
Какое животное — самый смертоносный охотник на Земле?
- Элла Дейвис
- BBC Earth
Автор фото, Thinkstock
Хищные животные зачастую возвращаются с охоты не солоно хлебавши. Но некоторые виды ставят рекорды результативности при поимке добычи, обнаружил корреспондент BBC Earth.
Владельцы кошек, регулярно таскающих в дом придушенные «гостинцы», конечно, станут утверждать, что у самого смертоносного хищника на Земле четыре лапы и усатая мордочка.
С этим мнением согласились бы и многочисленные жертвы Барсиков и Мурок: по оценке британского Общества изучения млекопитающих (Mammal Society), 9 млн домашних кошек Великобритании ежегодно приносят своим хозяевам 92 млн умерщвленных ими мелких животных, включая 27 млн птиц.
При такой результативности котиков их более крупные и опасные сородичи, наподобие тигра или леопарда, несомненно, должны быть самыми успешными хищниками в мире.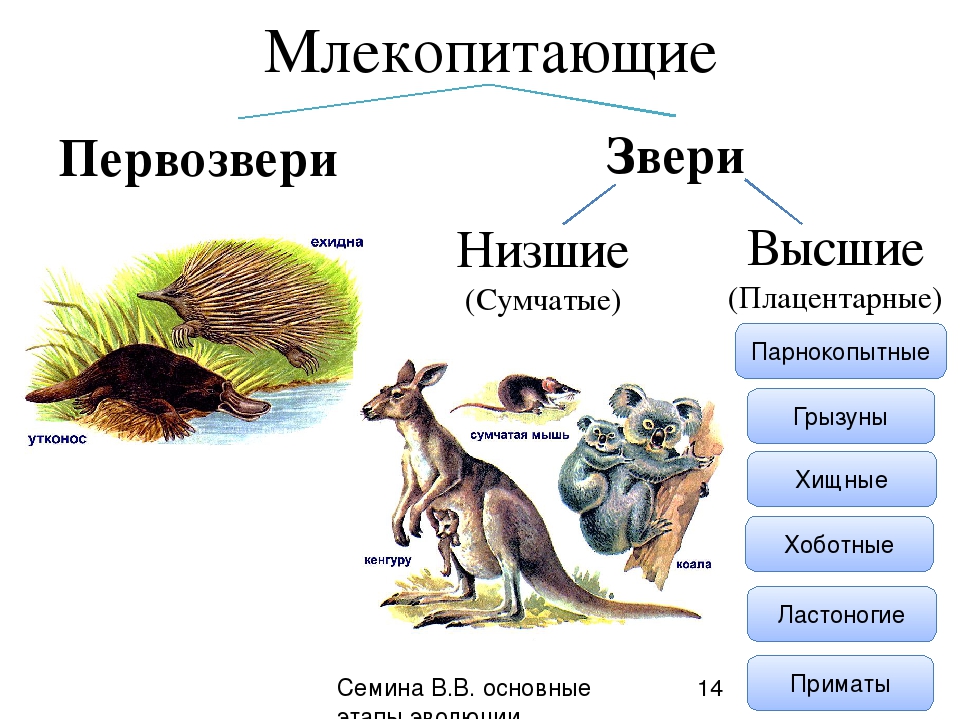
Несомненно, крупные представители семейства кошачьих — превосходные охотники. Это суперхищники с ужасными, мощными челюстями.
Тигры отлично плавают, а леопарды прекрасно лазают по деревьям, так что сложный рельеф местности во многих случаях не является для них препятствием при погоне за добычей.
Тем не менее, у леопардов, обитающих в южноафриканском Национальном парке Крюгера, в шести случаях из семи охота оканчивается неудачей. Тиграм удается поймать добычу еще реже.
Автор фото, Thinkstock
Подпись к фото,Тигры — отличные охотники, но далеко не лучшие
По оценке американского этолога Джорджа Шаллера, изучавшего бенгальских тигров, лишь в одном случае из 20-ти этот большой кот настигает и убивает свою жертву.
Данные основаны на наблюдениях и, разумеется, значительно разнятся в зависимости от количества дичи в данном районе и от опытности хищника. Скрытность кошачьих также весьма затрудняет изучение их охотничьих повадок.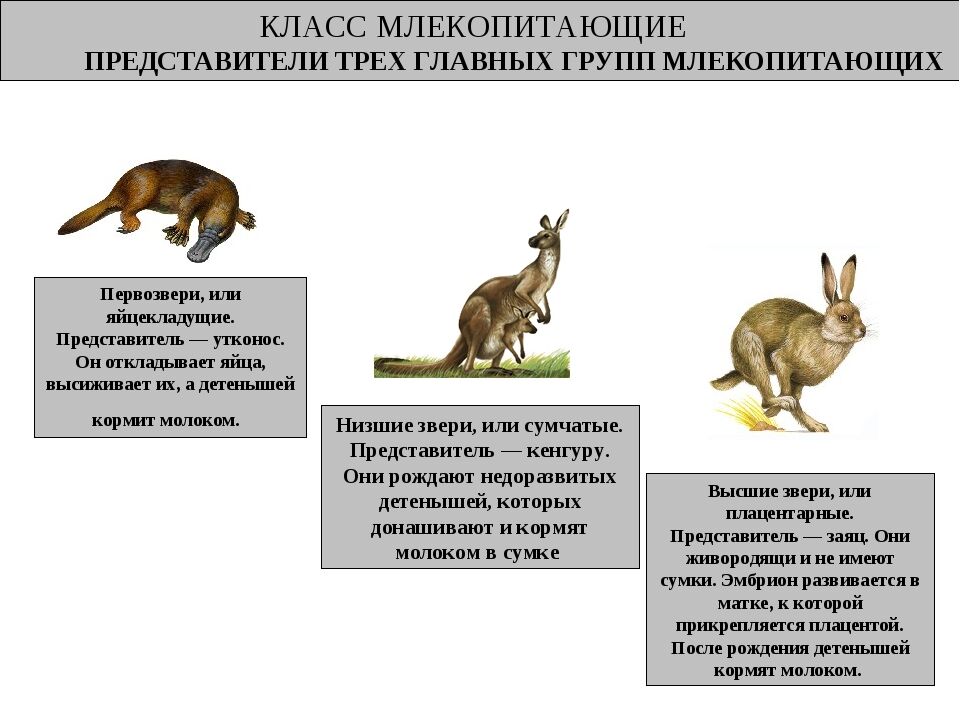
Несмотря на знаменитый пятнистый или полосатый камуфляж, крупным диким кошкам не всегда удается незаметно подобраться к добыче, поэтому при охоте они полагаются на элемент неожиданности — передвигаются тихо и скрытно, устраивают засады и используют ландшафт для маскировки.
Для пущей незаметности хищники семейства кошачьих охотятся ночью, но полная луна может их демаскировать.
Гепард, развивающий скорость до 93 км/ч, обладает несомненным преимуществом при погонях на равнинной местности. Но даже у гепардов успешна лишь каждая вторая охота.
Автор фото, Thinkstock
Подпись к фото,Убежать от гепарда трудно, и тем не менее только каждая вторая охота у него удачная
Командная работа повышает шансы на успех, как видно на примере львиных прайдов. Львицы, охотящиеся группой или в паре, в среднем вдвое больше результативны, чем добывающие пищу в одиночку.
Тем не менее, лишь в 30% случаев групповая охота львов бывает удачной.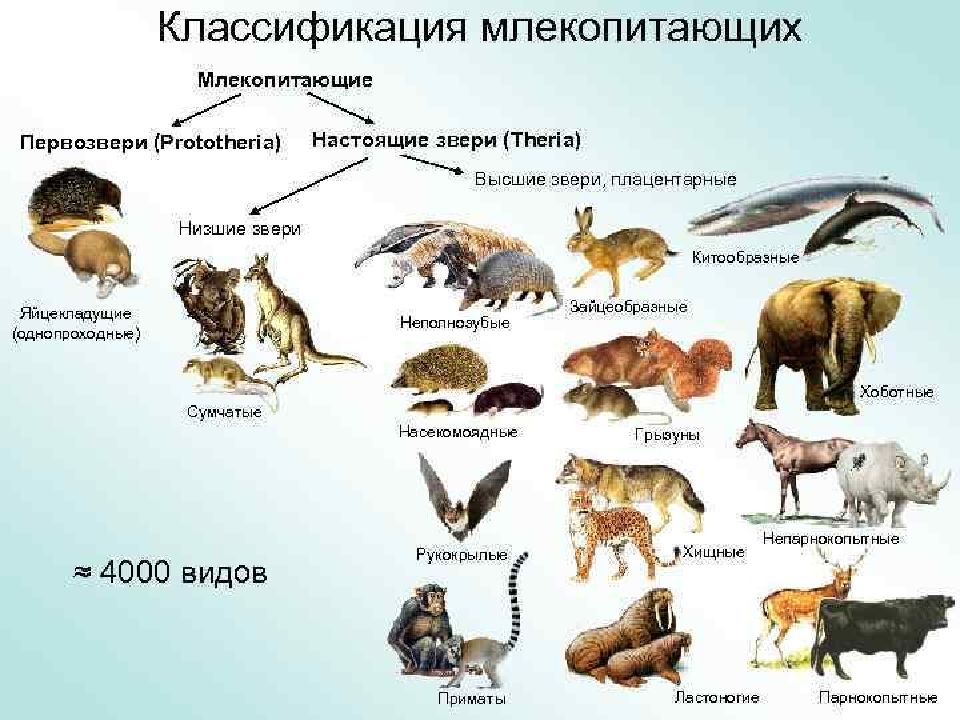
Охота стаей лучше получается у семейства псовых. Гиеновидные собаки, например, повышают шансы на успех до 67%, сбившись в стаю, насчитывающую около 20 особей. При этом они могут нападать на добычу, вдвое превышающую их собственные размеры.
Стая обыкновенных волков способна загнать и загрызть гигантского американского бизона весом до 900 кг.
Такой результат достигается благодаря эффективной командной работе: во время погони волки передают друг другу эстафету до тех пор, пока жертва окончательно не выдохнется — после чего стая атакует ее.
Автор фото, Thinkstock
Подпись к фото,Муравьи берут количеством — на охоте это помогает
Однако вся эта беготня выматывает и псовых. Они охотятся лишь один или два раза в сутки, при этом добытая пища делится на всех членов стаи.
По-своему впечатляющим являются ежесуточные объемы добычи гораздо более мелкого охотника — кочевого муравья.
Эти маленькие хищники также прибегают к помощи сородичей. Колония кочевых муравьев способна поймать за сутки до 30 000 тысяч насекомых.
Колония кочевых муравьев способна поймать за сутки до 30 000 тысяч насекомых.
Впрочем, нужно учесть, что колония может насчитывать до полумиллиона муравьев.
С точки зрения результативности самый удачливый хищник на Земле принадлежит к миру насекомых.
В 2012 г. исследователи из американского Гарвардского университета выяснили, что стрекозы ловят до 95% всей добычи, за которой охотятся.
Такой высокий результат объясняется тем, что стрекозы в процессе эволюции приобрели ряд адаптивных черт — включая сложное строение глаза, позволяющее видеть потенциальную жертву на фоне неба.
Крылья стрекозы приводятся в движение отдельными группами мышц, скоординированной работе которых насекомое обязано своими поразительными скоростью и маневренностью.
А вот нейробиолог Энтони Леонардо объясняет успешность охоты стрекоз особенностями их мозга.
«Мозг стрекозы задействует алгоритмы оптимизации, позволяющие насекомому просчитывать траекторию движения добычи и выдающие мышцам команды, облегчающие перехват цели»,- говорит он.
Автор фото, Thinkstock
Подпись к фото,Устройство стрекозы способствует успешности на охоте
«Стрекозы не особо разборчивы в еде — они охотятся на любую крылатую добычу, размеры которой примерно равны размерам стрекозиной головы, включая пчел, мотыльков и мух. По большей части стрекозы питаются мелкими насекомыми, такими как комары и мошки, но крупные виды могут даже ловить и поедать других стрекоз».
Леонардо продолжает: «У себя в лаборатории мы кормим стрекоз исключительно плодовыми мушками, поймать которых труднее, чем более представителей более распространенных в природе видов насекомых. Наши стрекозы ловят добычу примерно в 80% случаев. Все равно это весьма впечатляющий результат, поскольку плодовые мушки летают с огромной скоростью — примерно 1 м в секунду».
В лаборатории Леонардо, расположенной в Медицинском институте Говарда Хьюза, исследователи заняты чрезвычайно сложной и кропотливой работой — они изучают поведение насекомых в полете. На спину подопытным стрекозам прикрепляют миниатюрные «ранцы».
На спину подопытным стрекозам прикрепляют миниатюрные «ранцы».
«Эти крохотные устройства позволяют нам вести запись сигналов, посылаемых нейронами мышцам, отвечающим за направление движения и работу крыльев стрекозы во время охоты», — объясняет Леонардо.
Цель исследования — выяснить, каким образом мозг стрекозы обрабатывает информацию об окружающем пространстве, просчитывает траекторию движения цели и выдает соответствующие команды мышцам.
Впрочем, на титул наиболее успешного охотника имеется еще один претендент. Хоть и не всем из нас он покажется подходящим кандидатом.
Синие киты — крупнейшие из когда-либо населявших Землю животных. Они достигают 34 м в длину, что почти равняется длине крупного пассажирского самолета.
Такому гигантскому существу необходимо соответствующее питание: кит поедает до 4 тонн пищи в сутки.
Автор фото, Thinkstock
Подпись к фото,Синий кит (Balaenoptera musculus).
Излюбленное лакомство синих китов — мелкие планктонные ракообразные, известные под собирательным названием криль. Чтобы набрать необходимое количество калорий, один кит съедает 40 млн таких рачков в сутки.
Если же вы полагаете, что черпание морепродуктов широко разинутым ртом настоящей охотой не является, вы сильно ошибаетесь.
Только для того чтобы раскрыть гигантские челюсти, киту требуется израсходовать столько энергии, что он делает это лишь при обнаружении большого косяка криля.
В течение лета синие киты прочесывают океан в поисках скоплений криля, чтобы запастись энергией на голодную зиму.
Таким образом, итоговый выбор, кого считать самым смертоносным хищником, зависит от того, что мы вкладываем в понятие «смертоносный».
Картина стаи волков, загоняющих огромного бизона, несомненно, производит на зрителей большое впечатление. Не менее впечатляет и высокий показатель успешности на охоте.
С точки зрения людей, одновременное поедание миллионов живых существ делает весьма смертоносным и кита, хотя для него такое событие — не более чем обед.
Для всех вышеперечисленных диких хищников охота — вопрос выживания.
Домашних же кошек кормят, за ними ухаживают; их тяга к охоте на мышей и птиц объясняется остаточным инстинктом.
Осуждающим домашних питомцев за убийства невинных животных стоило бы задуматься о некоторых параллелях в поведении Барсика и Человека разумного.
Образовательный плакат «Вирус папилломы человека» (ВПЧ)
Вирусы папилломы (papilloma viruses) заражают клетки кожи и слизистых оболочек — кератиноциты, которые служат основным механическим барьером между внешней средой и внутренней средой организма. Эта группа вирусов очень разнообразна, и многие ее представители не вызывают никаких симптомов. Однако патогенные представители могут приводить не только к образованию папиллом и бородавок, но и к злокачественным опухолям.
История и распространенность ВПЧ
Бородавки и кожные заболевания упоминаются и описываются начиная с самых древних литературных источников. Первые подробные описания кожных новообразований составил древнеримский естествоиспытатель и врач Авл Корнелий Цельс, живший на рубеже старой и новой эры [7]. Предположения о том, что некоторые бородавки могут передаваться инфекционными агентами возникло еще раньше. Показать, что этими агентами являются вирусы удалось в начале ХХвека (обзорная информация по ссылке 4). Первый вирус папилломы был выделен в 1933 Ричардом Шоупом [8], американским вирусологом, который также впервые выделил и вирус гриппа [9].
Папилломавирусы втречаются у млекопитающих и существенно менее распространены среди птиц. Эволюционная история папилломавирусов началась тогда, когда появились первые высшие позвоночные (амниоты, к которым относятся пресмыкающиеся, птицы и млекопитающие) [10]. Более подходящими хозяевами для этой группы вирусов, в виду строения кожного покрова, оказались млекопитающие, у которых они представлены множеством подсемейств и родов [11].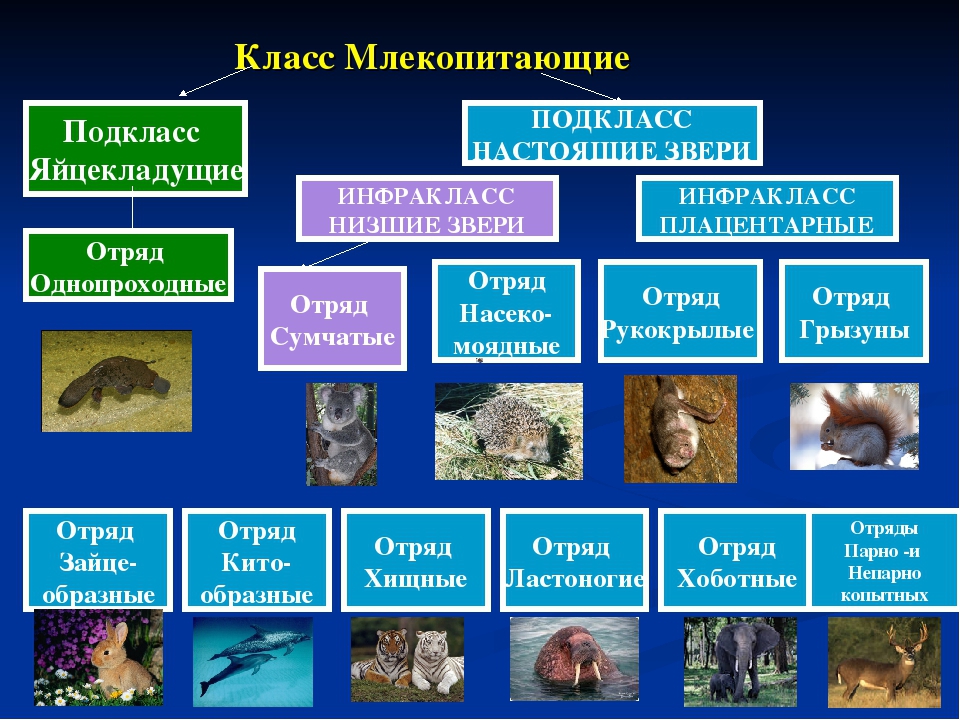
К злокачественным новообразованиям (в это касается рака шейки матки и аногенитальной зоны) приводят всего 18 штаммов. Передаются вирусы папилломы человека(ВПЧ) в основном половым путем и при контакте слизистых.
Считается, что ВПЧ низкого риска продуцируют больше вирионов, лучше распространяются, хотя и элиминируются из организма за один-два года, или даже быстрее [13]. По сравнению с ними, штаммы высокого риска заражают меньшее количество людей, но и иммунной системе с ними справиться сложнее [14]. Самыми канцерогенными являются штаммы ВПЧ 16 и ВПЧ 18, а за возникновение бородавок в аногенитальной зоне в 90% случаев ответственны штаммы ВПЧ 6 и ВПЧ 11 [15]. Именно эти представители папилломавирусов привлекают наибольшее внимание исследователей [16] [17].
Именно эти представители папилломавирусов привлекают наибольшее внимание исследователей [16] [17].
Строение и биология вируса
Папилломавирусы имеют сравнительно маленькие вирионы, лишенные мембранной оболочки. Их диаметр около 30 нм. Это примерно в четыре раза меньше диаметра ВИЧ или гриппа, имеющих мембранную оболочку, происходящую из мембраны зараженной клетки. Геном папилломавирусов представлен кольцевой двуцепочечной молекулой ДНК, которая упакована при помощи гистонов — клеточных белков, участвующих в компактизации ДНК в ядре. Геном папилломавирусов кодирует два типа белков. Ранние белки (early proteins, E-proteins) осуществляют регуляторные функции, обеспечивают размножение вируса и, в ряде случаев, ответственны за злокачественное перерождение клеток. Поздние белки (late proteins, L-proteins) выполняют структурную функцию, формируя капсид вирионов.
Папилломавирусы непросто культивировать в лабораторных условиях, так как нативные частицы образуются либо в живых организмах, либо в сложных органотипических клеточных культурах, в которых клетки проходят стадии дифференцировки, аналогичные существующим в живом организме [18, 19 и 20].
Активно делящиеся молодые кератиноциты выстилают базальную мембрану, находящуюся под верхними слоями эпидермиса, а по мере созревания и дифференцировки кератиноциты вытесняются к поверхности кожи. Вирусные частицы заражают еще не дифференциированные клетки, а новые частицы образуются в кератиноцитах, достигших терминальной стадии дифференцировки.
Ранние белки папилломавирусов обеспечивают нужное содержание ДНК вируса в ядре клетки и координируют экспрессию его генов. Белки E1 и E2 образуют комплекс с ДНК вируса, привлекая клеточные системы репликации. За канцерогенный эффект у штаммов ВПЧ высокого риска ответственны ранние белки E6 и E7. E6 способен связываться с опухолевым супрессором р53, провоцируя его убиквитинизацию и уничтожение [21]. E7 у таких штаммов связывается и влияет на работу целого ряда белков, среди которых регуляторные белки и опухолевые супрессоры, такие как ретинобластомный фактор. Активность E6 и E7 приводит к неконтролируемому клеточному делению [22 и 18].
Активность E6 и E7 приводит к неконтролируемому клеточному делению [22 и 18].
Поздние белки ВПЧ нужны для образования капсида и упаковки ДНК вируса. Основным структурным компонентом капсида папилломавирусов является белок L1, который образует пентамеры [23]. В состав одного капсида входит 72 пентамера. Капсид папилломавирусов имеет сферическую форму, однако в действительности он представляет собой икосаэдр с триангуляционным числом равным семи. Сферическая структура образуется не из комбинации гексамеров и пентамеров (как, например, в футбольном мяче), а из двух типов пентамеров. 60 пентамеров капсида ВПЧ образуют по 6 связей с соседними пентамерами, а 12 из них — по пять [2, глава 3]. Интересно, что строение L1 напоминает строение клеточных белков нуклеоплазминов, регулирующих упаковку нуклеосом [24]. На данный момент нет точного понимания, имеют ли L1 и нуклеоплазмины общего предка, или их схожесть результат конвергентной эволюции.
Минорный белок L2 также является структурным компонентом вирусной частицы.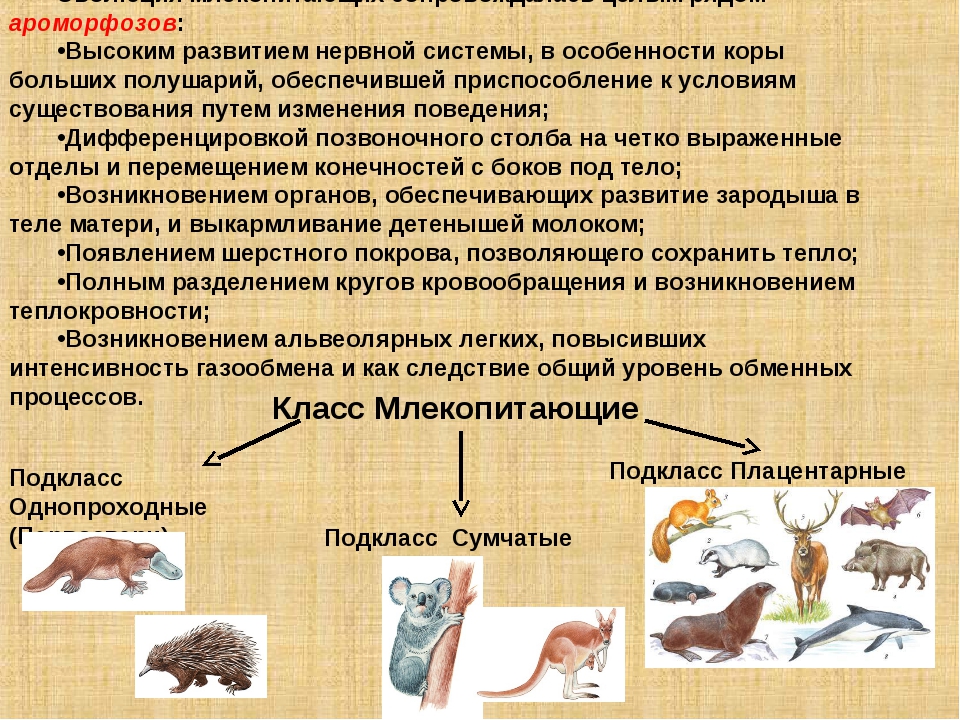 Один L2 приходится на один пентамер L1 [4]. По всей видимости, это белок важен для привлечения вирусных ДНК к собирающимся частицам. Существует гипотеза, что L1 и L2 могут взаимодействовать не непосредственно с нуклеиновой кислотой вируса, а с гистонами, которые ее компактизуют [4].
Один L2 приходится на один пентамер L1 [4]. По всей видимости, это белок важен для привлечения вирусных ДНК к собирающимся частицам. Существует гипотеза, что L1 и L2 могут взаимодействовать не непосредственно с нуклеиновой кислотой вируса, а с гистонами, которые ее компактизуют [4].
Каким конкретно образом ДНК уложена внутри вирсуной частицы, тоже не описано окончательно. Известно что в вирусоподобные частицы ВПЧ может быть упакована любая ДНК короче 8 тысяч пар нуклеотидов, что открывет перспективы для использовния ВПЧ в качестве трансформационных векторов. Интересно, что в распаковке капсида папилломавирусов участвует циклофилин — клеточный белок, также способствующий распаковке капсида, например ВИЧ [25].
Вакцины против ВПЧ
В настоящее время активно разрабатываются и внедряются вакцины против папилломавирусов. В основном они создаются против наиболее распространенных канцерогенных штаммов ВПЧ 16 и ВПЧ 18, а зтакже против ВПЧ 6 и ВПЧ 11, основной причины появления бородавок.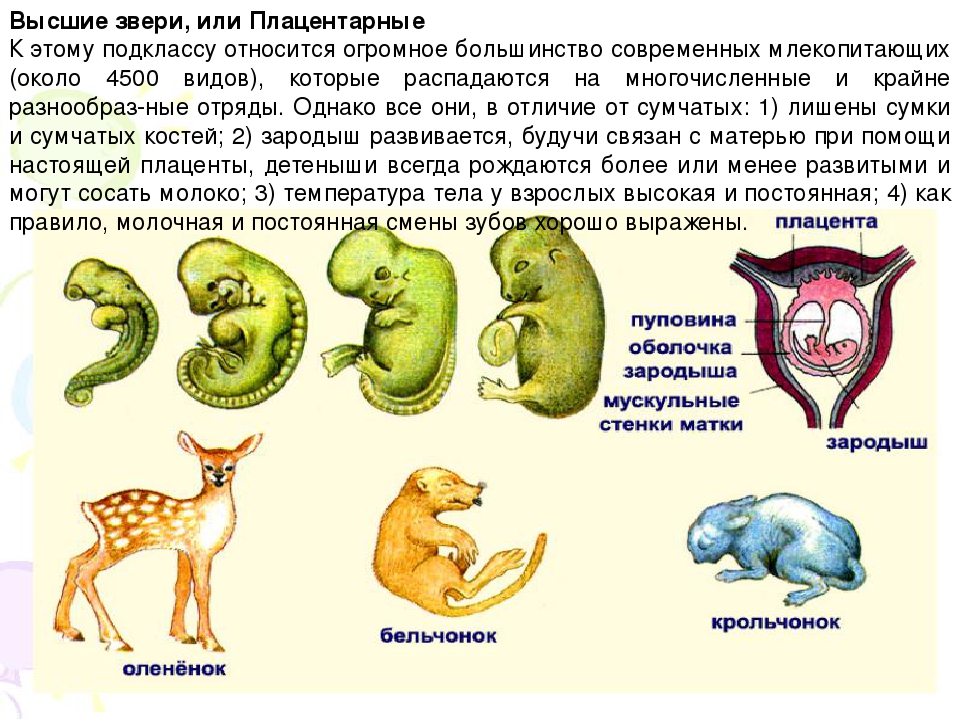
Имеющиеся вакцины от вирусов папилломы человека — это триумф прикладной структурной вирусологии. Однако, доступные вакцины, сделанные на основе рекомбинантных вирусоподобных частиц, собранных из белка основного капсидного L1, не защищают от всех типов патогенных ВПЧ. К счастью, новое поколение вакцин от ВПЧ, нацеленных на «Ахиллесову пяту» вируса — консервативные эпитопы, присутствующие на минорном белке L2, обещает предоставить широкую защиту от всех ВПЧ, включая все папилломавирусы, вызывающие рак, также как и вирусы, вызывающие доброкачественные кожные новообразования. Имеющиеся знания о структуре, динамике и функциях L2 все еще весьма ограничены. Эта информация крайне необходима для разработки универсальной вакцины от ВПЧ.
Доктор Кристофер Бак. Национальный института рака, США
Интерес к разработке вакцин был подогрет тем, что исследование связи папилломовирусов с раком шейки матки в 2008 году было удостоено Нобелевской премии в области физиологии и медицины. Немецкий ученый Харальд зур Хаузен (Harald zur Hausen) показал, что папилломавирусы являются причиной практически всех случаев рака шейки матки, второго по распространенности рака у женщин [1].
Немецкий ученый Харальд зур Хаузен (Harald zur Hausen) показал, что папилломавирусы являются причиной практически всех случаев рака шейки матки, второго по распространенности рака у женщин [1].
Интересно, что зур Хаузен исследовал среди прочего клетки культуры HeLa — первые человеческие клетки, которые удалось культивировать в 1951 году группе американских ученых [26]. Источником этой важнейшей клеточной линии стали клетки злокачественной опухоли американской гражданки Генриэтты Лакс, страдавшей раком шейки матки. Зур Хаузену удалось получить доступ к результатам биопсии Генриэтты и показать что женщина была заражена сразу несколькими разновидностями ВПЧ 18 одновременно, что и стало причиной возникновения у нее рака шейки матки. Эти важные исследования, а также история получения самой распространенной культуры человеческих клеток описаны в популярной форме в книге Ребекки Склут «Бессмертная жизнь Генриеэтты Лакс».
Показать ссылки
-
zur Hausen H.
 , Virology. 2009 Feb 20;384(2):260-5. Epub 2009 Jan 8.
, Virology. 2009 Feb 20;384(2):260-5. Epub 2009 Jan 8.
- Rossmann W.G. and Rao V.B. ed., ISBN-10: 1461409799; ISBN-13: 978-1461409793; Dec 26, 2011
- Belnap D.M., Olson N.H. et al., J Mol Biol. 1996 Jun 7;259(2):249-63.
-
Buck C.
