего промотора и транскрибируемых как единая и-РНК. Оперон – группа структурных генов управляемых одним геномоператором. Оперон – участок ДНК, на котором синтезируется и-РНК, определяющая синтез белка 18
Схема регуляции транскрипции у прокариот (гипотеза оперона) была предложена Ф. Жакобом и Ж. Моно в 1961 г. на примере лактозного оперона для объяснения регуляция генов у E. coli (Нобелевская премия по физиологии и медицине, 1965 г. ). Оперон – группа тесно сцепленных генов, находящихся под контролем общего промотора и транскрибируемых как единая и-РНК. Оперон – группа структурных генов управляемых одним геномоператором. Оперон – участок ДНК, на котором синтезируется и-РНК, определяющая синтез белка 18
 Регуляция транскрипции у прокариот В состав оперона входит: 1. Промотор 2. Инициатор 3. Ген-оператор 4. Структурные гены 5. Терминатор Ген-регулятор не является частью оперона, он активен постоянно и на основе его информации через и-РНК синтезируется особый белокрепрессор. Белок-репрессор связывается индуктором. Индуктор – вещество, инициирующее синтез фермента, который его разлагает. 19
Регуляция транскрипции у прокариот В состав оперона входит: 1. Промотор 2. Инициатор 3. Ген-оператор 4. Структурные гены 5. Терминатор Ген-регулятор не является частью оперона, он активен постоянно и на основе его информации через и-РНК синтезируется особый белокрепрессор. Белок-репрессор связывается индуктором. Индуктор – вещество, инициирующее синтез фермента, который его разлагает. 19
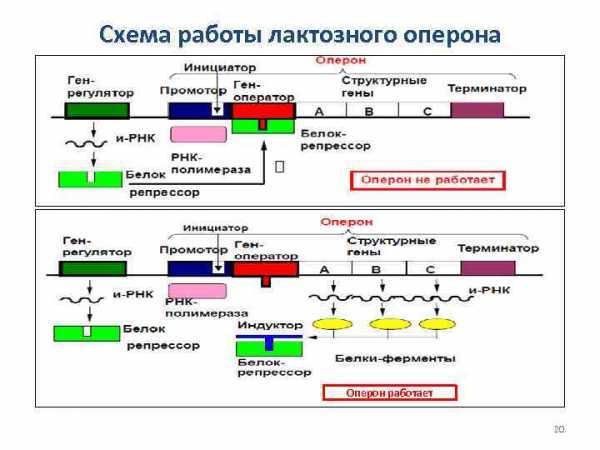 Схема работы лактозного оперона Оперон работает 20
Схема работы лактозного оперона Оперон работает 20
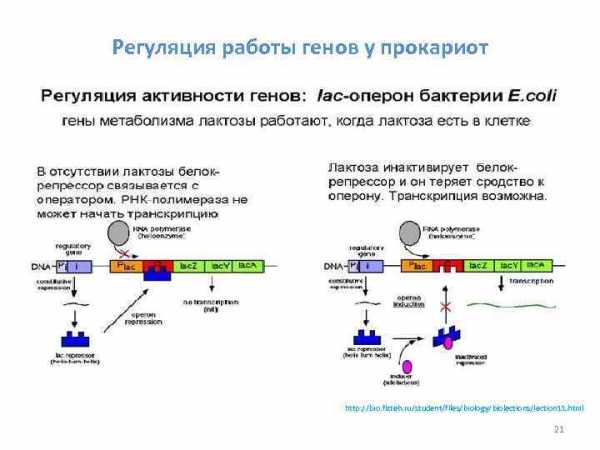 Регуляция работы генов у прокариот http: //bio. fizteh. ru/student/files/biology/biolections/lection 11. html 21
Регуляция работы генов у прокариот http: //bio. fizteh. ru/student/files/biology/biolections/lection 11. html 21
 4. Транскрипция у эукариот. Регуляция транскрипции у эукариот.
4. Транскрипция у эукариот. Регуляция транскрипции у эукариот.
 Схема регуляции транскрипции у эукариот разработана Георгием Павловичем Георгиевым (1972 г. ) и получила название гипотезы транскриптона. Единица транскрипции у эукариот также транскриптон Принцип регуляции (обратная связь) сохраняется, но механизмы ее более сложные. В прокариотической клетке наследственный материал и аппарат биосинтеза белка пространственно не разобщены, поэтому транскрипция и трансляция происходят почти одновременно. У эукариот транскрипция происходит в ядре и сопровождается процессингом пре-РНК транскрипта, трансляция — в 23 цитоплазме на рибосомах.
Схема регуляции транскрипции у эукариот разработана Георгием Павловичем Георгиевым (1972 г. ) и получила название гипотезы транскриптона. Единица транскрипции у эукариот также транскриптон Принцип регуляции (обратная связь) сохраняется, но механизмы ее более сложные. В прокариотической клетке наследственный материал и аппарат биосинтеза белка пространственно не разобщены, поэтому транскрипция и трансляция происходят почти одновременно. У эукариот транскрипция происходит в ядре и сопровождается процессингом пре-РНК транскрипта, трансляция — в 23 цитоплазме на рибосомах.
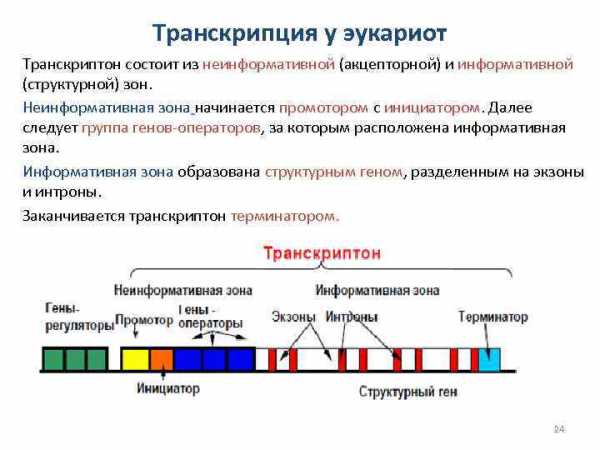 Транскрипция у эукариот Транскриптон состоит из неинформативной (акцепторной) и информативной (структурной) зон. Неинформативная зона начинается промотором с инициатором. Далее следует группа генов-операторов, за которым расположена информативная зона. Информативная зона образована структурным геном, разделенным на экзоны и интроны. Заканчивается транскриптон терминатором. 24
Транскрипция у эукариот Транскриптон состоит из неинформативной (акцепторной) и информативной (структурной) зон. Неинформативная зона начинается промотором с инициатором. Далее следует группа генов-операторов, за которым расположена информативная зона. Информативная зона образована структурным геном, разделенным на экзоны и интроны. Заканчивается транскриптон терминатором. 24
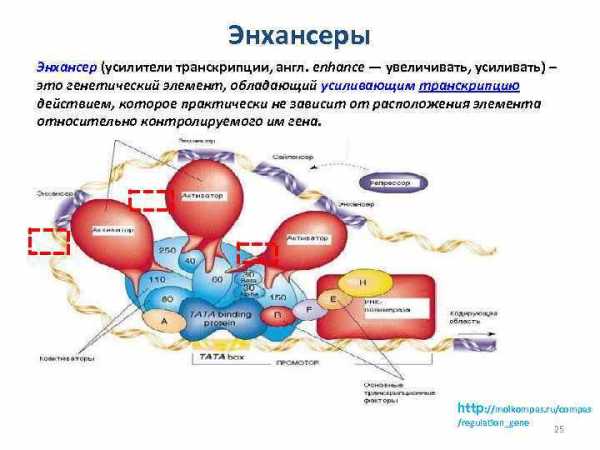 Энхансеры Энхансер (усилители транскрипции, англ. enhance — увеличивать, усиливать) – это генетический элемент, обладающий усиливающим транскрипцию действием, которое практически не зависит от расположения элемента относительно контролируемого им гена. http: //moikompas. ru/compas /regulation_gene 25
Энхансеры Энхансер (усилители транскрипции, англ. enhance — увеличивать, усиливать) – это генетический элемент, обладающий усиливающим транскрипцию действием, которое практически не зависит от расположения элемента относительно контролируемого им гена. http: //moikompas. ru/compas /regulation_gene 25
 Энхансеры представлены короткими последовательностями ДНК, включающих десятки нуклеотидных пар. Энхансер увеличивает эффективность транскрипции гена в десятки и сотни раз. Энхансеры способны активировать гены на больших расстояниях, достигающих нескольких десятков тысяч пар нуклеотидов. В некоторых случаях они могут активировать транскрипцию генов, расположенных на других хромосомах. Энхансер увеличивает эффективность транскрипции гена в десятки и сотни раз. Особенность энхансеров состоит в том, что они способны действовать на больших расстояниях (более чем 1000 п. н. ) и вне зависимости от ориентации по отношению к направлению транскрипции гена. Еще одной важной характеристикой энхансера является его способность активировать любой промотор, расположенный сравнительно недалеко. Энхансеры — основное средство регуляции транскрипции в клетках высших эукариот. Для взаимодействия энхансера с промотором необходимо приблизить эти сайты друг к другу. Возможность такого контакта может определяться другими регуляторными участками, формирующими петлевую укладку хроматина.
Энхансеры представлены короткими последовательностями ДНК, включающих десятки нуклеотидных пар. Энхансер увеличивает эффективность транскрипции гена в десятки и сотни раз. Энхансеры способны активировать гены на больших расстояниях, достигающих нескольких десятков тысяч пар нуклеотидов. В некоторых случаях они могут активировать транскрипцию генов, расположенных на других хромосомах. Энхансер увеличивает эффективность транскрипции гена в десятки и сотни раз. Особенность энхансеров состоит в том, что они способны действовать на больших расстояниях (более чем 1000 п. н. ) и вне зависимости от ориентации по отношению к направлению транскрипции гена. Еще одной важной характеристикой энхансера является его способность активировать любой промотор, расположенный сравнительно недалеко. Энхансеры — основное средство регуляции транскрипции в клетках высших эукариот. Для взаимодействия энхансера с промотором необходимо приблизить эти сайты друг к другу. Возможность такого контакта может определяться другими регуляторными участками, формирующими петлевую укладку хроматина.
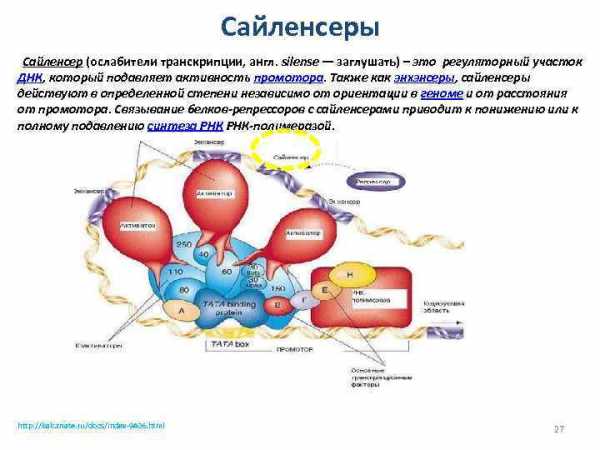 Сайленсеры Сайленсер (ослабители транскрипции, англ. silense — заглушать) – это регуляторный участок ДНК, который подавляет активность промотора. Также как энхэнсеры, сайленсеры действуют в определенной степени независимо от ориентации в геноме и от расстояния от промотора. Связывание белков-репрессоров с сайленсерами приводит к понижению или к полному подавлению синтеза РНК-полимеразой. http: //kak. znate. ru/docs/index-9406. html 27
Сайленсеры Сайленсер (ослабители транскрипции, англ. silense — заглушать) – это регуляторный участок ДНК, который подавляет активность промотора. Также как энхэнсеры, сайленсеры действуют в определенной степени независимо от ориентации в геноме и от расстояния от промотора. Связывание белков-репрессоров с сайленсерами приводит к понижению или к полному подавлению синтеза РНК-полимеразой. http: //kak. znate. ru/docs/index-9406. html 27
 Сайленсеры Ингибирование транскрипции с использованием регуляторных элементов, называемых сайленсерами, — активный процесс. В этом случае происходит прямое подавление инициации транскрипции путем разрушения транскрипционного комплекса на промоторе или посредством его инактивации иным способом. Первый из описанных в 1986 г. сайленсеров обладал классическими энхансероподобными свойствами, действуя на промоторы, расположенные в цис-положении (на той же молекуле ДНК) на большом расстоянии. При этом активность сайленсера, подобно энхансеру, не зависела от его ориентации по отношению к регулируемому промотору.
Сайленсеры Ингибирование транскрипции с использованием регуляторных элементов, называемых сайленсерами, — активный процесс. В этом случае происходит прямое подавление инициации транскрипции путем разрушения транскрипционного комплекса на промоторе или посредством его инактивации иным способом. Первый из описанных в 1986 г. сайленсеров обладал классическими энхансероподобными свойствами, действуя на промоторы, расположенные в цис-положении (на той же молекуле ДНК) на большом расстоянии. При этом активность сайленсера, подобно энхансеру, не зависела от его ориентации по отношению к регулируемому промотору.
