
Доказательства эволюции.
Научные факты, подтверждающие эволюционное развитие жизни на Земле.
Цитологические доказательства – все организмы построены из клеток, которые имеют общее строение и форму.
Сравнительно – анатомические доказательства:
— единство планов строения в пределах типов;
— рудименты– наличие у организмов остатков имевшихся ранее органов Пример: у человека это аппендикс – остаток слепой кишки, копчик – остаток хвоста, волосяной покров тела и др.
— атавизмы – внезапное появление у некоторых особей органов их предков. Пример: у человека – хвост, волосяной покров тела и др.
— наличие гомологических органов;
Гомологичные органы – органы, имеющие общее происхождения, одни и те же эмбриональные зачатки и сходство строения, но выполняющих разные функции. Пример: передняя конечность лошади и плавники кита; листья берёзы и колючки кактуса и т.д.
— аналогичные органы;
Аналогичные органы – органы, выполняющие сходные функции, внешне похожи, но имеют разное происхождение. Пример: крыло бабочки и птицы, роющие конечности крота и медведки, жабры рака и рыбы и т.д.
— конвергентная эволюция.
Конвергенция (схождение признаков) – процесс приобретения сходного строения в результате развития организмов в одинаковой среде.
Пример: при обитание в воде, рыба, ископаемая рептилия – ихтиозавр и млекопитающее – кит приобрели обтекаемую форму тела и плавники.
Биохимические доказательства – единое химическое строение организмов.
Установлена универсальность генетического кода – единого для всего органического мира Земли; универсальность биохимических принципов метаболизма; выявлено родство организмов различных таксонов по ДНК хромосом (ДНК человека имеет одинаковые гены с ДНК макаки – 66%, быка – 28%, крысы – 17 %), по белкам крови (у человека и человекообразных обезьян гемоглобин, белки, определяющие группу крови и резус-фактор идентичны).
Эмбриологические – сходство ранних стадий развития зародыша.
Закон зародышевого сходства (К.М. Бэр) – на ранних этапах эмбрионального развития зародыши разных видов в пределах типа сходны между собой. Пример: в эмбриональном периоде позвоночные последовательно проходят стадии оплодотворенного яйца, дробления, бластулы, гаструлы, трехслойного зародыша, закладки хорды, нервной трубки, пищеварительной трубки и др.
Биогенетический закон (Ф. Мюлер, Э. Геккель) – зародыш в процессе индивидуального развития (онтогенеза) кратко повторяет историю развития вида (филогенез). Пример: у зародышей млекопитающих и рыб закладываются жаберные дуги, на основе которых у рыб развиваются жабры, а у млекопитающих – хрящи гортани и трахеи.
Палеонтологические – добывание и изучение останков растений и животных, населяющих Землю в различные периоды ее истории. Обнаружены переходные формы между типами и классами.
Пример: стегоцефалов считают переходной формой от кистеперовых рыб к амфибиям; археоптерикс – между рептилиями и птицами; лиценопс – между рептилиями и млекопитающими; псилофиты (риниофиты) – между водорослями и наземными растениями.
На основе ископаемых форм построены палеонтологические ряды. Пример: филогенетический ряд лошади, хоботных и др.
Биогеографические – исследование закономерностей географического распространения живых форм на Земле.
По сходству растительного и животного мира в биосфере выделены биогеографические области (голарктическая, индо-малайская, эфиопская, нео-тропическая, австралийская).
Особенности флоры и фауны каждой области тесно связаны с геологической историей и климатическими условиями. Пример: флора и фауна Австралии, водоемов типа озера Байкал.
Микроэволюция.
Микроэволюция – эволюционные процессы, протекающие внутри вида и ведущие к образованию новых видов – начальный этап эволюции.
Вид– совокупность популяций особей, обладающих наследственным сходством морфологических, физиологических и биохимических особенностей, свободно скрещивающихся и дающих плодовитое потомство, приспособленных к сходным условиям жизни и занимающих в природе определённую область распространения – ареал.
Развитие взглядов на понятие « биологический вид» (основные тезисы):
Виды сотворены и неизменны. ( К. Линей).
Виды не существуют реально, любая изменчивость в природе представляет собой видообразование. (Ж. Сент-Илэр).
Все живые постепенно изменяются, виды не существуют реально; понятие видов придуман человеком для своего удобства (Ж. Ламарк).
Критерии вида (признаки, согласно которым организм относится к тому, или иному виду; не абсолютны, используются в совокупности).
Морфологический – сходство внешнего и внутреннего строения; не абсолютнен по многим причинам: имеются виды – двойники, внешне одинаковые, но никогда не скрещивающиеся (Пример: 6 видов малярийных комаров анофелес, ранее считавшихся одним видом). Имеются политипичные виды – с несколькими различающимися фенотипами (Пример: все разнообразные природы голубей является одним видом).
Многие виды имеют ярко выраженный половой диморфизм – отличие самцов от самок, из-за чего их ошибочно относят к разным видам (К. Линей описал самца и самку утки кряквы, как разные виды).
Генетический – видовая специфичностьь хромосомного набора, сходства нуклеотидного состава ДНК.
Вид генетически закрытая система, его генофонд защищен от притока новых генов; механизм защиты репродуктивная изоляция (несовпадения сроков и места размножения; строгий ритуал поведения при спаривании; разница в строении полового аппарата; невозможность проникновения «чужих» сперматозоидов в яйцеклетку; нежизнеспособность или стерильность межвидовых грибов).
Физиологический – сходство процессов жизнедеятельности (данный критерий можно объединить с морфологическим) .
Биохимический – сходство белкового состава базируюется на уникальной нуклеотидной последовательности ДНК.
Критерий не абсолютен, т.к. иногда разные виды имеют очень большую степень сходства. Пример: человек и шимпанзе имеют 98% общих белков, но по другим критериям относятся к разными видами).
Географический – определённый ареал занимаемый видом.
Есть виды космополиты (бактерии и др.), обитающие повсеместно и есть виды с разорванными ареалами – аллопатрические виды. Группировки особей внутри ареалов аллопатрических видов называют популяциями.
Экологический – сходство особей вида в потребностях и образе жизни, но внутри популяции могут быть группы (экологические расы), различающиеся по питанию, времени активности, срокам размножения и т.д.
Цитогенетический – сходство числа и строения хромосом, примерами его не абсолютности являются человек и шимпанзе.
Виды – сложная система, состоящая из мелких группировок особей – подвидов, популяций, экологических рас и других таксонов, приспособленных к разным условиям среды.
Популяция – совокупность особей данного вида, занимающая определённый ареал внутри ареала вида, свободно скрещивающихся между собой ( панмиксия) и частично изолированных от других популяций.
Экологические расы – группа внутри одной популяции, различающиеся по питанию в разных местах, разной пищей, по времени активности, срокам размножения и т.д.
Пример: в озере Севан обитают несколько экологических рас форели, различных по местам обитания (на дне, на поверхности) по срокам размножения (весна или осень).
Особи из разных популяций сохраняют способность периодически скрещиваться друг с другом, что обеспечивает единство генофонда вида и обогащение потомства новыми аллелями.
Особи вида не могут скрещиваются с особями другого вида и давать плодовитое потомство, потому генофонд видов является защищённым и стабильным.
Популяция является элементарной структурной единицей эволюции.
Термин «популяция» ввёл в 1904 г. датский генетик В. Иогансен.
Для каждого популяции характерна определённая генетическая структура, соотношение определеных генов и аллелей. Совокупность генов всех особей популяции, называется
Свойства генофонда:
— Разнообразие (генетическая гетерогенность). Одновременное наличие в генофондах разнообразных генотипов и аллельных вариантов генов, что обеспечивается: мутационным процессом, перекомбинации генов в процессах скрещивания, потоком генов из других популяций.
— Генофонды – неповторимы, уникальны.
— В каждой популяции имеется определённое соотношение частот встречаемости разных аллелей и генотипов (закон Харди-Вайнберга).
— Генетическое единство – общность генофондов популяций для всех составляющих их особей, обеспечивается свободным скрещиванием и механизмами наследования согласно законам Менделя.
Единство генофондов имеет важных следствия:
-Любые возникшие мутации имеют шанс распространится на всю популяцию.
-благодаря единству генофонда, популяции являются элементной единицей эволюции – они реагируют на изменение окружающей среды, как единое целое – направленным изменениям генофонда.