 B. and Trus B.L., Adv Exp Med Biol. 2012;726:403-22.
B. and Trus B.L., Adv Exp Med Biol. 2012;726:403-22.
- Matsukura T., Kanda T. et al., J Virol. 1986 Jun;58(3):979-82.
- Favre M., Breitburd F. et al., J Virol. 1977 Mar;21(3):1205-9.
-
Lutzner MA.
 , Arch Dermatol. 1983 Aug;119(8):631-5.
, Arch Dermatol. 1983 Aug;119(8):631-5.
- Shope R.E. and Hurst E.W., J Exp Med. 1933 Oct 31;58(5):607-24.
- Shope R.E., J Exp Med. 1931 Jul 31;54(3):373-85.
-
Bravo I.
 G. and Alonso A., Virus Genes. 2007 Jun;34(3):249-62. Epub 2006 Aug 22.
G. and Alonso A., Virus Genes. 2007 Jun;34(3):249-62. Epub 2006 Aug 22.
- Bravo I.G., de Sanjosé S. et al., Trends Microbiol. 2010 Oct;18(10):432-8. Epub 2010 Aug 24.
- Domingo E., Parrish C.R. et al. ed., ISBN-10: 012374153X; ISBN-13: 978-0123741530; Sep 26, 2008
-
Giuliano A.
 R., Lu B. et al., J Infect Dis. 2008 Sep 15;198(6):827-35.
R., Lu B. et al., J Infect Dis. 2008 Sep 15;198(6):827-35.
- Orlando P.A., Gatenby R.A. et al., J Infect Dis. 2012 Jan 15;205(2):272-9. Epub 2011 Nov 16.
- Gravitt P.E., J Clin Invest. 2011 Dec;121(12):4593-9. doi: 10.1172/JCI57149. Epub 2011 Dec 1.
-
Calleja-Macias I.
 E., Villa L.L. et al., J Virol. 2005 Nov;79(21):13630-40.
E., Villa L.L. et al., J Virol. 2005 Nov;79(21):13630-40.
- Burk R.D., Chen Z. et al., Acta Dermatovenerol Alp Panonica Adriat. 2011 Sep;20(3):113-23.
- Conway M.J. and Meyers C., J Dent Res. 2009 Apr;88(4):307-17.
-
Meyers C.
 , Frattini M.G. et al., Science. 1992 Aug 14;257(5072):971-3.
, Frattini M.G. et al., Science. 1992 Aug 14;257(5072):971-3.
- McLaughlin-Drubin M.E. and Meyers C., Methods Mol Med. 2005;119:171-86.
- Rolfe M., Beer-Romero P. et al., Proc Natl Acad Sci U S A. 1995 Apr 11;92(8):3264-8.
-
Dyson N.
 , Howley P.M. et al., Science. 1989 Feb 17;243(4893):934-7.
, Howley P.M. et al., Science. 1989 Feb 17;243(4893):934-7.
- Chen X.S., Casini G. et al., J Mol Biol. 2001 Mar 16;307(1):173-82.
- Ramos I., Martín-Benito J. et al.,, J Biol Chem. 2010 Oct 29;285(44):33771-8. Epub 2010 Aug 9.
-
Luban J.
 , Cell. 1996 Dec 27;87(7):1157-9.
, Cell. 1996 Dec 27;87(7):1157-9.
- Scherer W.F., Syverton J.T. et al, J Exp Med. 1953 May;97(5):695-710.
Плацентарные, или Высшие звери. Млекопитающие
Читайте также
Подотряд Человекопообные (Anthropoidea), или высшие приматы
Подотряд Человекопообные (Anthropoidea), или высшие приматы
Переходим к описанию самых интересных и высокоразвитых приматов — к вершине животного царства. В подотряд человекоподобные входят обезьяны и человек: семь семейств, 33 рода. Сюда включены и мелкие, и средние, и крупные
Сюда включены и мелкие, и средние, и крупные
Надсемейство гоминоиды (Hominoidea), или высшие обезьяны и человек
Надсемейство гоминоиды (Hominoidea), или высшие обезьяны и человек Последнее надсемейство отряда[50]. Относится к секции узконосых приматов. Хорошо изучены, однако систематика вызывает споры. По схеме 4 — три семейства: гиббонообразные (Hylobatidae), крупные человекообразные
ПОДЦАРСТВ0 ВЫСШИЕ РАСТЕНИЯ. ОТДЕЛ МОХОВИДНЫЕ
ПОДЦАРСТВ0 ВЫСШИЕ РАСТЕНИЯ. ОТДЕЛ МОХОВИДНЫЕ Выберите правильный ответ.1. Моховидные относятся к высшим растениям, так как они:A. Многолетние растенияБ. Образуют органические веществаB. Имеют стебель и листьяГ. Размножаются спорами2. Ризоиды у мхов служат:A. Для
Высшие растения
Высшие растения
34. Выберите верное утверждение.1. Высшие растения имеют хорошо выраженные ткани: образовательную, покровную, проводящую, механическую, выделительную, основную и запасающую.2. Секреторная ткань есть только у животных.3. К вегетативным органам растений
Выберите верное утверждение.1. Высшие растения имеют хорошо выраженные ткани: образовательную, покровную, проводящую, механическую, выделительную, основную и запасающую.2. Секреторная ткань есть только у животных.3. К вегетативным органам растений
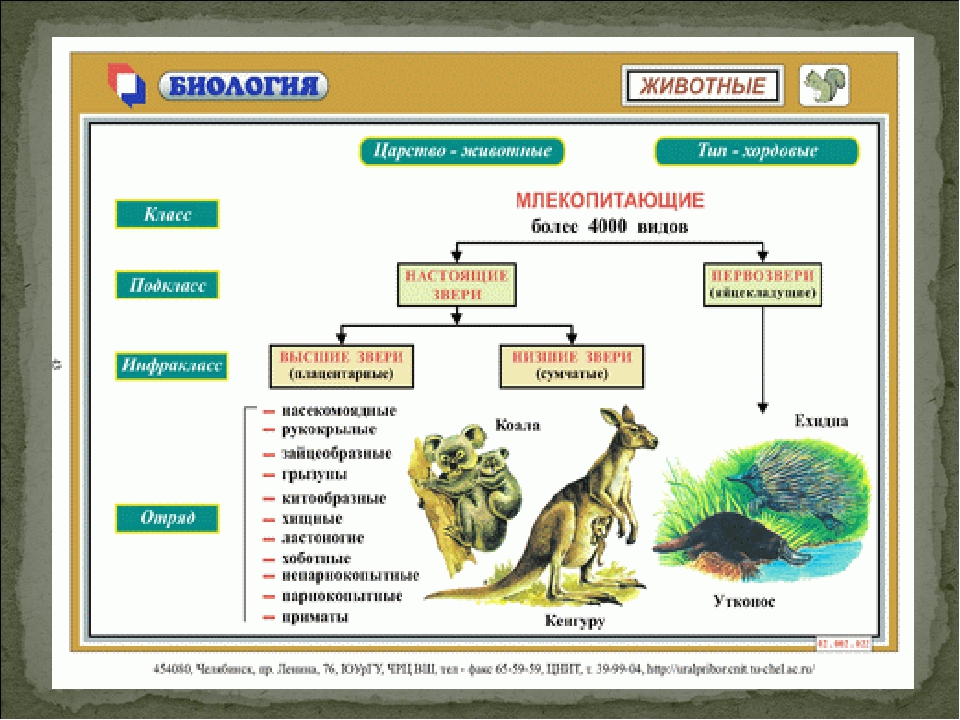
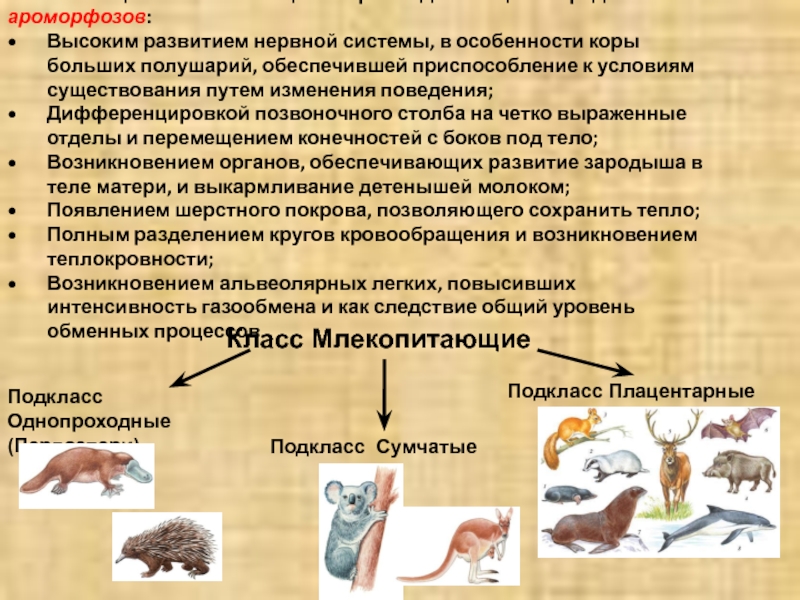
Подкласс Настоящие звери (Сумчатые и Плацентарные)
Подкласс Настоящие звери (Сумчатые и Плацентарные)
Птицы и звери
Птицы и звери Еще сравнительно недавно считали, что у птиц неважное обоняние. Теперь мы знаем, что, по крайней мере, некоторые виды птиц представляют исключение из этого правила.Знаменитый новозеландский киви — одна из тех птиц, у которых хорошее обоняние. Ноздри у киви не
Глава 7 ВЫСШИЕ ФУНКЦИИ НЕРВНОЙ СИСТЕМЫ
Глава 7
ВЫСШИЕ ФУНКЦИИ НЕРВНОЙ СИСТЕМЫ
Общепризнано, что нервная высшая деятельность человека и животных обеспечивается целым комплексом совместно работающих мозговых структур, каждая из которых вносит в этот процесс свой специфический вклад.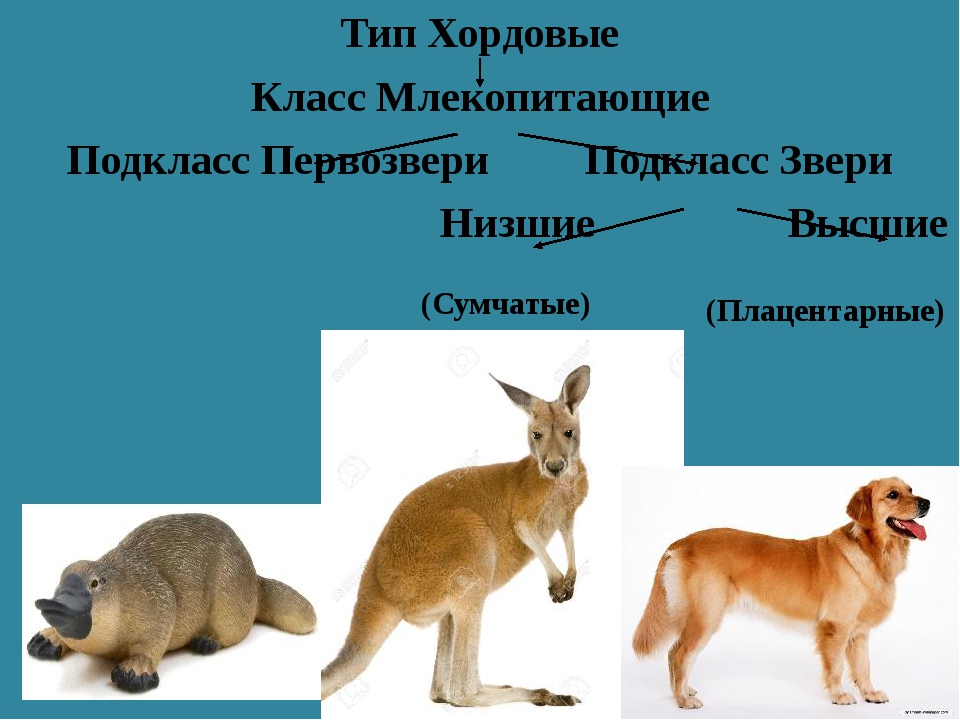 Это означает, что нервная
Это означает, что нервная
8.6. Высшие эстетические эмоции как следствие естественного отбора
8.6. Высшие эстетические эмоции как следствие естественного отбора Лишь убедившись в том, что наши элементарные эстетические эмоции действительно могли слагаться под действием естественного отбора, можно приступить к рассмотрению происхождения гораздо более сложных
Высшие формы игровой исследовательской деятельности животных
Высшие формы игровой исследовательской деятельности животных При всем многообразии форм игры их объединяет большая общая подвижность животного, большое разнообразие производимых им телодвижений и интенсивное перемещение в пространстве (рис. 23). Наиболее ярко это
2.10.1.1.4. Подкласс Высшие раки (Malacostraca)
2. 10.1.1.4. Подкласс Высшие раки (Malacostraca)
Отряд Мизиды (Mysidacca)Отряд из подкласса высших ракообразных. Длина тела у большинства от 0,5 до 2,5 см, у глубоководных – до 18 см. Внешне напоминают маленьких креветок. Развитие прямое (без метаморфоза), протекает в выводковой сумке матери.
10.1.1.4. Подкласс Высшие раки (Malacostraca)
Отряд Мизиды (Mysidacca)Отряд из подкласса высших ракообразных. Длина тела у большинства от 0,5 до 2,5 см, у глубоководных – до 18 см. Внешне напоминают маленьких креветок. Развитие прямое (без метаморфоза), протекает в выводковой сумке матери.
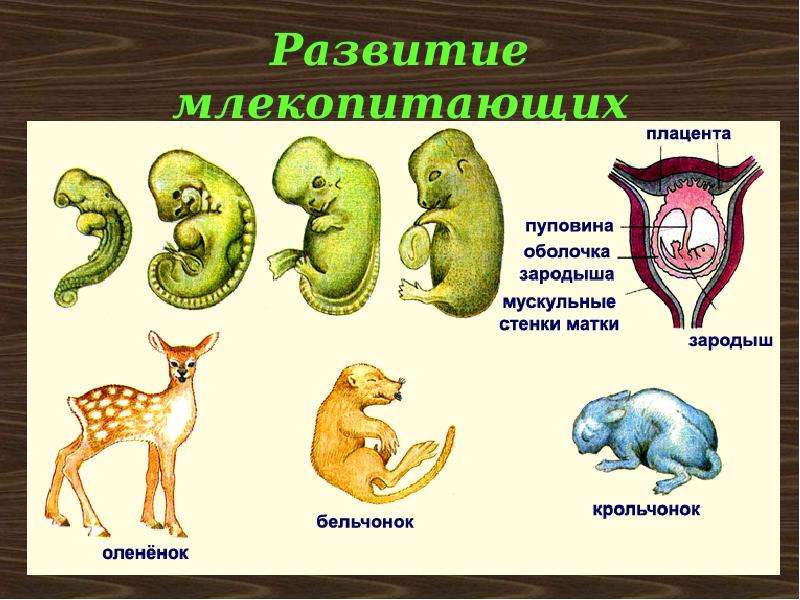
2.11.1.6.1. Подкласс Плацентарные (Eutheria)
2.11.1.6.1. Подкласс Плацентарные (Eutheria) К подклассу Плацентарных относится большинство современных млекопитающих. В организм зародыша питательные вещества и кислород у них поступают из тела матери через особый орган – плацента, которая образуется путем соединения хориона
Звери арсинои
Звери арсинои Лес внезапно расступился, и я очутился на краю обрыва, уступами спускавшегося в широкую низину, которая простиралась до горизонта. Перед моим изумленным взором в лучах низкого закатного солнца вставали на равнине кроваво-красные пирамиды, шпили, башни —
Глава 9.
 Высшие цивилизации
Высшие цивилизации
Глава 9. Высшие цивилизации Мы только что видели, что, по-видимому, в галактике, может, существует много других планет, имеющих на своей поверхности большое количество довольно водянистого раствора таких органических молекул, которые необходимы для того, чтобы служить в
Яйцекладущие звери
Яйцекладущие звери Клоачные, однопроходные, или птецеутробки, произошли, вероятно, от многобугорчатых древних зверей. Во всяком случае, об этом можно судить по строению их коренных зубов, которые есть только у эмбрионов птицеутробок. Рождают не живых детенышей, а
Звери хищные
Звери хищные
Несумчатые хищные звери обитают во всех странах мира. Только в Новой Зеландии и Австралии их никогда прежде не было. Но собак, кошек, лис люди завезли и туда. На Земле, по последним подсчетам, 252 вида хищных зверей. Многие из них разнообразят свою плотоядную
Многие из них разнообразят свою плотоядную
Хищные звери
Хищные звери Этот отряд класса млекопитающих дал человеку только трех домашних животных: из семейства псовых — собаку, из кошачьих — кошку и охотничьего хорька (фретта, или фреттхена) из семейства куньих. Сведения о времени приручения собаки очень
Риск рака у млекопитающих | Nature
Документирование рака у диких животных в большинстве случаев является чрезвычайно сложной задачей из-за отсутствия информации о возрасте особей, сложности извлечения тел для вскрытия и вероятности того, что рак негативно повлияет на выживаемость до того, как сам рак будет обнаружен. Хотя данные о заболеваемости раком среди диких популяций были бы необходимы для описания естественной заболеваемости злокачественными новообразованиями, такие данные, особенно с соответствующими возрастными и демографическими историями, к сожалению, все еще далеки от нашей досягаемости. Поэтому для оценки риска смертности от рака мы использовали данные, предоставленные Species360 и Системой управления зоологической информацией (ZIMS, номер разрешения на использование данных 73836), международной некоммерческой организацией, которая поддерживает централизованную базу данных животных, находящихся на попечении человека, в режиме реального времени. (перегруппировка информации из более чем 1200 зоопарков по всему миру). Хотя мы понимаем, что интерпретация данных, собранных о животных в зоопарках, требует осторожности из-за сильного человеческого контроля над рационом, здоровьем, факторами смертности, окружающей средой или стандартными биологическими функциями животных, зоопарки обеспечивают исключительно высокое разрешение данных о демографии и причинах болезней. гибель многих видов.Здесь мы полагаемся на высокую вероятность извлечения тел умерших животных из зоопарка и рутинное вскрытие большинства из них (если они не обнаружены на поздней стадии разложения) с целью выявления наиболее вероятной патологии, вызвавшей смерть животного.
Поэтому для оценки риска смертности от рака мы использовали данные, предоставленные Species360 и Системой управления зоологической информацией (ZIMS, номер разрешения на использование данных 73836), международной некоммерческой организацией, которая поддерживает централизованную базу данных животных, находящихся на попечении человека, в режиме реального времени. (перегруппировка информации из более чем 1200 зоопарков по всему миру). Хотя мы понимаем, что интерпретация данных, собранных о животных в зоопарках, требует осторожности из-за сильного человеческого контроля над рационом, здоровьем, факторами смертности, окружающей средой или стандартными биологическими функциями животных, зоопарки обеспечивают исключительно высокое разрешение данных о демографии и причинах болезней. гибель многих видов.Здесь мы полагаемся на высокую вероятность извлечения тел умерших животных из зоопарка и рутинное вскрытие большинства из них (если они не обнаружены на поздней стадии разложения) с целью выявления наиболее вероятной патологии, вызвавшей смерть животного. Эти исследования, вероятно, выявят большинство солидных опухолей, но (хотя и возможны) доброкачественные опухоли, жидкие опухоли (например, лейкемия) или рак на ранней стадии вряд ли будут здесь зарегистрированы либо из-за их диагностических трудностей, либо из-за их предполагаемой ограниченной роли в способствует гибели животного.
Эти исследования, вероятно, выявят большинство солидных опухолей, но (хотя и возможны) доброкачественные опухоли, жидкие опухоли (например, лейкемия) или рак на ранней стадии вряд ли будут здесь зарегистрированы либо из-за их диагностических трудностей, либо из-за их предполагаемой ограниченной роли в способствует гибели животного.
В частности, здесь мы используем модуль животноводства ZIMS, предоставляя информацию о рождении, смерти, поле и предварительно определенных категориях патологических изменений, включая новообразования (по определению опухоли, которые способствовали смерти, хотя и без возможности указать рак тип или другие данные). Статистические методы не использовались для предварительного определения размера выборки, но для сведения к минимуму систематической ошибки, вызванной потенциальной временной неоднородностью в методах управления данными и ведения записей о вскрытии 15 , здесь мы сосредоточились на людях, живших или родившихся после 1 января 2010 г. (извлечение данных: 30 мая 2010 г.). 2020).Эта выборка затем использовалась для характеристики ожидаемой продолжительности жизни и заболеваемости раком для конкретных видов, но только после исключения данных, которые не соответствовали ряду критериев, чтобы обеспечить максимально возможное и наиболее однородное качество данных. Во-первых, рак — это возрастное заболевание, которое редко встречается у подростков, а детский рак обычно отличается от рака у взрослых с медицинской точки зрения. Таким образом, различия в младенческой смертности, наблюдаемые у разных видов, могут значительно исказить оценки заболеваемости раком.Таким образом, мы собирали половые или видоспецифические (в тех случаях, когда первые были недоступны) возрасты половой зрелости, и мы рассматривали особей для анализа только в том случае, если они достигли зрелости до или во время нашего периода выборки. Для особей неизвестного пола (около 12 % всех особей в извлечении необработанных данных) в качестве возрастного порога включения использовался максимальный возраст наступления половой зрелости у самцов и самок.
(извлечение данных: 30 мая 2010 г.). 2020).Эта выборка затем использовалась для характеристики ожидаемой продолжительности жизни и заболеваемости раком для конкретных видов, но только после исключения данных, которые не соответствовали ряду критериев, чтобы обеспечить максимально возможное и наиболее однородное качество данных. Во-первых, рак — это возрастное заболевание, которое редко встречается у подростков, а детский рак обычно отличается от рака у взрослых с медицинской точки зрения. Таким образом, различия в младенческой смертности, наблюдаемые у разных видов, могут значительно исказить оценки заболеваемости раком.Таким образом, мы собирали половые или видоспецифические (в тех случаях, когда первые были недоступны) возрасты половой зрелости, и мы рассматривали особей для анализа только в том случае, если они достигли зрелости до или во время нашего периода выборки. Для особей неизвестного пола (около 12 % всех особей в извлечении необработанных данных) в качестве возрастного порога включения использовался максимальный возраст наступления половой зрелости у самцов и самок. Половой возраст наступления половой зрелости для каждого вида был получен из Conde et al. 19 или из опубликованных литературных ресурсов (см. источники данных в https://github.com/OrsolyaVincze/VinczeEtal2021Nature/blob/main/SupplementaryData.xls). Во-вторых, учитывая, что возраст является ключевым предиктором возникновения рака, мы рассматривали только лиц, для которых дата рождения была записана точно или в пределах узкого (максимум 30 дней) временного интервала. В-третьих, мы рассматривали только виды, у которых имелись патологоанатомические записи не менее 20 взрослых особей, независимо от причины смерти (например, инфекция, несчастный случай, гериатрическая болезнь и т. д.).Тем не менее, представленные модели были выполнены с повышенными пороговыми значениями 40, 60, 80 и 100 человек, чтобы проверить согласованность результатов (дополнительная таблица 2). В-четвертых, учитывая, что процесс одомашнивания широко рассматривается как основной фактор, способствующий подавлению инбридинга и более высокой заболеваемости раком 39 , мы исключили все виды, подвергшиеся одомашниванию, а также их диких предков (таксоны исключены из-за того, что они подвергались к одомашниванию перечислены в дополнительной таблице 1).
Половой возраст наступления половой зрелости для каждого вида был получен из Conde et al. 19 или из опубликованных литературных ресурсов (см. источники данных в https://github.com/OrsolyaVincze/VinczeEtal2021Nature/blob/main/SupplementaryData.xls). Во-вторых, учитывая, что возраст является ключевым предиктором возникновения рака, мы рассматривали только лиц, для которых дата рождения была записана точно или в пределах узкого (максимум 30 дней) временного интервала. В-третьих, мы рассматривали только виды, у которых имелись патологоанатомические записи не менее 20 взрослых особей, независимо от причины смерти (например, инфекция, несчастный случай, гериатрическая болезнь и т. д.).Тем не менее, представленные модели были выполнены с повышенными пороговыми значениями 40, 60, 80 и 100 человек, чтобы проверить согласованность результатов (дополнительная таблица 2). В-четвертых, учитывая, что процесс одомашнивания широко рассматривается как основной фактор, способствующий подавлению инбридинга и более высокой заболеваемости раком 39 , мы исключили все виды, подвергшиеся одомашниванию, а также их диких предков (таксоны исключены из-за того, что они подвергались к одомашниванию перечислены в дополнительной таблице 1). После этих ограничений извлечение данных о возрасте и причине смерти привело к получению информации о 110 148 (62 556 живых и 47 592 погибших) особях ( n = 191 вид). Для расчета ИКМ мы включили только виды, у которых правильно оценена доживаемость до старости (т.е. данные, позволяющие оценить возрастную доживаемость до возраста, в котором доживает только 10% особей, n = 172 вида ). Хотя эти ограничения устранили множественные источники систематической ошибки в наших оценках риска смертности от рака, мы не можем исключить возможность того, что некоторые виды (например, более харизматичные) подвергаются более частым или более подробным вскрытиям.Тем не менее, наш статистический подход, особенно полный анализ случаев, в значительной степени нечувствителен к таким предубеждениям, поскольку лица, не имеющие доступных посмертных диагностических записей, считаются подвергнутыми цензуре (см. ниже). Кроме того, несмотря на то, что глубина вскрытия может незначительно различаться у разных видов, новообразования, которые внесли значительный вклад в гибель животных (в центре внимания нашего исследования), обычно выявляются даже при макроскопическом вскрытии.
После этих ограничений извлечение данных о возрасте и причине смерти привело к получению информации о 110 148 (62 556 живых и 47 592 погибших) особях ( n = 191 вид). Для расчета ИКМ мы включили только виды, у которых правильно оценена доживаемость до старости (т.е. данные, позволяющие оценить возрастную доживаемость до возраста, в котором доживает только 10% особей, n = 172 вида ). Хотя эти ограничения устранили множественные источники систематической ошибки в наших оценках риска смертности от рака, мы не можем исключить возможность того, что некоторые виды (например, более харизматичные) подвергаются более частым или более подробным вскрытиям.Тем не менее, наш статистический подход, особенно полный анализ случаев, в значительной степени нечувствителен к таким предубеждениям, поскольку лица, не имеющие доступных посмертных диагностических записей, считаются подвергнутыми цензуре (см. ниже). Кроме того, несмотря на то, что глубина вскрытия может незначительно различаться у разных видов, новообразования, которые внесли значительный вклад в гибель животных (в центре внимания нашего исследования), обычно выявляются даже при макроскопическом вскрытии. Кроме того, крупные виды считаются ключевыми для зоопарков, что также подтверждается тем фактом, что доля погибших особей с посмертными патологическими записями больше у более крупных видов (корреляция Пирсона: r = 0.24, t = 3,35, df = 189, P = 0,001). Соответственно, если бы харизма играла роль в обнаружении рака, мы бы ожидали большего риска рака у крупных млекопитающих, в противоположность (незначительному) отрицательному эффекту массы тела в наших моделях. Следовательно, мы считаем, что харизма вряд ли представляет собой основной источник предвзятости в нашем анализе.
Кроме того, крупные виды считаются ключевыми для зоопарков, что также подтверждается тем фактом, что доля погибших особей с посмертными патологическими записями больше у более крупных видов (корреляция Пирсона: r = 0.24, t = 3,35, df = 189, P = 0,001). Соответственно, если бы харизма играла роль в обнаружении рака, мы бы ожидали большего риска рака у крупных млекопитающих, в противоположность (незначительному) отрицательному эффекту массы тела в наших моделях. Следовательно, мы считаем, что харизма вряд ли представляет собой основной источник предвзятости в нашем анализе.
Оценка ожидаемой продолжительности жизни во взрослом возрасте
Поскольку у нас нет оснований полагать, что подвергнутые цензуре лица не будут иметь такие же шансы на выживание, как и те, за которыми продолжают следить, мы оцениваем ожидаемую продолжительность жизни во взрослом возрасте на основе возрастной выживаемости, рассчитанной с использованием шкалы Каплана– Процедура Мейера (с использованием функции survfit в R-пакете выживания 40 ). Лица старше их возраста половой зрелости на 1 января 2010 г. были усечены слева в соответствии с их возрастом на эту дату; особи, достигшие половой зрелости после этой даты, были усечены влево в их возрасте половой зрелости. Особи, еще живые на момент извлечения данных, считались подвергнутыми правой цензуре (выборки на вид варьировались от 42 до 5816 особей), в то время как особи с известной судьбой были назначены мертвыми ( n = 47 592), независимо от того, была ли их причина смерти указано или нет.
Лица старше их возраста половой зрелости на 1 января 2010 г. были усечены слева в соответствии с их возрастом на эту дату; особи, достигшие половой зрелости после этой даты, были усечены влево в их возрасте половой зрелости. Особи, еще живые на момент извлечения данных, считались подвергнутыми правой цензуре (выборки на вид варьировались от 42 до 5816 особей), в то время как особи с известной судьбой были назначены мертвыми ( n = 47 592), независимо от того, была ли их причина смерти указано или нет.
Оценка ICM
ICM была рассчитана с использованием метода конкурирующих рисков, основанного на совокупной опасности смертей, связанных с раком, и вероятности выживания видов, находящихся под опекой человека. Во-первых, возрастная выживаемость S x в возрасте x была оценена с помощью анализа КМ, как указано выше. Однако здесь мы провели полный анализ случаев, используя только 11 840 человек, для которых причина смерти была указана вместе с выжившими, подвергнутыми правой цензуре. Анализ полного случая предполагает, что отсутствие в качестве причины отказа является случайным, но у нас не было оснований полагать, что это не так в нашем наборе данных. Посмертные исследования обычно проводятся для большинства обнаруженных тел в зоопарках, и после проведения исследований результаты с равной вероятностью будут внесены в базу данных независимо от выявленных патологий. Таким образом, оценки ICM были основаны на n = 74 396 особей, n = 179 видов. Во-вторых, риск смертности от рака h c x был оценен с использованием анализа KMx1, в котором в качестве события смерти учитывались только случаи смерти от рака.{{\rm{c}}}$$
Анализ полного случая предполагает, что отсутствие в качестве причины отказа является случайным, но у нас не было оснований полагать, что это не так в нашем наборе данных. Посмертные исследования обычно проводятся для большинства обнаруженных тел в зоопарках, и после проведения исследований результаты с равной вероятностью будут внесены в базу данных независимо от выявленных патологий. Таким образом, оценки ICM были основаны на n = 74 396 особей, n = 179 видов. Во-вторых, риск смертности от рака h c x был оценен с использованием анализа KMx1, в котором в качестве события смерти учитывались только случаи смерти от рака.{{\rm{c}}}$$
, где α — возраст наступления половой зрелости. Единственная разница с классической оценкой состоит в том, что мы извлекли ч с x (и S x ) для каждой единицы времени с дискретными скачками (и падениями) в момент времени события в возрасте t . {{\rm{c}}}/{n}_{t} \) на этих временных интервалах (где d c t — число умерших от рака в пределах интервала и n t — число выживших в начале интервала) .Мы выбрали этот метод, чтобы отразить истинную изменчивость данных для межвидового сравнения, когда виды сильно различаются по количеству событий и временному интервалу между ними (иногда треть продолжительности жизни взрослого организма), что редко встречается при сравнении человеческих групп.
