
Цитоплазма бактерий — Справочник химика 21
Цитоплазма бактерий. Все содержимое клетки, ограниченное клеточной стенкой, называется протопластом. Протопласт состоит пз цитоплазматической мембраны и живого вещества клетки — цитоплазмы, или протоплазмы. Цитоплазма бактерий является бесцветной, прозрачной, слегка вязкой. [c.249]
Плазмиды наиболее часто используют в качестве векторов. Плазмиды -небольшие кольцевые двухцепочечные ДНК из цитоплазмы бактерий, они могут содержать от 2 до 100 тыс. пар оснований. Каждая плазмида имеет гены, которые могут реплицироваться, транскрибироваться, транслироваться независимо от хромосомных генов, но одновременно с ними. Плазмиды можно перемещать из одной клетки в другую их можно встраивать в другие гены, которые затем переносятся вместе с плазмидой и становятся частью генома клетки-хозяина. [c.61]
Рибосомы находятся в цитоплазме клеток. Обычно они шаровидны, их размер составляет всего 15—35 нм. В рибосомах происходит биосинтез белка. В 1943 г. рибосомы были обнаружены в цитоплазме бактерий, а затем в цитоплазме животных, растений и дрожжей. Они находятся на поверхности мембраны (тогда они активны) либо свободно плавают в цитоплазме. В состав рибосом входят рибонуклеопротеиды, т. е. РНК и белковый комплекс. Молекулярная масса рибосом составляет около 10 . Белки и РНК в рибосомах содержатся в количестве примерно по 40—60%.
Достигнув определенных размеров, диктуемых соотношением объемов ядра и цитоплазмы, бактерии переходят к бесполому размножению путем простого деления, т. е. путем деления на две идентичные дочерние клетки (рис. 2.11). Клеточному делению предшествует репликация [c.27]
Не промывая, препараты подсушивают фильтровальной бумагой и докрашивают 0,25%-ным раствором светлого зеленого или хризоидина. Затем промывают водой и высушивают. На препарате К цитоплазма бактерий окрашена в желто-коричневый цвет, включения волютина — в вишнево-красный. На препарате Щ гранулы волютина обсцвечены. [c.51]
Ртутьредуктаза представляет собой флавопротеин, локализованный в цитоплазме бактерий. [c.460]
Цитоплазма бактерий окрашивается в желтый, а волютин — в темно-синий, почти черный цвет. [c.51]
В цитоплазме бактерий не обнаружены эндоплазматическая сеть и митохондрии, ио имеются рибосомы 703. У гетеротрофных бактерий есть мезосомы — мембранные структуры — производные цитоплазматической мембраны. Нуклеоид в отличие от ядра высших организмов не окружен ядерной мембраной и не содержит ядрышка, он может быть разветвленной формы, имеет одну хромосому, состоящую из кольцевой ДНК, не связанной с гистонами. У бактерий обнаружены дополнительные генетические элементы, содержащие ДНК в виде плазмид, которые могут быть автономны в цитоплазме или включены в хромосомы. Эта особенность плазмид используется в генной инженерии.
Матрикс митохондрии Или цитоплазма бактерии [c.78]
А хН-зависимая аккумуляция 5 в отсеке, заряженном отрицательно (например, в матриксе митохондрии или в цитоплазме бактерии), описывается уравнением (34) [c.146]
В клетке бактерий обособленное ядро отсутствует, хотя ядерные белки (нук-леопротеиды) находятся в цитоплазме бактерий в большом количестве. [c.20]
Бактерии настолько малы, что находятся на грани разрешения обычного светового микроскопа. Их линейные размеры достигают всего лишь порядка 1 мкм. Поэтому в течение долгого времени было трудно при непосредственном визуальном наблюдении получить информацию об их внутренней структуре. Однако с появлением электронного микроскопа оказалось возможным выявить детальное строение бактериальной клетки, как это можно видеть на приведенной электронной микрофотографии (фиг. 21). Следует отметить, что увеличение на этой микрофотографии в пять раз больше, чем на предыдущей микрофотографии (фиг. 20). Следовательно, размер всей бактериальной клетки не превышает размера митохондрий, находящихся в цитоплазме клеток эукариотов. Хотя в прокариотической клетке нет истинного ядра, ДНК в ней явно локализована в определенном участке клетки, которую иногда называют центральным телом. Окружающая это тело часть клетки o epжит много РНК. Как и в эукариотической клетке, основная масса РНК в клетке прокариотов сосредоточена в рибосомах — гранулярный фон на большей части клеток (фиг. 21). Эндоплазматической же сети в клетках прокариотов нет. По 4юрмальной аналогии с областью клетки эукариотов, в которой сосредоточена ДНК, содержащее ДНК пентральное тело бактерии часто называют ядром , остальную часть клетки обычно называют цитоплазмой бактерии. Это парадоксальное распространение терминов, используемых для эукариотов, на бактерии, отличающиеся от клеток высших форм отсутствием именно этих структур, настолько устоялось в молекулярной генетике, что в дальнейшем нельзя будет избежать употребления этих неточных слов.
Тесная связь между функциями ядра и цитоплазматическими эндосимбионтами была продемонстрирована в конце 60-х годов у Amoeba proteus К. Джеоном. Культура А. proteus была случайно заражена бактериями, которые проникли в цитоплазму простейшего и размножались там до численности 150 тыс. шт. на клетку. Большинство амеб погибло, однако часть из них выжила и активно делилась в присутствии инфицировавшей их цитоплазму бактерии. Число бактериальных клеток в амебе достигало теперь примерно 50 ООО. [c.249]
Трехмерная структура и расположение в мембране. Я+ — АТФ-синтазный комплекс так велик, что выдается в воду на довольно большое расстояние с одной стороны мембраны. Выступающая часть, которая представляет собой фактор Fi, обращена в цитоплазму бактерий, матрикс митохондрии или строму хлоропласта. [c.132]
Давайте обратимся к механизму транспорта К+ и Ыа+ через бактериальную мембрану. Известно, что между цитоплазмой бактерии и внешней средой существует разность электрических потенциалов, поддерживаемая работой белков-генераторов в бактериальной мембране. Откачивая протоны изнутри клетки наружу, белки-генераторы тем самым заряжают внутренность бактерии отрицательно. В этих условиях накопление ионов К+ внутри клетки могло бы происходить просто за счет электрофореза — движения Боложительно заряженного иона калия в отрицательно заряженную цитоплазму бактерии. [c.170]
Цитоплазматическая мембрана является трехслойной структурой и окружает наружную часть цитоплазмы бактерий. По структуре она похожа на цитоплазматическую мембрану клеток животных состоит из двойного слоя липидов, главным образом фосфолипидов со встроенными поверхностными и интефальны-ми белками, как бы пронизываюш.ими насквозь структуру мембраны. Некоторые из них являются пермеазами, участвующими в транспорте веществ. Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную, текучую структуру. Она участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки (за счет ферментов цепи переноса электронов, АТФ-азы и др.).
