
Место турбеллярий в эволюции беспозвоночных
Наиболее примитивным типом первичноротых, безусловно, надо считать плоских червей и среди них класс свободноживущих ресничных червей (Turbellaria, 10). Представители этого класса имеют еще ряд признаков, связывающих их с кишечнополостными.
Сюда относятся: отсутствие анального отверстия, элементы лучевой симметрии в строении ряда органов, ресничный покров. Часть плоских червей перешла к паразитизму, сперва, по-видимому, в качестве наружных паразитов, затем — внутренних (классы сосальщиков (Trematoda, 11) и ленточных червей (Cestoidea, 12). Переход к такому специфическому образу жизни вызвал ряд регрессивных изменений, например полное исчезновение кишечника у ленточных червей. Вместе с тем выработался ряд адаптивных признаков, тесно связанных с жизнью на хозяине или внутри него, например органы прикрепления, плотная кутикула.
Организмы, близкие к современным турбелляриям (их можно назвать первичными трехслойными), были, по всей вероятности, исходными формами для ряда более высокоорганизованных групп первичноротых.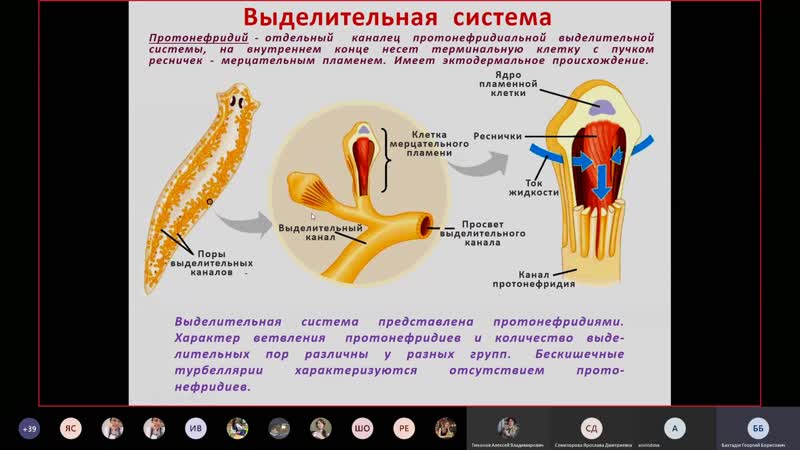
Прогрессивные изменения их выразились в появлении качественно новых признаков, поднявших организацию этих животных на новую ступень. Среди этих признаков одними из важнейших были: образование анального отверстия, большая концентрация нервной системы и усиление мускулатуры.
Безусловно, прогрессивным, но вместе с тем еще очень тесно связанным с плоскими червями является тип немертин (Nemertini, 18), представляющий особую ветвь развития первичноротых. У них имеется сквозной кишечник, хорошо развитый головной нервный узел, замкнутая кровеносная система и очень мощная мускулатура, достигающая у некоторых из них до 60% всей массы тела (Зенкевич, 1944). Вместе с тем у немертин имеются еще признаки плоских червей — ресничный покров, отсутствие настоящей полости тела и протонефридиальная выделительная система. Наличие этих признаков дает основание ряду авторов рассматривать немертин даже только как подтип плоских червей.
Еще интересные статьи по теме:
Тип Плоские черви — презентация онлайн
1.
 Типы Червей 7 класс
Типы Червей 7 класс2. Общая характеристика типа Плоские черви (15 000 видов)
Двусторонне-симметричные животныеТрехслойные (появляется мезодерма)
Тело имеет лентовидную или листовидную
форму, сплющено в спинно-брюшном
(дорзо-вентральном) направлении)
Кожно-мускульный мешок – однослойный
эпителий с ресничками + 3 слоя мышц
(кольцевые, продольные, диагональные)
Полости тела нет, пространство между
органами заполнено паренхимой
3. Общая характеристика типа Плоские черви
Пищеварительная система слепозамкнутая.Непереваренные остатки удаляются через рот
Газообмен через поверхность кожи.
Выделительная система –
протонефридиального типа
Нервная система лестничного типа – парные
(комиссуры)
Гермафродиты. Оплодотворение внутреннее,
развитие прямое или с метаморфозом
7. Класс Ресничные черви (планария молочная)
СвободноживущиеВодные, реже почвенные
Тело уплощенное и вытянутое, 2-3 см в
длину
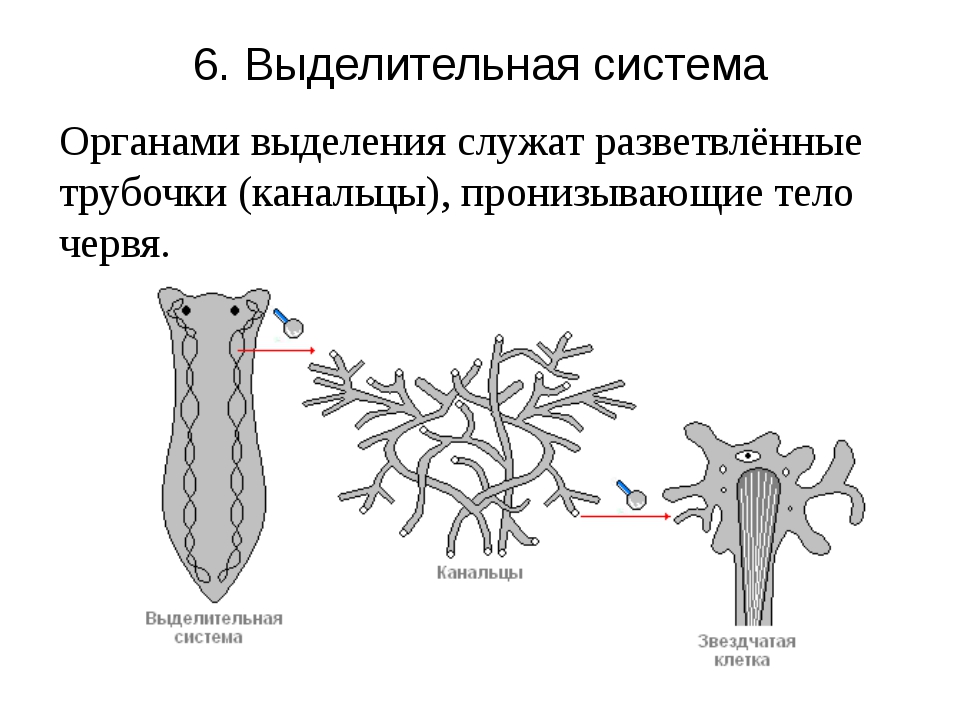
Хищник, питается мелкими животными
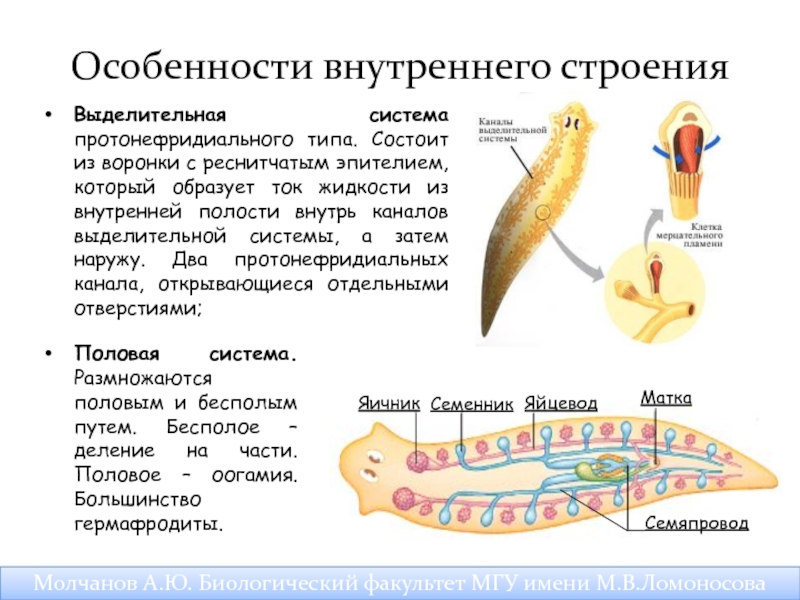
Протонефридий – это звездчатые клетки,
лежащие в паренхиме и выделительные
канальцы, сливающиеся в два канала
Помимо полового встречается бесполое
размножение – поперечным делением.
Недостающая часть достраивается
регенерацией
9. Системы органов плоских червей
14. Класс Сосальщики (Трематоды). 4000 видов
ЭндопаразитыТело листовидное, 4-5 см
Две присоски – ротовая и брюшная,
снабжены мощными мышцами
Кожно-мускульный мешок представлен
тегументом и мышцами
Пищеварительная система есть, но могут
поглощать пищу через поверхность тела
Анаэробное дыхание
Из органов чувств – осязание и химическое
чувство
15. Жизненный цикл
Промежуточный хозяин – организм, вкотором паразит не размножается или
размножается бесполым путем
Окончательный хозяин – это организм, в
котором происходит половое размножение
паразита
16. Цикл развития печеночного сосальщика
18. Меры профилактики заражения печеночным сосальщиком
Сокращение численности малогопрудовика
Изоляция и лечение больных животных
Запрет на питье сырой воды
Соблюдение правил личной гигиены (не
жевать травинки, сорванные на лугах)
Установление мест для водопоя скота в
строго определенных местах
19.
 Класс ленточные черви (Цестоды) – 3000 видов Эндопаразиты, окончательные хозяева
Класс ленточные черви (Цестоды) – 3000 видов Эндопаразиты, окончательные хозяевакоторых позвоночные животные
Очень длинные, на переднем конце тела
головка – сколекс с органами прикрепления
(крючья, присоски, ботрии – щелевидные
присоски), затем шейка и членистое
туловище (стробила), которая состоит из
незрелых члеников (мужских и
гермафродитных) и зрелых члеников
(проглоттид)
20. Класс ленточные черви
Мужские членики расположены ближе к шейке,первых члениках он недоразвит
В гермафродитных члениках хорошо развит
мужской и женский половой аппарат
Женская система состоит из одного яичника и
яйцевода, впадающего в оотип (происходит
оплодотворение)
Есть матка, в которой созревают яйца
Тело покрыто тегументом
Пищеварительной системы нет
Анаэробное дыхание
24. Сравнительная характеристика ленточных червей
признакБычий цепень
(невооружены
й)
Свиной
цепень
9вооруженны
й)
эхинококк
Размер тела
8-12 м, 1-2
тыс.
