
Параграф 5 — Броздецкий В.С. Рабочие матeриалы.
Глава 2
Явления и закономерности жизни на клеточном уровне
Изучив главу, вы сможете характеризовать:
• состав и строение клетки;
• свойства клеточных органоидов;
• процессы жизнедеятельности клетки.
Вы сумеете:
• определять различия в строении клеток эукариот и прокариот;
• оценивать роль автотрофов и гетеротрофов в природе;
• объяснять значение обмена веществ в жизнедеятельности клетки;
• сравнивать механизмы протекания процессов биосинтеза белка, фотосинтеза и дыхания.
Параграф 5. Многообразие клеток
Вспомните
• какой структурный уровень организации жизни характеризует клетка;
• что существуют одноклеточные и многоклеточные организмы.
Из истории изучения многообразия клеток. История изучения клетки неразрывно связана с развитием микроскопической техники. О существовании клеток стало известно лишь в XVII в. В 1665 г. английский естествоиспытатель Р. Гук, оценив значение увеличительного прибора, впервые применил его для исследования срезов некоторых растительных и животных тканей. Под микроскопом он обнаружил структуры, похожие по строению на пчелиные соты, и назвал их «ячейками», или «клетками». С тех пор этот термин прочно утвердился в биологии.
В 1674 г. голландский натуралист А. ван Левенгук впервые рассмотрел под самодельным микроскопом некоторых простейших и отдельные клетки животных (эритроциты, сперматозоиды).
В 30-х г. XIX в. шотландский учёный Р. Браун обнаружил в клетках растений круглое плотное образование, которое он назвал ядром.
В 1838 г., обобщая имевшиеся к тому времени сведения о клетке, немецкий ботаник М.Я. Шлейден первым пришёл к заключению о том, что ядро является обязательным структурным элементом всех растительных клеток. В 1839 г. немецкий физиолог Т. Шванн, основываясь на работах Шлейдена, изложил основы клеточной теории, согласно которой все ткани животных и растительных организмов состоят из клеток, клетки растений и животных имеют общий принцип строения, каждая отдельная клетка самостоятельна, а жизнедеятельность всего организма проявляется как совокупность жизнедеятельности отдельных групп клеток.
Появление клеточной теории Шлейдена и Шванна обусловило дальнейшее развитие учения о клетке. В 1858 г. немецкий патолог Р. Вирхов доказал, что клетки возникают только путём воспроизведения себе подобных. Ему принадлежит афористическое утверждение: «Каждая клетка — от клетки». В конце XIX в. была выдвинута гипотеза о том, что информация о наследственных свойствах организмов заключена в ядре.
Крупный вклад в развитие учения о клетке внесли русские учёные. В 1892 г. И.И. Мечников открыл явление фагоцитоза (от греч. phagos — «пожиратель», kytos — «клетка») — активного захватывания и поглощения различных частиц одноклеточными организмами и даже клетками многоклеточных организмов. В 1898 г. С.Г. Навашин описал особый тип оплодотворения — двойное оплодотворение, свойственное всем цветковым растениям.
В начале XX в. были разработаны методы культивирования клеток в пробирке и сконструирован первый электронный микроскоп. В результате наука обогатилась сведениями о мельчайших, ранее не известных клеточных структурах. Было доказано, что клетки всех организмов, несмотря на их разнообразие, сходны по строению, химическому составу и проявлениям своей жизнедеятельности.
Дальнейшие исследования показали, что ядерные структуры клетки служат основой передачи наследственных свойств организмов.
Мир клеток живой природы
. Клетки чрезвычайно разнообразны. Они различаются по своим размерам, структуре, форме и функциям. Размеры клеток варьируют от 0,1—0,25 мкм (некоторые бактерии) до 15—21 см (яйцо страуса в скорлупе).
Есть свободноживущие клетки, которые ведут себя как особи популяций и видов. Их жизнедеятельность зависит не только от слаженной работы внутриклеточных структур, но и от особенностей существования клетки как самостоятельного организма (добывание пищи, способ питания, размножение, подвижность в окружающей среде, активное и неактивное переживание неблагоприятных условий и пр.).
Клетки свободноживущие (1) и образующие ткани (2)
Одноклеточных организмов чрезвычайно много. Их представители встречаются среди всех царств живой природы и населяют все среды жизни на нашей планете.
У многоклеточных организмов разные клетки выполняют различные функции. Клетки, сходные по строению, расположенные рядом, объединённые межклеточным веществом и предназначенные для выполнения определённых (специализированных) функций в организме, образуют ткани. Ткани возникли в ходе эволюционного развития одновременно с появлением многоклеточности, так как специализация клеток и, следовательно, тканей способствовала лучшему обеспечению процессов жизнедеятельности целостного организма.
У животных различают четыре типа (группы) тканей: эпителиальную, соединительную, мышечную и нервную; у растений — пять типов (групп) тканей: образовательную, покровную, проводящую, механическую, основную.
Клетки всех организмов на Земле принципиально сходны по своему строению, химическому составу и основным проявлениям жизни. При этом процессы жизнедеятельности (дыхание, биосинтез, обмен веществ) происходят в клетках независимо от того, являются они одноклеточными организмами или составными частями многоклеточного организма.
Жизнь многоклеточного организма зависит от жизнедеятельности его отдельных клеток и их групп, выполняющих особые, специализированные функции.
Свойства клетки. Особенность клетки определяется специфичностью её составных компонентов, упорядоченностью происходящих в ней как в целостной системе процессов. Живая клетка осуществляет процессы, от которых зависит её жизнь: она поглощает пищу, извлекает из неё энергию, избавляется от отходов обмена веществ, поддерживает постоянство своего химического состава и воспроизводит саму себя. Всё это позволяет рассматривать клетку как особую единицу живой материи, как элементарную живую систему — биосистему клеточного уровня организации жизни.
Клетка — основная структурная и функциональная единица жизни.
Из клеток состоят все живые существа — от одноклеточных до крупных растений, животных и человека. У всех организмов клетки функционируют, с одной стороны, как самостоятельные биосистемы, а с другой — как взаимосвязанные части целого.
Два типа клеток.
В первой половине XX в. было обнаружено, что в клетках бактерий нет оформленного ядра, отделённого от цитоплазмы мембраной, хотя присутствует само ядерное вещество, несущее наследственную информацию. В клетках растений, животных и грибов ядро хорошо сформировано и отграничено от цитоплазмы.
Клетки, не имеющие оформленного ядра, называют прокариотическими (лат.pro — «перед», «раньше» и греч. karyon — «ядро»), а имеющие ядро — эукариотическими (лат. ей — «полностью» и греч. karyon — «ядро»). По этому признаку все организмы делят на две группы: доядерные (прокариоты) и ядерные (эукариоты).
Клетки прокариот имеют достаточно простое строение, так как сохраняют черты первых организмов, возникших на Земле. Эукариоты могут быть одноклеточными и многоклеточными, их клетки имеют более сложное строение, чем у прокариот, и отличаются большим разнообразием.
Клетки бактерий и цианобактерий
Разнообразие форм клеток прокариот: бактерии и цианобактерии
Жизнедеятельность организма зависит от согласованного функционирования всех его частей, мельчайшей из которых является клетка. За прошедшие миллиарды лет строение клетки не только усложнилось — она оказалась способной жить и активно функционировать в составе специализированных тканей многоклеточных организмов, сохраняя при этом свойства биосистемы и являясь основной структурной единицей жизни. В процессе воспроизведения клетки осуществляется передача наследственной информации, что обеспечивает непрерывность жизни на Земле.
Разнообразие форм клеток эукариот — растений и животных: 1 — спирогира; 2 — эвглена зелёная; 3 — клетка нервной ткани многоклеточного организма; 4 — клетка мышечной ткани многоклеточного организма; 5 — сувойка
Животная и растительная клетки
1. Назовите признаки сходства и различия клеток одноклеточных и многоклеточных организмов.
2. Что лежит в основе деления всех организмов на прокариот и эукариот?
3. Почему клетку называют структурной единицей жизни?