Цитоплазма бактерий занимает основной объем клетки и состоит из растворимых белков. Рибосомы бактерий имеют коэффициент седиментации 70 5 в отличие от рибосом, характерных для эукариотических клеток (80 8). Поэтому некоторые антибиотики, действие которых основано на подавлении синтеза белка путем связывания их с рибосомами бактерий, не оказывают влияния на синтез белка эукариотических клеток. В цитоплазме имеются различные включения — полисахариды, поли-р-масляная кислота и полифосфаты (волютин). Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей. Зерна волютина выявляются у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. [c.25]
Генетика микроорганизмов как учение о наследственности и изменчивости имеет характерные особенности, соответствующие их сфоению и биологии. Наиболее изучена генетика бактерий, характерными чертами которых являются малые размеры и большая скорость размножения бактериальной клетки, что позволяет проследить генетические изменения в течение небольшого промежутка времени на большом числе популяций. Бактериальная клетка имеет одинарный набор генов (нет аллелей). Хромосома бактерий является полинуклеотидом (две полинуклеотидные цепочки ДНК) длиной 1000 мкм и мол. массой около 1,5—2 10 Д. Она суперспирализована и замкнута в кольцо содержит от 3000 до 5000 генов. Аналогично хромосоме в цитоплазме бактерий располагаются ковалентно замкнутые кольца ДНК, называемые плазмидами (внехромосомные факторы наследственности). Масса плазмид значительно меньше массы хромосом. Хромосома и плазмида способны к автономному самокопированию — репликации, поэтому их называют репликонами. Свойства микроорганизмов, как и любых других организмов, определяются их генотипом, т.е. совокупностью генов данной особи. Термин геном в отношении микроорганизмов — почти синоним понятия генотип .
chem21.info
Цитоплазма, ядро, жгутики
Все содержимое клетки, за исключением ядра и клеточной стенки, называется цитоплазмой. В жидкой, бесструктурной фазе цитоплазмы (м а т р и к с е) находятся рибосомы, мембранные системы, митохондрии, пластиды и другие структуры, а также запасные питательные вещества. Цитоплазма обладает чрезвычайно сложной, тонкой структурой (слоистая, гранулярная). С помощью электронного микроскопа раскрыты многие интересные детали строения клетки.
Рис. Основные типы расположения жгутиков и направления движений бактерий.
Внешний липопротеидный слой протопласта бактерий, обладающий особыми физическими и химическими свойствами, называется цитоплазмат и ческой мембраной (рис. 2, 15).
Внутри цитоплазмы находятся все жизненно важные структуры и органеллы.
Цитоплазматическая мембрана выполняет очень важную роль — регулирует поступление веществ в клетку и выделение наружу продуктов обмена.
Через мембрану питательные вещества могут поступать в клетку в результате активного биохимического процесса с участием ферментов. Кроме того, в мембране происходит синтез некоторых составных частей клетки, в основном компонентов клеточной стенки и капсулы. Наконец, в цитоплазматической мембране находятся важнейшие ферменты (биологические катализаторы). Упорядоченное расположение ферментов на мембранах позволяет регулировать их активность и предотвращать разрушение одних ферментов другими. С мембраной связаны рибосомы — структурные частицы, на которых синтезируется белок. Мембрана состоит из липопротеидов. Она достаточно прочна и может обеспечить временное существование клетки без оболочки. Цитоплазматическая мембрана составляет до 20% сухой массы клетки.
На электронных фотографиях тонких срезов бактерий цитоплазматическая мембрана представляется в виде непрерывного тяжа толщиной около 75А, состоящего из светлого слоя (липиды), заключенного между двумя более темными (белки). Каждый слой имеет ширину 20—30 А. Такая мембрана называется элементарной (табл. 30, рис. 8).
Между плазматической мембраной и клеточной стенкой имеется связь в виде десмозов -мостиков. Цитоплазматическая мембрана часто дает инвагинации — впячивания внутрь клетки. Эти впячивания образуют в цитоплазме особые мембранные структуры, названные мезосомами. Некоторые виды мезосом представляют собой тельца, отделенные от цитоплазмы собственной мембраной. Внутри таких мембранных мешочков упакованы многочисленные пузырьки и канальцы (рис. 2). Эти структуры выполняют у бактерий самые различные функции. Одни из этих структур — аналоги митохондрий. Другие выполняют функции эндо-плазматической сети или аппарата Гольджи. Путем инвагинации цитоплазматической мембраны образуется также фотосинтезирующий аппарат бактерий. После впячивания цитоплазмы мембрана продолжает расти и образует стопки (табл. 30), которые по аналогии с гранулами хлоропластов растений называют стопками ти-лакоидов. В этих мембранах, часто заполняющих собой большую часть цитоплазмы бактериальной клетки, локализуются пигменты (бактериохлорофилл, каротиноиды) и ферменты (цитохромы), осуществляющие процесс фотосинтеза.
В цитоплазме бактерий содержатся рибосомы— белок-синтезирующие частицы диаметром 200А. В клетке их насчитывается больше тысячи. Состоят рибосомы из РНК и белка. У бактерий многие рибосомы расположены в цитоплазме свободно, некоторые из них могут быть связаны с мембранами.
Рибосомы являются центрами синтеза белка в клетке. При этом они часто соединяются между собой, образуя агрегаты, называемые полирибосомами или полисомами.
В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Однако их присутствие нельзя рассматривать как какой-то постоянный признак микроорганизма, обычно оно в значительной степени связано с физическими и химическими условиями среды. Многие цитоплазматические включения состоят из соединений, которые служат источником энергии и углерода. Эти запасные вещества образуются, когда организм снабжается достаточным количеством питательных веществ, и, наоборот, используются, когда организм попадает в условия, менее благоприятные в отношении питания.
У многих бактерий гранулы состоят из крахмала или других полисахаридов -гликогена и гранулезы. У некоторых бактерий при выращивании на богатой сахарами среде внутри клетки встречаются капельки жира. Другим широко распространенным типом гранулярных включений является волютин (метахроматиновые гранулы). Эти гранулы состоят из полиметафосфата (запасное вещество, включающее остатки фосфорной кислоты). Полиметафосфат служит источником фосфатных групп и энергии для организма. Бактерии чаще накапливают волютин в необычных условиях питания, например на среде, не содержащей серы. В цитоплазме некоторых серных бактерий находятся капельки серы.
Помимо различных структурных компонентов, цитоплазма состоит из жидкой части — растворимой фракции. В ней содержатся белки, различные ферменты, т-РНК, некоторые пигменты и низкомолекулярные соединения — сахара, аминокислоты.