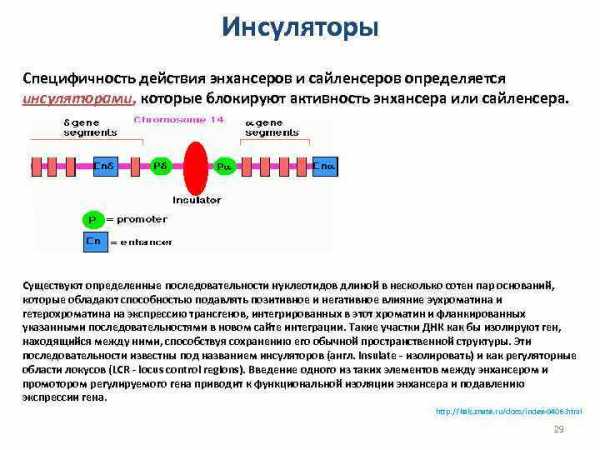 Инсуляторы Специфичность действия энхансеров и сайленсеров определяется инсуляторами, которые блокируют активность энхансера или сайленсера. Существуют определенные последовательности нуклеотидов длиной в несколько сотен пар оснований, которые обладают способностью подавлять позитивное и негативное влияние эухроматина и гетерохроматина на экспрессию трансгенов, интегрированных в этот хроматин и фланкированных указанными последовательностями в новом сайте интеграции. Такие участки ДНК как бы изолируют ген, находящийся между ними, способствуя сохранению его обычной пространственной структуры. Эти последовательности известны под названием инсуляторов (англ. insulate — изолировать) и как регуляторные области локусов (LCR — locus control regions). Введение одного из таких элементов между энхансером и промотором регулируемого гена приводит к функциональной изоляции энхансера и подавлению экспрессии гена. http: //kak. znate. ru/docs/index-9406. html 29
Инсуляторы Специфичность действия энхансеров и сайленсеров определяется инсуляторами, которые блокируют активность энхансера или сайленсера. Существуют определенные последовательности нуклеотидов длиной в несколько сотен пар оснований, которые обладают способностью подавлять позитивное и негативное влияние эухроматина и гетерохроматина на экспрессию трансгенов, интегрированных в этот хроматин и фланкированных указанными последовательностями в новом сайте интеграции. Такие участки ДНК как бы изолируют ген, находящийся между ними, способствуя сохранению его обычной пространственной структуры. Эти последовательности известны под названием инсуляторов (англ. insulate — изолировать) и как регуляторные области локусов (LCR — locus control regions). Введение одного из таких элементов между энхансером и промотором регулируемого гена приводит к функциональной изоляции энхансера и подавлению экспрессии гена. http: //kak. znate. ru/docs/index-9406. html 29
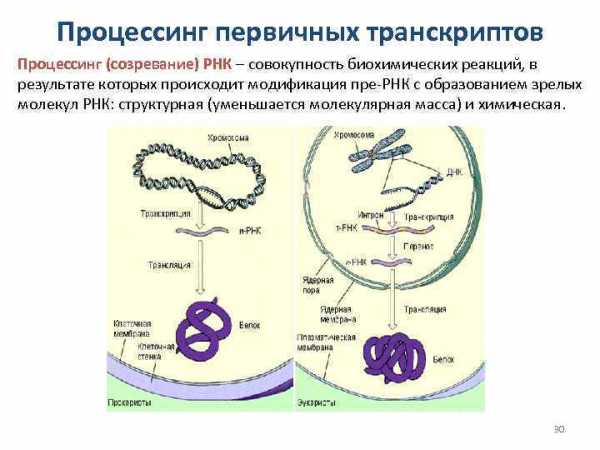 Процессинг первичных транскриптов Процессинг (созревание) РНК – совокупность биохимических реакций, в результате которых происходит модификация пре-РНК с образованием зрелых молекул РНК: структурная (уменьшается молекулярная масса) и химическая. 30
Процессинг первичных транскриптов Процессинг (созревание) РНК – совокупность биохимических реакций, в результате которых происходит модификация пре-РНК с образованием зрелых молекул РНК: структурная (уменьшается молекулярная масса) и химическая. 30
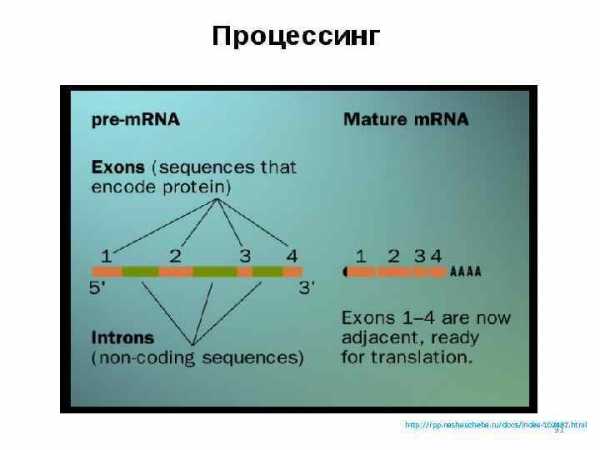 Процессинг первичных транскриптов http: //rpp. nashaucheba. ru/docs/index-102487. html 31
Процессинг первичных транскриптов http: //rpp. nashaucheba. ru/docs/index-102487. html 31
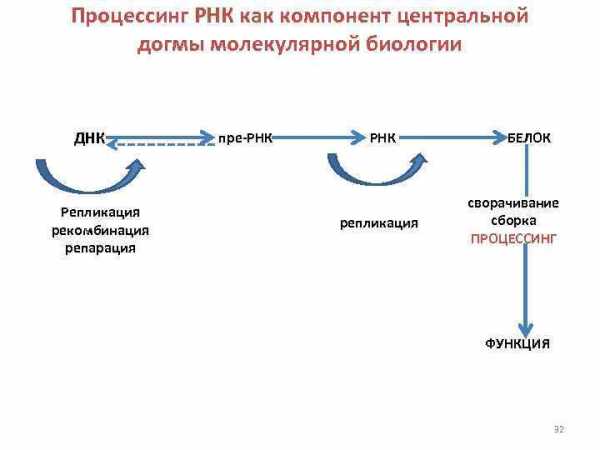 Процессинг РНК как компонент центральной догмы молекулярной биологии ДНК Репликация рекомбинация репарация пре-РНК репликация БЕЛОК сворачивание сборка ПРОЦЕССИНГ ФУНКЦИЯ 32
Процессинг РНК как компонент центральной догмы молекулярной биологии ДНК Репликация рекомбинация репарация пре-РНК репликация БЕЛОК сворачивание сборка ПРОЦЕССИНГ ФУНКЦИЯ 32
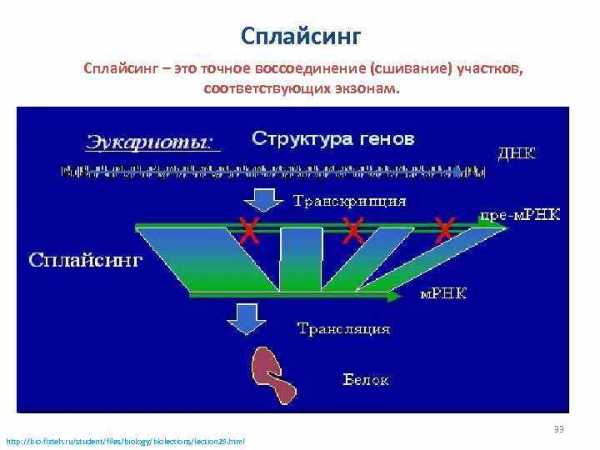 Сплайсинг – это точное воссоединение (сшивание) участков, соответствующих экзонам. 33 http: //bio. fizteh. ru/student/files/biology/biolections/lection 25. html
Сплайсинг – это точное воссоединение (сшивание) участков, соответствующих экзонам. 33 http: //bio. fizteh. ru/student/files/biology/biolections/lection 25. html
 Альтернативный сплайсинг • обеспечивает кодирование одним геном различных конечных продуктов (структурно и функционально различающихся полипептидов), что определяется спецификой клетки (ткани), то есть, один ген обеспечивает образование изоформ белка, специфичных для различных конкретных тканей; • является эффективным и экономичным способом кодирования множества продуктов ограниченным числом нуклеотидов; • служит одним из механизмов порождения белкового разнообразия у высших эукариот; 34
Альтернативный сплайсинг • обеспечивает кодирование одним геном различных конечных продуктов (структурно и функционально различающихся полипептидов), что определяется спецификой клетки (ткани), то есть, один ген обеспечивает образование изоформ белка, специфичных для различных конкретных тканей; • является эффективным и экономичным способом кодирования множества продуктов ограниченным числом нуклеотидов; • служит одним из механизмов порождения белкового разнообразия у высших эукариот; 34
 http: //www. biospsma. spb. ru/SZGMU_SITE/TL_Abstracts_of_l ectures/Structure_and_function_of_the_gene. html
http: //www. biospsma. spb. ru/SZGMU_SITE/TL_Abstracts_of_l ectures/Structure_and_function_of_the_gene. html
present5.com
Биология для студентов — 05. Молекулярные основы наследственности
Доказательства роли ДНК в наследственности
После того как было установлено, что гены находятся в хромосомах и расположены там в определенном порядке, возник вопрос об их химический природе. Ученым было известно, что в состав хромосом высших организмов входят ДНК и несколько типов гистоновых и негистоновых белков. До 40-х годов нашего столетия большинство ученых считали, что гены имеют белковую природу. Русский ученый Н. К. Кольцов высказал мысль, что хромосома это гигантская биологическая молекула, обладающая свойством самоудвоения, и что все свойства и признаки организма обусловлены строением белка и взаимодействием его молекул. Казалось вероятным, что именно в 6елках заключена наследственная информация о развитии всех признаков и свойств организма. Однако проведенные в последующем эксперименты на микроорганизмах с применением новейших методов исследований позволили установить, что генетическая информация сосредоточена в нуклеиновых кислотах. В 1944 году американский микробиолог Эвери из бактерий штамма S выделил ДНК и внес ее в питательную среду, на которой размножались бактерии авирулентного штамма R. Значительная часть авирулентных бескапсульных бактерий штамма R трансформировалась в капсульные вирулентные бактерии S -штамма. Это явление дало Эвери основание утверждать о ведущей роли ДНК в переносе наследственной информации от одного штамма бактерий к другому.
Другой эксперимент, подтверждающий роль ДНК в наследственности, провели американские ученые И. Чейз и Херши. Они размножали ДНК-содержащий вирус-бактериофаг на среде, содержащей радиоактивные фосфор и серу Р35 и S33. Радиоактивная сера включилась в серусодержащие белки оболочки фага, а радиоактивный фосфор — в ДНК. Далее мечеными радиоактивными изотопами фагами заражали бактерии. С помощью электронного микроскопа было установлено, что радиоактивная сера не проникала в клетку бактерии, внутри клетки был обнаружен только радиоактивный фосфор. Это свидетельствовало о том, что при заражении бактерии фагом внутрь клетки проникает только ДНК. В зараженной клетке образовалось множество вирионов фага. Следовательно, генетическая информация, необходимая для синтеза ДНК фагов, содержится в ДНК проникших в клетку вирусов. Доказательством ведущей роли ДНК в наследственности является и то, что она локализована главным образом в хромасомах, поэтому молекулярная генетика не противоречит хромосомной теории наследственности и законам классической генетики.
Строение и синтез ДНК.
В период с 1900 по 1932 год был выяснен химический состав ДНК. Было установлено, что в ее состав входят:
- остатки фосфорной кислоты,
- углеводный компонент,
- дезоксирибоза,
- четыре типа азотистых оснований, два производных пурина (аденин и гуанин) и два производных пиримидина (тимин и цитозин).
Э. Чаргафф установил, что в ДНК содержание аденина равно содержанию тимина (А=Т), а содержание гуанина равно содержанию цитозина (А=Ц). Отсюда: (А+Г) : (Т+Ц) = 1, т. е. сумма пуриновых нуклеотидов равна сумме пиримидиновых. Такая закономерность указывают на комплементарное соединение пуриновых и пиримидиновых оснований в молекуле ДНК. Приоритет в расшифровке структуры молекулы ДНК принадлежит Д. Уотсону и Ф. Крику. Согласно их модели, молекула ДНК имеет двойную спираль, состоящую из двух нуклеотидных цепей с общей осью. Диаметр двойной спирали ДНК равен 2 нм, а расстояние между витками 3,4 нм. На каждый виток спирали приходится 10 пар нуклеотидов, отсюда расстояние между азотистыми основаниями равно 0,34 нм. Каждая из цепей ДНК является полинуклеотидом и состоит из 4 типов нуклеотидов.

В состав нуклеотида входят:
- дезоксирибоза (Д),
- остаток фосфорной кислоты (Ф) ,
- одно из четырех азотистых оснований (А,Г,Ц и Т).