4. Сравните особенности жизнедеятельности свободноживущей клетки и клетки многоклеточного организма.
www.bvc56.ru
Тип простейшие. Общая характеристика и строение простейших
Тип простейшие включает примерно 25 тыс. видов одноклеточных животных, обитающих в воде, почве или организмах других животных и человека. Имея морфологическое сходство в строении клеток с многоклеточными организмами, простейшие существенно отличаются от них в функциональном отношении.
Если клетки многоклеточного животного выполняют специальные функции, то клетка простейшего является самостоятельным организмом, способным к обмену веществ, раздражимости, движению и размножению.
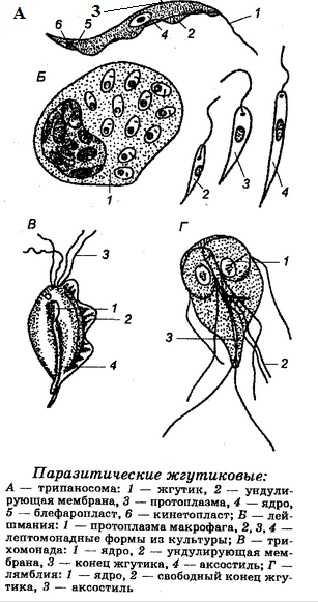 Представители простейших
Представители простейшихПростейшие — это организмы на клеточном уровне организации. В морфологическом отношении простейшее равноценно клетке, но в физиологическом представляет собой целый самостоятельный организм. Подавляющее большинство их — микроскопически малых размеров (от 2 до 150 мкм). Однако некоторые из ныне живущих простейших достигают 1см, а раковины ряда ископаемых корненожек имеют в диаметре до 5-6 см. Общее количество известных видов превышает 25 тыс.
Строение простейших
Строение простейших чрезвычайно разнообразно, но все они обладают чертами, характерными для организации и функции клетки. Общим в строении в строении простейших являются два основных компонента тела — цитоплазма и ядро.

Цитаплазма
Цитоплазма ограничена наружной мембраной, которая регулирует поступление веществ в клетку. У многих простейших она усложняется дополнительными структурами, увеличивающими толщину и механическую прочность наружного слоя. Таким образом возникают образования типа пелликулы и оболочки.
Цитоплазма простейших обычно распадается на 2 слоя — наружный более светлый и плотный — эктоплазму и внутренний, снабженный многочисленными включениями,— эндоплазму.
В цитоплазме локализуются общеклеточные органоиды. Кроме того, в цитоплазме многих простейших могут присутствовать разнообразные специальные органеллы. Особенно широко распространены различные фибриллярные образования — опорные и сократимые волоконца, сократительные вакуоли, пищеварительные вакуоли и др.
Ядро
Простейшие обладают типичным клеточным ядром, одним или несколькими. Ядро простейших имеет типичную двухслойную ядерную оболочку. В ядре распределен хроматиновый материал и ядрышки. Ядра простейших характеризуются исключительным морфологическим многообразием по размерам, числу ядрышек, количеству ядерного сока и т.д.
Особенности жизнедеятельности простейших
В отличие от соматических клеток многоклеточные простейшие характеризуются наличием жизненного цикла. Он слагается из ряда следующих друг за другом стадий, которые в существовании каждого вида повторяются с определенной закономерностью.
Чаще всего цикл начинается стадией зиготы, отвечающей оплодотворенному яйцу многоклеточных. За этой стадией следует однократно или многократно повторяющееся бесполое размножение, осуществляемое путем клеточного деления. Затем образуются половые клетки (гаметы), попарное слияние которых вновь дает зиготу.
Важной биологической особенностью многих простейших является способность к инцистированию. При этом животные округляются, сбрасывают или втягивают органеллы движения, выделяют на своей поверхности плотную оболочку и впадают в состояние покоя. В инцистированном состоянии простейшие могут переносить резкие изменения окружающей среды, сохраняя жизнеспособность. При возвращении благоприятных для жизни условий цисты раскрываются и простейшие выходят из них в виде активных, подвижных особей.
Простейшие обладают очень широким распространением. Множество их живет в море. Некоторые входят в состав пресноводных водоемов. Существуют виды, живущие во влажной почве. Широкое распространение среди всех классов простейших получил паразитизм. Многие виды паразитических простейших вызывают тяжелые заболевания человека, домашних и промысловых животных, растений.
По строению органоидов движения и особенностей размножения тип простейшие делится на 6 классов. Основные 4 класса: Саркодовые, Жгутиковые, Споровики и Инфузории.
animals-world.ru
(жр) Свободноживущие азотфиксирующие микроорганизмы — Zbio
Материал из Zbio
В. К. Шильникова[править] Азотобактер (Azotobacter)
В 1901 году Бейеринк выделил из почвы аэробную неспорообразующую грамотрицательную бактерию, фиксирующую молекулярный азот, и назвал ее Azotobacter chroococcum (в родовом названии отражена способность бактерии фиксировать азот, в видовом — способность синтезировать коричневый пигмент — chroo и образовывать кокковидные клетки — coccum). Азотобактер — типичный представитель свободноживущих микроорганизмов. Свободноживущие — это все те микроорганизмы, которые живут в почве независимо от того, развивается вблизи растение или нет.
 Рис. 172. Клетки Az. vinelandii (1), Az. chroococcum (2) и Az. agilis (3). Увел. ×1000. (По Г. Иенсену).
Рис. 172. Клетки Az. vinelandii (1), Az. chroococcum (2) и Az. agilis (3). Увел. ×1000. (По Г. Иенсену).
Культуры азотобактера в лабораторных условиях отличаются полиморфизмом. Клетки разных видов азотобактера в молодом возрасте показаны на рисунке 172. Молодые клетки азотобактера подвижны; они имеют многочисленные или единичные жгутики (рис. 173). У азотобактера обнаружены выросты, подобные фимбриям (рис. 174). В старых культурах клетки азотобактера покрываются плотной оболочкой, образуя цисты. Они могут прорастать, давая начало молодым клеткам (рис. 175).
Полиморфизм азотобактера зависит в значительной степени от состава среды, на которой он выращивается. На среде с этиловым спиртом (в качестве единственного источника углерода) азотобактер длительное время сохраняет подвижность и форму палочек. В то же время на многих других средах полиморфизм проявляется очень резко.
На плотных питательных средах, не содержащих азота, азотобактер образует крупные слизистые, иногда морщинистые колонии (рис. 176), окрашивающиеся при старении в желтовато-зеленоватый, розовый или коричнево-черный цвет. Колониям разных видов азотобактера присуща своя специфическая пигментация.
К настоящему времени известен ряд видов азотобактера: Azotobacter chroococcum, Az. beijerinckii, Az. vinelandii, Az. agilis, Az. nigricans, Az. galophilum.
Источником азота для азотобактера могут служить разнообразные минеральные (соли аммония, азотной и азотистой кислот) и органические (мочевина, различные аминокислоты) соединения. Однако если азотобактер развивается только за счет связанного в среде азота, он не выполняет своей основной функции — фиксации молекулярного азота. Азотобактер обычно фиксирует до 10-15 мг молекулярного азота на 1 г использованного источника углерода (например, глюкозы, сахарозы). Эта величина сильно колеблется в зависимости от условий выращивания культуры, состава питательной среды, ее кислотности, температуры, аэрации.
Рис. 173. Делящиеся клетки азотобактера (A. agilis), видны перитрихиальные жгутики (1), у A. macrocytogenes видны полярные жгутики (2) (по А. Бейлю и др.).По отношению к источникам углерода В. Л. Омелянский (1923) назвал азотобактер полифагом («всеядным»).
Азотобактер хорошо усваивает разнообразные углеводы (моно- и дисахара, некоторые полисахариды), органические кислоты, многоатомные спирты (глицерин, маннит) и другие вещества.
Многим исследователям удавалось выращивать азотобактер в чашках с питательной средой без азота и углерода, если чашки были помещены в камеру, где находились пары ацетона, этилового спирта или некоторых других органических соединений. В присутствии легкодоступных форм углеродсодержащих соединений азотобактер может частично использовать углекислоту из атмосферы. Повышение концентрации углекислого газа до 0,5% в воздухе несколько стимулирует развитие азотобактера. Но азотобактером лучше усваиваются легкодоступные формы углеродсодержащих органических соединений. В почве запас подвижного органического вещества невелик, поэтому именно недостаток легкодоступных соединений углерода в первую очередь ограничивает развитие азотобактера в природных условиях.