 члеников
члеников2-3 м., до 1
тыс. члеников
5-6 мм, 3-4
членика
Органы
фиксации на
сколексе
4 присоски
4 присоски и
хитиновые
крючья
4 присоски и
хитиновые
крючья
Половая
система
Матка с 17-35
парами
боковых
ответвлений
Матка с 7-12
парами
боковых
ответвлений
Мешковидная
матка
25. Цикл развития бычьего цепня
30. Жизненный цикл широкого лентеца
31. Меры профилактики и борьбы с ленточными червями
Термическая обработка мяса и рыбыСоблюдение санитарного контроля над
продуктами питания
Соблюдение личной гигиены
Изоляция и лечение больных животных
33. Общая характеристика Типа Круглые черви
Бывают как свободноживущие (моря,пресные водоемы и почва), так и паразиты
Круглые в поперечном сечении
Двусторонняя симметрия, трехслойность
Есть первичная полость тела, заполненная
жидкостью
Кожно-мускульный мешок.
 Мышцы только
Мышцы толькопродольные
В пищеварительной системе появляется
анальное отверстие
34. Общая характеристика Типа Круглые черви
Выделительная системапротонефридиальная или особые кожные
железы
Нервная система лестничного типа
(окологлоточное нервное кольцо и нервный
ствол с перемычками)
Раздельнополые
Размножение только половое
Способность к регенерации отсутствует
35. Класс Собственно Круглые черви (Нематоды) – человеческая аскарида
36. Общая характеристика на примере Человеческой аскариды
Тело веретенообразноепродольные мышцы
Изнутри гиподермы 4 валика – в них
каналы нервной и выделительной системы
Первичная полость заполнена жидкостью –
поддерживается повышенное давление
Первичная полость – гидроскелет,
транспортные функции, участвует в
движении
37. Общая характеристика на примере Человеческой аскарид
Питается содержимым кишечника хозяинаВыделительная система – одноклеточная
кожная железа и два выделительных
канала, открываются наружу общей порой
Нервная система – окологлоточное кольцо
и два нервных ствола
Органы чувств – осязательные и
обонятельные клетки
41.
 Жизненный цикл острицы
Жизненный цикл острицы43. Меры профилактики
Строгое соблюдение гигиены (мытье рукперед едой)
Соблюдение гигиены питания (тщательное
мытье овощей и фруктов)
Соблюдение санитарных норм:
недопущение загрязнения водоемов и почв
фекалиями, организация закрытых туалетов
и мусорных ям
Борьба с переносчиками яиц (мухами)
44. Меры профилактики глистных заболеваний. Борьба с червями-паразитами
Основоположник современнойгельминтологии К.И. Скрябин
1. Соблюдение личной гигиены и гигиены
питания
2. Санитарный контроль продуктов питания
3. Борьба с переносчиками яиц и личинок (с
мухами, тараканами, моллюсками и т.д.)
4. Термическая обработка рыбы и мяса
5. Изоляция и лечение больных животных
45. Тип Кольчатые черви насчитывает 12 000 видов
47. Общие признаки типа
Двусторонняя симметрия, трехслойностьПоявляется вторичная полость тела – целом,
выстланная эпителием
Тело разделено на сегменты – кольца.
 Такая
Такаязакономерность называется метаморфией
Сегментация делится:
А. гомономная – все членики, кроме первых
двух, построены одинаково
Б. гетерономная – все членики, за исключением
первых двух отличаются от других
48. Общие признаки типа
Кожно-мускульный мешок состоит изкутикулы и 2 слоев мышц (продольного и
кольцевого)
Появляется кровеносная система
замкнутого типа
Дыхательная система представлена
Выделительная система
метанефридиального типа. Метанефридий
состоит из воронки, извитого канальца и
выделительной поры
Нервная система состоит из пары спинных
ганглиев и брюшной нервной цепочки
51. Класс Многощетинковые
В основном морские свободноживущиеНа головном отделе находятся глаза и
чувствительные придатки – щупальца, усики и
пальпы.
На туловищных сегментах – боковые выросты с
щетинками – параподии (органы движения),
чувствительные усики и жабры
В пищеварительной системе нет зоба
В глотке есть хитиновые челюсти и шипы
Нереида – кожное дыхание, пескожил – жабры
Оплодотворение внешнее.
 Размножение
Размножениеполовое и бесполое (поперечное деление или
почкование) Развитие с личинкой (трохофорой)
54. Класс Малощетинковые (дождевой червь)
Тело длинное, червеобразное,сегментированное
Число сегментов от 100 до 180
На первом рот, на анальной лопасти –
анальное отверстие
На каждом сегменте 4 пары щетинок
На однослойном эпителии, входящем в
кожно-мускульный мешок много желез,
выделяющих слизь
Кожа выделяет тонкую кутикулу
56. Класс Малощетинковые (дождевой червь)
Под кожей первый слой кольцевые мышцы,второй слой – продольные, затем
целомический эпителий
Целом заполнен жидкостью – внутренняя
среда организма
В целоме находятся половая и
выделительная система
Целом выполняет функции: транспортную,
выделительную, половую, опорнодвигательную
57. Класс Малощетинковые (дождевой червь)
Стенки пищевода имеют железы, секреткоторых нейтрализует гуминовые кислоты
Кровеносная система представлена 2
продольными сосудами, соединенными
кольцевыми
Кровь красная, так как содержит
гемоглобин
Дыхание через кожу
Выделение через метанефридии
Способны к регенерации после
перерезания
63.
 Выделительная и нервная системы
Выделительная и нервная системы66. Размножение дождевого червя
Гермафродиты: семенники в 10-11От семенников отходят семяпроводы,
которые открываются на 15 сегменте
От яичников отходят яйцеводы,
открывающиеся на 14 сегменте
Семяприёмники на 9 и 10 сегменте
На 32-37 сегментах развивается поясок,
выделяющий слизистую муфту
Оплодотворение внутреннее
69. Общая характеристика класса Пиявки (медицинская пиявка)
Эктопаразиты (питаются кровью) илихищники
Тело сплющено в спинно-брюшном
направлении
Параподий и щетинок нет
Две присоски – на переднем (треугольная)
и заднем (округлая) конце тела
Сегментация ложная – каждому сегменту
соответствует 3-5 наружных колец
71. Общая характеристика класса Пиявки (медицинская пиявка)
Кожно-мускульный мешок представленоднослойным эпителием (на поверхности –
кутикула) с большим количеством желез и
пигментных клеток и мышцами
(продольными и кольцевыми)
Целом редуцирован и превращен в
лакунарную систему, содержащую кровь
Большая часть полости заполнена
паренхимой
73.
 Общая характеристика класса Пиявки (медицинская пиявка) В пищеварительной системе имеется
Общая характеристика класса Пиявки (медицинская пиявка) В пищеварительной системе имеетсяхоботок или челюсти для прокусывания
кожи жертвы
Слюнные железы содержат гирудин –
вещество, препятствующее свертыванию
крови
Кровеносная система сильно редуцирована,
ее функцию выполняет лакунарная система
Не способны к регенерации
Органы чувств – глаза, органы осязания и
химического чувства
78. Значение кольчатых червей
Важное звено в цепи питания рыб,млекопитающих и т.д.
Участвуют в почвообразовании, улучшают
структуру и аэрацию почвы. Рыхлят и
обогащают почву органическими
веществами
Участвуют в круговороте веществ
Выполняют очистку морской воды
Применяются в лечебных целях
(медицинская пиявка). Гирудотерапия –
лечение с помощью пиявок
Общая характеристика круглых червей
Тип круглые черви на сегодняшний день включает до 25 тысяч видов, но скорость, с которой ученые выявляют новые виды, заставляет предполагать, что в реальности круглых червей несоизмеримо больше, и впереди еще много открытий.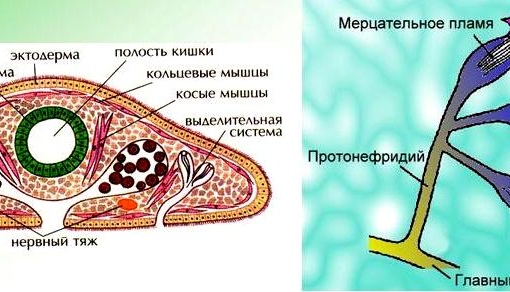 Эти животные обитают повсюду, в водоемах и мокрой почве, им свойственны как свободный образ жизни, так и паразитизм. С точки зрения паразитологии наиболее интересен класс собственно круглые черви, или же нематоды. Среди них уже обнаружено не менее 3 тысяч видов, ведущих паразитический образ жизни внутри разных животных и человека и вызывающих нематодозы — достаточно сложные заболевания.
Эти животные обитают повсюду, в водоемах и мокрой почве, им свойственны как свободный образ жизни, так и паразитизм. С точки зрения паразитологии наиболее интересен класс собственно круглые черви, или же нематоды. Среди них уже обнаружено не менее 3 тысяч видов, ведущих паразитический образ жизни внутри разных животных и человека и вызывающих нематодозы — достаточно сложные заболевания.
Происхождение круглых червей
Хотя вопрос о том, как и от кого эволюционировали круглые черви, до сих пор не закрыт окончательно, существует достаточно убедительная теория — их предками были морские ресничные плоские черви. Несмотря на отличия в строении плоских ресничных и круглых червей (в частности, отсутствие ресничек, кольцевых и диагональных мышц, круглое в разрезе тело с внутренней полостью там, где у ресничных сплошная желеобразная паренхима), все-таки есть связующее звено — примитивный круглый червь, принадлежащий к классу брюхоресничных, который живет в придонных слоях водоемов.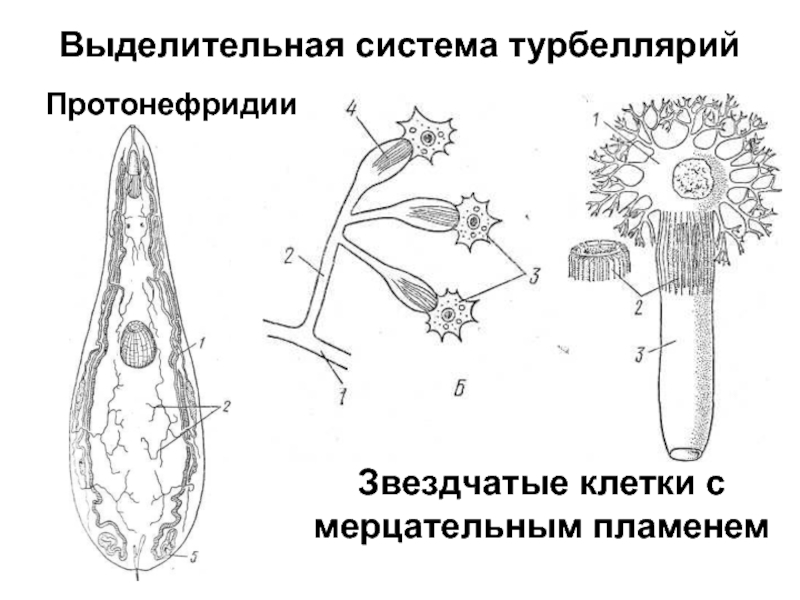 Он как раз имеет признаки и тех, и других животных.
