
Закон гомологических рядов наследственной изменчивости Н. И. Вавилова
Николай Иванович Вавилов имел научные интересы в широких сферах, но более всего он известен как ботаник-селекционер и генетик. Исследуя параллелизм в наследственной изменчивости, в 1920 году он открыл и обосновал закон гомологических рядов, который звучит так:
«Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости.
Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейства».
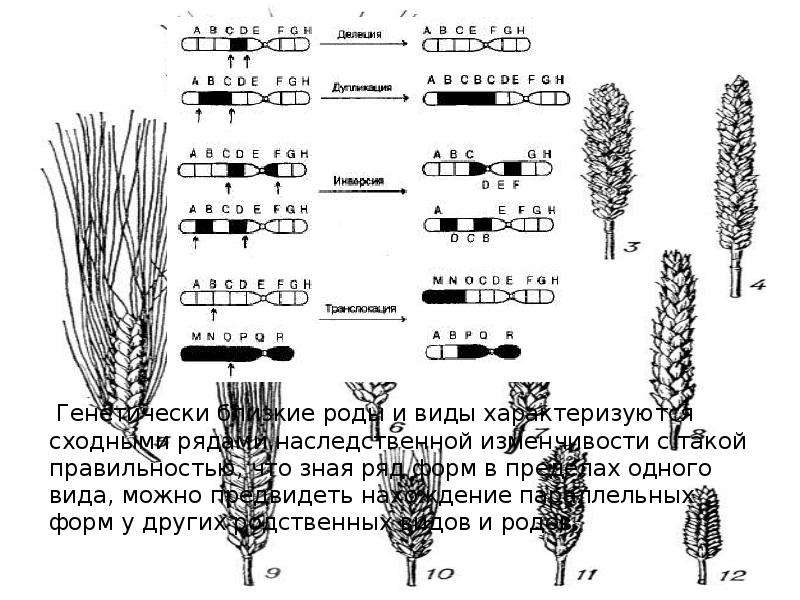
Для наглядной иллюстрации действия закона удобно взять растения семейства мятликовых. Например, зерновка черной окраски имеется у пшеницы, ячменя, ржи, кукурузы и ряда других представителей семейства.
С помощью этого закона Н. И. Вавилов обнаружил ряд форм ржи, еще не известных в то время: с остистыми и безостными колосьями, с черной, белой, фиолетовой и красной зерновкой, с зерном мучнистым и стекловидным и проч. Исследователь опирался на наличие данных признаков у пшеницы.
Знак «+» указывает на наличие наследственных форм, которые обладают названным признаком.
Николай Вавилов не зря был назван «Менделеевым от биологии». Он выявил базовые закономерности в биологии, сходные с теми строгими закономерностями, которые Менделеев обнаружил для химических элементов. Закон гомологических рядов наследственной изменчивости можно применять как для растений, так и для животных.
1. Например, короткопалость встречается и у человека, и у птиц, собак, крупного рогатого скота.
2. Альбинизм бывает у представителей разных групп млекопитающих, а также у птиц и прочих животных.
Альбинизм бывает у представителей разных групп млекопитающих, а также у птиц и прочих животных.
3. Перья могут отсутствовать у птиц, чешуя у рыб, волосистый покров у млекопитающих, и всё это производные кератина.
Закон, открытый Вавиловым, очень важен в селекции. Он дает возможность предугадать появление форм, доселе не обнаруженных у конкретного вида, однако имеющихся у близкородственных видов. Данную форму можно «вдруг» обнаружить в дикой природе, но можно и получить с помощью искусственного мутагенеза.
Закон гомологических рядов Н.И. Вавилова: VIKENT.RU
Закон гомологических рядов Н.
 И. Вавилова
И. Вавилова В 1920 году Н.И. Вавилов излагает основные идеи Закона гомологических рядов в докладе на III-м Всероссийском селекционном съезде в Саратове. Главная идея: родственные виды растений имеют схожие спектры изменчивости (часто это фиксированное число строго определённых вариаций).
«И Вавилов проделал такую штуку. Он у наилучше изученных, как я уже сказал, растений из числа культурных злаков собрал все известные наследственные признаки, расположил их в определённом порядке в таблицах и сравнил все известные в то время ему подвиды, формы и сорта. Таблиц было составлено много, конечно, материал огромный был. Он тогда же, ещё в Саратове, пристегнул к злакам и бобовые — различные горохи, вики, бобы, фасоли и т.д. — и ещё какие-то культурные растения. И оказалась в очень многих случаях параллельность у очень многих видов. Конечно, у каждого семейства, рода, вида растений все признаки имели свои особенности, свою форму, свой способ выражения. Например, цвет семян от почти белого до почти чёрного варьировал почти у всех культурных растений. Значит, ежели у лучше изученных злаков с огромным количеством уже известных, изученных сортов и форм описано несколько сот различных признаков, а у других, менее изученных или диких родственников культурных видов многих признаков нет, то их можно, так сказать, предсказать. Они будут-таки найдены на соответствующем большом материале.
Например, цвет семян от почти белого до почти чёрного варьировал почти у всех культурных растений. Значит, ежели у лучше изученных злаков с огромным количеством уже известных, изученных сортов и форм описано несколько сот различных признаков, а у других, менее изученных или диких родственников культурных видов многих признаков нет, то их можно, так сказать, предсказать. Они будут-таки найдены на соответствующем большом материале.
Вавилов показал, что в общем и целом наследственная изменчивость всех растений в очень сильной степени варьирует параллельно. Он назвал это гомологическими рядами изменчивости растений. И указал, что чем ближе виды друг к другу, тем больше эта гомология рядов изменчивости признаков. Целый ряд разных общих закономерностей был выявлен на этих гомологических рядах наследственной изменчивости растений. И это обстоятельство было взято Вавиловым в качестве одной из важнейших основ дальнейшей селекции и поисков хозяйственно полезных признаков у вводимых в культуру растений. Изучение гомологических рядов наследственной изменчивости прежде всего у культурных растений, затем у домашних животных является теперь уже само собой разумеющимся, одной из основ дальнейшей селекции нужных человеку сортов тех или иных видов изучаемых растений. Это было, может быть, одно из первых крупных достижений Вавилова мирового масштаба, которое очень быстро и создало ему мировое имя. Имя ежели не первого и лучшего, то одного из первых и лучших прикладных ботаников в мире.
Изучение гомологических рядов наследственной изменчивости прежде всего у культурных растений, затем у домашних животных является теперь уже само собой разумеющимся, одной из основ дальнейшей селекции нужных человеку сортов тех или иных видов изучаемых растений. Это было, может быть, одно из первых крупных достижений Вавилова мирового масштаба, которое очень быстро и создало ему мировое имя. Имя ежели не первого и лучшего, то одного из первых и лучших прикладных ботаников в мире.
Параллельно с этим Вавилов совершил по всему миру — по всей Европе, большей части Азии, по значительной части Африки, по Северной, Центральной и Южной Америке — большое количество экспедиций со сбором огромного материала, в основном по культурным растениям. В 20-м году, по-моему, Вавилов был сделан директором Бюро по прикладной ботанике и новым культурам. Это Бюро было несколько изменено и превращено в Институт по прикладной ботанике и новым культурам, потом в Институт прикладной ботаники, генетики и селекции растений. А к концу 30-х годов он стал уже Всесоюзным институтом растениеводства. Это название и до сих пор сохранилось, хотя мировой удельный вес его после гибели Вавилова, конечно, сильно упал. Но всё-таки многие вавиловские традиции и до сих пор поддерживаются, и часть огромной мировой живой коллекции сортов, подвидов и форм культурных растений буквально из всех групп культивируемых на земном шаре растений сохраняется в Пушкине, бывшем Детском Селе, бывшем Царском Селе. Это живой музей, каждый год пересевающийся заново, созданный Вавиловым. То же и на бесчисленных опытных станциях, разбросанных по всему Советскому Союзу.
Во время своих многочисленных поездок Вавилов опять-таки умудрился не потонуть в огромном материале, в данном случае уже географическом многообразии форм различных видов культурных растений. Он наносил всё на карты большого масштаба разноцветными карандашами, сперва играя, как малые дети, в географические карты, а потом все это переводя в сравнительно простые небольшие карты с чёрными значками различного типа для разных форм культурных растений. Так он обнаружил в мире, на земном шаре, в биосфере нашей планеты, несколько центров многообразия культурных растений. И показал, просто на картах, расползание, распространение на Земле не только отдельных видов, но определённых групп видов, окультуренных, по-видимому, впервые в определенном месте, ну, скажем, в Северном или Среднем Китае или в горной части Северной Африки, или, скажем, в районе Перу, в Южной Америке, в горах, в Андах. Оттуда обыкновенно не один вид каких-нибудь культурных растений, а группа хозяйственно друг с другом связанных видов, возникших как культурные растения и прижившихся как культурные растения в определённом месте, расползались по Земле. Некоторые недалеко, на небольшое расстояние, а другие завоевали полмира, как говорится, вроде той же пшеницы или гороха.
Так он обнаружил в мире, на земном шаре, в биосфере нашей планеты, несколько центров многообразия культурных растений. И показал, просто на картах, расползание, распространение на Земле не только отдельных видов, но определённых групп видов, окультуренных, по-видимому, впервые в определенном месте, ну, скажем, в Северном или Среднем Китае или в горной части Северной Африки, или, скажем, в районе Перу, в Южной Америке, в горах, в Андах. Оттуда обыкновенно не один вид каких-нибудь культурных растений, а группа хозяйственно друг с другом связанных видов, возникших как культурные растения и прижившихся как культурные растения в определённом месте, расползались по Земле. Некоторые недалеко, на небольшое расстояние, а другие завоевали полмира, как говорится, вроде той же пшеницы или гороха.
Вавилов, таким образом, установил центры многообразия и происхождения различных форм культурных растений в разных местах земного шара. И создал целую теорию происхождения культурных растений в различные эпохи древнейшего и древнего мира. Это было вторым большим достижением Вавилова, опять-таки мировым. Сейчас невозможно дальнейшее развитие истории мирового земледелия и истории очагов происхождения культурных растений без созданного Вавиловым фундамента. Существуют попытки, так сказать, некоторой реформы и видоизменения вавиловских воззрений, но можно сказать, что это частности по сравнению с общей мировой картиной, созданной Вавиловым.
Это было вторым большим достижением Вавилова, опять-таки мировым. Сейчас невозможно дальнейшее развитие истории мирового земледелия и истории очагов происхождения культурных растений без созданного Вавиловым фундамента. Существуют попытки, так сказать, некоторой реформы и видоизменения вавиловских воззрений, но можно сказать, что это частности по сравнению с общей мировой картиной, созданной Вавиловым.
Значит, я перечислил уже три огромных достижения: иммунитет растений, закон гомологических рядов и теория центров земледелия и возникновения различных форм культурных растений. Пожалуй, последнее, что хочется назвать из общих достижений Вавилова, это большое количество его работ и усилий, главным образом усилий, уже в смысле пропаганды на различных конгрессах, международных и всесоюзных, написание статей научно-популярных по проблеме продвижения земледелия на север в первую очередь и в области, занятые пустынями и пустошами, соединенное с охраной природы в совершенно современном и даже предназначенном для ближайшего будущего смысле: продвижение культуры вместе с разумным отношением к сообществам живых организмов биосферы. Вот в этих направлениях Вавилов является совершенно исключительным, я бы сказал, исключительно крупным ученым в мировом масштабе».
Вот в этих направлениях Вавилов является совершенно исключительным, я бы сказал, исключительно крупным ученым в мировом масштабе».
Тимофеев-Ресовский Н.В., Воспоминания, М., «Вагриус», 2008 г., с. 304-306.
Эта работа Н.И. Вавилова была посвящена Альфонсу Декандолю.
К столетию Закона гомологических рядов в наследственной изменчивости
В этом году исполняется сто лет доклада Николая Ивановича Вавилова о «Законе гомологических рядов в наследственной изменчивости». Этот доклад был озвучен Николаем Ивановичем на Третьем Всероссийском селекционном съезде в г. Саратове 4 июня 1920 г.
Закон гомологических рядов в наследственной изменчивости был напечатан в виде отдельной брошюры, изданной по случаю доклада1. Развернутая версия концепции закона опубликована в 1922 г. на английском языке в ведущем международном генетическом журнале.2
Это издание в обратном переводе на русский язык стало основой для последующих переизданий труда Н. И. Вавилова. Ссылки и тексты трех первых публикаций сведены в отдельном томе работ юбилейного пятитомного издания, выпущенного издательством Наука к 100-летию Н.И. Вавилова.
3
И. Вавилова. Ссылки и тексты трех первых публикаций сведены в отдельном томе работ юбилейного пятитомного издания, выпущенного издательством Наука к 100-летию Н.И. Вавилова.
3
Наш институт имеет исторические связи с периодом активной деятельности и наследием академика Н.И Вавилова. Мы изначально размещались в одном здании с группой академических институтов биолого-химического профиля, в том числе с Институтом генетики, руководимым Н.И. Вавиловым до злосчастного ареста в 1940 г., репрессий и гибели ученого в Саратовской тюрьме. Оттиск с дарственной надписью Вавилова первому директору нашего Института академику А.Н. Северцову хранится в академической библиотеке в здании на Ленинском проспекте, 33.
При следующем директоре академике И.И. Шмальгаузене в 1940 г. был напечатан труд зав. сектором эволюции животных профессора С.Н. Боголюбского4, имевший прямое отношение к Закону гомологических рядов Н.И. Вавилова и позже переиздававшийся.
Современным проблемам закона гомологических рядов были посвящены части книги молекулярного биолога Б.
Столетие самого Н.И. Вавилова, праздновавшееся в мире под эгидой ЮНЕСКО, дало повод директору ИЭМЭЖ, преобразованного при нем в ИПЭЭ РАН, академику В.Е. Соколову выразить восхищение гением Вавилова в вводной статье к юбилейному выпуску журнала «Природа» в 1987 г. Подобное высокое признание ученого ученым стоит знать и хранить в анналах институтской истории и не забывать.6
Прочитать статью в журнале «Природа» можно по ссылке.
1 Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. Доклад на III Всероссийском селекционном съезде в г. Саратове 4 июня 1920 г. Саратов, 1920, 16 с.
2 Vavilov N.I. The law of homologous series in variation. Journal of Genetics. 1922. V. 12(1): 47 – 89.
3 Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости.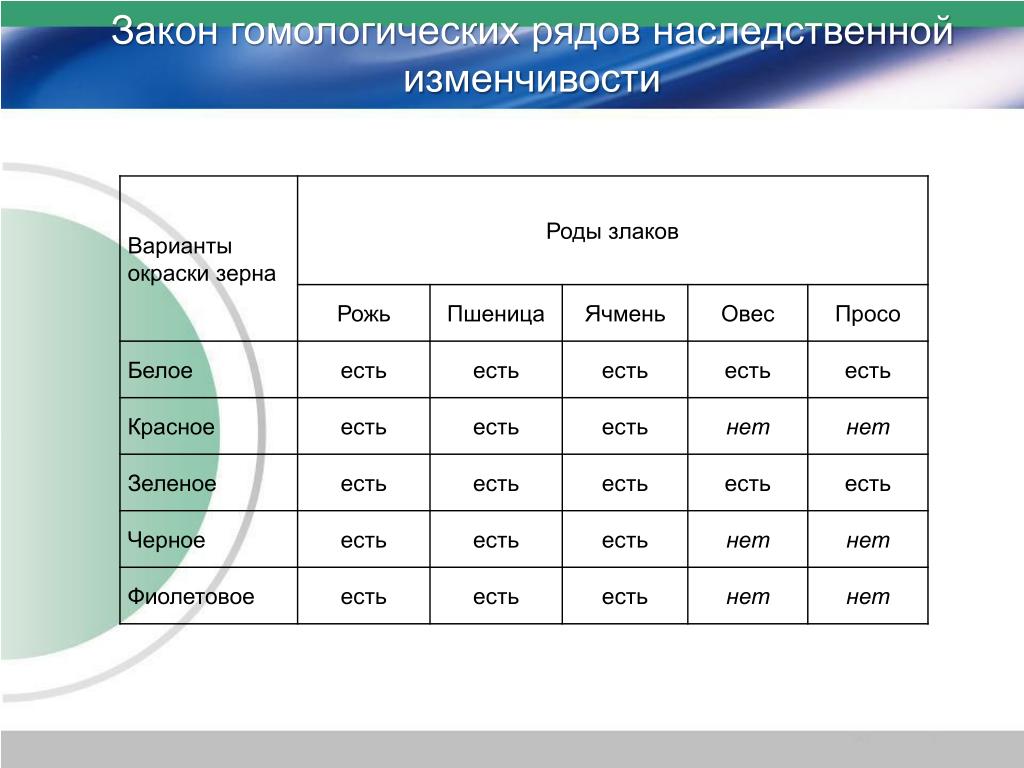 — Л.: Наука, 1987. — 256 с.
