
Локализация генов иммунного ответа (Ir-генов)
Выявление генов иммунного ответа определило необходимость локализации этих генов в геноме животных. В опытах с кон- генными, отличающимися только по гаплотипу комплекса Н-2 линиями мышей, обнаружена сцспленность Ir-генов с МНС. Использование значительного количества конгенных линий и большого набора антигенов узкой специфичности показало индивидуальную, зависящую от шплотипа реакцию на тот или иной антиген (рис. 10.7). Один и тот же антиген вызывал иммунный ответ разной силы у отличающихся по гаплотипу конгенных линий и, наоборот, мыши одной и той же конгенной линии формировали ответ разной силы в зависимости от предложенного антигена. Эти данные можно было трактовать как отражение либо множественности аллельных форм 1г-генов, либо значительного числа близ- косцепленных генов. Нерешенность вопроса не отменяла основного заключения о сцепленности Ir-генов с главным комплексом гистосовместимости.
Факт сцепления Ir-генов с МНС требовал установления конкретного места локализации этих генов. Конгенно-рекомбинант- ные линии мышей предоставляли такую возможность. Принципиальные подходы к выяснению этого вопроса лучше всего проиллюстрировать на конкретном примере (рис. 10.8).
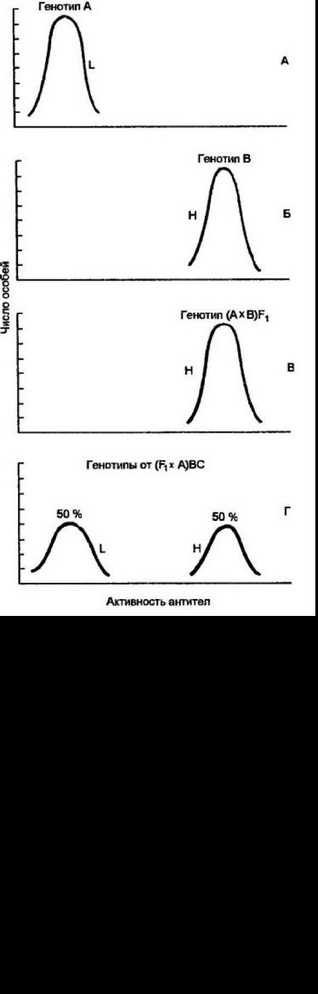
Рис. 10.7, Иммунный ответ мышей с разными Н-2галлотн1тми на тнму сне зависимые антигены трех групп.

Исследования проведены с кошенными, отличающимися только по комплексу Н-2 линиями мышей. Демонстрируется индивидуальная, свойственная только тому или иному га плоти ну мозаичность иммунного ответа на широкий набор антигенов узкой специфичности. Синтетические полипептиды: GLAs — линейный сополимер глутаминовой кислоты, лизина и аланина, составляющего 5% от всех аминокислотных остатков; GLAio —то же с 10% аланина; GLA^o — то же, с 30% аланина; GA — линейный сополимер глутаминовой кислоты, аланина; GATjo — линейный сополимер глутаминовой кислоты, аланина и тирозина, составляющего 10% от всех аминокислотных остатков; GLPros —линейный сополимер глутаминовой кислоты, лизина и про л и на. составляющего 5% от всех аминокислотных остатков; (T,G)-~A— L — разветвленный, многоцеттьсвой сополимер тирозина, глутаминовой кислоты, аланина, лизина; (Ф,С)—A—L — то же, но тирозин заменен на фенилаланин; (H,G)—A—L — то же, но тирозин заменен на гистидин. Лг/юан- тигены мышей; IgA, IgG — иммуноглобулины с аллотипом, отличающимся от аллотипа подопытных мышей; Н—Y — половой антиген самцов; Thy—1.1 — аллоантиген Т-клеток; Еа. 1 — антиген эритроцитов диких мышей; Thyroglob. — тирог- лобулин с аллотипом, отличающимся от аллотипа подопытных мышей. Чужеродные белковые антигены: RE — лекгик; Nase — рибонуклеазл; XI — белок сыворотки крови; 1-ОМ — овомукоид; 1-BGG — бычий у-глобулин; I-OVA — овальбумин; LDHB — лактагидрогенеза
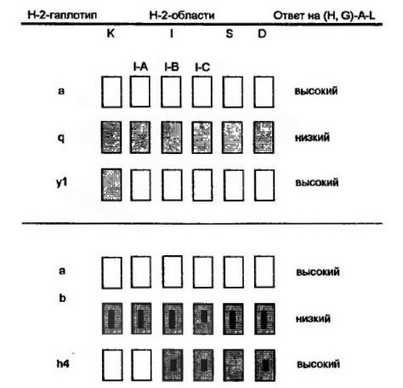
Рис. 10.8. Принцип определения локализации Jr-гена в главном комплексе гнето- совместим ости.