Он как раз имеет признаки и тех, и других животных.
Строение круглых червей
1. Круглые черви — животные трехслойные, обладающие двусторонней симметрией.
2. Тела их червей напоминают нити, веретена, бочонки или лимоны, в зависимости от вида.
3. Черви очень отличаются по размерам — от совсем крошечных, не достигающих миллиметра, до восьмиметровых гигантов.
4. Внешний покров — кутикула, она может иметь поперечные бороздки-кольца или быть оснащенной приспособлениями для прикрепления.
5. Следующий внутренний слой, гиподерма, состоит не из отдельных клеток, окруженных стенками, а из обособленных «кусочков» цитоплазмы с ядрами, между которыми построены цитоплазматические мостики.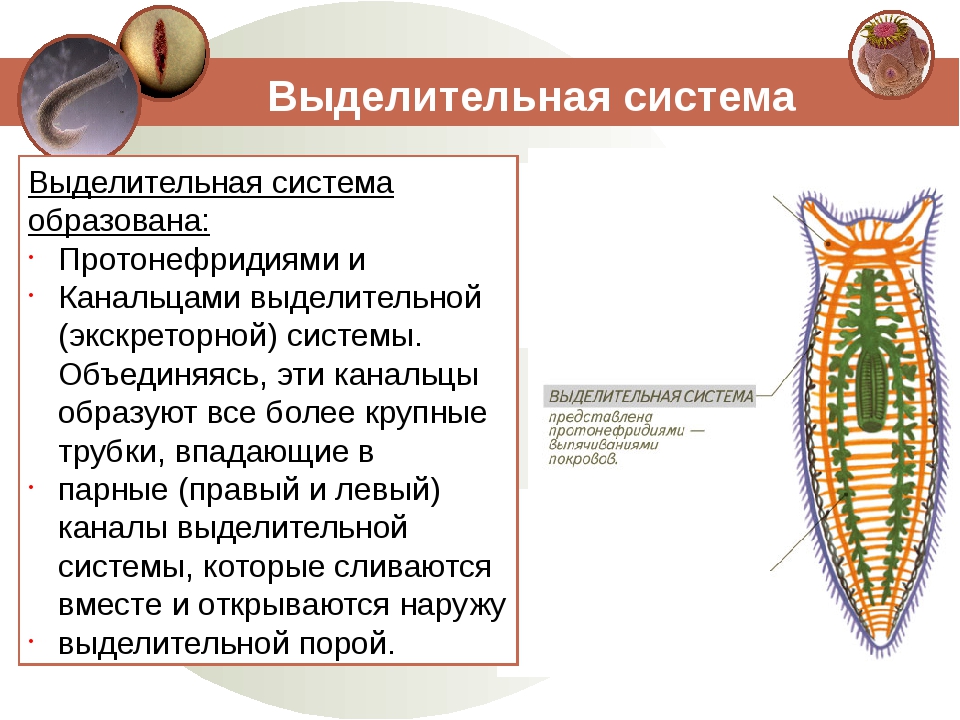 Такой тип ткани называется синтицием. В гиподерме тянутся специфические продольные валики: брюшной, спинной и пара боковых.
Такой тип ткани называется синтицием. В гиподерме тянутся специфические продольные валики: брюшной, спинной и пара боковых.
6. У круглых червей имеются только продольные мышцы. Таким образом, кутикула, гиподерма и внутренняя мускулатура составляют кожно-мускульный мешок.
7. Круглые черви впервые на эволюционном пути обзавелись первичной полостью тела — так называемой схизоцелью, которая еще не имеет эпителиальной выстилки. Внутри полости находится жидкость под давлением, с ее помощью перераспределяются питательные вещества.
Системы органов
1. Пищеварительная система, разомкнутая вовне, по сути, является сквозной трубочкой. Пища поступает сначала в рот, потом в глотку — это передняя кишка. Далее идут средняя кишка и задняя, завершающаяся анальным отверстием. У хищников и некоторых паразитирующих видов рот оснащен губами и зубами.
Пища поступает сначала в рот, потом в глотку — это передняя кишка. Далее идут средняя кишка и задняя, завершающаяся анальным отверстием. У хищников и некоторых паразитирующих видов рот оснащен губами и зубами.
2. Нет ни кровеносной, ни дыхательной систем. Обмен газами идет через поверхность тела. Паразиты, живущие во внутренних органах, — анаэробы, расщепляющие гликоген.
3. Выделительная система — протонефридиальная. Представлена она одно- или многоклеточными железами на шее червя, боковыми выводными протоками и клетками-псевдоцеломоцитами.
4. Нервная система лестничного типа состоит из ганглиозного кольца, расположенного около глотки, отходящих от него двух нервных стволов, и еще нескольких нервов, которые соединяются перемычками.
5. Органы чувств в основном развиты слабо, имеются хеморецепторы и разнообразные сенсиллы, отвечающие за осязание и обоняние.
6. Среди круглых червей обнаружены гермафродитные виды, но в подавляющем большинстве они раздельнополы с отчетливым половым диморфизмом.
7. Оплодотворение у нематод внутреннее, самки разных видов способны как откладывать яйца с личинками внутри, так и рожать «готовые» личинки. Интересно, что из яиц личинки могут вылупиться еще внутри хозяина, до выхода в окружающую среду.
Хочешь сдать экзамен на отлично? Жми сюда — онлайн тесты по биологии ОГЭ
Протонефридиальная выделительная система у Vestimentifera (Siboglinidae, Annelida)
Rouse, G.W. и Fauchald, K., Zool. Сценарий , 1997, том. 26, нет. 26, стр. 139–204.
Артикул Google ученый
Струк, Т.Х., в Эволюция ядовитых животных и их токсинов , 2015, стр. 1–12.
Книга Google ученый
 «>
«> Ван дер Ланд, Дж.и Норреванг, А., К. Дан. Виденск. Сельск. биол. скр. , 1977, том. 21, стр. 1–102.
Google ученый
Гардинер, С.Л. и Джонс, М.Л., Микроскопическая анатомия беспозвоночных , Нью-Йорк: Wiley-Liss, 1993, vol. 12.
Google ученый
Southward, EC, J. Mar. Biol. доц. Великобритания , 1988, том. 68, стр. 465–487.
Артикул Google ученый
Малахов В.В., Попеляев И.С., Галкин С.В., биол. Моря , 1996, вып. 22, нет. 5, стр. 275–284.
Google ученый
Schulze, A., J. Morphol. , 2001, том. 250, стр. 1–11.
КАС Статья пабмед Google ученый
 «>
«> Кох, М., Кваст, Б., и Бартоломеус, Т., в Глубокая филогения многоклеточных животных: основа древа жизни — новые идеи из анализа молекул, морфологии и теории анализа данных , Берлин: Де Грюйтер, 2014, стр.173–284.
Google ученый
Бартоломеус Т. и Кваст Б., в Морфология, молекулы, эволюция и филогения полихет и родственных таксонов , 2005, стр. 139–165.
Книга Google ученый
Карасева Н.П., Малахов В.В., Галкин С.В., Биол. Моря , 2012, том. 38, нет. 2, стр. 122–129.
Google ученый
Роуз Г.В., Зоол. Дж. Линн. соц. , 2001, том. 132, стр. 55–80.
Артикул Google ученый
Иванов А.В., Погонофоры (Pogonophora). Животный мир СССР. Новая серия (Фауна Советского Союза: Новая серия), Москва: Акад. Наук СССР, 1960, вып. 75.
Новая серия (Фауна Советского Союза: Новая серия), Москва: Акад. Наук СССР, 1960, вып. 75.
Google ученый
Southward, EC, in Microscopic Anatomy of Invertebrates, vol. 12: Onychophora, Chilopoda и Lesser Prostomata , Harrison, F.W. and Rice, ME, Eds., Wiley-Liss, New York, 1993, стр. 327–369.
Google ученый
Арп, А.Дж. и Чайлдресс, Дж. Дж., Science , 1981, vol. 213, нет. 4505, стр. 342–344.
КАС Статья пабмед Google ученый
Zal, F., Lallier, F.H., Green, B.N., Vinogradov, S.N., and Toulmond, A., J. Biol. хим. , 1996, том. 271, с.8869–8874.
КАС Статья пабмед Google ученый
Развитие выделительной системы у полиплакофорного моллюска: этапы развития метанефридиальной системы | Frontiers in Zoology
Сравнение с предыдущими данными о нефрогенезе моллюсков
Предыдущие отчеты о нефрогенезе ограничены только четырьмя из восьми классов моллюсков: Gastropoda, Bivalvia, Cephalopoda и Polyplacophora.
Данные по Gastropoda скудны и значительно различаются: сообщается, что зачатки почек и перикарда парные (e.g.[21]: Patella ) или непарные ([22]: Marisa ), быть отдельными (напр.[23]: Viviparus ) или общими (напр.[22, 24]: Marisa ). Описания сходятся в том факте, что изначально твердая закладка рядом с задней кишкой дает начало почке и перикарду. У брюхоногих реноперикардиальный комплекс дифференцируется до метаморфоза [21, 25, 26].
Реноперикардиальное развитие двустворчатых моллюсков изучалось в ряде исследований, но они датируются периодом между 80 и 130 годами (e.г. [27–32]). Эти исследования можно резюмировать следующим образом: парные твердые зачатки дают начало почкам, перикарду и (позже) гонадам. Дифференцировка почки с образованием просвета предшествует дифференцировке сердца; почка становится трубчатой и соприкасается с эпидермисом и, тем самым, образует пору наружу; большая часть оставшихся первоначальных закладок дает начало предполагаемому перикарду; две стороны впоследствии сливаются в сагиттальной плоскости, образуя непарный перикард и сердце.
Головоногие демонстрируют высокопроизводный способ развития.Данные по онтогенезу реноперикарда ограничены Sepia, Loligo и Octopus [33–37], у которых развитие почки с образованием просвета отчетливо предшествует развитию перикарда. С началом дифференцировки перикарда обнаруживается связь с почкой, которая становится будущим реноперикардиальным протоком.