Какие же органические соединения может использовать в почве азотобактер? Перегнойные вещества почвы азотобактером практически не усваиваются. Поэтому в почвах, даже очень богатых перегноем, при отсутствии свежих органических остатков интенсивного размножения азотобактера не происходит.
Однако, если в почве имеются органические соединения и продукты распада растительных и животных клеток, азотобактер развивается хорошо. В частности, он усиленно размножается в почвах, удобренных соломой и соломистым навозом, а также в разнообразных компостах, содержащих целлюлозу. Азотобактер хорошо ассимилирует вещества, образующиеся при распаде целлюлозы.
Развитие азотобактера и фиксация им азота в значительной степени зависят от наличия в среде фосфора. Источником фосфора могут служить как органические, так и минеральные фосфорсодержащие соединения. Высокая чувствительность азотобактера к фосфору позволила разработать микробиологический метод определения потребности почв в фосфорных удобрениях.
 Рис. 174. Фимбриоподобные образования у клеток азотобактера. Увел. ×24000. (По Э. В. Болтянской.)
Рис. 174. Фимбриоподобные образования у клеток азотобактера. Увел. ×24000. (По Э. В. Болтянской.)
В качестве тест-организма в этом методе используется азотобактер. Микробиологические методы определения потребности почвы в удобрении имеют ряд преимуществ перед химическими анализами, хотя, безусловно, уступают в точности.
Важную роль в обмене веществ азотобактера играет кальций. Этот элемент необходим азотобактеру при питании как молекулярным, так и аммонийным азотом (Г. Н. Зайцева, 1965). Недостаток кальция в среде приводит к сильной вакуолизации клеток и их вздутию.
Высокая чувствительность азотобактера к кальцию, так же как и к фосфору, используется для определения потребности почв в известковании.
Микроэлементы (молибден, бор, ванадий, железо, марганец) необходимы азотобактеру в первую очередь для осуществления процесса азотфиксации. Потребность в микроэлементах определяется в значительной степени геохимическими условиями существования азотобактера в почвах. Штаммы микроорганизма, выделенные из почв с высоким естественным содержанием того или иного микроэлемента, нуждаются, как правило, в более высоких концентрациях этих элементов.
Интересно, что радиоактивные элементы (радий, торий, уран) оказывают стимулирующий эффект на развитие азотобактера и процесс азотфиксации.
Азотобактер чрезвычайно чувствителен к реакции среды. Оптимальная для его развития область рН 7,2-8,2. Однако азотобактер способен развиваться и на средах с рН от 4,5 до 9,0; кислая реакция среды неблагоприятно действует на его развитие. Из кислых почв выделяются неактивные формы азотобактера, утратившие способность связывать молекулярный азот.
Большое влияние на развитие азотобактера оказывает влажность почвы. Клетки азотобактера имеют меньшее осмотическое давление, чем клетки грибов и актиномицетов; потребность во влаге аналогична потребности высших растений. Азотобактер распространен в пресных водоемах, илах, затопляемых рисовых полях, сточных водах, сильно увлажненных почвах, на водных растениях в прудах и водохранилищах. Это свидетельствует о его высокой степени гидрофильности. На основании высокой потребности во влаге почвенных форм азотобактера предполагается, что предки некоторых морских и почвенных видов азотобактера могли быть общими.
Рис. 175. Цисты азотобактера (по Й. Чану и др.). Зрелая циста, наполненная гранулами жира и окруженная толстой плотной оболочкой (справа), и прорастающая циста (растущая молодая клетка разрывает оболочку цисты — слева). Увел. ×35000.В отношении температуры азотобактер является типичным мезофильным организмом, с оптимумом развития около 25-30°С. Понижение температуры азотобактер переносит хорошо, поэтому зимой даже в северных широтах численность его клеток в почве заметно не уменьшается.
Из биологических факторов, влияющих на развитие азотобактера, следует прежде всего отметить почвенные микроорганизмы. Они могут оказывать влияние на жизнедеятельность азотобактера в почве косвенно, изменяя, например, рН или окислительно-восстановительные условия, и непосредственно, вырабатывая питательные и биологически активные вещества. Так, активирующее влияние целлюлозоразрушающих и маслянокислых микроорганизмов на развитие азотобактера и его антагонистические отношения с представителями почвенной микрофлоры отмечали многие советские и зарубежные исследователи. Биоценоз микроорганизмов, формирующийся в условиях той или иной почвы, меняется в значительной степени под влиянием растительного покрова. И азотобактер как член биоценоза также зависит от этого фактора. С помощью метода радиоавтографии установлено, что при нанесении меченных по фосфору клеток азотобактера на семена зерновых культур клетки обычно концентрируются вокруг растущей корневой системы проростков.
Имеются, однако, данные, что клеток азотобактера в ризосфере растений очень мало. В самом лучшем случае (при полном отсутствии антагонистов и благоприятных окружающих условиях) их количество не превышает 1% от общего числа ризосферной микрофлоры.
Культуры азотобактера, как правило, образуют значительное количество биологически активных веществ: витамины группы В, никотиновую и пантотеновую кислоты, биотин, гетероауксин и гиббереллин. Однако, несмотря на то что культуры азотобактера вырабатывают целую серию биологически активных веществ, внесение витаминов, гиббереллина и гетероауксина в среду ускоряет рост азотобактера. Реакция на дополнительное внесение витаминов в среду является индивидуальной особенностью штаммов.
Азотобактер может продуцировать ростовые вещества типа ауксинов. Это подтверждается опытами, в которых было установлено образование дополнительных корешков у черенков фасоли под влиянием ауксинов, вырабатываемых азотобактером. Биологический тест — карликовая форма гороха сорта Пионер — позволяет определить в культуре азотобактера гиббереллиноподобные соединения.
 Рис. 176. Развитие колоний A. chroococcum вокруг комочков почвы на безазотной среде.
Рис. 176. Развитие колоний A. chroococcum вокруг комочков почвы на безазотной среде.
Все эти соединения в совокупности способны стимулировать прорастание семян растений и ускорять их рост в тех, конечно, случаях, когда на корневой системе растений находится достаточное количество клеток азотобактера.
Кроме того, была обнаружена антагонистическая активность азотобактера по отношению к возбудителям бактериальных болезней растений. Азотобактер синтезирует фунгистатический (задерживающий развитие грибов) антибиотик группы анисомицина. Ряд грибных организмов, встречающихся на семенах и в почве (виды из родов Fusarium, Alternaria, Penicillium), может угнетать развитие многих видов растений, особенно в холодную погоду. Азотобактер, продуцируя противогрибные антибиотические вещества, помогает растениям расти и развиваться, что имеет особенно большое значение в ранние фазы развития.
К сожалению, способность азотобактера активно размножаться в почве и проявлять свои многогранные качества весьма ограничена из-за дефицита легкодоступных органических веществ в почве и высокой требовательности микроорганизма к окружающим условиям. Поэтому стимулирующий эффект азотобактера проявляется лишь на плодородных почвах.
Распространение азотобактера в почвах Советского Союза имеет определенные закономерности. В целинных подзолах и дерново-подзолистых почвах, характеризующихся кислой реакцией, условия для развития азотобактера неблагоприятны. Только окультуривание таких почв создает возможности для его развития. В почвах с повышенным увлажнением и преобладанием луговой растительности (почвы пойм) азотобактер обычно встречается в течение всего вегетационного периода в больших количествах. В торфяниках азотобактер или отсутствует, или развивается очень слабо. В зоне достаточно увлажненных северных мощных черноземов азотобактер развивается хорошо, а в зоне обычных и южных черноземов при отсутствии орошения, а также в целинных и неполивных окультуренных каштановых почвах только как весенний эфемер. Максимальное развитие азотобактера в весенний иериод наблюдается и в целинных и в богарных почвах сероземной зоны. В солонцах и солончаках распространены преимущественно солестойкие расы азотобактера. В основном в почвах нашей страны доминирует Az. chroococcum.
