
Ученые нашли в подледном озере Восток признаки жизни — Российская газета
Американские ученые сделали сенсационное открытие, которое позволяет предположить, что в знаменитом подледном озере Восток могут обитать сложные организмы.
Анализ проб озерного льда выявил наличие 3 507 уникальных последовательностей генов на 500 мл пресной воды. Примерно 94 процента генетического материала относится к роду Bacteria, а остальные 6 процентов — к более сложным эукариотам (одно-или многоклеточным организмам, клетки которых содержат ядра). При этом два процента фрагментов РНК совпали с археями, группой микроскопических одноклеточных организмов. Об этом говорится в отчете, опубликованном в журнале PLoS.
По словам ученых, многие виды бактерий, генетические последовательности которых совпали с обнаруженными в пробах льда, нередко сопутствуют моллюскам, ракообразным и рыбам.
Находка породила гипотезу, что в озере Восток могут обитать не только бактерии, но и сложные организмы, сообщает NEWSru.
Правда, откуда взялись бактерии, еще предстоит выяснить. Известно, что под толщей ледников расположена густая сеть рек. А выявленные организмы могли просто попасть в озеро из других подледных водоемов или из океана.
Озеро Восток скрыто под ледяным панцирем Антарктики толщиной почти в 4 км. Оно уникально прежде всего тем, что, возможно, находилось в изоляции от земной поверхности на протяжении нескольких миллионов лет. Восток считается самым крупным подледным озером Антарктиды: его длина составляет 230 км, глубина — около 1200 метров, а площадь — более 15 тысяч квадратных километров.
Озеро обладает необычными характеристиками. Например, известно, что концентрация кислорода в Востоке в 50 раз выше, чем в обычной пресной воде. А давление воды здесь, согласно расчетам, составляет более 300 атмосфер. Все это дает ученым право предположить, что микроорганизмы, приспособленные к жизни в таких удивительных условиях, изолированные от земной биосферы, могут обладать уникальными свойствами.
Почему 47 лет — самый несчастный возраст и кто хранит тайну многоклеточных
- Леонид Лунеев
- Би-би-си
В очередной подборке интересных научных новостей недели:
Вышли мы все из эукариотов
Автор фото, Thinkstock
Познакомьтесь: этот крошечный микроб, обнаруженный глубоко на дне Тихого океана — ваш дальний предок. Да, выглядит он невзрачно, но он может хранить тайну возникновения первых многоклеточных структур, а значит — и нас с вами.
Задолго до появления многоклеточных организмов миром правили простые одноклеточные бактерии и археи. Простые-то простые, но около 2 млрд лет назад они решили, что момент настал, и начали эволюционировать, создав домен эукариотов — живых организмов, клетки которых содержат ядро.
Но самое интересное кроется именно в процессе образования из этого биологического супа растений и животных, в том числе и мыслящих, а процесс этот изучен до обидного мало.
В попытке хотя бы отчасти воспроизвести этот процесс, ученые из Японии извлекли из донных пород Архею Асгарда, предка современных эукариотов, гены которого при этом похожи на гены более развитых организмов.
Микроорганизмы были помещены в биореактор, и долгие 10 лет исследователи наблюдали за их поведением. В итоге они неожиданно для себя вывели новый штамм архей Prometheoarchaeum syntrophicum, названный в честь греческого бога Прометея, по легенде сотворившего людей из глины.
Дальнейшие анализы подтвердили, что P. syntrophicum обладают генным набором, напоминающим гены эукариотов, содержащим инструкции по строительству белков, только вот белки эти, в отличие от эукариотов, не создавали органеллы вроде митохондрий.
Зато у новых микробов появились напоминающие щупальца отростки, которыми они успешно ловили находящиеся поблизости бактерии.
В результате авторы исследования выдвинули гипотезу, согласно которой нашему микробу, привыкшему обходиться без кислорода, очень не понравилось, когда около 2,7 млрд лет на планете стала расти кислородная атмосфера.
Ему пришлось приспосабливаться и для этого вступить в связь с бактериями, которые хорошо переносили кислород. Эти бактерии давали P. syntrophicum витамины и питательные вещества, а сами пользовались отходами его жизнедеятельности.
По мере повышения уровня кислорода P. syntrophicum могли стать более агрессивными и начать захватывать своими щупальцами находившиеся поблизости бактерии и затягивать их внутрь.
Постепенно они могли научиться переделывать эти захваченные бактерии в жизненно важные для всех эукариотов митохондрии — подобие органов, вырабатывающих энергию. Ну а дальше до человека разумного было уже рукой подать…
Как армия США намерена защитить Нью-Йорк от наводнений
Автор фото, Reuters
Подпись к фото,На восточное побережье США, включая Нью-Йорк, часто обрушиваются мощные ураганы
Наверняка все вы видели хотя бы отрывки из апокалиптических фильмов о недалеком будущем, когда гигантская волна обрушивается на Нью-Йорк, сметая небоскребы как спичечные коробки.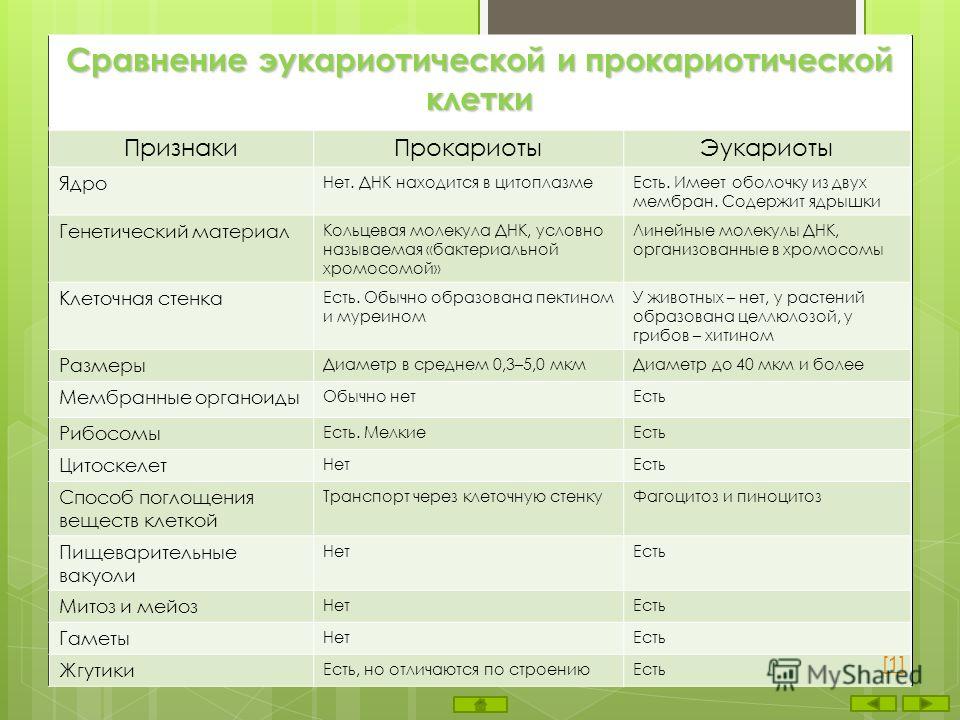
Казалось бы, кто всерьез будет рассматривать такой сценарий? Оказывается, весьма серьезная организация — американская армия.
Командование инженерных войск США изучает сразу пять возможных вариантов защиты береговой линии Нью-Йорка от необычно высоких приливных волн. Но самый амбициозный проект — это гигантская стена стоимостью 119 млрд долларов, которую планируется построить за 25 лет.
Не удивительно, что этот проект навлек критику со стороны американского президента, который не особо верит в угрозу климатических изменений.
«Огромная стена стоимостью в 200 млрд долларов, построенная вокруг Нью-Йорка для защиты от случайного шторма — это дорогая, глупая и неэкологичная затея, которая, быть может, и не сработает вовсе, — написал он, как обычно, в «Твиттере». — К тому же она будет выглядеть ужасно, так что лучше готовьте ведра и швабры».
Как ни странно, этот проект, получивший название «Альтернатива-2», пока не получил поддержки и у местных жителей, которые опасаются, что барьер сможет защитить город лишь от штормов, но не от поднимающегося уровня моря.
«Эти морские ворота не смогут защитить людей от наводнений, вызванных приливными волнами и подъемом уровня моря, а когда их построят, на этом все закончится, нам таких денег больше никто не даст», — заявил газете New York Times ревизор Совета Нью-Йорка Скотт Стрингер.
Критики проекта говорят также, что гигантский волнорез создаст собственные проблемы, как это происходило уже во многих портовых городах мира, в том числе с затруднением оттока канализации.
«Мы, по сути, будем сидеть в ванной с собственными экскрементами», — прямолинейно заявила газете Times представительница одной из экологических организаций старший прокурор Кимберли Онг.
Ожидается, что окончательное решение по этому вопросу будет принято в 2022 году, когда военные представят расширенный доклад на рассмотрение американскому конгрессу.
Но уже сейчас ясно одно: если ничего не предпринимать, то ничего хорошего для Нью-Йорка и других прибрежных мегаполисов из этого не выйдет.
В своем отчете Стрингер приводит научные выкладки, согласно которым к 2050 году уровень моря в районе Нью-Йорка поднимется на 53 сантиметра.
Любопытно, что в эти климатические прогнозы верит даже Дональд Трамп, иначе зачем бы он тогда отдал распоряжение возвести защитные сооружения в прибрежном гольф-парке в Ирландии, которым он владеет.
Куда вывозит нас кривая счастья
Автор фото, Getty Images
Говорят, в 40 лет жизнь только начинается. Но это ненадолго, успокаивают ученые, потому что уже в 47 по статистике у вас будет пик кризиса среднего возраста.
Согласно отчету американского Национального бюро экономических исследований (NBER), нижняя точка так называемой кривой счастья человеческой жизни приходится примерно на 47 лет в развитых и на 48 — в развивающихся странах.
По словам автора исследования профессора Дартмутского колледжа в Ганновере Дэвида Бланчфлауэра, изучившего данные по 132 странам, эта тенденция прослеживается вне зависимости от пола, образования, семейного положения или социального статуса: кривая счастья присутствует везде.
В своей работе профессор Бланчфлауэр использовал около 15 различных мерил несчастия, включая боль, фобии, отчаяние, одиночество, стресс и недосып.
Более того, Бланчфлауэр указывает на результаты исследования 2012 года, выявившие сходную кривую у орангутангов и шимпанзе.
«На этом основании мы можем сделать вывод, что кривая счастья — это не уникальная человеческая черта, и хотя частично ее можно объяснить особенностями человеческого общества и жизни, корни ее следует искать в биологии приматов», — подозревает профессор, предполагая даже, что наличие этой кривой в жизненном цикле запрограммировано где-то на генном уровне.
Главный же вывод этого исследования, по мнению Бланчфлауэра, заключается в том, что после самой низкой точки жизнь начинает налаживаться.
«Если вы переживаете кризис среднего возраста, полезно понимать, что вы не один такой, что это происходит со многими, — делится советом профессор. — И еще, старайтесь не оставаться в одиночестве, общайтесь с соседями, с семьей, проводите больше времени с близкими и не мешайте людям помогать вам.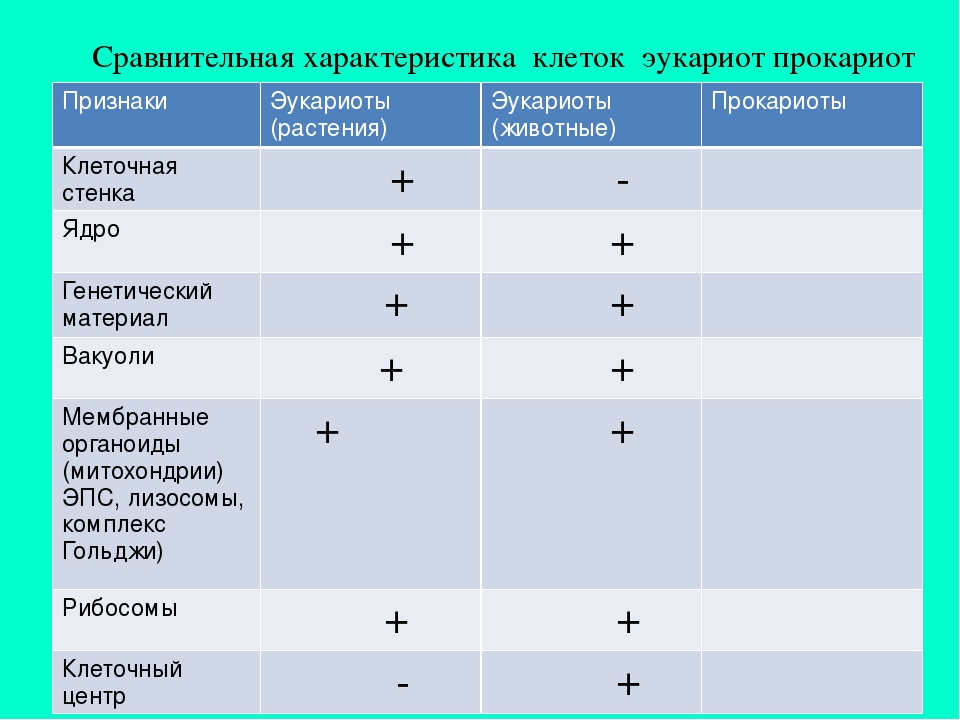 Тогда вам станет легче».
Тогда вам станет легче».
Дышите глубже: как добыть кислород из лунной пыли
Автор фото, Getty Images
Подпись к фото,Возможно, в будущем человеческая активность на Луне будет выглядеть так
Хотя у Луны нет атмосферы, она богата кислородом, просто он лежит под ногами в виде лунной пыли, реголита, который на 40-45% состоит из кислорода. На Земле уже предпринимались попытки извлекать кислород из искусственно созданной лунной пыли, но процесс оказался нерентабельным.
И вот в прошлом году ученые предложили новый способ извлечения кислорода из реголита, а сейчас на опытной фабрике попробуют впервые получить кислород в промышленных масштабах.
Правда, экспериментальная фабрика открыта пока не на Луне, а в центре Европейского космического агентства в Нидерландах.
Эта новая технология была разработана химиками из университета Глазго во главе с Бет Ломакс и получила название «электролиз расплавленных солей».
Реголит помещается в емкость, проложенную проволокой, туда добавляется в качестве электролита хлорид кальция, и смесь нагревается до 950 градусов — при такой температуре материал не плавится.
Затем через емкость пропускается электрический ток, при этом выделяется кислород, а соли перемещаются к аноду, где их можно удалить. Выход кислорода составляет до 96%, и вдобавок побочным продуктом являются сплавы металлов.
Если эксперимент удастся, люди получат жизненно необходимый для дальнейших лунных миссий источник кислорода, а там не за горами будут и первые долговременные лунные базы.
«Возможность получать кислород из лунных источников будет очень полезна для будущих лунных поселенцев, ведь он нужен как для дыхания, так и для производства ракетного топлива на местах», — говорит Бет Ломакс.
Древнейшие эукариоты лахандинской биоты (мезопротерозой, юго-восточная Сибирь) – морфологические и биогеохимические данные
ДОКЛАДЫ РОССИЙСКОЙ АКАДЕМИИ НАУК. НАУКИ О ЖИЗНИ том 500 2021
НАУКИ О ЖИЗНИ том 500 2021
ДРЕВНЕЙШИЕ ЭУКАРИОТЫ ЛАХАНДИНСКОЙ БИОТЫ 411
таллому аимонемы свойственно анастомозное
ветвление, не известное у желто-зеленых водо-
рослей, но встречающееся у филогенетически
удаленной группы хищных нематофаговых гри-
бов [8]. Кроме того, в отличие от палеовошерии,
таллом аимонемы полностью лишен перегоро-
док, а на окончаниях нитей таллома не встреча-
ются отверстия. Учитывая перечисленные разли-
чия, отождествление этих ископаемых таксонов
представляется необоснованным. Но подчерк-
нем, что эукариотная природа палеовошерии и
аимонемы сомнений не вызывает, поскольку хотя
среди прокариот и встречаются формы, образую-
щие ценоцитный ветвящийся мицелий, диаметр их
нитей не превышает двух микрометров [11].
Пример палеовошерии относится к разряду
редчайших. Большинство же протерозойских ис-
копаемых остатков лишены диагностических
признаков, позволяющих ассоциировать их с
конкретными современными организмами.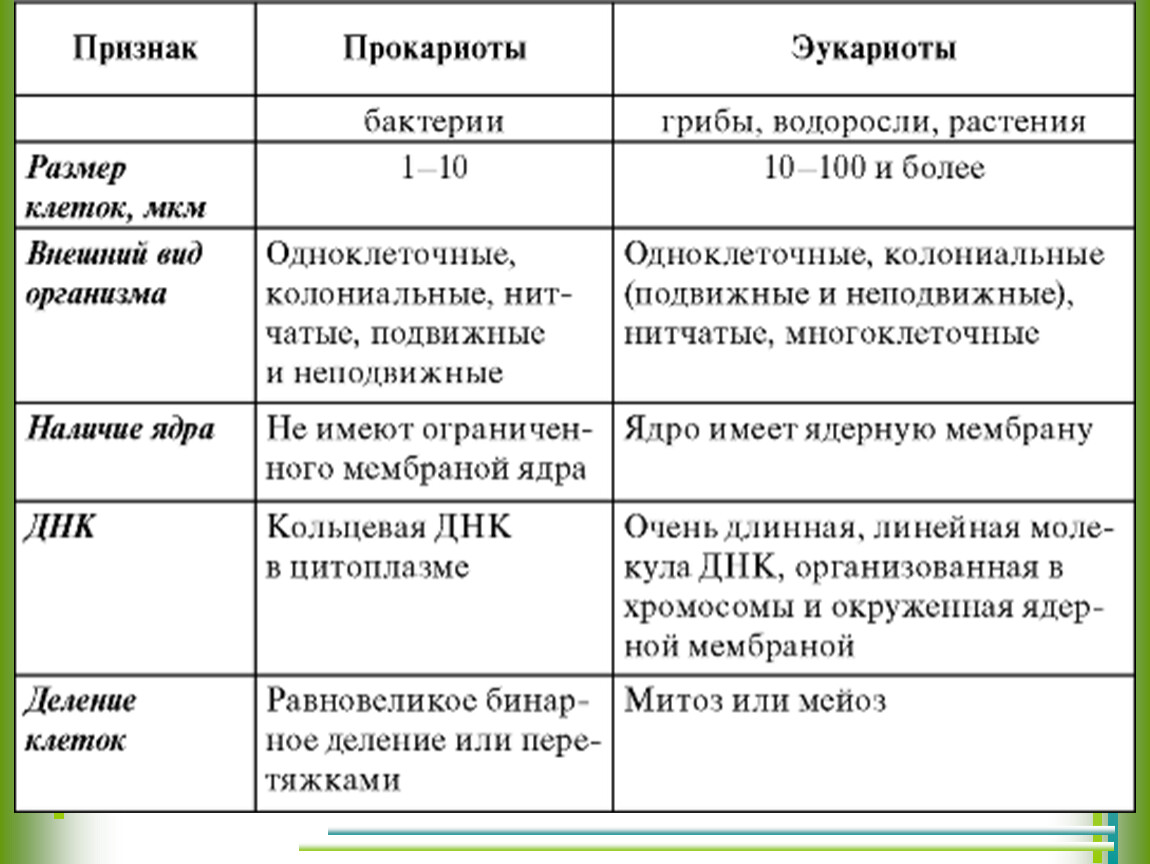 Это
Это
может быть связано либо с первичным отсутстви-
ем у них каких-либо синапоморфий, либо с их
утратой в процессе фоссилизации. Поэтому для
подавляющего большинства протерозойских
эуфоссилий возможна интерпретация лишь до
домена. Для этого достаточно выявить опреде-
ленные конструктивные детали морфологии или
их комбинации, возможные только при эффек-
тивной локализации основных внутриклеточных
процессов (компартментализация внутриклеточ-
ного пространства), свойственной эукариотам и
не встречающейся у прокариот. К таким деталям
строения, например, можно отнести характер ор-
наментации клеточной стенки и наличие выро-
стов, которые в сочетании с крупными размерами
данных элементов не встречаются у прокариот.
Так, к эукариотам следует относить лахандин-
ских стриатных акритарх Valeria lophostriata, акан-
томорфных акритарх Trachyhystrichosphaera aimi-
ka, спорангиоподобные структуры Caudosphaera
expansa и новую еще неописанную форму, а также
сферические и цилиндрические оболочки с еди-
ничными отростками ценоцитного уровня орга-
низации, напоминающие прорастающие споры
низших грибов, страменопил и зеленых водорос-
лей – Germinosphaera bispinosa и Jacutianema solubila.
Концентрически ребристые сферические обо-
лочки V. lophostriata диаметром более 60 мкм отно-
сятся к эукариотам на основании крупных разме-
ров элементов скульптуры – ребра имеют ширину
около 1 мкм (рис. 3г, 3д), в то время как у прока-
риот аналогичные структуры не выходят из нано-
метрового класса. Значительно более крупные
оболочки T. a i mika (диаметр > 200 мкм) покрыты
полыми нерегулярно расположенными выроста-
ми диаметром ~5 мкм и длиной более 40 мкм
(рис. 3е, 3ж). Несмотря на то что среди бактерий,
живущих в насыщенных питательными веще-
ствами условиях, встречаются клетки сравнимого
размера, они не орнаментированы, как и макро-
скопические колониальные оболочки цианобак-
териальных водорослей. Следует отметить, что
некоторые спорангиеподобные структуры бакте-
рий могут быть снабжены выростами, но их диа-
метр не выходит из нанометрового диапазона
[10]. Большая часть акритарх обычно относится к
одноклеточным протистам, по-видимому, расти-
тельного происхождения, хотя встречаются фор-
мы, демонстрирующие морфологию, характерную
для зооспоровых грибов. К сожалению, за недо-
К сожалению, за недо-
статком диагностических признаков, определение
их систематического положения внутри домена
эукариот в значительной мере спекулятивно.
Отдельную группу в лахандинской биоте обра-
зуют гладкие сферические оболочки с одиноч-
ным отростком. Среди этих форм выделяются
С. expansa – это сферы диаметром более 200 мкм,
снабженные хвостообразным отростком, состоя-
щим из более тонких нитей около 1 мкм толщи-
ной (рис. 3з, 3и), и еще неописанная форма,
представленная субсферическими оболочками
диаметром более 30 мкм, прикрепленными через
короткую ножку к нитям диаметром около 1 мкм
(рис. 3к, 3л). Морфология этих ископаемых форм
близка к виду Ourasphaira giraldae Loron et al., 2019
из (?) мезо- и неопротерозойских отложений
Арктической Канады. В составе стенки оболочки
O. giraldae присутствует хитин и отсутствует цел-
люлоза, что, учитывая внешний облик и относи-
тельно регулярное септирование таллома, являет-
ся диагностическим признаком грибов [3]. На
На
данный момент у нас пока нет данных о биохи-
мии новой лахандинской фоссилии, что затруд-
няет систематическую интерпретацию этих иско-
паемых остатков. Сходной морфологией обладает
G. bispinosa – это сферические оболочки диамет-
ром 30–40 мкм, несущие постепенно сужающий-
ся к свободному концу вырост, полость которого
свободно сообщается с полостью сферы.
Кроме того, к морфогруппе гладких сфер с
одиночным отростком могут быть отнесены
представители вида J. solubila – сферические до
цилиндрических оболочки 20–40 мкм в диаметре
с одиночным, выдержанным по ширине трубча-
тым выростом, имеющим единую со сферой по-
лость (рис. 3м, 3н). Хотя среди бактерий известны
формы, cпоры которых при прорастании образу-
ют длинные выросты, они значительно меньшего
размера [10]. Также некоторые бактерии способ-
ны образовывать сложные ветвящиеся структуры
со стеблями, увенчанными сферическими спо-
рангиеподобными плодовыми телами до 1000 мкм
[12]. Однако поверхность крупных плодовых тел
Однако поверхность крупных плодовых тел
не гладкая, а бугристая, повторяющая форму на-
ходящихся внутри спор [13].
Прокариоты
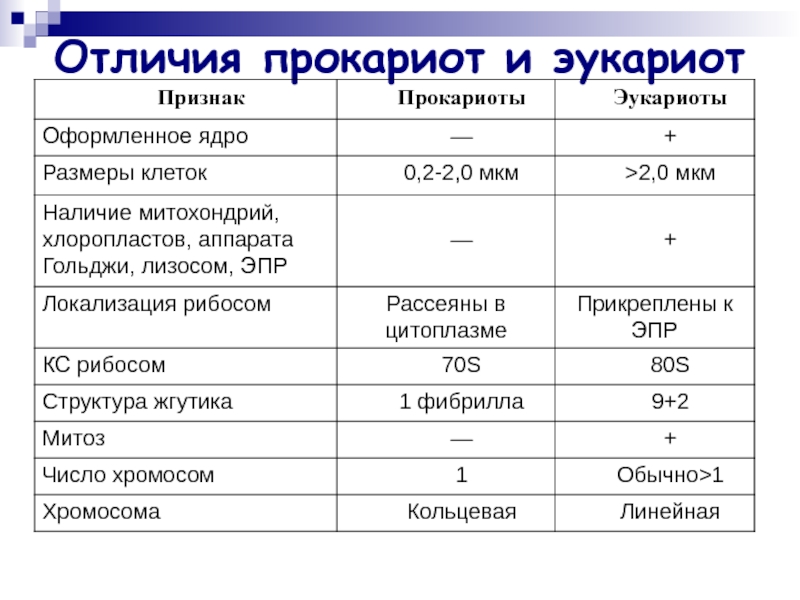
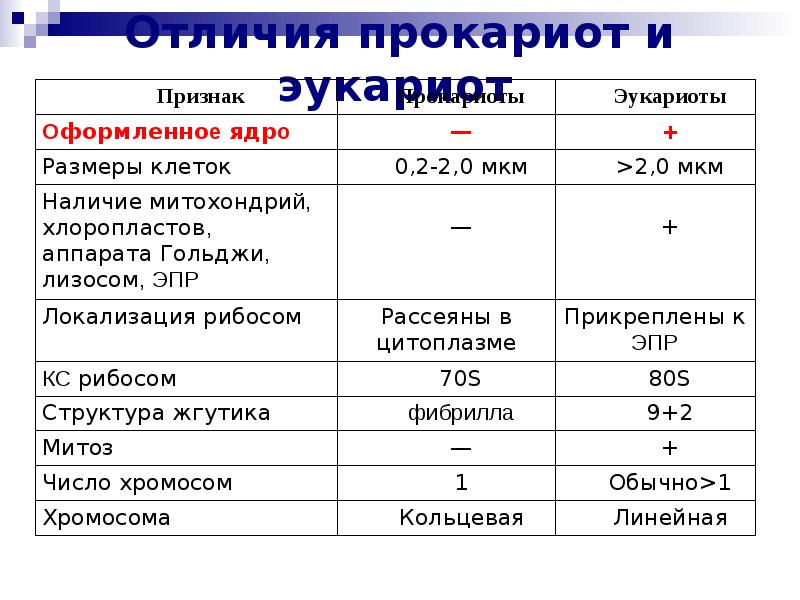
Все организмы, имеющие клеточное строение, делятся на две группы: предъядерные (прокариоты) и ядерные (эукариоты).
Клетки прокариот, к которым относятся бактерии, в отличии от эукариот, имеют относительно простое строение. В прокариотической клетке нет организованного ядра, в ней содержится только одна хромосома, которая не отделена от остальной части клетки мембраной, а лежит непосредственно в цитоплазме. Поскольку бактериальная хромосома содержит очень мало белков и представляет собой нить ДНК, она может быть только условно названа хромосомой. Однако в ней также записана все наследственная информация бактериальной клетки.
Цитоплазма прокариот по сравнению с цитоплазмой эукариотических клеток значительно беднее по составу структур. Там находятся многочисленные более мелкие, чем в клетках эукариот, рибосомы.
| Признаки | Прокариоты | Эукариоты |
|---|---|---|
| Ядерная оболочкаДНК | НетЗамкнута в кольцо(условно называетсябактериальная хромосома) | ЕстьЯдерная ДНК представляет собой линейнуюструктуруи находится в хромосомах |
| ХромосомыМитозМейозГаметыМитохондрииПластиды у автотрофовСпособ поглощенияпищиПищеварительные вакуолиЖгутики | НетНетНетНетНетНетАдсорбция черезклеточную мембрануНетЕсть | ЕстьЕстьЕстьЕстьЕстьЕстьФагоцитози пиноцитозЕстьЕсть |
Другие заметки по биологии
Тесты по биологии по теме «Признаки эукариот и прокариот»
ПРИЗНАКИ эукариот и прокариот (открытый банк ЕГЭ, ФИПИ)
1. Растения, грибы, животные – это эукариоты, так как их клетки 1) не имеют оформленного ядра 2) не делятся митозом 3) имеют оформленное ядро 4) имеют ядерную ДНК, замкнутую в кольцо
Растения, грибы, животные – это эукариоты, так как их клетки 1) не имеют оформленного ядра 2) не делятся митозом 3) имеют оформленное ядро 4) имеют ядерную ДНК, замкнутую в кольцо
2. Грибы, клетки которых имеют оболочку, ядро, цитоплазму с органоидами, относят к группе организмов 1) эукариот 2) хемотрофов 3) автотрофов 4) прокариот
3. Клетки растений, в отличие от клеток животных, содержат 1) ядра 2) митохондрии 3) хлоропласты 4) эндоплазматическую сеть
5.Почему бактерии относят к прокариотам? 1) содержат в клетке ядро, обособленное от цитоплазмы 2) состоят из множества дифференцированных клеток 3) имеют одну кольцевую хромосому 4) не имеют клеточного центра, комплекса Гольджи и митохондрий 5) не имеют обособленного от цитоплазмы ядра 6) имеют цитоплазму и плазматическую мембрану
7. Организмы растений, животных, грибов и бактерий состоят из клеток – это свидетельствует о 1) единстве органического мира 2) разнообразии строения живых организмов 3)связи организмов со средой обитания 4) сложном строении живых организмов
Организмы растений, животных, грибов и бактерий состоят из клеток – это свидетельствует о 1) единстве органического мира 2) разнообразии строения живых организмов 3)связи организмов со средой обитания 4) сложном строении живых организмов
8. Бактерии, в отличие от растений, имеют 1) специализированные половые клетки 2) одну кольцевую молекулу ДНК в клетке 3) ядро, обособленное от цитоплазмы ядерной оболочкой 4) две и более хромосом
9. Что свидетельствует о более высокой организации грибов по сравнению с бактериями? 1)способность вступать в симбиоз с растениями 2) наличие в клетках цитоплазмы и оболочки 3) наличие ядра и митохондрий в клетках 4) питание готовыми органическими веществами
10. Растительная клетка, в отличие от животной, имеет 1) плазматическую мембрану 2)аппарат Гольджи 3) митохондрии 4) пластиды
11. Сходство клеток прокариот и эукариот состоит в том, что они имеют 1) цитоплазму 2) комплекс Гольджи 3) митохондрии 4)ядро
12. Клетки животных относят к группе эукариотных, так как они имеют 1) хлоропласты 2) плазматическую мембрану 3) оболочку 4) ядро, отделенное от цитоплазмы оболочкой
Клетки животных относят к группе эукариотных, так как они имеют 1) хлоропласты 2) плазматическую мембрану 3) оболочку 4) ядро, отделенное от цитоплазмы оболочкой
1) митохондрии
2) плазматическую мембрану
3) клеточный центр
4) пищеварительные вакуоли
15. К прокариотам относят 1) вирусы и бактериофаги 2) бактерии и синезеленые 3)водоросли и простейшие 4) грибы и лишайники
16 . Сходство в строении клеток автотрофных и гетеротрофных организмов состоит в наличии у них 1)хлоропластов 2) плазматической мембраны 3) оболочки из клетчатки 4) вакуолей с клеточным соком
17Митохондрии, как и лизосомы, отсутствуют в клетках 1) бактерий 2) грибов 3) животных 4) Растений
18. Установите соответствие между особенностью строения клетки и царством, для которого оно характерно.
ОСОБЕННОСТЬ СТРОЕНИЯ КЛЕТОК ЦАРСТВА
А) наличие пластид 1) Грибы
Б) отсутствие хлоропластов 2. Растения
Растения
В) запасное вещество – крахмал
Г) наличие вакуолей с клеточным соком
Д) клеточная стенка содержит клетчатку
Е) клеточная стенка содержит хитин
19. Плотная оболочка отсутствует в клетках 1)бактерий 2) млекопитающих 3) земноводных 4) грибов 5) птиц 6) Растений
20. Организмы, клетки которых имеют хлоропласты, относят к царству 1) животных 2) растений 3)грибов4) бактерий
21. Особенности обмена веществ у растений по сравнению с животными состоят в том, что в их клетках происходит 1) хемосинтез 2) энергетический обмен 3) фотосинтез 4) биосинтез белка
22. . В клетках каких организмов содержится в десятки раз больше углеводов, чем в клетках животных? 1) бактерий-сапротрофов 2) одноклеточных 3) простейших 4) растений
23. К эукариотам относят 1) кишечную палочку 2)амебу 3) холерный вибрион 4) стрептококк
24. К прокариотным относят клетки 1) животных 2) цианобактерий 3) грибов 4) растений
К прокариотным относят клетки 1) животных 2) цианобактерий 3) грибов 4) растений
25. Клетки прокариот, в отличие от клеток эукариот, не имеют 1) хромосом 2) клеточной оболочки 3) ядерной мембраны 4) плазматической мембраны
26. Прокариоты – это организмы, 1) клетки которых не имеют оформленного ядра
2) содержащие в клетках одно или несколько ядер 3) состоящие из одинаковых клеток и не имеющие тканей 4) которые не имеют клеточного строения
27. Наследственное вещество находится в цитоплазме клетки у 1) дизентерийной амебы 2) малярийного паразита 3) кишечной палочки 4) эвглены зеленой
28. Запасным углеводом в животной клетке является 1) крахмал 2) гликоген 3) хитин 4) целлюлоза
29. Клетки эукариотных организмов, в отличие от прокариотных, имеют 1) цитоплазму 2) ядро, покрытое оболочкой 3) молекулы ДНК 4) митохондрии 5) плотную оболочку 6) эндоплазматическую сеть
30 В бактериальной клетке нет 1) обособленного ядра 2) клеточной стенки 3) цитоплазмы 4) рибосом.
31. Эукариоты – это организмы, в клетках которых 1) отсутствуют митохондрии 2) ядрышки находятся в цитоплазме 3) ядерная ДНК образует хромосомы 4) отсутствуют рибосомы
32. Клетка многоклеточного животного, в отличие от клетки простейшего, 1) покрыта оболочкой из клетчатки 2) выполняет все функции организма 3) выполняет определенную функцию 4) представляет собой самостоятельный организм
33. В клетках растений, в отличие от клеток человека, животных, грибов происходит
1) выделение 2) питание 3) дыхание 4) фотосинтез
30. Почему бактерии нельзя отнести к эукариотам?
31. К эукариотическим относят клетки 1) клубеньковых бактерий 2) цианобактерий
3) растений 4) кишечной палочки
32. В клетках грибов, как и в клетках животных, отсутствуют 1) лейкопласты и хлоропласты 2) оболочки из хитина 3) плазматические мембраны 4) митохондрии и рибосомы
В клетках грибов, как и в клетках животных, отсутствуют 1) лейкопласты и хлоропласты 2) оболочки из хитина 3) плазматические мембраны 4) митохондрии и рибосомы
33. Клетки грибов, в отличие от клеток бактерий, имеют 1) цитоплазму 2) ядр 3)плазматическую мембрану 4) рибосомы
34. К прокариотам относятся 1) растения 2) животные 3) грибы-паразиты 4) цианобактерии
35. При размножении прокариот происходит удвоение 1) кольцевой ДНК 2) хроматид 3) митохондрий 4) сестринских хромосом
36. На процессы жизнедеятельности животные расходуют 1) тепловую энергию, поступающую из окружающей среды 2) механическую энергию, которая освобождается при сокращении мышц 3) световую энергию, которую поглощают клетки их тела 4) энергию, которая освобождается при окислении органических веществ в клетках тела
37. Почему зелёную эвглену одни учёные относят к растениям, а другие – к животным? Укажите не менее трёх причин.
38. Главное отличие клеток прокариот от эукариот состоит в 1) отсутствии ядерного вещества в цитоплазме 2) наличии рибосом на эндоплазматической сети 3) наличии органоидов движения 4) отсутствии оформленного ядра
39. Клетка бактерии, как и растительная клетка, имеет 1) ядро 2) комплекс Гольджи 3) эндоплазматическую сеть 4) цитоплазму
40. Клетки каких организмов имеют клеточную оболочку (клеточную стенку)? 1) животных
2) растений 3) человека 4) грибов 5) вирусов 6) Бактерий
41. В животной клетке отсутствуют 1) пластиды 2) лизосомы 3) элементы комплекса Гольджи 4) центриоли клеточного центра
42. Чем бактерии отличаются от организмов других царств живой природы? Укажите не менее 3-х отличий.
43. Сходство бактериальной клетки с клетками организмов других царств состоит в наличии в ней 1) ядрышка 2) митохондрий 3) цитоплазмы 4) вакуоли с клеточным соком
44. Клетка – единица строения и жизнедеятельности
Клетка – единица строения и жизнедеятельности
1) вируса табачной мозаики
2) возбудителя СПИДа
3) бактерии кишечной палочки
4) белой планарии
5) обыкновенной амебы
6) бактериофага
45. Почему одноклеточных животных относят к эукариотам? 1) имеют оформленное ядро
2) содержат кольцевую хромосому 3) синтезируют на рибосомах белки
4) окисляют органические вещества и запасают АТФ
46. Вывод о родстве растений и животных можно сделать на основании 1) хромосомной теории
2) закона сцепленного наследования 3) теории гена 4) клеточной теории
47. По каким признакам строения можно отличить бактериальную клетку от растительной? Назовите не менее трёх признаков.
48. К эукариотам относят клетки 1) бактерий 2) вирусов 3) животных 4) бактериофагов
49. Почему бактерии относят к прокариотам? их тело состоит из одной клетки
Почему бактерии относят к прокариотам? их тело состоит из одной клетки
2)
они имеют микроскопические размеры
3)
они не имеют оформленного ядра
4)
они не имеют цитоплазмы и клеточной стенки
50.
Грибы, по сравнению с бактериями, имеют более высокий уровень организации, так как 1) по способу питания они являются гетеротрофами 2) их споры переносят неблагоприятные условия 3) их клетки имеют плотную оболочку 4) их клетки имеют оформленное ядро51. Эукариоты – это организмы, в клетках которых 1) ядерное вещество не отделено от цитоплазмы 2) одна кольцевая хромосома 3) отсутствуют многие органоиды 4) ядро отделено от цитоплазмы оболочкой
52. Прокариотические клетки отличаются от эукариотических 1) наличием рибосом 2)отсутствием митохондрий 3) отсутствием оформленного ядра 4) наличием плазматической мембраны 5) отсутствием органоидов движения 6)наличием кольцевой молекулы ДНК
53. В животной и растительной клетках имеются 1) вакуоли с клеточным соком 2) пластиды
В животной и растительной клетках имеются 1) вакуоли с клеточным соком 2) пластиды
3) митохондрии 4) пищеварительные вакуоли
54. Для прокариотической клетки характерно 1) отсутствие цитоплазмы и оболочки 2) отсутствие процесса фотосинтеза 3) деление путём митоза 4) наличие кольцевых ДНК в цитоплазме
55. Отсутствие в клетке митохондрий, комплекса Гольджи, ядра указывает на её принадлежность к 1) эукариотам 2) прокариотам 3) вирусам 4) бактериофагам
56. Сходное строение клеток растений и животных – доказательство 1) их родства 2) общности происхождения организмов всех царств 3) происхождения растений от животных 4) усложнения организмов в процессе эволюции 5) единства органического мира 6) многообразия организмов
57. Организмы, клетки которых не содержат оформленного ядра, митохондрий, аппарата Гольджи, относят к группе 1) прокариот 2)эукариот 3) автотрофов 4) гетеротрофов
58. Клетку бактерии относят к группе прокариот, так как она не содержит
Клетку бактерии относят к группе прокариот, так как она не содержит
1) органоидов движения 2) клеточной оболочки 3) многих органоидов и ядра
4) плазматической мембраны
59. Малярия – заболевание человека, в результате которого развивается малокровие. Кем оно вызывается? Объясните причину малокровия.
60. Найдите три ошибки в приведённом тексте. Укажите номера предложений, в которых они сделаны, исправьте их.
1. По типу организации различают про- и эукариотические клетки.
2. У эукариотических организмов размеры клеток значительно больше, чем
у прокариот. 3. В клетках эукариот имеется нуклеоид. 4. В клетках прокариот нет мембранных органоидов. 5. В клетках всех эукариот присутствуют митохондрии, эндоплазматическая сеть, хлоропласты и другие органоиды. 6. В синтезе белков эукариоты используют свободный азот атмосферы. 7. Растения, грибы, животные – это эукариоты.
61. Организмы, клетки которых имеют обособленное ядро, – это 1) вирусы 2) прокариоты
3) эукариоты 4) бактерии
62. Синтез молекул АТФ за счёт энергии солнечного света происходит в клетках
1) грибов 2) многоклеточных животных 3) растений 4) бактерий гниения
63. Клетку можно отнести и к клеточному, и к организменному уровням организации жизни. Объясните почему. Приведите соответствующие примеры.
64. Выберите одно из положений клеточной теории. 1) Зигота образуется в процессе оплодотворения. 2) В процессе мейоза образуется четыре гаплоидные клетки.
3) Клетки сходны по строению и химическому составу.
4) Наследственная информация клетки сосредоточена в хромосомах ядра.
65.
Установите соответствие между характеристикой клетки и её типом. наличие нуклеоида – АБ)
наличие хитина в клеточной стенке – Б
В)
отсутствие мембранных органоидов – В
Г)
способность к хемосинтезу – Г
Д)
запасающее вещество – крахмал — Д
бактериальная
2)
грибная
3)
растительная
Конец формы
66. В клетках животных, в отличие от клеток высших растений, содержатся 1) митохондрии 2) крупные вакуоли 3) центриоли 4) субъединицы рибосом
В клетках животных, в отличие от клеток высших растений, содержатся 1) митохондрии 2) крупные вакуоли 3) центриоли 4) субъединицы рибосом
67. В животной клетке отсутствует 1) клеточный центр 2) клеточная стенка 3) плазматическая мембрана 4) цитоскелет
68. Для эукариотической клетки, в отличие от прокариотической, характерно наличие
1) рибосом 2) цитоплазмы 3) лизосом 4) немембранных органоидов
69. Найдите три ошибки в приведённом тексте. Укажите номера предложений, в которых они сделаны, исправьте их.
1. Растения, как и другие организмы, имеют клеточное строение, питаются, дышат, растут, размножаются. 2. Как представители одного царства растения имеют признаки, отличающие их от других царств. 3. Клетки растений имеют клеточную стенку, состоящую из целлюлозы, пластиды, вакуоли с клеточным соком. 4. В клетках высших растений имеются центриоли.
5. В растительных клетках синтез АТФ осуществляется в лизосомах. 6. Запасным питательным веществом в клетках растений является гликоген. 7. По способу питания большинство растений автотрофы.
6. Запасным питательным веществом в клетках растений является гликоген. 7. По способу питания большинство растений автотрофы.
70. Установите соответствие между характеристикой клетки и типом организации этой клетки.
ХАРАКТЕРИСТИКА ТИП КЛЕТОЧНОЙ ОРГАНИЗАЦИИ
А) Хромосома образована кольцевой ДНК. 1) прокариотический
Б) Клетка делится митозом. 2) эукариотический
В) В цитоплазме находятся лизосомы.
Г) Мембрана образует мезосомы.
Д) Клеточный центр участвует в образовании веретена деления.