{{\rm{c}}}/{n}_{t} \) на этих временных интервалах (где d c t — число умерших от рака в пределах интервала и n t — число выживших в начале интервала) .Мы выбрали этот метод, чтобы отразить истинную изменчивость данных для межвидового сравнения, когда виды сильно различаются по количеству событий и временному интервалу между ними (иногда треть продолжительности жизни взрослого организма), что редко встречается при сравнении человеческих групп.
Ковариаты и статистический анализ
Для каждого вида мы получили данные о массе тела взрослых особей в зависимости от пола из ZIMS Species360 (см. https://github.com/OrsolyaVincze/VinczeEtal2021Nature/blob/main/SupplementaryData.xls). Видоспецифичную массу тела рассчитывали как среднее значение всех измерений массы тела, зарегистрированных в базе данных ZIMS у взрослых особей, тогда как видоспецифические значения получали путем усреднения масс тела самцов и самок. Они были рассчитаны только для видов, для которых было зарегистрировано не менее 100 записей о массе тела взрослых особей; в противном случае масса тела была взята из литературы и обзора базы данных Conde et al. 19 . Мы проверили, что существует однозначное соответствие в информации о массе тела для видов с записями в обоих наборах данных.
Они были рассчитаны только для видов, для которых было зарегистрировано не менее 100 записей о массе тела взрослых особей; в противном случае масса тела была взята из литературы и обзора базы данных Conde et al. 19 . Мы проверили, что существует однозначное соответствие в информации о массе тела для видов с записями в обоих наборах данных.
Информация о рационе была получена из глобального набора данных о рационе наземных млекопитающих 32 , предоставляющего информацию о составе рациона на четырех иерархических уровнях продуктов питания (никогда не потребляемые, иногда потребляемые, второстепенные продукты питания, основные продукты питания). Мы собрали информацию о содержании животных в рационе, а также о подкатегориях этого класса рациона, а именно о потреблении беспозвоночных или позвоночных, а также особенно о потреблении рыбы, рептилий, птиц и млекопитающих. Учитывая, что большинство продуктов питания имело несколько видов на промежуточных уровнях (употребление время от времени и второстепенный продукт питания), мы переклассифицировали переменные рациона на двух уровнях: никогда/редко потребляемые или представляющие собой основной/второстепенный продукт питания данного вида. Влияние диеты было проверено в регрессиях PGLS с использованием только видов с ненулевым риском смертности от рака. Модели запускались отдельно для каждого продукта питания, который был введен в базовую модель, включая массу тела и ожидаемую продолжительность жизни во взрослом возрасте в качестве ковариатов. Результаты представлены в Таблице 2 с расширенными данными. 41 , охватывающий 5 911 видов. Затем мы получили дерево консенсуса с корнями, используя библиотеку Python sumtrees 42 . Два вида, недавно повышенные до уровня видов, были вручную добавлены в дерево как родственные таксоны видов, от которых он был недавно отделен (то есть от Cervus canadensis до Cervus elaphus и от Gazella marica до Gazella subgutturosa ). Филогенетический сигнал риска рака оценивали с помощью функции phylosig из пакета R phytools 22 .Частные коэффициенты детерминаций рассчитывали с использованием функции R2.pred из пакета R rr2 (ref. 43 ) на основе моделей, представленных в таблице расширенных данных 3.
Влияние диеты было проверено в регрессиях PGLS с использованием только видов с ненулевым риском смертности от рака. Модели запускались отдельно для каждого продукта питания, который был введен в базовую модель, включая массу тела и ожидаемую продолжительность жизни во взрослом возрасте в качестве ковариатов. Результаты представлены в Таблице 2 с расширенными данными. 41 , охватывающий 5 911 видов. Затем мы получили дерево консенсуса с корнями, используя библиотеку Python sumtrees 42 . Два вида, недавно повышенные до уровня видов, были вручную добавлены в дерево как родственные таксоны видов, от которых он был недавно отделен (то есть от Cervus canadensis до Cervus elaphus и от Gazella marica до Gazella subgutturosa ). Филогенетический сигнал риска рака оценивали с помощью функции phylosig из пакета R phytools 22 .Частные коэффициенты детерминаций рассчитывали с использованием функции R2.pred из пакета R rr2 (ref. 43 ) на основе моделей, представленных в таблице расширенных данных 3.
логистические модели, которые позволяют делать выводы о вероятности обнаружения хотя бы одного случая рака у вида и, при условии обнаружения рака, выводы о CMR или ICM. Таким образом, первая часть этого состояла из филогенетической биномиальной регрессии (с использованием функции binaryPGLMM, в пакете R ape 44 ), где зависимая переменная объясняла наличие нулей и ненулевых значений в CMR или ICM.Эта модель содержала логарифмическое количество умерших с доступными посмертными патологическими записями в качестве объясняющей переменной из-за более высокой вероятности обнаружения рака при большем количестве осмотренных умерших. Кроме того, модель содержала массу тела и ожидаемую продолжительность жизни во взрослом возрасте как ковариаты. Вторая часть модели состояла из регрессии PGLS, которая исследовала дисперсию только при ненулевых рисках рака. ICM и CMR были логит-преобразованы во всех моделях PGLS в соответствии с рекомендациями при анализе пропорций 45 . Эти модели были взвешены по логарифмическому числу умерших с доступными посмертными патологическими записями, поскольку ожидается, что точность оценок риска смертности от рака будет увеличиваться с увеличением числа умерших, подвергнутых патологоанатомическому исследованию, но не ожидается, что это объяснит систематическую ошибку в оценке. зависимой переменной в любом конкретном направлении (как в случае биномиальных моделей). Эти модели также содержали массу тела и ожидаемую продолжительность жизни во взрослом возрасте в качестве объясняющих переменных.Учитывая ожидаемый аддитивный эффект массы тела и продолжительности жизни, взаимодействие между показателями массы тела и продолжительности жизни также было проверено во всех четырех моделях (биномиальная и логистическая регрессии для CMR и ICM), но эти взаимодействия ни в каком случае не улучшали соответствие модели и являются поэтому не представлены. Обе модели контролировались на предмет филогенетического родства между видами, где параметр масштабирования филогенетической зависимости (то есть s2/Pagel’s λ в PGLMM и PGLS соответственно) был установлен на наиболее подходящие значения, оцененные статикой отношения правдоподобия в каждой модели отдельно.
Эти модели были взвешены по логарифмическому числу умерших с доступными посмертными патологическими записями, поскольку ожидается, что точность оценок риска смертности от рака будет увеличиваться с увеличением числа умерших, подвергнутых патологоанатомическому исследованию, но не ожидается, что это объяснит систематическую ошибку в оценке. зависимой переменной в любом конкретном направлении (как в случае биномиальных моделей). Эти модели также содержали массу тела и ожидаемую продолжительность жизни во взрослом возрасте в качестве объясняющих переменных.Учитывая ожидаемый аддитивный эффект массы тела и продолжительности жизни, взаимодействие между показателями массы тела и продолжительности жизни также было проверено во всех четырех моделях (биномиальная и логистическая регрессии для CMR и ICM), но эти взаимодействия ни в каком случае не улучшали соответствие модели и являются поэтому не представлены. Обе модели контролировались на предмет филогенетического родства между видами, где параметр масштабирования филогенетической зависимости (то есть s2/Pagel’s λ в PGLMM и PGLS соответственно) был установлен на наиболее подходящие значения, оцененные статикой отношения правдоподобия в каждой модели отдельно. Модели PGLS, в которых λ Пейджа сходились к отрицательным значениям, были переоборудованы с λ Пейджа, фиксированным на 0. Три вида ( Lagurus lagurus , Cricetus cricetus и Dasyuroides byrnei ) были удалены. к их высокому рычагу, вызванному их очень низкой ожидаемой продолжительностью взрослой жизни по сравнению с остальными видами, и, следовательно, опасениями о сильном влиянии этих точек на соответствие модели. Тем не менее, все модели были выполнены с использованием всего набора данных, и результаты в высокой степени соответствовали исключениям и без них (дополнительная таблица 4 и расширенные данные, рис.7).
Модели PGLS, в которых λ Пейджа сходились к отрицательным значениям, были переоборудованы с λ Пейджа, фиксированным на 0. Три вида ( Lagurus lagurus , Cricetus cricetus и Dasyuroides byrnei ) были удалены. к их высокому рычагу, вызванному их очень низкой ожидаемой продолжительностью взрослой жизни по сравнению с остальными видами, и, следовательно, опасениями о сильном влиянии этих точек на соответствие модели. Тем не менее, все модели были выполнены с использованием всего набора данных, и результаты в высокой степени соответствовали исключениям и без них (дополнительная таблица 4 и расширенные данные, рис.7).
Различия порядков заболеваемости раком были проверены с использованием стандартных линейных регрессий, построенных с использованием только таксономических порядков, в которых по крайней мере два вида оценивали заболеваемость раком. Модель содержала CMR или ICM (непреобразованные) в качестве зависимых переменных и порядок в качестве единственного объясняющего фактора.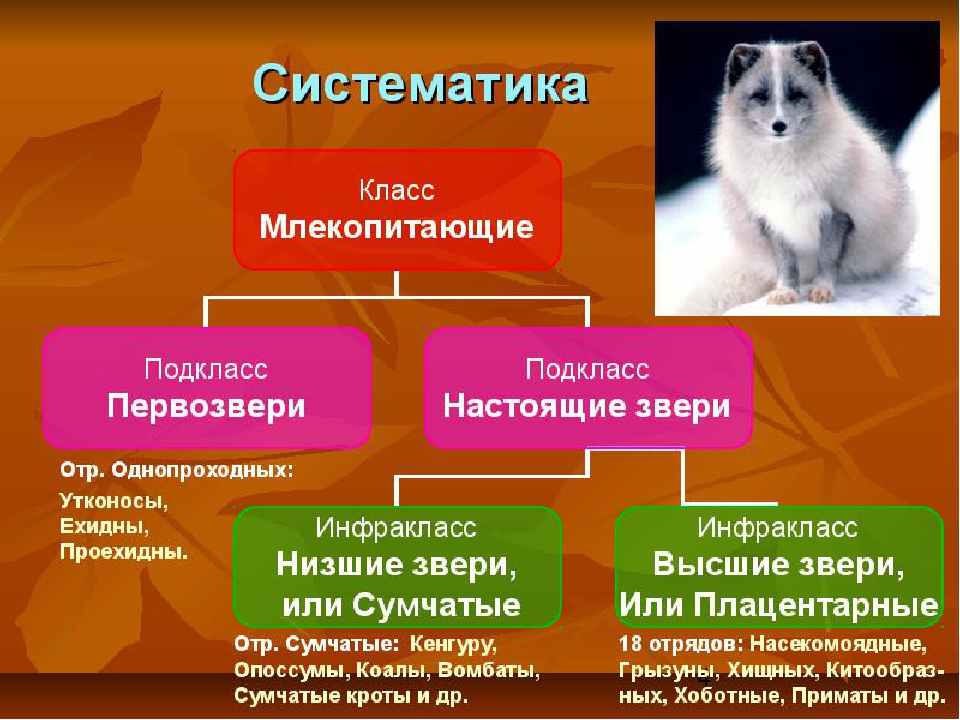 Различия в попарном порядке оценивались с использованием пакета R emmeans 46 . Весь анализ был выполнен в R Statistical and Programming Environment, версия 4.0.4 (ссылка. 47 ). Риски смертности от рака были преобразованы в проценты в цифрах и в анализе, выполненном на различиях порядка (расширенная таблица данных 1), для облегчения интерпретации. Модели, представленные в таблицах 2 и 3 с расширенными данными и в дополнительных таблицах 2–4, основаны на вероятностях.
Различия в попарном порядке оценивались с использованием пакета R emmeans 46 . Весь анализ был выполнен в R Statistical and Programming Environment, версия 4.0.4 (ссылка. 47 ). Риски смертности от рака были преобразованы в проценты в цифрах и в анализе, выполненном на различиях порядка (расширенная таблица данных 1), для облегчения интерпретации. Модели, представленные в таблицах 2 и 3 с расширенными данными и в дополнительных таблицах 2–4, основаны на вероятностях.
Сводка отчета
Дополнительную информацию о дизайне исследования можно найти в Сводке отчета об исследовании природы, связанной с этим документом.
Травоядные, подверженные наибольшему риску исчезновения среди млекопитающих, птиц и рептилий
ВВЕДЕНИЕ
За последние 500 лет вымерло не менее 368 видов позвоночных ( 1 ), большинство в результате прямого (например,ж., охота) и косвенное (например, потеря среды обитания) влияние человека ( 2 – 4 ).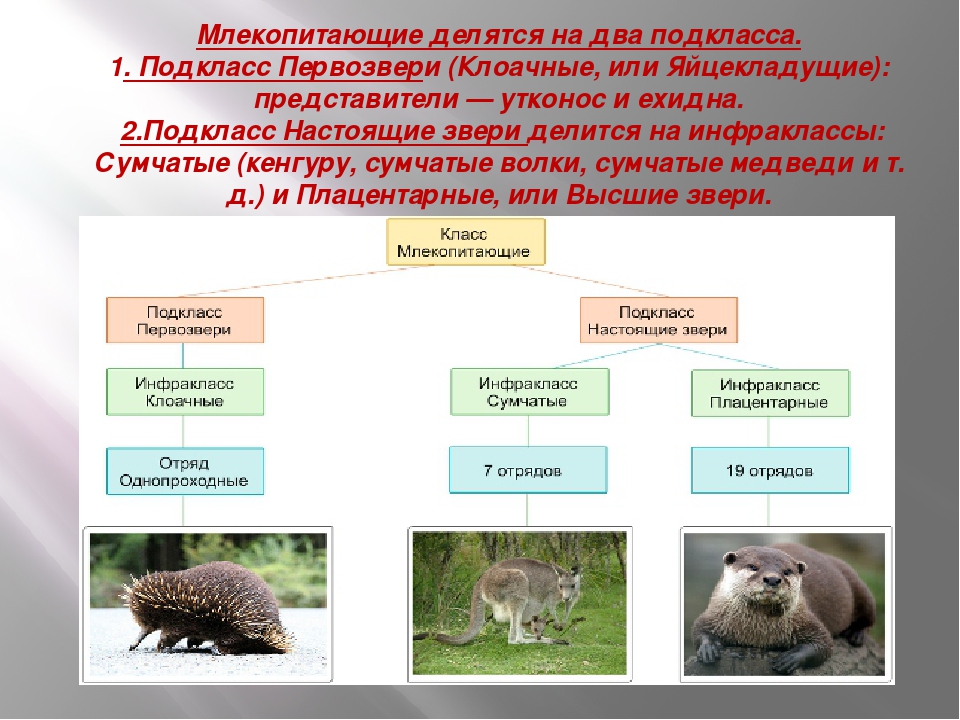 Кроме того, 18% существующих позвоночных животных находятся под угрозой исчезновения ( 1 ), и ожидается, что распространение антропогенных нарушений в сочетании с изменением климата усугубит сокращение видов ( 5 ). Поскольку виды управляют функциями экосистемы, эта потеря видов, как и предшествовавшие исторические события массового вымирания, вероятно, сильно повлияют на экологию нашей планеты ( 6 , 7 ).Исторические вымирания научили нас тому, что непропорциональная утрата крупных видов и видов в пределах определенной трофической группы (хищников, травоядных или всеядных) имеет существенные экологические и эволюционные последствия, выходящие далеко за рамки простой потери таксономического разнообразия ( 7 – 9 ). Например, вымирание мегатравоядных в позднем четвертичном периоде изменило взаимодействие растений и травоядных таким образом, что это привело к заметным сдвигам в растительных сообществах, которые, в свою очередь, изменили режимы пожаров и биогеохимический цикл ( 9 , 10 ).
Кроме того, 18% существующих позвоночных животных находятся под угрозой исчезновения ( 1 ), и ожидается, что распространение антропогенных нарушений в сочетании с изменением климата усугубит сокращение видов ( 5 ). Поскольку виды управляют функциями экосистемы, эта потеря видов, как и предшествовавшие исторические события массового вымирания, вероятно, сильно повлияют на экологию нашей планеты ( 6 , 7 ).Исторические вымирания научили нас тому, что непропорциональная утрата крупных видов и видов в пределах определенной трофической группы (хищников, травоядных или всеядных) имеет существенные экологические и эволюционные последствия, выходящие далеко за рамки простой потери таксономического разнообразия ( 7 – 9 ). Например, вымирание мегатравоядных в позднем четвертичном периоде изменило взаимодействие растений и травоядных таким образом, что это привело к заметным сдвигам в растительных сообществах, которые, в свою очередь, изменили режимы пожаров и биогеохимический цикл ( 9 , 10 ). Травоядные животные также производят значительное количество парникового газа метана, и исследования показали, что результирующее сокращение метана в результате вымирания мегатравоядных в позднем четвертичном периоде могло быть причиной понижения климата Земли на 0,08–0,20 ° C, что привело к позднему дриасу. ( 8 ). Хотя есть убедительные доказательства того, что текущая потеря видов продолжает непропорционально сильно влиять на крупных позвоночных ( 11 , 12 ), наше понимание того, какая трофическая группа подвергается наибольшему риску исчезновения, в основном основано на неофициальных данных и корреляциях со связанными видовыми признаками. к риску исчезновения.Предполагается, что трофическая группа, положение организма в пищевой цепи, связана с современным риском исчезновения ( 13 – 15 ). В частности, считается, что более высокие трофические группы (например, хищники) подвергаются большему риску, чем более низкие (например, травоядные) ( 13 – 16 ).
Травоядные животные также производят значительное количество парникового газа метана, и исследования показали, что результирующее сокращение метана в результате вымирания мегатравоядных в позднем четвертичном периоде могло быть причиной понижения климата Земли на 0,08–0,20 ° C, что привело к позднему дриасу. ( 8 ). Хотя есть убедительные доказательства того, что текущая потеря видов продолжает непропорционально сильно влиять на крупных позвоночных ( 11 , 12 ), наше понимание того, какая трофическая группа подвергается наибольшему риску исчезновения, в основном основано на неофициальных данных и корреляциях со связанными видовыми признаками. к риску исчезновения.Предполагается, что трофическая группа, положение организма в пищевой цепи, связана с современным риском исчезновения ( 13 – 15 ). В частности, считается, что более высокие трофические группы (например, хищники) подвергаются большему риску, чем более низкие (например, травоядные) ( 13 – 16 ). Прогнозируется, что риск исчезновения коррелирует с трофическим положением по нескольким причинам, включая более высокие потребности в энергии ( 14 ) и зависимость хищников от более низких трофических уровней в отношении пищи, которые сами по себе могут быть в упадке ( 15 , 17 ).Карбоне и Гиттлман ( 18 ) подсчитали, что 10 000 кг добычи необходимо, чтобы прокормить всего 90 кг данного вида хищников, а истощение добычи связано с сокращением численности многих крупных хищников, включая тигров, красных волков и несколько видов леопардов. ( 17 ). Сокращение популяций естественной добычи в сочетании с сокращением ареалов обитания также увеличивает конфликты между людьми и хищниками, поскольку хищники распространяются на территории, занятые людьми ( 19 ). Незначительные отношения между людьми и хищниками иллюстрируются тем фактом, что охота и отлов, по крайней мере частично, привели к исчезновению 80% исчезающих видов отряда хищников ( 1 ).
Прогнозируется, что риск исчезновения коррелирует с трофическим положением по нескольким причинам, включая более высокие потребности в энергии ( 14 ) и зависимость хищников от более низких трофических уровней в отношении пищи, которые сами по себе могут быть в упадке ( 15 , 17 ).Карбоне и Гиттлман ( 18 ) подсчитали, что 10 000 кг добычи необходимо, чтобы прокормить всего 90 кг данного вида хищников, а истощение добычи связано с сокращением численности многих крупных хищников, включая тигров, красных волков и несколько видов леопардов. ( 17 ). Сокращение популяций естественной добычи в сочетании с сокращением ареалов обитания также увеличивает конфликты между людьми и хищниками, поскольку хищники распространяются на территории, занятые людьми ( 19 ). Незначительные отношения между людьми и хищниками иллюстрируются тем фактом, что охота и отлов, по крайней мере частично, привели к исчезновению 80% исчезающих видов отряда хищников ( 1 ). Понимание того, как риск варьируется среди разных трофических групп, важно, потому что, независимо от признаков, определяющих риск исчезновения, неслучайные модели вымирания среди трофических групп могут привести к изменениям в трофической структуре, которые могут повлиять на многие аспекты экосистемы ( 13 ). Например, сокращение количества стервятников, питающихся падалью в Индии, привело к повышенному риску распространения таких заболеваний, как бешенство и сибирская язва ( 13 ). В другом примере сокращение численности крупных плодоядных в тропических лесах меняет форму древесных сообществ, что, в свою очередь, по прогнозам, приведет к общему сокращению запасов углерода в лесах ( 20 ).Риск исчезновения определяется историческими и современными социально-экологическими факторами, повлиявшими на регион (например, время с момента колонизации человеком, валовой национальный продукт и промышленное развитие) ( 2 , 16 ). Пространственная изменчивость человеческого следа ( 21 ) может в конечном итоге по-разному влиять на трофические группы, приводя к различиям в риске вымирания на географическом уровне или на уровне среды обитания среди хищников, всеядных и травоядных.
Понимание того, как риск варьируется среди разных трофических групп, важно, потому что, независимо от признаков, определяющих риск исчезновения, неслучайные модели вымирания среди трофических групп могут привести к изменениям в трофической структуре, которые могут повлиять на многие аспекты экосистемы ( 13 ). Например, сокращение количества стервятников, питающихся падалью в Индии, привело к повышенному риску распространения таких заболеваний, как бешенство и сибирская язва ( 13 ). В другом примере сокращение численности крупных плодоядных в тропических лесах меняет форму древесных сообществ, что, в свою очередь, по прогнозам, приведет к общему сокращению запасов углерода в лесах ( 20 ).Риск исчезновения определяется историческими и современными социально-экологическими факторами, повлиявшими на регион (например, время с момента колонизации человеком, валовой национальный продукт и промышленное развитие) ( 2 , 16 ). Пространственная изменчивость человеческого следа ( 21 ) может в конечном итоге по-разному влиять на трофические группы, приводя к различиям в риске вымирания на географическом уровне или на уровне среды обитания среди хищников, всеядных и травоядных. Кроме того, люди играли роль в вымирании видов, по крайней мере, с позднего плейстоцена ( 4 ), и закономерности исчезающих видов в трофических группах могут быть получены в результате прошлых вымираний.Эти процессы могут вызвать гистерезис угрозы исчезновения: если люди в прошлом активно преследовали хищников, то выжившие хищные виды, вероятно, будут наиболее устойчивыми к антропогенным воздействиям и, следовательно, могут с меньшей вероятностью подвергаться угрозе со стороны современных факторов. Мы исследовали возможные различия в риске исчезновения между трофическими группами с использованием трех различных подходов. Во-первых, мы изучили современные модели риска исчезновения среди травоядных, всеядных и хищников. Для этого мы сосредоточились на сравнении моделей угроз в трофических группах и рационах основных таксономических групп (млекопитающие, птицы и рептилии) в глобальном масштабе, в разных регионах суши и океана и в разных типах среды обитания.Во-вторых, мы изучили роль, которую прошлые вымирания могли сыграть в траектории современных паттернов, изучив пропорции недавно вымерших видов млекопитающих, птиц и рептилий, а также млекопитающих, вымерших в позднем плейстоцене, в каждой трофической группе.
Кроме того, люди играли роль в вымирании видов, по крайней мере, с позднего плейстоцена ( 4 ), и закономерности исчезающих видов в трофических группах могут быть получены в результате прошлых вымираний.Эти процессы могут вызвать гистерезис угрозы исчезновения: если люди в прошлом активно преследовали хищников, то выжившие хищные виды, вероятно, будут наиболее устойчивыми к антропогенным воздействиям и, следовательно, могут с меньшей вероятностью подвергаться угрозе со стороны современных факторов. Мы исследовали возможные различия в риске исчезновения между трофическими группами с использованием трех различных подходов. Во-первых, мы изучили современные модели риска исчезновения среди травоядных, всеядных и хищников. Для этого мы сосредоточились на сравнении моделей угроз в трофических группах и рационах основных таксономических групп (млекопитающие, птицы и рептилии) в глобальном масштабе, в разных регионах суши и океана и в разных типах среды обитания.Во-вторых, мы изучили роль, которую прошлые вымирания могли сыграть в траектории современных паттернов, изучив пропорции недавно вымерших видов млекопитающих, птиц и рептилий, а также млекопитающих, вымерших в позднем плейстоцене, в каждой трофической группе. Наконец, мы определили, является ли трофическая группа важным фактором, определяющим риск исчезновения, изучив, как размер тела и трофическая группа взаимодействуют, влияя на статус угрозы вида. Чтобы изучить наши цели, мы собрали базу данных, классифицирующую трофические группы (хищники, травоядные и всеядные) и рацион всех млекопитающих, птиц и рептилий, не имеющих дефицита данных (22 166 видов), оцененных Международным союзом охраны природы ( IUCN) ( 1 ), мировой орган по вымиранию видов и риску исчезновения.
Наконец, мы определили, является ли трофическая группа важным фактором, определяющим риск исчезновения, изучив, как размер тела и трофическая группа взаимодействуют, влияя на статус угрозы вида. Чтобы изучить наши цели, мы собрали базу данных, классифицирующую трофические группы (хищники, травоядные и всеядные) и рацион всех млекопитающих, птиц и рептилий, не имеющих дефицита данных (22 166 видов), оцененных Международным союзом охраны природы ( IUCN) ( 1 ), мировой орган по вымиранию видов и риску исчезновения.ОБСУЖДЕНИЕ
Многочисленные исследования помогли определить черты видов, которые коррелируют с риском вымирания, чтобы пролить свет на текущие модели антропоценового кризиса вымирания ( 12 , 14 , 15 ). Результаты некоторых из этих прошлых исследований, а также множество исследований отдельных видов, посвященных угрозе исчезновения харизматичных хищников, привели к общему предположению, что хищники по ряду причин подвергаются более высокому риску исчезновения, чем другие трофические группы. 14 – 16 , 19 ).Путем систематического изучения моделей подверженных риску видов в различных трофических группах наше исследование показало, что угроза исчезновения имеет трофическую асимметрию. Однако, вопреки многим из этих предыдущих ожиданий, мы обнаружили, что травоядные животные являются трофической группой с наибольшим риском среди млекопитающих, птиц и рептилий. Хотя география и среда обитания влияли на таксономические результаты для конкретных классов, травоядные неизменно имели самую высокую представленность видов, подвергающихся риску, в настоящее время, в недавнем прошлом и в позднем плейстоцене.Во многих случаях рептилии были основной группой, определяющей наблюдаемые нами модели риска исчезновения современных травоядных. Этот результат заслуживает внимания, учитывая, что многое еще предстоит определить в отношении их современной функциональной роли в экосистемах ( 23 ). Мы выявили несколько случаев, когда хищники демонстрируют повышенный риск исчезновения.
14 – 16 , 19 ).Путем систематического изучения моделей подверженных риску видов в различных трофических группах наше исследование показало, что угроза исчезновения имеет трофическую асимметрию. Однако, вопреки многим из этих предыдущих ожиданий, мы обнаружили, что травоядные животные являются трофической группой с наибольшим риском среди млекопитающих, птиц и рептилий. Хотя география и среда обитания влияли на таксономические результаты для конкретных классов, травоядные неизменно имели самую высокую представленность видов, подвергающихся риску, в настоящее время, в недавнем прошлом и в позднем плейстоцене.Во многих случаях рептилии были основной группой, определяющей наблюдаемые нами модели риска исчезновения современных травоядных. Этот результат заслуживает внимания, учитывая, что многое еще предстоит определить в отношении их современной функциональной роли в экосистемах ( 23 ). Мы выявили несколько случаев, когда хищники демонстрируют повышенный риск исчезновения. Когда были проанализированы более подробные категории рациона, как рыбоядные, так и падальщики имели повышенный риск по сравнению с фоновыми уровнями. В соответствии с более высоким риском вымирания рыбоядных мы обнаружили, что повышенный риск вымирания хищников возникает почти исключительно в морских средах обитания, что позволяет предположить, что давление вымирания, угрожающее хищникам, может быть выше в океане, чем на суше.Прошлые исследования показали, что во многих регионах люди преимущественно и неустойчиво эксплуатируют морские организмы на вершине пищевой цепи ( 24 ). Однако это исследование было сосредоточено в основном на рыбе ( 24 ), которую мы не включили в наше исследование. Вероятным продуктивным будущим исследованием будет определение того, как включение костистых и хрящевых рыб и других таксономических классов, не включенных в это исследование (например, беспозвоночных), повлияет на интерпретацию того, как трофическая группа формирует риск исчезновения в целом, а также как она влияет на наши наблюдения относительного риска для различных трофических групп в наземной и водной средах.
Когда были проанализированы более подробные категории рациона, как рыбоядные, так и падальщики имели повышенный риск по сравнению с фоновыми уровнями. В соответствии с более высоким риском вымирания рыбоядных мы обнаружили, что повышенный риск вымирания хищников возникает почти исключительно в морских средах обитания, что позволяет предположить, что давление вымирания, угрожающее хищникам, может быть выше в океане, чем на суше.Прошлые исследования показали, что во многих регионах люди преимущественно и неустойчиво эксплуатируют морские организмы на вершине пищевой цепи ( 24 ). Однако это исследование было сосредоточено в основном на рыбе ( 24 ), которую мы не включили в наше исследование. Вероятным продуктивным будущим исследованием будет определение того, как включение костистых и хрящевых рыб и других таксономических классов, не включенных в это исследование (например, беспозвоночных), повлияет на интерпретацию того, как трофическая группа формирует риск исчезновения в целом, а также как она влияет на наши наблюдения относительного риска для различных трофических групп в наземной и водной средах. Хотя нам не удалось выявить ни одного антропогенного фактора глобального сокращения численности травоядных, мы обнаружили, что некоторые факторы непропорционально больше влияют на некоторые группы травоядных по сравнению с другими трофическими группами. Например, мы обнаружили, что инвазивные виды поражают травоядных рептилий непропорционально сильнее, чем всеядных и хищников. Инвазивные позвоночные (например, крысы), насекомые (например, огненные муравьи) и растения (например, готтентотский инжир) были причастны к упадку и даже исчезновению некоторых рептилий ( 25 ).Кроме того, мы обнаружили, что инвазивные виды, загрязнение и изменение среды обитания непропорционально сильно влияют на мелких травоядных птиц. Однако в обоих этих случаях неясно, почему эти антропогенные факторы больше нацелены на травоядных, чем на другие трофические группы. Новой задачей природоохранной биологии будет выявление четких механизмов, ответственных за эту очевидную связь между трофической группой и риском исчезновения.