— Л.: Наука, 1987. — 256 с.
4 Боголюбский С.Н. Происхождение и эволюция домашних животных. М.: Сельхозиздат, 1940. 168 с.— Боголюбский С.Н. Происхождение и преобразование домашних животных. М.: Сов. Наука, 1959. 593 с.
5 Медников Б.М. Избранные труды. Организм, геном, язык. М.: Т-во научных изданий КМК. 2005. 452 с.
6 Соколов В.Е. Великий современник. Природа. 1987. № 10. С. 4-5.
Закон гомологических рядов Н.И.Вавилова и его значение в биологии
Рукопись, датированная 20 февраля 1957 г.
Работа Н.И.Вавилова «Закон гомологических рядов в наследственной изменчивости» была опубликована в 1920 г. на русском языке в «Трудах 3-го съезда по растениеводству» и в 1922 г. – на английском (Vavilov, 1922). Как известно, установление этого закона сопровождалось открытием предсказанной Н.И. Вавиловым на основе этого закона безлигульной формы ржи на Памире. Это открытие некоторыми лицами сравнивалось с открытием новых элементов на основе периодической системы Д.
Сам Н.И.Вавилов, будучи образованным и добросовестным биологом, нисколько не претендовал на то, что он первый сформулировал положение о гомологической изменчивости. Он указывал многих авторов, в частности самого Ч. Дарвина, в свою очередь называвшего Уолша (В.D. Walsh), установившего для насекомых в естественном состоянии закон «равнозначащей изменчивости» (law of equable variability). Ч. Дарвин (1952, с. 192) так формулирует это положение. «Виды, между собой различные, представляют аналогичные изменения, так что разновидность известного вида часто приобретает особенности, свойственные сродному виду, или возвращается к признакам более раннего предка»92.
Из этого определения, как и из последующего текста, ясно:
- это положение, по мысли Дарвина, не претендовало на звание строгого закона, так как в нем не было обязательности и не были указаны условия преимущественности;
- оно распространялось только на виды одного рода; как толковали это положение предшественники Дарвина по этому вопросу-Уолш, Ноден и другие, остается неясным;
- аналогичными признаками в собственном смысле слова он считает те, которые возникают самостоятельно у близких видов, что он считает естественным в силу происхождения всех видов одного рода от общего предка: от них он отличает возвратные признаки-восстановление утраченных.
Правда, он оговаривается, что так как нам неизвестны общие предки наших естественных групп, то мы не в состоянии отличить признаки аналогичные от признаков возвратных. Кроме того, Дарвин считает (с. 194), что «признаки, происходящие исключительно в силу аналогичных изменений, будут, по всей вероятности, несущественными, так как сохранение всех признаков, важных по отношению к отправлениям организма, будет ограждаться естественным отбором соответственно различному образу жизни видов».
Эта последняя оговорка очень важна: Дарвин, как честный ученый, зарегистрировал этот закон аналогичной изменчивости. но от него не укрылась враждебность этого закона его пониманию эволюции: дивергенции под руководством естественного отбора. Ведь возникновение аналогичных изменений во многих или во всех видах одного рода – это параллельная, а не дивергентная эволюция. Ясно, что если бы закон аналогичной изменчивости имел универсальное значение, то крайне ценимое самим Дарвином положение о господстве дивергенции в эволюции получило бы сильнейший удар.
Большое изумление вызывает у Дарвина и факт проявления признаков, исчезнувших за много поколений, так как Дарвин еще придерживался взгляда на наследственность, как на нечто, связанное с «кровью». Он пишет (с. 193): «После 12 поколений доля крови, если позволительно будет употребить это ходячее выражение, одного потомка будет 1/2048, и тем не менее, по общераспространенному мнению, этот остаток чужой крови выражается стремлением к возврату».
Ясно, что сам Дарвин и его ближайшие последователи потому не придавали большого значения этому закону, что он не гармонировал как с общей системой дарвиновских взглядов на эволюцию, так и с господствовавшими тогда взглядами на наследственность.
Однако факты систематики организмов постепенно приводили ко все большему числу высказываний, возрождавших забытые многими слова Дарвина и его предшественников по данному вопросу. При этом ясно было, что сформулированное Дарвином положение вовсе не касается только видов одного рода, а распространяется и на более высокие таксономические категории и что оно имеет гораздо большие претензии на закон, чем думал Дарвин. Некоторые авторы отмечали сходство системы организмов (конечно, в относительно узких рамках) со справедливо прошумевшей периодической системой химических элементов Д.И. Менделеева, и наш талантливый зоолог В.М. Шимкевич (Schimkewitsch, 1906) так озаглавил одну из своих работ: «О периодичности в системе пантопод», расположив роды этих животных в таблицу, внешне напоминающую периодическую систему. На самом деле сходство не так велико, и систему, предложенную Шимкевичем для пантопод, правильнее назвать решетчатой, а не периодической. Полезно при этом вспомнить, что еще Линней указывал, что в системе организмов мы имеем решетчатое расположение элементов. Сам В.М. Шимкевич, будучи убежденным дарвинистом, не придавал сколько-нибудь существенного значения обнаруженной им закономерности.
На самом деле сходство не так велико, и систему, предложенную Шимкевичем для пантопод, правильнее назвать решетчатой, а не периодической. Полезно при этом вспомнить, что еще Линней указывал, что в системе организмов мы имеем решетчатое расположение элементов. Сам В.М. Шимкевич, будучи убежденным дарвинистом, не придавал сколько-нибудь существенного значения обнаруженной им закономерности.
Всякому, много занимающемуся определением любых организмов, не может не броситься в глаза неудовлетворительность определительных таблиц. Со времен Ламарка они все строятся в форме дихотомических таблиц, т. е. как бы в виде деревцев с большим числом разветвлений. Однако форма наших определительных таблиц отнюдь не является наиболее экономной, из-за этого возникает много ошибок в определении. Именно определяющий, сделав ошибку в выборе антитез, дальше получает как бы подтверждение тому, что он стоит на правильном пути, так как в дальнейшем встречает такие противопоставления, которые соответствуют признакам определяемого им объекта.
Н.И.Вавилову и принадлежит поэтому бесспорная заслуга, что он не просто извлек из забвения старое положение Дарвина, но сделал крупный шаг вперед по пути познания одной из глубоких закономерностей, лежащих в основе формообразования организмов. По сравнению с Дарвином понимание Вавиловым характеризуется следующими особенностями.
- Закон приложим не только к близким видам одного рода, но и к родам семейств, не только близких, но и отдаленных.
- Этому закону придается такая универсальность, что по наличию ряда изменчивости в одном роде или в семействе можно предвидеть соответствующий ряд в другом роде или семействе: это и позволило Н.И. Вавилову сделать удачный прогноз.
- Н.И. Вавилов и его сотрудники не задумались сделать дальнейшие выводы из возрожденного на повышенном основании закона: они указали, что многие случаи миметизма прекрасно истолковываются как проявления этого закона. В одной из работ имеется прекрасная цветная таблица, показывающая великолепные случаи «миметизма» в семействе бобовых.
 С полным знанием дела использует закон Вавилова выдающийся знаток проблемы миметизма, талантливейший критик банального дарвинистического толкования этой проблемы австрийский ученый Ф. Гейкертингер (Heikertinger, 1954).
С полным знанием дела использует закон Вавилова выдающийся знаток проблемы миметизма, талантливейший критик банального дарвинистического толкования этой проблемы австрийский ученый Ф. Гейкертингер (Heikertinger, 1954).
В этой превосходной посмертной сводке Гейкертингер прекрасно показывает, что огромное число случаев миметизма только потому фигурирует как таковое, что оба партнера занимают тот же ареал. Но огромное число случаев «псевдомиметизма» просто не регистрируется, хотя можно привести большое число примеров, где представители одного семейства «подражают» представителям многих других семейств. Гейкертингер приводит в своей книге (с. 153) список видов семейства чернотелок (Tenebrionidae), которые «подражают» видам семейств – Carabidae, Dytiscidae, Elateridae, Silphidae, Chrysomelidae (подсемейства Chrysomelinae, Cassidinae), Cerambycidae, Curculionidae, Scarabeidae, Passalidae, Lucanidae, Languridae.
Могу привести из личного опыта, что при просмотре богатых коллекций Зоологического института АН СССР по подсемейству земляных блошек (Halticinae) я наткнулся на мелких жучков, которых я без всякого сомнения отнес к жужелицам подсемейства Bembidiini и потому передал специалисту по жужелицам О. Л. Крыжановскому, который их тоже принял без возражения, но вернул их на следующий день, так как при очень внимательном рассмотрении они оказались все-таки земляными блошками, а не жужелицами. Но среди земляных блошек (наибольшее разнообразие, конечно, дает Южная Америка) есть виды, поразительно напоминающие не только представителей соседнего подсемейства Galerucinae, но и жуков семейств Curculionidae, Cerambycidae, Helodidae (некоторые виды этого подсемейства были описаны как блошки).
Л. Крыжановскому, который их тоже принял без возражения, но вернул их на следующий день, так как при очень внимательном рассмотрении они оказались все-таки земляными блошками, а не жужелицами. Но среди земляных блошек (наибольшее разнообразие, конечно, дает Южная Америка) есть виды, поразительно напоминающие не только представителей соседнего подсемейства Galerucinae, но и жуков семейств Curculionidae, Cerambycidae, Helodidae (некоторые виды этого подсемейства были описаны как блошки).
По-видимому, эти случаи псевдомиметизма совершенно не зарегистрированы, так как о них только вскользь упоминает Гейкертингер (1917, с. 122), который, как известно, был не только знатоком литературы по миметизму, но и знатоком систематики земляных блошек.
То возражение, которое часто приводится защитниками дарвинистического толкования миметизма, что слишком часто миметические виды водятся вместе, парируется наличием географических закономерностей в изменчивости организмов: в этой области Вавилов сделал весьма существенный вклад в науку. В его работе «Ботанико-географические основы селекции» (1935) есть специальный раздел «Правильности географического распределения сортового разнообразия культурных растений». Приведу только два примера. В Китае аккумулируется мировое разнообразие голозерного ячменя, голозерного проса, крупно-зернового голого овса. Только на Памире найдены безлигульные формы ржи, мягкой пшеницы и карликовой пшеницы (твердой пшеницы – на Кипре).
В его работе «Ботанико-географические основы селекции» (1935) есть специальный раздел «Правильности географического распределения сортового разнообразия культурных растений». Приведу только два примера. В Китае аккумулируется мировое разнообразие голозерного ячменя, голозерного проса, крупно-зернового голого овса. Только на Памире найдены безлигульные формы ржи, мягкой пшеницы и карликовой пшеницы (твердой пшеницы – на Кипре).
В общем можно сказать, что закон гомологических рядов открывал широкие перспективы по изучению закономерностей, лежащих в основе эволюции. В этом же направлении двигалась и мысль нашего выдающегося ученого Л.С. Берга (1922), палеонтолога Д.Н. Соболева (1924) и гистолога А.А. Заварзина (1923). Несмотря на наличие ряда ошибок у всех этих авторов, сопутствующих каждому новому течению в науке, и разнообразие подходов к биологическим проблемам, общим выводом всех их было то, что морфологические закономерности, существование которых вынужден был допускать и Ч. Дарвин, играют в эволюции органического мира несравненно большую роль, чем это принимает ортодоксальный дарвинизм.
Чем же объяснить то, что это направление в науке сейчас, можно сказать, еле теплится (даже в мировой литературе)? Не касаясь чисто внешних причин, могу указать следующие.
- Сам Вавилов вскоре после формулировки своего закона оказался во главе прикладной ботаники. Здесь он проявил в полной мере свои блестящие организаторские способности и вложил всю свою большую душу в это дело. Только сейчас мы можем полностью оценить все величие его работы и как велика потеря вследствие его безвременной кончины. Но разнообразная административная работа, многочисленные экспедиции и прикладное направление его главных работ в этот период не могли не сказаться на снижении деятельности в том теоретическом секторе работы, который не сулил непосредственного практического приложения.
- За период двадцатых и последующих годов огромное количество ученых было отвлечено в область генетики. Результаты в этой области достигнуты блестящие, но они также отвлекли внимание биологов от тех теоретических проблем, которые недоступны в настоящее время экспериментальной проверке и не отличаются той точностью, которой достигла генетика.
 Как крайнее выражение этой переоценки экспериментального метода, можно вспомнить слова В. Бэтсона, что палеонтология вообще некомпетентна в вопросах о факторах эволюции, так как в палеонтологии эксперимент отсутствует.
Как крайнее выражение этой переоценки экспериментального метода, можно вспомнить слова В. Бэтсона, что палеонтология вообще некомпетентна в вопросах о факторах эволюции, так как в палеонтологии эксперимент отсутствует. - Видимо, для многих генетиков «периодичность» в систематике объяснялась гибридизацией, и, вероятно, многие из сторонников Н.И. Вавилова считали, что закон гомологических рядов исчерпывается той «решеткой», которая получается в результате скрещивания. Введен даже термин «ретикулатная» (т. е. сетчатая) эволюция. Однако при этом упускается из виду, что, как уже указывал Вавилов, сетчатость проявляется и на таком таксономическом уровне, на котором, по крайней мере по имеющимся данным, гибридизация невозможна. Еще важнее, пожалуй, то, что параллелизм распространяется и на, так сказать, «идейные» признаки, которые никак не могут быть выведены из единой морфологической или генетической основы. Чтобы дать понять, что это значит, приведу из обширнейшего запаса фактов этого рода только один пример: стридуляционные органы и органы слуха кузнечиков и саранчевых.

- Несомненно, что закон гомологических рядов является только началом выяснения номогенетического компонента эволюции. Это показывает, что здесь, видимо, не простые пробелы в наших знаниях, а действительное несоответствие закона с реальным миром. Крайнее разнообразие организмов, продолжающееся новоописание, отсутствие сколько-нибудь удовлетворительных сводок по большинству групп отвлекают внимание систематиков от теоретической работы. В области теории работы крайне редки. Недостаточно ясно осознается различие между комбинативной и периодической системами (см.: Любищев, 1923).
- Генетика первоначально развивалась, и это было вполне естественно, независимо от дарвинизма. Но вскоре выяснилось, что критическая мысль должна затронуть самые основы биологии и что эта работа по перестройке основ биологии является исключительно сложной и трудной. А при наличии той переоценки экспериментального метода, о которой уже говорилось, проявилась тенденция к «примирению» с дарвинизмом у тех ученых, как, например, Иоганнсен, которые первоначально резонно критиковали дарвинизм.