Под изменение генофондов понимают изменение часто встречающихся в них отдельных, аллельных вариантов генов и образованных им генотипов.
Причины, способные изменить генофонд популяции, внешние и внутренние факторы, изменяющие её генетическую структуру, называется элементарным фактором эволюции. Длительное и направленное изменение генофонда получило название элементарного эволюционного процесса.
Пример: «Индустриальный меланизм» – распространение тёмной окраски (определяемой меланином) у нескольких видов бабочек в Англии.
До середины 19 века в популяции этих бабочек преобладали особи со светлой окраской, хорошо маскирующей их на стволах деревьев, покрытых серым лишайником. Число бабочек с тёмной окраской (определяемой мутантным доминантным вариантом гена) было небольшим, поскольку они были заметны на стволах и их быстро склёвывали птицы.
Но с середины 19 века, с началом быстрого развития и широкого применения каменного угля воздух загрязнился. В результате лишайники погибли, обнажая чёрные стволы на которые малозаметны тёмно-коричневые бабочки.
Дальнейшие, более глубокие исследования показали, что различия тёмных и светлых бабочек не сводится только к их окраске, они отличались предпочтением и разным фонам (меланисты предпочитают садится на тёмную фон, а светлые – на светлый).
Генофонд складывается из всего разнообразия генов и аллелей, которые имеются в популяции, размножающейся половым путём.
Состав генофонда изменяется из поколения в поколение. Новые сочетания генов образуют уникальные генотипы, которые в своем физическом выражении, т.е. в форме фенотипов, подвергаются давлению со стороны факторов среды, что ведёт к непрерывному отбору.
Популяция генофонд, которой непрерывно изменяется из поколения в поколение, претерпевает эволюционное изменение.
Частоты аллелей.
Любой физический признак организма, например окраска шерсти у мышей, определяется одним или несколькими генами.
Каждый ген может существовать в нескольких разных формах, которые называются аллелями.Число организмов в данной популяции, несущих определённый аллель, определяли частоту данного аллели.
Пример: у человека частота данного аллеля, определяющего нормальную пигментацию кожи, волос, глаз = 99%. Рецессивный аллель отсутствие пигментации – альбинизм, встречается с частотой в 1%.
Это значит, что из общего числа аллелей, контролирующих синтез этого пигмента, 1% не способен обеспечивать его, а 99% делают это.
Пример:
P – Частота доминантного,
Q – Частота рецессивных аллелей.
В примере с пигментацией кожи у человека p=0,99, а Q=0,01.
P+Q=1
0,99+0,01=1
Пример: частота рецессивного аллели = 25% или 0,25. Тогда поскольку P+Q=1
P+0,25=1
P=0,75
Таким образом, частота доминантного аллели равна 0,75 или 75%.
Похожие статьи:
poznayka.org
Доказательства эволюции | Биология
Макроэволюция
Основные эволюционные преобразования в природе происходят в популяциях. Этот процесс называют микроэволюцией. Преобразования, происходящие на надвидовых уровнях, которые приводят к формированию систематических групп более высокого ранга, чем вид (род, семейство, отряд и т. д.), называют макроэволюцией. Макроэволюция — надвидовая эволюция живых форм. Между макроэволюцией и микроэволюцией нельзя провести принципиальную границу, поскольку макроэволюция совершается посредством микроэволюционных процессов. В макроэволюции действуют те же процессы, что и в микроэволюции — дивергенция, борьба за существование, естественный отбор, — и связанное с ними вымирание.
Эволюционные преобразования, приводящие к возникновению систематических групп более высокого порядка, чем вид, называют микроэволюцией.
Макроэволюция недоступна непосредственному наблюдению, хотя ее результаты окружают нас повсеместно, потому что она происходит на протяжении грандиозного промежутка времени, исчисляемого сотнями тысяч, миллионами лет. Однако наука накопила множество фактов, доказывающих реальность мароэволюционных процессов.
Имеется огромное количество фактов, подтверждающих существование процесса эволюции. Среди них видное место занимают материалы палеонтологии, эмбриологии, биогеографии, экологии, генетики, биохимии, молекулярной биологии, анатомии, физиологии, морфологии и многих других областей биологической науки. Рассмотрим некоторые примеры из них.
Палеонтология как доказательство эволюции
Палеонтология — это наука об ископаемых животных, растениях и микроорганизмах, с помощью которых воссоздается картина поэтапного развития живого мира с доисторических времен.
Обычно ископаемые формы представлены в виде минерализованных скелетов (окаменелостей), хотя возможны находки останков с сохранившимися мягкими тканями. Например, в Сибири в слоях вечной мерзлоты находят останки мамонтов (взрослых и детенышей) с сохранившимися мышечными и другими тканями, кожей с волосяным покровом, внутренними органами, наполненными растительной массой. В сланцах, каменном угле встречаются ископаемые останки многих древних растений и животных.
По найденным окаменелостям удалось восстановить картину появления позвоночных животных и этапы усложнения их организации.
Сопоставление ископаемых останков из земных пластов различных геологических эпох убедительно свидетельствует об изменениях органического мира во времени. В самых глубоких, древних пластах встречаются останки типов беспозвоночных животных, а в более поздних пластах — останки типа хордовых. В поверхностных, более молодых геологических пластах содержатся останки организмов, относящихся к видам, похожим на современные.
 Останки раковин в осадочных породах
Останки раковин в осадочных породах
Палеонтологам удалось на основе большого фактического ископаемого материала найти переходные формы организмов. Например, скелеты зверозубых рептилий, совмещающих в себе признаки пресмыкающихся и млекопитающих, первоптицы — археоптерикса, имеющего признаки пресмыкающихся и птиц. Эти и многие другие находки позволили установить исторический путь развития класса Птицы и класса Млекопитающие, определить их родство между собой и время, когда они стали расходиться.
По ископаемым находкам выявляются особенности происхождения и этапы эволюции человека.
Данные палеонтологии дают большой материал о преемственных связях между различными систематическими группами. В одних случаях удается установить переходные формы, в других — филогенетические ряды, то есть ряды видов, последовательно сменяющих друг друга во времени. Примером филогенетических рядов могут служить находки, подтверждающие происхождение перьев птиц от роговых чешуй рептилий и демонстрирующие эволюцию лошади.
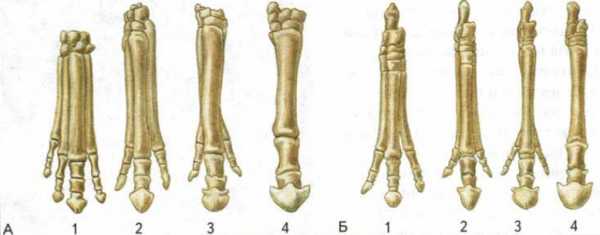 Эволюция конечностей лошадиных: А — передняя конечность; Б — задняя конечность; 1 — эогиппус; 2 — миогиппус; 3 — мерикгиппус; 4 — лошадь (Equus)
Эволюция конечностей лошадиных: А — передняя конечность; Б — задняя конечность; 1 — эогиппус; 2 — миогиппус; 3 — мерикгиппус; 4 — лошадь (Equus)
Русский ученый-зоолог В.О. Ковалевский (1842-1883) по ископаемым находкам с помощью переходных форм воссоздал филогенетический ряд лошади. Метод создания филогенетических рядов, введенный в палеонтологию В.О. Ковалевским, широко используется и в настоящее время.
Эмбриологические доказательства эволюции
Убедительные доказательства родства между организмами представляет эмбриология — наука, изучающая зародышевое развитие организмов.
Сравнение эмбриогенеза (зародышевого развития) различных позвоночных животных, например, рыб, амфибий, птиц и млекопитающих, показывает, что их ранние стадии развития очень сходы между собой. Но на поздних стадиях эмбрионы этих животных уже различаются, и довольно сильно. Эти факты сходства эмбрионов у позвоночных животных были впервые описаны в первой половине XIX века отечественным ученым-эмбриологом К.М. Бэром.
В книге «Об истории развития животных» (1828) он сопоставил зародышевую организацию представителей различных классов позвоночных животных и пришел к важным выводам, согласно которым все зародыши позвоночных воспроизводят общие признаки типа (то есть как бы исходят их общего типа организации), развиваясь в дальнейшем в разных направлениях, «обособляясь друг от друга». При этом, утверждал Бэр, в ходе эмбриогенеза у разных классов позвоночных общие признаки типа формируются раньше, чем видовые признаки вида. Эмпирические обобщения, сделанные Бэром о закономерностях эмбрионального развития, Дарвин рассматривал как одно из существенных доказательств эволюции и назвал их законом зародышевого сходства. Позже это было названо законом Бэра.