[править] Бейеринкия (Beijerinckia)
Впервые аэробные бактерии рода Beijerinckia были выделены из кислых почв рисовых полей в Индии (в 1939 г.). Г. Деркс (1950), обнаружив эту бактерию в почве Ботанического сада в Богоре (Ява), предложил назвать ее именем М. Бейеринка — одного из первых исследователей фиксаторов азота.
 Рис. 177. Колонии бейеринкии разных типов (гладкие и складчатые варианты): 1-4, 6-8, 10 — по Н. И. Гогорикидзе; 5, 9, 11 — по Дж. Бекингу.
Рис. 177. Колонии бейеринкии разных типов (гладкие и складчатые варианты): 1-4, 6-8, 10 — по Н. И. Гогорикидзе; 5, 9, 11 — по Дж. Бекингу.
Клетки бактерий рода Beijerinckia имеют круглую, овальную или палочковидную формы; палочки иногда искривлены. Размеры молодых клеток 0,5-2,0 х 1,0-4,5 мкм. Встречаются подвижные и неподвижные формы. Цист и спор не образуют. Культуры характеризуются медленным ростом. Типичные колонии формируются обычно через 3 недели при 30°С. Большинство культур Beijerinckia образует на безазотном агаре с глюкозой выпуклые, нередко складчатые, блестящие слизистые колонии очень вязкой консистенции (рис. 177). При старении культуры, как правило, образуют темноокрашенный пигмент.
На 1 г использованного энергетического материала организмы рода Beijerinckia фиксируют 16-20 мг молекулярного азота. Спектр углеродсодержащих соединений, доступных бейеринкии, значительно уже, чем у азотобактера. Хорошо используются моно- и дисаха-риды, хуже — крахмал, органические кислоты, ароматические вещества не усваиваются. Минеральный азот и многие аминокислоты бактерии рода Beijerinckia предпочитают молекулярному азоту.
Основные отличия бейеринкии от азотобактера — высокая кислотоустойчивость (могут расти даже при рН 3,0), кальцефобность (ничтожные дозы кальция подавляют рост), устойчивость к высоким концентрациям железа, алюминия.
Бактерии рода Beijerinckia широко распространены в почвах южной и тропической зоны, реже встречаются в зоне умеренного климата. Beijerinckia часто встречается на поверхности листьев тропических растений в Индонезии.
Раньше полагали, что бактерии рода Beijerinckia могут существовать только в кислых почвах. Сейчас установлено, что они неплохо развиваются и в нейтральных и щелочных почвах. Тем не менее следует полагать, что Beijerinckia играют значительную роль в азотном балансе главным образом кислых почв (латеритах, красноземах), не имея существенного агрономического значения для нейтральных почв.
[править] Клостридиум (Clostridium)
Первый анаэробный микроорганизм, усваивающий молекулярный азот, был выделен и описан С. Н. Виноградским в 1893 г. Он оказался спорообразующей бактерией, которой было дано наименование Clostridium pasteurianum (родовое название происходит от латинского слова clostrum — веретено; видовое — pasteurianum — дано в честь Луи Пастера).
 Рис. 178. Клетки Clostridium pasteurianum со спорами. Увел. ×3500 (по В. И. Дуде).
Рис. 178. Клетки Clostridium pasteurianum со спорами. Увел. ×3500 (по В. И. Дуде).
Клетки Cl. pasteurianum крупные, их длина 2,5-7,5 мкм, ширина 0,7-1,3 мкм. Располагаются они поодиночке, парами или образуют короткие цепочки. Молодые клетки подвижны, имеют перитрихиально расположенные жгутики, плазма их гомогенна. При старении клетки плазма становится гранулированной, в ней накапливается гранулеза (вещество типа крахмала). В центре клетки или ближе к ее концу формируется спора, которая в поперечнике значительно шире, чем вегетативная клетка, и поэтому клетка в этот период приобретает форму веретена. Размер спор 1,3 × 1,6 мкм. На рисунке 178 изображены клетки Cl. pasteurianum со спорами. Морфология спор и поведение ядерного вещества в процессе спорообразования у клостридиев детально описаны в разделе Спорообразование у анаэробных бактерий В. И. Дудой.
Азотфиксирующая функция выявлена у многих представителей рода Clostridium: Cl. pasteurianum, Cl. butyricum, Cl. butylicum, Cl. beijerinckia, Cl. pectinovorum, Cl. acetobutylicum и других видов. Наиболее энергичный азотонакопитель — Cl. pasteurianum — фиксирует 5-10 мг азота на 1 г потребленного источника углерода.
Наряду с молекулярным азотом бактерии рода Clostridium хорошо усваивают минеральные и органические азотсодержащие соединения. В качестве источника углеродного питания бактерии рода Clostridium используют различные соединения, которые обычно одновременно служат для них и источником энергии. К фосфору, калию и кальцию они значительно менее чувствительны, чем азотобактер. Однако удобрение почв фосфорно-калийными солями, известкование почв или компостов всегда приводит к возрастанию численности.
Клостридии относительно устойчивы к кислой и щелочной реакции среды. Область рН, при которой их развитие протекает нормально, довольно широка; минимальное значение рН ниже 4,5, максимальное — выше 8,5.
Влияние воздушно-водного режима на развитие бактерий рода Clostridium изучено достаточно полно. Будучи анаэробными, они хорошо переносят высокое насыщение почвы влагой. Однако оптимальная степень увлажнения для них определяется типом почвы и обеспеченностью органическим веществом. Лучше всего клостридии развивается при влажности почвы, равной 60-80% от полной влагоемкости.
Больше всего бактерий рода Clostridium в верхних слоях почвы, которые богаты органическими веществами.
Бактерии рода Clostridium по-разному относятся к температуре, встречаются как мезофильные, так и термофильные бактерии. Молекулярный азот фиксируют лишь мезофилы.
У мезофильных форм оптимальная температура развития чаще всего находится в диапазоне 25-30°С. Предел максимальных температур 37-45°С.
Споры клостридиев весьма устойчивы к высоким температурам. Они выдерживают нагревание при 75°С в течение 5 ч и в течение 1 ч нагревание при 80°С. Споры термофильных клостридиев погибают при кипячении через 30 мцн. Более высокая температура (110°С) быстро убивает их.
Со многими микроорганизмами в почве Clostridium находится в метабиотических отношениях, при которых предполагается обмен продуктами метаболизма. Так, азотобактер улучшает условия жизни клостридия, поглощая кислород, а клостридии вырабатывает из органических соединений, недоступных азотобактеру, органические кислоты, которые может ассимилировать азотобактер.
Было бы трудно ответить на вопрос: в каких почвах нет Clostridium? «Всеядность» Clostridium, малая взыскательность к условиям внешней среды, а также способность в неблагоприятных условиях переходить в состояние спор объясняют их широкое, практически повсеместное распространение.
Накопления азота в почвах за счет деятельности Clostridium, однако, невелики и не превышают, как правило, нескольких килограммов на один гектар почвы.
[править] Другие роды азотфиксирующих бактерий
До недавнего времени считалось, что монополия азотфиксации принадлежит только представителям родов Azotobacter, Clostridium и Rhizobium.
Правда, еще до выделения М. Бейеринком (1901) азотобактера и почти одновременно с С. Н. Виноградским (1895) клостридия французский исследователь Бертло (1885-1892) описал большое количество разнообразных почвенных микроорганизмов, обладающих азотфиксирующей функцией. Некоторые из них повышали содержание азота в среде на 80% (от исходного). Однако отсутствие в ряде случаев идентификации выделенных культур и несовершенство используемых Бертло методик не давали права согласиться с утверждениями о значительной широте распространения азотфиксирующей функции среди почвенных микроорганизмов.
За последнее время использование высокочувствительного изотопного метода и других точных приемов исследований позволило установить способность многих почвенных микроорганизмов связывать атмосферный азот. Список азотоусвоителей пополнился новыми видами бактерий: Azotomonas insolita, Azotomonas fluorescens, Pseudomonas azotogensis. Установлена азотфиксирующая способность у ряда известных бактерий, таких, как актиномицеты (Actinomycetes), фотосинтезирующие бактерии (Rhodospirillum rubrum), представители эубактерий (семейств Thiorhodaceae, Athiorhodaceae, а также родов: Pseudomonas, Bacterium, Bacillus). Наконец, выявлено, что некоторые виды микроорганизмов, не фиксирующие азот в обычных условиях культивирования, могут стать энергичными азотфиксаторами в ассоциациях с другими видами микроорганизмов.