В результате наличия в цитоплазме низкомолекулярных соединений возникает разность в осмотическом давлении клеточного содержимого и наружной среды, причем у разных микроорганизмов это давление может быть различным. Наибольшее осмотическое давление отмечено у грамположительных бактерий — 30 атм, у грамотрицательных бактерий оно гораздо ниже — 4—8 атм.
Ядерный аппарат.
В центральной части клетки локализовано ядерное вещество — дезоксирибонуклеиновая кислота (ДНК).
У бактерий нет такого ядра, как у высших организмов (эукариотов), а есть его аналог — «ядерный эквивалент» — нуклеоид (см. рис. 2, 8), который является эволюционно более примитивной формой организации ядерного вещества. Микроорганизмы, не имеющие настоящего ядра, а обладающие его аналогом, относятся к прокариотам. Все бактерии -прокариоты. В клетках большинства бактерий основное количество ДНК сконцентрировано в одном или нескольких местах. В клетках эукариотов ДНК находится в определенной структуре — ядре. Ядро окружено оболочкой— мембраной.
У бактерий ДНК упакована менее плотно, в отличие от истинных ядер; нуклеоид не обладает мембраной, ядрышком и набором хромосом. Бактериальная ДНК не связана с основными белками — гистонами — ив нуклео-иде расположена в виде пучка фибрилл.
Жгутики.
На поверхности некоторых бактерий имеются придаточные структуры; наиболее широко распространенными из них являются жгутики — органы движения бактерий.
Жгутик закрепляется под цитоплазматической мембраной с помощью двух пар дисков (см. на стр. 206). У бактерий может быть один, два или много жгутиков. Расположение их различно: на одном конце клетки, на двух, по всей поверхности и т. д. (рис. выше). Жгутики бактерий имеют диаметр 0,01—0,03 мкм, длина их может во много раз превосходить длину клетки. Бактериальные жгутики состоят из белка — флагеллина — и представляют собой скрученные винтообразные нити.
На поверхности некоторых бактериальных клеток имеются тонкие ворсинки — фимбрии.
www.berl.ru
Структурные элементы бактериальной клетки:
|
ПОСТОЯННЫЕ |
НЕПОСТОЯННЫЕ |
|
клет.стенка |
капсула |
|
цитоплазматич. мембрана |
жгутики |
|
цитоплазма |
споры |
|
рибосомы |
включения |
|
мезосомы |
пили (ворсинки) |
|
нуклеоид |
плазмиды |
№11 Цитоплазматическая мембрана, цитоплазма, рибосомы, мезосомы, генофор, их строение, функции и значение для бактериальной клетки.
-
Цитоплазматическая мембрана
Цитоплазма бактериальной клетки ограничена от клеточной стенки тонкой полупроницаемой структурой толщиной 5-10 нм, называемой цитоплазматической мембраной (ЦПМ). ЦПМ состоит из двойного слоя фосфолипидов, пронизанных белковыми молекулами (рис. 6).
С ЦПМ связаны многие ферменты и белки, участвующие в транслокации питательных веществ, а также ферменты и переносчики электронов конечных стадий биологического окисления (дегидрогеназы, цитохромная система, АТФ-аза). На ЦМП локализуются ферменты, катализирующие синтез пептидогликана, белков клеточной стенки, собственных структур. Мембрана является также местом превращения энергии при фотосинтезе, окислительном фосфорилировании.
-
Периплазматическое пространство
Периплазматическое пространство (периплазма) представляет собой зону между клеточной стенкой и ЦПМ. Толщина периплазмы составляет около 10 нм, объем зависит от условий среды и, прежде всего, от осмотических свойств раствора. Периплазма может включать до 20% всей находящейся в клетке воды, в ней локализуются некоторые ферменты (фосфатазы, пермеазы, нуклеазы и др.) и транспортные белки – переносчики соответствующих субстратов.
Цитоплазма
Содержимое клетки, окруженное ЦПМ, составляет цитоплазму бактерий. Та часть цитоплазмы, которая имеет гомогенную коллоидную консистенцию и содержит растворимые РНК, ферменты, субстраты и продукты обмена веществ, обозначается как цитозоль. Другая часть цитоплазмы представлена различными структурными элементами: мезосомами, рибосомами, включениями, нуклеоидом, плазмидами.
Рибосомы —
Рибосомы — субмикроскопические рибонуклеопротеидные гранулы диаметром 15-20 нм. В рибосомах находится примерно 80-85% всей бактериальной РНК. Рибосомы прокариот имеют константу седиментации 70 S. Они построены из двух частиц: 30 S (малая субъединица) и 50 S (большая субъединица) (рис. 8). Рибосомы служат местом синтеза белка.
Рис. 8. Рибосома (а) и ее субчастицы-большая (б) и малая (в) (Блинов Н.П., 1989).
Некоторые бактерии способны накапливать фосфорную кислоту в виде гранул полифосфата (зерна волютина, метахроматические зерна, зерна Бабеша-Эрнста). Они играют роль фосфатных депо и регулярно выявляются у коринебактерий, микобактерий и спирилл в виде плотных, хорошо контурированных образований в форме шара или эллипса, располагающихся, в основном, у полюсов клетки. Обычно на полюсах бывает по одной грануле.
Наличие зерен волютина у бактерий определяют методом Нейссера
Мезосомы
Мезосомы представляют собой мембранные структуры, образуемые при закручивании ЦПМ. Морфологически мезосомы выглядят как ламеллярные стопки или спирально упакованные ламеллы, везикулярные или тубулярные структуры, а также смешанные мембранные системы, образованные трубочками, пузырьками и ламеллами (рис. 7). По расположению в клетке различают: мезосомы, образующиеся в зоне клеточного деления и формирования клеточной перегородки (септальныемезосомы) и мезосомы, сформированные в результате инвагинации периферических участков ЦПМ (латеральные мезосомы).
Предполагается, что мезосомыполифункциональны, содержат различные ферментные системы и играют определенную роль в энергетическом метаболизме. Считают, что они являются сайтом для формирования клеточной стенки бактерий и прикрепления нуклеоида в процессе репликации ДНК. Септальныемезосомы участвуют в построении поперечной перегородки при делении бактерий.
бактериальная хромосома, или генофор)
studfiles.net
Ультраструктура бактериальной клетки
Бактериальная клетка, несмотря на внешнюю простоту строения, представляет собой весьма сложный организм, для которого характерны процессы, свойственные всем живым существам.
Ультраструктуру бактерий удалось детально изучить после создания электронных микроскопов с большой разрешающей способностью, разработки техники получения ультратонких срезов клеток, появления фазово-контрастной микроскопии, усовершенствования методов микрохимических анализов. Разнообразные методы исследований дали возможность определить различные поверхностные и внутренние структуры у бактерий, (рис. 5).