Хотя нам не удалось выявить ни одного антропогенного фактора глобального сокращения численности травоядных, мы обнаружили, что некоторые факторы непропорционально больше влияют на некоторые группы травоядных по сравнению с другими трофическими группами. Например, мы обнаружили, что инвазивные виды поражают травоядных рептилий непропорционально сильнее, чем всеядных и хищников. Инвазивные позвоночные (например, крысы), насекомые (например, огненные муравьи) и растения (например, готтентотский инжир) были причастны к упадку и даже исчезновению некоторых рептилий ( 25 ).Кроме того, мы обнаружили, что инвазивные виды, загрязнение и изменение среды обитания непропорционально сильно влияют на мелких травоядных птиц. Однако в обоих этих случаях неясно, почему эти антропогенные факторы больше нацелены на травоядных, чем на другие трофические группы. Новой задачей природоохранной биологии будет выявление четких механизмов, ответственных за эту очевидную связь между трофической группой и риском исчезновения. Эти исследования, вероятно, будут включать исследования взаимодействующих эффектов нескольких антропогенных факторов и их влияния на присущие видам черты, связанные с травоядностью.Наше исследование показало, что трофическая группа является важным фактором риска исчезновения рептилий, млекопитающих и птиц. Прошлые исследования, изучавшие влияние трофической группы или рациона на риск вымирания, либо не выявили влияния трофической группы ( 26 ), либо наибольшему риску вымирания подвергаются хищники более высокого порядка ( 14 , 16 ). Эти прошлые исследования, однако, были сосредоточены на избранных группах организмов, таких как животные из отряда Carnivora ( 16 ) или чешуйчатые рептилии ( 26 ), предполагая, что эти более целенаправленные исследования маскируют более общую роль трофической группы как важной переменная, которая вместе влияет на модели вымирания млекопитающих, птиц и рептилий.Важно отметить, что наши результаты показывают, что любые будущие попытки объяснить закономерности риска исчезновения у млекопитающих, птиц и рептилий должны учитывать трофическую группу.
Эти исследования, вероятно, будут включать исследования взаимодействующих эффектов нескольких антропогенных факторов и их влияния на присущие видам черты, связанные с травоядностью.Наше исследование показало, что трофическая группа является важным фактором риска исчезновения рептилий, млекопитающих и птиц. Прошлые исследования, изучавшие влияние трофической группы или рациона на риск вымирания, либо не выявили влияния трофической группы ( 26 ), либо наибольшему риску вымирания подвергаются хищники более высокого порядка ( 14 , 16 ). Эти прошлые исследования, однако, были сосредоточены на избранных группах организмов, таких как животные из отряда Carnivora ( 16 ) или чешуйчатые рептилии ( 26 ), предполагая, что эти более целенаправленные исследования маскируют более общую роль трофической группы как важной переменная, которая вместе влияет на модели вымирания млекопитающих, птиц и рептилий.Важно отметить, что наши результаты показывают, что любые будущие попытки объяснить закономерности риска исчезновения у млекопитающих, птиц и рептилий должны учитывать трофическую группу. Помимо трофической группы, наше исследование показало, что размер тела является важной чертой, которая независимо влияет на риск исчезновения. у млекопитающих и птиц и взаимодействует с трофической группой, влияя на риск исчезновения у рептилий. Начиная с позднего плейстоцена средняя масса тела организмов снижалась из-за угроз, связанных с размером тела, которые тесно связаны с деятельностью человека ( 27 ).Наши результаты согласуются с выводами прошлых исследований о том, что селекция против крупных организмов, вероятно, продолжится при обычных сценариях управления ( 11 , 12 ). Наше исследование дополняет этот массив литературы, показывая, что, за исключением хищных рептилий, крупные организмы во всех трех трофических группах подвержены высокому риску исчезновения, но травоядные являются непропорционально наиболее подверженной трофической группе среди этих крупнотелых животных. разновидность. Экологические последствия исчезновения крупных травоядных будут в некоторой степени зависеть от способности более мелких видов компенсировать это численно и/или функционально ( 28 ).
Помимо трофической группы, наше исследование показало, что размер тела является важной чертой, которая независимо влияет на риск исчезновения. у млекопитающих и птиц и взаимодействует с трофической группой, влияя на риск исчезновения у рептилий. Начиная с позднего плейстоцена средняя масса тела организмов снижалась из-за угроз, связанных с размером тела, которые тесно связаны с деятельностью человека ( 27 ).Наши результаты согласуются с выводами прошлых исследований о том, что селекция против крупных организмов, вероятно, продолжится при обычных сценариях управления ( 11 , 12 ). Наше исследование дополняет этот массив литературы, показывая, что, за исключением хищных рептилий, крупные организмы во всех трех трофических группах подвержены высокому риску исчезновения, но травоядные являются непропорционально наиболее подверженной трофической группе среди этих крупнотелых животных. разновидность. Экологические последствия исчезновения крупных травоядных будут в некоторой степени зависеть от способности более мелких видов компенсировать это численно и/или функционально ( 28 ). Однако многие функции, зависящие от размера тела (например, разбрасывание ленивцами плодов с крупными семенами), не могут быть воспроизведены простым увеличением численности видов с меньшим телом ( 28 ). Доисторическое вымирание мегатравоядных резко изменило структуру и функционирование Экосистемы Земли за счет изменения динамики растительности, режимов пожаров, круговорота углерода и биогеохимического круговорота ( 7 – 9 ). Наши результаты показывают, что Земля снова переживает упадок и вымирание, которое непропорционально сильно затрагивает крупных травоядных.Однако то, как эти сокращения и вымирание травоядных могут повлиять на траекторию жизни на Земле, пока неизвестно, но исследования связывают современных травоядных с такими разнообразными экосистемными процессами, как эволюция черт растений и хищников, устойчивость/сопротивление экосистемы, питательные вещества. цикличность, режимы пожаров, динамика парниковых газов, регенерация растений и первичная продукция ( 29 ).
Однако многие функции, зависящие от размера тела (например, разбрасывание ленивцами плодов с крупными семенами), не могут быть воспроизведены простым увеличением численности видов с меньшим телом ( 28 ). Доисторическое вымирание мегатравоядных резко изменило структуру и функционирование Экосистемы Земли за счет изменения динамики растительности, режимов пожаров, круговорота углерода и биогеохимического круговорота ( 7 – 9 ). Наши результаты показывают, что Земля снова переживает упадок и вымирание, которое непропорционально сильно затрагивает крупных травоядных.Однако то, как эти сокращения и вымирание травоядных могут повлиять на траекторию жизни на Земле, пока неизвестно, но исследования связывают современных травоядных с такими разнообразными экосистемными процессами, как эволюция черт растений и хищников, устойчивость/сопротивление экосистемы, питательные вещества. цикличность, режимы пожаров, динамика парниковых газов, регенерация растений и первичная продукция ( 29 ).
. МАТЕРИАЛЫ И МЕТОДЫ. 1
), а также недавно вымершие (вымершие в дикой природе и вымершие в Красном списке видов, находящихся под угрозой исчезновения МСОП) млекопитающие, птицы и рептилии, а также вымершие млекопитающие позднего плейстоцена, полученные из McCauley et al. ( 2 ) и Smith и др. ( 27 ). Во-первых, мы присвоили бинарные оценки угроз каждому виду. Во-вторых, мы охарактеризовали рацион каждого вида на основе доступной информации. В-третьих, мы классифицировали трофические группы каждого вида на основе характеристик нашего рациона. Затем мы объединили эти данные с информацией о географическом местоположении, среде обитания и угрозах конкретных видов из Красного списка видов, находящихся под угрозой исчезновения МСОП. Мы использовали заполненную базу данных для количественной оценки наблюдаемой доли находящихся под угрозой исчезновения или вымерших видов в определенных трофических группах (т.э., хищник, всеядное и травоядное) по отношению друг к другу и фоновым фракциям исчезающих или вымерших видов.
Бинарная классификация угроз
Для каждого существующего вида, оцененного МСОП, мы присвоили бинарную классификацию угроз «находящиеся под угрозой исчезновения» или «не находящиеся под угрозой исчезновения». Виды с оценками Красного списка МСОП как «находящиеся под угрозой исчезновения», «находящиеся под угрозой исчезновения» и «уязвимые» считались находящимися под угрозой исчезновения, а виды с оценками «находящиеся под угрозой исчезновения», «наименее опасные» и «низкий риск» считались не находящимися под угрозой исчезновения.Эта система бинарной классификации представляет два возможных исхода события исчезновения (то есть вымирание или выживание) и согласуется с аналитическими подходами, ранее использовавшимися для изучения риска исчезновения ( 11 ). Виды, оцененные как «исчезнувшие» или «исчезнувшие в дикой природе», не были включены в анализ статуса, находящегося под угрозой исчезновения. Вместо этого они были классифицированы как недавно вымершие при анализе исторических моделей трофических групп. Виды, отнесенные МСОП к категории данных, по которым недостаточно данных, в нашем анализе не использовались.Млекопитающие из списков вымерших видов из McCauley et al. ( 2 ) и Smith и др. ( 27 ) были классифицированы как вымершие в позднем плейстоцене и использовались только при анализе исторических моделей трофических групп.
Виды, отнесенные МСОП к категории данных, по которым недостаточно данных, в нашем анализе не использовались.Млекопитающие из списков вымерших видов из McCauley et al. ( 2 ) и Smith и др. ( 27 ) были классифицированы как вымершие в позднем плейстоцене и использовались только при анализе исторических моделей трофических групп. Категории рациона
Мы классифицировали рацион всех видов млекопитающих, птиц и рептилий, оцененных МСОП, используя опубликованную литературу, справочные тексты, базы данных или экстраполяцию из соответствующего таксономического класса. Рационы видов определялись по разным типам данных (т.д., процент, ранжированное наличие-отсутствие и неранжированное наличие-отсутствие конкретных пищевых продуктов; таблицу S9). Мы классифицировали категории диеты, используя бинарную реакцию присутствия или отсутствия. Использовалась информация только о рационе взрослых особей из диких популяций. Основными категориями диеты (с подкатегориями в скобках) были эндотермы (млекопитающие и птицы), герптилии (земноводные и рептилии), рыбы, беспозвоночные (насекомые), падаль, растения (листва, корень и древесина), фрукты, нектар и зерно. ( 30 , 31 ).Падальеды (то есть падальщики) были определены как виды, которые потребляют падаль животного происхождения. Чтобы быть отнесенным к категории растений, вид должен потреблять водоросли, грибы, листья, побеги, корни, древесину, цветы, пыльцу, чашелистики или другой разнообразный растительный материал (например, растительное вещество) ( 30 ).
( 30 , 31 ).Падальеды (то есть падальщики) были определены как виды, которые потребляют падаль животного происхождения. Чтобы быть отнесенным к категории растений, вид должен потреблять водоросли, грибы, листья, побеги, корни, древесину, цветы, пыльцу, чашелистики или другой разнообразный растительный материал (например, растительное вещество) ( 30 ). Для диет, описанных в литературе как проценты, мы использовали определенные процентные отсечки, чтобы классифицировать категории диеты как имеющиеся или отсутствующие. Процентное отсечение было скорректировано на основе количества уникальных элементов рациона, включенных в рацион вида, потому что по мере увеличения количества элементов рациона увеличивается вес категории рациона.Например, если вид потреблял только два элемента рациона, а один элемент рациона составлял ≤20% рациона, то этот элемент классифицировался как отсутствующий. Однако, если в рационе вида было более двух элементов рациона, то любой элемент с вкладом ≥20% классифицировался как присутствующий.
Для диет, описанных как данные о присутствии-отсутствии, категории диет были классифицированы в соответствии с ключевыми словами. Мы включили элементы диеты, связанные со следующими ключевыми словами: «основной», «вторичный», «в основном», «также», «часто», «регулярно», «обычно» и «в достаточном количестве».Мы исключили продукты питания, связанные с ключевыми словами «третичный», «иногда», «иногда», «редко», «небольшие количества», «и даже», «по возможности» и «иногда с добавками». Если какой-либо элемент диеты был связан с неопределенными утверждениями «вероятно», «возможно», «может быть» или «предположительно», то он исключался, если не было никакой другой информации о диете. Для диет, описанных с использованием неранжированных данных присутствия-отсутствия (например, «питается фруктами, насекомыми и семенами»), все перечисленные элементы диеты были классифицированы как присутствующие.
Трофическая группа
Используя информацию о питании, мы разделили виды на три трофические группы: хищники, всеядные и травоядные. Для ранжированных данных о присутствии-отсутствии хищники были определены как животные, которые в основном потребляли продукты животного происхождения и могли потреблять растительные продукты в небольших количествах; травоядные в основном потребляли продукты растительного происхождения и могли потреблять продукты животного происхождения в небольших количествах, а всеядные в первую очередь или вторично потребляли смесь обоих типов продуктов питания.Для неранжированных данных о присутствии-отсутствии хищники потребляли только продукты животного происхождения, травоядные потребляли только растительные продукты, а рационы всеядных включали продукты из обеих категорий. Чтобы изучить чувствительность результатов к нашей классификации трофических групп, трофические группы были классифицированы тремя различными способами, в зависимости от того, как менялся вклад рациона травоядных или хищников: 70% отсечка, 80% отсечка и 90% отсечка. Результаты, описанные в основном тексте, представляют собой наиболее консервативный взгляд на хищников и травоядных, при этом хищники определяются как виды, потребляющие ≥90 % животного рациона, травоядные как виды, потребляющие ≥90 % растительного рациона, и всеядные как виды.
Для ранжированных данных о присутствии-отсутствии хищники были определены как животные, которые в основном потребляли продукты животного происхождения и могли потреблять растительные продукты в небольших количествах; травоядные в основном потребляли продукты растительного происхождения и могли потреблять продукты животного происхождения в небольших количествах, а всеядные в первую очередь или вторично потребляли смесь обоих типов продуктов питания.Для неранжированных данных о присутствии-отсутствии хищники потребляли только продукты животного происхождения, травоядные потребляли только растительные продукты, а рационы всеядных включали продукты из обеих категорий. Чтобы изучить чувствительность результатов к нашей классификации трофических групп, трофические группы были классифицированы тремя различными способами, в зависимости от того, как менялся вклад рациона травоядных или хищников: 70% отсечка, 80% отсечка и 90% отсечка. Результаты, описанные в основном тексте, представляют собой наиболее консервативный взгляд на хищников и травоядных, при этом хищники определяются как виды, потребляющие ≥90 % животного рациона, травоядные как виды, потребляющие ≥90 % растительного рациона, и всеядные как виды. которые потребляют от 11 до 89% животного или растительного рациона.Рацион 80 % определяет хищников как виды, потребляющие ≥80 % животного рациона, травоядных как виды, потребляющие ≥80 % растительного рациона, и всеядных как виды, потребляющие от 21 до 79 % животного или растительного рациона. . Рацион 70 % определяет хищников как виды, потребляющие ≥70 % животного рациона, травоядных как виды, потребляющие ≥70 % растительного рациона, и всеядных как виды, потребляющие от 31 до 69 % животного или растительного рациона. . В целом, наши выводы о том, как трофические группы формируют риск вымирания, были нечувствительны к нашим методам определения трофических групп (рис.С1).
которые потребляют от 11 до 89% животного или растительного рациона.Рацион 80 % определяет хищников как виды, потребляющие ≥80 % животного рациона, травоядных как виды, потребляющие ≥80 % растительного рациона, и всеядных как виды, потребляющие от 21 до 79 % животного или растительного рациона. . Рацион 70 % определяет хищников как виды, потребляющие ≥70 % животного рациона, травоядных как виды, потребляющие ≥70 % растительного рациона, и всеядных как виды, потребляющие от 31 до 69 % животного или растительного рациона. . В целом, наши выводы о том, как трофические группы формируют риск вымирания, были нечувствительны к нашим методам определения трофических групп (рис.С1).
 Чтобы подтвердить этот подход, мы сначала удалили все экстраполированные виды из нашего набора данных, а затем разделили оставшиеся данные на обучающие данные (75% данных) и тестовые данные (25%). Затем мы присвоили трофическую группу видам в нашем наборе тестовых данных, используя трофическую группу ближайшего живого родственника в наших обучающих данных. Мы повторили этот процесс для 1000 бутстрепных филогений, чтобы обеспечить среднюю точность (± SE), и сравнили наши экстраполированные результаты по трофическим группам с известными трофическими группами в исходном наборе данных.Мы обнаружили, что точность предсказания трофической группы млекопитающих, птиц и рептилий составила 94 % (±0,02), 82 % (±0,03) и 88 % (±0,05) соответственно. Смит и др. ( 27 ), была классифицирована только трофическая группа, а не диета из-за нехватки информации о конкретной диете. Трофическая группа была классифицирована на основе информации, собранной из первичной литературы, баз данных и ближайших существующих сородичей, с использованием протоколов, аналогичных описанной ранее диетической категоризации.
Чтобы подтвердить этот подход, мы сначала удалили все экстраполированные виды из нашего набора данных, а затем разделили оставшиеся данные на обучающие данные (75% данных) и тестовые данные (25%). Затем мы присвоили трофическую группу видам в нашем наборе тестовых данных, используя трофическую группу ближайшего живого родственника в наших обучающих данных. Мы повторили этот процесс для 1000 бутстрепных филогений, чтобы обеспечить среднюю точность (± SE), и сравнили наши экстраполированные результаты по трофическим группам с известными трофическими группами в исходном наборе данных.Мы обнаружили, что точность предсказания трофической группы млекопитающих, птиц и рептилий составила 94 % (±0,02), 82 % (±0,03) и 88 % (±0,05) соответственно. Смит и др. ( 27 ), была классифицирована только трофическая группа, а не диета из-за нехватки информации о конкретной диете. Трофическая группа была классифицирована на основе информации, собранной из первичной литературы, баз данных и ближайших существующих сородичей, с использованием протоколов, аналогичных описанной ранее диетической категоризации.
Регионы, местообитания и антропогенные глобальные изменения движут
Для изучения антропогенного воздействия на паттерны трофических групп и изменчивость классификаций угроз мы загрузили все внешние переменные, характерные для конкретных видов (т. е. места обитания, регионы и связанные с ними антропогенные угрозы) из Красного Список видов, находящихся под угрозой исчезновения. Основные географические (наземные и морские) регионы, места обитания и антропогенные угрозы были классифицированы как присутствующие или отсутствующие для каждого вида. Основные антропогенные факторы глобальных изменений, классифицированные МСОП, были упрощены до пяти основных категорий антропогенных угроз ( 3 ): изменение среды обитания (жилая и коммерческая застройка, сельское хозяйство и аквакультура, производство энергии и добыча полезных ископаемых, транспортные и сервисные коридоры, человеческое вторжение и нарушение или модификации природных систем), использование биологических ресурсов (например,г. , чрезмерная эксплуатация), изменение климата, загрязнение и инвазивные виды.
, чрезмерная эксплуатация), изменение климата, загрязнение и инвазивные виды. Размер тела
Мы использовали размер тела взрослого человека для классификации. Когда были даны размеры мужского и женского тела, мы использовали среднее геометрическое двух значений. Когда был задан диапазон размеров тела, мы использовали среднее геометрическое наименьшего и наибольшего значения ( 30 ). Масса тела была наиболее доступна для птиц и млекопитающих. Однако измерения размеров тела рептилий не были стандартизированы для разных таксономических отрядов. Размер тела Squamata обычно указывался как общая длина или длина рыла, в то время как размер тела Testudines указывался как стандартная длина панциря.Чтобы преодолеть нестандартные методы измерения размера тела, мы преобразовали все измерения длины в массу тела, используя ранее опубликованные регрессии длины тела и массы на уровне семьи. Ссылки на регрессии длины и массы приведены в дополнительных материалах. Если ранее опубликованные регрессии были недоступны, то мы разрабатывали регрессии по видам в наших данных, которые содержали данные как о массе тела, так и о длине. Для видов с отсутствующими данными о размерах тела мы экстраполировали размер тела, используя среднее значение всех сородичей (за исключением сородичей, которые ранее были экстраполированы таким образом).
Для видов с отсутствующими данными о размерах тела мы экстраполировали размер тела, используя среднее значение всех сородичей (за исключением сородичей, которые ранее были экстраполированы таким образом).Анализы
Красный список видов, находящихся под угрозой исчезновения МСОП, состоит из неполных данных, поскольку многие виды классифицируются как неполные данные, а другие вообще не описаны. Из-за несовершенства данных МСОП мы использовали имеющиеся данные для получения средних оценок и 95% доверительного интервала пропорций видов, находящихся под угрозой исчезновения, для различных трофических групп. Поэтому, хотя мы можем не знать точную истинную долю видов, находящихся под угрозой исчезновения, для каждой трофической группы, наши доверительные интервалы указывают границы, в пределах которых, вероятно, находится истинное количество.Мы исследовали закономерности доли видов, находящихся под угрозой исчезновения, для каждой трофической группы (травоядные, всеядные и хищники) в глобальном масштабе для существующих видов позвоночных (млекопитающих, птиц и рептилий), недавно вымерших видов позвоночных (млекопитающих, птиц и рептилий). и плейстоценовые вымершие виды позвоночных (только млекопитающие). Различия в долях видов, находящихся под угрозой исчезновения, среди трофических групп были проанализированы с использованием GLM, при этом статус угрозы использовался в качестве переменной реакции, а трофическая группа — в качестве переменной-предиктора.Мы использовали биномиальную структуру ошибки (угрожаемая или неугрожаемая) и функция логит-ссылки. Каждый вид рассматривался как единственная повторность и регистрировался либо как находящийся под угрозой исчезновения, либо как вымерший или нет, если были изучены исторические закономерности. Вслед за GLM были использованы апостериорные тесты Тьюки для выявления различий между трофическими группами в доле исчезающих или вымерших видов. Для каждой трофической группы мы произвели средние оценки и 95% ДИ доли видов, находящихся под угрозой исчезновения, используя функцию «прогнозировать» в R.Эти 95% доверительные интервалы сравнивались с долей находящихся под угрозой исчезновения или вымерших видов на фоновых уровнях, которые рассчитывались путем деления количества находящихся под угрозой исчезновения видов на общее количество видов в этом анализе.
и плейстоценовые вымершие виды позвоночных (только млекопитающие). Различия в долях видов, находящихся под угрозой исчезновения, среди трофических групп были проанализированы с использованием GLM, при этом статус угрозы использовался в качестве переменной реакции, а трофическая группа — в качестве переменной-предиктора.Мы использовали биномиальную структуру ошибки (угрожаемая или неугрожаемая) и функция логит-ссылки. Каждый вид рассматривался как единственная повторность и регистрировался либо как находящийся под угрозой исчезновения, либо как вымерший или нет, если были изучены исторические закономерности. Вслед за GLM были использованы апостериорные тесты Тьюки для выявления различий между трофическими группами в доле исчезающих или вымерших видов. Для каждой трофической группы мы произвели средние оценки и 95% ДИ доли видов, находящихся под угрозой исчезновения, используя функцию «прогнозировать» в R.Эти 95% доверительные интервалы сравнивались с долей находящихся под угрозой исчезновения или вымерших видов на фоновых уровнях, которые рассчитывались путем деления количества находящихся под угрозой исчезновения видов на общее количество видов в этом анализе. Если 95% ДИ не пересекал фоновую фракцию видов, находящихся под угрозой исчезновения (т. е. фоновый уровень), то трофическая группа определялась как имеющая более высокую или меньшую долю исчезающих видов, чем фоновая фракция. После проведения анализа по всем таксонам из Красного списка исчезающих видов МСОП мы разделили данные на основе таксономического класса и запустили отдельные модели для каждого класса отдельно.
Если 95% ДИ не пересекал фоновую фракцию видов, находящихся под угрозой исчезновения (т. е. фоновый уровень), то трофическая группа определялась как имеющая более высокую или меньшую долю исчезающих видов, чем фоновая фракция. После проведения анализа по всем таксонам из Красного списка исчезающих видов МСОП мы разделили данные на основе таксономического класса и запустили отдельные модели для каждого класса отдельно.
Чтобы понять взаимосвязь между конкретными типами рациона и долей исчезающих видов, мы провели последующие анализы, в которых каждый тип рациона рассматривался как бинарная описательная переменная. Этот метод означал, что каждый вид можно было классифицировать по нескольким категориям рациона (т. Е. Травоядное животное можно было классифицировать как потребляющее нектар и зерно), а это означало, что категории рациона не были взаимоисключающими. Мы использовали GLM, чтобы понять, как различные категории рациона сравниваются с фоновой долей видов, находящихся под угрозой исчезновения, и снова использовали функцию прогнозирования для получения средних значений и 95% ДИ для каждого типа рациона. Затем эти 95% доверительные интервалы для каждого конкретного рациона сравнивали с фоновой долей исчезающих видов.
Затем эти 95% доверительные интервалы для каждого конкретного рациона сравнивали с фоновой долей исчезающих видов.
После глобального анализа мы разделили наши данные на основе типа среды обитания и географического региона и повторили анализ, чтобы понять, как модели состояния угрозы различных трофических групп различаются в зависимости от географических регионов и типов среды обитания. Для каждого места обитания или географического региона мы повторили методы, описанные выше для глобального анализа. Во-первых, мы использовали GLM и апостериорный тест Тьюки для оценки различий в доле видов, находящихся под угрозой исчезновения, среди трофических групп для каждого местообитания или региона.Затем мы использовали функцию прогнозирования для получения средних долей и 95% ДИ для каждого таксономического класса и сравнили их с долей видов, находящихся под угрозой исчезновения в этом таксономическом классе (всех видов, млекопитающих, птиц или рептилий) по всему миру. Все анализы проводились в R.
 Мы взяли 1000 филогений для каждой таксономической группы — Faurby и Svenning ( 32 ) для млекопитающих, Jetz et al. ( 34 ) для птиц и Tonini et al. ( 33 ) для рептилий — и повторил все анализы для них, сообщив оценки средних параметров для всех этих моделей. Чтобы убедиться, что наш признак и филогенетические данные совместимы, мы сопоставили все филогенетические данные с таксономией, используемой МСОП (то есть с основой наших данных о признаке; см. выше), используя Taxize ( 40 ).Мы использовали информационный критерий Акаике для выбора наиболее экономичной модели. Коды R для повторения наших анализов приведены в дополнительных материалах.
Мы взяли 1000 филогений для каждой таксономической группы — Faurby и Svenning ( 32 ) для млекопитающих, Jetz et al. ( 34 ) для птиц и Tonini et al. ( 33 ) для рептилий — и повторил все анализы для них, сообщив оценки средних параметров для всех этих моделей. Чтобы убедиться, что наш признак и филогенетические данные совместимы, мы сопоставили все филогенетические данные с таксономией, используемой МСОП (то есть с основой наших данных о признаке; см. выше), используя Taxize ( 40 ).Мы использовали информационный критерий Акаике для выбора наиболее экономичной модели. Коды R для повторения наших анализов приведены в дополнительных материалах. Острота зрения у млекопитающих: влияние размера глаза и экология — Полный текст — Мозг, поведение и эволюция 2014, Vol. 83, No. 1
Предыдущие сравнительные исследования связывали межвидовые различия в размере глаз у млекопитающих с отбором, связанным с остротой зрения. Также было высказано предположение, что виды млекопитающих различаются по остроте зрения отчасти из-за различий в экологии. В то время как в ряде предыдущих исследований изучались экологические и филогенетические эффекты на форму глаза, в настоящее время отсутствует широкий сравнительный анализ взаимосвязей между остротой зрения, размером глаза и экологией у млекопитающих. Здесь мы используем филогенетические сравнительные методы для изучения этих отношений в таксономически и экологически разнообразной выборке из 91 вида млекопитающих. Эти данные подтверждают, что осевая длина глаза и острота зрения значительно положительно коррелируют у млекопитающих. Это соотношение соответствует ожиданиям, основанным на теоретической оптике и предварительном анализе меньших сравнительных образцов.Наши данные также показывают, что более высокая острота зрения у млекопитающих связана с: (1) дневным образом жизни и (2) хищническими привычками, если влияние размера глаза и филогенеза статистически контролируется. Эти результаты предполагают, что межвидовая изменчивость остроты зрения млекопитающих является результатом сложного взаимодействия между филогенетической историей, визуальной анатомией и экологией.
В то время как в ряде предыдущих исследований изучались экологические и филогенетические эффекты на форму глаза, в настоящее время отсутствует широкий сравнительный анализ взаимосвязей между остротой зрения, размером глаза и экологией у млекопитающих. Здесь мы используем филогенетические сравнительные методы для изучения этих отношений в таксономически и экологически разнообразной выборке из 91 вида млекопитающих. Эти данные подтверждают, что осевая длина глаза и острота зрения значительно положительно коррелируют у млекопитающих. Это соотношение соответствует ожиданиям, основанным на теоретической оптике и предварительном анализе меньших сравнительных образцов.Наши данные также показывают, что более высокая острота зрения у млекопитающих связана с: (1) дневным образом жизни и (2) хищническими привычками, если влияние размера глаза и филогенеза статистически контролируется. Эти результаты предполагают, что межвидовая изменчивость остроты зрения млекопитающих является результатом сложного взаимодействия между филогенетической историей, визуальной анатомией и экологией.
© 2014 S. Karger AG, Базель
Введение
Млекопитающие демонстрируют значительные различия в остроте зрения (т.е. способность различать пространственные детали), начиная от низкой остроты зрения у рукокрылых летучих мышей и мелких грызунов (0,4–1,0 циклов на градус, или имп/д) до очень острого зрения (30–64 имп/д) у дневных человекообразных приматов [Walls, 1942]. ; Петтигрю и др., 1988; Кирк и Кей, 2004]. Предполагается, что эволюция более высокой остроты зрения у некоторых клад (например, приматов) отражает возросшую зависимость от зрения при удовлетворении основных потребностей, таких как поиск пищи и избегание хищников [Walls, 1942; Хьюз, 1977; Кей и Кирк, 2000 г .; Росс, 2000; Кирк и Кей, 2004 г.; Вейе и Кирк, 2009 г .; Холл и др., 2012]. В то же время более низкая острота зрения у некоторых других ветвей млекопитающих (например, у рукокрылых летучих мышей, грызунов) может отражать повышенную зависимость от незрительных органов чувств [Walls, 1942; Хатчеон и др. , 2002; Даймонд и др., 2008 г.; Хайден и др., 2010]. Таким образом, выявление факторов, влияющих на остроту зрения, имеет решающее значение для понимания сенсорной экологии млекопитающих.
, 2002; Даймонд и др., 2008 г.; Хайден и др., 2010]. Таким образом, выявление факторов, влияющих на остроту зрения, имеет решающее значение для понимания сенсорной экологии млекопитающих.