Сейчас известно свыше 80 видов и разновидностей бактерий, несколько видов актиномицетов, дрожжей, дрожжеподобных организмов и плесневых грибов, способных фиксировать азот. Они населяют почву, дно морей и пресных водоемов.
Остановимся на характеристике лишь основных представителей бактерий.
Способность фиксировать азот присуща ряду представителей семейства Pseudomonadaceae. Они довольно широко распространены в природе. К наиболее интересным представителям этого семейства относится несколько видов: Azotomonas insolita, фиксирующий до 12 мг азота на 1 г использованного сахара и встречающийся нередко не только в почве, но и как эпифит на растениях; Azotomonas fluorescens, выделенный впервые из компостов Н. А. Красильниковым (1945), продуктивность азотфиксации которого не меньше, чем у первого вида; Pseudomonas azotocolligans, населяющий кислые и щелочные почвы Америки; Pseudomonas azotogensis, широко распространенный в почвах Канады и легко выделяющийся из парниковых почв, и, наконец, Pseudomonas methanitrificens, который встречается в почвах, имеющих выход источников природных газов. Последняя бактерия особенно интересна тем, что она использует метан и другие газообразные углеводороды в качестве единственного источника углерода, за счет энергии разложения которых она усваивает азот атмосферы.
Представители семейства Spirillaceae, фиксирующие молекулярный азот, распространены преимущественно в рыбоводных прудах, озерах, морской воде, морских отложениях. Возможно, они играют немаловажную роль в фиксации азота в водоемах.
Не только азотобактеру, как члену семейства Azotobacteriaceae, свойственна азотфиксирующая функция. В почвах Индии встречаются еще два представителя этого семейства — Derxia gummosa и Derxia indica — активные, хотя и медленнодействующие, азотфиксаторы. В азотном балансе почв они, как и бактерии Agrobacterium radiobacter из семейства Rhizobiaceae, по-видимому, не играют сколько-нибудь значительной роли. Молекулярному азоту они предпочитают связанные источники азота, такие, как мочевина, пептон, аминокислоты и минеральные соединения.
Среди спорообразующих грамположительных бактерий семейства Bacillaceae азотфиксирующая способность выявлена у факультативных анаэробов Bacillus polymyxa, аэробов Bacillus megaterium и Thermobacillus azotofigens. Последняя бактерия, выделенная из удобренной навозом дерново-карбонатной почвы Эстонской ССР, оказалась термофильной с оптимумом роста 45-50°С и максимумом 60-65°С. При температурах ниже 20°С она не развивается.
Распространение функции азотфиксации в ряде семейств фотосинтезирующих бактерий (семейства Thiorhodaceae, Athiorhodaceae, Chlorobacteriaceae, Hyphomicrobiaceae) не случайно, так как, по-видимому, они являются представителями одной из древнейших групп азотфиксаторов на Земле.
Небольшие количества молекулярного азота способны усваивать почвенные микобактерии. Усвоенный микобактериями азот в сочетании с азотом, ассимилированным другими олигонитрофильными микроорганизмами, сине-зелеными водорослями, лишайниками и мхами, настолько обогащает примитивную почву азотом, что ее могут заселять высшие растения.
molbiol.ru
Строение и жизнедеятельность одноклеточных
Строение тела Простейших
Замечание 1
Клетка простейших – это самостоятельный организм со всеми необходимыми жизненными функциями.
Жизненные функции одноклеточного животного представлены обменом веществ, раздражимостью, размножением и перемещенем в пространстве.
Определение 1
Простейшие – эукариотические организмы, клетки которых имеют оформленное ядро с наличием генетической информацией.
Клетка животного ограничена от внешней среды мембранной. Мембрана выполняет таким образом барьерную и защитную функцию. Цитоплазма имеет два слоя: эктоплазма и эндоплазма, внешний и внутренний соответственно.
Эндоплазма клетки содержит органеллы, которые имеют постоянную структуру и выполняют определенные функции. Органеллы эукариотических клеток представлены митохондриями, эндоплазматической сеткой, рибосомами, аппаратом Гольджи, лизосомами и ядром. Также цитоплазме присущи органеллы — это характерно только для простейших: это сократительные и пищеварительные вакуоли, базальное тельце – у жгутиковых представителей, и глазки.
Определение 2
Сократительная вакуоль – это органелла осморегуляции, которая непосредственно контролирует поток воды в клетку.
Сократительная вакуоль важна для пресноводных организмов. Вода накапливается в сократительной вакуоли в спокойном состоянии, а вовремя ее сокращения вода выбрасывается наружу. Также функциями сократительной вакуоли является: выделение продуктов обмена веществ вместе с водой, дыхание – с водой кислород попадает вовнутрь. Для морских видов интенсивность работы вакуоли зависит от солености воды, она также может отсутствовать вообще.
Определение 3
Пищеварительная вакуоль – органелла, выполняющая функцию пищеварения.
Определение 4
В пищеварительной вакуоли имеются ферменты, способные перерабатывать поглощенные частички пищи. Способ захвата твердых частиц клеткой называется фагоцитозом.
При помощи ложноножек у амеб, жгутиков у эвглены и ресничек у инфузории осуществляется их передвижение в пространстве. У автотрофов имеется органелла (хроматофор), которая содержит фотосинтезирующие пигменты, например у вольвокса.
Раздражимость
Как и всему живому на Земле, простейшим присуща раздражимость.
Определение 5
Раздражимость – способность реагировать на изменения окружающей среды.
Раздражимость подразделяется на позитивный таксис и негативный таксис.
Определение 6
Позитивный таксис – перемещение организма к источнику раздражения.
Определение 7
Негативный таксис – перемещение организма от источника раздражения.
По типу раздражения различают фототаксис и хемотаксис.
Фототаксис – раздражение на свет. Хемотаксис – раздражение на химические вещества.
Экология Одноклеточных животных
По типу питания Простейшие, или Одноклеточные животные, могут быть фототрофами или гетеротрофами. Встречается и смешанный тип питания (Эвглена зеленая).
Пример 1
Например, Эвглена зеленая, в условиях яркого освещения ведет себя как фототроф, а в темноте, при наличие органики, переходит в гетеротроф.
Среди гетеротрофов встречаются сапрофиты и паразиты. Среди простейших встречаются свободноживущие и паразитические формы. Свободноживущие организмы способны к активному передвижению, что обеспечивается псевдоподиями.
Замечание 2
Псевдоподии – непостоянные выросты клетки.
Многие свободноживущие животные образуют колонии. В колонии каждая клеточка является самостоятельным организмом. К свободноживущим организмам относятся: амеба, радиолярий, инфузория, хламидомонада, вольвокс.
Паразитические формы живут засчет других организмов, находятся в их внутренней среде (многие Простейшие живут в организме человека). К паразитам относят: амеба дизентерийная (вызывает дизентерию), малярийный плазмодий (вызывает малярию).
Обитают простейшие в водоемах, морях, почве и живых организмах. Большинство простейших имеют способность образовывать на стадии покоя в неблагоприятных условиях цисты. Циста покрыта защитной оболочкой, что обеспечивает выживание при неблагоприятных условиях и размножения животного. При наступлении благоприятных условий организм покидает оболочку цисты и продолжает привычную для себя жизнедеятельность. Размножение одноклеточных организмов происходит делением клетки пополам при бесполом процессе. Многие одноклеточные размножаются половым способом, а некоторые смешанным.
spravochnick.ru
Свободноживущие микроорганизмы, фиксирующие молекулярный азот
К настоящему времени установлено, что многие свободноживущие бактерии — представители около 30 видов — могут фиксировать молекулярный азот. Большое значение в фиксации азота имеет семейство Azotobacteriaceae (бактерия Azotobacter chroococcum, открытая М. Бейеринком и др.).
Молодые клетки азотобактера представляют собой палочки размером 2—3X4—6 мкм. Позже они превращаются в крупные кокки диаметром до 4 мкм. Кокковидные клетки обычно покрываются капсулой и содержат разные включения (жир, крахмал и т. д.).