Информация о нефрогенезе Polyplacophora скудна. В исследовании общего развития Acanthochiton disrepans Hammarsten и Runnström [17] сообщают, что почки формируются как выросты предварительно сформированного перикарда; впоследствии они контактируют с эпидермисом и образуют нефропор.Более поздние исследования Bartolomaeus [38] и Salvini-Plawen & Bartolomaeus [13] на Lepidochitona cinerea добавили ультраструктурные детали развития перикарда и сердца. В этом исследовании наблюдения ограничены тремя стадиями (длина 250, 550 и 1000 мкм). В то время как мы можем подтвердить основной способ развития Lepidochitona (т. е. вырастание выводящего протока из перикарда при минимальной поддержке со стороны эктодермы), мы отмечаем ряд различий в деталях нефрогенеза: мкм длины тела, Bartolomaeus [38] и Salvini-Plawen & Bartolomaeus [13] описали парную закладку эпителиальных трубок перикарда слева и справа от прямой кишки с уже дифференцированными подоцитами и ресничками.Напротив, наше исследование показывает непарную массу клеток, которые дифференцируются в сторону эпителиальной полости, в которой отсутствуют ультрафильтрационные клетки (подоциты). У экземпляра с длиной тела около 1 мм Бартоломеус описал большой перикард с сердцем и каудально выходящие перикардиодукты со слепыми окончаниями в стенке тела. У наших образцов такого размера вся метанефридиальная система полностью развита и показывает нефропоры, а перикардиодукты выходят относительно далеко вперед по бокам перикарда.
е. вырастание выводящего протока из перикарда при минимальной поддержке со стороны эктодермы), мы отмечаем ряд различий в деталях нефрогенеза: мкм длины тела, Bartolomaeus [38] и Salvini-Plawen & Bartolomaeus [13] описали парную закладку эпителиальных трубок перикарда слева и справа от прямой кишки с уже дифференцированными подоцитами и ресничками.Напротив, наше исследование показывает непарную массу клеток, которые дифференцируются в сторону эпителиальной полости, в которой отсутствуют ультрафильтрационные клетки (подоциты). У экземпляра с длиной тела около 1 мм Бартоломеус описал большой перикард с сердцем и каудально выходящие перикардиодукты со слепыми окончаниями в стенке тела. У наших образцов такого размера вся метанефридиальная система полностью развита и показывает нефропоры, а перикардиодукты выходят относительно далеко вперед по бокам перикарда.
Оба вида, Lepidochitona cinereus и L. corrugata , являются близкородственными, тем не менее нельзя полностью исключить гетерохронные эффекты, т.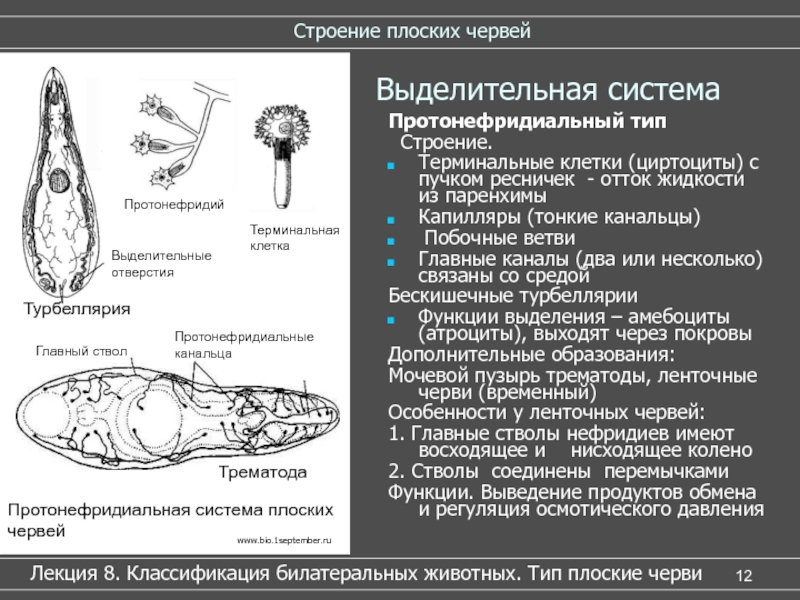 е. сдвиги во времени нефрогенеза. С другой стороны, мы основывали наше исследование на 16 образцах разных стадий нефрогенеза, тогда как в первом исследовании изучались только образцы разных стадий. Соответственно, исследования нефрогенеза других видов, в частности представителей Lepidopleurida, представляются необходимыми, чтобы продвинуться к выводу об основном образце нефрогенеза у Polyplacophora.
е. сдвиги во времени нефрогенеза. С другой стороны, мы основывали наше исследование на 16 образцах разных стадий нефрогенеза, тогда как в первом исследовании изучались только образцы разных стадий. Соответственно, исследования нефрогенеза других видов, в частности представителей Lepidopleurida, представляются необходимыми, чтобы продвинуться к выводу об основном образце нефрогенеза у Polyplacophora.
Соображения гомологии
Общие аспекты:
Вывод о гомологии всегда является вопросом вероятностей, и даже идентичный генетический фон фенотипического субъекта не решает вопрос, т.е. «гомократические» структуры;[39]. Любая гомологизация есть сравнение, и необходимо четко различать различные иерархические уровни[40, 41]. Что касается выделительной системы, следует учитывать следующие уровни; (1) Итеративная гомология ультрафильтрационных клеток, т. е. протонефридиального цитоцита с метанефридиальным подоцитом; (2) Серийная гомология личиночных протонефридиев с метанефридиальной системой взрослых особей; (3) Надвидовая гомология обеих систем у моллюсков и за их пределами. Современное понимание модульной организации генома позволяет рассматривать гомологии (сходства, основанные на общей экспрессии генов) независимо от синапоморфий (появление признаков на филогенетическом дереве).
Современное понимание модульной организации генома позволяет рассматривать гомологии (сходства, основанные на общей экспрессии генов) независимо от синапоморфий (появление признаков на филогенетическом дереве).
Прото- и метанефридиальная система в Lepidochitona corrugata :
(Развивающаяся) метанефридиальная система напоминает протонефридии [14] Lepidochitona corrugata в нескольких аспектах: (1) Обе системы органов начинаются изнутри с участков ультрафильтрации.За ними следует транспортный проток, затем модификационная часть и, наконец, эфферентный проток, который открывается через пору наружу. Нематоды или членистоногие имеют принципиально разные экскреторные структуры, так что это сходство не является тривиальным. (2) В обоих случаях эпителиальные ультрафильтрационные клетки фильтруют молекулы из первичной (гемоцель) во вторичную (эндотелиальную) полость тела. (3) тонкая структура самого ультрафильтра идентична и сложна; она состоит из извилистых щелей, образующих щели между элементами ножки, и эти щели соединены между собой тонкой диафрагмой. (4) В транспортном канале, следующем за ультрафильтрационной частью, видны плотные нижние реснички. (5) Оба органа имеют абсорбирующую часть, почку, которая имеет идентичную клеточную организацию с базальными складками, апикальной границей микроворсинок и сильно вакуолизированной цитоплазмой. Случай с почками позвоночных снова показывает, что это сходство не является тривиальным.
(4) В транспортном канале, следующем за ультрафильтрационной частью, видны плотные нижние реснички. (5) Оба органа имеют абсорбирующую часть, почку, которая имеет идентичную клеточную организацию с базальными складками, апикальной границей микроворсинок и сильно вакуолизированной цитоплазмой. Случай с почками позвоночных снова показывает, что это сходство не является тривиальным.
Эти две системы также имеют различия:
(1) Ультрафильтрация протонефридиев осуществляется терминальными клетками, тогда как в случае метанефридиев она осуществляется подоцитами.По существу, разница между этими двумя типами клеток заключается в наличии (терминальные клетки) и отсутствии (подоциты) реснитчатых клеток. Тем не менее, гомология между этими типами клеток кажется вероятной [6], так как сообщалось о промежуточных формах, например. «циртоподоциты» у amphioxus Branchiostomma и встречаются также у ранних ювенильных двустворчатых моллюсков (Ruthensteiner et al., неопубликованные). Кроме того, оба типа встречаются впоследствии в онтогенезе у некоторых полихет (например, [42]). Обнаружение реснички в подоците Lepidochitona corrugata — примечательно первое сообщение о подоците, несущем ресничку у моллюска — существенно поддерживает эту концепцию.
Кроме того, оба типа встречаются впоследствии в онтогенезе у некоторых полихет (например, [42]). Обнаружение реснички в подоците Lepidochitona corrugata — примечательно первое сообщение о подоците, несущем ресничку у моллюска — существенно поддерживает эту концепцию.
(2) В метанефридиальной системе фильтрационное давление (частично) создается сердечным сокращением, а (частично) создается реснитчатыми терминальными клетками протонефридиев. Однако это различие, по-видимому, имеет ограниченное значение, так как в обеих системах основная часть генерации отрицательного давления обеспечивается одними и теми же структурами, расположенными ниже по течению реснитчатым отделом выводного протока (см. [43, 44] для обзора метанефридиальных систем моллюсков) .
В заключение мы предполагаем, что существует существенная гомология между протонефридиальной и метанефридиальной системами у Lepidochitona , по крайней мере, на клеточном уровне.Для Lepidochitona представляется возможным, что метанефридиальная система в целом является серийным гомологом протонефридиальной системы. Однако общие данные пока фрагментарны и не позволяют окончательно прояснить этот вопрос. Например, остается неясным, являются ли абсорбирующие части (почки) протонефридиев гомологичными у моллюсков [14].
Однако общие данные пока фрагментарны и не позволяют окончательно прояснить этот вопрос. Например, остается неясным, являются ли абсорбирующие части (почки) протонефридиев гомологичными у моллюсков [14].
Прото- и метанефридиальные системы у моллюсков и родственных таксонов:
Текущее эволюционное понимание нефридиальных систем у моллюсков следующее: («голова-почки» sensu [45], «архинефридии» sensu [46] полихет) — синапоморфная система органов, по крайней мере, для Trochozoa (т.е. Entoprocta, Mollusca, Sipuncula и Annelida; сомнительно также Nemertinea) [4, 5, 9, 47] и, таким образом, плезиоморфен для Mollusca (см. также [14]).
(2) Метанефридиальная система: Сальвини-Плавен (например, [11–13, 48–50]) считал, что метанефридиальные нефропродукты («почки») развились у моллюсков на эволюционном уровне Testaria (Polyplacophora и Conchifera). Эти «эмунктории» интерпретируются как специализированные части ранее недифференцированных «перикардиодуктов», которые изначально не участвуют в экскреции. Сальвини-Плавен приводит две основные причины своей гипотезы: (а) отсутствие системы ультрафильтрации/реабсорбции (метанефридиальной) у аплакофорных моллюсков; (б) негомология с органами выделения других типов, такими как метанефридии кольчатых червей. Тем не менее, оба аргумента кажутся сомнительными: (ad a) результаты ТЭМ предполагают регулярную функцию метанефридиев также среди аплакофор: подоциты были описаны в перикарде обоих таксонов аплакофор, Solenogastres [51] и Caudofoveata [52], что свидетельствует об ультрафильтрации.Морфологические доказательства реабсорбции были обнаружены у каудофовеата Falcidens crossotus : Клетки нижних перикардиальных протоков (высвобождающие также гаметы) демонстрируют все ультраструктурные характеристики абсорбирующих клеток, включая отчетливую базальную систему складок (рис. 23 G из [53]). Соответственно, метанефридиальная система, вероятно, существует и у Solenogastres. (ad b) Salvini-Plawen (например, [12]) и Salvini-Plawen & Bartolomaeus ([13], а также [38]) также отвергли гомологию между моллюском и кольчатыми червями/сипункулановыми метанефридиями, предполагая различное происхождение зародышевого листка и другой способ формирование: в то время как почки моллюсков представляют собой мезодермальный отросток перикарда, почки кольчатых червей формируются как врастание эктодермального эпидермиса.