Теоретическая оптика предсказывает, что для глаз млекопитающих абсолютный размер глаза будет влиять на ряд различных аспектов зрительной функции [Walls, 1942; Хьюз, 1977].В частности, осевой диаметр глаза («длина глаза») играет ключевую роль в определении размера изображения на сетчатке [Hughes, 1977]. Если форма глаза остается неизменной, увеличение длины глаза приводит к увеличению расстояния до заднего узла (PND) и увеличению размера изображения на сетчатке (рис. 1) [Hughes, 1977; Петтигрю и др., 1988; Росс, 2000]. Более крупные изображения сетчатки, как правило, получают большее количество независимых единиц выборки (т. е. рецептивных полей ганглиозных клеток), так что длина глаза должна положительно коррелировать с остротой зрения [Walls, 1942; Киршфельд, 1976; Хьюз, 1977; Хизи и Холл, 2010]. В результате в исследованиях, в которых острота зрения рассчитывается на основе PND и пиковой плотности ганглиозных клеток, для оценки PND обычно используются измерения длины глаза [Hughes, 1977; Петтигрю и др., 1988; Арресе и др., 1999; Петтигрю и др., 2010]. Поэтому ожидается, что виды с абсолютно более длинными глазами будут иметь более высокую остроту зрения, чем виды с более короткими глазами. Некоторое подтверждение этого предсказания было обеспечено сравнительным анализом остроты зрения и длины глаза в смешанных выборках млекопитающих и птиц [Kiltie, 2000; Хизи и Холл, 2010].Однако в этих исследованиях использовались относительно небольшие образцы млекопитающих с ограниченным таксономическим и экологическим разнообразием. Например, Kiltie [2000] продемонстрировал положительную корреляцию между остротой зрения и длиной глаза в выборке из 14 ночных млекопитающих и 3 дневных антропоидов. Аналогичный результат был получен Heesy and Hall [2010] с использованием выборки из 14 ночных и 11 дневных видов млекопитающих, но связь между остротой зрения и длиной глаза была менее линейной, чем показанная Kiltie [2000].
В результате в исследованиях, в которых острота зрения рассчитывается на основе PND и пиковой плотности ганглиозных клеток, для оценки PND обычно используются измерения длины глаза [Hughes, 1977; Петтигрю и др., 1988; Арресе и др., 1999; Петтигрю и др., 2010]. Поэтому ожидается, что виды с абсолютно более длинными глазами будут иметь более высокую остроту зрения, чем виды с более короткими глазами. Некоторое подтверждение этого предсказания было обеспечено сравнительным анализом остроты зрения и длины глаза в смешанных выборках млекопитающих и птиц [Kiltie, 2000; Хизи и Холл, 2010].Однако в этих исследованиях использовались относительно небольшие образцы млекопитающих с ограниченным таксономическим и экологическим разнообразием. Например, Kiltie [2000] продемонстрировал положительную корреляцию между остротой зрения и длиной глаза в выборке из 14 ночных млекопитающих и 3 дневных антропоидов. Аналогичный результат был получен Heesy and Hall [2010] с использованием выборки из 14 ночных и 11 дневных видов млекопитающих, но связь между остротой зрения и длиной глаза была менее линейной, чем показанная Kiltie [2000].
Рис.1
Влияние размера глаза на размер изображения на сетчатке. В этом схематическом примере два глаза имеют одинаковую форму, но отличаются осевым диаметром. Здесь больший глаз имеет осевой диаметр примерно в два раза больше, чем меньший глаз. Оба глаза расположены так, что визуальная цель (двунаправленная стрелка) длины Y находится на одинаковом расстоянии от задней узловой точки (черный кружок; положение приблизительное). В этом примере изображение цели на сетчатке большого глаза (2X) в два раза длиннее изображения на сетчатке меньшего глаза (X).
Сравнительные исследования длины глаза и других аспектов зрительной морфологии связывают повышенную остроту зрения с рядом экологических факторов, в том числе с характером диальной активности, питанием и скоростью передвижения [Walls, 1942; Хьюз, 1977; Петтигрю и др., 1988; Арресе и др., 1999; Кирк и Кей, 2004 г.; Пайхл, 2005; Херд-Бут и Кирк, 2012]. Характер активности диэля (т. е. время суток, когда вид обычно активен), в частности, предсказывает тесную связь с остротой зрения, поскольку адаптации, повышающие зрительную чувствительность при слабом освещении, обычно несовместимы с высокой остротой зрения [Walls, 1942; Хьюз, 1977; Лэнд и Нильссон, 2012]. Следовательно, ожидается, что дневные млекопитающие будут иметь более высокую остроту зрения, чем катемерные или ночные виды [Walls, 1942; Хьюз, 1977; Петтигрю и др., 1988; Хизи и Холл, 2010]. Сравнительные исследования в некоторой степени подтвердили это предсказание у сумчатых [Arrese et al., 1999; Арресе и др., 2000; Arrese et al., 2002] и стрепсириновых приматов [Veilleux, Kirk, 2009]. Kiltie [2000] также сообщил, что паттерн активности diel оказывает сильное влияние на связь между остротой зрения и длиной глаза, но выборка дневных млекопитающих в этом анализе полностью состояла из антропоидных приматов.Дневные антропоиды имеют сильно развитую зрительную анатомию и значительно более высокую остроту зрения, чем другие дневные млекопитающие [Ross, 2000; Кирк, 2004 г.; Кирк и Кей, 2004 г.; Росс и Кирк, 2007 г .; Hall et al., 2012], предполагая, что разница в остроте зрения между дневными и ночными млекопитающими, наблюдаемая Kiltie [2010], может быть связана с факторами, отличными от паттерна активности diel как такового.
Следовательно, ожидается, что дневные млекопитающие будут иметь более высокую остроту зрения, чем катемерные или ночные виды [Walls, 1942; Хьюз, 1977; Петтигрю и др., 1988; Хизи и Холл, 2010]. Сравнительные исследования в некоторой степени подтвердили это предсказание у сумчатых [Arrese et al., 1999; Арресе и др., 2000; Arrese et al., 2002] и стрепсириновых приматов [Veilleux, Kirk, 2009]. Kiltie [2000] также сообщил, что паттерн активности diel оказывает сильное влияние на связь между остротой зрения и длиной глаза, но выборка дневных млекопитающих в этом анализе полностью состояла из антропоидных приматов.Дневные антропоиды имеют сильно развитую зрительную анатомию и значительно более высокую остроту зрения, чем другие дневные млекопитающие [Ross, 2000; Кирк, 2004 г.; Кирк и Кей, 2004 г.; Росс и Кирк, 2007 г .; Hall et al., 2012], предполагая, что разница в остроте зрения между дневными и ночными млекопитающими, наблюдаемая Kiltie [2010], может быть связана с факторами, отличными от паттерна активности diel как такового. Действительно, данные, представленные Heesy and Hall [2010], по-видимому, показывают небольшую разницу между дневными и ночными неантропоидными млекопитающими в соотношении между длиной глаза и остротой зрения.Соответственно, взаимосвязь между паттерном диальной активности и остротой зрения у млекопитающих остается малоизученной.
Действительно, данные, представленные Heesy and Hall [2010], по-видимому, показывают небольшую разницу между дневными и ночными неантропоидными млекопитающими в соотношении между длиной глаза и остротой зрения.Соответственно, взаимосвязь между паттерном диальной активности и остротой зрения у млекопитающих остается малоизученной.
Также предполагается, что другие экологические факторы влияют на изменение остроты зрения млекопитающих. Например, предсказывается, что млекопитающие, участвующие в зрительной охоте, будут иметь более высокую остроту зрения, чем более травоядные виды [Arrese et al., 1999, 2000; Росс, 2000; Тетро и др., 2004 г.; Кирк, 2006а; Росс и Кирк, 2007 г .; Хизи, 2008 г .; Veilleux and Kirk, 2009]. Наблюдение, что максимальная скорость бега (MRS) и длина глаза сильно положительно коррелируют у млекопитающих, также привело к предположению, что скорость передвижения влияет на остроту зрения [Hughes, 1977; Херд-Бут и Кирк, 2012].Однако эти гипотезы о влиянии диеты и передвижения на остроту зрения не были проверены в широком сравнительном анализе.
В то время как предыдущие сравнительные исследования задокументировали филогенетические и экологические влияния на размер глаз млекопитающих [Ross and Kirk, 2007; Heard-Booth and Kirk, 2012], в этих исследованиях в целом предполагалось, что эволюция увеличения размеров глаз функционально связана с отбором на более высокую остроту зрения. Здесь мы проверяем это ожидание, исследуя взаимосвязь между длиной глаза и остротой зрения в таксономически и экологически разнообразной выборке из 91 вида из 14 отрядов млекопитающих.После статистического контроля филогении и межвидовых различий в размере глаз мы затем оцениваем взаимосвязь между остротой зрения и характером диетической активности, диетой и MRS. При этом мы преследуем две основные цели. Во-первых, мы стремимся точно определить влияние размера глаза на остроту зрения у млекопитающих, используя большой сравнительный набор данных, который менее ограничен предвзятостью выборки, чем предыдущие анализы. Во-вторых, мы стремимся определить, связаны ли дневной образ жизни, активное хищничество и высокая скорость бега с более высокой остротой зрения даже после учета влияния размера глаза на остроту зрения.
Материалы и методы
Сравнительный набор данных
Мы собрали опубликованные данные по максимальной остроте зрения, длине глаза и массе тела для 91 вида млекопитающих (онлайн-приложение, таблица 1; см. www.karger.com/doi/10.1159/000357830 для всех онлайн-приложение). Мы включили как остроту зрения, измеренную на основе поведения, так и оценки остроты зрения, основанные на анатомии сетчатки и размере глаза. Когда для вида были доступны как анатомическая, так и поведенческая острота зрения, мы использовали поведенческие измерения, потому что они лучше отражают зрительную функцию [Arrese et al., 2000]. Анатомические оценки остроты зрения представляют собой теоретический максимум и имеют тенденцию слегка завышать остроту зрения, измеренную по поведенческим признакам [Pettigrew et al., 1988; Arrese и др., 1999].
Для 14 видов (Bos taurus, Capra hircus, Dama dama, Sus scrofa, Acinonyx jubatus, Canis lupus, Sarcophilus harrisi, Didelphis virginiana, Macropus fuliginosus, Setonix brachyurus, Alouatta caraya, Chlorocebus aethiops, Saguinus midas ) , мы модифицировали установленные протоколы для расчета анатомической остроты зрения на основе опубликованных данных о длине глаза и пиковой плотности ганглиозных клеток сетчатки или пиковой плотности колбочек [Pettigrew et al. , 1988; Arrese и др., 1999]. Вслед за Петтигрю и соавт. [1988] мы сначала оценили PND по длине глаза, используя множимое k , которое различается в зависимости от паттерна диэльной активности. Разные исследователи предлагали разные значения для k . Hughes [1977] первоначально использовал k = 0,60 для всех таксонов, которые Pettigrew et al. [1988] впоследствии были пересмотрены для учета различий в форме глаз с характером активности ( k = 0,52, 0,57 и 0,67 для ночных, катемерных и дневных видов соответственно) на основе сравнительной выборки 14 видов позвоночных.Этот метод оценки PND объясняет тот факт, что морфология глаз у млекопитающих предсказуемо меняется в зависимости от характера диальной активности [Kirk, 2004; Холл и др., 2012]. Совсем недавно Schmitz [2009] использовал расширенный набор данных по 23 видам, чтобы пересмотреть эти значения k до 0,55, 0,63 и 0,66 для ночных, катемерных и дневных видов соответственно. Однако этот источник неправильно классифицировал страусов (Struthio camelus) как катемерных, а не дневных [Williams, 1993], а европейских кроликов (Oryctolagus cuniculus) как ночных, а не катемерных [Lombardi et al.
, 1988; Arrese и др., 1999]. Вслед за Петтигрю и соавт. [1988] мы сначала оценили PND по длине глаза, используя множимое k , которое различается в зависимости от паттерна диэльной активности. Разные исследователи предлагали разные значения для k . Hughes [1977] первоначально использовал k = 0,60 для всех таксонов, которые Pettigrew et al. [1988] впоследствии были пересмотрены для учета различий в форме глаз с характером активности ( k = 0,52, 0,57 и 0,67 для ночных, катемерных и дневных видов соответственно) на основе сравнительной выборки 14 видов позвоночных.Этот метод оценки PND объясняет тот факт, что морфология глаз у млекопитающих предсказуемо меняется в зависимости от характера диальной активности [Kirk, 2004; Холл и др., 2012]. Совсем недавно Schmitz [2009] использовал расширенный набор данных по 23 видам, чтобы пересмотреть эти значения k до 0,55, 0,63 и 0,66 для ночных, катемерных и дневных видов соответственно. Однако этот источник неправильно классифицировал страусов (Struthio camelus) как катемерных, а не дневных [Williams, 1993], а европейских кроликов (Oryctolagus cuniculus) как ночных, а не катемерных [Lombardi et al. , 2003]. Поэтому мы повторно классифицировали эти два вида, получив общую сравнительную выборку из 6 ночных, 6 катемерных и 11 дневных видов позвоночных, для которых как PND, так и длину глаза можно использовать для расчета k [ср. таблица 1 в Schmitz, 2009]. Таким образом, наши пересмотренные значения для k , основанные на этой выборке, составляют 0,547 для ночных видов, 0,623 для катемерных видов и 0,643 для дневных видов. После оценки PND на основе длины глаза мы рассчитали коэффициент увеличения сетчатки (RMF) как (2π·PND)/360.У большинства млекопитающих множественные фоторецепторы колбочек могут объединять свои входы в каждую ганглиозную клетку (т. е. суммировать сетчатку) даже в центральной части сетчатки. В результате плотность ганглиозных клеток представляет собой фактор, ограничивающий остроту зрения [Pettigrew et al., 1988; Кей и Кирк, 2000]. Напротив, дневные гаплорины не обнаруживают суммирования сетчатки в центральной ямке [Kirk and Kay, 2004], поэтому острота зрения ограничивается плотностью колбочек [Pettigrew et al.
, 2003]. Поэтому мы повторно классифицировали эти два вида, получив общую сравнительную выборку из 6 ночных, 6 катемерных и 11 дневных видов позвоночных, для которых как PND, так и длину глаза можно использовать для расчета k [ср. таблица 1 в Schmitz, 2009]. Таким образом, наши пересмотренные значения для k , основанные на этой выборке, составляют 0,547 для ночных видов, 0,623 для катемерных видов и 0,643 для дневных видов. После оценки PND на основе длины глаза мы рассчитали коэффициент увеличения сетчатки (RMF) как (2π·PND)/360.У большинства млекопитающих множественные фоторецепторы колбочек могут объединять свои входы в каждую ганглиозную клетку (т. е. суммировать сетчатку) даже в центральной части сетчатки. В результате плотность ганглиозных клеток представляет собой фактор, ограничивающий остроту зрения [Pettigrew et al., 1988; Кей и Кирк, 2000]. Напротив, дневные гаплорины не обнаруживают суммирования сетчатки в центральной ямке [Kirk and Kay, 2004], поэтому острота зрения ограничивается плотностью колбочек [Pettigrew et al. , 1988]. Соответственно, мы рассчитали остроту зрения для дневных гаплоринов по формуле (RMF·√пиковая плотность колбочек)/2, а для всех других таксонов — по формуле (RMF·√пиковая плотность ганглиозных клеток)/2 [Pettigrew et al., 1988].
, 1988]. Соответственно, мы рассчитали остроту зрения для дневных гаплоринов по формуле (RMF·√пиковая плотность колбочек)/2, а для всех других таксонов — по формуле (RMF·√пиковая плотность ганглиозных клеток)/2 [Pettigrew et al., 1988].
Там, где это было возможно, мы собрали данные о характере диетической активности, MRS и рационе для каждого вида (онлайн-приложение, таблица 1). Паттерн диэльной активности был классифицирован как дневной, катемерный или ночной [Ross and Kirk, 2007; Холл и др., 2012]. Мы использовали самый высокий MRS, зарегистрированный для доступных видов. Мы ограничили наш диетический анализ сравнением травоядных видов («травоядных») и видов, описанных в литературе как активно ловящих движущуюся добычу («активный хищник»), следуя категориям, аналогичным Garamszegi et al.[2002]. Мы исключили виды, для которых стиль охоты был недоступен.
Анализ остроты зрения и длины глаза
Мы использовали филогенетический обобщенный метод наименьших квадратов (PGLS) для исследования связи между длиной глаза и остротой зрения [Garland and Ives, 2000]. Анализ PGLS выполнялся в R v.2.15.2 [R Development Core Team, 2011] с использованием пакетов geiger и caper [Orme et al., 2010; Хармон и др., 2008]. Топология дерева и длина ветвей соответствуют Bininda-Emonds et al.[2007]. Поскольку длина глаза участвует в расчетах анатомической остроты зрения [хотя и преобразуется с коэффициентом k в соответствии с паттерном диэльной активности; Pettigrew et al., 1988], существует некоторая озабоченность по поводу цикличности при использовании анатомически полученных оценок для изучения взаимосвязи между остротой зрения и длиной глаза. Ранее Килти [2000, с. 229] обнаружили, что результаты «значительно не отличались» при использовании анатомических или поведенческих оценок для изучения взаимосвязи между длиной глаза и остротой зрения.Чтобы сравнить наши наборы поведенческих и анатомических данных, мы рассчитали отдельные регрессии PGLS: (1) поведенческая острота по длине глаза, (2) анатомическая острота по длине глаза и (3) комбинированный набор данных поведенческой и анатомической остроты по длине глаза.
Анализ PGLS выполнялся в R v.2.15.2 [R Development Core Team, 2011] с использованием пакетов geiger и caper [Orme et al., 2010; Хармон и др., 2008]. Топология дерева и длина ветвей соответствуют Bininda-Emonds et al.[2007]. Поскольку длина глаза участвует в расчетах анатомической остроты зрения [хотя и преобразуется с коэффициентом k в соответствии с паттерном диэльной активности; Pettigrew et al., 1988], существует некоторая озабоченность по поводу цикличности при использовании анатомически полученных оценок для изучения взаимосвязи между остротой зрения и длиной глаза. Ранее Килти [2000, с. 229] обнаружили, что результаты «значительно не отличались» при использовании анатомических или поведенческих оценок для изучения взаимосвязи между длиной глаза и остротой зрения.Чтобы сравнить наши наборы поведенческих и анатомических данных, мы рассчитали отдельные регрессии PGLS: (1) поведенческая острота по длине глаза, (2) анатомическая острота по длине глаза и (3) комбинированный набор данных поведенческой и анатомической остроты по длине глаза. Для всех регрессий PGLS мы рассчитали лямбду Пагеля [Freckleton et al., 2002] в каперсах, чтобы оценить влияние филогенетических отношений на наши данные.
Для всех регрессий PGLS мы рассчитали лямбду Пагеля [Freckleton et al., 2002] в каперсах, чтобы оценить влияние филогенетических отношений на наши данные.
Поскольку приматы haplorhine проявляют специализацию по высокой остроте зрения, уникальную среди млекопитающих [e.г., ямка сетчатки; Kirk, Kay, 2004], мы ожидаем, что гаплорины будут отличаться от других млекопитающих по соотношению между длиной глаза и остротой зрения [Kiltie, 2000; Хизи и Холл, 2010]. Соответственно, мы сравнили остатки haplorhine приматов, strepsirrhine приматов и млекопитающих, не являющихся приматами, из комбинированной линии регрессии остроты зрения PGLS по длине глаза. Различия в остатках этих трех групп оценивались с использованием критерия Крускала-Уоллиса с последующим односторонним апостериорным критерием Вилкоксона.
Анализы остроты зрения и экологии
Затем мы изучили влияние характера диеты, диеты и MRS на взаимосвязь между остротой зрения и длиной глаза.В частности, мы проверили, влияют ли эти экологические факторы на изменение остроты зрения, когда филогения и длина глаза статистически контролировались. Мы ограничили этот экологический анализ набором данных о поведенческой остроте. Несколько групп таксонов, которые были включены в наше исследование длины и остроты зрения, были исключены из нашего экологического анализа. Во-первых, полуводные и роющие виды были исключены из всех экологических анализов, потому что они обитают в визуальной среде, принципиально отличной от той, с которой сталкиваются другие виды млекопитающих в нашей выборке.Во-вторых, мы исключили haplorhine приматов из всех экологических анализов, потому что они демонстрируют набор производных адаптаций для очень высокой остроты зрения, которые не обнаружены у других млекопитающих [Ross, 2000; Кирк, 2004 г.; Кирк и Кей, 2004 г.; Росс и Кирк, 2007 г .; Холл и др., 2012]. В-третьих, мы исключили летучих мышей-микрокрылых из нашего анализа рациона, потому что у них развились альтернативные сенсорные приспособления (активная эхолокация) для охоты на движущуюся добычу [Schnitzler and Kalko, 2001]. С этими исключениями мы попытались ограничить наши экологические сравнения группами видов, которые принципиально не различаются по: (1) визуальному плану или (2) визуальной экологии.
Мы ограничили этот экологический анализ набором данных о поведенческой остроте. Несколько групп таксонов, которые были включены в наше исследование длины и остроты зрения, были исключены из нашего экологического анализа. Во-первых, полуводные и роющие виды были исключены из всех экологических анализов, потому что они обитают в визуальной среде, принципиально отличной от той, с которой сталкиваются другие виды млекопитающих в нашей выборке.Во-вторых, мы исключили haplorhine приматов из всех экологических анализов, потому что они демонстрируют набор производных адаптаций для очень высокой остроты зрения, которые не обнаружены у других млекопитающих [Ross, 2000; Кирк, 2004 г.; Кирк и Кей, 2004 г.; Росс и Кирк, 2007 г .; Холл и др., 2012]. В-третьих, мы исключили летучих мышей-микрокрылых из нашего анализа рациона, потому что у них развились альтернативные сенсорные приспособления (активная эхолокация) для охоты на движущуюся добычу [Schnitzler and Kalko, 2001]. С этими исключениями мы попытались ограничить наши экологические сравнения группами видов, которые принципиально не различаются по: (1) визуальному плану или (2) визуальной экологии. Кроме того, важно отметить, что некоторые фауноядные и всеядные млекопитающие могут в большей степени полагаться на незрительные органы чувств во время кормодобывания [Walls, 1942; Хатчеон и др., 2002; Даймонд и др., 2008 г.; Хайден и др., 2010]. Соответственно, мы ограничили наши диетические анализы сравнением травоядных с хищниками, которые активно ловят движущуюся добычу. Ожидается, что в последней группе зрение будет играть важную роль в компенсации уклончивых движений добычи.
Кроме того, важно отметить, что некоторые фауноядные и всеядные млекопитающие могут в большей степени полагаться на незрительные органы чувств во время кормодобывания [Walls, 1942; Хатчеон и др., 2002; Даймонд и др., 2008 г.; Хайден и др., 2010]. Соответственно, мы ограничили наши диетические анализы сравнением травоядных с хищниками, которые активно ловят движущуюся добычу. Ожидается, что в последней группе зрение будет играть важную роль в компенсации уклончивых движений добычи.
Мы использовали два подхода для изучения воздействия окружающей среды на остроту зрения: (1) традиционные непараметрические тесты и (2) многомерное моделирование PGLS.Для непараметрических тестов мы рассчитали остаточную остроту зрения для каждого вида, используя регрессии остроты зрения по длине глаза PGLS для набора поведенческих данных. Ожидается, что остаточная острота будет в основном зависеть от различий в анатомии сетчатки, поскольку она представляет собой часть межвидовых вариаций остроты зрения, которая не может быть объяснена длиной глаза. Затем мы сравнили вариации остаточной остроты зрения между экологическими категориями. Мы также исследовали, как исходная острота зрения (без поправки на длину глаза или филогению) зависит от экологии.Следуя предсказаниям визуальной анатомии, мы проверили вариации остаточной и исходной остроты зрения с характером диэльной активности (дневная, катемерная, ночная) с использованием теста Крускала-Уоллиса с последующим односторонним апостериорным тестом Уилкоксона. Аналогичным образом мы сравнили остаточную и исходную остроту зрения в двух диетических группах (травоядные и активные хищники) с помощью одностороннего критерия Уилкоксона. Мы использовали тесты χ 2 , чтобы подтвердить, что доля видов в каждой модели диетической активности существенно не различалась между категориями рациона.Мы исследовали взаимосвязь между MRS и исходной остротой зрения, а также остаточной остротой зрения, используя ранговые корреляции Спирмена.
Затем мы сравнили вариации остаточной остроты зрения между экологическими категориями. Мы также исследовали, как исходная острота зрения (без поправки на длину глаза или филогению) зависит от экологии.Следуя предсказаниям визуальной анатомии, мы проверили вариации остаточной и исходной остроты зрения с характером диэльной активности (дневная, катемерная, ночная) с использованием теста Крускала-Уоллиса с последующим односторонним апостериорным тестом Уилкоксона. Аналогичным образом мы сравнили остаточную и исходную остроту зрения в двух диетических группах (травоядные и активные хищники) с помощью одностороннего критерия Уилкоксона. Мы использовали тесты χ 2 , чтобы подтвердить, что доля видов в каждой модели диетической активности существенно не различалась между категориями рациона.Мы исследовали взаимосвязь между MRS и исходной остротой зрения, а также остаточной остротой зрения, используя ранговые корреляции Спирмена.
Мы также рассчитали три отдельные многомерные модели PGLS с остротой зрения в качестве переменной ответа и с длиной глаза и одной из трех экологических переменных (модель диетической активности, диета или MRS) в качестве копредикторов. Для каждой из этих многомерных моделей PGLS с экологической переменной в качестве сопредиктора мы также рассчитали нулевую двумерную регрессию остроты зрения и длины глаза PGLS с использованием того же подмножества видов.Мы сравнили соответствие многомерных моделей и нулевых моделей, используя значения информационного критерия Акаике (AIC) и ANOVA. При ΔAIC <2 модели считались эквивалентными [Burnham et al., 2011; Саймондс и Муссалли, 2011]. Многомерные модели с ΔAIC от 3 до 7 считались, возможно, лучше, чем нулевая модель, в то время как многомерные модели с ΔAIC >10 считались значительно лучше, чем нулевая модель [Burnham et al., 2011; Саймондс и Муссалли, 2011]. Мы также сравнили нулевую и многомерную модели, используя последовательную сумму квадратов с анова.Функция pgls в каперсах.
Для каждой из этих многомерных моделей PGLS с экологической переменной в качестве сопредиктора мы также рассчитали нулевую двумерную регрессию остроты зрения и длины глаза PGLS с использованием того же подмножества видов.Мы сравнили соответствие многомерных моделей и нулевых моделей, используя значения информационного критерия Акаике (AIC) и ANOVA. При ΔAIC <2 модели считались эквивалентными [Burnham et al., 2011; Саймондс и Муссалли, 2011]. Многомерные модели с ΔAIC от 3 до 7 считались, возможно, лучше, чем нулевая модель, в то время как многомерные модели с ΔAIC >10 считались значительно лучше, чем нулевая модель [Burnham et al., 2011; Саймондс и Муссалли, 2011]. Мы также сравнили нулевую и многомерную модели, используя последовательную сумму квадратов с анова.Функция pgls в каперсах.
Результаты
Острота зрения и длина глаза
Наша сравнительная выборка документирует существенные различия в остроте зрения как внутри, так и между таксономическими группами млекопитающих (рис. 2). В целом, микрорукокрылые летучие мыши демонстрируют наименьшую остроту зрения в нашей выборке (медиана 0,6 имп/д, диапазон 0,1-1,9 имп/д), в то время как гаплориновые приматы имеют самую высокую остроту зрения (медиана 46 имп/д, диапазон 8,3-64,3 имп/д). Острота зрения для большинства (~91%) негаплориновых таксонов колебалась в пределах 0.1 и 10,4 циклов в день (± 1 стандартное отклонение). Мы также идентифицировали несколько выпадающих видов, у которых острота зрения значительно выше, чем у других видов в их таксономических группах (рис. 2), в том числе жираф ( Giraffa camelopardalis ; 25,5 имп/д), гепард ( Acinonyx jubatus ; 23 имп/д) и западный серый кенгуру ( Macropus fuliginosus ; 11,2 экземпляра в сутки).
2). В целом, микрорукокрылые летучие мыши демонстрируют наименьшую остроту зрения в нашей выборке (медиана 0,6 имп/д, диапазон 0,1-1,9 имп/д), в то время как гаплориновые приматы имеют самую высокую остроту зрения (медиана 46 имп/д, диапазон 8,3-64,3 имп/д). Острота зрения для большинства (~91%) негаплориновых таксонов колебалась в пределах 0.1 и 10,4 циклов в день (± 1 стандартное отклонение). Мы также идентифицировали несколько выпадающих видов, у которых острота зрения значительно выше, чем у других видов в их таксономических группах (рис. 2), в том числе жираф ( Giraffa camelopardalis ; 25,5 имп/д), гепард ( Acinonyx jubatus ; 23 имп/д) и западный серый кенгуру ( Macropus fuliginosus ; 11,2 экземпляра в сутки).
Рис. 2
Межпорядковая изменчивость остроты зрения млекопитающих.
PGLS-регрессия остроты поведения в зависимости от длины глаза у 42 видов выявляет значительную положительную связь между двумя переменными (рис.3а; Таблица 1). Таким образом, с увеличением размера глаз острота зрения также имеет тенденцию к увеличению. Согласно этой регрессии, около 35% вариаций остроты зрения между видами млекопитающих связаны с разницей в длине глаз (таблица 1). Семь видов с маленькими глазами расположены значительно ниже линии поведенческой регрессии (рис. 3а), что позволяет предположить, что острота зрения у них ниже, чем ожидалось для их длины глаза. Эти таксоны включают 4 микохирокрылых летучих мышей (Myotis mystacinus, M. daubentonii, Artibeus jamaicensis, Phyllostomus hastatus) и 3 ночных или роющих грызунов (Mesocricetus auratus, Mus musculus, Peromyscus maniculatus) .Дневные гаплоринные приматы также наносятся на кластер над линиями регрессии, что указывает на то, что у них высокая острота зрения для их длины глаза. Тем не менее, таксоны в этих группах с низкой остротой и высокой остротой не были идентифицированы как выбросы в модели [определяемой как наличие студенческого остатка ±3; Джонс и Первис, 1997]. Отдельная регрессия PGLS анатомической остроты зрения по длине глаза у 49 видов указывает на аналогичную значимую положительную связь между двумя переменными (таблица 1).