К внешним структурам обычно относят капсулы, жгутики, фимбрии и пили, а также клеточную стейку, под которой расположена цитоплазматическая мембрана. Внутреннее содержимое бактерий представлено цитоплазмой, в которой находятся нуклеоид, рибосомы и мембранные структуры, а также разнообразные включения. Бациллы и некоторые другие бактерии образуют споры.
Капсула. Большинство бактерий окружены слоем вещества, расположенного поверх клеточной стенки. Это слизистое образование — капсула (рис. 6). Бывают макрокапсулы (толщина слоя 0,2 мкм), микрокапсулы (менее 0,2 мкм), слизистый слой и растворимая слизь.
По химическому составу капсулы бактерий можно разделить на два типа. Одни представлены полисахаридами, другие — полипептидами. Однако встречаются капсулы, состоящие из липидов, (у туберкулезных бактерий), гетерополисахаридов и других веществ. Капсулы содержат до 98% воды. Поэтому они создают дополнительный осмотический барьер, а также защищают клетку от механических повреждений и высыхания. Капсулы защищают клетки и от других неблагоприятных воздействий окружающей среды.
Замечено, что бактерии, имеющие капсулы, могут жить в такой среде, в которой рост бактерий без капсул ограничен.
Жгутики. Существуют два типа подвижных бактерий: скользящие и плавающие. Скольжение наблюдается у миксобактерий. и серных бактерий. Эти организмы могут совершать скольжение по поверхности в результате волнообразных сокращений, вызывающих периодическое изменение формы клетки.
Плавающие палочковидные бактерии передвигаются с помощью особых нитевидных придатков — жгутиков. За счет жгутиков передвигается большинство спирилл. Кокки, за исключением отдельных видов, не имеют жгутиков.
Бактерия с одним жгутиком называется монотрихом; бактерия с пучком жгутиков на одном конце клетки — лофотрихом; на обоих концах — амфитрихом; бактерия со жгутиками, расположенными по всей поверхности клетки, называется перитрихом (рис. 7).
Число жгутиков различно у разных видов бактерий. Например, спириллы (Spirillum) имеют от 5 до 30 жгутиков, вибрионы (Vibrio) — 1 или 2—3 жгутика на полюсе клетки, а у палочковидных бактерий Proteus vulgaris и Clostridium tetani обнаружено от 50 до 100 жгутиков. Толщина жгутиков колеблется от 10 до 20 нм, длина — от 3 до 15 .мкм, причем у одной и той же бактериальной клетки длина может изменяться в зависимости от состояния культуры и факторов внешней среды. В химическом отношении жгутики представляют собой белок флагеллин.
Белковые молекулы, из которых состоят жгутики, собраны в спиральные цепи, закругленные вокруг полой сердцевины.
Жгутики хорошо видны в электронном микроскопе, для наблюдения через оптический микроскоп требуется их специальная обработка. Жгутики не относятся к жизненно важным структурам бактериальной клетки. Так, бактерии, обладающие жгутиками, можно вырастить в условиях, при которых эти структуры у них не развиваются. У подвижных бактерий наблюдаются «фазовые вариации», то есть в течение одной фазы развития жгутики имеются, в другой — отсутствуют. Жгутики можно разрушить, а клетка останется жизнеспособной.
Жгутики прикрепляются к особой структуре — базальному тельцу, расположенному под цитоплазматической мембраной. Движение жгутику сообщает через его основание жгутиковый «мотор» — базальное тельце, состоящее из центрального стержня, вставленного в систему колец, которые вращаются относительно друг друга.
Бактериальные клетки со жгутиками двигаются со скоростью, которая зависит от особенностей их аппарата движения и свойств среды – вязкости, температуры рН, осмотического давления и др. Большинство бактерий за секунду проходят расстояние, равное размерам их клетки.

Однако некоторые бактерии при благоприятных условиях за то же время могут передвигаться на расстояние, превышающее размеры клетки в 50 раз и более.
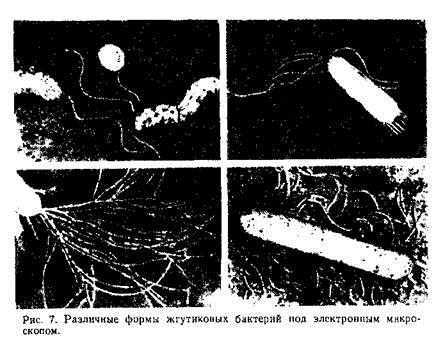
Бактерии передвигаются беспорядочно, но способны и к направленным движениям, так называемым таксисам. В зависимости от внешних факторов, под воздействием которых происходит движение, различают хемотаксис, обусловленный разницей в концентрации химических веществ в среде, аэротаксис, связанный с разницей в содержании кислорода, и, наконец, фототаксис, когда условием направленного движения бактерий является различие в интенсивности освещения.
Фимбрии и пили. Кроме жгутиков, клетки бактерий могут иметь длинные, тонкие, прямые нити — фимбрии. Фимбрии значительно короче и тоньше жгутиков, но более многочисленны. Обнаружены они как у подвижных, так и у неподвижных организмов. Размеры фимбрий 0,3—4 мкм в длину и 5—10 нм в ширину. Число их достигает 100—200 и более (до нескольких тысяч) на одну бактериальную клетку.
Фимбрии состоят из белка — пилина.
В настоящее время известно несколько типов фимбрий, которые отличаются своими функциями. Наиболее изучены функции фимбрий первого и второго типов. Фимбрии первого типа имеют многие бактерии,
Что дало возможность назвать их «фимбриями общего типа». Наличие фимбрий первого типа помогает бактериальной клетке прилипать к другим клеткам или инертному субстрату, или способствовать образованию пленок на поверхности жидкостей, и поэтому считают, что фимбрии этого типа — органы прикрепления (рис. 8).
Большой интерес представляют фимбрии второго типа, так называемые половые фимбрии, или пили, имеющие внутри канал, через который передается генетический материал от одной клетки к другой при конъюгации бактерий. Пили также могут служить для прикрепления патогенных бактерий к тканям животных и человека.
Клеточная стенка — один из главных элементов структуры бактериальной клетки. Клеточная стенка обладает определенной ригидностью (жесткостью), но вместе с тем эластичностью и может изгибаться. Ее можно разрушить ультразвуком, ферментом лизоцимом и другими способами. В результате разрушения клеточной стенки содержимое клетки — цитоплазма с ее включениями, окруженная цитоплазматической мембраной, приобретает шаровидную форму. Такая округлившаяся клетка, образовавшаяся после удаления клеточной стенки у бактерии, называется протопластом (сферопластом). Отсюда следует, что стенка придает ‘бактериальной клетке определенную форму.
Клеточная стенка имеет и другие функции. Она защищает внутреннее содержимое клетки от действия механических и осмотических сил внешней среды, ей принадлежит важная роль в регуляции роста и деления бактерий и распределении генетического материала.