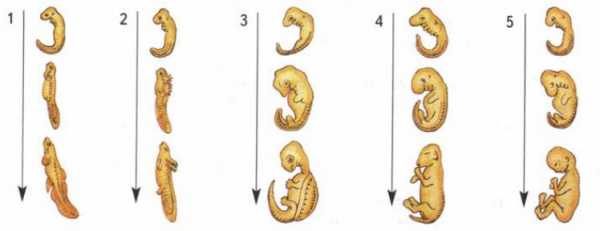 Сходство стадий эмбрионального развития позвоночных: 1 — рыба; 2 — амфибия; 3 — рептилия; 4 — млекопитающее; 5 — человек
Сходство стадий эмбрионального развития позвоночных: 1 — рыба; 2 — амфибия; 3 — рептилия; 4 — млекопитающее; 5 — человек
К.М. Бэр первым в науке исследовал и описал яйцеклетку млекопитающих. Интересно, что рассматривая эту яйцеклетку, он очень удивился, найдя ее необычайно большое сходство с яйцеклеткой птиц.
Зародышевое сходство разных видов является следствием их филогенетического родства и указывает на общность происхождения.
По мере развития (в онтогенезе) сходство между зародышами ослабевает и начинают проявляться признаки тех классов позвоночных, к которым принадлежат данные животные. В ходе эмбрионального развития у эмбрионов происходит расхождение признаков: у них появляются признаки, характеризующие класс, затем отряд, род и, наконец, вид, к которому они принадлежат. Такое явление Дарвин назвал «свидетельством общности происхождения позвоночных животных».
Идеи, высказанные Бэром и Дарвином, получили развитие в трудах немецких ученых-зоологов Ф. Мюллера и Э. Геккеля (1866). Рассматривая взаимоотношения между онтогенезом (индивидуальное развитие организма) и филогенезом (историческое развитие систематических групп), они сформулировали эмпирическое обобщение, которое было названо биогенетическим законом или законом Мюллера-Геккеля.
Ф. Мюллер в 1864 году установил закономерность, а Э. Геккель в 1866 году сформулировал биогенетический закон: «Онтогенез всякого организма есть краткое и сжатое повторение филогенеза данного вида». Филогенез, по Геккелю, осуществляется главным образом путем появления («наращивания») новых стадий в конце онтогенеза.
В эволюционно-морфологическом отношении связи онтогенеза и филогенеза были наиболее полно рассмотрены и развиты позже А.Н. Северцовым в его «Теории филэмбриогенезов» (1939). По концепции Северцова, филогенез представляет собой генетический ряд онтогенезов, который не следует рассматривать как ряд сменяющихся состояний организмов последовательных поколений. Дело в том, что эволюционируют не только фенотипы, представляющие совокупность всех внешних и внутренних признаков организма, но и их онтогенезы. Изменчивость организмов проявляется на всех стадиях индивидуального развития (онтогенеза) и служит материалом для действия естественного отбора. Поэтому Северцов уточняет кратко: «Филогенез есть эволюция онтогенеза». Изменения темпов индивидуального развития, особенно на стадии дифференцировки различных органов у эмбриона могут служить одной из форм эмбриональной изменчивости, которая дает богатый материал для различных эволюционных преобразований.
Биогенетический закон позволяет использовать данные эмбриологии для воссоздания хода эволюции.
Биогенетический закон применим и для растений.
blgy.ru
1. Доказательства эволюции органического мира
Эволюцией называется необратимый процесс развития любой системы, в результате которого возникают новые структуры и новые функции. В биологии термин «эволюция» (от лат. evolutio – раз-витие, развертывание) впервые использовал швейцарский натуралист Шарль Бонне в 1762 г. в одной из эмбриологических работ.
По современным представлениям, биологическая эволюция – это необратимое и, в известной мере, направленное историческое развитие живой природы, сопровождающееся изменением генетического состава популяций, формированием адаптаций, образованием и вымиранием видов, преобразованиями биогеоценозов и биосферы в целом.
Таким образом, само понятие «эволюция» включает, как минимум, два момента: адаптациогенез и формирование таксонов.
Существуют многочисленные доказательства эволюции органического мира Земли, которые одновременно являются и методами изучения эволюции. К классическим доказательствам эволюции относятся палеонтологические, сравнительно-анатомические и сравнительно-эмбриологические.
1. Палеонтологические. Ранее существовавшие организмы оставляют после себя различные формы ископаемых остатков: окаменелости, отпечатки, скелеты, следы деятельности. По этим остаткам можно проследить изменение групп организмов во времени. Реконструированы филогенетические ряды лошадиных, хоботных, некоторых моллюсков. Обнаружено множество переходных форм между современными группами организмов. Однако из-за неполноты палеонтологической летописи не всегда удается реконструировать ход эволюции.
2. Сравнительно-морфологические. Системы органов современных организмов образуют ряд последовательных изменений. Например, на современных организмах можно проследить судьбу отдельных костей мозгового и висцерального черепа. К сравнительно-морфологическим доказательствам близки сравнительно-биохимические. Например, на современных организмах можно проследить изменение структуры гемоглобина. Однако в этих рядах имеются и пробелы, поскольку далеко не все переходные формы дожили до нашего времени.
3. Сравнительно-эмбриологические. В ходе эмбрионального развития у зародышей часто наблюдаются черты сходства с зародышами предковых форм. Например, у всех позвоночных на ранних стадиях развития появляются внутренние жабры (или их зачатки – жаберные карманы).
На основании закона зародышевого сходства был сформулирован биогенетический закон Мюллера–Геккеля, который в краткой формулировке гласит: «Онтогенез (индивидуальное развитие) есть быстрое и краткое повторение филогенеза (исторического развития)». Однако в этих рядах эмбрионального развития сходство между зародышами лишь самое общее, проявляются не все признаки. Например, у зародышей амниот (рептилий, птиц и млекопитающих) не появляются наружные жабры, характерные для личинок анамний (рыб и земноводных), а развитие жаберных щелей останавливается на стадии жаберных карманов. Поэтому биогенетический закон в трактовке Мюллера-Геккеля носит ограниченный характер.
В ходе эволюции наблюдаются эволюционные преобразования процессов онтогенеза, связанные с адаптациями взрослых (половозрелых) организмов. В ходе таких преобразований могут появляться новые органы, но могут и утрачиваться старые органы (полностью или превращаться в рудименты). При этом могут изменяться: начальная масса зачатка органа, место и время закладки органа. Эти преобразования могут происходить на разных стадиях онтогенеза: на самых ранних (закладка хорды, нервной трубки), средних (закладка чешуи у рыб, перьев у птиц, видоизменение побегов растений) и поздних (редукция хвоста у головастиков, формирование четырехкамерного сердца у птиц и млекопитающих, изменение формы листьев). При изменениях органов на поздних стадиях онтогенеза и может действовать филогенетический закон.
В настоящее время для изучения эволюционного развития той, или иной группы организмов используется целый комплекс методов: биогеографические, экологические, генетические, молекулярно-биологические, иммунологические, биохимические, а также методы палеоэкологии, сравнительной физиологии и этологии; широко используются методы компьютерного моделирования.
studfiles.net
Реферат: Доказательства эволюции
Доказательства эволюции
Эмбриологическое доказательство эволюции
Все многоклеточные животные проходят в ходе индивидуального развития стадии бластулы и гаструлы. С особой отчетливостью выступает сходство эмбриональных стадий в пределах отдельных видов и классов. Например у всех наземных позвоночных так же как и у рыб обнаруживается закладка жаберных дуг, хотя эти образования не имеют функционального значения у взросл организмов. Подобное сходство эмбриональных эмбриональных стадий объясняется единством происхождения всех живых организмов.