Согласно этой регрессии, около 35% вариаций остроты зрения между видами млекопитающих связаны с разницей в длине глаз (таблица 1). Семь видов с маленькими глазами расположены значительно ниже линии поведенческой регрессии (рис. 3а), что позволяет предположить, что острота зрения у них ниже, чем ожидалось для их длины глаза. Эти таксоны включают 4 микохирокрылых летучих мышей (Myotis mystacinus, M. daubentonii, Artibeus jamaicensis, Phyllostomus hastatus) и 3 ночных или роющих грызунов (Mesocricetus auratus, Mus musculus, Peromyscus maniculatus) .Дневные гаплоринные приматы также наносятся на кластер над линиями регрессии, что указывает на то, что у них высокая острота зрения для их длины глаза. Тем не менее, таксоны в этих группах с низкой остротой и высокой остротой не были идентифицированы как выбросы в модели [определяемой как наличие студенческого остатка ±3; Джонс и Первис, 1997]. Отдельная регрессия PGLS анатомической остроты зрения по длине глаза у 49 видов указывает на аналогичную значимую положительную связь между двумя переменными (таблица 1). Наклоны поведенческих и анатомически полученных линий регрессии имеют перекрывающиеся 95% доверительные интервалы (поведенческий 0.58-1,36; анатомический 0,85-1,32), предполагая, что существует постоянная связь между остротой зрения и длиной глаза независимо от того, как измеряется острота зрения (таблица 1). Кроме того, многомерная модель остроты зрения и длины глаза PGLS с типом измерения (поведенческий, анатомический) в качестве копредиктора показала, что тип измерения не является значимым фактором в модели (p = 0,072). Когда все данные об остроте зрения объединены, регрессия остроты зрения PGLS по длине глаза аналогична регрессиям, полученным с использованием поведенческих и анатомических подмножеств (таблица 1), и полностью попадает в их 95% доверительные интервалы.
Наклоны поведенческих и анатомически полученных линий регрессии имеют перекрывающиеся 95% доверительные интервалы (поведенческий 0.58-1,36; анатомический 0,85-1,32), предполагая, что существует постоянная связь между остротой зрения и длиной глаза независимо от того, как измеряется острота зрения (таблица 1). Кроме того, многомерная модель остроты зрения и длины глаза PGLS с типом измерения (поведенческий, анатомический) в качестве копредиктора показала, что тип измерения не является значимым фактором в модели (p = 0,072). Когда все данные об остроте зрения объединены, регрессия остроты зрения PGLS по длине глаза аналогична регрессиям, полученным с использованием поведенческих и анатомических подмножеств (таблица 1), и полностью попадает в их 95% доверительные интервалы.
Таблица 1
PGLS-регрессия остроты зрения и длины глаза
Рис. 3
Влияние длины глаза и филогенеза на остроту зрения у 91 млекопитающего. a PGLS-регрессии остроты зрения и длины глаза для поведенческих, анатомических и комбинированных наборов данных. b Квартильные диаграммы остаточной остроты зрения, рассчитанные для гаплориновых приматов, стрепсириновых приматов и млекопитающих, не являющихся приматами, с использованием комбинированной регрессии остроты зрения. Усы представляют самые высокие и самые низкие значения, которые не являются выбросами (закрашенные кружки; определены как> 1.5-кратный межквартильный размах). Гаплориновые выбросы: совиные обезьяны (Aotus azarae, A. trivirgatus) и долгопяты (Tarsius syrichta) . Млекопитающие, не относящиеся к приматам: фруктовая летучая мышь (A. jamaicensis) и большая копьеносая летучая мышь (P. hastatus) .
b Квартильные диаграммы остаточной остроты зрения, рассчитанные для гаплориновых приматов, стрепсириновых приматов и млекопитающих, не являющихся приматами, с использованием комбинированной регрессии остроты зрения. Усы представляют самые высокие и самые низкие значения, которые не являются выбросами (закрашенные кружки; определены как> 1.5-кратный межквартильный размах). Гаплориновые выбросы: совиные обезьяны (Aotus azarae, A. trivirgatus) и долгопяты (Tarsius syrichta) . Млекопитающие, не относящиеся к приматам: фруктовая летучая мышь (A. jamaicensis) и большая копьеносая летучая мышь (P. hastatus) .
Наши оценки лямбда Пагеля позволяют предположить, что существует сильное филогенетическое влияние на взаимосвязь между размером глаза и остротой зрения (таблица 1). Лямбда равна 1,00 в регрессии остроты поведения по длине глаза PGLS, что указывает на то, что распределение состояний признаков среди таксонов сильно коррелирует с выбранной нами филогенией. Однако лямбда значительно меньше 1 как для анатомических (λ = 0,75, p = 0,007), так и для комбинированных наборов данных (λ = 0,71, p <0,0001). Влияние филогении особенно очевидно для гаплориновых приматов. С учетом размера глаз остаточная острота зрения значительно различается между гаплориновыми приматами, стрепсириновыми приматами и млекопитающими, не являющимися приматами (Крускал-Уоллис: χ 2 (2) = 30,90, p <0,0001). В частности, гаплорины имеют значительно более высокую остаточную остроту зрения по сравнению с обоими млекопитающими, не являющимися приматами (Wilcoxon: W = 958, p < 0.0001) и стрепсиррин (Wilcoxon: W = 79, p = 0,0005; рис. 3б). Три выброса с наименьшими остаточными значениями для группы гаплоринов представляют единственные ночные таксоны гаплорхинов (Aotus и Tarsius) . Однако стрепсирины не имеют более высокой остаточной остроты зрения, чем другие млекопитающие, не являющиеся приматами (Wilcoxon: W = 270, p = 0,142). Из-за их производной остроты зрения по отношению к размеру глаза гаплорины были исключены из всех экологических анализов.
Однако лямбда значительно меньше 1 как для анатомических (λ = 0,75, p = 0,007), так и для комбинированных наборов данных (λ = 0,71, p <0,0001). Влияние филогении особенно очевидно для гаплориновых приматов. С учетом размера глаз остаточная острота зрения значительно различается между гаплориновыми приматами, стрепсириновыми приматами и млекопитающими, не являющимися приматами (Крускал-Уоллис: χ 2 (2) = 30,90, p <0,0001). В частности, гаплорины имеют значительно более высокую остаточную остроту зрения по сравнению с обоими млекопитающими, не являющимися приматами (Wilcoxon: W = 958, p < 0.0001) и стрепсиррин (Wilcoxon: W = 79, p = 0,0005; рис. 3б). Три выброса с наименьшими остаточными значениями для группы гаплоринов представляют единственные ночные таксоны гаплорхинов (Aotus и Tarsius) . Однако стрепсирины не имеют более высокой остаточной остроты зрения, чем другие млекопитающие, не являющиеся приматами (Wilcoxon: W = 270, p = 0,142). Из-за их производной остроты зрения по отношению к размеру глаза гаплорины были исключены из всех экологических анализов.
Влияние окружающей среды на изменение остроты зрения
Непараметрические тесты
Влияние экологических переменных на исходную остроту зрения показано на рисунке 4.Острота зрения значительно различается в зависимости от паттернов диальной активности (рис. 4а; Крускал-Уоллис: χ 2 (2) = 13,23, p = 0,001). И дневные, и катемерные виды проявляют более высокую остроту зрения, чем ночные виды (дневные и ночные по Уилкоксону: W = 112,5, p = 0,001; катемерные и ночные W = 127, p = 0,003). Однако острота зрения катемерных и дневных видов существенно не различается (Wilcoxon: W = 68, p = 0,086). Хотя мы обнаружили, что у активных хищников медиана остроты зрения выше, чем у травоядных (рис.2b), эти различия не были статистически значимыми (Wilcoxon: W = 40,5, p = 0,241). Наконец, мы обнаружили, что MRS значительно положительно коррелирует с остротой зрения (рис. 4c; Spearman: S = 56,6, r = 0,80, p = 0,002).
Рис. 4
Экологические факторы, потенциально влияющие на остроту зрения для набора данных по поведенческой остроте. и Модель активности Diel. Выбросы: ночные галаго (Galago senegalensis, Otolemur crassicaudatus) , катемерная лошадь (Equus caballus) и дневной верблюд (Camelus bactrianus) . б Сейм. Выброс: E. caballus (травоядное). c МРС. Все анализы исключают haplorhines, полуводные и роющие виды. Анализы питания исключают летучих мышей-микрокрылых. Блочные диаграммы являются квартильными, с усами, представляющими самые высокие и самые низкие значения, которые не являются выбросами (закрашенные кружки; определяется как более чем 1,5-кратный межквартильный диапазон).
и Модель активности Diel. Выбросы: ночные галаго (Galago senegalensis, Otolemur crassicaudatus) , катемерная лошадь (Equus caballus) и дневной верблюд (Camelus bactrianus) . б Сейм. Выброс: E. caballus (травоядное). c МРС. Все анализы исключают haplorhines, полуводные и роющие виды. Анализы питания исключают летучих мышей-микрокрылых. Блочные диаграммы являются квартильными, с усами, представляющими самые высокие и самые низкие значения, которые не являются выбросами (закрашенные кружки; определяется как более чем 1,5-кратный межквартильный диапазон).
Влияние экологии на остаточную остроту зрения (т. е. остроту зрения относительно длины глаза) показано на рисунке 5.Эти остаточные анализы показывают, что только паттерн диельной активности (рис. 5а) и диета (рис. 5б) оказывают значительное влияние на остроту зрения после того, как филогения и межвидовые вариации длины глаза статистически контролируются. Как и ожидалось, остаточная острота зрения зависит от характера активности (Краскал-Уоллис: χ 2 (2) = 7,02, p = 0,03), при этом дневные виды демонстрируют значительно более высокую остаточную остроту, чем ночные виды (Wilcoxon: W = 102, p = 0,006). ). Катемерные виды имеют значительно более высокую остаточную остроту зрения, чем ночные виды (Wilcoxon: W = 111, p = 0.033), но не отличаются от дневных таксонов (W = 59, p = 0,251). Остаточная острота зрения также зависит от рациона: у активных хищников остаточная острота зрения значительно выше, чем у травоядных (Wilcoxon: W = 55, p = 0,014). Наконец, в отличие от результатов для исходной остроты зрения, остаточная острота зрения не имеет значимой корреляции с MRS (рис. 5c; Spearman: S = 250, r = 0,13, p = 0,70).
). Катемерные виды имеют значительно более высокую остаточную остроту зрения, чем ночные виды (Wilcoxon: W = 111, p = 0.033), но не отличаются от дневных таксонов (W = 59, p = 0,251). Остаточная острота зрения также зависит от рациона: у активных хищников остаточная острота зрения значительно выше, чем у травоядных (Wilcoxon: W = 55, p = 0,014). Наконец, в отличие от результатов для исходной остроты зрения, остаточная острота зрения не имеет значимой корреляции с MRS (рис. 5c; Spearman: S = 250, r = 0,13, p = 0,70).
Рис. 5
Экологические факторы, потенциально влияющие на остаточную остроту зрения для набора данных поведенческой остроты зрения.Остаточная острота зрения рассчитана по поведенческой регрессии PGLS. и Модель активности Diel. б Сейм. Выброс: северный кволл ( Dasyurus hallucatus ; активный хищник). c МРС. Все анализы исключают haplorhines, полуводные и роющие виды. Анализы питания исключают летучих мышей-микрокрылых. Блочные диаграммы являются квартильными, с усами, представляющими самые высокие и самые низкие значения, которые не являются выбросами (закрашенные кружки; определяется как более чем 1,5-кратный межквартильный диапазон).
Блочные диаграммы являются квартильными, с усами, представляющими самые высокие и самые низкие значения, которые не являются выбросами (закрашенные кружки; определяется как более чем 1,5-кратный межквартильный диапазон).
Многомерные модели PGLS
Результаты всех многомерных моделей PGLS показаны в таблице 2. Многомерные модели PGLS, включающие в себя схему диетической активности и диету, точно отражают наши результаты в отношении остаточной остроты зрения. Эти многомерные модели демонстрируют, что как характер активности, так и диета оказывают значительное влияние на остроту зрения, которая не зависит от длины глаза (таблица 2). Включение модели активности в качестве сопредиктора приводит к модели, которая значительно лучше соответствует данным, чем нулевая модель, измеренная с помощью ANOVA (F 2 = 3.95, p = 0,03) и увеличивает объяснительную способность модели на ∼3,8%. Как и в случае остаточного анализа, дневные и катемерные виды имеют значительно более высокую остроту зрения по отношению к размеру глаза, чем ночные виды (таблица 2).
Таблица 2
Результаты многовариантных моделей питания показывают, что у активных хищников острота зрения относительно размера глаз значительно выше, чем у травоядных видов (таблица 2). Включение диеты в качестве сопредиктора значительно улучшает соответствие моделей данным (ANOVA: F 1 = 5.57, р = 0,033). Включение диеты в качестве сопредиктора увеличивает объяснительную силу многомерной модели на 5,19%.
В отличие от результатов для модели диетической активности и диеты, многофакторные модели не показывают значительного влияния MRS на остроту зрения, если принять во внимание влияние размера глаза (таблица 2). Кроме того, добавление MRS к нулевой модели не улучшает соответствие модели (ANOVA: F 1 = 1,325, p = 0,279), а только увеличивает объяснительную способность модели на 0.8%. Этот результат согласуется с нашим выводом о том, что MRS не оказывает существенного влияния на остаточную остроту зрения при непараметрическом анализе.
Обсуждение
Это исследование представляет собой первый филогенетически контролируемый анализ влияния размера глаза на остроту зрения в широкой сравнительной выборке млекопитающих. Наши результаты показывают, что острота зрения сильно положительно коррелирует с длиной глаза, и что только длина глаза может объяснить существенную долю межвидовой изменчивости остроты зрения (~35% на основе выборки поведенческой остроты, ~50% на основе комбинированной остроты зрения). образец).Другими словами, млекопитающие с абсолютно большими глазами, как правило, имеют более высокую остроту зрения, чем млекопитающие с абсолютно меньшими глазами. Этот результат согласуется с ожиданиями, основанными на оптических соображениях [Walls, 1942; Киршфельд, 1976; Хьюз, 1977; Pettigrew et al., 1988] и предыдущие результаты, основанные на меньших сравнительных выборках [например, Килти, 2000 г.; Хизи и Холл, 2010]. Вместе эти результаты показывают, что наличие абсолютно больших глаз облегчает более детальную выборку абсолютно большего изображения на сетчатке с помощью мозаики фоторецепторов.Хотя длина глаза является одной из переменных, которая часто используется для расчета анатомических оценок остроты зрения [например, Хьюз, 1977; Pettigrew et al.
Наши результаты показывают, что острота зрения сильно положительно коррелирует с длиной глаза, и что только длина глаза может объяснить существенную долю межвидовой изменчивости остроты зрения (~35% на основе выборки поведенческой остроты, ~50% на основе комбинированной остроты зрения). образец).Другими словами, млекопитающие с абсолютно большими глазами, как правило, имеют более высокую остроту зрения, чем млекопитающие с абсолютно меньшими глазами. Этот результат согласуется с ожиданиями, основанными на оптических соображениях [Walls, 1942; Киршфельд, 1976; Хьюз, 1977; Pettigrew et al., 1988] и предыдущие результаты, основанные на меньших сравнительных выборках [например, Килти, 2000 г.; Хизи и Холл, 2010]. Вместе эти результаты показывают, что наличие абсолютно больших глаз облегчает более детальную выборку абсолютно большего изображения на сетчатке с помощью мозаики фоторецепторов.Хотя длина глаза является одной из переменных, которая часто используется для расчета анатомических оценок остроты зрения [например, Хьюз, 1977; Pettigrew et al. , 1988], мы также обнаружили, что взаимосвязь между остротой зрения и длиной глаза существенно не меняется в зависимости от того, измеряется ли острота зрения анатомически или поведенчески [Kiltie, 2000].
, 1988], мы также обнаружили, что взаимосвязь между остротой зрения и длиной глаза существенно не меняется в зависимости от того, измеряется ли острота зрения анатомически или поведенчески [Kiltie, 2000].
Наши результаты подтверждают гипотезу о том, что одним из непосредственных факторов, влияющих на эволюцию размера глаз у млекопитающих, является отбор на повышенную остроту зрения.Например, предполагается, что большие глаза млекопитающих с высокой скоростью бега представляют собой приспособления для повышения остроты зрения, чтобы избежать столкновений с препятствиями в окружающей среде [Heard-Booth and Kirk, 2012]. Точно так же предполагается, что большие глаза некоторых таксонов млекопитающих (например, кошачьих плотоядных, долгопятов и совиных обезьян) являются результатом селекции, направленной на повышение остроты зрения без ущерба для зрительной чувствительности в ночное время [Ross, 2000; Кирк и Кей, 2004 г.; Росс и Кирк, 2007].Здесь мы подтверждаем ключевой компонент этих адаптивных сценариев, показывая, что большие глаза связаны с более высокой остротой зрения, предполагая, что эти два фактора часто развиваются в тандеме. Хотя наши данные не дают прямых доказательств того, что ослабленный отбор по остроте зрения может привести к уменьшению размера глаза, такое ожидание разумно, поскольку рост и поддержание глаза требует метаболических затрат [Schmidt et al., 2003; Нивен и Лафлин, 2008]. Действительно, генетические данные пещерных рыб показывают, что отбор (а не дрейф) может уменьшать размер глаз, когда зрение не требуется [Protas et al., 2007].
Хотя наши данные не дают прямых доказательств того, что ослабленный отбор по остроте зрения может привести к уменьшению размера глаза, такое ожидание разумно, поскольку рост и поддержание глаза требует метаболических затрат [Schmidt et al., 2003; Нивен и Лафлин, 2008]. Действительно, генетические данные пещерных рыб показывают, что отбор (а не дрейф) может уменьшать размер глаз, когда зрение не требуется [Protas et al., 2007].
Экология и острота зрения млекопитающих
Тот факт, что длина глаза объясняет лишь часть межвидовой изменчивости остроты зрения млекопитающих, неудивителен, учитывая большое количество других анатомических факторов, которые, как известно, влияют на остроту зрения [Walls, 1942; Хьюз, 1977; Арресе и др., 1999; Росс, 2000; Кирк и Кей, 2004]. Конфигурация диоптрийного аппарата влияет на остроту зрения, влияя на размер изображения на сетчатке, в то время как аномалии рефракции, площадь зрачка и плотность ганглиозных клеток сетчатки могут устанавливать верхний предел максимальной остроты зрения, которая может быть достигнута в любом глазу млекопитающего [Walls , 1942; Хьюз, 1977; Лэнд и Нильссон, 2012]. В дополнение к этим непосредственным анатомическим детерминантам остроты зрения, большинство основных эволюционных факторов, определяющих различия в остроте зрения между видами, являются экологическими [Walls, 1942; Хьюз, 1977; Лэнд и Нильссон, 2012]. В этом контексте наши результаты дают убедительные доказательства того, что характер диальной активности и диета являются важными селективными факторами, влияющими на межвидовые различия в остроте зрения млекопитающих.
В дополнение к этим непосредственным анатомическим детерминантам остроты зрения, большинство основных эволюционных факторов, определяющих различия в остроте зрения между видами, являются экологическими [Walls, 1942; Хьюз, 1977; Лэнд и Нильссон, 2012]. В этом контексте наши результаты дают убедительные доказательства того, что характер диальной активности и диета являются важными селективными факторами, влияющими на межвидовые различия в остроте зрения млекопитающих.
Модель активности Diel оказала значительное влияние на исходную остроту зрения в нашей сравнительной выборке, при этом дневные и катемерные виды имели более высокую исходную остроту, чем ночные виды.Этот результат отчасти ожидаем, поскольку млекопитающие, ведущие дневной образ жизни, отличаются от ночных млекопитающих морфологией глаза и анатомией сетчатки, обеспечивающей более высокую остроту зрения [Walls, 1942; Хьюз, 1977; Петтигрю и др., 1988; Кирк, 2006b; Хизи и Холл, 2010 г .; Холл и др., 2012; Лэнд и Нильссон, 2012]. Однако вопреки ожиданию, что катемерные виды должны обладать зрительными адаптациями, промежуточными между таковыми у дневных и ночных видов [Walls, 1942; Кей и Кирк, 2000 г .; Кирк, 2006b; Veilleux, Kirk, 2009], дневные и катемерные млекопитающие в нашей выборке достоверно не различались по сырой остроте.Этот результат, вероятно, связан с тем, что многие из самых крупных видов в нашей выборке являются катемерными (например, жираф, лошадь и носорог; онлайн-приложение, таблица 1). Размер глаз у млекопитающих сильно положительно коррелирует с размером тела [Ross and Kirk, 2007], и, таким образом, катемерные таксоны также имеют самые большие глаза в нашей выборке (онлайн-приложение, рис. 1a). Эти более крупные глаза катемерных видов, вероятно, компенсируют повышенную суммацию сетчатки и относительно более короткую PND по сравнению с дневной выборкой, что приводит к сопоставимой исходной остроте зрения в двух группах.
Однако вопреки ожиданию, что катемерные виды должны обладать зрительными адаптациями, промежуточными между таковыми у дневных и ночных видов [Walls, 1942; Кей и Кирк, 2000 г .; Кирк, 2006b; Veilleux, Kirk, 2009], дневные и катемерные млекопитающие в нашей выборке достоверно не различались по сырой остроте.Этот результат, вероятно, связан с тем, что многие из самых крупных видов в нашей выборке являются катемерными (например, жираф, лошадь и носорог; онлайн-приложение, таблица 1). Размер глаз у млекопитающих сильно положительно коррелирует с размером тела [Ross and Kirk, 2007], и, таким образом, катемерные таксоны также имеют самые большие глаза в нашей выборке (онлайн-приложение, рис. 1a). Эти более крупные глаза катемерных видов, вероятно, компенсируют повышенную суммацию сетчатки и относительно более короткую PND по сравнению с дневной выборкой, что приводит к сопоставимой исходной остроте зрения в двух группах.
После учета различий в длине глаз анализ остаточной остроты зрения и многомерные модели PGLS показали, что дневные и катемерные млекопитающие имеют самую высокую остроту зрения в нашей сравнительной выборке. Другими словами, при сравнении видов с одинаковым абсолютным размером глаз дневные и катемерные таксоны имеют значительно более высокую остроту зрения, чем ночные таксоны. Хотя дневные и катемерные виды существенно не различаются по остаточной остроте зрения, катемерные виды, как правило, имеют значения остаточной остроты зрения, промежуточные между дневными и ночными видами (рис.5а). Это изменение остроты зрения, не зависящее от наблюдаемого эффекта размера глаза, вероятно, достигается за счет адаптации диоптрийного аппарата (для увеличения размера изображения на сетчатке) и/или сетчатки (для увеличения плотности выборки) и подтверждается множеством сравнительных данных о млекопитающих. Морфология глаза и анатомия сетчатки. У млекопитающих, ведущих дневной образ жизни, часто наблюдаются меньшие относительные размеры роговицы и хрусталика, более низкое отношение палочек к колбочкам и более низкая суммация сетчатки, чем у ночных видов (все эти факторы имеют тенденцию к увеличению остроты зрения) [Walls, 1942; Хьюз, 1977; Арресе и др.
Другими словами, при сравнении видов с одинаковым абсолютным размером глаз дневные и катемерные таксоны имеют значительно более высокую остроту зрения, чем ночные таксоны. Хотя дневные и катемерные виды существенно не различаются по остаточной остроте зрения, катемерные виды, как правило, имеют значения остаточной остроты зрения, промежуточные между дневными и ночными видами (рис.5а). Это изменение остроты зрения, не зависящее от наблюдаемого эффекта размера глаза, вероятно, достигается за счет адаптации диоптрийного аппарата (для увеличения размера изображения на сетчатке) и/или сетчатки (для увеличения плотности выборки) и подтверждается множеством сравнительных данных о млекопитающих. Морфология глаза и анатомия сетчатки. У млекопитающих, ведущих дневной образ жизни, часто наблюдаются меньшие относительные размеры роговицы и хрусталика, более низкое отношение палочек к колбочкам и более низкая суммация сетчатки, чем у ночных видов (все эти факторы имеют тенденцию к увеличению остроты зрения) [Walls, 1942; Хьюз, 1977; Арресе и др. , 1999; Кей и Кирк, 2000 г .; Росс, 2000; Кирк, 2004 г.; Кирк и Кей, 2004 г.; Сильвейра, 2004 г.; Пайхл, 2005; Кирк, 2006b; Росс и Кирк, 2007 г .; Холл и др., 2012]. Кроме того, дневные гаплорины имеют значительно более высокую остроту зрения, чем другие млекопитающие, из-за узкоспециализированных специализаций сетчатки (например, полностью колбочковая ямка без суммирования сетчатки) и морфологии глаза (например, очень маленькая роговица по сравнению с размерами глаза) [Kay and Kirk, 2000; Росс, 2000; Кирк, 2004 г.; Кирк и Кей, 2004 г.; Кирк, 2006а, б; Росс и Кирк, 2007 г .; Холл и др., 2012].
, 1999; Кей и Кирк, 2000 г .; Росс, 2000; Кирк, 2004 г.; Кирк и Кей, 2004 г.; Сильвейра, 2004 г.; Пайхл, 2005; Кирк, 2006b; Росс и Кирк, 2007 г .; Холл и др., 2012]. Кроме того, дневные гаплорины имеют значительно более высокую остроту зрения, чем другие млекопитающие, из-за узкоспециализированных специализаций сетчатки (например, полностью колбочковая ямка без суммирования сетчатки) и морфологии глаза (например, очень маленькая роговица по сравнению с размерами глаза) [Kay and Kirk, 2000; Росс, 2000; Кирк, 2004 г.; Кирк и Кей, 2004 г.; Кирк, 2006а, б; Росс и Кирк, 2007 г .; Холл и др., 2012].
Мы также обнаружили, что диета оказала значительное влияние на остроту зрения после статистического контроля длины глаз. Как в остаточных анализах, так и в многомерных моделях PGLS активные хищники имели более высокую остроту зрения, чем травоядные. Эти результаты показывают, что хищные млекопитающие, которые частично полагаются на зрение, чтобы поймать движущуюся добычу, как правило, имеют сетчатку, поддерживающую более высокое визуальное разрешение, чем сетчатка травоядных млекопитающих. В то время как несколько исследований изучали влияние диеты на морфологию глаза млекопитающих и анатомию сетчатки, более фаунистические млекопитающие обычно демонстрируют более низкое соотношение палочек и колбочек, более высокую плотность ганглиозных клеток и более конвергентные орбиты (отражающие большее перекрытие бинокулярного поля), чем травоядные виды [Peichl et al. ., 2004; Тетро и др., 2004 г.; Хизи, 2008]. Каждый из этих факторов должен приводить к повышению остроты зрения у животных, если размер и морфология глаз остаются постоянными. Однако мы не обнаружили существенной разницы в исходной остроте зрения между двумя диетическими группами. Как и в случае с нашими результатами для катемерных таксонов, этот вывод не совсем удивителен, потому что наша группа травоядных включает много видов с крупным телом и большими глазами (онлайн-приложение, рис. 1b; онлайн-приложение, таблица 1) 1 .
В то время как несколько исследований изучали влияние диеты на морфологию глаза млекопитающих и анатомию сетчатки, более фаунистические млекопитающие обычно демонстрируют более низкое соотношение палочек и колбочек, более высокую плотность ганглиозных клеток и более конвергентные орбиты (отражающие большее перекрытие бинокулярного поля), чем травоядные виды [Peichl et al. ., 2004; Тетро и др., 2004 г.; Хизи, 2008]. Каждый из этих факторов должен приводить к повышению остроты зрения у животных, если размер и морфология глаз остаются постоянными. Однако мы не обнаружили существенной разницы в исходной остроте зрения между двумя диетическими группами. Как и в случае с нашими результатами для катемерных таксонов, этот вывод не совсем удивителен, потому что наша группа травоядных включает много видов с крупным телом и большими глазами (онлайн-приложение, рис. 1b; онлайн-приложение, таблица 1) 1 .
Наши необработанные результаты по остроте зрения подтверждают предположение, что виды с более быстрой MRS должны демонстрировать более высокую остроту зрения, чем виды с более медленной MRS (рис. 4c) [Уоллс, 1942; Хьюз, 1977; Херд-Бут и Кирк, 2012]. Это открытие можно было бы объяснить результатом отбора на повышенную остроту зрения у быстро бегающих млекопитающих, чтобы избежать столкновений с препятствиями окружающей среды [Walls, 1942; Хьюз, 1977; Херд-Бут и Кирк, 2012]. Однако наши результаты также показывают, что при постоянной длине глаза острота зрения не имеет существенной связи с MRS. Качество данных о максимальной скорости передвижения может значительно различаться в зависимости от методологии, используемой разными авторами, поэтому к интерпретации этих результатов следует относиться с осторожностью [Garland, 1983].Тем не менее, наши результаты показывают, что, хотя MRS может влиять на межвидовые различия в длине глаз у млекопитающих, это не связано с дополнительной адаптацией сетчатки или диоптрий для повышения остроты зрения. Если эта интерпретация верна и у быстро бегающих млекопитающих большие глаза, но отсутствует четкая адаптация сетчатки для повышения остроты зрения, то можно предположить, что наблюдаемая взаимосвязь между размером глаза и MRS у млекопитающих [Heard-Booth and Kirk, 2012] может быть результатом отбора, благоприятствующего факторам, отличным от повышенной максимальной остроты зрения.
4c) [Уоллс, 1942; Хьюз, 1977; Херд-Бут и Кирк, 2012]. Это открытие можно было бы объяснить результатом отбора на повышенную остроту зрения у быстро бегающих млекопитающих, чтобы избежать столкновений с препятствиями окружающей среды [Walls, 1942; Хьюз, 1977; Херд-Бут и Кирк, 2012]. Однако наши результаты также показывают, что при постоянной длине глаза острота зрения не имеет существенной связи с MRS. Качество данных о максимальной скорости передвижения может значительно различаться в зависимости от методологии, используемой разными авторами, поэтому к интерпретации этих результатов следует относиться с осторожностью [Garland, 1983].Тем не менее, наши результаты показывают, что, хотя MRS может влиять на межвидовые различия в длине глаз у млекопитающих, это не связано с дополнительной адаптацией сетчатки или диоптрий для повышения остроты зрения. Если эта интерпретация верна и у быстро бегающих млекопитающих большие глаза, но отсутствует четкая адаптация сетчатки для повышения остроты зрения, то можно предположить, что наблюдаемая взаимосвязь между размером глаза и MRS у млекопитающих [Heard-Booth and Kirk, 2012] может быть результатом отбора, благоприятствующего факторам, отличным от повышенной максимальной остроты зрения. Например, увеличение размера глаза может иметь важные последствия для цепей сетчатки, связанных с восприятием движения и оптического потока на периферии сетчатки. Как минимум, наши результаты предполагают, что современные интерпретации взаимосвязи между размером глаза и максимальной скоростью передвижения требуют дальнейшего уточнения на основе анализа дополнительных данных [см. также Hall and Heesy, 2011].
Например, увеличение размера глаза может иметь важные последствия для цепей сетчатки, связанных с восприятием движения и оптического потока на периферии сетчатки. Как минимум, наши результаты предполагают, что современные интерпретации взаимосвязи между размером глаза и максимальной скоростью передвижения требуют дальнейшего уточнения на основе анализа дополнительных данных [см. также Hall and Heesy, 2011].