 У сторонников же точного мышления теоретическая мысль пошла по разработке методов приложения математики, к нейтральным областям систематики и теории эволюции. Я имею в виду работы Вольтерра и др. по математической теории борьбы за существование, Р. Фишера, Д. Холдена, С. Райта и других по математической теории естественного отбора и, наконец, что имеет особенное значение для систематики, по разработке методики комплексных признаков или дискриминантных функций, связанной опять-таки с именем Р. Фишера. Это последнее направление имеет исключительно важные методические и практические перспективы.
У сторонников же точного мышления теоретическая мысль пошла по разработке методов приложения математики, к нейтральным областям систематики и теории эволюции. Я имею в виду работы Вольтерра и др. по математической теории борьбы за существование, Р. Фишера, Д. Холдена, С. Райта и других по математической теории естественного отбора и, наконец, что имеет особенное значение для систематики, по разработке методики комплексных признаков или дискриминантных функций, связанной опять-таки с именем Р. Фишера. Это последнее направление имеет исключительно важные методические и практические перспективы. - Закон гомологических рядов оказался, таким образом, лишенным разработанной общебиологической основы, он не имел «объяснений». Как уже указано, это возражение формально совершенно справедливо, и закон гомологических рядов для сколько-нибудь полного понимания требует весьма радикального пересмотра наших общебиологических воззрений. Но, беря в качестве методологического руководства историю точных наук, мы видим, что требование «объяснения» часто служило тормозом на пути самых блестящих научных достижений.
 Великому Ньютону говорили, что его принцип всемирного тяготения непонятен. Он отвечал примерно так: «Несомненно, нелепо считать, что тело может действовать там, где его нет, но все происходит так, как если бы такое действие имело место…».
Великому Ньютону говорили, что его принцип всемирного тяготения непонятен. Он отвечал примерно так: «Несомненно, нелепо считать, что тело может действовать там, где его нет, но все происходит так, как если бы такое действие имело место…».
После Эйнштейна теория тяготения сделала большой шаг вперед (общую теорию относительности часто называют теорией тяготения), но, насколько мне известно, и сейчас там далеко не все ясно. Нечто подобное случилось и со знаменитыми уравнениями электромагнитной теории света К. Максвелла. Наконец, беря наиболее близкий пример, укажу, что в момент своего появления периодическая система элементов Д. И. Менделеева была совершенно «непонятной». Известно, что когда один из предшественников Д. И. Менделеева, Ньюлендс, делал доклад о первой несовершенной попытке найти связь свойств элементов с атомным весом в научном химическом обществе, то председатель, известный химик, сделал такое «убийственное» возражение: «А вы не пробовали находить связь свойств элементов с алфавитным расположением их названий?». Сейчас мы знаем, что под периодическую систему подведено весьма прочное основание, но отсутствие такого основания не дает нам права отвергать теорию из-за ее «непонятности».
Сейчас мы знаем, что под периодическую систему подведено весьма прочное основание, но отсутствие такого основания не дает нам права отвергать теорию из-за ее «непонятности».
Мы можем подвести итог. Работа Н. И. Вавилова по закону гомологических рядов представляет собой очень крупный шаг по пути проникновения в закономерности систематики и эволюции. Однако сравнение в смысле высоты научного достижения с периодической системой Менделеева было бы неправильно. Система Менделеева представляет уже весьма совершенное решение проблемы систематизации химических элементов, хотя, конечно, далеко не идеальное. Попытка же Вавилова, как ни почтенна она сама по себе, представляет собой только маленький отрезок грандиозной проблемы биологической системы. Здесь совершенная несоизмеримость задач. Биологическая систематика в своем полном здании неизмеримо труднее химической как по подавляющему количественному и качественному многообразию форм, так и по осложнению проблемы системы проблемой органической целесообразности, отсутствующей в химической систематике. Мы знаем, что очень большое число умнейших и талантливейших биологов искренне думают, что вся морфология и физиология организмов подчинена проблеме целесообразности. Заслуга Н.И. Вавилова и заключается кроме всего прочего, в том, что он с определенной точки зрения указал на самостоятельность систематики. В этом отношении он оказался выше не только своих противников, но и большинства своих последователей и почитателей, которые часто выдвигают, например, миметизм как одно из бесспорнейших доказательств теории естественного отбора, забывая, что таким высказыванием они обнаруживают или незнакомство, или игнорирование весьма обоснованного мнения Н.И. Вавилова.
Мы знаем, что очень большое число умнейших и талантливейших биологов искренне думают, что вся морфология и физиология организмов подчинена проблеме целесообразности. Заслуга Н.И. Вавилова и заключается кроме всего прочего, в том, что он с определенной точки зрения указал на самостоятельность систематики. В этом отношении он оказался выше не только своих противников, но и большинства своих последователей и почитателей, которые часто выдвигают, например, миметизм как одно из бесспорнейших доказательств теории естественного отбора, забывая, что таким высказыванием они обнаруживают или незнакомство, или игнорирование весьма обоснованного мнения Н.И. Вавилова.
ЛИТЕРАТУРА
Берг Л.С. Номогенез. Пг., 1922, с. I–VIII. 1–306. (Тр. Геогр. ин-та; Т. 1).
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. Саратов, 1920. 16 с.
Вавилов Н.И. Ботанико-географические основы селекции. – В кн.: Теоретические основы селекции. М., 1935, т. 1, с. 17–74.
Дарвин Ч. Происхождение видов. М.: Сельхозгиз, 1952. 483 с.
М.: Сельхозгиз, 1952. 483 с.
Заварзин А.А. Параллелизм структур как основной принцип морфологии. – Изв. Биол. н.-и. ин-та при Перм. ун-те, 1923, т. 2, вып. 4, с. 135–140.
Любищев А.А. О форме естественной системы организмов. – Изв. Биол. н.-и. ин-та при Перм. ун-те, 1923, т. 2, вып. 4, с. 99–100,
Соболев Д.Н. Начала исторической биогенетики. Симферополь: Госиздат Украины, 1923. 203 с.
Heikertinger F. Das Ratsel der Mimikry und seine Losung. Jena: G. Fischer, 1954. 208 S.
Schirnkewitsch W. Uber die Periodizitat in dem System der Pantopoda. – Zool. Anz., 1906, Bd. 30, N 1/2, S. 1–22.
Vavilov N.I. The law of homologous series in variation. – J. Genet., 1922, vol. 12, N 1, p. 47–89.
Вавилов Николай Иванович. Открытие закона гомологических рядов в наследственной изменчивости
Николай
Иванович Вавилов родился 13 (25) ноября 1887 года в Москве в семье купца. В
1906 году Вавилов окончил Московское коммерческое училище, в стенах которого
постигал естественные науки, и поступил в сельскохозяйственный институт, где
изучал селекцию растений. По окончании института Вавилов перебрался в Петербург
и устроился на работу в Бюро прикладной ботаники, а затем отправился на
Полтавщину проходить научную практику. В последующие годы Вавилов в рамках
научной командировки много путешествовал — в 1913 году несколько месяцев провел
в Англии в лаборатории знаменитого биолога У. Бетсона, затем направился в
Париж, потом — в германский город Иена, где посетил лабораторию известного
биолога-эволюциониста Э. Геккеля. Первая мировая война прервала командировку, и
с 1917-го Вавилов постоянно жил в Саратове и преподавал в университете. Вскоре
увидела свет фундаментальная работа ученого «Иммунитет растений к инфекционным
заболеваниям» — и Вавилов вошел в число ведущих биологов мира. В дальнейшем
Николай Иванович объездил Среднее и Нижнее Поволжье и собрал ценный научный
материал. В 1920 году на съезде селекционеров он впервые рассказал об открытии
закона гомологических рядов. Последние двадцать лет жизни Вавилова связаны с
Ленинградом. Здесь он создал получивший всемирную известность Всесоюзный
институт растениеводства.
По окончании института Вавилов перебрался в Петербург
и устроился на работу в Бюро прикладной ботаники, а затем отправился на
Полтавщину проходить научную практику. В последующие годы Вавилов в рамках
научной командировки много путешествовал — в 1913 году несколько месяцев провел
в Англии в лаборатории знаменитого биолога У. Бетсона, затем направился в
Париж, потом — в германский город Иена, где посетил лабораторию известного
биолога-эволюциониста Э. Геккеля. Первая мировая война прервала командировку, и
с 1917-го Вавилов постоянно жил в Саратове и преподавал в университете. Вскоре
увидела свет фундаментальная работа ученого «Иммунитет растений к инфекционным
заболеваниям» — и Вавилов вошел в число ведущих биологов мира. В дальнейшем
Николай Иванович объездил Среднее и Нижнее Поволжье и собрал ценный научный
материал. В 1920 году на съезде селекционеров он впервые рассказал об открытии
закона гомологических рядов. Последние двадцать лет жизни Вавилова связаны с
Ленинградом. Здесь он создал получивший всемирную известность Всесоюзный
институт растениеводства. В 1929-м его избирают академиком, а затем и
президентом Академии сельскохозяйственных наук. Трагические события жизни
Вавилова тесно связаны с широко известной личностью, фамилия которой устойчиво ассоциируется
с ликвидацией советской школы генетиков и репрессиями в отношении многих
ученых, — Трофимом Лысенко. В 1940 году Вавилова арестовали во время одной из
научных экспедиций. Бесконечное следствие по его делу сократило жизнь великого
ученого — Николай Вавилов умер в тюрьме 26 января 1943 года.
В 1929-м его избирают академиком, а затем и
президентом Академии сельскохозяйственных наук. Трагические события жизни
Вавилова тесно связаны с широко известной личностью, фамилия которой устойчиво ассоциируется
с ликвидацией советской школы генетиков и репрессиями в отношении многих
ученых, — Трофимом Лысенко. В 1940 году Вавилова арестовали во время одной из
научных экспедиций. Бесконечное следствие по его делу сократило жизнь великого
ученого — Николай Вавилов умер в тюрьме 26 января 1943 года.
Николай Иванович Вавилов — крупнейший советский биолог, прославившийся прежде всего как автор одного из ключевых законов живой природы — гомологических рядов.
Чтобы
попытаться образно раскрыть его суть, следует обратиться к примеру, который
стал классической иллюстрацией закона гомологических рядов. Если все известные
варианты хорошо изученного вида, например мягкой пшеницы, расположить в ряд, а
под ним поместить другой ряд из вариантов генетически близкого вида, например
твердой пшеницы, то можно легко заметить закономерность в изменениях этих видов. Прорыв и важность открытия заключались в том, что если бы вдруг во втором ряду
не нашлось аналога, то на основании закона можно сделать вывод, что этот аналог
может и должен существовать. Это дало широчайшее поле для деятельности
ученых-селекционеров.
Прорыв и важность открытия заключались в том, что если бы вдруг во втором ряду
не нашлось аналога, то на основании закона можно сделать вывод, что этот аналог
может и должен существовать. Это дало широчайшее поле для деятельности
ученых-селекционеров.
Вавилов
для доказательства найденной им закономерности привел красноречивый пример. У
пшеницы, ржи, кукурузы, овса были найдены формы с листьями, у которых отсутствовал
особый «язычок». Такие формы долго не могли найти только лишь у ячменя. Но
закон гомологических рядов утверждал: такие формы у ячменя должны существовать
в природе или могут быть получены искусственным путем. Удивительно, но факт —
такой вариант ячменя был получен при обработке семян рентгеновскими лучами.
Сейчас, когда мы знаем о ДНК и принципах передачи наследственной информации в
деталях, закон гомологических рядов очевиден. Однако в 20-е годы прошлого века
генетика лишь зарождалась, а закон Вавилова вступил в кажущееся противоречие с
теорией Дарвина. Действительно, теория эволюции утверждает, что все живое
изменяется под действием внешней среды в том направлении, которое обеспечит
лучшие шансы на выживание. А тут оказывается, что список возможных вариаций
довольно предсказуем, то есть изменения запрограммированы в самом живом
организме, а не зависят только от внешней среды.
А тут оказывается, что список возможных вариаций
довольно предсказуем, то есть изменения запрограммированы в самом живом
организме, а не зависят только от внешней среды.
Это противоречие было разрешено позже с появлением представлений о мутагенезе: в обычном случае набор генов и вызываемые ими признаки действительно стабильны, однако иногда в генах могут возникать мутации, которые, если они выгодны в данной среде, закрепляются в новых поколениях.
Таким
образом, закон Вавилова существенным образом повлиял на развитие генетики.
Кроме того, он имел огромное практическое значение, так как активно применялся
в селекции для предсказания возможных свойств растений, поиском которых стоит
заняться. Основываясь на данном законе, селекционеры целенаправленно искали
коров, дающих больше молока, морозостойкую пшеницу, урожайный сорт картофеля.
Такие интенсивные поиски осуществлялись в течение нескольких десятилетий
буквально по всему свету. Вавилов лично организовал большое количество
экспедиций (около 110!) по сбору культурных растений и участвовал в них.
Благодаря закону гомологических рядов и последующей огромной работе, проделанной Николаем Вавиловым, в стране произошел значительный рост целенаправленной селекции и создания новых сортов, что имело огромное практическое значение в то непростое время. Саратовский съезд селекционеров 1920 года вошел в историю науки, так как именно на нем Вавилов поделился своим открытием с коллегами. А спустя год на Международном конгрессе по сельскому хозяйству, который проходил в США, ученый представил закон гомологических рядов на суд заокеанской публике.
Реакция
тамошних коллег была ошеломляющей: портреты Николая Вавилова украшали первые
страницы крупнейших американских изданий. Закон Вавилова в биологии сравним по
значимости с открытием периодической системы Менделеевым. Как периодическая
таблица Менделеева позволяет предсказывать свойства элементов и их соединений
(и даже существование новых элементов), так и закон гомологических рядов
предсказывает, какие признаки могут быть у вида, а также какие виды могут быть
в родах.
Николаю Вавилову также принадлежит идея «переписать» все сорта окультуренных растений и хранить собранные материалы живыми. Это начинание было поддержано советскими учеными, поэтому часть собранных растений ежегодно высевалась в самых разных условиях на специально созданных по всей стране опытных станциях.
Существенно
облегчило поиски нужных форм учение о центрах происхождения культурных
растений, которое также было сформулировано Николаем Ивановичем Вавиловым.
Основываясь на богатом коллекционном материале, собранном советскими ботаниками
в многочисленных экспедициях, он выделил семь основных географических центров
происхождения культурных растений, где и были сосредоточены основные поиски.
Последователи ученого до конца сохранили преданность ему, ведь даже в самые
голодные времена блокады Ленинграда ни одно зернышко из обширной коллекции не
было съедено сотрудниками Всесоюзного института растениеводства. Благодаря этим
экспедициям и созданию опытных станций Вавилову удалось собрать не имеющую
аналогов в мире коллекцию семян растений.
Коллекция семян до сих пор хранится в Санкт-Петербурге. Ее берегли, несмотря ни на что, и ценили больше жизни как уникальную реликвию государства. Даже на сегодняшний день на основании этой коллекции семян можно создавать новые сорта растений. Более того, по мнению биологов, данная коллекция обладает огромным потенциалом и способна обеспечить выживание всему миру. К слову, четверть тех растений, семена которых находятся в коллекции Вавилова, уже считаются вымершими. Однако потенциально растения живы, пока невредима коллекция.
Доказательство на примере культурных растений семейства Злаковые закона гомологических рядов в наследственной изменчивости Н.И. Вавилова
Доказательство на примере культурных растений семейства Злаковые закона гомологических рядов в наследственной изменчивости Н.И. Вавилова
«Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство».
Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство».
Зако́н гомологи́ческих рядов — закон наследственной изменчивости, сформулированный Н.И. Вавиловым в 1920 году. Закон устанавливает параллелизм в изменчивости родственных групп растений, в основе которого лежит гомология генов (их одинаковое молекулярное строение) и гомология (сходство) в порядке их расположения в хромосомах у родственных видов.
Н.И. Вавилов обнародовал этот закон в докладе «Закон гомологических рядов в наследственной изменчивости» в день открытия в Саратове III Всероссийского селекционного съезда 4 июня 1920 года. Позднее ботаник-физиолог профессор Заленский сказал о законе Вавилова:
Главная ценность закона гомологических рядов в наследственной изменчивости состоит в его прогностической сущности, позволяющей вести целенаправленный поиск гендоноров адаптивно значимых и хозяйственно ценных признаков.
На основе закона растениеводы и животноводы могут целенаправленно искать и находить нужные признаки и варианты у различных видов в мировом многообразии форм как культурных растений и домашних животных, так и у их диких родичей. Например, если черная окраска зерна встречается у ржи, пшеницы и других злаков, то растения с такой же окраской зерна могут быть получены у овса и проса. Так Н.И. Вавилов предсказал наличие безалкалоидного (неядовитого) люпина, учитывая существование безалкалоидных форм в родах гороха, фасоли и других растений семейства бобовых.
Цель: влияние климатических факторов (солнечный свет и вода) на злаковые культуры, произрастающие на одном участке (биотоп).
Задачи: наблюдать за воздействием абиотических факторов за культурами одного семейства.
Гипотеза: если на культуры одного семейства (злаковые) воздействуют одинаковые абиотические факторы, то результативность будет одинаковой.
В исследовании использовали 4 вида злаковых растений: рожь, ячмень, овес, пшеница.
Первые всходы
17.05-20.05
В зависимости от вида
-Зерновка овса, словно панцирем, плотно прикрыта цветковыми чешуями, и они оказывают воздействие на ход прорастания:появляющееся колеоптиле не в силах пробить этот пленчатый панцирь и проходит вдоль него, пока не найдет выход у верхушки зерновки, где между соприкасающимися чешуями имеется щель; выйдя из нее, оно направляется кверху.
Продолжающее расти колеоптиле пронзает почву и выходит на дневную поверхность;
—Ячмень:начинается с прорастания семян, заканчивается появлением всходов и образованием третьего листа. Общая продолжительность ее 15-30 дней.
Вначале развивается главный побег и в пазухах зародышевых листьев закладываются почки боковых побегов. Главный зародышевый корешок к концу фазы всходов проникает в почву на глубину до 40 см.
—Пшеница: При температуре почвы на глубине заделки семян 15 °С всходы появляются на 7-ой день (19.05)
—Рожь: для прорастания зародышевых корней требуется 3,8°С, а для появления первого листа требуется более высокая температура. Первым трогается в рост главный корешок и углубляясь в почву поглощает воду и минеральные вещества почвенного раствора. Всходы на 6-8 день.
Первым трогается в рост главный корешок и углубляясь в почву поглощает воду и минеральные вещества почвенного раствора. Всходы на 6-8 день.
Кущение: Кущение — это биологический процесс, имеющий важное значение в жизни растений, так как в фазу кущения закладываются органы, определяющие урожай.
С 20.05
-У ржи кущение начинается, когда образуется 4-й лист. В это время появляется первый боковой побег из почки в пазухе первого зародышевого листа. Затем появляются следующие побеги из почек в пазухах следующих зародышевых листьев. Одновременно развиваются узловые корни. Каждый новый побег имеет свои корни. Таким путем формируется узел кущения, который располагается близко к поверхности почвы. Узел кущения — важная часть растения, так как в нем формируются боковые побеги и узловые корни. Степень кущения зависит от многих факторов — плодородия почвы, погодных условий, сроков посева, норм высева и глубины заделки семян, а также от биологических особенностей сорта. Кущение продолжается 35-37 дней.
Кущение продолжается 35-37 дней.
-Кущение яровой пшеницы лучше проходит при температуре 10-12°С.
Недостаток влаги в почве в этот период увеличивает количество бесплодных колосков
—овес:Эта фаза начинается обычно через 10- 15 дней после появления всходов, в момент развития 3- 4-го листа. Число всех стеблей на одно растение обозначается как общая кустистость, а число стеблей с нормально развитой метелкой — как продуктивная. Последняя обычно составляет 2-4 стебля. В разреженных посевах кустистость возрастает. Сорта голозерного овса кустятся сильнее пленчатого. В фазе полного кущения у сортов овса различают следующие формы куста: прямостоячую (сомкнутую), распластанную (лежачую) и промежуточную.
— Кустится ячмень сильнее, чем яровая пшеница и овес. При появлении третьего листа ячмень начинает куститься, то есть образовывать боковые побеги. Главный узел кущения расположен в почве на глубине 1 — 3 см, важно, чтобы узел кущения находился в увлажненном слое почвы.
выход в трубку
С 10-24.06
— Ячмень сильно страдает от быстрого наступления высокой температуры в фазе выхода в трубку, когда формируется продуктивность колоса. В период выхода в трубку — колошения наиболее благоприятна среднесуточная температура 20—22°С, Фаза выхода в трубку наступает примерно через 3—4 недели после появления полных всходов.
— пшеница:Период от выхода в трубку до колошения — очень важный этап в развитии зерновых хлебов. В это время усиленно растут листья и соломина, формируется колос. Растение испытывает повышенную потребность во влаге и питательных веществах. В фазе выхода в трубку у яровой пшеницы можно легко различить узлы, междоузлия, язычки и ушки. Стеблевой узел представляет перехват на тех частях стебля, где имеются сплошные перегородки. Междоузлием является часть стебля между двумя узлами. Междоузлия бывают полые, реже выполненные вверху рыхлой тканью. Над стеблевым узлом располагается листовой узел — небольшое кольцевое утолщение, с помощью которого лист прикреплен к стеблю. Лист плотно охватывает междоузлие, что придает стеблю прочность и защищает его нежные растущие части от внешних повреждений. В месте перехода листового влагалища в листовую пластинку находится язычок, представляющий тонкую пленку, прилегающую к стеблю. Около язычка по краям листового влагалища помещаются два полулунных рожка, или ушка, которые способствуют закреплению влагалища листа на стебле.
Лист плотно охватывает междоузлие, что придает стеблю прочность и защищает его нежные растущие части от внешних повреждений. В месте перехода листового влагалища в листовую пластинку находится язычок, представляющий тонкую пленку, прилегающую к стеблю. Около язычка по краям листового влагалища помещаются два полулунных рожка, или ушка, которые способствуют закреплению влагалища листа на стебле.
— Овес:Начинается фаза через 10-15 дней после кущения и означает начало образования соломины. На практике можно определить прощупыванием узла на стебле от поверхности почвы. Обычно с этого момента начинается быстрый рост надземных органов и корней, который продолжается до начала цветения; позднее процессы роста замедляются и постепенно затухают. После выхода в трубку появляются органы полового размножения — цветки, собранные в колоски и в соцветие — метелку.
—Рожь: Вытягивание первого междоузлия на 5-6 см и появление нижнего стеблевого узла на поверхности почвы принято считать началом фазы выхода в трубку.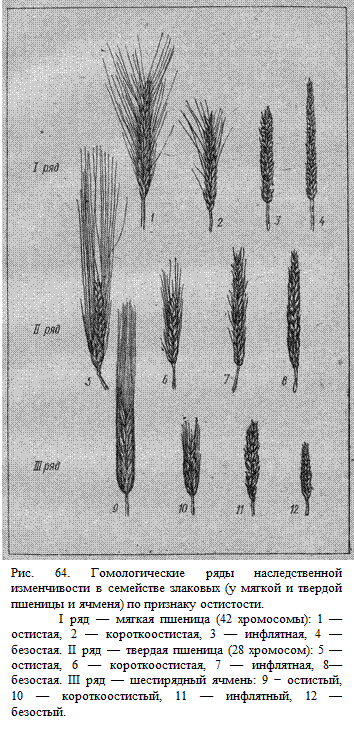 С этого момента начинается быстрый рост стеблей и развитие колоса, заключенного внутри трубки листовых влагалищ. В это время заканчивается образование основной массы листьев. К концу фазы растения достигают высоты, соответствующей особенностям данного сорта.
С этого момента начинается быстрый рост стеблей и развитие колоса, заключенного внутри трубки листовых влагалищ. В это время заканчивается образование основной массы листьев. К концу фазы растения достигают высоты, соответствующей особенностям данного сорта.
колошение
С 10.07
— Рожь: Фаза отмечается при выдвижении колоса из влагалища верхнего листа. Листья раздвигаются в результате вытягивания междоузлий, листья увеличиваются в размере. Стебель укрепляется механически и выдерживает увеличивающуюся массу листьев и колоса. Колошение начинается в среднем через 40-50 дней после весеннего возобновления вегетации. Период от кущения до колошения у яровой ржи- 25-30 дней.
С 10.07
—Овес: определяют по появлению первого колоска из влагалища верхнего листа.
С 15.07
—Пшеница: Фаза колошения начинается выходом из влагалища последнего листа. Колошение у яровой пшеницы наступает через 50-60 дней после посева и длится 10-12 дней. Эта фаза продолжается 1-4 дня в зависимости от сорта и погодных условий. Наиболее благоприятная температура в этот период 20-25 ºС. В период выхода в трубку и колошения происходит самый интенсивный рост вегетативной массы растений, а также расходуется большое количество влаги (50-60 % потребляемого количества за вегетацию)
Эта фаза продолжается 1-4 дня в зависимости от сорта и погодных условий. Наиболее благоприятная температура в этот период 20-25 ºС. В период выхода в трубку и колошения происходит самый интенсивный рост вегетативной массы растений, а также расходуется большое количество влаги (50-60 % потребляемого количества за вегетацию)
С 13.07
—Ячмень: Фаза колошения наступает, когда колос наполовину выходит из влагалища листа. Обычно это случается на 38-42 день после появления всходов. К наступлению колошения ячмень полностью сформировывает генеративные органы— пыльники и пестик с рыльцами.
цветение
С 13.07
—Рожь: При созревании генеративных органов цветковые чешуи раздвигаются вследствии набухания лодикул. При этом наружу выбрасываются тычинки и рыльце пестика. В каждом пыльнике у ржи до 20 тыс. пыльцевых зерен.. Цветение яровой ржи наступает через 3-6 дней после колошения и продолжается 6-7 дней. Цветение начинается со средней части колоса. Пыльца переносится ветром на рыльце другого цветка и прорастает, проникает в завязь и сливается с яйцеклеткой. На рыльце своего цветка пыльца плохо прорастает. Для цветения требуется 12,5°С. Цветение каждого цветка продолжается 12-25 минут.
Пыльца переносится ветром на рыльце другого цветка и прорастает, проникает в завязь и сливается с яйцеклеткой. На рыльце своего цветка пыльца плохо прорастает. Для цветения требуется 12,5°С. Цветение каждого цветка продолжается 12-25 минут.
С 15.07
—Овес: Овес считается самоопылителем, однако у гексаплоидных видов нередки случаи перекрестного опыления. Цветение овса начинается одновременно с выходом метелки из влагалища с растрескиванием пыльников самых верхних ее колосков и концов отдельных веточек. Затем цветение последовательно переходит к основанию веточек и мутовок метелки. В колоске оно начинается с нижнего цветка и идет в восходящем порядке, поэтому колоски в метелке разновозрастные: если в нижней части метелки еще наблюдается цветение, в верхушечных колосках происходит налив зерна.
С 25.07
—Пшеница: Цветение яровой пшеницы протекает интенсивнее в утренние и вечерние часы. Продолжительность цветения одного колоса 3-5 дней, а всего поля обычно 5-7 дней. Яровая пшеница относится к самоопыляющимся культурам, но не исключено и перекрестное опыление. Недостаточная влажность и повышенная температура воздуха в это время снижают степень оплодотворения цветков, что уменьшает число зерен в колосе
Яровая пшеница относится к самоопыляющимся культурам, но не исключено и перекрестное опыление. Недостаточная влажность и повышенная температура воздуха в это время снижают степень оплодотворения цветков, что уменьшает число зерен в колосе
С 20.07
—Ячмень: Цветение — важный период в жизни растений, так как к концу его прекращается рост вегетативных органов, а в большинстве случаев — и рост корневой системы. Ячмень — самоопыляющееся растение, однако в ряде случаев опыляется и перекрестно. В каждом развитом цветке у него находятся мужские и женские органы. Цветение ячменя чаще всего совпадает с началом колошения и реже (через 1—3 дня) после него. В засушливые годы ячмень цветет во влагалище листа.
Цветение начинается со средних колосков и одновременно распространяется на верхние и нижние части колоса. Наиболее интенсивное цветение и оплодотворение наблюдается в утренние часы. Весь процесс оплодотворения продолжается 6—8 ч.
созревание
С 20. 05
05
—Рожь:Поскольку колошение происходит не одновременно, то зерно от разных побегов формируется при различных условиях. В пределах каждого колоса зерно также не однородно. Кроме внешних условий на зерно влияют внутренние условия, поэтому зерно отличается по морфологии, биохимии. Скорость созревания зерновки зависит от погодных условий. Сильное влияние оказывают ветры, температура воздуха и влажность. При молочной спелости зерно имеет зеленую окраску, содержит 60-40% воды, в которой растворены органические вещества. При восковой спелости растение желтеет. Зерно становится желтым, имеет консистенцию воска. В эту фазу зерно отделяется от материнского растения. Влаги в зерне 20-40%. При полной спелости растение полностью желтеет, зерно твердеет, имеет 14-16% влаги. В период созревания в зерне увеличивается содержание крахмала, в стебле уменьшается содержание сахара. В зерне до конца молочной спелости увеличивается содержание азота. В начале созревания в зерне активны различные ферменты, но в стадии восковой спелости зерно устойчиво на влияние ферментов. При полной спелости зерно имеет слабую ферментативную активность. В молочной и восковой спелости в зерне много аскорбиновой кислоты, каротина и каротиноидов. При полной спелости их количество резко уменьшается. В период восковой спелости в зерновке накапливаются все жизненно необходимые вещества, обеспечивающие нормальную всхожесть.
При полной спелости зерно имеет слабую ферментативную активность. В молочной и восковой спелости в зерне много аскорбиновой кислоты, каротина и каротиноидов. При полной спелости их количество резко уменьшается. В период восковой спелости в зерновке накапливаются все жизненно необходимые вещества, обеспечивающие нормальную всхожесть.
С 22.07
—Овес: При послеуборочном дозревании в зерне протекают сложные биохимические процессы. Содержание азотистых веществ и крахмала в зерне по мере хранения увеличивается, количество сахаров уменьшается. Семена при хранении от — 1 до 25 °С имеют малорастворимые белки, которые распадаются до аминокислот, тормозится дыхание. При покое у семян ржи влажность должна быть 10%. Если влажность увеличивается дыхание усиливается, что приводит к самосогреванию зерна.
С 28.07
—пшеница: созревание зерна наступает после оплодотворения завязи, когда начинается приток в нее питательных веществ и постепенное ее разрастание. Различают три фазы созревания: молочную, восковую и полное созревание.
Различают три фазы созревания: молочную, восковую и полное созревание.
С 22.07
—Ячмень: Формирование зерна у ярового ячменя продолжается 10-11 дней. В конце этого этапа зерно достигает окончательной длины, но остается щуплым. Содержание влаги в нем снижается с 80 до 65%. Растение и зерно имеют зеленый цвет. Налив зерна длится 11-14 дней, от начала молочной до восковой спелости. Во время налива происходит максимальное накопление в зерне минеральных и органических веществ, при снижении влаги до 40%. При сдавливании зерна в молочной спелости выделяется белая жидкость.