Морфологические доказательство эволюции
Существование форм сочетаются признаки нескольких систематических единиц указывает на то, что в прежние геологические эпохи жили организмы, являющиеся родоначальниками нескольких систематических групп. На основании исследований Ковалевского все группу животных, присоединили к позвоночным и дали этому типу название хордовых. Связь между разными классами животных так же хорошо иллюстрирует общность их происхождения. Строение передних конечностей некоторых позвоночных несмотря на выполнение этими органами совершенно разных функций, в общем сходны. Некоторые кости в скелете конечностей могут отсутствовать, другие — срастаться, но их гомология совершенно очевидна. Гомологичным называются Органы, которые развиваются из одинаковых эмбриональных зачатков сходным образом. Некоторые органы не функционируют у взрослых животных и являются лишними — это рудименты. Наличие рудиментов так же как и гомологических органов — свидетельство общности происхождения
Палеонтологические признаки
Палеонтологические данные указывают на смену животных и растений во времени. Палеонтология так же указывает на причины эволюционных преобразований. Богатейший палеонтологический материал — одно из наиболее убедительных доказательств эволюционного процесса.
Биогеографические доказательства эволюции
Ярким свидетельством произошедших и происходящих эволюционных изменений является распространение различных животных и растений по всей территории планеты. А.Уоллесу удалось составить биогеографию областей:
1) Палеарктическую
2) Неоарктическую
3) Индо-Малайскую
4) Эфиопскую
5) Неотропическую
6) Австралийскую.
Сравнение животного и растительного мира раз зон дает богатейший материал для доказательства эволюционного процесса. Распределение видов животных и растений по поверхности планеты и их группировка в биогеографические зоны отражает процесс исторического развития Земли и эволюции животного
Островные флора и фауна.
Для понимания эволюционного процесса интерес представляют фауна и флора островов. Состав их Ф и Ф полностью зависит от происхождения этих островов. Острова могут быть материкового происхождения или океанического. Материковые острова характеризуются флорой и фауной, близкой по составу к материковой. Чем древнее остров и чем более значительная водная преграда, тем больше обнаруживается отличий. При рассмотрении океанических островов обнаружено, что их видовой состав очень беден. Отсутствуют наземные млекопитающие и амфибии. Вся фауна океанических островов — результат случайного заселения. Огромное кол-во разнообразных факторов указывает на то, что особенности распределения живых существ на планете тесно связанны с преобразованием земной коры и с эволюционным изменением видов.
www.referatmix.ru
Доказательства эволюции органического мира
Экология Доказательства эволюции органического мира просмотров — 432
Проблемы эволюции видов с низким уровнем разнообразия
По уровню внутривидового разнообразия можно выделить две крайние группы видов: с высоким и низким уровнем внутривидового полиморфизма. Первая группа — ϶ᴛᴏ политипические эврибионтные виды с широким ареалом и сложной внутривидовой структурой, с высоким уровнем внутрипопуляционной и межпопуляционной изменчивости. Вторая группа — ϶ᴛᴏ эндемики с низким уровнем изменчивости. Очевидно, что первая группа видов обладает высоким эволюционным потенциалом, ᴛ.ᴇ. может дать начало множеству новых видов (а в дальнейшем и таксонам более высокого ранга). Вторая группа характеризуется низким эволюционным потенциалом; вероятность того, что она даст начало новым видам (а тем более таксонам более высокого ранга), значительно меньше.
Тема 10: Основные закономерности макроэволюции
- Доказательства эволюции органического мира
- Макроэволюция. Связь макроэволюции с микроэволюцией
- Общие закономерности эволюции
Эволюцией принято называть необратимый процесс развития любой системы, в результате которого возникают новые структуры и новые функции. В биологии термин «эволюция» (от лат. evolutio – раз-витие, развертывание) впервые использовал швейцарский натуралист Шарль Бонне в 1762 ᴦ. в одной из эмбриологических работ.
По современным представлениям, биологическая эволюция — ϶ᴛᴏ необратимое и, в известной мере, направленное историческое развитие живой природы, сопровождающееся изменением генетического состава популяций, формированием адаптаций, образованием и вымиранием видов, преобразованиями биогеоценозов и биосферы в целом.
Τᴀᴋᴎᴍ ᴏϬᴩᴀᴈᴏᴍ, само понятие «эволюция» включает, как минимум, два момента: адаптациогенез и формирование таксонов.
Существуют многочисленные доказательства эволюции органического мира Земли, которые одновременно являются и методами изучения эволюции. К классическим доказательствам эволюции относятся палеонтологические, сравнительно-анатомические и сравнительно-эмбриологические.
1. Палеонтологические. Ранее существовавшие организмы оставляют после себя различные формы ископаемых остатков: окаменелости, отпечатки, скелеты, следы деятельности. По этим остаткам можно проследить изменение групп организмов во времени. Реконструированы филогенетические ряды лошадиных, хоботных, некоторых моллюсков. Обнаружено множество переходных форм между современными группами организмов. При этом из-за неполноты палеонтологической летописи не всегда удается реконструировать ход эволюции.
2. Сравнительно-морфологические. Системы органов современных организмов образуют ряд последовательных изменений. К примеру, на современных организмах можно проследить судьбу отдельных костей мозгового и висцерального черепа. К сравнительно-морфологическим доказательствам близки сравнительно-биохимические. К примеру, на современных организмах можно проследить изменение структуры гемоглобина. При этом в этих рядах имеются и пробелы, поскольку далеко не все переходные формы дожили до нашего времени.
3. Сравнительно-эмбриологические. В ходе эмбрионального развития у зародышей часто наблюдаются черты сходства с зародышами предковых форм. К примеру, у всех позвоночных на ранних стадиях развития появляются внутренние жабры (или их зачатки – жаберные карманы).
На основании закона зародышевого сходства был сформулирован биогенетический закон Мюллера–Геккеля, который в краткой формулировке гласит: «Онтогенез (индивидуальное развитие) есть быстрое и краткое повторение филогенеза (исторического развития)». При этом в этих рядах эмбрионального развития сходство между зародышами лишь самое общее, проявляются не все признаки. К примеру, у зародышей амниот (рептилий, птиц и млекопитающих) не появляются наружные жабры, характерные для личинок анамний (рыб и земноводных), а развитие жаберных щелей останавливается на стадии жаберных карманов. По этой причине биогенетический закон в трактовке Мюллера-Геккеля носит ограниченный характер.
В ходе эволюции наблюдаются эволюционные преобразования процессов онтогенеза, связанные с адаптациями взрослых (половозрелых) организмов. В ходе таких преобразований могут появляться новые органы, но могут и утрачиваться старые органы (полностью или превращаться в рудименты). При этом могут изменяться: начальная масса зачатка органа, место и время закладки органа. Эти преобразования могут происходить на разных стадиях онтогенеза: на самых ранних (закладка хорды, нервной трубки), средних (закладка чешуи у рыб, перьев у птиц, видоизменение побегов растений) и поздних (редукция хвоста у головастиков, формирование четырехкамерного сердца у птиц и млекопитающих, изменение формы листьев). При изменениях органов на поздних стадиях онтогенеза и может действовать филогенетический закон.
Сегодня для изучения эволюционного развития той, или иной группы организмов используется целый комплекс методов: биогеографические, экологические, генетические, молекулярно-биологические, иммунологические, биохимические, а также методы палеоэкологии, сравнительной физиологии и этологии; широко используются методы компьютерного моделирования.
Читайте также
Подумайте! 1.Приведите примеры видов, находящихся на пути биологического прогресса; биологического регресса. 2.Может ли упрощение строения способствовать биологическому прогрессу вида? Поясните на конкретных примерах. Эволюция — это очень длительный исторический… [читать подробенее]
Биологические науки накопили огромный материал, доказывающий единство происхождения и историческое развитие органического мира. Сравнительная анатомия — наука о сравнительном строении живых организмов — показывает общность строения и происхождения живых… [читать подробенее]
Проблемы эволюции видов с низким уровнем разнообразия По уровню внутривидового разнообразия можно выделить две крайние группы видов: с высоким и низким уровнем внутривидового полиморфизма. Первая группа – это политипические эврибионтные виды с широким ареалом и… [читать подробенее]
oplib.ru
Доказательства эволюции — WiKi
Наблюдаемые сегодня изменения в популяциях доказывают не только существование эволюции, но и существование ряда механизмов, необходимых для эволюционного происхождения всех видов от общего предка. Было установлено, что геномы подвержены разнообразным мутациям, среди которых перемещение интронов, дупликация генов, рекомбинации, транспозиции, ретровирусные вставки (горизонтальный перенос генов), замена, удаление и вставка отдельных нуклеотидов, а также хромосомные перестройки. Известны такие хромосомные перестройки как дупликация генома (полиплоидия), неравный кроссинговер, хромосомная инверсия, транслокация, деление, слияние, дупликация и удаление хромосом[8][9]. Также наблюдается изменение строения организмов[10][11] и функциональные изменения[10][12] — различные адаптации[13], появление способности усваивать новый вид пищи (в том числе — нейлон и пентахлорфенол, производство которых началось в 1930-х годах)[14] и т. д. Кроме того, были обнаружены всевозможные промежуточные этапы возникновения новых видов, что свидетельствует о плавном характере видообразования.