Толщина клеточной стенки колеблется от 10 до 80 нм и составляет около 20% сухого вещества бактериальной клетки. Клеточная стенка относительно проницаема для крупных молекул. Она связана с цитоплазматической мембраной соединительными тяжами — «мостиками».
Считают, что клеточная стенка ответственна за окрашивание бактерий по Граму. Способность (или, наоборот, неспособность) окрашиваться по Грамму связана с различием в химическом составе клеточных стенок бактерий.
Главным структурным компонентом клеточных стенок большиства исследованных бактерий является пептидогликан (муреин), представляющий собой гетерополимер, который построен из чередующихся остатков N-ацетил-N-глюкозамина и N-ацетилмурамовой кислоты, соединенных в-1,4-связями. Полисахаридные цепи связаны между собой короткими пептидными мостиками. Пептидогликан придает клеточной стенке ригидные свойства, благодаря чему бактериальная клетка способна сохранять свою форму.
У грамположительных бактерий клеточная стенка состоит главным образом из многослойного пептидогликана, в который сложным образом вплетены такие сопутствующие компоненты, как белки, полисахариды, а также так называемые тейхоевые кислоты (полимеры рибитфосфорной и глицеринфосфорной кислот).
У грамотрицательных бактерий пептидогликан однослойный. Сверху пептидогликана располагается структура, получившая название наружной мембраны. Она имеет мозаичное строение и состоит из фосфолипидов, липопротеидов, белков и сложного липополисахарида (ЛПС).
Содержание пептидогликана в клеточных стенках колеблется от 50 до 90% у грамположительных бактерий и от 1 до 10% у грамотрицательных.
Таким образом, неодинаковое отношение бактерий к окраске по Граму может быть объяснено различием в количестве пептидогликана и его локализацией в клеточной стенке.
В настоящее время выявлено, что поверхность клеточной стенки некоторых палочковидных форм бактерий покрыта выростами, шипами или буграми.
Клеточной стенки нет у микоплазм, а также у L-форм бактерий. Наименование L-форм (от названия Института имени Листера (Великобритания), где впервые изучены эти формы) получили бактерии, полностью или частично лишенные клеточной стенки (под влиянием антибиотиков или спонтанно без видимой причины) и сохранившие способность к размножению. Для L-форм разных бактерий характерно образование крупных и мелких шаровидных клеток. L-формы описаны у многих болезнетворных и сапрофитных бактерий.
Цитоплазматическая мембрана (ЦПМ). К клеточной стенке бактериальной клетки тесно прилегает внешний слой цитоплазмы — цитоплазматическая мембрана, состоящая обычно из двойного слоя липидов, каждая из поверхностей которого покрыта мономолекулярным слоем белка. Мембрана составляет около 8—15% сухого вещества клетки и содержит до 70—90% липидов клетки. Общая толщина мембраны равняется приблизительно 9 нм.
Цитоплазматическая мембрана играет роль осмотического барьера, контролирующего транспорт веществ в бактериальную клетку и из нее. Нередко мембрана дает внутрицитоплазматические впячивания (инвагинации), приводящие к образованию особых телец—мезосом.
Цитоплазматическая мембрана и мезосомы выполняют функции, свойственные мембранам и митохондриям высших организмов, в которых или на которых локализованы ферментные системы-поставщики энергии. В отличие от митохондрий в цитоплазматической мембране и мезосомах бактерий наряду с дыхательными системами ферментов и механизмом регуляции проницаемости располагаются специфичные ферментные системы, участвующие в таких процессах, как азотфиксация, хемосинтез и др.
С цитоплазматической мембраной, мезосомами и близкими структурами бактерий связаны также многие другие функции — биосинтез клеточной стенки и капсулы, выделение экзоферментов, деление и спорообразование и т. д.
Цитоплазма. Под цитоплазматической мембраной у бактерий находится цитоплазма. Это коллоидная система, состоящая из воды, белков, жиров, углеводов, минеральных соединений и других веществ, соотношение которых варьирует в зависимости от вида бактерий и их возраста. Цитоплазма бактерий имеет различные структурные элементы — внутрицитоплазматические мембраны, генетический аппарат, рибосомы и включения, остальная часть ее представлена цитозолем.
Цитозоль — это фракция цитоплазмы, которая имеет гомогенную консистенцию и состоит главным образом из белковых макромолекул (растворимых РНК, ферментных белков, продуктов и субстратов различных реакций) и служит поддерживающей средой для клеточных гранул. Изучение структуры цитоплазмы выявило ее мелкогранулярный характер, она представлена цитоплазматическими гранулами диаметром 10—20 нм. Многие из этих гранул являются рибосомами — частицами, состоящими из РНК (60%) и белка (40%). Каждая бактерия содержит от 5000 до 50 000 рибосом, которые служат центрами синтеза белков. Рибосомы совместно с молекулами информационной и транспортных РНК участвуют в синтезе белка в форме не изолированных частиц, а их агрегатов, называемых полирибосомами, или полисомами.
В цитоплазме цианобактерий имеются так называемые Тилакоиды (или Фикобилисомы) — мембранные фотосинтезирующие структуры, содержащие хлорофилл и каротиноиды, при помощи которых осуществляется фотосинтез. У пурпурных серобактерий фотосинтезирующие пигменты (бактериохлорофилл и каротиноиды) локализованы в хроматофорах, которые составляют от 40 до 50% массы клетки. Тилакоиды в основном состоят из белков и липидов. Предполагают, что тилакоиды связаны с цитоплазматической или внутренними мембранами. У зеленых бактерий пигменты, участвующие в фотосинтезе, содержатся в мембранных структурах, называемых хлоросомами. Клетки ряда водных бактерий содержат наполненные газом структуры — газовые вакуоли (аэросомы). Некоторые бактерии имеют структуры, называемые полиэдральными телами (в виде многоугольника), или карбоксисомами, где осуществляется процесс связывания С02 (у автотрофных бактерий).
Включения. В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Их присутствие нельзя рассматривать как постоянный признак микроорганизма, обычно они в значительной степени связаны с физическими и химическими условиями среды обитания.
Многие внутрицитоплазматические гранулы состоят из соединений, которые служат для микроорганизмов источником энергии и углерода. Такие соединения обычно образуются, когда микроорганизм снабжается достаточным количеством питательных веществ, и используются, когда он попадает в неблагоприятные в отношении питания условия. В качестве резервных питательных веществ в клетках бактерий могут накапливаться вещества, состоящие из углеводов — гранулы гликогена (крахмала) или гранулезы (близкого к крахмалу полисахарида). При недостаточном поступлении углеродсодержащих веществ в питательную среду гранулы гликогена или гранулезы постепенно исчезают из клеток бактерий.