Кошенные и рекомбинантные линии мышей, опозитно реагирующие на синтетический сополимер (H,G)-A-L. Во всех случаях присутствие у рекомбинантных мышей в Н-2-комплексе локуса I-A, контролирующего молекулы II класса, от высокореактивнот гаплотипа определяет сильный иммунный ответ. Локус К не оказывает влияния на силу ответа, так как рекомбинанты (Н-211), включающие этот локус от ареактивной линии, развивают высокий ответ (подробнее см. текст)
аланин-лизин]. В то же время конгенная линия мышей с гаплоти- пом Н-2Ч характеризуется низким ответом. Рекомбинант Н-2у1 между двумя этими линиями, который имеет только один обший Н-2К- локус с Н-2Ч, развивает сильный ответ на анализируемый антиген. Из этого следует, что локус Н-2К не принимает участия в контроле силы иммунного ответа. В противном случае рекомбинанты ответили бы по слабому типу. Во второй серии опытов проведены исследования с мышами, имеющими гаплотип Н-2\ — сильными продуцентами, и мышами с Н-2Ь — слабыми продуцентами антител к данному антигену. Рекомбинант Н-2М между Н-2а и Н-2Ь, гаплотип которого включает локусы Н-2К и I-A (контроль 1а-антигена), а все остальные — от Н-2Ь, характеризуется сильной продукцией антител. Поскольку локус Н-2К не участвует в контроле силы иммунного ответа, следует заключить, что 1г-ген для использованного антигена сцеплен с I-A локусом. Этот локус ответствен за синтез 1а-антигена (Ара)- и Ер-цепи Ера-антигена. Никаких других продуктов, контролируемых данных локусом, до сих
пор нс обнаружено. Из этих фактов было сделано предположение, что фенотипическим продуктом 1г-гснов являются 1а-антигены (молекулы II класса МНС).
Пониятие об иммунитете и иммунной системе. Генетический контроль иммунного ответа.
Иммунитет — невосприимчивость, сопротивляемость организма к инфекциям и инвазиям чужеродных организмов (в том числе — болезнетворных микроорганизмов), а также воздействию чужеродных веществ, обладающих антигенными свойствами.
Иммунная система это совокупность органов, тканей и клеток, работа которых направлена непосредственно на защиту организма от различных заболеваний и на истребление уже попавших в организм чужеродных веществ.
Контроль иммунного ответа генетический – высокоспецифическая форма реакции организма на чужеродные вещества(антигены. При имммунном ответе происходят распознавание чужеродного агента и его элиминация.. Он осуществляется рядом генетических систем. К таким системам относятся гены иммуноглобулинов, Т-клеточного рецептора, молекул ГКГ, цитокинов, системы комплемента и другие.