Быстрая эволюция ящериц в новых условиях
Согласно опубликованному в 2008 году исследованию, у популяции стенных ящериц Podarcis sicula (англ.)русск. всего лишь за 36 лет (крайне короткий для эволюции период) изменились размер и форма головы, увеличилась сила укуса и развились новые структуры в пищеварительном тракте. Эти физические перемены произошли одновременно с серьёзными изменениями в популяционной плотности и социальной структуре.
В 1971 году пять пар взрослых особей Podarcis sicula с острова Под Кописте были перевезены на соседний остров Под Маркару (эти хорватские острова находятся на юге Адриатического моря, недалеко от Ластово). Условия на новом месте были близки к привычной среде обитания, наземные хищники отсутствовали, и за три десятилетия ящерицы естественным образом распространились по острову (вытеснив при этом местную популяцию Podarcis melisellensis). Когда исследователи вернулись на Под Маркару, они обнаружили, что ящерицы на этом острове существенно отличаются от ящериц на Под Кописте. Сравнение митохондриальной ДНК показало, что популяции генетически неразличимы, но в то же время в популяции с Под Маркару средний размер особей увеличен, задние конечности короче, максимальная скорость бега меньше, а также отличается реакция на нападение хищников. Эти изменения строения и поведения объясняются меньшей активностью хищников на Под Маркару и наличием более густой растительности, способствующей маскировке. Кроме того, в новой популяции существенно изменились размер и форма головы, она стала более массивной, увеличилась сила укуса. Эти физические перемены соответствуют изменению рациона: P. sicula с Под Кописте преимущественно насекомоядные, а диета ящериц на Под Маркару в зависимости от сезона почти на две трети состоит из растительной пищи. Растения на новом месте обитания жёсткие и волокнистые, а мощные челюсти позволяют качественнее откусывать и размельчать растительную еду.
Наиболее неожиданным[15] различием оказалось наличие у новой популяции илеоцекального (подвздошно-слепокишечного) клапана, предназначенного для замедления прохода пищи за счёт создания в кишке камер брожения, в которых микробы могут разбивать трудно перевариваемые кусочки растительности. Эти клапаны никогда ранее не были обнаружены у этой разновидности ящериц, включая исходную популяцию с острова Под Кописте. Специалисты называют их «эволюционным новшеством, новым свойством, отсутствовавшим у предковой популяции и развившемся в ходе эволюции у этих ящериц»[16]. Кроме того, в кишечнике новой популяции присутствуют нематоды, отсутствующие у исходной популяции.
Изменения в диете также вызвали изменения в плотности популяции и социальной структуре. Из-за того, что растения — это намного больший, предсказуемый и надёжный источник пропитания, чем насекомые, количество ящериц увеличилось. Кроме того, пропала необходимость гоняться за добычей, что привело к тому, что ящерицы перестали агрессивно защищать свои территории[17].
Развитие устойчивости к антибиотикам и пестицидам
Развитие и распространение устойчивости к антибиотикам у бактерий и устойчивости к пестицидам у растений и насекомых является доказательством эволюции видов. Появление устойчивых к ванкомицину форм золотистого стафилококка и та опасность, которую они представляют для пациентов больниц, — это прямой результат эволюции путём естественного отбора. Ещё один пример — развитие штаммов шигеллы, устойчивых к антибиотикам из группы сульфаниламидов[18]. Появление малярийных комаров, устойчивых к ДДТ, и развитие у австралийских популяций кроликов устойчивости к миксоматозу демонстрируют действие эволюции в условиях давления отбора при быстрой смене поколений.
За последние 50 лет зарегистрировано более 2500 случаев адаптации насекомых-вредителей к различным пестицидам. Поэтому для борьбы с вредителями вместо обычных ядов стараются использовать биологические пестициды — энтомопатогенные вирусы. В частности, для борьбы с яблонной плодожоркой Cydia pomonella (личинки которой — это те самые «червяки» в червивых яблоках) активно используется Cydia pomonella грануровирус (англ.)русск. (CpGV). Но в 2003 году были зафиксированы первые случаи снижения эффективности вируса, и к 2005 году появились популяции яблонной плодожорки, у которых устойчивость к вирусу выросла в 100 000 раз по сравнению с исходной. Установлено, что причиной устойчивости является мутация в половой Z-хромосоме[19].
Примеры видообразования
Согласно синтетической теории эволюции, образование новых видов происходит в результате разделения особей одного вида на группы, не скрещивающиеся между собой. Такое разделение может быть вызвано различными причинами, такими как наличие непреодолимых географических барьеров (аллопатрическое видообразование) или разделение по экологическим нишам внутри одного ареала (симпатрическое видообразование). После возникновения между популяциями различных типов изоляции популяции начинают эволюционировать независимо, в результате чего между популяциями постепенно накапливаются генетические различия, и через некоторое время достигается генетическая несовместимость — скрещивание оказывается невозможным.
Кроме того, новые виды могут возникать практически мгновенно (в геологическом масштабе времени) путём полиплоидии (кратного увеличения количества хромосом в клетке) и гибридизации (скрещивания различных видов).
Наблюдения за современными видами показывают, что видообразование непрерывно происходит в существующих популяциях.
В качестве наблюдаемых человеком исторических примеров видообразования (в результате искусственного отбора) можно назвать выведение из одомашненной популяции дикого тутового шелкопряда тутового шелкопряда домашнего (Bombyx mori). Скрещивания между этими формами практически не происходят и практически не дают способного размножаться потомства, следовательно, имеет место репродуктивная изоляция.
Промежуточные этапы видообразования
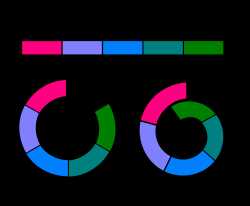 Диаграмма, демонстрирующая варианты скрещивания соседних популяций одного вида. В третьем случае ареалы отдельных популяций образуют кольцо, в результате две соседние популяции имеют генетическое родство, однако друг с другом не скрещиваются и ведут себя как отдельные виды
Диаграмма, демонстрирующая варианты скрещивания соседних популяций одного вида. В третьем случае ареалы отдельных популяций образуют кольцо, в результате две соседние популяции имеют генетическое родство, однако друг с другом не скрещиваются и ведут себя как отдельные виды
В определении понятия биологического вида существенное значение имеет практически полная репродуктивная изоляция в природных условиях[20]. Если видообразование происходит постепенно, в результате небольших последовательных изменений, то и сегодня мы должны наблюдать все возможные стадии видообразования, а значит, и все возможные стадии репродуктивной изоляции: свободное скрещивание между популяциями, затруднённое скрещивание, снижение плодовитости потомства, бесплодное потомство, и наконец — полную генетическую изоляцию. Если ничего подобного среди современных видов обнаружить не удаётся, то тем самым опровергаются основы эволюционного учения, то есть это пример фальсифицируемости эволюционного учения[21].
Существует множество примеров того, как разные виды могут скрещиваться в исключительных условиях. Например, у чёрной вороны и серой вороны разные ареалы, но на пересечении этих ареалов виды могут образовывать гибриды. Ещё один пример — различные виды рыбы рода чукучанов из реки Платт, которые живут рядом, но редко скрещиваются[22].
Неполное видообразование можно наблюдать у различных кольцевых видов[23]: представители вида обитают вокруг гор, водоёмов и т. п., при этом соседние популяции скрещиваются между собой и незначительно различаются, а крайние формы различаются значительно и не скрещиваются (см. диаграмму). Пример кольцевого вида — безлёгочная саламандра Ensatina eschscholtzii, обитающая на склонах гор близ тихоокеанского побережья Северной Америки. В зависимости от района обитания вокруг гор саламандры образуют различные формы, постепенно изменяющие свои морфологические и экологические характеристики. Крайние формы Ensatina eschscholtzii eschscholtzii и Ensatina eschscholtzii klauberi, первая из которых имеет красноватую окраску, а вторая чёрно-белую, сосуществуют в узкой зоне гор на юге Калифорнии, однако между собой не скрещиваются[23][24]. Другие примеры кольцевых видов — зелёная пеночка (Phylloscopus trochiloides), оленья мышь (Peromyscus maniculatus), наземная улитка Partula olympia, дрозофила Drosophila paulistorum.