Сальвини-Плавен приводит две основные причины своей гипотезы: (а) отсутствие системы ультрафильтрации/реабсорбции (метанефридиальной) у аплакофорных моллюсков; (б) негомология с органами выделения других типов, такими как метанефридии кольчатых червей. Тем не менее, оба аргумента кажутся сомнительными: (ad a) результаты ТЭМ предполагают регулярную функцию метанефридиев также среди аплакофор: подоциты были описаны в перикарде обоих таксонов аплакофор, Solenogastres [51] и Caudofoveata [52], что свидетельствует об ультрафильтрации.Морфологические доказательства реабсорбции были обнаружены у каудофовеата Falcidens crossotus : Клетки нижних перикардиальных протоков (высвобождающие также гаметы) демонстрируют все ультраструктурные характеристики абсорбирующих клеток, включая отчетливую базальную систему складок (рис. 23 G из [53]). Соответственно, метанефридиальная система, вероятно, существует и у Solenogastres. (ad b) Salvini-Plawen (например, [12]) и Salvini-Plawen & Bartolomaeus ([13], а также [38]) также отвергли гомологию между моллюском и кольчатыми червями/сипункулановыми метанефридиями, предполагая различное происхождение зародышевого листка и другой способ формирование: в то время как почки моллюсков представляют собой мезодермальный отросток перикарда, почки кольчатых червей формируются как врастание эктодермального эпидермиса. Однако есть моллюски, показывающие зачаток метанефридиев без связи с перикардом [54], а также некоторые кольчатые черви (например, [55, 56]), у которых метанефридии происходят из целотелия ранее сформированных целомических карманов (см. также [57]). Таким образом, из-за вариабельности нефрогенеза как у Mollusca, так и у Annelida это рассуждение не может быть подтверждено. Также второй аргумент против гомологии моллюсков и кольчатых червей нефридиев по меньшей мере слаб: у моллюсков отсутствует воронка в начале нефропродукта, как у кольчатых червей [12].Однако воронка — это всего лишь выпячивание метанефридиального протока в объемистую целомическую полость. Опять же, кольчатые черви различаются в этом отношении, и особенно те, у которых ограниченная целомическая полость, такие как некоторые пиявки, также не имеют воронки [58].
Однако есть моллюски, показывающие зачаток метанефридиев без связи с перикардом [54], а также некоторые кольчатые черви (например, [55, 56]), у которых метанефридии происходят из целотелия ранее сформированных целомических карманов (см. также [57]). Таким образом, из-за вариабельности нефрогенеза как у Mollusca, так и у Annelida это рассуждение не может быть подтверждено. Также второй аргумент против гомологии моллюсков и кольчатых червей нефридиев по меньшей мере слаб: у моллюсков отсутствует воронка в начале нефропродукта, как у кольчатых червей [12].Однако воронка — это всего лишь выпячивание метанефридиального протока в объемистую целомическую полость. Опять же, кольчатые черви различаются в этом отношении, и особенно те, у которых ограниченная целомическая полость, такие как некоторые пиявки, также не имеют воронки [58].
Соответственно, и на основании существенного сходства в тонкой структуре, мы полагаем, что метанефридиальная система, состоящая из терминальных подоцитов на стенках перикарда (участок ультрафильтрации), реоперикардиального протока (транспорт и создание давления ресничками) и почечного протока ( «почки»: реабсорбция) принадлежит к основному типу моллюсков и гомологичен таковым других типов трохозой.
Следует признать, что база данных для этих предположений все еще скудна. Необходимы дальнейшие комплексные исследования с использованием 4-мерного подхода (3D-анализ последовательных стадий развития) нефридиальных органов большего количества таксонов моллюсков и других беспозвоночных для лучшего понимания поднятых выше вопросов гомологии и синапоморфии и, следовательно, в рамках эволюции. фильтрационных органов выделения у Metazoa.
Нефридий — обзор | Темы ScienceDirect
7.1 Анатомический фон
Хотя Cladocera обладают функционирующим органом секреции, Peters (1987, p. 219) полагает, что выделение происходит в основном через поверхность тела Cladocera: «растворимые экскременты Daphnia выделяются через общую поверхность тела. ” Орган выделения состоит из парных верхнечелюстных желез (раковинных желез), включающих нефридий и его извитые выводящие протоки (рис. 1.5 и 7.1). Верхнечелюстная железа расположена в гемоцеле и поэтому открыта для крови, под передней частью панциря и открывается наружу в передней части тела (Сгаиз, 1875а). Судя по подробным рисункам Клауса, отверстие наружу находится в пределах передней части выводкового мешка. Подробную схему строения нефридия дает Гиклхорн (1931а) (рис. 1.5).
Судя по подробным рисункам Клауса, отверстие наружу находится в пределах передней части выводкового мешка. Подробную схему строения нефридия дает Гиклхорн (1931а) (рис. 1.5).
Рисунок 7.1. Нефридиум Дафния.
C, целомический мешок; I-V, его протоки; различной окраски при прижизненном окрашивании.
Источник: Gicklhorn (1931a). Hérouard (1905) описал различные стадии усложнения верхнечелюстной железы, происходящие с возрастом, и заявил, что у Eurycercus она относительно проста.Он также отметил, что приводящие мышцы панциря прикрепляются на внутренней поверхности его извилин. Длина и расположение извилин этой железы несколько различны у разных представителей отдельных семейств (Сгаиз, 1875а, табл. XI; Нероуар, 1905). Например, у Daphnia и Sida извилины многочисленны, а у Ceriodaphnia и Moina железа короче. Он самый короткий у Macrothrix и Acroperus , но все они имеют извитой эфферентный проток.
Верхнечелюстная железа Penilia (морской вид) сильно отличается от таковой у других кладоцер (Leder, 1915): ее проток короткий, не имеет извилин, расширен в дистальной части, образуя своеобразный мочевой пузырь. мочевого пузыря (рис. 7.2).
Рисунок 7.2. Нефридий Penilia.
Ag, выводящий проток; Hb, мочевой пузырь; п, нерв; э, открытие; Вакуоли, вакуоли с жидкостью.
Источник: Leder (1915)Нефридий верхнечелюстной железы и усиковая железа (последняя не имеет канала и, вероятно, нефункциональна) — это все, что осталось от целома Cladocera.Хотя прозрачная верхнечелюстная железа различима без какой-либо обработки, целомические мешочки можно сделать хорошо видимыми с помощью прижизненной окраски (Fischel, 1908; Dejdar, 1930; Gicklhorn, 1931a, 1931c; Gicklhorn, 1931a; Gicklhorn, 1931c) нейтральным красным, метиленовым синим, нилом. голубой сульфат, или бисмарк коричневый.
Активный способ выделения через пищеварительные ткани предшествует возникновению органов выделения
Рис 2.
Экскреция у акоеломорфов.
(a) WMISH резус , v-ATPase , nka и hcn в I . pulchra и M . стихопы . (b) Скорость выделения аммиака I . pulchra до (Ctrl) и после воздействия в течение 2 часов в 50, 100, 200 и 500 мкМ и после воздействия в течение 7 дней в 1 мМ NH 4 Cl (коробчатая диаграмма). Экскрецию измеряли в течение 2 часов после обработки HEA по меньшей мере в трех независимых биологических повторах, каждый из которых был разделен на два отдельных образца (всего шесть измерений).Жирные горизонтальные полосы в прямоугольниках обозначают медиану; нижняя и верхняя границы прямоугольника обозначают нижний и верхний квартиль; и усы указывают на минимум и максимум. Звездочками отмечены значительные изменения ( p < 0,02 в непарном двустороннем тесте t с неравной дисперсией). (c) Количественная относительная экспрессия резус , нка , v-АТФазы B , amts , aq и ca после 7 дней воздействия в HEA (1 мМ) . Каждый кружок указывает среднее значение трех независимых биологических повторов, в каждом из которых по четыре технических повторения. Столбики погрешностей указывают минимум и максимум биологических повторов (усредненные технические повторы). 1-кратное изменение означает отсутствие изменений; ≥2 указывает на значительно повышенный уровень экспрессии; ≤0,5 указывает на значительное снижение уровня экспрессии (красные метки). (d) Влияние различных ингибиторов на скорость выделения аммиака в I . pulchra (блочная диаграмма, с иллюстрацией и повторениями, аналогичными рис. 2b).Используемые концентрации составляли 5 мкМ Con-C в качестве ингибитора v-АТФазы A/B, 1 мМ азетазоламида в качестве ингибитора CA, 1 мМ квабаина в качестве ингибитора NKA и 2 мМ колхицина для ингибирования сети микротрубочек. Con-C разбавляли 0,5% ДМСО, для чего мы использовали соответствующий контроль с 0,5% ДМСО. (д) Белковая локализация резуса в I . pulchra и M .