Большая часть бактерий в качестве резервного вещества синтезирует полимер из в — оксимасляной кислоты (поли — в — оксимасляная кислота). У некоторых видов бактерий в клетках накапливаются гранулы жира и волютина. Волютиновые гранулы, называемые еще метахроматическими гранулами, состоят преимущественно из полифосфатов и служат запасным источником фосфора. Волютин обнаруживается в виде крупных, хорошо видимых гранул, образующихся в больших количествах на средах, богатых глицерином или углеводами. В клетках серных бактерий в качестве включений встречается сера, которая образуется в результате окисления сероводорода и видна в виде расположенных непосредственно в цитоплазме блестящих полужидких капелек. Включения серы для аэробных тионовых бактерий, окисляющих сероводород, служат источником энергии. Некоторые серные бактерии наряду с капельками серы имеют зернышки аморфного карбоната кальция, роль которого пока не выяснена.
В цитоплазматическом матриксе содержатся также растворимые белки, различные ферменты, РНК, пигменты и низкомолекулярные соединения — углеводы, аминокислоты и нуклеотиды. Наличие в цитоплазме низкомолекулярных соединений обусловливает разность в осмотическом давлении клеточного содержимого и внешней среды. Величина внутриклеточного осмотического давления значительно изменяется у разных микроорганизмов.
Нуклеоид. В цитоплазме бактериальных клеток расположен эквивалент ядра, называемый нуклеоидом. Нуклеоид бактериальной клетки находится в ее центральной части. Предполагают, что в зависимости от стадии развития клетки нуклеоид может быть либо дискретным (прерывистым, состоящим из отдельных форменных структур), либо в виде хроматиновой сети — ядерного вещества, дисперсно распределенного в цитоплазме.
Ядерные структуры, присутствующие в цитоплазме бактерий в дискретном состоянии, имеют палочковидную форму (рис. 9).
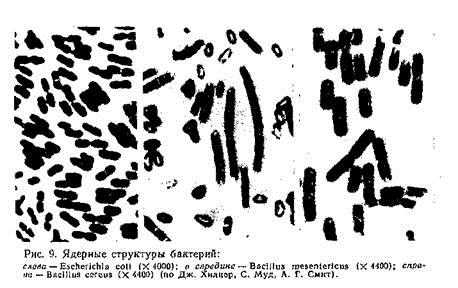
Нуклеоиды бактерий содержат ДНК с молекулярной массой 2—3-109. Установлено, что бактериальная ДНК имеет форму свернутой в кольцо нити длиной 1,1 — 1,4 мм, называемой также бактериальной хромосомой и генофорой.
В покоящейся бактериальной клетке обычно содержится один нуклеоид; клетки, находящиеся в фазе, предшествующей делению, имеют два нуклеоида; в фазе логарифмического роста — размножения — до четырех и более нуклеоидов. В условиях, которые отрицательно сказываются на росте бактерий, могут возникать нитевидные многоядерные клетки. Это объясняется нарушением синхронизации между скоростью роста клетки и скоростью деления клетки и нуклеоида.
Нуклеоид бактерий — основной носитель информации о свойствах клетки и основной фактор передачи этих свойств потомству.
Кроме нуклеоида, в цитоплазме бактериальной клетки могут — находиться в сотни раз более короткие нити ДНК — так называемые внехромосомные факторы наследственности, получившие название плазмид. Как выяснено, плазмиды необязательно имеются у бактерий, но они придают организму дополнительные, полезные для него свойства, в частности связанные с размножением, устойчивостью к лекарственным препаратам, болезнетворностью и др.
Споры и спорообразование. Бактерии рода Bacillus, Clostridium и Desulfotomaculum, так же как и отдельные кокки и спириллы, способны образовывать споры (эндоспоры) — тельца сферической или эллиптической формы, устойчивые к воздействию неблагоприятных факторов (рис.10).
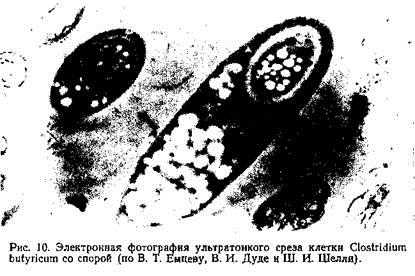
Споры преломляют свет и четко видны в световом микроскопе. Как правило, внутри бактериальной клетки образуется только одна спора. Однако в последнее время у отдельных видов Clostridium обнаружены клетки с двумя и более спорами. Обычно спорообразование начинается, когда бактерии испытывают недостаток питательных веществ или когда в среде в большом количестве накапливаются продукты обмена веществ бактерий. Поэтому споры можно рассматривать как приспособление организма для выживания в неблагоприятных условиях среды.
Формирование спор зависит от условий роста. Споры могут оставаться живыми в условиях, когда вегетативные клетки, то есть клетки, не образовавшие споры, погибают. Большинство спор хорошо переносит высушивание, многие споры нельзя убить даже кипячением в течение нескольких часов. Для их уничтожения требуется температура пара 120° С при давлении его 1 атм. (1,01* 105 Па). При этих условиях споры погибают через 20 мин. В сухом состоянии они погибают лишь при сильном нагревании (до 150—160°С) в течение нескольких часов. Споры отдельных видов бактерий отличаются особенной термоустойчивостью.
В процессе образования спор осуществляется синтез особого соединения — дипиколиновой кислоты (пиридин-2,6-дикарбоновая кислота), обычно отсутствующей у вегетативных клеток бактерий. Дипиколиновая кислота может составлять 10—15% массы сухой споры. Показано, что это вещество накапливается в центральной части споры — ее цитоплазме, образуя с ионами кальция комплекс, который вместе с повышенным содержанием других катионов (магния, марганца и калия)
Обеспечивает пребывание спор в состоянии покоя и их термоустойчивость.
Общая схема спорообразования может быть представлена в следующем виде. В результате неравномерного деления бактериальной клетки, сопровождающегося впячиванием цитоплазматической мембраны, наблюдается обособление части нуклеоида с небольшой частью цитоплазмы. Образовавшаяся проспора затем покрывается цитоплазматической мембраной бактериальной клетки.
Таким образом, внутри клетки бактерии возникает новая клетка — проспора, окруженная двумя мембранами. Затем между мембранами образуется кортикальный слой, или кортекс, состоящий из особых молекул пептидогликана.
Дальнейшее развитие споры заключается в образовании нескольких слоев споровых покровов и ее созревании. Споровые покровы формируются в основном из вновь синтезированных особых белков, а также липидов и гликолипидов. Электронномикроскопическое изучение ультратонких срезов спор многих бактерий показало, что поверх покровов споры образуется еще одна структура — экзоспориум, часто состоящий из ряда слоев и имеющий подчас разнообразную «лепную» форму. Диаметр споры приблизительно равен диаметру клетки, в которой она образовалась, или несколько превышает его. У некоторых бактерий спора формируется на конце клетки, которая при этом несколько расширяется, приобретая вид барабанной палочки. У других бактерий спора образуется в центре клетки, и последняя либо не меняет формы (род Bacillus), либо расширяется в середине, принимая вид веретена (род Clostridium).