Е) Отсутствуют мембранные органоиды.
71. В бактериальной клетке нет 1) обособленного ядра 2) клеточной стенки 3) цитоплазмы 4) рибосом
73. На рисунке изображена клетка1) грибная
2) животная
3) бактериальная
4) растительная
74. Все прокариотические и эукариотические клетки имеют 1) митохондрии и ядро 2)вакуоли и комплекс Гольджи 3) ядерную мембрану и хлоропласты 4)плазматическую мембрану и рибосомы
Все прокариотические и эукариотические клетки имеют 1) митохондрии и ядро 2)вакуоли и комплекс Гольджи 3) ядерную мембрану и хлоропласты 4)плазматическую мембрану и рибосомы
Как на Земле появились клетки с ядрами
Архея под названием Локи показала, как древние безъядерные микробы могли дать начало эукариотическим клеткам.
Микроба Lokiarchaea нашли на глубине 2 352 метров, в системе чёрных курильщиков под названием «Замок Локи». (Фото R.B. Pedersen, Centre for Geobiology (University of Bergen, Norge)).
Колония метанпродуцирующих архей Methanosarcina mazei. (Фото Ralph Robinson / Visuals Unlimited / Corbis.)
Архея Methanoculleus nigri с отпочковывающейся клеткой. (Фото Dr. Terry Beveridge / Visuals Unlimited / Corbis.)
‹
›
Жизнь на Земле делится на три домена: эукариоты (птицы, звери, рыбы, черви и т. д., все многоклеточные и ещё масса одноклеточных организмов), бактерии и археи. Эукариоты отличаются от двух остальных по целому ряду важных признаков – в их клетках есть ядро, в котором хранится генетический материал, а также целый набор клеточных органоидов, сделанных из липидных мембран; кроме того, у эукариот есть масса отличий на уровне самых базовых молекулярных процессов, как-то: синтез белка, репликация ДНК и т. д.
Эукариоты отличаются от двух остальных по целому ряду важных признаков – в их клетках есть ядро, в котором хранится генетический материал, а также целый набор клеточных органоидов, сделанных из липидных мембран; кроме того, у эукариот есть масса отличий на уровне самых базовых молекулярных процессов, как-то: синтез белка, репликация ДНК и т. д.
Бактерий и архей (которых раньше называли архебактериями) долгое время объединяли вместе, поскольку ни у бактерий, ни у архей нет ядра и мембранных органелл, однако впоследствии оказалось, что одни довольно сильно отличаются от других. Перечислять отличия мы не будем, скажем лишь, что у архей обнаружили множество особенностей, общих с эукариотами. И с археями же связана одна из главных гипотез происхождения эукариот. Бактерии на Земле, очевидно, появились раньше, и долгое время биологи не могли понять, как и откуда рядом с бактериями появились клетки с ядрами и внутриклеточными сложноорганизованными мембранами. И вот, когда археями занялись во всеоружии молекулярно-генетических методов, возникла версия, что эукариоты есть результат симбиоза между обычной бактерией (или эубактерией) и археей.
Но в каких именно родственных отношениях находятся эукариоты и археи, всё равно оставалось неясным. С одной стороны, и те, и другие могли произойти от одного общего предка. С другой стороны, эукариотическая клетка могла возникнуть внутри какой-то уже сформировавшейся архейной группы. Сделать выбор здесь можно было бы, если бы нашёлся ещё какой-нибудь вид архей, с помощью которого можно было бы глубже прояснить генетические взаимосвязи между двумя доменами.
И такой вид был найден: в статье в Nature группа исследователей из Уппсальского университета описывает архею Lokiarchaea, названную так потому, что обнаружили её в грунте рядом с «Замком Локи» – системой гидротермальных источников на Срединно-Атлантическом хребте, между Норвегией и Гренландией.
Первоначальный анализ показал, что Lokiarchaea принадлежат к одной из самых крупных групп морских архей, которых, правда, до сих пор не удаётся культивировать в лабораторных условиях (что для архей, да и для бактерий, совсем не редкость). Метагеномный анализ позволил собрать воедино 92% генетического материала, после чего оказалось, что около 175 белков Lokiarchaea весьма близки эукариотическим аналогам, участвующим в формировании мембран, изменении формы клетки и фагоцитозе (то есть мембранном захвате частичек из внешней среды). Более того, эукариоты, по-видимому, сформировались как отдельный кластер уже внутри этой группы морских микробов. Сам Lokiarchaea, несомненно, относится к археям, однако в прошлом у него и у эукариот был общий архейный предок.
Метагеномный анализ позволил собрать воедино 92% генетического материала, после чего оказалось, что около 175 белков Lokiarchaea весьма близки эукариотическим аналогам, участвующим в формировании мембран, изменении формы клетки и фагоцитозе (то есть мембранном захвате частичек из внешней среды). Более того, эукариоты, по-видимому, сформировались как отдельный кластер уже внутри этой группы морских микробов. Сам Lokiarchaea, несомненно, относится к археям, однако в прошлом у него и у эукариот был общий архейный предок.
Одна из проблем, возникающих в связи с гипотезой о слиянии архей и бактерий в одну эукариотическую клетку – отсутствие и у бактерий, и у архей способности к фагоцитозу. То есть они не могут образовать впячивание в своей внешней оболочке и заключить в себя некий объект, а ведь если мы придерживаемся гипотезы о симбиозе, то одна клетка должна как бы спрятать в себя другую.
У большинства бактерий и архей, кроме наружной мембраны, есть ещё и прочная клеточная стенка, которая помешала бы фагоцитозу, задумай клетка его осуществить; но, кроме того, здесь нужно уметь ещё и довольно тонко манипулировать собственной мембраной, для чего нужен соответствующий белковый аппарат. И вот, как оказалось, такой аппарат, по крайней мере, частично, есть у Lokiarchaea.
И вот, как оказалось, такой аппарат, по крайней мере, частично, есть у Lokiarchaea.
По словам Тийса Эттема (Thijs Ettema), этот микроб как бы стоит в начале эволюционного пути, ведущего к полноценному фагоцитозу. Миллиарды лет назад похожая архея могла бы, спустя какое-то время, в конце концов захватить настоящую бактерию – и не съесть её, а оставить в себе в качестве симбионта; впоследствии «гость» мог бы превратиться, например, в митохондрию или хлоропласт. (В скобках заметим, что появление ядра часто объясняют тем, что, поселив у себя симбионта, клетке-хозяину пришлось дополнительно защитить от него свой собственный геном, чтобы гены симбионта не влезли в хозяйскую ДНК, и как раз ядерная мембрана стала одной из линий такой защиты.)
Среди других особенностей Lokiarchaea авторы работы отмечают гены скелетного белка актина, которые больше похожи на эукариотические актиновые гены, чем на архейные, и несколько десятков последовательностей, соответствующих особым регуляторным сигнальным белкам – их количество у Loki (как ласково называют нового микроба) примерно такое же, как у многих одноклеточных эукариотических видов. Кроме того, у Lokiarchaea в геноме нашли кусок, похожий на эукариотический ген ESCRT, необходимый для внутриклеточного перемещения мембранных пузырьков-везикул.
Кроме того, у Lokiarchaea в геноме нашли кусок, похожий на эукариотический ген ESCRT, необходимый для внутриклеточного перемещения мембранных пузырьков-везикул.
Теперь перед биологами стоит задача выяснить, что всё-таки делают эукариотоподобные гены в клетках археи. (Загрязнение посторонним генетическим материалом исследователи уже исключили.) Очевидно, что у Lokiarchaea они появились не зря, в противном случае в них накопилось бы множество мутаций, они утратили бы функциональность и стали бы генетическим мусором. Возможно, что у микроба есть какие-то зачатки, какие-то элементы эндомембранных структур и белкового скелета, напоминающие таковые у эукариот. Но для того, чтобы в этом убедиться, капризную архею придётся научиться выращивать в лабораториях. Хотя в целом и так понятно, что Lokiarchaea стала одним из крупнейших открытий последних десятилетий: с её помощью мы можем понять, что происходило на одном из важнейших этапов развития жизни, во время появления на Земле эукариот.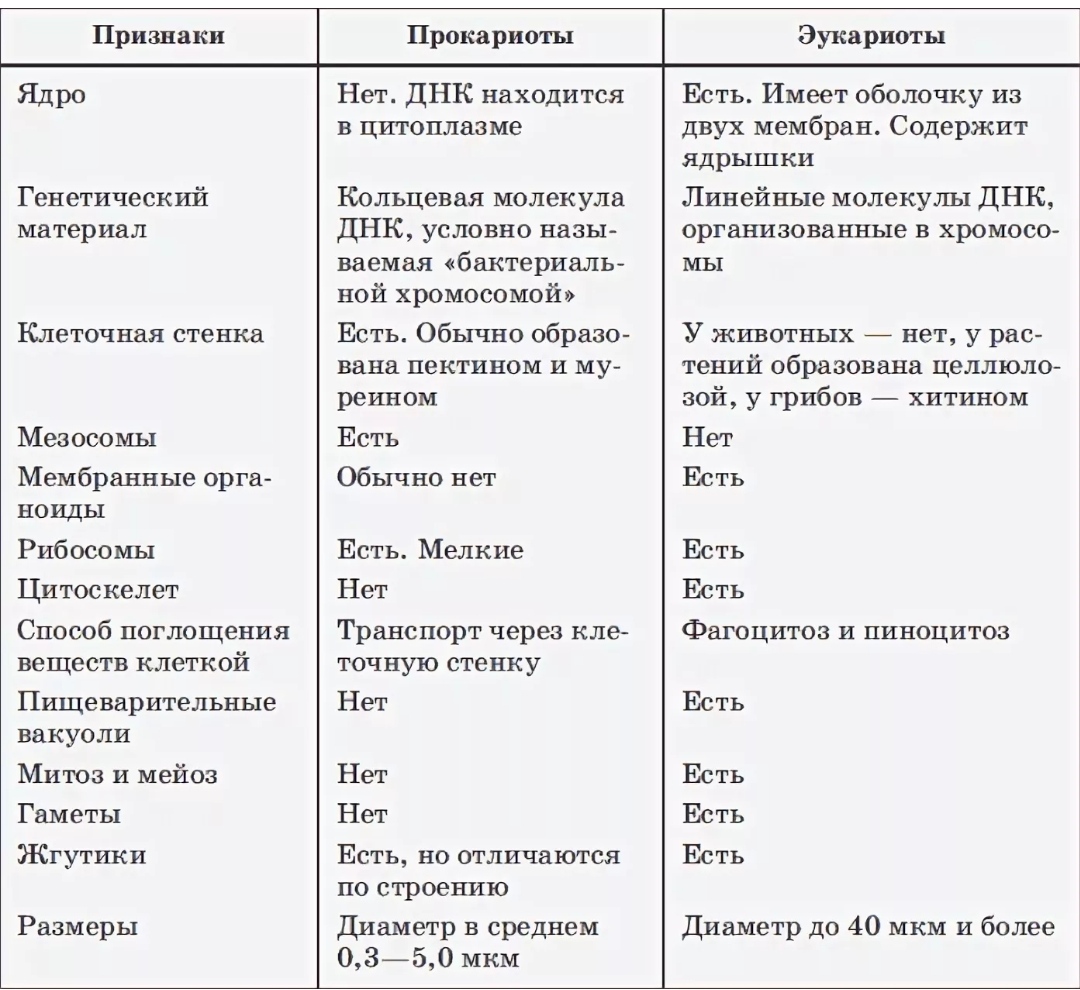
2.7. Различия в строении клеток эукариот и прокариот
Вопрос 1. Какие признаки примитивности прокариот по сравнению с эукариотами вы можете назвать?
У прокариот отсутствуют ядро и хромосомы. Наследственная информация прокариот представлена единственной молекулой ДНК, которая замкнута в виде кольца и расположена в цитоплазме.
Рибосомы у прокариот очень мелкие. Л органоидов, окруженных мембранами (эндоплазматическая сеть, аппарат Гольджи, митохондрии, пластиды), прокариоты и вовсе не имеют. В эукариотической клетке эти структуры предназначены для синтеза разнообразных соединений. В силу их отсутствия у прокариот реакции обмена веществ протекают на выступах клеточной мембраны.
Среди прокариот много анаэробов, т. е. организмов, не использующих кислород. Это тоже следует отметить как признак примитивности, так как при кислородном расщеплении органических веществ выделяется много больше энергии, чем при анаэробном расщеплении.
Прокариоты в большинстве своем размножаются бесполым путем, половой процесс, основанный на обмене генетической информацией, у них встречается редко.
Вопрос 2. Для чего бактериям споры?
Споры у бактерий формируются при наступлении неблагоприятных условий. При этом содержимое клетки сжимается, вокруг него образуется плотная оболочка. В таком виде бактерии способны выдерживать высокие и низкие температуры, сухость воздуха, переноситься с помощью воды и ветра на дальние расстояния.
2.7. Различия в строении клеток эукариот и прокариот
5 (100%) 2 votesНа этой странице искали :
- какие признаки примитивности прокариот по сравнению с эукариотами
- какие признаки примитивности прокариот по сравнению с эукариотами вы можете назвать
- для чего бактериям споры
- Различия в строении клеток эукариот и прокариот
- какие признаки примитивности прокариот
Сохрани к себе на стену!
Когда впервые появились эукариотические клетки (клетки с ядрами и другими внутренними органеллами)? Что мы знаем о том, как они произошли от более ранних форм жизни?
Происхождение эукариот — царство жизни, включающее в себя все высшие растения и животные, включая нас самих, — происходили в сильно затемненной ранней истории земной шар. Следовательно, есть еще много предположений, связанных с ответом
этот вопрос. Карл
Вёз, профессор микробиологии Иллинойского университета в г.
Урбана-Шампейн и первооткрыватель архебактерий дает один ответ:
Следовательно, есть еще много предположений, связанных с ответом
этот вопрос. Карл
Вёз, профессор микробиологии Иллинойского университета в г.
Урбана-Шампейн и первооткрыватель архебактерий дает один ответ:
«Свидетельства микроокаменелостей убедительно свидетельствуют о том, что жизнь возникла на Земле
давным-давно, вероятно, в течение нескольких сотен миллионов лет после того, как планета
формирование.Осадочные породы возрастом 3,5 миллиарда лет (и, возможно, те 3,8
миллиардов лет) содержат то, что похоже на ископаемые строматолиты,
которые представляют собой естественные колонии, образованные фотосинтезирующими бактериями; внутри
stromalites можно увидеть микроскопические формы, напоминающие бактерии. Если эти
предполагаемые бактерии являются прямыми предками существующих фотосинтезирующих бактерий,
жизнь была к тому времени уже хорошо развита, пройдя этапы
что привело к самому последнему универсальному предку и расколу
наследственной линии в первичные линии происхождения.
«Сравнивая молекулярные последовательности, чтобы сделать вывод о генеалогии, молекулярные филогенетики говорят нам, что две основные линии происхождения ведут к эубактерии (или обычные бактерии, в том числе фотосинтезирующие бактерии) и ко второй общей линии, которая впоследствии разделилась, чтобы сформировать археи. (которые, как и эубактерии, являются прокариотами) и эукариотами (которые включают все высшие растения и животные). Все эти события, по-видимому, предшествовали древнейшим ископаемым строматолитам.Так появляется эукариотическая линия быть очень древним, примерно таким же древним, как две прокариотические линии.
«Ключевой вопрос без ответа здесь касается того, когда в эукариотической линии
сформировался эукариотический тип клеток. Эукариотические клетки кажутся структурно гораздо более
сложны, чем их прокариотические аналоги (из которых они произошли), поэтому
биологи обычно считают, что многие этапы эволюции должны
разделил двоих. Тем не менее эукариотический ствол на филогенетическом
древо жизни порождает множество ветвей, прежде чем одна из них дойдет до раздвоения, которое
отделяет предков растений от предков животных, что кажется
произошло более миллиарда лет назад. Кажется, было много
более ранние ответвления от эукариотического ствола, представленные одноклеточными
эукариоты (например, слизевики, жгутиконосцы, трихомонады,
дипломонады, микроспоридии и др.).
Кажется, было много
более ранние ответвления от эукариотического ствола, представленные одноклеточными
эукариоты (например, слизевики, жгутиконосцы, трихомонады,
дипломонады, микроспоридии и др.).
«Очевидно, что история эукариот восходит к эпохе, когда Атмосфера содержала мало кислорода или вообще не содержала его более двух миллиардов лет назад. Наша нынешняя концепция происхождения эукариотической клетки постоянно меняется. однако, и эволюционная последовательность, которая кажется простой, когда концептуально на диаграмме филогенетического дерева может быть гораздо более сложным и интересно на самом деле.Мы знаем, что эукариотическая клетка имеет древнее происхождение. происхождения, но мы еще не знаем эволюционной динамики, которая лежит в основе его формирование».
Дж. Петер Гогартен на кафедре молекулярной и клеточной биологии Университет Коннектикута в Сторрсе дает более широкий обзор:
«Этот вопрос является предметом непрекращающихся и оживленных споров.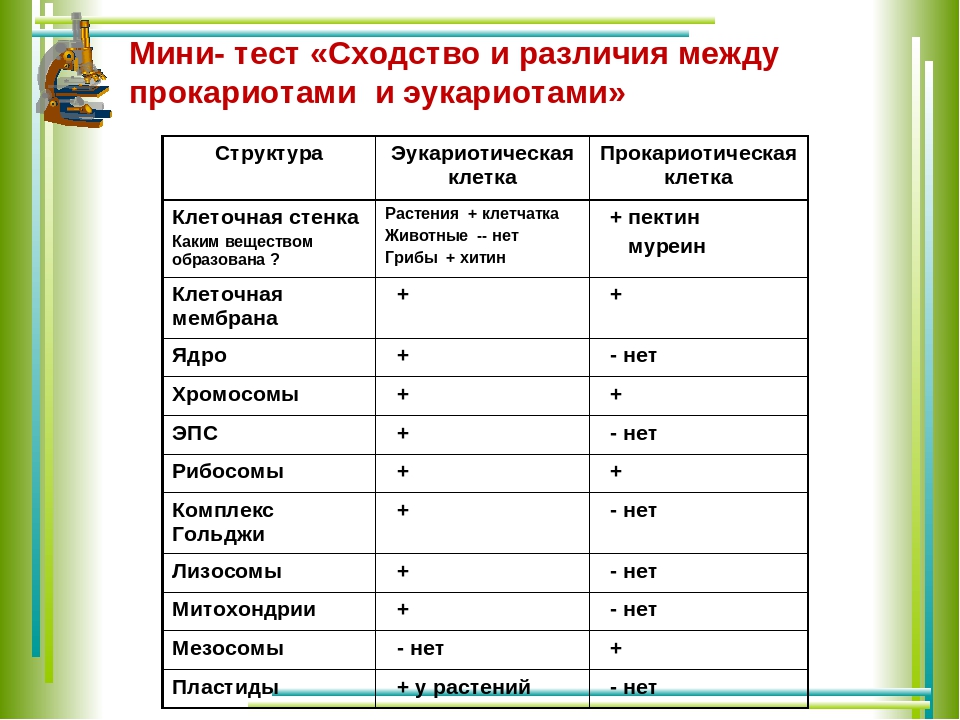 Лучший
предположения о времени эволюции эукариот колеблются чуть ниже 2,0
миллиардов лет до примерно 3.5 миллиардов лет до настоящего времени.
Лучший
предположения о времени эволюции эукариот колеблются чуть ниже 2,0
миллиардов лет до примерно 3.5 миллиардов лет до настоящего времени.
«Один из менее двусмысленных источников информации — летопись окаменелостей. Работа Гонсало Видал из Университета Упсалы в Швеции указывает на то, что одноклеточные планктонные эукариоты определенно датируются назад к 1,7 миллиарда лет до н.э. и очень вероятно, по крайней мере, 2,2 миллиарда лет Б.П. Однако ранняя летопись окаменелостей очень скудна, и небольшие эукариотические клетки, присутствующие в летописи окаменелостей, не обязательно были бы положительно идентифицирован.Мои коллеги в целом согласны с тем, что летопись окаменелостей дает только самую последнюю оценку того времени, когда эукариоты были уже обильный; они, возможно, были вокруг задолго до того, как они сделали его в летопись окаменелостей в узнаваемой форме.
«Другой подход состоит в том, чтобы датировать филогении, глядя на «часы»
молекулярные изменения, накапливающиеся в генетическом коде. Хотя это
подход был успешно использован для расшифровки отношений между организмами,
калибровать его для измерения времени, прошедшего с момента расхождения
филогенетические ветви проблематичны; относительно ранней эволюции
жизни, общепринятого подхода нет.Большинство попыток раннего свидания
молекулярные филогенетические деревья использовали появление эукариот (около 2,0
миллиардов лет назад) в качестве точки калибровки. Рассел
Ф. Дулиттл и его сотрудники из Калифорнийского университета в Сан-Франциско.
Недавно Диего попытался продлить калибровку в прошлое.
но эта работа оспаривается. Споры ведутся вокруг возможных случаев
горизонтальный перенос генов по филогенетическим ветвям, который был проигнорирован
этих авторов и о недостаточной поправке на множественные замены.Кроме того, эти обратные экстраполяции предполагают, что скорость молекулярного
изменения во время возникновения эукариот такие же, как и во время
эволюция многоклеточных животных, хотя на самом деле она, вероятно, была намного быстрее.
Хотя это
подход был успешно использован для расшифровки отношений между организмами,
калибровать его для измерения времени, прошедшего с момента расхождения
филогенетические ветви проблематичны; относительно ранней эволюции
жизни, общепринятого подхода нет.Большинство попыток раннего свидания
молекулярные филогенетические деревья использовали появление эукариот (около 2,0
миллиардов лет назад) в качестве точки калибровки. Рассел
Ф. Дулиттл и его сотрудники из Калифорнийского университета в Сан-Франциско.
Недавно Диего попытался продлить калибровку в прошлое.
но эта работа оспаривается. Споры ведутся вокруг возможных случаев
горизонтальный перенос генов по филогенетическим ветвям, который был проигнорирован
этих авторов и о недостаточной поправке на множественные замены.Кроме того, эти обратные экстраполяции предполагают, что скорость молекулярного
изменения во время возникновения эукариот такие же, как и во время
эволюция многоклеточных животных, хотя на самом деле она, вероятно, была намного быстрее.
«Типичная эукариотическая клетка возникла в результате симбиоза между различными прокариотических предков. Три прокариотических компонента можно проследить по сравнение молекул у современных прокариот и эукариот. Эти компоненты митохондрии (полученные из пурпурных бактерий), пластиды (из цианобактерии) и ядерно-цитоплазматический компонент (из архебактерий).Другие особенности эукариотических клеток, например цитоскелет, также могут иметь бактериальное происхождение, но до сих пор молекулярные записи не дали недвусмысленные указания на их происхождение.
«Ядерно-цитоплазматический компонент эукариотической клетки отходит очень
в начале эволюционного излучения архебактерий. Есть
активный спор о том, являются ли некоторые из архебактерий более близкими
связаны с эукариотической нуклеоцитоплазмой, чем другие (сторонники
разные взгляды — Джеймс Лейк из Калифорнийского университета в Лос-Анджелесе.
Анхелес и Карл Вёзе из Университета Иллинойса в Урбана-Шампейн).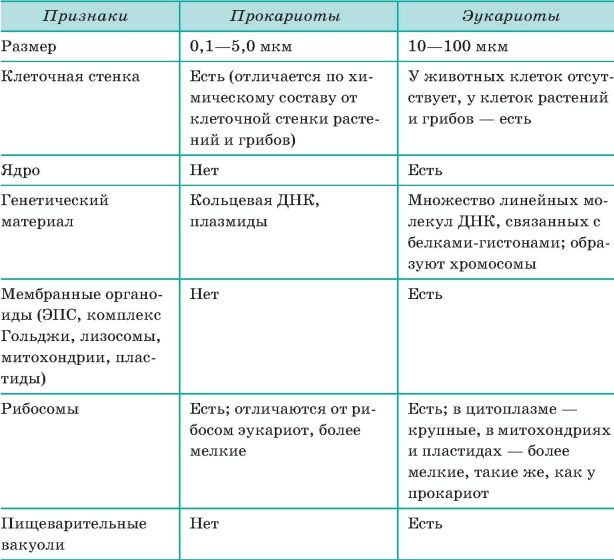 Независимо от того, как разрешится спор, предок эукариотического
нуклеоцитоплазма должна была отделиться от архебактерий на ранней стадии или
еще раньше, эпохи возникновения основных групп архебактерий. В
напротив, бактериальные предки митохондрий и пластид отделились от
линия эубактерий только после эволюционной радиации, которая породила
к основным эубактериальным царствам. Поэтому вполне вероятно, что примитив
эукариоты, лишенные митохондрий и пластид, существовали задолго до
они попали в летопись окаменелостей.Я был бы удивлен, если бы такие эукариоты
не датируется по крайней мере 3,0 миллиарда лет до настоящего времени.
Независимо от того, как разрешится спор, предок эукариотического
нуклеоцитоплазма должна была отделиться от архебактерий на ранней стадии или
еще раньше, эпохи возникновения основных групп архебактерий. В
напротив, бактериальные предки митохондрий и пластид отделились от
линия эубактерий только после эволюционной радиации, которая породила
к основным эубактериальным царствам. Поэтому вполне вероятно, что примитив
эукариоты, лишенные митохондрий и пластид, существовали задолго до
они попали в летопись окаменелостей.Я был бы удивлен, если бы такие эукариоты
не датируется по крайней мере 3,0 миллиарда лет до настоящего времени.
Происхождение эукариот – концепции биологии
Глава 13: Разнообразие микробов, грибов и простейших
Цели обучения
К концу этого раздела вы сможете:
- Описать эндосимбиотическую теорию
- Объясните происхождение митохондрий и хлоропластов
Летопись окаменелостей и генетические данные позволяют предположить, что прокариотические клетки были первыми организмами на Земле. Эти клетки возникли примерно 3,5 миллиарда лет назад, то есть примерно через 1 миллиард лет после образования Земли, и были единственными формами жизни на планете, пока примерно 2,1 миллиарда лет назад не появились эукариотические клетки. Во время правления прокариот появились фотосинтезирующие прокариоты, способные использовать энергию солнечного света для синтеза органических материалов (например, углеводов) из углекислого газа и источника электронов (например, водорода, сероводорода или воды).
Эти клетки возникли примерно 3,5 миллиарда лет назад, то есть примерно через 1 миллиард лет после образования Земли, и были единственными формами жизни на планете, пока примерно 2,1 миллиарда лет назад не появились эукариотические клетки. Во время правления прокариот появились фотосинтезирующие прокариоты, способные использовать энергию солнечного света для синтеза органических материалов (например, углеводов) из углекислого газа и источника электронов (например, водорода, сероводорода или воды).
Фотосинтез с использованием воды в качестве донора электронов потребляет углекислый газ и высвобождает молекулярный кислород (O 2 ) в качестве побочного продукта.Функционирование фотосинтезирующих бактерий на протяжении миллионов лет постепенно насыщало земную воду кислородом, а затем насыщало кислородом атмосферу, которая ранее содержала гораздо более высокие концентрации углекислого газа и гораздо более низкие концентрации кислорода. Старые анаэробные прокариоты той эпохи не могли функционировать в своей новой аэробной среде. Некоторые виды погибли, а другие выжили в оставшихся на Земле анаэробных средах. Другие ранние прокариоты развили механизмы, такие как аэробное дыхание, чтобы использовать насыщенную кислородом атмосферу, используя кислород для хранения энергии, содержащейся в органических молекулах.Аэробное дыхание — более эффективный способ получения энергии из органических молекул, что способствовало успеху этих видов (о чем свидетельствует количество и разнообразие аэробных организмов, живущих сегодня на Земле). Эволюция аэробных прокариот была важным шагом на пути эволюции первых эукариот, но также должны были развиться некоторые другие отличительные черты.
Некоторые виды погибли, а другие выжили в оставшихся на Земле анаэробных средах. Другие ранние прокариоты развили механизмы, такие как аэробное дыхание, чтобы использовать насыщенную кислородом атмосферу, используя кислород для хранения энергии, содержащейся в органических молекулах.Аэробное дыхание — более эффективный способ получения энергии из органических молекул, что способствовало успеху этих видов (о чем свидетельствует количество и разнообразие аэробных организмов, живущих сегодня на Земле). Эволюция аэробных прокариот была важным шагом на пути эволюции первых эукариот, но также должны были развиться некоторые другие отличительные черты.
Происхождение эукариотических клеток было в значительной степени загадкой, пока в 1960-х годах Линн Маргулис всесторонне не исследовала революционную гипотезу.Эндосимбиотическая теория утверждает, что эукариоты являются продуктом поглощения одной прокариотической клетки другой, одной жизни внутри другой и эволюции вместе с течением времени до тех пор, пока отдельные клетки не перестали распознаваться как таковые.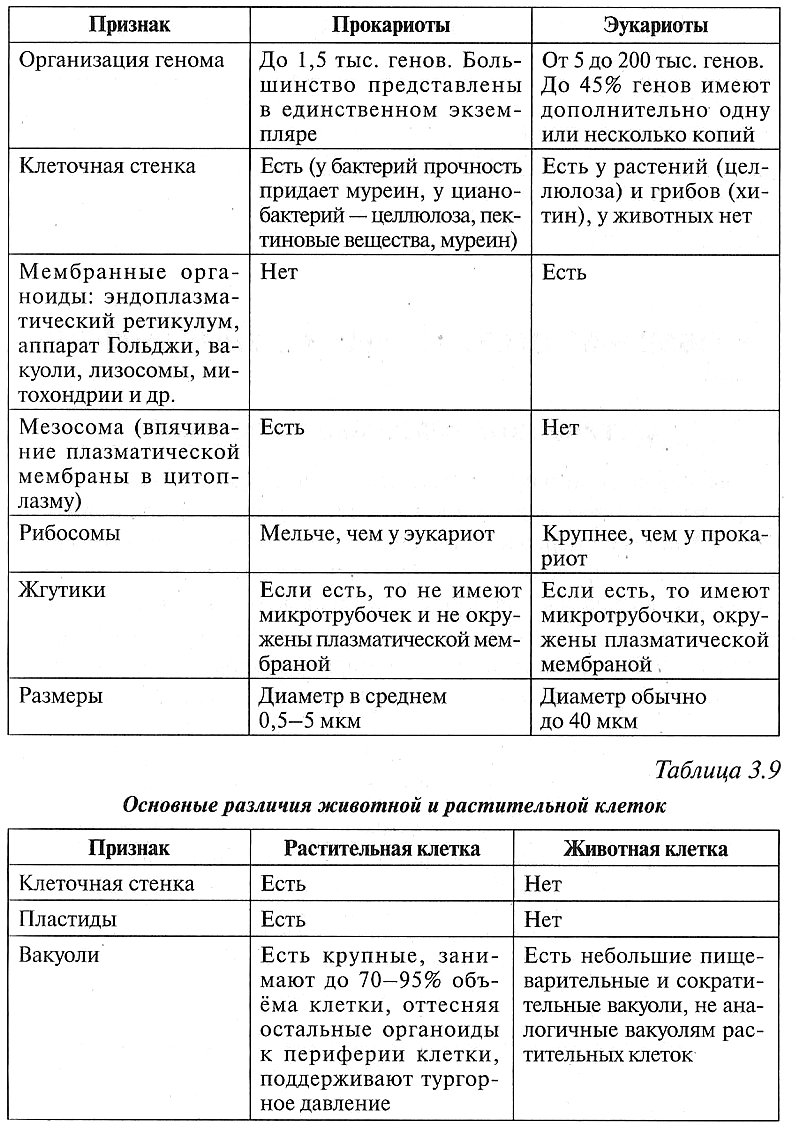 Эта когда-то революционная гипотеза сразу же стала убедительной и теперь широко принята, и работа по раскрытию шагов, вовлеченных в этот эволюционный процесс, а также ключевых игроков продолжается. Стало ясно, что многие ядерные эукариотические гены и молекулярные механизмы, ответственные за репликацию и экспрессию этих генов, по-видимому, тесно связаны с археями.С другой стороны, метаболические органеллы и гены, ответственные за многие процессы сбора энергии, произошли от бактерий. Многое еще предстоит выяснить о том, как возникли эти отношения; это продолжает оставаться захватывающей областью открытий в биологии. Несколько эндосимбиотических событий, вероятно, способствовали возникновению эукариотической клетки.
Эта когда-то революционная гипотеза сразу же стала убедительной и теперь широко принята, и работа по раскрытию шагов, вовлеченных в этот эволюционный процесс, а также ключевых игроков продолжается. Стало ясно, что многие ядерные эукариотические гены и молекулярные механизмы, ответственные за репликацию и экспрессию этих генов, по-видимому, тесно связаны с археями.С другой стороны, метаболические органеллы и гены, ответственные за многие процессы сбора энергии, произошли от бактерий. Многое еще предстоит выяснить о том, как возникли эти отношения; это продолжает оставаться захватывающей областью открытий в биологии. Несколько эндосимбиотических событий, вероятно, способствовали возникновению эукариотической клетки.
Митохондрии
Эукариотические клетки могут содержать от одной до нескольких тысяч митохондрий, в зависимости от уровня потребления энергии клеткой.Каждая митохондрия имеет длину от 1 до 10 микрометров и существует в клетке в виде движущегося, сливающегося и делящегося продолговатого сфероида ([Рисунок 1]). Однако митохондрии не могут выжить вне клетки. Поскольку атмосфера насыщалась кислородом в результате фотосинтеза и по мере эволюции успешных аэробных прокариот, данные свидетельствуют о том, что предковая клетка поглотила и поддерживала жизнь свободноживущих аэробных прокариот. Это дало клетке-хозяину возможность использовать кислород для высвобождения энергии, хранящейся в питательных веществах. Несколько линий доказательств подтверждают, что митохондрии произошли от этого эндосимбиотического события.Митохондрии имеют форму определенной группы бактерий и окружены двумя мембранами, которые возникают, когда один мембраносвязанный организм поглощается другим мембраносвязанным организмом. Внутренняя мембрана митохондрий состоит из существенных складок или крист, которые напоминают текстурированную внешнюю поверхность некоторых бактерий.
Однако митохондрии не могут выжить вне клетки. Поскольку атмосфера насыщалась кислородом в результате фотосинтеза и по мере эволюции успешных аэробных прокариот, данные свидетельствуют о том, что предковая клетка поглотила и поддерживала жизнь свободноживущих аэробных прокариот. Это дало клетке-хозяину возможность использовать кислород для высвобождения энергии, хранящейся в питательных веществах. Несколько линий доказательств подтверждают, что митохондрии произошли от этого эндосимбиотического события.Митохондрии имеют форму определенной группы бактерий и окружены двумя мембранами, которые возникают, когда один мембраносвязанный организм поглощается другим мембраносвязанным организмом. Внутренняя мембрана митохондрий состоит из существенных складок или крист, которые напоминают текстурированную внешнюю поверхность некоторых бактерий.
 (кредит: модификация работы Луизы Ховард; данные масштабной линейки от Мэтта Рассела)
(кредит: модификация работы Луизы Ховард; данные масштабной линейки от Мэтта Рассела) Митохондрии делятся сами по себе в процессе, напоминающем бинарное деление у прокариот. Митохондрии имеют собственную кольцевую ДНК-хромосому, которая несет гены, сходные с экспрессируемыми бактериями. Митохондрии также имеют специальные рибосомы и транспортные РНК, которые напоминают эти компоненты у прокариот. Все эти особенности подтверждают, что митохондрии когда-то были свободноживущими прокариотами.
Хлоропласты
Хлоропласты представляют собой один из типов пластид, группу родственных органелл в растительных клетках, которые участвуют в хранении крахмалов, жиров, белков и пигментов.Хлоропласты содержат зеленый пигмент хлорофилл и играют роль в фотосинтезе. Генетические и морфологические исследования предполагают, что пластиды произошли от эндосимбиоза предковой клетки, поглотившей фотосинтетическую цианобактерию. Пластиды похожи по размеру и форме на цианобактерии и окружены двумя или более мембранами, соответствующими внутренней и внешней мембранам цианобактерий.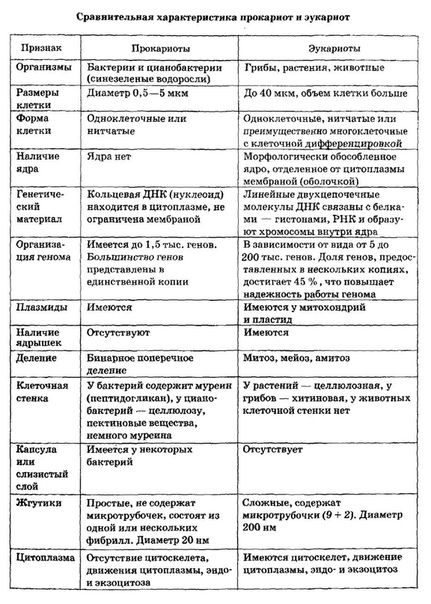 Подобно митохондриям, пластиды также содержат кольцевые геномы и делятся процессом, напоминающим деление прокариотических клеток.Хлоропласты красных и зеленых водорослей демонстрируют последовательности ДНК, которые тесно связаны с фотосинтезирующими цианобактериями, что позволяет предположить, что красные и зеленые водоросли являются прямыми потомками этого эндосимбиотического события.
Подобно митохондриям, пластиды также содержат кольцевые геномы и делятся процессом, напоминающим деление прокариотических клеток.Хлоропласты красных и зеленых водорослей демонстрируют последовательности ДНК, которые тесно связаны с фотосинтезирующими цианобактериями, что позволяет предположить, что красные и зеленые водоросли являются прямыми потомками этого эндосимбиотического события.
Митохондрии, вероятно, эволюционировали раньше пластид, потому что все эукариоты имеют либо функциональные митохондрии, либо митохондриоподобные органеллы. Напротив, пластиды встречаются только у подмножества эукариот, таких как наземные растения и водоросли. Одна из гипотез об эволюционных шагах, ведущих к появлению первого эукариота, резюмирована на [Рис. 2].
Рисунок 2: Первый эукариот, возможно, произошел от предка прокариот, который претерпел мембранную пролиферацию, разделение клеточных функций (на ядро, лизосомы и эндоплазматический ретикулум) и установление эндосимбиотических отношений с аэробными прокариотами и, в некоторых случаях, случаях фотосинтезирующий прокариот с образованием митохондрий и хлоропластов соответственно.
Точные шаги, ведущие к первой эукариотической клетке, можно только предполагать, и существуют некоторые разногласия относительно того, какие события действительно произошли и в каком порядке.Было высказано предположение, что бактерии спирохет дали начало микротрубочкам, а жгутиковые прокариоты, возможно, предоставили сырье для эукариотических жгутиков и ресничек. Другие ученые предполагают, что пролиферация и компартментализация мембран, а не эндосимбиотические события, привели к развитию митохондрий и пластид. Однако подавляющее большинство исследований поддерживает эндосимбиотическую гипотезу эукариотической эволюции.
Ранние эукариоты были одноклеточными, как и большинство современных протистов, но по мере того, как эукариоты становились более сложными, эволюция многоклеточности позволяла клеткам оставаться маленькими, но при этом выполнять специализированные функции.Считается, что предки современных многоклеточных эукариот появились примерно 1,5 миллиарда лет назад.
Первые эукариоты произошли от предковых прокариот в результате процесса, который включал пролиферацию мембран, потерю клеточной стенки, эволюцию цитоскелета, а также приобретение и эволюцию органелл.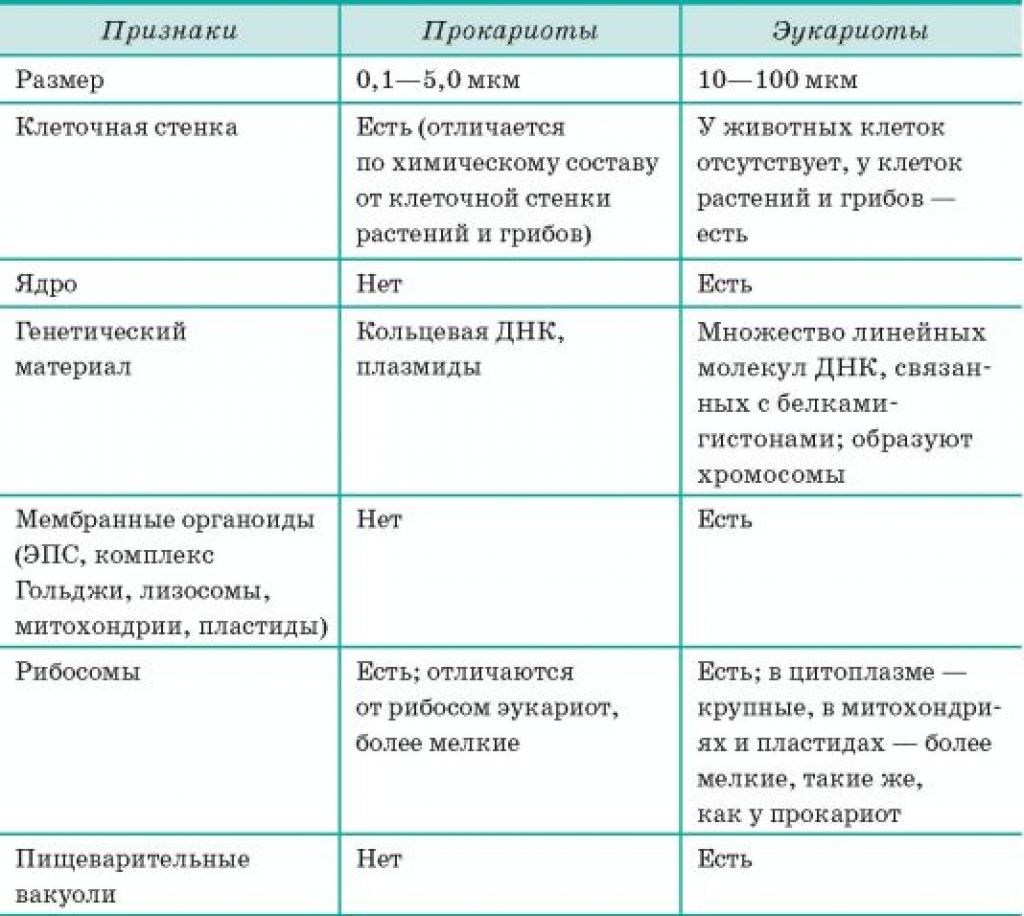 Ядерные эукариотические гены, по-видимому, произошли от архей, тогда как энергетический аппарат эукариотических клеток, по-видимому, имеет бактериальное происхождение. Митохондрии и пластиды возникли в результате эндосимбиотических событий, когда клетки предков поглотили аэробную бактерию (в случае митохондрий) и фотосинтетическую бактерию (в случае хлоропластов).Эволюция митохондрий, вероятно, предшествовала эволюции хлоропластов. Имеются данные о вторичных эндосимбиотических событиях, в которых пластиды кажутся результатом эндосимбиоза после предшествующего эндосимбиотического события.
Ядерные эукариотические гены, по-видимому, произошли от архей, тогда как энергетический аппарат эукариотических клеток, по-видимому, имеет бактериальное происхождение. Митохондрии и пластиды возникли в результате эндосимбиотических событий, когда клетки предков поглотили аэробную бактерию (в случае митохондрий) и фотосинтетическую бактерию (в случае хлоропластов).Эволюция митохондрий, вероятно, предшествовала эволюции хлоропластов. Имеются данные о вторичных эндосимбиотических событиях, в которых пластиды кажутся результатом эндосимбиоза после предшествующего эндосимбиотического события.
Какое событие, как считается, способствовало эволюции эукариот?