Иногда у кокковидных клеток появляется толстая оболочка, и они превращаются в цисты. На одних питательных средах палочки быстро приобретают кокковидную форму, на других — по истечении длительного времени. Палочковидные клетки микроба имеют жгутики и обладают подвижностью. При переходе палочек в кокки жгутики обычно теряются (рис. 34).
Из описанных видов азотобактера наиболее изучены Az. chroococcum, Az. vinelandii и Az. agilis. Эти виды различаются между собой размерами и формой клетки, а также некоторыми другими признаками, в частности пигментацией колоний. Так, колонии Az. chroococcum имеют бурый, почти черный цвет. Для Az. agilis характерны бесцветные колонии, Az. vinelandii дает флюоресцирующую желтовато-зеленоватую окраску среды. Наиболее обычен для почвы Аz. chroococcum.
Все виды азотобактера аэробы. В качестве источника азота могут ассимилировать соли аммония, нитриты, нитраты и аминокислоты. При отсутствии связанных форм азота фиксируют молекулярный азот. Небольшие дозы азотсодержащих соединений не депрессируют фиксацию азота, а иногда даже стимулируют ее. Увеличение дозировки связанного азота в среде полностью подавляет усвоение молекулярного азота.
Энергия усвоения азота у отдельных культур азотобактера колеблется в широком диапазоне. Активные культуры связывают 15—20 мг азота на 1 г потребленного органического вещества.
Азотобактер способен использовать огромный набор органических соединений — моно — и дисахариды, некоторые полисахариды (декстрин, крахмал), многие спирты, органические кислоты, в том числе ароматические. Вообще азотобактер проявляет высокую потребность в органических веществах. Поэтому он в больших количествах встречается в почвах, хорошо заправленных органическими удобрениями.

Для роста азотобактер нуждается в элементах минерального питания, особенно в фосфоре и кальции. Потребность азотобактера в этих элементах столь значительна, что его используют в качестве биологического индикатора на наличие фосфора и кальция в почве.
Для энергичной фиксации молекулярного азота азотобактеру и другим фиксаторам азота нужны микроэлементы. Важное значение имеет молибден, который входит в состав ферментов, катализирующих процесс усвоения азота.
Отмеченные физиологические особенности азотобактера определяют экологию данного организма. Он обитает в высокоплодородных, достаточно влажных почвах с нейтральной или близкой к ней реакцией среды. При дефиците увлажнения большинство клеток этого микроорганизма отмирают. Во многих черноземах, каштановых и сероземных почвах, благоприятных для азотобактера, его обнаруживают в значительных количествах лишь весной. При летнем иссушении почвы в ней остаются немногочисленные его зародыши. В зоне подзолистых и дерново-подзолистых почв азотобактер можно найти в огородных и пойменных почвах, богатых органическими соединениями, с благоприятным значением pH.
Отметим, что из ризосферы тропического злака Paspalum notatum был выделен Azotobacter, дающий колонии зеленого тона и имеющий некоторые физиологические отличия от типовой культуры. Этот микроорганизм назван Azotobacter paspali.
К семейству Azotobacteriaceae относятся бактерии рода Beijerinckia, близкие по свойствам к азотобактеру. От азотобактера они отличаются значительной кислотоустойчивостью, кальцифобностью и некоторыми другими свойствами. Они могут расти даже в среде с pH около 3.
Впервые бактерия рода Beijerinckia была получена из кислых почв рисовых полей Индии Р. Старки и П. Де (1939). Ее назвали Azotobacter indicum. Материал, накопленный позднее, показал, что рассматриваемый микроорганизм относится к особому роду, названному в честь известного голландского ученого М. Бейеринка — Beijerinckia. К настоящему времени описан ряд видов бактерий этого рода.
Клетки Beijerinckia имеют различную форму — палочковидную, овальную или круглую. У некоторых видов они подвижны, у других неподвижны. Иногда наблюдается образование капсул. Цисты и эндоспоры отсутствуют.
Большинство культур бактерий рода Beijerinckia дают на безазотной среде с глюкозой выпуклые, блестящие, нередко складчатые слизистые колонии вязкой консистенции. При старении колонии окрашиваются в красноватый или темно-коричневый цвет. В отличие от азотобактера, Beijerinckia не усваивает ароматические соединения и хуже ассимилирует органические кислоты. При развитии на углеводах в среде накапливаются кислые продукты (уксусная и другие органические кислоты). Бактерии рода Beijerinckia менее требовательны по сравнению с азотобактером к концентрации фосфорных соединений в среде. Небольшие дозы соединений кальция тормозят рост Beijerinckia. Значительно менее, чем азотобактер, Beijerinckia чувствительна к повышенной концентрации солей железа и алюминия. Beijerinckia нуждается в молибдене, но довольствуется меньшими его дозами.
Бактерии рода Beijerinckia широко распространены в кислых почвах субтропической и тропической зон. Реже они встречаются в почвах зоны умеренного климата. Бактерии рода Beijerinckia найдены в красноземах Грузии. В окультуренных кислых почвах юга содержится больше клеток Beijerinckia, чем в целинных. Целинные луговые почвы богаче бактериями рода Beijerinckia, чем лесные.
К свободноживущим фиксаторам молекулярного азота семейства Azotobacteriaceae относятся также виды рода Derxia, получившего свое наименование в честь голландского микробиолога Г. Деркса. Это медленно растущие на безазотных средах палочковидные бактерии, обладающие в определенной стадии развития жгутиком. Колонии могут быть пленочными или слизистыми. При старении они становятся желтовато-коричневыми. Derxia обнаружена в почвах тропической зоны, как и бактерии рода Azomonas, также близкие к азотобактеру. Последние имеют округлые клетки и образуют колонии с флюоресцирующим пигментом.
Как показали исследования последнего времени, в почве могут находиться азотфиксирующие бактерии рода Klebsiella (семейство Enterobacteriaceae), представляющие собой грамотрицательные, подвижные с перитрихиальным расположением жгутиков, факультативно-анаэробные палочки. Они выносят довольно низкое значение pH и в большом количестве имеются в лесных подзолистых почвах. В зоне умеренного климата они встречаются и под травянистой растительностью.
К данному семейству относится род Erwinia, представители которого, населяющие надземную часть травянистых растений, могут усваивать атмосферный азот.
К азотфиксаторам относятся и некоторые расы Pseudomonas fluorescens (семейство Pseudomonadaceae). Этот аэробный микроб богаче представлен в почвах северной зоны.
Среди бактерий, фиксирующих молекулярный азот, имеются вибрионы и спириллы. Недавно большой интерес вызвали работы Ж. Доберейнер (Бразилия), показавшей, что в тропических почвах на корнях травянистых растений присутствуют аэробные фиксирующие молекулярный азот спириллы. В то время как большинство сапрофитных азотфиксаторов населяют почву прикорневой зоны (ризосферу), спириллы находятся на самой поверхности корня (ризоплане) и подчас даже проникают в корневую систему. Таким образом, эти микроорганизмы тесно связаны с растениями, и можно ожидать, что они оказывают существенное влияние на их рост.
Изучение физиологических свойств отмеченных спирилл показало, что существует, по крайней мере, две их группы — Azospirillum lipoferum и Azospirillum brasilense, отличающиеся некоторыми физиологическими свойствами. Растения с типом С4 –фотосинтеза (кукуруза, просо, сорго и т. д.) имеют на корневой системе преимущественно A. lipoferum, а растения с типом С3-фотосинтеза (рис, пшеница и др.) —A. brasilense. Исключение составляет сахарный тростник, заражаемый преимущественно A. brasilense.
Достаточно широкие исследования ряда микробов позволяют заключить, что бактерии рода Azospirillum встречаются на корневой системе растений и в более северной зоне, доминируя в зоне южных почв.
Обнаружены и другие аэробные азотфиксирующие бактерии, экология и роль которых в почвенных процессах еще достаточно не изучена. Это представители рода Arthrobacter, относящегося к коринебактериям — микробам, на определенных этапах развития образующим искривленные, неправильной формы клетки и т. д.
Из анаэробных азотфиксаторов довольно хорошо изучены представители рода Clostridium (семейство Bacillaceae).