Соединение пуриновых и пиримидиновых оснований с дезоксирибозой приводит к образованию нуклеозида. При присоединении фосфорного остатка к углеводной части нуклеозида образуется нуклеотид. Дезоксирибоза в нуклеотидах соединяется с основаниями гликозидной связью, а с остатками фосфорной кислоты — эфирными связями. Азотистые основания нуклеотидов обоих цепей заключены внутри между витками спирали и соединены водородными связями. Причем аденин одной цепи всегда связан только с тимином другой цепи, а гуанин — только с цитозином. Пара А — Т соединена двумя водородными связями, а пара Г-Ц — тремя. Такой порядок азотистых оснований называется комплементарностью. Коэффициентом видовой специфичности называют отношение (А+Т): (Г+Ц).
Репликация ДНК.
ДНК является веществом, количество которого строго постоянно во всех клетках организма. ДНК находится в хромосомах, и репликация ее происходит перед каждым удвоением хромосом и делением клетки. На отдельных участках молекулы ДНК образуются так называемые вилки репликации. В этих местах водородные связи между азотистыми основаниями под действием ферментов разрываются, комплементарные нити разъединяются и каждая из них становится матрицей, на которой происходит синтез дочерних нитей. Такой тип репликации ДНК получил название полуконсервативного. Процесс синтеза протекает при участии комплекса ферментов, главнейшим из которых является ДНК-полимераза. Участок ДНК в том месте, где начали расплетаться комплементарные нити, называется вилкой репликации. Она образуется у прокариот в одной определенной, генетически фиксированной точке. В молекуле ДНК эукариот таких «стартовых точек» бывает несколько. Синтез новых комплементарных цепей при репликации ДНК происходит по частям. Эти отрезки, состоящие из 1000-2000 нуклеотидов, называют фрагментами Оказаки. Структура, способная к репликации (хромосома, плазмида, вирусный геном), называется репликоном. Репликация обеспечивает материальную непрерывность наследственного вещества клетки.
Строение, синтез и типы РНК
Молекулы рибонуклеиновой кислоты имеют одну полинуклеотидную цепь. В состав молекулы РНК входят четыре типа азотистых оснований (аденин, гуанин, цитозин и урацил), сахар рибоза и остатки фосфорной кислоты. По составу от ДНК она отличается тем, что вместо дезоксирибозы содержит рибозу и вместо пуринового основания тимина — урацил. Схему строения молекулы РНК можно представить следующим образом? У где, А,Г,Ц,У — азотистые основания, Р — рибоза и Ф — остатки фосфорной кислоты. Синтез молекулы РНК происходит на одной из цепей молекулы ДНК. Этот процесс протекает с участием большого числа ферментов и называется транскрипцией. Причем двойная цепь ДНК раскручивается и на одной из ее цепей, которая называется смысловой синтезируется РНК. В организме существуют три основных типа РНК:
- информационная (и-РНК), или матричная (м-РНК),
- рибосомальная (р-РНК),
- транспортная (т-РНК).
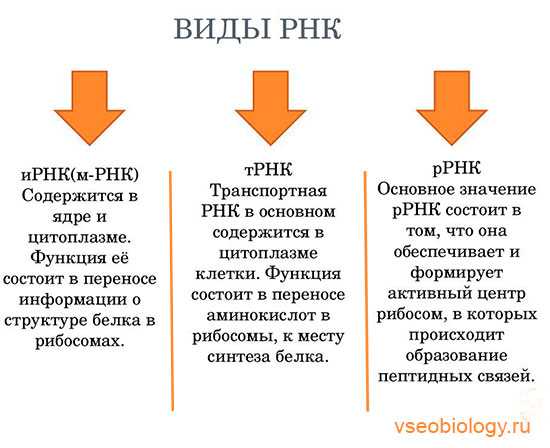
Эти типы РНК различаются по величине молекул и функциям.
Информационная РНК. Роль информационной РНК заключается в том, что она переписывает информацию с молекулы ДНК и переносит ее к месту синтеза белка. В рибосомах и-РНК выполняет роль матрицы в процессе биосинтеза белка. Транспортные РНК выполняют функцию переноса аминокислот к месту синтеза белка. Молекула т-РНК напоминает форму клеверного листа. На конце одной цепи находится акцепторный участок — триплет ЦЦА, к которому прикрепляется аминокислота. В центре средней петли находится антикодон — триплет, состоящий из трех нуклеотидов комплементарных генетическому коду и — РНК. Рибосомальная РНК синтезируется в ядрышках, затем поступает в цитоплазму. Объединяясь с особыми белками, она образует рибосомы, в которых осуществляется биосинтез белков. Количество рибосомальной РНК составляет около 80 процентов.
Генетический код является триплетным. Кроме того, к свойствам генетического кода относят:
- триплетность
- вырожденность,
- неперекрываемость,
- универсальность.

Вырожденность генетического кода заключается в том, что, как правило, одну аминокислоту кодируют не один, а несколько триплетов. В генетическом коде есть аминокислоты, кодируемые одним, двумя, тремя, четырьмя и шестью триплетами. Неперекрываемость генетического кода связана с тем, что каждый из нуклеотидов входит только в один из кодонов и считывание идет в одном направлении — триплет за триплетом. Генетический код универсален. Это значит, что у животных, растений, бактерий и вирусов одну и ту же аминокислоту кодируют одинаковые сочетания. Процесс реализации наследственной информации в биосинтезе белка осуществляется при участии трех видов РНК, ферментов, АТФ и других компонентов. Передачу наследственной информации с ДНК на белок можно представить следующим образом: ДНК → и-РНК → белок. Процесс биосинтеза сложный и включает ряд этапов:
- транскрипцию,
- сплайсинг,
- трансляцию.
Первый этап называется транскрипцией. Он происходит в ядре клетки. В результате транскрипции наследственная информация с ДНК переписывается на и-РНК. Этот процесс осуществляется при участии ряда ферментов, главным из которых является РНК-полимераза. Исследования показали, что в результате транскрипции синтезируется проматричная РНК, которая значительно больше по размеру и содержит фрагменты не несущие наследственной информации. Они получили название интронов в отличие от кодирующих фрагментов, которые называются экзонами. Интроны считываются с молекулы ДНК одновременно с экзонами, поэтому про-м-РНК значительно длиннее, чем зрелая м-РНК. В дальнейшем интроны «вырезаются» из молекулы РНК, а фрагменты экзонов «сращиваются» между собой в строгом порядке. Этот процесс называется сплайсингом. В процессе сплайсинга образуется зрелая м-РНК, которая содержит только ту информацию, которая необходима для синтеза белков.
Следующий этап биосинтеза — трансляция. Этот процесс происходит на рибосомах при участии т-РНК. Молекула и-РНК после сплайсинга через поры ядра выходит в цитоплазму и прикрепляется к рибосоме. Трансляция начинается с так называемого стартового кодона — АУТ. Активированные аминокислоты прикрепляются к т-РНК и переносятся к рибосомам. Здесь они в соответствии с генетическим кодом соединяются в полипептидную цепь. Молекула и-РНК обычно работает на нескольких рибосомах (5-20), соединенных в полисомы. Начало синтеза полипептидной цепи называется инициацией. Последовательность аминокислот в молекуле белка определяется последовательностью кодонов в и-РНК. Синтез полипептидной цепи прекращается, когда, на и-РНК появляется один из кодонов -терминаторов (УАА, УАГ или УГА).
Строение хромосом: хроматида, хромомеры, эухроматические и гетерохроматические районы хромосом.
Хромосомы состоят из двух хроматид, объединенных первичной перетяжкой. По положению центромеры хромосомы делятся на:
- метацентрические (равноплечие),
- субметацентрические (неравноплечие),
- акроцентрические (центромера лежит у одного из концов хромосомы, последняя представляет собой палочку с очень коротким или даже незаметным вторым плечом),
- телоцентрические — палочковидные хромосомы с центромерой, расположенной на проксимальном конце.
Хромомеры, по мнению одних исследователей, представляют собой плотно спирализованные участки, по мнению других — уплотнения нуклеопротеидного материала. Промежутки между хромомерами называются межхромомерными нитями.
Политения — редупликация хромонем в хромосомах, приводящая к увеличению числа хромонем без увеличения числа хромосом и без реорганизации ядра. Этот процесс, протекающий внутри хромосом, приводит к полиплоидизации количества.
Эухроматин, активный хроматин, участки хроматина (вещества хромосом), сохраняющие деспирализованное состояние элементарных дезоксирибонуклеопротеидных нитей (ДНП) в покоящемся ядре, т. е. в интерфазе. Эухроматин отличается от гетерохроматина также способностью к интенсивному синтезу рибонуклеиновой кислоты (РНК) и большим содержанием негистоновых белков.
Гетерохроматин, участки хромосом, остающиеся в промежутке между делениями клетки, т. е. в интерфазе, уплотненными (в отличие от др. участков — эухроматина). Гетерохроматин иногда тесно связан с ядрышком, образуя вокруг него подобие кольца или оболочки. Во время митоза Гетерохроматин окрашивается сильнее или слабее, чем эухроматин (явление положительного или отрицательного гетеропикноза).
Изменения в организации морфологии хромосом в ходе митоза и мейоза. Репликация хромосом. Политения. Онтогенетическая изменчивость хромосом. Хромосомы в период митоза и мейоза
При переходе клетки к делению синтез ДНК и РНК в хромосомах прекращается, хромосомы приобретают всё более плотную упаковку (например, в одной хромосоме человека цепочка ДНК длиной 160 мм укладывается в объёме всего 0,5´10 мкм), ядерная мембрана разрушается и хромосомы выстраиваются на экваторе клетки. Основная структурная единица метафазныххромасом, так же как и интерфазных, — нить ДНП диаметром 100—200, уложенная в плотную спираль. Каждая метафазная хромасома состоит из хроматид, образовавшихся в результате репликации исходной интерфазной хромосомы. Использование меченых и модифицированных предшественников ДНК позволило четко различать в хромосоме, находящейся в метафазе митоза, дифференциально окрашенные хроматиды, благодаря чему было установлено, что при репликации хромосом нередко происходит обмен участками между сестринскими хроматидами (кроссинговер). Современные цитологи рассматривают матрикс метафазных хромосом, как остаточный материал разрушающегося ядрышка; часто он вовсе не обнаруживается.
Политения — редупликация хромонем в хромосомах, приводящая к увеличению числа хромонем без увеличения числа хромосом и без реорганизации ядра. Этот процесс, протекающий внутри хромосом, приводит к полиплоидизации количества.
Хроматин — основной компонент клеточного ядра; его достаточно легко получить из выделенных интерфазных ядер и из выделенных митотических хромосом. Фракции хроматина, полученные из разных объектов, обладают довольно однообразным набором компонентов. Было найдено, что по суммарному химическому составу хроматин из интерфазных ядер мало отличается от хроматина из митотических хромосом. Главными компонентами хроматина являются ДНК и белки, среди которых основную массу составляют гистоны и негистоновые белки. В среднем в хроматине около 40% приходится на ДНК и около 60% — на белки, среди которых специфические ядерные белки-гистоны составляют от 40 до 80% от всех белков, входящих в состав выделенного хроматина. Кроме того, в состав хроматиновой фракциям входят мембранные компоненты, РНК, углеводы, липиды, гликопротеиды. В структурном отношении хроматин представляет собой нитчатые комплексные молекулы дезоксирибонуклеопротеида (ДНП), которые состоят из ДНК, ассоциированной с гистонами.
Различают четыре уровня организации ядерного хроматина. Первый — уровень нуклеосомной фибриллы. В нуклеосоме различают сердцевинную часть и линкерную область. Сердцевинная часть соответствует «бусинам», а линкерная — связывающему «бусины» участку базовой ДНК. Сердцевинная и линкерная области образуют полную нуклеосому. Размеры полной нуклеосомы могут варьироваться у разных видов. Второй уровень пространственной структуры хроматина — соленоид позволяет «сложить» ДНК с ее спутниками — белками еще более компактно. Соленоидная структура образуется в результате свертывания (на манер спирали) нуклеосомной нити и в одних местах носит более-менее регулярный характер, в других — неравномерный — здесь наблюдается как бы «сгущение» витков. Третий уровень организации (компактизации) ДНК в хроматине определяется укладкой соленоидной структуры в петли, опирающиеся, как полагают, на скелетные осевые образования хромосом. Длина петли — до 90 тысяч пар нуклеотидов. Материалом для скелетных нитей (для ядерного скелета) служат белки. Эти нити получили название «нуклеонемы». Четвертый уровень организации хроматина представлен хромосомами. Механизм формирования этих органелл еще не ясен. Но несомненно, что активация генов подразумевает глубокие изменения пространственных структур хромосом: налицо связь организации хроматина и регуляции работы генов. Кольцевые ДНК могут образовывать особую пространственную структуру — суперспираль. Суперспирализация опять же зависит от биохимии плазмы клетки.
vseobiology.ru
Лекция 2 Молекулярные основы генетики
С этим файлом связано 19 файл(ов). Среди них: Shmidt_R_Tevs_G_Fiziologia_cheloveka_tom_3.pdf, Shmidt_R_Tevs_G_Fiziologia_cheloveka_tom_2.pdf, Тесты для подготовки к Крок-1.doc, Gayvoronskiy_Metodichka_Osteologia.pdf, Lektsia_2_Molekulyarnye_osnovy_genetiki_Prezentatsia.pdf и ещё 9 файл(а).