У многих гибридов снижена фертильность. У гибридов землероек из рода бурозубок размножение затруднено вследствие разницы хромосомных наборов. Тот же эффект наблюдался в лаборатории при скрещивании Drosophila pseudoobscura (англ.)русск. из Юты и Калифорнии. Фертильность также снижена у гибридов лягушек из семейства Bombinatoridae жерлянки краснобрюхой (Bombina bombina) и жерлянки желтобрюхой (Bombina variegata)[25].
Многие другие виды могут давать бесплодное гибридное потомство. В их числе — различные земноводные (например — некоторые виды лягушек из рода Rana) и млекопитающие (например, мул — гибрид от скрещивания лошади и осла — как правило, бесплоден). Гибриды гребенчатого тритона и мраморного тритона бесплодны вследствие наличия непарных хромосом[26].
При скрещивании ряда других видов достигается оплодотворение, но позднее зародыш погибает. В качестве примера можно привести леопардовую лягушку (Rana pipiens) и лесную лягушку (Rana sylvatica) из рода настоящих лягушек[27]. Аналогичная ситуация наблюдается у дрозофил и у некоторых растений, таких как хлопчатник обыкновенный (Gossypium hirsutum) и хлопчатник барбадосский (Gossypium barbadense)[28].
- Белый медведь
Конкретным примером крупномасштабных эволюционных изменений при неполном видообразовании является белый медведь (Ursus maritimus), который, несмотря на родство с бурым медведем (Ursus arctos), очевидное в силу того факта, что эти виды могут скрещиваться и давать плодовитое потомство[29], приобрёл значительные физиологические различия с бурым медведем. Эти различия позволяют белому медведю комфортно жить в условиях, в которых бурый медведь не выжил бы. В частности, белый медведь способен проплыть десятки километров в ледяной воде, его окраска сливается со снегом, он не замерзает в Арктике. Всё это возможно благодаря конкретным изменениям: белая окраска способствует маскировке хищника при охоте на тюленей; полые волоски увеличивают плавучесть и сохраняют тепло; слой подкожного жира, толщина которого к зиме доходит до 10 сантиметров, обеспечивает дополнительную теплоизоляцию; удлинённая, по сравнению с другими медведями, шея позволяет легче держать голову над водой во время плавания; увеличенные лапы с перепонками действуют как весла; небольшие бугорки и полости-присоски на подошвах уменьшают опасность поскользнуться на льду, а плотная шерсть на подошвах защищает лапы от сильного холода и обеспечивает трение; уши меньше, чем у других медведей, и уменьшают потери тепла; веки действуют как солнечные очки; зубы острее, чем у других медведей, и больше подходят для полностью мясного рациона; увеличенный объём желудка позволяет голодному хищнику съесть сразу целого тюленя, кроме того, белый медведь способен обходиться без пищи до девяти месяцев за счет переработки мочевины[30][31].
Наблюдаемое видообразование
Судя по палеонтологической летописи и по измерениям скорости мутаций, полная несовместимость геномов, делающая невозможным скрещивание, достигается в природе в среднем за 3 млн лет[32]. А значит, наблюдение образования нового вида в естественных условиях в принципе возможно, но это редкое событие. В то же время, в лабораторных условиях скорость эволюционных изменений может быть увеличена, поэтому есть основания надеяться увидеть видообразование у лабораторных животных[33][34].
Известны многие случаи видообразования посредством гибридизации и полиплоидизации у таких растений как конопля, крапива, первоцвет, редька, капуста, а также у различных видов папоротников. В ряде случаев видообразование у растений происходило без гибридизации и полиплоидизации (кукуруза[35], стефаномерия (англ.)русск. Stephanomeria malheurensis из семейства астровых[36]).
Дрозофилы, также известные как плодовые мухи, входят в число наиболее изученных организмов. С 1970-х годов зафиксированы многие случаи видообразования у дрозофил. Видообразование происходило, в частности, за счёт пространственного разделения, разделения по экологическим нишам в одном ареале, изменения поведения при спаривании, дизруптивного отбора, а также за счет сочетания эффекта основателя с эффектом бутылочного горлышка (в ходе экспериментов founder-flush).
Видообразование наблюдалось в лабораторных популяциях комнатных мух, мух Eurosta solidaginis, яблонных мух-пестрокрылок, мучных жуков, комаров и других насекомых.
Известны случаи, когда в результате давления отбора (в присутствии хищников) одноклеточные зелёные водоросли из рода хлорелла образовывали многоклеточные колониальные организмы, а у бактерий в аналогичных условиях менялось строение и увеличивались размеры (c 1,5 до 20 микрометров за 8—10 недель). Являются ли эти случаи примерами видообразования, зависит от того, какое используется определение вида (при бесполом размножении нельзя использовать критерий репродуктивной изоляции)[37][38].
Видообразование также наблюдалось и у млекопитающих. Шесть случаев видообразования у домовых мышей на острове Мадейра за последние 500 лет были следствием исключительно географической изоляции, генетического дрейфа и слияния хромосом. Слияние двух хромосом — это наиболее заметное различие геномов человека и шимпанзе, а у некоторых популяций мышей на Мадейре за 500 лет было девять подобных слияний[2][39].
- Яблонная муха
Яблонная пестрокрылка Rhagoletis pomonella является примером наблюдаемого симпатрического видообразования (то есть видообразования в результате разделения по экологическим нишам)[40][41]. Первоначально вид обитал в восточной части США. До появления европейцев личинки этих мух развивались только в плодах боярышника. Однако с завозом в Америку яблонь (первое упоминание яблонь в Америке — 1647 год), открылась новая экологическая ниша. В 1864 году личинки Rhagoletis pomonella были обнаружены в яблоках, тем самым зафиксирована яблонная раса этого вида[42]. За полтора века наблюдений они очень сильно разошлись. Теперь мухи почти не скрещиваются друг с другом (уровень гибридизации не превышает 4—6 %). Яблоневая муха спаривается почти исключительно на яблонях, а боярышниковая — на боярышнике, что, учитывая разное время созревания плодов, приводит к репродуктивной изоляции. В скором времени возможно полное превращение этих подвидов в самостоятельные виды.
У пестрокрылок известно ещё несколько видов-двойников, которые живут на разных видах растений, — предположительно, видообразование у них протекало именно по описанной схеме[43][44][45].
Экспериментальная эволюция и искусственный отбор
Опыты Г. Шапошникова по искусственной эволюции
ru-wiki.org
Доказательства эволюции
Ч. Дарвин доказал историческое развитие живой природы, но в последующие годы сбор прямых и косвенных доказательств эволюции продолжался.
Филогенетические ряды
Выдающиеся достижения в накоплении прямых доказательств эволюции принадлежат отечественным ученым, прежде всего В. О. Ковалевскому. Работы В. О. Ковалевского были первыми палеонтологическими исследованиями, которыми удалось показать, что одни виды происходят от других.
Исследуя историю развития лошадей, В. О. Ковалевский показал, что современные однопалые животные происходят от мелких пятипалых всеядных предков, живших 60—70 млн. лет назад в лесах. Изменение климата Земли, повлекшее за собой сокращение площадей лесов и увеличение размеров степей, привело к тому, что предки современных лошадей начали осваивать новую среду обитания — степи. Необходимость защиты от хищников и передвижений на большие расстояния в поисках хороших пастбищ привела к преобразованию конечностей — уменьшению числа фаланг вплоть до одной (рис. 14). Параллельно изменению конечностей происходило преобразование всего организма: увеличение размеров тела, изменение формы черепа и усложнение строения зубов, возникновение свойственного травоядным млекопитающим пищеварительного тракта и многое другое.