Как отмечалось выше, первым из свободноживущих фиксаторов азота был открыт Clostridium pasteurianum, Этот микроорганизм имеет палочковидные клетки длиной 1,5—8 мкм и шириной 0,8— 1,3 мкм. Молодые клетки несут перитрихиально расположенные жгутики, старые — образуют споры (см. рис. 29). При спорообразовании клетки утолщаются посередине или на конце. В присутствии кислорода воздуха Cl. pasteurianum может развиваться только при наличии в среде аэробных бактерий, поглощающих кислород, малочувствителен к реакции среды и встречается как в кислых (pH 4,5—5,5), так и в щелочных (pH 8—9) почвах. Споры Cl. pasteurianum довольно устойчивы: они выдерживают нагревание до 96° С в течение нескольких минут.
В качестве источника азотного питания бактерии рода Clostridium могут использовать соли аммония, азотной кислоты и многие содержащие азот органические соединения. При дефиците этого элемента бактерии усваивают молекулярный азот. Источником углерода для Cl. pasteurianum может быть широкий набор углеродсодержащих соединений — моносахариды, дисахариды, некоторые полисахариды (декстрин, крахмал) и органические кислоты. Развиваясь на питательных средах, содержащих углеводы, Cl. pasteurianum разлагает их с образованием масляной и уксусной кислот, углекислого газа и водорода. Освобождающаяся при сбраживании углеводов энергия частично идет на усвоение газообразного азота атмосферы.
Cl. pasteurianum обычно считался слабоактивным фиксатором азота. Пределом его активности было связывание от 1 до 3 мг азота на 1 г сброженного сахара. Однако, используя питательные среды, наиболее отвечающие физиологическим потребностям Cl. pasteurianum, удалось повысить его активность до 10—12 мг азота на 1 г сброженного сахара, а в некоторых случаях и более.
Способность фиксировать азот атмосферы свойственна и другим видам рода Clostridium (Cl. butyricum, Cl. acetobutylicum, Cl. pectinovorum, Cl. felsineum и т. д.). Эти бактерии близки в систематическом отношении к Cl. pasteurianum, но отличаются от него ферментативными свойствами. Так, Cl. acetobutylicum вызывает ацетонобутиловое брожение, Cl. pectinovorum может разлагать пектин и т. д. Отдельные виды Clostridium приурочены к определенным почвам (см. главу 14).
Из других анаэробных азотфиксирующих бактерий в почвах нередко обнаруживается Bacillus polymyxa, относящаяся к семейству Bacillaceae. Этот микроорганизм в качестве углеродного питания использует простые сахара и некоторые полисахариды, а также пектин.
Небольшие количества молекулярного азота могут фиксировать некоторые олиготрофные бактерии.
Весьма своеобразная обстановка для микроорганизмов вообще и азотфиксаторов в частности создается на затопленных полях под посевами риса. При разложении массы растительных остатков в почве под слоем воды наряду с другими веществами образуются газообразные соединения — водород, метан, СО2. Установлено, что Н2 и СН4 могут быть источниками энергии и питания для некоторых азотфиксирующих бактерий. Имеются корине — и микобактерии, способные жить автотрофно, окисляя водород и ассимилируя углекислоту. Одновременно они фиксируют молекулярный азот.
Метан используется специфическими азотфиксирующими бактериями из рода Methylomonas (семейство Methylomonadaceae), которые в аэробных условиях могут жить, окисляя только метан или метиловый спирт. Это небольшие монотрихи, не образующие спор.
В воде рисовых полей, а также в Других водоемах можно обнаружить и других азотфиксаторов, представляющих собой анаэробные бактерии. К ним относятся фототрофные пурпурные серобактерии (Thiocapsa, Chromatium, Thiocystis и др.), пурпурные несерные бактерии (Rhodospirillum, Rhodopseudomonas и др.) и зеленые серобактерии (Chlorobium, Pelodiction).
В почвах, загрязненных нефтью, развивается специфический ценоз микроорганизмов, усваивающих молекулярный азот. В основном здесь размножаются ассимилирующие углеводороды коринебактерии (Arthrobacter).
К активным азотфиксаторам относятся аэробные цианобактерии (сине-зеленые водоросли). Можно считать, что все микроорганизмы этой группы, обладающие гетероцистами (клетки с толстой оболочкой), способны фиксировать N2, Они входят в порядки Mastigocladales, Stigonematales и Nostocales класса Hormogoniophyсеае. В почвах СНГ обнаружено около 130 видов и разновидностей гетероцистных форм цианобактерий. Из них к азотусвоителям принадлежат роды Anabaena, Nostoc, Cylindrospermum, Calothris, Tolypothrix и Scytonema, причем наиболее распространены в почвах представители рода Nostoc.
Усвоение молекулярного азота происходит в гетероцистах, то есть в клетках, куда ограничен доступ кислорода. Тем не менее, ферментный аппарат, связывающий N2, обнаружен и в вегетативных клетках гетероцистных форм цианобактерий. Это послужило основанием для поиска негетероцистных азотфиксирующих форм. В последнее время они найдены — это представители родов Plectonema и Phormidium, которые, не обладая гетероцистами, могут связывать N2. В СНГ подобные формы найдены Е. М. Панкратовой.
Цианобактерии распространены во всех почвенно-климатических зонах. Однако они предпочитают нейтральную среду, и. поэтому их численность и видовой состав существенно возрастают в нейтральных почвах южной зоны. Отдельные их виды приурочены к определенным местам обитания. Многие цианобактерии живут в оимбиозе с другими растительными организмами, например,— с грибами, образуя при этом лишайники. В результате адаптации к местным условиям они приобрели способность фиксировать азот при температуре, близкой к 0°С, иногда азотфиксация происходит даже при —5°С, оптимальная температура для этого процесса 15—20°С. В тропиках цианобактерии находятся в симбиозе с саговниками и печеночниками, а также с водным папоротником Azolla и т. д.
В природной обстановке цианобактерии всегда сожительствуют с другими микроорганизмами — бактериями и грибами. В местах разрастания водорослей особенно много олиготрофных бактерий. Массовое развитие цианобактерий отмечается в сильно увлажненных почвах, где они нередко образуют «цветение» почв. Аналогичное явление имеет место в водоемах при обильном размножении водорослей. В неорошаемых окультуренных почвах наиболее благоприятные условия для роста цианобактерий бывают весной и осенью, то есть в периоды увлажнения почвы, а в поливных, кроме того, и после орошения пашни.
Вклад свободноживущих азотфиксаторов в азотный фонд почвы весьма существен. Вопрос о роли свободноживущих и симбиотических азотфиксаторов в азотном балансе почв рассмотрен в главе 16.
agroinf.com
КЛЕТКА — это… Что такое КЛЕТКА?