Показать все связанные файлы
Тезисы лекции
Тема лекции: МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ. РЕАЛИЗАЦИЯ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ.
Что такое наследственная информация?
Под наследственной информацией мы понимаем информацию о строении белков и характере синтеза белков в организме человека. Синоним – генетическая информация.
В хранении и реализации наследственной информации ведущую роль играют нуклеиновые кислоты. Нуклеиновые кислоты – это полимеры, мономерами которых являются нуклеотиды. Впервые нуклеиновые кислоты были открыты Ф. Мишером в 1869 г в ядрах лейкоцитов из гноя. Название происходит от латинского nucleus –ядро. Различают два вида нуклеиновых кислот: ДНК и РНК
Функции нуклеиновых кислот
ДНК хранит генетическую информацию. В ДНК находятся гены. РНК принимают участие в биосинтезе белка (т.е. в реализации наследственной информации)
Открытие роли ДНК в хранении наследственной информации. В 1944 г. Oswald Avery, Macklin McCarty, and Colin MacLeod представили доказательства того, что гены находятся в ДНК. Они работали с пневмококками, у которых есть два штамма: патогенный (S-штамм) и непатогенный (R- штамм). Заражение S-штаммом мышей приводит к их гибели
Если вводят R- штамм, то мыши выживают. Из убитых бактерий S-штамма выделили ДНК, белки и полисахариды и добавляли к R- штамму. Добавление ДНК вызывает трансформацию непатогенного штамма в патогенный.
История открытия строения ДНК.
Строение ДНК открыли в 1953 г Дж.Уотсон и Ф.Крик. В своей работе они использовали данные, которые получили биохимик Е.Чаргафф и биофизики Р.Франклин, М.Уилкинс.
Работа Е.Чаргаффа: В 1950 г. биохимик Ервин Чаргафф установил, что в молекуле ДНК:
1) А=Т и Г=Ц
2) Сумма пуриновых оснований (А и Г) равна сумме пиримидиновых оснований (Т и Ц): А+Г=Т+Ц
Или А+Г/Т+Ц=1
Работа Р.Франклин и М.Улкинс: В начале 50-х г.г. биофизики Р.Франклин и М.Уилкинс получили рентгенограммы ДНК, которые показали, что ДНК имеет форму двойной спирали. В 1962 г. Ф.Крик, Дж.Уотсон и Морис Уилкинс получили Нобелевскую премию по физиологии и медицине за расшифровку строения ДНК
Строение ДНК
ДНК – это полимер, который состоит из мономеров – нуклеотидов. Строение нуклеотида ДНК: нуклеотид ДНК состоит из остатков трех соединений:
1) Моносахарида дезоксирибозы
2) Фосфата — остатка фосфорной кислоты
3) Одного из четырех азотистых оснований – аденина (А), тимина (Т), гуанина (Г) и цитозина (Ц).
Азотистые основания: А и Г – производные пурина (два кольца), Т и Ц- производные пиримидина (одно кольцо).
А комплементарен Т
Г комплементарен Ц
Между А и Т образуется 2 водородные связи, между Г и Ц — 3
В нуклеотиде атомы карбона в дезоксирибозе пронумерованы от 1’ до 5’.
К 1’-карбону присоединяется азотистое основание, а к 5’-карбону – фосфат. Нуклеотиды соединяются между собой фосфодиэфирными связями. В результате образуется полинуклеотидная цепьСкелет цепи состоит из чередующихся молекул фосфата и сахара дезоксирибозы.
Азотистые основания расположены сбоку молекулы. Один из концов цепи обозначают 5’, а другой — 3’ (по обозначению соответствующих атомов карбона). На 5’ – конце находится свободный фосфат, это начало молекулы. На 3’- конеце находится ОН-группа. Это хвост молекулы. Новые нуклеотиды могут присоединяться к 3’- концу.
Строение ДНК:
Согласно модели Крика –Уотсона, ДНК состоит из двух полинуклеотидных цепей, которые свернуты в спираль. Спираль правая (В-форма)
Цепи в ДНК расположены антипараллельно. 5’-конец одной полинуклеотидной цепи соединяется с 3’-концом другой.
В молекуле ДНК видны маленькая и большая борозды.
К ним присоединяются разные регуляторные белки.
В двух цепях азотистые основания расположены по принципу комплементарности и соединены водородными связями
А и Т – двумя водородными связями
Г и Ц — тремя
Размеры ДНК: толщина молекулы ДНК составляет 2 нм, расстояние между двумя витками спирали – 3,4 нм, в одном полном витке — 10 пар нуклеотидов. Средняя длина одной пары нуклеотидов 0,34 нм. Длина молекулы варьирует. В бактерии кишечная палочка кольцевидная ДНК имеет длину 1,2 мм. У человека суммарная длина 46 ДНК, выделенных из 46 хромосом составляет около 190 см. Следовательно, средняя длина 1 молекулы ДНК человека более 4 см.
Линейное изображение ДНК. Если цепи ДНК изображают в виде линии, то принято вверху изображать цепь в направлении от 5‘ к 3‘.
5‘ АТТГТЦЦГАГТА 3‘
3‘ ТААЦАГГЦТЦАТ 5′
Локализация ДНК в клетках эукариот:
Ядро – входит в состав хромосом;
Митохондрии;
У растений – пластиды.
Функция ДНК: хранит наследственную (генетическую) информацию. В ДНК находятся гены. У человека в клетке менее 30 000 генов.
Свойства ДНК
Способность к самоудвоению (редупликации) Редупликация – синтез ДНК.
Способность к репарации – восстановлению повреждений ДНК.
Способность к денатурации и ренатурации. Денатурация – под действием высокой температуры и щелочей разрываются водородные связи между цепями ДНК и ДНК становится однонитевой. Ренатурация – обратный процесс. Это свойство используется в ДНК-диагностике.
Редупликация – это синтез ДНК.
Процесс идет перед делением клетки в синтетическом периоде интерфазы.
Суть процесса: Фермент геликаза разрывает водородные связи между двумя цепями ДНК и раскручивает ДНК. На каждой материнской цепи по принципу комплементарности синтезируется дочерняя цепь. Процесс катализирует фермент ДНК-полимераза.
В результате редупликации образуется две дочерние ДНК, которые имеют такое же строение как и материнская молекула ДНК.
Рассмотрим процесс редупликации более подробно
1) Редупликация – полуконсервативный процесс, т.к. дочерняя молекула получает одну нить от материнской ДНК, а вторую синтезирует вновь
2) ДНК синтезируется из нуклеотидов с тремя фосфатами – АТФ, ТТФ,ГТФ,ЦТФ. При образовании фосфодиэфирной связи два фосфата выщепляются.
3) Синтез ДНК начинается в определенных точках – точках инициации репликации. В этих участках много А-Т пар. Специальные белки присоединяются к точке инициации.
Фермент геликаза начинает раскручивать материнскую ДНК. Нити ДНК расходятся.
Редупликацию катализирует фермент ДНК-полимераза.
От точки инициации фермент ДНК-полимераза движется в двух противоположных направлениях. Между расходящимися цепями образуется угол- репликационная вилка.
3) Цепи материнской ДНК антипараллельны. Дочерние цепи синтезируются антипараллельно материнским, поэтому синтез дочерних цепей в области репликационной вилки идет в двух противоположных направлениях. Синтез одной цепи идет в направлении движения фермента. Эта цепь синтезируется быстро и непрерывно (лидирующая). Вторая синтезируется в противоположном направлении маленькими фрагментами – фрагментами Оказаки (отстающая цепь).
4) Фермент ДНК-полимераза не может сам начать синтез дочерней цепи ДНК.
Синтез лидирующей цепи и любого фрагмента Оказаки начинается с синтеза праймера. Праймер — кусочек РНК длиной 10-15 нуклеотидов. Праймер синтезирует фермент праймаза из нуклеотидов РНК. К праймеру ДНК-полимераза присоединяет нуклеотиды ДНК.
В последующем праймеры вырезаются, брешь застраивается нуклеотидами ДНК.
Фрагменты сшиваются ферментами — лигазами
5) Ферменты, участвующие в редупликации: геликаза, топоизомераза, дестабилизирующие белки, ДНК-полимераза, лигаза.
6) Молекула ДНК длинная. В ней образуется большое число точек начала репликации.
ДНК синтезируется фрагментами – репликонами. Репликон – участок между двумя точками инициации репликации. В соматической клетке человека в 46 хромосомах более 50000 репликонов. Синтез ДНК 1 соматической клетки человека длится более 10 часов.
Самокоррекция ДНК (ДНК-редактирование)
В процессе редупликации ДНК полимераза иногда делает ошибки (неправильно включает нуклеотиды). Она проверяет свою работу. Если обнаруживает ошибку, то вырезает последние нуклеотиды и включает в ДНК новые.
Это процесс называется самокоррекция ДНК. Она уменьшает частоту ошибок при редупликации (неправильно включенные нуклеотиды) в 10 раз – с 1/100000 нуклеотидов до 10/1000000
Значение редупликации: в результате редупликации образуется две дочерние ДНК, которые как две капли воды похожи на материнскую молекулу ДНК. При делении клеток дочерние ДНК расходятся в дочерние клетки. Таким образом, редупликация обеспечивает передачу наследственной информации в дочерние клетки.
Строение РНК
РНК – это полимер, состоящий из мономеров – нуклеотидов. Главные отличия РНК от ДНК:
ДНК состоит из двух полинуклеотидных цепей, РНК — из одной;
ДНК содержит моносахарид дезоксирибозу, РНК — рибозу;
ДНК содержит Тимин, РНК — Урацил
Виды РНК и функции
| иРНК | Переносит информацию о строении белка из ядра в цитоплазму |
| рРНК | Структурная функция. Входит в состав рибосом. Синтезируется в ядрышках. |
| тРНК | Транспортирует аминокислоты в рибосомы для синтеза белка. Играет важную роль в переводе последовательности нуклеотидов в иРНК в последовательность аминокислот в белке |
| Малые ядерные РНК | Принимают участие в процессинге (созревание иРНК) |
| Малые ядрышко-вые РНК | Принимают участие в созревании рРНК |
Все перечисленные РНК закодированы в ДНК и синтезируются в ядре клетки. Общая функция всех РНК – обеспечивают синтез белка.
Что такое ген?
Термин «ген» предложил В. Йогансен в 1909 г. Ген ( в узком смысле слова) – это участок ДНК, в котором закодирована информация о строении одного белка.
Однако, в ДНК закодированы не только белки, но и строение всех видов РНК. В ДНК также находятся регуляторные участки, которые регулируют процессы транскрипции: ускоряют или замедляют транскрипцию, блокируют транскрипцию или, наоборот, активируют.
Ген в более широком смысле слова – это участок ДНК, который кодирует первичную структуру белка, рРНК, тРНК, или регулирует транскрипцию другого гена.
Классификация генов. В зависимости от выполняемых функций выделяют две группы генов:
1. Структурные гены – это гены, которые кодируют белок или РНК (рРНК, тРНК или др. вид РНК).
2. Регуляторные гены – гены, которые регулируют процессы биосинтеза белка (у эукариот – это промоторы –место присоединения РНК-полимеразы, энхансеры – ускоряют транскрипцию, сайленсеры — тормозят)
Что такое ген?
Строение гена эукариот, кодирующего белок:
Промотор –участок гена, к которому присоединяется фермент РНК-полимераза. Определенные участки промотора (ГЦ-мотивы, ЦААТ-бокс) нужны для присоединения регуляторных белков. ТАТА-бокс – участок, где много АТ-пар. Здесь ДНК начинает раскручиваться.
Транскриптон – транскрибируемый участок гена. Он включает :
лидер- нужен для соединения иРНК с рибосомой,
участок гена, кодирующий полипептид начинается с инициального триплета и заканчивается стоп-кодоном. У эукариот он состоит из экзонов и интронов. Экзоны кодируют белки, а интроны – нет. Интроны в последующем вырезаются из иРНК.
трейлер- необходим для отсоединения иРНК от рибосомы,
Терминатор – место окончания транскрипции.
Типичный ген человека состоит примерно из 28 000 оснований и имеет 8 экзонов. Он кодирует полипептид, состоящий в среднем из 447 аминокислот.
Самый длинный ген, найденный в геноме человека, это ген мышечного белка дистрофина, содержащий 2,4 · 106 п.н.
Что такое генетический код?