- глобальное потепление
- оледенение
- вулканическая активность
- оксигенация атмосферы
[reveal-answer q=»371406″]Показать ответ[/reveal-answer]
[hidden-answer a=»371406″]4[/hidden-answer]
Митохондрии, скорее всего, произошли от _____________.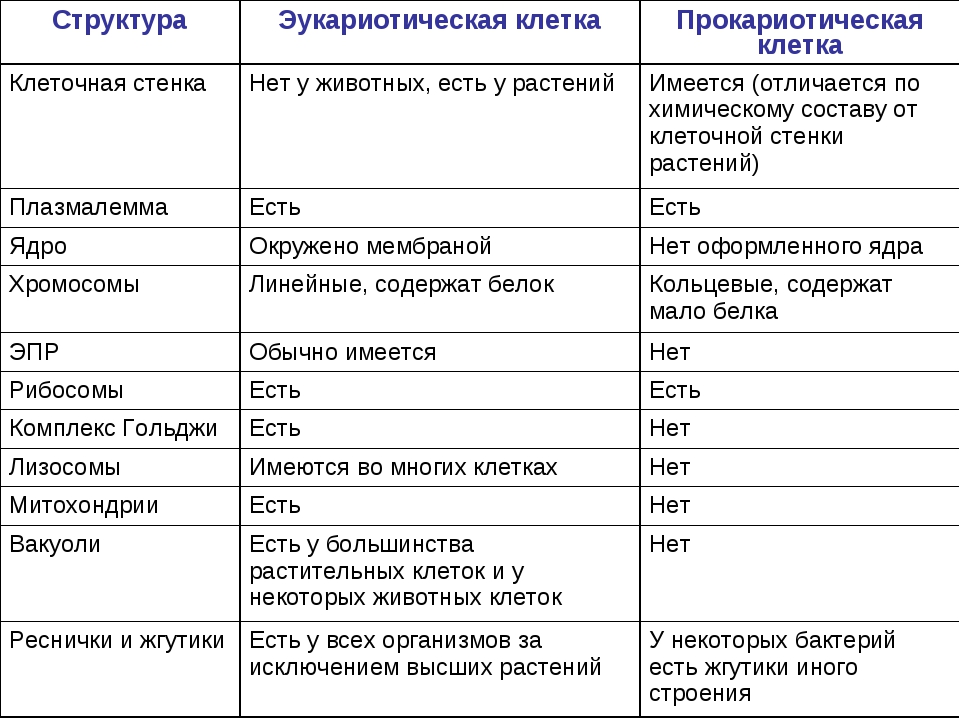
- фотосинтетическая цианобактерия
- элементы цитоскелета
- аэробные бактерии
- мембранная пролиферация
[reveal-answer q=»261309″]Показать ответ[/reveal-answer]
[hidden-answer a=»261309″]3[/hidden-answer]
Опишите предполагаемые этапы происхождения эукариотических клеток.
Эукариотические клетки возникли в результате эндосимбиотических событий, которые дали начало производящим энергию органеллам внутри эукариотических клеток, таким как митохондрии и пластиды.Ядерный геном эукариот наиболее тесно связан с археями, поэтому, возможно, это были ранние археи, поглотившие бактериальную клетку, которая превратилась в митохондрию. Митохондрии, по-видимому, произошли от альфа-протеобактерий, тогда как хлоропласты произошли от цианобактерий. Имеются также свидетельства вторичных эндосимбиотических событий. Другие клеточные компоненты могли возникнуть в результате эндосимбиотических событий.
Глоссарий
- эндосимбиоз 90–108 поглощение одной клетки другой таким образом, что поглощенная клетка выживает, и обе клетки извлекают выгоду; процесс, ответственный за эволюцию митохондрий и хлоропластов у эукариот
- пластид
- один из группы родственных органелл в растительных клетках, которые участвуют в хранении крахмалов, жиров, белков и пигментов
Эукариотические клетки – обзор
4 Эукариотические клетки подразделяются на компартменты
Геном эукариотической клетки находится внутри отдельного компартмента – ядра.На самом деле эукариотические клетки имеют несколько внутренних клеточных компартментов, окруженных мембранами. Само ядро окружено двойной мембраной, ядерной оболочкой , которая отделяет ядро от цитоплазмы, но обеспечивает некоторую связь с цитоплазмой через ядерные поры (рис.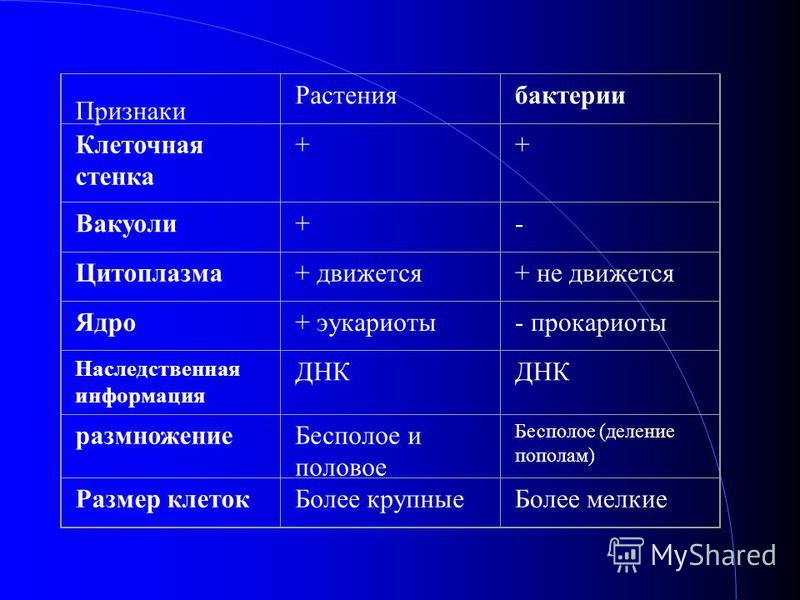 1.09). Геном эукариот состоит из 10 000–50 000 генов, находящихся на нескольких хромосомах. Хромосомы эукариот линейны, в отличие от кольцевых хромосом бактерий. Большинство эукариот диплоидны, с двумя копиями каждой хромосомы.Следовательно, они обладают по крайней мере двумя копиями каждого гена. Кроме того, эукариотические клетки часто имеют множественные копии определенных генов в результате дупликации генов.
1.09). Геном эукариот состоит из 10 000–50 000 генов, находящихся на нескольких хромосомах. Хромосомы эукариот линейны, в отличие от кольцевых хромосом бактерий. Большинство эукариот диплоидны, с двумя копиями каждой хромосомы.Следовательно, они обладают по крайней мере двумя копиями каждого гена. Кроме того, эукариотические клетки часто имеют множественные копии определенных генов в результате дупликации генов.
Рисунок 1.09. Эукариот имеет несколько клеточных компартментов
Просвечивающая электронная микрофотография плазматической клетки из костного мозга в искусственных цветах. Множественные компартменты, окруженные мембранами, включая ядро, обнаружены в эукариотических клетках. Для плазматических клеток характерно расположение гетерохроматина (оранжевого цвета) в ядре, где он прилегает к внутренней ядерной мембране.Также типична сеть шероховатого эндоплазматического ретикулума ( желтых пунктирных линий ) в цитоплазме. Овальные или округлые малиновые образования в цитоплазме — митохондрии (увеличение × 4500).
Эукариоты обладают множеством других органелл . Это субклеточные структуры, выполняющие определенные задачи. Одни отделены от остальной клетки мембранами (так называемые 90–105 мембраносвязанные органеллы 90–106 ), а другие (например, 90–106).г., рибосомы) не являются. Эндоплазматический ретикулум представляет собой мембранную систему, которая является продолжением ядерной оболочки и пронизывает цитоплазму. Аппарат Гольджи представляет собой стопку уплощенных мембранных мешочков и связанных с ними пузырьков, которые участвуют в секреции белков или других материалов наружу клетки. Лизосомы представляют собой связанные с мембраной структуры, содержащие ферменты деградации и специализированные для пищеварения.
Все эукариоты, за исключением очень немногих, содержат митохондрий (единственное число, митохондрия; рис.1.10). Обычно это палочковидные органеллы, ограниченные двойной мембраной. По своим размерам и форме они напоминают бактерии. Как будет обсуждаться более подробно (см. главу 4: Гены, геномы и ДНК), считается, что митохондрии действительно произошли от бактерий, которые поселились у первобытных предков эукариотических клеток. Как и бактерии, каждая митохондрия содержит кольцевую молекулу ДНК. Митохондриальный геном похож на бактериальную хромосому, но намного меньше.В митохондриальной ДНК есть несколько генов, необходимых для функционирования митохондрий. Митохондрии также имеют рибосомы, которые по структуре напоминают типичные прокариотические рибосомы.
По своим размерам и форме они напоминают бактерии. Как будет обсуждаться более подробно (см. главу 4: Гены, геномы и ДНК), считается, что митохондрии действительно произошли от бактерий, которые поселились у первобытных предков эукариотических клеток. Как и бактерии, каждая митохондрия содержит кольцевую молекулу ДНК. Митохондриальный геном похож на бактериальную хромосому, но намного меньше.В митохондриальной ДНК есть несколько генов, необходимых для функционирования митохондрий. Митохондрии также имеют рибосомы, которые по структуре напоминают типичные прокариотические рибосомы.
Рисунок 1.10. Митохондрия
Митохондрия окружена двумя концентрическими мембранами. Внутренняя мембрана загибается внутрь, образуя крист , которые содержат белки дыхательной цепи, генерирующие энергию для клетки.
Митохондрии специализируются на выработке энергии при дыхании и встречаются у всех эукариот.(Известно несколько эукариот, которые не могут дышать; тем не менее, они сохраняют остатки митохондриальных органелл — обсуждается позже. ) У эукариот ферменты дыхания расположены на внутренней митохондриальной мембране, которая имеет многочисленные изгибы, увеличивающие площадь мембраны. Это контрастирует с бактериями, у которых дыхательная цепь расположена в цитоплазматической мембране, поскольку митохондрии отсутствуют.
) У эукариот ферменты дыхания расположены на внутренней митохондриальной мембране, которая имеет многочисленные изгибы, увеличивающие площадь мембраны. Это контрастирует с бактериями, у которых дыхательная цепь расположена в цитоплазматической мембране, поскольку митохондрии отсутствуют.
Хлоропласты — мембраносвязанные органеллы, специализированные для фотосинтеза (рис. 1.11). Они обнаружены только у растений и некоторых одноклеточных эукариот. Они имеют форму от овальной до палочковидной и содержат сложные наборы внутренних мембран, которые содержат зеленый светопоглощающий пигмент хлорофилл и другие компоненты, необходимые для улавливания световой энергии. Как и митохондрии, хлоропласты содержат кольцевую молекулу ДНК и собственные рибосомы и, как полагают, произошли от фотосинтезирующей бактерии.
Рисунок 1.11. Хлоропласт
Хлоропласт связан двойной мембраной и содержит свернутые стопки мембран, предназначенных для фотосинтеза.Хлоропласт также содержит рибосомы и ДНК.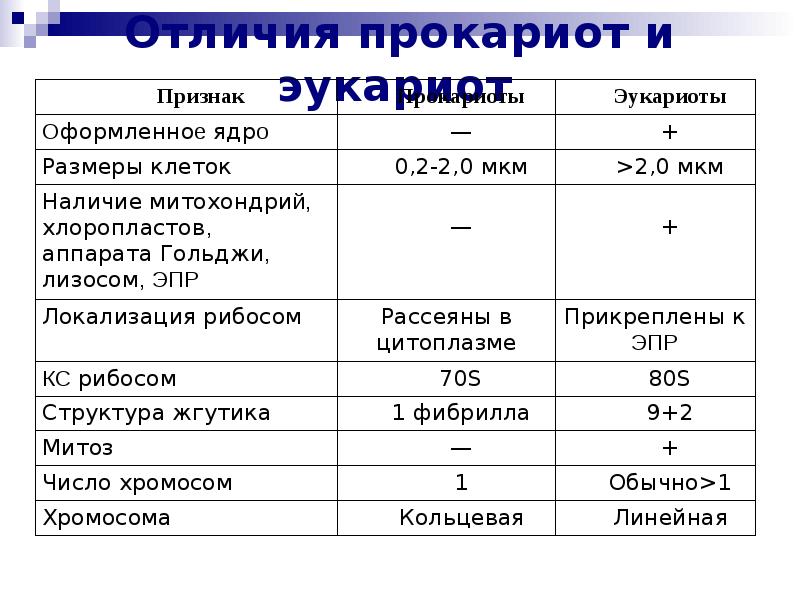
Эукариотические клетки имеют разветвленную внутриклеточную архитектуру для поддержания своей формы и перемещения материалов и органелл вокруг клеток. Цитоскелет представляет собой сложную сеть филаментов, состоящую из белков, таких как актин , винкулин и фибронектин (рис. 1.12). Помимо поддержания формы клетки, цитоскелет важен для клеточного транспорта. Например, цитоскелетные волокна проходят через длинные аксоны нейронов, а везикулы, заполненные нейротрансмиттерами, перемещаются вверх и вниз по аксону, облегчая связь между ядром и нервными волокнами.Цитоскелет также инициирует клеточные движения. Увеличивая длину волокон на одной стороне клетки и уменьшая их длину на противоположной стороне, клетка может физически двигаться. Это особенно верно для более мелких одноклеточных эукариот и для движений в процессе развития многоклеточного организма. Наконец, эти движения цитоскелета важны для таких процессов, как деление клеток, поскольку те же самые волокна составляют веретено.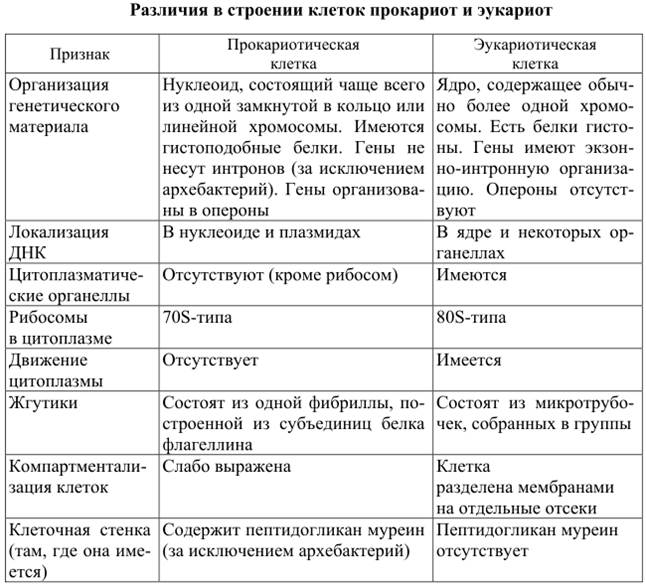
Рисунок 1.12. Цитоскелет
Актин, винкулин и фибронектин — это три белка цитоскелета, которые придают этой клетке уплощенный край.Этот край имеет спайки, которые соединяют клетку с чашкой in vitro , но служат для удержания клетки прикрепленной к другим клеткам в органах многоклеточного организма.
(Источник: Байрон и др. (2010) Curr Biol 20 (24): R1063–R1067.)Жизнь модульна. Сложные организмы подразделяются на органы. Крупные и сложные клетки делятся на органеллы.
Эукариоты имеют множество связанных с мембраной органелл, выполняющих такие функции, как дыхание (митохондрии), расщепление ферментов (лизосомы), процессинг и секреция белков (аппарат Гольджи и эндоплазматический ретикулум).
Эукариотические клетки имеют внутренние структурные элементы, называемые цитоскелетом.
Доказательства синцитиального происхождения эукариот из реконструкции государства предков | Геномная биология и эволюция
 Одноядерное состояние последнего эукариотического общего предка (LECA) редко, если вообще когда-либо, подвергается сомнению, даже несмотря на то, что клетки, содержащие множественные (синцитии, ценоциты и поликарионы), удивительно распространены среди эукариотических супергрупп. Здесь мы представляем обзор многоядерных форм. Реконструкция состояния предковых признаков для представителей 106 эукариотических таксонов с использованием 16 различных возможных корней и сестринских отношений супергруппы указывает на то, что LECA, помимо того, что он был митохондриальным, половым и мейотическим, был многоядерным.LECA демонстрировал закрытый митоз, что является правилом для современных синцитиальных форм, что проливает свет на механику его хромосомной сегрегации. Простая математическая модель показывает, что в многоядерном цитозоле LECA отношения между митохондриями и ядрами не были ни один-к-одному, ни один-ко-многим, а многие-ко-многим, что ставит митоядерные взаимодействия и цитоядерную совместимость в эволюционную основу эукариотических клеточное происхождение.
Одноядерное состояние последнего эукариотического общего предка (LECA) редко, если вообще когда-либо, подвергается сомнению, даже несмотря на то, что клетки, содержащие множественные (синцитии, ценоциты и поликарионы), удивительно распространены среди эукариотических супергрупп. Здесь мы представляем обзор многоядерных форм. Реконструкция состояния предковых признаков для представителей 106 эукариотических таксонов с использованием 16 различных возможных корней и сестринских отношений супергруппы указывает на то, что LECA, помимо того, что он был митохондриальным, половым и мейотическим, был многоядерным.LECA демонстрировал закрытый митоз, что является правилом для современных синцитиальных форм, что проливает свет на механику его хромосомной сегрегации. Простая математическая модель показывает, что в многоядерном цитозоле LECA отношения между митохондриями и ядрами не были ни один-к-одному, ни один-ко-многим, а многие-ко-многим, что ставит митоядерные взаимодействия и цитоядерную совместимость в эволюционную основу эукариотических клеточное происхождение.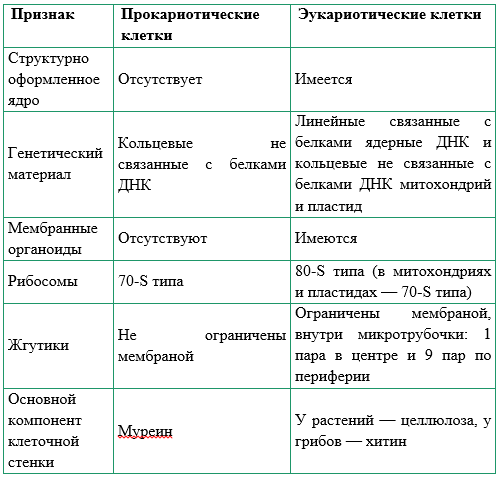 Внутри синцития отдельные ядра и отдельные митохондрии функционируют как начальные эволюционные единицы более низкого уровня отбора, в отличие от отдельных клеток, во время эукариогенеза.Ядра внутри синцития спасают друг друга от летальных мутаций, тем самым откладывая отбор жизнеспособных ядер и цитоядерной совместимости до генерации спор, буферизуя переходные узкие места в эукариогенезе. Традиционно считается, что переход от прокариот к эукариотам не оставил промежуточных звеньев, однако, если эукариогенез происходил через синцитиального общего предка, то промежуточные формы сохранились до настоящего времени на всем эукариотическом древе в виде синцитиев, но до сих пор оставались нераспознанными.
Внутри синцития отдельные ядра и отдельные митохондрии функционируют как начальные эволюционные единицы более низкого уровня отбора, в отличие от отдельных клеток, во время эукариогенеза.Ядра внутри синцития спасают друг друга от летальных мутаций, тем самым откладывая отбор жизнеспособных ядер и цитоядерной совместимости до генерации спор, буферизуя переходные узкие места в эукариогенезе. Традиционно считается, что переход от прокариот к эукариотам не оставил промежуточных звеньев, однако, если эукариогенез происходил через синцитиального общего предка, то промежуточные формы сохранились до настоящего времени на всем эукариотическом древе в виде синцитиев, но до сих пор оставались нераспознанными.
Переход от прокариот к эукариотам сопровождался эндосимбиозом и резким увеличением сложности внутриклеточных клеток. В то время как большинство теорий эукариогенеза рассматривают и иллюстрируют последнего эукариотического общего предка (LECA) как одноядерную, половую, жгутиковую популяцию клеток, происхождение скоординированного деления ядер и органелл, связанного с клеточным циклом, редко обсуждается. Используя реконструкцию наследственного состояния, мы показываем, что LECA, скорее всего, включал многоядерную стадию, которая также допускала посредничество в конфликте между митохондриальным и ядерным геномами, вызванное эндосимбиотическим переносом генов.Почти повсеместное присутствие синцитиальной стадии жизни во всех основных группах эукариот предполагает, что многоядерный LECA является жизнеспособным промежуточным продуктом, который позволил проводить внутриклеточные эксперименты и эволюцию сложных эукариотических процессов, которые мы наблюдаем сегодня.
Используя реконструкцию наследственного состояния, мы показываем, что LECA, скорее всего, включал многоядерную стадию, которая также допускала посредничество в конфликте между митохондриальным и ядерным геномами, вызванное эндосимбиотическим переносом генов.Почти повсеместное присутствие синцитиальной стадии жизни во всех основных группах эукариот предполагает, что многоядерный LECA является жизнеспособным промежуточным продуктом, который позволил проводить внутриклеточные эксперименты и эволюцию сложных эукариотических процессов, которые мы наблюдаем сегодня.
 2016). В летописи окаменелостей они появляются примерно на 2 миллиарда лет позже, чем прокариоты (Javaux et al. 2001; Javaux and Lepot 2018). Среди специалистов существует консенсус в отношении того, что эукариоты произошли от прокариот, но вопрос о том, как они произошли от прокариот, активно обсуждается. Все современные теории происхождения эукариот так или иначе влекут за собой концепцию симбиогенеза (Mereschkowsky 1910; английский перевод Kowallik and Martin 2021), поскольку митохондрии прослеживаются до последнего общего предка эукариот LECA (Embley and Martin 2006; Tria et al.2021), и нет надежного способа объяснить структуру, ДНК и биоэнергетические свойства митохондрий (и хлоропластов) без их эндосимбиотического происхождения. Различия между современными теориями происхождения эукариот (рассмотренные в Martin et al. 2015; López-García and Moreira 2015; Dacks et al. 2016) в основном касаются предположений о биологической природе и клеточной сложности хозяина, который приобрел митохондрию.
2016). В летописи окаменелостей они появляются примерно на 2 миллиарда лет позже, чем прокариоты (Javaux et al. 2001; Javaux and Lepot 2018). Среди специалистов существует консенсус в отношении того, что эукариоты произошли от прокариот, но вопрос о том, как они произошли от прокариот, активно обсуждается. Все современные теории происхождения эукариот так или иначе влекут за собой концепцию симбиогенеза (Mereschkowsky 1910; английский перевод Kowallik and Martin 2021), поскольку митохондрии прослеживаются до последнего общего предка эукариот LECA (Embley and Martin 2006; Tria et al.2021), и нет надежного способа объяснить структуру, ДНК и биоэнергетические свойства митохондрий (и хлоропластов) без их эндосимбиотического происхождения. Различия между современными теориями происхождения эукариот (рассмотренные в Martin et al. 2015; López-García and Moreira 2015; Dacks et al. 2016) в основном касаются предположений о биологической природе и клеточной сложности хозяина, который приобрел митохондрию.
В симбиогенных теориях предполагается, что хозяин является типичным археоном с точки зрения его клеточной сложности, с происхождением митохондрий, вызывающих генетические, клеточные биологические и биоэнергетические изменения в консорциуме хозяин-симбионт, которые в конечном итоге привели к LECA (Мартин и Мюллер). 1998; Лейн и Мартин 2012; Гулд и др.2016; Имачи и др. 2020). В градуалистских теориях предполагается, что хозяин является потомком архейной линии, которая, однако, преодолела порог сложности от прокариотической к эукариотической клетке посредством эволюционных механизмов, отличных от симбиоза, тем самым преодолев разрыв между прокариотической и эукариотической сложностью (Martijn and Ettema 2013; Spang et al., 2015) до возникновения митохондрий, что, следовательно, мало повлияло на сложность эукариот. В гибридных теориях переход от прокариот к эукариотам включал один или несколько дополнительных симбиозов, которые предшествовали возникновению митохондрий, таких как жгутики (Sagan 1967), пероксисомы (de Duve 1969), ядро (López-García and Moreira 2020) или ER (Gupta and Golding, 1996) или был вызван латеральным переносом генов (LGT) в линию хозяина, так что многие характерные черты эукариот происходят от генов, которые были изобретены в чужих линиях и переданы в LECA через LGT (Pittis and Gabaldón). 2016; Воссеберг и др.2021), хотя методы, лежащие в основе таких заявлений, были поставлены под сомнение (Martin et al., 2017a; Tria et al., 2021; Nagies et al., 2020). Градуалистские и гибридные теории обычно постулируют происхождение фаготрофного питания в линии архей-хозяев до возникновения митохондрий (Doolittle, 1998; Spang et al., 2015; Zaremba-Niedzwiedzka et al., 2017; Vosseberg et al. 2021), что однако это очень проблематичное предположение с физиологической точки зрения (Martin et al. 2017b) и расходится с данными из летописи микрофоссилий, указывающими на позднее происхождение фагоцитоза (Mills, 2020).Эукариоты, несомненно, являются генетическими химерами, причем большинство эукариотических генов происходит от бактерий, а не от архей (Brueckner and Martin 2020), при этом бактериальные гены у эукариот восходят к LECA, а не к наследственно-специфическим приобретениям в ходе эукариотической эволюции (Nagies et al., 2020).
2016; Воссеберг и др.2021), хотя методы, лежащие в основе таких заявлений, были поставлены под сомнение (Martin et al., 2017a; Tria et al., 2021; Nagies et al., 2020). Градуалистские и гибридные теории обычно постулируют происхождение фаготрофного питания в линии архей-хозяев до возникновения митохондрий (Doolittle, 1998; Spang et al., 2015; Zaremba-Niedzwiedzka et al., 2017; Vosseberg et al. 2021), что однако это очень проблематичное предположение с физиологической точки зрения (Martin et al. 2017b) и расходится с данными из летописи микрофоссилий, указывающими на позднее происхождение фагоцитоза (Mills, 2020).Эукариоты, несомненно, являются генетическими химерами, причем большинство эукариотических генов происходит от бактерий, а не от архей (Brueckner and Martin 2020), при этом бактериальные гены у эукариот восходят к LECA, а не к наследственно-специфическим приобретениям в ходе эукариотической эволюции (Nagies et al., 2020).
Несмотря на их разнообразие и различные предпосылки, теории происхождения эукариот единообразно предполагают, обычно неявно, что LECA был одноклеточным и одноядерным (Gould and Dring 1979; Cavalier-Smith 1987; Lake and Rivera 1994; Gupta and Golding 1996; Horiike и другие. , 2004; Имачи и др., 2020 г.; Мартин и Эттема, 2013 г.; Martin et al., 2015), предположение, которое почти никогда не подвергалось сомнению (Garg and Martin 2016). Единообразие взглядов на одноядерную природу LECA настолько распространено, что принимается как данность, то есть редко, если вообще когда-либо, упоминается в качестве предположения. Более того, теории происхождения эукариот, если они вообще проиллюстрированы схематической диаграммой, неизменно представляют LECA как одноядерную клетку. Такие изображения часто носят символический характер, изображая признаки, а не живые клетки, но в то же время они влияют на то, как мы осмысляем проблему происхождения эукариот.Модели эукариогенеза, в которых митохондрии играют механистическую роль, обычно подразумевают отношения «один к одному» или «многие к одному» (Lane and Martin, 2012) между митохондриями и ядром, в силу чего природа ядерной динамики LECA, гетерогенность ядер в LECA , его координация ядерного деления с клеточным делением, его клеточный цикл (мейотический или митотический) и эволюционная последовательность, связывающая деление органелл, ядерное и клеточное деление, редко обсуждаются (Cavalier-Smith 2010; Garg and Martin 2016).
, 2004; Имачи и др., 2020 г.; Мартин и Эттема, 2013 г.; Martin et al., 2015), предположение, которое почти никогда не подвергалось сомнению (Garg and Martin 2016). Единообразие взглядов на одноядерную природу LECA настолько распространено, что принимается как данность, то есть редко, если вообще когда-либо, упоминается в качестве предположения. Более того, теории происхождения эукариот, если они вообще проиллюстрированы схематической диаграммой, неизменно представляют LECA как одноядерную клетку. Такие изображения часто носят символический характер, изображая признаки, а не живые клетки, но в то же время они влияют на то, как мы осмысляем проблему происхождения эукариот.Модели эукариогенеза, в которых митохондрии играют механистическую роль, обычно подразумевают отношения «один к одному» или «многие к одному» (Lane and Martin, 2012) между митохондриями и ядром, в силу чего природа ядерной динамики LECA, гетерогенность ядер в LECA , его координация ядерного деления с клеточным делением, его клеточный цикл (мейотический или митотический) и эволюционная последовательность, связывающая деление органелл, ядерное и клеточное деление, редко обсуждаются (Cavalier-Smith 2010; Garg and Martin 2016).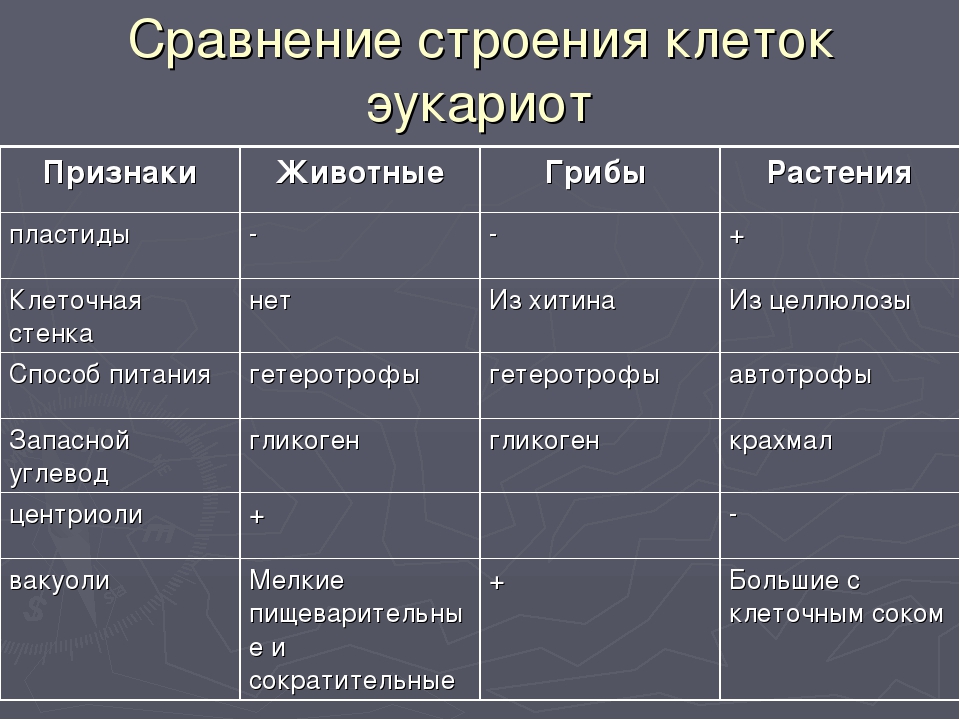
Чем интересна возможность многоядерного состояния LECA? Основное эволюционное преимущество, которое многоядерное состояние дало бы LECA, очевидно: генные мутации или даже серьезные хромосомные мутации, включая анеуплоидии, которые в противном случае были бы летальными в одноядерной клетке, могут быть дополнены мРНК из других ядер в том же цитозоле, что обеспечивает выживание. (многоядерный) индивидуум как набор гетерогенных ядер, стабильная отправная точка, из которой могут развиваться бесчисленные различия между прокариотической и эукариотической сегрегацией хромосом и обращением в клеточных делениях (Garg and Martin 2016).Таким образом, многоядерное состояние должно буферизовать переход от прокариотического к эукариотическому делению хромосом и, кроме того, отделить его от эволюционного препятствия, связанного с преодолением перехода от прокариотического к эукариотическому клеточному делению, а также от прокариотической к эукариотической организации хроматина во время клеточного цикла (Бранк и Мартин 2019).
Наличие многоядерных таксонов было зарегистрировано у членов всех эукариотических супергрупп и у многочисленных высших таксонов, как древних, так и производных (Archibald et al.2017; Адл и др. 2019; см. дополнительную таблицу 3, Дополнительный материал онлайн). Хорошо известные примеры многоядерных форм встречаются в надгруппе амебозойных: миксомицеты (миксогастридные амебы), протоспорангииды, диктиостелиды, вампиреллиды и шизоплазмодиды (рис. 1). Грибы, пожалуй, самые распространенные ценоциты на Земле, причем большинство классов и порядков имеют многоядерных представителей, а одноклеточные формы, как правило, редки и часто имеют вторичное происхождение (Kiss et al.2019). Помимо грибов, внутри опистоконтов многоядерны также ядерные амебы (Dirren, Posch 2016) и ихтиоспореи, а среди животных очень хорошо известны синцитии, например, тело гексактинеллидных губок (Leys 2003), мышцы всех остальных животных, и личинки голометаболических насекомых, в том числе Drosophila . Более того, давно предполагалось, что общий предок Metazoa мог быть многоядерным (Hadži 1953). В Rhizaria, самой глубоководной ветви ЮАР, есть многочисленные примеры многоядерных представителей (наиболее примечательным является Xenophyophorea).Кроме того, Opalinata и Apicomplexa также имеют многоядерные формы в рамках своего жизненного цикла (Archibald et al. 2017; Adl et al. 2019). Среди гетеротрофных эукариот обнаружены не только синцитии, но и многочисленные примеры многоядерных водорослей, как красных ( Florideophyceae ), так и зеленых ( Ulvophyceae ), а также различных многоядерных тканей наземных растений (Niklas et al. 2013). Многоядерные формы также встречаются среди эукариот со вторичными пластидами, например, у Chlorarachniophyceae , Phaeophyceae и Xanthophyceae (Niklas et al.2013). Распространение и эволюция многоядерных тканей среди эукариот с пластидами обнаруживает большое разнообразие форм в 60 семействах архепластид и пяти различных линиях водорослей (Niklas et al.
Более того, давно предполагалось, что общий предок Metazoa мог быть многоядерным (Hadži 1953). В Rhizaria, самой глубоководной ветви ЮАР, есть многочисленные примеры многоядерных представителей (наиболее примечательным является Xenophyophorea).Кроме того, Opalinata и Apicomplexa также имеют многоядерные формы в рамках своего жизненного цикла (Archibald et al. 2017; Adl et al. 2019). Среди гетеротрофных эукариот обнаружены не только синцитии, но и многочисленные примеры многоядерных водорослей, как красных ( Florideophyceae ), так и зеленых ( Ulvophyceae ), а также различных многоядерных тканей наземных растений (Niklas et al. 2013). Многоядерные формы также встречаются среди эукариот со вторичными пластидами, например, у Chlorarachniophyceae , Phaeophyceae и Xanthophyceae (Niklas et al.2013). Распространение и эволюция многоядерных тканей среди эукариот с пластидами обнаруживает большое разнообразие форм в 60 семействах архепластид и пяти различных линиях водорослей (Niklas et al. 2013).
2013).
Рис. 1.
Представление разнообразия групп, содержащих многоядерных представителей. (A) Фораминиферы: Filosa, глубоководный ценоцитарный ксенофиофор; (B) Endomyxa: Lateromyxa gallica , многоядерная хищная амеба; (C) Tubulinea: Chaos sp.многоядерная амеба; (D) Mesomycetozoea: Sphaeroforma arctica , ценоцит с синими ядрами; (E) Protostomia: Drosophila melanogaster , многоядерный эмбрион; (F) Hexactinellida: Euplectella aspergillum ценоцитарная гексактинеллидная губка; (G) Дейтеростомия: многоядерные мышечные клетки мыши; (H) Heterolobosea: Acrasis rosea , плодовое тело; (I) Chloroplastida: Ulvophyceae: Cladophora sp. сифонный таллом; (J) — Rhodophyta: Florideophyceae: Lithophyllum sp.; (K) — миксомицеты: многоядерный плазмодий члена Physaraceae ; (L) Ascomycota: Eremothecium gossypii , асептированные гифы. Авторы фотографий и общий домен Creative Commons (CC): A и F.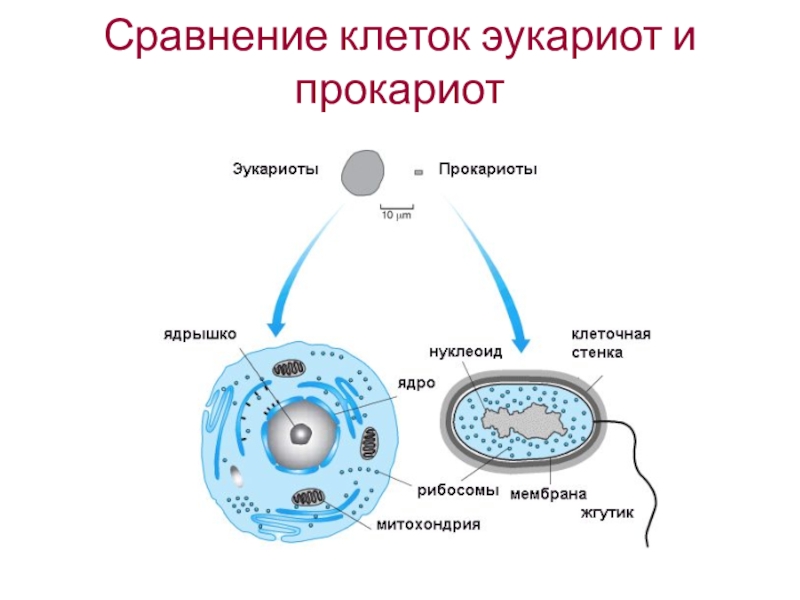 NOAA, общественное достояние; Б. Норберт Хюльсманн, BY-NC-SA 2.0; С. и И. Proyecto Agua, BY-NC-SA 2.0; D. Лаборатория мультиклеточного генома, BY 2.0; Э. Билли Лиар, BY-NC-SA 2.0; Г. Кевин А. Мурах, Галерея изображений NIH, BY-CN 2.0; Х. Ширли Чио, Лаборатория биологии грибов, Калифорнийский университет в Беркли, Калифорния, BY-SA 3.0; Дж.Кристоф Куинтин, BY-NC 2.0; К. Андре Амарал, распространяется под лицензией CC BY-NC 4.0; Лаборатория Л. Джасперсена, общественное достояние. Масштабная линейка является приблизительной.
NOAA, общественное достояние; Б. Норберт Хюльсманн, BY-NC-SA 2.0; С. и И. Proyecto Agua, BY-NC-SA 2.0; D. Лаборатория мультиклеточного генома, BY 2.0; Э. Билли Лиар, BY-NC-SA 2.0; Г. Кевин А. Мурах, Галерея изображений NIH, BY-CN 2.0; Х. Ширли Чио, Лаборатория биологии грибов, Калифорнийский университет в Беркли, Калифорния, BY-SA 3.0; Дж.Кристоф Куинтин, BY-NC 2.0; К. Андре Амарал, распространяется под лицензией CC BY-NC 4.0; Лаборатория Л. Джасперсена, общественное достояние. Масштабная линейка является приблизительной.
Рис. 1.
Представление разнообразия групп, содержащих многоядерных представителей. (A) Фораминиферы: Filosa, глубоководный ценоцитарный ксенофиофор; (B) Endomyxa: Lateromyxa gallica , многоядерная хищная амеба; (C) Tubulinea: Chaos sp. многоядерная амеба; (D) Mesomycetozoea: Sphaeroforma arctica , ценоцит с синими ядрами; (E) Protostomia: Drosophila melanogaster , многоядерный эмбрион; (F) Hexactinellida: Euplectella aspergillum ценоцитарная гексактинеллидная губка; (G) Дейтеростомия: многоядерные мышечные клетки мыши; (H) Heterolobosea: Acrasis rosea , плодовое тело; (I) Chloroplastida: Ulvophyceae: Cladophora sp. сифонный таллом; (J) – Rhodophyta: Florideophyceae: Lithophyllum sp.; (K) — миксомицеты: многоядерный плазмодий члена Physaraceae ; (L) Ascomycota: Eremothecium gossypii , асептированные гифы. Авторы фотографий и общий домен Creative Commons (CC): A и F. NOAA, общественное достояние; Б. Норберт Хюльсманн, BY-NC-SA 2.0; С. и И. Proyecto Agua, BY-NC-SA 2.0; D. Лаборатория мультиклеточного генома, BY 2.0; Э. Билли Лиар, BY-NC-SA 2.0; Г. Кевин А. Мурах, Галерея изображений NIH, BY-CN 2.0; Х. Ширли Чио, Лаборатория биологии грибов, Калифорнийский университет в Беркли, Калифорния, BY-SA 3.0; Дж. Кристоф Куинтин, BY-NC 2.0; К. Андре Амарал, распространяется под лицензией CC BY-NC 4.0; Лаборатория Л. Джасперсена, общественное достояние. Масштабная линейка является приблизительной.
сифонный таллом; (J) – Rhodophyta: Florideophyceae: Lithophyllum sp.; (K) — миксомицеты: многоядерный плазмодий члена Physaraceae ; (L) Ascomycota: Eremothecium gossypii , асептированные гифы. Авторы фотографий и общий домен Creative Commons (CC): A и F. NOAA, общественное достояние; Б. Норберт Хюльсманн, BY-NC-SA 2.0; С. и И. Proyecto Agua, BY-NC-SA 2.0; D. Лаборатория мультиклеточного генома, BY 2.0; Э. Билли Лиар, BY-NC-SA 2.0; Г. Кевин А. Мурах, Галерея изображений NIH, BY-CN 2.0; Х. Ширли Чио, Лаборатория биологии грибов, Калифорнийский университет в Беркли, Калифорния, BY-SA 3.0; Дж. Кристоф Куинтин, BY-NC 2.0; К. Андре Амарал, распространяется под лицензией CC BY-NC 4.0; Лаборатория Л. Джасперсена, общественное достояние. Масштабная линейка является приблизительной.
Некоторые исследователи различают термины синцитий и ценоцит на основании механизма, лежащего в основе многоядерного состояния, когда синцитии возникают в результате слияния клеток, а ценоциты возникают в результате сегрегации хромосом и ядерных делений без цитокинеза (Daubenmire 1936).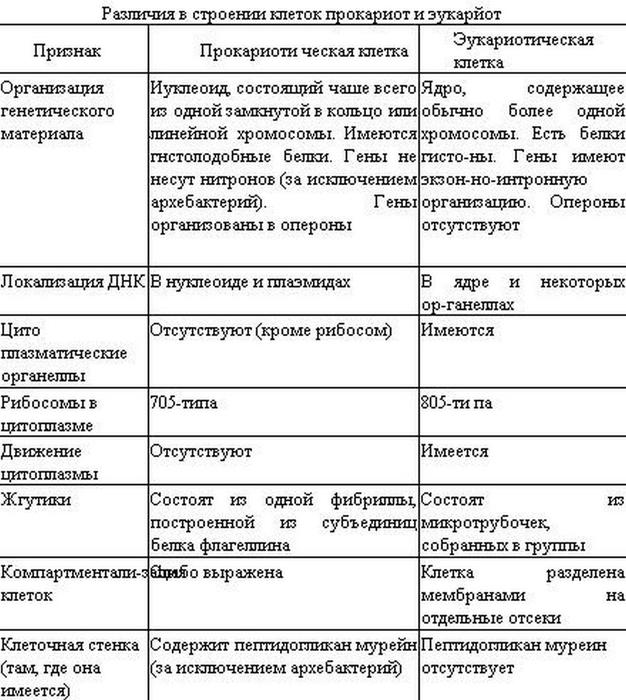 Оба приводят к многоядерному состоянию и не исключают друг друга. Мы используем термин «многоядерный» для описания состояния наличия более двух (обычно четырех или более) ядер в одной и той же клетке, независимо от механизма, вызвавшего это состояние. Стандартные митотические и мейотические промежуточные продукты, очевидно, не учитываются здесь как многоядерные состояния, поскольку это сделало бы признак тривиальным, сделав его столь же универсальным, как и наличие самих ядер. Изображения на рисунке 1 передают впечатление многоядерного состояния в том смысле, в каком это подразумевается в этой статье.