Сбор семян
—Рожь: При послеуборочном дозревании в зерне протекают сложные биохимические процессы. Содержание азотистых веществ и крахмала в зерне по мере хранения увеличивается, количество сахаров уменьшается. Семена при хранении от — 1 до 25 °С имеют малорастворимые белки, которые распадаются до аминокислот, тормозится дыхание. При покое у семян ржи влажность должна быть 10%. Если влажность увеличивается дыхание усиливается, что приводит к самосогреванию зерна.
Если влажность увеличивается дыхание усиливается, что приводит к самосогреванию зерна.
— Овес:
—Пшеница: Молочная спелость наступает через 8-18 дней после начала цветения. В эту фазу зерно достигает нормальной длины, при надавливании из него выступает белая, густой консистенции жидкость. Влажность зерна при этом высокая – 72-47 %.
Восковая спелость наступает через 10-14 дней после молочной. Зерно приобретает желтоватую окраску, его содержимое, как воск, хорошо режется ногтем. В этой фазе спелости в зерновке содержится 32-25 % воды.
Полная спелость характеризуется потерей воды в зерне до 18-15 %, оно приобретает характерную для сорта окраску, твердость, ногтем не режется.
—Ячмень:
Подводя итоги рассмотренным закономерностям, мы приходим к следующим положениям.
Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и линнеоны, тем полнее сходство в рядах их изменчивости.
Чем ближе генетически расположены в общей системе роды и линнеоны, тем полнее сходство в рядах их изменчивости.
Гомологическая изменчивость обусловлена генетическим родством видов и родов, проявляется лучше всего в переделах одного семейства, тогда как в аналогичной и общей изменчивости генетическое родство не играет роли.
Позже было установлено, что этот закон распространяется на животные и микроорганизмы. Так, альбинизм встречается у всех классов позвоночных животных, коротколапость наблюдается у всех пород крупного рогатого скота, овец и собак.
Подготовка почвы к посадке
Посадка семян и полив
Первые всходы и кущение
Созревание
Закон гомологических рядов
Общее строение клеток различных организмов, отмеченное -еще Т. Шваном (1839), было научно обосновано в 1920 г. академиком Н. И. Вавиловым в открытом им законе гомологических рядов в наследственной изменчивости.[ …]
Шваном (1839), было научно обосновано в 1920 г. академиком Н. И. Вавиловым в открытом им законе гомологических рядов в наследственной изменчивости.[ …]
Ограничение числа и форм взаимодействия подсистем одного системного уровня ограничивает «фантазию природы», укладывает ее в строго лимитированный закономерный ряд образований. Периодический закон химических элементов Д. И. Менделеева (свойства химических элементов, проявляющиеся в простых веществах и соединениях, находятся в периодической зависимости от заряда ядер их атомов) был первой формализацией «третьего измерения» таблицы «системы систем» (см. табл. 2.1). Родственные виды, роды, семейства и др.[ …]
Вавилов Николай Иванович (1887—1943) — русский, советский ученый, основоположник современного учения о биологических основах селекции и учения о центрах происхождения культурных растений. Сформулировал закон гомологических рядов в наследственной изменчивости близких видов и родов (1920), который биологи сравнивают с периодической системой Д. И. Менделеева. С одинаковым основанием В. можно считать и генетиком, и растениеводом, и селекционером, и географом.[ …]
И. Менделеева. С одинаковым основанием В. можно считать и генетиком, и растениеводом, и селекционером, и географом.[ …]
Подробные сведения о генетике фотопериодической реакции цветения различных растительных видов приведены в сводке В. В. Скрипчинского [Скрипчинский, 1971, 1975]. Анализируя большой материал по генетике фотопериодизма, он приходит к выводу, что изменчивость фотопериодических биотипов подчиняется закону гомологических рядов, как это было установлено Н. И. Вавиловым в 1935 г. для озимых, полуозимых и яровых форм.[ …]
Поскольку с некоторой вероятностью каждый рецессивный аллель иногда оказывается в гомозиготном состоянии, т. е. в результате свободного скрещивания оба аллеля данного гена оказываются у определенного организма одинаковыми, соответствующий вариант признака проявляется в фенотипе, как называется весь комплекс проявившихся у конкретного организма признаков. Таким образом, у родственных видов, например у видов одного семейства растений, варианты тех признаков, которые не являются строго видовыми, оказываются в значительной мере общими, как и сами признаки, и эта общность имеющихся у близких видов вариантов признаков в виде запаса рецессивных аллелей унаследована ими от общего предка и сохранена в течение миллионов лет независимой эволюции. В этом и состоит смысл открытого Н. И. Вавиловым в 1920 г. Закона гомологических рядов наследственной изменчивости.[ …]
В этом и состоит смысл открытого Н. И. Вавиловым в 1920 г. Закона гомологических рядов наследственной изменчивости.[ …]
Закон гомологических рядов Вавилова к пятидесятилетию его формулировки
Андерсон Э. 1949. Интрогрессивная гибридизация. Нью-Йорк.
Бахтеев Х. 1962. Новое звено в дикорастущем виде ячменя. Бот. журнал. 47, 844–847 (рус.).
Google Scholar
Аль-Бассам, Х. 1966. Биохимические изменения, сопряженные с мутациями потери антоциановой окраски цвета у некоторых растений.Докл. акад. Наук С.С.С.Р. 168, 1405–1408 (рус.).
Google Scholar
Крейн, М. Б. и Лоуренс В. Дж. К. 1952. Генетика садовых растений. Лондон.
Филипченко Ю. А. 1924. Параллелизм в жизни природы. Успехи эксперимент. биологии 3, 242–258.
Google Scholar
Харлан, Т. Р. , Дж. М. де Вет и Г. Прайс. 1973.Сравнительная эволюция злаков. Эволюция 27 : 311–325.
, Дж. М. де Вет и Г. Прайс. 1973.Сравнительная эволюция злаков. Эволюция 27 : 311–325.
Артикул Google Scholar
Кляйншмидт, О. 1936. Parallelentwicklung und Wiederholungerscheinung in der Tierwelt. Нова Акта Леопольдина 4, 367–391.
Google Scholar
Купзов, А. Дж. 1959. Параллелизм в изменчивости видов растений с некоторыми общими признаками. Zeitschr.шерсть. Pflanzenzüchtung 41, 313–325.
Google Scholar
Купзов, А. Дж. 1969. Амфимиксис и апомиксис в эволюции и размножении гуаюлы. Zeitschr. für Pflanzenzüchtung 61, 121–140.
Google Scholar
Lamprecht, H. 1964. Видовая концепция и происхождение видов. Две категории генов: внутри- и межвидовые. Agri-hortique Genetica 22 : 272–280.
Google Scholar
Леппик Э. Е. 1969. Гомологические и аналогичные ряды в эволюции типов цветков (на русском языке с англ. резюме) Генетика СССР, 5 :12–23.
Е. 1969. Гомологические и аналогичные ряды в эволюции типов цветков (на русском языке с англ. резюме) Генетика СССР, 5 :12–23.
Google Scholar
Матус, Дж. 1965. Самцы стерилита у культурныч ростлин а ежи выузити при следствии гетерозныч одруд. Вед. праче Вызк-уставу окраски заградництви в Пругоничичей, 3, 13–88.
Google Scholar
Michaelis, P. 1954. Цитоплазматическое наследование у Epilobium и его теоретическое значение. Успехи генетики 6, 287–401.
ПабМед КАС Google Scholar
Напп-Цинн, К. 1961. Uber Bedeutung genetischer Untersuchungen an Kälte bedürftigen Pflanzen für Aufklärung von Vernalisationsercheinungen. Цюхтер 31, 128–135.
Артикул Google Scholar
Пармасто, Э. 1970. Академия наук Эстонской ССР, Тарту, Эстония. Личное письмо в редакцию.
Личное письмо в редакцию.
Roemer, Th. 1936. Die Bedeutung des Gesetzes der Parrallelvariation für die Pflanzenzüchtung. Нова Акта Леопольдина, 4, 351–365.
Google Scholar
Синская Е. Н. 1927. Генетико-систематические исследования культурных Brassica .Бык. заявл. бот. и пл. порода. 17, 3–166.
Google Scholar
Вавилов Н. И. 1920. Закон гомологических рядов в наследственной изменчивости. Доклад на 3 Всероссийском селекционном выезде в Саратово, 4 июня 1920. Саратов.
Вавилов Н. И. 1922. Закон гомологических рядов в вариации. Журнал. генетики 12, 47–89.
Артикул Google Scholar
Вавилов Н.I. 1935. Закон гомологических рядов в наследственной изменчивости. Теоретические основы селекции растений I, 75–128.
Google Scholar
Заварзин, А. 1925. Der Parallelismus der Strukturen als Grundprinzip der Morphologie. Zeitschr. für wissenschaft. Зоология 124, 118–212.
1925. Der Parallelismus der Strukturen als Grundprinzip der Morphologie. Zeitschr. für wissenschaft. Зоология 124, 118–212.
Google Scholar
4. Закон гомологических рядов Вавилова — актуален ли он для картофеля? | Биологический журнал Линнеевского общества
Получить помощь с доступом
Институциональный доступ
Доступ к контенту с ограниченным доступом в Oxford Academic часто предоставляется посредством институциональных подписок и покупок.Если вы являетесь членом учреждения с активной учетной записью, вы можете получить доступ к контенту следующими способами:
Доступ на основе IP
Как правило, доступ предоставляется через институциональную сеть к диапазону IP-адресов. Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с проверкой подлинности IP.
Войдите через свое учреждение
Выберите этот вариант, чтобы получить удаленный доступ за пределами вашего учреждения.
Технология Shibboleth/Open Athens используется для обеспечения единого входа между веб-сайтом вашего учебного заведения и Oxford Academic.
- Щелкните Войти через свое учреждение.
- Выберите свое учреждение из предоставленного списка, после чего вы перейдете на веб-сайт вашего учреждения для входа.
- При посещении сайта учреждения используйте учетные данные, предоставленные вашим учреждением.Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если вашего учреждения нет в списке или вы не можете войти на веб-сайт своего учреждения, обратитесь к своему библиотекарю или администратору.
Вход с помощью читательского билета
Введите номер своего читательского билета, чтобы войти в систему. Если вы не можете войти в систему, обратитесь к своему библиотекарю.
Члены общества
Многие общества предлагают своим членам доступ к своим журналам с помощью единого входа между веб-сайтом общества и Oxford Academic. Из журнала Oxford Academic:
- Щелкните Войти через сайт сообщества.
- При посещении сайта общества используйте учетные данные, предоставленные этим обществом. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если у вас нет учетной записи сообщества или вы забыли свое имя пользователя или пароль, обратитесь в свое общество.
Некоторые общества используют личные аккаунты Oxford Academic для своих членов.
Личный кабинет
Личную учетную запись можно использовать для получения оповещений по электронной почте, сохранения результатов поиска, покупки контента и активации подписок.
Некоторые общества используют личные учетные записи Oxford Academic для предоставления доступа своим членам.
Институциональная администрация
Для библиотекарей и администраторов ваша личная учетная запись также предоставляет доступ к управлению институциональной учетной записью. Здесь вы найдете параметры для просмотра и активации подписок, управления институциональными настройками и параметрами доступа, доступа к статистике использования и т. д.
Просмотр ваших зарегистрированных учетных записей
Вы можете одновременно войти в свою личную учетную запись и учетную запись своего учреждения.Щелкните значок учетной записи в левом верхнем углу, чтобы просмотреть учетные записи, в которые вы вошли, и получить доступ к функциям управления учетной записью.
Выполнен вход, но нет доступа к содержимому
Oxford Academic предлагает широкий ассортимент продукции. Подписка учреждения может не распространяться на контент, к которому вы пытаетесь получить доступ.
Если вы считаете, что у вас должен быть доступ к этому контенту, обратитесь к своему библиотекарю.
Подписка учреждения может не распространяться на контент, к которому вы пытаетесь получить доступ.
Если вы считаете, что у вас должен быть доступ к этому контенту, обратитесь к своему библиотекарю.
Закон гомологических рядов в вариации (Н.И. Вавилов)
Нина Булатова / Comparative Cytogenetics 14: 329–338 (2020)
334
Еще большее разнообразие наблюдается у растений, размножающихся вегетативно или апогамно, таких как розы, картофель, яблони, Hierac ), и Далия.
Мы точно не знаем, существуют ли в природе действительно монотипные линнеевские виды,
достаточно специфичные и отделенные от других линнеевских видов и представленные одной
разновидностью, только одной джорданоновской.В целом создается впечатление, что чем больше мы изучаем наши растения
и животных, тем более они изменчивы, тем больше разновидностей мы находим среди линнеевских
видов. Некоторые линнеевские виды растений и животных, такие как розы, пшеница, индийская кукуруза, рис, кабачки, дрозофилы, кажутся чрезвычайно изменчивыми, но они привлекли больше внимания, чем другие. Мы легко замечаем резкие различия в цвете, размере и форме
Мы легко замечаем резкие различия в цвете, размере и форме
одних органов и весьма невнимательны к другим.
Отличия Иорданонов в пределах одного и того же Линнеона в форме
и окраске их цветков, форме и размерах листьев, плодов и других органов весьма
часто не менее заметны, чем у
различия между самими Линнеонами. Например,
некоторые сорта Cucurbita Pepo характеризуются плодами размером с куриное яйцо; другие сорта
, растущие в тех же условиях, приносят плоды диаметром три и четыре фута.У некоторых разновидностей Sesamum indicum листья и плоды супротивные, у других
очередные. Некоторые сорта пшеницы и ржи имеют простые листья, без дифференциации
на влагалища и пластинки, не имеющие ни «язычков», ни «ушных раковин»; у других обычные сложные листья, с «язычком» и «ушком».
Растения самоопыляемые, такие как пшеница, ячмень, горох, соя и т. д., и перекрестно опыляемые, такие как рожь,
кукуруза, свекла, эль, подобные полиморфные. Кажущаяся однородность нескольких перекрестно опыленных
Кажущаяся однородность нескольких перекрестно опыленных
дикорастущих и культурных растений становится очевидной только тогда, когда они тщательно не изучаются.Различие состоит только в гомозиготности многих признаков у перекрестно-опыленных
растений и в гомозиготности самоопыленных растений. Некоторые рецессивные признаки могут быть скрыты в растениях с перекрестным опылением за счет доминирования других признаков,
, но при искусственном самоопылении этих растений и путем инбридинга возможно
восстановить их . Из того, что мы знаем в настоящее время при изучении индийской кукурузы (Эм-
,, Эрсон, Коллинз и др.), ржи, свеклы, дрозофилы, самого человека, перекрестно-оплодотворенные
организмы не менее изменчивы, чем самоопыляемые.
Вышеупомянутое количество иорданцев в действительности еще больше, потому что до настоящего времени африканские и азиатские разновидности даже самых важных культурных
растений, таких как пшеница, овес, ячмень, горох, чечевица, Крестоцветные почти неизвестны.
Проблемы будущего
Существует реальная потребность в изучении и систематизации этих иорданов, особенно
у культурных растений и домашних животных, в интересах генетиков, а также
как систематиков и агрономов.Только самое пристальное изучение Джорданонов и генотипов
даст реальное представление о том, что такое Линнеон. Для построения общих генетических
схем необходимо знать состав линнеевских видов. Прежде чем создавать
новых сорта путем скрещивания, надо знать, что существует в природе. Даже для злаков, Le-
guminoseae и других важнейших растений мы не располагаем достаточными знаниями даже о
легко узнаваемых ботанических разновидностях.Области древней культуры в Азии, Африке и
Америке еще сохраняют множество сортов, неизвестных систематикам и селекционерам.