После созревания споры клеточная стенка вегетативной части клетки разрушается, и спора выходит в окружающую среду. При попадании в благоприятные условия спора начинает прорастать.
Прорастанию предшествует поглощение спорой воды и последующее набухание. Затем оболочка под влиянием давления, вызванного ростом, разрывается, возникает ростовая трубка. В дальнейшем происходит удлинение освободившегося бактериального организма и, наконец, деление уже удлиненной клетки.
Споры бактерий могут длительное время (десятки, сотни и даже тысячи лет) существовать в покоящемся состоянии.
Имеются микроорганизмы, образующие относительно устойчивые к неблагоприятным условиям среды (температура, кислотность, аэрация и др.) покоящиеся клетки — цисты, не являющиеся спорами. Например, азотобактер образует цисты, устойчивые к высушиванию и теплу.
Известны и другие группы покоящихся клеток (миксоспоры миксобактерий, акинеты цианобактерий, эндоспоры актиномицетов и др.).
agroinf.com
15. Морфология бактерий
Компартменты (части) бактериальной клетки
Клеточная стенка
Цитоплазматическая мембрана
Цитоплазма
(нуклеоид, рибосомы, включения)
16. Цитоплазма бактериальной клетки
Цитоплазма – гелеобразная субстанция, окруженная ЦПМ, содержит рибосомы, белки и ферменты.
В цитоплазме cтруктурно различимы: 1. Рибосомы 2. Нуклеоид 3. Включения
Рибосомы бактерий
Прокариотические Р относятся к 70S типу.
Эукариотические Р — к 80S типу.
Р имеют четкие контуры.
Р – мембраной не окружены.
Количество Р строго регулируется и составляет ~ от 1-100 x 1000 на клетку.
В клетке E.coli – 1000 Р.
Р распределены диффузно в цитоплазме
Иногда Р расположены вблизи ЦПМ.
Бактериальные Р содержат 55 белков и 3 типа рРНК
30S — малая субъединица:
21 белок + 16S РНК
50S — большая субъединица:
34 белка + 5S, 23S РНК
Функции рибосом
Р выполняют функцию трансляции генетической информации – синтез белка на матрице иРНК.
Р, нанизанные на иРНК образуют полисомы.
Нуклеоид
2-й структурный компонент цитоплазмы
четко различима область цитоплазмы с ДНК, не окруженная мембраной.
Н включает хромосому и лишен рибосом.
Хромосома и плазмиды составляют геном
Геном – совокупность генов.
17. Строение и функции цитоплазматической мембраны
Цитоплазма бактерии окружена ЦПМ
ЦПМ — билипидная универсальная структура — физиологически активное образование
ЦПМ – высокоселективный барьер, обеспечивает существование клетки
Жизнеспособность бактерий обусловлена свойствами ЦПМ:
Текучесть – белки способны свободно перемещаться в толще мембраны
Флексибильность — способность изгибаться
Стабильность — за счет ионов Ca2+ и Mg2+
Строение ЦПМ
У большинства бактерий снаружи от ЦПМ есть особая структура – клеточная стенка (КС)
КС отсутствует у микоплазм (р. Mycoplasma)
ЦПМ – единственное мембранное образование бактерий, определяет ее жизнедеятельность
У бактерий нет мембран ядра, митохондрий, АГ и ЭПС
ЦПМ образована двумя слоями фосфолипидов (ФЛ) с комплексами белков
ФЛ есть во внешнем и внутреннем листке ЦПМ, холестерины отсутствуют
Белки ЦПМ
В состав мембран входят белки и белковые комплексы
Белки интегральные могут несколько раз пронизывать мембрану
Белки гидрофобные – внутри мембран
Белки гидрофильные – снаружи на поверхности мембран
Белки периферические – находятся на мембране, не в цитоплазме – в основном, ферменты.
Жирные кислоты ЦМП бактерий
ЖК состоят из 16-18 атомов углерода
У бактерий, в отличие от эукариот, практически отсутствуют двойные (ненасыщенные) связи в ЖК
Степень насыщенности ЖК определяет свойства бактериальных мембран
У бактерий мембраны должны находиться в переходном подвижном состоянии, чтобы активно реагировать на воздействия извне
Подвижное состояние мембран бактерий обеспечивает широкие температурные границы их существования
Функции ЦПМ
Строение ЦПМ определяет ее функции
ЦПМ — полифункциональная структура, вместилище различных ферментов.
Ферменты участвуют в самых различных процессах жизнедеятельности бактерий.
Все функции мембраны связаны и плавно перетекают друг в друга.
Условно выделим 5 групп функций ЦПМ
Регуляция осмотического давления -главный осмотический барьер.
Энергетическая функция.
Транспортная функция.
Сенсорная функция.
Регуляция деления бактериальной клетки.
Регуляция осмотического давления
1. Неспецифическая простая диффузия по градиенту концентрации без затраты энергии.
2. Облегченная диффузия – за счет фермента – субстрат-специфической пермеазы (транспортный белок) по градиенту концентрации не требует затрат энергии.
3. Активный транспорт — с участием специфических транспортных белков – пермеаз. В отличие от облегченной диффузии требуется затрата энергии.
Энергетическая функция
Система первичной протонной помпы или протондвижущая сила (ПДС) возникает:
1. В результате дыхания.
2. Источником может быть энергия света.
3. ПДС возникает за счет работы белкового комплекса АТФ-азы (включает 7 разных белков).
4. За счет ПДС протоны Н+ поступают внутрь клетки.
ПДС складывается за счет:
Электрического мембранного потенциала
Разности рН между наружной и внутренней сторонами мембраны.
Или тем и другим одновременно.
Процесс идет за счет энергии АТФ.
Другие варианты первичной помпы
Вместо протонов (Н+) могут работать другие ионы, например, K+, Na+.
K+ первичная помпа.
Na+ первичная помпа.
В этих случаях происходит поступление K+, Na+ за счет энергии АТФ.
Например: морские бактерии, термофилы, бактерии в рубце жвачных животных.
Т.о., ПДС может создаваться за счет разных ионов.
Транспортная функция ЦПМ
Бактерии могут существовать только во влажной среде, поглощая растворенные вещества.
Все вещества должны проходить через ЦПМ.
Перенос веществ через ЦПМ
Существует несколько вариантов переноса веществ через ЦПМ:
1. Активный транспорт
2. Вторичная помпа
1. Первый вариант переноса — активный транспорт Участвуют специфические транспортные белки – пермеазы, отличаются друг от друга по ряду показателей: по степени сродства к субстрату, по специфичности к определенным веществам, по эффективности определения концентрации веществ в клетке и вне клетки
2. Второй вариант переноса -вторичная помпа при участии энергетического протонного потенциала – вторичной помпы.