Оба приводят к многоядерному состоянию и не исключают друг друга. Мы используем термин «многоядерный» для описания состояния наличия более двух (обычно четырех или более) ядер в одной и той же клетке, независимо от механизма, вызвавшего это состояние. Стандартные митотические и мейотические промежуточные продукты, очевидно, не учитываются здесь как многоядерные состояния, поскольку это сделало бы признак тривиальным, сделав его столь же универсальным, как и наличие самих ядер. Изображения на рисунке 1 передают впечатление многоядерного состояния в том смысле, в каком это подразумевается в этой статье.
Вышеизложенные наблюдения приводят к вопросу о том, как далеко назад в эволюции эукариот можно проследить синцитиальное состояние. Являются ли многоядерные формы во всех эукариотических супергруппах результатом конвергенции или они отражают состояние предков? Здесь мы исследуем наличие многоядерных форм во всем разнообразии эукариот, вероятность многоядерного синцитиального LECA с использованием реконструкции наследственного состояния и последствия для образа жизни LECA.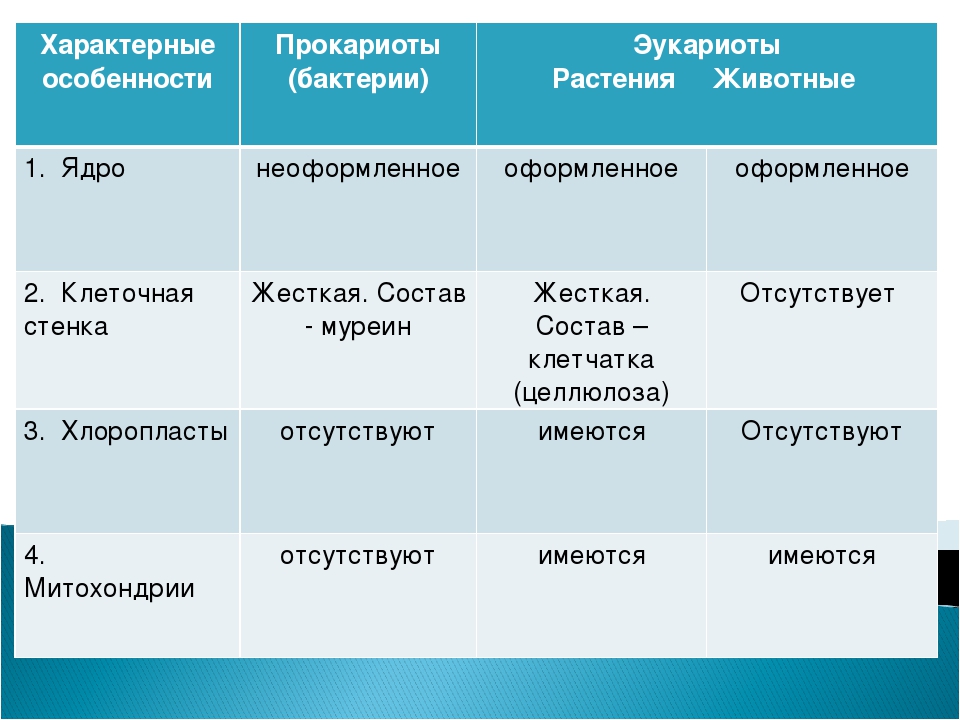
 Как упоминалось в разделе «Введение», многоядерное состояние обычно достигается одним из двух путей, а именно за счет отсутствия клеточного деления после деления ядра, что приводит к образованию клеток, обычно обозначаемых как ценоциты, и слияния одноядерных клеток, что приводит к образованию клеток, обычно обозначаемых как ценоциты. как синцитий. Хотя онтогенетическая разница между двумя состояниями очевидна, в отсутствие тщательных исследований клеточной биологии и клеточного цикла, которые отсутствуют для многих из рассмотренных здесь таксонов, невозможно точно закодировать две формы по отдельности, и поэтому они рассматриваются здесь. вместе как многоядерные.Обратите внимание, что наше эволюционное исследование касается свойств многоядерных клеток и взаимодействия в них ядер и митохондрий, независимо от процесса, породившего многоядерное состояние.
Как упоминалось в разделе «Введение», многоядерное состояние обычно достигается одним из двух путей, а именно за счет отсутствия клеточного деления после деления ядра, что приводит к образованию клеток, обычно обозначаемых как ценоциты, и слияния одноядерных клеток, что приводит к образованию клеток, обычно обозначаемых как ценоциты. как синцитий. Хотя онтогенетическая разница между двумя состояниями очевидна, в отсутствие тщательных исследований клеточной биологии и клеточного цикла, которые отсутствуют для многих из рассмотренных здесь таксонов, невозможно точно закодировать две формы по отдельности, и поэтому они рассматриваются здесь. вместе как многоядерные.Обратите внимание, что наше эволюционное исследование касается свойств многоядерных клеток и взаимодействия в них ядер и митохондрий, независимо от процесса, породившего многоядерное состояние.
Кладограмма для эукариот была создана на основе обширной литературы (Archibald et al. 2017; Cavalier-Smith 2018; Adl et al. 2019; Kiss et al.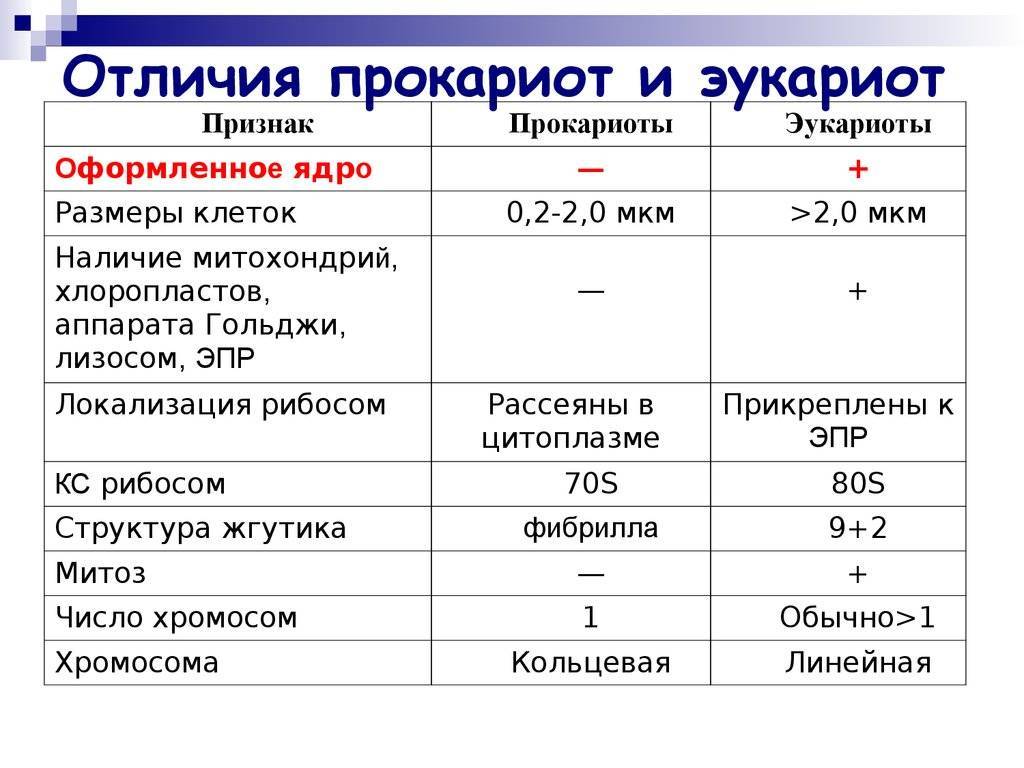 2019, полный список см. в дополнительных таблицах 2 и 3, дополнительный онлайн-материал) и дополнительно уточнены за счет учета различных конфигураций политомий и различных принятых положений корня (всего 16), что привело к 30 различным топологиям эукариот (дополнительная таблица 4, дополнительный онлайн-материал).Хотя дерево, показанное на рисунке 2, среди доступных в настоящее время альтернатив является наименее противоречивым и, возможно, наиболее надежным деревом для изучаемых здесь линий эукариот, все три различные топологии использовались для реконструкции состояния предков (дополнительная таблица 4, Дополнительный материал онлайн).
2019, полный список см. в дополнительных таблицах 2 и 3, дополнительный онлайн-материал) и дополнительно уточнены за счет учета различных конфигураций политомий и различных принятых положений корня (всего 16), что привело к 30 различным топологиям эукариот (дополнительная таблица 4, дополнительный онлайн-материал).Хотя дерево, показанное на рисунке 2, среди доступных в настоящее время альтернатив является наименее противоречивым и, возможно, наиболее надежным деревом для изучаемых здесь линий эукариот, все три различные топологии использовались для реконструкции состояния предков (дополнительная таблица 4, Дополнительный материал онлайн).
Рис. 2.
Схематическое изображение эволюции клеток. Дерево уходит корнями в физиологию и геохимию с несвободноживущим последним универсальным общим предком (LUCA).Происхождение эукариот изображается как полифилетическое (симбиогенное) событие, когда две прокариотические линии, линия архей и линия альфапротеобактерий, дали начало эукариотической линии через LECA — последнего эукариотического общего предка.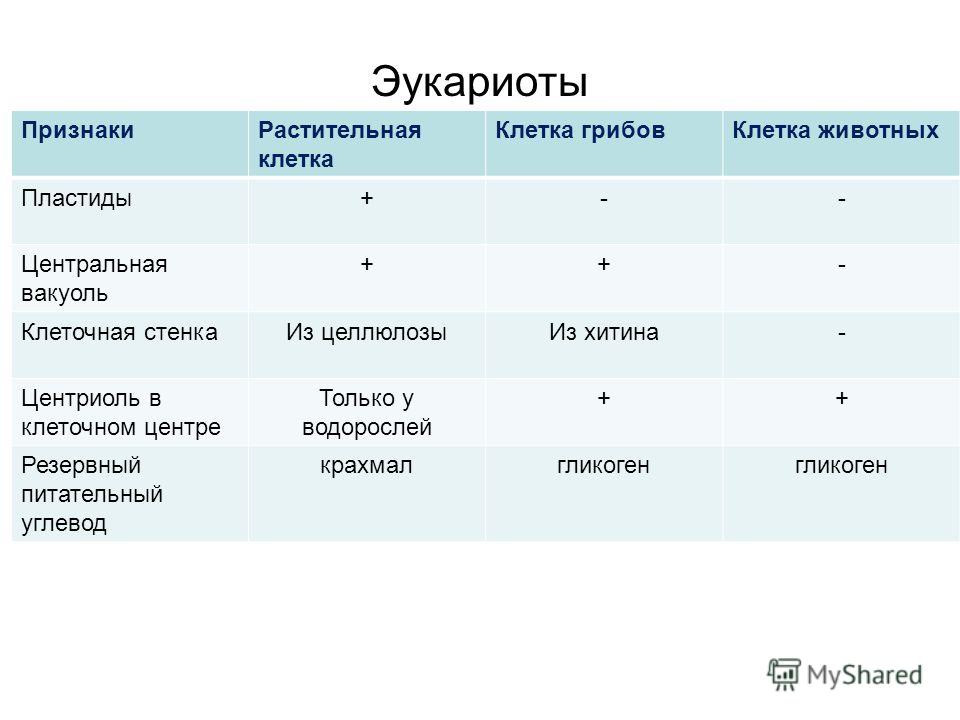 Показаны основные прокариотические группы (внутри архей и бактерий) и эукариотические супергруппы, всего в этот анализ включено 106 таксонов. Сравнение числа известных видов показано в логарифмической шкале. Квадраты на концах определенных ветвей обозначают, в каких группах секвенированы геномы.Справочное дерево эукариот см. в разделе «Материалы и методы». Схематическое прокариотическое древо жизни было построено на основе литературы. Дерево было нарисовано с помощью iTol. LUCA изображен как возникший в гидротермальном источнике, в то время как LECA, который мог также возникнуть вблизи гидротермальных источников как геологический источник H 2 (15, 23), изображен как многоядерный организм, в котором ядра делятся, а их оболочки остаются неповрежденными. . Показан первичный эндосимбиоз с цианобактериями, давший начало Archaeplastida.Вторичные эндосимбиотические события, множественное происхождение вторичных пластид, показаны стрелками. 6-минутный анимационный видеоролик, иллюстрирующий происхождение эукариот от симбиоза и роль синцитиального состояния в жизненном цикле LECA, можно посмотреть по адресу (https://www.
Показаны основные прокариотические группы (внутри архей и бактерий) и эукариотические супергруппы, всего в этот анализ включено 106 таксонов. Сравнение числа известных видов показано в логарифмической шкале. Квадраты на концах определенных ветвей обозначают, в каких группах секвенированы геномы.Справочное дерево эукариот см. в разделе «Материалы и методы». Схематическое прокариотическое древо жизни было построено на основе литературы. Дерево было нарисовано с помощью iTol. LUCA изображен как возникший в гидротермальном источнике, в то время как LECA, который мог также возникнуть вблизи гидротермальных источников как геологический источник H 2 (15, 23), изображен как многоядерный организм, в котором ядра делятся, а их оболочки остаются неповрежденными. . Показан первичный эндосимбиоз с цианобактериями, давший начало Archaeplastida.Вторичные эндосимбиотические события, множественное происхождение вторичных пластид, показаны стрелками. 6-минутный анимационный видеоролик, иллюстрирующий происхождение эукариот от симбиоза и роль синцитиального состояния в жизненном цикле LECA, можно посмотреть по адресу (https://www. youtube.com/watch?v=mmh_IpdgWvw&t=2s).
youtube.com/watch?v=mmh_IpdgWvw&t=2s).
Рис. 2.
Схематическое изображение эволюции клеток. Дерево уходит корнями в физиологию и геохимию с несвободноживущим последним универсальным общим предком (LUCA). Происхождение эукариот изображается как полифилетическое (симбиогенное) событие, когда две прокариотические линии, линия архей и линия альфапротеобактерий, дали начало эукариотической линии через LECA — последнего эукариотического общего предка.Показаны основные прокариотические группы (внутри архей и бактерий) и эукариотические супергруппы, всего в этот анализ включено 106 таксонов. Сравнение числа известных видов показано в логарифмической шкале. Квадраты на концах определенных ветвей обозначают, в каких группах секвенированы геномы. Справочное дерево эукариот см. в разделе «Материалы и методы». Схематическое прокариотическое древо жизни было построено на основе литературы. Дерево было нарисовано с помощью iTol. LUCA изображен как возникший в гидротермальном источнике, в то время как LECA, который мог также возникнуть вблизи гидротермальных источников как геологический источник H 2 (15, 23), изображен как многоядерный организм, в котором ядра делятся, а их оболочки остаются неповрежденными. .Показан первичный эндосимбиоз с цианобактериями, давший начало Archaeplastida. Вторичные эндосимбиотические события, множественное происхождение вторичных пластид, показаны стрелками. 6-минутный анимационный видеоролик, иллюстрирующий происхождение эукариот от симбиоза и роль синцитиального состояния в жизненном цикле LECA, можно посмотреть по адресу (https://www.youtube.com/watch?v=mmh_IpdgWvw&t=2s).
.Показан первичный эндосимбиоз с цианобактериями, давший начало Archaeplastida. Вторичные эндосимбиотические события, множественное происхождение вторичных пластид, показаны стрелками. 6-минутный анимационный видеоролик, иллюстрирующий происхождение эукариот от симбиоза и роль синцитиального состояния в жизненном цикле LECA, можно посмотреть по адресу (https://www.youtube.com/watch?v=mmh_IpdgWvw&t=2s).
В качестве контрольных наборов данных для реконструкции состояния предков мы включили несколько признаков, которые уже аннотированы для многих линий эукариотического домена.Помимо наличия митохондрий, первый эукариот был половым и имел мейотическую рекомбинацию (Speijer et al. 2015; Fu et al. 2019; Hofstatter and Lahr 2019). Известно, что гидрогеносомы и митосомы возникли из митохондрий в результате потери дыхательной цепи и экологической специализации в нескольких независимых линиях (Embley and Martin 2006; Müller et al. 2012; Maciszewski and Karnkowska 2019; Gould et al. 2019), что первичные пластиды возникли однажды (Sánchez-Baracaldo et al.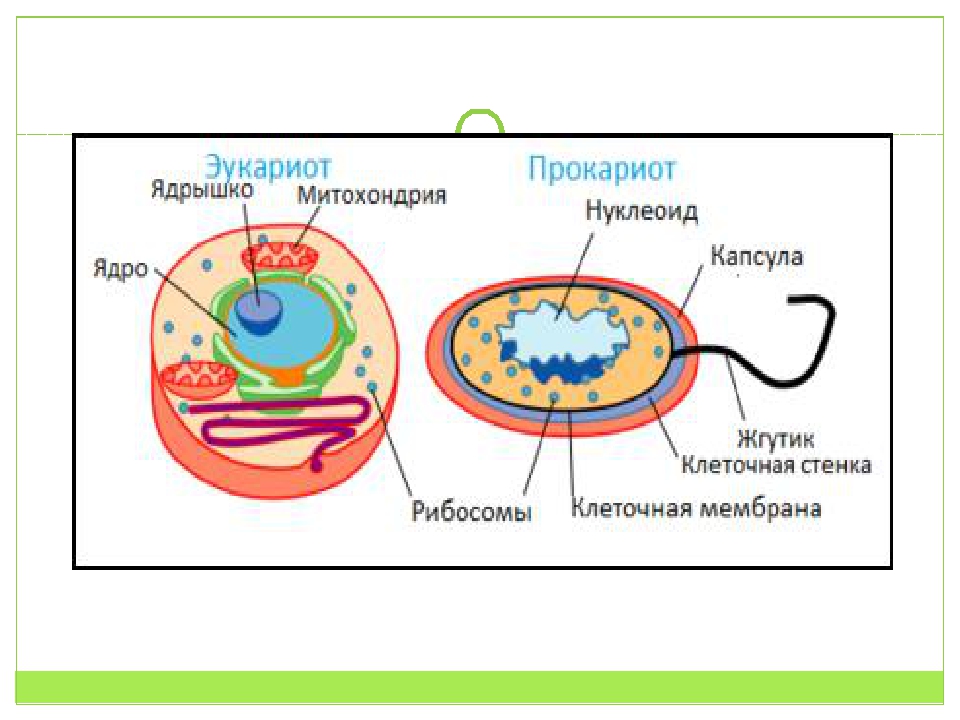 2017) и что вторичные пластиды несколько раз возникали независимо от эукариот, содержащих первичную пластиду (Maciszewski and Karnkowska 2019; Keeling 2004; Gould et al.2008). Реконструкция состояния предков должна соответствующим образом отображать эти черты.
2017) и что вторичные пластиды несколько раз возникали независимо от эукариот, содержащих первичную пластиду (Maciszewski and Karnkowska 2019; Keeling 2004; Gould et al.2008). Реконструкция состояния предков должна соответствующим образом отображать эти черты.
Общая схема родства прокариот с эукариотами, включая симбиоз и количество описанных видов в каждой группе, представлена на рисунке 2. Используя реконструкцию состояния предков, мы обнаружили, что последний общий предок эукариот (LECA) был половым, митохондриальный и гетеротрофный организм с закрытым делением ядра (митоз) и, вероятно, с гаплоидным ядром (рис. 3). Метод и дерево прослеживают половое размножение и митохондрии до происхождения эукариотической сложности, что согласуется с ранее опубликованными исследованиями (Speijer et al.2015 г.; Хофштеттер и Лар, 2019). Линии с гидрогеносомами, митосомами и типичными митохондриями (рис. 3А) представляют экологические специализации от общей предковой органеллы (Müller et al. 2012). В соответствии с предыдущими отчетами установлено, что пол встречается повсеместно в домене Eukarya, а мейотические гены присутствуют во всех супергруппах в высокой степени консервативно (Ramesh et al. 2005; Speijer et al. 2015; Hofstatter and Lahr 2019).
2012). В соответствии с предыдущими отчетами установлено, что пол встречается повсеместно в домене Eukarya, а мейотические гены присутствуют во всех супергруппах в высокой степени консервативно (Ramesh et al. 2005; Speijer et al. 2015; Hofstatter and Lahr 2019).
Рис. 3.
Обзор эукариотического разнообразия с реконструкцией наследственного состояния.(A) Признаки аннотированы на эталонном дереве эукариот (1. наличие многоядерного состояния, 2. половое размножение, 3. тип деления ядра, 4. полиплоидия, 5. тип митохондрий и 6. тип пластид). (B) Показана реконструкция состояния предков многоядерного состояния, а также (C). Эволюция пластид. Схематично изображены представители основных фотосинтезирующих эукариот. Полученные данные свидетельствуют о том, что LECA был многоядерным половым гетеротрофом с закрытым митозом.
Рис.3.
Обзор эукариотического разнообразия с реконструкцией наследственного состояния. (A) Признаки аннотированы на эталонном дереве эукариот (1. наличие многоядерного состояния, 2. половое размножение, 3. тип деления ядра, 4. полиплоидия, 5. тип митохондрий и 6. тип пластид). (B) Показана реконструкция состояния предков многоядерного состояния, а также (C). Эволюция пластид. Схематично изображены представители основных фотосинтезирующих эукариот. Полученные данные свидетельствуют о том, что LECA был многоядерным половым гетеротрофом с закрытым митозом.
наличие многоядерного состояния, 2. половое размножение, 3. тип деления ядра, 4. полиплоидия, 5. тип митохондрий и 6. тип пластид). (B) Показана реконструкция состояния предков многоядерного состояния, а также (C). Эволюция пластид. Схематично изображены представители основных фотосинтезирующих эукариот. Полученные данные свидетельствуют о том, что LECA был многоядерным половым гетеротрофом с закрытым митозом.
Последний общий предок архепластид был первым организмом, имеющим первичную пластиду, и первым общим предком всех (вторичных) пластид, обнаруженных у Euglenida , Hacrobia и SAR. Несмотря на широкое распространение в эукариотическом дереве, пластиды не восходят к LECA с наследственной реконструкцией, которая служит формой внутреннего контроля (рис. 3С). Поскольку у LECA не было пластиды, он не мог быть фотосинтезирующим автотрофным эукариотом — он был гетеротрофом.Первичные пластиды произошли от цианобактерий в результате симбиогенного события, в котором, вероятно, также участвовал предок пресноводных архепластид, который был многоядерным, по крайней мере, в части своего жизненного цикла (Sánchez-Baracaldo et al.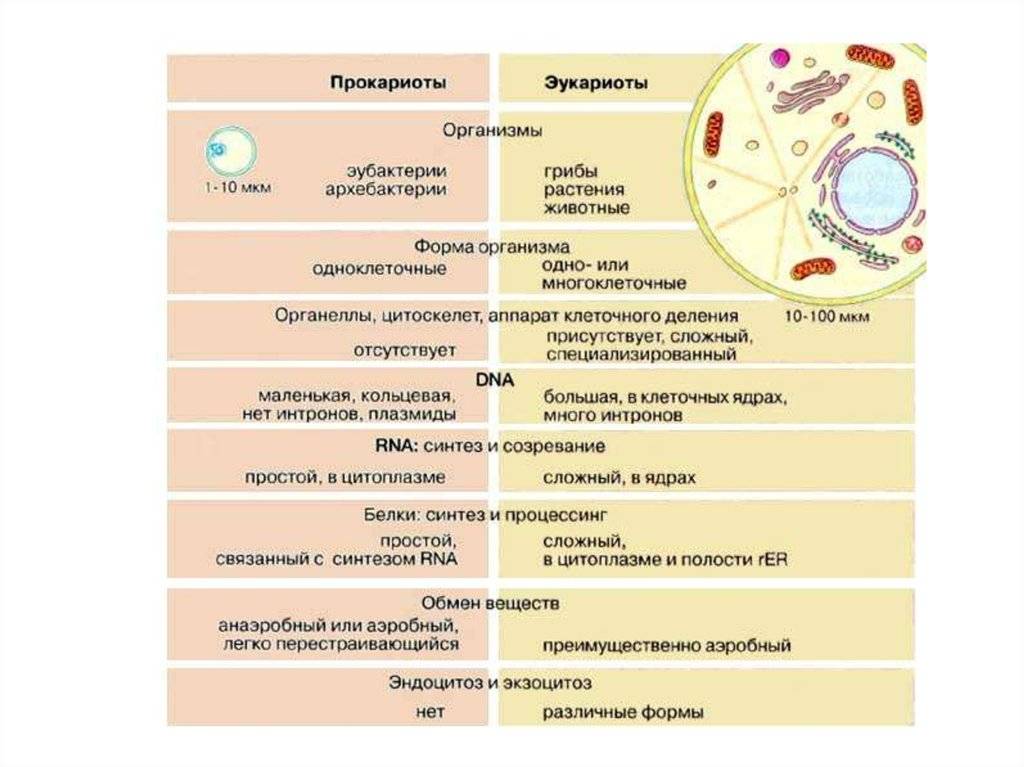 2017) (рис. 3B). Полиплоидия (>2 n ) также возникала несколько раз независимо друг от друга. Хотя полиплоидные эукариоты неоднократно возникали в эволюционно хорошо обособленных группах, LECA, скорее всего, имели гаплоидные ядра (рис. 3А). Полиплоидные трофобионты (стадии питания) среди эукариот встречаются редко.Единственными полиплоидными фазами у большинства эукариот является диплоидная зигота (особенно ее тетраплоидная фаза перед первым делением), которая наследственно подвергается мейотической рекомбинации до образования четырех различных ядер (трофических клеток, спор или гамет). В тех линиях, чьи трофобионты являются диплоидными, каждое соматическое ядро, имеющее ДНК, реплицируемую до сегрегации хромосом, можно рассматривать как временно тетраплоидное, хотя, как и в случае образования зиготы (см. Материалы и методы), они не были оценены как полиплоидные.Соответственно, LECA не был полиплоидным, но поскольку он был мейотическим, он содержал некоторую форму кариогамной стадии.
2017) (рис. 3B). Полиплоидия (>2 n ) также возникала несколько раз независимо друг от друга. Хотя полиплоидные эукариоты неоднократно возникали в эволюционно хорошо обособленных группах, LECA, скорее всего, имели гаплоидные ядра (рис. 3А). Полиплоидные трофобионты (стадии питания) среди эукариот встречаются редко.Единственными полиплоидными фазами у большинства эукариот является диплоидная зигота (особенно ее тетраплоидная фаза перед первым делением), которая наследственно подвергается мейотической рекомбинации до образования четырех различных ядер (трофических клеток, спор или гамет). В тех линиях, чьи трофобионты являются диплоидными, каждое соматическое ядро, имеющее ДНК, реплицируемую до сегрегации хромосом, можно рассматривать как временно тетраплоидное, хотя, как и в случае образования зиготы (см. Материалы и методы), они не были оценены как полиплоидные.Соответственно, LECA не был полиплоидным, но поскольку он был мейотическим, он содержал некоторую форму кариогамной стадии.
В то время как контрольные признаки были реконструированы, как и ожидалось, тот же анализ показывает, что LECA был многоядерным и/или имел многоядерную стадию в течение своего жизненного цикла (рис. 3A и B). Это не был одноядерный протистоподобный жгутиковый эукариот того типа, который характерен для большинства теорий, хотя нельзя исключать, что некоторые фазы жизненного цикла могли быть одноядерными, протистоподобными и жгутиковыми, например, подвижные споры.Предковая реконструкция указывает на то, что состояние LECA могло быть многоядерным с ядрами, разделенными закрытым митозом, при котором ядерная оболочка оставалась интактной (рис. 3А). Наличие у предков закрытого митоза (закрытая сегрегация хромосом, а не цитокинез) имеет важное значение, поскольку сегрегация хромосом в синцитиальных формах требует интактной ядерной мембраны, соответствующей наследственной многоядерной стадии. В наших анализах вероятность того, что LECA была многоядерной, так же высока, как и вероятность того, что она была половой и обладала митохондриями (дополнительная таблица 7, дополнительный онлайн-материал). Полные подробные результаты реконструкции состояния предков см. в дополнительной таблице 6, Дополнительный материал онлайн.
Полные подробные результаты реконструкции состояния предков см. в дополнительной таблице 6, Дополнительный материал онлайн.
Независимо от того, где мы укореняли эукариотическое дерево и сколько неразрешенных ветвей мы допускали, LECA всегда реконструировался как многоядерный. Более того, что очень важно, не только предок эукариот был многоядерным, но и общие предки всех надгрупп эукариот также были реконструированы как многоядерные, за исключением последнего общего предка Hacrobia (рис.3Б). Был ли предок Hacrobia одноядерным или отсутствует информация, которая приводит к выводу об одноядерном предке Hacrobia, в то время как все другие предки надгруппы реконструируются как многоядерные, остается нерешенным. Реконструкция предков изображает LECA как многоядерный поликарион, синцитиальный или ценоцитный, популяцию взаимодействующих митохондрий и ядер в пределах одной клеточной мембраны. Множественные ядра в одной и той же цитоплазме — нередкое явление у эукариот.Синцитии и ценоциты встречаются в большинстве групп высших эукариот (дополнительная таблица 2, дополнительный онлайн-материал). Свободно плавающие ядра в цитозоле, а не привязанные к клеточным стенкам, означают, что в синцитии они могут делиться только в том случае, если ядерное деление — и, следовательно, сегрегация хромосом — закрыто, при этом ядерная мембрана остается неповрежденной на протяжении всего митоза. Открытый митоз в ценоците потенциально может привести к прикреплению веретенообразного аппарата к хромосомам из разных ядер и их сегрегации аберрантным и, вероятно, летальным образом.Реконструкция (рис. 3Б; табл. 1) свидетельствует о том, что открытое деление ядра (растворение ядерной оболочки при митозе, как хорошо известно у позвоночных) возникло из закрытого митоза путем полуоткрытого деления, при котором растворяются части ядерной оболочки, как промежуточное состояние (Boettcher and Barral, 2013). Открытое деление ядра характерно для некоторых одноядерных (как одноклеточных, так и многоклеточных) и большинства наземных эукариот (рис. 3А).
Свободно плавающие ядра в цитозоле, а не привязанные к клеточным стенкам, означают, что в синцитии они могут делиться только в том случае, если ядерное деление — и, следовательно, сегрегация хромосом — закрыто, при этом ядерная мембрана остается неповрежденной на протяжении всего митоза. Открытый митоз в ценоците потенциально может привести к прикреплению веретенообразного аппарата к хромосомам из разных ядер и их сегрегации аберрантным и, вероятно, летальным образом.Реконструкция (рис. 3Б; табл. 1) свидетельствует о том, что открытое деление ядра (растворение ядерной оболочки при митозе, как хорошо известно у позвоночных) возникло из закрытого митоза путем полуоткрытого деления, при котором растворяются части ядерной оболочки, как промежуточное состояние (Boettcher and Barral, 2013). Открытое деление ядра характерно для некоторых одноядерных (как одноклеточных, так и многоклеточных) и большинства наземных эукариот (рис. 3А).
Таблица 1 Сводка результатов реконструкции состояния предков с максимальной экономией по 30 различным топологиям
.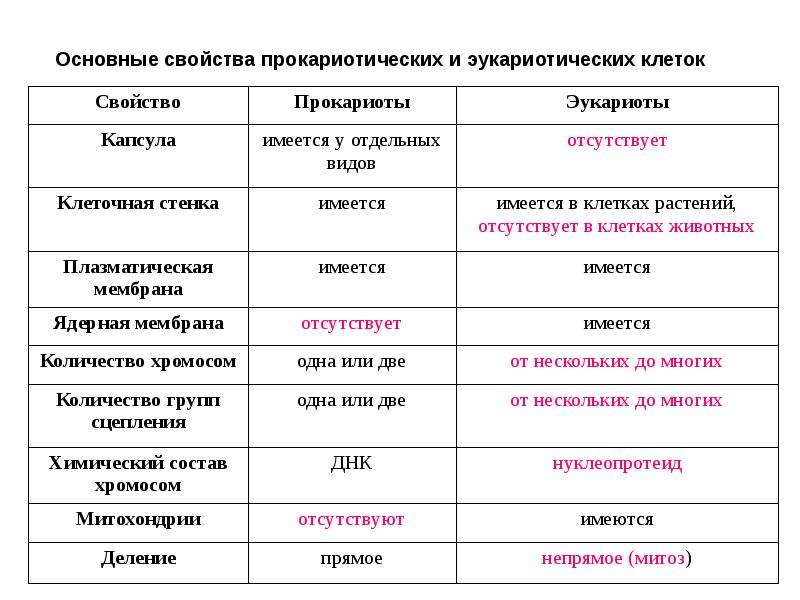
. % Деревья
.
.
. Отсутствует
. Настоящее
. Неоднозначный
. синцити 0 0,8 0,2 Пол 0 0,867 0,133 полиплоидия 1 0 0 Закрытое деление 0 1 0 Митохондрии 0 0.867 0,133 Пластид 1 0 0 2 8
.
. % Деревья
.
.
. Отсутствует
. Настоящее
. Неоднозначный
. Синцитий 0 0. 8
8 0,2 Пол 0 0,867 0,133 полиплоидия 1 0 0 Закрытое деление 0 1 0 Mitochondria 0 0 0 0 0,867 0.133 Plastid 1 0 0 9042 0
Таблица 1 Краткое изложение результатов реконструкции состояния предков максимальной парисийкой, через 30 разных топологий
.
. % Деревья
.
.
. Отсутствует
. Настоящее
. Неоднозначный
. синцити 0 0,8 0,2 Пол 0 0,867 0,133 полиплоидия 1 0 0 Закрытое деление 0 1 0 Митохондрии 0 0. 867
867 0,133 Пластид 1 0 0 2 8
.
. % Деревья
.
.
. Отсутствует
. Настоящее
. Неоднозначный
. Синцитий 0 0.8 0,2 Пол 0 0,867 0,133 полиплоидия 1 0 0 Закрытое деление 0 1 0 Mitochondria 0 0 0 0.867 0.133 Plastid 1 0 0 2  Симбиоз, давший начало эукариотам, произошел более 1,5 миллиардов лет назад (Knoll et al. 2006). В то время как происхождение эукариот не может произойти в лаборатории, эндосимбиоз может произойти (Mehta et al. 2018). Контуры эукариогенеза, промежуточные стадии и последовательность вовлеченных событий могут быть рассмотрены посредством вывода из сравнительного исследования современных линий. Первый эукариот был результатом взаимодействия между археями и бактериями, двумя сильно расходящимися клеточными линиями, которые в результате взаимодействия и сотрудничества породили новый вид организма, LECA, с новыми свойствами, новой биоэнергетикой, химерными хромосомами, клеточным циклом, новым генетика, реципрокная рекомбинация и клеточная сложность.Потомки этих симбиотических партнеров сохраняются в виде бактериальных рибосом в митохондриях и архейных рибосом в эукариотическом цитозоле.
Симбиоз, давший начало эукариотам, произошел более 1,5 миллиардов лет назад (Knoll et al. 2006). В то время как происхождение эукариот не может произойти в лаборатории, эндосимбиоз может произойти (Mehta et al. 2018). Контуры эукариогенеза, промежуточные стадии и последовательность вовлеченных событий могут быть рассмотрены посредством вывода из сравнительного исследования современных линий. Первый эукариот был результатом взаимодействия между археями и бактериями, двумя сильно расходящимися клеточными линиями, которые в результате взаимодействия и сотрудничества породили новый вид организма, LECA, с новыми свойствами, новой биоэнергетикой, химерными хромосомами, клеточным циклом, новым генетика, реципрокная рекомбинация и клеточная сложность.Потомки этих симбиотических партнеров сохраняются в виде бактериальных рибосом в митохондриях и архейных рибосом в эукариотическом цитозоле.
LECA имели половое размножение, которое включало слияние гаплоидных ядер, отобранных для размножения (гаметы), и рекомбинацию их генетического материала (мейоз).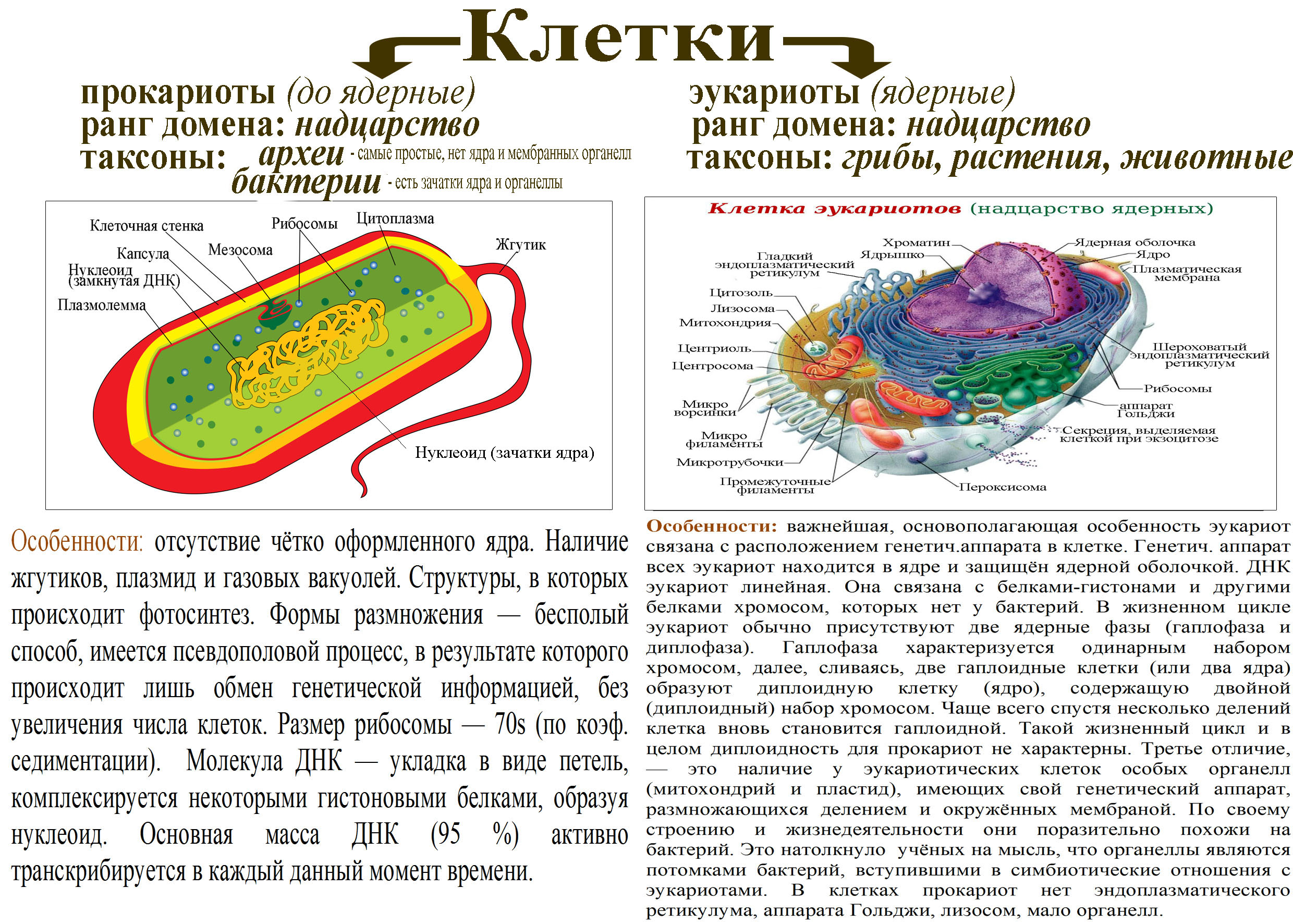 Митохондрии, пол и несколько ядер являются сигнатурами состояния LECA с синергетическими взаимодействиями. В отличие от митохондрий, ядро имеет большой сложный геном с небольшими ограничениями по размеру.Генетическая совместимость ядер и митохондрий, населяющих одну и ту же цитоплазму, имеет решающее значение для выживания эукариотических клеток. Внутренняя конкуренция или цитоядерная несовместимость могут привести к летальному исходу (Blackstone and Green 1999; Pesole et al. 2012; Rand and Mossman 2020) или привести к дисфункции организма. Наследование митохондрий часто однородительское. Однако наследование ядерного генома бывает би-, три- или мультиродительским. Однородительское наследование митохондрий указывает на существование жесткого контроля на совместимость.Мейотическая рекомбинация, происходящая у предков во время фазы зиготы, является контрольной точкой совместимости. В начале эволюции эукариот была необходима совместимость митохондрий с вновь возникшими ядрами. В одноядерных клетках естественный отбор пережили только совместимые комбинации.
Митохондрии, пол и несколько ядер являются сигнатурами состояния LECA с синергетическими взаимодействиями. В отличие от митохондрий, ядро имеет большой сложный геном с небольшими ограничениями по размеру.Генетическая совместимость ядер и митохондрий, населяющих одну и ту же цитоплазму, имеет решающее значение для выживания эукариотических клеток. Внутренняя конкуренция или цитоядерная несовместимость могут привести к летальному исходу (Blackstone and Green 1999; Pesole et al. 2012; Rand and Mossman 2020) или привести к дисфункции организма. Наследование митохондрий часто однородительское. Однако наследование ядерного генома бывает би-, три- или мультиродительским. Однородительское наследование митохондрий указывает на существование жесткого контроля на совместимость.Мейотическая рекомбинация, происходящая у предков во время фазы зиготы, является контрольной точкой совместимости. В начале эволюции эукариот была необходима совместимость митохондрий с вновь возникшими ядрами. В одноядерных клетках естественный отбор пережили только совместимые комбинации. В синцитиях взаимодействия «многие ко многим» между митохондриями и ядрами обеспечивают буферную совместимость в пределах окружающей среды одной цитоплазмы. Споры, порожденные синцитиальным LECA, представляли сильное узкое место отбора на цитоядерную совместимость (Garg and Martin 2016).
В синцитиях взаимодействия «многие ко многим» между митохондриями и ядрами обеспечивают буферную совместимость в пределах окружающей среды одной цитоплазмы. Споры, порожденные синцитиальным LECA, представляли сильное узкое место отбора на цитоядерную совместимость (Garg and Martin 2016).
Интригующий аспект многоядерного состояния LECA касается перехода от прокариотической к эукариотической сегрегации хромосом. У прокариот сегрегация хромосом связана с делением клеток посредством прикрепления хромосом к клеточной стенке. У эукариот зависящая от микротрубочек сегрегация конденсированных хромосом и клеточное деление (цитокинез) не связаны ни физически, ни механически, хотя часто противопоставлены во времени. То есть хромосомы могут и часто реплицируются и сегрегируют в неделящихся клетках без образования веретен деления самого ядра (Geitler 1953), процессы, которые в более ранней литературе назывались Amitose (Strasburger 1908).Если начало деления ядра (репликация с последующей сегрегацией) предшествовало возникновению клеточного деления у эукариот (обратное вряд ли могло бы быть верным), то образовавшийся синцитий не обязательно обладал хорошо регулируемой сегрегацией хромосом в начале.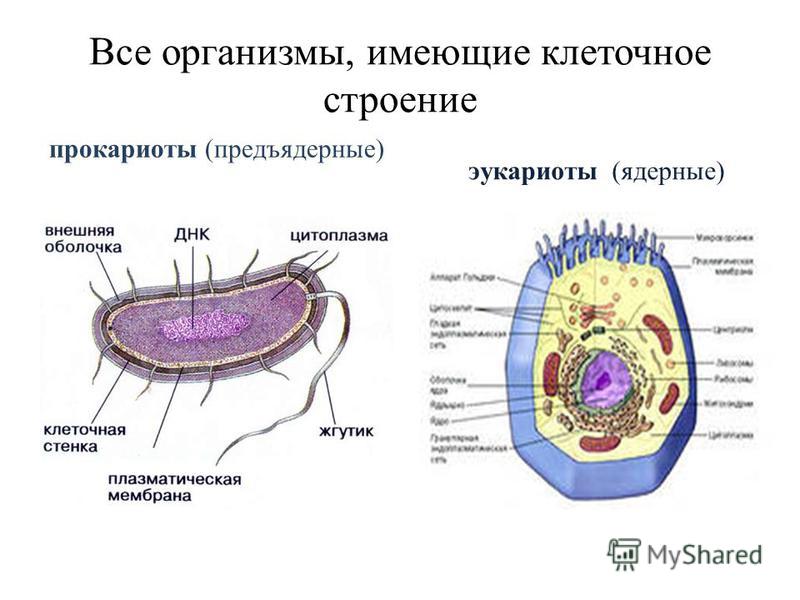 Он мог генерировать ядра с аберрантным числом хромосом или анеуплоидными гаплоидами. Такие дефектные ядра были бы летальными для одноядерной клетки, но не для синцития, потому что даже сильно дефектные ядра могут свободно дополнять друг друга через мРНК в цитозоле.Таким образом, многоядерное состояние должно амортизировать практически все вредные эффекты ядер, возникающие как продукты неправильного разделения хромосом во время закрытого протомитоза в начале сегрегации хромосом эукариот. Это могло бы сохранить жизнь синцитию как единице вегетативной пролиферации, в то же время укрывая ядра с очень разными наборами хромосом, ядра, которые поддерживали бы жизнеспособность друг друга в синцитии за счет комплементации через мРНК в цитозоле. Это участие рибосом, чей синтез требует массивной экспрессии генов рРНК, для комплементации может объяснить, почему соотношение объемов ядра и цитоплазмы ( Kern-Plasmarelation ) имеет тенденцию приближаться к примерно постоянному значению (Klieneberger 1917) 1:10 даже в синцитиальных клетках.