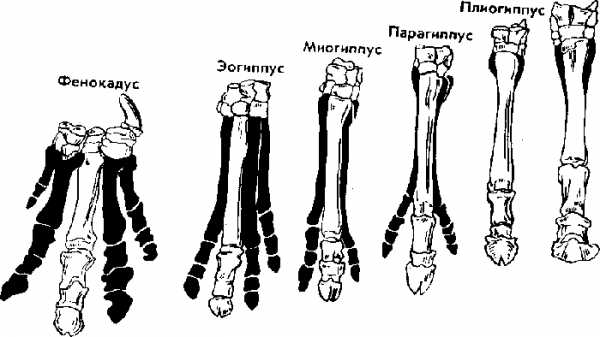
Рис. 14. Преобразование конечностей предков лошадей из пятипалой в трехпалую и затем в однопалую. Последняя конечность — современная лошадь.
В. О. Ковалевский обнаружил последовательные ряды ископаемых форм лошадиных, эволюция которых совершалась в указанных направлениях. Такие ряды видов, последовательно сменяющих друг друга, называются филогенетическими и свидетельствуют о существовании эволюционного процесса.
Переходные формы
Установление факта постепенного эволюционного развития в линиях невысокого систематического ранга (ряды ископаемых лошадей, слонов, моллюсков) показало существование преемственности между современными и ископаемыми видами. Не в силах опровергнуть эти факты, противники эволюционной теории утверждали, что систематические группы более высокого ранга не могли произойти друг от друга, а явились результатом отдельного акта творения. Поэтому особый интерес представляют ископаемые формы, сочетающие признаки древних и более молодых групп высокого систематического ранга. Такие формы называются переходными. Примером их могут служить кистеперые рыбы, связывающие рыб с вышедшими на сушу четвероногими земноводными; семенные папоротники — переходная группа между папоротиикообразными и голосеменными и др.
Существование переходных форм между разными типами, классами, отрядами показывает, что постепенный характер исторического развития свойствен не только низшим, но и высшим систематическим категориям.
Гомологнчные и аналогичные органы, рудименты и атавизмы
Сравнительно-анатомические исследования показали, что строение передних конечностей некоторых позвоночных, например ласты кита, лапы крота, крокодила, крылья птицы, летучей мыши, руки человека, несмотря на выполнение совершенно разных функций, в принципиальных чертах строения сходны. Некоторые кости в скелете конечностей могут отсутствовать, другие срастаться, относительные размеры костей могут меняться, но их гомология, т. е. сходство, основанное на общности происхождения, совершенно очевидна. Гомологичными называются такие органы, которые развиваются из одинаковых эмбриональных зачатков сходным образом (рис. 15).
Наличие у организмов разных групп (классов, семейств и т. д.) гомологичных органов дает возможность установить степень родства между ними, проследить их эволюцию. Видоизменение органов, имеющих общее происхождение, объясняется дивергенцией по признаку строения данного органа в связи с приспособлением к среде обитания.
Не всякое сходство органов свидетельствует в пользу их родства. Крыло бабочки и крыло птицы выполняют сходную функцию, но их строение совершенно различно. Сходство вызвано образом жизни, приспособлением к полету, возникшим независимо друг от друга у бабочек и птиц, а не родственным происхождением этих форм. Органы, имеющие внешнее сходство, вызванное сходными приспособлениями к сходным условиям жизни, но различное строение, называются аналогичными. Аналогичные органы возник-
Рис. 15. Сходство строения передних конечностей обезьяны, летучей мыши, тюленя и лошади свидетельствует об их происхождении от единой предковой формы или в результате конвергенции — схождения признаков и не свидетельствуют о родстве между организмами.
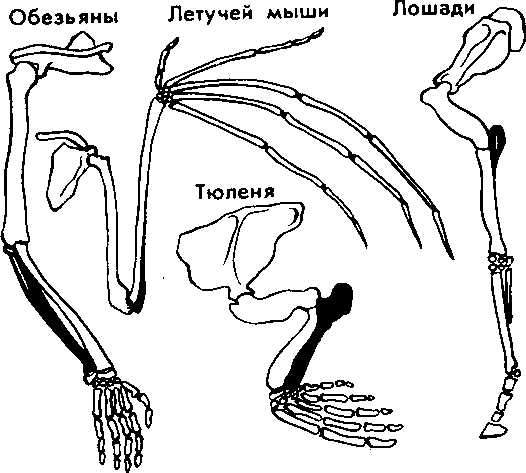
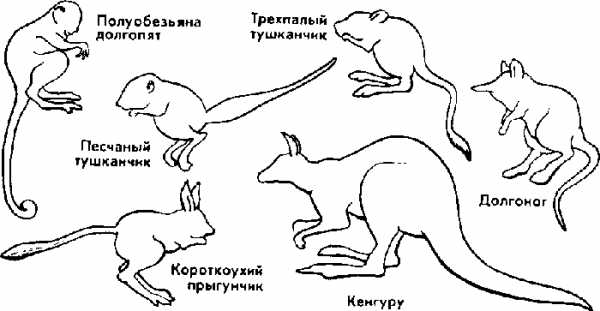
Рис. 16. Конвергенция по форме тела у прыгающих млекопитающих.
Некоторые органы или их части не функционируют у взрослых животных и являются для них лишними — это рудиментарные органы, или рудименты. Наличие рудиментов, так же как и гомо-логичных органов, свидетельствует об общности происхождения живых форм. Задние конечности у кита, скрытые внутри тела,— рудимент, доказывающий наземное происхождение его предков. У человека тоже известны рудиментарные органы: мышцы, двигающие ушную раковину, рудимент третьего века и т. п.
У некоторых организмов рудиментарные органы могут развиться до органов нормальных размеров. Такой возврат к строению органа предковых форм называется атавизмом. Среди тысяч однопалых лошадей изредка попадаются особи, у которых развиты маленькие копытца II и IV пальцев. Известны случаи появления атавистических признаков и у человека: рождение детей с первичным волосяным покровом, с длинным хвостиком и т. д. Возникновение атавизмов указывает на возможное строение того или иного органа у предковых форм.
Сходство зародышевого развития позвоночных
Факт единства происхождения живых организмов был установлен на основе эмбриологических исследований. Все многоклеточные животные развиваются из одной оплодотворенной яйцеклетки. В процессе индивидуального развития они проходят стадии дробления, образования двух- и трехслойного зародышей, формирования органов из зародышевых листков. Сходство зародышевого развития животных свидетельствует о единстве их происхождения.
С особой отчетливостью сходство эмбриональных стадий выступает в пределах отдельных типов и классов. Так, на ранних стадиях развития у зародышей позвоночных (рыбы, ящерицы, кролика, человека) наблюдается поразительное сходство: все они имеют головной, туловищный и хвостовой отделы, зачатки конечностей, по бокам тела — зачатки жабр (рис. 17).
По мере развития зародышей черты различия выступают все более явственно. Причем вначале проявляются признаки класса, к которому относятся зародыши, затем признаки отряда и на еще более поздних стадиях — признаки рода и вида. Эта закономерность в развитии зародышей указывает на их родство, происхождение от одного ствола, который в ходе эволюции распался на множество ветвей.
Биогенетический закон
Основываясь на приведенных выше, а также множестве других фактов, немецкие ученые Ф. Мюллер и Э. Геккель во второй половине XIX в. установили закон соотношения онтогенеза, который получил название биогенетического закона. Согласно этому закону каждая особь в индивидуальном развитии (онтогенезе) повторяет историю развития своего вида (филогенез), или, короче, онтогенез есть краткое повторение филогенеза.
Однако за короткий период индивидуального развития особь не может повторить все этапы эволюции, которая совершалась тысячи или миллионы лет. Поэтому повторение стадий исторического развития вида в зародышевом развитии происходит в сжатой форме, с выпадением ряда этапов.