Каждый кружок указывает среднее значение трех независимых биологических повторов, в каждом из которых по четыре технических повторения. Столбики погрешностей указывают минимум и максимум биологических повторов (усредненные технические повторы). 1-кратное изменение означает отсутствие изменений; ≥2 указывает на значительно повышенный уровень экспрессии; ≤0,5 указывает на значительное снижение уровня экспрессии (красные метки). (d) Влияние различных ингибиторов на скорость выделения аммиака в I . pulchra (блочная диаграмма, с иллюстрацией и повторениями, аналогичными рис. 2b).Используемые концентрации составляли 5 мкМ Con-C в качестве ингибитора v-АТФазы A/B, 1 мМ азетазоламида в качестве ингибитора CA, 1 мМ квабаина в качестве ингибитора NKA и 2 мМ колхицина для ингибирования сети микротрубочек. Con-C разбавляли 0,5% ДМСО, для чего мы использовали соответствующий контроль с 0,5% ДМСО. (д) Белковая локализация резуса в I . pulchra и M .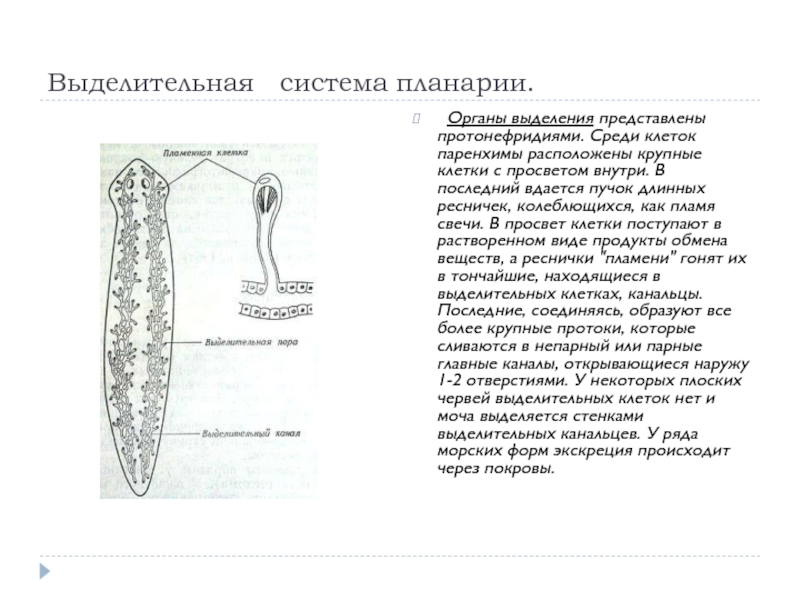 стихопы . Синцитий и кишка обозначены серым цветом, а пурпурное окрашивание просвета M . stichopi — ложноположительное окрашивание содержимого кишечника. Флуоресцентные изображения представляют собой проекции объединенных конфокальных стеков. Нервная система окрашивается в зеленый цвет тиртубулином. (f) Двойной флуоресцентный Wmish из V-ATPASE и NKA , AQ C и NKA , V-ATPASE и AQ B и V-ATPASE и RHESUS в . пульхра . Белые области на первой панели являются результатом слияния стеков, а не перекрывающихся выражений.Ядра окрашены DAPI в синий цвет. Передний слева. Шкала баров составляет 50 мкм для I . pulchra и 100 мкм для M . стихопы . Значения, лежащие в основе панелей b и d, представлены в таблице S6, а значения, лежащие в основе панели c, представлены в таблице S4. amt, переносчик аммиака; водный, аквапорин; CA, карбоангидраза; Con-C, конканамицин C; Ctrl, управление; DAPI, 4′,6-диамидино-2-фенилиндол; ds, пищеварительный синцитий; gwc, обволакивающая кишечник клетка; HCN, K + [Nh5 + ] канал; HEA, высокое содержание аммиака в окружающей среде; NKA, Na + /K + [Nh5 + ] АТФаза; Rh, гликопротеин резуса; slc, переносчик растворенных веществ; тир, тирозинированный; v-АТФаза, вакуолярная H + -АТФаза протонный насос; WMISH, полная гибридизация in situ.
стихопы . Синцитий и кишка обозначены серым цветом, а пурпурное окрашивание просвета M . stichopi — ложноположительное окрашивание содержимого кишечника. Флуоресцентные изображения представляют собой проекции объединенных конфокальных стеков. Нервная система окрашивается в зеленый цвет тиртубулином. (f) Двойной флуоресцентный Wmish из V-ATPASE и NKA , AQ C и NKA , V-ATPASE и AQ B и V-ATPASE и RHESUS в . пульхра . Белые области на первой панели являются результатом слияния стеков, а не перекрывающихся выражений.Ядра окрашены DAPI в синий цвет. Передний слева. Шкала баров составляет 50 мкм для I . pulchra и 100 мкм для M . стихопы . Значения, лежащие в основе панелей b и d, представлены в таблице S6, а значения, лежащие в основе панели c, представлены в таблице S4. amt, переносчик аммиака; водный, аквапорин; CA, карбоангидраза; Con-C, конканамицин C; Ctrl, управление; DAPI, 4′,6-диамидино-2-фенилиндол; ds, пищеварительный синцитий; gwc, обволакивающая кишечник клетка; HCN, K + [Nh5 + ] канал; HEA, высокое содержание аммиака в окружающей среде; NKA, Na + /K + [Nh5 + ] АТФаза; Rh, гликопротеин резуса; slc, переносчик растворенных веществ; тир, тирозинированный; v-АТФаза, вакуолярная H + -АТФаза протонный насос; WMISH, полная гибридизация in situ.
Подробнее »
Глава 44
Глава 44 ГЛАВА 44УПРАВЛЕНИЕ
ВНУТРЕННЯЯ СРЕДА
Авторское право © 1999 г. The Бенджамин/Каммингс Паблишинг Компани, Инк.
ЭЛЕКТРОННАЯ ДЕЯТЕЛЬНОСТЬ (IOSU: В порядке
Скорость и полезность)
1. «Интерактивное исследование
Partner» CD ROM Quiz (или веб-сайт Campbell http://occ.awlonline.com/bookbind/pubbooks/campbell_awl/chapter44/deluxe.HTML)
2. «Интерактивное исследование
Диск «Партнер»
А. 44.1 Структура выделительной системы
B. 44.2 Функция нефрона
C. 44.3 Контроль реабсорбции воды
3. Биология
Разместите индивидуальную викторину «TestFlight».
4. Кэмпбелл (http://www.biology.com/campbell)
Веб-адреса (http://occ.awlonline.com/bookbind/pubbooks/campbell_awl/chapter44/deluxe.html)
5.Место биологии (http://www.biology.com/home/home.html)
Выберите главу 44, затем нажмите «Перейти», чтобы найти текущие действия.
КОНТУР
I. Регуляция температуры тела
A. Учет четырех физических процессов
для притока или потери тепла
B. Эктотермы в основном выделяют тепло тела
из своего окружения; эндотермы получают его в основном за счет метаболизма
C. Терморегуляция включает физиологическую
и поведенческие корректировки
Д.Большинство животных экзотермичны, но
эндотермия широко распространена
E. Оцепенение сохраняет энергию во время
крайности окружающей среды
II. Водный баланс и утилизация отходов
A. Водный баланс и удаление отходов
зависит от транспортного эпителия
B. Азотистые выделения животных
коррелируют с его филогении и средой обитания
C. Ячейкам требуется баланс между
осмотическое приращение и потеря воды
D. Осморегуляторы расходуют энергию на
контролировать их внутреннюю осмолярность; осмоконформеры изоосмотичны
окрестности
III.Выделительные системы
A.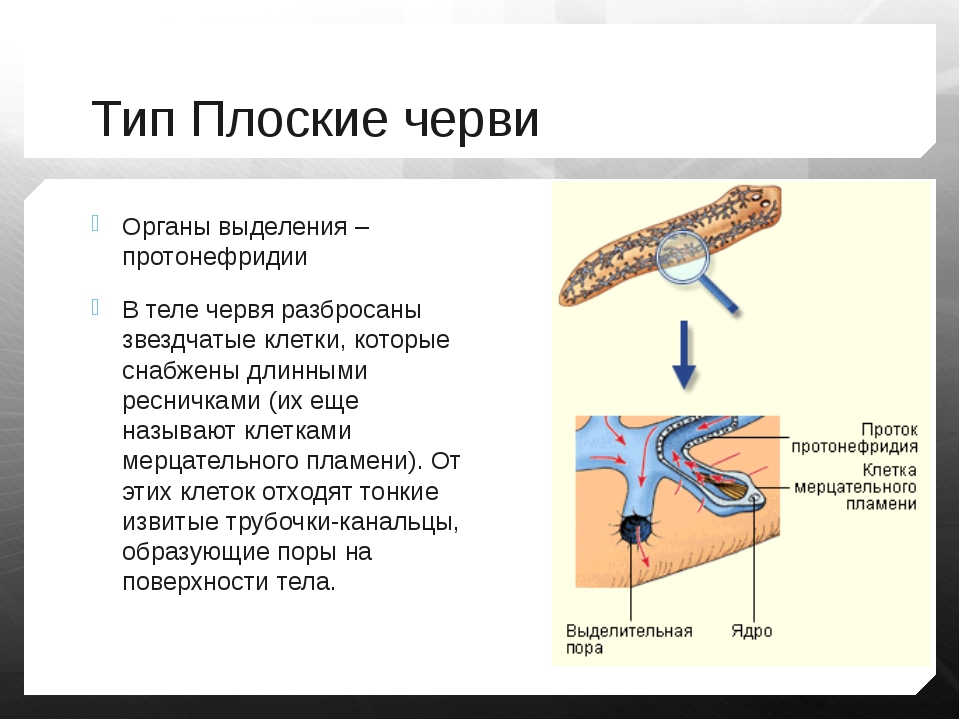 Большинство выделительных систем производят
моча путем очистки фильтрата, полученного из биологических жидкостей: обзор
Большинство выделительных систем производят
моча путем очистки фильтрата, полученного из биологических жидкостей: обзор
B. Различные выделительные системы являются вариациями
на трубчатую тему
C. Нефроны и связанные с ними кровеносные сосуды
являются функциональными единицами почек млекопитающих.
D. Из фильтрата крови в мочу: a
пристальный взгляд
E. Способность почек млекопитающих
сохранение воды является ключевой наземной адаптацией
Ф.Нервные и гормональные цепи обратной связи
регулировать
функции почек
G. Разнообразные приспособления позвоночных
почки развились
H. Взаимодействующие регуляторные системы
поддерживать гомеостаз
ЦЕЛИ
Прочитав эту главу и посетив лекцию, вы
должны быть в состоянии:
1. Различают осморегуляторы и осмоконформеры.
2. Обсудите проблемы, с которыми сталкиваются морские организмы, пресноводные
организмы, и наземные организмы сталкиваются в поддержании гомеостаза, и
объясните, какие осморегуляторные приспособления служат решению этих проблем.
3. Объясните роль транспортных эпителиев в осморегуляции.
4. Опишите, как пламенная (протонефридиальная) экскреторная
системные функции.
5. Объясните, как метанефридиальный выводной каналец
функции кольчатых червей и описывают любые структурные улучшения по сравнению с протонефридиальными
система.
6. Объясните, как выделительная система мальпигиевых канальцев
способствовал успеху насекомых в наземной среде.
7. Используя схему, определите и укажите функцию
каждая структура выделительной системы млекопитающих.
8. Используя схему, определите и укажите функцию
каждой части нефрона.
9. Описать и показать взаимосвязь между процессами
фильтрации, секреции и реабсорбции.
10. Объясните значение того факта, что юкстамедуллярный
нефроны есть только у птиц и млекопитающих.
11. Объясните, как петля Генле способствует сохранению воды.
почкой.
12. Опишите механизмы гормонального
регуляция почек.
13. Описать структурные и физиологические адаптации
в почках немлекопитающих, что позволяет им осморегулируть
в разных средах.
14. Объясните взаимосвязь между типом азотистых
отходы, производимые (аммиак, мочевина или мочевая кислота) организмом и средой его обитания.
15. Опишите адаптивные преимущества эндотермии.