Генетический код – система записи генетической информации о строении белков в ДНК в виде определенной последовательности нуклеотидов
Основные свойства генетического кода:
Триплетность
Вырожденность (избыточность)
Специфичность
Неперекрываемость
Однонаправленность
Наличие инициирующего кодона (АУГ) и нонсенс-кодонов
Колинеарность
Универсальность
Экспрессия гена
Под экспрессией гена понимают реализацию записанной в нем наследственной информации. Синтез белка – это процесс, который обеспечивает реализацию наследственной информации в клетке. Согласно центральной догме молекулярной биологии он идет в следующем направлении:
ДНК → иРНК → белок →признак.
Этапы синтеза белка
Транскрипция – синтез иРНК
Активация аминокислот и соединение с тРНК
Трансляция — синтез первичной структуры белка в рибосоме
Посттрансляционные процессы образование пространственных структур белка (вторичной, третичной, четвертичной), модификация аминокислот.
Транскрипция.
Транскрипция – это синтез иРНК. У эукариот трнаскрипция имеет свои осрбенности.
Ген эукариот состоит из экзонов и интронов. Интроны – не кодируют белок. Они вырезаются из иРНК. Таким образом, транскрипция у эукариот включает два этапа:
Синтез про-иРНК (незрелой иРНК), которая полностью комплементарна гену.
Процессинг-созревание иРНК. Процессинг включает:
сплайсинг (вырезание интронов и сшивание экзонов),
образование кэпа и поли-А-хвоста. Кэп (модифицированный гуанин) прикрепляется к начальному концу иРНК, поли-А-хвост – большое количество А-нуклеотидов прикрепляются к концу иРНК. Кэп и хвост обеспечивают стабильность иРНК в цитоплазме.
Активация аминокислот и соединение с тРНК.
В клетках эукариот около 50 видов РНК (в связи с избыточностью генетического кода). Каждая тРНК имеет антикодон (для взаимодействия с кодоном иРНК) и акцепторный участок (куда присоединяется аминокислота). Соединение тРНК с аминокислотой катализирует фермент аминоацил-тРНК –синтетаза. Процессу предшествует активация аминокислот (соединение с остатком АТФ -АМФ).
Аминокислота+АТФ= Аминокислота+АМФ (АК+АМФ)
АК+АМФ +ТРНК =АК+ТРНК +АМФ
Трансляция.
Трансляция – синтез первичной структуры белка в рибосоме. Этапы трансляции:
Инициация – начало трансляции. Рибосома соединяется с иРНК и захватывает два кодона (первый – инициальный -оказывается в пептидильном центре). К инициальному триплету подходит тРНК с инициальным метионином. Образуется инициальный комплекс- рибосома, инициальный триплет, тРНК
Элонгация – синтез полипептида. Ко второму кодону иРНК подходит вторая тРНК с аминокислотой. Если антикодон тРНК комплементарен кодону иРНК, две аминокислоты соединяются пептидной связью. Затем первая тРНК выходит из рибосомы, рибосома перемещается на один триплет вперед. К этому триплету подходит новая тРНК с аминокислотой. Если антикодон тРНК комплементарен кодону иРНК, то между двумя последними аминокислотами вновь образуется пептидная связь и процесс повторяется. Процесс продолжается до тех пор, пока рибосома не дойдет до стоп-кодона
Терминация транскрипции – окончание. Рибосома доходит до стоп-кодона. Синтез полипептида останавливается.
Посттрансляционные процессы- образование вторичной, третичной, четвертичной структуры белка, модификация аминокислот Процесс может идти в цитоплазме, гранулярной ЭПС, комплексе Гольджи. После того как белок образовал третичную или четвертичную структуру, он может выполнить свои функции.
Регуляция экспрессии генов у прокариот. Оперон.
У прокариот кольцевидная ДНК, которая кодирует небольшое количество белков (у кишечной палочки более 4000). Для многих генов характерна оперонная регуляция активности.
Оперон – это группа структурных генов, которые кодируют белки-ферменты одного метаболического процесса и работы которых находится под контролем общих регуляторных генов. Опероны позволяют маленькой ДНК кодировать много белков.
Оперон был открыт в 1961 г. французскими учеными Жакобом и Моно. Они открыли лактозный оперон у кишечной палочки. Если кишечную палочку поместить в среду, содержащую лактозу, то она начинает вырабатывать три фермента, участвующих в метаболизме лактозы.
Ферменты кодируют три структурных гена:
lacZ — галактозидаза – расщепляет лактозу на глюкозу и галактозу
Lac Y –фермент пермеаза (обеспечивает поступление лактозы в клетку)
lacA – трансацетилаза, участвует в удалении из клетки токсичных продуктов расщепления лактозы.
Структурные гены находятся в окружении регуляторных генов:
Ген-регулятор – кодирует белок-репрессор
Ген-промотор – место присоединения РНК-полимеразы для начала транскрипции
Ген-оператор. Если к нему присоединен белок-репрессор, то он блокирует транскрипцию.
Терминатор – на нем заканчивается транскрипция.
Оперон инактивирован, если белок-репрессор соединен с геном-оператором. Оперон в активном состоянии если в клетку попадает лактоза. Она соединяется с белком-репрессором и инактивирует его. Начинается синтез трех ферментов.
Отличия организации генома и экспрессии генов
у прокариот и эукариот
| Прокариоты | Эукариоты |
| ДНК кольцевидной формы, не соединена с белками, расположена в цитоплазме | ДНК линейная,соединяется с гистоновыми и негистоновыми белками, находится в ядре клетки |
| В генах нет интронов | Есть интроны |
| Мало генов (у кишечной палочки около 4000) | Много генов (у человека до 30000) |
| Есть опероны | Нет оперонов Каждый ген окружен группой регуляторных генов |
Регуляция экспрессии гена у эукариот.
В каждой клетке у эукариот экспрессируется 7-10% всех генов. Остальные гены находятся в репрессированном (неактивном) состоянии. У эукариот преобладает так называемый позитивный генетический контроль, при котором основная часть генома репрессирована, и регуляция идет путем активации необходимых генов.
На уровне транскрипции регуляция может идти следующими путями:
Амплификация (увеличение числа копий) гена;
Связывание с промотором факторов транскрипции — белков, облегчающих или затрудняющих транскрипцию;
С помощью регуляторных генов -энхансеров и сайленсеров;
Влияние гормонов, которые часто служат активаторами транскрипции; например, стероидные гормоны проникают в цитоплазму клетки, соединяются со специальным белком-рецептором, поступаю в ядро и активируют несколько генов.
Альтернативный сплайсинг – из одной про-иРНК могут вырезаться разные интроны
Метилирование нуклеотидов ДНК, в основном, в области промотора, богатой ГЦ-парами; это делает невозможным присоединение факторов транскрипции к промотору и выключает ген;
Ацетилирование белков — гистонов, что уменьшает степень связывания с ними ДНК и облегчает транскрипцию.
Контроль на уровне трансляции
Идет путем регуляции образования комплекса мРНК — стартовая тРНК— рибосома и изменении времени жизни иРНК за счет различных цитоплазматических факторов.
С помощью микроцитоплазматических РНК – маленьки РНК, которые соединяются с иРНК и блокируют трансляцию
Регуляция образования белков возможна и путем изменения быстроты и активности посттрансляционной модификации полипептидной цепи
Репарация ДНК
Репарация ДНК – это исправление ошибок ДНК. Если ошибки остаются, то они могут привести к генным мутациям и генным болезням. Репарация поддерживает генетическую целостность организма и их выживание
1) Фоторепарация у прокариот. Облучение клетки ультрафиолетовыми лучами вызывает образование в ДНК тиминовых димеров. УФ лучи активируют фермент фотореактивации, который связывается с тиминовыми димерами и разрывают их.
2) Эксцизионная репарация у прокариот и эукариот — ферменты-нуклеазы вырезают ошибочное основание или участок поврежденной цепи ДНК, фермент ДНК-полимера 1 типа встраивает нормальные нуклеотиды, ферменты лигазы сшивают фрагменты.
3) Репарация во время репликации – самокоррекция ДНК
4) Пострепликационная репарация – если не удалены ошибочные нуклеотиды во время репликации, то происходит рекомбинация поврежденной цепи с цепью ДНК во второй дочерней молекуле и ошибка устраняется
5) SOS-репарация – при репликации ДНК- полимераза перескакивает место повреждения и продолжает репликацию без разрывов, но последовательность нуклеотидов меняется
Болезни репарации ДНК.
При нарушении репарации ДНК в клетках накапливаются мутации, что со временем приводит: 1) к развитию опухолей, 2) преждевременному старению, 3) наследственным заболеваниям – болезням репарации.
Наследственные болезни, которые обусловлены мутацией генов репарации ДНК, называются болезнями репарации ДНК. Пример — пигментная ксеродерма – генная болезнь с аутосомно-рецессивным типом наследования. У больных нарушена эксцизионная репарация ДНК, которые повреждены УФ лучами и др. мутагенами. Под действием солнечного света на коже появляются веснушки, пигментные пятна, со временем у 100% больных развивается рак кожи
Схема переноса генетической информации в клетке – центральная догма молекулярной биологии
От ДНК к ДНК – редупликация ДНК.
От ДНК к РНК – транскрипция.
Возможна передача информации от РНК на ДНК – обратная транскрипция (в жизненном цикле вирусов и у эукариот)
С РНК на белок – трансляция
перейти в каталог файлов
biologo.ru
Молекулярные основы наследственности
Структура гена, его функция. Характеристика генома человека. ДНК — молекула наследственности. Химические и структурные особенности
Уже из школьной программы биологии известно, что ген — это отрезок молекулы ДНК. Только эта макромолекула из довольно обширного спектра макромолекул, существующих в каждой клетке каждого живого организма, способна самовоспроизводиться, а значит, передавать в поколениях клеток или организмов содержащуюся в ней информацию. Способность ДНК к самовоспроизведению обусловлена особенностями ее химической структуры. Молекула ДНК построена из трех компонентов: сахара, представленного дезоксирибозой, фосфатных групп и 4 типов азотистых оснований — цитозина (Ц), тимина (Т), которые еще называют пуринами, аденина (А) и гуанина (Г). Это — пиримидины.
В 1953 г. Уотсон и Крик опубликовали свою историческую статью о физической структуре ДНК. Согласно модели Уотсона и Крика, молекула ДНК представляет собой двойную спираль. Каждая спираль обвивается вокруг другой спирали вдоль общей оси. Цепи этой спирали образуют дезоксирибоза и фосфатные группы. Через определенные промежутки к каждой цепи крепится азотистое основание, обращенное внутрь спирали. Два основания каждой цепи, расположенные на одном и том же уровне, соединяются между собой.
Самое замечательное в молекуле ДНК то, что каждое азотистое основание может соединиться только с другим строго определенным и комплементарным (подходящим только для него) ему основанием, а именно аденин с тимином, а гуанин с цитозином.
Это свойство нуклеотидов комплементарно спариваться обеспечивает основу для точного воспроизведения последовательности нуклеотидов каждой цепи ДНК. Нуклеотидные цепи ДНК полярны. Полярность определяется тем, как соединяются между собой сахара (дезоксирибозы). Фосфатная группа, присоединенная к С5 (5-углерод) одного сахара, соединяется с гидроксильной группой в положении С, (3-углерод) следующего сахара с помощью фосфодиэфирной связи. В результате концевой нуклеотид на одном конце цепи имеет свободную 5-, а на другом — свободную 3-группу. Последовательность нуклеотидных оснований принято записывать в направлении от 5- к 3-концу. Две нити ДНК антипараллельны друг другу, так как идут в противоположных направлениях и 5-концу одной цепи соответствует 3-конец другой цепи и наоборот.
Модель ДНК Уотсона и Крика объяснила к тому времени хорошо известное правило английского биохимика Чаргаффа, согласно которому в любой молекуле ДНК количество пуринов строго соответствует количеству пиримидинов.
В двойной спирали ДНК пурины-(аденин, гуанин) всегда соединяются с пиримидинами (тимин и цитозин). Между цитозином и гуанином образуется три водородные связи, а между тимином и аденином — две, поэтому иным способом основания соединиться просто не могут.
Элементарной единицей ДНК является нуклеотид, в состав которого входит одна дезоксирибоза, одна фосфатная группа и одно азотистое основание.
Репликация ДНК
Поскольку ДНК является молекулой наследственности, то для реализации этого качества она должна точно копировать саму себя и таким образом сохранять всю имеющуюся в исходной молекуле ДНК информацию в виде определенной последовательности нуклеотидов. Это обеспечивается за счет особого процесса, предшествующего делению любой клетки организма, который называется репликацией ДНК. Суть этого процесса заключается в том, что специальный фермент разрывает слабые водородные связи, которые соединяют между собой нуклеотиды двух цепей. В результате цепи ДНК разъединяются и из каждой цепи «торчат» свободные азотистые основания (возникновение так называемой вилки репликации). Особый фермент ДНК-полимераза начинает двигаться вдоль свободной цепи ДНК от 5- к 3-концу молекулы, помогая присоединиться свободным нуклеотидам, постоянно синтезируемым в клетке, к 3-концу вновь синтезируемой цепи ДНК.
В результате репликации образуются две новые, абсолютно идентичные молекулы ДНК, идентичные также исходной молекуле ДНК до начала ее редупликации.