100 лет закону Вавилова о гомологических рядах в вариации
Прошел век с тех пор, как впервые проявился закон гомологических рядов в вариации. Это событие произошло в 1920 г. в Саратове, на третьем послереволюционном году, в рамках III Всероссийской конференции по селекции, тогда мобилизованной с учетом актуальных потребностей сельскохозяйственной практики, науки и образования.Сенсацию произвел доклад 33-летнего профессора Николая Вавилова, которого сопровождали его студенты из Саратовского университета. Обобщение Вавилова о явлении гомологического ряда в изменчивости культурных растений было доложено 4 июня 1920 г. и с энтузиазмом оценено квалифицированными селекционерами как великое научное достижение, сравнимое с периодическим законом химических элементов Менделеева. 21 июня 1920 года сообщение губернского Саратовского отделения Российского телеграфного агентства распространило на международном уровне информацию о «величайшем открытии мирового значения», которое решением собрания было адресовано Государственному правительству.
Это событие произошло в 1920 г. в Саратове, на третьем послереволюционном году, в рамках III Всероссийской конференции по селекции, тогда мобилизованной с учетом актуальных потребностей сельскохозяйственной практики, науки и образования.Сенсацию произвел доклад 33-летнего профессора Николая Вавилова, которого сопровождали его студенты из Саратовского университета. Обобщение Вавилова о явлении гомологического ряда в изменчивости культурных растений было доложено 4 июня 1920 г. и с энтузиазмом оценено квалифицированными селекционерами как великое научное достижение, сравнимое с периодическим законом химических элементов Менделеева. 21 июня 1920 года сообщение губернского Саратовского отделения Российского телеграфного агентства распространило на международном уровне информацию о «величайшем открытии мирового значения», которое решением собрания было адресовано Государственному правительству.
Очень скоро после первой русской публикации (Вавилов, 1920) в «Журнале генетики» под редакцией В. Бейтсона и Р.К. Пеннетт, старейшие государственные деятели генетики (Вавилов, 1922).
Бейтсона и Р.К. Пеннетт, старейшие государственные деятели генетики (Вавилов, 1922).
В 1925 году Уильям Бейтсон, директор Садоводческого института Джона Иннеса, с коллегами посетил опытные поля и лаборатории Николая Вавилова, директора Бюро прикладной ботаники и селекции растений (будущий Н.И. Вавилова) в пригороде Петрограда — Ленинграда (ныне Санкт-Петербург) (рис. 1). Статья заняла 42 страницы тома XII (1) (апрель 1922 г., стр. 47–89). Суть этой работы Вавилова была отражена в главах Н. Тимофеева-Ресовского (1940) и Н. Вавилова (1940) в монографии «Новая систематика» (Хаксли, 1940), обзорной книге, предшествовавшей публикации о новый синтез теории эволюции (Huxley 1942).
С тех пор и до сих пор генетическая природа гомологичной изменчивости, сущность закона Вавилова, находится в центре внимания различных дисциплин, от земледелия до палеонтологии, обновляясь с прогрессом молекулярной биологии.В настоящее время молекулярная гомология может быть установлена универсально на различных уровнях, от уникальных генов до порядков генов в хромосомах с помощью генетического, цитогенетического и молекулярного анализов (Захаров, 1987) до генных сетей, изучаемых биоинформатикой (Суслов и др. , 2008). Представляется рациональным встретить 100-летие этого знаменательного события молодой наследственной науки цифровой копией, сохранив замысел автора для будущих читателей и исследователей. Здесь текст воспроизводится в архивном формате с печатных страниц Введения (стр.48–53) и заключительный раздел (стр. 86–89) оригинальной английской версии (Вавилов, 1922). На титульном листе, скопированном на рис. 2, представлено все содержание этой работы. Детали пунктуации и цитирования обычно сохраняются.
, 2008). Представляется рациональным встретить 100-летие этого знаменательного события молодой наследственной науки цифровой копией, сохранив замысел автора для будущих читателей и исследователей. Здесь текст воспроизводится в архивном формате с печатных страниц Введения (стр.48–53) и заключительный раздел (стр. 86–89) оригинальной английской версии (Вавилов, 1922). На титульном листе, скопированном на рис. 2, представлено все содержание этой работы. Детали пунктуации и цитирования обычно сохраняются.
Н. Булатова
Цитаты:
Вавилов Н.И. (1922) Закон гомологических рядов в вариации. Журнал генетики 12 (1): 47–89. https://doi.org/10.1007/BF02983073
Булатова Н. (2020) Архив: Закон гомологических рядов в вариации (Н.И. Вавилов). Сравнительная цитогенетика 14 (3): 329–338. https://doi.org/10.3897/CompCytogen.v14i3.54511
————————————————— ——-
Рисунок 1.
Н.И. Вавилов (слева внизу) и российские генетики В.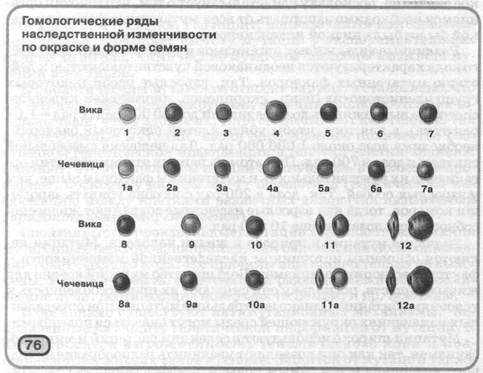 А. Догель, Ю.А. Филипченко с прибывшей европейской делегацией: Х. Федерли, О. Фогт и У. Бейтсон (слева направо во втором ряду). 1925 г., Ленинград (Вавилов 2012).
А. Догель, Ю.А. Филипченко с прибывшей европейской делегацией: Х. Федерли, О. Фогт и У. Бейтсон (слева направо во втором ряду). 1925 г., Ленинград (Вавилов 2012).
Рисунок 2.
Титульный лист статьи Вавилова в «Журнале генетики», 12(1), 1922 г.
Ботаника онлайн: Классическая генетика — Хромосомные числа — Генные центры
Ботаника онлайн: Классическая генетика — Хромосомные числа — Генные центры — АтавизмыБотаника онлайн 1996-2004. Никаких дальнейших обновлений, только исторический документ ботанической науки !
Географическое происхождение культивируемых Растения (генные центры) и атавизмы
Генные центры – это регионы, содержащие экстраординарное ассортимент дикорастущих собратьев культурных видов, полезные тропические растения или географическое происхождение некультивируемых родов.Генетический откат признака, считающегося примитивным у культивируемых животных.
разновидности называется атавизмом.
Российский генетик Н. И. ВАВИЛОВ (Московский сельскохозяйственный институт, Москва — потом работал в Саратове, умер как политический заключенный) обнаружил в своих многочисленных экспедициях, что некоторые географические районы характеризуются необычайной ряд диких сортов, которые можно рассматривать как дикие аналоги культурных видов. Он назвал эти районы главными центрами происхождения культурных растений или мировых центров сортового разнообразия (гены) культурных растений.
Идея была разработана с тех пор, так что теперь это не относится только к диким эквивалентам культурных растений во всем мире, но и полезным тропическим растениям, а также к географическому происхождению некультурные роды или семейства.
ВАВИЛОВ обнаружил сходные изменения или параллельные мутации у многих
близкородственные виды («Закон гомологических рядов в наследственном
Вариация», 1920). Подводя итоги своих исследований, ВАВИЛОВ
пришли к следующим выводам:
Подводя итоги своих исследований, ВАВИЛОВ
пришли к следующим выводам:
-
Генетически близкородственные виды и роды характеризуется сходными рядами наследственной изменчивости с такими закономерность, которая, зная ряды форм внутри одного вида, можно предсказать существование параллельных форм у других видов и роды.Чем генетически ближе роды и Линнеоны внутри общей системы, тем больше сходство в серии их вариаций.
-
Целые семейства растений обычно характеризуются определенным закономерность изменчивости, пронизывающая все роды и виды в составе семьи.
Все злаки, например рожь, пшеница, просо, саго или кукуруза, имеют
сорта с шелухой или без нее, например, с устойчивыми или ломкими колосьями или
бесязычковые формы (пшеница и рожь).Идеальные предпосылки для создания
Таким образом, были заданы оптимальные для человека комбинации и поставлена задача
человеку оставалось только отобрать наиболее благоприятные сорта
и начать их выращивание.
«Хрупкие початки» злаков считаются примитивным признаком, встречается лишь в исключительных случаях у культурных сортов. Крест некоторые сорта со стабильными колосьями иногда дают потомство с ломкими ушами. Такие отступления называются атавизмов (песн. атавизм ).
Важная информация (фотографии, распространение, выход) о
количество культурных растений можно найти в разделе:
http://www.mpiz-koeln.mpg.de/pr/garten/schau/index.html
Мировые центры сортового разнообразия (гены) культурных растений
- Южноазиатская тропическая, включающая следующие очаги: (1.) Индийская, (2.) Индокитайская, (3.) Островная, включая всю Малайский архипелаг
- Восточноазиатский: (1.) Китайский, (2.) Японский
- Юго-Западная Азия: (1.) Кавказ, (2.) Ближний Восток, (3.
 )
Северо-Западный Индийский
)
Северо-Западный Индийский - Средиземное море
- Эфиопский: Рядом с ним находится горный Аравийский, или Йемен, фокус
- Центральная Америка, включая Южную Мексику. Это делится на (1.) Горный район Южной Мексики, (2.) Центральноамериканский, (3.) Вест-Индийские островные очаги
- Андианский, в Южной Америке. Он включает (1.) Андийский, (2.) хилоанские или арауканские и (3.) боготские очаги.
Некоторые растения, писал ВАВИЛОВ, введены в культуру из дикая флора в прошлом, независимая от перечисленных выше семи основные центры. Так, в оазисах Аравии, Южной Месопотамии и может Сахара финиковая пальма была представил. В ЮАР, в районах, прилегающих к Калахари пустыня, арбуз был введен; во внутренних тропических Южная Америка маниок, ананас, арахис, а недавно (у европейцев) каучуковое дерево.в Северной Америке индейцы еще до Колумба ввели топинамбур и подсолнух.
Некоторые растения Старого Света, такие как конопля, сорго или яблоня
были введены в разное время и в разных регионах, так что
невозможно локализовать их исходное видообразование и
выращивание.
© Peter v. Sengbusch — Impressum
Архивы: Закон гомологических рядов в вариации (Н. И. Вавилов)
Журнальная статья Открытый доступ
Булатова Нина
Архивы: Закон гомологических рядов в вариации (Н.И. Вавилов)
Предварительный просмотр Файлы (1,2 МБ)| Имя | Размер | |
|---|---|---|
|
CCG_article_54511.pdf
md5: af11c8d6c5295b8a434bed0f416c994c  «/> «/>
|
1,2 МБ | Предварительный просмотр Скачать |
| Имя | |
|---|---|
|
XML-файл TaxPub
md5:20cf715300e69768e8e4624c6e324e07 |
Скачать |
Бета
ЦитатыГомоплазия в полногеномном анализе замен редких аминокислот: молекулярно-эволюционная основа закона гомологических рядов Вавилова | Biology Direct
Отчет рецензента 1
Алекс Кондрашов, Мичиганский университет, Анн-Арбор, США
Интересная бумага. Изучение гомоплазии некоторых из наименее склонных к гомоплазии признаков, связанных с белками, важно, потому что только признаки белков могут быть использованы для вывода о взаимоотношениях между отдаленными формами жизни. Связывание параллелизма в эволюции белков с гомологическими рядами Вавилова также имеет смысл, отчасти. И вклад в дебаты о Coelomata-Ecdysozoa также имеет смысл — хотя я не уверен, что этот вопрос действительно хочет быть решенным (что, если два последовательных события ветвления были разделены 10 млн лет назад или 1000 млн лет назад?).
Изучение гомоплазии некоторых из наименее склонных к гомоплазии признаков, связанных с белками, важно, потому что только признаки белков могут быть использованы для вывода о взаимоотношениях между отдаленными формами жизни. Связывание параллелизма в эволюции белков с гомологическими рядами Вавилова также имеет смысл, отчасти. И вклад в дебаты о Coelomata-Ecdysozoa также имеет смысл — хотя я не уверен, что этот вопрос действительно хочет быть решенным (что, если два последовательных события ветвления были разделены 10 млн лет назад или 1000 млн лет назад?).
Ответ автора: Последний пункт хорошо известен: загадка Coelomata-Ecdysozoa, подобно другим филогенетическим проблемам, относящимся к эпохам «Большого взрыва», может не особо хотеть сдаваться. интересный момент в эволюции эукариот для лучшего понимания потенциала и ограничений различных филогенетических методов .
У меня нет жестких критических замечаний, но есть несколько моментов, на которые стоит обратить внимание:
1. Редкие геномные изменения (RGC) — это, безусловно, правильный путь, пока у нас есть данные в масштабе генома, чтобы мы могли их наблюдать в достаточном количестве. Действительно, гомоплазия — злейший враг филогенетических реконструкций, и RGC менее склонны к ней.
Редкие геномные изменения (RGC) — это, безусловно, правильный путь, пока у нас есть данные в масштабе генома, чтобы мы могли их наблюдать в достаточном количестве. Действительно, гомоплазия — злейший враг филогенетических реконструкций, и RGC менее склонны к ней.
2. Я считаю, что разговоры о кладистике, синапоморфиях и маркерах Хеннига в контексте RGC могут ввести в заблуждение. Кладистика или нет, признаки без гомоплазии лучше всего подходят для филогенетики (Дарвин знал это), и если мы можем быть уверены, что эволюция была свободна от гомоплазии, самое экономное дерево будет правильным — см. книгу Фельзенштейна.Это верно даже в том случае, если мы не знаем, какие состояния RGC являются производными (укоренение дерева — отдельная проблема) — правильное неукорененное дерево по-прежнему остается максимально экономным. А в простейшем случае бинарных признаков очень легко увидеть, согласуются ли данные с отсутствием гомоплазии — это так, пока между признаками нет попарных конфликтов (конфликт между двумя признаками возникает, если все 4 комбинации их состояний – см. теорему о попарной совместимости в книге Фельзенштейна).
теорему о попарной совместимости в книге Фельзенштейна).
Ответ автора: Мы полагаем, что упоминание кладистики и производных признаков не введет читателя в заблуждение, и это, действительно, довольно традиционно в исследованиях, связанных с RGC. Читатель, заинтересованный в дальнейшем понимании, оценит этот комментарий .
3. Наиболее свободные от гомоплазии RGC связаны с крупномасштабными событиями – делециями, инсерциями и инверсиями (в частности, см. Chaisson M.J., Raphael B.J., and Pevzner P.A. Microinversions in mlinevolution.PNAS 2006 103: 19824–19829), потому что в этом случае маловероятно множественное происхождение одного и того же состояния. Замены (нуклеотидные или даже аминокислотные) не так хороши, потому что число возможных состояний признака (4 или 20) достаточно мало, чтобы обеспечить существенную гомоплазию. Тем не менее, виды замен, используемые авторами (RGC-CAM, 2- или 3-нуклеотидные аминокислотные замены в консервативных сайтах), наиболее близки к RGC среди всех замен. Их использование имеет смысл по двум причинам. Во-первых, изменения, связанные с некодирующими последовательностями, не подходят для сравнения отдаленных организмов, где такие последовательности слишком непохожи.Во-вторых, вставки в консервативных участках белков, вероятно, слишком редки даже на уровне всего протеома.