В этом случае специфические белки катализируют перенос различных субстратов за счет ПДС.
Как и в случае первичной помпы это перенос, но различных веществ (не только ионов Н+, K+, Na+) в клетку за счет разности мембранного потенциала, обеспечивающего
1-й вариант вторичной помпы
Унипорт – втягивание вещества отрицательным зарядом за счет разности потенциалов на мембране. Например, электрофоретический вариант переноса вещества.
2-й вариант вторичной помпы
Синпорт – белок катализирует одновременный и однонаправленный перенос веществ (двух или сразу нескольких) вместе с протоном за счет ПДС. Например, Н+ и лактоза.
3-й вариант вторичной помпы
Антипорт – белки вторичной помпы катализируют одновременный и встречный перенос двух различных веществ. Например, Н+ и иона Са+ или Na+.
Сенсорная функция ЦПМ
Бактерии способны улавливать и определять малейшие изменения в окружающей среде
Сенсорные системы бактерий похожи на подобные системы в клетках высших организмов.
studfiles.net
Цитоплазма бактерий
Количество просмотров публикации Цитоплазма бактерий — 320
Функция цитоплазматической мембраны
Строение цитоплазматической мембраны бактерий
Безоболочечные формы бактерий
Оболочка Грам(-) бактерий
Оболочка Грам(+) бактерий
Строение пептидогликана бактерий
Свойства клеточной стенки бактерий
Функция клеточной стенки бактерий
-Формообразующая
-Защита от внешних воздействий
-Защита от внутреннего давления клетки
-Обеспечение диффузии питательных веществ
-Ригидность
-Растяжимость
-Эластичность
-Полупроницаемость
-Многослойность
-Волокнистая структуру
Пептидогликан – (муреин, мукопептид) – параллельно расположенные молекулы гликана, состоящие из остатков N-ацетилгликозамина и N-ацетилмурамовая кислота͵ связанных пептидной связью.
Основу пептидной связи составляют тетрапептиды, отличающиеся у Гр(+) и (-) бактерий.
Тетрапептиды Гр(-) бактерий: L-аланин, D-глютаминовая кислота͵ L-мезодиаминопимелиновая кислота͵ D-аланин
Тетрапептиды Гр(+) бактерий: L-аланин, D-глютаминовая кислота͵ L-диаминопимелиновая кислота͵ D-аланин.
- Пептидогликан – 40-90%
- Тейхоловые кислоты, ковалентно связанные с пептидогликанами
- Трехслойная наружная оболочка(похожая на ЦПМ), связанная с пептидогликаном
- Пептидогликан
- Цитоплазмаическая мембрана
Оболочка представляет собой эндотоксин бактерий.
— Протопласты – бактерии, полностью лишенные клеточной стенки, нежизнеспособны
— Сферопласты – бактерии, частично лишенные клеточной стенки, нежизнеспособные
— L-формы – утратившие клеточную стенку под влиянием антибиотиков. Маловирулентны, способны к размножению, длительной персистенции(долгое выживание в организме) в организме человека и к реверсии в исходные высоковирулентное состояние. Вызывают вялотекущие хронические инфекции.
— Микоплазмы – мелкие полиморфные Грам(-) бактерии
3х-слойное (2 слоя липидов, один слой белков) незамкнутые постоянные и временные(мезозомы) структуры в виде сложно закрученных структур.
Мезозома вызывает репликацию ДНК, она действует на нуклеоид, а с другой стороны она тесно прилегает к поперечной перегородке клетки – ᴛ.ᴇ. синтез перегородки клетки при размножении.
- Транспортная(селективная проницаемость и транспорт веществ в бактерию)
- Энергетическая(перенос электронов и окислительные реакции у анаэробов) – аналог митохондрий
- Пластическая(биосинтез белка, биополимер оболочки, цитоплазматической мембраны, цитоплазмы)
- Информационная(биосинтез ДНК)
Коллоидная система, содержащая аналог ядра – нуклеоид, РНК(16S рРНК, 23 рРНК), ДНК, другие биополимеры, рибосомы, включения органических(крахмал, Гликоген) и минеральных веществ(сера, карбонаты, фосфаты, волютин-по методу Нейссера), вакуоли.
referatwork.ru
Цитоплазма — Микробиология и биотехнологии
Цитоплазма –содержимое клетки, за исключением ядра. Цитоплазма имеет сложный изменяющийся химический состав. Основными химическими соединениями являются белки, нуклеиновые кислоты, липиды, Н2О. В цитоплазме содержатся рибосомы, мезосомы, включения (липиды, углеводы, сера и др.)
Поверхностный слой цитоплазмы более плотный, обладает полупроницаемостью — цитоплазмотическая мембрана. Выполняет важную роль в обмене веществ между клеткой и окружающей средой. Цитоплазмотическая мембрана состоит из трех слоев: одного липидного и двух, примыкающих к нему с обеих сторон, белковых. Содержит 60-65% белка и 35-40% липидов, локализовано ферменты. Цитоплазма пегомогенна. Помимо бесструктурной полужидкой массы колоидного состояния, она местами пронизана мембранами, в которой находятся различные по форме и величине частички — рибосомы. Рибосомы состоят на 60% из РНК и на 40% из белка. В одной бак. клетке содержится десятки тысяч рибосом. В рибосоме осуществляется
синтез белка. Кроме рибосом в цитоплазме имеются пластинчатые структуры – мезосомы, они образуются путем ответвления и впячивания в полость клетки цитоплазмотической мембраны. В мезосомах происходят процессы окисления органических веществ. Помимо этих образований в цитоплазме также содержатся разнообразные включения, являющиеся запасными питательными веществами: крупинки гликогена, капли жира, гранулы волютина.
Жгутики – это органы движения бактерий. Представляют собой вращающиеся полужесткие спирально изогнутые нити из белка флагеллина, который обладает способностью сокращаться. Длина жгутиков больше самих бактерий и колеблется от 5 до 10 мкм в длину. По типу расположения и числу жгутиков бактерии делят на четыре группы: монотрихи- имеют один жгутик на полюсе клетки; лофотрихи — с пучком жгутиков на одном из концов палочки; амфитрихи — с двумя пучками жгутиков на полюсах; перитрихи — с моножеством жгутиков вокруг бактерии. Жгутикование характерно, например, для кишечных бактерий, столбняка и ботулизма, холерного вибриона. Характер и скорость движения неодинаковы у отдельных видов бактерий. Подвижность бактерий может быть утрачена под влиянием неблагоприятных условий жизни, при старении клеток и при механических воздействий.
mikrobiki.ru