Наконец, также стоит отметить, что анатомические данные свидетельствуют о том, что дополнительные экологические факторы, не рассмотренные здесь, могут дополнительно влиять на межвидовые различия в остроте зрения млекопитающих.Например, Veilleux and Lewis [2011] недавно обнаружили, что форма глаз млекопитающих предсказуемо менялась в зависимости от интенсивности окружающего света в разных типах среды обитания. В частности, млекопитающие из более открытых местообитаний имели меньшие относительные размеры роговицы по сравнению с видами из лесов с закрытым пологом. Таким образом, мы могли бы предсказать, что с учетом характера активности млекопитающие, обитающие в более открытой среде обитания, демонстрируют более высокую остроту зрения. Соответственно, не может быть случайным, что 3 млекопитающих, не являющихся приматами, с наивысшей остротой зрения в настоящем анализе (т.е. лошади, гепарды, жирафы) обычно обитают в более открытой среде [Nowak, 1999].
Соответственно, не может быть случайным, что 3 млекопитающих, не являющихся приматами, с наивысшей остротой зрения в настоящем анализе (т.е. лошади, гепарды, жирафы) обычно обитают в более открытой среде [Nowak, 1999].
Выводы
Межвидовые различия в размере глаз млекопитающих имеют предсказуемые последствия для остроты зрения. Длина глаза и острота зрения значительно положительно коррелируют у млекопитающих, и поэтому виды с абсолютно большими глазами, как правило, имеют более высокую остроту зрения, чем виды с абсолютно маленькими глазами. Некоторые ночные летучие мыши и грызуны имеют более низкую остроту зрения, чем ожидалось, для их размеров глаз, в то время как дневные человекообразные приматы имеют высокую остроту зрения для их размеров глаз.Когда различия между видами в абсолютном размере глаз статистически контролируются, у дневных видов острота зрения выше, чем у ночных, а у активных хищников острота зрения выше, чем у травоядных. Эти результаты свидетельствуют о том, что как дневные виды, так и активные хищники демонстрируют адаптацию диоптрий и/или сетчатки для повышения остроты зрения. Хотя виды с высоким MRS также, как правило, имеют абсолютно высокую остроту зрения по сравнению с видами с более низким MRS, нет никакой связи между MRS и остротой зрения, если влияние длины глаза остается постоянным.Это открытие предполагает, что у быстро бегающих млекопитающих может отсутствовать специфическая адаптация сетчатки для повышения остроты зрения. Если это так, современные функциональные объяснения наблюдаемой взаимосвязи между размером глаза и MRS у млекопитающих могут потребовать пересмотра.
Хотя виды с высоким MRS также, как правило, имеют абсолютно высокую остроту зрения по сравнению с видами с более низким MRS, нет никакой связи между MRS и остротой зрения, если влияние длины глаза остается постоянным.Это открытие предполагает, что у быстро бегающих млекопитающих может отсутствовать специфическая адаптация сетчатки для повышения остроты зрения. Если это так, современные функциональные объяснения наблюдаемой взаимосвязи между размером глаза и MRS у млекопитающих могут потребовать пересмотра.
Благодарности
Мы хотели бы поблагодарить Эндрю Барра, Эмбер Хёрд-Бут, Маргарет Холл, Аддисон Кемп, Чарли Нанна и Клару Скарри за полезные обсуждения. Мы также благодарим трех анонимных рецензентов за предоставление полезных комментариев для пересмотра этой статьи.
Авторское право: Все права защищены. Никакая часть данной публикации не может быть переведена на другие языки, воспроизведена или использована в любой форме и любыми средствами, электронными или механическими, включая фотокопирование, запись, микрокопирование или любую систему хранения и поиска информации, без письменного разрешения издателя. .
.
Дозировка препарата: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор препарата и дозировка, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации.Тем не менее, в связи с продолжающимися исследованиями, изменениями в правительственных постановлениях и постоянным потоком информации, касающейся лекарственной терапии и реакций на лекарства, читателю настоятельно рекомендуется проверять вкладыш в упаковке для каждого лекарства на предмет любых изменений в показаниях и дозировке, а также для дополнительных предупреждений. и меры предосторожности. Это особенно важно, когда рекомендуемый агент является новым и/или редко используемым лекарственным средством.
Отказ от ответственности: заявления, мнения и данные, содержащиеся в этой публикации, принадлежат исключительно отдельным авторам и участникам, а не издателям и редакторам.Появление рекламы и/или ссылок на продукты в публикации не является гарантией, одобрением или одобрением рекламируемых продуктов или услуг или их эффективности, качества или безопасности. Издатель и редактор(ы) отказываются от ответственности за любой ущерб, нанесенный людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в содержании или рекламе.
Издатель и редактор(ы) отказываются от ответственности за любой ущерб, нанесенный людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в содержании или рекламе.
Мелкие млекопитающие забираются выше, спасаясь от потепления в Скалистых горах | CU Boulder Today
Изображение баннера: Златошейный суслик набивает щеки едой.(Фото: CC photo Zachary S.L. Foster через Wikimedia Commons)
Суслик с золотым плащом ( Callospermophilus lateralis ) — популярное зрелище среди туристов в Скалистых горах — маленький грызун — фотогеничное существо с полосатой спиной и пухлыми щеками, в которых хранятся семена и другая еда.
Но есть реальность, которую фотографии в Instagram не передают, говорит Кристи Маккейн, эколог из Университета Колорадо в Боулдере. В новом исследовании, продолжавшемся почти 13 лет, она и ее коллеги обнаружили, что суслик присоединился ко многим другим мелким млекопитающим в Скалистых горах Колорадо, которые совершают зловещий путь: они взбираются в гору, чтобы избежать повышения температуры в штате, вызванного изменение климата.
«Это пугает, — сказал Маккейн, доцент кафедры экологии и эволюционной биологии. «Мы давно говорим об изменении климата в Скалистых горах, но я думаю, мы можем сказать, что это признак того, что сейчас все реагирует и реагирует довольно резко».
Златошейный суслик, которого часто путают с бурундуком, обитает в хвойных лесах в Скалистых горах и некоторых других западных горных хребтах.
«Вероятно, это одно из самых фотографируемых млекопитающих в Национальном парке Роки-Маунтин, поскольку оно позирует и прихорашивается на камнях у обочины дороги и в кемпингах», — сказал Маккейн, также куратор отдела позвоночных в Музее естественной истории CU.«Зимой они впадают в спячку, летом остаются территориальными и издают характерные сигналы тревоги, чтобы уведомить друг друга о ближайших опасностях».
Ее последнее исследование, опубликованное на этой неделе в журнале Ecology , глубоко погружается в судьбу белки и 46 других видов мелких млекопитающих из Переднего хребта и Скалистых гор Сан-Хуана — от мышей до землероек и даже желтых пузатый сурок ( Marmota flaviventris ).
Группа сообщает, что с 1980-х годов ареал обитания этих тварей в среднем сместился более чем на 400 футов вверх по высоте.Горные млекопитающие или те, которые уже живут на больших высотах, такие как суслики, приняли на себя самый большой удар, поднявшись в среднем на 1100 футов. Это значительное изменение, которое, если оно продолжится, может привести к полному вытеснению многих из этих животных из Колорадо.
И хотя многие из этих маленьких существ никогда не появятся на открытках Колорадо, Маккейн сказал, что они могут быть инициаторами более крупных и все более срочных изменений в Скалистых горах.
Страсть к горам
Для Маккейна этот проект во многом стал кульминацией ее любви к горам на протяжении всей ее жизни.
Это началось в середине 1990-х, когда она служила в Корпусе мира в Гондурасе. Там Маккейн была поражена тем, насколько горы тропической страны напоминали слоеные пироги: по мере того, как она поднималась вверх, горные экосистемы вокруг нее трансформировались, иногда резко.
«Везде, куда бы вы ни пошли, были совершенно разные птицы», — сказала она. «Изменения настолько резкие».
Но эта природа слоеного пирога может также быть причиной гибели горных экосистем перед лицом изменения климата.Колорадо потеплело почти на 3,5 градуса по Фаренгейту с 1980-х годов из-за антропогенного изменения климата. По мере того, как штат нагревается, многие ученые предсказывают, что сосновые леса пондероза и другие горные экосистемы должны будут двигаться выше, чтобы найти более прохладную погоду. Некоторые животные и даже целые сообщества могут быть вытеснены на вершины гор, где им больше некуда идти.
Маккейн и ее коллеги хотели выяснить, происходит ли уже этот сдвиг вверх в Скалистых горах.
Начиная с 2008 года, ее команда посетила несколько участков в Переднем хребте Колорадо и в горах Сан-Хуан, чтобы собрать записи о нынешних ареалах 47 видов грызунов и землероек.Среди них были редкие животные, такие как карликовая землеройка ( Sorex hoyi ), которая весит менее четверти и считается самым маленьким млекопитающим Северной Америки. Находящаяся под угрозой исчезновения прыгающая мышь Пребла ( Zapus hudsonius preblei ), известная своим длинным хвостом и большими ногами, которые полезны для плавания, также попала в список. В некоторых случаях, по словам Маккейна, места обитания были настолько удаленными, что исследователи путешествовали на конных поездах для перевозки своего оборудования.
«Нам пришлось привыкать к действительно неприятным условиям, — сказал Маккейн.«Мы миримся с сыростью в горах и жарой в пустыне, внезапными наводнениями и грозами».
Затем группа сравнила результаты своих опросов с примерно 4500 историческими записями из музейных коллекций в горах Фронт-Рейндж и Сан-Хуан, датируемых 1880-ми годами. Среди них были животные, хранящиеся в собственном музее CU Boulder, в котором хранится около 12 000 образцов млекопитающих из Колорадо.
Резкое действие
Результаты показали сложную реакцию на изменение климата и другие нагрузки.По словам Маккейна, некоторые из животных, участвовавших в исследовании, двигались вниз по высоте, а не вверх, в то время как у других с 1980-х годов их ареалы действительно увеличились.
Но большинство исследованных млекопитающих сдвинулись в гору — и не на малую величину. Особенно это касалось тех, кто уже жил на больших высотах. До 1980 года карликовую землеройку, например, никогда не обнаруживали на высоте более 9800 футов. Сегодня его максимальная протяженность составляет более 11 800 футов. Суслик с золотым плащом также поднялся на 650 футов, или 200 метров, в Переднем хребте и на 2300 футов, или 700 метров, в Сан-Хуане.
«Я ожидал, что мы увидим что-то на расстоянии от 100 до 200 метров, но мы увидели намного больше», — сказал Маккейн. «Это намного больше, чем изменения, которые были зафиксированы в других горных регионах по всему миру».
Исследование, по словам Маккейна, рисует суровую картину горного хребта, переживающего кризис. Но у этого есть и положительная сторона: еще может быть время, чтобы защитить знаковые западные сосновые леса пондерозы, альпийские луга и тощую тундру, но только если колорадцы и люди во всем мире начнут действовать сейчас, чтобы сократить выбросы парниковых газов.
— Это тревожный звонок, — сказал Маккейн. «Мы должны немедленно начать серьезно относиться к этому, если мы хотим иметь здоровые горы и экосистемы».
Среди других соавторов нового исследования Тим Шевчик, бывший аспирант Калифорнийского университета в Боулдере, ныне работающий в Лозаннском университете, и Сара Кинг из Университета штата Колорадо.
границ | Вирусы H9N2, выделенные от млекопитающих, реплицируются в мышах на более высоких уровнях, чем вирусы птичьего происхождения
Введение
H9N2 считался вирусом птичьего гриппа (AIV) с низкой патогенностью, поскольку он вызывает легкое клиническое течение заболевания (Cauthen et al., 2000). Домашняя птица, инфицированная ВГП H9N2, является важным инкубатором для новых появляющихся пандемических ВГП (Liu et al., 2014; Sun and Liu, 2015), которые способствуют рекомбинации новых вирусов гриппа H7N9 или h20N8 человека (Pu et al., 2015). В материковом Китае вирус H9N2 был впервые выделен у кур в провинции Гуандун в 1994 г. (Zhao et al., 2015), а затем 5 лет спустя у людей в Гонконге (Guo et al., 1999). Вирусы H9N2 повторно объединяются с множеством других подтипов с образованием новых генотипов вируса гриппа.Кроме того, сообщалось, что вирусы H9N2 обладают эволюционной идентичностью рецепторов, подобных человеческим, с заменой Q226L в гликопротеине гемагглютинина (HA) и адаптировались к связыванию с рецепторами хозяина млекопитающих, что привело к увеличению восприимчивости человека (Matrosovich et al., 2001). . Таким образом, предполагалось, что этот подтип представляет значительную угрозу для здоровья населения (Lin et al., 2000; Saito et al., 2001; Butt et al., 2005).
вируса гриппа H9N2 эволюционировали путем рекомбинации и мутации в течение 25 лет распространенности среди домашней птицы.Наиболее «приспособленный» генотип, G57, изменил антигенность, что привело к ускользанию вируса от иммунизации и вызвало широкомасштабные вспышки в 2010–2015 гг., и, наконец, стал единственным преобладающим генотипом у кур, выращиваемых на китайских фермах. Кроме того, генотип G57 подтипа H9N2 AIV внес свои шесть внутренних сегментов в новые вирусы H7N9 путем рекомбинации в 2013 г. Широкое использование вакцин против вирусов H9N2 у кур оказывает постоянное иммунное давление на гликопротеин гемагглютинина (HA) с антигенным дрейфом ( Пу и др., 2015).
На сегодняшний день имеется мало информации о реакции хозяина и выработке цитокинов в ответ на инфекцию вирусом H9N2. В этой статье патогенность вирусов H9N2 была исследована на мышах и цыплятах для определения иммунологических паттернов с различными генотипами.
Материалы и методы
Заявление об этике использования животных
Все исследования на животных проводились в строгом соответствии с рекомендациями руководства Шанхайского института ветеринарных исследований, CAAS (ID: SHVRI-PO-2014-0098) и одобрены Комиссией Научно-технической ассоциации животных Шанхайского муниципалитета, Китай. (Номер разрешения: 2013-11).
Культура клеток и реагенты
Клетки эмбриональной почки человека 293T (HEK 293T) и клетки почки собаки Madin Darby (MDCK) содержали в среде Игла, модифицированной Дульбекко (DMEM; HyClone, США), содержащей 5% эмбриональной бычьей сыворотки (FBS; PAN Biotech, Германия) и 1% раствор пенициллина-стрептомицина (PS; Thermo Fisher Scientific., США) при 37°C с 5% CO 2 .
Вирусы
Пять штаммов вируса гриппа А H9N2 были выделены от разных хозяев.Штамм Sw/08 был выделен на свиноферме Цзянсу в 2008 г. с легким респираторным заболеванием и назван A/swine/Jiangsu/Sw/08/08 (аббр. Sw/08). Штамм Ck/15 был выделен на птицеферме Аньхой в 2015 г. и назван A/chicken/Anhui/120/2015 (аббр. Ck/15). Штамм Gs/14 был выделен на гусиной ферме Цзянсу в 2014 г. и назван A/goose/Jiangsu/4539/2014 (аббр. Gs/14). Штамм Hu/04 был выделен от человека в 2004 г. и назван A/Guangdong/Hu/04/2004 сокр. Ху/04). Штамм Mi/14 был выделен из норки в 2014 году и назван A/mink/China/01/2014 (сокр.Ми/14). Вирусы H9N2 выращивали путем инокуляции в 9-дневных куриных яйцах, свободных от специфических патогенов (Merialvital, Co., Пекин). Аллантоисные жидкости собирали через 48 ч и хранили при температуре -80°C до использования (Jin et al., 2018). Титры вируса определяли и рассчитывали по методу Рида-Мюнча (Reed and Muench, 1938; Brown, 1964) для 50% инфекционной дозы тканевой культуры (TCID 50 ) на клетках MDCK с МЕМ (Hyclone Laboratories, San Angelo , Техас, США) с 1 мкг/мл трипсина, обработанного L -(тозиламидо-2-фенил)этилхлорметилкетоном (TPCK) (Chen et al., 2014а).
Секвенирование и филогенетический анализ вирусов H9N2
Тотальную РНК экстрагировали с использованием набора RNeasy (Qiagen, Inc., Gaithersburg, MD, США) в соответствии с инструкциями производителя. Обратную транскрипцию проводили с использованием праймера Uni12 (5′-AGCAAAAGCAAGG-3′) и обратной транскриптазы вируса миелобластоза птиц (AMV) (Takara) в соответствии с инструкциями производителя, а продукты кДНК хранили при -80°C до использования. Восемь сегментов вируса гриппа А амплифицировали методом ПЦР с использованием универсальных праймеров (Wang et al., 2017) и высокоточной ДНК-полимеразы Phusion (Vazyme, Inc., Нанкин, Китай). Продукты ПЦР клонировали в вектор pHW2000 с использованием подхода рекомбинации in vitro (Wang et al., 2017). Клонированные плазмиды подтверждали секвенированием по методу Сэнгера (Genewiz, Inc., Сучжоу, Китай) с праймерами: 5′-CGCAAATGGGCGGTAGGCGTG-3′ (CMV-Forward) и 5′-TAGAAGGCACAGTCGAGG-3′ (BGH-Reverse). Восемь сегментов вируса H9N2 были проанализированы и сопоставлены с помощью программного обеспечения MEGA6 (Tamura et al., 2013).
Патогенность вирусов H9N2 у кур
Для определения патогенности вирусов H9N2 у цыплят восьми цыплятам SPF (4-недельного возраста) на группу интраназально инокулировали в равной дозе 1 × 10 6 TCID 50 на 100 мкл, разведенных в буфере PBS. Куры отрицательного контроля получали 100 мкл PBS. После заражения цыплят SPF ежедневно контролировали в течение 14 дней. Мазки из ротоглотки и клоаки собирали через 1, 3, 5 дней после инокуляции (dpi), а затем хранили при -80°C в 1 мл буфера PBS, содержащего 100 ЕД/мл пенициллина и 0.05 мг/мл стрептомицина. Три цыпленка из каждой контрольной группы подвергали случайной эвтаназии через 3 dpi, образцы трахеи собирали и фиксировали в 10% формалине, а затем заливали в парафин для окрашивания гематоксилином и эозином (H&E). Тем временем образцы легких собирали и гомогенизировали с помощью Tissue Lyser со скоростью 70 раз 1/с в течение 2 мин (Chen et al., 2014a) и центрифугировали при 13, 200 × g в течение 10 мин, аликвоты по 100 мкл собирали супернатанты и определяли титры на клетках MDCK.Патогенные баллы измерялись на основе распределения поражений трахеи (Gibson-Corley et al., 2013). Остальных цыплят ежедневно контролировали на наличие клинических признаков, а затем подвергали эвтаназии через 14 точек на дюйм.
Инфицированные ткани легких гомогенизировали и выделяли тотальную РНК (Sun et al., 2017). Синтез кДНК осуществляли транскриптазой AMV (Takara) с Oligo(dT) 18 в объеме 20 мкл при 30°С в течение 10 мин, 42°С в течение 2 ч и затем 70°С в течение 15 мин. Праймеры для qRT-PCR были разработаны с использованием PrimerQuest Tool.Последовательности праймеров были следующими: qckβ-актин-F: TCCCTGGAGAAGAGCTATGAA; qckβ-актин-R: CAGGACTCCATACCCAAGA AAG. qckIL-1β-F: CTCACAGTCCTTCGACATCTTC; qckIL-1β-R: TGTTGAGCC TCACTTTCTGG. qckIL-6-F: TCTGTTCGCCTTTCAGACCTA; qckIL-6-R: GACCACCCTCATCGGGATTTAT. qckTNF-α-F: CCGCCCAGTTCAGATGAGTT; qckTNF-α-R: GCAACAACCAGCTATGCACC. qckMCP-1α-F: CCCACAACTGCC ATAGATGAA; qckMCP-1α-R: GCTTTCCCGAGGGCAATAA; qckMIP-1-F: GGGCTCTGTTTCTCTCTCTCTC; qckMIP-1-R: CATTTGCTGCTGGTGATGTAG. Реакции проводили с помощью qRT-PCR анализа с AceQ ® qPCR SYBR ® Green Master Mix (Vazyme) по следующему протоколу циклов: 95°C в течение 5 мин, 40 циклов при 95°C в течение 10 с и 60°С в течение 30 с или с последующей стадией кривой плавления (95°С в течение 15 с, 60°С в течение 1 мин и 95°С в течение 15 с).Результаты реакции были представлены значениями порогового цикла (Ct). Изменение кратности рассчитывали с использованием метода 2 -ΔΔCt (Huang et al., 2017). Средние значения Ct определяли на основе трех повторностей.
Проточная цитометрия
Мононуклеарные лейкоциты периферической крови цыплят (PBML) выделяли из вен крыльев цыплят с использованием Ficoll-paque (Amersham Pharmacia Biotech, Piscataway, NJ, United States) с градиентом плотности 50×g при 4°C в течение 30 мин и промывали три раза с PBS, содержащим 1% FBS.Клетки PBML инкубировали с FITC-меченым антителом против куриного CD3 (1:50) (Southern Biotech, Кембридж, Соединенное Королевство), PE-меченым антителом против куриного CD4 (1:50) (Southern Biotech) и APC-меченым -антитела против куриного CD8 (1:50) (Southern Biotech) в течение 20 мин при 4°C, промывали PBA (PBS, 0,5% BSA и 0,0001% азида натрия) и затем инкубировали с козьим антикуриным IgG (Southern Biotech). Biotech) в течение 20 мин при 4°C. Окрашенные клетки промывали буфером PBS и фиксировали 2% параформальдегидом (Merck, Inc., Германия) в течение 10 мин при комнатной температуре. Клетки однократно промывали буфером PBA. Не менее 10 000 событий были получены с использованием проточного цитометра FACS Calibur (BD Biosciences, Сан-Хосе, Калифорния, США), а данные были проанализированы с использованием FlowJO (Three Star, Inc., Ашленд, штат Орегон, США).
Патогенность вирусов H9N2 у мышей
Для определения вирулентности вирусов H9N2 in vivo восемь 5-недельных самок мышей BALB/c (Vital-River, Co., Пекин) инфицировали интраназально.Мыши BALB/c были анестезированы сухим льдом перед интраназальной инокуляцией (Jones et al., 1999; Kohler et al., 1999), каждая из которых получила 1 × 10 6 TCID 50 на 50 мкл интраназально (13 мышей в группе). ), с 50 мкл PBS, введенными мышам отрицательного контроля. Для определения титров вируса в легких по три мыши на группу умерщвляли через 3 dpi (Chen et al., 2014a). Образцы легких собирали и гомогенизировали с помощью Tissue Lyser со скоростью 70 раз 1/с в течение 2 мин (Chen et al., 2014a) и центрифугировали при 13, 200 × g в течение 10 мин. собрал.Массу тела животных контролировали ежедневно до 14 дней после инокуляции, и животных с потерей массы тела более чем на 25% подвергали эвтаназии. Титры вируса титровали на клетках MDCK. Уровни IL-1β, IL-6, MIP-1α, MCP-1 и TNF-α в аликвотах по 100 мкл гомогенизированных образцов легких измеряли с использованием наборов для твердофазного иммуноферментного анализа (sELISA) (eBioscience, Inc. ., Сан-Диего, Калифорния, США) в соответствии с рекомендациями производителя.
Статистический анализ
Статистический анализ проводили с использованием программного обеспечения Prism 6 (GraphPad, La Jolla, CA, США).Парные сравнения между средними группами были проведены с помощью двухэтапной повышающей процедуры с помощью теста Бенджамини, тогда как множественные сравнения были проведены с помощью дисперсионного анализа (метод однофакторного дисперсионного анализа). Различия считались достоверными при p -значениях < 0,05 или < 0,01.
Результаты
Филогенетический анализ генотипов IAV подтипа H9N2
Для дальнейшей характеристики различных генотипов вирусов гриппа А подтипа H9N2 восемь сегментов каждого выбранного вируса были амплифицированы, субклонированы и секвенированы.Последовательности депонированы в GenBank вместе с образцами изолята KK (KX867822–KX867829), изолята Ck/15 (KX867830–KX867837), изолята Gs/14 (KX867838–KX867845) и изолята Hu/04 (KX867846–KX867853). ). Характеристика вируса Mi/14 (MF996796–MF996803) была подтверждена в недавнем отчете (Xue et al., 2018).
Для отслеживания вирусов H9N2 генетическое происхождение подтипа было охарактеризовано на основе филогенетического анализа восьми сегментов (рис. 1), который был построен с использованием метода максимального правдоподобия в MEGA6.0 (Tamura et al., 2013) с 1000 бутстреп-реплицированием. Эталонные последовательности, используемые для генетического сравнения, были получены из GenBank. На основании филогенетического анализа три вируса (Gs/14, Ck/15 и Mi/14) принадлежали к генотипу B69, который является преобладающим генотипом, циркулирующим в Китае и демонстрировал более эффективную репликацию у кур, изолят Sw/08 принадлежал к генотипу B35 и изолят Hu/04 попали в генотип G9 (рис. 2) (Dong et al., 2011; Pu et al., 2015; Teng et al., 2016).
Рис. 1. Филогенетический анализ вирусов гриппа H9N2. Филогенетический анализ всех восьми сегментов был построен с использованием метода максимального правдоподобия с помощью MEGA6.0 с 1000 повторами начальной загрузки. (A–H) Филогенетические деревья для генов PB2, PB1, PA, HA, NP, NA, M и NS, с анализом на основе сегмента соответствующего гена, указанного в нуклеотидах. Эталонные последовательности, используемые для генетического сравнения, были получены из GenBank. АХ, Аньхой; ск, курица; Дк, Утка; Гусь, Гс; Человек, Ху; FJ, Фуцзянь; GD, Гуандун; HN, Хунань; Гонконг, Гонконг; JS, Цзянсу; Ка, перепел; Ш, Шанхай; СТ, Шаньтоу; Св, Свинья; ZJ, Чжэцзян; 03, 2003; 04, 2004; 06, 2006; 09, 2009; 11, 2011; 12, 2012; 13, 2013; 14, 2014; 15, 2015.
Рисунок 2. Генотипы вирусов гриппа H9N2. В овалах горизонтальные полосы сверху вниз обозначают PB2, PB1, PA, HA, NP, NA, M и NS. A/Quail/Hong Kong/G1/1997 (Qa/HK/G1/97) был выделен из перепелов в 1997 году в Гонконге, идентификатор таксона NCBI был 97352 и представлял генотип G1. A/Chicken/Beijing/1/1994 (Ck/BJ/94) был выделен из курицы в 1994 году в Пекине, идентификатор таксона NCBI был 1342391 и представлял генотип BJ94. A/Chicken/Hong Kong/G9/1997 (Ck/HK/G9/97) был выделен из курицы в 1997 году в Гонконге, идентификатор таксона NCBI был 1342394 и представлял генотип G9.A/Chicken/Korea/38349-p96323/1996 (Ck/KR/96323/96) был выделен из курицы в 1996 году в Корее, идентификатор таксона NCBI был 97358 и представлял генотип C0. A/Chicken/Shandong/01/2009 (Ck/SD/01/09) был выделен из курицы в 2009 году в Шаньдуне, идентификатор таксона NCBI был 1005638 и представлял генотип B65. A/Chicken/Shanghai/F/1998 (Ck/SH/F/98) был выделен из курицы в 1998 году в Шанхае, идентификатор таксона NCBI был 183741 и представлял генотип F/98. A/Chicken/Zhejiang/B2013/2012 (Ck/ZJ/B2013/12) был выделен из курицы в 2013 году в провинции Чжэцзян, идентификатор таксона NCBI был 1621010 и представлял генотип B69.A/Duck/Hong Kong/289/1978 (Dk/HK/289/78) был выделен из утки в 1978 году в Гонконге, идентификатор таксона NCBI был 210667 и представлял генотип E0. A/Duck/Hong Kong/Y439/1997 (Dk/HK/Y439/97) был выделен из утки в 1997 году в Гонконге, идентификатор таксона NCBI был 1342412 и представлял генотип Y439. A/Duck/Germany/113/1995 (Dk/GE/113/95) был выделен из утки в 1978 г. в Германии, идентификатор таксона NCBI был 1175984 и представлял генотип D1. A/Duck/ Fujian/ C2246 (DK/FJ/C2246/09) был выделен из утки в 2009 г. в провинции Фуцзянь, идентификатор таксона NCBI был 1521103 и представлял генотип B67.A/Duck/Shantou/163/2004 (Dk/ST/163/04) был выделен из утки в 2004 г. в Шаньтоу, идентификатор таксона NCBI был 456814 и представлял генотип ST/163. A/Chicken/Hong Kong/715.5/2001 (H5N1) (H5N1/01) был выделен из курицы в 2001 г. в Гонконге, идентификатор таксона NCBI был 196434 (Bi et al., 2011; Dong et al., 2011; Teng и др., 2016).
Анализ мутацийаминокислот показал, что все вирусы имели 226G и 228R в белке HA в области связывания рецептора, и у этих пяти вирусов H9N2 не наблюдалось делеции в областях стебля NA.Однако Gs/14, Ck/15 и Mi/14 имеют сходные аминокислотные последовательности по сравнению с Sw/08 и Hu/04 (таблица 1). Детально ПБ2 (598В, 676М), ПА (356Р), НП (377Н, 406В), НА (80Н, 125С, 184Я, 199Т, 356Д), М1 (37А, 95К, 140А, 142Г, 224Н, 242Н, 247Л ), M2 (16E, 21G, 31N, 68M, 82N), NS1 (47S, 80S, 103L) и NEP (14T, 21R, 49A) были показаны в таблице 1. Кроме того, мутации, связанные с адаптацией млекопитающих, наблюдались в Ген PB2 Mi/14 (A588V и D701N) и Sw/08 (D701N) (Sediri et al., 2015; Xiao et al., 2016) вирусы. R340K наблюдался в PB2 вирусов Mi/14 и Sw/08, 522V наблюдался в PB1 вируса Mi/14 (Neumann et al., 2014), который является потенциально связанным с вирулентностью сайтом. PA K356R, который увеличивает репликацию и патогенность, наблюдался у Gs/14, Ck/15 и Mi/14 (Xu et al., 2016).
Таблица 1. Аминокислотные мутации в пяти охарактеризованных вирусах H9N2.
Патогенность вирусов H9N2 у кур
Все пять вирусов H9N2 хорошо реплицировались в образцах ротоглотки при 1 и 3 dpi; однако очень низкие уровни вируса были обнаружены в ротоглоточных образцах вирусов Sw/08, Gs/14 и Ck/15 при 5 dpi.Вирусы Hu/04, Gs/14 и Mi/14 выделялись с фекалиями (рис. 3А–Е). Между тем, вирусные титры образцов легких при 3 dpi также гомогенизировали и титровали на клетках MDCK, все пять вирусов H9N2, кроме Hu/04, хорошо реплицировались в легких. По сравнению с Sw/08 и Gs/14, Ck/15 и Mi/14 демонстрируют более высокую способность к репликации в куриных легких (рис. 3F).