. Хромосомная теория определения пола. Балансовая теория пола
У растений и животных наиболее распространён хромосомный механизм определения пола. В зависимости от того, какой пол является гетерогаметным, выделяют следующие типы хромосомного определения пола:
самки XX; самцы XY
самки XX; самцы X0
самки гетерогаметны, самцы гомогаметны
самки ZW; самцы ZZ
самки Z0; самцы ZZ
Гинандроморфизм — аномалия развития организма, выражающаяся в том, что в одном организме крупные участки тела имеют генотип и признаки разных полов. Является результатом наличия в мужских и женских клетках организма наборов половых хромосом с разным количеством последних, как например у многих насекомых. Гинандроморфизм происходит как результат неправильного распределения половых хромосом по клеткам в ходе нарушенного созревания яйцеклетки, её оплодотворения или дробления.
-Андрогенез — развитие яйцеклетки с мужским ядром, привнесённым в неё спермием в процессе оплодотворения.
-Гиногенез — частный случай партеногенеза, особая форма полового размножения, при которой после проникновения спермия в яйцеклетку их ядра не сливаются, и в последующем развитии участвует только ядро яйцеклетки, либо не происходит оплодотворения. При этом нет объединения наследственного материала родителей посредством слияния ядер их половых клеток.
В популяциях сельскохозяйственных животных постоянно изменяются частоты генов, что можно наблюдать при анализе смежных поколений. Такие изменения составляют суть генетической эволюции.
— Основные факторы эволюции: мутации, естественный и искусственный отбор, миграции, дрейф генов. Одна из основных причин генетической изменчивости в популяции- мутации. Спонтанные мутации каждого гена происходят с низкой частотой, однако общая частота мутаций всех генов, которые содержат популяции, очень велика. Распространение мутаций в разных популяциях животных может произойти в результате миграций. Генетическая структура популяций формируется и изменяется под действием естественного и искусственного отбора. При естественном отборе преимущество у животных с высокой жизнеспособностью, а при искусственном определяющее значение имеют признаки продуктивности. Власов отвечает, что естеств. Отбор идет на всех этапах онтогенеза популяции. Генетическая структура популяций может изменяться в силу случайных генетико-автоматических процессов или дрейфу генов. Наиболее интенсивно дрейф протекает в малых популяциях.
Генетическое разнообразие по устойчивости к болезням конечностей разных форм неодинаково. Определенную роль в скорости роста, устойчивости к стиранию, количестве роговых трубочек на единице площади копыт, влагопоглощающей способности копытного рога и т. д. играет наследственность. Так, наследуемость количества роговых трубо чек равна 0,38—0,75, диаметра трубочек — 0,63, степени стира ния копыт — 0,79, влагоемкости — 0,82. Поражения копыт связа ны с наследственной предрасположенностью к образованию винтообразного рога и язв подошвы. Серая местная и болгарская породы скота отличаются креп кой конституцией и особенно крепкими ногами и копытами. Заболеваемость коров черно-пестрой эстонской породы в 2 раза выше, чем красной эстонской.
Сущность законов Г. Менделя.
Законы: единообразия гибридов первого поколения, расщепления по фенотипу во втором поколении моногибридного скрещивания, независимого расщепления аллелей и чистоты гамет.
Сущность правила единообразия гибридов первого поколения заключается в том, что при скрещивании гомозиготных родительских форм, различающихся по своим признакам, первое поколение получается единообразным.
Суть правила расщепления заключается в следующем: во втором поколении моногибридного скрещивания наблюдается расщепление по фенотипу в соотношении 3:1, по генотипу в соотношении 1:2:1 ( одна часть особей, гомозиготных по доминантному признаку, две части гетерозиготных и одна часть гомозиготных по рецессивному признаку). Правило частоты гамет: сущность которого в том, что у гетерозиготной особи наследственные задатки не смешиваются друг с другом, а передаются в половые клетки «в чистом» (неизменном) виде.