16. Обсудите четыре основные категории физиологических
и поведенческие приспособления, используемые наземными млекопитающими для поддержания относительно
постоянная температура тела.
17. Различать два терморегуляторных центра
гипоталамуса.
18. Опишите терморегуляторные приспособления, обнаруженные у
животных, кроме наземных млекопитающих.
19. Опишите несколько механизмов физиологической акклиматизации
к новым диапазонам температур.
20. Различайте спячку и сон.
ОСНОВНЫЕ ТЕРМИНЫ
акклиматизация
приносящая артериола
альдостерон
аммиак
ангиотензин II
ангидробиоз
антидиуретический гормон
предсердный натрийуретический фактор
Капсула Боумена
бурый жир
собирательный канал
проводимость
конвекция
кортикальные нефроны
Противоточный теплообменник
дистальный каналец
экзотерм
выносящая артериола
эндотерм
оценка
эвригалин
испарение
экскреция
фильтрация
клубочек
белка теплового шока
спящий режим
юкстагломерулярный аппарат
юкстамедуллярные нефроны
петля Генле
Мальпигиевы канальцы
метанефридий
нефрон
недрожательный термогенез
осмоконформер
осмолярность
осморегуляция
перитубулярные капилляры
подоцитов
протонефридий
проксимальный каналец
излучение
реабсорбция
почечная артерия
корковое вещество почки
мозговое вещество почки
почечная вена
ренин
ренин-ангиотензин-альдостероновая система
секреция
стеногалин
стресс-индуцированных белка
терморегуляция
оцепенение
транспортный эпителий
мочеточник
уретра
мочевая кислота
мочевой пузырь
прямые сосуды
сосудосуживающий осморегулятор
расширение сосудов
| Тип публикации | диссертация |
| Школа или колледж | Школа медицины |
| Отдел | Нейробиология и анатомия |
| Автор | Ву, Хан Тхи Ким |
| Титул | Дивергентная эволюция выделительных систем животных: данные молекулярных и функциональных исследований планарий |
| Дата | 2015-05 |
| Описание |  For instance, with respect to salt and water homeostasis, freshwater animals continuously eliminate excess water while conserving solutes, whereas land-dwelling organisms have to conserve water and solutes as much as possible. Comparative morphological studies suggest that animals have tackled the problems of excretion and osmoregulation by evolving a specialized structure: the excretory organ. Animal excretory organs are extremely diverse. Some are unicellular, such as the excretory cell in nematodes. Others are multicellular and highly specialized, such as the protonephridia/metanephridia in invertebrates or the kidneys in vertebrates. In light of such anatomical and functional diversity, the evolutionary origins of animal excretory systems pose an interesting question in biology. However, the hypotheses proposed thus far remain highly controversial for two main reasons. First, many evolutionary arguments are based solely on morphology in organisms for which no molecular data are available, precluding rigorous genetic comparisons. For instance, with respect to salt and water homeostasis, freshwater animals continuously eliminate excess water while conserving solutes, whereas land-dwelling organisms have to conserve water and solutes as much as possible. Comparative morphological studies suggest that animals have tackled the problems of excretion and osmoregulation by evolving a specialized structure: the excretory organ. Animal excretory organs are extremely diverse. Some are unicellular, such as the excretory cell in nematodes. Others are multicellular and highly specialized, such as the protonephridia/metanephridia in invertebrates or the kidneys in vertebrates. In light of such anatomical and functional diversity, the evolutionary origins of animal excretory systems pose an interesting question in biology. However, the hypotheses proposed thus far remain highly controversial for two main reasons. First, many evolutionary arguments are based solely on morphology in organisms for which no molecular data are available, precluding rigorous genetic comparisons. Second, while invertebrates are critical elements of this evolutionary puzzle, the molecularly tractable ones studied to date display highly derived excretory systems. C. elegans possesses a single excretory cell, while the ultrafiltration of nephrocytes is uncoupled from the absorption/secretion of Malpighian tubules in D. melanogaster. Comparative morphological studies have demonstrated the existence of more complex excretory organs amongst many other invertebrates, including planarians. Planarians have a protonephridial excretory system in which each protonephridial unit consists of a tubule, opening distally via a nephridiopore at the surface of the animal and ending proximally in one or more terminal structures called flame cells. Protonephridia are commonly found amongst many invertebrates. Since protonephridia combine ultrafiltration with filtrate modification, planarians close an "invertebrate gap" in the study of excretory system biology. Taking advantage of a rapidly expanding list of molecular tools in recent years, this dissertation project aims to perform a comprehensive molecular and functional study of planarian protonephridia in order to provide new insights into the longstanding question on the evolutionary relationship between vertebrate and invertebrate excretory systems and gauge planarians’ potential as a novel invertebrate model for human kidney development and disease. Second, while invertebrates are critical elements of this evolutionary puzzle, the molecularly tractable ones studied to date display highly derived excretory systems. C. elegans possesses a single excretory cell, while the ultrafiltration of nephrocytes is uncoupled from the absorption/secretion of Malpighian tubules in D. melanogaster. Comparative morphological studies have demonstrated the existence of more complex excretory organs amongst many other invertebrates, including planarians. Planarians have a protonephridial excretory system in which each protonephridial unit consists of a tubule, opening distally via a nephridiopore at the surface of the animal and ending proximally in one or more terminal structures called flame cells. Protonephridia are commonly found amongst many invertebrates. Since protonephridia combine ultrafiltration with filtrate modification, planarians close an "invertebrate gap" in the study of excretory system biology. Taking advantage of a rapidly expanding list of molecular tools in recent years, this dissertation project aims to perform a comprehensive molecular and functional study of planarian protonephridia in order to provide new insights into the longstanding question on the evolutionary relationship between vertebrate and invertebrate excretory systems and gauge planarians’ potential as a novel invertebrate model for human kidney development and disease. «> Животные развили исключительную способность поддерживать гомеостаз перед лицом серьезных осморегуляторных проблем со стороны окружающей среды.Например, что касается гомеостаза соли и воды, пресноводные животные постоянно выделяют избыток воды, сохраняя при этом растворенные вещества, тогда как наземные организмы должны максимально сохранять воду и растворенные вещества. Сравнительные морфологические исследования показывают, что животные решили проблемы экскреции и осморегуляции, развив специализированную структуру: орган выделения. Органы выделения животных чрезвычайно разнообразны. Некоторые из них одноклеточные, например экскреторная клетка нематод.Другие являются многоклеточными и узкоспециализированными, например, протонефридии/метанефридии у беспозвоночных или почки у позвоночных. В свете такого анатомического и функционального разнообразия эволюционное происхождение выделительной системы животных ставит перед биологией интересный вопрос. Однако гипотезы, предложенные до сих пор, остаются весьма спорными по двум основным причинам. «> Животные развили исключительную способность поддерживать гомеостаз перед лицом серьезных осморегуляторных проблем со стороны окружающей среды.Например, что касается гомеостаза соли и воды, пресноводные животные постоянно выделяют избыток воды, сохраняя при этом растворенные вещества, тогда как наземные организмы должны максимально сохранять воду и растворенные вещества. Сравнительные морфологические исследования показывают, что животные решили проблемы экскреции и осморегуляции, развив специализированную структуру: орган выделения. Органы выделения животных чрезвычайно разнообразны. Некоторые из них одноклеточные, например экскреторная клетка нематод.Другие являются многоклеточными и узкоспециализированными, например, протонефридии/метанефридии у беспозвоночных или почки у позвоночных. В свете такого анатомического и функционального разнообразия эволюционное происхождение выделительной системы животных ставит перед биологией интересный вопрос. Однако гипотезы, предложенные до сих пор, остаются весьма спорными по двум основным причинам. Во-первых, многие эволюционные аргументы основаны исключительно на морфологии организмов, для которых нет доступных молекулярных данных, что исключает строгое генетическое сравнение.Во-вторых, в то время как беспозвоночные являются важнейшими элементами этой эволюционной головоломки, изученные к настоящему времени молекулярно-послушные животные демонстрируют высокоразвитые выделительные системы. C. elegans обладает одной экскреторной клеткой, в то время как ультрафильтрация нефроцитов не связана с абсорбцией/секрецией мальпигиевых канальцев у D. melanogaster. Сравнительно-морфологические исследования показали наличие более сложных органов выделения у многих других беспозвоночных, в том числе у планарий. Планарии имеют протонефридиальную выделительную систему, в которой каждая протонефридиальная единица состоит из канальца, открывающегося дистально через нефридиопор на поверхности животного и оканчивающегося проксимально в одной или нескольких концевых структурах, называемых пламенными клетками. Во-первых, многие эволюционные аргументы основаны исключительно на морфологии организмов, для которых нет доступных молекулярных данных, что исключает строгое генетическое сравнение.