Он мог генерировать ядра с аберрантным числом хромосом или анеуплоидными гаплоидами. Такие дефектные ядра были бы летальными для одноядерной клетки, но не для синцития, потому что даже сильно дефектные ядра могут свободно дополнять друг друга через мРНК в цитозоле.Таким образом, многоядерное состояние должно амортизировать практически все вредные эффекты ядер, возникающие как продукты неправильного разделения хромосом во время закрытого протомитоза в начале сегрегации хромосом эукариот. Это могло бы сохранить жизнь синцитию как единице вегетативной пролиферации, в то же время укрывая ядра с очень разными наборами хромосом, ядра, которые поддерживали бы жизнеспособность друг друга в синцитии за счет комплементации через мРНК в цитозоле. Это участие рибосом, чей синтез требует массивной экспрессии генов рРНК, для комплементации может объяснить, почему соотношение объемов ядра и цитоплазмы ( Kern-Plasmarelation ) имеет тенденцию приближаться к примерно постоянному значению (Klieneberger 1917) 1:10 даже в синцитиальных клетках.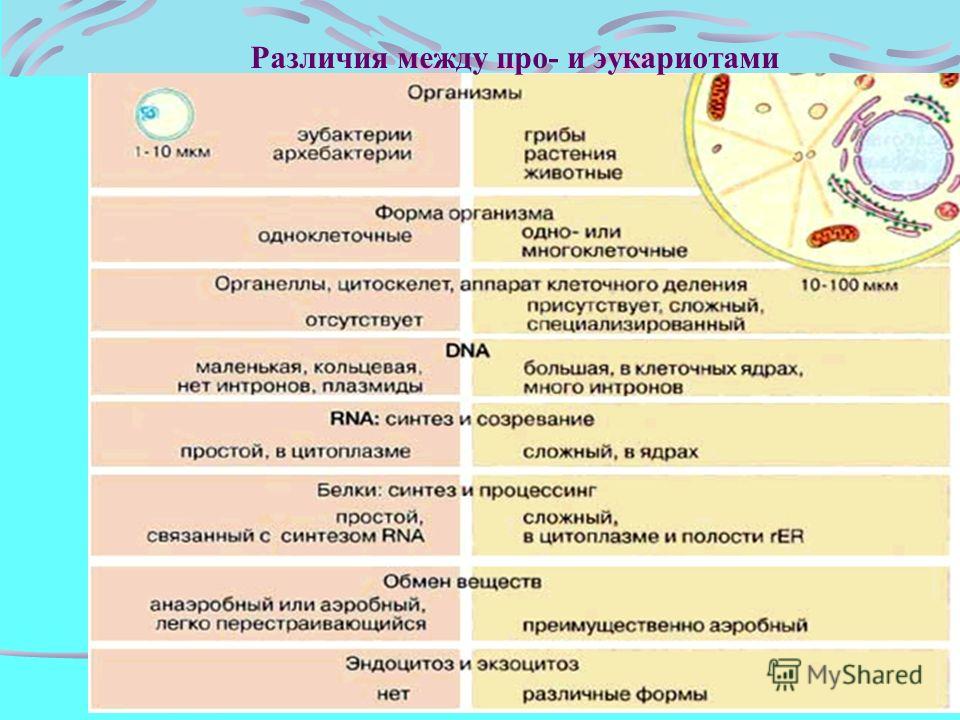 (Ситте и др.1991). Как выразился Страсбургер (1908): «У Characeae амитотическое деление ядер в интернодиальных клетках не является дегенеративным процессом, а скорее является средством амплификации определенных компонентов ядерного вещества в связи с увеличением цитоплазматической массы» (стр. 40). , перевод авторов).
(Ситте и др.1991). Как выразился Страсбургер (1908): «У Characeae амитотическое деление ядер в интернодиальных клетках не является дегенеративным процессом, а скорее является средством амплификации определенных компонентов ядерного вещества в связи с увеличением цитоплазматической массы» (стр. 40). , перевод авторов).
Физическое слияние ядер, примитивный и нерегулируемый предшественник кариогамии (присутствующий в LECA, потому что LECA занимается сексом), будет генерировать новые комбинации хромосом одновременно с переносом генов из митохондрий в ядра (Lane and Martin 2012; Гарг и Мартин, 2016 г.).Это породило гетерогенную популяцию ядер, взаимодействующих с гетерогенной популяцией митохондрий внутри одного и того же синцития. Синцитий также может быть физически разорван, генерируя сегменты или фрагменты, которые при наличии средств запечатывания концов могут давать потомство (в виде диаспор) без потребности в регулируемом клеточном делении. Синцитиальные фрагменты обеспечивают механизм распространения популяций ядер и митохондрий. Но главным эволюционным препятствием, которое необходимо было преодолеть, была эволюция регулируемой симметричной сегрегации хромосом, которая учитывала состояние питания клетки (Brunk and Martin 2019) на пути к клеточному циклу — основе биологии эукариотических клеток.
Но главным эволюционным препятствием, которое необходимо было преодолеть, была эволюция регулируемой симметричной сегрегации хромосом, которая учитывала состояние питания клетки (Brunk and Martin 2019) на пути к клеточному циклу — основе биологии эукариотических клеток.
Внутри синцития и ядра, и митохондрии были единицами отбора и единицами эволюции. Они были промежуточным состоянием при переходе от прокариот к эукариотам. Они сосуществовали в одном и том же цитозоле. Ядра стали наследственным набором генов, способных влиять на непосредственно окружающий их цитозоль и способных взаимодействовать друг с другом и с митохондриями посредством экспортируемой мРНК. Многоядерные клетки широко распространены среди эукариот, как живых (рис. 1 и 3А), так и ископаемых, таких как недавно обнаруженная ценоцитарная зеленая водоросль возрастом 1 миллиард лет (Tang et al.2020).
 Чтобы сравнить относительную приспособленность одноядерной клетки (монокариона) и многоядерной клетки (поликариона), можно рассмотреть разницу между вероятностью выживания популяции одноклеточных одноядерных эукариот и одиночного синцития. Для монокарионов вероятность выживания популяции зависит от индивидуальных вероятностей выживания, которые, в свою очередь, зависят от приспособленности соответствующей митонуклеарной пары.Однако в случае синцития, поскольку митохондрии и ядра сосуществуют в одной клетке, вероятность выживания зависит от кумулятивной приспособленности всех возможных комбинаций митоядерных пар. Это, в свою очередь, позволяет синцитию вести себя аналогично популяции, позволяя отбору независимо разрешать внутренние мито-ядерные конфликты. Это схематически показано на рисунке 4 и математически описано в дополнительной информации 11, Дополнительный материал онлайн. Синцитий ведет себя скорее как популяция ядер и митохондрий, чем как отдельная клетка.Таким образом, у синцития больше шансов на выживание, чем у популяции монокарионов.
Чтобы сравнить относительную приспособленность одноядерной клетки (монокариона) и многоядерной клетки (поликариона), можно рассмотреть разницу между вероятностью выживания популяции одноклеточных одноядерных эукариот и одиночного синцития. Для монокарионов вероятность выживания популяции зависит от индивидуальных вероятностей выживания, которые, в свою очередь, зависят от приспособленности соответствующей митонуклеарной пары.Однако в случае синцития, поскольку митохондрии и ядра сосуществуют в одной клетке, вероятность выживания зависит от кумулятивной приспособленности всех возможных комбинаций митоядерных пар. Это, в свою очередь, позволяет синцитию вести себя аналогично популяции, позволяя отбору независимо разрешать внутренние мито-ядерные конфликты. Это схематически показано на рисунке 4 и математически описано в дополнительной информации 11, Дополнительный материал онлайн. Синцитий ведет себя скорее как популяция ядер и митохондрий, чем как отдельная клетка.Таким образом, у синцития больше шансов на выживание, чем у популяции монокарионов. Конечно, существуют древние линии эукариот, несущие одноядерные формы, в том числе и раскопки. Однако многоядерная LECA объясняет, почему разнообразие современных эукариот легче происходит от синцитиального предка, чем от популяции одноядерных одноклеточных предков (монокарионов). Популяция монокарионов, особенно гаплоидных монокарионов, вряд ли будет накапливать генетическое разнообразие.С другой стороны, синцитий легко накапливает генетическое разнообразие в пределах одного цитозоля, поскольку ядра с полезными аллелями дополняют недостатки других ядер, а кариогамия, на которую способна мейотическая LECA, внутри синцития может генерировать новые комбинации хромосом (рис. 4). .
Конечно, существуют древние линии эукариот, несущие одноядерные формы, в том числе и раскопки. Однако многоядерная LECA объясняет, почему разнообразие современных эукариот легче происходит от синцитиального предка, чем от популяции одноядерных одноклеточных предков (монокарионов). Популяция монокарионов, особенно гаплоидных монокарионов, вряд ли будет накапливать генетическое разнообразие.С другой стороны, синцитий легко накапливает генетическое разнообразие в пределах одного цитозоля, поскольку ядра с полезными аллелями дополняют недостатки других ядер, а кариогамия, на которую способна мейотическая LECA, внутри синцития может генерировать новые комбинации хромосом (рис. 4). .
Рис. 4.
Синцитии буферируют хромосомные дефекты, в отличие от монокарионов. Схематическое изображение популяции одноклеточных протистов и синцитиальной клетки. Геномы каждого ядра схематично показаны серыми линиями, дефицитные аллели — красными прямоугольниками, тогда как полезный аллель показан зеленым цветом. Если применить те же эволюционные ограничения, популяция монокарионов с большей вероятностью вымрет, чем синцитий, поскольку ядра из разных клеток не могут ни покрывать дефекты друг друга, ни буферировать митонуклеарную несовместимость, в то время как в синцитии они могут.
Если применить те же эволюционные ограничения, популяция монокарионов с большей вероятностью вымрет, чем синцитий, поскольку ядра из разных клеток не могут ни покрывать дефекты друг друга, ни буферировать митонуклеарную несовместимость, в то время как в синцитии они могут.
Рис. 4.
Синцитии буферируют хромосомные дефекты, в отличие от монокарионов. Схематическое изображение популяции одноклеточных протистов и синцитиальной клетки. Геномы каждого ядра схематично показаны серыми линиями, дефицитные аллели — красными прямоугольниками, тогда как полезный аллель показан зеленым цветом.Если применить те же эволюционные ограничения, популяция монокарионов с большей вероятностью вымрет, чем синцитий, поскольку ядра из разных клеток не могут ни покрывать дефекты друг друга, ни буферировать митонуклеарную несовместимость, в то время как в синцитии они могут.
Эволюционные переходы в индивидуальности включают сотрудничество и конфликт (Buss, 1987; Maynard Smith and Szathmáry, 1996; Michod, 1999).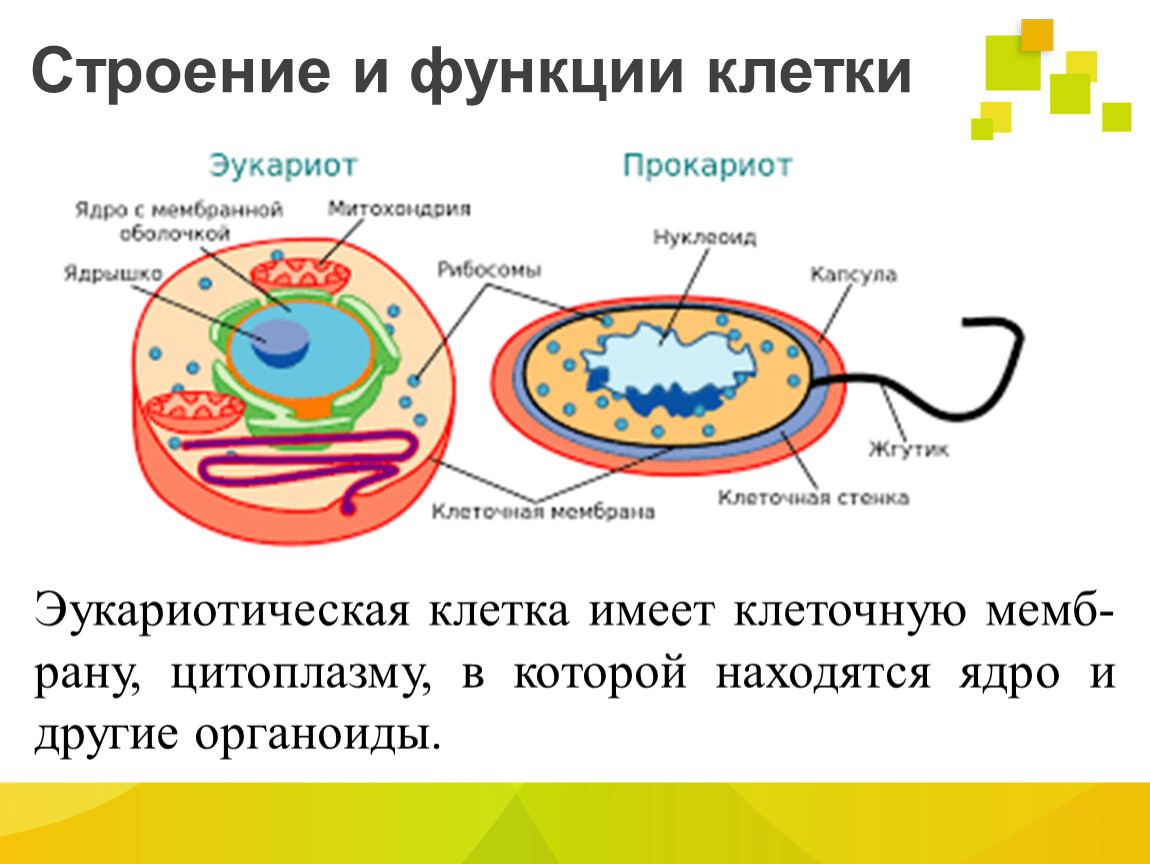 Без механизмов посредничества в конфликтах сотрудничество не может выжить (Nowak 2006) и не может возникнуть единица более высокого уровня (Radzvilavicius and Blackstone 2018).В ходе эволюции популяционная структура всегда признавалась одним из наиболее общих механизмов, благоприятствующих кооперации. Даже если отбор благоприятствует несотрудничающим перебежчикам, как это обычно и бывает, сотрудничество все же может развиваться в структурированной популяции. Рассмотрим популяцию, состоящую из особей (нижний уровень), разделенных на группы (верхний уровень). В то время как перебежчики пользуются преимуществом на более низком уровне, кооператоры пользуются преимуществом на более высоком уровне. Если популяция представляла собой одну большую группу, то отбор на более высоком уровне слаб, и преобладают перебежчики.Однако в популяции со многими небольшими группами отбор потенцируется на более высоких уровнях. Группы кооператоров могут образовываться случайно и вытеснять группы перебежчиков (Szathmáry and Demeter 1987).
Без механизмов посредничества в конфликтах сотрудничество не может выжить (Nowak 2006) и не может возникнуть единица более высокого уровня (Radzvilavicius and Blackstone 2018).В ходе эволюции популяционная структура всегда признавалась одним из наиболее общих механизмов, благоприятствующих кооперации. Даже если отбор благоприятствует несотрудничающим перебежчикам, как это обычно и бывает, сотрудничество все же может развиваться в структурированной популяции. Рассмотрим популяцию, состоящую из особей (нижний уровень), разделенных на группы (верхний уровень). В то время как перебежчики пользуются преимуществом на более низком уровне, кооператоры пользуются преимуществом на более высоком уровне. Если популяция представляла собой одну большую группу, то отбор на более высоком уровне слаб, и преобладают перебежчики.Однако в популяции со многими небольшими группами отбор потенцируется на более высоких уровнях. Группы кооператоров могут образовываться случайно и вытеснять группы перебежчиков (Szathmáry and Demeter 1987). Таким образом, более крупные группы (например, синцитиальная LECA) вызывают больше конфликтов, в то время как меньшие группы (особенно половые гаметы) влекут за собой меньше. Следовательно, при больших размерах клеток стохастические процессы могли быть менее важными в опосредовании эволюционного конфликта.
Таким образом, более крупные группы (например, синцитиальная LECA) вызывают больше конфликтов, в то время как меньшие группы (особенно половые гаметы) влекут за собой меньше. Следовательно, при больших размерах клеток стохастические процессы могли быть менее важными в опосредовании эволюционного конфликта.
 В синцитии сегрегация хромосом, вероятно, включает молекулярную самоорганизацию механизма разделения хромосом, который не требует точек закрепления на плазматической мембране.
В синцитии сегрегация хромосом, вероятно, включает молекулярную самоорганизацию механизма разделения хромосом, который не требует точек закрепления на плазматической мембране.
Закрытый митоз, при котором ядерная оболочка остается в основном интактной, считается предком открытого митоза (Cavalier-Smith 2010), что согласуется с нашими собственными результатами (рис. 3). Все варианты митоза имеют общую сеть на основе микротрубочек, которая может быть собрана в пучки или рассыпаться звездообразным образом, тянуться к хромосомам и прикрепляться к кинетохоре, которая присутствовала в LECA (Tromer et al.2019). Однако центросомы не являются существенными для разделения хромосом (Heald et al. 1996). Важно отметить, что эукариотические хромосомы разделяются в основном под действием силы, толкающей вдоль микротрубочек, в которых эукариот-специфичное семейство белков кинезинов играет существенную роль (Shimamoto et al. 2015). Эти механизмы органично согласуются с биологией синцитиальной клетки, поскольку митоз отдельных ядер может происходить независимо от локализованных точек фиксации плазматической мембраны.
Следовательно, происхождение одноядерных жгутиковых простейших можно рассматривать с новой точки зрения.Изображения ближайшего живого родственника архейной клетки-хозяина и бактериального партнера изображают двух сидячих, неподвижных партнеров (Imachi et al. 2020), предлагаемая нами синцитиальная LECA также была сидячей. Центр организации микротрубочек (MTOC), или базальное тело, и способность образовывать жгутики присутствовали у LECA. Эта черта диверсифицировалась среди эукариотических супергрупп и периодически терялась (Yubuki and Leander 2013). Эукариотические жгутики непосредственно связаны с базальными тельцами или формируются de novo зависимым от центриолей образом (Schrøder et al.2011). Комплекс пор жгутиков имеет ряд общих белков с комплексом ядерных пор (Dishinger et al. 2010; Kee et al. 2012; Gould et al. 2016). Мы предполагаем, что жгутик развился на основе (дублированной) структуры, происходящей из центросомы, которая охватывает область плазматической мембраны. Таким образом, одноядерные, жгутиковые споры могли появиться из синцития с актиновым цитоскелетом, поддерживающим окончательное расщепление (Heidstra 2007).
Только споры, содержащие жизнеспособные митонуклеарные взаимодействия и способные к жгутиковому движению, обладали бы свойствами подвижных гамет при условии, что они были способны сливаться с другими в своем роде, что возможно, учитывая склонность самих архей к слиянию (Lange et al. .2011 г.; Гарг и Мартин, 2016 г.; Шалев и др. 2017). Такие споры представляют собой подвижные единицы отбора. Ядро многих жгутиковых простейших расположено в непосредственной близости от базального тела, если не связано с ним, как у многочисленных Archamoebea, Chytridiomycota, Olpidium , Pelagophyceae, Bacillariophyceae, Rhizaria и других (обзор в ссылке 2). Возможно, такие гаметоподобные клетки стали основателями эукариотических супергрупп, каждая из которых содержит жгутиковых представителей, способных генерировать синцитии (рис.3А). У нас нет никаких предположений о физических размерах LECA как синцития, хотя мы предполагаем, что он обитал в морских отложениях (Мартин и Мюллер, 1998), где анаэробная синтрофия необходима для симбиотических взаимодействий (Имачи и др. , 2020). Гифы современных грибов могут занимать площади в квадратные мили (Anderson et al. 2018). LECA мог быть крупной неделящейся многоядерной одноклеточной клеткой, которая порождала супергруппы посредством вытеснения митохондриальных жгутиковых спор. 6-минутный анимационный видеоролик, иллюстрирующий происхождение эукариот от симбиоза и роль синцитиального состояния в жизненном цикле LECA, можно посмотреть по адресу (https://www.youtube.com/watch?v=mmh_IpdgWvw&t=2s)
, 2020). Гифы современных грибов могут занимать площади в квадратные мили (Anderson et al. 2018). LECA мог быть крупной неделящейся многоядерной одноклеточной клеткой, которая порождала супергруппы посредством вытеснения митохондриальных жгутиковых спор. 6-минутный анимационный видеоролик, иллюстрирующий происхождение эукариот от симбиоза и роль синцитиального состояния в жизненном цикле LECA, можно посмотреть по адресу (https://www.youtube.com/watch?v=mmh_IpdgWvw&t=2s)
 Однако наши результаты предполагают, что при происхождении эукариот ядра и митохондрии были единицами отбора и единицами эволюции в пределах синцитиального LECA. Реконструкция состояния предкового признака на основе богатой таксонами выборки, охватывающей все супергруппы, предполагает, что LECA был 1) митохондриальным, 2) многоядерным (синцитиальным, ценоцитным), 3) гаплоидным, 4) с закрытым делением ядра и 5) с половым размножением. Часто утверждается, в том числе во многих работах авторов, что переход от прокариот к эукариотам не оставил промежуточных форм.Однако, если наши текущие мысли примерно соответствуют цели, синцитии на самом деле являются промежуточным состоянием в переходе от прокариот к эукариотам, хотя до сих пор не признавались таковыми. В этом свете синцитии, присутствующие во всех надгруппах эукариот, могут содержать ранее непризнанные формы доказательств эукариотического происхождения и перехода от прокариот к эукариотам.
Однако наши результаты предполагают, что при происхождении эукариот ядра и митохондрии были единицами отбора и единицами эволюции в пределах синцитиального LECA. Реконструкция состояния предкового признака на основе богатой таксонами выборки, охватывающей все супергруппы, предполагает, что LECA был 1) митохондриальным, 2) многоядерным (синцитиальным, ценоцитным), 3) гаплоидным, 4) с закрытым делением ядра и 5) с половым размножением. Часто утверждается, в том числе во многих работах авторов, что переход от прокариот к эукариотам не оставил промежуточных форм.Однако, если наши текущие мысли примерно соответствуют цели, синцитии на самом деле являются промежуточным состоянием в переходе от прокариот к эукариотам, хотя до сих пор не признавались таковыми. В этом свете синцитии, присутствующие во всех надгруппах эукариот, могут содержать ранее непризнанные формы доказательств эукариотического происхождения и перехода от прокариот к эукариотам. «> Отбор таксонов
На основе изучения литературы (дополнительная таблица 4, дополнительный материал онлайн) был создан богатый таксонами (Katz and Grant 2014) набор эукариотических данных, включающий 106 высших таксонов (дополнительная таблица 1, дополнительный материал онлайн). Включены представители шести супергрупп эукариот. Мы используем недавно определенную номенклатуру надгрупп эукариот: Amoebozoa, Archaeplastida, Excavata, Hacrobia, Opisthokonta и SAR , хотя для ясности мы сохранили здесь более знакомый термин Opisthokonta вместо Obazoa. Набор состоит в основном из высших категорий, но в некоторых случаях были включены семейства и роды (табл. 2).
Включены представители шести супергрупп эукариот. Мы используем недавно определенную номенклатуру надгрупп эукариот: Amoebozoa, Archaeplastida, Excavata, Hacrobia, Opisthokonta и SAR , хотя для ясности мы сохранили здесь более знакомый термин Opisthokonta вместо Obazoa. Набор состоит в основном из высших категорий, но в некоторых случаях были включены семейства и роды (табл. 2).
Таблица 2 Обзор таксонов эукариот, рассматриваемых в этом анализе
. Фила
. Классы
. Заказы
. Семьи
. Роды
. ПерСупергруппа
. амёбозои — 7 1 — — 8 архепластиды 7 14 4 — 1 26 Excavata 1 4 3 1 4 13 Hacrobia — 3 5 — — 8 заднежгутиковые 13 9 1 — 1 24 SAR 6 17 3 1 — 27 за ранг 27 54 17 2 6
.
Фила
. Классы
. Заказы
. Семьи
. Роды
. ПерСупергруппа
. амёбозои — 7 1 — — 8 архепластиды 7 14 4 — 1 26 Excavata 1 4 3 1 4 13 Hacrobia — 3 5 — — 8 заднежгутиковые 13 9 1 — 1 24 SAR 6 17 3 1 — 27 за ранг 27 54 17 2 6
Таблица 2 Обзор таксонов эукариот, рассматриваемых в этом анализе
.
Фила
. Классы
. Заказы
. Семьи
. Роды
. ПерСупергруппа
. амёбозои — 7 1 — — 8 архепластиды 7 14 4 — 1 26 Excavata 1 4 3 1 4 13 Hacrobia — 3 5 — — 8 заднежгутиковые 13 9 1 — 1 24 SAR 6 17 3 1 — 27 за ранг 27 54 17 2 6
.
Фила
. Классы
. Заказы
. Семьи
. Роды
. ПерСупергруппа
. амёбозои — 7 1 — — 8 архепластиды 7 14 4 — 1 26 Excavata 1 4 3 1 4 13 Hacrobia — 3 5 — — 8 заднежгутиковые 13 9 1 — 1 24 SAR 6 17 3 1 — 27 за ранг 27 54 17 2 6
 Кладограммы, представленные в этом исследовании, основаны на опубликованных отношениях внутри каждой супергруппы эукариот (дополнительные таблицы 3 и 4, дополнительный материал онлайн). Если мы обозначаем отношения как «разрешенные», это означает, что мы включили соответствующий шаблон ветвления для нашего дерева из 106 таксонов. Паттерны ветвления, которые были «неразрешенными», были преобразованы в политомии.
Кладограммы, представленные в этом исследовании, основаны на опубликованных отношениях внутри каждой супергруппы эукариот (дополнительные таблицы 3 и 4, дополнительный материал онлайн). Если мы обозначаем отношения как «разрешенные», это означает, что мы включили соответствующий шаблон ветвления для нашего дерева из 106 таксонов. Паттерны ветвления, которые были «неразрешенными», были преобразованы в политомии.  Поскольку нет единого мнения о том, где находится корень эукариот Eukarya, был подготовлен набор эталонных деревьев с коллекцией опубликованных предложений по корню эукариот: Excavata или внутри раскопок (Cavalier-Smith 2002; He et al.2014; Триа и др. 2021), Opisthokonta , Fungi или внутри Fungi (например, Microsporidia ) (Vossbrinck et al. 1987), Amoebozoa или внутри (Stechmann and Cavalier-Smith 34 102-Smith 2 Amorphaea ( Opisthokonta + Amoebozoa ) (Derelle et al. 2015). Также был подготовлен нерутированный набор. Подробные данные, лежащие в основе всех параметров и деревьев, представлены в дополнительной таблице 3 «Дополнительный материал онлайн».
Поскольку нет единого мнения о том, где находится корень эукариот Eukarya, был подготовлен набор эталонных деревьев с коллекцией опубликованных предложений по корню эукариот: Excavata или внутри раскопок (Cavalier-Smith 2002; He et al.2014; Триа и др. 2021), Opisthokonta , Fungi или внутри Fungi (например, Microsporidia ) (Vossbrinck et al. 1987), Amoebozoa или внутри (Stechmann and Cavalier-Smith 34 102-Smith 2 Amorphaea ( Opisthokonta + Amoebozoa ) (Derelle et al. 2015). Также был подготовлен нерутированный набор. Подробные данные, лежащие в основе всех параметров и деревьев, представлены в дополнительной таблице 3 «Дополнительный материал онлайн».  Все признаки были численно закодированы для реконструкции наследственного состояния (дополнительная таблица 5, дополнительный материал онлайн).Для многоядерного состояния признак кодировался как 1, когда имелось указание на многоядерное состояние, присутствующее во всей группе или части жизненного цикла многих (> 2 родов) членов, или если оно присутствовало в неразрешенных группах. Признак считался неоднозначным (0/1) для группы, когда либо существовал консенсус в отношении того, что многоядерное состояние присутствует у одного производного вида в группе, либо когда были доказательства наличия стадий жизненного цикла, которые очень напоминают синцитии. -подобные структуры без четкого описания у членов группы.Наконец, признак был закодирован как 0, когда вообще не было указаний на существование многоядерного состояния в пределах известного разнообразия определенного таксона. Источники информации обобщены в дополнительных таблицах 2 и 4, Дополнительные материалы онлайн.
Все признаки были численно закодированы для реконструкции наследственного состояния (дополнительная таблица 5, дополнительный материал онлайн).Для многоядерного состояния признак кодировался как 1, когда имелось указание на многоядерное состояние, присутствующее во всей группе или части жизненного цикла многих (> 2 родов) членов, или если оно присутствовало в неразрешенных группах. Признак считался неоднозначным (0/1) для группы, когда либо существовал консенсус в отношении того, что многоядерное состояние присутствует у одного производного вида в группе, либо когда были доказательства наличия стадий жизненного цикла, которые очень напоминают синцитии. -подобные структуры без четкого описания у членов группы.Наконец, признак был закодирован как 0, когда вообще не было указаний на существование многоядерного состояния в пределах известного разнообразия определенного таксона. Источники информации обобщены в дополнительных таблицах 2 и 4, Дополнительные материалы онлайн. 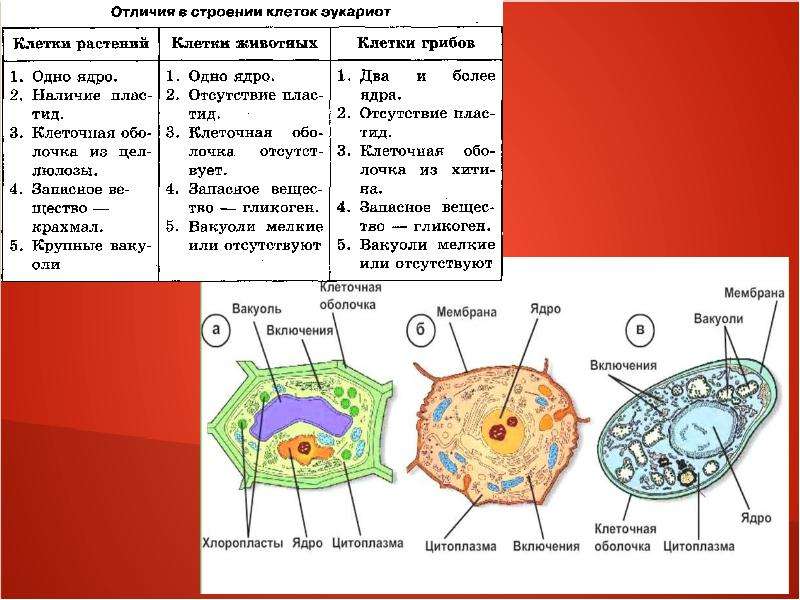 6 (https://www.mesquiteproject.org/). Всего было отобрано 106 таксонов эукариот (от рода до уровня типа) (см. Выбор таксонов, дополнительная таблица 1, Дополнительный материал онлайн). Затем был построен набор из 30 эталонных деревьев с разным положением корня и разной степенью свободы в отношении полифилии (см. Установка топологии и корня дерева, дополнительная таблица 3, Дополнительный онлайн-материал). Матрица признаков была подготовлена на основе литературных данных по шести признакам: наличие многоядерного состояния, наличие полиплоидов, наличие пола/мейоза, поведение ядерной оболочки при делении, тип митохондрий и тип пластиды (дополнительные таблицы 2 и 5, Дополнительный материал онлайн).В некоторых группах одни члены группы проявляют одну черту, а другие — другую. В подобных случаях для этой группы кодировались оба признака (0/1 или 1/2).
6 (https://www.mesquiteproject.org/). Всего было отобрано 106 таксонов эукариот (от рода до уровня типа) (см. Выбор таксонов, дополнительная таблица 1, Дополнительный материал онлайн). Затем был построен набор из 30 эталонных деревьев с разным положением корня и разной степенью свободы в отношении полифилии (см. Установка топологии и корня дерева, дополнительная таблица 3, Дополнительный онлайн-материал). Матрица признаков была подготовлена на основе литературных данных по шести признакам: наличие многоядерного состояния, наличие полиплоидов, наличие пола/мейоза, поведение ядерной оболочки при делении, тип митохондрий и тип пластиды (дополнительные таблицы 2 и 5, Дополнительный материал онлайн).В некоторых группах одни члены группы проявляют одну черту, а другие — другую. В подобных случаях для этой группы кодировались оба признака (0/1 или 1/2).  F.M. и С.Г.Г. WFM также хотел бы поблагодарить ERC (666053) и VW Foundation (
F.M. и С.Г.Г. WFM также хотел бы поблагодарить ERC (666053) и VW Foundation (
и 96742) за финансирование. S.B.G благодарит VolkswagenStiftung (Life) за финансирование.Мы благодарим профессора Клауса В. Коваллика за полезные обсуждения.
«> Доступность данных
Все данные, связанные с рукописью, представлены в дополнительной информации.
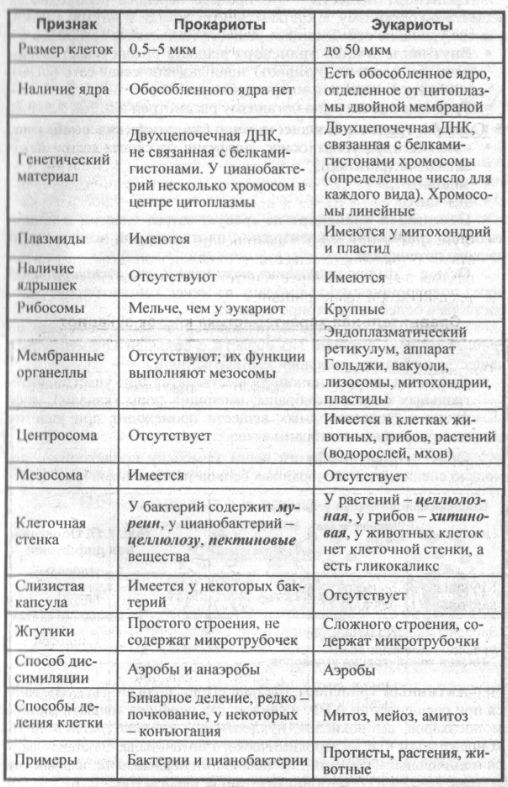
Proc Biol Sci
. 285
( 1893
): 20182233
.
Арчибальд
ДМ
, Симпсон
АГБ
, Сламовиц
Ч
, редакторы. 2017
. Справочник протистов. Спрингер Природа.
Блэкстоун
СЗ
, Зеленый
ДР.
1999
. Эволюция механизма клеточного самоубийства
. Биоэссе
. 21
( 1
): 84
– 88
.
Бетчер
Б
, Барраль
Ю.
2013
. Клеточная биология открытого и закрытого митоза
. Ядро
4
( 3
): 160
– 165
.
Брюкнер
Дж
, Мартин
ВФ.
2020
. Генов бактерий больше, чем генов архей в эукариотических геномах
. Геном Биол Эвол
. 12
( 4
): 282
– 292
.
Брунк
CF
, Мартин
ВФ.
2019
.
Вклад гистонов архей в происхождение эукариот
. Trends Microbiol
. 27
( 8
): 703
– 714
.
Шина
LW.
1987
. Эволюция индивидуальности. Нью-Джерси: Издательство Принстонского университета.
Кавалер-Смит
Т.
1987
. Происхождение клеток эукариот и архебактерий
. Ann NY Acad Sci
. 503
: 17
– 54
.
Кавалер-Смит
Т.
2002
. Фаготрофное происхождение эукариот и филогенетическая классификация простейших
. Int J Syst Evol Micro
. 52
( 2
): 297
– 354
.
Кавалер-Смит
Т.
2010
. Происхождение клеточного ядра, митоза и пола: роль внутриклеточной коэволюции
. Биол Директ
. 5
: 7
.
Кавалер-Смит
Т.
2018
. Царство Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание перипластидных белков, эволюцию цитоскелета и перипластид и древние расхождения
.
Протоплазма
255
( 1
): 297
– 357
.
Сумки
JB
, и другие. 2016
. Меняющийся взгляд на эукариогенез – окаменелости, клетки, родословные и то, как они все объединяются
. J Cell Sci
. 129
( 20
): 3695
– 3703
.
Добенмайр
РФ.
1936
. Использование терминов ценоцит и синцитий в биологии
. Наука
. 84
( 2189
): 533
– 533
.
де Дюв
К.
1969
. Эволюция пероксисомы
. Ann NY Acad Sci
. 168
( 2
): 369
– 381
.
Дерелле
Р
, и другие. 2015
. Бактериальные белки указывают на единственный эукариотический корень
. Proc Natl Acad Sci U S A
. 112
( 7
): E693
– E699
.
Диррен
С
, Почта
Т.
2016
. Беспорядочное и специфическое приобретение симбионтов бактериями из рода амебоидных Nuclearia (Opisthokonta)
. FEMS Microbiol Ecol
. 92
( 8
): fiw105
.
Посудомоечная машина
J
, и другие. 2010
. Вход в реснички мотора кинезина-2 KIF17 регулируется импортином-β2 и RanGTP
. Nat Cell Biol
. 12
( 7
): 703
– 710
.
Дулиттл
ВФ.
1998
. Вы то, что вы едите: механизм переноса генов может объяснить бактериальные гены в ядерных геномах эукариот
. Тренды Genet
. 14
( 8
): 307
– 311
.
Эмбли
ТМ
, Мартин
ВФ.
2006
. Эволюция эукариот, изменения и вызовы
. Природа
440
( 7084
): 623
– 630
.
Фу
С
, Коэльо
МА
, Давид-Пальма
М
, Священник
СЖ
, Хейтман
Дж.
2019
. Генетическая и геномная эволюция полового размножения: эхо от LECA до царства грибов
. Curr Opin Genet Dev
. 58-59
: 70
– 75
.
Гарг
СГ
, Мартин
ВФ.
2016
. Митохондрии, клеточный цикл и происхождение пола через синцитиального общего предка эукариот
. Геном Биол Эвол
. 8
( 6
): 1950
– 1970
.
Гейзен
С
, и другие. 2017
. Перезагрузка почвенной протистологии: 30 фундаментальных вопросов для начала
. Почвенный Биол Биохим
. 111
: 94
– 103
.
Гейтлер
Л.
1953
. Эндомитоза и амитополиплоидизация. Протоплазматология. Справочник Der Protoplasmaforschung Bd. VI: Kern- Und Zellteilung. Том. 6:С. Спрингер, Вена. п. 89
.
Гулд
ГВт
, Дринг
ГДж.
1979
.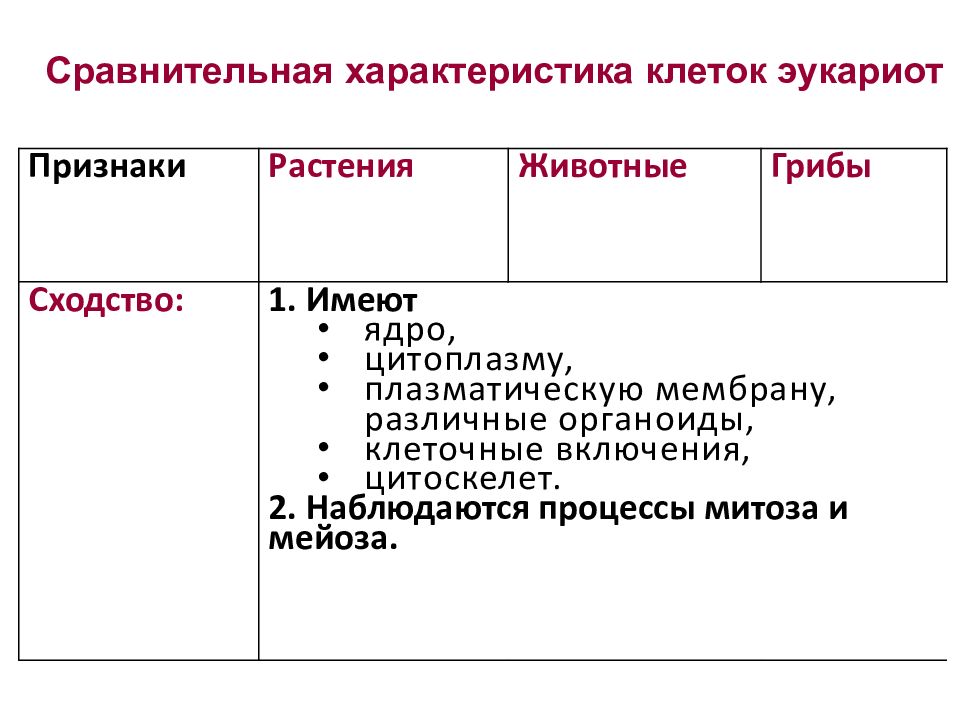
О возможной связи образования эндоспор бактерий с происхождением эукариотических клеток
. Дж Теор Биол
. 81
( 1
): 47
– 53
.
Гулд
СБ
, Уоллер
РФ
, Макфадден
GI.
2008
. Эволюция пластид
. Annu Rev Plant Biol
. 59
: 491
– 517
.
Гулд
СБ
, Гарг
СГ
, Мартин
ВФ.
2016
. Секреция бактериальных пузырьков и эволюционное происхождение эукариотической эндомембранной системы
. Trends Microbiol
. 24
( 7
): 525
– 534
.
Гулд
СБ
, и другие. 2019
. Адаптация к жизни на суше и высокому содержанию кислорода путем перехода от ферредоксина к НАДН-зависимому окислительно-восстановительному балансу
. Proc R Soc B
. 286
( 2019149
).
Гупта
RS
, Голдинг
ГБ.
1996
. Происхождение эукариотической клетки
. Trends Biochem Sci
. 21
( 5
): 166
– 171
.
Хаджи
Дж.
1953
. Попытка реконструировать систему классификации животных
. Сист Зоол
. 2
: 145
– 154
.
Он
Д
, и другие. 2014
. Альтернативный корень эукариотического древа жизни
. Карр Биол
. 24
( 4
): 465
– 470
.,
Хильда
Р
, и другие. 1996
. Самоорганизация микротрубочек в биполярные веретена вокруг искусственных хромосом в экстрактах яиц Xenopus
. Природа
382
( 6590
): 420
– 425
.
Хейдстра
Р.
2007
. Асимметричное деление клеток в развитии растений. В: Macieira-Coelho
A
, редактор. Асимметричное деление клеток
. Verlag Berlin Heidelberg:
Verlag Berlin Heidelberg: Springer
. п. 1
– 37
.