Рисунок 3. Репликация вирусов H9N2 у кур. Образцы ротоглотки и клоаки штаммов Sw/08 (A) , Hu/04 (B) , Gs/14 (C) и Ck/15 (D) и Mi/14 (E) вирусов были титрованы с помощью TCID 50 .Вирусная нагрузка в образцах легких при инфицировании на 3 dpi (F) . гр, грамм.
Для определения провоспалительной реакции пяти вирусов образцы легких 3 dpi гомогенизировали для количественного определения уровней мРНК IL-1β, IL-6, TNF-α, MIP-1 и MCP-1α (рис. 4A). . В частности, уровень мРНК IL-1β Ck/15 и Mi/14 увеличился более чем в 400 раз по сравнению с отрицательным контролем PBS, а также был значительно выше, чем у трех других штаммов H9N2 ( p <0,01).Уровни мРНК IL-6 ( p < 0,05), TNF-α ( p < 0,05), MIP-1 ( p < 0,05) и MCP-1α ( p < 0,01) также увеличились. минимум в 10 раз. Все вирусы H9N2 вызывали тяжелые поражения трахеи с увеличением бокаловидных клеток и экссудативным воспалением со средним баллом патогенности 4 (рис. 4В). Среднее количество CD 8 + Т-клеток увеличилось до 0,70% с 0,13%, когда цыплят заражали Sw/08 или Mi/14 (рис. 4C) (таблица 2).
Рисунок 4. Провоспалительные реакции и патогенность вирусов H9N2 у кур. (A) В гомогенизированных образцах легких, инфицированных пятью вирусами H9N2, определяли уровни мРНК IL-1β, IL-6, TNF-α, MIP-1 и MCP-1α. (B) Гистопатологическое исследование инфицированных образцов трахеи с окрашиванием H&E. (C) Анализ FACS для количественного определения CD 4 + и CD 8 + Т-клеток.Данные представляют собой средние значения ± SD из трех независимых экспериментов в трех повторностях. ∗ р < 0,05, ∗∗ р < 0,01.
Таблица 2. Анализ FACS для количественного определения CD 4 + и CD 8 + Т-клеток.
Патогенность вирусов H9N2 у мышей
Для оценки репликации вируса в образцах легких три мыши из каждой группы были подвергнуты эвтаназии через 3 dpi (рис. 5).До вскрытия вирус Mi/14 вызывал клинические признаки взъерошенной шерсти и депрессии. Примечательно, что 20% смертность и значительная потеря массы тела наблюдались при 8 dpi ( p <0,01). Однако другие вирусы не вызывали серьезных клинических признаков (рис. 5А). Все вирусы H9N2 реплицировались в легких без предварительной адаптации, кроме Gs/14. В частности, при 3 днях на дюйм легкие мышей, инфицированных штаммом Sw/08, содержали высокую вирусную нагрузку со средним титром 1,48 × 10 5 TCID 50 /г, штамм Hu/04 со средним титром 6.01 × 10 4 TCID 50 /г, вирус Ck/15 со средним титром 1,41 × 10 2 TCID 50 /г, вирус Mi/14 со средним титром 8,60 × 5 0 6 10 49000 TCID 50 /гр. Вирусы Sw/08 и Mi/14 показали более высокую репликацию в легких мышей, которая была в 10-1000 раз выше, по сравнению с Hu/04 и Ck/15 ( p < 0,01) (рис. 5C). ), которые представляли генотипы B35 (изолят Sw/08), B69 (изоляты Gs/14, Ck/15 и Mi/14) и G9 (изолят Hu/04) соответственно.
Рисунок 5. Патогенность вирусов H9N2 у мышей. Потеря массы тела (А) и выживаемость (В) отслеживались ежедневно в течение 14 дней после инокуляции. Вирусная нагрузка в образцах легких при инфицировании на 3 dpi (C) . Уровни провоспалительных цитокинов в образцах легких мышей при 3 днях на дюйм анализировали с помощью коммерческих наборов для сэндвич-ELISA (D) . Данные представлены как средние значения ± стандартное отклонение из трех независимых экспериментов в трех повторностях. ∗ р < 0,05, ∗∗ р < 0,01. гр, грамм.
Между тем, образцы легких мышей на 3 dpi использовались для проверки уровня цитокинов. Результаты показали, что Sw/08, Hu/04 и Mi/14 млекопитающих индуцировали более сильные провоспалительные реакции, на что указывало количественное определение IL-1β, IL-6, TNF-α, MCP-1 и MIP- Уровни 1α с помощью ELISA по сравнению с повышением уровня цитокинов, индуцированным Gs/14 и Ck/15 (фиг. 5D).
Обсуждение
Для изучения взаимосвязи между генотипами и патотипами были выбраны различные изоляты ВГП H9N2 из разных генотипов (В69, G1 и В35).С 2010 г. генотип B69 преобладает у кур и уток (Teng et al., 2016), и было показано, что генотип B69 обеспечивает внутренние гены для новых вирусов H7N9 и h20N8 при зоонозных инфекциях (Gao et al., 2013; Chen et al., 2013). и др., 2014б). У генотипа В69 отмечена высокая генетическая совместимость по сравнению с другими ВГА.
вируса H9N2 инфицируют многие виды (Group, 2013). У мышей штаммы Mi/14 (генотип B69) и Sw/08 (генотип B35) проявляли более высокую патогенность и более сильную провоспалительную реакцию (рис. 5).Очевидно, что инфекция Mi/14 показала значительную потерю веса при 8 dpi. Более того, 20% мышей погибли при 8 dpi. Это говорит о том, что вирус Mi/14 был более вирулентным у мышей. У кур изоляты Mi/14 (генотип B69) и Ck/15 (генотип B69) проявляли более высокую патогенность и более сильную провоспалительную реакцию (рис. 3, 4). Интересно, что более высокая вирусная нагрузка в легких показала более высокий провоспалительный ответ. Вероятно, антигенная нагрузка усиливала воспалительный ответ на вирусную инфекцию.Hu/08 показал более высокую репликацию у мышей, но более низкую у цыплят. Человеческий вирус реплицировался на более низких уровнях у цыплят, вероятно, адаптация к людям снизит репликацию вируса у цыплят. Более того, Gs/14 не может реплицироваться в легких мышей, но хорошо реплицируется в куриных. Таким образом, эти данные указывают на то, что один и тот же вирус в разных хозяевах демонстрировал сложную патотипическую картину. Предполагается, что патогенность вирусов H9N2 зависит от штамма.
Наши данные показывают, что Sw/08, Hu/04 и Mi/14 хорошо реплицируются в легких мышей, указывая на то, что вирусы млекопитающих реплицируются на более высоких уровнях, чем птичьи вирусы у мышей (рис. 5C).Более высокая адаптация указывает на то, что это важный этап передачи вируса от человека к человеку. Поразительно, но мутация D701N, связанная с адаптацией млекопитающих, наблюдалась в гене PB2 вирусов Mi/14 и Sw/08. Последовательности PA Sw/08, Hu/04, Ck/15 и Mi/14 были 70A и 594S, тогда как Gs/14 были 70V и 594S. Кроме того, R340K впервые обнаружен в гене PB2 вирусов Mi/14 и Sw/08. Это могут быть потенциальные детерминанты вирулентности, требующие подтверждения.
Также были проведены обширные исследования того, как вирусы H9N2 влияют на экспрессию предвоспалительных цитокинов (Geiss et al., 2001, 2002; Каш и др., 2004; Деген и др., 2006; Кобаса и др., 2007). Значительное увеличение IL-1β, IL-6, TNF-α, MCP-1 и MIP-1α в легких при инфицировании H9N2. Однако результаты снижения уровня IL-1β, индуцированного вирусом Hu/04, у цыплят сравнивают с Sw/08, Ck/15 и Mi/14. Однако вирус Hu/04 у мышей продуцировал тот же уровень продукции IL-1β, что и вирусы Mi/14 и Sw/08, что указывает на различия в реакциях хозяина на вирусный антиген. В совокупности эти пять цитокинов могут быть ключевыми факторами легочной патологии.Одной из причин различий в уровнях продукции IL-1β является способность к активации инфламмасомного комплекса при распознавании вирусного антигена, такого как PB1-F2 (McAuley et al., 2013). Интересно, что сайт мутации 79R, связанный с активацией воспаления, наблюдался в PB1-F2 всех пяти вирусов H9N2, кроме вируса Hu/04 (Alymova et al., 2018) (табл. 1). Наши результаты показали, что даже вирусы LPAI H9N2 хорошо реплицируются у мышей и цыплят и могут вызывать значительное увеличение экспрессии провоспалительных цитокинов (рис. 4A, 5D).Будущие обратные генетические анализы для создания точечных мутаций могут раскрыть молекулярный механизм вирулентности вирусов H9N2.
Вклад авторов
HC задумал и разработал исследование. HS, KW и WY разработали методологию. QL, JY, QT и XL получили данные (предоставили животных, материалы, регентов, оборудование и т. д.). HS, KW и HC проанализировали и интерпретировали данные. HS и HC написали, рассмотрели и/или отредактировали рукопись. HC и ZL руководили исследованием.
Финансирование
Работа выполнена при поддержке Национального фонда естественных наук Китая (грант No.31572502), Национальная ключевая программа исследований и разработок Китая (гранты № 2017YFD0500702 и 2017YFD0502302) и Ключевой проект по сельскому хозяйству Шанхайской сельскохозяйственной комиссии (грант № 201702080008F00068).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Каталожные номера
Алымова И.V., McCullers, J.A., Kamal, R.P., Vogel, P., Green, A.M., Gansebom, S., et al. (2018). Вирулентные остатки PB1-F2: влияние на приспособленность вируса гриппа А h2N1 у мышей и изменения в ходе эволюции вирусов гриппа А человека. Науч. Респ. 8:7474. doi: 10.1038/s41598-018-25707-y
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Bi, Y., Lu, L., Li, J., Yin, Y., Zhang, Y., Gao, H., et al. (2011). Новые генетические реассортанты вирусов гриппа А H9N2 и их разнообразная патогенность для мышей. Вирол. Дж. 8:505. дои: 10.1186/1743-422X-8-505
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Браун, В.Ф. (1964). Оценка дисперсии при определении пятидесятипроцентной конечной точки Рида-Мюнха. утра. Дж. Хиг. 79, 37–46.
Реферат PubMed | Академия Google
Butt, K.M., Smith, G.J., Chen, H., Zhang, L.J., Leung, Y.H., Xu, K.M., et al. (2005). Заражение человека вирусом птичьего гриппа А H9N2 в Гонконге в 2003 г. Дж. Клин. микробиол. 43, 5760–5767. doi: 10.1128/JCM.43.11.5760-5767.2005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Cauthen, A.N., Swayne, D.E., Schultz-Cherry, S., Perdue, M.L., and Suarez, D.L. (2000). Продолжающаяся циркуляция в Китае высокопатогенных вирусов птичьего гриппа, кодирующих ген гемагглютинина, связана со вспышкой гриппа H5N1 1997 года среди домашних птиц и людей. Дж. Вирол. 74, 6592–6599. doi: 10.1128/ОВИ.74.14.6592-6599.2000
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чен, Х., Энджел, М., Ли, В., Финч, К., Гонсалес, А.С., Саттон, Т., и соавт. (2014а). Бакмиды «все в одном»: эффективная стратегия обратной генетики для вакцин против вируса гриппа А. Дж. Вирол. 88, 10013–10025. doi: 10.1128/ОВИ.01468-14
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Chen, H., Yuan, H., Gao, R., Zhang, J., Wang, D., Xiong, Y., et al. (2014б). Клинико-эпидемиологическая характеристика смертельного случая заражения вирусом птичьего гриппа A h20N8: описательное исследование. Ланцет 383, 714–721. doi: 10.1016/S0140-6736(14)60111-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Деген, В.Г., Смит, Дж., Симмелинк, Б., Гласс, Э.Дж., Берт, Д.В., и Шейнс, В.Е. (2006). Молекулярное иммунофенотипирование легких и селезенки интактных и вакцинированных цыплят в ранние сроки после инфицирования вирусом легочного птичьего гриппа A (H9N2). Вакцина 24, 6096–6109. doi: 10.1016/j.vaccine.2006.05.027
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Донг, Г., Luo, J., Zhang, H., Wang, C., Duan, M., Deliberto, T.J., et al. (2011). Филогенетическое разнообразие и генотипическая сложность вирусов гриппа А H9N2, выявленные с помощью анализа геномной последовательности. PLoS ONE 6:e17212. doi: 10.1371/journal.pone.0017212
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Р. Гао, Б. Цао, Ю. Ху, З. Фэн, Д. Ван, В. Ху и др. (2013). Заражение человека новым вирусом птичьего гриппа A (H7N9). Н. англ. Дж. Мед. 368, 1888–1897 гг. дои: 10.1056/NEJMoa1304459
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гейсс, Г.К., Ан, М.С., Бумгарнер, Р.Е., Хаммерсмарк, Э., Каннингем, Д., и Катце, М.Г. (2001). Глобальное воздействие вируса гриппа на клеточные пути опосредуется как зависимыми от репликации, так и независимыми от репликации событиями. Дж. Вирол. 75, 4321–4331. doi: 10.1128/ОВИ.75.9.4321-4331.2001
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гейсс, Г.К., Сальваторе М., Тампи Т.М., Картер В.С., Ван X., Баслер С.Ф. и соавт. (2002). Профилирование клеточной транскрипции в эпителиальных клетках легких, инфицированных вирусом гриппа А: роль неструктурного белка NS1 в уклонении от врожденной защиты хозяина и его потенциальный вклад в пандемический грипп. Проц. Натл. акад. науч. США 99, 10736–10741. doi: 10.1073/pnas.112338099
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гибсон-Корли, К.N., Olivier, A. K., and Meyerholz, D. K. (2013). Principles for valid histopathologic scoring in research. Vet. Pathol. 50, 1007–1015. doi: 10.1177/0300985813485099
PubMed Abstract | CrossRef Full Text | Google Scholar
Guo, Y., Li, J., and Cheng, X. (1999). [Discovery of men infected by avian influenza A (H9N2) virus]. Zhonghua shi yan he lin chuang bing du xue za zhi = Zhonghua shiyan he linchuang bingduxue zazhi. Chin. J. Exp. Clin. Virol. 13, 105–108.
Реферат PubMed | Академия Google
Хуанг, К., Ван, К., Пан, Л., Ци, К., Лю, Х., и Чен, Х. (2017). Коинфицирование вирусом птичьего гриппа подтипа H9N2 и вирусом инфекционного бронхита снижает уровень экспрессии SP-A у кур. Вет. микробиол. 203, 110–116. doi: 10.1016/j.vetmic.2017.02.015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джин, Х., Ван, В., Ян, X., Су, Х., Фань, Дж., Чжу, Р., и другие. (2018). Эволюция вируса птичьего гриппа H9N2 в эмбрионах куриных яиц с гомологичными вакцинными антителами или без них. BMC Вет. Рез. 14:71. doi: 10.1186/s12917-018-1391-6
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джонс, Д.М., Артерс, Дж., и Бергер-Суини, Дж. (1999). Анестезия, индуцированная углекислым газом, не влияет на концентрацию биогенных аминов в головном мозге мышей. Лаб. Аним. науч. 49, 316–318.
Реферат PubMed | Академия Google
Каш, Дж. К., Баслер, С. Ф., Гарсия-Састре, А., Картер, В., Биллхарц, Р., Суэйн, Д. Е., и соавт.(2004). Глобальный иммунный ответ хозяина: патогенез и профилирование транскрипции вирусов гриппа типа А, экспрессирующих гены гемагглютинина и нейраминидазы из пандемического вируса 1918 года. Дж. Вирол. 78, 9499–9511. doi: 10.1128/ОВИ.78.17.9499-9511.2004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кобаса Д., Джонс С. М., Шинья К., Каш Дж. К., Коппс Дж., Эбихара Х. и соавт. (2007). Аберрантный врожденный иммунный ответ при летальном заражении макак вирусом гриппа 1918 года. Природа 445, 319–323. doi: 10.1038/nature05495
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Колер И., Мейер Р., Бусато А., Нейгер-Эшбахер Г. и Шацманн У. (1999). Является ли двуокись углерода (CO2) полезным анестетиком короткого действия для мелких лабораторных животных? Лаб. Аним. 33, 155–161.
Реферат PubMed | Академия Google
Лин, Ю.П., Шоу, М., Грегори, В., Кэмерон, К., Лим, В., Климов, А., и др. (2000).Передача от птиц к человеку вирусов гриппа А подтипа H9N2: взаимосвязь между человеческими изолятами H9N2 и H5N1. Проц. Натл. акад. науч. США 97, 9654–9658. doi: 10.1073/pnas.160270697
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Матросович, М. Н., Краусс, С., и Вебстер, Р. Г. (2001). Вирусы гриппа А H9N2 от домашней птицы в Азии обладают специфичностью рецептора, подобной человеческому вирусу. Вирусология 281, 156–162. doi: 10.1006/viro.2000.0799
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Маколи, Дж.L., Tate, M.D., MacKenzie-Kludas, C.J., Pinar, A., Zeng, W., Stutz, A., et al. (2013). Активация инфламмасомы NLRP3 белком вирулентности IAV PB1-F2 способствует тяжелой патофизиологии и заболеванию. PLoS Pathog. 9:e1003392. doi: 10.1371/journal.ppat.1003392
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нойманн, Г., Макен, К.А., и Каваока, Ю. (2014). Идентификация изменений аминокислот, которые могли иметь решающее значение для генезиса вирусов гриппа A(H7N9). Дж. Вирол. 88, 4877–4896. doi: 10.1128/ОВИ.00107-14
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Pu, J., Wang, S., Yin, Y., Zhang, G., Carter, R. A., Wang, J., et al. (2015). Эволюция генотипа гриппа H9N2, которая способствовала возникновению нового вируса H7N9. Проц. Натл. акад. науч. США 112, 548–553. doi: 10.1073/pnas.1422456112
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сайто, Т., Lim, W., Suzuki, T., Suzuki, Y., Kida, H., Nishimura, S.I., et al. (2001). Характеристика человеческого вируса гриппа H9N2, выделенного в Гонконге. Вакцина 20, 125–133. doi: 10.1016/S0264-410X(01)00279-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Седири, Х., Швальм, Ф., Габриэль, Г., и Кленк, Х. Д. (2015). Адаптивная мутация PB2 D701N способствует ядерному импорту вРНП вируса гриппа в клетки млекопитающих. евро. Дж. Клеточная биология. 94, 368–374.doi: 10.1016/j.ejcb.2015.05.012
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сунь Х., Яо В., Ван К., Цянь Ю., Чен Х. и Юнг Ю. С. (2017). Ингибирование пути неддиляции подавляет репликацию вируса гриппа и провоспалительные реакции. Вирусология 514, 230–239. doi: 10.1016/j.virol.2017.11.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тамура К., Стечер Г., Петерсон Д., Филипски А. и Кумар С.(2013). MEGA6: Молекулярно-эволюционный генетический анализ версии 6.0. Мол. биол. Эвол. 30, 2725–2729. doi: 10.1093/molbev/mst197
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Teng, Q., Xu, D., Shen, W., Liu, Q., Rong, G., Li, X., et al. (2016). Единственная мутация в положении 190 в гемагглютинине увеличивает аффинность связывания с рецептором сиаловой кислоты человеческого типа и репликацию вируса птичьего гриппа H9N2 у мышей. Дж. Вирол. 90, 9806–9825.doi: 10.1128/ОВИ.01141-16
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ван К., Хуанг К., Ян З., Ци К., Лю Х. и Чен Х. (2017). Альтернативная система обратной генетики вирусов гриппа на основе синтезированного свиного промотора 45S рРНК. Гены вируса 53, 661–666. doi: 10.1007/s11262-017-1457-8
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сяо, К., Ма, В., Сунь, Н., Хуанг, Л., Ли, Ю., Цзэн, З., и другие.(2016). PB2-588 V способствует адаптации вирусов птичьего гриппа h20N8, H7N9 и H9N2 к млекопитающим. Науч. Респ. 6:19474. дои: 10.1038/srep19474
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Xu, G., Zhang, X., Gao, W., Wang, C., Wang, J., Sun, H., et al. (2016). Преобладающая мутация PA K356R в вирусе птичьего гриппа H9N2 увеличивает репликацию и патогенность млекопитающих. Дж. Вирол. 90, 8105–8114. doi: 10.1128/ОВИ.00883-16
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сюэ, Р., Tian, Y., Hou, T., Bao, D., Chen, H., Teng, Q., et al. (2018). Вирус гриппа H9N2, выделенный из норок, обладает повышенной вирулентностью у мышей. Трансграничный. Эмердж. Дис. 65, 904–910. doi: 10.1111/tbed.12805
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Zhao, Y.X., Li, S., Zhou, Y.F., Song, W.G., Tang, Y.J., Pang, Q.H., et al. (2015). Филогенетический анализ генов гемагглютинина вирусов птичьего гриппа H9N2, выделенных от кур в провинции Шаньдун, Китай, в период с 1998 по 2013 год. Биомед Рез. Междунар. 2015:6. дои: 10.1155/2015/267520
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
90 000 крупных млекопитающих находятся под повышенным риском исчезновения в беднейших странах мира — ScienceDaily нашел.Обзор, в котором рассматривалось 81 исследование, проведенное в период с 1980 по 2020 год, показал, что незаконная охота вызывает тревожное сокращение популяций крупных млекопитающих в охраняемых районах по всему миру, особенно в более бедных странах.
На четырех континентах, включенных в исследование, было обнаружено, что 294 различных вида млекопитающих стали объектом незаконной охоты в национальных парках, созданных для их защиты.
Угрожающие виды, с которыми сталкиваются более бедные страны, могут быть связаны с бумом торговли мясом диких животных и нехваткой ресурсов для сохранения.
Чтобы защитить виды, правительства и политики должны сосредоточиться на борьбе с бедностью среди людей, настаивают исследователи.
Ведущий автор исследования, д-р Альфан Риджа из Сельскохозяйственного университета Сокойн в Танзании, проводивший обзор, будучи аспирантом Йоркского университета, сказал: «Нам уже несколько лет известно, что незаконная охота сокращает популяции млекопитающих, но наш обзор показывает, что это происходит даже внутри охраняемых территорий и особенно затрагивает крупных млекопитающих (с массой тела более 100 кг) в беднейших странах.В более бедных странах, таких как Танзания, где я живу, мясо диких животных является ценным источником дохода и белка в некоторых районах, а также меньше ресурсов, доступных для инвестирования в безопасность и охрану национальных парков.
«Угроза незаконной охоты особенно опасна для крупных млекопитающих, потому что они имеют медленные темпы роста, и поэтому чрезмерная охота может привести к сокращению популяции.
«Помимо опасений по поводу того, что будущее многих из этих видов находится в опасности, потеря млекопитающих из-за незаконной охоты связана со значительной потерей важных функциональных характеристик в экосистеме.Они поддерживают многие экологические взаимодействия, такие как распространение семян и регенерация, и их сокращение угрожает более широкому биоразнообразию».
Исследование показало, что в целом по всему миру более строго охраняемые территории демонстрируют более низкие темпы сокращения популяции крупных млекопитающих. Однако этого не произошло в Азии, где более строгие национальные парки имели более высокие показатели незаконной охоты и сокращения видов.
Старший автор исследования, доктор Колин Бил с факультета биологии Йоркского университета, сказал: «Наше исследование дополняет растущее число исследований, которые предполагают, что Азия в настоящее время является особым центром незаконной торговли частями тела диких животных.Несмотря на строгие законы, нелегальные охотники могут быть вынуждены проникнуть в охраняемые районы, где все еще обитают наиболее востребованные виды, такие как снежный барс, тигр, панголин, орангутанг и солнечный медведь.
«Повышение эффективности азиатских охраняемых территорий будет иметь важное значение для укрепления сохранения биоразнообразия в континентальной Азии и, вероятно, потребует ряда мер, включая обеспечение эффективного правоприменения, а также работу с сообществами в и вокруг ценных районов дикой природы».
Исследование проводилось в сотрудничестве с Департаментом управления дикой природой Сельскохозяйственного университета Сокойн в Танзании.
Источник истории:
Материалы предоставлены Университетом Йорка . Примечание. Содержимое можно редактировать по стилю и длине.
Приводит ли инвазивность плаценты к более высоким показателям злокачественной трансформации у млекопитающих? | Эволюция, медицина и общественное здравоохранение
В нашем исследовании Распространенность рака в течение жизни и особенности жизненного цикла у млекопитающих мы сообщили о распространенности новообразований и злокачественных новообразований в избранной группе млекопитающих, содержавшихся в глобальном зоопарке Сан-Диего с 1964 по 1978 и с 1987 по 2015 годы [1].Мы также использовали эти данные для оценки связи между чертами истории жизни и показателями здоровья населения. Наш анализ показал, что инвазивность плаценты не может предсказать долю животных с диагнозом неоплазия или злокачественное новообразование. В ответ на нашу статью доктор Вагнер и его коллеги описывают другой расчет для проверки взаимосвязи между плацентарной инвазивностью и злокачественностью. Они идентифицировали и включили ранее опубликованные ветеринарные данные о новообразованиях и злокачественных новообразованиях в наш опубликованный набор данных и предложили положительную связь между плацентарной инвазивностью и развитием злокачественных новообразований (называемых уровнем злокачественных новообразований в ответе Вагнера и его коллег).Эти данные подтвердили гипотезу эволюционных уровней инвазивности (ELI) [2]. Мы рады, что другие исследователи находят наши данные полезными, и полностью согласны с доктором Вагнер и его коллегами в необходимости получения дополнительных данных о раке у самых разных видов. Тем не менее, этот обновленный анализ поднимает ряд тем, которые мы хотели бы рассмотреть.
В соответствии с гипотезой Evolved Levels of Invasiveness [2], доктор Вагнер и его коллеги предполагают связь между скоростью развития злокачественных новообразований и плацентацией в подгруппе животных, у которых уже диагностирована неоплазия.Хотя мы согласны с тем, что это интересный вопрос, который требует дальнейшего изучения, это не был первоначальный вопрос, на который было направлено наше исследование. Наш подход рассчитывал распространенность злокачественных новообразований среди животных, подвергшихся вскрытию, как меру бремени болезни и здоровья популяции. Действительно, эти два вопроса (и методы расчета этих вопросов) дополняют друг друга и могут улучшить наше понимание злокачественных новообразований у млекопитающих. Доктора Вагнер и его коллеги проводят важное различие между развитием неоплазии и переходом от доброкачественного новообразования к злокачественному новообразованию, называемому «трансформацией» в биологии рака.С помощью своих расчетов они проверяют, предсказывает ли инвазивность плаценты злокачественную трансформацию у млекопитающих. Мы согласны с важностью этого перехода в неопластической прогрессии, и, учитывая определение злокачественного новообразования как инвазивного и/или метастатического новообразования, кажется разумным, что трансформация может быть связана с биологией плацентарной инвазии. Кроме того, следует уточнить, что не все злокачественные опухоли являются метастатическими (т. е. перемещаются в отдаленные органы), и могут также существовать различные механизмы, лежащие в основе развития метастазирования.
Объединяя результаты двух описанных выше подходов, текущие данные позволяют предположить, что тип плаценты не объясняет различий в распространенности неоплазии или злокачественных новообразований у млекопитающих. Однако из животных, у которых развиваются доброкачественные опухоли, виды с более инвазивной плацентой (гемохориальные) могут иметь более высокие показатели злокачественной трансформации. Кроме того, процессы, вызывающие превращение доброкачественных опухолей в злокачественные, различаются для разных подтипов рака. Например, предыдущие данные свидетельствуют о том, что злокачественная трансформация при нежелезистом эпителиальном раке высока (> 83%) для всех типов плаценты, но скорости трансформации между видами (корова/лошадь по сравнению с собакой/кошкой) от доброкачественных до злокачественных различаются для кожи и кожи. железисто-эпителиальный рак [3, 4].Эти результаты позволяют предположить, что клеточное и тканевое происхождение опухоли может быть важным компонентом связи между риском злокачественной трансформации и плацентарной инвазивностью.
Мы хотели бы сделать одну оговорку в отношении данных, использованных в ответах доктора Вагнер и ее коллег. В повторном анализе авторы добавили данные о злокачественных новообразованиях и новообразованиях четырех дополнительных видов (кошка, собака, лошадь и корова) из 12 колледжей ветеринарной медицины США и Канады, собранные с 1964 по 1969 год и опубликованные в журнале Priester and Mantel. 4] в 1971 году.Мы рекомендуем проявлять осторожность при использовании данных о домашних видах, поскольку искусственное селективное давление одомашнивания могло превзойти естественное селективное давление рака. Также представляется, что данные, добавленные в повторном анализе Вагнера и его коллег, были ограничены раком кожи, железистого эпителия, нежелезистого эпителия и соединительной ткани (например, 208 злокачественных опухолей крупного рогатого скота [3] по сравнению с 401, о которых сообщил Пристер). и Мантель [4]). Наши данные о раке не ограничивались анатомическим типом или типом ткани.Доктор Вагнер и его коллеги пришли к выводу, что текущий набор данных слишком мал, чтобы явно проверить их прогнозы. Более крупный набор данных, включая высококачественные данные из контролируемых популяций, проанализированные в сотрудничестве с сертифицированными ветеринарными патологоанатомами, поможет продвинуть вперед сравнительную онкологию и эволюцию злокачественных новообразований.
Конфликт интересов: Доктор Шиффман является соучредителем, акционером и сотрудником компании PEEL Therapeutics, Inc., разрабатывающей основанные на эволюции лекарства, основанные на устойчивости к раку у слонов.Доктор Абегглен является акционером и консультантом PEEL Therapeutics, Inc.
ССЫЛКИ
1
Кузов
AM
Абегглен
LM
Пессье
АП
и другие.Распространенность рака в течение жизни и особенности жизненного цикла млекопитающих
.Evol Med Public Health
doi:10.1093/emph/eoaa015.2
Kshitiz
Afzal J
Мазиарз
JD
и другие.Развитие плацентарной инвазии и метастазирования рака причинно связаны
.Nat Ecol Evol
2019
;3
:1743
–53
.3
Д’Суза
AW
Вагнер
ГП.
Злокачественный рак и инвазивная плацентация: случай положительной плейотропии между эндометриальным и злокачественным фенотипами
.Evol Med Общественное здравоохранение
2014
;2014
:136
–45
.4
Священник
WA
Каминная полка
№
Возникновение опухолей у домашних животных. Данные 12 колледжей ветеринарной медицины США и Канады
.J Natl Cancer Inst
1971
;47
:1333
–44
.© Автор(ы), 2020. Опубликовано Oxford University Press от имени Фонда эволюции, медицины и общественного здравоохранения.
Это статья в открытом доступе, распространяемая в соответствии с лицензией Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/), которая разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии, что оригинал работа цитируется правильно. .