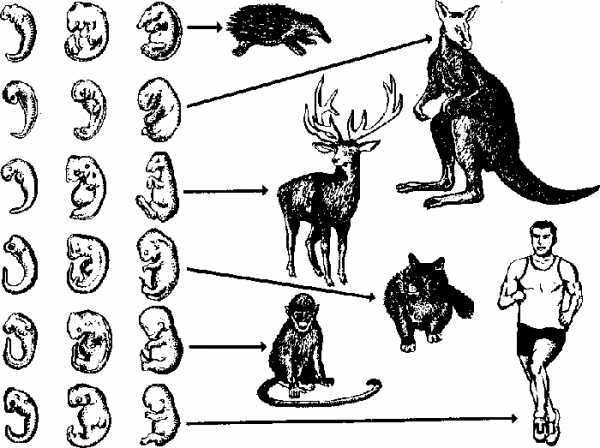
Рис. 17. Сходство начальных стадий онтогенеза позвоночных свидетельствует о родстве и указывает на пройденные в процессе филогенеза этапы
Кроме того, эмбрионы имеют сходство не со взрослыми формами предков, а с их зародышами. Так, в онтогенезе млекопитающих и рыб имеется этап, на котором у зародышей образуются жаберные дуги. У зародыша рыбы на основании этих дуг образуется орган дыхания — жаберный аппарат. В онтогенезе млекопитающих повторяется не строение жаберного аппарата взрослых рыб, а строение закладок жаберного аппарата зародыша, на основе которых у млекопитающих развиваются совершенно иные органы (хрящи гортани и трахеи). В разработке теории онтогенеза выдающуюся роль сыграли исследования академика А. Н. Северцова. Он доказал, что изменения исторического развития обусловлены изменениями хода зародышевого развития. Наследственные изменения затрагивают все стадии жизненного цикла, в том числе и зародышевый период. Мутации, возникающие в ходе развития зародыша, как правило, нарушают взаимодействие в организме и ведут к его гибели. Однако мелкие мутации могут оказаться полезными и тогда сохранятся естественным отбором. Они передадутся потомству, включатся в историческое развитие, влияя на его ход.
Биогеографические доказательства эволюции
Сравнение фаун и флор разных континентов. Яркое свидетельство эволюционного процесса — распространение животных и растений по поверхности нашей планеты. Во все времена путешественников и натуралистов поражало своеобразие растительного и животного мира Австралии, Южной Америки, океанических островов и вызывало интерес сходство фауны некоторых материков, например Северной Америки и Евразии. А. Уоллес привел все сведения в систему и выделил шесть зоогеографических областей:
1) Палеоарктическую, охватывающую Европу, Северную Африку, Северную и Среднюю Азию, Японию;
2) Неоарктическую, включающую Северную Америку;
3) Эфиопскую, включающую Африку к югу от пустыни Сахара;
4) Индомалайскую, охватывающую Южную Азию и Малайский архипелаг;
5) Неотропическую, занимающую Южную и Центральную Америку;
6) Австралийскую, включающую Австралию, Новую Гвинею, Новую Зеландию, Тасманию, Соломоновы острова и Новую Каледонию.
Степень сходства и различия между разными зоогео-графическимн областями неодинакова. Фауна и флора Палеоарк-тической и Неоарктической областей имеют много общего, хотя и изолированы Беринговым проливом- Неоарктическая и Неотропическая области, наоборот, существенно отличаются, хотя и соединены сухопутной связью (Панамским перешейком). В чем же причины этого сходства и различия? Очевидно, они связаны с историей формирования материков, временем их изоляции. Так, глубокое различие в фауне Неотропической и Неоарктической областей определяется тем, что сухопутная связь между ними установилась совсем недавно. Об этом свидетельствуют геологические данные. После возникновения Панамского моста лишь немногим южноамериканским видам удалось проникнуть на север (например, дикобраз, броненосец, опоссум). Североамериканские виды преуспели в освоении южноамериканской области несколько больше. Олени, лисы, выдры, медведи проникли в Южную Америку, но не оказали существенного влияния на ее уникальный видовой состав. Только здесь живут представители отряда неполнозубых (муравьеды и ленивцы), сохранились птицы гоацины, которые могут лазать по деревьям благодаря когтям на пальце крыла (как археоптерикс), и другие животные. Своеобразие фауны Южной Америки было одним из фактов, натолкнувших Ч. Дарвина на мысль об эволюции. Сходство фауны Неоарктической и Палеоарктической областей обусловлено тем, что в прошлом между ними существовал сухопутный мост — Берингов перешеек.
Наиболее отличен от других континентов животный мир Австралии. Известно, что Австралия обособилась от Южной Азии свыше 100 млн. лет назад, еще до возникновения высших млекопитающих. Лишь в Ледниковый период сюда через острова Зонд-ского архипелага перебрались немногие плацентарные — мыши и собаки (потомок последних—австралийская собака динго). В остальном фауна Австралии очень своеобразна.
Таким образом, чем теснее связь континентов, тем более родственные формы там обитают, чем древнее изоляция частей света друг от друга, тем больше различия между их населением.
Фауна островов
Видовой состав фауны и флоры островов целиком определяется историей их происхождения. Острова могут быть материковыми, представлять собой результат обособления части материка и океаническими (вулканические и коралловые острова). Растительный и животный мир первых близок по составу к материковому. На Британских островах и на Сахалине большинство видов сходно с близлежащими районами континента. Это объясняется тем, что острова лишь несколько тысяч лет назад отделились от суши. Однако чем древнее остров и чем более значительна водная преграда, тем больше обнаруживается отличий. На Мадагаскаре нет типичных для Африки крупных копытных:
быков, антилоп, зебр, нет и крупных хищников (львов, леопардов, гиен), высших обезьян. Однако Мадагаскар — последнее убежище лемуров. Когда-то, до появления обезьян, лемуры были доминирующими приматами. Но они не могли соперничать со своими более развитыми сородичами и исчезли повсюду, кроме Мадагаскара, который отделился от материка прежде, чем эволюционировали обезьяны. Другая большая группа мадагаскарских животных — тенреки и виверры — древние, мало изменившиеся за длительный период изоляции примитивные плацентарные млекопитающие. Мадагаскар обладает широким разнообразием мест обитания, и неудивительно, что он приютил большое количество различных птиц, 46 родов которых не встречаются больше нигде в мире. Трудно представить себе более причудливое животное, чем мадагаскарский хамелеон. Хамелеоны обитают и в Африке, но мадагаскарские виды крупнее и разнообразнее. К ним принадлежит самый большой хамелеон Устале длиной 60 см. У некоторых хамелеонов Мадагаскара на конце морды расположены рога, что делает их похожими на грозных миниатюрных динозавров. Любопытно, что при изобилии ядовитых змей на африканском континенте они отсутствуют на острове. Здесь широко представлены питоны и другие неядовитые змеи. Согласно истории живого мира змеи появились довольно поздно по сравнению с другими рептилиями, причем ядовитые змеи — самые из них молодые. Не значит ли это, что Мадагаскар отделился от континента до появления там змей? Мадагаскар — прекрасное место для лягушек, которых здесь насчитывается около 150 видов.
Иная картина обнаруживается при рассмотрении фауны океанических островов. Ее видовой состав беден и является результатом случайного занесения некоторых видов, обычно птиц, рептилий, насекомых. Наземные млекопитающие, амфибии и другие животные, не способные преодолевать значительные водные преграды, на большинстве таких островов отсутствуют. Приведем пример. Галапагосские острова удалены от берегов Южной Америки на 700 км. Это расстояние могут преодолеть только хорошо летающие формы. Оказалось, что 15% видов птиц Галапагоса представлены южноамериканскими видами, а 85% видов птиц отличны от материковых и нигде, кроме этого архипелага, не встречаются. Именно это с удивлением и наблюдал Ч. Дарвин на Галапагосских островах. Он обнаружил там птиц, которые хотя и напоминали вьюрков, виденных им в Южной Америке, но все были специализированы и мало похожи друг на друга. У этих 14 видов птиц, имевших общего предка, конкурентная борьба за корм и места гнездования была ослаблена или исчезла вовсе. Ч. Дарвин провел лишь три недели на раскаленных скальных островах. однако увиденное послужило ему материалом для размышлений на долгие годы. Особенно пристальное внимание он обратил на клюв вьюрков: у одних клюв был толстый, дробящий, у других — длинный, у третьих — острый, как ножницы. К одному из шести родов относится дятловый древесный вьюрок, одно ия редких в мире животных, использующих орудия. К сожалению, Ч. Дарвин не увидел эту замечательную птицу за работой, когда она колючкой кактуса выковыривает из древесной коры насекомых и личинок, подобно тому, как это делают дятлы клювом. Галапагосские вьюрки вошли в историю как первый пример влияния изоляции на развитие видов. Сегодня таких примеров известно много. Ч. Дарвин не побывал на Гавайских островах, где эффект изоляции проявляется еще более наглядно на гавайских цветочницах и улитках, прежде всего потому, что они обитают на этих отдаленных островах много дольше, чем вьюрки на Галапагосах.
biofile.ru