Хофштеттер
PG
, Лар
DJ.
2019
. Все эукариоты являются половыми, если не доказано обратное: многие так называемые асексуалы имеют мейотический аппарат и могут заниматься сексом
. Bioessays
41(6):e1800246.
Хориике
Т
, Хамада
К
, Мията
Д
, Шинозава
Т.
2004
. Происхождение эукариот предполагается как симбиоз Pyrococcus с γ-протеобактериями по филогенетическому древу на основе содержания генов
. Дж Мол Эвол
. 59
( 5
): 606
– 619
.
Имачи
Н
, и другие. 2020
. Изоляция археона на границе прокариотов и эукариот
. Природа
577
( 7791
): 519
– 525
.
Жаво
EJ
, Холм
АХ
, Вальтер
МР.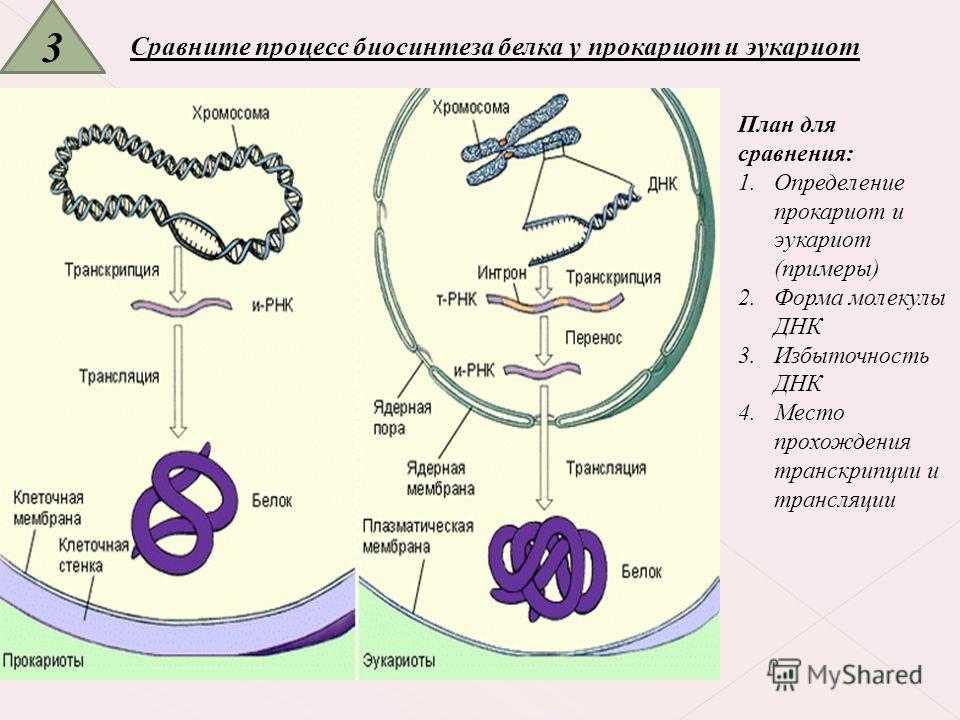
2001
. Морфологическая и экологическая сложность ранних эукариотических экосистем
. Природа
412
( 6842
): 66
– 69
.
Жаво
EJ
, Лепо
К.
2018
. Палеопротерозойская летопись окаменелостей: значение для эволюции биосферы в средние века Земли
. Earth-Sci Rev
. 176
: 68
– 86
.
Камикава
Р
, и другие. 2014
. Эволюция содержания генов в дискобидных митохондриях, выведенная из филогенетического положения и полного митохондриального генома Tsukubamonas globosa
. Геном Биол Эвол
. 6
( 2
): 306
– 315
.
Кац
ЛА
, Грант
Младший.
2015
. Филогеномный анализ, богатый таксонами, разрешает эукариотическое древо жизни и раскрывает силу подвыборки по участкам
. Сист Биол
.
64
( 3
): 406
– 415
.
Ки
HL
, и другие. 2012
. Барьер проницаемости исключения размера и нуклеопорины характеризуют комплекс цилиарных пор, который регулирует транспорт в реснички
. Nat Cell Biol
. 14
( 4
): 431
– 437
.,
Килинг
ПЖ.
2004
. Разнообразие и эволюционная история пластид и их хозяев
. Am J Bot
. 91
( 10
): 1481
– 1493
.
Поцелуй
E
, и другие. 2019
. Сравнительная геномика раскрывает происхождение грибных гиф и многоклеточность
. Нац Коммуна
. 10
( 1
): 13
.
Клинебергер
Е.
1917
. Über die Größe und Beschaffenheit der Zellkerne mit besonderer Berücksichtigung der Systematik (Inaug.-Diss. Frankfurt)
. Beihefte Бот. Збл
. 35 I
: 219
– 278
.
Холм
АХ
, Жаво
ЭДЖ
, Хьюитт
Д
, Коэн
стр.
2006
. Эукариотические организмы в протерозойских океанах
. Philos Trans R Soc Lond B Biol Sci
. 361
( 1470
): 1023
– 1038
.
Коваллик
КВ
, Мартин
ВФ.
2021
. Рассвет симбиогенеза: аннотированный английский перевод статьи Мерешковского 1910 года
. БиоСистемс
199
: 104281
.
Озеро
JA
, Ривера
MC.
1994
. Было ли ядро первым эндосимбионтом?
Proc Natl Acad Sci U S A
. 91
( 8
): 2880
– 2881
.
Пер.
N
, Мартин
ВФ.
2012
. Происхождение мембранной биоэнергетики
. Сотовый
151
( 7
): 1406
– 1416
.
Линдос
AC
, и другие.
2008
. Уникальный механизм деления клеток у архей
. Proc Natl Acad Sci U S A
. 105
( 48
): 18942
– 18946
.
Ланге
С
, Зерулла
К
, Брейерт
С
, Соппа
Дж.
2011
. Конверсия генов приводит к выравниванию копий генома у полиплоидных галоархей Haloferax volcanii
. Мол Микробиол
. 80
( 3
): 666
– 677
.
Лейс
СП.
2003
. Значение синцитиальных тканей для положения Hexactinellida в Metazoa
. Интегр Комп Биол
. 43
( 1
): 19
– 27
.
Лопес-Гарсия
P
, Морейра
Д.
1999
. Метаболический симбиоз у эукариот
. Trends Biochem Sci
. 24
( 3
): 88
– 93
.
Лопес-Гарсия
P
, Верещака
А
, Морейра
Д.
2007
. Эукариотическое разнообразие, связанное с карбонатами и поверхностью раздела флюид-морская вода в гидротермальном поле Лост-Сити
. Энвайрон Микробиол
. 9
( 2
): 546
– 554
.
Лопес-Гарсия
P
, Морейра
Д.
2015
. Открытые вопросы о происхождении эукариот
. Тренды Ecol Evol
. 30
( 11
): 697
– 708
.
Лопес-Гарсия
P
, Морейра
Д.
2020
. Пересмотр гипотезы синтрофии происхождения эукариот
. Нат Микробиол
. 5
( 5
): 655
– 667
.
Мацишевский
К
, Карнковска
А.
2019
. Мне остаться или уйти? Задержка и потеря компонентов в рудиментарных эндосимбиотических органеллах
. Curr Opin Genet Dev
. 58-59
: 33
– 39
.
Мартин
Дж
, Эттема
TJG.
2013
. От археи к эукариоту: темные века эволюции эукариотической клетки
. Биохим Сок Т
. 41
( 1
): 451
– 457
.
Мартин
WF
, Мюллер
М.
1998
. Водородная гипотеза первого эукариота
. Природа
392
( 6671
): 37
– 41
.
Мартин
WF
, Гарг
С
, Зиморский
В.
2015
. Эндосимбиотические теории происхождения эукариот
. Philos Trans R Soc Lond B Biol Sci
. 370
( 1678
): 20140330
.
Мартин
WF
, и другие. 2017а
. Позднее митохондриальное происхождение является артефактом
. Геном Биол. Эвол
. 9
( 2
): 373
– 379
.
Мартин
WF
, Тиленс
АГМ
, Ментел
М
, Гарг
СГ
, Гулд
СБ.
2017b
. Физиология фагоцитоза в контексте митохондриального происхождения
. Микробиол Мол Биол Ред.
. 81
( 3
): 17
.
Мейнард Смит
Дж
, Сатмари
E.
1995
. Основные переходы в эволюции. Оксфорд: Издательство Оксфордского университета.
Мехта
АП
, и другие. 2018
. Создание эндосимбионтов дрожжей как шаг к эволюции митохондрий
. Proc Natl Acad Sci U S A
. 115
( 46
): 11796
– 11801
.
Мерешковский
С.
1905
. Über Natur und Ursprung der Chromatophoren im Pflanzenreiche. Биол Центральный бл. 25:293-604. [Английский перевод в Martin W, Kowallik K. 1999 Аннотированный английский перевод статьи Мерешковского 1905 года «Über Natur und Ursprung der Chromatophoren im Pflanzenreiche. евро. Дж. Фикол. 34, 287–295].
Мерешковский
С.
1910
. Английский перевод в Коваллик К.В., Мартин В.Ф. 2021. Происхождение симбиогенеза: аннотированный английский перевод статьи Мерешковского 1910 года о теории двух плазменных линий. БиоСист. биол. Центральный бл. 30:104281. Theorie der zwei Plasmaarten als Grundlage der Symbiogenesis, einer neuen Lehre von der Entstehung der Organismen. 278–288; 289–303; 321–347; 353–367. [199].
Английский перевод в Коваллик К.В., Мартин В.Ф. 2021. Происхождение симбиогенеза: аннотированный английский перевод статьи Мерешковского 1910 года о теории двух плазменных линий. БиоСист. биол. Центральный бл. 30:104281. Theorie der zwei Plasmaarten als Grundlage der Symbiogenesis, einer neuen Lehre von der Entstehung der Organismen. 278–288; 289–303; 321–347; 353–367. [199].
Мишо
RE.
1999
. Дарвиновская динамика. Оксфорд: Издательство Оксфордского университета.
Мельницы
ДБ.
2020
. Происхождение фагоцитоза в истории Земли
. Интерфейс Фокус
. 10
( 4
): 20200019
.
Мора
С
, Титтензор
P
, Адл
СМ
, Симпсон
АГ
, Червь
Б.
2011
. Сколько видов существует на Земле и в океане?
ПЛОС Биол
. 9
( 8
): e1001127
.
Мюллер
М
, и другие.
2012
. Биохимия и эволюция анаэробного энергетического обмена у эукариот
. Микробиол Мол Биол Ред.
. 76
( 2
): 444
– 495
.
Нагиес
FSP
, Брюкнер
Дж
, Триа
ФДК
, Мартин
ВФ.
2020
. Спектр вертикальности генов
. ПЛОС Жене
. 16
( 11
): e1009200
.
Никлас
КДж
, Кобб
ЭД
, Кроуфорд
ДР.
2013
. Эво-дево многоядерных клеток, тканей и организмов и альтернативный путь к многоклеточности
. Эвол Дев
. 15
( 6
): 466
– 474
.
Новак
Массачусетс.
2006
. Эволюционная динамика: исследование уравнений жизни. Кембридж: Издательство Гарвардского университета.
Песоле
Г
, и другие. 2012
. Забытый геном
.
EMBO Реп.
. 13
( 6
): 473
– 474
.
Питтис
АА
, Габальдон
Т.
2016
. Позднее приобретение митохондрий хозяином химерного прокариотического происхождения
. Природа
531
( 7592
): 101
– 104
.
Радзвилавичюс
АЛ
, Блэкстоун
СЗ.
2018
. Новый взгляд на эволюцию индивидуальности
. Biol Rev Camb Philos Soc
. 93
( 3
): 1620
– 1633
.
Рамеш
МА
, Малик
СБ
, Логсдон
Дж.М.
младший
2005
. Филогеномный перечень мейотических генов: свидетельство пола у Giardia и раннее эукариотическое происхождение мейоза
. Карр Биол
. 15
( 2
): 185
– 191
.
Рэнд
Немецкая марка
, Моссман
Дж. А.
А.
2020
. Митоядерный конфликт и сотрудничество управляют интеграцией генотипов, фенотипов и окружающей среды
. Philos Trans R Soc B
. 375
( 1790
): 201
.
Саган
Л.
1967
. О происхождении митозирующих клеток
. J Теорет Биол
. 14
( 3
): 225
–274.
Санчес-Баракальдо
P
, Ворон
ДЖА
, Пизани
Д
, Нолл
АХ.
2017
. Ранние фотосинтезирующие эукариоты населяли слабосоленые местообитания
. Proc Natl Acad Sci U S A
. 114
( 37
): E7737
– E7745
.
Шредер
JM
, и другие. 2011
. EB1 и EB3 способствуют биогенезу ресничек с помощью нескольких механизмов, связанных с центросомами
. J Cell Sci
. 124
( Pt 15
): 2539
– 2551
.,
Шалев
Y
, Тургеман-Гротт
I
, Тамир
А
, Эйхлер
Дж
, Гофна
У.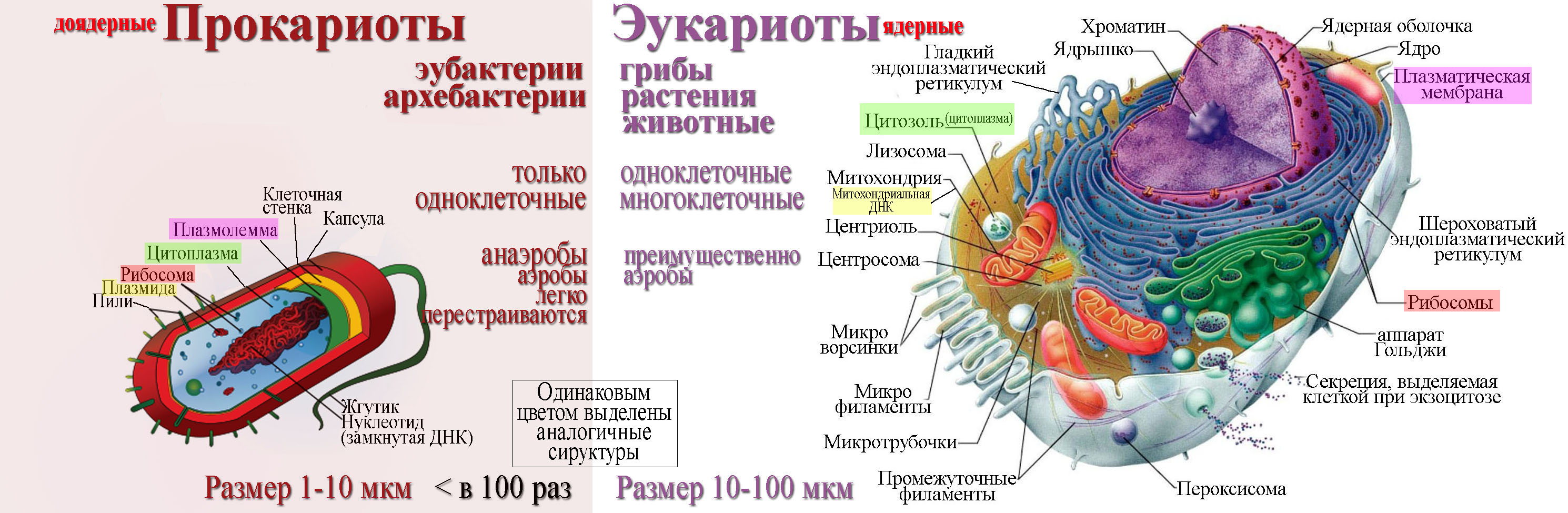
2017
. Гликозилирование клеточной поверхности необходимо для эффективного спаривания Haloferax volcanii
. Фронт Микробиол
. 8
: 1253
.
Шимамото
Y
, Форт
С
, Капур
ТМ.
2015
. Измерение сил толкания и торможения, генерируемых ансамблями кинезина-5, сшивающими две микротрубочки
. Dev Cell
. 34
( 6
): 669
– 681
.
Ситте
Р
, Циглер
Н
, Эрендорфер
Ф
, Бресинский
А.
1991
. Strasburger, Lehrbuch der Botanik für Hochschulen
. 33-е изд. Штутгарт
: Густав Фишер
.
Спанг
А
, и другие. 2015
. Сложные археи, занимающие промежуточное положение между прокариотами и эукариотами
. Природа
. 521
( 7551
): 173
– 179
.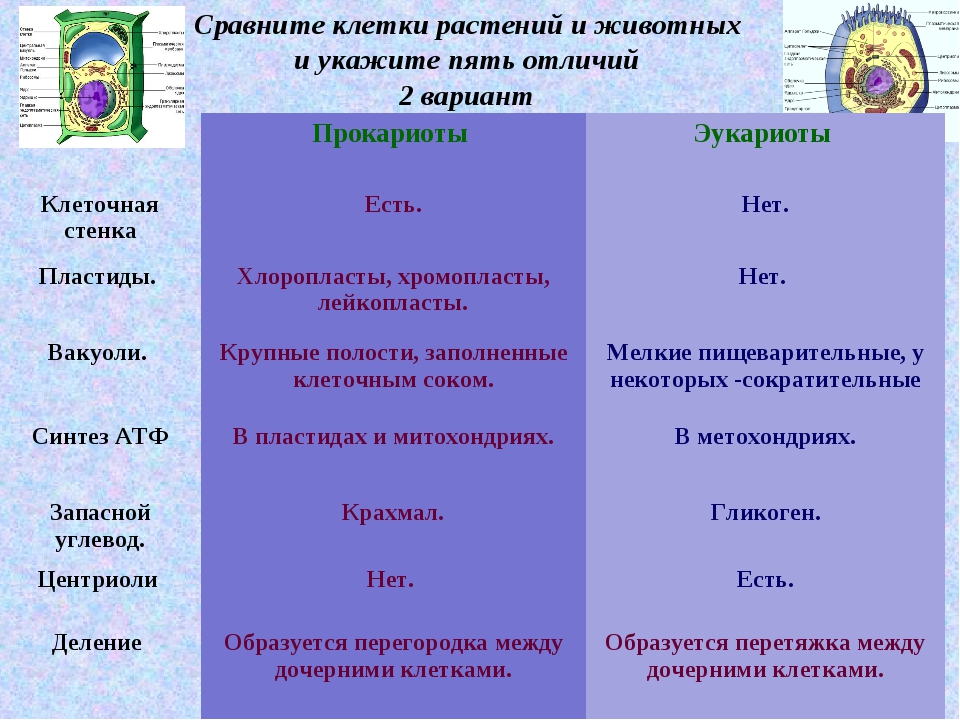
Шпейер
Д
, Лукеш
Дж
, Элиаш
М.
2015
. Секс — вездесущий, древний и неотъемлемый атрибут эукариотической жизни
. Proc Natl Acad Sci U S A
. 112
( 29
): 8827
– 8834
.
Штехманн
А
, Кавалер-Смит
Т.
2002
. Укоренение дерева эукариот с использованием производного слияния генов
. Наука
297
( 5578
): 89
– 91
.
Страсбургер
E.
1908
. Einiges über Characeen und Amitose. Wiesner Festschrift im Auftrage Des Festkomitees Redigiert Von K. Linsbauer
. Вена: К. Конеген.
р. 24
– 47
.
Сатмари
E
, Деметра
Л.
1987
. Модель стохастического корректора
. Дж Теор Биол
. 128
( 4
): 463
– 486
.
Тан
Q
, Панг
К
, юаней
XL
, Сяо
S-H.
2020
. Многоклеточный хлорофит возрастом один миллиард лет
. Нат Экол Эвол
. 4
( 4
): 543
– 547
.
Тромер
ЕС
, Ван Хофф
JJE
, Копс
GJPL
, Снел
Б.
2019
. Мозаичное происхождение эукариотических кинетохор
. Proc Natl Acad Sci U S A
. 116
( 26
): 12873
– 12882
.
Триа
ФДК
и другие. 2021
. Дупликации генов прослеживают митохондрии до начала сложности эукариот. Геном Биол Эвол.
Воссбринк
CR
, и другие. 1987
. Последовательность рибосомной РНК предполагает, что микроспоридии являются очень древними эукариотами
. Природа
326
( 6111
): 411
– 414
.,
Воссеберг
Дж
, и другие. 2021
. Определение времени происхождения эукариотической клеточной сложности с древними дупликациями
.
Нат Экол Эвол
. 5
( 1
): 92
– 99
.
Юбуки
N
, Леандер
БС.
2013
. Эволюция центров организации микротрубочек у эукариот
. Завод J
. 75
( 2
): 230
– 244
.
Заремба-Недзведска
К
, и другие. 2017
. Археи Асгарда проливают свет на происхождение эукариотической клеточной сложности
. Природа
541
( 7637
): 353
– 358
.
© Автор(ы), 2021 г. Опубликовано Oxford University Press от имени Общества молекулярной биологии и эволюции.
Это статья в открытом доступе, распространяемая в соответствии с условиями некоммерческой лицензии Creative Commons Attribution (http://creativecommons.org/licenses/by-nc/4.0/), которая разрешает некоммерческое повторное использование, распространение, и воспроизведение на любом носителе при условии надлежащего цитирования оригинальной работы. Для коммерческого повторного использования, пожалуйста, свяжитесь с [email protected]
Для коммерческого повторного использования, пожалуйста, свяжитесь с [email protected]
Происхождение эукариот – эволюционная биология
Обзоры эукариогенеза см. в Martin, et al.2015 и Baum 2015. Martin 2005 представляет более старый, но все еще полезный обзор, тогда как Harold 2014 представляет собой доступное исследование эволюции клеток длиной в книгу. Гулд и др. 2008 фокусируется на приобретении пластид и последующих дополнительных эндосимбиотических событиях. Кунин и др. 2010 и Ломбард и др. 2012 обсуждают регуляторные сети белков и химию мембран в трех сферах жизни и их значение для эукариогенеза. Эме и др. 2014 связывает молекулярные данные, которые определяют большую часть поля, с ископаемыми свидетельствами.Уильямс и др. 2013 обобщает филогенетические аргументы в пользу филогенетической модели, которая доминирует в современном мышлении, а именно, что археи/эукариоты и бактерии представляют собой два основных домена жизни. Эме и др. 2017 строится на отношениях между археями и моделями эукариогенеза.
Баум, Д. А. 2015. Сравнение аутогенных теорий происхождения эукариотических клеток. Американский журнал ботаники 102:1954–1965.
Сравнивает различные аутогенные теории происхождения ядра и эукариогенеза в целом в контексте клеточной топологии, соответствия современной клеточной биологии и времени приобретения митохондрий.
Эме, Л., С. К. Шарп, М. В. Браун и др. 2014. О возрасте эукариот: оценка данных окаменелостей и молекулярных часов. Перспективы Колд-Спринг-Харбор в биологии 6:a016139.
Обзор филогенетических и ископаемых данных о возрасте эукариот.
Эме, Л., А. Спанг, Дж. Ломбард, К. У. Стейрс и Т. Дж. Г. Эттема. 2017. Археи и происхождение эукариот. Nature Reviews Microbiology 15:711–723.
Обсуждает модели эукариогенеза в свете недавно открытых и охарактеризованных линий архей.
Гулд С. Б., Р. Ф. Уоллер и Г. И. Макфадден. 2008. Эволюция пластид. Ежегодный обзор биологии растений 59:491–517.
Обеспечивает всесторонний обзор эволюции пластид, включая первичные и вторичные эндосимбиозы, нацеливание белков на пластиды и метаболизм пластид.
Гарольд Ф. М. 2014. В поисках клеточной истории: эволюция строительных блоков жизни .Чикаго: Чикагский ун-т. Нажмите.
Дает синтетический обзор происхождения и эволюции клеток, уделяя особое внимание происхождению эукариот.
Кунин Е. В., Дэкс Дж., Дулитл В. и др. 2010. Происхождение и ранняя эволюция эукариот в свете филогеномики. Биология генома 11:209.
Анализ происхождения ключевых регуляторных модулей эукариотических белков с использованием сравнительной геномики.
Ломбард, Дж., П.Лопес-Гарсия и Д. Морейра. 2012. Ранняя эволюция липидных мембран и три домена жизни. Nature Reviews Microbiology 10:507–515.
Обзор молекулярного состава фосфолипидных мембран архей и бактерий и последствия для моделей эукариогенеза.
Martin, W. F. 2005. Архебактерии (археи) и происхождение эукариотического ядра. Текущее мнение по микробиологии 8:630–637.
F. 2005. Архебактерии (археи) и происхождение эукариотического ядра. Текущее мнение по микробиологии 8:630–637.
Обобщает разнообразие моделей происхождения ядерного компартмента, выступая против ядерных эндосимбиотических моделей.
Мартин В. Ф., С. Гарг и В. Зиморски. 2015. Эндосимбиотические теории происхождения эукариот. Philosophical Transactions of the Royal Society B: Biological Science 370.1678: 20140318.
Обзор моделей эукариогенеза с историческим уклоном, уделяя особое внимание происхождению ядерного и митохондриального компартментов, а также метаболическим соображениям.
Williams, T. A., P. G. Foster, C. J. Cox, et al. 2013. Архейное происхождение эукариот поддерживает только два основных домена жизни. Природа 504:231–236.
Обобщает поддержку существования только двух основных доменов жизни, при этом эукариоты встроены в парафилетические археи.
Мембраны, энергетика и эволюция между прокариотами и эукариотами
Любая попытка определить значение мембран для клеточной эволюции должна учитывать высокую стоимость биосинтеза липидных молекул. Существует два способа количественной оценки таких затрат. Во-первых, с эволюционной точки зрения стоимость синтеза молекулы принимается как сумма прямого использования АТФ в пути биосинтеза плюс косвенная потеря АТФ в результате использования метаболических предшественников, которые в противном случае были бы преобразованы в АТФ и доступны для альтернативных клеточных функций (Akashi and Gojobori, 2002; Lynch, Marinov, 2015).Во-вторых, для простой количественной оценки прямого вклада в общую потребность клетки в АТФ затраты на отклонение метаболических предшественников игнорируются.
Существует два способа количественной оценки таких затрат. Во-первых, с эволюционной точки зрения стоимость синтеза молекулы принимается как сумма прямого использования АТФ в пути биосинтеза плюс косвенная потеря АТФ в результате использования метаболических предшественников, которые в противном случае были бы преобразованы в АТФ и доступны для альтернативных клеточных функций (Akashi and Gojobori, 2002; Lynch, Marinov, 2015).Во-вторых, для простой количественной оценки прямого вклада в общую потребность клетки в АТФ затраты на отклонение метаболических предшественников игнорируются.
Путем суммирования общих затрат всех молекул, лежащих в основе клеточного признака, и масштабирования по расходу энергии клетки в течение жизни можно получить меру относительного расхода энергетического бюджета клетки, связанного с формированием и поддержанием признака. Затем это измерение, sc, можно рассматривать как частичное увеличение энергетического бюджета клетки, которое могло бы быть направлено на рост, размножение и выживание в отсутствие таких вложений, игнорируя прямые преимущества приспособленности, связанные с выражением признака, sa. Чтобы отбор был эффективным, чистое селективное преимущество признака sn=sa-sc должно превышать мощность случайного генетического дрейфа, 1/Ne у гаплоидных видов и 1/(2Ne) у диплоидных, где Ne — эффективная численность населения.
Чтобы отбор был эффективным, чистое селективное преимущество признака sn=sa-sc должно превышать мощность случайного генетического дрейфа, 1/Ne у гаплоидных видов и 1/(2Ne) у диплоидных, где Ne — эффективная численность населения.
Большинство клеточных мембран преимущественно состоят из глицерофосфолипидов, которые, несмотря на наличие различных головных групп (например, глицерин, холин, серин и инозитол), имеют общую стоимость биосинтеза на молекулу (в единицах гидролиза АТФ, включая стоимость отведения промежуточные метаболиты)
(2а)cL≃320+[38⋅(NL−16)]+(6⋅NU), (2b)cL≃340+[40⋅(NL−16)]+(6⋅NU),
у бактерий и эукариот соответственно, где NL — средняя длина цепи жирной кислоты, а NU — среднее количество ненасыщенных атомов углерода в цепи жирной кислоты (дополнительный материал).Хотя варианты глицерофосфолипидов используются у различных видов (Guschina and Harwood, 2006; Geiger et al., 2010), они структурно достаточно сходны, так что предыдущие выражения должны по-прежнему обеспечивать превосходные приближения первого порядка. Приведенные (прямые) затраты, которые не учитывают потерю АТФ-генерирующего потенциала из-за утечки метаболических предшественников, составляют
Приведенные (прямые) затраты, которые не учитывают потерю АТФ-генерирующего потенциала из-за утечки метаболических предшественников, составляют
.
(3a)cL′≃110+[7⋅(NL−16)]+(6⋅NU), (3b)cL′≃120+[9⋅(NL−16)]+(6⋅NU),
у бактерий и эукариот соответственно.С точки зрения общего энергетического баланса клетки эволюционная стоимость липидной молекулы равна cL/CT.
Для большинства липидов в биологических мембранах 14≤NL≤22 и 0≤NU≤6, поэтому стоимость молекулы липида обычно находится в диапазоне cL≃200–600 АТФ, хотя среднее значение для всех липидов, развернутых в видоспецифичных мембран значительно уже (внизу). Кардиолипин, который редко составляет более 20% мембранных липидов, является исключительным, имея эволюционную стоимость ~ 640 АТФ / молекула (и уменьшенную стоимость ~ 240 АТФ).Чтобы представить эти расходы в перспективе, эволюционные затраты на биосинтез каждого из четырех нуклеотидов составляют ≈ 50 гидролизов АТФ на молекулу (Lynch and Marinov, 2015), тогда как средняя стоимость аминокислоты составляет ≈30 АТФ (Atkinson, 1970; Akashi).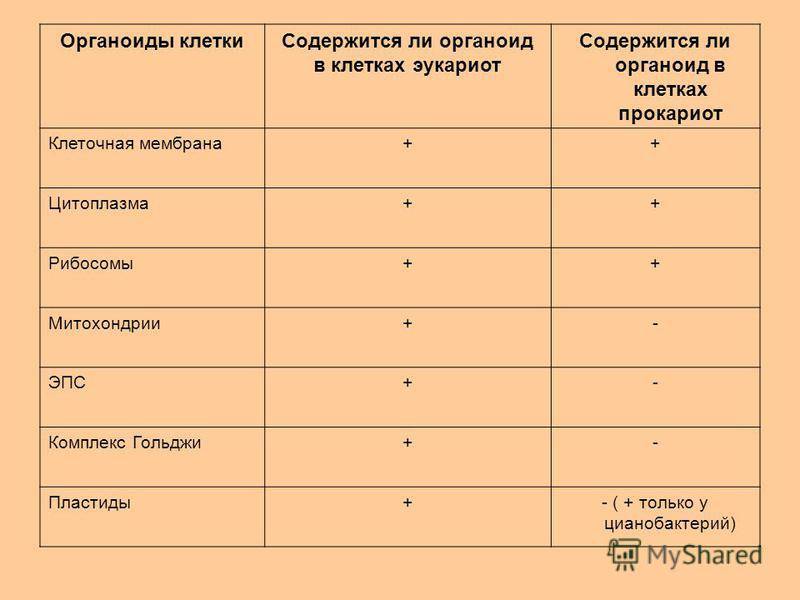 и Годжобори, 2002).
и Годжобори, 2002).
Применение предыдущих выражений к известным композициям мембран клеток показывает, что затраты на биосинтез эукариотических липидов выше, чем у бактерий (дополнительная таблица). Например, для различных видов бактерий средняя прямая стоимость молекулы липида в плазматической мембране составляет 123 (SE = 3) АТФ, тогда как для эукариот она составляет 143 (2).Последняя оценка идентична средней, полученной для целых эукариотических клеток, но стоимость митохондриальных липидов особенно высока 155 (5). Эти повышенные затраты у эукариот являются совместным эффектом стоимости митохондриального экспорта оксалоацетата для образования ацетил-КоА и тенденции эукариотических липидов иметь более длинные цепи, содержащие больше ненасыщенных атомов углерода.
Чтобы понять общие биоэнергетические затраты, связанные с мембранами, нам нужна информация о количестве липидных молекул, необходимых для образования мембраны, что эквивалентно общей площади поверхности мембраны, деленной на количество липидных молекул на единицу площади поверхности и умноженной на на два для учета липидного двойного слоя.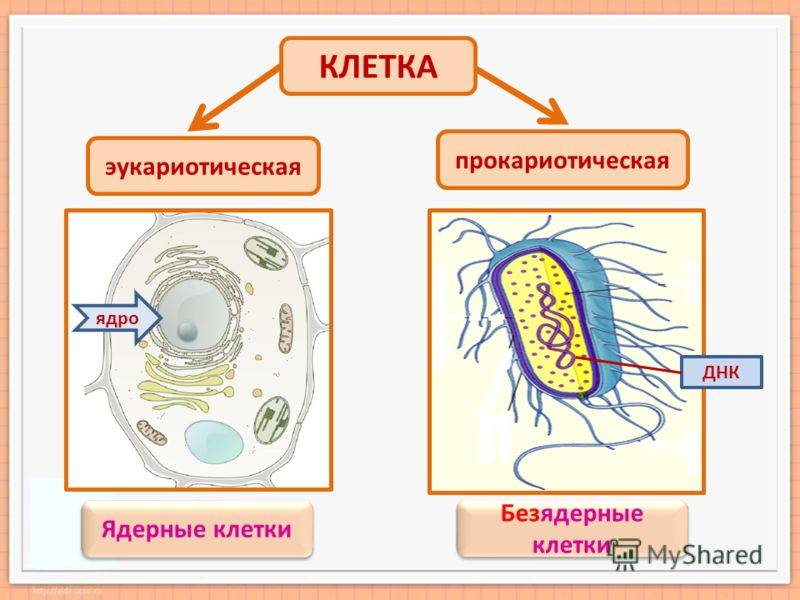 Оценки площадей головной группы липидов мембран в основном находятся в пределах 10% от среднего значения 6,5 × 10 −7 мкм 2 (Petrache et al., 2000; Kučerka et al., 2011), поэтому стоимость мембраны (в единицах АТФ, без учета липидного обмена и пространства, занимаемого трансмембранными белками) составляет
Оценки площадей головной группы липидов мембран в основном находятся в пределах 10% от среднего значения 6,5 × 10 −7 мкм 2 (Petrache et al., 2000; Kučerka et al., 2011), поэтому стоимость мембраны (в единицах АТФ, без учета липидного обмена и пространства, занимаемого трансмембранными белками) составляет
(4)
CL≃(3,1×106)⋅c¯L⋅A,
, где A — площадь поверхности мембраны в единицах мкм 2 , а c¯L — средняя стоимость липида.
Имеется достаточно информации об общих инвестициях в митохондриальные мембраны, чтобы можно было сделать общее заявление.В эукариотическом домене общая площадь поверхности митохондрий (внутренняя плюс внешняя мембрана, суммированная по всем митохондриям, в мкм 2 ) зависит от объема клетки (V, в единицах мкм 3 ) как 3,0 В 0,99 ( Рисунок 1c; SE точки пересечения и наклона на логарифмических графиках составляют 0,22 и 0,08 соответственно). Применяя это к уравнению (4) со средней общей стоимостью митохондриальных липидов (c¯L=440 АТФ/молекула; Приложение 1 – таблица 4) и используя выражение для общих потребностей роста клетки, уравнение (1b) , относительная стоимость липидов митохондриальных мембран составляет
(5)
ск≃0. 15V0.02,
15V0.02,
или ~15% от общего бюджета роста эукариотической клетки минимального размера (1 мкм 3 ) и почти не зависит от размера клетки в пределах диапазона, обычно встречающегося у эукариот (SE экспоненты составляет 0,08). Прямой вклад митохондриальных мембранных липидов в бюджет роста клетки составляет ~36% от этой общей стоимости. Эти затраты на митохондриальные мембраны представляют собой базовую цену, не понесенную прокариотами, связанную с перемещением биоэнергетики внутрь эукариотических клеток, то есть ~ 5%.К сожалению, дополнительные затраты на поддержание митохондриальных липидов неизвестны, но для быстрорастущих клеток подавляющая часть энергетического бюджета клетки направляется на рост (Lynch and Marinov, 2015), поэтому вышеуказанные затраты по-прежнему должны применяться как первоочередные. приближения; для медленно растущих клеток затраты будут выше или ниже в зависимости от того, выше или ниже стоимость обслуживания митохондриальной мембраны по сравнению с общим обслуживанием клетки. Белки не занимают> 50% мембран, поэтому учет этого изменит предыдущие результаты в <2 раза.
Белки не занимают> 50% мембран, поэтому учет этого изменит предыдущие результаты в <2 раза.
Ожидается, что для прокариотических клеток без внутренних мембран относительный вклад клеточной мембраны в общий энергетический баланс клетки будет уменьшаться с увеличением размера клетки из-за снижения отношения площади поверхности к объему. Для крошечных клеток Leptospira interrogans и Mycoplasma pneumoniae (средний объем 0,03 и 0,22 мкм 3 соответственно) ~ 63 и 43% бюджета роста клетки должно быть отнесено к плазматической мембране, но для крупнее Bacillus subtilis и Escherichia coli (в среднем 1.4 и 1,0 мкм 3 соответственно), эти вклады падают до ~ 14 и 19%, и можно ожидать, что они будут продолжать снижаться с дальнейшим увеличением размера клетки, масштабируясь обратно пропорционально линейному размеру клетки.
Напротив, из-за увеличения инвестиций во внутренние мембраны доля энергетического бюджета эукариотической клетки, направляемая на мембраны, не уменьшается с увеличением размера клетки. Хотя есть только несколько типов эукариотических клеток, для которых эта проблема может быть оценена количественно (таблица 1), данные охватывают три порядка по объему клеток и единообразно предполагают, что от 10 до 30% общего бюджета роста приходится на липиды. биосинтеза, и что все большая доля таких затрат связана с внутренними мембранами в клетках с увеличивающимся размером.Пикопланктонная водоросль Ostreococcus , имеющая размер клетки всего 0,22 мкм 3 (меньше, чем у многих прокариот), расходует ~32% своего энергетического бюджета на мембраны, и 44% этих затрат (~18% общий клеточный бюджет) связаны с внутренними мембранами. Клетка млекопитающего среднего размера тратит примерно 30% своего энергетического бюджета на мембраны, но 96% этих затрат (~29% общего клеточного бюджета) связаны с внутренними мембранами.
Хотя есть только несколько типов эукариотических клеток, для которых эта проблема может быть оценена количественно (таблица 1), данные охватывают три порядка по объему клеток и единообразно предполагают, что от 10 до 30% общего бюджета роста приходится на липиды. биосинтеза, и что все большая доля таких затрат связана с внутренними мембранами в клетках с увеличивающимся размером.Пикопланктонная водоросль Ostreococcus , имеющая размер клетки всего 0,22 мкм 3 (меньше, чем у многих прокариот), расходует ~32% своего энергетического бюджета на мембраны, и 44% этих затрат (~18% общий клеточный бюджет) связаны с внутренними мембранами. Клетка млекопитающего среднего размера тратит примерно 30% своего энергетического бюджета на мембраны, но 96% этих затрат (~29% общего клеточного бюджета) связаны с внутренними мембранами.
Вклад мембран в общие затраты на рост клеток.
Ot обозначает зеленую водоросль Ostreococcus tauri , Sc — дрожжи Saccharomyces cerevisiae , Ds — зеленую водоросль Dunaliella salina и Ss — клетку поджелудочной железы свиньи ( Sus scrofa ); ссылки даны в дополнительном материале.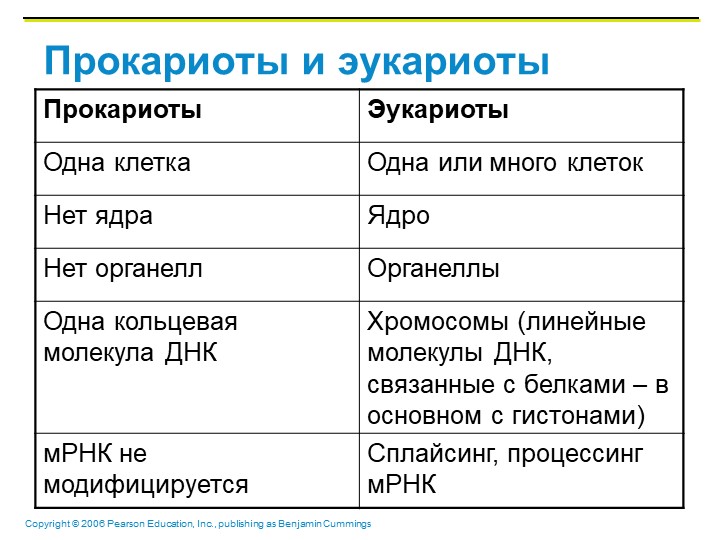 Объемы клеток и общая площадь мембран выражены в мкм 3 и мкм 2 соответственно, причем последние исключают мембраны, связанные с пластидами у видов водорослей.Доля общего бюджета роста клеток, выделяемая на мембраны, получается соотношением уравнений (1b) и (4) с использованием видово-специфических приведенных затрат в таблице 1, где они доступны, и в противном случае с применением средних значений для эукариотических видов; эта общая стоимость затем распределяется на пять различных дробных взносов в следующих строках.
Объемы клеток и общая площадь мембран выражены в мкм 3 и мкм 2 соответственно, причем последние исключают мембраны, связанные с пластидами у видов водорослей.Доля общего бюджета роста клеток, выделяемая на мембраны, получается соотношением уравнений (1b) и (4) с использованием видово-специфических приведенных затрат в таблице 1, где они доступны, и в противном случае с применением средних значений для эукариотических видов; эта общая стоимость затем распределяется на пять различных дробных взносов в следующих строках.
https://doi.org/10.7554/eLife.20437.004
+ От Sc Ds Сс объем ячейки 1 44 591 1060 Всего мембран 15 204 2299 12952 Доля абсолютного бюджета роста клеток 0.324 0.096 0,094 0,302 Плазменная мембрана 0,556 0,328 0,134 0,044 Митохондрии 0,243 0,359 0,197 0,223 Nucleus 0.113 0.085 0,034 0,034 0,034 0.008 Эндоплазматический ретикулум + Golgi 0,034 0,11 0.318 0,706 везикулы / вакуулы 0,055 0.114 0.316 0.019
Взятые вместе, эти наблюдения подразумевают, что использование внутренних мембран представляет собой значительную утечку общего энергетического бюджета эукариотических клеток, гораздо большую, чем можно было бы ожидать у бактерий сопоставимого размера. Более того, поскольку липиды, связанные только с митохондриями, составляют от 20% до 35% инвестиций эукариотической клетки в мембраны (таблица 1), энергетическая нагрузка локализации мембранной биоэнергетики в митохондриях является существенной.
Наконец, учитывая, что наблюдения, обобщенные на рис. 1a,b, получены из различных исследований, вероятно, с множеством уникальных неточностей, стоит рассмотреть, согласуются ли общие выводы с известной способностью АТФ-синтазы. Во-первых, следует отметить, что лишь часть энергии, затрачиваемой на биосинтез, получается непосредственно за счет хемиосмотической активности АТФ-синтазы. Например, биосинтез аминокислот включает примерно 1,5 окисления НАДН и НАДФН на каждый гидролиз АТФ (Akashi and Gojobori, 2002).Если предположить, что каждый из первых эквивалентен ~ 3 гидролизам АТФ, это означает, что только ~ 18% энергии, затрачиваемой на биосинтез аминокислот, связано с гидролизом АТФ. Как отмечено в дополнительном тексте, отношение использования НАДН/НАДФН к АТФ больше порядка 2,0 в биосинтезе липидов, что снижает прямые инвестиции в АТФ до ~14%. клетка связана с синтезом мономерных строительных блоков белков и мембран, только ~ 15% биосинтетической энергии может быть получено из гидролиза АТФ.