Можно сказать, несколько упрощая, что феномен точного удвоения молекулы ДНК, в основе которого лежит комплементарность оснований этой молекулы, составляет молекулярную основу наследственности.
Скорость репликации ДНК у человека относительно низкая, и для того чтобы обеспечить репликацию ДНК любой хромосомы человека, требовались бы недели, если бы репликация начиналась из одной точки. На самом деле в молекуле ДНК любой хромосомы, а каждая хромосома человека содержит только одну молекулу ДНК, имеется множество мест инициации репликации (репликонов). От каждого репликона репликация идет в обоих направлениях до тех пор, пока соседние репликоны не сливаются. Поэтому репликация ДНК в каждой хромосоме протекает относительно быстро.
Понятие «генетический код»
Для молекулы наследственности, которой является ДНК, мало того, что она сама способна самовоспроизводиться, — это только часть наследственности. ДНК должна каким-то образом кодировать все признаки организма. Большинство признаков любого организма, одноклеточного или многоклеточного, определяется белками: ферментами, структурными белками, белками-переносчиками, белками-каналами, белками-рецепторами. Следовательно, ДНК должна каким-то образом кодировать строение белков и непосредственно порядок расположения в них аминокислот.
Аминокислоты соединяются друг с другом с помощью пептидной связи, которая образуется в результате конденсации аминогруппы (NH2) одной аминокислоты с карбоксильной группой (СООН) другой аминокислоты. Последовательность аминокислот в полипептидной цепи записывают от аминокислоты со свободной NН2-группой до аминокислоты со свободной СООН-группой.
Ученые установили, что код является триплетным, это означает, что каждая аминокислота кодируется тройкой нуклеотидов. Действительно, так как для построения белков используется 20 различных аминокислот, код не может быть однонуклеотидным, поскольку существует всего 4 нуклеотида. Код не может быть также динуклеотидным, так как возможно всего 16 комбинаций из 2 нуклеотидов. При 3 нуклеотидах число комбинаций возрастает до 64, и этого вполне достаточно, чтобы кодировать 20 различных аминокислот. Кроме того, из этого также следует, что генетический код должен быть вырожденным, а именно одна аминокислота может кодироваться более чем одной тройкой нуклеотидов. Еще одним важным свойством генетического кода является то, что он неперекрывающийся, при этом каждую последовательно новую аминокислоту полипептидной цепи кодирует последовательно новый триплет ДНК. Генетический код не содержит знаков препинания, и кодирующие триплеты следуют один за другим. Генетический код является универсальным и используется одинаково как прокариотами, так и эукариотами. Кодирующие триплеты нуклеотидов получили название кодонов.
Наиболее важны первые два нуклеотида каждого кодона. Третий нуклеотид неспецифичен. Три кодона определяют сигнал прекращения синтеза полипептидной цепи (терминация трансляции): УАА, УАГ и УГА. Это означает, что в том месте информационной РНК (мРНК), где находится любой из этих кодонов, синтез полипептидной цепи белка прекращается. Кодоны, указывающие на терминацию синтеза полипептидной цепи, называют стоп-кодонами.
Информационная РНК и процесс транскрипции
Следует объяснить, зачем было так необходимо вводить понятие информационной РНК. Как известно, ДНК содержится в хромосомах клеток и, следовательно, в ядре, а белок синтезируется в цитоплазме клеток. Для того чтобы информация о структуре белка, записанная на языке ДНК, попала в цитоплазму клетки, он сначала переписывается (транскрибируется) на молекулу мРНК.
РНК отличается от ДНК тем, что в цепи РНК остаток сахара представлен рибозой (отсюда ее название), тимин замещен на урацил, который обладает примерно такой же комплементарностью к аденину, как и тимин.
Для того чтобы произошло списывание последовательности нуклеотидов гена, кодирующих первичную структуру определенной полипептидной цепи белка на мРНК, к цепи ДНК на некотором расстоянии от гена, к специальной последовательности нуклеотидов, называемых промотором, присоединяется специальный фермент — РНК-полимераза.
Стартовой точкой транскрипции служит основание ДНК, соответствующее основанию РНК, которое первым включается в транскрипт. Транскрипция мРНК продолжается до тех пор, пока РНК-полимеразе II не встретится сигнал терминации (окончания) транскрипции.
Биосинтез полипептидной цепи
В полипептидной цепи происходят расшифровка информации, закодированной с помощью генетического кода, и построение на матрице мРНК полипептидной цепи определенного белка. В этом процессе участвуют еще два вида РНК — рибосомальная (рРНК) и транспортная (тРНК). Для обоих видов РНК в геноме имеются многочисленные гены, на матрице которых эти РНК синтезируются.
Образование полипептидной цепи из последовательно доставляемых к мРНК тРНК с соответствующими аминокислотами происходит на рибосомах.
Структура гена у высших организмов достаточно сложная. В нее входят промотор, содержащий сайт инициации транскрипции, экзоны и интроны. Экзоны содержат кодирующие последовательности гена, функция интронов остается неизвестной. На границе экзонов и интронов располагается консенсусная последовательность, которая распознается ферментами сплайсинга, т. е. ферментами для вырезания интронов из первичного транскрипта мРНК. На 3-конце гена уже в некодирующей части расположен сайт, обеспечивающий добавление 100—200 остатков аденина к мРНК для обеспечения ее стабильности. Для гена характерна так называемая открытая рамка считывания, т. е. наличие достаточно длинной последовательности триплетов, кодирующих аминокислоты, не перебиваемые стоп-кодонами или бессмысленными триплетами.
Похожие статьи
medn.ru
Лекция 7 (2ч ) молекулярные основы наследственности — Лекция
Лекция 7.(2ч.).
МОЛЕКУЛЯРНЫЕ ОСНОВЫ
НАСЛЕДСТВЕННОСТИ
Хромосома представляет собой нуклеопротеидную структуру (дезоксинуклеопротеид), в состав которой входит дезоксирибо-нуклеиновая кислота, основные белки — гистоны, негистоновые белки и небольшое количество рибонуклеиновой кислоты. Ведущая роль в наследственности принадлежит ДНК, которая является носителем наследственной информации практически у всех организмов, как прокариот, так и эукариот, за исключением некоторых РНК-содержащих вирусов.
Наследственная информация осуществляется при участии рибонуклеиновых кислот.
Нуклеиновые кислоты — материальные носители наследственной информации. Нуклеиновые кислоты были открыты Фридрихом Мимером (1844—1895 гг.) в 1869 г. Из ядер клеток человека он выделил вещество, названное им нуклеином (от лат» nucleus— ядро). В дальнейшем были изучены строение и молекулярная структура нуклеина и установлено, что он представлен двумя типами нуклеиновых кислот — дезоксирибонуклеиновой кислотой (ДНК), локализованной преимущественно в ядре, и рибонуклеиновой кислотой (РНК), находящейся в ядре и цитоплазме.
Раскрытию ведущей роли нуклеиновых кислот, особенно ДНК, в наследственности предшествовали экспериментальные работы Ф. Гриффитса. Он наблюдал в 1928 г. изменение наследственного признака у бактерий пневмококков (Diplococcuspneumoniae). У этого вида имеется несколько штаммов, в том числе штаммы Sи R. Штамм S вызывает гибель животных от пневмонии. Он имеет полисахаридную слизистую капсулу и образует гладкие колонии. Штамм R— авирулентный, капсулы не имеет и образует шероховатые колонии. Ф. Гриффите заражал мышей смесью живых бескапсульных бактерий R-штамма и убитых нагреванием капсульных пневмококков S-штамма. Мыши заболевали пневмонией и погибали, а выделенные из них живые клетки были как R -, так и S-штаммов. Следовательно, произошло превращение (трансформация) некоторых бескапсульных бактерий R-штамма в вирулентные капсульные бактерии S-штамма,
В 1944 г. американский микробиолог О. Эвери с сотрудниками повторил эксперимент Ф. Гриффитса. Из бактерий штамма S он выделил ДНК и внес ее в питательную среду, на которой размножались бактерии авирулентного штамма R.
Значительная часть авирулентных бескапсульных бактерий R -штамма трансформировалась в капсульные вирулентные бактерии S-штамма. Это явление дало Эвери основание утверждать о ведущей роли ДНК в переносе наследственной информации от одного штамма бактерий к другому и послужило началом разработки молекулярной теории наследственности.
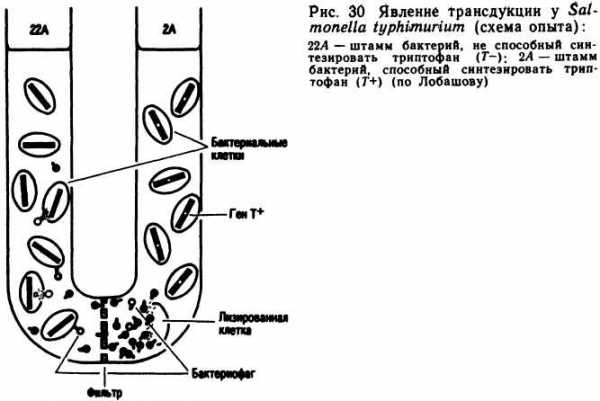
Явление трансформации в последующие годы было доказано у самых различных видов бактерий, у дрожжей, а также у многоклеточных организмов. Так, доминантный ген темной окраски яиц (грены) от одной породы тутового шелкопряда был трансформирован насекомым другой породы, имеющим белую окраску яиц.
В 1952 г. Н. Циндлер и Дж. Ледерберг открыли явление трансдукции. Трансдукцией называют перенос наследственной информации в виде фрагмента ДНК вирусами (бактериофагами) от одного штамма бактерий (донора) другому (реципиенту) и включение этого фрагмента в генотип реципиента. Явление трансдукции было открыто на тифозных бактериях, (Salmonellatyphimurium). В U-образную трубку, разделенную посредине бактериальным фильтром (рис. 30), были помещены бактерии штамма 22А, не способные синтезировать аминокислоту триптофан (Т—), и бактерии штамма 2 А, способные синтезировать данную аминокислоту (Т +). В среду был внесен бактериофаг, и после совместного культивирования некоторые клетки штамма 22А приобрели способность синтезировать триптофан. Таким образом было доказано, что бактериофаги могут являться переносчиками наследственной информации от бактерий одного штамма бактериям другого штамма.
Американский ученый Хейши совместно с М. Чейзомв 1952 г. размножали ДНК-содержащий вирус — бактериофаг Т-2 на среде, содержащей радиоактивные серу и фосфор, что позволило пометить ДНК фага радиоактивным фосфоромР35, а белковую оболочку фага — радиоактивной серой S33. Призаражении бактерий данными фагами в них попадала толькомеченая ДНК, а белковая оболочка, меченная радиоактивной серой, оставалась снаружи. В зараженной клетке образовалось множество вирионов фага, Следовательно, генетическая информация, необходимая для синтеза ДНК фагов, содержится в ДНК проникших в клетку вирусов.
Дезоксирибонуклеиновая кислота — ДНК — является уникальным носителем наследственной информации как у прокариотов, так и у эукариотов. Только у некоторых форм простейших вирусов наследственная информация закодирована не в ДНК, а в рибонуклеиновой кислоте.
Доказательством ведущей роли ДНК в наследственности является то, что, она локализована главным образом в хромосомах, поэтому молекулярная генетика не противоречит хромосомной теории наследственности и законам классической генетики. Количество ДНК в клетках каждого организма относительнопостоянно, причем в половых клетках — гаметах — количество ее в два раза меньше, чем в соматических, что соответствует поведению хромосом в мейозе и при оплодотворении. Одним из главнейших свойств ДНК является ее способность самоудваиваться (реплицироваться) в интерфазе митотического цикла, благодаря чему в каждой клетке многоклеточного организма сохраняется полный объем наследственной информации. ДНК относительно константна.
Особенности строения молекулы ДНК свидетельствуют об ее исключительном многообразии, видовой и индивидуальной специфичности. Изменение в строении молекулы ДНК обусловливает изменение признака или свойства организма.
Строение молекулы ДНК. ДНК — сложный биополимер, состоящий из 108 нуклеотидов и более. Каждый нуклеотид включает три компонента — остаток фосфорной кислоты (фосфат), пентозный сахар — дезоксирибозу и одно из четырех азотистых оснований: пуриновых — аденин или гуанин — либо пиримидиновых — тимин или цитозин.