Независимое расщепление аллелей состоит в том, что во втором поколении каждая пара аллелей и признаков, определяемых ими, ведет себя независимо от других пар аллелей и признаков соответственно. При этом возникают всевозможные сочетания в определенных числовых соотношениях по фенотипу и генотипу. При дигибридном скрещивании при полном доминировании соотношение по фенотипу будет 9:3:3:1 по генотипу 1:2:1:2:4:2:1:2:1
cyberpedia.su
Генетический контроль иммунного ответа: заключительный обзор
Генетический контроль иммунного ответа: заключительный обзор
Исследования механизмов генетического контроля силы иммунного ответа привели к достаточно конкретному заключению: сила иммунного ответа на специфический антиген зависит от работы одного аутосомного доминантного гена; фенотипическим продуктом такого гена являются молекулы II класса МНС ; клеточным типом, экспрессирующим этот ген, являются антигенпрезентирующие клетки .
В тех случаях, когда конформационные особенности антигенраспознающего участка молекул II класса MHC соответствуют структуре антигена (точнее антигенным эпитопам ), образуется иммуногенный комплекс, экспрессируюшийся на поверхности антигенпрезентирующих клеток, что и обеспечивает развитие иммунного ответа.
Напротив, неспособность молекул II класса MHC особей определенного генотипа взаимодействовать с антигенными пептидами будет причиной иммунной ареактивности.
Однако, достигнутые к настоящему времени значительные успехи в выяснении вопросов регуляции иммунного ответа показывают, что генетически иммунный ответ зависит не только от относящихся к MHC генов. К числу факторов, влияющих на иммунный ответ, относятся и гены, не сцепленные с MHC. К примеру, тяжелый комбинированный иммунодефицит обусловлен отсутствием гена рекомбиназы, а недостаточность адгезии лейкоцитов возникает вследствие мутаций гена, кодирующего субъединицу бета1-интегрина , и вызванного ими нарушения экспрессии LFA-1 , CR3 и CR4 . Крупным успехом в изучении генетических основ иммунной патологии стало картирование локусов, регулирующих предрасположенность к инсулинзависимому сахарному диабету (в основном на мышах линии NOD). У этих мышей картировано по меньшей мере 15 генетических локусов, и только один из них сцеплен с MHC в хромосоме 17. Предполагается, что этот ген кодирует молекулы MHC класса II.
Макрофаги играют важнейшую роль в иммунной системе. Поэтому гены, регулирующие их активность, могут также определять результат многих иммунных реакций.
В целом гены, расположенные вне области MHC и регулирующие иммунный ответ, отличаются меньшим полиморфизмом, в связи с чем их вклад в определение чувствительности к заболеваниям не столь велик, как генов MHC.
Ссылки:
medbiol.ru
N генетического контроля силы иммунного ответа
N генетики антигенов
HLA – система – это система генов и кодируемых ими антигенов, участвующая в ряде важнейших биологических процессов:
n распознавание «своего» и «чужого» (обеспечение противоопухолевого, противоинфекционного, трансплантационного иммунитета и др.)
N обеспечение межклеточных взаимодействий
N генетический контроль иммунного ответа
N генетический контроль активности системы комплемента и др.
N обеспечение предрасположенности к различным заболеваниям (онкологическим, аутоиммунным, аллергическим и др.)
Установлено, что HLA-система участвуют в таких физиологических феноменах, как продукция гормонов, миграция клеток, процессы старения. Эти важнейшие функции обеспечиваются, в первую очередь, двумя особенностями HLA-системы:
крайне выраженным полиморфизмом, проявляющимся в том, что она имеет около 700 аллельных вариантов и тем, что HLA-антигены, кодируемые этими аллелями обеспечивают взаимодействие всех клеток организма, включая клетки иммунной системы.
Отдельно взятая молекула антигена гистосовместимости способна связывать ограниченный круг пептидов. Чтобы чужеродные белки не могли избежать иммунологического распознавания, необходимо присутствие на клеточной мембране целого набора антигенов гистосовместимости, включающих в гетерозиготном состоянии по 2 антигена каждого из локусов системы HLA. Полиморфизм классических генов МНС (I и II классов) означает наличие в популяции множества аллелей – вариантов одноименного гена у разных особей.