КЛЕТКА — (cellula, cytus), основная структурно функциональная единица всех живых организмов, элементарная живая система. Может существовать как отд. организм (бактерии, простейшие, нек рые водоросли и грибы) или в составе тканей многоклеточных животных,… … Биологический энциклопедический словарь
КЛЕТКА — КЛЕТКА. Содержание: Исторический очерк…………… 40 Строение К……………….. 42 Форма и величина К…………. 42 Клеточное тело……………. 42 Ядро…………………. 52 Оболочка……………….. 55 Жизнедеятельность К … Большая медицинская энциклопедия
Клетка — Начиная знакомство с животным миром, необходимо сначала в самых общих чертах остановиться на строении и отправлениях клетки. Клетка представляет собой структурную и функциональную единицу, лежащую в основе строения и развития… … Биологическая энциклопедия
клетка — ячейка, элемент, секция; моноспора, квадрат, клеточка, бластомер, клетушка, конура, макромер, базофил, зигота, каморка, ооцит, конурка Словарь русских синонимов. клетка сущ. • каморка • клетушка • конура • конурка Словарь русских синонимов. К … Словарь синонимов
клетка — Клеть, клетка. Наряду с процессом обогащения основного словарного фонда, между прочим и за счет народно областной лексики, наряду с процессом образования новых идущих от него словарных групп протекают и процессы иного характера, состоящие в… … История слов
КЛЕТКА — КЛЕТКА, в биологии основной компонент, из которого состоят все растительные и животные ткани. Клетка является наименьшей живой частицей, способной существовать независимо и обладающей собственной саморегулирующейся химической системой.… … Научно-технический энциклопедический словарь
КЛЕТКА — КЛЕТКА, клетки, жен. 1. Помещение для птиц и мелких животных в форме коробки из металлических или деревянных прутьев. Канарейка в клетке. || Огороженное решеткой место, закрытое со всех сторон, для животных. Тигр беспокойно бегал по клетке. 2.… … Толковый словарь Ушакова
клетка — КЛЕТКА, и, жен. 1. Помещение со стенками из поставленных с промежутками прутьев. К. для птиц, для зверей. 2. Отдельный квадрат разграфлённого пространства. Клетки шахматной доски. Ткань в крупную клетку. • Грудная клетка часть туловища,… … Толковый словарь Ожегова
Клетка-2 — The Cell 2 Режиссёр Тим Якафано Продюсер Алекс Бандер Лоуренс Сильверстайн В главных ролях Тесси Сантьяго Крис Бруно Френк Уэйли … Википедия
Клетка — * клетка * cell основная структурно функциональная единица всех живых организмов, способная к самовоспроизводству путем амитоза (см.), митоза (см.) и мейоза (см.). Все живые К. содержат цитоплазму, окруженную мембраной, а также генетический… … Генетика. Энциклопедический словарь
клетка — 1. КЛЕТКА, и; мн. род. ток, дат. ткам; ж. 1. Помещение для птиц и животных со стенками из металлических или деревянных прутьев. Тигр в клетке. К. для канареек. Грудная к. (часть скелета, ограниченная рёбрами, грудиной и позвонками, заключающая в… … Энциклопедический словарь
dic.academic.ru
Крупные овальные или палочковидные клетки, свободноживущие аэробные азотфиксаторы
Семейство Azotbacteriaceae
Клетки крупные, большей частью от палочковидных с тупыми концами до овальных, но их морфология резко меняется со временем или при изменении условий роста. Клетки часто образуют пары. Подвижные перетрихи или лофотрихи, встречаются неподвижные формы. Грамотрицательные, могут быть грамвариабельными. Спор не образуют, но некоторые виды образуют покоящиеся формы — цисты. Гетеротрофы. Способны фиксировать молекулярный азот в безазотистой среде с органическим источником углерода. Образуют слизистые капсулы. Некоторые штаммы образуют флуоресцирующие пигменты. Аэробы, но способны расти и фиксировать азот и при пониженном давлении кислорода. Каталазоположительные за исключением р. Derxia. Мезофильные обитатели почвы, воды и поверхности листьев.
Род Azotobacter
Крупные овальные клетки 2 мкм и более в поперечнике, длина варьирует вплоть до кокковидной формы. Встречаются поодиночке, парами, неправильными группами, редко образуют цепочки более чем из четырех клеток. Отмечается выраженный полиморфизм. Образуют толстостенные цисты. Могут вырабатывать большое количество капсульной слизи. Перетрихи или неподвижны. Грамотрицательны или грамвариабельны. Некоторые штаммы синтезируют водорастворимый пигмент, дающий в ультрафиолетовых лучах зелёную флуоресценцию. Фиксируют молекулярный азот в присутствии органического источника углерода. Для фиксации азота требуется молибден, который может быть заменен ванадием. Желатин не разжижают. Каталазоположительные. Оптимальная температура роста 20-30°С. Диапазон рН 4,8-8,5 при оптимуме 7,0-7,5.
Встречаются в почве и в воде.
Содержание Г+Ц в ДНК от 63 до 66 мол. %.
Род Beijerinckia
Прямые, слегка изогнутые или грушевидные палочки с явно закругленными концами, иногда клетки могут раздваиваться или ветвиться. На каждом конце клетки имеются характерные крупные, сильно преломляющие свет липоидные (поли-b-оксибутират) тельца. Перетрихи или неподвижны. Некоторые виды образуют цисты и капсулы. Грамотрицательные. В жидких средах не образуют поверхностной пленки, но вся среда превращается в гомогенную, очень вязкую, полупрозрачную массу. При выращивании некоторых видов среда становится опалесцирующей и мутной, но клейкая слизь не образуется. На агаровой среде вырастают огромные колонии с гладкой, складчатой или ребристой поверхностью. Слизь очень плотная, клейкая или эластичная, она затрудняет удаление части колонии петлей.
Аэробы, но могут расти и фиксировать азот при пониженной концентрации кислорода. Каталазоположительные. Температурный предел роста 10-35°С, при оптимуме 20-30°С. Клетки толерантны к кислоте, растут в пределах рН от 3 до 10. В нейтральных и щелочных средах образуется кислота, в сильнокислых средах образуются щелочные вещества, которые повышают рН среды. Используют глюкозу, фруктозу и сахарозу с образованием СО2 и небольших количеств уксусной кислоты, поэтому при росте рН среды снижается. На пептонном агаре или бульоне не растут. При старении колонии принимают рыжевато-розовый-светлокоричневый цвет.
Содержание Г + Ц в ДНК от 554,7 до 60,7 мол. %.
Встречаются в почвах, особенно часто — в тропических.
Палочковидные бактерии азотфиксаторы, симбионты бобовых растений.
Семейство Rhizobiaceae
Палочковидные клетки, спор не образуют, грамотрицательные, подвижные монотрихи или перетрихи. Аэробы. Используют углеводы с образованием кислоты без образования газа. Во время роста на углеводных средах образуют много внеклеточной слизи.
Род I. Rhizobium
Бактерии имеют форму палочек, в неблагоприятных условиях полиморфны, подвижны, перетрихи или монотрихи, грамотрицательны. Спор не образуют, при росте на углеводных средах образуют много слизи. Гетеротрофы, аэробы, желатин не разжижают или разжижают очень медленно в течение 2-х месяцев. Казеин и агар не гидролизуют. Не образуют или почти не образуют сероводорода. Целлюлозу и крахмал не гидролизуют. Моно- и дисахариды и спирты усваивают с образованием кислоты без газа. Источники азота — соли аммония, нитраты и большинство аминокислот. Температурный оптимум 25-30°С, оптимум рН 5,0-8,5.
Для представителей рода характерна способность инфицировать корневые волоски бобовых растений и вызывать образование корневых клубеньков, в которых бактерии растут как внутриклеточные симбионты. Все штаммы обнаруживают специфичность в отношении растений-хозяев. Бактерии обычно находятся в клубеньках в виде полиморфных форм — бактероидов, заключенных по одной или небольшими группами в образованные растением пленчатые мешки. Для бактероидов характерна фиксация молекулярного азота и включение его в соединения, пригодные для использования растением-хозяином. Содержание Г + Ц в ДНК от 59,1 до 65,5 мол. %.
Представители этого рода — обычные обитатели почвы.
Род II. Agrobacterium
Палочки, подвижны, передвигаются при помощи 1-4 жгутиков. Спор не образуют. Грамотрицательны. Гетеротрофы. Аэробы. Углеводы усваивают с образованием кислоты и иногда газа. Целлюлозу, крахмал, агар, хитин и казеин не гидролизуют. Желатин не разжижают или разжижают медленно за несколько недель. На висмут-сульфитном агаре обычно образуют сероводород. Рост на средах с углеводами обычно сопровождается образование обильной внеклеточной полисахаридной слизи. Колонии не пигментированы. Источник азота — соли аммония, нитраты, некоторые аминокислоты. Температурный оптимум 25-30°С, рН 6,0-9,0.
Каталазо- и оксидазоположительные, расщепляют уреазу.
Представители этого рода вызывают разрастание стебля у различных растений, в которых они обитают как внутриклеточные паразиты. Бактерии внедряются в ткань хозяина через существующие повреждения, способность проникать непосредственно в ткани хозяина отсутствует. Предпочтение хозяина выражено нерезко. Вызываемые бактериями заболевания растений известны как галл корончатый (бактериальный), корень волосяной и рак стеблей бактериальный. Опухоли саморазмножаются и способны трансплантироваться. Индукция опухолей коррелирует с присутствием в клетках бактерий Т — плазмиды. Агробактерии обитают в почве, онкогенные штаммы встречаются в тех почвах, куда ранее попадал пораженный растительный материал.
biofile.ru