Специфичность каждого нуклеотида в молекуле ДНК определяется наличием соответствующего азотистого основания, поэтому нуклеотиды принято обозначать начальными буквами азотистых оснований: А — аденин, Г — гуанин, Т — тимин, Ц — цитозин. Нуклеотиды соединяются между собой, образуя длинную цепочку,

химическим остовом, которой служат остатки фосфорной кислоты, которые связаны фосфодиэфирными связями с 5′ углеродом одной молекулы пентозного сахара и 3′ углеродом другой (рис. 31). К первому атому углерода каждой молекулы пентозного сахара присоединяется одно из четырех азотистых оснований. Благодаря такому соединению нуклеотидов молекула ДНК обладает полярностью: репликация ДНК на матричной нити идет в направлении от 5’к 3′.Структурная формула молекулы ДНК была установлена в 1953 г. Д. Уотсоном и Ф. Криком (рис. 32). Молекула ДНК состоит из двух цепочек нуклеотидов, соединенных комплементарно. Каждый нуклеотид одной цепочки соединяется водородными связями с нуклеотидом другой цепочки строго закономерно: аденин соединяется с тимином, гуанин — с цитозином. Аденин соединяется с тимином двумя, а цитозин с гуанином — тремя водородными связями. Число пурииовых нуклеотидов (А+ +Г) равно числу пиримидиновых Ц+Т, то есть отношение (А+ +Г) : (Т+Ц) =1. Две комплементарные нити образуют правовинтовую спираль, каждый виток которой имеет длину 3,4 нм, расстояние между нуклеотидами 0,34 нм. Азотистые основания ориентированы к середине спирали. Для хромосом эукариотов характерно линейное строение молекулы ДНК, у прокариотов, плазмид, митохондрий и пластид молекулы ДНК бывают замкнуты в кольцо.
Видовая специфичность молекулы ДНК. Число нуклеотндов и их последовательность в молекуле ДНК специфичны для каждого вида и частично — для каждой особи. Д. Уотсон ввел понятие о видовой специфичности ДНК. Коэффициентом видовой
специфичности называют соотношение .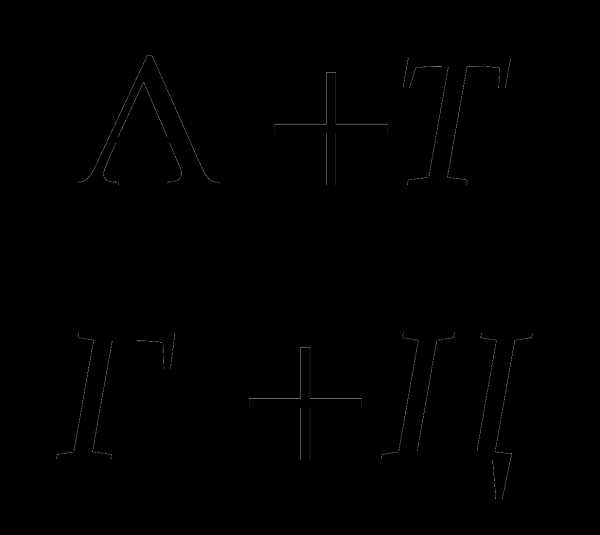 Молекула ДНК
Молекула ДНК
обладает исключительным многообразием. Если, предположить, что у млекопитающих в ДНК содержится 108 нуклеотидов, то число молекул ДНК, различающихся по порядку чередования нуклеотидов, будет 4 в степени 108. Таким образом, в молекуле ДНК доожет быть записан практически любой объем наследственной информации и у каждой особи эта запись уникальна и специфична.
Репликация молекулы ДНК. Репликацией называют процесс самокопирования молекулы ДНК с точным соблюдением порядка чередования нуклеотидов, присущего исходным комплементарным нитям.
У многоклеточных организмов в результате слияния гамет при оплодотворении образуется зигота, в которой содержится наследственная информация гаплоидных геномов родительских особей. В онтогенезе из зиготы в результате митоза образуются миллиарды клеток, каждая из которых несет в себе всю генетическую информацию. Об этом свидетельствует тот факт, что у многих видов растений — табака, картофеля, бегонии, моркови и других — из одной клетки, взятой из корня, листа, клубня или другого органа, можно получить целое растение, имеющее характерные признаки исходного. Это возможно благодаря тому, что молекула ДНК способна к самовоспроизведению в процессе репликации.
Репликация происходит в период синтеза (5-период) интерфазы митотического цикла. На отдельных участках молекулы ДНК образуются так называемые вилки репликации. В этих местах водородные связи между азотистыми основаниями под действием соответствующих ферментов разрываются, комплементарные нити разъединяются и каждая из них становится матрицей, на которой происходит синтез дочерних нитей. Такой тип репликации получил название полуконсервативного; он был убедительно доказан в опытах М. Мезельсон и,Ф. Сталь (1958) на бактериях Е. coli и Дж. Тейлором на проростках семян кормовых бобов Vicia faba.
Бактерии кишечной палочки сначала выращивали на среде, содержащей тяжелый изотоп азота —N15. Затем они были перенесены на среду, содержащую только N14. Через определенные промежутки времени из клеток выделяли ДНК и с помощью центрифугирования в соответствующем растворителе разделяли нафракций. Молекулы ДНК, выделенные из исходных бактерий, содержали изотоп N15. По мере роста бактерий на среде, содержащей N14, сначала в ДНК содержались нити, одна из которых включала N14, другая — N15, а затем наряду с молекулами, нити которых содержали N14+N16, появились молекулы ДНК, содержащие только N14 (рис. 33).
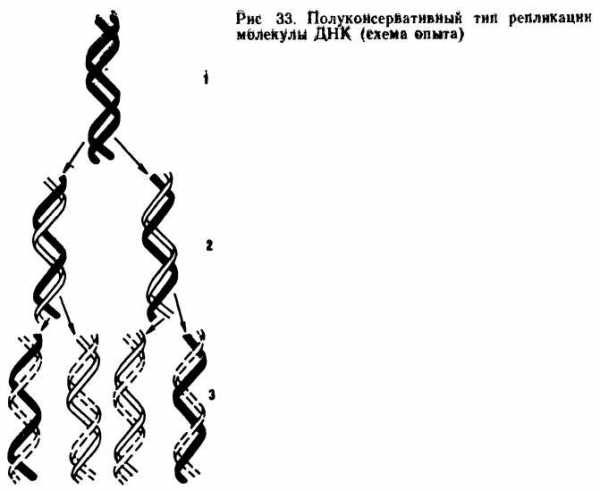
Дж. Тейлор для доказательства полуконсервативного типа репликации ДНК использовал метод радиоавтографии. Сначала семена конских бобов проращивали на среде, содержащей тимидин, меченный тритием — Н3. Затем корешки проростков, в клетках которых содержались хромосомы, меченные Н3, переносили в среду, не содержащую радиоактивный тимидин. В первом митозе каждая хроматида содержала Н3, во втором одна из двух хроматид была меченой, другая не содержала Н3. В третьем митозе только часть хромосом содержала одну хроматиду, меченную Н3. Впоследствии эти данные были подтверждены исследованиями, проведенными на культуре клеток растений, животных и человека.
Процесс репликации протекает при участии комплекса ферментов, главнейшие из которых получили название ДНК-полимераз, одновременно на двух комплементарных нитях. Участок молекулы ДНК в том месте, где начали расплетаться комплементарные нити, называются вилкой репликации.Она образуется у прокариот, плазмид, митохондрий и пластид в одной определенной, генетически фиксированной точке. В молекуле ДНК у эукариот таких «стартовых точек» бывает несколько.
У эукариот на каждой комплементарной нити ДНК процесс репликации идет неодинаково, так как они антипараллельны, поэтому одна из нитей называется «лидирующей», другая — «запаздывающей». «Лидирующая» нить синтезируется от 5′ конца к 3′ при участии фермента ДНК-полимеразы в виде сплошной комплементарной нити (рис. 34).
Синтез «запаздывающей» нцти протекает сложнее с участием комплекса ферментов. Вначале образуются отрезки — реплики новой дочерней нити ДНК, прочное соединение которых осуществляет фермент лигаза. Эти отрезки новой нити ДНК содержат у эукариот 100 — 200 нуклеотидов, у прокариот — 1000 — 2000 нуклеотидов. Их называют фрагментами Оказаки по имени описавшего их японского ученого.

Репликация кольцевых молекул ДНК у прокариот, а также ДНК плазмид, митохондрий и пластид протекает по типу, получившему название «катящегося обруча». При этом одна из нитей молекулы ДНК разрывается и ее конец прикрепляется к клеточной мембране, а на противоположном конце, как на матрице, происходит синтез дочерней нити ДНК.
Репликация ДНК протекает довольно быстро, У бактерий она составляет около 30 мкм в минуту; за это время к нити-матрице присоединяется около 500 нуклеотидов дочерней нити. У вирусов — около 900 нуклеотидов в минуту. У эукариот репликация протекает медленнее — дочерняя нить удлиняется на 1,5 — 2,5 мкм в минуту.
Таким образом, ДНК способна самовоспроизводиться (реплицироваться, самокопироваться) и сохранять наследственную информацию, закодированную в ней в виде последовательности чередования нуклеотидных оснований, во множестве поколений клеток, образующихся в онтогенезе многоклеточного организма.
Реализация наследственной информации. Наследственная информация, закодированная в молекуле ДНК, реализуется на всех этапах жизнедеятельности клетки и многоклеточного организма в процессе биосинтеза. Основанием для доказательства реализации наследственной информации в процессе биосинтеза послужили исследования американских ученых Дж. Бидла и Э. Л. Татума в 1941—1944 гг., которыми были получены мутантные штаммы плесневого гриба нейроспоры. Их различие состояло в утрате способности к синтезу той или иной аминокислоты и потере свойства синтезировать соответствующий ферментативный белок. Исследования показали, что каждый ген контролирует синтез одного соответствующего фермента («один ген — один фермент») и реализация наследственной информации осуществляется в процессе синтеза. Ген, локализованный на определенном участке молекулы ДНК, контролирует синтез первичной молекулы белка, представляющей собой полипептидную цепь, специфичность которой зависит от порядка чередования в ней аминокислот.
Белкам принадлежит исключительно важная роль в жизнедеятельности каждой клетки и всего многоклеточного организма. Они участвуют в построении мембран, хроматина, рибосом, митохондрий, являются составной частью сложных белков. В качестве ферментов и гормонов они управляют всеми процессами в клетке и в многоклеточном организме. Подавляющее большинство метаболических реакций, от которых зависит развитие признака или свойства, находится под контролем ферментов и, следовательно, генов.
Химическая структура белковых молекул. Первичная молекула белка представляет собой цепочку, состоящую из 100 — 300 различных аминокислот и более, порядок чередования которых определяет специфичность данной молекулы (рис. 35): каждая из 20 аминокислот может встречаться многократно, но местонахождение контролируется ДНК. В настоящее время для многих молекул белка установлена их первичная структура, то есть порядок чередования аминокислот в полипептидной цепи.
Вторичная структура белковой молекулы зависит от первичной: аминокислоты в полипептидной цепи соединяются водородными связями между NH- и СО — группами, в результате чего она свертывается в так называемую альфа-спираль. Образование больших альфа-спиральных участков характерно для фибриллярных белков. В молекулах ферментов спиралеобразных участков значительно меньше (рис. 36)
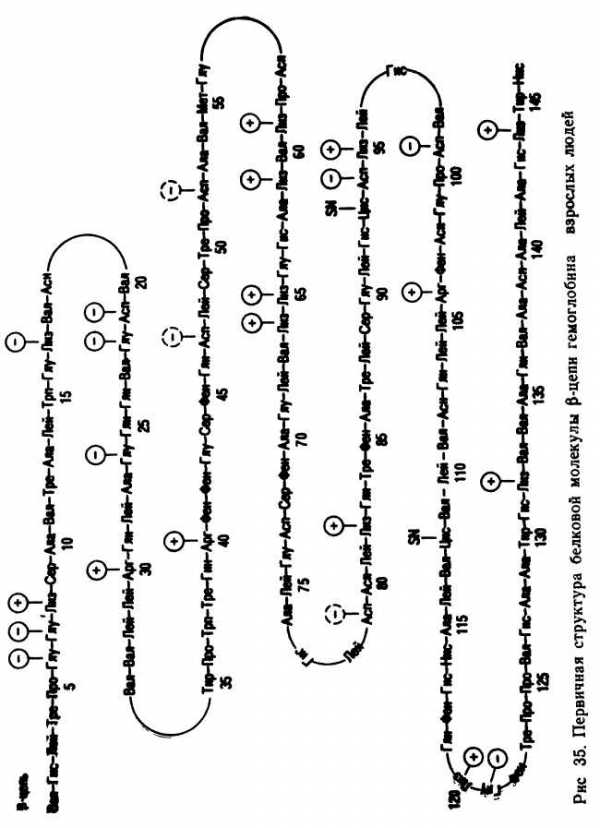
Третичная структура белковых молекул образуется в результате связывания так называемыми дисульфидными мостиками (S—S) двух цистеиновых остатков аминокислот. Это определяет специфическое пространственное расположение полипептидных цепей.
Четвертичная структура белковых молекул характеризуется тем, что они состоят из двух — четырех различных, стабильно соединенных полипептидных цепей. Такая структура характерна для глобулярных белков, в том числе и для многих ферментов.