Их использование имеет смысл по двум причинам. Во-первых, изменения, связанные с некодирующими последовательностями, не подходят для сравнения отдаленных организмов, где такие последовательности слишком непохожи.Во-вторых, вставки в консервативных участках белков, вероятно, слишком редки даже на уровне всего протеома.
4. Что насчет конвергентной эволюции – поскольку для RGC-CAM существует более двух возможных состояний признака, конвергенция возможна.
Ответ автора
Трудно изучать конвергентные изменения для текущего набора данных, потому что плотность деревьев (рис. 1) недостаточна. Некоторые конвергентные изменения могут считаться параллельными изменениями .
5. Вопрос о гомологических рядах Вавилова понятен. Однако параллельная эволюция может объяснить лишь часть этого явления. Что еще более важно, сходные (не обязательно гомологичные; см. Wooding S. et al. Независимая эволюция чувствительности к горькому вкусу у людей и шимпанзе.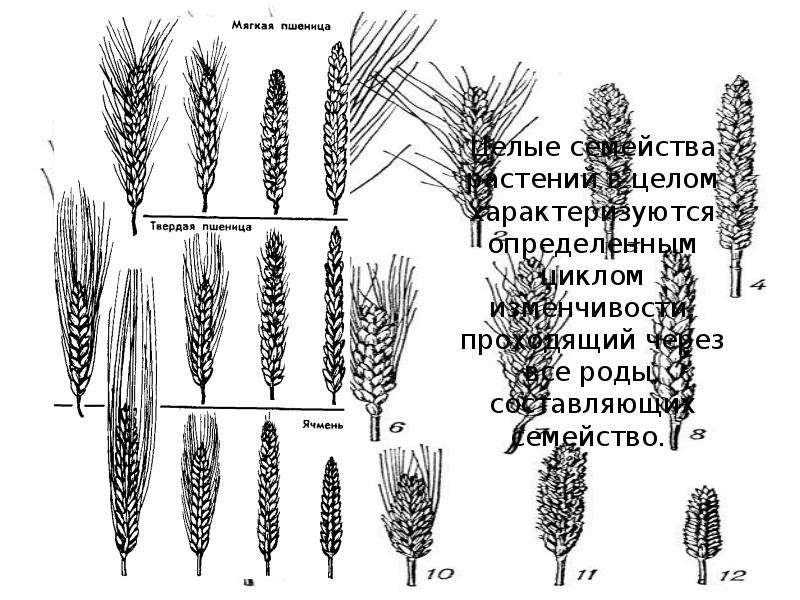 Nature 440, 930 – 934, 2006) мутации и полиморфизмы вызывают сходные («гомологичные») фенотипы у близкородственные виды (об этом говорится в книге Медникова; Медников Б.М. Закон гомологических рядов.К 60 -летию открытия закона Н. И. Вавиловым. Москва, Знание, 1980). Этот второй фактор, вероятно, является ключевой причиной гомологичной изменчивости внутри близкородственных видов.
Nature 440, 930 – 934, 2006) мутации и полиморфизмы вызывают сходные («гомологичные») фенотипы у близкородственные виды (об этом говорится в книге Медникова; Медников Б.М. Закон гомологических рядов.К 60 -летию открытия закона Н. И. Вавиловым. Москва, Знание, 1980). Этот второй фактор, вероятно, является ключевой причиной гомологичной изменчивости внутри близкородственных видов.
Ответ автора: Здесь предполагается, что наблюдаемые Вавиловым параллельные фенотипические изменения могли быть вызваны различными мутациями в ортологичных генах. Однако следует отметить, что в захватывающей статье Вудинга и др. вовлеченные мутации по-разному нарушают рассматриваемый ген.Это, действительно, могло быть достаточно распространенным и могло вызвать параллельное возникновение некоторых общих фенотипов. Однако объяснение более высокой распространенности параллельных мутаций у близкородственных видов, основанных на коварионной модели, как предлагается здесь, применимо ко всем видам мутаций, а не только к тем, которые нарушают функцию, и в этом отношении представляется значительно более общим . .
.
Отчет рецензента 2
Николя Галтье, CNRS, Университет Монпелье 2, Франция
Эта рукопись основана на наблюдении, что редкие геномные изменения (здесь аминокислотные замены, наблюдаемые только во внутренней группе, а не во внешних, и требующие замены двух или трех нуклеотидов), которые были отобраны из-за их предполагаемой очень малой вероятности появления дважды независимо друг от друга в ходе эволюции фактически обнаруживают значительную гомоплазию, так что порядок ветвления между членистоногими, нематодами и позвоночными до сих пор остается нерешенным.Здесь Рогозин и др. спросите, какие события множественных изменений повлияли на эволюцию этих персонажей и в каких линиях они произошли. Обнаружив больше гомоплазии в старых, чем в новых ветвях, авторы ссылаются на ковариационный процесс как на возможную причину этой неожиданной закономерности. Мне нравится это предложение, которое, я думаю, имеет смысл, и я желаю, чтобы мы получили дальнейшую поддержку в будущем. Наконец, авторы утверждают, что с поправкой на эти случаи гомоплазии гипотеза Coelomata (нематоды, (членистоногие, вторичноротые)) лучше всего подтверждается.
Наконец, авторы утверждают, что с поправкой на эти случаи гомоплазии гипотеза Coelomata (нематоды, (членистоногие, вторичноротые)) лучше всего подтверждается.
Я приветствую публикацию этой рукописи в Biology Direct, особенно потому, что она принадлежит к серии статей, анализирующих эти данные. Я надеюсь, что система обсуждения, предлагаемая Biology Direct, позволит сгруппировать аргументы, относящиеся к (важным) Coelomata/ Обсуждение Ecdysozoa в одном редакционном месте для удобства читателей.
Теперь технические комментарии к статье:
1. Выбор внешней группы
В этом исследовании опробованы все возможные комбинации восьми внешних групп, и сообщаются средние модели.Комбинациям внешних групп присваиваются равные априорные веса и неясные апостериорные веса — события параллелизма или обращения, поддерживаемые несколькими вариантами выбора внешней группы, учитываются один раз, так что комбинации, дающие аналогичные результаты, будут иметь заниженный вес по сравнению с комбинациями, поддерживающими уникальные события. Тем не менее ясно, что различные аутгруппы не имеют одинаковой ценности: одни гораздо ближе к ингруппе, чем другие, и поэтому более надежны. Как отмечают Иримия и Рой, игра с внешними группами значительно меняет картину в том, что касается спора о Coelomata/Ecdysozoa.Если, например, сосредоточить внимание на 6-й строке дополнительной таблицы 1, в которую включены только две ближайшие внешние группы (базальные многоклеточные Trichoplax и Nematostella), окажется, что (приблизительно оцененное) ожидаемое количество параллельных нематод/насекомых изменяется ( 5.8) значительно меньше, чем количество RGC_CAM, поддерживающих Ecdysozoa (18 против 7 для Coelomata), в отличие от средней картины (по внешним группам).
Тем не менее ясно, что различные аутгруппы не имеют одинаковой ценности: одни гораздо ближе к ингруппе, чем другие, и поэтому более надежны. Как отмечают Иримия и Рой, игра с внешними группами значительно меняет картину в том, что касается спора о Coelomata/Ecdysozoa.Если, например, сосредоточить внимание на 6-й строке дополнительной таблицы 1, в которую включены только две ближайшие внешние группы (базальные многоклеточные Trichoplax и Nematostella), окажется, что (приблизительно оцененное) ожидаемое количество параллельных нематод/насекомых изменяется ( 5.8) значительно меньше, чем количество RGC_CAM, поддерживающих Ecdysozoa (18 против 7 для Coelomata), в отличие от средней картины (по внешним группам).
Ответ автора: Использование только одного набора видов внешней группы (например,г. в цитируемой статье Irimia et al.) может ввести в заблуждение, поскольку количество общих RGC_CAM невелико и на него могут сильно повлиять статистические ошибки. Таким образом, к оценке рецензентом количества параллельных изменений следует относиться с большой осторожностью. При этом мы не можем исключить возможность того, что некоторые очевидные филогенетические сигналы в поддержку Ecdysozoa являются «реальными», т. е. из-за дупликации всего генома и последующей потери гена (Ref. [14]). См. также наш ответ на аналогичный комментарий рецензента №3 .
При этом мы не можем исключить возможность того, что некоторые очевидные филогенетические сигналы в поддержку Ecdysozoa являются «реальными», т. е. из-за дупликации всего генома и последующей потери гена (Ref. [14]). См. также наш ответ на аналогичный комментарий рецензента №3 .
2. Сосредоточение внимания на внутренних ветвях
Параллельные изменения и развороты в основном исследуются во внутренних ветвях. Только один раз авторы исследуют пару концевых ветвей Caenorhabditis/Mosquito для контроля. Я не уверен, почему. При выдвинутой в обсуждении интересной гипотезе, утверждающей, что параллельные изменения более вероятны в близкородственных линиях благодаря большему перекрытию их наборов вариабельных участков (коварионов), можно было бы ожидать высокой степени параллелизма у Apis/Aedes и Apis. /Anopheles пары терминальных ветвей, например.
Ответ автора: Рецензент предлагает анализ трех паттернов:
Am_ В : Am = Y, Dm = X, Ag = Y, Aa = Y, все другие виды = X.
Am_Ag: Am = Y, Dm = X, Ag = Y, Aa = X, все другие виды = X.
Am_Aa: Am = Y, Dm = X, Ag = X, Aa = Y, все остальные виды = X.
Предполагается, что параллельные изменения в ветви, ведущей к Am, и в другой ветви насекомых могут быть правдоподобным объяснением этих паттернов.Мы проанализировали эти паттерны:
Число RGC_CAM невелико, и поэтому его трудно интерпретировать, однако RGC_CA действительно сильно поддерживает высокую скорость параллельных изменений в кладе насекомых, как и предполагал рецензент. Необработанное количество RGC_CA для Am_In (62) в 4 раза больше, чем для In_ Dn ([16], см. Таблицу 1) , хотя длины ветвей почти идентичны (рис. 1 и результаты не показаны).Это согласуется с предсказанием, что параллельные изменения более вероятны в близкородственных линиях. Однако существует также потенциальная проблема с этим анализом, поскольку существуют альтернативные сценарии, которые могут объяснить наблюдаемые закономерности. Например, паттерн Am_In можно объяснить одной мутацией X->Y в ветви, ведущей от анализируемой трифуркации к насекомым, и другой мутацией Y->X в ветви, ведущей к Dm. Мы не можем оценить влияние таких альтернативных сценариев на данном этапе .
Например, паттерн Am_In можно объяснить одной мутацией X->Y в ветви, ведущей от анализируемой трифуркации к насекомым, и другой мутацией Y->X в ветви, ведущей к Dm. Мы не можем оценить влияние таких альтернативных сценариев на данном этапе .
3. Использование максимальной экономии (MP)
Я нашел несколько противоречивым использование MP для раскрытия истории этих персонажей, когда предпосылка исследования состоит в том, что гомоплазия существенна, т. Е. MP вводит в заблуждение. Если гомоплазия сильна, то истинный сценарий может отличаться от сценария MP, что усложняет задачу количественной оценки и локализации в дереве событий обращения или параллелизма. В частности, мне интересно, могут ли ошибки метода MP способствовать удивительной закономерности, о которой сообщается в этом исследовании, — более высокому количеству гомоплазии в старых ветвях.
Приведу пример параллельных изменений. Параллельные изменения вызываются, когда (i) две разные линии, скажем, A и B, имеют общее производное состояние Y, в то время как другие линии несут внешнегрупповое состояние X, и (ii) A и B разделены по крайней мере двумя внутренними ветвями в дереве. Это так, потому что, если A и B разделены одной внутренней ветвью (например, Apis и Drosophila), то модель также может быть объяснена инверсией ветвления между ними (комары). Это строгое депутатское обоснование.Теперь предположим, что эволюция не является строго экономной, и присвоим ненулевую вероятность субоптимальным сценариям. Если А и В разделены ровно двумя внутренними ветвями, то сценарий с двумя инверсиями (в двух линиях, разветвляющихся между А и В) требует всего на один шаг больше, чем сценарий МП. Итак, вы ожидаете, что определенная часть таких признаков будет ложно истолкована как старые параллелизмы, тогда как на самом деле они соответствуют молодым инверсиям. Вероятность того, что множественные инверсии вызовут паттерн, подобный параллелизму, уменьшается по мере увеличения топологического расстояния между A и B (поскольку количество требуемых инверсий увеличивается), имитируя ковариационный эффект.
Это так, потому что, если A и B разделены одной внутренней ветвью (например, Apis и Drosophila), то модель также может быть объяснена инверсией ветвления между ними (комары). Это строгое депутатское обоснование.Теперь предположим, что эволюция не является строго экономной, и присвоим ненулевую вероятность субоптимальным сценариям. Если А и В разделены ровно двумя внутренними ветвями, то сценарий с двумя инверсиями (в двух линиях, разветвляющихся между А и В) требует всего на один шаг больше, чем сценарий МП. Итак, вы ожидаете, что определенная часть таких признаков будет ложно истолкована как старые параллелизмы, тогда как на самом деле они соответствуют молодым инверсиям. Вероятность того, что множественные инверсии вызовут паттерн, подобный параллелизму, уменьшается по мере увеличения топологического расстояния между A и B (поскольку количество требуемых инверсий увеличивается), имитируя ковариационный эффект.
Вот как это можно перевести в отношении набора данных Metazoa. Предположим на мгновение, что (вторичноротые, (нематода, насекомое)) — это настоящее дерево (гипотеза Ecdysozoa).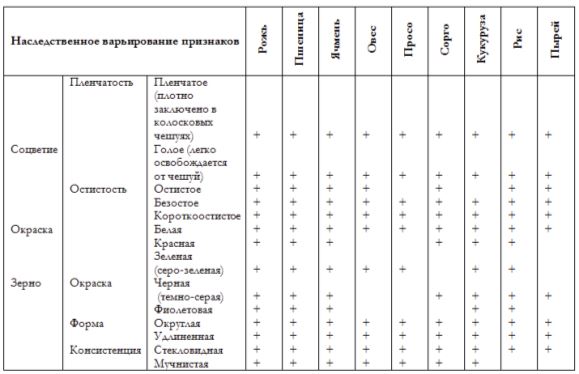 Паттерны, поддерживающие параллельное изменение N-Io, можно объяснить одним изменением X->Y у предка Ecdysozoa и двумя инверсиями Y->X: одним у Apis, одним у Brugia. Паттерны, поддерживающие параллельное изменение N-In, потребуют трех реверсий (у Apis, Drosophila и Brugia). Это может частично объяснить, почему последние наблюдаются реже, чем первые.
Паттерны, поддерживающие параллельное изменение N-Io, можно объяснить одним изменением X->Y у предка Ecdysozoa и двумя инверсиями Y->X: одним у Apis, одним у Brugia. Паттерны, поддерживающие параллельное изменение N-In, потребуют трех реверсий (у Apis, Drosophila и Brugia). Это может частично объяснить, почему последние наблюдаются реже, чем первые.
MP может быть введен в заблуждение многими способами, и я не хочу утверждать, что неэкономная эволюция всегда приводит к более старым реконструированным, чем реальные изменения — иногда вы также можете пропустить старые изменения. Я полагаю, однако, что при интерпретации этих цифр необходима некоторая осторожность — их не следует воспринимать как безошибочные наблюдения, и приветствуется тщательное изучение потенциальных погрешностей. Я отмечаю, что эти потенциальные погрешности могут распространяться на дальнейшие этапы анализа, когда длина ветвей, оцененная с помощью MP, используется для грубой количественной оценки ожидаемых закономерностей в ветвях стебля нематод, насекомых и вторичноротых.