Зная известные потребности в энергии для поддержания и роста клетки, время клеточного деления и количество комплексов АТФ-синтазы на клетку, можно оценить требуемую скорость превращения АДФ → АТФ на комплекс. Используя ранее представленные клеточные энергетические данные (Lynch and Marinov, 2015) и распространенность АТФ-синтазы в Приложении 1-таблице 2, после дисконтирования максимальных значений на 85%, предполагаемые необходимые скорости производства АТФ/комплекс/сек составляют: 2109, 221 и 19 соответственно для бактерий B.subtilis , E. coli и M. pneumoniae , а также 1440 и 329 для дрожжей S. cerevisiae и S. pombe . Было предпринято несколько попыток оценить максимальную скорость оборота (в секунду) АТФ-синтазы F0F1, обычно в реконструированных липосомах, и в среднем она составляет 195/с у бактерий (Etzold et al., 1997; Slooten and Vandenbranden, 1989; Toei et al. ., 2007), 295 в пластидах сои (Schmidt, Gräbe, 1985; Junesch, Gräber, 1991), 120 в S.cerevisiae (Förster et al., 2010) и 440 в сердце крупного рогатого скота (Matsuno-Yagi and Hatefi, 1988). Таким образом, учитывая, что значительная часть комплексов, вероятно, будет неправильно собрана в искусственных мембранах, оценки количества оборотов АТФ, генерируемых на клетку, на основе энергетического баланса, по-видимому, согласуются с известной способностью АТФ-синтазы.
Этот странный микроб может ознаменовать один из больших скачков жизни
Причудливый микроб с щупальцами, обнаруженный на дне Тихого океана, может помочь объяснить происхождение сложной жизни на этой планете и решить одну из самых глубоких тайн биологии, сообщили ученые в среду .
Два миллиарда лет назад простые клетки дали начало гораздо более сложным клеткам. Биологи десятилетиями пытались понять, как это произошло.
Ученым давно известно, что на пути эволюции должны были быть предшественники. Но, судя по летописи окаменелостей, сложные клетки возникли просто из ниоткуда.
Новый вид, названный Prometheoarchaeum, оказывается именно такой переходной формой, помогающей объяснить происхождение всех животных, растений, грибов — и, конечно же, человека.Об исследовании сообщается в журнале Nature.
«На самом деле это довольно круто — это окажет большое влияние на науку», — сказала Криста Шлепер, микробиолог из Венского университета, которая не участвовала в новом исследовании.
Наши камеры забиты контейнерами. Например, они хранят ДНК в ядре и вырабатывают топливо в отсеках, называемых митохондриями. Они разрушают старые белки внутри крошечных хозяйственных машин, называемых лизосомами.
Наши клетки также строят себе скелет из нитей, построенных из строительных блоков, подобных Lego.Растягивая одни нити и разрывая другие, клетки могут менять свою форму и даже перемещаться по поверхности.
Виды, которые разделяют эти сложные клетки, известны как эукариоты, и все они происходят от общего предка, жившего примерно два миллиарда лет назад.
До этого мир был домом только для бактерий и группы маленьких простых организмов, называемых археями. Бактерии и археи не имеют ядер, лизосом, митохондрий или скелета.
Биологи-эволюционисты долго ломали голову над тем, как эукариоты могли развиться из таких простых предшественников.
В конце 1900-х годов исследователи обнаружили, что митохондрии когда-то были свободноживущими бактериями. Каким-то образом их затянуло внутрь другой клетки, обеспечив своим хозяином новое топливо.
В 2015 году Тийс Эттема из Упсальского университета в Швеции и его коллеги обнаружили фрагменты ДНК в отложениях, извлеченных из Северного Ледовитого океана. Фрагменты содержали гены вида архей, которые, по-видимому, были тесно связаны с эукариотами.
Доктор Эттема и его коллеги назвали их археями Асгарда.(Асгард — дом скандинавских богов.) ДНК этих таинственных микробов была обнаружена в реке в Северной Каролине, горячих источниках в Новой Зеландии и других местах по всему миру.
Асгардские археи полагаются на ряд генов, которые ранее были обнаружены только у эукариот. Возможно, эти микробы использовали эти гены для тех же целей — или для чего-то еще.
«Пока у вас нет организма, вы не можете быть уверены», — сказал доктор Шлепер.
Масару К. Нобу, микробиологу из Национального института передовых промышленных наук и технологий в Цукубе, Япония, и его коллегам удалось вырастить эти организмы в лаборатории.Усилия заняли более десяти лет.
Микробы, приспособленные к жизни на холодном морском дне, живут в замедленном темпе. Prometheoarchaeum может делиться до 25 дней. Напротив, кишечная палочка делится каждые 20 минут.
Проект начался в 2006 году, когда исследователи подняли отложения со дна Тихого океана. Первоначально они надеялись изолировать микробы, питающиеся метаном, которые можно было бы использовать для очистки сточных вод.
В лаборатории исследователи воспроизвели условия на морском дне, поместив осадок в камеру без доступа кислорода.Они закачивали метан и извлекали смертоносные отработанные газы, которые могли убить живущих там микробов.
Грязь содержала много видов микробов. Но к 2015 году исследователи выделили новый интригующий вид архей. И когда доктор Эттема и его коллеги объявили об открытии ДНК архей Асгарда, японские исследователи были потрясены. Их новый живой микроб принадлежал к этой группе.
Затем исследователи предприняли более кропотливые исследования, чтобы понять новый вид и связать его с эволюцией эукариот.
Исследователи назвали микроб Prometheoarchaeum syntrophicum в честь Прометея, греческого бога, который дал людям огонь, вылепив их из глины.
«Двенадцать лет микробиологии, которые потребовались, чтобы дойти до точки, когда вы можете увидеть ее под микроскопом, просто поразительны», — сказал Джеймс Макинерни, биолог-эволюционист из Ноттингемского университета, который не участвовал в исследовании.
Под микроскопом Prometheoarchaeum оказался странным зверем.Поначалу микроб выглядит как крошечная сфера, но в течение нескольких месяцев он отрастает длинными ветвящимися щупальцами и выпускает флотилию покрытых мембраной пузырей.
Это оказалось еще более странным, когда исследователи исследовали внутреннюю часть камеры. Доктор Шлепер и другие исследователи ожидали, что археи Асгарда использовали свои эукариотоподобные белки для создания некоторых эукариотоподобных структур внутри своих клеток. Но это не то, что обнаружила японская команда.
«Внутри нет никакой структуры, только ДНК и белки», — сказал доктор.Нобу.
Это открытие позволяет предположить, что белки, которые эукариоты использовали для построения сложных клеток, сначала выполняли другие функции, и только позже им поручили новую работу.
Доктор Нобу и его коллеги сейчас пытаются выяснить, что это были за рабочие места. Возможно, сказал он, Prometheoarchaeum создает свои щупальца с генами, которые позже использовались эукариотами для построения клеточных скелетов.
Доктор Шлепер хотел увидеть больше доказательств этой идеи. «У других архей очень красивые руки», — заметила она.Но эти другие виды не используют белки, столь похожие на наши.
До открытия Prometheoarchaeum некоторые исследователи подозревали, что предки эукариот жили как хищники, поедая более мелких микробов. Таким образом они могли поглотить первые митохондрии.
Но Prometheoarchaeum не подходит под это описание. Команда доктора Нобу часто обнаруживала, что микроб прилипает к бокам бактерий или других архей.
Вместо того, чтобы охотиться на добычу, Prometheoarchaeum, кажется, зарабатывает на жизнь, поедая фрагменты белков, проплывающих мимо.Его партнеры питаются его отходами. Они, в свою очередь, обеспечивают Prometheoarchaeum витаминами и другими необходимыми соединениями.
Доктор Нобу предположил, что вид архей Асгарда на морском дне затягивает бактерии паутиной щупалец, втягивая их в еще более тесную связь. В конце концов, он поглотил бактерии, которые превратились в митохондрии, питающие каждую сложную клетку.

Используя реконструкцию наследственного состояния, мы показываем, что LECA, скорее всего, включал многоядерную стадию, которая также допускала посредничество в конфликте между митохондриальным и ядерным геномами, вызванное эндосимбиотическим переносом генов.Почти повсеместное присутствие синцитиальной стадии жизни во всех основных группах эукариот предполагает, что многоядерный LECA является жизнеспособным промежуточным продуктом, который позволил проводить внутриклеточные эксперименты и эволюцию сложных эукариотических процессов, которые мы наблюдаем сегодня. 2016). В летописи окаменелостей они появляются примерно на 2 миллиарда лет позже, чем прокариоты (Javaux et al. 2001; Javaux and Lepot 2018). Среди специалистов существует консенсус в отношении того, что эукариоты произошли от прокариот, но вопрос о том, как они произошли от прокариот, активно обсуждается. Все современные теории происхождения эукариот так или иначе влекут за собой концепцию симбиогенеза (Mereschkowsky 1910; английский перевод Kowallik and Martin 2021), поскольку митохондрии прослеживаются до последнего общего предка эукариот LECA (Embley and Martin 2006; Tria et al.2021), и нет надежного способа объяснить структуру, ДНК и биоэнергетические свойства митохондрий (и хлоропластов) без их эндосимбиотического происхождения. Различия между современными теориями происхождения эукариот (рассмотренные в Martin et al. 2015; López-García and Moreira 2015; Dacks et al. 2016) в основном касаются предположений о биологической природе и клеточной сложности хозяина, который приобрел митохондрию. 2016; Воссеберг и др.2021), хотя методы, лежащие в основе таких заявлений, были поставлены под сомнение (Martin et al., 2017a; Tria et al., 2021; Nagies et al., 2020). Градуалистские и гибридные теории обычно постулируют происхождение фаготрофного питания в линии архей-хозяев до возникновения митохондрий (Doolittle, 1998; Spang et al., 2015; Zaremba-Niedzwiedzka et al., 2017; Vosseberg et al. 2021), что однако это очень проблематичное предположение с физиологической точки зрения (Martin et al. 2017b) и расходится с данными из летописи микрофоссилий, указывающими на позднее происхождение фагоцитоза (Mills, 2020).Эукариоты, несомненно, являются генетическими химерами, причем большинство эукариотических генов происходит от бактерий, а не от архей (Brueckner and Martin 2020), при этом бактериальные гены у эукариот восходят к LECA, а не к наследственно-специфическим приобретениям в ходе эукариотической эволюции (Nagies et al., 2020). , 2004; Имачи и др., 2020 г.; Мартин и Эттема, 2013 г.; Martin et al., 2015), предположение, которое почти никогда не подвергалось сомнению (Garg and Martin 2016). Единообразие взглядов на одноядерную природу LECA настолько распространено, что принимается как данность, то есть редко, если вообще когда-либо, упоминается в качестве предположения. Более того, теории происхождения эукариот, если они вообще проиллюстрированы схематической диаграммой, неизменно представляют LECA как одноядерную клетку. Такие изображения часто носят символический характер, изображая признаки, а не живые клетки, но в то же время они влияют на то, как мы осмысляем проблему происхождения эукариот.Модели эукариогенеза, в которых митохондрии играют механистическую роль, обычно подразумевают отношения «один к одному» или «многие к одному» (Lane and Martin, 2012) между митохондриями и ядром, в силу чего природа ядерной динамики LECA, гетерогенность ядер в LECA , его координация ядерного деления с клеточным делением, его клеточный цикл (мейотический или митотический) и эволюционная последовательность, связывающая деление органелл, ядерное и клеточное деление, редко обсуждаются (Cavalier-Smith 2010; Garg and Martin 2016). Более того, давно предполагалось, что общий предок Metazoa мог быть многоядерным (Hadži 1953). В Rhizaria, самой глубоководной ветви ЮАР, есть многочисленные примеры многоядерных представителей (наиболее примечательным является Xenophyophorea).Кроме того, Opalinata и Apicomplexa также имеют многоядерные формы в рамках своего жизненного цикла (Archibald et al. 2017; Adl et al. 2019). Среди гетеротрофных эукариот обнаружены не только синцитии, но и многочисленные примеры многоядерных водорослей, как красных ( Florideophyceae ), так и зеленых ( Ulvophyceae ), а также различных многоядерных тканей наземных растений (Niklas et al. 2013). Многоядерные формы также встречаются среди эукариот со вторичными пластидами, например, у Chlorarachniophyceae , Phaeophyceae и Xanthophyceae (Niklas et al.2013). Распространение и эволюция многоядерных тканей среди эукариот с пластидами обнаруживает большое разнообразие форм в 60 семействах архепластид и пяти различных линиях водорослей (Niklas et al. 2013). NOAA, общественное достояние; Б. Норберт Хюльсманн, BY-NC-SA 2.0; С. и И. Proyecto Agua, BY-NC-SA 2.0; D. Лаборатория мультиклеточного генома, BY 2.0; Э. Билли Лиар, BY-NC-SA 2.0; Г. Кевин А. Мурах, Галерея изображений NIH, BY-CN 2.0; Х. Ширли Чио, Лаборатория биологии грибов, Калифорнийский университет в Беркли, Калифорния, BY-SA 3.0; Дж.Кристоф Куинтин, BY-NC 2.0; К. Андре Амарал, распространяется под лицензией CC BY-NC 4.0; Лаборатория Л. Джасперсена, общественное достояние. Масштабная линейка является приблизительной. сифонный таллом; (J) – Rhodophyta: Florideophyceae: Lithophyllum sp.; (K) — миксомицеты: многоядерный плазмодий члена Physaraceae ; (L) Ascomycota: Eremothecium gossypii , асептированные гифы. Авторы фотографий и общий домен Creative Commons (CC): A и F. NOAA, общественное достояние; Б. Норберт Хюльсманн, BY-NC-SA 2.0; С. и И. Proyecto Agua, BY-NC-SA 2.0; D. Лаборатория мультиклеточного генома, BY 2.0; Э. Билли Лиар, BY-NC-SA 2.0; Г. Кевин А. Мурах, Галерея изображений NIH, BY-CN 2.0; Х. Ширли Чио, Лаборатория биологии грибов, Калифорнийский университет в Беркли, Калифорния, BY-SA 3.0; Дж. Кристоф Куинтин, BY-NC 2.0; К. Андре Амарал, распространяется под лицензией CC BY-NC 4.0; Лаборатория Л. Джасперсена, общественное достояние. Масштабная линейка является приблизительной. Оба приводят к многоядерному состоянию и не исключают друг друга. Мы используем термин «многоядерный» для описания состояния наличия более двух (обычно четырех или более) ядер в одной и той же клетке, независимо от механизма, вызвавшего это состояние. Стандартные митотические и мейотические промежуточные продукты, очевидно, не учитываются здесь как многоядерные состояния, поскольку это сделало бы признак тривиальным, сделав его столь же универсальным, как и наличие самих ядер. Изображения на рисунке 1 передают впечатление многоядерного состояния в том смысле, в каком это подразумевается в этой статье. Как упоминалось в разделе «Введение», многоядерное состояние обычно достигается одним из двух путей, а именно за счет отсутствия клеточного деления после деления ядра, что приводит к образованию клеток, обычно обозначаемых как ценоциты, и слияния одноядерных клеток, что приводит к образованию клеток, обычно обозначаемых как ценоциты. как синцитий. Хотя онтогенетическая разница между двумя состояниями очевидна, в отсутствие тщательных исследований клеточной биологии и клеточного цикла, которые отсутствуют для многих из рассмотренных здесь таксонов, невозможно точно закодировать две формы по отдельности, и поэтому они рассматриваются здесь. вместе как многоядерные.Обратите внимание, что наше эволюционное исследование касается свойств многоядерных клеток и взаимодействия в них ядер и митохондрий, независимо от процесса, породившего многоядерное состояние. 2019, полный список см. в дополнительных таблицах 2 и 3, дополнительный онлайн-материал) и дополнительно уточнены за счет учета различных конфигураций политомий и различных принятых положений корня (всего 16), что привело к 30 различным топологиям эукариот (дополнительная таблица 4, дополнительный онлайн-материал).Хотя дерево, показанное на рисунке 2, среди доступных в настоящее время альтернатив является наименее противоречивым и, возможно, наиболее надежным деревом для изучаемых здесь линий эукариот, все три различные топологии использовались для реконструкции состояния предков (дополнительная таблица 4, Дополнительный материал онлайн). Показаны основные прокариотические группы (внутри архей и бактерий) и эукариотические супергруппы, всего в этот анализ включено 106 таксонов. Сравнение числа известных видов показано в логарифмической шкале. Квадраты на концах определенных ветвей обозначают, в каких группах секвенированы геномы.Справочное дерево эукариот см. в разделе «Материалы и методы». Схематическое прокариотическое древо жизни было построено на основе литературы. Дерево было нарисовано с помощью iTol. LUCA изображен как возникший в гидротермальном источнике, в то время как LECA, который мог также возникнуть вблизи гидротермальных источников как геологический источник H 2 (15, 23), изображен как многоядерный организм, в котором ядра делятся, а их оболочки остаются неповрежденными. . Показан первичный эндосимбиоз с цианобактериями, давший начало Archaeplastida.Вторичные эндосимбиотические события, множественное происхождение вторичных пластид, показаны стрелками. 6-минутный анимационный видеоролик, иллюстрирующий происхождение эукариот от симбиоза и роль синцитиального состояния в жизненном цикле LECA, можно посмотреть по адресу (https://www. youtube.com/watch?v=mmh_IpdgWvw&t=2s). .Показан первичный эндосимбиоз с цианобактериями, давший начало Archaeplastida. Вторичные эндосимбиотические события, множественное происхождение вторичных пластид, показаны стрелками. 6-минутный анимационный видеоролик, иллюстрирующий происхождение эукариот от симбиоза и роль синцитиального состояния в жизненном цикле LECA, можно посмотреть по адресу (https://www.youtube.com/watch?v=mmh_IpdgWvw&t=2s). 2017) и что вторичные пластиды несколько раз возникали независимо от эукариот, содержащих первичную пластиду (Maciszewski and Karnkowska 2019; Keeling 2004; Gould et al.2008). Реконструкция состояния предков должна соответствующим образом отображать эти черты. 2012). В соответствии с предыдущими отчетами установлено, что пол встречается повсеместно в домене Eukarya, а мейотические гены присутствуют во всех супергруппах в высокой степени консервативно (Ramesh et al. 2005; Speijer et al. 2015; Hofstatter and Lahr 2019). наличие многоядерного состояния, 2. половое размножение, 3. тип деления ядра, 4. полиплоидия, 5. тип митохондрий и 6. тип пластид). (B) Показана реконструкция состояния предков многоядерного состояния, а также (C). Эволюция пластид. Схематично изображены представители основных фотосинтезирующих эукариот. Полученные данные свидетельствуют о том, что LECA был многоядерным половым гетеротрофом с закрытым митозом. 2017) (рис. 3B). Полиплоидия (>2 n ) также возникала несколько раз независимо друг от друга. Хотя полиплоидные эукариоты неоднократно возникали в эволюционно хорошо обособленных группах, LECA, скорее всего, имели гаплоидные ядра (рис. 3А). Полиплоидные трофобионты (стадии питания) среди эукариот встречаются редко.Единственными полиплоидными фазами у большинства эукариот является диплоидная зигота (особенно ее тетраплоидная фаза перед первым делением), которая наследственно подвергается мейотической рекомбинации до образования четырех различных ядер (трофических клеток, спор или гамет). В тех линиях, чьи трофобионты являются диплоидными, каждое соматическое ядро, имеющее ДНК, реплицируемую до сегрегации хромосом, можно рассматривать как временно тетраплоидное, хотя, как и в случае образования зиготы (см. Материалы и методы), они не были оценены как полиплоидные.Соответственно, LECA не был полиплоидным, но поскольку он был мейотическим, он содержал некоторую форму кариогамной стадии. Полные подробные результаты реконструкции состояния предков см. в дополнительной таблице 6, Дополнительный материал онлайн. Свободно плавающие ядра в цитозоле, а не привязанные к клеточным стенкам, означают, что в синцитии они могут делиться только в том случае, если ядерное деление — и, следовательно, сегрегация хромосом — закрыто, при этом ядерная мембрана остается неповрежденной на протяжении всего митоза. Открытый митоз в ценоците потенциально может привести к прикреплению веретенообразного аппарата к хромосомам из разных ядер и их сегрегации аберрантным и, вероятно, летальным образом.Реконструкция (рис. 3Б; табл. 1) свидетельствует о том, что открытое деление ядра (растворение ядерной оболочки при митозе, как хорошо известно у позвоночных) возникло из закрытого митоза путем полуоткрытого деления, при котором растворяются части ядерной оболочки, как промежуточное состояние (Boettcher and Barral, 2013). Открытое деление ядра характерно для некоторых одноядерных (как одноклеточных, так и многоклеточных) и большинства наземных эукариот (рис. 3А). 8 867 Симбиоз, давший начало эукариотам, произошел более 1,5 миллиардов лет назад (Knoll et al. 2006). В то время как происхождение эукариот не может произойти в лаборатории, эндосимбиоз может произойти (Mehta et al. 2018). Контуры эукариогенеза, промежуточные стадии и последовательность вовлеченных событий могут быть рассмотрены посредством вывода из сравнительного исследования современных линий. Первый эукариот был результатом взаимодействия между археями и бактериями, двумя сильно расходящимися клеточными линиями, которые в результате взаимодействия и сотрудничества породили новый вид организма, LECA, с новыми свойствами, новой биоэнергетикой, химерными хромосомами, клеточным циклом, новым генетика, реципрокная рекомбинация и клеточная сложность.Потомки этих симбиотических партнеров сохраняются в виде бактериальных рибосом в митохондриях и архейных рибосом в эукариотическом цитозоле.
LECA имели половое размножение, которое включало слияние гаплоидных ядер, отобранных для размножения (гаметы), и рекомбинацию их генетического материала (мейоз). Митохондрии, пол и несколько ядер являются сигнатурами состояния LECA с синергетическими взаимодействиями. В отличие от митохондрий, ядро имеет большой сложный геном с небольшими ограничениями по размеру.Генетическая совместимость ядер и митохондрий, населяющих одну и ту же цитоплазму, имеет решающее значение для выживания эукариотических клеток. Внутренняя конкуренция или цитоядерная несовместимость могут привести к летальному исходу (Blackstone and Green 1999; Pesole et al. 2012; Rand and Mossman 2020) или привести к дисфункции организма. Наследование митохондрий часто однородительское. Однако наследование ядерного генома бывает би-, три- или мультиродительским. Однородительское наследование митохондрий указывает на существование жесткого контроля на совместимость.Мейотическая рекомбинация, происходящая у предков во время фазы зиготы, является контрольной точкой совместимости. В начале эволюции эукариот была необходима совместимость митохондрий с вновь возникшими ядрами. В одноядерных клетках естественный отбор пережили только совместимые комбинации. В синцитиях взаимодействия «многие ко многим» между митохондриями и ядрами обеспечивают буферную совместимость в пределах окружающей среды одной цитоплазмы. Споры, порожденные синцитиальным LECA, представляли сильное узкое место отбора на цитоядерную совместимость (Garg and Martin 2016).
Интригующий аспект многоядерного состояния LECA касается перехода от прокариотической к эукариотической сегрегации хромосом. У прокариот сегрегация хромосом связана с делением клеток посредством прикрепления хромосом к клеточной стенке. У эукариот зависящая от микротрубочек сегрегация конденсированных хромосом и клеточное деление (цитокинез) не связаны ни физически, ни механически, хотя часто противопоставлены во времени. То есть хромосомы могут и часто реплицируются и сегрегируют в неделящихся клетках без образования веретен деления самого ядра (Geitler 1953), процессы, которые в более ранней литературе назывались Amitose (Strasburger 1908).Если начало деления ядра (репликация с последующей сегрегацией) предшествовало возникновению клеточного деления у эукариот (обратное вряд ли могло бы быть верным), то образовавшийся синцитий не обязательно обладал хорошо регулируемой сегрегацией хромосом в начале. Он мог генерировать ядра с аберрантным числом хромосом или анеуплоидными гаплоидами. Такие дефектные ядра были бы летальными для одноядерной клетки, но не для синцития, потому что даже сильно дефектные ядра могут свободно дополнять друг друга через мРНК в цитозоле.Таким образом, многоядерное состояние должно амортизировать практически все вредные эффекты ядер, возникающие как продукты неправильного разделения хромосом во время закрытого протомитоза в начале сегрегации хромосом эукариот. Это могло бы сохранить жизнь синцитию как единице вегетативной пролиферации, в то же время укрывая ядра с очень разными наборами хромосом, ядра, которые поддерживали бы жизнеспособность друг друга в синцитии за счет комплементации через мРНК в цитозоле. Это участие рибосом, чей синтез требует массивной экспрессии генов рРНК, для комплементации может объяснить, почему соотношение объемов ядра и цитоплазмы ( Kern-Plasmarelation ) имеет тенденцию приближаться к примерно постоянному значению (Klieneberger 1917) 1:10 даже в синцитиальных клетках. (Ситте и др.1991). Как выразился Страсбургер (1908): «У Characeae амитотическое деление ядер в интернодиальных клетках не является дегенеративным процессом, а скорее является средством амплификации определенных компонентов ядерного вещества в связи с увеличением цитоплазматической массы» (стр. 40). , перевод авторов).
Физическое слияние ядер, примитивный и нерегулируемый предшественник кариогамии (присутствующий в LECA, потому что LECA занимается сексом), будет генерировать новые комбинации хромосом одновременно с переносом генов из митохондрий в ядра (Lane and Martin 2012; Гарг и Мартин, 2016 г.).Это породило гетерогенную популяцию ядер, взаимодействующих с гетерогенной популяцией митохондрий внутри одного и того же синцития. Синцитий также может быть физически разорван, генерируя сегменты или фрагменты, которые при наличии средств запечатывания концов могут давать потомство (в виде диаспор) без потребности в регулируемом клеточном делении. Синцитиальные фрагменты обеспечивают механизм распространения популяций ядер и митохондрий. Но главным эволюционным препятствием, которое необходимо было преодолеть, была эволюция регулируемой симметричной сегрегации хромосом, которая учитывала состояние питания клетки (Brunk and Martin 2019) на пути к клеточному циклу — основе биологии эукариотических клеток.
Внутри синцития и ядра, и митохондрии были единицами отбора и единицами эволюции. Они были промежуточным состоянием при переходе от прокариот к эукариотам. Они сосуществовали в одном и том же цитозоле. Ядра стали наследственным набором генов, способных влиять на непосредственно окружающий их цитозоль и способных взаимодействовать друг с другом и с митохондриями посредством экспортируемой мРНК. Многоядерные клетки широко распространены среди эукариот, как живых (рис. 1 и 3А), так и ископаемых, таких как недавно обнаруженная ценоцитарная зеленая водоросль возрастом 1 миллиард лет (Tang et al.2020).
Чтобы сравнить относительную приспособленность одноядерной клетки (монокариона) и многоядерной клетки (поликариона), можно рассмотреть разницу между вероятностью выживания популяции одноклеточных одноядерных эукариот и одиночного синцития. Для монокарионов вероятность выживания популяции зависит от индивидуальных вероятностей выживания, которые, в свою очередь, зависят от приспособленности соответствующей митонуклеарной пары.Однако в случае синцития, поскольку митохондрии и ядра сосуществуют в одной клетке, вероятность выживания зависит от кумулятивной приспособленности всех возможных комбинаций митоядерных пар. Это, в свою очередь, позволяет синцитию вести себя аналогично популяции, позволяя отбору независимо разрешать внутренние мито-ядерные конфликты. Это схематически показано на рисунке 4 и математически описано в дополнительной информации 11, Дополнительный материал онлайн. Синцитий ведет себя скорее как популяция ядер и митохондрий, чем как отдельная клетка.Таким образом, у синцития больше шансов на выживание, чем у популяции монокарионов. Конечно, существуют древние линии эукариот, несущие одноядерные формы, в том числе и раскопки. Однако многоядерная LECA объясняет, почему разнообразие современных эукариот легче происходит от синцитиального предка, чем от популяции одноядерных одноклеточных предков (монокарионов). Популяция монокарионов, особенно гаплоидных монокарионов, вряд ли будет накапливать генетическое разнообразие.С другой стороны, синцитий легко накапливает генетическое разнообразие в пределах одного цитозоля, поскольку ядра с полезными аллелями дополняют недостатки других ядер, а кариогамия, на которую способна мейотическая LECA, внутри синцития может генерировать новые комбинации хромосом (рис. 4). .
Рис. 4.
Синцитии буферируют хромосомные дефекты, в отличие от монокарионов. Схематическое изображение популяции одноклеточных протистов и синцитиальной клетки. Геномы каждого ядра схематично показаны серыми линиями, дефицитные аллели — красными прямоугольниками, тогда как полезный аллель показан зеленым цветом. Если применить те же эволюционные ограничения, популяция монокарионов с большей вероятностью вымрет, чем синцитий, поскольку ядра из разных клеток не могут ни покрывать дефекты друг друга, ни буферировать митонуклеарную несовместимость, в то время как в синцитии они могут.
Рис. 4.
Синцитии буферируют хромосомные дефекты, в отличие от монокарионов. Схематическое изображение популяции одноклеточных протистов и синцитиальной клетки. Геномы каждого ядра схематично показаны серыми линиями, дефицитные аллели — красными прямоугольниками, тогда как полезный аллель показан зеленым цветом.Если применить те же эволюционные ограничения, популяция монокарионов с большей вероятностью вымрет, чем синцитий, поскольку ядра из разных клеток не могут ни покрывать дефекты друг друга, ни буферировать митонуклеарную несовместимость, в то время как в синцитии они могут.
Эволюционные переходы в индивидуальности включают сотрудничество и конфликт (Buss, 1987; Maynard Smith and Szathmáry, 1996; Michod, 1999). Без механизмов посредничества в конфликтах сотрудничество не может выжить (Nowak 2006) и не может возникнуть единица более высокого уровня (Radzvilavicius and Blackstone 2018).В ходе эволюции популяционная структура всегда признавалась одним из наиболее общих механизмов, благоприятствующих кооперации. Даже если отбор благоприятствует несотрудничающим перебежчикам, как это обычно и бывает, сотрудничество все же может развиваться в структурированной популяции. Рассмотрим популяцию, состоящую из особей (нижний уровень), разделенных на группы (верхний уровень). В то время как перебежчики пользуются преимуществом на более низком уровне, кооператоры пользуются преимуществом на более высоком уровне. Если популяция представляла собой одну большую группу, то отбор на более высоком уровне слаб, и преобладают перебежчики.Однако в популяции со многими небольшими группами отбор потенцируется на более высоких уровнях. Группы кооператоров могут образовываться случайно и вытеснять группы перебежчиков (Szathmáry and Demeter 1987). Таким образом, более крупные группы (например, синцитиальная LECA) вызывают больше конфликтов, в то время как меньшие группы (особенно половые гаметы) влекут за собой меньше. Следовательно, при больших размерах клеток стохастические процессы могли быть менее важными в опосредовании эволюционного конфликта.
В синцитии сегрегация хромосом, вероятно, включает молекулярную самоорганизацию механизма разделения хромосом, который не требует точек закрепления на плазматической мембране.
Закрытый митоз, при котором ядерная оболочка остается в основном интактной, считается предком открытого митоза (Cavalier-Smith 2010), что согласуется с нашими собственными результатами (рис. 3). Все варианты митоза имеют общую сеть на основе микротрубочек, которая может быть собрана в пучки или рассыпаться звездообразным образом, тянуться к хромосомам и прикрепляться к кинетохоре, которая присутствовала в LECA (Tromer et al.2019). Однако центросомы не являются существенными для разделения хромосом (Heald et al. 1996). Важно отметить, что эукариотические хромосомы разделяются в основном под действием силы, толкающей вдоль микротрубочек, в которых эукариот-специфичное семейство белков кинезинов играет существенную роль (Shimamoto et al. 2015). Эти механизмы органично согласуются с биологией синцитиальной клетки, поскольку митоз отдельных ядер может происходить независимо от локализованных точек фиксации плазматической мембраны.
Следовательно, происхождение одноядерных жгутиковых простейших можно рассматривать с новой точки зрения.Изображения ближайшего живого родственника архейной клетки-хозяина и бактериального партнера изображают двух сидячих, неподвижных партнеров (Imachi et al. 2020), предлагаемая нами синцитиальная LECA также была сидячей. Центр организации микротрубочек (MTOC), или базальное тело, и способность образовывать жгутики присутствовали у LECA. Эта черта диверсифицировалась среди эукариотических супергрупп и периодически терялась (Yubuki and Leander 2013). Эукариотические жгутики непосредственно связаны с базальными тельцами или формируются de novo зависимым от центриолей образом (Schrøder et al.2011). Комплекс пор жгутиков имеет ряд общих белков с комплексом ядерных пор (Dishinger et al. 2010; Kee et al. 2012; Gould et al. 2016). Мы предполагаем, что жгутик развился на основе (дублированной) структуры, происходящей из центросомы, которая охватывает область плазматической мембраны. Таким образом, одноядерные, жгутиковые споры могли появиться из синцития с актиновым цитоскелетом, поддерживающим окончательное расщепление (Heidstra 2007).
Только споры, содержащие жизнеспособные митонуклеарные взаимодействия и способные к жгутиковому движению, обладали бы свойствами подвижных гамет при условии, что они были способны сливаться с другими в своем роде, что возможно, учитывая склонность самих архей к слиянию (Lange et al. .2011 г.; Гарг и Мартин, 2016 г.; Шалев и др. 2017). Такие споры представляют собой подвижные единицы отбора. Ядро многих жгутиковых простейших расположено в непосредственной близости от базального тела, если не связано с ним, как у многочисленных Archamoebea, Chytridiomycota, Olpidium , Pelagophyceae, Bacillariophyceae, Rhizaria и других (обзор в ссылке 2). Возможно, такие гаметоподобные клетки стали основателями эукариотических супергрупп, каждая из которых содержит жгутиковых представителей, способных генерировать синцитии (рис.3А). У нас нет никаких предположений о физических размерах LECA как синцития, хотя мы предполагаем, что он обитал в морских отложениях (Мартин и Мюллер, 1998), где анаэробная синтрофия необходима для симбиотических взаимодействий (Имачи и др. , 2020). Гифы современных грибов могут занимать площади в квадратные мили (Anderson et al. 2018). LECA мог быть крупной неделящейся многоядерной одноклеточной клеткой, которая порождала супергруппы посредством вытеснения митохондриальных жгутиковых спор. 6-минутный анимационный видеоролик, иллюстрирующий происхождение эукариот от симбиоза и роль синцитиального состояния в жизненном цикле LECA, можно посмотреть по адресу (https://www.youtube.com/watch?v=mmh_IpdgWvw&t=2s)
Однако наши результаты предполагают, что при происхождении эукариот ядра и митохондрии были единицами отбора и единицами эволюции в пределах синцитиального LECA. Реконструкция состояния предкового признака на основе богатой таксонами выборки, охватывающей все супергруппы, предполагает, что LECA был 1) митохондриальным, 2) многоядерным (синцитиальным, ценоцитным), 3) гаплоидным, 4) с закрытым делением ядра и 5) с половым размножением. Часто утверждается, в том числе во многих работах авторов, что переход от прокариот к эукариотам не оставил промежуточных форм.Однако, если наши текущие мысли примерно соответствуют цели, синцитии на самом деле являются промежуточным состоянием в переходе от прокариот к эукариотам, хотя до сих пор не признавались таковыми. В этом свете синцитии, присутствующие во всех надгруппах эукариот, могут содержать ранее непризнанные формы доказательств эукариотического происхождения и перехода от прокариот к эукариотам. «> Отбор таксонов
На основе изучения литературы (дополнительная таблица 4, дополнительный материал онлайн) был создан богатый таксонами (Katz and Grant 2014) набор эукариотических данных, включающий 106 высших таксонов (дополнительная таблица 1, дополнительный материал онлайн). Включены представители шести супергрупп эукариот. Мы используем недавно определенную номенклатуру надгрупп эукариот: Amoebozoa, Archaeplastida, Excavata, Hacrobia, Opisthokonta и SAR , хотя для ясности мы сохранили здесь более знакомый термин Opisthokonta вместо Obazoa. Набор состоит в основном из высших категорий, но в некоторых случаях были включены семейства и роды (табл. 2).
Обзор таксонов эукариот, рассматриваемых в этом анализе
| . | Фила . | Классы . | Заказы . | Семьи . | Роды . | ПерСупергруппа . |
|---|---|---|---|---|---|---|
| амёбозои | — | 7 | 1 | — | — | 8 |
| архепластиды | 7 | 14 | 4 | — | 1 | 26 |
| Excavata | 1 | 4 | 3 | 1 | 4 | 13 |
| Hacrobia | — | 3 | 5 | — | — | 8 |
| заднежгутиковые | 13 | 9 | 1 | — | 1 | 24 |
| SAR | 6 | 17 | 3 | 1 | — | 27 |
| за ранг | 27 | 54 | 17 | 2 | 6 |
|
. | Фила . | Классы . | Заказы . | Семьи . | Роды . | ПерСупергруппа . |
|---|---|---|---|---|---|---|
| амёбозои | — | 7 | 1 | — | — | 8 |
| архепластиды | 7 | 14 | 4 | — | 1 | 26 |
| Excavata | 1 | 4 | 3 | 1 | 4 | 13 |
| Hacrobia | — | 3 | 5 | — | — | 8 |
| заднежгутиковые | 13 | 9 | 1 | — | 1 | 24 |
| SAR | 6 | 17 | 3 | 1 | — | 27 |
| за ранг | 27 | 54 | 17 | 2 | 6 |
Обзор таксонов эукариот, рассматриваемых в этом анализе
|
. | Фила . | Классы . | Заказы . | Семьи . | Роды . | ПерСупергруппа . |
|---|---|---|---|---|---|---|
| амёбозои | — | 7 | 1 | — | — | 8 |
| архепластиды | 7 | 14 | 4 | — | 1 | 26 |
| Excavata | 1 | 4 | 3 | 1 | 4 | 13 |
| Hacrobia | — | 3 | 5 | — | — | 8 |
| заднежгутиковые | 13 | 9 | 1 | — | 1 | 24 |
| SAR | 6 | 17 | 3 | 1 | — | 27 |
| за ранг | 27 | 54 | 17 | 2 | 6 |
|
. | Фила . | Классы . | Заказы . | Семьи . | Роды . | ПерСупергруппа . |
|---|---|---|---|---|---|---|
| амёбозои | — | 7 | 1 | — | — | 8 |
| архепластиды | 7 | 14 | 4 | — | 1 | 26 |
| Excavata | 1 | 4 | 3 | 1 | 4 | 13 |
| Hacrobia | — | 3 | 5 | — | — | 8 |
| заднежгутиковые | 13 | 9 | 1 | — | 1 | 24 |
| SAR | 6 | 17 | 3 | 1 | — | 27 |
| за ранг | 27 | 54 | 17 | 2 | 6 |
Кладограммы, представленные в этом исследовании, основаны на опубликованных отношениях внутри каждой супергруппы эукариот (дополнительные таблицы 3 и 4, дополнительный материал онлайн). Если мы обозначаем отношения как «разрешенные», это означает, что мы включили соответствующий шаблон ветвления для нашего дерева из 106 таксонов. Паттерны ветвления, которые были «неразрешенными», были преобразованы в политомии. Поскольку нет единого мнения о том, где находится корень эукариот Eukarya, был подготовлен набор эталонных деревьев с коллекцией опубликованных предложений по корню эукариот: Excavata или внутри раскопок (Cavalier-Smith 2002; He et al.2014; Триа и др. 2021), Opisthokonta , Fungi или внутри Fungi (например, Microsporidia ) (Vossbrinck et al. 1987), Amoebozoa или внутри (Stechmann and Cavalier-Smith 34 102-Smith 2 Amorphaea ( Opisthokonta + Amoebozoa ) (Derelle et al. 2015). Также был подготовлен нерутированный набор. Подробные данные, лежащие в основе всех параметров и деревьев, представлены в дополнительной таблице 3 «Дополнительный материал онлайн». Все признаки были численно закодированы для реконструкции наследственного состояния (дополнительная таблица 5, дополнительный материал онлайн).Для многоядерного состояния признак кодировался как 1, когда имелось указание на многоядерное состояние, присутствующее во всей группе или части жизненного цикла многих (> 2 родов) членов, или если оно присутствовало в неразрешенных группах. Признак считался неоднозначным (0/1) для группы, когда либо существовал консенсус в отношении того, что многоядерное состояние присутствует у одного производного вида в группе, либо когда были доказательства наличия стадий жизненного цикла, которые очень напоминают синцитии. -подобные структуры без четкого описания у членов группы.Наконец, признак был закодирован как 0, когда вообще не было указаний на существование многоядерного состояния в пределах известного разнообразия определенного таксона. Источники информации обобщены в дополнительных таблицах 2 и 4, Дополнительные материалы онлайн. 6 (https://www.mesquiteproject.org/). Всего было отобрано 106 таксонов эукариот (от рода до уровня типа) (см. Выбор таксонов, дополнительная таблица 1, Дополнительный материал онлайн). Затем был построен набор из 30 эталонных деревьев с разным положением корня и разной степенью свободы в отношении полифилии (см. Установка топологии и корня дерева, дополнительная таблица 3, Дополнительный онлайн-материал). Матрица признаков была подготовлена на основе литературных данных по шести признакам: наличие многоядерного состояния, наличие полиплоидов, наличие пола/мейоза, поведение ядерной оболочки при делении, тип митохондрий и тип пластиды (дополнительные таблицы 2 и 5, Дополнительный материал онлайн).В некоторых группах одни члены группы проявляют одну черту, а другие — другую. В подобных случаях для этой группы кодировались оба признака (0/1 или 1/2). F.M. и С.Г.Г. WFM также хотел бы поблагодарить ERC (666053) и VW Foundation (
Поскольку нет единого мнения о том, где находится корень эукариот Eukarya, был подготовлен набор эталонных деревьев с коллекцией опубликованных предложений по корню эукариот: Excavata или внутри раскопок (Cavalier-Smith 2002; He et al.2014; Триа и др. 2021), Opisthokonta , Fungi или внутри Fungi (например, Microsporidia ) (Vossbrinck et al. 1987), Amoebozoa или внутри (Stechmann and Cavalier-Smith 34 102-Smith 2 Amorphaea ( Opisthokonta + Amoebozoa ) (Derelle et al. 2015). Также был подготовлен нерутированный набор. Подробные данные, лежащие в основе всех параметров и деревьев, представлены в дополнительной таблице 3 «Дополнительный материал онлайн». Все признаки были численно закодированы для реконструкции наследственного состояния (дополнительная таблица 5, дополнительный материал онлайн).Для многоядерного состояния признак кодировался как 1, когда имелось указание на многоядерное состояние, присутствующее во всей группе или части жизненного цикла многих (> 2 родов) членов, или если оно присутствовало в неразрешенных группах. Признак считался неоднозначным (0/1) для группы, когда либо существовал консенсус в отношении того, что многоядерное состояние присутствует у одного производного вида в группе, либо когда были доказательства наличия стадий жизненного цикла, которые очень напоминают синцитии. -подобные структуры без четкого описания у членов группы.Наконец, признак был закодирован как 0, когда вообще не было указаний на существование многоядерного состояния в пределах известного разнообразия определенного таксона. Источники информации обобщены в дополнительных таблицах 2 и 4, Дополнительные материалы онлайн. 6 (https://www.mesquiteproject.org/). Всего было отобрано 106 таксонов эукариот (от рода до уровня типа) (см. Выбор таксонов, дополнительная таблица 1, Дополнительный материал онлайн). Затем был построен набор из 30 эталонных деревьев с разным положением корня и разной степенью свободы в отношении полифилии (см. Установка топологии и корня дерева, дополнительная таблица 3, Дополнительный онлайн-материал). Матрица признаков была подготовлена на основе литературных данных по шести признакам: наличие многоядерного состояния, наличие полиплоидов, наличие пола/мейоза, поведение ядерной оболочки при делении, тип митохондрий и тип пластиды (дополнительные таблицы 2 и 5, Дополнительный материал онлайн).В некоторых группах одни члены группы проявляют одну черту, а другие — другую. В подобных случаях для этой группы кодировались оба признака (0/1 или 1/2). F.M. и С.Г.Г. WFM также хотел бы поблагодарить ERC (666053) и VW Foundation (
6 (https://www.mesquiteproject.org/). Всего было отобрано 106 таксонов эукариот (от рода до уровня типа) (см. Выбор таксонов, дополнительная таблица 1, Дополнительный материал онлайн). Затем был построен набор из 30 эталонных деревьев с разным положением корня и разной степенью свободы в отношении полифилии (см. Установка топологии и корня дерева, дополнительная таблица 3, Дополнительный онлайн-материал). Матрица признаков была подготовлена на основе литературных данных по шести признакам: наличие многоядерного состояния, наличие полиплоидов, наличие пола/мейоза, поведение ядерной оболочки при делении, тип митохондрий и тип пластиды (дополнительные таблицы 2 и 5, Дополнительный материал онлайн).В некоторых группах одни члены группы проявляют одну черту, а другие — другую. В подобных случаях для этой группы кодировались оба признака (0/1 или 1/2). F.M. и С.Г.Г. WFM также хотел бы поблагодарить ERC (666053) и VW Foundation (
и 96742) за финансирование. S.B.G благодарит VolkswagenStiftung (Life) за финансирование.Мы благодарим профессора Клауса В. Коваллика за полезные обсуждения.