Во-вторых, в то время как беспозвоночные являются важнейшими элементами этой эволюционной головоломки, изученные к настоящему времени молекулярно-послушные животные демонстрируют высокоразвитые выделительные системы. C. elegans обладает одной экскреторной клеткой, в то время как ультрафильтрация нефроцитов не связана с абсорбцией/секрецией мальпигиевых канальцев у D. melanogaster. Сравнительно-морфологические исследования показали наличие более сложных органов выделения у многих других беспозвоночных, в том числе у планарий. Планарии имеют протонефридиальную выделительную систему, в которой каждая протонефридиальная единица состоит из канальца, открывающегося дистально через нефридиопор на поверхности животного и оканчивающегося проксимально в одной или нескольких концевых структурах, называемых пламенными клетками.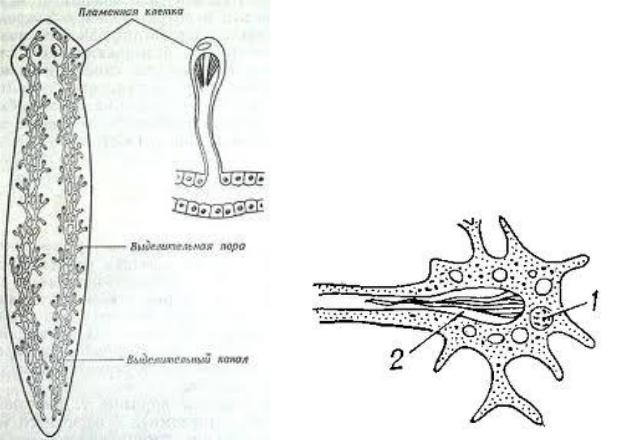 Протонефридии обычно встречаются среди многих беспозвоночных. Поскольку протонефридии сочетают ультрафильтрацию с модификацией фильтрата, планарии закрывают «беспозвоночный пробел» в изучении биологии выделительной системы. Используя в последние годы быстро расширяющийся список молекулярных инструментов, этот проект диссертации направлен на проведение всестороннего молекулярного и функционального исследования протонефридиев планарии, чтобы дать новый взгляд на давний вопрос об эволюционных отношениях между выделительной системой позвоночных и беспозвоночных. оценить потенциал планарий как новой модели беспозвоночных для развития и заболеваний почек человека. Протонефридии обычно встречаются среди многих беспозвоночных. Поскольку протонефридии сочетают ультрафильтрацию с модификацией фильтрата, планарии закрывают «беспозвоночный пробел» в изучении биологии выделительной системы. Используя в последние годы быстро расширяющийся список молекулярных инструментов, этот проект диссертации направлен на проведение всестороннего молекулярного и функционального исследования протонефридиев планарии, чтобы дать новый взгляд на давний вопрос об эволюционных отношениях между выделительной системой позвоночных и беспозвоночных. оценить потенциал планарий как новой модели беспозвоночных для развития и заболеваний почек человека. |
| Тип | Текст |
| Издатель | Университет Юты |
| Тема MESH | планарии; Caenorhabditis elegans; биологическая эволюция; Морфогенез; биологические процессы; физиологические процессы; Распространение клеток; беспозвоночные; мальпигиевы канальцы; нефроны; почечные канальцы; Гомеостаз; Генная частота; Осморегуляция; водно-электролитный баланс; ультрафильтрация; почечная ликвидация; Заболевания почек, кистозные; Барьер клубочковой фильтрации; Реснички; интерференция РНК; Гибридизация на месте; Гомология последовательности |
| Диссертационное учреждение | Университет Юты |
| Название диссертации | Доктор философии |
| Язык | англ |
| Отношение — это версия | Цифровая репродукция книги «Дивергентная эволюция выделительных систем животных: данные молекулярных и функциональных исследований планарий» |
| Управление правами | Copyright © Хан Тхи Ким Ву 2015 |
| Носитель формата | заявка/pdf |
| Формат Экстента | 9 449 772 байта |
| Источник | 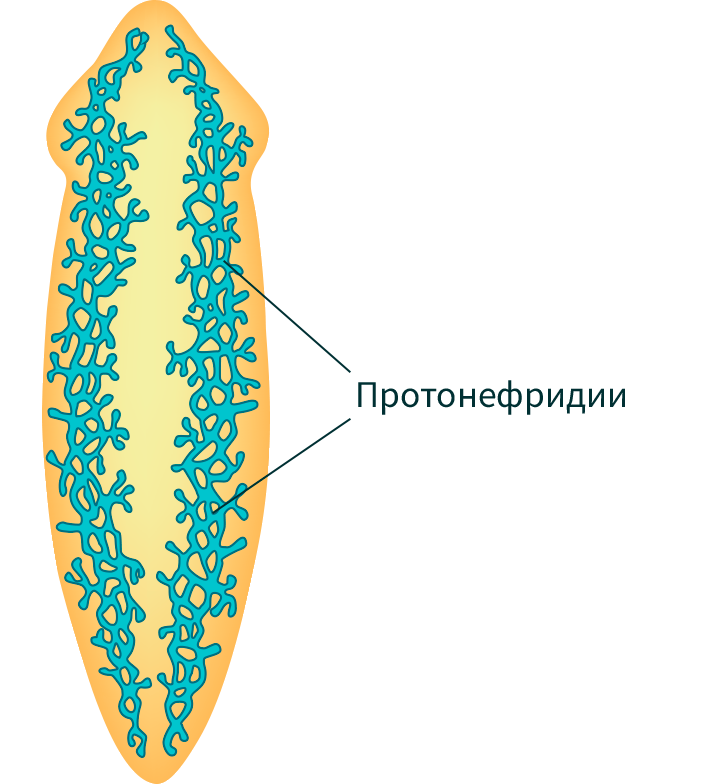 «> Оригинал из специальных коллекций Marriott Library. «> Оригинал из специальных коллекций Marriott Library. |
| КОВЧЕГ | ковчег:/87278/s6rc0cwb |
| Имя набора | ir_etd |
| Дата создания | 14.04.2016 |
| Дата изменения | 06.05.2021 |
| ID | 197345 |
| Ссылочный URL-адрес | https://collections.lib.utah.edu/ark:/87278/s6rc0cwb |
В чем разница между протонефридией и метанефридией
Основное различие между протонефридиями и метанефридиями заключается в том, что протонефридии представляют собой сеть тупиковых канальцев без внутренних отверстий, тогда как метанефридии представляют собой разновидность экскреторных желез с реснитчатой воронкой, открывающейся в полость тела.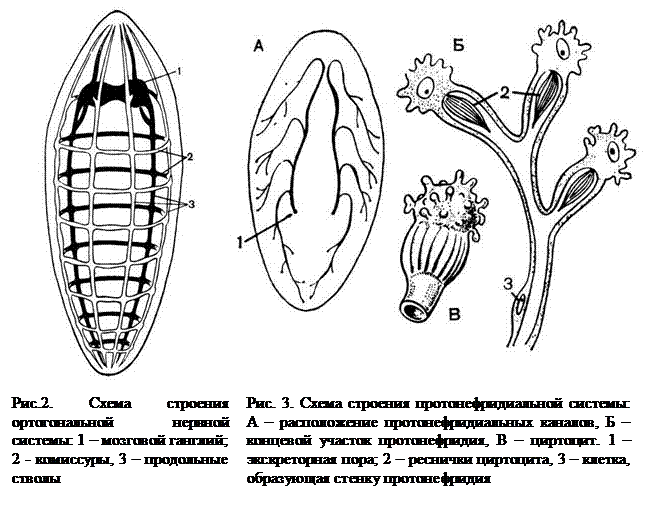 . Кроме того, протонефридии встречаются в типах Platyhelminthes, Nemertea, Rotifera и Chordata (ланцетники), а метанефридии встречаются в типах Annelida, Arthropoda и Mollusca.
. Кроме того, протонефридии встречаются в типах Platyhelminthes, Nemertea, Rotifera и Chordata (ланцетники), а метанефридии встречаются в типах Annelida, Arthropoda и Mollusca.
Протонефридии и метанефридии — это два типа нефридиев, которые в основном встречаются у беспозвоночных. Как правило, они функционально аналогичны почкам позвоночных и отвечают за удаление метаболических отходов из организма.
Ключевые области охвата
1. Что такое протонефридии
– определение, структура, значение
2. Что такое метанхридия
9069 — определение, структура, важность
3. Что такое сходство между протонафридией и метанфридией
— Схема распространенных особенностей
4. Что такое разница между протонамидийными и метанфидиями
9069 – Сравнение ключевых отличий
Ключевые термины
Акоеломаты, Целоматы, Метанефридии, Нефридии, Протонефридии
Что такое протонефридии
Протонефридии — это тип нефридиев, которые встречаются у беспозвоночных, включая типы Platyhelminthes, Nemertea, Rotifera и Chordata (ланцетники). Однако протонефридии являются гомологичными органами у билатеральных. Основная характерная черта протонефридиев состоит в том, что они закрыты пламенной клеткой. Поэтому они не открываются в полость тела. По сути, они могут выглядеть как пара моноресничных органов, происходящих из эктодермы. Кроме того, каждый протонефридий содержит единственный терминальный проток, а также клетку нефрона.
Однако протонефридии являются гомологичными органами у билатеральных. Основная характерная черта протонефридиев состоит в том, что они закрыты пламенной клеткой. Поэтому они не открываются в полость тела. По сути, они могут выглядеть как пара моноресничных органов, происходящих из эктодермы. Кроме того, каждый протонефридий содержит единственный терминальный проток, а также клетку нефрона.
Рисунок 1: Протонефидий
Кроме того, терминальной клеткой протонефридии является пламенная клетка.Хотя клетка пламени имеет только один корешок, она окружена восемью длинными и сильными микроворсинками. Напротив, клетка пламени соединяется с соседней клеткой протока десмосомами.
Метанефридии — это другой тип нефридиев, которые встречаются у беспозвоночных, включая типы Annelida, Arthropoda и Mollusca. Обычно метанефридии представляют собой реснитчатую воронку, открывающуюся в полость тела. Это отверстие соединяется с воздуховодом, который складывается или расширяется. После этого воздуховод открывается наружу.Более того, основная функция этих реснитчатых канальцев состоит в том, чтобы направлять к воронке избыток ионов, токсинов, бесполезных гормонов и метаболических отходов.
После этого воздуховод открывается наружу.Более того, основная функция этих реснитчатых канальцев состоит в том, чтобы направлять к воронке избыток ионов, токсинов, бесполезных гормонов и метаболических отходов.
Рисунок 2: Метанефридий
Кроме того, мешковидные нефридии представляют собой тип метанефридиев, встречающихся у членистоногих: тазовые железы паукообразных, усики (или зеленые) железы и верхнечелюстные железы ракообразных и т. д. Основная функция метанефридиев этого типа заключается в фильтрации жидкости гемоцель.
- Протонефридии и метанефридии представляют собой два типа нефридиев, представляющих собой выделительные железы беспозвоночных.
- Оба встречаются у билатеральных.
- Более того, они встречаются парами.

- Морфологически аналогичны почкам позвоночных.
- Основной функцией нефридии является удаление метаболических отходов.
- Кроме того, они сначала фильтруют, а затем модифицируют жидкости организма.
Определение
Протонефридии относятся к трубчатым экскреторным структурам у некоторых беспозвоночных, обычно оканчивающимся внутри пламенных клеток и имеющим внешнюю пору, в то время как метанефридии относятся к примитивным экскреторным органам, присутствующим у многих беспозвоночных и происходящим из реснитчатой целомической воронки.Таким образом, в этом основное отличие протонефридии от метанефридии.
Структура
В то время как протонефридии представляют собой сеть тупиковых канальцев без внутренних отверстий, метанефридии представляют собой тип экскреторных желез с реснитчатой воронкой, открывающейся в полость тела.
Пламенная ячейка
Кроме того, еще одно важное различие между протонефридиями и метанефридиями заключается в том, что протонефридии закрыты пламенной клеткой, тогда как метанефридии имеют отверстие в полость тела и не закрыты пламенными клетками.
Возникновение
Кроме того, протонефридии встречаются в типах Platyhelminthes, Nemertea, Rotifera и Chordata (ланцетники), а метанефридии встречаются в типах Annelida, Arthropoda и Mollusca.
Целоматы и акоеломаты
Более того, протонефридии встречаются как у целоматов, так и у акоеломатов, тогда как метанефридии встречаются у целоматов.
Заключение
Протонефридии — это тип примитивного выделительного органа, встречающийся у Platyhelminthes, немертин, коловраток и ланцетников.Более того, это сеть тупиковых канальцев. Кроме того, он не содержит внутреннего отверстия. С другой стороны, метанефридии представляют собой еще один тип примитивных органов выделения, которые встречаются у кольчатых червей, членистоногих и моллюсков.