Ответ автора: Любой филогенетический метод может ввести в заблуждение. Мы не думаем, что RGC_CAM в этом отношении отличаются от других методов. Однако, принимая во внимание, что RGC_CAMs являются редкими событиями, мы не ожидаем, что неэкономная эволюция будет столь же частой, как и для всех возможных аминокислотных замен (см. также новый Ref. [43]). Мы согласны с тем, что интерпретация RGC_CAMs требует некоторой осторожности ввиду использования экономии, и добавили к тексту следующее:
» Конечно, интерпретация этих результатов требует осторожности, как и любого результата, полученного с использованием принципа экономии.Мы сделали вывод о параллельных изменениях и/или инверсиях в эволюционных сценариях, которые, согласно максимальной экономии, требовали двух замен (рис. 3 и 6). Мы не можем исключить, что неэкономные сценарии, включающие более двух параллельных изменений и/или разворотов, могут оказать некоторое влияние на анализ RGC_CAs и RGC_CAMs. Однако ожидается, что RGC_CAM (и, в меньшей степени, RGC_CA) в значительной степени невосприимчивы к этой проблеме, потому что вероятность трех независимых редких событий намного меньше, чем двух независимых редких событий .»
Однако ожидается, что RGC_CAM (и, в меньшей степени, RGC_CA) в значительной степени невосприимчивы к этой проблеме, потому что вероятность трех независимых редких событий намного меньше, чем двух независимых редких событий .»
4. Выборка таксонов
В целом, у меня есть ощущение, что выборка таксонов ограничивает текущие исследования редких геномных изменений многоклеточных животных. Наш коллективный опыт в других группах/временных масштабах показывает, что методологические дебаты продолжались до тех пор, пока не был получен набор данных разумных Размер (по таксономии) был доступен, после чего возник консенсус. Это имело место, например, для млекопитающих. Могу поспорить, что добавление еще десяти геномов к этому набору данных решит основные вопросы, как топологические (Ecdysozoa или Coelomata?) и механистической (коварион-зависимая гомоплазия?).
Ответ автора: По нашему мнению, решающий аргумент в пользу клады грызунов-приматов был выдвинут Thomas et al.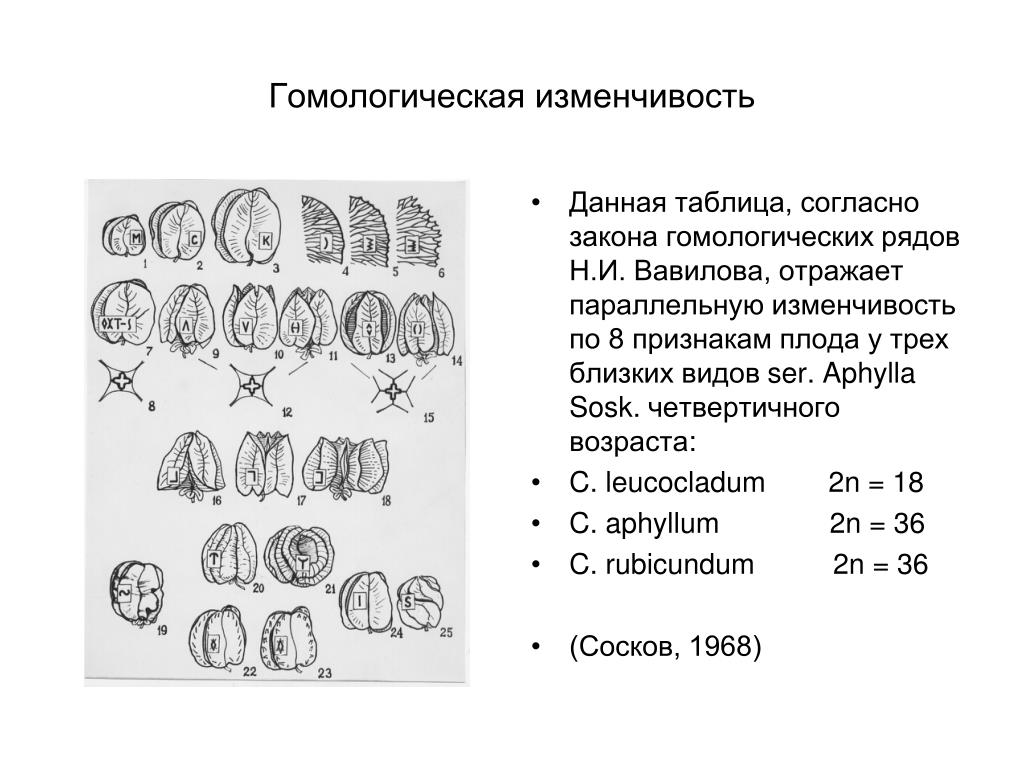 (Thomas JW, Touchman JW, Blakesley RW, et al. (71 соавтор). 2003. Сравнительный анализ многовидовых последовательностей из целевых областей генома. Nature. 424: 788–793) с использованием действительно необратимых RGC (вставки мобильных элементы) для относительно небольшого числа видов. В целом для истинно необратимых РГК количество видов не имеет значения.К сожалению, такие RGC недоступны для отдаленно родственных видов. Таким образом, если недавно секвенированные геномы не помогут решить проблему резких различий в скорости эволюции между нематодами, насекомыми и млекопитающими (рис. 1), мы не ожидаем, что добавление еще нескольких геномов действительно поможет решить проблему .
(Thomas JW, Touchman JW, Blakesley RW, et al. (71 соавтор). 2003. Сравнительный анализ многовидовых последовательностей из целевых областей генома. Nature. 424: 788–793) с использованием действительно необратимых RGC (вставки мобильных элементы) для относительно небольшого числа видов. В целом для истинно необратимых РГК количество видов не имеет значения.К сожалению, такие RGC недоступны для отдаленно родственных видов. Таким образом, если недавно секвенированные геномы не помогут решить проблему резких различий в скорости эволюции между нематодами, насекомыми и млекопитающими (рис. 1), мы не ожидаем, что добавление еще нескольких геномов действительно поможет решить проблему .
Отчет рецензента 3
Роберт Ланфир и Максимилиан Дж. Телфорд, Университетский колледж Лондона, Лондон, Великобритания (выдвинут Лоуренсом Херстом)
Очень интересная и провокационная газета.В нем есть два основных сообщения:
Во-первых, тщательное рассмотрение их базы данных RGC-CAM, по-видимому, поддерживает гипотезу Coelomata ((членистоногие, позвоночные) нематоды) по сравнению с широко принятой гипотезой Ecdysozoa ((членистоногие, нематоды) позвоночные).
Во-вторых, количество параллельных изменений в сестринских таксонах гораздо чаще наблюдается вблизи точки их дивергенции, чем позже, намного позже их дивергенции. Предполагается, что это происходит из-за эффектов ковариационной модели, которая предполагает, что сходные гены (в сходном генетическом фоне?) будут вынуждены подвергаться сходным заменам, и это будет менее верно для более непохожих генов.
У нас есть ряд моментов, которые, по нашему мнению, необходимо решить.
В соответствии с анализом Irimia, авторы приложили некоторые усилия, чтобы включить разнообразие внешних групп. Они анализируют влияние всех возможных комбинаций этих внешних групп и подсчитывают, сколько раз их RGC-CAM поддерживали Coelomata и Ecdysozoa. Их вывод состоит в том, что существует очень небольшая разница в поддержке Ceolomata и Ecdysozoa.
Однако мы знаем, что некоторые внешние группы (близкие) априори должны превосходить более дальние.Из их дополнительного файла 1 мы извлекли эксперименты, которые содержат 1 или обе близкие внешние группы (т. е. многоклеточные животные Trichoplax и Nematostella ), и сравнили их с экспериментами без внешних групп многоклеточных животных. В оптимальных случаях (включая обе внешние группы многоклеточных) 34 из 64 экспериментов подтверждают Ecdysozoa и 0 поддерживают Coelomata. В целом, включая по крайней мере один анализ внешней группы метаозоев, 88 анализов (из 192) поддерживают Ecdysozoa и 22 поддерживают Coelomata. Поддержка Ecdysozoa ниже только для случаев Trichoplax и выше для случаев только Nematostella .В худших случаях (ни одно из многоклеточных животных в анализе) ВСЕ анализы подтверждают Coelomata. Для нас это говорит о том, что, по крайней мере, при отсутствии каких-либо поправок на длину ветвей, их набор данных поддерживает Ecdysozoa.
е. многоклеточные животные Trichoplax и Nematostella ), и сравнили их с экспериментами без внешних групп многоклеточных животных. В оптимальных случаях (включая обе внешние группы многоклеточных) 34 из 64 экспериментов подтверждают Ecdysozoa и 0 поддерживают Coelomata. В целом, включая по крайней мере один анализ внешней группы метаозоев, 88 анализов (из 192) поддерживают Ecdysozoa и 22 поддерживают Coelomata. Поддержка Ecdysozoa ниже только для случаев Trichoplax и выше для случаев только Nematostella .В худших случаях (ни одно из многоклеточных животных в анализе) ВСЕ анализы подтверждают Coelomata. Для нас это говорит о том, что, по крайней мере, при отсутствии каких-либо поправок на длину ветвей, их набор данных поддерживает Ecdysozoa.
Ответ автора
1) Корректировка длины ветвей абсолютно необходима. Это рассматривается более подробно в Ref. [21].
2) Рецензенты считают, что Nematostella a priori является высшей внешней группой. Однако внутренняя ветвь между Nematostella и анализируемой трифуркацией чрезвычайно короткая (0 RGC_CAMs, , рис. 1), , поэтому Nematostella находится слишком близко к трифуркации, чтобы быть надежной внешней группой. Например, если была полная дупликация генома и дифференциальные потери генов, Nematostella становится неподходящей внешней группой, которая не допускает правильной поляризации RGC_CAMs .
Однако внутренняя ветвь между Nematostella и анализируемой трифуркацией чрезвычайно короткая (0 RGC_CAMs, , рис. 1), , поэтому Nematostella находится слишком близко к трифуркации, чтобы быть надежной внешней группой. Например, если была полная дупликация генома и дифференциальные потери генов, Nematostella становится неподходящей внешней группой, которая не допускает правильной поляризации RGC_CAMs .
Результаты, подтверждающие ковариионную модель, поразительны и очень интересны. Мы считаем, что эту идею можно было бы лучше проверить на наборе данных, который имеет менее противоречивую филогению и с хорошими ограничениями на длину ветвей / время расхождения.В частности, кажется вероятным, что часто очень маленькие длины ветвей, оцениваемые с использованием подхода RGC_CAM, могут быть особенно подвержены ошибкам. Возможно, на самом деле мы имеем в виду, что данные текущего анализа нуждаются в дальнейшем тестировании на наборе данных с меньшим количеством свободных параметров.
Ответ автора: Мы, безусловно, ценим это замечание и вполне можем вернуться к этому вопросу с другими наборами данных. Тем временем мы добавили дополнительный анализ параллельных изменений в кладе насекомых в наш ответ рецензенту № 2; выводы сильно армированы .
Ранее авторы использовали статистические тесты для сравнения поддержки гипотез Coelomata и Ecdysozoa. Эти статистические тесты были основаны на предположении, что «RGC_CAM внутри гена развиваются независимо друг от друга» (Ref. [14]). Мы обеспокоены тем, что если текущая статья верна и применима модель ковариации, то это предположение не выполняется. Если это так, то предыдущие проверки между гипотезами могут быть недействительными. Не могли бы авторы включить какое-то обсуждение этого пункта?
Ответ автора: Во-первых, кажется, что если существование коварионов нарушает предположение о независимости, это повлияет на любые статистические модели и тесты, используемые в филогенетическом анализе, а не только на те, которые применялись в наших предыдущих исследованиях RGC_CAM. Во-вторых, далеко не очевидно, что если применима ковариионная модель, то предположение о независимости сайтов — нет. Нет оснований полагать, что замены внутри набора ковариаций, существующих в данное время, не являются независимыми (хотя никакие замены не допускаются за пределами набора ковариаций), и поскольку набор ковариаций быстро меняется в процессе эволюции, проходя через белковую последовательность, весь сайт независимость кажется разумным предположением. Конечно, желательна строгая проверка ковариационной модели и ее влияния на независимость сайтов, и мы надеемся, что эта статья может привлечь некоторое дополнительное внимание к изучению коварионов, которое в настоящее время не так популярно, как этот предмет, возможно, заслуживает . .
Во-вторых, далеко не очевидно, что если применима ковариионная модель, то предположение о независимости сайтов — нет. Нет оснований полагать, что замены внутри набора ковариаций, существующих в данное время, не являются независимыми (хотя никакие замены не допускаются за пределами набора ковариаций), и поскольку набор ковариаций быстро меняется в процессе эволюции, проходя через белковую последовательность, весь сайт независимость кажется разумным предположением. Конечно, желательна строгая проверка ковариационной модели и ее влияния на независимость сайтов, и мы надеемся, что эта статья может привлечь некоторое дополнительное внимание к изучению коварионов, которое в настоящее время не так популярно, как этот предмет, возможно, заслуживает . .
«Анализ RGC_CAM был объединен с несколькими статистическими тестами конкурирующих филогенетических гипотез, и было показано, что он устойчив к различиям в длине ветвей и выборке таксонов в широком диапазоне вариаций [14]»
Мы утверждаем, что недавние исследования показывают что подход RGC_CAM на самом деле очень чувствителен к выборке таксонов — как в недавней критике подхода, так и в текущем ответе авторов на эту критику выборка таксонов оказалась фундаментальной проблемой, поскольку разные наборы таксонов дают значительно разные ответы на Coelomata. /Ecdysozoa дебаты.Наш повторный анализ текущего набора данных также поддерживает мнение о том, что выборка таксонов имеет фундаментальное значение для выводов, сделанных с использованием этого подхода.
/Ecdysozoa дебаты.Наш повторный анализ текущего набора данных также поддерживает мнение о том, что выборка таксонов имеет фундаментальное значение для выводов, сделанных с использованием этого подхода.
Ответ автора: Мы полностью согласны с рецензентами в отношении важности выборки таксонов. Поэтому здесь и в Ref. [21], , мы использовали все комбинации из большого набора видов внешней группы. Мы считаем, что это помогает получить надежные результаты .
Стр. 6:
«Более того, в настоящее время в сообществе эво-дево предпочтение отдается топологии экдизозоя, на основании предполагаемых глубоких общностей в процессах развития различных линяющих животных [30, 31]»
Это введение в заблуждение сообщества evodevo.На самом деле нет «ощущаемой глубокой общности» в основе развития линьки — просто линька — это единственное, что, по-видимому, объединяет экдизозойных животных, и, как таковая, это скорее предполагаемая, чем предполагаемая общность.
Ответ автора: Если так говорят некоторые лидеры сообщества эво-дево… исправлено, как было предложено .
Стр. 7:
«Однако, как показано ранее, если принять во внимание длину ветвей, поддержка целоматной клады становится значительно больше, чем у Ecdysozoa [21].»
Можно это уточнить. Неясно, исправили ли авторы длину ветвей в настоящем анализе, или они просто ссылаются на свой предыдущий анализ, в котором они сделали поправку на длину ветвей (но использовали более ограниченный набор данных). если авторы сделали поправку на длину ветвей, можно ли уточнить приведенное выше предложение, чтобы указать количество тестов, в которых гипотеза Coelomata получила значительную поддержку («существенно больше» немного двусмысленно — имеет ли это какое-то отношение к значимости?), и число, в котором Ecdysozoa получают значительную поддержку.
Ответ автора: В приведенном выше предложении мы ссылаемся на предыдущие результаты, где значение слова «существенный» является явным.