«> Доступность данных
Все данные, связанные с рукописью, представлены в дополнительной информации.
Proc Biol Sci
.285
(1893
):20182233
.
Арчибальд
ДМ
,Симпсон
АГБ
,Сламовиц
Ч
, редакторы.2017
. Справочник протистов. Спрингер Природа.
Блэкстоун
СЗ
,Зеленый
ДР.
1999
.Эволюция механизма клеточного самоубийства
.Биоэссе
.21
(1
):84
–88
.
Бетчер
Б
,Барраль
Ю.
2013
.Клеточная биология открытого и закрытого митоза
.Ядро
4
(3
):160
–165
.
Брюкнер
Дж
,Мартин
ВФ.
2020
.Генов бактерий больше, чем генов архей в эукариотических геномах
.Геном Биол Эвол
.12
(4
):282
–292
.
Брунк
CF
,Мартин
ВФ.
2019
. Вклад гистонов архей в происхождение эукариот
.Trends Microbiol
.27
(8
):703
–714
.
Шина
LW.
1987
. Эволюция индивидуальности. Нью-Джерси: Издательство Принстонского университета.
Кавалер-Смит
Т.
1987
.Происхождение клеток эукариот и архебактерий
.Ann NY Acad Sci
.503
:17
–54
.
Кавалер-Смит
Т.
2002
.Фаготрофное происхождение эукариот и филогенетическая классификация простейших
.Int J Syst Evol Micro
.52
(2
):297
–354
.
Кавалер-Смит
Т.
2010
.Происхождение клеточного ядра, митоза и пола: роль внутриклеточной коэволюции
.Биол Директ
.5
:7
.
Кавалер-Смит
Т.
2018
.Царство Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание перипластидных белков, эволюцию цитоскелета и перипластид и древние расхождения
. Протоплазма
255
(1
):297
–357
.
Сумки
JB
, и другие.2016
.Меняющийся взгляд на эукариогенез – окаменелости, клетки, родословные и то, как они все объединяются
.J Cell Sci
.129
(20
):3695
–3703
.
Добенмайр
РФ.
1936
.Использование терминов ценоцит и синцитий в биологии
.Наука
.84
(2189
):533
–533
.
де Дюв
К.
1969
.Эволюция пероксисомы
.Ann NY Acad Sci
.168
(2
):369
–381
.
Дерелле
Р
, и другие.2015
.Бактериальные белки указывают на единственный эукариотический корень
.Proc Natl Acad Sci U S A
.112
(7
):E693
–E699
.
Диррен
С
,Почта
Т.
2016
.Беспорядочное и специфическое приобретение симбионтов бактериями из рода амебоидных Nuclearia (Opisthokonta)
.FEMS Microbiol Ecol
.92
(8
):fiw105
.
Посудомоечная машина
J
, и другие.2010
.Вход в реснички мотора кинезина-2 KIF17 регулируется импортином-β2 и RanGTP
.Nat Cell Biol
.12
(7
):703
–710
.
Дулиттл
ВФ.
1998
.Вы то, что вы едите: механизм переноса генов может объяснить бактериальные гены в ядерных геномах эукариот
.Тренды Genet
.14
(8
):307
–311
.
Эмбли
ТМ
,Мартин
ВФ.
2006
.Эволюция эукариот, изменения и вызовы
.Природа
440
(7084
):623
–630
.
Фу
С
,Коэльо
МА
,Давид-Пальма
М
,Священник
СЖ
,Хейтман
Дж.
2019
.Генетическая и геномная эволюция полового размножения: эхо от LECA до царства грибов
.Curr Opin Genet Dev
.58-59
:70
–75
.
Гарг
СГ
,Мартин
ВФ.
2016
.Митохондрии, клеточный цикл и происхождение пола через синцитиального общего предка эукариот
.Геном Биол Эвол
.8
(6
):1950
–1970
.
Гейзен
С
, и другие.2017
.Перезагрузка почвенной протистологии: 30 фундаментальных вопросов для начала
.Почвенный Биол Биохим
.111
:94
–103
.
Гейтлер
Л.
1953
. Эндомитоза и амитополиплоидизация. Протоплазматология. Справочник Der Protoplasmaforschung Bd. VI: Kern- Und Zellteilung. Том. 6:С. Спрингер, Вена. п.89
.
Гулд
ГВт
,Дринг
ГДж.
1979
. О возможной связи образования эндоспор бактерий с происхождением эукариотических клеток
.Дж Теор Биол
.81
(1
):47
–53
.
Гулд
СБ
,Уоллер
РФ
,Макфадден
GI.
2008
.Эволюция пластид
.Annu Rev Plant Biol
.59
:491
–517
.
Гулд
СБ
,Гарг
СГ
,Мартин
ВФ.
2016
.Секреция бактериальных пузырьков и эволюционное происхождение эукариотической эндомембранной системы
.Trends Microbiol
.24
(7
):525
–534
.
Гулд
СБ
, и другие.2019
.Адаптация к жизни на суше и высокому содержанию кислорода путем перехода от ферредоксина к НАДН-зависимому окислительно-восстановительному балансу
.Proc R Soc B
.286
(2019149
).
Гупта
RS
,Голдинг
ГБ.
1996
.Происхождение эукариотической клетки
.Trends Biochem Sci
.21
(5
):166
–171
.
Хаджи
Дж.
1953
.Попытка реконструировать систему классификации животных
.Сист Зоол
.2
:145
–154
.
Он
Д
, и другие.2014
.Альтернативный корень эукариотического древа жизни
.Карр Биол
.24
(4
):465
–470
.,
Хильда
Р
, и другие.1996
.Самоорганизация микротрубочек в биполярные веретена вокруг искусственных хромосом в экстрактах яиц Xenopus
.Природа
382
(6590
):420
–425
.
Хейдстра
Р.
2007
. Асимметричное деление клеток в развитии растений. В:Macieira-Coelho
A
, редактор.Асимметричное деление клеток
. Verlag Berlin Heidelberg: Springer
. п.1
–37
.
Хофштеттер
PG
,Лар
DJ.
2019
.Все эукариоты являются половыми, если не доказано обратное: многие так называемые асексуалы имеют мейотический аппарат и могут заниматься сексом
.Bioessays
41(6):e1800246.
Хориике
Т
,Хамада
К
,Мията
Д
,Шинозава
Т.
2004
.Происхождение эукариот предполагается как симбиоз Pyrococcus с γ-протеобактериями по филогенетическому древу на основе содержания генов
.Дж Мол Эвол
.59
(5
):606
–619
.
Имачи
Н
, и другие.2020
.Изоляция археона на границе прокариотов и эукариот
.Природа
577
(7791
):519
–525
.
Жаво
EJ
,Холм
АХ
,Вальтер
МР.
2001
.Морфологическая и экологическая сложность ранних эукариотических экосистем
.Природа
412
(6842
):66
–69
.
Жаво
EJ
,Лепо
К.
2018
.Палеопротерозойская летопись окаменелостей: значение для эволюции биосферы в средние века Земли
.Earth-Sci Rev
.176
:68
–86
.
Камикава
Р
, и другие.2014
.Эволюция содержания генов в дискобидных митохондриях, выведенная из филогенетического положения и полного митохондриального генома Tsukubamonas globosa
.Геном Биол Эвол
.6
(2
):306
–315
.
Кац
ЛА
,Грант
Младший.
2015
.Филогеномный анализ, богатый таксонами, разрешает эукариотическое древо жизни и раскрывает силу подвыборки по участкам
.Сист Биол
. 64
(3
):406
–415
.
Ки
HL
, и другие.2012
.Барьер проницаемости исключения размера и нуклеопорины характеризуют комплекс цилиарных пор, который регулирует транспорт в реснички
.Nat Cell Biol
.14
(4
):431
–437
.,
Килинг
ПЖ.
2004
.Разнообразие и эволюционная история пластид и их хозяев
.Am J Bot
.91
(10
):1481
–1493
.
Поцелуй
E
, и другие.2019
.Сравнительная геномика раскрывает происхождение грибных гиф и многоклеточность
.Нац Коммуна
.10
(1
):13
.
Клинебергер
Е.
1917
.Über die Größe und Beschaffenheit der Zellkerne mit besonderer Berücksichtigung der Systematik (Inaug.-Diss. Frankfurt)
.Beihefte Бот. Збл
.35 I
:219
–278
.
Холм
АХ
,Жаво
ЭДЖ
,Хьюитт
Д
,Коэн
стр.
2006
.Эукариотические организмы в протерозойских океанах
.Philos Trans R Soc Lond B Biol Sci
.361
(1470
):1023
–1038
.
Коваллик
КВ
,Мартин
ВФ.
2021
.Рассвет симбиогенеза: аннотированный английский перевод статьи Мерешковского 1910 года
.БиоСистемс
199
:104281
.
Озеро
JA
,Ривера
MC.
1994
.Было ли ядро первым эндосимбионтом?
Proc Natl Acad Sci U S A
.91
(8
):2880
–2881
.
Пер.
N
,Мартин
ВФ.
2012
.Происхождение мембранной биоэнергетики
.Сотовый
151
(7
):1406
–1416
.
Линдос
AC
, и другие. 2008
.Уникальный механизм деления клеток у архей
.Proc Natl Acad Sci U S A
.105
(48
):18942
–18946
.
Ланге
С
,Зерулла
К
,Брейерт
С
,Соппа
Дж.
2011
.Конверсия генов приводит к выравниванию копий генома у полиплоидных галоархей Haloferax volcanii
.Мол Микробиол
.80
(3
):666
–677
.
Лейс
СП.
2003
.Значение синцитиальных тканей для положения Hexactinellida в Metazoa
.Интегр Комп Биол
.43
(1
):19
–27
.
Лопес-Гарсия
P
,Морейра
Д.
1999
.Метаболический симбиоз у эукариот
.Trends Biochem Sci
.24
(3
):88
–93
.
Лопес-Гарсия
P
,Верещака
А
,Морейра
Д.
2007
.Эукариотическое разнообразие, связанное с карбонатами и поверхностью раздела флюид-морская вода в гидротермальном поле Лост-Сити
.Энвайрон Микробиол
.9
(2
):546
–554
.
Лопес-Гарсия
P
,Морейра
Д.
2015
.Открытые вопросы о происхождении эукариот
.Тренды Ecol Evol
.30
(11
):697
–708
.
Лопес-Гарсия
P
,Морейра
Д.
2020
.Пересмотр гипотезы синтрофии происхождения эукариот
.Нат Микробиол
.5
(5
):655
–667
.
Мацишевский
К
,Карнковска
А.
2019
.Мне остаться или уйти? Задержка и потеря компонентов в рудиментарных эндосимбиотических органеллах
.Curr Opin Genet Dev
.58-59
:33
–39
.
Мартин
Дж
,Эттема
TJG.
2013
.От археи к эукариоту: темные века эволюции эукариотической клетки
.Биохим Сок Т
.41
(1
):451
–457
.
Мартин
WF
,Мюллер
М.
1998
.Водородная гипотеза первого эукариота
.Природа
392
(6671
):37
–41
.
Мартин
WF
,Гарг
С
,Зиморский
В.
2015
.Эндосимбиотические теории происхождения эукариот
.Philos Trans R Soc Lond B Biol Sci
.370
(1678
):20140330
.
Мартин
WF
, и другие.2017а
.Позднее митохондриальное происхождение является артефактом
.Геном Биол. Эвол
.9
(2
):373
–379
.
Мартин
WF
,Тиленс
АГМ
,Ментел
М
,Гарг
СГ
,Гулд
СБ.
2017b
.Физиология фагоцитоза в контексте митохондриального происхождения
.Микробиол Мол Биол Ред.
.81
(3
):17
.
Мейнард Смит
Дж
,Сатмари
E.
1995
. Основные переходы в эволюции. Оксфорд: Издательство Оксфордского университета.
Мехта
АП
, и другие.2018
.Создание эндосимбионтов дрожжей как шаг к эволюции митохондрий
.Proc Natl Acad Sci U S A
.115
(46
):11796
–11801
.
Мерешковский
С.
1905
. Über Natur und Ursprung der Chromatophoren im Pflanzenreiche. Биол Центральный бл. 25:293-604. [Английский перевод в Martin W, Kowallik K. 1999 Аннотированный английский перевод статьи Мерешковского 1905 года «Über Natur und Ursprung der Chromatophoren im Pflanzenreiche. евро. Дж. Фикол. 34, 287–295].
Мерешковский
С.
1910
. Английский перевод в Коваллик К.В., Мартин В.Ф. 2021. Происхождение симбиогенеза: аннотированный английский перевод статьи Мерешковского 1910 года о теории двух плазменных линий. БиоСист. биол. Центральный бл. 30:104281. Theorie der zwei Plasmaarten als Grundlage der Symbiogenesis, einer neuen Lehre von der Entstehung der Organismen. 278–288; 289–303; 321–347; 353–367. [199].
Мишо
RE.
1999
. Дарвиновская динамика. Оксфорд: Издательство Оксфордского университета.
Мельницы
ДБ.
2020
.Происхождение фагоцитоза в истории Земли
.Интерфейс Фокус
.10
(4
):20200019
.
Мора
С
,Титтензор
P
,Адл
СМ
,Симпсон
АГ
,Червь
Б.
2011
.Сколько видов существует на Земле и в океане?
ПЛОС Биол
.9
(8
):e1001127
.
Мюллер
М
, и другие. 2012
.Биохимия и эволюция анаэробного энергетического обмена у эукариот
.Микробиол Мол Биол Ред.
.76
(2
):444
–495
.
Нагиес
FSP
,Брюкнер
Дж
,Триа
ФДК
,Мартин
ВФ.
2020
.Спектр вертикальности генов
.ПЛОС Жене
.16
(11
):e1009200
.
Никлас
КДж
,Кобб
ЭД
,Кроуфорд
ДР.
2013
.Эво-дево многоядерных клеток, тканей и организмов и альтернативный путь к многоклеточности
.Эвол Дев
.15
(6
):466
–474
.
Новак
Массачусетс.
2006
. Эволюционная динамика: исследование уравнений жизни. Кембридж: Издательство Гарвардского университета.
Песоле
Г
, и другие.2012
.Забытый геном
. EMBO Реп.
.13
(6
):473
–474
.
Питтис
АА
,Габальдон
Т.
2016
.Позднее приобретение митохондрий хозяином химерного прокариотического происхождения
.Природа
531
(7592
):101
–104
.
Радзвилавичюс
АЛ
,Блэкстоун
СЗ.
2018
.Новый взгляд на эволюцию индивидуальности
.Biol Rev Camb Philos Soc
.93
(3
):1620
–1633
.
Рамеш
МА
,Малик
СБ
,Логсдон
Дж.М.
младший2005
.Филогеномный перечень мейотических генов: свидетельство пола у Giardia и раннее эукариотическое происхождение мейоза
.Карр Биол
.15
(2
):185
–191
.
Рэнд
Немецкая марка
,Моссман
Дж. А.
2020
.Митоядерный конфликт и сотрудничество управляют интеграцией генотипов, фенотипов и окружающей среды
.Philos Trans R Soc B
.375
(1790
):201
.
Саган
Л.
1967
.О происхождении митозирующих клеток
.J Теорет Биол
.14
(3
):225
–274.
Санчес-Баракальдо
P
,Ворон
ДЖА
,Пизани
Д
,Нолл
АХ.
2017
.Ранние фотосинтезирующие эукариоты населяли слабосоленые местообитания
.Proc Natl Acad Sci U S A
.114
(37
):E7737
–E7745
.
Шредер
JM
, и другие.2011
.EB1 и EB3 способствуют биогенезу ресничек с помощью нескольких механизмов, связанных с центросомами
.J Cell Sci
.124
(Pt 15
):2539
–2551
.,
Шалев
Y
,Тургеман-Гротт
I
,Тамир
А
,Эйхлер
Дж
,Гофна
У.
2017
.Гликозилирование клеточной поверхности необходимо для эффективного спаривания Haloferax volcanii
.Фронт Микробиол
.8
:1253
.
Шимамото
Y
,Форт
С
,Капур
ТМ.
2015
.Измерение сил толкания и торможения, генерируемых ансамблями кинезина-5, сшивающими две микротрубочки
.Dev Cell
.34
(6
):669
–681
.
Ситте
Р
,Циглер
Н
,Эрендорфер
Ф
,Бресинский
А.
1991
.Strasburger, Lehrbuch der Botanik für Hochschulen
. 33-е изд.Штутгарт
:Густав Фишер
.
Спанг
А
, и другие.2015
.Сложные археи, занимающие промежуточное положение между прокариотами и эукариотами
.Природа
.521
(7551
):173
–179
.
Шпейер
Д
,Лукеш
Дж
,Элиаш
М.
2015
.Секс — вездесущий, древний и неотъемлемый атрибут эукариотической жизни
.Proc Natl Acad Sci U S A
.112
(29
):8827
–8834
.
Штехманн
А
,Кавалер-Смит
Т.
2002
.Укоренение дерева эукариот с использованием производного слияния генов
.Наука
297
(5578
):89
–91
.
Страсбургер
E.
1908
.Einiges über Characeen und Amitose. Wiesner Festschrift im Auftrage Des Festkomitees Redigiert Von K. Linsbauer
.Вена: К. Конеген.
р.24
–47
.
Сатмари
E
,Деметра
Л.
1987
.Модель стохастического корректора
.Дж Теор Биол
.128
(4
):463
–486
.
Тан
Q
,Панг
К
,юаней
XL
,Сяо
S-H.
2020
.Многоклеточный хлорофит возрастом один миллиард лет
.Нат Экол Эвол
.4
(4
):543
–547
.
Тромер
ЕС
,Ван Хофф
JJE
,Копс
GJPL
,Снел
Б.
2019
.Мозаичное происхождение эукариотических кинетохор
.Proc Natl Acad Sci U S A
.116
(26
):12873
–12882
.
Триа
ФДК
и другие.2021
. Дупликации генов прослеживают митохондрии до начала сложности эукариот. Геном Биол Эвол.
Воссбринк
CR
, и другие.1987
.Последовательность рибосомной РНК предполагает, что микроспоридии являются очень древними эукариотами
.Природа
326
(6111
):411
–414
.,
Воссеберг
Дж
, и другие.2021
.Определение времени происхождения эукариотической клеточной сложности с древними дупликациями
. Нат Экол Эвол
.5
(1
):92
–99
.
Юбуки
N
,Леандер
БС.
2013
.Эволюция центров организации микротрубочек у эукариот
.Завод J
.75
(2
):230
–244
.
Заремба-Недзведска
К
, и другие.2017
.Археи Асгарда проливают свет на происхождение эукариотической клеточной сложности
.Природа
541
(7637
):353
–358
.© Автор(ы), 2021 г. Опубликовано Oxford University Press от имени Общества молекулярной биологии и эволюции.
Это статья в открытом доступе, распространяемая в соответствии с условиями некоммерческой лицензии Creative Commons Attribution (http://creativecommons.org/licenses/by-nc/4.0/), которая разрешает некоммерческое повторное использование, распространение, и воспроизведение на любом носителе при условии надлежащего цитирования оригинальной работы. Для коммерческого повторного использования, пожалуйста, свяжитесь с [email protected]
Происхождение эукариот – эволюционная биология
Обзоры эукариогенеза см. в Martin, et al.2015 и Baum 2015. Martin 2005 представляет более старый, но все еще полезный обзор, тогда как Harold 2014 представляет собой доступное исследование эволюции клеток длиной в книгу. Гулд и др. 2008 фокусируется на приобретении пластид и последующих дополнительных эндосимбиотических событиях. Кунин и др. 2010 и Ломбард и др. 2012 обсуждают регуляторные сети белков и химию мембран в трех сферах жизни и их значение для эукариогенеза. Эме и др. 2014 связывает молекулярные данные, которые определяют большую часть поля, с ископаемыми свидетельствами.Уильямс и др. 2013 обобщает филогенетические аргументы в пользу филогенетической модели, которая доминирует в современном мышлении, а именно, что археи/эукариоты и бактерии представляют собой два основных домена жизни. Эме и др. 2017 строится на отношениях между археями и моделями эукариогенеза.
Баум, Д. А. 2015. Сравнение аутогенных теорий происхождения эукариотических клеток. Американский журнал ботаники 102:1954–1965.
Сравнивает различные аутогенные теории происхождения ядра и эукариогенеза в целом в контексте клеточной топологии, соответствия современной клеточной биологии и времени приобретения митохондрий.
Эме, Л., С. К. Шарп, М. В. Браун и др. 2014. О возрасте эукариот: оценка данных окаменелостей и молекулярных часов. Перспективы Колд-Спринг-Харбор в биологии 6:a016139.
Обзор филогенетических и ископаемых данных о возрасте эукариот.
Эме, Л., А. Спанг, Дж. Ломбард, К. У. Стейрс и Т. Дж. Г. Эттема. 2017. Археи и происхождение эукариот. Nature Reviews Microbiology 15:711–723.
Обсуждает модели эукариогенеза в свете недавно открытых и охарактеризованных линий архей.
Гулд С. Б., Р. Ф. Уоллер и Г. И. Макфадден. 2008. Эволюция пластид. Ежегодный обзор биологии растений 59:491–517.
Обеспечивает всесторонний обзор эволюции пластид, включая первичные и вторичные эндосимбиозы, нацеливание белков на пластиды и метаболизм пластид.
Гарольд Ф. М. 2014. В поисках клеточной истории: эволюция строительных блоков жизни .Чикаго: Чикагский ун-т. Нажмите.
Дает синтетический обзор происхождения и эволюции клеток, уделяя особое внимание происхождению эукариот.
Кунин Е. В., Дэкс Дж., Дулитл В. и др. 2010. Происхождение и ранняя эволюция эукариот в свете филогеномики. Биология генома 11:209.
Анализ происхождения ключевых регуляторных модулей эукариотических белков с использованием сравнительной геномики.
Ломбард, Дж., П.Лопес-Гарсия и Д. Морейра. 2012. Ранняя эволюция липидных мембран и три домена жизни. Nature Reviews Microbiology 10:507–515.
Обзор молекулярного состава фосфолипидных мембран архей и бактерий и последствия для моделей эукариогенеза.
Martin, W. F. 2005. Архебактерии (археи) и происхождение эукариотического ядра. Текущее мнение по микробиологии 8:630–637.
Обобщает разнообразие моделей происхождения ядерного компартмента, выступая против ядерных эндосимбиотических моделей.
Мартин В. Ф., С. Гарг и В. Зиморски. 2015. Эндосимбиотические теории происхождения эукариот. Philosophical Transactions of the Royal Society B: Biological Science 370.1678: 20140318.
Обзор моделей эукариогенеза с историческим уклоном, уделяя особое внимание происхождению ядерного и митохондриального компартментов, а также метаболическим соображениям.
Williams, T. A., P. G. Foster, C. J. Cox, et al. 2013. Архейное происхождение эукариот поддерживает только два основных домена жизни. Природа 504:231–236.
Обобщает поддержку существования только двух основных доменов жизни, при этом эукариоты встроены в парафилетические археи.
Мембраны, энергетика и эволюция между прокариотами и эукариотами
Любая попытка определить значение мембран для клеточной эволюции должна учитывать высокую стоимость биосинтеза липидных молекул. Существует два способа количественной оценки таких затрат. Во-первых, с эволюционной точки зрения стоимость синтеза молекулы принимается как сумма прямого использования АТФ в пути биосинтеза плюс косвенная потеря АТФ в результате использования метаболических предшественников, которые в противном случае были бы преобразованы в АТФ и доступны для альтернативных клеточных функций (Akashi and Gojobori, 2002; Lynch, Marinov, 2015).Во-вторых, для простой количественной оценки прямого вклада в общую потребность клетки в АТФ затраты на отклонение метаболических предшественников игнорируются.
Путем суммирования общих затрат всех молекул, лежащих в основе клеточного признака, и масштабирования по расходу энергии клетки в течение жизни можно получить меру относительного расхода энергетического бюджета клетки, связанного с формированием и поддержанием признака. Затем это измерение, sc, можно рассматривать как частичное увеличение энергетического бюджета клетки, которое могло бы быть направлено на рост, размножение и выживание в отсутствие таких вложений, игнорируя прямые преимущества приспособленности, связанные с выражением признака, sa. Чтобы отбор был эффективным, чистое селективное преимущество признака sn=sa-sc должно превышать мощность случайного генетического дрейфа, 1/Ne у гаплоидных видов и 1/(2Ne) у диплоидных, где Ne — эффективная численность населения.
Большинство клеточных мембран преимущественно состоят из глицерофосфолипидов, которые, несмотря на наличие различных головных групп (например, глицерин, холин, серин и инозитол), имеют общую стоимость биосинтеза на молекулу (в единицах гидролиза АТФ, включая стоимость отведения промежуточные метаболиты)
(2а)cL≃320+[38⋅(NL−16)]+(6⋅NU), (2b)cL≃340+[40⋅(NL−16)]+(6⋅NU),
у бактерий и эукариот соответственно, где NL — средняя длина цепи жирной кислоты, а NU — среднее количество ненасыщенных атомов углерода в цепи жирной кислоты (дополнительный материал).Хотя варианты глицерофосфолипидов используются у различных видов (Guschina and Harwood, 2006; Geiger et al., 2010), они структурно достаточно сходны, так что предыдущие выражения должны по-прежнему обеспечивать превосходные приближения первого порядка. Приведенные (прямые) затраты, которые не учитывают потерю АТФ-генерирующего потенциала из-за утечки метаболических предшественников, составляют
(3a)cL′≃110+[7⋅(NL−16)]+(6⋅NU), (3b)cL′≃120+[9⋅(NL−16)]+(6⋅NU),
у бактерий и эукариот соответственно.С точки зрения общего энергетического баланса клетки эволюционная стоимость липидной молекулы равна cL/CT.
Для большинства липидов в биологических мембранах 14≤NL≤22 и 0≤NU≤6, поэтому стоимость молекулы липида обычно находится в диапазоне cL≃200–600 АТФ, хотя среднее значение для всех липидов, развернутых в видоспецифичных мембран значительно уже (внизу). Кардиолипин, который редко составляет более 20% мембранных липидов, является исключительным, имея эволюционную стоимость ~ 640 АТФ / молекула (и уменьшенную стоимость ~ 240 АТФ).Чтобы представить эти расходы в перспективе, эволюционные затраты на биосинтез каждого из четырех нуклеотидов составляют ≈ 50 гидролизов АТФ на молекулу (Lynch and Marinov, 2015), тогда как средняя стоимость аминокислоты составляет ≈30 АТФ (Atkinson, 1970; Akashi). и Годжобори, 2002).
Применение предыдущих выражений к известным композициям мембран клеток показывает, что затраты на биосинтез эукариотических липидов выше, чем у бактерий (дополнительная таблица). Например, для различных видов бактерий средняя прямая стоимость молекулы липида в плазматической мембране составляет 123 (SE = 3) АТФ, тогда как для эукариот она составляет 143 (2).Последняя оценка идентична средней, полученной для целых эукариотических клеток, но стоимость митохондриальных липидов особенно высока 155 (5). Эти повышенные затраты у эукариот являются совместным эффектом стоимости митохондриального экспорта оксалоацетата для образования ацетил-КоА и тенденции эукариотических липидов иметь более длинные цепи, содержащие больше ненасыщенных атомов углерода.
Чтобы понять общие биоэнергетические затраты, связанные с мембранами, нам нужна информация о количестве липидных молекул, необходимых для образования мембраны, что эквивалентно общей площади поверхности мембраны, деленной на количество липидных молекул на единицу площади поверхности и умноженной на на два для учета липидного двойного слоя. Оценки площадей головной группы липидов мембран в основном находятся в пределах 10% от среднего значения 6,5 × 10 −7 мкм 2 (Petrache et al., 2000; Kučerka et al., 2011), поэтому стоимость мембраны (в единицах АТФ, без учета липидного обмена и пространства, занимаемого трансмембранными белками) составляет
(4) CL≃(3,1×106)⋅c¯L⋅A,
, где A — площадь поверхности мембраны в единицах мкм 2 , а c¯L — средняя стоимость липида.
Имеется достаточно информации об общих инвестициях в митохондриальные мембраны, чтобы можно было сделать общее заявление.В эукариотическом домене общая площадь поверхности митохондрий (внутренняя плюс внешняя мембрана, суммированная по всем митохондриям, в мкм 2 ) зависит от объема клетки (V, в единицах мкм 3 ) как 3,0 В 0,99 ( Рисунок 1c; SE точки пересечения и наклона на логарифмических графиках составляют 0,22 и 0,08 соответственно). Применяя это к уравнению (4) со средней общей стоимостью митохондриальных липидов (c¯L=440 АТФ/молекула; Приложение 1 – таблица 4) и используя выражение для общих потребностей роста клетки, уравнение (1b) , относительная стоимость липидов митохондриальных мембран составляет
(5)
ск≃0. 15V0.02,
или ~15% от общего бюджета роста эукариотической клетки минимального размера (1 мкм 3 ) и почти не зависит от размера клетки в пределах диапазона, обычно встречающегося у эукариот (SE экспоненты составляет 0,08). Прямой вклад митохондриальных мембранных липидов в бюджет роста клетки составляет ~36% от этой общей стоимости. Эти затраты на митохондриальные мембраны представляют собой базовую цену, не понесенную прокариотами, связанную с перемещением биоэнергетики внутрь эукариотических клеток, то есть ~ 5%.К сожалению, дополнительные затраты на поддержание митохондриальных липидов неизвестны, но для быстрорастущих клеток подавляющая часть энергетического бюджета клетки направляется на рост (Lynch and Marinov, 2015), поэтому вышеуказанные затраты по-прежнему должны применяться как первоочередные. приближения; для медленно растущих клеток затраты будут выше или ниже в зависимости от того, выше или ниже стоимость обслуживания митохондриальной мембраны по сравнению с общим обслуживанием клетки. Белки не занимают> 50% мембран, поэтому учет этого изменит предыдущие результаты в <2 раза.
Ожидается, что для прокариотических клеток без внутренних мембран относительный вклад клеточной мембраны в общий энергетический баланс клетки будет уменьшаться с увеличением размера клетки из-за снижения отношения площади поверхности к объему. Для крошечных клеток Leptospira interrogans и Mycoplasma pneumoniae (средний объем 0,03 и 0,22 мкм 3 соответственно) ~ 63 и 43% бюджета роста клетки должно быть отнесено к плазматической мембране, но для крупнее Bacillus subtilis и Escherichia coli (в среднем 1.4 и 1,0 мкм 3 соответственно), эти вклады падают до ~ 14 и 19%, и можно ожидать, что они будут продолжать снижаться с дальнейшим увеличением размера клетки, масштабируясь обратно пропорционально линейному размеру клетки.
Напротив, из-за увеличения инвестиций во внутренние мембраны доля энергетического бюджета эукариотической клетки, направляемая на мембраны, не уменьшается с увеличением размера клетки. Хотя есть только несколько типов эукариотических клеток, для которых эта проблема может быть оценена количественно (таблица 1), данные охватывают три порядка по объему клеток и единообразно предполагают, что от 10 до 30% общего бюджета роста приходится на липиды. биосинтеза, и что все большая доля таких затрат связана с внутренними мембранами в клетках с увеличивающимся размером.Пикопланктонная водоросль Ostreococcus , имеющая размер клетки всего 0,22 мкм 3 (меньше, чем у многих прокариот), расходует ~32% своего энергетического бюджета на мембраны, и 44% этих затрат (~18% общий клеточный бюджет) связаны с внутренними мембранами. Клетка млекопитающего среднего размера тратит примерно 30% своего энергетического бюджета на мембраны, но 96% этих затрат (~29% общего клеточного бюджета) связаны с внутренними мембранами.
Вклад мембран в общие затраты на рост клеток.
Ot обозначает зеленую водоросль Ostreococcus tauri , Sc — дрожжи Saccharomyces cerevisiae , Ds — зеленую водоросль Dunaliella salina и Ss — клетку поджелудочной железы свиньи ( Sus scrofa ); ссылки даны в дополнительном материале. Объемы клеток и общая площадь мембран выражены в мкм 3 и мкм 2 соответственно, причем последние исключают мембраны, связанные с пластидами у видов водорослей.Доля общего бюджета роста клеток, выделяемая на мембраны, получается соотношением уравнений (1b) и (4) с использованием видово-специфических приведенных затрат в таблице 1, где они доступны, и в противном случае с применением средних значений для эукариотических видов; эта общая стоимость затем распределяется на пять различных дробных взносов в следующих строках.
| От | Sc | Ds | Сс | |||
|---|---|---|---|---|---|---|
| объем ячейки | 1 | 44 | 591 | 1060 | ||
| Всего мембран | 15 | 204 | 2299 | 12952 | ||
| Доля абсолютного бюджета роста клеток | 0.324 | 0.096 | 0,094 | 0,302 | ||
| Плазменная мембрана | 0,556 | 0,328 | 0,134 | 0,044 | ||
| Митохондрии | 0,243 | 0,359 | 0,197 | 0,223 | ||
| Nucleus | 0.113 | 0.085 | 0,034 | 0,034 | 0,034 | 0.008 |
| Эндоплазматический ретикулум + Golgi | 0,034 | 0,11 | 0.318 | 0,706 | ||
| везикулы / вакуулы | 0,055 | 0.114 | 0.316 | 0.019 |
Взятые вместе, эти наблюдения подразумевают, что использование внутренних мембран представляет собой значительную утечку общего энергетического бюджета эукариотических клеток, гораздо большую, чем можно было бы ожидать у бактерий сопоставимого размера. Более того, поскольку липиды, связанные только с митохондриями, составляют от 20% до 35% инвестиций эукариотической клетки в мембраны (таблица 1), энергетическая нагрузка локализации мембранной биоэнергетики в митохондриях является существенной.
Наконец, учитывая, что наблюдения, обобщенные на рис. 1a,b, получены из различных исследований, вероятно, с множеством уникальных неточностей, стоит рассмотреть, согласуются ли общие выводы с известной способностью АТФ-синтазы. Во-первых, следует отметить, что лишь часть энергии, затрачиваемой на биосинтез, получается непосредственно за счет хемиосмотической активности АТФ-синтазы. Например, биосинтез аминокислот включает примерно 1,5 окисления НАДН и НАДФН на каждый гидролиз АТФ (Akashi and Gojobori, 2002).Если предположить, что каждый из первых эквивалентен ~ 3 гидролизам АТФ, это означает, что только ~ 18% энергии, затрачиваемой на биосинтез аминокислот, связано с гидролизом АТФ. Как отмечено в дополнительном тексте, отношение использования НАДН/НАДФН к АТФ больше порядка 2,0 в биосинтезе липидов, что снижает прямые инвестиции в АТФ до ~14%. клетка связана с синтезом мономерных строительных блоков белков и мембран, только ~ 15% биосинтетической энергии может быть получено из гидролиза АТФ.
Зная известные потребности в энергии для поддержания и роста клетки, время клеточного деления и количество комплексов АТФ-синтазы на клетку, можно оценить требуемую скорость превращения АДФ → АТФ на комплекс. Используя ранее представленные клеточные энергетические данные (Lynch and Marinov, 2015) и распространенность АТФ-синтазы в Приложении 1-таблице 2, после дисконтирования максимальных значений на 85%, предполагаемые необходимые скорости производства АТФ/комплекс/сек составляют: 2109, 221 и 19 соответственно для бактерий B.subtilis , E. coli и M. pneumoniae , а также 1440 и 329 для дрожжей S. cerevisiae и S. pombe . Было предпринято несколько попыток оценить максимальную скорость оборота (в секунду) АТФ-синтазы F0F1, обычно в реконструированных липосомах, и в среднем она составляет 195/с у бактерий (Etzold et al., 1997; Slooten and Vandenbranden, 1989; Toei et al. ., 2007), 295 в пластидах сои (Schmidt, Gräbe, 1985; Junesch, Gräber, 1991), 120 в S.cerevisiae (Förster et al., 2010) и 440 в сердце крупного рогатого скота (Matsuno-Yagi and Hatefi, 1988). Таким образом, учитывая, что значительная часть комплексов, вероятно, будет неправильно собрана в искусственных мембранах, оценки количества оборотов АТФ, генерируемых на клетку, на основе энергетического баланса, по-видимому, согласуются с известной способностью АТФ-синтазы.
Этот странный микроб может ознаменовать один из больших скачков жизни
Причудливый микроб с щупальцами, обнаруженный на дне Тихого океана, может помочь объяснить происхождение сложной жизни на этой планете и решить одну из самых глубоких тайн биологии, сообщили ученые в среду .
Два миллиарда лет назад простые клетки дали начало гораздо более сложным клеткам. Биологи десятилетиями пытались понять, как это произошло.
Ученым давно известно, что на пути эволюции должны были быть предшественники. Но, судя по летописи окаменелостей, сложные клетки возникли просто из ниоткуда.
Новый вид, названный Prometheoarchaeum, оказывается именно такой переходной формой, помогающей объяснить происхождение всех животных, растений, грибов — и, конечно же, человека.Об исследовании сообщается в журнале Nature.
«На самом деле это довольно круто — это окажет большое влияние на науку», — сказала Криста Шлепер, микробиолог из Венского университета, которая не участвовала в новом исследовании.
Наши камеры забиты контейнерами. Например, они хранят ДНК в ядре и вырабатывают топливо в отсеках, называемых митохондриями. Они разрушают старые белки внутри крошечных хозяйственных машин, называемых лизосомами.
Наши клетки также строят себе скелет из нитей, построенных из строительных блоков, подобных Lego.Растягивая одни нити и разрывая другие, клетки могут менять свою форму и даже перемещаться по поверхности.
Виды, которые разделяют эти сложные клетки, известны как эукариоты, и все они происходят от общего предка, жившего примерно два миллиарда лет назад.
До этого мир был домом только для бактерий и группы маленьких простых организмов, называемых археями. Бактерии и археи не имеют ядер, лизосом, митохондрий или скелета.
Биологи-эволюционисты долго ломали голову над тем, как эукариоты могли развиться из таких простых предшественников.
В конце 1900-х годов исследователи обнаружили, что митохондрии когда-то были свободноживущими бактериями. Каким-то образом их затянуло внутрь другой клетки, обеспечив своим хозяином новое топливо.
В 2015 году Тийс Эттема из Упсальского университета в Швеции и его коллеги обнаружили фрагменты ДНК в отложениях, извлеченных из Северного Ледовитого океана. Фрагменты содержали гены вида архей, которые, по-видимому, были тесно связаны с эукариотами.
Доктор Эттема и его коллеги назвали их археями Асгарда.(Асгард — дом скандинавских богов.) ДНК этих таинственных микробов была обнаружена в реке в Северной Каролине, горячих источниках в Новой Зеландии и других местах по всему миру.
Асгардские археи полагаются на ряд генов, которые ранее были обнаружены только у эукариот. Возможно, эти микробы использовали эти гены для тех же целей — или для чего-то еще.
«Пока у вас нет организма, вы не можете быть уверены», — сказал доктор Шлепер.
Масару К. Нобу, микробиологу из Национального института передовых промышленных наук и технологий в Цукубе, Япония, и его коллегам удалось вырастить эти организмы в лаборатории.Усилия заняли более десяти лет.
Микробы, приспособленные к жизни на холодном морском дне, живут в замедленном темпе. Prometheoarchaeum может делиться до 25 дней. Напротив, кишечная палочка делится каждые 20 минут.
Проект начался в 2006 году, когда исследователи подняли отложения со дна Тихого океана. Первоначально они надеялись изолировать микробы, питающиеся метаном, которые можно было бы использовать для очистки сточных вод.
В лаборатории исследователи воспроизвели условия на морском дне, поместив осадок в камеру без доступа кислорода.Они закачивали метан и извлекали смертоносные отработанные газы, которые могли убить живущих там микробов.
Грязь содержала много видов микробов. Но к 2015 году исследователи выделили новый интригующий вид архей. И когда доктор Эттема и его коллеги объявили об открытии ДНК архей Асгарда, японские исследователи были потрясены. Их новый живой микроб принадлежал к этой группе.
Затем исследователи предприняли более кропотливые исследования, чтобы понять новый вид и связать его с эволюцией эукариот.
Исследователи назвали микроб Prometheoarchaeum syntrophicum в честь Прометея, греческого бога, который дал людям огонь, вылепив их из глины.
«Двенадцать лет микробиологии, которые потребовались, чтобы дойти до точки, когда вы можете увидеть ее под микроскопом, просто поразительны», — сказал Джеймс Макинерни, биолог-эволюционист из Ноттингемского университета, который не участвовал в исследовании.
Под микроскопом Prometheoarchaeum оказался странным зверем.Поначалу микроб выглядит как крошечная сфера, но в течение нескольких месяцев он отрастает длинными ветвящимися щупальцами и выпускает флотилию покрытых мембраной пузырей.
Это оказалось еще более странным, когда исследователи исследовали внутреннюю часть камеры. Доктор Шлепер и другие исследователи ожидали, что археи Асгарда использовали свои эукариотоподобные белки для создания некоторых эукариотоподобных структур внутри своих клеток. Но это не то, что обнаружила японская команда.
«Внутри нет никакой структуры, только ДНК и белки», — сказал доктор.Нобу.
Это открытие позволяет предположить, что белки, которые эукариоты использовали для построения сложных клеток, сначала выполняли другие функции, и только позже им поручили новую работу.
Доктор Нобу и его коллеги сейчас пытаются выяснить, что это были за рабочие места. Возможно, сказал он, Prometheoarchaeum создает свои щупальца с генами, которые позже использовались эукариотами для построения клеточных скелетов.
Доктор Шлепер хотел увидеть больше доказательств этой идеи. «У других архей очень красивые руки», — заметила она.Но эти другие виды не используют белки, столь похожие на наши.
До открытия Prometheoarchaeum некоторые исследователи подозревали, что предки эукариот жили как хищники, поедая более мелких микробов. Таким образом они могли поглотить первые митохондрии.
Но Prometheoarchaeum не подходит под это описание. Команда доктора Нобу часто обнаруживала, что микроб прилипает к бокам бактерий или других архей.
Вместо того, чтобы охотиться на добычу, Prometheoarchaeum, кажется, зарабатывает на жизнь, поедая фрагменты белков, проплывающих мимо.Его партнеры питаются его отходами. Они, в свою очередь, обеспечивают Prometheoarchaeum витаминами и другими необходимыми соединениями.
Доктор Нобу предположил, что вид архей Асгарда на морском дне затягивает бактерии паутиной щупалец, втягивая их в еще более тесную связь. В конце концов, он поглотил бактерии, которые превратились в митохондрии, питающие каждую сложную клетку.