Конкретные варианты МНС закрепляются в эволюции естественным отбором (в отличие от TCR и Ig) и каждая отдельная особь оказывается приспособленной к регионарным видам и штаммам инфекционных микроорганизмов, на защиту от которых шел отбор МНС у предков.
Нарушение или полное отсутствие какой-либо из функций HLA лежит в основе аутоиммунных, онкологических, иммунодефицитных заболеваний
Гомозиготы по антигенам HLA являются чрезвычайно невыгодным для организма с физиологической точки зрения.
СТРОЕНИЕ СИСТЕМЫ HLA.
HLA локализуется на коротком плече VI хромосомы справа от центромеры, между генами, кодирующими гипоксалазу (GLO) и мочевой пепсиноген 5 (Pg5) и занимает расстояние около 2 сантиморган.
Все гены HLA разделяются на три группы, экспрессирующие охарактеризованные гены, псевдогены и гены с неустановленной функцией.
На сегодняшний день расстояние между условными границами HLA расширено более чем в 2 раза (в 1987 году оценивалось в 2000kb), причем протяженность отдельных его элементов – генных кластеров – колеблется в широких пределах в зависимости от HLA-гаплотипа.
Похожие статьи:
poznayka.org
Генетический контроль иммунного ответа.
Иммунология
Генетический контроль иммунного ответа. просмотров — 68
Исследования механизмов генетического контроля силы иммунного ответа привели к достаточно конкретному заключению: сила иммунного ответа на специфический антиген зависит от работы одного аутосомного доминантного гена; фенотипическим продуктом такого гена являются молекулы II класса МНС ; клеточным типом, экспрессирующим данный ген, являются антигенпрезентирующие клетки . В тех случаях, когда конформационные особенности антигенраспознающего участка молекул II класса MHC соответствуют структуре антигена (точнее антигенным эпитопам ), образуется иммуногенный комплекс, экспрессируюшийся на поверхности антигенпрезентирующих клеток, что и обеспечивает развитие иммунного ответа. Напротив, неспособность молекул II класса MHC особей определенного генотипа взаимодействовать с антигенными пептидами будет причиной иммунной ареактивности. При этом, достигнутые к настоящему времени значительные успехи в выяснении вопросов регуляции иммунного ответа показывают, что генетически иммунный ответ зависит не только от относящихся к MHC генов. К числу факторов, влияющих на иммунный ответ, относятся и гены, не сцепленные с MHC. К примеру, тяжелый комбинированный иммунодефицит обусловлен отсутствием гена рекомбиназы, а недостаточность адгезии лейкоцитов возникает вследствие мутаций гена, кодирующего субъединицу бета1-интегрина , и вызванного ими нарушения экспрессии LFA-1 , CR3 и CR4 . Крупным успехом в изучении генетических основ иммунной патологии стало картирование локусов, регулирующих предрасположенность к инсулинзависимому сахарному диабету (в основном на мышах линии NOD). У этих мышей картировано по меньшей мере 15 генетических локусов, и только один из них сцеплен с MHC в хромосоме 17. Предполагается, что данный ген кодирует молекулы MHC класса II. Макрофаги играют важнейшую роль в иммунной системе. По этой причине гены, регулирующие их активность, могут также определять результат многих иммунных реакций. В целом гены, расположенные вне области MHC и регулирующие иммунный ответ, отличаются меньшим полиморфизмом, в связи с чем их вклад в определение чувствительности к заболеваниям не столь велик, как генов MHC.
100. Предотвращение иммунологического конфликта матери и плода.Самое главное –предотвращение и записывание иммунной системы матери попавших в материнский орагнизм эритроцитов плода. Для этого в теч. 3-х суток(72 часов) после родов, аборта(выкидыша) или операции по выводу внематочной беременностиженщине однократно вводят 300мкг антирезусного иммуноглобулина. Механизм действия препарата основан на его связывании с антигеном резус не попавших к матери плодовых эритроцитов. По этой причине материнская иммунная система их не запоминает и в последующем реагировать на них не будет. Естественно, что такую профилактику нужно повторять после каждых следующих родов или абортов. Но перед введением антирезусного иммуноглобулина нужно определить резус новорожденного. Ведь у резус отрицательной женщины и резус положительного мужчины может появиться резус-отрицательный ребенок. Тогда проводить профилактику нет крайне важности.
medic.oplib.ru
23.Исп.Групп крови и биохимического полиморфизма в практике.