Вторичная, третичная и четвертичная структуры белковых молекул зависят от числа и порядка чередования аминокислот в полипептидной цепи, то есть от первичной структуры. Процесс синтеза белка в клетке называется биосинтезом. Он осуществляется под контролем молекулы ДНК, которая таким образом реализует закодированную в ней наследственную информацию. Схематично реализацию наследственной информации можно представить следующим образом:
Биосинтез. Процесс реализации наследственной информации в биосинтезе осуществляется при участии трех видов рибонуклеиновых кислот: информационной (матричной) — иРНК (мРНК), рибосомальной — рРНК и транспортной — тРНК. Все рибонуклеиновые кислоты синтезируются на соответствующих участках молекулы ДНК. Они имеют значительно меньшие размеры, чем ДНК, и представляют собой одинарную цепь нуклеотидов. Нуклеотиды содержат остаток фосфорной кислоты (фосфат), пентозный сахар (рибозу) и одно из четырех азотистых оснований— аденин, цитозин, гуанин и урацил. Азотистое основание— урацил — комплементарно аденину.
Процесс биосинтеза сложный и включает ряд этапов — транскрипцию, сплайсинг и трансляцию.
Первый этап называется транскрипцией. Он происходит в ядре клетки: на участке определенного гена молекулы ДНК синтезируется мРНК. Этот синтез осуществляется при участии комплекса ферментов, главным из которых является ДНК-зависимая РНК-полимераза, которая прикрепляется к начальной (инициальной) точке молекулы ДНК, расплетает двойную спираль и, перемещаясь вдоль одной из нитей, синтезирует рядом с ней комплементарную нить мРНК. В результате транскрипции мРНК содержит генетическую информацию в виде последовательного чередования нуклеотидов, порядок которых точно скопирован с соответствующего участка (гена) молекулы ДНК.
Дальнейшие исследования показали, что в процессе транскрипции синтезируется так называемая про-мРНК — предшественник зрелой мРНК, участвующей в трансляции. Про-мРНК имеет значительно большие размеры и содержит фрагменты, не кодирующие синтез соответствующей полипептидной цепи. В ДНК наряду с участками, кодирующими рРНК, тРНК и полипептиды, имеются фрагменты, не содержащие генетической информации. Они получили название интронов в отличие от кодирующих фрагментов, которые называются экзонами. Интроны обнаружены на многих участках молекул ДНК. Так, например, в одном гене — участке ДНК, кодирующем овальбумин курицы, содержится 7 интронов, в гене сывороточного альбумина крысы — 13 интронов. Длина интрона бывает различной — от двухсот до тысячи пар нуклеотидов ДНК. Интроны считываются (транскрибируются) одновременно с экзонами, поэтому про-мРНК значительно длиннее, чем зрелая мРНК. В ядре в про-мРНК специальными ферментами вырезаются интроны, а фрагменты экзона «сращиваются» между собой в строгом порядке. Этот процесс называется сплайсингом. В процессе сплайсинга образуется зрелая мРНК, которая содержит только ту информацию, которая необходима для синтеза соответствующего полипептида, то есть информативную часть структурного гена.
Значение и функции интронов до сих пор еще не совсем выяснены, но установлено, что, если в ДНК считываются только участки экзонов, зрелая мРНК не образуется. Процесс сплайсинга изучен на примере работы гена овальбумина. Он содержит один экзон и 7 интронов. Сначала на ДНК синтезируется про-мРНК, содержащая 7700 нуклеотидов. Затем в про-мРНК
число нуклеотидов уменьшается до 6800, затем — до 5600, 4850, 3800, 3400 и т. д. до 1372 нуклеотидов, соответствующих экзону. Содержащая 1372 нуклеотида мРНК выходит из ядра в цитоплазму, попадает на рибосому и синтезирует соответствующий полипептид.
Следующий этап биосинтеза — трансляция — происходит в цитоплазме на рибосомах при участии тРНК.
Транспортные РНК синтезируются в ядре, но функционируют в свободном состоянии в цитоплазме клетки. Одна молекула тРНК содержит 76 — 85 нуклеотидов и имеет довольно сложную структуру, напоминающую клеверный лист (рис. 37). Три участка тРНК имеют особо важное значение 1) антикодон, состоящийиз трех нуклеотидов, определяющий место прикрепления тPHK к соответствующему комплементарному кодону (мРНК) на рибосоме; 2) участок, определяющий специфичность тРНК, способность данной молекулы прикрепляться только к определенной аминокислоте; 3) акцепторный участок, к которому прикрепляется аминокислота. Он одинаков для всех тРНК и состоит из трех нуклеотидов — Ц—Ц—А. Присоединению аминокислоты к тРНК предшествует ее активация ферментом аминоацил-тРНК-синтетазой. Этот фермент специфичен для каждой аминокислоты Активированная аминокислота прикрепляется к соответствующей тРНК и доставляется ею на рибосому.
Центральное место в трансляции принадлежит рибосомам— рибонуклеопротеиновым органоидам цитоплазмы, во множестве в ней присутствующим. Размеры рибосом у прокариот в среднем 30X30x20 нм, у эукариот — 40X40X20 нм. Обычно их размеры определяют в единицах седиментации (S) — скорости осаждения при центрифугировании в соответствующей среде. У бактерии кишечной палочки рибосома имеет
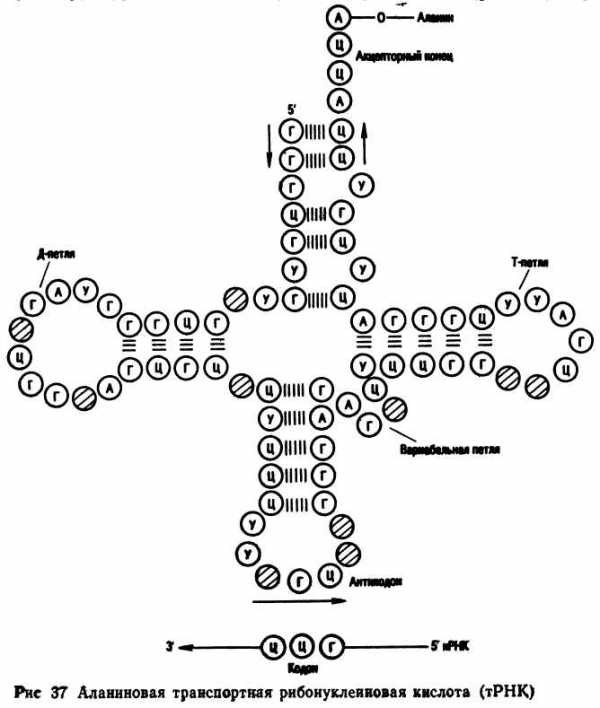
величину 70S и состоит из двух субчастиц, одна из которых имеет константу 30S, вторая 505, и содержит 64% рибосомальной РНК и 36% белка.
Молекула мРНК выходит из ядра в цитоплазму и прикрепляется к малой субчастице рибосомы. Трансляция начинается с так называемого стартового кодона (инициатора синтеза) — А —У—Г— Когда тРНК доставляет к рибосоме активированную аминокислоту, ее антикодон соединяется водородными связями с нуклеотидами комплементарного кодона мРНК. Акцепторный конец тРНК с соответствующей аминокислотой прикрепляется кповерхности большой субчастицы рибосомы. После первой аминокислоты другая тРНК доставляет следующую аминокислоту, и таким образом на рибосоме синтезируется полипептидная цепь. Молекула мРНК обычно работает сразу на нескольких (5—20) рибосомах, соединенных в полисомы. Начало синтеза полипептидной цепи называют инициацией, рост ее — элонгацией. Последовательность аминокислот в полипептидной цепи определяется последовательностью кодонов в мРНК. Синтез полипептидной цепи прекращается, когда на мРНК появляется один из кодонов-терминаторов — УАА, УАГ или УГА. Окончание синтеза данной полипептидной цепи называется терминацией (рис. 38).
Установлено, что в клетках животных полипептидная цепь за одну секунду удлиняется на 7 аминокислот, а мРНК продвигается на рибосоме на 21 нуклеотид.
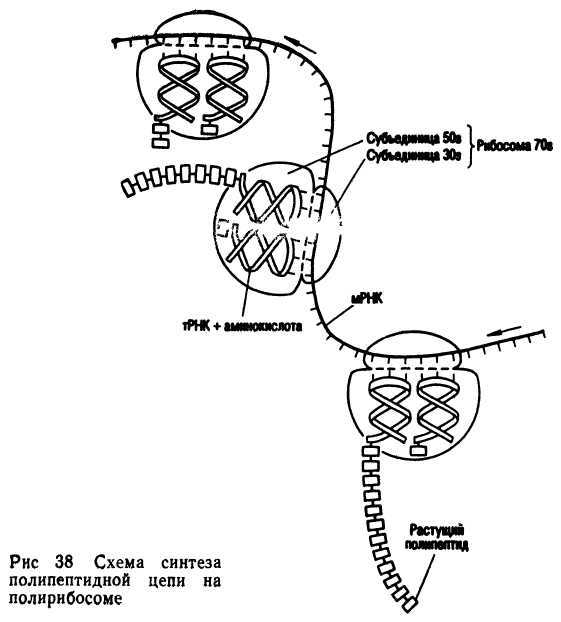
У бактерий этот процесс протекает в два-три раза быстрее.
Следовательно, синтез первичной структуры белковой молекулы— полипептидной цепи — происходит на рибосоме в соответствии с порядком чередования нуклеотидов в матричной рибонуклеиновой кислоте — мРНК. Она не зависит от строения рибосомы.
Генетический код. Кодом наследственности или генетическим кодом называется процесс перевода триплетной последовательности нуклеотидов молекулы ДНК в последовательность аминокислот в белковой молекуле. Одним из важнейших свойств генетического кода является его колинеарность — четкое соответствие между последовательностями кодонов нуклеиновых кислот и аминокислотами полипептидных цепей (табл.2).
Важное значение для раскрытия генетического кода имели исследования М. Ниренберга и Дж. Маттеи, а затем С. Очао с сотрудниками, начатые ими в 1961 г. в США. Они разработали метод и экспериментально установили последовательность нуклеотидов в кодонах мРНК, контролирующая местоположение данной аминокислоты в полипептидной цепи. В бесклеточную среду, содержащую все аминокислоты, рибосомы, тРНК, АТФ и ферменты, Ниренберг и Маттеи вводили искусственно синтезированный биополимер типа мРНК, представляющий собдй цепочку одинаковых нуклеотидов — УУУ — УУУ — УУУ — УУУ — и т. д. Биополимер кодировал синтез полипептидной цепи, содержащей только одну аминокислоту — фенилаланин; такая цепь называется полифенилаланином. Если мРНК состояла из кодонов, содержащих нуклеотиды с азотистым основанием ци-тозин, то синтезируется полипептидная цепь, содержащая аминокислоту пролин, — полипролин. Искусственные полимеры мРНК, содержащие кодоны АГУ, синтезировали полипептидную цепь из аминокислоты серии — полисерин и т. д. Этот метод позволил в начале 60-х годов полностью расшифровать генетический код и определить его свойства (табл. 3),
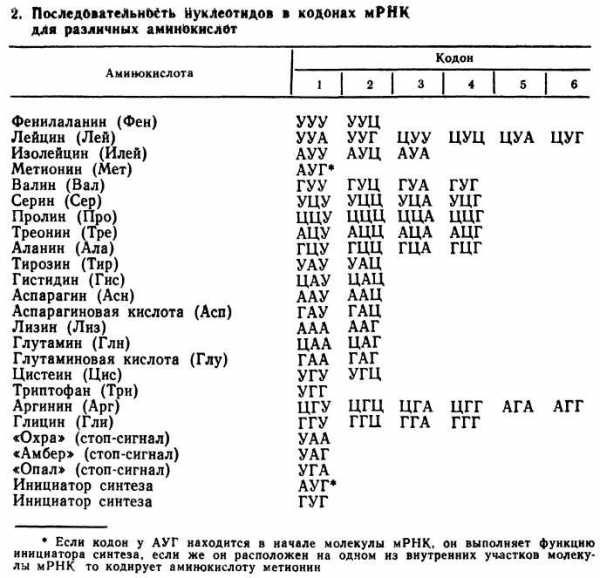
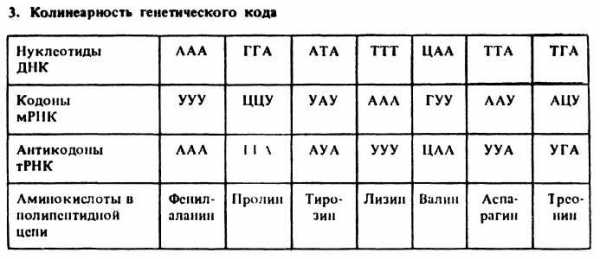
Свойства генетического кода
Генетический код универсален — един для всех организмов (вирусов, бактерий, растений, животных и человека).
Код триплетный. Местоположение каждой аминокислоты кодируется сочетанием строго определенных трех нуклеотидов в мРНК, образующих один специфический кодон.
Код вырожденный. Одна аминокислота может кодироваться несколькими (от одного до шести) кодонами. Только две аминокислоты кодируются одним триплетом — метионин (АУГ) и триптофан (УГГ).
textarchive.ru