Как и группы крови, так и полиморфные системы используются для следующих целей:
1) Изучение причин и динамики генотипической изменчивости, составляющей основу эволюционной генетики.
2) Уточнения происхождения отдельных животных и филогенеза целых пород.
3) Описание межпородной и внутрипородной дифференциации.
4)Определение моно- и дизиготных двоен.
5) Построение генетических карт хромосом.
6) Подбора гетерозисной сочетаемости.
7) Выявления связи с резистентностью к заболеваниям, продуктивностью и использования биохимических систем в качестве генетических маркеров в селекции животных.
Использование полиморфных систем белков вместе с группами крови повышает точность определения происхождения животных. Так, по группам крови отцовство можно установить в 81% случаев, а дополнительные анализы типов трансферрина повышают точность до 90%.
24. Понятие об иммунитете и иммунной системе организма. Иммунитет – это невосприимчивость организма к инфекционнм агентам и генетически чужеродным веществам антигенной природы. По Р.В. Петрову – Иммунитет – способ защиты организма от живых тел и веществ, несущих на себе признаки генетической чужеродности.
Главная функция – иммунологический надзор за внутренним постоянством (гомеостазом) организма.
Следствием этой функции является распознование, а потом специфическое блокирование, нейтрализация или уничтожение генетически чужеродных веществ.
За сохранение генетически обусловленной биологической индивидуальности отвечает иммунная система организма – совокупность всех лимфоидных органов и скопление лифоидных клеток. Она состоит из центральных и периферических органов. Центральные органы иммунной системы включают тимус (у млекопитающих), сумку Фабриция (у птиц), костный мозг, пейеровы бляшки и миндалины. К периферическим органам относят лимфатические узлы, селезёнку и кровь. Иммунная система и её главнее исполнители – лимфоциты обеспечивают специфическую реакцию организма на чужеродные антигены.
25. Генетический контроль иммунного ответа.
Иммунный ответ, или иммунологическая реактивность, — высокоспецифическая форма реакции организма на чужеродные вещества (антигены). Можно выделить несколько основных вопросов: 1) строение и функция главного комплекса гистосовместимости.
2) генетический контроль антителогенеза.
Во втором вопросе можно выделить несколько аспектов: 1) ген.контроль синтеза иммуноглобулинов и природа их разнообразия.
2) ген.контроль силы иммунного ответа, в основном осуществляемый Ir-генами, локализованными в пределах главного комплекса гистосовместимости.
3) ген.контроль первичных (врождённых) дефектов иммунной системы (иммунодефициты) Предполагают, что мутации любых локусов, кодирующих разные звенья иммунной системы организма, в той или иной степени влияют на иммунный ответ.
Чем больше изучают ген.контроль иммунного ответа, тем очевиднее становится сложность этой проблемы. Однако можно сделать некоторые обобщения:
Ir-гены определяют количество синтезируемых антител против определённых генов.
Ir-гены не сцеплены с локусами, кодирующими синтез иммуноглобулинов.
Ir-гены высокоспецифичны. Организмы с одним и тем же генотипом могут обладать высоким иммунным ответом против одного антигена и низким против другого антигена. Не обнаружены гаплотипы, определяющие общую высокую или низкую иммунологическую реактивность.
Генетически обусловленные различия в высоте иммунного ответа сохраняются в различные возрастные периоды.
Между генами, контролирующими высокий или низкий иммунный ответ против различных антигенов, в основном не существует никакой связи.
Эффекты генов, отвечающих за иммунологическую реактивность, реализуются на уровне популяции лимфоидных клеток.
studfiles.net
